
Histopathological Changes after Induced Injury in Leeches 1
15
Histopathological Changes after Induced Injury in Leeches 1 Magda de Eguileor, Gianluca Tettamanti, Annalisa Grimaldi, Angelo Boselli, Giorgio Scarı `, Roberto Valvassori, Edwin L. Cooper,* and Giulio Lanzavecchia DBSF, University of Insubria, Via J. H. Dunant 3, 21100 Varese, Italy; and *Laboratory of Comparative Immunology, Department of Neurobiology, UCLA School of Medicine, California 90095-1763 Received June 15, 1998; accepted January 28, 1999 Cells involved in leech inflammatory responses have been characterized by morphological, histochemical, and immunohistochemical methods. Macrophage-like cells, NK-like cells, and granulocytes migrated shortly after injury by pricking with bacterial lipopolysaccha- ride. Inflammatory responses increased progressively and provoked cell migration to the body wall and then to wound surfaces. Macrophages, NK cells, and granu- locytes display similar features and behavior traits in invertebrates and vertebrates. r 1999 Academic Press Key Words: leeches; wound healing; macrophage-like cells; NK-like cells; granulocytes; LPS. INTRODUCTION The process of lipopolysaccharide (LPS) induction of wound healing in leeches is of interest because of observed histopathological changes that accompany repair after injury. In mammalian and in some inverte- brate cells, LPS is known for the direct and indirect roles it plays in immune responses (Blanco et al., 1997). Although wound healing in leeches is similar to that of vertebrates, it is still easier to study the processes in leeches because of their relative anatomical simplicity (Huguet and Molinas, 1994). Previous studies of inju- ries inflicted on leeches revealed that healing involves the formation of a temporary cellular clump, known as the pseudoblastema, reepithelialization, and wound contraction (Molinas et al., 1988; Huguet and Molinas, 1988, 1992, 1994, 1996). Moreover Huguet and Molinas (1994, 1996) report a migration of vasocentral cells in relation to regeneration. While studies of wound healing and inflammation in leeches are fragmentary and scarce (Cornec, 1990; Le Gore and Sparks, 1971, 1973; Mann, 1962), it is well known that macrophage-like and NK-like cells have been found in oligochaetes (terrestrial annelids, e.g., earthworms) (Cooper, 1996) and polychaetes (marine annelids, e.g., ‘‘blood worms’’) (Porchet-Hennere ´ et al., 1992). The purpose of the present work was to reveal histopathological changes in leeches after injury with the view toward further understanding the inflamma- tory response. MATERIALS AND METHODS Leeches Leeches (Glossiphonia complanata, Hirudinea) were collected from the Adda river, near Milan, Italy. They were kept for a month before experiments in water at 18°C in aerated tanks and fed with Planorbidae snails. Leeches were subdivided into five groups (10 leeches): group 1 served as controls; group 2 served as test subjects for reactions to pricking alone. Leeches in groups 3, 4, and 5 were pricked with a needle (1.20 3 40 mm) and given 10 mg/ml of lipopolysaccharide (used for immune stimulation). Leeches were soaked in the same solution to enhance the effect of LPS and then sacrificed at 10 min, 30 min, and 4 h. All leeches were anesthe- tized with a saturated solution of Mephenesin (3-o- toloxy-1,2 propandiol). This is a well known muscle relaxant (The Merck Index) that is advantageous since leech body walls are characterized by heavy bands of striated muscle. Light Microscopy and Transmission Electron Microscopy For routine transmission electron microscopy (TEM), leeches were sectioned (longitudinally from oral to anal sucker), fixed in 2% glutaraldehyde in 0.1 M cacodylate buffer for 2 h, rinsed in buffer, and postfixed for 2 h with 1% osmic acid in cacodylate buffer (pH 7.2). After standard dehydration in an ethanol series, specimens were embedded in an Epon-Araldite 812 mixture and sectioned with a Reichert Ultracut E ultrotome (Leica, Nussloch, Germany). Semi-thin sections were stained by conventional methods according to Moore et al. (1960) and observed with an optical microscope (Olym- pus, Tokyo, Japan). Thin sections were stained with 1 This work was supported by grants from MURST, 40%, 1996. Journal of Invertebrate Pathology 74, 14–28 (1999) Article ID jipa.1999.4850, available online at http://www.idealibrary.com on 14 0022-2011/99 $30.00 Copyright r 1999 by Academic Press All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Histopathological Changes after Induced Injury in Leeches 1

bacaratli
c
worbrAvl(rttc1(r
lGkbe
Journal of Invertebrate Pathology 74, 14–28 (1999)Article ID jipa.1999.4850, available online at http://www.idealibrary.com on
0CA
Histopathological Changes after Induced Injury in Leeches1
Magda de Eguileor, Gianluca Tettamanti, Annalisa Grimaldi, Angelo Boselli, Giorgio Scarı, Roberto Valvassori,Edwin L. Cooper,* and Giulio Lanzavecchia
DBSF, University of Insubria, Via J. H. Dunant 3, 21100 Varese, Italy; and *Laboratory of Comparative Immunology,Department of Neurobiology, UCLA School of Medicine, California 90095-1763
Received June 15, 1998; accepted January 28, 1999
a1htt
L
cw1Lgsgmisattrls
L
lsb1swsNb(
Cells involved in leech inflammatory responses haveeen characterized by morphological, histochemical,nd immunohistochemical methods. Macrophage-likeells, NK-like cells, and granulocytes migrated shortlyfter injury by pricking with bacterial lipopolysaccha-ide. Inflammatory responses increased progressivelynd provoked cell migration to the body wall and theno wound surfaces. Macrophages, NK cells, and granu-ocytes display similar features and behavior traits innvertebrates and vertebrates. r 1999 Academic Press
Key Words: leeches; wound healing; macrophage-likeells; NK-like cells; granulocytes; LPS.
INTRODUCTION
The process of lipopolysaccharide (LPS) induction ofound healing in leeches is of interest because ofbserved histopathological changes that accompanyepair after injury. In mammalian and in some inverte-rate cells, LPS is known for the direct and indirectoles it plays in immune responses (Blanco et al., 1997).lthough wound healing in leeches is similar to that ofertebrates, it is still easier to study the processes ineeches because of their relative anatomical simplicityHuguet and Molinas, 1994). Previous studies of inju-ies inflicted on leeches revealed that healing involveshe formation of a temporary cellular clump, known ashe pseudoblastema, reepithelialization, and woundontraction (Molinas et al., 1988; Huguet and Molinas,988, 1992, 1994, 1996). Moreover Huguet and Molinas1994, 1996) report a migration of vasocentral cells inelation to regeneration.While studies of wound healing and inflammation in
eeches are fragmentary and scarce (Cornec, 1990; Leore and Sparks, 1971, 1973; Mann, 1962), it is wellnown that macrophage-like and NK-like cells haveeen found in oligochaetes (terrestrial annelids, e.g.,arthworms) (Cooper, 1996) and polychaetes (marine
1
pThis work was supported by grants from MURST, 40%, 1996.14022-2011/99 $30.00opyright r 1999 by Academic Pressll rights of reproduction in any form reserved.
nnelids, e.g., ‘‘blood worms’’) (Porchet-Hennere et al.,992). The purpose of the present work was to revealistopathological changes in leeches after injury withhe view toward further understanding the inflamma-ory response.
