
Phylogenetic relationships of Mexican minnows of the genus Notropis (Actinopterygii, Cyprinidae):...
16
Biological Journal of the Linnean Society , 2003, 80 , 323–337. With 4 figures © 2003 The Linnean Society of London, Biological Journal of the Linnean Society, 2003, 80 , 323–337 323 Blackwell Science, LtdOxford, UKBIJBiological Journal of the Linnean Society0024-4066The Linnean Society of London, 2003? 2003 802 323337 Original Article PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO S. SCHÖNHUTH and I. DOADRIO * Corresponding author. E-mail: [email protected] Phylogenetic relationships of Mexican minnows of the genus Notropis (Actinopterygii, Cyprinidae) SUSANA SCHÖNHUTH* and IGNACIO DOADRIO Museo Nacional de Ciencias Naturales. C.S.I.C., Dpto. Biodiversidad y Biología Evolutiva, José Gutiérrez Abascal 2, Madrid 28006, Spain Received 27 August 2002; accepted for publication 2 April 2003 We conducted phylogenetic analyses based on complete mitochondrial cytochrome b gene sequences among southern and central Mexican cyprinid species, included in the genera Notropis and Hybopsis . In addition 15 northern species of the genera Notropis and Hybopsis were included in the analyses in order to place the Mexican species into a larger phylogenetic framework. The phylogenetic relationships supported the existence of five major clades: (1) including species of the subgenus Alburnops of the genus Notropis plus N. shumardi ; (2) species of the subgenus Notropis ; (3) species of the genus Hybopsis ; (4) species of the N. texanus + N. volucellus species group of the genus Notropis ; (5) Mexican endemic species of the genus Notropis plus the genus Yuriria . Previous phylogenetic inferences based on morphological characters resolved the Mexican minnows analysed as N. sallaei , N. calientis, N. boucardi and Y. alta , non-monophyletic. According to our cytochrome b evidence all Mexican minnows of the genera Notropis and Yuriria formed a monophyletic group with respect to the northern species of the genera Notropis and Hybopsis . Within the Mexican clade, three well-supported clades were identified: the first included the closely related species N. moralesi and N. boucardi , which occur in three independent drainages of south Mexico; the second consisted of two different lineages, N. imeldae and an undescribed species of Notropis , inhabiting two independent drainages of south Mexico; the third comprised two central Mexican Notropis species ( N. calientis and N. sallaei ) and the Y. alta populations. Based on this study and pending a more extensive taxonomic revision of the genus Notropis , we adopt the conser- vative criterion of considering all Notropis species from southern and central Mexico examined, including Y. alta, as belonging to the genus Notropis . © 2003 The Linnean Society of London, Biological Journal of the Linnean Society , 2003, 80 , 323–337. ADDITIONAL KEYWORDS: Bayesian inference – cyprinids – cytochrome b – Mexico – mitochondrial DNA – trans-Mexican volcanic belt. INTRODUCTION With close to 100 species, the genus Notropis s.l. is among the most species-rich genera of Nearctic fresh- water fishes. It occurs in rivers from Canada and Alaska to southern Mexico. The taxonomy and phylo- genetic relationships of the genus in the United States and Canada have been the subject of extensive research (Buth, 1979; Stein, Rogers & Cashner. 1985; Dimmick, 1987; Mayden, 1989, 1991; Coburn & Cav- ender, 1992; Warren, Burr & Grady, 1994; Simons & Mayden, 1999; Bielawski & Gold, 2001; Raley & Wood, 2001). Unfortunately, few studies have focused on Mexican species south of the Río Bravo. Notropis is currently represented in Mexico by at least 20 species (Burr & Mayden, 1981; Espinosa, Gaspar & Fuentes, 1993), but only the following five recognized species are distributed across central and southern Mexico: N. boucardi (Günther, 1868) , N. sallaei (Günther, 1868), N. calientis Jordan & Sny- der, 1900, N. moralesi De-Buen, 1955 and N. imeldae Cortés, 1966. Taxonomy of these species has been con- troversial. For example, N. sallaei was included in eight different genera and 16 nominal species. Cher- noff & Miller (1981) considered it as a highly variable taxon ‘provisionally’ assigned to Notropis , which was followed by different authors (Chernoff & Miller, 1986; Espinosa et al ., 1993), or it was considered as several subspecies (Vallejo de Aquino, 1988), while some authors consider it to be the only member of the
Transcript of Phylogenetic relationships of Mexican minnows of the genus Notropis (Actinopterygii, Cyprinidae):...

