Hormones and the auditory system: A review of physiology and pathophysiology
20
REVIEW HORMONES AND THE AUDITORY SYSTEM: A REVIEW OF PHYSIOLOGY AND PATHOPHYSIOLOGY D. AL-MANA, a,d B. CERANIC, b * O. DJAHANBAKHCH c AND L. M. LUXON a,d a Department of Neuro-Otology, The National Hospital for Neurology and Neurosurgery, Queen Square, London WC1N 3BG, UK b Department of Audiology, St. George’s Hospital, Blackshaw Road, London SW17 0QT, UK c Newham University Hospital, Academic Department of Obstetrics & Gynaecology, Barts and The London Queen Mary’s School of Medi- cine and Dentistry, Glen Road, London E13 8SL, UK d Academic Unit of Audiological Medicine, University College London, Institute of Child Health, 30 Guilford Street, London WC1N 1EH, UK Abstract—This review explores the potential role of hor- mones in modulating the auditory function. The review de- scribes four groups of hormones (the hormones of the circa- dian cycle, reproduction, stress response and the fluid and electrolyte balance), their physiological variations, interac- tions, as well as the physiological basis for their effect on the auditory system. Possible contribution of hormones to pathophysiology of auditory dysfunctions, including hyper- acusis, tinnitus, Menière’s disease and pre-menstrual audi- tory dysfunction, has also been discussed. Published by Elsevier Ltd on behalf of IBRO. Key words: auditory, hearing, hormones. Contents Functional anatomy of the auditory system 882 Anatomy and physiology 882 Modulation of the auditory system 882 How the different hormones may influence the auditory system 883 Reproductive steroids 883 Estrogen 884 Progesterone 884 Stress-related hormones 884 Glucocorticoids 884 Catecholamines and endogenous opioids 884 Hormones regulating fluid and electrolyte balance 885 Aldosterone 885 Vasopressin 885 Melatonin 885 Physiological variation in hormones and the effects on auditory function 885 Circadian cycle and auditory function 886 Gender differences in auditory function 886 The ovarian cycle and auditory function 887 Auditory thresholds 889 SOAEs 889 Transient OAEs 890 ABR 890 Menopause and auditory function 890 Pregnancy and auditory function 891 Stress and auditory function 891 Auditory pathology and hormones 892 Hyperacusis and tinnitus 892 Menière’s disease and endocrine system 893 Reproductive hormones 893 Stress-related hormones 893 Fluid and electrolyte related hormones 893 Pre-menstrual syndrome and auditory function 893 Conclusion 894 References 894 Considerable anecdotal evidence and limited information from previous studies suggest that auditory function may be influenced by hormones. Recent advances in the fields of neuroendocrinology and neuropharmacology, together with the development of new assessment techniques have pro- vided insights into the contribution of hormones in the regu- lation of the circadian cycle, reproduction, stress responses and the maintenance of fluid and electrolyte balance. In this context, this review examines the potential role of hormones in modulating the auditory system and in the development of pathological conditions in the auditory system. Four areas will be considered: The functional anatomy of the auditory system. A description of the groups of hormones and their interac- tions, together with the physiological basis and proposed mechanisms of effects of hormones on auditory function. Physiological hormonal variations, including gender differ- ences, and possible implications for the auditory sys- tem. Possible contribution of hormones to pathophysiology of auditory dysfunction, including hyperacusis, tinnitus, *Corresponding author. Tel: 44-0-20-8725-1195; fax: 44-2-8725- 1874. E-mail address: [email protected] (B. Ceranic). Abbreviations: ABR, auditory brain stem– evoked response; ADH, an- tidiuretic hormone; ART, acoustic reflex threshold; CN, cochlear nu- cleus; DPOAE, distortion product otoacoustic emission; ER, estrogen receptor; FSH, follicular stimulating hormone; GnRH, gonadotrophin releasing hormone; GR, glucocorticoid receptor; HIOMT, hydroxyin- dole-O-methyltransferase; HRT, hormone replacement therapy; IHC, inner hair cell; LH, luteinizing hormone; LL, lateral leminiscus; MGB, medial geniculate body; MOC, medial olivocochlear system; NAT, N-acetyltransferase; OAE, otoacoustic emission; OC, oral contracep- tives; OCB, olivary cochlear bundle; OHC, outer hair cell; OVX, oo- phorectomized; PMDD, premenstrual dysphoric disorder; PMS, pre- menstrual syndrome; SCN, suprachiasmatic nucleus; SOAE, sponta- neous otoacoustic emission; SOC, superior olivary complex. Neuroscience 153 (2008) 881–900 0306-4522/08$32.000.00 Published by Elsevier Ltd on behalf of IBRO. doi:10.1016/j.neuroscience.2008.02.077 881
Transcript of Hormones and the auditory system: A review of physiology and pathophysiology

R
HP
DAa
ab
Lc
Gcd
I
AmsdetapatE
K
F
H
*1EAtcrrdimNtpmn
Neuroscience 153 (2008) 881–900
0d
EVIEW
ORMONES AND THE AUDITORY SYSTEM: A REVIEW OF
HYSIOLOGY AND PATHOPHYSIOLOGYP
A
CR
Cfintvlacip
TA
P
P
. AL-MANA,a,d B. CERANIC,b* O. DJAHANBAKHCHc
ND L. M. LUXONa,d
Department of Neuro-Otology, The National Hospital for Neurologynd Neurosurgery, Queen Square, London WC1N 3BG, UK
Department of Audiology, St. George’s Hospital, Blackshaw Road,ondon SW17 0QT, UK
Newham University Hospital, Academic Department of Obstetrics &ynaecology, Barts and The London Queen Mary’s School of Medi-ine and Dentistry, Glen Road, London E13 8SL, UK
Academic Unit of Audiological Medicine, University College London,nstitute of Child Health, 30 Guilford Street, London WC1N 1EH, UK
bstract—This review explores the potential role of hor-ones in modulating the auditory function. The review de-
cribes four groups of hormones (the hormones of the circa-ian cycle, reproduction, stress response and the fluid andlectrolyte balance), their physiological variations, interac-ions, as well as the physiological basis for their effect on theuditory system. Possible contribution of hormones toathophysiology of auditory dysfunctions, including hyper-cusis, tinnitus, Menière’s disease and pre-menstrual audi-ory dysfunction, has also been discussed. Published bylsevier Ltd on behalf of IBRO.
ey words: auditory, hearing, hormones.
Contentsunctional anatomy of the auditory system 882Anatomy and physiology 882Modulation of the auditory system 882
ow the different hormones may influence the auditory system883
Reproductive steroids 883Estrogen 884Progesterone 884
Stress-related hormones 884Glucocorticoids 884
Corresponding author. Tel: �44-0-20-8725-1195; fax: �44-2-8725-874.-mail address: [email protected] (B. Ceranic).bbreviations: ABR, auditory brain stem–evoked response; ADH, an-
idiuretic hormone; ART, acoustic reflex threshold; CN, cochlear nu-leus; DPOAE, distortion product otoacoustic emission; ER, estrogeneceptor; FSH, follicular stimulating hormone; GnRH, gonadotrophineleasing hormone; GR, glucocorticoid receptor; HIOMT, hydroxyin-ole-O-methyltransferase; HRT, hormone replacement therapy; IHC,
nner hair cell; LH, luteinizing hormone; LL, lateral leminiscus; MGB,edial geniculate body; MOC, medial olivocochlear system; NAT,-acetyltransferase; OAE, otoacoustic emission; OC, oral contracep-
ives; OCB, olivary cochlear bundle; OHC, outer hair cell; OVX, oo-horectomized; PMDD, premenstrual dysphoric disorder; PMS, pre-
enstrual syndrome; SCN, suprachiasmatic nucleus; SOAE, sponta-eous otoacoustic emission; SOC, superior olivary complex.306-4522/08$32.00�0.00 Published by Elsevier Ltd on behalf of IBRO.oi:10.1016/j.neuroscience.2008.02.077
881
Catecholamines and endogenous opioids 884Hormones regulating fluid and electrolyte balance 885
Aldosterone 885Vasopressin 885
Melatonin 885hysiological variation in hormones and the effects on auditory
function 885Circadian cycle and auditory function 886Gender differences in auditory function 886The ovarian cycle and auditory function 887
� Auditory thresholds 889� SOAEs 889� Transient OAEs 890� ABR 890
Menopause and auditory function 890Pregnancy and auditory function 891Stress and auditory function 891
uditory pathology and hormones 892Hyperacusis and tinnitus 892Menière’s disease and endocrine system 893Reproductive hormones 893Stress-related hormones 893Fluid and electrolyte related hormones 893Pre-menstrual syndrome and auditory function 893onclusion 894eferences 894
onsiderable anecdotal evidence and limited informationrom previous studies suggest that auditory function may benfluenced by hormones. Recent advances in the fields ofeuroendocrinology and neuropharmacology, together with
he development of new assessment techniques have pro-ided insights into the contribution of hormones in the regu-
ation of the circadian cycle, reproduction, stress responsesnd the maintenance of fluid and electrolyte balance. In thisontext, this review examines the potential role of hormones
n modulating the auditory system and in the development ofathological conditions in the auditory system.
Four areas will be considered:
he functional anatomy of the auditory system. description of the groups of hormones and their interac-
tions, together with the physiological basis and proposedmechanisms of effects of hormones on auditory function.
hysiological hormonal variations, including gender differ-ences, and possible implications for the auditory sys-tem.
ossible contribution of hormones to pathophysiology of
auditory dysfunction, including hyperacusis, tinnitus,
A
Tsip
fibtfiStcaoct
I
bdbtW
ntias
M
TuleCFc
1
a
m
2
FaltnpatpcdfeOIE
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900882
Menière’s disease and pre-menstrual auditory dys-function.
FUNCTIONAL ANATOMY OF THE AUDITORYSYSTEM
natomy and physiology
he auditory system consists of a variety of anatomicaltructures, from the cochlea to the auditory cortex, includ-ng both ascending (afferent) and descending (efferent)athways (Ceranic B, Luxon LM, in press) (Fig. 1).
The inner hair cells (IHC) of the organ of Corti trans-orm the mechanical energy of sound waves to electricalnformation that passes to the type I auditory afferent fi-ers. The outer hair cells (OHC) on the other hand, are
hought to act as a modulator and amplifier capable ofne-tuning the receptive function of the cochlea (Santos-acchi, 2001). The auditory signal from the organ of Corti
hen travels through the auditory nerve to the ipsilateralochlear nucleus (CN) and from there the majority of thefferent auditory fibers project to the contralateral superiorlivary complex (SOC), the lateral lemniscus (LL), inferiorolliculus (IC), medial geniculate body (MGB) to the audi-ory cortex (Chermak and Musiek, 1997).
The efferent auditory pathway is less well understood.
ig. 1. Schematic illustration of the afferent (left) and efferent (right)uditory pathways: The afferent pathway originates at the cochlear
evel, from the outer cells (OHCs), which feed mechanical oscillation tohe inner hair cells (IHCs. Neural signals are conveyed via the auditoryerve to the cochlear nuclei (CN). The fibers from the CN projectredominantly contralaterally to the superior olivary complex (SOC)nd further via the lateral lemniscus (LL), the inferior colliculus (IC) andhe medial geniculate body (MGB) to the auditory cortex. The efferentathway runs in parallel to the afferent pathway, from the cortex to theochlea, allowing multiple feedback loops, with illustrated in moreetails the most distal part: the olivocochlear bundle (OCB) consistsrom the medial bundle (MB), which projects predominantly contralat-ral fibers from the superior olivary complex (SOC) directly onto theHCs, and the lateral bundle (LB), which projects indirectly to the
HCs. (Ceranic B, Luxon LM, in press. Reproduced by permission ofdward Arnold, Publishers, Ltd.)
t arises in the auditory cortex and descends into the
rainstem to reach the cochlea (Suga et al., 2000). Theistal pathway is its best known part, the olivary cochlearundle (OCB), which originates from the SOC and projectso the cochlea through two main pathways (reviewed by
arr, 1992):
● Medial olivocochlear system (MOC) which projectsmainly to the contralateral cochlea and terminatesdirectly on the OHCs.
● Lateral olivocochlear system (LOC) which projectsmainly to the ipsilateral cochlea, and ends on the typeI afferent dendrites that terminate at the IHCs.
The exact function of the efferent auditory system isot fully understood, but it seems to act as an autoregula-ory feedback mechanism, with a predominantly inhibitorynfluence, but it may also be excitatory at different levelsnd so adjusts and improves the processing of the auditoryignal (Suga et al., 2000).
odulation of the auditory system
he baseline activity of the auditory system can be mod-lated by different mechanisms, such as the cardiovascu-
ar system, drugs, various neurotransmitter systems andxtra-auditory structures, i.e. the other structures of theNS with direct or indirect inputs to the auditory system.or the purpose of this review only the following will beonsidered:
. Neurotransmitters
The neurotransmitters identified in the auditory systemre summarized in Table 1.
These neurotransmitters are a potential target for hor-onal modulation of auditory function (see Hormones).
. Extra-auditory structures with inputs to the auditorysystem
The main extra-auditory connections are:
● The limbic system regulates instinctive behavior andemotions and connects with the auditory system viathe MGB. It is thought to be important in attachingemotional significance to acoustic stimuli (LeDoux etal., 1984; LeDoux, 1993). The limbic system alsoexpresses hormone receptors which include stress-related hormone and reproductive hormone receptors(Gray and Bingaman, 1996; Jennes and Langub,2000).
● The hypothalamus, is the integrator center for theendocrine and autonomic systems and is linked withthe auditory system through the IC (Adams, 1980),although, its effect on the auditory function is unclear.The hypothalamus contains the suprachiasmatic nu-cleus (SCN) which is thought to regulate the circadianrhythm (Halasz, 2000; Levine, 2000), and expressesa wide range of hormone receptors (reviewed byJennes and Langub, 2000).
● The reticular system is concerned with the behavioral
state of arousal and alertness and projects serotoner-
tc
1234
R
Ttla
T
L
I
O
C
S
LI
M
A
r system
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 883
gic fibers to most levels of the auditory system fromthe cochlea (Gil-Loyzaga et al., 2000) to the auditorycortex (Juckel et al., 1997). The ascending reticularsystem reacts more to “important” than to “unimpor-tant” stimuli, and this may be related to hearing innoise and selective attention (Chermak and Musiek,1997). The reticular formation is involved in the stressresponse and expresses adrenal steroid receptors(Jennes and Langub, 2000). The presence of noise orother stressful stimuli was found to modulate theserotonergic system, by increasing the release of5-HT (Singewald et al., 1998). 5-HT was also found tocontrol the gain the IC neural responses, either in apositive or negative direction depending on the typeof auditory stimuli, thus influencing auditory process-ing (reviewed by Hurley et al., 2002). Animal andhuman studies suggest that the serotonergic systemis sexually dimorphic, i.e. there is a gender difference.It has been reported that there is an increase in 5-HTactivity in the female rat brain compared with the male(Carlsson and Carlsson, 1988) and a decrease in thewhole brain 5-HT synthesis in women compared with
able 1. Neurotransmitters of the afferent and efferent auditory syste
evel Afferent auditory system
Neurotransmitter Possible function
HC Glutamate Physiologically excitatory (Puel, 1Le Prell et al., 2001)
and pathologically neurotoxic(acoustic trauma and/or ischeminjury) (Janssen et al., 1991;Eybalin, 1993)
HC ? ?