MATERIALS AND METHODS
eeches
Leeches (Glossiphonia complanata, Hirudinea) wereollected from the Adda river, near Milan, Italy. Theyere kept for a month before experiments in water at8°C in aerated tanks and fed with Planorbidae snails.eeches were subdivided into five groups (10 leeches):roup 1 served as controls; group 2 served as testubjects for reactions to pricking alone. Leeches inroups 3, 4, and 5 were pricked with a needle (1.20 3 40m) and given 10 mg/ml of lipopolysaccharide (used for
mmune stimulation). Leeches were soaked in the sameolution to enhance the effect of LPS and then sacrificedt 10 min, 30 min, and 4 h. All leeches were anesthe-ized with a saturated solution of Mephenesin (3-o-oloxy-1,2 propandiol). This is a well known muscleelaxant (The Merck Index) that is advantageous sinceeech body walls are characterized by heavy bands oftriated muscle.
ight Microscopy and Transmission ElectronMicroscopy
For routine transmission electron microscopy (TEM),eeches were sectioned (longitudinally from oral to analucker), fixed in 2% glutaraldehyde in 0.1 M cacodylateuffer for 2 h, rinsed in buffer, and postfixed for 2 h with% osmic acid in cacodylate buffer (pH 7.2). Aftertandard dehydration in an ethanol series, specimensere embedded in an Epon-Araldite 812 mixture and
ectioned with a Reichert Ultracut E ultrotome (Leica,ussloch, Germany). Semi-thin sections were stainedy conventional methods according to Moore et al.1960) and observed with an optical microscope (Olym-
us, Tokyo, Japan). Thin sections were stained with
lm
S
saTts
H
ggBVstaPaafbsuoe
H
mtffits(miamccrrttst
S
tnco1sf1
S
fiprTmGle1sdb(a
icm2cgtc2gMs
C
iHc2tcsA(
15WOUND HEALING IN LEECHES
ead citrate and observed with a Jeol 1010 EX electronicroscope (Jeol, Tokyo, Japan).
canning Electron Microscopy
Six leeches were fixed, post-fixed (as previously de-cribed), dehydrated in an increasing series of ethanolnd hexamethyldisilazane (HMDS), and then air dried.he specimens were gold coated with a Sputter Nano-ech instrument and observed in a Cambridge Stereo-can 250 MK.
istochemistry
Frozen sections (10 µm thick) of 20 leeches from eachroup were cut in a cryostat at 220°C, mounted onlass slides, and stained according to Dubowitz androoke (1973) using a histoenzymatic kit (Bio-optica).arious cytoplasmic characteristics were assayed usingeveral histochemical procedures: NADH-TR (NADH-etrazolium reductase) attributable to mitochondrialctivity (i.e., to the source of energy in cell metabolism);eriodic Acid Schiff (PAS) for polysaccharides; alkalinend acid phosphatase for microsomal and lysosomalctivity; and nonspecific esterase and Oil red O (ORO)or lipid metabolism. Controls for all reactions wereased on the omission of the enzyme, except for the PAStain, whose specificity for glycogen was confirmed bysing diastase digestion. Negative results were alwaysbtained, confirming the specificity of reactions; eachxperiment was repeated three to four times.
RESULTS
istological Features of Macrophage-like Cells,NK-like Cells, and Granulocytes and theirMigrations after Wounding
Normal uninjured body wall of leeches showed no celligration (Fig. 1). Pricked leeches showed cell migra-
ion from the central region of the body to its surface. Aew cells were observed in the area between the muscleelds just 30 min after injury (Fig. 2) and at the sameime leeches pricked with the addition of LPS showedubstantial cell migration toward the wounded regionFig. 3). Migrating cells from botryoidal tissue betweenuscle layers reached the healing area at which they
ncreased numerically in relation to the time elapsedfter the lesions were inflicted. In fact, after 4 h,ultiple cell layers formed plugs (Fig. 4). Distinct
ellular populations included: (a) irregularly shapedells up to 25 3 10 µm (their dimensions varied inelation to the emission of pseudopodia) (Fig. 5), (b)oundish cells up to 10 µm in diameter (Fig. 6), and (c)wo types of granulocytes (Figs. 7 and 8). The first cellype was designated as macrophage-like cells, theecond cell type was referred to as NK-like cells, and
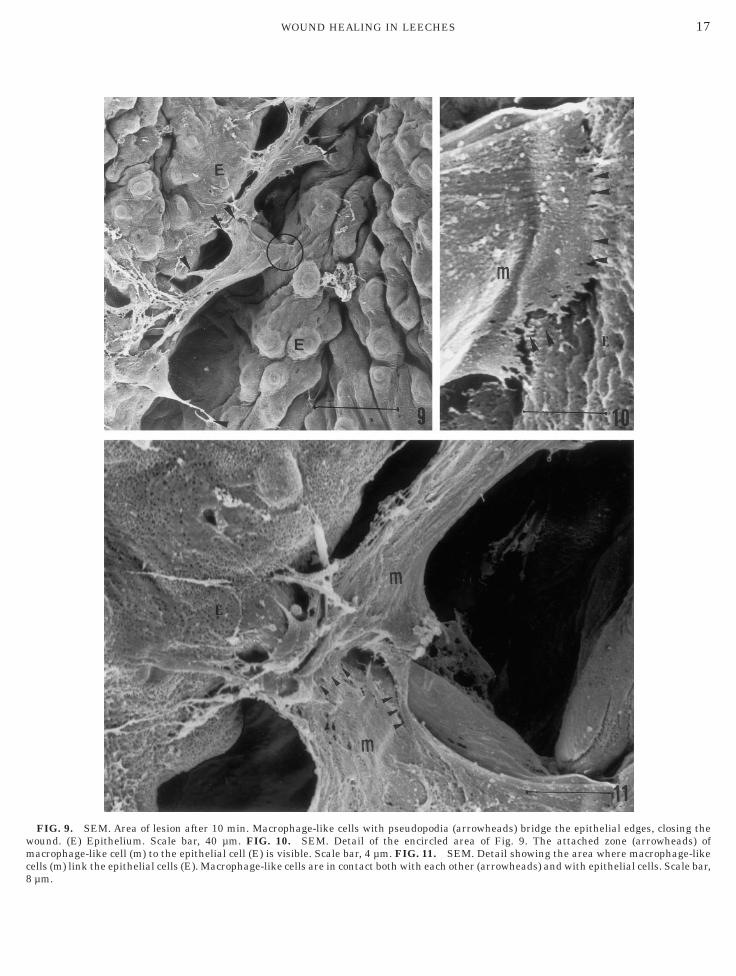
he third cell type was classified as granulocytes. 3canning Electron Microscopy Revealed the Shape ofMigrating Cells
After 10 min, we observed irregularly shaped cellshat reduced the width of the lesion, bringing one edgeear to the other with their pseudopodia (Fig. 9). Theseells were in contact either with epithelial cells (Fig. 10)r with pseudopodia of similar neighboring cells (Fig.1). After 15 min, other cells, smaller and roundish inhape, were observed in the healing area where theyormed a plug, filling the area of the lesion (Figs. 12 and3).
