Homocysteine and oxidative stress in Egyptian children with Down syndrome
13
Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study Ragaa R. Hamed a , Tahany M. Maharem b , Nagwa Abdel-Meguid c , Gilane M. Sabry b , Abdel-Monem Abdalla a , Rasha A. Guneidy a, * a Department of Molecular Biology, National Research Center, Dokki, Cairo, Egypt b Department of Biochemistry, Faculty of Science, Ain-Shams University, Cairo, Egypt c Department of Research on Children with Special Needs, National Research Center, Dokki, Cairo, Egypt 1. Introduction Down syndrome (DS) is the most frequent genetic cause of mental retardation, contributing to about 30% of all moderate to severe cases of mental retardation (Rachidia & Lopes, 2008). DS is one of the most common genetic abnormalities in live born children (1 in 700–1000), leading to an early mental decline and premature aging. In DS, there is an extra copy of Research in Developmental Disabilities xxx (2011) xxx–xxx ARTICLE INFO Article history: Received 14 October 2010 Received in revised form 10 January 2011 Accepted 11 January 2011 Keywords: Down syndrome Antioxidants Oxidative stress Glutathione Glutathione S-transferase ABSTRACT Down syndrome (DS) is the phenotypic manifestation of trisomy 21. Our study was concerned with the characterization and purification of glutathione S-transferase enzyme (GST) from normal and Down syndrome (DS) erythrocytes to illustrate the difference in the role of this enzyme in the cell. Glutathione S-transferase and glutathione (GSH) was determined in ten DS and ten healthy children matched for age (3–10 years). DS group exhibited significantly lower GST value (2.7 units/g Hb) as compared to controls (6.6 units/ g Hb) (40.9%). GST activity was significantly decreased to 40.9% in the DS group as compared to controls. Also GSH concentration was significantly decreased to 60.6% in the DS group compared to the controls. Glutathione transferase was purified from erythrocytes of normal and DS pooled blood samples by affinity chromatography with specific activity of 23.7% and 7.9%, respectively. The effect of freezing and thawing, storage time of freezing and GSH concentration on the stability of the enzyme were examined. Normal GST exhibited a pH optimum at pH 7 followed by sharp decrease, however DS GST exhibited pH optimum between pH 7.5 and 8. The K m values for 1-chloro-2,4- dinitrobenzene (CDNB) and GSH were 0.205 mM and 0.786 mM, respectively, for normal GST, and 0.318 mM and 1.307 mM, respectively for DS GST. The activation energy (E a ) was calculated to be 2.25 and 4.25 cal/mol for normal GST and 3.8 cal/mol for DS GST. Normal and DS GST were inhibited by the same inhibitors (hematin, bromosulfophthalein and cibacron blue), but with different degree. On kinetic basis, the individuals with lower overall GST activity and slight differences in some kinetic characters are at greater risk from xenobiotic contamination as compared to those with higher overall GST activity observed in normal individuals. ß 2011 Elsevier Ltd. All rights reserved. * Corresponding author at: Molecular Biology Department, Genetic Engineering and Biotechnology Division, National Research Center, Tahrir St., Cairo, Egypt. Tel.: +20 2 2714033; mobile: +20 012 4322461. E-mail address: [email protected] (R.A. Guneidy). G Model RIDD-1117; No. of Pages 13 Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S- transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental Disabilities (2011), doi:10.1016/j.ridd.2011.01.013 Contents lists available at ScienceDirect Research in Developmental Disabilities 0891-4222/$ – see front matter ß 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.ridd.2011.01.013
Transcript of Homocysteine and oxidative stress in Egyptian children with Down syndrome

Research in Developmental Disabilities xxx (2011) xxx–xxx
G Model
RIDD-1117; No. of Pages 13
Contents lists available at ScienceDirect
Research in Developmental Disabilities
Purification and biochemical characterization of glutathioneS-transferase from Down syndrome and normal children erythrocytes:A comparative study
Ragaa R. Hamed a, Tahany M. Maharem b, Nagwa Abdel-Meguid c, Gilane M. Sabry b,Abdel-Monem Abdalla a, Rasha A. Guneidy a,*a Department of Molecular Biology, National Research Center, Dokki, Cairo, Egyptb Department of Biochemistry, Faculty of Science, Ain-Shams University, Cairo, Egyptc Department of Research on Children with Special Needs, National Research Center, Dokki, Cairo, Egypt
A R T I C L E I N F O
Article history:
Received 14 October 2010
Received in revised form 10 January 2011
Accepted 11 January 2011
Keywords:
Down syndrome
Antioxidants
Oxidative stress
Glutathione
Glutathione S-transferase
A B S T R A C T
Down syndrome (DS) is the phenotypic manifestation of trisomy 21. Our study was
concerned with the characterization and purification of glutathione S-transferase enzyme
(GST) from normal and Down syndrome (DS) erythrocytes to illustrate the difference in
the role of this enzyme in the cell. Glutathione S-transferase and glutathione (GSH) was
determined in ten DS and ten healthy children matched for age (3–10 years). DS group
exhibited significantly lower GST value (2.7 units/g Hb) as compared to controls (6.6 units/
g Hb) (40.9%). GST activity was significantly decreased to 40.9% in the DS group as
compared to controls. Also GSH concentration was significantly decreased to 60.6% in the
DS group compared to the controls. Glutathione transferase was purified from
erythrocytes of normal and DS pooled blood samples by affinity chromatography with
specific activity of 23.7% and 7.9%, respectively. The effect of freezing and thawing, storage
time of freezing and GSH concentration on the stability of the enzyme were examined.
Normal GST exhibited a pH optimum at pH 7 followed by sharp decrease, however DS GST
exhibited pH optimum between pH 7.5 and 8. The Km values for 1-chloro-2,4-
dinitrobenzene (CDNB) and GSH were 0.205 mM and 0.786 mM, respectively, for normal
GST, and 0.318 mM and 1.307 mM, respectively for DS GST. The activation energy (Ea) was
calculated to be 2.25 and 4.25 cal/mol for normal GST and 3.8 cal/mol for DS GST. Normal
and DS GST were inhibited by the same inhibitors (hematin, bromosulfophthalein and
cibacron blue), but with different degree. On kinetic basis, the individuals with lower
overall GST activity and slight differences in some kinetic characters are at greater risk
from xenobiotic contamination as compared to those with higher overall GST activity
observed in normal individuals.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Down syndrome (DS) is the most frequent genetic cause of mental retardation, contributing to about 30% of all moderateto severe cases of mental retardation (Rachidia & Lopes, 2008). DS is one of the most common genetic abnormalities in liveborn children (1 in 700–1000), leading to an early mental decline and premature aging. In DS, there is an extra copy of
* Corresponding author at: Molecular Biology Department, Genetic Engineering and Biotechnology Division, National Research Center, Tahrir St., Cairo,
Egypt. Tel.: +20 2 2714033; mobile: +20 012 4322461.
E-mail address: [email protected] (R.A. Guneidy).