Biological Journal of the Linnean Society
2003
80
323ndash337 With 4 figures
copy 2003 The Linnean Society of London
Biological Journal of the Linnean Society
2003
80
323ndash337
323
Blackwell Science LtdOxford UKBIJBiological Journal of the Linnean Society0024-4066The Linnean Society of London 2003 2003802323337Original Article
PHYLOGENETIC RELATIONSHIPS OF
NOTROPIS
IN MEXICOS SCHOumlNHUTH and I DOADRIO
Corresponding author E-mail mcns137mncncsices
Phylogenetic relationships of Mexican minnows of the genus
Notropis
(Actinopterygii Cyprinidae)
SUSANA SCHOumlNHUTH and IGNACIO DOADRIO
Museo Nacional de Ciencias Naturales CSIC Dpto Biodiversidad y Biologiacutea Evolutiva Joseacute Gutieacuterrez Abascal 2 Madrid 28006 Spain
Received 27 August 2002 accepted for publication 2 April 2003
We conducted phylogenetic analyses based on complete mitochondrial cytochrome
b
gene sequences among southernand central Mexican cyprinid species included in the genera
Notropis
and
Hybopsis
In addition 15 northern speciesof the genera
Notropis
and
Hybopsis
were included in the analyses in order to place the Mexican species into a largerphylogenetic framework The phylogenetic relationships supported the existence of five major clades (1) includingspecies of the subgenus
Alburnops
of the genus
Notropis
plus
N shumardi
(2) species of the subgenus
Notropis
(3)species of the genus
Hybopsis
(4) species of the
N texanus
+
N volucellus
species group of the genus
Notropis
(5)Mexican endemic species of the genus
Notropis
plus the genus
Yuriria
Previous phylogenetic inferences based onmorphological characters resolved the Mexican minnows analysed as
N sallaei
N calientis N boucardi
and
Y alta
non-monophyletic According to our cytochrome
b
evidence all Mexican minnows of the genera
Notropis
and
Yuriria
formed a monophyletic group with respect to the northern species of the genera
Notropis
and
Hybopsis
Within theMexican clade three well-supported clades were identified the first included the closely related species
N moralesi
and
N boucardi
which occur in three independent drainages of south Mexico the second consisted of two differentlineages
N imeldae
and an undescribed species of
Notropis
inhabiting two independent drainages of south Mexicothe third comprised two central Mexican
Notropis
species (
N calientis
and
N sallaei
) and the
Y alta
populationsBased on this study and pending a more extensive taxonomic revision of the genus
Notropis
we adopt the conser-vative criterion of considering all
Notropis
species from southern and central Mexico examined including
Y alta
asbelonging to the genus
Notropis
copy 2003 The Linnean Society of London
Biological Journal of the Linnean Society
2003
80
323ndash337
ADDITIONAL KEYWORDS
Bayesian inference ndash cyprinids ndash cytochrome
b
ndash Mexico ndash mitochondrial DNA
ndash trans-Mexican volcanic belt
INTRODUCTION
With close to 100 species the genus
Notropis sl
isamong the most species-rich genera of Nearctic fresh-water fishes It occurs in rivers from Canada andAlaska to southern Mexico The taxonomy and phylo-genetic relationships of the genus in the United Statesand Canada have been the subject of extensiveresearch (Buth 1979 Stein Rogers amp Cashner 1985Dimmick 1987 Mayden 1989 1991 Coburn amp Cav-ender 1992 Warren Burr amp Grady 1994 Simons ampMayden 1999 Bielawski amp Gold 2001 Raley amp Wood2001) Unfortunately few studies have focused onMexican species south of the Riacuteo Bravo
Notropis
is currently represented in Mexico by atleast 20 species (Burr amp Mayden 1981 EspinosaGaspar amp Fuentes 1993) but only the following fiverecognized species are distributed across centraland southern Mexico
N boucardi
(Guumlnther 1868)
N sallaei
(Guumlnther 1868)
N calientis
Jordan amp Sny-der 1900
N moralesi
De-Buen 1955 and
N imeldae
Corteacutes 1966 Taxonomy of these species has been con-troversial For example
N sallaei
was included ineight different genera and 16 nominal species Cher-noff amp Miller (1981) considered it as a highly variabletaxon lsquoprovisionallyrsquo assigned to
Notropis
which wasfollowed by different authors (Chernoff amp Miller1986 Espinosa
et al