patial Relations of Migrating Macrophage-like Cells,NK-like Cells, and Granulocytes
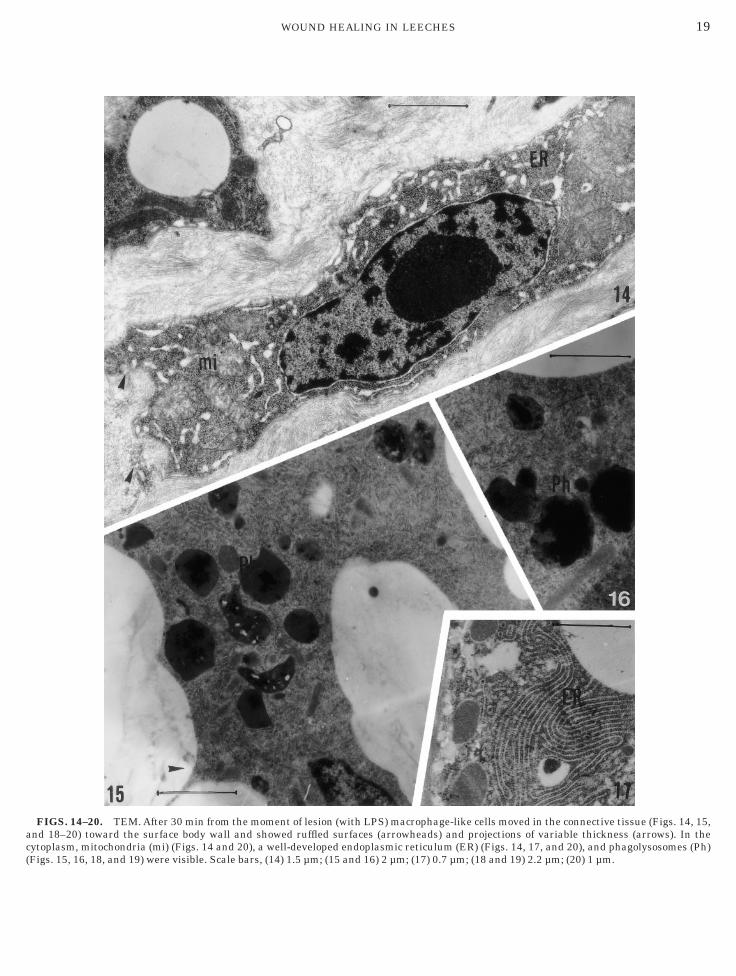
Transmission electron micrographs revealed that therst cell type (macrophage-like cells), characterized byseudopodia, possessed a higher cytoplasmic/nuclearatio (Fig. 14) than the second cell type (NK-like cells).he cytoplasm of macrophage-like cells contained nu-erous mitochondria, lipid droplets, a well-developedolgi complex, cisternae of endoplasmic reticulum, and
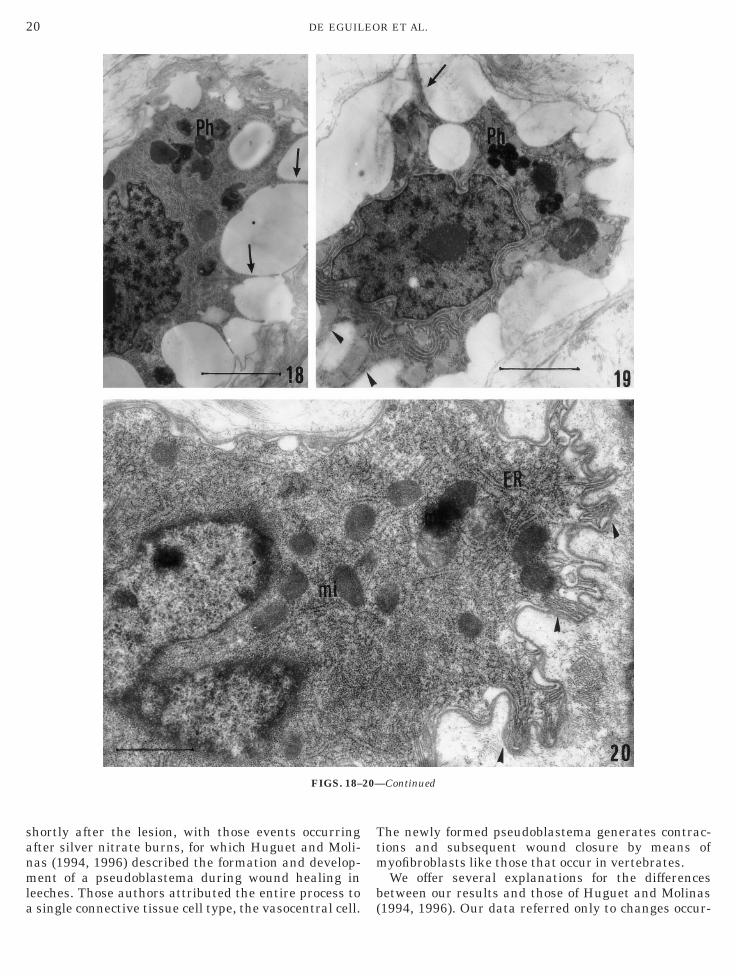
ysosomes (Figs. 14–20). The phagolysosomes werespecially numerous 4 h after the injury (Figs. 15 and6). Moreover, these migrating cells changed theirhape, showing, in the connective tissue, a certainegree of surface ruffling (Figs. 14–20). The ruffledorders consisted of short, blunt, finger-like projectionsFigs. 