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013
0891-4222/$ – see front matter � 2011 Elsevier Ltd. All rights reserved.
doi:10.1016/j.ridd.2011.01.013

R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx2
G Model
RIDD-1117; No. of Pages 13
chromosome 21 (Chr 21). Full trisomy of Chr 21 accounts for approximately 95% of the cases, and the remainder involvesmosaicism or translocation to Chr 14 or 21. DS can affect many aspects of development, producing a wide and variable set ofclinical features in a given person. It has been generally accepted that individuals with DS are exposed to an excessiveoxidative stress, arising from the presence of three chromosomes 21. The excessive synthesis of multiple gene productsderived from overexpression of the genes present on Chr 21 is thought to underlie both the dysmorphic features and thepathogenesis of the neurological, immunologic, endocrine and biochemical deviations that are characteristic of DS (Satge &Benard, 2008; Zana, Janka, & Kalman, 2007).
In mammalian cells, glutathione (GSH) is the major source of available nucleophilic thiol equivalents. Thiol homeostasis iscarefully maintained, and thiol:disulfide exchange reactions are important to the functional status of many proteins. Arelative deficiency of GSH may therefore predispose to diseases mediated by reactive oxygen species (ROS) and increase ROSinduced tissue damage (Hall, Ali, Pierro, & Eaton, 2005). Maintaining optimal GSH/GSSG ratios in the cell are critical tosurvival, hence, tight regulation of the system is imperative. Deficiency of GSH puts the cell at risk for oxidative damage. It isnot surprising that an imbalance of GSH is observed in a wide range of pathologies, including, cancer and neurodegenerativedisorders, cystic fibrosis (CF), HIV and aging (Meister, 1983). Catalytic conjugations of glutathione to low molecular weightacceptors frequently involve glutathione S-transferases (GSTs). It is now clear that the GST gene family is extensive, withdistinct isozymes expressing different functional properties. The family is characterized by promiscuous substratespecificity with low ‘‘catalytic efficiency’’, characteristics integral to the evolution of GSTs as detoxifiers of a broad spectrumof endogenous and environmental chemicals (Ruscoe et al., 2001).
Glutathione S-transferases (EC 2.5.1.18) are a multifunctional super family of detoxification proteins that catalyze theconjugation of glutathione to chemical toxins. They play an important role in the detoxification of electrophilic xenobiotics suchas, drugs, toxins and carcinogens allowing the products to be exported from the cell through the GS-X pump in an ATP-dependent manner. The GST enzyme system functions as an adaptive response to chemical challenge since they are induced bya variety of chemopreventive agents (Hayeshi, Mutingwende, Mavengere, Masiyanise, & Mukanganyama, 2007). The humancytosolic GST’s are dimeric proteins that can be grouped into at least seven gene-independent classes (Alpha, Mu, Pi, Sigma,Theta, Omega, and Zeta) on the basis of their amino acid sequence (Hegazy, Mannervik, & Stenberg, 2004; Parker et al., 2008).The cytosolic isoforms are differentially expressed in the different organs in the human body. It is also possible that one organexpresses more than one isoform of GST. Human erythrocytes only express the pi isoform of the enzyme. The pi class isoenzyme(GSTP1) appears the most widely distributed GST isoenzyme and is the most abundant form found in many tissues (Van-Haaften and Haenen, 2003). These GST enzymes are often over expressed in drug-resistant cell lines and particularly GSTP1 isover expressed in cancer cells, hence it is regarded as a prognostic factor in cancer treatment (Hayeshi et al., 2007). The GSTP1enzyme has been the subject of many studies because of its potential role in disease (Parker et al., 2008).
In the present study, we focused on characterization of the purified GSTP1 enzyme from erythrocytes as circulatingbioreactors, conjugating electrophiles near or in the tissues where they originate in normal and DS patients and illustratingthe role of this enzyme in protection of the cell.
2. Materials and methods
2.1. Subjects
This study was carried out on 10 DS (1.6–6.8 years) children referred to the Children’s Clinic with Special Needs, NationalResearch Centre and 10 healthy children matched for age (5–10 years). Cytogenetic analysis (comprehensive history, clinicalexamination and karotyping by G-banding technique) showed free non-disjunction trisomy 21 in the studied DS patients.
2.2. Chemicals
Reduced glutathione (GSH), bromosulfophthalein, cibacron blue, cumene hydroperoxide, epoxy-activated Sepharose-6B,ethacrynic acid and hematin, were purchased from Sigma Company. 1-Chloro-2,4-dinitrobenzene (CDNB) and 1,2-dichloro-4-nitrobenzene (DCNB) were purchased from Fluka Company. All the reagents used in electrophoresis were purchased fromPharmacia Company. All other chemicals used were of analytical grade.
2.3. Preparation of blood samples
Venous blood was collected in ethylene diaminetetraacetic acid, sodium salt (EDTA) containing tube (1 mg/1 ml blood)and centrifuged within 4 h of sampling at 1000� g for 10 min. The plasma and buffy layer were then removed; theerythrocytes were washed three times with a 9.0 g/l NaCl solution, and hemolyzed by the addition of an equal volume of icecold distilled water to yield a 50% hemolysate. The erythrocytes lysate was saved at �20 8C until later analysis.
2.4. Assay of GST activity
Unless other wise stated GST was assayed spectrophotometrically at 340 nm over 3 min at 37 8C, according to the methodof Habig, Pabst, and Jakoby (1974). All reactions were corrected for non-enzymatic conjugation, with reaction mixtures
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx 3
G Model
RIDD-1117; No. of Pages 13
without enzyme serving as controls. Under standard assay conditions, the reaction mixture contained 100 mM potassiumphosphate buffer, pH 6.5, 1.0 mM 1-chloro-2,4-dinitrobenzene (CDNB) in ethanol (final concentration of ethanol less than4%), 1.0 mM glutathione (GSH) and the enzyme solution. A unit of enzyme activity is equivalent to the amount of enzymeconjugating 1 mmol of CDNB in 1 min at 37 8C, using a molar extinction coefficient of 9.6 mM�1 cm�1 for CDNB. GST activitiestowards 1,2-dichloro-4-nitrobenzene (DCNB), bromosulfophthalein and ethacrynic acid were measured as described byHabig et al. (1974). The GST activity towards cumene hydroperoxide was determined as described by Lawrence and Burk(1976).
2.5. Determination of protein
Protein was estimated by the method of Bradford (1976) using bovine serum albumin as standard.
2.6. Determination of hemoglobin (Hb)
Hemoglobin concentration in the whole blood samples was determined spectrophotometrically according to the methoddescribed by Van-Kampen and Zijlstra (1961).
2.7. Determination of glutathione (GSH)
The total GSH was measured colorimetrically according to the method described by Saville (1958).
2.8. Purification of normal and DS erythrocytes
Preparation of erythrocytes hemolysate from pooled samples of DS and normal children previously examined for GSTactivity was performed according to the method described by Awasthi and Singh (1984). The hemolysate was dialyzedagainst 22 mM potassium phosphate buffer, pH 7.0, containing 5 mM b-mercaptoethanol (dialysis buffer) and centrifugedat 10,000� g for 1 h. The dialyzed hemolysates were mixed individually with 15 cm of GSH-Sepharose matrix previouslyequilibrated with the dialysis buffer, and allowed to proceed for 24 h at 4 8C with shaking. The matrix with GST wascollected by filtration through centered glass funnel and extensively washed with dialysis buffer containing 0.15 M NaCluntil no absorbance at 280 nm was spectrophotometrically detected. The matrix with bound proteins was packed to acolumn (15 cm� 1 cm i.d.) and the bound GST was eluted with 50 mM Tris–HCl buffer, pH 9.6 containing 10 mM GSH at aflow rate of 15 ml/h. Two-milliliter column fractions were collected and monitored for protein at 280 nm and for GSTactivity.
