
Histopathological and ultrastructural observations of metacercarial infections of Diplostomum...
9
DISEASES OF AQUATIC ORGANISMS Dis Aquat Org Vol. 75: 51–59, 2007 Published March 29 INTRODUCTION Members of the strigeid digenean genus Diplosto- mum Nordmann, 1832 and the closely related genus Tylodelphys Diesing, 1850 are important parasitic pathogens of farmed and wild stocks of fish, and infec- tions are known to induce blindness, emaciation, cra- nial deformation, poor growth and, in some species, host mortality (Shariff et al. 1980, Chappell 1995, Pylkkö et al. 2006). A parasite’s affinity or specificity for particular tis- sue/organs sites may result from a number of factors that may be driven by the specific nutritional and/or environmental requirements of the parasite (Crompton 1976), the consequences of inter-specific competition for resources and subsequent niche separation be- tween species (Holmes & Price 1985), or by attempts to evade the host’s chemical and cellular immune defences (Price 1987, Sukhdeho & Mettrick 1987, Sukhdeho & Sukhdeho 1994). The current study on a population of Diplostomum phoxini (Faust, 1989)-infected Phoxinus phoxinus (L.) in the River Endrick, Scotland, found metacercariae principally within the cerebellum, the medulla oblon- © Inter-Research 2007 · www.int-res.com *Email: [email protected] Histopathological and ultrastructural observations of metacercarial infections of Diplostomum phoxini (Digenea) in the brain of minnows Phoxinus phoxinus Bahram Sayyaf Dezfuli 1, *, Silvia Capuano 1 , Edi Simoni 1 , Luisa Giari 1 , Andrew Paul Shinn 2 1 Department of Biology, University of Ferrara, Via Borsari, 46, 44100 Ferrara, Italy 2 Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, UK ABSTRACT: The spatial distribution and histopathological changes induced by metacercariae of the digenean trematode Diplostomum phoxini (Faust, 1918) in the brains of European minnows Phoxinus phoxinus (L.) from the River Endrick, Scotland, were studied by light and electron microscopy. Post- mortem examination of a sample of 34 minnows revealed that 50% (n = 17) of the population was infected with 13.7 ± 2.6 (mean ± SE; range 1 to 38) metacercariae per infected host. Serial histologi- cal sections of the infected minnow brains revealed that the metacercariae were unevenly distributed throughout the brain, with aggregations occurring in the cerebellum, the medulla oblongata and the optic lobes. In fish with highest intensities of infection, over 40% of the cerebellar area and about 30% of the medulla oblongata area were occupied by larvae. Metacercariae disrupt the integrity of brain tissue, with individuals being found in small pockets surrounded by cellular debris. Metacer- cariae were rarely encountered on the surface of the brain. Electron microscopic examination of infection sites revealed that the granular layer surrounding metacercariae was necrotic, exhibited nuclear degradation and was marked by vacuolation of the cytoplasm. Rodlet cells, the only inflam- matory cell types recorded in this study, were found only in parasitized brains and in close proximity to the teguments of metacercariae. It is hypothesised that secretions released from the teguments of metacercariae are a counter response to protect the metacercariae from the fish brain’s cellular defence mechanisms. KEY WORDS: Brain infection · Digenean trematode · Phoxinus phoxinus · Rodlet cells · Histopathology · Ultrastructure Resale or republication not permitted without written consent of the publisher
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Histopathological and ultrastructural observations of metacercarial infections of Diplostomum...

DISEASES OF AQUATIC ORGANISMSDis Aquat Org
Vol. 75: 51–59, 2007 Published March 29
INTRODUCTION
Members of the strigeid digenean genus Diplosto-mum Nordmann, 1832 and the closely related genusTylodelphys Diesing, 1850 are important parasiticpathogens of farmed and wild stocks of fish, and infec-tions are known to induce blindness, emaciation, cra-nial deformation, poor growth and, in some species,host mortality (Shariff et al. 1980, Chappell 1995,Pylkkö et al. 2006).
A parasite’s affinity or specificity for particular tis-sue/organs sites may result from a number of factors
that may be driven by the specific nutritional and/orenvironmental requirements of the parasite (Crompton1976), the consequences of inter-specific competitionfor resources and subsequent niche separation be-tween species (Holmes & Price 1985), or by attempts toevade the host’s chemical and cellular immunedefences (Price 1987, Sukhdeho & Mettrick 1987,Sukhdeho & Sukhdeho 1994).
The current study on a population of Diplostomumphoxini (Faust, 1989)-infected Phoxinus phoxinus (L.)in the River Endrick, Scotland, found metacercariaeprincipally within the cerebellum, the medulla oblon-
© Inter-Research 2007 · www.int-res.com*Email: [email protected]
Histopathological and ultrastructural observationsof metacercarial infections of Diplostomum phoxini
(Digenea) in the brain of minnowsPhoxinus phoxinus
Bahram Sayyaf Dezfuli1,*, Silvia Capuano1, Edi Simoni1, Luisa Giari 1, Andrew Paul Shinn2
1Department of Biology, University of Ferrara, Via Borsari, 46, 44100 Ferrara, Italy2Institute of Aquaculture, University of Stirling, Stirling FK9 4LA, UK
ABSTRACT: The spatial distribution and histopathological changes induced by metacercariae of thedigenean trematode Diplostomum phoxini (Faust, 1918) in the brains of European minnows Phoxinusphoxinus (L.) from the River Endrick, Scotland, were studied by light and electron microscopy. Post-mortem examination of a sample of 34 minnows revealed that 50% (n = 17) of the population wasinfected with 13.7 ± 2.6 (mean ± SE; range 1 to 38) metacercariae per infected host. Serial histologi-cal sections of the infected minnow brains revealed that the metacercariae were unevenly distributedthroughout the brain, with aggregations occurring in the cerebellum, the medulla oblongata and theoptic lobes. In fish with highest intensities of infection, over 40% of the cerebellar area and about30% of the medulla oblongata area were occupied by larvae. Metacercariae disrupt the integrity ofbrain tissue, with individuals being found in small pockets surrounded by cellular debris. Metacer-cariae were rarely encountered on the surface of the brain. Electron microscopic examination ofinfection sites revealed that the granular layer surrounding metacercariae was necrotic, exhibitednuclear degradation and was marked by vacuolation of the cytoplasm. Rodlet cells, the only inflam-matory cell types recorded in this study, were found only in parasitized brains and in close proximityto the teguments of metacercariae. It is hypothesised that secretions released from the teguments ofmetacercariae are a counter response to protect the metacercariae from the fish brain’s cellulardefence mechanisms.