14–20). The extracellular matrix was less denseround these projections (Figs. 18–20).The second cell type (NK-like cells) was character-
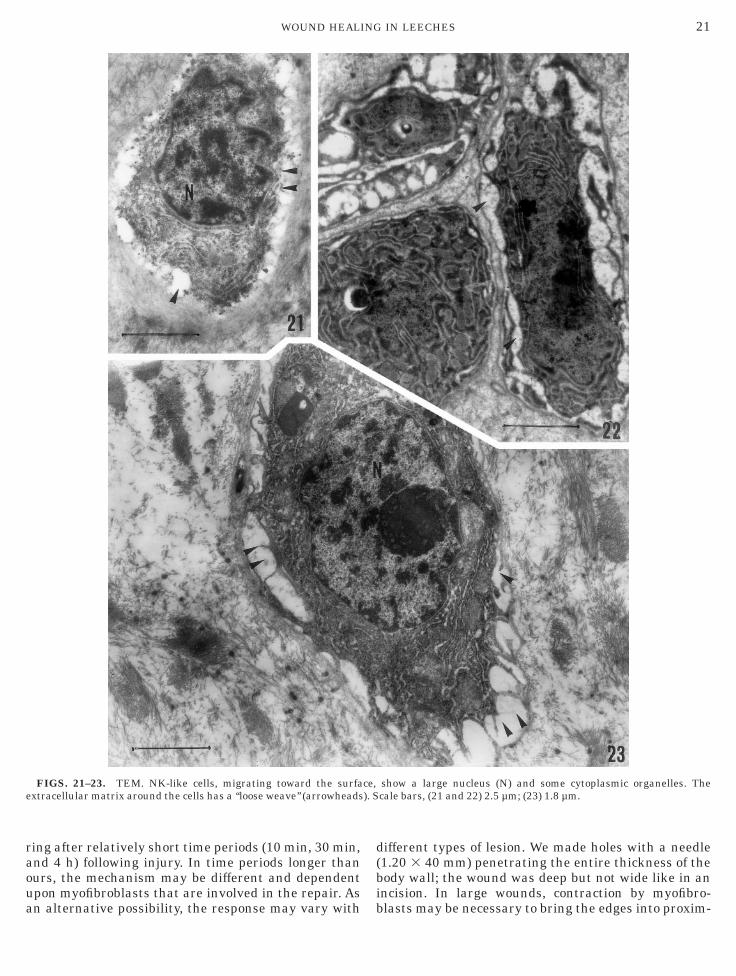
zed by a low nuclear/cytoplasmic ratio. The cytoplasmontained a well-developed endoplasmic reticulum, nu-erous mitochondria, and dense granules (Figs. 21–
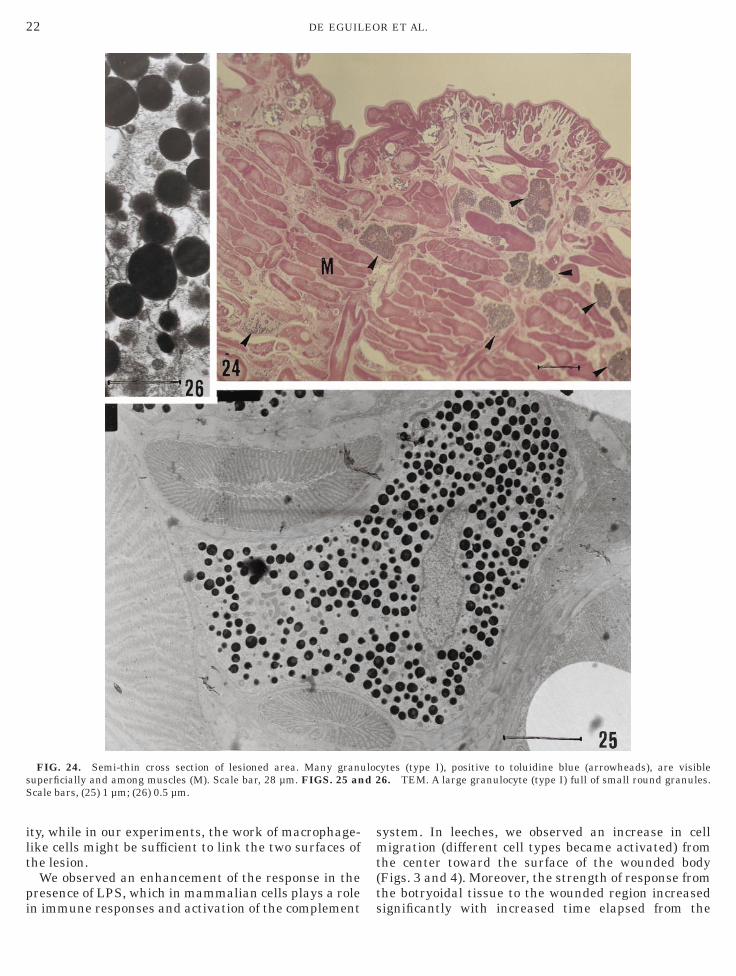
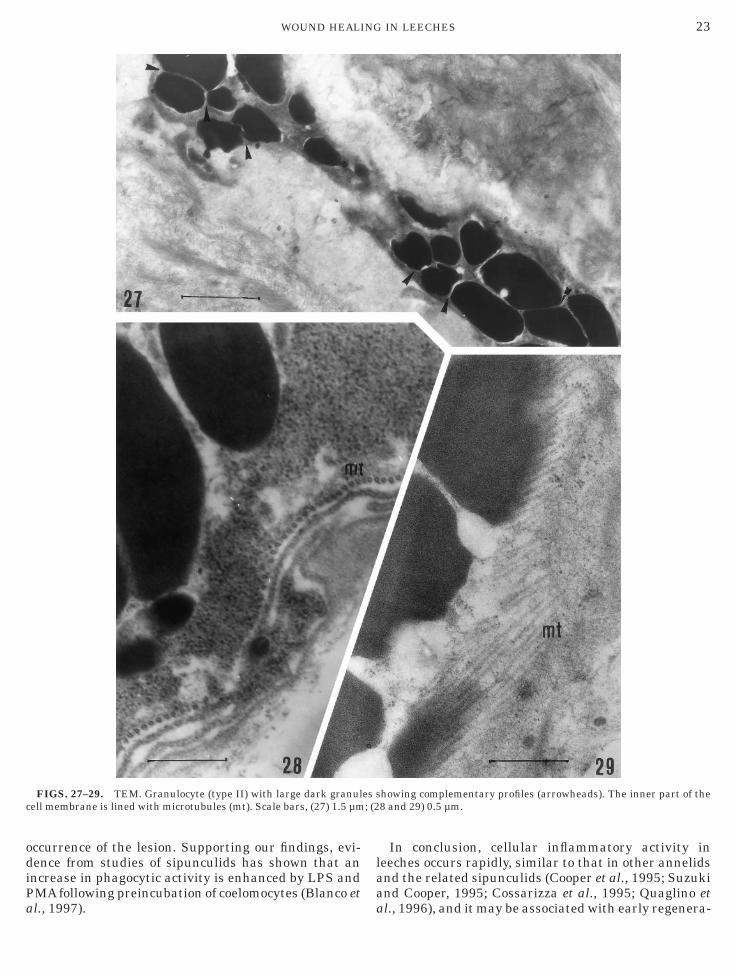
3). Granulocytes, the third cell type, contained denseytoplasmic granules. On the basis of size and shape ofranules, we distinguished two different granulocyticypes. Granulocyte type I was large (up to 30 µm) andontained numerous, small round granules (Figs. 24–6). Granulocyte type II possessed irregularly shapedranules showing complementary profiles (Fig. 27).icrotubules lined the inner region of granulocyte
urfaces (Figs. 28 and 29).