2.9. Polyacrylamide gel electrophoresis (PAGE)
Native PAGE (7%) was carried out according to the method described by Davis (1964). About 14 mg of the purified proteinwas loaded on to the gel. The gel was stained with 0.1% Coomassie brilliant blue R-250.
2.10. Kinetic studies
The apparent Km and Vmax values for GSH were determined at pH 6.5 using a GSH range from 0.1 to 5 mM and a fixed CDNBconcentration of 1.0 mM. The apparent Km and Vmax values for CDNB were determined using a CDNB range from 0.1 to2.5 mM and a fixed GSH concentration of 1.0 mM at pH 6.5. All assays were carried out at 37 8C.
3. Results
3.1. GST catalytic activity and GSH concentration of normal and DS
The GST activity in units/g Hb in DS group exhibited significantly lower value (2.7 units/g Hb) as compared to control(6.6 units/g Hb) (40.9%, p< 0.001) (Table 1). Also the GSH concentration was significantly decreased by 60.6% in the DS groupcompared to control group (p< 0.001).
3.2. Purification of normal and DS erythrocytes
The dialyzed normal (specific activity of 0.0015 units/mg protein) and DS (specific activity of 0.0011 units/mgprotein) hemolysates exhibited almost the same chromatographic behavior on GSH-Sepharose affinity column (Fig. 1aand b). One peak of GST bound to the column and eluted with 10 mM GSH. The specific activity increased to 23.7 units/mg protein with 114% recovery for normal GST (Table 1) compared to 7.9 units/mg protein with 82.1% recovery for DSenzyme (Table 2).
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

Table 1
The catalytic activities of GST and GSH concentrations in erythrocytes hemolysate of DS and control groups, the values are expressed as mean� SE (U/g Hb).
Group GST (U/g Hb) GSH (mmol/g Hb)
Control, n = 10 6.60� 0.69 (3.10–9.68) 3.86� 0.34 (2.10–5.30)
DS, n = 10 2.70� 0.47 (0.62–5.05) 2.34� 0.15 (1.30–3.10)
p p< 0.001 p< 0.001
% Change relative to control �59.10 �39.40
[()TD$FIG]
0
1
2
3
4
5
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 10 20 30 40
Act
ivit
y (
un
it /
ml)
Ab
sorb
an
ce a
t 280 n
m
Elution volume (ml)
Absorbance at 280 nm
Activity (unit/ml)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0 10 20 30 40
Act
ivit
y (
un
it /
ml)
Ab
sorb
an
ce a
t 280 n
m
Elution volume (ml)
Absorbance at 280 nm
Activity(unit/ml)
(b)
(a)
Fig. 1. Typical elution profile of erythrocytes GST on GSH-Sepharose affinity column chromatography of (a) normal and (b) DS. Absorbance at 280 nm (– � –)
and GST activity using CDNB (- - ~ - -).
R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx4
G Model
RIDD-1117; No. of Pages 13
3.3. Polyacrylamide gel electrophoresis
Affinity purified erythrocyte GST from normal and DS were almost pure as judged by 7% PAGE, where one band could bedetected (Fig. 2).
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

[()TD$FIG]
Fig. 2. Polyacrylamide gel electrophoresis (PAGE, 7%) of (a) normal and (b) DS, purified erythrocytes GST, stained for protein by Coomassie brilliant blue (R-
250).
Table 2
Purification of normal erythrocytes GST using GSH-Sepharose affinity chromatography.
Purification step Activity (units) Protein (mg) Specific activity (unit/mg protein) % Recovery
Hemolysate 7.28 4960 0.0015 100
Dialyzed lysate 6.97 4750 0.0015 96
Affinity purified fraction 8.30 0.35 23.7 114
R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx 5
G Model
RIDD-1117; No. of Pages 13
3.4. Stability studies
The affinity purified GST in 0.05 M Tris–HCl buffer, pH 9.6 containing 10 mM GSH lost, 42.5% and 47.5% of the activity innormal and DS, respectively by freezing and thawing for the second time (Table 3).
Samples of the purified GST were stored at �4 8C for 7, 15, 21, 28 and 60 days. The normal and DS GST activity were notaffected by freezing for 7 days. The normal GST activity was decreased to 54% by storage for 28 days at �4 8C; however, DSGST activity was decreased to 67%. By storage for 60 days, GST activity was decreased to 43% for normal and 64% for DS. Thestability studies indicated that normal GST was further affected by the freezing storage time (Table 4).
Normal purified GST containing 25 mM GSH was dialyzed for 2 h against 50 mM Tris–HCl buffer, pH 8 containing 5 mM b-mercaptoethanol. Enzyme activity in sample containing GSH was moderately affected by incubation for 1, 2 and 24 h at 4 8C,where only 21% of GST activity was lost after 24 h. However, in the absence of GSH, normal GST activity decreased to 17.9% byincreasing the incubation time for 24 h. DS GST activity was not affected by incubation for 24 h at 4 8C in the presence of GSH,however 34% of its activity was lost in the absence of GSH (Tables 5 and 6).
Table 3
Purification of DS erythrocytes GST using GSH-Sepharose affinity chromatography.
Purification step Activity (units) Protein (mg) Specific activity (unit/mg protein) % Recovery
Hemolysate 4.16 4862 0.0011 100
Dialyzed lysate 4.10 4752 0.0011 89.3
Affinity purified fraction 4.6 0.58 7.9 82.1
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

Table 4
Effect of freezing and thawing on the purified GST activity of normal and DS erythrocytes
hemolysate.
Step % Residual activity
Normal DS
Fresh 100 100
Freezing 1 87.3 80.6
Freezing 2 57.5 52.5
Table 5
Effect of storage time of freezing at �4 8C on the purified GST activity of normal and DS erythrocytes
hemolysate.
Time of freezing (days) % Residual activity
Normal DS
0 100 100
7 91 98
15 63 70
21 61 69
28 54 67
60 43 64
Table 6
Effect of GSH on the stability of GST purified from normal and DS erythrocytes.
Time (h) % Residual activity
Normal DS
(+ GSH) (� GSH) (+ GSH) (�GSH)
0 100 100 100 100
1 86.0 62.0 100 98
2 80.7 56.6 100 98
24 78.9 17.9 100 66
R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx6
G Model
RIDD-1117; No. of Pages 13
3.5. Characterization of the purified normal and DS erythrocytes GST, effect of pH
The effect of pH on the purified enzyme activity from normal and DS erythrocytes was examined between pH 4.5 and 9,using 0.1 M sodium acetate buffer for pH values 4.5 and 5.7, 0.1 M potassium phosphate buffer for pH values from pH 5.7 topH 7.5 and 0.1 M Tris–HCl buffer for pH values from pH 7.5 to pH 9. The pH profile of the normal GST exhibited a bell shapewith a pH optimum at pH 7 and a sharp decrease at the alkaline side of the pH to 20% (Fig. 3a). However the DS purifiedenzyme exhibited pH optimum between pH 7.5 and 8, this was followed by a decrease in the relative activity at pH 9 to 80%(Fig. 3a and b).