1993) or it was considered asseveral subspecies (Vallejo de Aquino 1988) whilesome authors consider it to be the only member of the
324
S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London
Biological Journal of the Linnean Society
2003
80
323ndash337
genus
Aztecula
(Mayden 1991 Coburn amp Cavender1992) The remaining Mexican species analysed
N boucardi
N moralesi
N imeldae
and
N calientis
have been assigned to the genus
Notropis
(Chernoff ampMiller 1986 Vallejo 1988 Espinosa
et al
1993) or tothe genus
Hybopsis
(Mayden 1991)
N calientis
shows a high degree of population structure and hasbeen recognized as a species complex (Chernoff ampMiller 1986) On the other hand
Yuriria
was consid-ered as a subgenus of the genus
Hybopsis
(Bailey1951) until Mayden (1989) elevated
Yuriria
to genericstatus
Based on morphology two principal phylogenetichypotheses placed the species analysed in differentphylogenetic clades Coburn amp Cavenderrsquos (1992)hypothesis considers the genus
Notropis
(
sl
includ-ing genus
Hybopsis
) as a member of the Shiner cladeand the genus
Yuriria
as a member of the Westernclade Maydenrsquos (1989) hypothesis places the Mexicanspecies analysed in different clades
N boucardi
and
N calientis
as members of the genus
Hybopsis
andthe genus
Yuriria
in a different clade both of themincluded in the open posterior miodome (OPM) cladewhile
N sallaei
was excluded from this principalclade Mayden (1989) considers the genus
Notropis
tobe polyphyletic (comprising different subgenus andspecies groups)
Mitochondrial DNA sequences provide an objectiveframework for the analysis of population processesand evolutionary patterns (Bermingham amp Martin1998) and have been useful in studies of phylogeneticrelationships of several groups of North American cyp-rinids (Dowling amp Naylor 1997 Simons amp Mayden1997 1998 1999) In particular cytochrome
b
hasbeen shown to be an appropriate gene for inferringphylogenetic relationships in several phylogeneticstudies on freshwater fishes (Martin amp Bermingham1998 Zardoya amp Doadrio 1999 Bielawski amp Gold2001 Machordom amp Doadrio 2001) Our study is thefirst to address relationships among Mexican
Notropis
using mtDNA sequences and to provide a phylogeneticframework with the northern species of the genera
Notropis
and
Hybopsis
The purpose of this work was thus to use the com-
plete sequence of the cytochrome
b
gene to infer phy-logenetic affinities of Mexican endemic species of thegenus
Notropis
with species that represent differentsubgenera or species groups of the genus
Notropis
from the USA and Canada and the genus
Hybopsis
and to test previous hypotheses of paraphyletic lin-eages within Mexican endemic species In particularwe used molecular sequence data from 30 geographi-cally distinct populations of seven Mexican endemicspecies of
Notropis
to test evolutionary hypothesesregarding their patterns of diversification in centraland southern Mexico
MATERIAL AND METHODS
The taxa sequenced in this study were previouslyassigned to four different genera (Mayden 1991) ndash
Hybopsis
Aztecula
Notropis
and
Yuriria
ndash and rep-resent the different clades proposed by Mayden (1989)and Coburn amp Cavender (1992) Due to the controver-sial taxonomy of the Mexican species analysed wehave followed the classification of Eschmeyer (httpwwwfishbaseorg) We sequenced 34 specimens of 30populations representing seven Mexican endemic spe-cies of minnows (
N moralesi
N boucardi
N imeldae
N sallaei
N calientis Y alta
and one undescribedspecies of the genus
Notropis
referred to here as
N
sp 1) distributed over central and southern Mexicoplus three species of the genus
Notropis
from Canada(
N rubellus
N anogenus
and
N heterodon
obtainedfrom the Royal Ontario Museum) Seventeen cyprinidsequences were obtained from GenBank and includedin the phylogenetic analyses
N atherinoides
(AF352272)
N volucellus
(AF352268)
N boops
(AF352261)
N suttkusi
(AF352287)
N jemezanus
(AF352277)
N amabilis
(AF352269)
N stilbius(AF352286) N shumardi (AF117200) N potteri(AF117192) N blennius (AF117171) Hybopsisamblops (AF117153) Hybopsis winchelli (AF117165)Cyprinella spiloptera (L07753) Pteronotropissignipinnis (AF261230) Moapa coriacea (AF452075)Barbus sclateri (AF334083) and Tinca tinca (Y10451)The last three taxa were used as outgroups