ellular Characteristics Revealed by Histochemistry
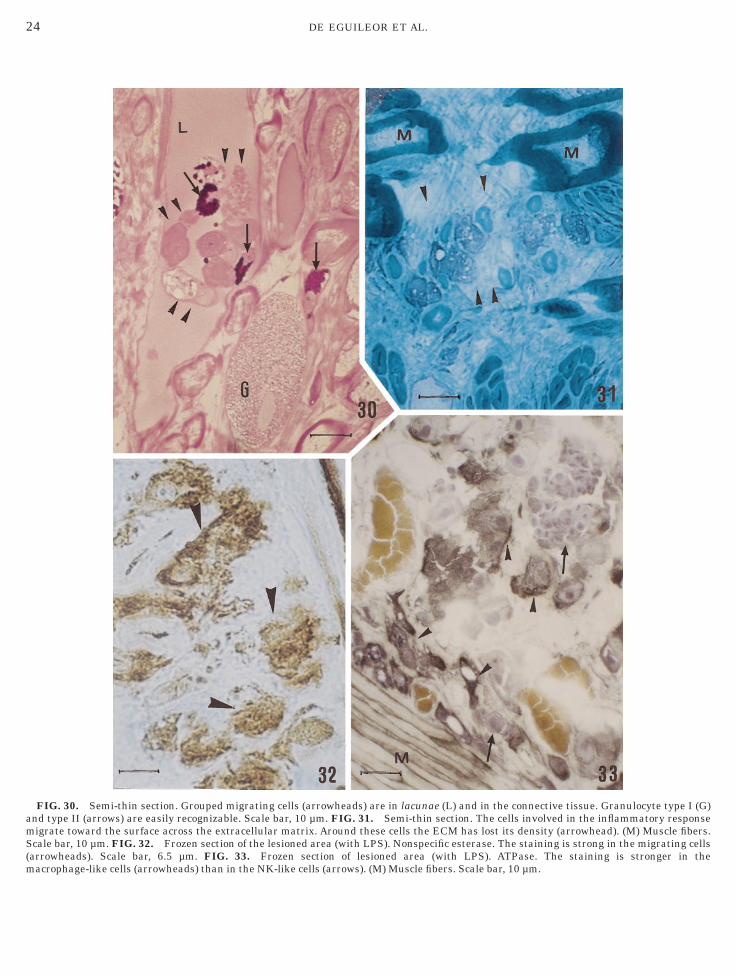
All cell types were generally visible in blood vessels,n lacunae (Fig. 30), and in connective tissue (Fig. 31).owever, when cells migrate, they increase numeri-
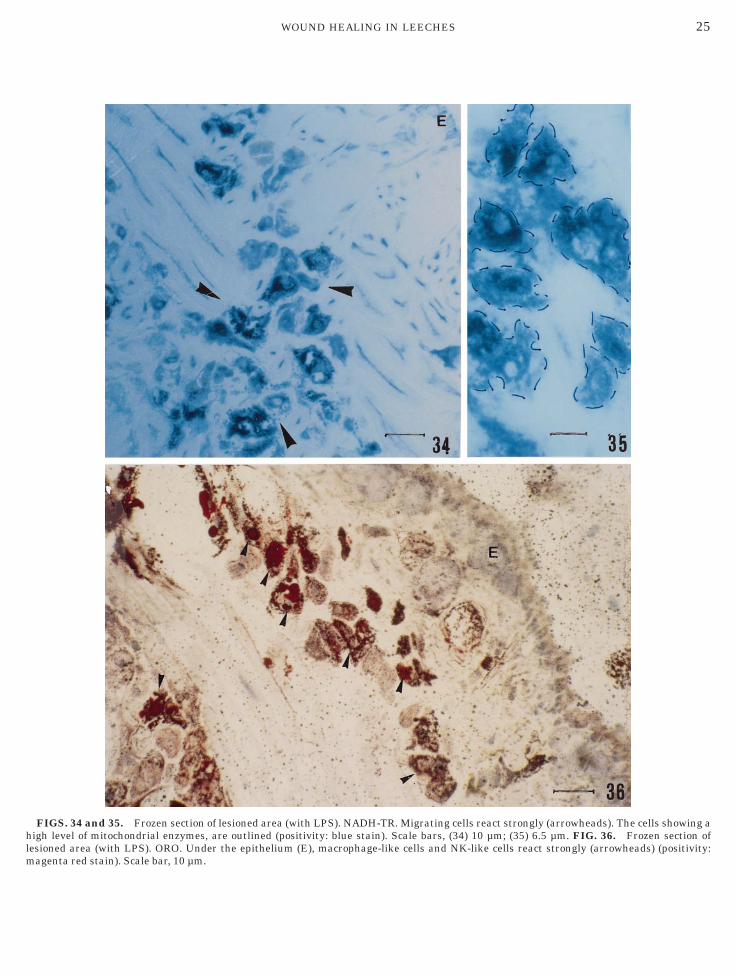
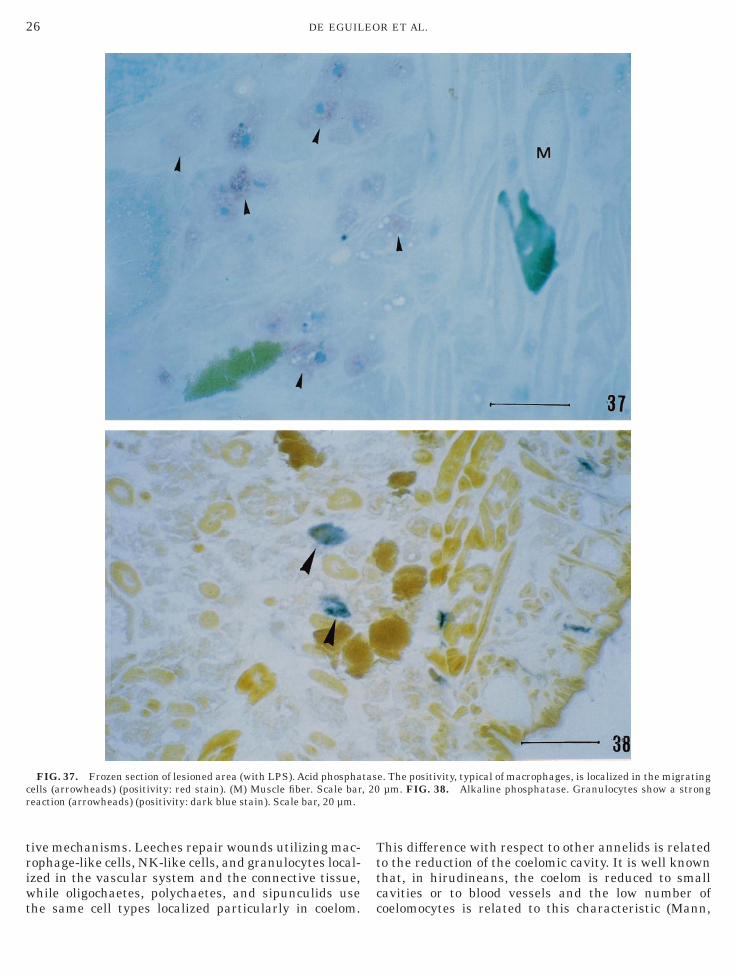
ally in the connective tissue of the lesioned areas (Figs.–4). Using histochemistry, we observed several charac-eristics of the different cell types. The first and secondell type (macrophage-like cells and NK-like cells) weretrongly positive for nonspecific esterase (Fig. 32),TPase (Fig. 33), NADH-TR (Figs. 34 and 35), ORO
Fig. 36), acid and alkaline phosphatase (Figs. 37 and
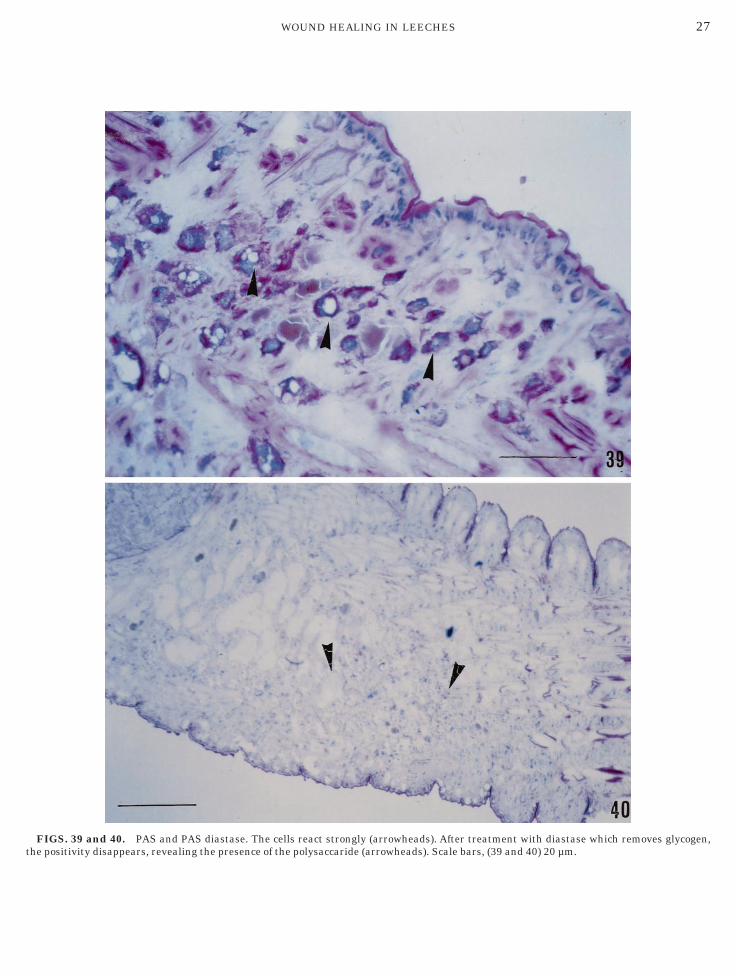
8), and PAS (Figs. 39 and 40). Granulocytes were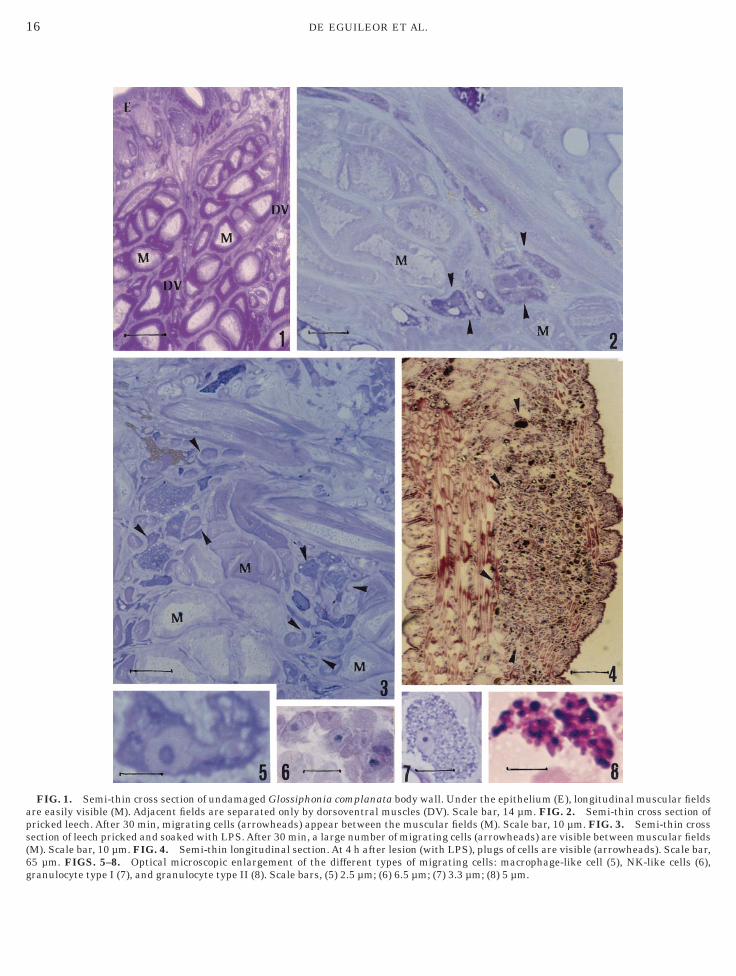
aps(6g m; (6) 6.5 µm; (7) 3.3 µm; (8) 5 µm.
16 DE EGUILEOR ET AL.
FIG. 1. Semi-thin cross section of undamaged Glossiphonia compre easily visible (M). Adjacent fields are separated only by dorsovenricked leech. After 30 min, migrating cells (arrowheads) appear betwection of leech pricked and soaked with LPS. After 30 min, a large nuM). Scale bar, 10 µm. FIG. 4. Semi-thin longitudinal section. At 4 h5 µm. FIGS. 5–8. Optical microscopic enlargement of the differeranulocyte type I (7), and granulocyte type II (8). Scale bars, (5) 2.5 µ