3.6. Effect of substrate concentration
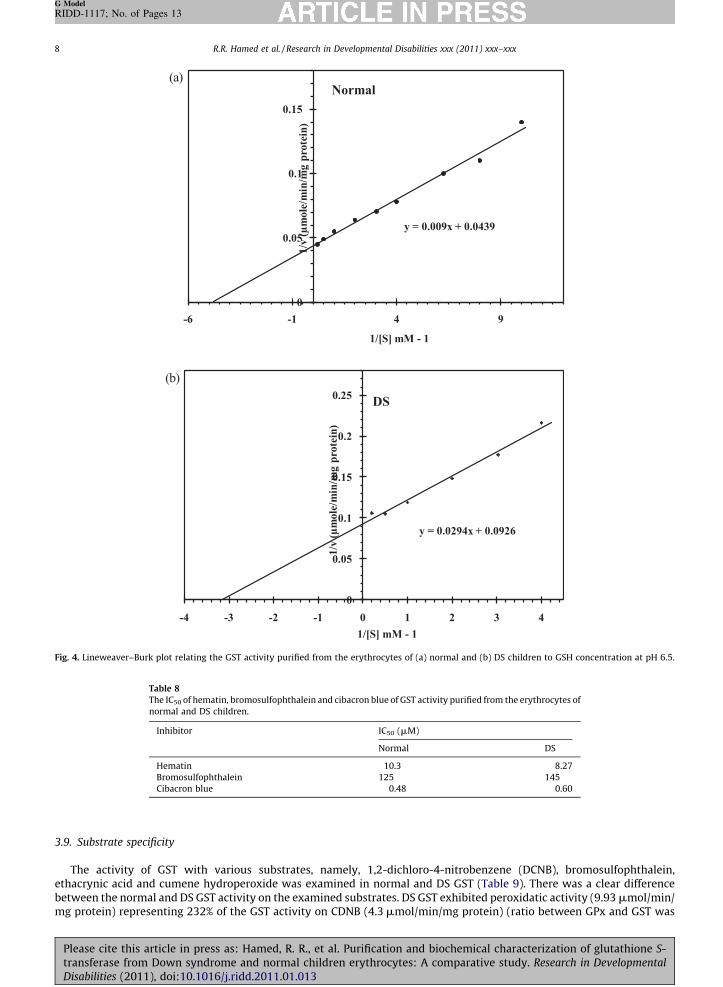
Effect of the substrates GSH and CDNB on the enzyme reaction was investigated at 37 8C at pH 6.5. The enzyme exhibited atypical Michaelian at GSH concentration ranging from 0.1 to 5 mM, i.e. a linear relationship was obtained when 1=v wasplotted against 1/[S] in both normal and DS (Fig. 4a and b). The Km value was calculated to be 0.205 mM with a Vmax equal22.78 mmol/min/mg protein, and 0.318 mM with Vmax equal 10.8 mmol/min/mg protein for normal and DS GST, respectively.In normal and DS, a linear relationship was obtained when 1=v was plotted against 1/[S], with Km equal 0.786 mM and Vmax
equal 49.75 mmol/min/mg protein at pH 6.5 for normal GST compared to the Km value (1.31 mM with Vmax equal 11.3 mmol/min/mg protein) for DS GST (Table 7).
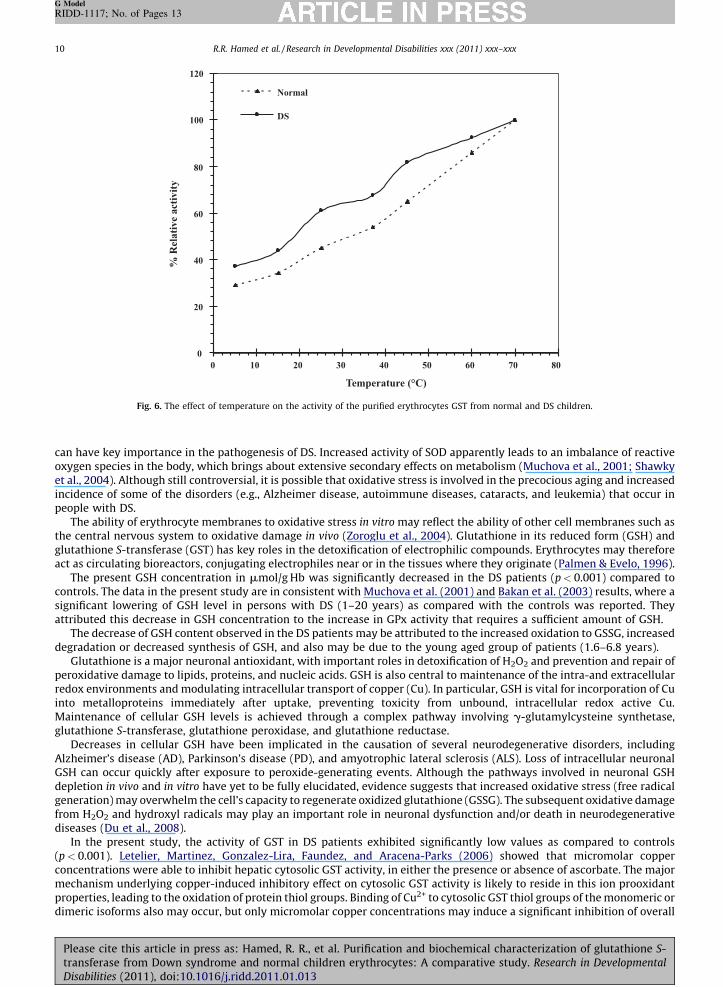
3.7. Effect of temperature
GST activity was examined at temperature ranging from 5 8C to 70 8C. The GST activity was increased by increasing thetemperature up to 70 8C in normal and DS (Figs. 5 and 6). In Arrhenius plot of DS GST, a single straight line is observed.However, the graph has a discontinuity of slope and approximates to two straight lines meeting at an angle in normal GST.
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

[()TD$FIG]
0
20
40
60
80
100
120
140
160
4 4.5 5 5.5 6 6.5 7 7.5 8 8.5 9 9.5
% R
elati
ve
act
ivit
y
pH
Normal
0
20
40
60
80
100
120
140
160
4 4.5 5 5.5 6 6.5 7 7.5 8 8.5 9 9.5
% R
elati
ve
act
ivit
y
pH
DS
(a)
(b)
Fig. 3. The effect of pH on the enzymatic activity of the purified erythrocytes GST from (a) normal and (b) DS, between pH 4.5 and 9, using 0.1 M sodium
acetate buffer for pH values 4.5 and 5.7, 0.1 M potassium phosphate buffer for pH values from pH 5.7 to pH 7.5 and 0.1 M Tris–HCl buffer for pH values from
pH 7.5 to pH 9.
Table 7
The kinetic parameters (Km and Vmax) of the purified normal and DS erythrocytes GST using the substrates GSH and CDNB at pH 6.5.