N Glutamate, aspartate Excitatory
Acetylcholine
GABA, glycine Inhibitory (Musiek and Hoffman,1990)
OC GABA, glycine Inhibitory
Glutamate, NMDA Excitatory (Musiek and Hoffman,1990)
L GABA ? Inhibitory (Moore and Moore, 1C Glycine, GABA Inhibitory
Glutamate Excitatory (Faingold et al., 1989;Musiek and Hoffman, 1990)
GB ? ?
C Acetylcholine, opioids Not clear (Musiek and Hoffman,1990)
AC, auditory cortex; IC, inferior colliculus; LOC, lateral olivocochlea
men (Nishizawa et al., 1997). It seems that estrogen e
contributes to dimorphism, as it may either enhanceor decrease 5-HT binding, depending on the site ofthe receptors in the brain (reviewed by Rubinow et al.,1998).
Hormones may modulate auditory function indirectlyhrough their action on the extra-auditory structures thatonnect with the auditory system.
HOW THE DIFFERENT HORMONES MAYINFLUENCE THE AUDITORY SYSTEM
. Reproductive steroids
. Stress-related hormones
. Hormones regulating fluid and electrolyte balance
. Melatonin
eproductive steroids
he reproductive (gonadal) hormones regulate reproduc-ive behavior, but also modulate the activity of hypotha-amic and extrahypothalamic noradrenergic, dopaminergicnd serotonergic neurons (Scholtz et al., 1998; Genazzani
Efferent auditory system
Neurotransmitter Possible function
Acetylcholine Excitatory (Felix and Ehrenberger, 1992)
GABA Inhibitory (Eybalin, 1993; Le Prell et al.,2001)
Dopamine, enkephalin
Dynorphin (via LOC) Inhibitory (Gil-Loyzaga, 1995; Pujol,1994; Le Prell et al., 2001)
Excitatory (Sahley and Nodar, 1994;Sahley et al., 1999)
Acetylcholine Mainly inhibitory (Eybalin, 1993; Dalloset al., 1997; Le Prell et al., 2001)
GABA (via MOC) Inhibitory (Eybalin, 1993; Le Prell et al.,2001)
GABA Inhibitory (Thompson and Schofield,2000)
Glutamate Excitatory (Thompson and Schofield,2000)
Glutamate Excitatory (Thompson and Schofield,2000)
? GABA Inhibitory (Campos et al., 2001)
? ?Glutamate Excitatory (Thompson and Schofield,
2000)GABA, glycine Inhibitory (Huffman and Henson, 1990)
Glutamate Excitatory (Thompson and Schofield,2000)
? Glutamate Excitatory (Thompson and Schofield,2000)
; NMDA, N-methyl-D-aspartate; ?, not known; N/A, not applicable.
m
995;
ic
987)
t al., 2000).

te2vm
l
acc21ertmgisc
mb
tE3rsattTaTlatwrsfheadbbg((rstgM
beaakn
nnia
stat(btaistsoa2(ahbl
S
iic(aahtpssh(flcesg
ao
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900884
Estrogen. Estrogens influence physiological func-ions of many organs and systems in both females (Nilssont al., 2001) and males (Sharpe, 1998; Lombardi et al.,001), including the skeletal, cardiovascular and the ner-ous systems, as well as the male urogenital tract, mam-ary glands and female reproductive organs.
Estrogen may influence the auditory system at differentevels, at the cochlear and more proximal levels:
At the cochlear level, estrogen receptors alpha (ER�)nd beta (ER�) have been identified in the inner ear (in-luding spiral ganglion type I cells, the stria vascularis andochlear blood vessels) in both humans (Stenberg et al.,001) and animal models (rats and mice) (Stenberg et al.,999) by immunohistochemistry, while previously, Nathant al. (1999) did not find the mRNA that encodes ERs in theat cochlea. These conflicting results may be the result ofhe small number of receptors and/or the low stability of theRNA. The presence of the receptors in the spiral gan-lion and outer and IHCs, suggests that estrogen may
nfluence auditory transmission, while the receptors in thetria vascularis may affect fluid electrolyte balance in theochlear fluids (Lee and Marcus, 2001).
Additionally, the ERs in the cochlear blood vesselsay influence auditory function by modulating cochlearlood flow (Laugel et al., 1987).
Stenberg and co-workers (2003) have reported thatreating oophorectomized rats with tamoxifen (a specificR modulator and generally acts as an ER antagonist) fordays did not affect the ER content of the cochlea. A
ecent study by Thompson and colleagues (2006) demon-trated that tamoxifen treatment for 3 months in youngdult female CBA mice have led to changes in their audi-ory function, although this does not necessarily mean thathese changes occurred through the cochlear receptors.he potential difference between the two studies could bettributed to the difference in the length of treatment.hompson et al. (2006) speculated that blocking ER had
ed to a decline in suppression of distortion product oto-coustic emissions (DPOAEs), as a result of a reduction ofhe MOC suppression. This decline could be similar tohat occurs with aging and precedes the onset of age-
elated hearing loss (Guimaraes et al., 2004) which mayuggest that estrogen has a protective role on auditoryunction, in conjunction with its neuroprotective effect. Itas been observed that the decrease in estrogen levels,.g. after menopause or in Turner’s syndrome, is associ-ted with an increased frequency of neurodegenerativeisorders. One mechanism of this protective effect coulde based on the “genomic” (steroid hormone actions cane delayed in onset and prolonged in duration and involveene expression and thus described as a genomic effectMcEwen and Alves, 1999)) action of estrogen on its alphaER�) and beta (ER�), possibly membrane receptors, as aegulator of the electrical activity of neurons, promotingynaptogenesis and the expression of nerve growth fac-ors. Another proposed mechanism would implicate estro-en as a free radical scavenging antioxidant (Behl and
anthey, 2000; Garcia-Segura et al., 2001). aIn the proximal auditory system, specific ERs have noteen reported. However, from what is known about theffect of estrogen on the CNS (Kuiper et al., 1998; McEwennd Alves, 1999), theoretically, estrogen may influence theuditory function at different levels of the CNS, through itsnown action as a modulator of the GABA-ergic, seroto-ergic, and glutamatergic systems (Woolley et al., 1997).
Estrogen may have an excitatory action on auditoryerve fibers, as it has been found to be mainly excitatory toeurons in other areas of the CNS (Smith et al., 2002), and
t could have a neuroprotective effect, as mentionedbove.
Progesterone. Progesterone is an ovarian hormoneecreted by the corpus luteum during the luteal phase ofhe ovarian cycle (see below) and it is also produced in thedrenal glands and the CNS. Progesterone is a precursoro other steroid hormones and acts as a neurosteroidBaulieu, 1998). Specific progesterone receptors have noteen identified in the in the auditory system, but proges-erone may cross-react with other steroid receptors (suchs glucocorticoid and mineralocorticoid receptors) present
n the cochlea or more proximal areas of the auditoryystem (Lang et al., 1990; Nathan et al., 1999). Proges-erone and its metabolites may also influence the auditoryystem through its interaction with the steroid binding sitesn GABA-A receptors acting as a GABA-A agonist, whichre present throughout the auditory system (Follesa et al.,001). Progesterone was found to decrease 5-HT levelsreviewed by Birzniece et al., 2006) and this may affectuditory processing indirectly. In general, progesteroneas a mainly inhibitory action on the CNS which mayalance the mainly excitatory action of estrogen (Katzenel-
enbogen, 2000).
tress-related hormones
Glucocorticoids. Cortisol (a steroid-based hormone)s the main glucocorticoid secreted from the adrenal cortexn response to stress, and acts through glucocorticoid re-eptors (GR) which are widely distributed in the CNSJennes and Langub, 2000) and other organs. GR havelso been identified in the inner ear of animals (Rarey etl., 1993; Zuo et al., 1995; Shimazaki et al., 2002) andumans (Rarey and Curtis, 1996), more in the cochlearhan the vestibular tissue. Within the cochlea, they areresent in the sensory (the organ of Corti’s hair cells andupporting cells) and non-sensory (spiral ligament andtria vascularis) tissues, suggesting both a possible role inomeostasis of inner ear fluids and signal transductionreviewed by Horner, 2003). Glucocorticoids may also in-uence the auditory function by interacting with their re-eptors found in the brainstem nuclei, including the mes-ncephalic raphe nuclei and locus ceruleus, which containerotonergic and noradrenergic neurons (Jennes and Lan-ub, 2000).
Catecholamines and endogenous opioids. Catechol-mines (adrenalin and nor-adrenalin) and endogenouspioids (such as �-endorphin, enkephalins, dynorphins)
re released in response to stress from the adrenal me-
dep(f(
pombdEt2nwta(o2ta
H
icha(titcrAthhaseae
itv2acsni
u
ssraewwVtgc(ecmad
M
Mt(asiodttc(pGarwstc1o(t
a((ia
12345
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 885
ulla and act mainly through the nervous system. Cat-cholamines are the main neurotransmitters of the sym-athetic nervous system, which is activated during stressBrook and Marshall, 2001). The opioids are also releasedrom the pituitary and the limbic system during stressSapolsky, 2002).
Endorphins and enkephalins act as analgesics androduce a euphoric state (Brook and Marshall, 2001). Opi-id binding of mu (�) and delta (�) opioid receptors com-only leads to neural inhibition (Crain and Shen, 1998),ut may also lead to neural excitation especially throughynorphin sensitive kappa (�) opioid receptors (Mains andipper, 1999), and these receptors have been identified in
he mammalian cochlea (Jongkamonwiwat et al., 2003,006). Enkephalins and dynorphins are thought to act aseurotransmitters in the auditory system (see Table 1),hile the nor-adrenalin fibers of the sympathetic innerva-
ion of the cochlea, that surround the labyrinthine arterynd the modiolar branches, control cochlear blood flowBrown, 2001). Sympathetic innervation is also seen inther auditory structures including the CN (Thompson,003) and the SOC (Mulders and Robertson, 2001) and,hus, has a potential to modulate higher levels of theuditory system.
ormones regulating fluid and electrolyte balance
Aldosterone. Aldosterone (a steroid-based hormone)s the main mineralocorticoid secreted from the adrenalortex. It is predominantly involved in regulating sodiumomeostasis, and indirectly volume homeostasis, but islso released during stress. Mineralocorticoid receptorsMR) are distributed in the CNS, but to a lesser degreehan GR (Jennes and Langub, 2000), and have beendentified within the cochlea (Rarey and Luttge, 1989) inhe marginal cells of the stria vascularis and spiral ganglionells (Furuta et al., 1994). Aldosterone increases sodiumeabsorption mainly by regulating the enzyme Na�, K�-TPase (Brook and Marshall, 2001), which has been iden-
ified in the inner ear epithelium. This suggests possibleormonal modulation of endolymph secretion and fluidomeostasis (Ferrary and Sterkers, 1998), although thection on spiral ganglion cells remains unknown. The ab-ence of circulating adrenal hormones does not lead tolectrophysiological changes in inner ear fluids (Ferrary etl., 1996) but to a decrease in endolymph volume (Lohuist al., 2000).
Aldosterone treatment in mice with autoimmune hear-ng loss has been reported to demonstrate a similar effecto prednisolone in improving hearing thresholds and re-ersing the pathology in the stria vascularis (Trune et al.,000). Tadros and colleagues (2005) found that aged mennd women with lower levels of aldosterone (but still in thelinically normal range) had worse hearing thresholds thanubjects with aldosterone levels in the upper middle of theormal range. Thus, aldosterone may play a protective role
n the cochlea.
Vasopressin. Vasopressin, also known as antidi-
retic hormone (ADH), is involved in fluid homeostasis in a 6imilar way to aldosterone and is also released duringtress (Halasz, 2000). Vasopressin may play a role inegulating the cochlear fluids by regulating the enzymedenylate cyclase which has been identified in the innerar epithelium (Ferrary and Sterkers, 1998). Vasopressinas found to stimulate potassium secretion and increaseater permeability in the cochlea (Ferrary et al., 1996).asopressin receptors V1a and V2 have been identified
hroughout the developing rat cochlea, but only in the spiralanglion and spiral ligament of the adult cochlea. It isurrently thought to be important in cochlear developmentFuruta et al., 1998). The vasopressin receptor V1a isxpressed widely in the CNS, in the frontal and piriformortex, olfactory system, hippocampus, and throughout theidbrain, pons and medulla (Jennes and Langub, 2000)nd may therefore, directly or indirectly modulate the au-itory system.
elatonin
elatonin is a neuroactive substance derived from 5-HT byhe melatonin forming enzyme 5-HT N-acetyltransferaseNAT) and hydroxyindole-O-methyltransferase (HIOMT)nd secreted from the pineal gland. Melatonin is alsoynthesized in the guinea-pig cochlea by melatonin-form-
ng enzymes, NAT and HIOMT, and is detectable in thergan of Corti, the basilar membrane and to a lesseregree in the cochlear nerve and stria vascularis, includinghe spiral ligament (Biesalski et al., 1988). The concentra-ion of melatonin in the cochlea is affected by light, andorrelates with the peripheral concentration of melatoninLopez-Gonzalez et al., 1997). Melatonin was found torolong the postmortem activity of OHC in rats (Lopez-onzalez et al., 1999). Otoacoustic emissions (OAEs), asn expression of the OHC activity, can be recorded fromats for a few minutes postmortem, i.e. after clinical death,hen the cells of different tissues, including OHCs, stillhow activity. The treatment with melatonin before killinghe animal seems to prolong this activity up to seven timesompared with control animals (Lopez-Gonzalez et al.,999). It was also observed that melatonin ameliorates thetotoxicity of aminoglycosides and cisplatinum in ratsLopez-Gonzalez et al., 2000a,b). These findings suggesthat melatonin may have a protective role on the cochlea.
Melatonin has multiple effects on the CNS including annticonvulsant and anxiolytic action by enhancing GABAGolombek et al., 1996) and benzodiazepine functionGuardiola-Lemaitre et al., 1992). Therefore, theoretically,t may also have an effect on GABA-ergic fibers of theuditory system.
PHYSIOLOGICAL VARIATION IN HORMONESAND THE EFFECTS ON AUDITORY FUNCTION
. Circadian cycle
. Gender differences
. The ovarian cycle
. Menopause
. Pregnancy
. Stress

nftv
tritps
C
Mpttpdwteetmisccac
fgo
dawaicHat
bpw1
iamt
G
Rd(steuabfeetadrhc“pp
FcsGttiha
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900886
The levels of hormones vary in response to endoge-ous and exogenous stimuli and many vary in a cyclic
ashion. The hypothalamus acts as the neural control cen-er of the endocrine system and regulates the physiologicalariation in hormones (Halasz, 2000).
The endocrine changes related to reproductive func-ion (ovarian cycle, pregnancy, and menopause), dailyhythm (circadian) and exposure to stressful stimuli couldn turn affect auditory function. Additionally, there are mul-iple interactions between the hormones involved in thesehysiological changes (Fig. 2) and this enhances the pos-ible multidirectional effects on the auditory system.
ircadian cycle and auditory function
elatonin, cortisol and vasopressin show a clear circadianattern. The circadian pacemaker in mammals is found inhe paired SCN of the hypothalamus, which also controlshe rhythm of melatonin synthesis through a multi synapticathway (Urbanski, 2000). Cortisol levels are highest atawn and low at dusk (Despopoulos and Silbernagl, 1991),hile vasopressin levels are higher during the night (Kos-
oglou-Athanassiou et al., 1998a). The SCN rhythm isntrained by light via the retino-hypothalamic tract (Blockt al., 2000). In subterranean animals, such as the mole,
he IC (with auditory information) projects to the SCN, anday have a similar function to the retino-hypothalamic tract
n other mammals (Kudo et al., 1997). In humans, a recenttudy has found that auditory stimuli led to a phase shift inircadian rhythms (Goel, 2005). This finding implies that aonnection between the auditory pathways and the SCNlso exists in humans, which may enable a direct effect ofircadian cycle on auditory function and vice versa.