lanata body wall. Under the epithelium (E), longitudinal muscular fieldstral muscles (DV). Scale bar, 14 µm. FIG. 2. Semi-thin cross section ofeen the muscular fields (M). Scale bar, 10 µm. FIG. 3. Semi-thin cross
mber of migrating cells (arrowheads) are visible between muscular fieldsafter lesion (with LPS), plugs of cells are visible (arrowheads). Scale bar,nt types of migrating cells: macrophage-like cell (5), NK-like cells (6),
wmc8
17WOUND HEALING IN LEECHES
FIG. 9. SEM. Area of lesion after 10 min. Macrophage-like cellsound. (E) Epithelium. Scale bar, 40 µm. FIG. 10. SEM. Detaiacrophage-like cell (m) to the epithelial cell (E) is visible. Scale bar,
ells (m) link the epithelial cells (E). Macrophage-like cells are in contaµm.
with pseudopodia (arrowheads) bridge the epithelial edges, closing thel of the encircled area of Fig. 9. The attached zone (arrowheads) of4 µm. FIG. 11. SEM. Detail showing the area where macrophage-likect both with each other (arrowheads) and with epithelial cells. Scale bar,

pa
iimtpntlb1MidN
baIcgbwpdf
pltdmtil
1 arr
18 DE EGUILEOR ET AL.
ositive for toluidine blue (Fig. 24), and acid andlkaline phosphatase (Figs. 37 and 38).