Substrates Concentration range Normal DS
Km (mM) Vmax Km (mM) Vmax
GSH 0.1–5 mM 0.205 22.78 0.318 10.8
CDNB 0.1–2.5 mM 0.786 49.75 1.307 11.3
R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx 7
G Model
RIDD-1117; No. of Pages 13
The activation energy (Ea) was calculated from the Arrhenius plot to be 2.25 and 4.25 cal/mol for normal GST and 3.8 cal/molfor DS GST (Fig. 3a and b).
3.8. Effect of inhibitors
Under the standard assay conditions, the effect of hematin, bromosulfophthalein and cibacron blue, were tested for theirability to inhibit CDNB conjugating activity of the normal and DS erythrocytes purified GST. These inhibitors were examinedin the range of 2–20, 25–200 and 0.1–4 mmol/assay, respectively. In normal GST, the IC50 were found to be 10.3, 125 and0.48 mM for hematin, bromosulfophthalein and cibacron blue, respectively. However, the IC50 were calculated to be 8.27,145 and 0.6 mM for DS GST (Table 8).
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013
[()TD$FIG]
y = 0.0294x + 0.0926
0
0.05
0.1
0.15
0.2
0.25
-4 -3 -2 -1 0 1 2 3 4
1/v
(µ
mole
/min
/mg
pro
tein
)
1/[S] mM - 1
DS
y = 0.009x + 0.0439
0
0.05
0.1
0.15
-6 -1 4 9
1/v
(µ
mole
/min
/mg
pro
tein
)
1/[S] mM - 1
Normal(a)
(b)
Fig. 4. Lineweaver–Burk plot relating the GST activity purified from the erythrocytes of (a) normal and (b) DS children to GSH concentration at pH 6.5.
Table 8
The IC50 of hematin, bromosulfophthalein and cibacron blue of GST activity purified from the erythrocytes of
normal and DS children.
Inhibitor IC50 (mM)
Normal DS
Hematin 10.3 8.27
Bromosulfophthalein 125 145
Cibacron blue 0.48 0.60
R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx8
G Model
RIDD-1117; No. of Pages 13
3.9. Substrate specificity
The activity of GST with various substrates, namely, 1,2-dichloro-4-nitrobenzene (DCNB), bromosulfophthalein,ethacrynic acid and cumene hydroperoxide was examined in normal and DS GST (Table 9). There was a clear differencebetween the normal and DS GST activity on the examined substrates. DS GST exhibited peroxidatic activity (9.93 mmol/min/mg protein) representing 232% of the GST activity on CDNB (4.3 mmol/min/mg protein) (ratio between GPx and GST was
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

[()TD$FIG]
y = 0.1155x + 0.0884
0
0.2
0.4
0.6
-2 -1 0 1 2 3 4 5
1/v
(µ
mole
/min
/mg p
rote
in)
1/[S] mM - 1
DS
y = 0.0158x + 0.0201
0
0.02
0.04
0.06
0.08
-2 -1 0 1 2 3 4 5
1/v
(µ
mole
/min
/mg p
rote
in)
1/[S] mM - 1
Normal(a)
(b)
Fig. 5. Lineweaver–Burk plot relating the GST activity purified from the erythrocytes of (a) normal and (b) DS children to CDNB concentration at pH 6.5.
Table 9
Substrate specificity of the purified GST from the erythrocytes of normal and DS children.
% Relative activity Substrate
Normal DS
1-Chloro-2,4-dinitrobenzene (CDNB) 100 100
1,2-Dichloro-4-nitrobenzene (DCNB) 2 NDa
Bromosulfophthalein (BSP) 1.65 11
Ethacrynic acid 3.35 6.10
Cumene hydroperoxide 103 232a ND: not detected.
R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx 9
G Model
RIDD-1117; No. of Pages 13
2.3:1) compared to 103% peroxidatic activity (13 mmol/min/mg protein) of normal GST activity on CDNB (12.8 mmol/min/mg protein) (GPx/GST ratio was 1:1) (Table 9).
4. Discussion
It has been generally accepted that individuals with DS are exposed to an excessive oxidative stress arising from thepresence of three chromosomes 21. The excessive genetic dose of Cu/Zn SOD gives rise to an increased SOD activity, which
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013
[()TD$FIG]
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70 80
Temperature (°C)
% R
elati
ve
act
ivit
y
Normal
DS
Fig. 6. The effect of temperature on the activity of the purified erythrocytes GST from normal and DS children.
R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx10
G Model
RIDD-1117; No. of Pages 13
can have key importance in the pathogenesis of DS. Increased activity of SOD apparently leads to an imbalance of reactiveoxygen species in the body, which brings about extensive secondary effects on metabolism (Muchova et al., 2001; Shawkyet al., 2004). Although still controversial, it is possible that oxidative stress is involved in the precocious aging and increasedincidence of some of the disorders (e.g., Alzheimer disease, autoimmune diseases, cataracts, and leukemia) that occur inpeople with DS.
The ability of erythrocyte membranes to oxidative stress in vitro may reflect the ability of other cell membranes such asthe central nervous system to oxidative damage in vivo (Zoroglu et al., 2004). Glutathione in its reduced form (GSH) andglutathione S-transferase (GST) has key roles in the detoxification of electrophilic compounds. Erythrocytes may thereforeact as circulating bioreactors, conjugating electrophiles near or in the tissues where they originate (Palmen & Evelo, 1996).
The present GSH concentration in mmol/g Hb was significantly decreased in the DS patients (p< 0.001) compared tocontrols. The data in the present study are in consistent with Muchova et al. (2001) and Bakan et al. (2003) results, where asignificant lowering of GSH level in persons with DS (1–20 years) as compared with the controls was reported. Theyattributed this decrease in GSH concentration to the increase in GPx activity that requires a sufficient amount of GSH.
The decrease of GSH content observed in the DS patients may be attributed to the increased oxidation to GSSG, increaseddegradation or decreased synthesis of GSH, and also may be due to the young aged group of patients (1.6–6.8 years).
Glutathione is a major neuronal antioxidant, with important roles in detoxification of H2O2 and prevention and repair ofperoxidative damage to lipids, proteins, and nucleic acids. GSH is also central to maintenance of the intra-and extracellularredox environments and modulating intracellular transport of copper (Cu). In particular, GSH is vital for incorporation of Cuinto metalloproteins immediately after uptake, preventing toxicity from unbound, intracellular redox active Cu.Maintenance of cellular GSH levels is achieved through a complex pathway involving g-glutamylcysteine synthetase,glutathione S-transferase, glutathione peroxidase, and glutathione reductase.
Decreases in cellular GSH have been implicated in the causation of several neurodegenerative disorders, includingAlzheimer’s disease (AD), Parkinson’s disease (PD), and amyotrophic lateral sclerosis (ALS). Loss of intracellular neuronalGSH can occur quickly after exposure to peroxide-generating events. Although the pathways involved in neuronal GSHdepletion in vivo and in vitro have yet to be fully elucidated, evidence suggests that increased oxidative stress (free radicalgeneration) may overwhelm the cell’s capacity to regenerate oxidized glutathione (GSSG). The subsequent oxidative damagefrom H2O2 and hydroxyl radicals may play an important role in neuronal dysfunction and/or death in neurodegenerativediseases (Du et al., 2008).