Diurnal changes in OAE, which reflect cochlear OHCunction, have been reported (Wit, 1985; Bell, 1992; Hag-erty et al., 1993). Wit (1985) measured spontaneous
ig. 2. Hypothalamic pituitary adrenal and gonadal axis and the cir-adian cycle. Light effect on the SCN leads to inhibition of melatoninynthesis from the pineal gland. * The inhibitory effect of melatonin onnRH is only seen before puberty and in certain pathological condi-
ions in humans (Silman, 1991; Kadva et al., 1998). Solid lines: Exci-atory effect, Dashed lines: inhibitory effect. CRF: corticotropin releas-ng factor, E2: estrogen, Prog: progesterone, ACTH: adrenocorticotropicormone, Endo: �-endorphin, Dyn: dynorphin, ACh: acetylcholine, Ad:drenalin and, NA: nor adrenalin. (Italics indicate inhibitory action.)
toacoustic emissions (SOAEs) in two volunteers on 10
ifferent days, twice and three times daily for the femalend male volunteer respectively. The SOAE frequencyas shown to shift significantly to a lower frequency in thefternoon compared with the morning sessions. This find-
ng was confirmed by Bell (1992) who calculated a 1%hange in SOAE frequency, with no amplitude fluctuation.aggerty et al. (1993) also noted a significant 24-h vari-bility in SOAEs frequency but reported that there was lesshan 1% change.
These changes suggest that the SOAE generator maye synchronized with the biological circadian rhythm. Theeripheral auditory structures may be less inhibited alongith other subcortical structures during sleep (Lancel,993), resulting in an increase in SOAE levels.
The auditory evoked potentials arising from more prox-mal, cortical structures, such as the P300 seem to beffected by the time of day (Polich and Kok, 1995). Thisay be attributed to circadian changes in cognitive func-
ion that contribute to the P300 (Wesensten et al., 1990).
ender differences in auditory function
eproductive hormones have been implicated in genderifferences in sensorimotor (Becker, 2002) and cognitiveHampson, 2002) functions in animals and humans. Expo-ure to reproductive hormones during development leadso sexual dimorphism in CNS structures (de Courten-My-rs, 1999; Rhodes and Rubin, 1999). This may be thenderlying cause for gender differences and may alsoffect the auditory system. Indeed, sexual dimorphism haseen noted in auditory structures, such as the cochlea inemales which may have a larger number of OHC (Wrightt al., 1987) and is shorter than the cochlea in males (Satot al., 1991). The sexually dimorphic structure of the sero-onergic system (Rubinow et al., 1998) as mentionedbove, may also modulate neural transmission in the au-itory brain stem and cortical structures. The differences ineproductive hormones during development and in adult-ood may explain gender effects in auditory function, spe-ifically the theoretical possibility that auditory function isbetter” in females than males due to the excitatory androtective effects of estrogen. This notion could be sup-orted by the following:
� Several studies have reported that adult femaleshave more sensitive hearing in higher frequencies(measured by pure tone audiometry) compared withmales (Jerger and Hall, 1980; Davis, 1995; McFadden,1993, 1998) which is also noted in carefully screenedpopulations for noise exposure (Johansson and Ar-linger, 2002). It was also observed in school-agedchildren, that girls tend to have lower audiometricthreshold compared with boys, however the differ-ence was not statistically significant (Roberts andHuber, 1970; Haapaniemi, 1996).
� OAE are thought to originate from the OHCs reflect-ing cochlear function (Kemp et al., 1990), and areassociated with good hearing sensitivity (Probst et
al., 1991). Females tend to have OAEs with larger
fc
T
Tddrssa
ibpdr(flbapm
fpheiawo
aa
ltaTc�sslpwm(ffiie
e(p�o2
motcCtdnvmomdeom
hifseflb
tli
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 887
amplitudes compared with men (McFadden, 1993;Hall, 2000) and are more likely to have recordableSOAEs (75% of females compared with 58% ofmales (Penner and Zhang, 1997). The gender differ-ence is also seen in neonates (Kei et al., 1997) andolder children (Lamprecht-Dinnesen et al., 1998;O’Rourke et al., 2002) which may be due to prenatalhormonal exposure.
� Oral contraceptives (OC) suppress the endogenouslevels of estrogen, and females using OC displayOAEs which are closer to males’ results than fe-males not using OC (McFadden, 2000).
� The excitatory effect of estrogen and sexual dimor-phism in the CNS and auditory system may affect theneural transmission in the auditory brainstem leadingto gender differences in auditory brain stem–evokedresponses (ABR). Female adults were found to haveshorter ABR wave latencies and larger wave V am-plitude compared with males (Jerger and Hall, 1980;Jerger and Johnson, 1988; Trune et al., 1988; Dehanand Jerger, 1990). The same findings were alsonoted in neonates (Chiarenza et al., 1988; Stuart andYang, 2001), but to a lesser extent in older children(Trune et al., 1988; Hall, 1992).
The effect of reproductive hormones on the auditoryunction may be more clearly noted in the endocrinehanges that occur during the menstrual cycle.
he ovarian cycle and auditory function
he secretion of estrogen and progesterone fluctuatesuring the ovarian cycle, with the estrogen level risinguring the first, proliferative (follicular) phase of the cycleeaching a peak just before the luteinizing hormone (LH)urge, while progesterone secretion starts to rise in theecond, luteal phase of the cycle, following the LH surgend ovulation (Djahanbakhch et al., 1981) (Fig. 3).
The higher level of estrogen during the follicular phases also associated with a rise in other hormone levels. Theasal ACTH plasma level seems to rise in the late follicularhase (Genazzani et al., 1975; Mauri et al., 1990) possiblyue to the enhancing effect of estrogen on corticotropineleasing factor gene transcription in the hypothalamusKirschbaum et al., 1999). The rise of ACTH during the lateollicular phase is not associated with higher free cortisolevel, due to estrogen induced changes in corticosteriod-inding protein levels (Kirschbaum et al., 1999; Altemus etl., 2001). The lower level of free cortisol may affect thehysiological response to stress during this phase of theenstrual cycle.
Vasopressin levels also tend to be higher during theollicular phase of the cycle compared with the mid-lutealhase (Kostoglou-Athanassiou et al., 1998b). The en-ancement of vasopressin secretion is possibly due tostrogen, which was found to increase vasopressin levels
n females who had undergone oophorectomy (Forsling etl., 1996). This may lead to fluid retention or redistributionhich occurs in some women in the pre-menstrual period
f the menstrual cycle (Tollan et al., 1993) and may also tffect the fluid balance in the cochlea and thus affectuditory function.
The level of �-endorphin peaks 2–4 days before ovu-ation followed by a dip in levels postovulation and thenhere is a gradual rise again during the late luteal phase,bout 24 h before the next menses (Vrbicky et al., 1982;ang et al., 1987; Ferrer et al., 1997). Women with poly-ystic ovarian disease and amenorrhea have levels of-endorphin lower than in normal women (Martinez-Guisa-ola et al., 1999). There is also evidence that estrogentimulates opioid receptor expression and stabilizes the
evels of �-endorphins that tend to decreases after meno-ause (surgical or spontaneous). This may be associatedith mood changes that can be helped by estrogen treat-ent, which increases �-endorphin levels in plasma
Genazzani et al., 2000). Estrogen also affects mood byacilitating the function of enkephalin that is also importantor reproductive behavior (Pfaff et al., 2000). It is not clearf the changes seen in �-endorphin are the result of anntrinsic role in the regulation of the ovarian cycle or are theffect of estrogen.
Progesterone alone had no effect on �-endorphin lev-ls in the hypothalamus and pituitary of oophorectomizedOVX) rats. However, treatment with both estrogen androgesterone reverses the effect of estrogen alone on-endorphin levels in the pituitary, and increases the levelsf �-endorphin in the hypothalamus (Genazzani et al.,000).
The fluctuation of hormones during the ovarian cycleay potentially lead to fluctuation in auditory function andther sensory processes (Parlee, 1983). The auditory sys-em may be more sensitive during the peak of estrogenirculation due to its excitatory and protective effect in theNS. Correspondingly, the low levels of hormones during
he pre-menstrual phase may relate to less sensitive au-itory function. A recent study (Sisneros and Bass, 2003)oted that the auditory nerve fibers of a midshipman fish (aocal seasonally breeding fish) are more responsive toale mating calls during the breeding season and not atther times. Treatment with estradiol of female midship-an fish during the non-breeding season makes their au-itory nerves respond more to male mating calls (Sisnerost al., 2004). No studies have reported these changes inther vertebrates, but these findings may imply auditoryodulation in the presence of reproductive steroids.
Auditory function in women during the ovarian cycleas been previously investigated, with inconsistent find-
ngs (Table 2). Fluctuation in auditory function during dif-erent stages of the cycle has been demonstrated, but thetudies lacked precise timing of the cycle, together withxact correlation between hormonal levels and auditoryunction. Only a few studies monitored ovarian steroidevels during the cycle, and these examined only auditoryrainstem-evoked responses.
No studies have reported auditory symptoms duringhe ovarian cycle. However, a few case reports in theiterature have described women who had fluctuating hear-ng loss associated with the ovarian cycle, that occurred in
he late luteal phase and improved after the onset of men-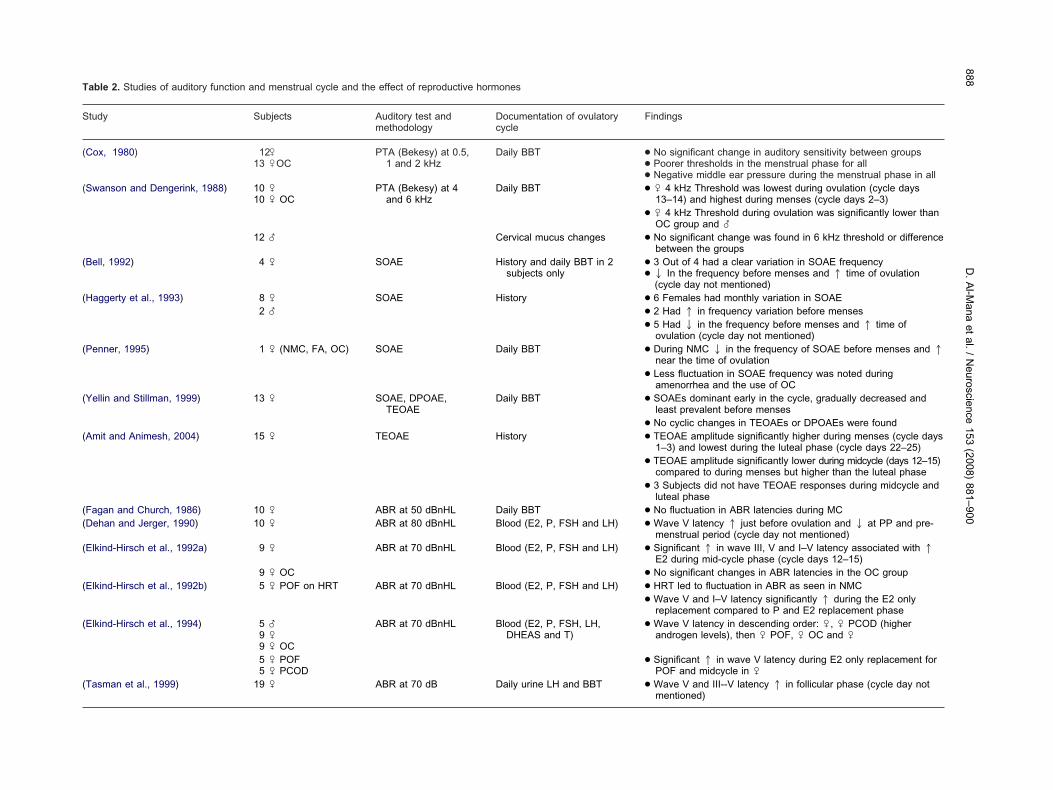
Table 2. Studies of auditory function and menstrual cycle and the effect of reproductive hormones
Study Subjects Auditory test andmethodology
Documentation of ovulatorycycle
Findings
(Cox, 1980) 12 �13 �OC
PTA (Bekesy) at 0.5,1 and 2 kHz
Daily BBT ● No significant change in auditory sensitivity between groups● Poorer thresholds in the menstrual phase for all● Negative middle ear pressure during the menstrual phase in all
(Swanson and Dengerink, 1988) 10 �10 � OC
PTA (Bekesy) at 4and 6 kHz
Daily BBT ● � 4 kHz Threshold was lowest during ovulation (cycle days13–14) and highest during menses (cycle days 2–3)
● � 4 kHz Threshold during ovulation was significantly lower thanOC group and �
12 � Cervical mucus changes ● No significant change was found in 6 kHz threshold or differencebetween the groups
(Bell, 1992) 4 � SOAE History and daily BBT in 2subjects only
● 3 Out of 4 had a clear variation in SOAE frequency● 2 In the frequency before menses and 1 time of ovulation
(cycle day not mentioned)(Haggerty et al., 1993) 8 � SOAE History ● 6 Females had monthly variation in SOAE
2 � ● 2 Had 1 in frequency variation before menses● 5 Had 2 in the frequency before menses and 1 time of
ovulation (cycle day not mentioned)(Penner, 1995) 1 � (NMC, FA, OC) SOAE Daily BBT ● During NMC 2 in the frequency of SOAE before menses and 1
near the time of ovulation● Less fluctuation in SOAE frequency was noted during
amenorrhea and the use of OC(Yellin and Stillman, 1999) 13 � SOAE, DPOAE,
TEOAEDaily BBT ● SOAEs dominant early in the cycle, gradually decreased and
least prevalent before menses● No cyclic changes in TEOAEs or DPOAEs were found
(Amit and Animesh, 2004) 15 � TEOAE History ● TEOAE amplitude significantly higher during menses (cycle days1–3) and lowest during the luteal phase (cycle days 22–25)
● TEOAE amplitude significantly lower during midcycle (days 12–15)compared to during menses but higher than the luteal phase
● 3 Subjects did not have TEOAE responses during midcycle andluteal phase
(Fagan and Church, 1986) 10 � ABR at 50 dBnHL Daily BBT ● No fluctuation in ABR latencies during MC(Dehan and Jerger, 1990) 10 � ABR at 80 dBnHL Blood (E2, P, FSH and LH) ● Wave V latency 1 just before ovulation and 2 at PP and pre-
menstrual period (cycle day not mentioned)(Elkind-Hirsch et al., 1992a) 9 � ABR at 70 dBnHL Blood (E2, P, FSH and LH) ● Significant 1 in wave III, V and I–V latency associated with 1
E2 during mid-cycle phase (cycle days 12–15)9 � OC ● No significant changes in ABR latencies in the OC group
(Elkind-Hirsch et al., 1992b) 5 � POF on HRT ABR at 70 dBnHL Blood (E2, P, FSH and LH) ● HRT led to fluctuation in ABR as seen in NMC● Wave V and I–V latency significantly 1 during the E2 only
replacement compared to P and E2 replacement phase(Elkind-Hirsch et al., 1994) 5 �
9 �9 � OC
ABR at 70 dBnHL Blood (E2, P, FSH, LH,DHEAS and T)
● Wave V latency in descending order: �, � PCOD (higherandrogen levels), then � POF, � OC and �
5 � POF5 � PCOD
● Significant 1 in wave V latency during E2 only replacement forPOF and midcycle in �
(Tasman et al., 1999) 19 � ABR at 70 dB Daily urine LH and BBT ● Wave V and III--V latency 1 in follicular phase (cycle day notmentioned)
D.