KEY WORDS: Brain infection · Digenean trematode · Phoxinus phoxinus · Rodlet cells ·Histopathology · Ultrastructure
Resale or republication not permitted without written consent of the publisher

Dis Aquat Org 75: 51–59, 2007
gata and the optic lobes, centres of the brain that areresponsible for host motor activity, sensory functionsand vision (Barber et al. 2000, Shirakashi & Goater2005). Until recently, metacercariae were generallyregarded as immature, non-feeding and metabolicallyinactive stages waiting for transmission to the next/definitive host (Bush et al. 2001, Poulin & Latham2003). However, a recent study by Goater et al. (2005)demonstrated via experimental infection that themetacercariae of Ornithodiplostomum ptychocheilusFaust, 1917 in Pimephales promelas Rafinesque, 1820undergo a series of complex developmental changesassociated with their feeding on host brain tissue.
A histological examination of Phoxinus phoxinusheads in the current study revealed the presence ofrodlet cells only in infected brains, notably in theepithelium lining the ventricles of the optic lobes. It isbelieved that rodlet cells play a role within the piscineinflammatory response (Dezfuli et al. 2000, Manera &Dezfuli 2004, Reite 2005, Reite & Evensen 2006). Wefound rodlet cells only within the infected brain of afish, and our findings are briefly compared with theinformation available on their presence in other fish-parasite systems (Dezfuli et al. 1998, 2000, 2002, 2003,Reite 1998, Bosi et al. 2005). While there have beennumerous ultrastructural studies on metacercariae(Stein & Lumsden 1971, Strong & Cable 1972, So &Wittrock 1982, Faliex & Biagianti 1987, Wittrock et al.1991, Goater et al. 2005), most focused on the develop-ment of the larval epithelial layer within the intermedi-ate host, and only a few of these studies focused on theinteractions between host tissues and the parasite(Baghaei 1981, So & Wittrock 1982, Faliex & Biagianti1987, Wittrock et al. 1991). Fewer again used transmis-sion electron microscopy (TEM) to examine the cysts ofstrigeoid digeneans (Bibby & Rees 1971, Mitchell 1974,Baghaei 1981, Goater et al. 2005).
The present study provides information on (1) theoccurrence of Diplostomum phoxini metacercariae inthe brains of minnows Phoxinus phoxinus, with partic-ular comments on the interface region between thehost and the parasite; (2) light and electron micro-scopic observations on histopathological changes inbrain tissue; (3) the location of metacercariae, the dam-age they cause in key regions of the brain and theireffect on host vision and motor activity; and (4) therelationship between brain cells, inflammatory cellsand D. phoxini metacercariae.
MATERIALS AND METHODS
Thirty-four minnows Phonixus phoxinus, were hand-netted from the River Endrick, Stirlingshire, Scotland(56° 03’ N, 4° 27’ W) and transferred to the laboratory in
river water where they were maintained in aeratedaquaria until being processed. Fish were subsequentlysacrificed by exposure to a lethal dose of the anaes-thetic MS-222 (Sandoz), and were then weighed (meanweight ± SE: 0.21 ± 0.01 g), measured (mean totallength ± SE: 30.57 ± 0.55 mm) and processed for histol-ogy. After severing the spinal cord, 28 heads wereremoved and fixed in chilled (4°C) Bouin’s fluid for 7 hbefore being transferred into chilled 70% alcohol forstorage until being processed for paraffin embeddingby routine methods using an automatic tissue proces-sor (Shandon Citadel 2000 Tissue Processor, ShandonSouthern Products). Histological sections (5 µm thick)were cut and stained with a haematoxylin and eosin(H&E), Azan-Mallory, periodic acid Schiff (PAS) or analcian blue/PAS stain. Light photomicrographs weretaken using a Nikon microscope Eclipse 80i. Thedimensions of Diplostomum phoxini key features in theacquired images were subsequently determined usingcomputerised image analysis software (Lucia G 4.8,Laboratory Imaging). For the TEM study, 6 P. phoxinusheads were fixed for 2 h in a chilled (4°C) 2% glu-taraldehyde solution buffered at pH 7.2 with 0.1 Msodium cacodylate, and were then rinsed in 0.1 Msodium cacodylate buffer containing 6% sucrose forapproximately 12 h. The minnow heads were thenpost-fixed in 1% osmium tetroxide in the same bufferfor 2 h, dehydrated through a graded ethanol series,transferred to propylene oxide and then embedded inan Epoxy-Araldite mixture. Semi-thin sections (1.5 µm)were cut on a Reichert Om U2 ultramicrotome usingglass knives and stained with toluidine blue. Ultra-thinsections (90 nm) were contrasted in a 50% alcohol-uranyl acetate solution and lead citrate, and examinedusing a Hitachi H-800 electron microscope operated at80 kV.