In the present study, the activity of GST in DS patients exhibited significantly low values as compared to controls(p< 0.001). Letelier, Martinez, Gonzalez-Lira, Faundez, and Aracena-Parks (2006) showed that micromolar copperconcentrations were able to inhibit hepatic cytosolic GST activity, in either the presence or absence of ascorbate. The majormechanism underlying copper-induced inhibitory effect on cytosolic GST activity is likely to reside in this ion prooxidantproperties, leading to the oxidation of protein thiol groups. Binding of Cu2+ to cytosolic GST thiol groups of the monomeric ordimeric isoforms also may occur, but only micromolar copper concentrations may induce a significant inhibition of overall
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx 11
G Model
RIDD-1117; No. of Pages 13
cytosolic GST activity (Letelier et al., 2006). Neve, Vertongen, Cauchie, Gnat, and Molle (1984) reported that Cu level inplasma is normal while an elevated level was observed in serum and erythrocytes in DS patients.
The observed significant decrease in GST activity in the DS patients may be due to: (1) young aged group of our patients(1.6–6.8 years), (2) increased generation of reactive oxygen radicals, (3) imbalance in the antioxidant defense system, (4)decreased levels of GSH and inactivation of enzymes and (5) inhibition by copper.
Comparison between DS and normal GST regarding its physical, chemical, kinetic and stability of the erythrocytes GSTmay through more light on its role.
A simple reproducible procedure for the purification of GST from normal and DS erythrocytes using affinitychromatography on GSH-Sepharose column was carried out. The specific activity increased to 23.7 units/mg protein with114% recovery for normal children, compared to 7.9 units/mg protein with 82.1% recovery for DS children. The affinitypurified enzyme from normal and DS erythrocytes hemolysates were found to be almost pure as judged by PAGE.
Instability of normal and DS GST was observed after freezing and thawing and storage time of freezing. This observedinstability of the enzyme may be due to the very low concentration of protein observed in the affinity purified fraction and/ordegradation of GSH that may act as reducing agent and as a substrate stabilizing the enzyme structure. Letelier et al. (2006)reported that binding of GSH to the GST active site seems to protect GST activity. The difference between normal and DS GSTwith respect to stability may be explained by the difference in the accessibility of the GST thiol groups to GSH. GSH may besaturating GST molecules under physiological conditions, protecting its activity. If the enzyme exists in an active –SH formand an inactive/less active –S–S– form arising due to intra- or inter-subunit disulfide bonds, the active (–SH) and the inactive(–S–S–) forms may be in equilibrium and in vivo this equilibrium may be regulated by GSH (Singh & Awashti, 1986). Thedecrease in GSH concentration in DS patients may affect this equilibrium. On the other hand, it has been reported that rat GSTp form is inactivated by H2O2. This result suggested that cysteine 47 and cysteine 101 may be involved in the formation ofintra- or inter-subunit disulfides and these residues may be located in an important region for GSH binding, disulfide bondformation between these residues resulting in steric hindrance. Similar results were observed with human placenta GST pform and treatment with 2,4-dithiotreitol (DTT) reversed this oxidative inactivation (Letelier et al., 2006).
The optimum pH values for GST with a variety of different substrates ranged from 6.0 to 9.5. When CDNB is considered,narrower range of pH 7.0–9.0 is obtained, but the most is in the vicinity of pH 8.0 (Clark, 1989). The pH optimum for normaladult erythrocyte GST was between 6.8 and 6.9 (Galli et al., 1999). A shift in pH optimum from pH 7.0 for normal erythrocyteGST to 7.5–8 for DS was observed. Also DS GST retained 80% of its activity at pH 9.0. This may suggest the higher stability ofthe DS enzyme in the alkaline conditions compared to normal. The slightly different pH profiles of normal and DS GSTsuggested that, pH will adversely affect substrate binding and thus may retard catalysis.
Kinetic studies with purified GST show high differences in the specific activity for a same substrate as also for differentsubstrates, Km values for CDNB can differ 100 or more fold depending on the GST isoform. Likewise, the GST isoforms Km
values for GSH described seem to be lower than 1 mM. One has to note that comparative values of these kinetic constants aredifficult to obtain because the assay conditions used by the investigators are not the same (Letelier et al., 2006).
The present investigation is the first report to our knowledge on purification and kinetic properties of erythrocyte DS GST.The affinity of the normal and DS enzyme towards GSH are almost different, normal and DS GST displayed a distinctMichaelis–Menten constant, a linear double reciprocal plot with an observed increase in the Km values for DS GST withrespect to GSH and CDNB as substrates at pH 6.5.
The present Km (GSH) values invite comparison with those carried out by other workers on the human erythrocyteenzyme purified from adults, 0.24 mM (Galli et al., 1999) and higher than 0.12 mM (Thumser & Ivanetich, 1993).However, the present type of Km was observed in Hirrell, Collins, Nimmo, and Strange (1987) study where Km values forGSH were 0.14 in the lower range between 0.01 and 0.1 mM GSH, and 1.02 in the higher range between 1 and 3 mM GSHat pH 6.5. The present Km (CDNB) values for normal GST were 0.78 mM compared to the purified erythrocyte enzyme,0.7 mM (Thumser & Ivanetich, 1993), 0.6 mM (Awasthi & Singh, 1984) and 0.5 mM (Galli et al., 1999). A comparison oferythrocyte GST activity from human fetus and adult carried out by (Strange, Johnston, Coghill, & Hume, 1980), Km
(GSH) was 0.14 and 0.1 mM for adult and fetal, respectively. While Km values for CDNB were 0.75 mM and 1 mM foradult and fetal, respectively.
Biochemical and structural investigations revealed that the a2 loop (The a2 loop form one wall of the G-site and has anumber of amino acid residues) is highly flexible in the absence of GSH and is stabilized by GSH binding to the active site.Therefore, perturbation of the conformational structure of the a2 loop may be responsible for increasing the Km value. Thusany mutation, which disturbs the conformational flexibility of helix a2 and possibly changes the G-site geometry, may beexpected to lead to increasing Km GSH values (54-fold increase observed at 30 8C) and a decrease in GSH affinity (Hegazyet al., 2004).
Raising the temperature increases the rate of both uncatalyzed and enzyme-catalyzed reactions by increasing the kineticenergy and the collision frequency of the reacting molecules. However, heat energy can also increase the kinetic energy ofthe enzyme to a point that exceeds the energy barrier for disrupting the non-covalent interactions that maintain theenzymes three-dimensional structure (Rodwell & Kennelly, 2003). The polypeptide chain then begins to unfold, or denature,with an accompanying rapid loss of catalytic activity. DS GST is more active since it exhibited lower activation energy thannormal GST. A maximum activity of both fetal and adult GST was occurred at 45 8C and data give linear plots between 11 8Cand 36 8C, the energies of activation of the reaction were 37–42.7 and 44.7 kJ mol�1 for fetal and adult GST, respectively(Strange et al., 1980).