DNA SOURCES
Fish samples were collected by electrofishing in thesouthern and central region of Mexico from Sierrade Miahuatlaacuten in Oaxaca at about 16degN latitude tothe Lerma drainage in Guanajuato State some21degN These collections (Table 1) cover the Atlanticand the Pacific slopes and represent five riverbasins Papaloapan Balsas Atoyac Lerma andPaacutenuco (Fig 1) The specimens analysed correspondto populations inhabiting the entire distributionarea known for N moralesi N boucardi N imeldaeand N sp 1 We obtained N sallaei specimens fromthe Paacutenuco and Lerma drainages N calientis andY alta were fished from Lerma river drainage andadjacent lakes
DNA EXTRACTION
Total DNA was extracted from muscle tissue Frozenpieces of tissue were ground in liquid N2 and homog-enized in 600 microL extraction buffer (NaCl 014 M mag-nesium acetate 15 mM KCl 5 mM 1 SDS) Theresulting homogenate was extracted using standardproteinase Kphenolndashchloroform protocols (Sambrook
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 325
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 1
S
peci
men
s u
sed
in t
his
stu
dy a
nd
loca
lity
in
form
atio
n
Pop
ula
tion
de
sign
atio
nS
peci
es
MN
CN
ID
Dra
inag
eR
iver
Loc
alit
yS
tate
Gen
Ban
k R
ef
1 F
ran
cisc
oN
otro
pis
imel
dae
353
Ato
yac
San
Fra
nci
sco
San
Pab
lo C
oatl
an O
axac
aA
F46
9131
2 G
ruta
sN
im
eld
ae 1
095
Ato
yac
De
las
Gru
tas
rive
r (S
prin
g)S
ola
de V
ega
Oax
aca
AF
4691
323
Sab
ino
N i
mel
dae
113
8A
toya
cA
gua
del
Sab
ino
Sol
a de
Veg
a O
axac
aA
F46
9130
4 V
erde
N m
oral
esi
97A
toya
cG
ran
de-V
erde
Noc
hix
tlan
Oax
aca
AF
4691
535
Bra
voN
mor
ales
i 10
67A
toya
cB
ravo
San
Ju
an T
eita
Tla
xiac
o O
axac
aA
F46
9152
6 I
gual
ites
N m
oral
esi
5059
Bal
sas
Igu
alit
esT
lapa
Gu
erre
roA
F46
9151
7 C
oico
yan
N m
oral
esi
1086
Bal
sas
Coi
coya
n d
e la
s F
lore
sC
oico
yan
Ju
xtla
hu
aca
Oax
aca
AF
4691
458
Ch
iqu
ito
N m
oral
esi
133
Bal
sas
Ch
iqu
ito
(Spr
ing)
Tla
xiac
o O
axac
aA
F46
9150
9 S
alad
oN
mor
ales
i 99
0B
alsa
sS
alad
oH
uaj
uap
an O
axac
aA
F46
9156
10 I
sabe
lN
mor
ales
i 19
99B
alsa
sS
ta Isa
bel
Ch
ilap
a G
uer
rero
AF
4691
4811
Mix
teco
N m
oral
esi
5048
Bal
sas
Tec
oloy
anX
och
ihu
ehu
etla
n G
uer
rero
AF
4691
4912
Mix
teco
-1N
mor
ales
i 10
6B
alsa
sM
ixte
coA
sun
cioacuten
Tla
xiac
o O
axac
aA
F46
9144
13 T
elol
oapa
nN
mor
ales
i 20
09B
alsa
sT
elol
oapa
nT
elol
oapa
n G
uer
rero
AF
4691
4714
Hon
doN
mor
ales
i 91
4P
apal
oapa
nH
ondo
Con
cepc
ioacuten
Bu
ena
Vis
ta O
axac
aA
F46
9157
15 G
ran
de
N m
oral
esi
150
154
Pap
aloa
pan
Gra
nde
Tep
eln
eme
de M
orel
os O
axac
aA
F46
9154
ndash55
16 A
mac
uya
cN
mor
ales
i 34
67B
alsa
sA
mac
uya
cG
ruta
s de
Cac
ahoa
mil
pa G
uer
rero
AF
4691
4617
Pol
lo
N b
ouca
rdi
3487
Bal
sas
del
Pol
lo C
ol L
agu
nil
la
Cu
ern
avac
a M
orel
osA
F46
9158
18 H
uel
lapa
nN
bou
card
i 34
74B
alsa
sH
uel
lapa
n l
agoo
nE
l T
exal
Cu
ern
avac
a M
orel
osA
F46
9159
19 T
axin
gu
N s
alla
ei 3
125
Paacuten
uco
Tax
ingu
Dam
Acu
lco
Meacutex
ico
DF
A
F46
9135
20 A
lmoy
aN
sal
laei
351
7L
erm
aA
lmoy
a de
l R
iacuteo l
agoo
nA
lmoy
a de
l R
iacuteo M
eacutexic
o D
F
AF
4691
3621
SM
Cu
evas
N s
p 4
17 4
19B
alsa
sS
an M
igu
el C
uev
as (
Spr
ing)
Juxt
lah
uac
a O
axac
aA
F46
9133
ndash34
22 P
iru
les
N c
alie
nti
s 33
26L
erm
aP
iru
les
Dam
San
Ju
an R
ayas
Qu
eret
aro
AF
4691
3723
Zac
apu
N c
alie
nti
s 36
663
717
Zac
apu
Zac
apu
lag
oon
Zac
apu
Mic
hoa
caacuten
AF
4691
41ndash4
224
Ch
arco
N c
alie
nti
s 39
18L
erm
aC
har
co d
el I
nge
nio
San
Mig
uel
All
ende
Gu
anaj
uat
oA
F46
9143
25 M
inzi
taN
cal
ien
tis
3813
Cu
itze
oL
a M
inzi
ta D
amL
a M
inzi
ta M
ich
oacaacute
nA
F46
9138
26 S
Mig
uel
N c
alie
nti
s 38
29 3
831
Ler
ma
San
Mig
uel
(S
prin
g)S
an M
igu
el 2
033
m M
ich
oacaacute
nA
F46
9139
ndash40
27 C
eja
Yuri
ria
alta
333
4L
erm
aC
eja
de B
ravo
Dam
La
Cej
a de
Bra
vo Q
uer
etar
oA
F46
9161
28 Z
acap
uY
alt
a 34
49L
erm
aZ
acap
u l
agoo
n o
utl
edP
anin
dicu
aro
Mic
hoa
caacuten
AF
4691
6329
Laj
asY
alt
a 33
70L
erm
aL
ajas
tri
buta
ryS
an M
igu
el A
llen
de G
uan
aju
ato
AF
4691
6230
Min
zita
Y a
lta
3809
Cu
itze
oL
a M
inzi
ta D
amL
a M
inzi
ta M
ich
oacaacute
nA
F46
9160
31 C
anad
aN
ru
bell
us
1703