RESULTS
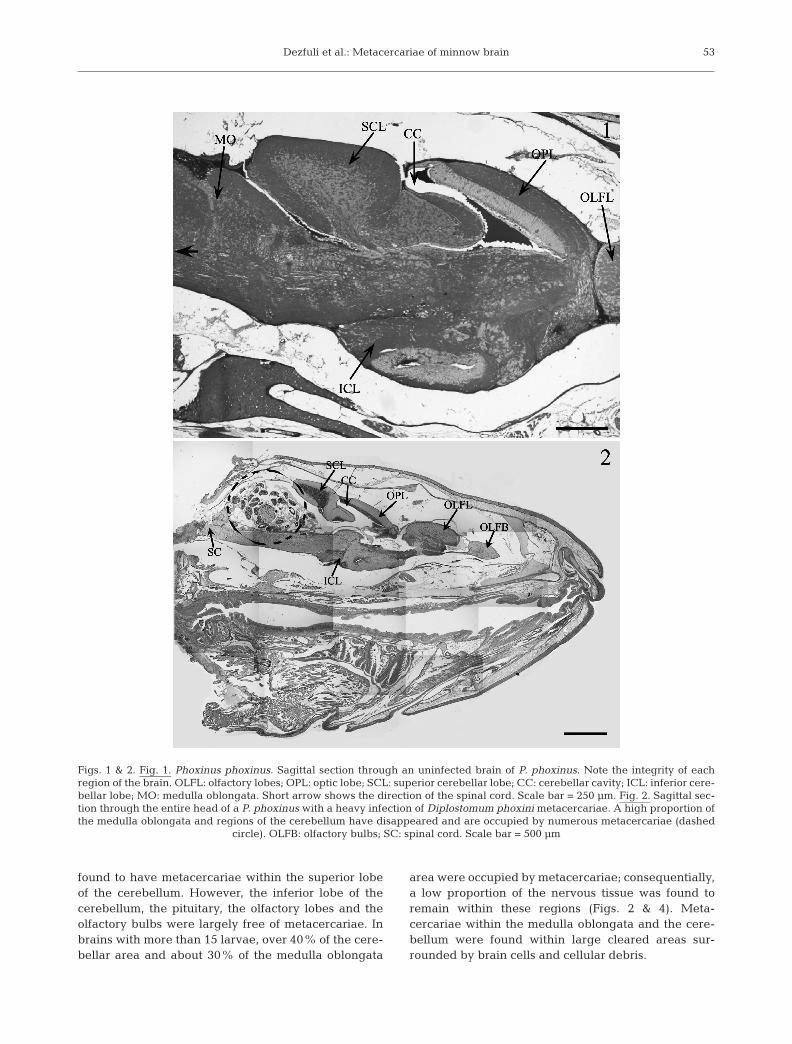
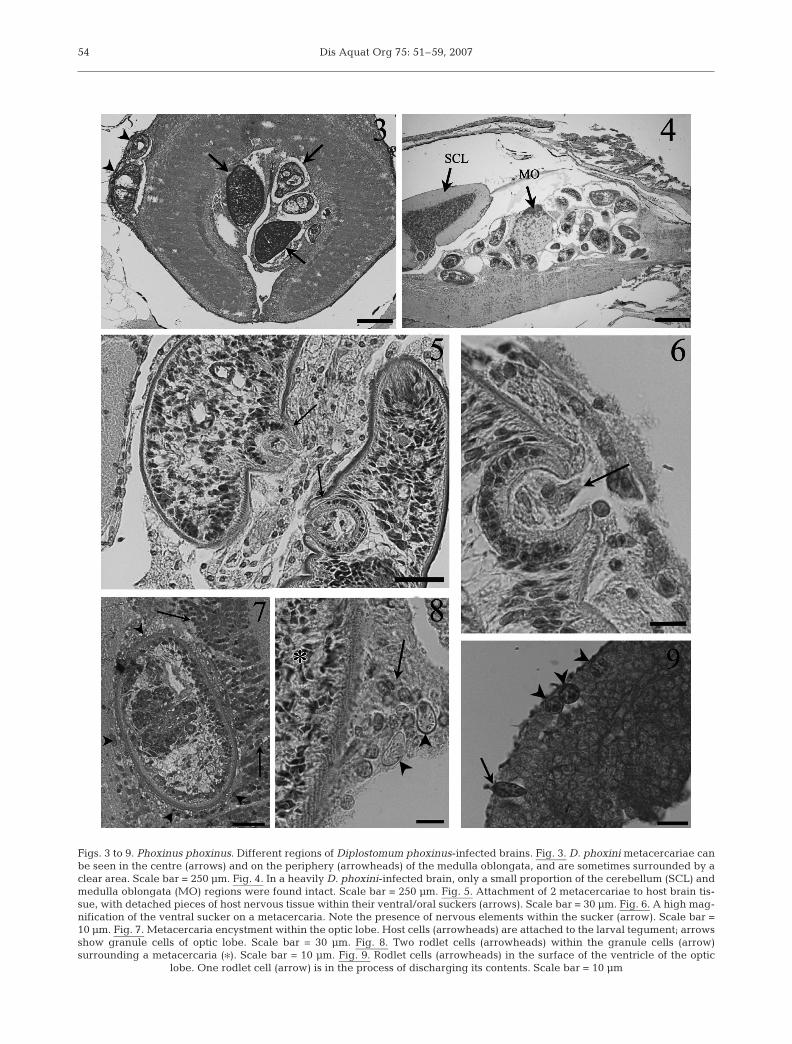
Post-mortem histological examination of 34 minnowheads revealed that 50% (n = 17) were infected with13.7 ± 2.6 (mean ± SE; range 1 to 38) Diplostomumphoxini metacercariae per infected fish. Measurement(mean ± SE) of the metacercariae (239.75 ± 11.51 µmlong by 103.05 ± 5.88 µm wide) indicated that theinfections were well established (as opposed to beingcharacterised by diplostomules that had recentlyentered the brain). Fig. 1 & 2 shows uninfected and D.phoxini-infected brains. The most frequently occupiedregions of the brain included the medulla oblongata(cortex and centre, Fig. 3), the optic lobes, the cerebel-lum and its superior lobe, and the cavity between thesuperior lobe and the anterior part of the medullaoblongata. In addition to these areas, 3 fish were also
52
Dezfuli et al.: Metacercariae of minnow brain
found to have metacercariae within the superior lobeof the cerebellum. However, the inferior lobe of thecerebellum, the pituitary, the olfactory lobes and theolfactory bulbs were largely free of metacercariae. Inbrains with more than 15 larvae, over 40% of the cere-bellar area and about 30% of the medulla oblongata
area were occupied by metacercariae; consequentially,a low proportion of the nervous tissue was found toremain within these regions (Figs. 2 & 4). Meta-cercariae within the medulla oblongata and the cere-bellum were found within large cleared areas sur-rounded by brain cells and cellular debris.