Al-M
anaet
al./
Neuroscience
153(2008)
881–900888

sMcpai(ntrhodhaTcipttlfiafl
tt
t(ppsflafmtdts
scgowetfflmlnwle
2.co
ntin
ued
dyS
ubje
cts
Aud
itory
test
and
met
hodo
logy
Doc
umen
tatio
nof
ovul
ator
ycy
cle
Fin
ding
s
send
eet
al.,
2000
)15
�A
BR
at85
dBH
isto
ryof
chan
ges
inva
gina
lsec
retio
n●
No
sign
ifica
ntdi
ffere
nce
inA
BR
late
ncie
sin
the
thre
ete
stin
gse
ssio
nsda
vet
al.,
2002
)20
�A
BR
at70
dBnH
LD
aily
BB
T●
Atr
end
of1
inA
BR
wav
ela
tenc
ies
and
inte
rpea
kin
terv
als
durin
gth
em
id-c
ycle
phas
e,bu
tno
tsi
gnifi
cant
.rr
aet
al.,
2003
)94
�A
BR
at10
0dB
spl
Ultr
ason
ogra
phy
and
seru
mP
leve
l●
AB
Rw
ave
late
ncie
san
din
terp
eak
inte
rval
ssi
gnifi
cant
ly2
inth
epe
riovu
lato
ryph
ase
(cyc
leda
yno
tm
entio
ned)
ruso
etal
.,20
03b)
94�
Bef
ore
and
afte
rO
CA
BR
at10
0dB
spl
Ultr
ason
ogra
phy
and
seru
mP
leve
l●
AB
Rw
ave
late
ncie
san
din
terp
eak
inte
rval
ssi
gnifi
cant
ly2
inth
epe
riovu
lato
ryph
ase
(cyc
leda
ys13
–16)
●N
osi
gnifi
cant
diffe
renc
ein
AB
Rw
ave
late
ncie
san
din
terp
eak
inte
rval
sdu
ring
OC
MC
,no
rmal
men
stru
alcy
cle;
FA
,fu
nctio
nal
amen
orrh
ea;
BB
T,
basa
lbo
dyte
mpe
ratu
re;
PO
F,
prem
atur
eov
aria
nfa
ilure
;P
CO
D,
poly
cyst
icov
aria
ndi
seas
e;E
2,oe
stro
gen;
P,
prog
este
rone
;E
AS
,de
hydr
oepi
andr
oter
one
sulfa
te;
T,
test
oste
rone
;P
P,
prog
este
rone
phas
e1
,in
crea
se;2
,de
crea
se;
�,
mal
es;
�,
fem
ales
with
norm
alm
enst
rual
cycl
e.
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 889
es (Miller and Gould, 1967; Andreyko and Jaffe, 1989).iller and Gould (1967) described two women with this
ondition, one of whom had worse hearing when givenrogesterone. A similar case was reported by Andreykond Jaffe, (1989). Their patient’s fluctuating hearing loss
mproved during pregnancy, while she was breast-feedingshe had amenorrhea) and with the use of nafarelin (go-adotrophin releasing hormone (GnRH) agonist that leadso ablation of sex steroids by downregulation of GnRHeceptors and decreases secretion of follicular stimulatingormone (FSH) and LH leading to anovulation and amen-rrhea). On the other hand, Souaid and Rappaport (2000)escribed a case of a 45-year-old woman who had bilateralearing loss with the onset of menses with right ear block-ge and tinnitus that improved later on during the cycle.hey also reported that she had an abnormal ABR re-orded in the middle of her ovarian cycle (prolonged III–V
nter-peak interval on the left and delayed wave V withrolonged I–V inter-peak interval on the right). She wasreated with diuretics, which improved her symptoms, andhe ABR results of the right ear but the ABR results of theeft ear were the same as before, and her hormonal pro-le was within normal limits. These cases suggest that theuditory system in some women is more sensitive to theuctuation of hormones during the ovarian cycle.
The following alterations in the auditory system duringhe ovarian cycle have been reported (as summarized inhe Table 2):
� Auditory thresholds. The less sensitive auditoryhresholds during the menstrual phase observed by Cox1980) and Swanson and Dengerink (1988) are unex-lained. However, the associated negative middle earressure observed during the menstrual phase in someubjects could contribute to this. An increase in interstitialuids (possibly due to progesterone; Pechere-Bertschi etl., 2002; Tollan et al., 1993), affects the eustachian tubeunction and, thus may lead to poorer thresholds during theenstrual phase of the cycle. Another explanation may be
hat a fluctuation in hormones affects higher areas of au-itory processing and thus leads to changes in auditoryhresholds, similar to the findings documented in otherensorimotor and cognitive functions (Hampson, 2002).
� SOAEs. The fluctuation in SOAEs with the men-trual cycle implies that there may be a link betweenochlear function and the pituitary–gonadal axis. Thereater frequency variation of SOAEs near the time ofvulation (Bell, 1992; Haggerty et al., 1993; Penner, 1995),ith a shift toward a higher frequency, may suggest anxcitatory effect of estrogen on the SOAE generator andhe frequency shift toward a lower frequency and lessrequency variation in SOAEs near menstruation may re-ect low levels of estrogen and progesterone. Estrogenay be implicated in this frequency change, as the level is
owest in the late luteal phase (before menses) and peaksear ovulation. Moreover the peak has been associatedith an increase in neuronal excitability and inhibition of
GABA function (Wagner et al., 2001).Tab
Stu
(Re
(Ya
(Se
(Ca N
DH

daoctcacss
asatai1siteHimpmmbilwfb
of
M
Cmo
m
es
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900890
� Transient OAEs. Only twostudiesevaluatedTEOAEsuring the ovarian cycle (Yellin and Stillman, 1999; Amitnd Animesh, 2004). Yellin and Stillman (1999) did notbserve any change in TEOAE responses during the cy-le. On the other hand, Amit and Animesh (2004) reportedhat the TEOAE responses tend to decrease during theycle and that three of their subjects did not have record-ble TEOAEs in two testing sessions (see Table 2). Thisould have been confounded their results because theirubjects could have had a cochlear dysfunction. Moretudies are needed to clarify TEOAE results.
� ABR. The studies that measured hormone levelsnd ABR produced conflicting findings (see Table 2). Thehorter ABR latencies reported by Caruso et al. (2003a)nd Serra et al. (2003) during the periovulatory phase ofhe cycle may suggest that the high estrogen level isssociated with shorter ABR latencies, as had been found
n estrogen treatment of young OVX rats (Coleman et al.,994). The higher level of estrogen may alter the speed ofensory neurotransmission in the brain stem by modulat-ng glutamate transmission (Behl and Manthey, 2000). Onhe other hand, the possible effect on neurosteroids mayxplain the results of Dehan and Jerger (1990) and Elkind-irsch et al. (1992a,b, 1994) of longer ABR latencies dur-
ng the periovulatory phase. The higher levels of estrogenay result in an increase in neurosteroids (such as allo-regnalone) which facilitate GABA inhibition in the auditoryidbrain (Disney and Calford, 2001). The estrogen treat-ent of OVX rats increases the level of allopregnaloneoth centrally and peripherally in a dose dependent fash-
on (Stomati et al., 2002). The difference between the ABRatencies in the periovulatory phase and luteal phaseould be compatible with progesterone blunting the ef-
ect of estrogen, by inhibiting its action in the auditoryrainstem.
Further larger studies on the effect of the ovarian cyclen auditory functions are needed to clarify the results
ound in the previous studies.
enopause and auditory function
hanges in auditory function have been noted in post-enopausal women attributed in part to the lower levels ofvarian hormones.
The following findings raise the suspicion that theenopause leads to a decline of the auditory function:
� The onset of age-related hearing loss is later inwomen compared with men (Pearson et al., 1995)and seem to coincide with the menopause (Murphyand Gates, 1997). The sex difference in age relatedhearing loss is also seen in mice models of agerelated hearing loss (CBA mice) (Guimaraes et al.,2004), with a decline in DPOAE amplitudes occur-ring earlier in the males. In female CBA mice thegreatest decline in DPOAE amplitudes occurred inolder age mice following the mouse menopause.This finding suggests that estrogen may have a pro-
tective role on the cochlea and OHC function.� A few studies explored the possible association ofhearing loss assessed by pure tone audiometry inpost-menopausal women. Kim et al. (2002) sug-gested that a lower level of serum estradiol impedeshearing sensitivity in post-menopausal women. Kilic-dag and co-workers (2004) reported that post-meno-pausal women using estrogen therapy had betterpure tone thresholds compared with those who didnot use it. A recent study by Hederstierna et al.(2007) evaluated the hearing thresholds in a group ofwomen around the time of menopause and foundthat 40% had some degree of hearing loss. Theyreported that a subgroup of women who were notusing hormone replacement therapy (HRT) had atendency of poorer hearing threshold levels com-pared with pre- and peri-menopausal and post-menopausal women who were using HRT.
These studies suggests that HRT and in particularstrogen therapy may have a beneficial effect on hearingensitivity.
� In a recent study, Guimaraes and colleagues (2006)reported that post-menopausal women who weretaking the combined estrogen and progesteroneHRT had poorer pure tone auditory thresholds, lowerlevels of high frequency DPOAE, and poorer perfor-mance in the hearing-in-noise test compared withpost-menopausal women who were either on estro-gen only replacement therapy or not taking any HRT.These results suggests that progestin (a syntheticprogesterone in combined HRT) may have a nega-tive effect on both peripheral (DPOAE) and central(hearing-in-noise test) auditory function and supportthe positive role of estrogen on hearing function.
� The low levels of estrogen and progesterone mayalter nerve conduction times as seen in carpal tunnelsyndrome (Pascual et al., 1991) and, thus, may alsoaffect ABR latencies. ABR latencies tend to increasewith age, and post-menopausal women have pro-longed ABR wave latencies and interpeak latenciescompared with younger females of a significantlygreater degree than that seen in men of matchedage groups (Jerger and Hall, 1980; Wharton andChurch, 1990). These findings in women cannot besolely age-related changes and hormones maytherefore be of importance. Indeed, HRT, in post-menopausal women, brings ABR latencies closer tothe values seen in pre-menopausal women (Carusoet al., 2000). ABR latencies were also found to besignificantly shorter in postmenopausal women afterthe use of different forms of HRT; tibolone (a syn-thetic steroid with combined estrogenic, progesto-genic and androgenic action) (Sator et al., 1999),combined estrogen and progesterone (Khaliq et al.,2003), trans-dermal estrogen therapy (Carusoet al., 2003a), and oral estrogen therapy (Khaliq et al.,2005). Moreover, the effect of HRT in post-meno-pausal women on ABR latencies is similar to what is
seen in females with premature ovarian failure (POF)
P
Dhirph
aaath(rfppaPartnsciiu
m
smms2lcptnwowdts
S
Terptacft2arwwslm
b(t
Fmot(hc
Fs
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 891
after treatment (Elkind-Hirsch et al., 1992b) and es-trogen treatment in animal studies (Coleman et al.,1994; Cooper et al., 1999). These findings suggestthat ovarian hormones may have an effect on syn-aptic transmission in the auditory brainstem.
regnancy and auditory function
uring pregnancy the levels of both ovarian hormones areigher than usual, and there are other complex physiolog-
cal changes (Hadley, 2000). These changes lead to fluidetention and a hyperdynamic circulation, which may im-act upon the circulation in the cochlea and cochlear fluidomeostasis.
Auditory symptoms such as aural fullness, changes inuditory sensitivity or tinnitus (Gurr et al., 1993; Tsunoda etl., 1999) have been reported in pregnancy. The results ofpostal questionnaire by Gurr et al. (1993) showed that
he prevalence of tinnitus in pregnant females (25%) wasigher than non-pregnant women (11%). Tsunoda et al.1999) noted that 25% of pregnant women in their surveyeported otologic symptoms, including tinnitus and auralullness that resolved after giving birth but no changes inure tone or impedance audiometry were noted comparingregnant and non-pregnant women. However, Sennaroglund Belgin (2001) found that pregnant women had higherTA thresholds at 125, 250 and 500 Hz, than postpartumnd non-pregnant women, but were still within the normalange (less than 20 dB HL), and pregnant women in thehird trimester had significantly lower uncomfortable loud-ess level compared with postpartum and non-pregnantubjects. These findings suggest that the physiologicalhanges in pregnancy may affect auditory function mim-cking the auditory dysfunction seen in Menière’s disease,.e. low frequency elevated thresholds, with lowering ofncomfortable loudness levels.
The higher levels of ovarian hormones in pregnancy
ig. 3. Schematic representation of the changes in reproductive hor-ones during the average menstrual cycle, and the associated rise ofther hormones in the periovulatory phase. The graph does not reflecthe pulsatile release of estrogen and LH in the periovulatory phaseDjahanbakhch et al., 1984) but the smoothed out overall changes inormones levels. E2: estrogen, P: progesterone, ACTH: adrenocorti-otropic hormone ADH: ADH (vasopressin), Endo: �-endorphin.
ay shorten ABR wave latencies as seen in previousmH
tudies (see above), but the higher level of progesteroneay blunt this effect. Neural conduction in the brain stemay be slower during pregnancy due to the higher levels of
teroids that facilitate GABA inhibition (Disney and Calford,001). Tandon et al. (1990) found that the absolute wave
atencies were slightly shorter but did not reach signifi-ance in eight pregnant women in the third trimester com-ared with age-matched non-pregnant women. However,he I–III, III–V, and I–V interpeak latencies which indicateeural conduction were significantly longer in pregnantomen. An ABR study of 38 women, 20 in different stagesf pregnancy (including postpartum) and 18 non-pregnantomen by Sennaroglu and Belgin (2001) did not find anyifference. This inconsistency in findings may be attributedo difference in methodology and sample sizes and varyingtates of pregnancy.
tress and auditory function
he presence of emotional or physical stress initiates thendocrine stress response by the release of corticotrophineleasing factor (CRF) which activates the hypothalamicituitary adrenal axis and this leads to the release of cor-isol and adrenalin from the adrenal gland (Fig. 4) and thectivation of the sympathetic nervous system. Acute orhronic stress leads to the inhibition of the reproductiveunction, with the suppression of LH, which is modulatedhrough GRs (McGivern and Redei, 1994; Pfaff et al.,000). GnRH release is suppressed by CRF (Saketos etl., 1993) and dynorphin and �-endorphin through � and �eceptors in the hypothalamus (Petraglia et al., 1986),hich reduces plasma levels of LH, and this may occur inomen with functional hypothalamic amenorrhea. Theuppression of the hypothalamic–pituitary–gonadal axis,
eading to the lower levels of estrogen and progesterone,ay also affect auditory function.