32G
ran
d R
iver
Bra
ntf
ord
On
tari
o C
anad
aA
F46
9164
32 C
anad
aN
het
erod
on 1
7033
1E
ire
Lak
eR
onde
au P
rovi
nci
al P
ark
On
tari
o C
anad
aA
Y14
0697
33 C
anad
aN
an
ogen
us
1703
36S
t C
lair
Lak
eM
itch
ell
Bay
On
tari
o C
anad
aA
Y14
0698
Nu
mbe
rs a
ssig
ned
to
popu
lati
ons
corr
espo
nd
to t
hos
e of
th
e m
ap (
see
Fig
1)
MN
CN
ID
nu
mbe
rs
nu
mbe
rs o
f vo
uch
er s
peci
men
s de
posi
ted
in t
he
ich
thyo
logy
coll
ecti
on o
f th
e M
use
o N
acio
nal
de
Cie
nci
as N
atu
rale
s (S
pain
) G
enB
ank
Ref
acc
essi
on n
um
ber
for
nu
cleo
tide
seq
uen
ces
depo
site
d in
Gen
Ban
kT
opot
ypes
or
spec
imen
s fo
un
d cl
ose
to t
hei
r te
rra
typi
ca
326 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Fritsch amp Maniatis 1989) The DNA was resuspendedin ultrapure water and used for amplification by PCR
AMPLIFICATION AND CLONING
Combinations of two sets of versatile primers (Zardoyaamp Doadrio 1998) were used in PCR amplifications toisolate two contiguous and overlapping fragments (of660 and 521 bp) that covered the entire cytochrome bgene The PCR reactions were conducted in 25 microl vol-umes (67 mM Tris-HCl pH 83 15 mM MgCl2 04 mM
each primer 1 microl DNA extraction and 1 unit Taq DNApolymerase (Promega)) After an initial denaturationstep at 94degC for 2 min 35 cycles were performed as fol-lows denaturation at 94degC (1 min) annealing at 48degC(1 min) and extension at 72degC (145 min) with a finalextension of 7 min at 72degC
After PCR amplification the DNA was precipitatedcloned into p-GEMT vectors and transformed intocompetent E coli (JM109) Positive clones weresequenced using the FS-Taq Dye Deoxy terminatorcycle sequencing kit (Applied Biosystems Inc) usingan automated DNA sequencer (Applied Biosystems3700) following the manufacturerrsquos instructions TheDNA sequences of both strands were obtained using
M13 universal (forward and reverse) sequencing prim-ers The sequences determined here have been depos-ited in GenBank under accession numbers AF469130ndashAF469164 AY140697 and AY140698
CYTOCHROME b SEQUENCE ANALYSES
Forty-nine complete minnow cytochrome b genesequences (1141 bp) were aligned manually withsequences of N atherinoides H amblops Cyprinellaspiloptera Tinca tinca and Barbus sclateri No ambig-uous alignments or gaps were found All codon posi-tions were included in the phylogenetic analyses
All Mexican specimens representing 30 populationsof seven minnow species were used to determine theirpopulation structure (Table 1) The intrapopulationaldivergence levels were corrected for ancestral poly-morphisms to avoid the effect of inflating divergencebetween phylogroups or species We applied the netsequence divergence correction described by Nei(1987) and the index of nucleotide diversity (π) (Nei ampLi 1979) using DnaSP ver 350 (Rozas amp Rozas 1999)Higher level relationships were inferred using 22 spe-cies of Mexican USA and Canadian NotropisHybop-sis and two species of the phylogenetically related
Figure 1 Localities from which specimens of the genera Notropis and Yuriria were sampled Numbers indicate collectionsample (see Table 1) Drainages are indicated by letters A Atoyac B Balsas C Lerma D Paacutenuco E PapaloapanDistribution range of each species is presented in different grey patterns
115degW
30degN
25deg
20deg
15deg
110deg 105deg 100deg 95deg 0 200 km
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 327
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
genera Cyprinella and Pteronotropis included in theNotropin clade (Phoxinini lineage Shiner clade)(Coburn amp Cavender 1992) The genus Moapa fromthe Western clade Phoxinini was selected as a taxo-nomic related outgroup whereas Tinca was chosen asa more distant outgroup