53
Figs. 1 & 2. Fig. 1. Phoxinus phoxinus. Sagittal section through an uninfected brain of P. phoxinus. Note the integrity of eachregion of the brain. OLFL: olfactory lobes; OPL: optic lobe; SCL: superior cerebellar lobe; CC: cerebellar cavity; ICL: inferior cere-bellar lobe; MO: medulla oblongata. Short arrow shows the direction of the spinal cord. Scale bar = 250 µm. Fig. 2. Sagittal sec-tion through the entire head of a P. phoxinus with a heavy infection of Diplostomum phoxini metacercariae. A high proportion ofthe medulla oblongata and regions of the cerebellum have disappeared and are occupied by numerous metacercariae (dashed
circle). OLFB: olfactory bulbs; SC: spinal cord. Scale bar = 500 µm
Dis Aquat Org 75: 51–59, 200754
Figs. 3 to 9. Phoxinus phoxinus. Different regions of Diplostomum phoxinus-infected brains. Fig. 3. D. phoxini metacercariae canbe seen in the centre (arrows) and on the periphery (arrowheads) of the medulla oblongata, and are sometimes surrounded by aclear area. Scale bar = 250 µm. Fig. 4. In a heavily D. phoxini-infected brain, only a small proportion of the cerebellum (SCL) andmedulla oblongata (MO) regions were found intact. Scale bar = 250 µm. Fig. 5. Attachment of 2 metacercariae to host brain tis-sue, with detached pieces of host nervous tissue within their ventral/oral suckers (arrows). Scale bar = 30 µm. Fig. 6. A high mag-nification of the ventral sucker on a metacercaria. Note the presence of nervous elements within the sucker (arrow). Scale bar =10 µm. Fig. 7. Metacercaria encystment within the optic lobe. Host cells (arrowheads) are attached to the larval tegument; arrowsshow granule cells of optic lobe. Scale bar = 30 µm. Fig. 8. Two rodlet cells (arrowheads) within the granule cells (arrow)surrounding a metacercaria (�). Scale bar = 10 µm. Fig. 9. Rodlet cells (arrowheads) in the surface of the ventricle of the optic
lobe. One rodlet cell (arrow) is in the process of discharging its contents. Scale bar = 10 µm

Dezfuli et al.: Metacercariae of minnow brain
Parasites were commonly observed attached to thehost tissue by their ventral and/or oral suckers, or wereobserved with detached pieces of brain tissue withinthem (Figs. 5 & 6). Single metacercaria, in 2 separatefish, were also noted embedded within the tissues of anolfactory bulb (Fig. 7). Although infections in the opticlobes were observed to be lower than those seen in thecerebellum or the medulla oblongata, metacercariaefound here and within the medulla oblongata (Fig. 7)were observed to be in intimate contact with granulecells of the brain. Granule cells, which are typicallydark in appearance and possess circular-shaped nucleiand a diffuse cytoplasm, form the granular layer of theoptic lobe ventricles (Fig. 7).
The presence of rodlet cells scattered among thegranule cells of the optic lobes of infected fish only(Figs. 8 & 12) is of particular interest. In each case, therodlet cells were observed on the surface of the ventri-cles of the optic lobes and in close proximity to ametacercaria, where some rodlet cells were seen to bedischarging their contents (Fig. 9). Rodlet cells werealso commonly encountered within the periphery ofthe cerebellum in all Diplostomum phoxini-infectedbrains. The rodlet cells, which appeared to be large indimension and measured 12.12 ± 0.19 µm (major axis)by 8.59 ± 0.16 µm (minor axis), typically contained ahigh number of club-shaped structures (rodlets) thatpossess a crystalline core and a vesicular, foam-likecytoplasm that did not contain any other evidentorganelles (Fig. 12).