The endocrine response to stress can also be initiatedy strong auditory stimuli such as excessive noiseMichaud et al., 2003; Spreng, 2000). The pathway may behrough the MGB interface between the auditory system
ig. 4. Possible interaction between stress, endocrine and auditoryystems, which may lead to pathology. Rep. H�reproductive hor-
ones, Stress. H�cortisol, catecholamines and opioids, Fluid&E.�aldosterone and vasopressin.
aam(ifdat
ptSstntsatd9AtSsd
o(aaccweb
aWsltastwAbrb
Hl
H
MA
H
Taaonbroaf
aapdota
cKGsnanfc1rpbaaba2wsZ
osemlbadsm
op
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900892
nd the stress-responsive limbic system. This non-primaryrea of the auditory cortex was found to have more CRFRNA than other areas of the rat’s central auditory system
Imaki et al., 1991). On the other hand, the hormonesnvolved in the response to stress may affect auditoryunction directly by an excitatory effect, which may lead toamage through glutamate induced neurotoxicity (Pujol etl., 1993). The possible interactions between stress and
he auditory system are demonstrated in Fig. 4.Emotional stress has been associated with auditory
henomena such as hyperacusis and an increase in tinni-us and dizziness (Holgers et al., 2000; Katzenell andegal, 2001; Sahley and Nodar, 2001). These phenomenauggest that stress may lead to stimulation and sensitiza-ion of the auditory system, resulting in heightened aware-ess and, thus, more vulnerability to acoustic trauma dueo excitotoxicity (Sahley and Nodar, 2001). The effect ofympathectomy in awake guinea pigs would support thebove hypothesis, as it was found to reduce temporaryhreshold shift by 8.5 dB (Hildesheimer et al., 1991) to 40B (Horner et al., 2001) and permanent threshold shift by.8 dB (Hildesheimer et al., 2002) after sound exposure.n alternative hypothesis is that activation of the sympa-
hetic system causing vasoconstriction (Despopoulos andilbernagl, 1991) may alter the blood flow in the auditoryystem and affect cochlear blood flow leading to ischemicamage following noise exposure.
In animal experiments, supra-physiological amountsf adrenalin were infused to mimic the stress responseJuhn et al., 1991) and this led to an elevation of serumnd perilymph osmolarity and an increase in compoundction potential threshold especially at higher frequen-ies. Chronic administration of adrenalin was also found toause a transient 20–40 dB threshold shift that increasedith time across all frequencies (Juhn et al., 1999). Theseffects are thought to be the result of adrenalin on cochlearlood vessels and inner ear fluid homeostasis.
The effects of stress and glucocorticoid treatment onuditory function have been studied in humans. Fehm-olfsdorf and co-workers (1993) noted that a subset of
ubjects, who presented with raised salivary free cortisolevels after being confronted by stress, had higher con-ralateral acoustic reflex thresholds (ART), while the sameuthors (Fehm-Wolfsdorf and Nagel, 1996) found a non-ignificant trend to higher contralateral ART with glucocor-icoid treatment in a double blind crossover study. Thereas no effect of glucocorticoid treatment on ipsilateralRT, pure tone thresholds and ABR. These findings maye explained on the basis of a multisynaptic contralateraleflex pathway in which both GR and 5-HT are present,oth or either of these may alter function.
AUDITORY PATHOLOGY AND HORMONES
ormones may play a role in the development of patho-ogical conditions of the auditory system, including:
yperacusis and tinnitus s
enière’s diseaseuditory dysfunction in pre-menstrual syndrome
yperacusis and tinnitus
innitus can be defined as an auditory perception in thebsence of any external auditory stimulation, while hyper-cusis has been described as a reduced tolerance tordinary environmental sounds, often associated with tin-itus (Anari et al., 1999; Andersson et al., 2001). It haseen considered that both tinnitus and hyperacusis areesults of a raised rate and/or altered pattern of spontane-us neural activity (Eggermont, 2003). Hence, the mech-nisms which alter spontaneous auditory activity may alsoacilitate the occurrence of tinnitus and/or hyperacusis.
Tinnitus may result from an abnormality/dysfunction atny level of the auditory system. Tinnitus-related activity,s any auditory signal, is, probably, subject to the normalrocess of habituation/adaptation, which protects the au-itory system from overstimulation through the attenuationf repetitive signals. This is supported by the observationhat the intrusiveness of tinnitus declines over time (Tylernd Baker, 1983; Andersson et al., 2001).
The process of habituation involves complex neuronalircuits and multiple transmitter system (Mesulam, 1990;andel, 2001), including acetylcholinergic, dopaminergic,ABA-ergic, nitric oxide and serotonergic systems. The
erotonergic system is thought to play a role in modulatingeuronal responses to repetitive stimulation and to act as“gain-control” between facilitating and inhibitory mecha-
isms (Hegerl and Juckel, 1993). There is also evidenceor the involvement of the serotonergic system in the pro-ess of habituation of sensory stimuli (Gottfries et al.,976; Hegerl and Juckel, 1993). Alterations in these neu-otransmitter systems may lead to a dysfunction of therocess of habituation. Indeed, their dysregulation haseen associated with the pathophysiology of depressionnd mood disorders (Owens and Nemeroff, 1998; Resslernd Nemeroff, 2000; Meyer et al., 2001). 5-HT has alsoeen suggested to play a role in the generation of tinnitusnd hyperacusis (Marriage and Barnes, 1995; Gopal et al.,000; Simpson and Davies, 2000), which may explain theell-recognized co-morbidity of severe tinnitus/hyperacu-is and psychological disorders (Forsling et al., 1996;oger et al., 2001).
The contribution of stress, which activates various bi-logical functions, including the above neurotransmitterystems, may influence the auditory system through differ-nt pathways, particularly in individuals with impairedechanisms of habituation, e.g. in individuals with psycho-
ogical/psychiatric disorders, as outlined above. The linketween stress and tinnitus is well recognized (Erlandssonnd Hallberg, 2000; Holgers et al., 2000). In addition,ysfunction of adrenal stress hormones, such as in Addi-on disease (Henkin et al., 1967; Henkin and Daly, 1968)ay lead to hyperacusis.
The reproductive hormones may also have a role in theccurrence of tinnitus and hyperacusis: both estrogen androgesterone could influence the auditory system, as de-
cribed earlier in this text. Estrogen has been generally
crtaotalt
M
Mppp
bea
R
S
F
P
Wam
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 893
onsidered to have an excitatory role and to have a neu-oprotective effect, and there is a strong relationship be-ween estrogen and serotoninergic pathways (Rubinow etl., 1998). On the other hand, progesterone and its metab-lites are known to have a potent inhibitory effect, through
he interaction with the GABA receptors. Therefore, alter-tions in these hormones, both physiological and patho-
ogical, may lead to increased susceptibility to developinginnitus.
enière’s disease and endocrine system
enière’s disease is a disease of poorly understood etio-athophysiology. The assumption that hormones couldlay is role in pathogenesis of the disease may be sup-orted by the following data:
● The female to male ratio shows a slight female pre-ponderance (1.3:1) (Schessel et al., 1998), and thepeak incidence for the disease is in the fourth to sixthdecade of life (Paparella, 1991), which spans meno-pause. This may suggest that the alterations in repro-ductive hormone levels may aggravate or initiate thedisease.
● The symptoms of Menière’s disease often becomemanifest during periods of stress (American Psychi-atric Association, 1994) and the disorder is morecommon in professionals and management occupa-tions (Watanabe et al., 1995) implying that the endo-crine and neural changes associated with the stressresponse may be involved in pathogenesis of thedisease or in its exacerbation.
● Endolymphatic hydrops associated with Menière’sdisease may involve hormones maintaining fluid andelectrolyte balance (Juhn et al., 1991; Naftalin, 1994).
A few studies have tried to explore the relationshipetween the endocrine system and Menière’s disease,specially the possible role of reproductive, stress and fluidnd electrolyte related hormones.
eproductive hormones
� The possible role of a sharp fall in the levels ofreproductive hormones pre-menstrually with an ex-acerbation of Menière’s disease has been noted insome studies. Andrews et al. (1992) found that only6 of 109 women reported this relationship, whileMorse and House (2001) noted that in 11 out of the13 females more symptoms were reported pre-men-strually. Price et al. (1994) reported a case in whichthe patient’s symptoms were exacerbated pre-men-strually but disappeared with leuprolide (GnRH ago-nist that leads to ablation of sex steroids by down-regulation of GnRH receptors and decreases secre-tion of FSH and LH leading to anovulation andamenorrhea) treatment. The symptoms recurredwhen she was prescribed progesterone, but notwhen given estrogen. The fluid retention or redistri-bution that occurs in some women in the pre-men-strual period (Tollan et al., 1993) may also play a role
in exacerbating symptoms. The fluctuation of symp- ltoms during the menstrual cycle in some patientswith Ménière’s disease may imply that variation ineither the reproductive or stress-related hormonesmight be involved in the pathophysiology of this dis-ease.
� The specific influence of menopause, when there isa sharp fall in reproductive hormones, on the onsetor worsening Menière’s disease has not been re-ported in the literature.
� The fluid electrolyte changes and higher levels ofprogesterone seen in pregnancy may also affectMenière’s disease. More attacks have been reportedduring early pregnancy, when the serum osmolarityis low, in a case report by Uchide et al. (1997), whichis possibly related to the sharp rise of hormonesduring the first trimester.
tress-related hormones
� Exacerbation of Menière’s disease tends to occur inrelation to stress and adrenal hormones are thoughtto be involved (Mateijsen et al., 2001).
� Patients with Menière’s disease were found to havehigher levels of cortisol compared with healthy con-trols (van Cruijsen et al., 2005). However, the higherlevels of cortisol are possibly the result of the chroniccourse of the disease, not the cause, as those with alonger history of Menière’s disease had the highestlevels of cortisol.
� Patients with Menière’s disease may have alteredresponse to stress. In comparison with patients withfacial spasm, which is assumed to cause stress butis not contributed to/triggered by stress, the patientswith Menière’s disease displayed a strong positivecorrelation between cortisol and adrenocorticotropichormone, as well as a significant correlation betweencortisol and prolactin (Horner and Cazals, 2005).
luid and electrolyte related hormones
� Altered inner ear fluid homeostasis may be the un-derlying mechanism, as seen in animal models ofendolymphatic hydrops (Juhn et al., 1991; Naftalin,1994; Juhn et al., 1999).
� The possible effect of ADH (vasopressin) inMenière’s disease has also been studied and anincrease in the level of plasma ADH in patients withMenière’s disease has been related to the vertigi-nous attacks (Takeda et al., 1995; Aoki et al., 2005).However, patients with Menière’s were not found tohave abnormal levels of aldosterone compared withcontrols (Mateijsen et al., 2001). Aldosterone levelsmaybe abnormally elevated before or after an attack,which are difficult to measure clinically.
re-menstrual syndrome and auditory function
omen who suffer from premenstrual syndrome (PMS),lso known as premenstrual dysphoric disorder (PMDD),ay have changes in their auditory function.
PMDD is a syndrome occurring specially during the
ate luteal phase of the ovarian cycle and is characterized
bpar1pvcS
lphDcehsa(
p(e(ei
ehlealocfltnEPpttg
Tmmarhb
spccepc
matdshpvflatlp
ds
a
n
A
A
A
A
A
A
A
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900894
y moderate to severe alteration in mood, behavior andhysical well-being that impairs the personal, professionalnd/or social functions in 5–8% of women during theireproductive years (American Psychiatric Association,994; Yonkers, 1997). The underlying pathophysiology isoorly understood, but may be related to progesterone iniew of the cyclic nature of the symptoms, although noonsistent finding has been documented (Rubinow andchmidt, 1995).
Dysregulation of the stress response and stress-re-ated hormones such as ACTH, �-endorphin and cortisollay a role in mood disorders, such as depression, so theyave also been implicated in the pathogenesis of PMS.ifferences in basal levels of these hormones betweenontrol women and those with PMS have not been firmlystablished (Bloch et al., 1998). However, a recent studyas found that women with PMS have dysregulation of thetress response (activation of the HPA axis in response ton external stressor) compared with those without PMSRoca et al., 2003).
Allopregnalone, a metabolite of progesterone and aotent GABA-A agonist, is also associated with stressBaulieu, 1998), and has a profound anxiolytic effect (Brott al., 1997). Women with PMS may have altered GABAEpperson et al., 2002) and/or allopregnalone (Monteleonet al., 2000; Girdler et al., 2001) functions, thus contribut-
ng to the mood disorder.These alterations in women with PMS have a potential
ffect on the auditory system. Changes in auditory functionave been documented in a study that compared ABR
atencies between women with and without PMS (Howardt al., 1992). Women with severe PMS had longer wave IIInd wave V latencies, while those with moderate PMS had
onger wave III latencies only, compared with women with-ut PMS. Howard et al. (1992) also noted that ABR laten-ies did not change after successful treatment of PMS withuoxetine (a selective 5-HT reuptake inhibitor) suggestinghat the difference in ABR latencies could be due to aeural abnormality at the brainstem level. In addition,hlers et al. (1996) reported that women diagnosed withMS had longer P3 latency of the auditory event–relatedotentials compared with normal controls. The presence ofhese differences in ABR and auditory event–related po-entials between women with and without PMS may sug-est a neurobiological predisposition to PMS.
CONCLUSION
his review highlights the complexity of the effects of hor-ones on the auditory system, with multidirectional andultidimensional interactions. An understanding of the ex-ct mechanism of hormonal influence on auditory functionemains limited. The reproductive steroids, stress-relatedormones and hormones regulating fluid and electrolytealance may all influence auditory function by:
● Direct action through the receptors within the auditorysystem
● Indirect actions
Modulating blood supply to the auditory systemRegulating the fluid electrolyte balance of the co-chlear fluidsModulating the effects of neurotransmitters presentalong the auditory pathwaysEffect through the extra auditory structures in theCNS, through their links with the auditory system
The same hormone may exert its action on the auditoryystem through multiple pathways (modulating blood sup-ly, inner ear fluids, sensory neurotransmission in the co-hlea, brain stem and cortex or through extra-auditoryonnections). For example, estrogen could lead to neuralxcitation and thus facilitate auditory transmission, but theossible increase in neurosteroids in the brainstem mayounteract this effect.
The difficulty in pinpointing the exact effect of a hor-one on the auditory system arises from the complexnatomy of the auditory system and the interactions be-ween the various hormones that occur as the result ofynamic nature of the physiological processes. However,ome general effects can be observed. The reproductiveormones, in general, facilitate auditory function, with arotective effect on the auditory system. Aldosterone andasopressin are mainly involved in maintaining cochlearuid balance. Melatonin possibly has a protective effect onuditory system. The hormones involved in the responseo stress are, generally, excitatory, but in the long run mayead to damage and may play an important role in auditoryathology.
Undoubtedly, there is a need for further studies withetailed analysis of auditory function in different hormonaltates, which may help in the development of:
new pharmacological strategy for the hormone-basedtreatment of some patients with auditory dysfunction;and
on-invasive monitoring of the hormone variations usingauditory tests such as the SOAE during the menstrualcycle.
REFERENCES
dams JC (1980) Crossed and descending projections to the inferiorcolliculus. Neurosci Lett 19(1):1–5.
ltemus M, Roca C, Galliven E, Romanos C, Deuster P (2001) In-creased vasopressin and adrenocorticotropin responses to stressin the mid luteal phase of the menstrual cycle. J Clin EndocrinolMetab 86(6):2525–2530.
merican Psychiatric Association (1994) Diagnostic and statisticalmanual of mental disorders, 4th ed. Washington, DC: AmericanPsychiatric Association.
mit G, Animesh B (2004) Effect of hormones on TEOAEs. Indian JOtol 10(1):24–26.
nari M, Axelsson A, Eliasson A, Magnusson L (1999) Hypersensitivityto sound-questionnaire data, audiometry and classification. ScandAudiol 28(4):219–230.
ndersson G, Vretblad P, Larsen HC, Lyttkens L (2001) Longitudinalfollow-up of tinnitus complaints. Arch Otolaryngol Head Neck Surg127(2):175–179.
ndrews JC, Ator GA, Honrubia V (1992) The exacerbation of symp-toms in Meniere’s disease during the premenstrual period. Arch
Otolaryngol Head Neck Surg 118(1):74–78.