DISCUSSION
This investigation was concerned with characteriz-ng the cells responsible for wound healing followingnjury in leeches. Shortly after injury, numerous cells
igrated from botryoidal tissue and from the circula-ory system, reaching the healing area and forming alug. Using morphological and histochemical tech-iques, we observed and characterized three main cellypes, macrophage-like cells, NK-like cells, and granu-ocytes, which share similar features with their verte-rate counterparts (Grossi et al., 1982; Meseguer et al.,994; Mikkelsen and Rumessen, 1992; Milicevic andilicevic, 1984). Macrophage-like cells, which migrate
n connective tissue, showed typical extensions of pseu-opodia (Figs. 14–20). Both macrophage-like cells and
FIG. 12. SEM. In the wound area, after about 15 min, numero3. SEM. Detail of Fig. 12. NK-like cells are roundish or oval (white
K-like cells showed intense metabolism, as revealed i
y histochemical analysis, as well as strong lysosomalctivity (Figs. 32–40). The two granulocytic types (I andI) were different in form and content. It was possible toompare only type I (Figs. 24–26), containing smallranules, with neutrophilic granulocytes of verte-rates. Leech granulocytes, like those of vertebrates,ere positive for toluidine blue and for alkaline phos-hatase. The granulocyte type II (Figs. 27–29), neverescribed before, was typical of the leeches but theirunction is unknown.
Macrophage-like cells were the first cells that ap-eared in the lesioned area, reducing the aperture byinking the wound edges with pseudopodia. Initially,he macrophage-like cells acted as bridges connectingirectly with epithelial cells or indirectly with otheracrophage-like cells (Figs. 9–11). Subsequently, addi-
ional macrophage-like cells, granulocytes, and round-sh NK-like cells crowded under the surface, filling theesion and forming a plug (Figs. 3, 4, 12, and 13). It is
NK-like cells (arrowheads) fill the lesion. Scale bar, 10 µm. FIG.owheads). Scale bar, 7 µm.
us
nteresting to compare these events, which occurred
ac(
19WOUND HEALING IN LEECHES
FIGS. 14–20. TEM. After 30 min from the moment of lesion (with LPS) macrophage-like cells moved in the connective tissue (Figs. 14, 15,nd 18–20) toward the surface body wall and showed ruffled surfaces (arrowheads) and projections of variable thickness (arrows). In theytoplasm, mitochondria (mi) (Figs. 14 and 20), a well-developed endoplasmic reticulum (ER) (Figs. 14, 17, and 20), and phagolysosomes (Ph)Figs. 15, 16, 18, and 19) were visible. Scale bars, (14) 1.5 µm; (15 and 16) 2 µm; (17) 0.7 µm; (18 and 19) 2.2 µm; (20) 1 µm.
sanmla
Ttm
b
20 DE EGUILEOR ET AL.
hortly after the lesion, with those events occurringfter silver nitrate burns, for which Huguet and Moli-as (1994, 1996) described the formation and develop-ent of a pseudoblastema during wound healing in
eeches. Those authors attributed the entire process to
FIGS. 18–
single connective tissue cell type, the vasocentral cell. (
he newly formed pseudoblastema generates contrac-ions and subsequent wound closure by means ofyofibroblasts like those that occur in vertebrates.We offer several explanations for the differences
etween our results and those of Huguet and Molinas
Continued
20—1994, 1996). Our data referred only to changes occur-
raoua
d(bi
e ). S
21WOUND HEALING IN LEECHES
ing after relatively short time periods (10 min, 30 min,nd 4 h) following injury. In time periods longer thanurs, the mechanism may be different and dependentpon myofibroblasts that are involved in the repair. As
FIGS. 21–23. TEM. NK-like cells, migrating toward the surfaxtracellular matrix around the cells has a ‘‘loose weave’’ (arrowheads
n alternative possibility, the response may vary with b
ifferent types of lesion. We made holes with a needle1.20 3 40 mm) penetrating the entire thickness of theody wall; the wound was deep but not wide like in anncision. In large wounds, contraction by myofibro-
show a large nucleus (N) and some cytoplasmic organelles. Thecale bars, (21 and 22) 2.5 µm; (23) 1.8 µm.
ce,
lasts may be necessary to bring the edges into proxim-
ilt
pi
smt(t
sS
22 DE EGUILEOR ET AL.
ty, while in our experiments, the work of macrophage-ike cells might be sufficient to link the two surfaces ofhe lesion.
We observed an enhancement of the response in theresence of LPS, which in mammalian cells plays a role
FIG. 24. Semi-thin cross section of lesioned area. Many granuperficially and among muscles (M). Scale bar, 28 µm. FIGS. 25 ancale bars, (25) 1 µm; (26) 0.5 µm.
n immune responses and activation of the complement s
ystem. In leeches, we observed an increase in celligration (different cell types became activated) from
he center toward the surface of the wounded bodyFigs. 3 and 4). Moreover, the strength of response fromhe botryoidal tissue to the wounded region increased
ytes (type I), positive to toluidine blue (arrowheads), are visible6. TEM. A large granulocyte (type I) full of small round granules.
ulocd 2
ignificantly with increased time elapsed from the
odiPa
laa
c ; (2
23WOUND HEALING IN LEECHES
ccurrence of the lesion. Supporting our findings, evi-ence from studies of sipunculids has shown that anncrease in phagocytic activity is enhanced by LPS andMA following preincubation of coelomocytes (Blanco et
FIGS. 27–29. TEM. Granulocyte (type II) with large dark granulell membrane is lined with microtubules (mt). Scale bars, (27) 1.5 µm
l., 1997). a
In conclusion, cellular inflammatory activity ineeches occurs rapidly, similar to that in other annelidsnd the related sipunculids (Cooper et al., 1995; Suzukind Cooper, 1995; Cossarizza et al., 1995; Quaglino et
howing complementary profiles (arrowheads). The inner part of the8 and 29) 0.5 µm.
es s
l., 1996), and it may be associated with early regenera-
amS(m
24 DE EGUILEOR ET AL.
FIG. 30. Semi-thin section. Grouped migrating cells (arrowheads) are in lacunae (L) and in the connective tissue. Granulocyte type I (G)nd type II (arrows) are easily recognizable. Scale bar, 10 µm. FIG. 31. Semi-thin section. The cells involved in the inflammatory responseigrate toward the surface across the extracellular matrix. Around these cells the ECM has lost its density (arrowhead). (M) Muscle fibers.cale bar, 10 µm. FIG. 32. Frozen section of the lesioned area (with LPS). Nonspecific esterase. The staining is strong in the migrating cells
arrowheads). Scale bar, 6.5 µm. FIG. 33. Frozen section of lesioned area (with LPS). ATPase. The staining is stronger in theacrophage-like cells (arrowheads) than in the NK-like cells (arrows). (M) Muscle fibers. Scale bar, 10 µm.