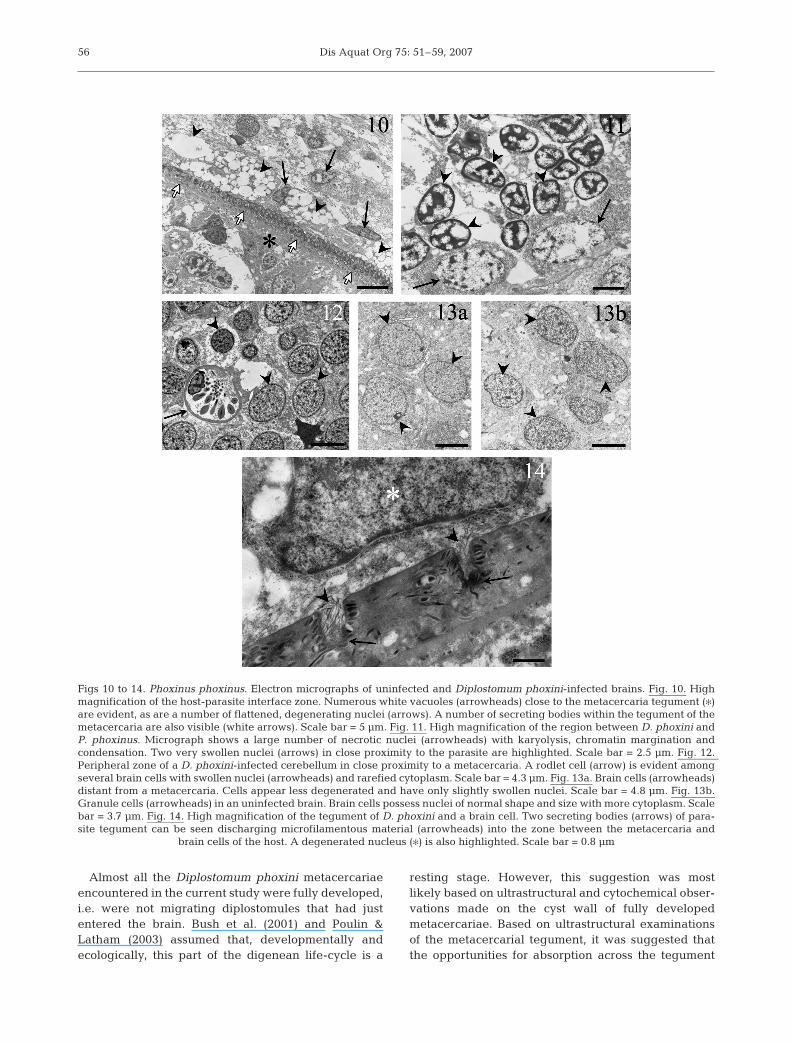
Thirteen of the 28 minnows heads prepared for waxembedding and 4 of the 6 heads prepared for the TEMstudy were infected with metacercariae: 1 to 38 parasitesper infected head were observed in the former study,and 1 to 3 in the latter. From these samples, only metac-ercariae within the optic and olfactory lobes wereobserved to make intimate contact with host tissue. ATEM study of these regions suggested that the regionwhere the parasite interfaces with the host can be bro-ken down into 2 zones. The inner zone, which is adjacentto the metacercaria, possesses a large number of whitevacuoles (Fig. 10). In histological sections prepared forlight microscopy, this zone appears as large white spacessurrounding metacercariae (Figs. 3 & 4). Within the op-tic lobe, the inner zone of granule cells, which are inclose proximity to a metacercaria, are notably flattenedand in a degenerative state typified by severe vacuola-tion and organelle regression (Figs. 10 & 11). The nucleiof the cells in this region also appear necrotic, showkaryolysis, chromatin margination and condensation(Fig. 11), and in some cases nuclear swelling (Fig. 11).
By contrast, the outer zone of the host-parasite inter-face appears more compact in density and containsseveral granule cells in a state of degeneration. Theultrastructural changes observed in the olfactory lobe
were similar to those observed for the optic lobe, withcells in a degenerative state that decreased in severitywith distance from the metacercariae (Fig. 13a). Thecharacteristics of the granule cells that are located at adistance from metacercariae and show few histopatho-logical changes (Fig. 13a) are similar to the granulecells observed in the brains of uninfected fish(Fig. 13b). The only observed difference betweenthese 2 groups of cells was that the granule cells in theuninfected fish brains contained more glycogen.
Close examination of the host-parasite interactionzone also revealed the occurrence of a high number ofsecretory vesicles (Fig. 14). These vesicles, which arearranged regularly within the tegument of the metac-ercariae, open into the interfacing zone, where theycan be seen releasing their cellular contents, whichconsist of a loose microfilamentous material (Fig. 14).In many instances, the secreted material was observedin contact with the host brain cells, which appeared tobe in a partial or total degenerative state (Fig. 14).
DISCUSSION
The current study found that the sub-sample ofPhoxinus phoxinus collected from the River Endrickpopulation were infected by Diplostomum phoxini thatfavor the cerebellum, the medulla oblongata and theoptic lobe regions of the brain. Metacercariae coloniz-ing these sites are suitably placed to alter the sensoryand motor functions of the host. For example, the cere-bellum of fish is believed to be linked to a variety ofcognitive functions (Rodriguez et al. 2005), and itsremoval from a dogfish Mustelus canis Mitchillresulted in a marked reduction in motor and sensoryperformance (Karamyan 1956). The medulla oblongatais known to control a range of involuntary responsessuch as breathing, digestion and heart rate (Bernstein1970), while the optic lobes are the primary vision-processing centre in teleost fishes (Guthrie 1986);herein, visual information regarding movement, colorand shape are analyzed, and stimuli from the lateralline system are received (Springer et al. 1977,Kortrschal et al. 1991).
An investigation by Barber & Crompton (1997) thatexamined the distribution of Diplostomum phoxini inthe brains of 2 populations of minnows found that the 2populations favoured particular regions of the brainand almost totally neglected others. The finding fromBarber & Crompton’s (1997) study suggests that indi-vidual D. phoxini or populations of D. phoxini mayfavour certain sites over others, which—once dam-aged—facilitates their transmission to the final host bycompromising the anti-predatory behaviour and per-formance of their fish host.