Phylogenetic trees were recovered using maximumparsimony (MP) neighbour joining (NJ) maximumlikelihood (ML) and Bayesian inference (BI) We usedfour different methods for inferring phylogenies to testthe congruence among methods and compare for thebest approximation for our data There has recentlybeen controversy about the overcredibility of molecu-lar phylogenies obtained with Bayesian inference(Suzuki Glazco amp Nei 2002) while others have sug-gested posterior probability as a better indicator ofstatistical confidence than bootstrap probability (Wil-cox et al 2002) Both bootstrap and posterior proba-bility were included It is well known that MP givesgood results in the Felsenstein inverse zone and MLin both zones when more than 1000 nucleotides areanalysed However accuracy decreases when theinternal nodes are short (Swofford et al 2001) Theanalysis of 1141 nucleotides resulted in short internalnodes within trees so we decided to include the fourmethods of analysis to infer the phylogeny
The MP analysis was performed through heuristicsearches with ten random stepwise additions of taxaMULTREE option and TBR branch swapping For theMP analyses equal weight of transversions (Tv) andtransitions (Ts) or Tv five times the weight of Ts basedon empirical evidence were used For ML BI and NJanalyses we used the hierarchical likelihood ratio test(LRT) implemented in Modeltest V304 (Posada ampCrandall 1998) to find the evolutionary model thatbest fit our data The HKY + I + G model (base fre-quencies A 03294 C 02805 G 01433 T 02469Nst 2 TsTv ratio 56889 Γ 09381 proportion ofinvariable sites 04383) was selected when Mexicanminnow populations were taken into account and theGTR + I + G model (base frequencies A 02853 C03302 G 01125 T 02720 Nst 6 Γ 07619 propor-tion of invariable sites 04915) was selected when onespecimen per species was analysed
All phylogenetic analyses were performed usingPAUP version 40b8 (Swofford 2001) and MR BAYES(Huelsenbeck amp Ronquist 2001) Robustness of theinferred trees was tested by bootstrapping (Felsen-stein 1985) with 100 pseudoreplications for MP and1000 for NJ with 1000 quartet puzzling steps forML and 500 000 generations for Bayesian posteriorprobability
Our two alternative phylogenetic hypotheses forN imeldae-N sp 1 clade were tested using the Shimo-dairandashHasegawa test (Shimodaira amp Hasegawa 1999)as implemented in PAUP
The genetic divergences widely discussed throughthe text were based on the uncorrected p-distances Toavoid confusion between derived divergences based onthe different evolutionary models selected (HKY +I + G and GTR + I + G) we considered more appropri-ate the uncorrected p genetic distances which allowscomparisons among our two data sets and could permita broad comparison with other related taxa thatshowed a different model of evolution
RESULTS
The complete cytochrome b gene was successfullyamplified and sequenced in 37 specimens of the generaNotropis and Yuriria Of the 1141 bp aligned for alltaxa 488 sites were variable and 373 were parsimonyinformative when only Mexican minnow populationswere taken into account Informative characters roseto 387 when one specimen per species for all NorthAmerican minnows analysed was considered
The nucleotide composition in the cytochrome bgene indicates a bias against guanine (see Modeltestparameters in methods) which is particularly strongin third codon positions (9) Most of the variabilityamong cytochrome b sequences was detected at thirdcodon positions No saturation was detected at anycodon position (Fig 2) However separate plots of AndashG
Figure 2 Relationships between uncorrected meansequence divergence (p-distance) and number of transi-tions () and transversions () at all codon positions (A)and at third codon positions (B) in the cytochrome b genefor all pairwise comparisons for Mexican populationsamples
020406080
100120140160180
005 01 015 02p
Cha
nges
020406080
100120140160
005 01 015 02
p
Cha
nges
0
0
A
B
328 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
and CndashT transitional divergences at third codon posi-tions suggested that AndashG transitions changed fasterthan CndashT ones (data not shown)
Pairwise sequence divergence (uncorrected p-distance) among northern species of the genus Notro-pis analysed ranged from 39 (between N blenniusand N potteri) to 155 (N potteri and N volucellus)Among Mexican species net sequence divergenceranged from 39 (between specimens of N moralesiand N boucardi) to 115 (between specimens ofN sallaei and N boucardi) (Table 2) Within Mexicanspecies the mean divergence among specimensrepresenting different populations was lt1 (amongN imeldae specimens and among specimens of geo-graphically close populations of N moralesi) to 3(among specimens of regionally separated populationsof N moralesi) Nucleotide diversity (π) within mostMexican species ranged from 0005 to 0015 Howeverfour specimens from two different sample sites ofN calientis fell outside this range showing over 7sequence divergence and nucleotide diversity up to0046 (Table 2)