A
A
B
B
B
B
B
B
B
B
B
B
B
C
C
C
C
C
C
C
C
C
C
C
C
D
Dd
D
D
D
D
D
E
E
E
E
E
E
E
E
F
F
F
F
F
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 895
ndreyko JL, Jaffe RB (1989) Use of a gonadotropin-releasing hor-mone agonist analogue for treatment of cyclic auditory dysfunction.Obstet Gynecol 74(3 Pt 2):506–509.
oki M, Ando K, Kuze B, Mizuta K, Hayashi T, Ito Y (2005) Theassociation of antidiuretic hormone levels with an attack of Me-niere’s disease. Clin Otolaryngol 30(6):521–525.
aulieu EE (1998) Neurosteroids: A novel function of the brain. Psy-choneuroendocrinology 23(8):963–987.
ecker JB (2002) Hormonal influences on sensorimotor function. In:Behavioral endocrinology (Becker JB, Breedlove SM, Crews D,McCarthy MM, eds), pp 497–525. Cambridge: MIT Press.
ehl C, Manthey D (2000) Neuroprotective activities of estrogen: Anupdate. J Neurocytol 29(5–6):351–358.
ell A (1992) Circadian and menstrual rhythms in frequency variationsof spontaneous otoacoustic emissions from human ears. Hear Res58(1):91–100.
iesalski HK, Welker HA, Thalmann R, Vollrath L (1988) Melatoninand other serotonin derivatives in the guinea pig membranouscochlea. Neurosci Lett 91(1):41–46.
irzniece V, Backstrom T, Johansson IM, Lindblad C, Lundgren P,Lofgren M, et al. (2006) Neuroactive steroid effects on cognitivefunctions with a focus on the serotonin and GABA systems. BrainRes Rev 51(2):212–239.
loch M, Schmidt PJ, Su TP, Tobin MB, Rubinow DR (1998) Pituitary-adrenal hormones and testosterone across the menstrual cycle inwomen with premenstrual syndrome and controls. Biol Psychiatry43(12):897–903.
lock GD, Kerbeshian M, Herzog ED (2000) Chronobiology. In: Neu-roendocrinology in physiology and medicine (Conn PM, FreemanME, eds), pp 391–403. Totowa, NJ: Humana Press Inc.
rook CGD, Marshall NJ (2001) The adrenal gland. In: Essentialendocrinology, Vol. 4 (Brook CGD, Marshall NJ, eds), pp 57–77.Oxford: Blackwell Science.
rot MD, Akwa Y, Purdy RH, Koob GF, Britton KT (1997) The anxio-lytic-like effects of the neurosteroid allopregnanolone: Interactionswith GABA(A) receptors. Eur J Pharmacol 325(1):1–7.
rown MC (2001) Functional neuroanatomy of the cochlea. In: Phys-iology of the ear (Jahn AF, Santos-Sacchi J, eds), pp 529–548.Australia: Singular.
ampos ML, de Cabo C, Wisden W, Juiz JM, Merlo D (2001) Expres-sion of GABA(A) receptor subunits in rat brainstem auditory path-ways: Cochlear nuclei, superior olivary complex and nucleus of thelateral lemniscus. Neuroscience 102(3):625–638.
arlsson M, Carlsson A (1988) A regional study of sex differences inrat brain serotonin. Prog Neuropsychopharmacol Biol Psychiatry12(1):53–61.
aruso S, Cianci A, Grasso D, Agnello C, Galvani F, Maiolino L, et al.(2000) Auditory brainstem response in postmenopausal womentreated with hormone replacement therapy: A pilot study. Meno-pause 7(3):178–183.
aruso S, Maiolino L, Agnello C, Garozzo A, Di Mari L, Serra A(2003a) Effects of patch or gel estrogen therapies on auditorybrainstem response in surgically postmenopausal women: A pro-spective, randomized study. Fertil Steril 79(3):556–561.
aruso S, Maiolino L, Rugolo S, Intelisano G, Farina M, Cocuzza S,et al. (2003b) Auditory brainstem response in premenopausalwomen taking oral contraceptives. Hum Reprod 18(1):85–89.
eranic B, Luxon LM. Tinnitus and other dysacuses. In: Scott-Brown’sOtorhinolaryngology, Head and Neck Surgery, 7th ed (Gleeson M,Luxon LM, eds). London: Edward Arnold (Publishers) Ltd, in press.
hermak GD, Musiek FE (1997) Neurobiology of the central auditorynervous system relevant to central auditory processing. In: Centralauditory processing disorders new prespectives (Chermak GD,Musiek FE, eds), pp 27–70. San Diego: Singular Publishing Group.
hiarenza GA, D’Ambrosio GM, Cazzullo AG (1988) Sex and eardifferences of brain-stem acoustic evoked potentials in a sample ofnormal full-term newborns. Normative study. Electroencephalogr
Clin Neurophysiol 71(5):357–366.oleman JR, Campbell D, Cooper WA, Welsh MG, Moyer J (1994)Auditory brainstem responses after ovariectomy and estrogen re-placement in rat. Hear Res 80(2):209–215.
ooper WA, Ross KC, Coleman JR (1999) Estrogen treatment andage effects on auditory brainstem responses in the post-breedinglong-evans rat. Audiology 38(1):7–12.
ox JR (1980) Hormonal influence on auditory function. Ear Hear1(4):219–222.
rain SM, Shen KF (1998) Gm1 ganglioside-induced modulation ofopioid receptor-mediated functions. Ann N Y Acad Sci 845:106–125.
allos P, He DZ, Lin X, Sziklai I, Mehta S, Evans BN (1997) Acetyl-choline, outer hair cell electromotility, and the cochlear amplifier.J Neurosci 17(6):2212–2226.
avis A (1995) Hearing in adults. London: Whurr.e Courten-Myers GM (1999) The human cerebral cortex: Gender
differences in structure and function. J Neuropathol Exp Neurol58(3):217–226.
ehan CP, Jerger J (1990) Analysis of gender differences in theauditory brainstem response. Laryngoscope 100(1):18–24.
espopoulos A, Silbernagl S (1991) Autonomic nervous system. In:Color atlas of physiology, Vol. 4 (Despopoulos A, Silbernagl S,eds), pp 50–59. Stuttgart: Thieme.
isney A, Calford MB (2001) Neurosteroids mediate habituation andtonic inhibition in the auditory midbrain. J Neurophysiol 86(2):1052–1056.
jahanbakhch O, Swanton IA, Corrie JE, McNeilly AS (1981) Predic-tion of ovulation by progesterone. Lancet 2(8256):1164–1165.
jahanbakhch O, Warner P, McNeilly AS, Baird DT (1984) Pulsatilerelease of LH and oestradiol during the periovulatory period inwomen. Clin Endocrinol (Oxf) 20(5):579–589.
ggermont JJ (2003) Central tinnitus. Auris Nasus Larynx 30(Suppl):7–12.
hlers CL, Phillips E, Parry BL (1996) Electrophysiological findingsduring the menstrual cycle in women with and without late lutealphase dysphoric disorder: Relationship to risk for alcoholism?BiolPsychiatry 39(8):720–732.
lkind-Hirsch KE, Stoner WR, Stach BA, Jerger JF (1992a) Estrogeninfluences auditory brainstem responses during the normal men-strual cycle. Hear Res 60(2):143–148.
lkind-Hirsch KE, Wallace E, Stach BA, Jerger JF (1992b) Cyclicsteroid replacement alters auditory brainstem responses in youngwomen with premature ovarian failure. Hear Res 64(1):93–98.
lkind-Hirsch KE, Wallace E, Malinak LR, Jerger JJ (1994) Sexhormones regulate ABR latency. Otolaryngol Head Neck Surg110(1):46 –52.
pperson CN, Haga K, Mason GF, Sellers E, Gueorguieva R, ZhangW, et al. (2002) Cortical gamma-aminobutyric acid levels acrossthe menstrual cycle in healthy women and those with premenstrualdysphoric disorder: A proton magnetic resonance spectroscopystudy. Arch Gen Psychiatry 59(9):851–858.
rlandsson SI, Hallberg LR (2000) Prediction of quality of life inpatients with tinnitus. Br J Audiol 34(1):11–20.
ybalin M (1993) Neurotransmitters and neuromodulators of the mam-malian cochlea. Physiol Rev 73(2):309–373.
agan PL, Church GT (1986) Effect of the menstrual cycle on theauditory brainstem response. Audiology 25(6):321–328.
aingold CL, Hoffmann WE, Caspary DM (1989) Effects of excitantamino acids on acoustic responses of inferior colliculus neurons.Hear Res 40(1–2):127–136.
ehm-Wolfsdorf G, Nagel D (1996) Differential effects of glucocorti-coids on human auditory perception. Biol Psychol 42(1–2):117–130.
ehm-Wolfsdorf G, Soherr U, Arndt R, Kern W, Fehm HL, Nagel D(1993) Auditory reflex thresholds elevated by stress-induced cor-tisol secretion. Psychoneuroendocrinology 18(8):579–589.
elix D, Ehrenberger K (1992) The efferent modulation of mammalian
inner hair cell afferents. Hear Res 64(1):1–5.
F
F
F
F
F
F
F
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
H
H
H
H
H
H
H
H
H
H
H
H
H
H
H
H
H
H
H
H
I
J
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900896
errary E, Bernard C, Teixeira M, Julien N, Bismuth P, Sterkers O, etal. (1996) Hormonal modulation of inner ear fluids. Acta Otolaryn-gol 116(2):244–247.
errary E, Sterkers O (1998) Mechanisms of endolymph secretion.Kidney Int Suppl 65:S98–S103.
errer J, Mtnez-Guisasola J, Diaz F, Alonso F, Guerrero M, Marin B(1997) Plasma levels of beta-endorphin during the menstrual cycle.Gynecol Endocrinol 11(2):75–82.
ollesa P, Concas A, Porcu P, Sanna E, Serra M, Mostallino MC, et al.(2001) Role of allopregnanolone in regulation of GABA(A) receptorplasticity during long-term exposure to and withdrawal from pro-gesterone. Brain Res Brain Res Rev 37(1–3):81–90.
orsling ML, Anderson CH, Wheeler MJ, Raju KS (1996) The effect ofoophorectomy and hormone replacement on neurohypophysealhormone secretion in women. Clin Endocrinol (Oxf) 44(1):39–44.
uruta H, Luo L, Ryan AF, Mori N (1998) Expression of mRNA en-coding vasopressin V1a, vasopressin V2, and ANP-B receptors inthe rat cochlea. Hear Res 117(1–2):140–148.
uruta H, Mori N, Sato C, Hoshikawa H, Sakai S, Iwakura S, et al.(1994) Mineralocorticoid type I receptor in the rat cochlea: mRNAidentification by polymerase chain reaction (PCR) and in situ hy-bridization. Hear Res 78(2):175–180.
arcia-Segura LM, Azcoitia I, DonCarlos LL (2001) Neuroprotectionby estradiol. Prog Neurobiol 63(1):29–60.
enazzani AR, Lemarchand-Beraud T, Aubert ML, Felber JP (1975)Pattern of plasma ACTH, hGH, and cortisol during menstrual cycle.J Clin Endocrinol Metab 41(3):431–437.
enazzani AR, Stomati M, Morittu A, Bernardi F, Monteleone P,Casarosa E, et al. (2000) Progesterone, progestagens and thecentral nervous system. Hum Reprod 15(Suppl 1):14–27.
il-Loyzaga P, Bartolome V, Vicente-Torres A, Carricondo F (2000)Serotonergic innervation of the organ of corti. Acta Otolaryngol120(2):128–132.
il-Loyzaga PE (1995) Neurotransmitters of the olivocochlear lateralefferent system: With an emphasis on dopamine. Acta Otolaryngol115(2):222–226.
irdler SS, Straneva PA, Light KC, Pedersen CA, Morrow AL (2001)Allopregnanolone levels and reactivity to mental stress in premen-strual dysphoric disorder. Biol Psychiatry 49(9):788–797.
oel N (2005) Late night presentation of an auditory stimulus phasedelays human circadian rhythms. Am J Physiol Regul Integr CompPhysiol 289:R209–R216.
olombek DA, Pevet P, Cardinali DP (1996) Melatonin effects onbehavior: Possible mediation by the central gabaergic system.Neurosci Biobehav Rev 20(3):403–412.
opal KV, Daly DM, Daniloff RG, Pennartz L (2000) Effects of selec-tive serotonin reuptake inhibitors on auditory processing: Casestudy. J Am Acad Audiol 11(8):454–463.
ottfries CG, von Knorring L, Perris C (1976) Neurophysiologicalmeasures related to levels of 5-hydroxyindoleacetic acid, ho-movanillic acid and tryptophan in cerebrospinal fluid of psychiatricpatients. Neuropsychobiology 2(1):1–8.
ray TS, Bingaman EW (1996) The amygdala: Corticotropin-releasingfactor, steroids, and stress. Crit Rev Neurobiol 10(2):155–168.
uardiola-Lemaitre B, Lenegre A, Porsolt RD (1992) Combined ef-fects of diazepam and melatonin in two tests for anxiolytic activityin the mouse. Pharmacol Biochem Behav 41(2):405–408.
uimaraes P, Frisina ST, Mapes F, Tadros SF, Frisina DR, Frisina RD(2006) Progestin negatively affects hearing in aged women. ProcNatl Acad Sci U S A 103(38):14246–14249.
uimaraes P, Zhu X, Cannon T, Kim S, Frisina RD (2004) Sexdifferences in distortion product otoacoustic emissions as a func-tion of age in CBA mice. Hear Res 192(1–2):83–89.
urr P, Owen G, Reid A, Canter R (1993) Tinnitus in pregnancy. ClinOtolaryngol 18(4):294–297.
aapaniemi JJ (1996) The hearing threshold levels of children at
school age. Ear Hear 17(6):469–477.adley ME (2000) Endocrinology of pregnancy, parturition, annd lac-tation. In: Endocrinology, 5th ed (Hadley ME, ed), pp 474–495.New Jersey: Prentice-Hall, Inc.
aggerty HS, Lusted HS, Morton SC (1993) Statistical quantification of24-h and monthly variabilities of spontaneous otoacoustic emis-sion frequency in humans. Hear Res 70(1):31–49.
alasz B (2000) The hypothalamus as an endocrine organ: The sci-ence of neuroendocrinology. In: Neuroendocrinology in physiologyand medicine (Conn PM, Freeman ME, eds), pp 3–21. Totowa, NJ:Humana Press Inc.
all JW III (1992) Effect of nonpathological subject characteristics. In:Handbook of auditory evoked responses (Hall JW III, ed), pp163–220. Boston: Allyn and Bacon.
all JW (2000) Distortion product and transient evoked OAEs: Non-pathologic factors influencing measurement. In: Handbook of oto-acoustic emissions (Hall JW III, ed), pp 163–220. San Diego:Singular Publishing Group.
ampson E (2002) Sex differences in human brain and cognition: Theinfluence of sex steroids in early and adult life. In: Behavioralendocrinology, 2nd ed (Becker JB, Breedlove SM, Crews D,McCarthy MM, eds), pp 579–628. Cambridge: MIT Press.
ederstierna C, Hultcrantz M, Collins A, Rosenhall U (2007) Hearingin women at menopause. Prevalence of hearing loss, audiometricconfiguration and relation to hormone replacement therapy. ActaOtolaryngol 127(2):149–155.
egerl U, Juckel G (1993) Intensity dependence of auditory evokedpotentials as an indicator of central serotonergic neurotransmission: Anew hypothesis. Biol Psychiatry 33(3):173–187.
enkin RI, Daly RL (1968) Auditory detection and perception in normalman and in patients with adrenal cortical insufficiency: Effect ofadrenal cortical steroids. J Clin Invest 47(6):1269–1280.
enkin RI, McGlone RE, Daly R, Bartter FC (1967) Studies on auditorythresholds in normal man and in patients with adrenal corticalinsufficiency: The role of adrenal cortical steroids. J Clin Invest46(3):429–435.
ildesheimer M, Henkin Y, Pye A, Heled S, Sahartov E, Shabtai EL,et al. (2002) Bilateral superior cervical sympathectomy and noise-induced, permanent threshold shift in guinea pigs. Hear Res163(1–2):46–52.
ildesheimer M, Sharon R, Muchnik C, Sahartov E, Rubinstein M(1991) The effect of bilateral sympathectomy on noise inducedtemporary threshold shift. Hear Res 51(1):49–53.
olgers KM, Erlandsson SI, Barrenas ML (2000) Predictive factors forthe severity of tinnitus. Audiology 39(5):284–291.
orner KC (2003) The emotional ear in stress. Neurosci Biobehav Rev27(5):437–446.
orner KC, Cazals Y (2005) Stress hormones in Meniere’s diseaseand acoustic neuroma. Brain Res Bull 66(1):1–8.
orner KC, Giraudet F, Lucciano M, Cazals Y (2001) Sympathectomyimproves the ear’s resistance to acoustic trauma: could stressrender the ear more sensitive? Eur J Neurosci 13(2):405–408.
oward R, Mason P, Taghavi E, Spears G (1992) Brainstem auditoryevoked responses (BAERS) during the menstrual cycle in womenwith and without premenstrual syndrome. Biol Psychiatry 32(8):682–690.
uffman RF, Henson OW Jr (1990) The descending auditory pathwayand acousticomotor systems: Connections with the inferior collicu-lus. Brain Res Brain Res Rev 15(3):295–323.
urley LM, Thompson AM, Pollak GD (2002) Serotonin in the inferiorcolliculus. Hear Res 168(1–2):1–11.
maki T, Nahan JL, Rivier C, Sawchenko PE, Vale W (1991) Differen-tial regulation of corticotropin-releasing factor mRNA in rat brainregions by glucocorticoids and stress. J Neurosci 11(3):585–599.
anssen R, Schweitzer L, Jensen KF (1991) Glutamate neurotoxicityin the developing rat cochlea: Physiological and morphological
approaches. Brain Res 552(2):255–264.