55
Dis Aquat Org 75: 51–59, 2007
Almost all the Diplostomum phoxini metacercariaeencountered in the current study were fully developed,i.e. were not migrating diplostomules that had justentered the brain. Bush et al. (2001) and Poulin &Latham (2003) assumed that, developmentally andecologically, this part of the digenean life-cycle is a
resting stage. However, this suggestion was mostlikely based on ultrastructural and cytochemical obser-vations made on the cyst wall of fully developedmetacercariae. Based on ultrastructural examinationsof the metacercarial tegument, it was suggested thatthe opportunities for absorption across the tegument
56
Figs 10 to 14. Phoxinus phoxinus. Electron micrographs of uninfected and Diplostomum phoxini-infected brains. Fig. 10. Highmagnification of the host-parasite interface zone. Numerous white vacuoles (arrowheads) close to the metacercaria tegument (�)are evident, as are a number of flattened, degenerating nuclei (arrows). A number of secreting bodies within the tegument of themetacercaria are also visible (white arrows). Scale bar = 5 µm. Fig. 11. High magnification of the region between D. phoxini andP. phoxinus. Micrograph shows a large number of necrotic nuclei (arrowheads) with karyolysis, chromatin margination andcondensation. Two very swollen nuclei (arrows) in close proximity to the parasite are highlighted. Scale bar = 2.5 µm. Fig. 12. Peripheral zone of a D. phoxini-infected cerebellum in close proximity to a metacercaria. A rodlet cell (arrow) is evident amongseveral brain cells with swollen nuclei (arrowheads) and rarefied cytoplasm. Scale bar = 4.3 µm. Fig. 13a. Brain cells (arrowheads)distant from a metacercaria. Cells appear less degenerated and have only slightly swollen nuclei. Scale bar = 4.8 µm. Fig. 13b.Granule cells (arrowheads) in an uninfected brain. Brain cells possess nuclei of normal shape and size with more cytoplasm. Scalebar = 3.7 µm. Fig. 14. High magnification of the tegument of D. phoxini and a brain cell. Two secreting bodies (arrows) of para-site tegument can be seen discharging microfilamentous material (arrowheads) into the zone between the metacercaria and
brain cells of the host. A degenerated nucleus (�) is also highlighted. Scale bar = 0.8 µm

Dezfuli et al.: Metacercariae of minnow brain
and for ingestion via the mouth were likely to be low(Stein & Lumsden 1971, Erasmus 1972, Bock 1988, Wit-trock et al. 1991). The recent research of Goater et al.(2005) provided morphological evidence to suggestthat certain species of metacercariae undergo complexdevelopmental changes associated with feeding intheir intermediate host. The observation of detachedbrain tissue within the oral suckers of attached metac-ercariae in the present study would suggest that thisspecies feeds on host tissues. The work of Bibby & Rees(1971) on extracorporeal digestion in D. phoxini metac-ercariae and the finding of acid phosphatase through-out the metacercaria would suggest that intracellulardigestion is occurring and that the parasites are feed-ing. Based on the results of an experimentally infectedsample of minnows, Goater et al. (2005) suggested thatminnows are either able to tolerate a certain level ofdamage to their brains or are able to regenerate orrepair brain tissue. Post-mortem examination of min-nows in the current study revealed that the minnowshad light infections (range 1 to 38) compared withthose (1300 metacercariae) reported by Rees (1955).Therefore, the observed levels of infection and theresults from the current study correspond to the find-ings of Goater et al. (2005).
Only a few records relating to the host’s immunereaction in response to Diplostomum spp. infectionsexist (Bortz et al. 1984, Stables & Chappell 1986). Sim-ilarly, the host cellular reaction in Diplostomum-basedhistopathological studies has been largely overlooked(Rees 1955, Hoffman & Hoyme 1958, Shariff et al. 1980,Baghaei 1981). One of the main objectives of the cur-rent study was to look closely at the host–parasiteinteraction and to characterise the cells present withinthe interfacing zone. One interesting observation wasthe presence of rodlet cells within this zone, the onlyhost inflammatory cells documented in this sub-sampleof D. phoxini-infected Phoxinus phoxinus brains.Rodlet cells are, as far as we are aware, exclusive tofish and are associated with the epithelial tissues of vir-tually every fish species (Morrison & Odense 1978).Recent research by a number of workers lends supportto the theory that rodlet cells have an endogenous fishorigin (Manera & Dezfuli 2004, Giari et al. 2006, Reite& Evensen 2006, Dezfuli et al. 2007). Although theirprecise function is unknown, it is thought that rodletcells are a type of inflammatory cell akin to otherpiscine inflammatory cells (e.g. mast cells/eosinophilicgranule cells, epithelioid cells and mesothelial cells)(Dezfuli et al. 2000, Reite 2005, Reite & Evensen 2006).Most of the rodlet cells that were found in the currentstudy were observed in the epithelial lining of the ven-tricles of the optic lobes and in sites close to the tegu-ment of metacercariae. This observation fits well withLeino’s (1996) suggestion that rodlet cells react to the
presence of parasites on the epithelial surface, and thatcellular secretions are possibly antibiotic in nature.This theory is also supported by the work of others thathave investigated fish protozoan (Leino 1996, Dezfuliet al. 2004) and metazoan parasitic agents, all of whichappear to induce the recruitment of rodlet cells to thesite of infection and/or to adjacent tissues (Dezfuli et al.1998, 2000, 2003, Palenzuela et al. 1999, Bosi et al.2005, Reite 2005, Reite & Evensen 2006, the presentstudy).