Cytochrome b sequences were analysed using fourcommon methods of phylogenetic inference (MP NJML BI) and recovered congruent topologies (Figs 3and 4) Topologies recovered from unweighted andweighted most parsimonious trees were similar butthe weighted analyses yielded a more highly resolvedtopology congruent to the remaining recovered analy-ses When one specimen per species was taken intoaccount in order to place the Mexican species into alarger phylogenetic framework all cyprinids of thegenera Notropis and Hybopsis were grouped togetherwith Pteronotropis and Cyprinella in a basal positionAccording to MP ML NJ and BI all Mexican speciesof the genera Notropis and Yuriria formed a well-supported clade with respect to the remaining speciesof the genus Notropis from the USA and Canada North-ern species of the genus Notropis formed three differ-ent well-supported clades while basal relationshipsamong clades remained unresolved The phylogeneticposition of the genus Hybopsis remained unresolved(Fig 3) Interestingly the genus Yuriria groupedtogether with all Mexican Notropis in all analysesThe robustness of these results was confirmed by highbootstrap values and Bayesian posterior probability
When populations were taken into account all Mex-ican Notropis plus Yuriria clustered together withrespect to the northern Notropis Hybopsis and Cyp-rinella forming a well-supported clade with bootstrapvalues (MP 94 ML 72 NJ 86) and BI (100)(Fig 4) Within this clade four well-supported lin-eages could be distinguished (Fig 4) (A) grouped allspecimens representing populations of N boucardiand N moralesi (B) grouped N imeldae and N sp 1(C) was formed by all specimens of N sallaei and
N calientis (D) included the Y alta group The genusYuriria was grouped with N calientis-N sallaei as sis-ter species using NJ (85) and ML (74) (Figs 3 4)The relative position of the clade N imeldae-N sp 1was different depending on the phylogenetic analysisaccording to MP (79) ML (58) and BI (100) it wasconsidered a sister group of the N moralesi-N boucardi clade whereas it appeared as a sistergroup of the clade (Yuriria (N calientis N sallaei)) inthe NJ analysis (66) (Fig 4) In order to test the twoalternative hypotheses we performed a ShimodairandashHasegawa test no statistically significant differenceswere observed between the two hypotheses
SOUTHERN MEXICAN NOTROPIS
The populations analysed which have been tradition-ally assigned to N boucardi and N moralesi formedtwo well-supported clades (mean sequence divergenceamong them was 507 ranging from 43 to 60 andnet sequence divergence (dA) was 395) One cladecontained 14 specimens belonging to 13 populations ofN moralesi which inhabit three different drainages(Atoyac Balsas and Papaloapan) and include speci-mens sampled in the type locality of the species Meansequence divergence among specimens of N moralesi(137) ranged from 02 to 30 (nucleotide diversity(π) was 0013) This clade was the sister group of spec-imens of N boucardi (Cuernavaca Balsas drainage)(π 0008) (Fig 4)
The remaining members of the southern MexicanNotropis formed another well-supported clade whichincluded all specimens of N imeldae from differentrivers of the Atoyac drainage and a population of Not-ropis sp 1 from an isolated spring of the Balsas drain-age (Oaxaca State) N sp 1 was identified astaxonomically distinct on the basis of isozyme data(Schoumlnhuth et al 2001) and awaits a formal descrip-tion (Schoumlnhuth 2002) Mean sequence divergencebetween N imeldae and N sp 1 was 7 (dA 67) Netsequence divergence between both species and othersouth Mexican Notropis ranged from 99 to 118 Netsequence divergence between the N imeldae-N sp 1clade and central Mexican cyprinids ranged from 80to 96 (Table 2)
CENTRAL MEXICAN CYPRINIDS