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx12
G Model
RIDD-1117; No. of Pages 13
Enzymes from human generally exhibit stability at temperatures up to 45–55 8C, by contrast, enzymes from thethermophilic microorganisms that reside in volcanic hot springs or undersea hydrothermal vents may be stable up to orabove 100 8C (Rodwell & Kennelly, 2003). Equilibrium unfolding studies with p GSTP1 indicate that there is a tight couplingbetween dissociation and association and unfolding/refolding for that protein. These findings suggest that dimericquaternary structure is essential and that interactions at the dimmer interface are the main contributions to protein foldingand stability. However, all these unfolding/folding studies were performed using chemical denaturants as urea andguanidinium chloride while thermal denaturations mechanisms for GSTs have not been sufficiently examined to date(Quesada-Soriano, Garcia-Maroto, & Garcia-Fuentes, 2006). The efficiency of the enzyme for catalyzing a particular chemicalreaction relies on the manner of the binding of the substrates to the active site and the dynamic structure of the enzyme.Enzymes accelerate the rate of the chemical reaction by reducing the activation energy, which is the difference in the freeenergy between the substrates in the ground state and the transition state for the reaction. The activation energy of thereaction can be reduced either by stabilization of the transition state, or by finding some other lower energy pathway for thereaction (Gutteridge and Thornton, 2005).
GSTP1 as a whole somewhat behaves as a thermodynamic system. In fact, whenever a physical factor like temperature orviscosity forces the G-site to assume a low affinity conformation for GSH binding, GSTP1 opposes this perturbation bydeveloping positive cooperativity and vice versa (Hegazy et al., 2004).
Przybylska, Bryszewska, and Kedziora (2000) experimental data suggest that trisomy of 21 chromosomes significantlyaffects thermostability of red blood cells, although the mechanisms of lowered thermosensitivity of erythrocytes frompersons suffering from DS still remain unclear. DS red blood cells were more resistant, to a certain extent, to disruptioninduced by heat which was indicated by the considerably reduced fraction of altered cells, compared to normal ones,especially for longer incubation time (>1 h). At each temperature from 48 8C to 54 8C, the fractions of hemoglobin retained byred blood cells from DS patients were greater than for erythrocytes of healthy individuals. The hemolysis sensitivity ratio(HSR) showed that DS erythrocytes were, in average, 1.6 times more resistant to heat injury than those from normal subjects(Przybylska et al., 2000).
Normal and DS GST were inhibited by the same inhibitors; however DS GST was inhibited with different degree. The effectof various compounds on GSTP1 activity was reported in a study by Mannervik and Widersten (1995) where the inhibitionconcentrations required for 50% inhibition were 5, 100 and 0.5 mM for hematin, BSP and cibacron blue, respectively.Erythrocyte and hepatic forms of GST have been shown to bind hematin with variable affinities. Harvey and Beutler (1982)suggested that human erythrocyte GST activity was inhibited, probably competitively, by hemin with a Ki of 10�7 mol/l. Itwas postulated that GST functions physiologically as a hemin-binding and/or transport protein in developing erythrocytecells. The IC50 for BSP reported by Hirrell et al. (1987) was 100.4� 2.6 mmol/l, while in Awasthi and Singh (1984) study, themajor erythrocytes GST, is not inhibited by BSP. DS GST not acts at all on DCNB while 11% and 6.1% relative activity were observedfor BSP and ethacrynic acid. Unexpected substrate specificity was observed against cumene hydroperoxide which was notdetected in other studies (Awasthi & Singh, 1984; Guthenberg & Mannervik, 1981). Using (DCNB) as a substrate, 0.067 mmol/min/mg protein in normal GST was observed compared to 0.025 mmol/min/mg protein reported in (Guthenberg & Mannervik, 1981)study on adults. While, 0.056 and 0.99 mmol/min/mg protein of BSP were observed in normal and DS children, adult GST purifiedfrom erythrocytes had no activity on this substrate. Using ethacrynic acid as a substrate, 0.11 and 0.55 mmol/min/mg protein wereobserved in normal and DS GST, respectively compared to 2.9 and 8.84 mmol/min/mg protein reported in Guthenberg andMannervik (1981) and Awasthi and Singh (1984) studies on adults. The reported specific activities of human GSTP1 with CDNB,DCNB, ethacrynic acid and cumene hydroperoxide were 105, 0.1, 0.9 and 0.03 mmol/min/mg protein, respectively (Mannervik &Widersten, 1995). It has been reported that H2O2 leads to the formation of intra- or inter-subunit disulfide bonds betweenparticular cysteine residues within GST amino acid sequence, leading to the inactivation of the cytosolic enzymes. The effect ofother reactive oxygen species on GST activity, however, has not been thoroughly investigated (Letelier et al., 2006).
To conclude, on kinetic basis, DS GST differs from normal GST regarding Km, pH, activation energy, substrate specificityand the degree of inhibition suggesting mutation. Also, with regard to stability, the individuals with lower overall GSTactivity and slightly different in some kinetic characteristics may be at greater risk from xenobiotic contamination ascompared to those with higher overall GST activity observed in normal individuals. The determination of erythrocyte GSTactivity is a promising indicator of oxidative stress conditions that occur in DS individuals. Measurement of GST activitymight be useful for the evaluation of prophylactic treatment in trials of antioxidant strategies. It is important to understandthe differences and similarities in the abilities of a species to undergo electrophilic detoxification in order to accuratelyquantify risk in different habitats.
References
Awasthi, Y. C., & Singh, S. V. (1984). Purification and characterization of a new form of glutathione S-transferase from human erythrocytes. Biochemical andBiophysical Research Communications, 125(3), 1053–1060.
Bakan, N., Taysi, S., Yilmaz, O., Bakan, E., Kuskay, S., Uzun, N., et al. (2003). Glutathione peroxidase, glutathione reductase, Cu–Zn superoxide dismutase activities,glutathione, nitric oxide, and malondialdehyde concentrations in serum of patients with chronic lymphocytic leukemia. Clinica Chimica Acta, 338, 143–149.
Bradford, M. M. (1976). A rapid and sensitive method for quantization of microgram quantities of protein utilizing the principle of protein-dye-binding. AnalyticalBiochemistry, 72, 248–254.
Clark, A. G. (1989). The comparative enzymology of the glutathione S-transferases from non-vertebrate organisms. Comparative Biochemistry and Physiology, 92B,419–446.
Davis, B. Y. (1964). Disc electrophoresis: Method and application to human serum protein. Annals of the New York Academy of Sciences, 121, 404–427.
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013

R.R. Hamed et al. / Research in Developmental Disabilities xxx (2011) xxx–xxx 13
G Model
RIDD-1117; No. of Pages 13
Du, T., Ciccotosto, G. D., Cranston, G. A., Kocak, G., Masters, C. L., Crouch, P. J., et al. (2008). Neurotoxicity from glutathione depletion is mediated by Cu-dependentp53 activation. Free Radical Biology & Medicine, 44, 44–55.
Galli, F., Rovidati, S., Benedetti, S., Buoncristiani, U., Covarelli, C., Floridi, A., et al. (1999). Overexpression of erythrocyte glutathione S-transferase in uremia anddialysis. Clinical Chemistry, 45, 1781–1788.
Guthenberg, C., & Mannervik, B. (1981). Glutathione S-transferase (transferase pi) from human placenta is identical or closely related to glutathione S-transferase(transferase rho) from erythrocytes. Biochimica et Biophysica Acta, 661, 255–260.