While there have been numerous studies on theultrastructure of metacercariae (Stein & Lumsden1971, Strong & Cable 1972, Higgins 1980, Bock 1988,Wittrock et al. 1991), only a few have focused on that ofdiplostomatids (Bibby & Rees 1971, Mitchell 1974,Goater et al. 2005). Of these, the detailed study byBibby & Rees (1971) on the epidermis of Diplostomumphoxini metacercariae described 3 types of secretorybodies that were regularly arranged in the outer epi-dermal layer, but did not comment on the possiblefunction of the secretions. The numerous secretorybodies found in the tegument of D. phoxini in the pre-sent study were observed to release a loose, microfila-mentous material into the zone between the host andthe parasite. In addition to these bodies, many bodiesthat had already emptied their contents were also evi-dent, usually in that part of the parasite’s tegumentadjacent to a host cell (Fig. 3f). The brain cells in thisregion characteristically bore flattened nuclei andwere in a partial or complete degenerative state. How-ever, without knowing the precise chemical nature ofthe secreted product (which we are investigatingwithin our laboratory at the time of writing), we canonly hypothesise that the compound is secreted to pro-tect the metacercaria from the host’s cellular response.
Acknowledgements. We thank the technical staff of theElectron Microscopy Centre at the University of Ferrara fortheir assistance and expertise. This study was supported inpart by grants from the Italian Ministry of the University andScientific Research and Technology.
LITERATURE CITED
Baghaei M (1981) Experimentally induced encapsulation ofDiplostomum phoxini (Faust) in the fish host. Experientia37:1283–1285
Barber I, Crompton DWT (1997) The distribution of the metac-ercariae of Diplostomum phoxini in the brain of minnow,Phoxinus phoxinus. Folia Parasitol 44:19–25
Barber I, Hoare D, Krause J (2000) Effects of parasites on fishbehaviour: a review and evolutionary perspective. RevFish Biol Fish 10:131–165
Bernstein JJ (1970) Anatomy and physiology of the centralnervous system. In: Hoar WS, Randall DJ (eds) Fish physi-ology, Vol IV. Academic Press, New York, p 1–90
Bibby MC, Rees G (1971) The ultrastructure of the epidermis
57

Dis Aquat Org 75: 51–59, 2007
and associated structures in the metacercaria cercaria andsporocyst of Diplostomum phoxini (Faust, 1918). Z Para-sitenkd 37:169–186
Bock D (1988) Formation, histochemistry, and ultrastructureof the metacercarial cyst wall of Plagiorchis species 1(Trematoda: Plagiorchiidae). Int J Parasitol 18:379–388
Bortz BM, Kenny GE, Pauley GB, Garcia-Ortigoza E, Ander-son DP (1984) The immune response in immunised andnaturally infected rainbow trout (Salmo gairdneri) toDiplostomum spathaceum as described by the enzyme-linked immunosorbent assay (ELISA). Dev Comp Immunol8:813–822
Bosi G, Shinn AP, Giari L, Simoni E, Pironi F, Dezfuli BS (2005)Histopathology and alteration to neuromodulators of thediffuse endocrine system of the alimentary canal of farmedOncorhynchus mykiss (Walbaum) naturally infected withEubothrium crassum (Cestoda). J Fish Dis 28:703–711
Bush AO, Fernandez JC, Esch GW, Seed JR (2001) Parasitism:the diversity and ecology of animal parasites. CambridgeUniversity Press, Cambridge
Chappell LH (1995) The biology of diplostomatid eyeflukes offishes. J Helminthol 69:97–101
Crompton DWT (1976) Entry into the host and site selection.In: Kennedy CR (ed) Ecological aspects of parasitology.North-Holland Publishing, Oxford, p 41–73
Dezfuli BS, Capuano S, Manera M (1998) A description ofrodlet cells of alimentary canal of Anguilla anguilla (L.)and their relationship with parasitic helminths. J Fish Biol53:1084–1095
Dezfuli BS, Simoni E, Rossi R, Manera M (2000) Rodlet cells andother inflammatory cells of Phoxinus phoxinus infected withRaphidascaris acus (Nematoda). Dis Aquat Org 43:61–69
Dezfuli BS, Giari L, Simoni E, Bosi G, Manera M (2002)Histopathology, immunohistochemistry and ultrastructureof the intestine of Leuciscus cephalus naturally infectedwith Pomphorhynchus laevis (Acanthocephala). J Fish Dis25:7–14
Dezfuli BS, Giari L, Konecny R, Jaeger P, Manera M (2003)Immunohistochemistry, ultrastructure and pathology ofgills of Abramis brama (L.) from lake Mondsee (Austria)due to Ergasilus sieboldi (Copepoda). Dis Aquat Org 53:257–262
Dezfuli BS, Giari L, Simoni E,. Shinn A, Bosi G (2004)Immunohistochemistry, histopathology and ultrastructureof Gasterosteus aculeatus (L.) tissues infected with Glugeaanomala (Moniez 1887). Dis Aquat Org 58:193–202
Dezfuli BS, Capuano S, Simoni E, Previati M, Giari L (2007)Rodlet cells and the sensory systems in zebrafish (Daniorerio). Anat Rec (in press)
Erasmus DA (1972) Pathology and host responses. In: ArnoldE (ed) The biology of trematodes. The University Press,Belfast, p 188–210
Faliex E, Biagianti S (1987) Metacercarial infection of marinefish by Labratrema minimus (Digenea, Bucephalidae):histo-cytological analysis of host-parasite relationship.Aquaculture 67:229–232
Giari L, Manera M, Simoni E, Dezfuli BS (2006) Changes tochloride and rodlet cells in gills, kidney and intestine ofDicentrarchus labrax (L.) exposed to reduced salinities.J Fish Biol 69:590–600