J
J
J
J
J
J
J
J
J
K
K
K
K
K
K
K
K
K
K
K
K
K
K
K
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
M
M
M
M
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 897
ennes L, Langub C (2000) Endocrine targets in the brain. In: Neu-roendocrinology in physiology and medicine (Conn PM, FreemanME, eds), pp 95–104. Totowa, NJ: Humana Press Inc.
erger J, Hall J (1980) Effects of age and sex on auditory brainstemresponse. Arch Otolaryngol 106(7):387–391.
erger J, Johnson K (1988) Interactions of age, gender, and sensori-neural hearing loss on ABR latency. Ear Hear 9(4):168–176.
ohansson MS, Arlinger SD (2002) Hearing threshold levels for anotologically unscreened, non-occupationally noise-exposed popu-lation in Sweden. Int J Audiol 41(3):180–194.
ongkamonwiwat N, Phansuwan-Pujito P, Casalotti SO, Forge A, Dod-son H, Govitrapong P (2006) The existence of opioid receptors inthe cochlea of guinea pigs. Eur J Neurosci 23(10):2701–2711.
ongkamonwiwat N, Phansuwan-Pujito P, Sarapoke P, ChetsawangB, Casalotti SO, Forge A, et al. (2003) The presence of opioidreceptors in rat inner ear. Hear Res 181(1–2):85–93.
uckel G, Molnar M, Hegerl U, Csepe V, Karmos G (1997) Auditory-evoked potentials as indicator of brain serotonergic activity- firstevidence in behaving cats. Biol Psychiatry 41(12):1181–1195.
uhn SK, Ikeda K, Morizono T, Murphy M (1991) Pathophysiology ofinner ear fluid imbalance. Acta Otolaryngol Suppl 485:9–14.
uhn SK, Li W, Kim JY, Javel E, Levine S, Odland RM (1999) Effect ofstress-related hormones on inner ear fluid homeostasis and func-tion. Am J Otol 20(6):800–806.
adva A, Djahanbakhch O, Monson J, Di WL, Silman R (1998) Ele-vated nocturnal melatonin is a consequence of gonadotropin-re-leasing hormone deficiency in women with hypothalamic amenor-rhea. J Clin Endocrinol Metab 83(10):3653–3662.
andel ER (2001) The molecular biology of memory storage: Adialogue between genes and synapses. Science 294(5544):1030 –1038.
atzenell U, Segal S (2001) Hyperacusis: Review and clinical guide-lines. Otol Neurotol 22(3):321–326.
atzenellenbogen BS (2000) Mechanisms of action and cross-talkbetween estrogen receptor and progesterone receptor pathways. JSoc Gynecol Investig 7(1 Suppl):S33–S37.
ei J, McPherson B, Smyth V, Latham S, Loscher J (1997) Transientevoked otoacoustic emissions in infants: Effects of gender, earasymmetry and activity status. Audiology 36(2):61–71.
emp DT, Ryan S, Bray P (1990) A guide to the effective use ofotoacoustic emissions. Ear Hear 11(2):93–105.
haliq F, Tandon OP, Goel N (2003) Auditory evoked responses inpostmenopausal women on hormone replacement therapy. IndianJ Physiol Pharmacol 47(4):393–399.
haliq F, Tandon OP, Goel N (2005) Differential effects of exogenousestrogen versus a estrogen-progesterone combination on auditoryevoked potentials in menopausal women. Indian J Physiol Phar-macol 49(3):345–352.
ilicdag EB, Yavuz H, Bagis T, Tarim E, Erkan AN, Kazanci F (2004)Effects of estrogen therapy on hearing in postmenopausal women.Am J Obstet Gynecol 190(1):77–82.
im SH, Kang BM, Chae HD, Kim CH (2002) The association betweenserum estradiol level and hearing sensitivity in postmenopausalwomen. Obstet Gynecol 99(5 Pt 1):726–730.
irschbaum C, Kudielka BM, Gaab J, Schommer NC, Hellhammer DH(1999) Impact of gender, menstrual cycle phase, and oral contra-ceptives on the activity of the hypothalamus-pituitary-adrenal axis.Psychosom Med 61(2):154–162.
ostoglou-Athanassiou I, Treacher DF, Wheeler MJ, Forsling ML(1998a) Melatonin administration and pituitary hormone secretion.Clin Endocrinol (Oxf) 48(1):31–37.
ostoglou-Athanassiou I, Athanassiou P, Treacher DF, Wheeler MJ,Forsling ML (1998b) Neurohypophysial hormone and melatoninsecretion over the natural and suppressed menstrual cycle inpremenopausal women. Clin Endocrinol (Oxf) 49(2):209–216.
udo M, Moriya M, Mizuno N (1997) Auditory projections from the ICto the SCN by way of the lg in the mole, mogera. Neuroreport
8(15):3405–3409.uiper GG, Shughrue PJ, Merchenthaler I, Gustafsson JA (1998) Theestrogen receptor beta subtype: A novel mediator of estrogenaction in neuroendocrine systems. Front Neuroendocrinol 19(4):253–286.
amprecht-Dinnesen A, Pohl M, Hartmann S, Heinecke A, Ahrens S,Muller E, et al. (1998) Effects of age, gender and ear side on SOAEparameters in infancy and childhood. Audiol Neurootol 3(6):386–401.
ancel M (1993) Cortical and subcortical EEG in relation to sleep-wakebehavior in mammalian species. Neuropsychobiology 28(3):154 –159.
ang I, Zielinski CC, Templ H, Spona J, Geyer G (1990) Medroxypro-gesterone acetate lowers plasma corticotropin and cortisol butdoes not suppress anterior pituitary responsiveness to humancorticotropin releasing factor. Cancer 66(9):1949–1953.
augel GR, Dengerink HA, Wright JW (1987) Ovarian steroid andvasoconstrictor effects on cochlear blood flow. Hear Res31(3):245–251.
eDoux JE (1993) Emotional memory systems in the brain. BehavBrain Res 58(1–2):69–79.
eDoux JE, Sakaguchi A, Reis DJ (1984) Subcortical efferent projec-tions of the medial geniculate nucleus mediate emotional re-sponses conditioned to acoustic stimuli. J Neurosci 4(3):683–698.
ee JH, Marcus DC (2001) Estrogen acutely inhibits ion transport byisolated stria vascularis. Hear Res 158(1–2):123–130.
e Prell CG, Bledsoe SC, Bobbin RP, Puel JL (2001) Neurotransmis-sion in the inner ear: Functional and molecular analysis. In: Phys-iology of the ear, 2nd ed (Jahn A, Santos-Sacchi J, eds), pp575–611. Australia: Singular.
evine JE (2000) The hypothalamus as a major integrating center. In:Neuroendocrinology in physiology and medicine (Conn PM, Free-man ME, eds), pp 75–93. Totowa, NJ: Humana Press Inc.
ohuis PJ, Borjesson PK, Klis SF, Smoorenburg GF (2000) The ratcochlea in the absence of circulating adrenal hormones: An elec-trophysiological and morphological study. Hear Res 143(1–2):189–196.
ombardi G, Zarrilli S, Colao A, Paesano L, Di Somma C, Rossi F,et al. (2001) Estrogens and health in males. Mol Cell Endocrinol178(1–2):51–55.
opez-Gonzalez MA, Guerrero JM, Delgado F (1997) Presence of thepineal hormone melatonin in rat cochlea: Its variations with lightingconditions. Neurosci Lett 238(1–2):81–83.
opez-Gonzalez MA, Guerrero JM, Rojas F, Delgado F (2000a) Oto-toxicity caused by cisplatin is ameliorated by melatonin and otherantioxidants. J Pineal Res 28(2):73–80.
opez-Gonzalez MA, Guerrero JM, Torronteras R, Osuna C, DelgadoF (2000b) Ototoxicity caused by aminoglycosides is ameliorated bymelatonin without interfering with the antibiotic capacity of thedrugs. J Pineal Res 28(1):26–33.
opez-Gonzalez MA, Guerrero JM, Rojas F, Osuna C, Delgado F(1999) Melatonin and other antioxidants prolong the postmortemactivity of the outer hair cells of the organ of corti: Its relation to thetype of death. J Pineal Res 27(2):73–77.
ains RE, Eipper BA (1999) Peptides. In: Basic neurochemistry, 6thed (Seigel GJ, Agranoff BW, Fisher SK, Uhler MD, eds), pp 363–382. New York: Lippincott-Raven Press.
arriage J, Barnes NM (1995) Is central hyperacusis a symptomof 5-hydroxytryptamine (5-HT) dysfunction?J Laryngol Otol109(10):915–921.
artinez-Guisasola J, Ferrer J, Guerrero M, Diaz F, Alonso F, BodegaA, et al. (1999) Circulating levels of immunoreactive beta-endor-phin in polycystic ovary syndrome. Gynecol Endocrinol 13(1):26–35.
ateijsen DJ, Kingma CM, De Jong PE, Wit HP, Albers FW (2001)Aldosterone assessment in patients with Meniere’s disease. ORL J
Otorhinolaryngol Relat Spec 63(5):280–286.
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
N
N
N
N
O
O
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
R
R
R
R
R
R
R
R
R
R
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900898
auri A, Martellotta MC, Melis MR, Caminiti F, Serri F, Fratta W (1990)Plasma alpha-melanocyte-stimulating hormone during the men-strual cycle in women. Horm Res 34(2):66–70.
cEwen BS, Alves SE (1999) Estrogen actions in the central nervoussystem. Endocr Rev 20(3):279–307.
cFadden D (1993) A speculation about the parallel ear asymmetriesand sex differences in hearing sensitivity and otoacoustic emis-sions. Hear Res 68(2):143–151.
cFadden D (1998) Sex differences in the auditory system. DevNeuropsychol 14(2/3):261–298.
cFadden D (2000) Masculinizing effects on otoacoustic emissionsand auditory evoked potentials in women using oral contracep-tives. Hear Res 142(1–2):23–33.
cGivern RF, Redei E (1994) Adrenalectomy reverses stress-inducedsuppression of luteinizing hormone secretion in long-term ovariec-tomized rats. Physiol Behav 55(6):1147–1150.
esulam MM (1990) Large-scale neurocognitive networks and distrib-uted processing for attention, language, and memory. Ann Neurol28(5):597–613.
eyer JH, Kruger S, Wilson AA, Christensen BK, Goulding VS, Schaf-fer A, et al. (2001) Lower dopamine transporter binding potential instriatum during depression. Neuroreport 12(18):4121–4125.
ichaud DS, McLean J, Keith SE, Ferrarotto C, Hayley S, Khan SA,et al. (2003) Differential impact of audiogenic stressors on Lewisand Fischer rats: Behavioral, neurochemical, and endocrine vari-ations. Neuropsychopharmacology 28:1068–1081.
iller MH, Gould WJ (1967) Fluctuating sensorineural hearing impair-ment associated with the menstrual cycle. J Auditory Res 7:373–385.
onteleone P, Luisi S, Tonetti A, Bernardi F, Genazzani AD, Luisi M,et al. (2000) Allopregnanolone concentrations and premenstrualsyndrome. Eur J Endocrinol 142(3):269–273.
oore JK, Moore RY (1987) Glutamic acid decarboxylase-like immu-noreactivity in brainstem auditory nuclei of the rat. J Comp Neurol260(2):157–174.
orse GG, House JW (2001) Changes in Meniere’s disease re-sponses as a function of the menstrual cycle. Nurs Res 50(5):286–292.
ulders WH, Robertson D (2001) Origin of the noradrenergic inner-vation of the superior olivary complex in the rat. J Chem Neuroanat21(4):313–322.
urphy MP, Gates GA (1997) Hearing loss: Does gender play a role?Medscape Womens Health 2(10):2.
usiek FE, Hoffman DW (1990) An introduction to the functionalneurochemistry of the auditory system. Ear Hear 11(6):395–402.
aftalin L (1994) The possible roles of hormones and enzymes in theproduction of the acute attack in Meniere’s disease. Eur ArchOtorhinolaryngol 251(3):173–177.
athan CA, Kim TS, Harris JP, Koutnouyan HA, Ryan AF (1999)Absence of mRNA encoding estrogen receptor in the rat cochlea.Acta Otolaryngol 119(8):853–857.
ilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G,et al. (2001) Mechanisms of estrogen action. Physiol Rev 81(4):1535–1565.
ishizawa S, Benkelfat C, Young SN, Leyton M, Mzengeza S, deMontigny C, et al. (1997) Differences between males and femalesin rates of serotonin synthesis in human brain. Proc Natl Acad SciU S A 94(10):5308–5313.