Gutteridge, A., & Thornton, J. M. (2005). Understanding nature’s catalytic toolkit. Trends in Biochemical Sciences, 30, 622–629.Habig, W. H., Pabst, M. J., & Jakoby, W. B. (1974). Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. Journal of Biological Chemistry,
249, 7130–7139.Hall, N. J., Ali, J., Pierro, A., & Eaton, S. (2005). Total glutathione is not decreased in infants with necrotizing enterocolitis. Journal of Pediatric Surgery, 40, 769–773.Harvey, J. W., & Beutler, E. (1982). Binding of heme by glutathione S-transferase: A possible role of the erythrocyte enzyme. Blood, 60, 1227–1230.Hayeshi, R., Mutingwende, I., Mavengere, W., Masiyanise, V., & Mukanganyama, S. (2007). The inhibition of human glutathione S-transferases activity by plant
polyphenolic compounds ellagic acid and curcumin. Food and Chemical Toxicology, 45, 286–295.Hegazy, U. M., Mannervik, B., & Stenberg, G. (2004). Functional role of the lock and key motif at the subunit interface of glutathione S-transferase P1-1. Journal of
Biological Chemistry, 279(10), 9586–9596.Hirrell, P. A., Collins, M. F., Nimmo, I. A., & Strange, R. C. (1987). The human glutathione S-transferases, studies on the kinetic, stability and inhibition characteristics
of the erythrocyte enzyme. Biochimica et Biophysica Acta, 913, 92–96.Lawrence, R. A., & Burk, R. F. (1976). Glutathione peroxidase activity in selenium-deficient rat liver. Biochemical and Biophysical Research Communications, 71(4),
952–958.Letelier, M. E., Martinez, M., Gonzalez-Lira, V., Faundez, M., & Aracena-Parks, P. (2006). Inhibition of cytosolic glutathione S-transferase activity from rat liver by
copper. Chemico-Biological Interactions, 164, 39–48.Mannervik, B., & Widersten, M. (1995). Human glutathione transferases: Classification, tissue distribution and functional properties. In G. M. Pacifici & G. N.
Fracchia (Eds.), Advances in drug metabolism in man (pp. 408–459). Luxembourg: European Commission.Meister, M. A. A. (1983). Glutathione. Annual Review of Biochemistry, 52, 711–760.Muchova, J., Sustrova, M., Garaiova, I., Liptakova, A., Blazicek, P., & Kvasnicka, et al. (2001). Influence of age on activities of antioxidant enzymes and lipid
peroxidation products in erythrocytes and neutrophils of Down syndrome patients. Free Radical Biology and Medicine, 31(4), 499–508.Neve, J., Vertongen, F., Cauchie, P., Gnat, D., & Molle, L. (1984). Selenium and glutathione peroxidase in plasma and erythrocytes of Down syndrome (trisomy 21)
patients. Journal of Mental Deficiency Research, 28(4), 261–268.Palmen, N. G. M., & Evelo, C. T. A. (1996). Glutathione depletion in human erythrocytes and rat liver: A study on the interplay between bioactivation and
inactivation functions of liver and blood. Toxicology In Vitro, 10, 273–281.Parker, L. J., Ciccone, S., Louis, C., Italiano, L. C., Primavera, A., Oakley, A. J., et al. (2008). The anti-cancer drug chlorambucil as a substrate for the human
polymorphic enzyme glutathione transferase P1-1: Kinetic properties and crystallographic characterisation of allelic variants. Journal of Molecular Biology,380, 131–144.
Przybylska, M., Bryszewska, M., & Kedziora, J. (2000). Thermosensitivity of red blood cells from Down’s syndrome individuals. Bioelectrochemistry, 52(2), 239–249.Quesada-Soriano, I., Garcia-Maroto, F., & Garcia-Fuentes, L. (2006). Kinetic study on the irreversible thermal denaturation of Schistosoma japonicum glutathione S-
transferase. Biochimica et Biophysica Acta, 1764, 979–984.Rachidia, M., & Lopes, C. (2008). Mental retardation and associated neurological dysfunctions in Down syndrome: A consequence of dysregulation in critical
chromosome 21 genes and associated molecular pathways. The European Journal of Paediatric Neurology, 12, 168–182.Rodwell, & Kennelly, (2003). Enzymes: Kinetics. In R. K. Murray, D. K. Granner, P. A. Mayes, & V. W. Rodwell (Eds.), Harper’s illustrated biochemistry (twenty-sixth
edition, pp. 60–71). London: McGraw-Hill Companies, A LANGE Medical Book.Ruscoe, J. E., Rosario, L. A., Wang, T., Gate, L., Arifoglu, P., Roland wolf, C., et al. (2001). Pharmacologic or genetic manipulation of glutathione S-transferase p1-1
(GSTp) influences cell proliferation pathways. Journal of Pharmacology and Experimental Therapeutics, 298(1), 339–345.Satge, D., & Benard, J. (2008). Carcinogenesis in Down syndrome, what can be learned from trisomy 21? Seminars in Cancer Biology, 18(5), 365–371.Saville, B. (1958). Colorimetric method for total thiol determination. Analyst, 83, 670–672.Shawky, R. M., Abdelaziz, E. A., Elhossiny, R. M., & Ahmed, M. E. (2004). Glutathione peroxidase enzyme and selenium level in patients with Down syndrome. The
Egyptian Journal of Medical Human Genetics, 5(1), 103–109.Singh, S. V., & Awashti, Y. C. (1986). Cationic glutathione S-transferase of human erythrocytes has unique kinetic characteristics among human glutathione S-
transferases. Biochemical and Biophysical Research Communications, 137(93), 1174–1180.Strange, R. C., Johnston, J. D., Coghill, D. R., & Hume, R. (1980). A comparison of erythrocyte glutathione S-transferase activity from human fetuses and adults.
Biochemical Journal, 188, 475–479.Thumser, A. E. A., & Ivanetich, K. M. (1993). Kinetic mechanism of human erythrocyte acidic isoenzyme p. Biomedica Biochimica Acta, 1203, 115–120.Van-Haaften, R. I. M., Haenen, G. R. M. M. , van Bladeren, P. J., Bogaards, J. J. P., Evelo, C. T. A., & Aalt, B. (2003). Inhibition of various glutathione S-transferase
isoenzymes by RRR-a-tocopherol. Toxicology In Vitro, 17, 245–251.Van-Kampen, E., & Zijlstra, W. G. (1961). Standardization of hemoglobinometry. II. The hemiglobincyanide method. Clinica Chimica Acta, 6, 538–544.Zana, M., Janka, Z., & Kalman, J. (2007). Oxidative stress: A bridge between Down’s syndrome and Alzheimer’s disease. Neurobiology of Aging, 28, 648–676.Zoroglu, S. S., Armutcu, F., Ozen, S., Gurel, A., Sivasli, E., Yetkin, O., et al. (2004). Increased oxidative stress and altered activities of erythrocyte free radical
scavenging enzymes in autism. European Archives of Psychiatry and Clinical Neuroscience, 254, 143–147.
Please cite this article in press as: Hamed, R. R., et al. Purification and biochemical characterization of glutathione S-transferase from Down syndrome and normal children erythrocytes: A comparative study. Research in Developmental
Disabilities (2011), doi:10.1016/j.ridd.2011.01.013