Goater CP, Douglas B, Conn DB (2005) Cellular aspects ofearly development of Ornithodiplostomum ptychocheilusmetacercariae in the brain of fathead minnows,Pimephales promelas. J Parasitol 91:814–821
Guthrie DM (1986) Role of vision in fish behaviour. In: PitcherTJ (ed) The behaviour of teleost fishes. Croom Helm, Lon-don, p 111–121
Higgins JC (1980) Formation of the cyst wall and relatedchanges in the structure of the tegument of Bucephalushaimeanus (Lacaze-Duthiers, 1854) during its metamor-phosis from the cercarial to the metacercarial stage. Para-sitology 81:47–59
Hoffman GL, Hoyme JB (1958) The experimental histopathol-ogy of the ‘tumor’ on the brain of the stickleback causedby Diplostomum baeri eucaliae Hoffman and Hundley,1957 (Trematoda: Strigeoidea). J Parasitol 44:374–378
Holmes JC, Price PW (1985) Communities of parasites. In:Anderson DJ, Kikkawa J (eds) Community ecology: pat-tern and processes. Blackwell Scientific Publications,Oxford, p 187–213
Karamyan AI (1956) Evolution of the function of the cerebel-lum and cerebral hemispheres. Fiziol Zh SSSR 25 (3). Eng-lish translation (1962) US Dept Comm Office Tech Serv PBRep 31014, TT 1–162
Kortrschal K, Brandstatter R, Gomahr A, Junger K, Palzen-berger M, Zaunreiter M (1991) Brain and sensory systems.In: Winfield IJ, Nelson JS (eds) Cyprinid fishes: systemat-ics, biology and exploitation. Chapman & Hall, London,p 284–331
Leino RL (1996) Reaction of rodlet cells to a myxosporeaninfection in kidney of the bluegill, Lepomis macrochirus.Can J Zool 74:217–225
Manera M, Dezfuli BS (2004) Rodlet cells in teleosts: a newinsight into their nature and functions. J Fish Biol 65:597–619
Mitchell CW (1974) Ultrastructure of the metacercarial cyst ofPosthodiplostomum minimum (MacCallum, 1921). J Para-sitol 60:67–74
Morrison CM, Odense PH (1978) Distribution and morphol-ogy of the rodlet cell in fish. J Fish Res Board Can 35:101–116
Palenzuela O, Alvarez-Pellitero P, Sitjà-Bobadilla A (1999)Glomerular disease associated with Polysporoplasmasparis (Myxozoa) infection in cultured gilthead sea bream,Sparus aurata L. (Pisces: Teleostei). Parasitology 118:245–256
Poulin R, Latham DM (2003) Effects of initial (larval) size andhost body temperature on growth in trematodes. CanJ Zool 81:574–581
Price PW (1987) Evolution in parasite communities. Int J Par-asitol 17:209–214
Pylkkö P, Suomalainen LR, Tiirola M, Valtonen ET (2006) Evi-dence of enhanced bacterial invasion during Diplostomumspathaceum infection in European grayling, Thymallusthymallus (L.). J Fish Dis 29:79–86
Rees G (1955) The adult and Diplostomulum stage (Diplosto-mulum phoxini [Faust]) of Diplostomum pelmatoidesDubois and an experimental demonstration of part of thelife cycle. Parasitology 45:295–313
Reite OB (1998) Mast cells/eosinophilic granule cells ofteleostean fish: a review focusing on staining propertiesand functional responses. Fish Shellfish Immunol 8:489–513
Reite OB (2005) The rodlet cells of teleostean fish: their poten-tial role in host defence in relation to the role of mastcells/eosinophilic granule cells. Fish Shellfish Immunol 19:253–267
Reite OB, Evensen Ø (2006) Inflammatory cells of teleosteanfish: a review focusing on mast cells/eosinophilic granulecells and rodlet cells. Fish Shellfish Immunol 20:192–208
Rodriguez F, Duran E, Gomez A, Ocana FM, Alvarez E,Jimenez-Moya F, Broglio C, Salas C (2005) Cognitive andemotional functions of the teleost fish cerebellum. BrainRes Bull 66:365–370
58

Dezfuli et al.: Metacercariae of minnow brain
Shariff M, Richards R, Sommerville C (1980) The histopathol-ogy of acute and chronic infections of rainbow trout Salmogairdneri Richardson with eye flukes, Diplostomum spp.J Fish Dis 3:455–465
Shirakashi S, Goater CP (2005) Chronology of parasite-induced alteration of fish behaviour: effects of parasitematuration and host experience. Parasitology 130:177–183
So FW, Wittrock DD (1982) Ultrastructure of the metacercarialcyst of Ornithodiplostomum ptychocheilus (Trematoda:Diplostomatidae) from the brains of fathead minnows.Trans Am Microsc Soc 101:181–185
Springer AD, Easter SS, Agranoff BW (1977) The role of optictectum in various visually mediated behaviors of goldfish.Brain Res 128:393–404
Stables JN, Chappell LH (1986) Putative immune responseof rainbow trout, Salmo gairdneri, to Diplostomumspathaceum infections. J Fish Biol 29:115–122
Stein PC, Lumsden RD (1971) An ultrastructural and cyto-chemical study of metacecarial cyst development in Asco-cotyle pachycystis Schroeder and Leigh, 1965. J Parasitol57:1231–1246
Strong PL, Cable RM (1972) Fine structure and developmentof the metacercarial cyst in Microphallus opacus (Ward,1894). J Parasitol 58:92–98
Sukhdeho MVK, Mettrick DF (1987) Parasite behaviour:understanding platyhelminth responses. Adv Parasitol 26:73–144
Sukhdeho MVK, Sukhdeho SC (1994) Optimal habitat selec-tion by helminths within the host environment. Parasitol-ogy 109:S41–S55
Wittrock DD, Bruce CS, Johnson AD (1991) Histochemistryand ultrastructure of the metacercarial cysts of blackspottrematodes Uvulifer ambloplitis and Neascus pyriformis.J Parasitol 77:454–460
59
Editorial responsibility: David Marcogliese,Montreal, Quebec, Canada
Submitted: October 12, 2006; Accepted: February 7, 2007Proofs received from author(s): March 26, 2007




