’Rourke C, Driscoll C, Kei J, Smyth V (2002) A normative study ofdistortion-product otoacoustic emissions in 6-year-old schoolchil-dren. Int J Audiol 41(3):162–169.
wens MJ, Nemeroff CB (1998) The serotonin transporter and de-pression. Depress Anxiety 8(Suppl 1):5–12.
aparella MM (1991) Pathogenesis and pathophysiology of Meniere’sdisease. Acta Otolaryngol Suppl 485:26–35.
arlee MB (1983) Menstrual rhythms in sensory processes: A reviewof fluctuations in vision, olfaction, audition, taste, and touch. Psy-
chol Bull 93(3):539–548.ascual E, Giner V, Arostegui A, Conill J, Ruiz MT, Pico A (1991)Higher incidence of carpal tunnel syndrome in oophorectomizedwomen. Br J Rheumatol 30(1):60–62.
earson JD, Morrell CH, Gordon-Salant S, Brant LJ, Metter EJ, KleinLL, et al. (1995) Gender differences in a longitudinal study ofage-associated hearing loss. J Acoust Soc Am 97(2):1196–1205.
echere-Bertschi A, Maillard M, Stalder H, Brunner HR, Burnier M(2002) Renal segmental tubular response to salt during the normalmenstrual cycle. Kidney Int 61(2):425–431.
enner MJ (1995) Frequency variation of spontaneous otoacousticemissions during a naturally occurring menstrual cycle, amenor-rhea, and oral contraception: A brief report. Ear Hear 16(4):428–432.
enner MJ, Zhang T (1997) Prevalence of spontaneous otoacousticemissions in adults revisited. Hear Res 103(1–2):28–34.
etraglia F, Vale W, Rivier C (1986) Opioids act centrally to modulatestress-induced decrease in luteinizing hormone in the rat. Endo-crinology 119(6):2445–2450.
faff DW, Vasudevan N, Attardi B (2000) Neuroendocrine regulation ofsexual behavior. In: Neuroendocrinology in physiology and medi-cine (Conn PM, Freeman ME, eds), pp 287–299. Totowa, NJ:Humana Press Inc.
olich J, Kok A (1995) Cognitive and biological determinants of p300:An integrative review. Biol Psychol 41(2):103–146.
rice TM, Allen TC, Bowyer DL, Watson TA (1994) Ablation of lutealphase symptoms of Meniere’s disease with leuprolide. Arch Oto-laryngol Head Neck Surg 120(2):209–211.
robst R, Lonsbury-Martin BL, Martin GK (1991) A review of otoacous-tic emissions. J Acoust Soc Am 89(5):2027–2067.
uel JL (1995) Chemical synaptic transmission in the cochlea. ProgNeurobiol 47(6):449–476.
ujol R (1994) Lateral and medial efferents: A double neurochemicalmechanism to protect and regulate inner and outer hair cell func-tion in the cochlea. Br J Audiol 28(4–5):185–191.
ujol R, Puel JL, de Gervais AC, Eybalin M (1993) Pathophysiology ofthe glutamatergic synapses in the cochlea. Acta Otolaryngol113(3):330–334.
arey KE, Curtis LM (1996) Receptors for glucocorticoids in the hu-man inner ear. Otolaryngol Head Neck Surg 115(1):38–41.
arey KE, Curtis LM, ten Cate WJ (1993) Tissue specific levels ofglucocorticoid receptor within the rat inner ear. Hear Res 64(2):205–210.
arey KE, Luttge WG (1989) Presence of type I and type II/Ib recep-tors for adrenocorticosteroid hormones in the inner ear. Hear Res41(2–3):217–221.
esende LA, Silva MD, Impemba F, Achoa NB, Schelp AO (2000)Multimodal evoked potentials and the ovarian cycle in young ovu-lating women. Arq Neuropsiquiatr 58(2B):418–423.
essler KJ, Nemeroff CB (2000) Role of serotonergic and noradren-ergic systems in the pathophysiology of depression and anxietydisorders. Depress Anxiety 12(Suppl 1):2–19.
hodes ME, Rubin RT (1999) Functional sex differences (“sexualdiergism”) of central nervous system cholinergic systems, vaso-pressin, and hypothalamic-pituitary-adrenal axis activity in mam-mals: A selective review. Brain Res Brain Res Rev 30(2):135–152.
oberts J, Huber P (1970) Hearing levels of children by age and sex:United States. Vital Health Stat 11(102):1–51.
oca CA, Schmidt PJ, Altemus M, Deuster P, Danaceau MA, PutnamK, et al. (2003) Differential menstrual cycle regulation of hypotha-lamic-pituitary-adrenal axis in women with premenstrual syndromeand controls. J Clin Endocrinol Metab 88(7):3057–3063.
ubinow DR, Schmidt PJ (1995) The neuroendocrinology of menstrualcycle mood disorders. Ann N Y Acad Sci 771:648–659.
ubinow DR, Schmidt PJ, Roca CA (1998) Estrogen-serotonin inter-actions: Implications for affective regulation. Biol Psychiatry
44(9):839–850.
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
S
T
T
T
T
T
T
T
T
T
T
T
T
T
U
U
v
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900 899
ahley TL, Nodar RH (1994) Improvement in auditory function follow-ing pentazocine suggests a role for dynorphins in auditory sensi-tivity. Ear Hear 15(6):422–431.
ahley TL, Nodar RH (2001) A biochemical model of peripheral tinni-tus. Hear Res 152(1–2):43–54.
ahley TL, Nodar RH, Musiek FE (1999) Endogenous dynorphins:Possible role in peripheral tinnitus. Int Tinnitus J 5(2):76–91.
aketos M, Sharma N, Santoro NF (1993) Suppression of the hypo-thalamic-pituitary-ovarian axis in normal women by glucocorti-coids. Biol Reprod 49(6):1270–1276.
antos-Sacchi J (2001) Cochlear physiology. In: Physiology of the ear,2nd ed (Jahn A, Santos-Sacchi J, eds), pp 357–391. Australia:Singular.
apolsky RM (2002) Endocrinology of the stress-response. In: Behav-ioral endocrinology, 2nd ed (Becker JB, Breedlove SM, Crews D,McCarthy MM, eds), pp 409–449. Cambridge: MIT Press.
ato H, Sando I, Takahashi H (1991) Sexual dimorphism and devel-opment of the human cochlea. Computer 3-d measurement. ActaOtolaryngol 111(6):1037–1040.
ator MO, Franz P, Egarter C, Gruber DM, Wolfl G, Nagele F (1999)Effects of tibolone on auditory brainstem responses in postmeno-pausal women-a randomized, double-blind, placebo-controlledtrial. Fertil Steril 72(5):885–888.
chessel DA, Minor LB, Nedzelski J (1998) Ménière’s disease andother peripheral vestibular disorders. In: Otolaryngology head andneck surgery, 3rd ed, Vol. 4 (Cummings CW, Fredrickson JM,Harker LA, Krause CJ, Richardson MA, Schuller DE, eds), pp2672–2705. St Louis: Mosby.
choltz AW, Kanonier G, Schrott-Fischer A (1998) Immunohistochem-ical investigation of enkephalins in the human inner ear. Hear Res118(1–2):123–128.
ennaroglu G, Belgin E (2001) Audiological findings in pregnancy.J Laryngol Otol 115(8):617–621.
erra A, Maiolino L, Agnello C, Messina A, Caruso S (2003) Auditorybrain stem response throughout the menstrual cycle. Ann OtolRhinol Laryngol 112(6):549–553.
harpe RM (1998) The role of oestrogen in the male. Trends Endo-crinol Metab 9(9):371–377.
himazaki T, Ichimiya I, Suzuki M, Mogi G (2002) Localization ofglucocorticoid receptors in the murine inner ear. Ann Otol RhinolLaryngol 111(12 Pt 1):1133–1138.
ilman R (1991) Melatonin and the human gonadotrophin-releasinghormone pulse generator. J Endocrinol 128(1):7–11.
impson JJ, Davies WE (2000) A review of evidence in support of arole for 5-HT in the perception of tinnitus. Hear Res 145(1–2):1–7.
ingewald N, Kaehler ST, Hemeida R, Philippu A (1998) Influence ofexcitatory amino acids on basal and sensory stimuli-induced re-lease of 5-ht in the locus coeruleus. Br J Pharmacol 123(4):746–752.
isneros JA, Bass AH (2003) Seasonal plasticity of peripheral auditoryfrequency sensitivity. J Neurosci 23(3):1049–1058.
isneros JA, Forlano PM, Deitcher DL, Bass AH (2004) Steroid-dependent auditory plasticity leads to adaptive coupling of senderand receiver. Science 305(5682):404–407.
mith MJ, Adams LF, Schmidt PJ, Rubinow DR, Wassermann EM(2002) Effects of ovarian hormones on human cortical excitability.Ann Neurol 51(5):599–603.
ouaid J, Rappaport JM (2000) Fluctuating sensorineural hearing lossassociated with the menstrual cycle. J Otolaryngol 30:246–250.
preng M (2000) Central nervous system activation by noise. NoiseHealth 2(7):49–58.
tenberg AE, Simonoska R, Stygar D, Sahlin L, Hultcrantz M (2003)Effect of estrogen and antiestrogens on the estrogen receptorcontent in the cochlea of ovariectomized rats. Hear Res182(1–2):19 –23.
tenberg AE, Wang H, Fish J III, Schrott-Fischer A, Sahlin L, Hult-
crantz M (2001) Estrogen receptors in the normal adult and devel-oping human inner ear and in Turner’s syndrome. Hear Res157(1–2):87–92.
tenberg AE, Wang H, Sahlin L, Hultcrantz M (1999) Mapping ofestrogen receptors alpha and beta in the inner ear of mouse andrat. Hear Res 136(1–2):29–34.
tomati M, Bernardi F, Luisi S, Puccetti S, Casarosa E, Liut M, et al.(2002) Conjugated equine estrogens, estrone sulphate and estra-diol valerate oral administration in ovariectomized rats: Effects oncentral and peripheral allopregnanolone and beta-endorphin. Ma-turitas 43(3):195–206.
tuart A, Yang EY (2001) Gender effects in auditory brainstem re-sponses to air- and bone-conducted clicks in neonates. J CommunDisord 34(3):229–239.
uga N, Gao E, Zhang Y, Ma X, Olsen JF (2000) The corticofugalsystem for hearing: Recent progress. Proc Natl Acad Sci U S A97(22):11807–11814.
wanson SJ, Dengerink HA (1988) Changes in pure-tone thresholdsand temporary threshold shifts as a function of menstrual cycle andoral contraceptives. J Speech Hear Res 31(4):569–574.
adros SF, Frisina ST, Mapes F, Frisina DR, Frisina RD (2005) Higherserum aldosterone correlates with lower hearing thresholds: Apossible protective hormone against presbycusis. Hear Res209(1–2):10 –18.
akeda T, Kakigi A, Saito H (1995) Antidiuretic hormone (ADH) andendolymphatic hydrops. Acta Otolaryngol Suppl 519:219–222.
andon OP, Misra R, Tandon I (1990) Brainstem auditory evokedpotentials (BAEPs) in pregnant women. Indian J Physiol Pharma-col 34(1):42–44.
ang LC, Chan SY, Tang GW, Ma HK (1987) Serum immunoreactivebeta-endorphin in the human ovulatory cycle. Int J Fertil32(2):149 –151.
asman A, Hahn T, Maiste A (1999) Menstrual cycle synchronizedchanges in brain stem auditory evoked potentials and visualevoked potentials. Biol Psychiatry 45(11):1516–1519.
hompson AM (2003) A medullary source of norepinephrine in catcochlear nuclear complex. Exp Brain Res 153(4):486–490.
hompson AM, Schofield BR (2000) Afferent projections of the supe-rior olivary complex. Microsc Res Tech 51(4):330–354.
hompson SK, Zhu X, Frisina RD (2006) Estrogen blockade reducesauditory feedback in CBA mice. Otolaryngol Head Neck Surg135(1):100–105.
ollan A, Oian P, Fadnes HO, Maltau JM (1993) Evidence for alteredtranscapillary fluid balance in women with the premenstrual syn-drome. Acta Obstet Gynecol Scand 72(4):238–242.
rune DR, Kempton JB, Kessi M (2000) Aldosterone (mineralocorti-coid) equivalent to prednisolone (glucocorticoid) in reversing hear-ing loss in MRL/MPJ-Fas1pr autoimmune mice. Laryngoscope110(11):1902–1906.
rune DR, Mitchell C, Phillips DS (1988) The relative importance ofhead size, gender and age on the auditory brainstem response.Hear Res 32(2–3):165–174.
sunoda K, Takahashi S, Takanosawa M, Shimoji Y (1999) Theinfluence of pregnancy on sensation of ear problems: ear problemsassociated with healthy pregnancy. J Laryngol Otol 113(4):318–320.
yler RS, Baker LJ (1983) Difficulties experienced by tinnitus suffer-ers. J Speech Hear Disord 48(2):150–154.
chide K, Suzuki N, Takiguchi T, Terada S, Inoue M (1997) Thepossible effect of pregnancy on Meniere’s disease. ORL J Otorhi-nolaryngol Relat Spec 59(5):292–295.
rbanski HF (2000) Influence of light and the pineal gland on biologicalrhythms. In: Neuroendocrinology in physiology and medicine(Conn PM, Freeman ME, eds), pp 405–420. Totowa, NJ: HumanaPress Inc.
an Cruijsen N, Dullaart RP, Wit HP, Albers FW (2005) Analysis ofcortisol and other stress-related hormones in patients with Me-
niere’s disease. Otol Neurotol 26(6):1214–1219.
V
W
W
W
W
W
W
W
W
Y
Y
Y
Z
Z
D. Al-Mana et al. / Neuroscience 153 (2008) 881–900900
rbicky KW, Baumstark JS, Wells IC, Hilgers TW, Kable WT, Elias CJ(1982) Evidence for the involvement of beta-endorphin in the hu-man menstrual cycle. Fertil Steril 38(6):701–704.
agner EJ, Ronnekleiv OK, Bosch MA, Kelly MJ (2001) Estrogenbiphasically modifies hypothalamic gabaergic function concomi-tantly with negative and positive control of luteinizing hormonerelease. J Neurosci 21(6):2085–2093.
arr WB (1992) Organization of the olivocochlear efferent systems inmammals. In: Mamalian auditory pathway: neuroanatomy (Web-ster DB, Popper AN, Fay RR, eds), pp 410–448. New York:Springer-Verlang.
atanabe Y, Mizukoshi K, Shojaku H, Watanabe I, Hinoki M, KitaharaM (1995) Epidemiological and clinical characteristics of Meniere’sdisease in Japan. Acta Otolaryngol Suppl 519:206–210.
esensten NJ, Badia P, Harsh J (1990) Time of day, repeated testing,and interblock interval effects on P300 amplitude. Physiol Behav47(4):653–658.
harton JA, Church GT (1990) Influence of menopause on the audi-tory brainstem response. Audiology 29(4):196–201.
it HP (1985) Diurnal cycle for spontaneous oto-acoustic emission
frequency. Hear Res 18(2):197–199.oolley CS, Weiland NG, McEwen BS, Schwartzkroin PA (1997)Estradiol increases the sensitivity of hippocampal ca1 pyramidalcells to nmda receptor-mediated synaptic input: Correlation withdendritic spine density. J Neurosci 17(5):1848–1859.
right A, Davis A, Bredberg G, Ulehlova L, Spencer H (1987) Hair celldistributions in the normal human cochlea. Acta Otolaryngol Suppl444:1–48.
adav A, Tandon OP, Vaney N (2002) Auditory evoked responsesduring different phases of menstrual cycle. Indian J Physiol Phar-macol 46(4):449–456.
ellin MW, Stillman RD (1999) Otoacoustic emissions in normal-cy-cling females. J Am Acad Audiol 10(7):400–408.
onkers KA (1997) The association between premenstrual dysphoricdisorder and other mood disorders. J Clin Psychiatry 58(Suppl15):19–25.
oger S, Svedlund J, Holgers KM (2001) Psychiatric disorders intinnitus patients without severe hearing impairment: 24-month fol-low-up of patients at an audiological clinic. Audiology 40(3):133–140.
uo J, Curtis LM, Yao X, ten Cate WJ, Bagger-Sjoback D, HultcrantzM, et al. (1995) Glucocorticoid receptor expression in the postnatal
rat cochlea. Hear Res 87(1–2):220–227.(Accepted 2 February 2008)(Available online 15 March 2008)