hlm
25WOUND HEALING IN LEECHES
FIGS. 34 and 35. Frozen section of lesioned area (with LPS). NADH-TR. Migrating cells react strongly (arrowheads). The cells showing aigh level of mitochondrial enzymes, are outlined (positivity: blue stain). Scale bars, (34) 10 µm; (35) 6.5 µm. FIG. 36. Frozen section of
esioned area (with LPS). ORO. Under the epithelium (E), macrophage-like cells and NK-like cells react strongly (arrowheads) (positivity:agenta red stain). Scale bar, 10 µm.
triwt
Tttc
cr
26 DE EGUILEOR ET AL.
ive mechanisms. Leeches repair wounds utilizing mac-ophage-like cells, NK-like cells, and granulocytes local-zed in the vascular system and the connective tissue,hile oligochaetes, polychaetes, and sipunculids use
FIG. 37. Frozen section of lesioned area (with LPS). Acid phosphaells (arrowheads) (positivity: red stain). (M) Muscle fiber. Scale bareaction (arrowheads) (positivity: dark blue stain). Scale bar, 20 µm.
he same cell types localized particularly in coelom. c
his difference with respect to other annelids is relatedo the reduction of the coelomic cavity. It is well knownhat, in hirudineans, the coelom is reduced to smallavities or to blood vessels and the low number of
e. The positivity, typical of macrophages, is localized in the migratingµm. FIG. 38. Alkaline phosphatase. Granulocytes show a strong
tas, 20
oelomocytes is related to this characteristic (Mann,
t
27WOUND HEALING IN LEECHES
FIGS. 39 and 40. PAS and PAS diastase. The cells react strongly (arrowheads). After treatment with diastase which removes glycogen,
he positivity disappears, revealing the presence of the polysaccaride (arrowheads). Scale bars, (39 and 40) 20 µm.
1crc
B
C
C
C
C
D
G
H
H
H
H
L
L
M
M
M
M
M
M
P
Q
S
S
28 DE EGUILEOR ET AL.
962; Sawyer, 1986). For this reason, although oligo-haetes and polychaetes can repair any type of lesion in aelatively short time by means of a flow of cells from theoelom, leeches do not survive amputation or large wounds.
REFERENCES
lanco, G. A. C., Escalada, A. M., Alvarez, E., and Hajos, S. 1997.LPS-induced stimulation of phagocytosis in the Sipunculan wormThemiste petricola: Possible involvement of human CD14, CD11Band CD11C cross-reactive molecules. Dev. Comp. Immunol. 21,349–362.ooper, E. L., Cossarizza, A., Suzuki, M. M., Salvioli, S., Capri, M.,Quaglino, D., and Franceschi, C. 1995. Autogeneic but not alloge-neic earthworm effector coelomocytes kill the mammalian tumorcell target K562. Cell. Immunol. 166, 113–122.
ooper, E. L. 1996. Earthworm immunity. Progr. Mol. Subcell. Biol.15, 10–45.
ornec, J. P. 1984. Modifications ultrastructurales apres amputationdans le territoire de regeneration posterieure de l’Hirudinee Ryn-chobdelle Helobdella stagnalis. Arch. Anat. Microsc. 73, 269–289.ossarizza, A., Cooper, E. L., Suzuki, M. M., Salvioli, S., Capri, M.,Gri, G., Quaglino, D., and Franceschi, C. 1996. Earthworm leuko-cytes that are not phagocytic and cross-react with several humanepitopes can kill human tumor cell lines. Exp. Cell Res. 224,174–182.ubowitz, V., and Brooke, M. H. 1973. ‘‘Muscle Biopsy: A ModernApproach.’’ Saunders, London.rossi, C. E., Cadoni, A., Zicca, A., Leprini, A., and Ferrarini, M.1982. Large granular lymphocytes in human peripheral blood:Ultrastructural and cytochemical characterization of the granules.Blood 59, 277–283.uguet, G., and Molinas, M. 1988. Estudi estrutural del proces deregeneracio de ferides a Hirudo medicinalis. Sci. Gerund. 14,123–132.uguet, G., and Molinas, M. 1992. Changes in epithelial cells in Hirudomedicinalis during wound healing. J. Invertebr. Pathol. 59, 11–17.uguet, G., and Molinas, M. 1994. The pseudoblastema in the woundhealing process on the leech Hirudo medicinalis L. (Hirudinea):Changes in cell junctions. J. Exp. Zool. 269, 23–36.
uguet, G., and Molinas, M. 1996. Myofibroblast-like cells andwound contraction in leech wound healing. J. Exp. Zool. 275,308–316.
e Gore, R. S., and Sparks, A. L. 1971. Repair of body incision in theRhynchobdellid leech Piscicola salmositica. J. Invertebr. Pathol.18, 40–45.
e Gore, R. S., and Sparks, A. L. 1973. Repair of body wall burns inthe Rhyncobdellid leech Piscicola salmositica. J. Invertebr. Pathol.22, 298–299.ann, K. H. 1962. ‘‘Leeches (Hirudinea): Their Structure, Physiology,Ecology and Embryology.’’ Pergamon, Oxford.eseguer, J., Lopez-Ruiz,A., andAngles Esteban, M. 1994. Cytochemi-cal characterization of leukocytes from the seawater teleost, gilt-head seabream (Sparus aurata L.) Histochemistry 102, 37–44.ikkelsen, H. B., and Rumessen, J. J. 1992. Characterization ofmacrophage-like cells in the external layers of human small andlarge intestine. Cell Tissue Res. 270, 273–279.ilicevic, N. M., and Milicevic, Z. J. 1984. Enzyme-histochemicalcharacterization of macrophages in the rat thymus, with specialreferences to metallophilic cells of corticomedullary zone. J. Leuko-cyte Biol. 36, 761–769.olinas, M., Dalmau, J., and Huguet, G. 1988. Cicatrizacio de lesferides per cauteritzacio del tegumento d’Hirudo medicinalis L.Sci. Gerund. 14, 113–122.oore, R. D., Mumaw, V., and Shonberg, M. D. 1960. Opticalmicroscopy of ultrathin tissue sections. J. Ultrastruct. Res. 4,113–116.
orchet-Hennere, H., Dugimont, T., and Fischer, A. 1992. Naturalkiller cells in a lower invertebrate, Nereis diversicolor. Eur. J. CellBiol. 58, 99–207.uaglino, D., Cooper, E. L., Salvioli, S., Capri, M., Suzuki, M.,Ronchetti, I. P., Franceschi, C., and Cossarizza, A. 1996. Earth-worm coelomocytes in vitro: Cellular features and ‘‘granuloma’’formation during cytofryuvtoxic activity against the mammaliantumor cell target K562. Eur. J. Cell Biol. 70, 278–288.
awyer, R. T. 1986. ‘‘Leech Biology and Behaviour. 1: Anatomy,Physiology and Behaviour.’’ Oxford Science, Oxford.
uzuki, M. M., and Cooper, E. L. 1995. Killing of intrafamilialleukocytes by earthworm effector cells. Immunol. Lett. 44, 45–49.




