N calientis N sallaei and Y alta are distributed overthe trans-Mexican volcanic belt in central Mexicoinhabiting the Riacuteo Lerma drainage the lagoons of theMesa Central and the Riacuteo Paacutenuco headwaters A cladecomposed of these species was well-supported in theML (74) and NJ (85) analyses Mean sequencedivergence between N sallaei and N calientis (921)ranged from 85 to 101 (dA 608) and mean
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 329
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 2
R
ange
an
d m
ean
(S
D)
of p
erce
nta
ge g
enet
ic d
iver
gen
ce f
or t
he
intr
aspe
cifi
c (d
iago
nal
) an
d in
ters
peci
fic
(abo
ve d
iago
nal
) co
mpa
riso
ns
amon
g sp
ecim
ens
anal
ysed
for
cyt
och
rom
e b
base
d on
un
corr
ecte
d p-
dist
ance
s L
ast
row
in
dia
gon
al (
bold
) sh
ows
nu
cleo
tide
div
ersi
ty (
π) w
ith
in s
peci
es
Net
seq
uen
ce d
iver
gen
ce(d
A)
is b
elow
th
e di
agon
al
Mex
ican
min
now
sM
inn
ows
from
US
A
and
Can
ada
N b
ouca
rdi
N m
oral
esi
N i
mel
dae
N s
p 1
N c
alie
nti
sN
sal
laei
Y a
lta
Not
ropi
sH
ybop
sis
N b
ouca
rdi
08
43ndash
60
111
ndash11
412
1ndash1
23
128
ndash13
712
5ndash1
29
121
ndash12
813
8ndash1
63
142
ndash15
10
875
07 (
04)
112
7 (0
1)
122
7 (0
1)
133
7 (0
3)
127
5 (0
1)
125
2 (0
1)
152
3 (0
6)
147
8 (0
4)
000
8N
mor
ales
i3
950
2ndash3
010
2ndash1
18
113
ndash11
913
1ndash1
52
122
ndash13
511
9ndash1
36
129
ndash16
413
9ndash1
48
137
(0
6)10
95
(03
)11
65
(02
)14
26
(04
)12
86
(03
)12
61
(03
)14
96
(07
)14
30
(03
)0
013
N i
mel
dae
105
59
980
1ndash0
80
58 (
03)
000
5
70ndash
71
706
(0
1)10
8ndash1
17
114
0 (0
2)
99ndash
104
100
9 (0
2)
94ndash
100
984
(0
2)11
7ndash1
46
134
5 (0
9)
123
ndash12
912
65
(02
)
N s
p 1
118
410
97
677
010
3ndash1
11
106
9 (0
3)
95ndash
98
968
(0
1)10
6ndash1
12
109
8 (0
2)
122
ndash15
013
79
(08
)13
7ndash1
39
138
5 (0
1)
N c
alie
nti
s10
59
112
38
068
351
4ndash7
04
69 (
22)
004
6
85ndash
101
921
(0
4)10
4ndash1
23
110
8 (0
5)
121
ndash16
014
04
(09
)12
7ndash1
40
133
8 (0
4)
N s
alla
ei11
53
113
99
69
296
081
51
570
015
90ndash
94
927
(0
1)12
0ndash1
43
128
7 (0
6)
126
13
312
98
(02
)
Y a
lta
113
511
19
859
923
87
750
9ndash1
91
47 (
03)
001
4
113
ndash15
113
25
(09
)13
5ndash1
43
139
6 (0
2)
Not
ropi
s3
9ndash15
512
25
(24
)12
2ndash1
53
137
8 (0
8)
Hyb
opsi
s7
47
45
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

324
S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London
Biological Journal of the Linnean Society
2003
80
323ndash337
genus
Aztecula
(Mayden 1991 Coburn amp Cavender1992) The remaining Mexican species analysed
N boucardi
N moralesi
N imeldae
and
N calientis
have been assigned to the genus
Notropis
(Chernoff ampMiller 1986 Vallejo 1988 Espinosa
et al
1993) or tothe genus
Hybopsis
(Mayden 1991)
N calientis
shows a high degree of population structure and hasbeen recognized as a species complex (Chernoff ampMiller 1986) On the other hand
Yuriria
was consid-ered as a subgenus of the genus
Hybopsis
(Bailey1951) until Mayden (1989) elevated
Yuriria
to genericstatus
Based on morphology two principal phylogenetichypotheses placed the species analysed in differentphylogenetic clades Coburn amp Cavenderrsquos (1992)hypothesis considers the genus
Notropis
(
sl
includ-ing genus
Hybopsis
) as a member of the Shiner cladeand the genus
Yuriria
as a member of the Westernclade Maydenrsquos (1989) hypothesis places the Mexicanspecies analysed in different clades
N boucardi
and
N calientis
as members of the genus
Hybopsis
andthe genus
Yuriria
in a different clade both of themincluded in the open posterior miodome (OPM) cladewhile
N sallaei
was excluded from this principalclade Mayden (1989) considers the genus
Notropis
tobe polyphyletic (comprising different subgenus andspecies groups)
Mitochondrial DNA sequences provide an objectiveframework for the analysis of population processesand evolutionary patterns (Bermingham amp Martin1998) and have been useful in studies of phylogeneticrelationships of several groups of North American cyp-rinids (Dowling amp Naylor 1997 Simons amp Mayden1997 1998 1999) In particular cytochrome
b
hasbeen shown to be an appropriate gene for inferringphylogenetic relationships in several phylogeneticstudies on freshwater fishes (Martin amp Bermingham1998 Zardoya amp Doadrio 1999 Bielawski amp Gold2001 Machordom amp Doadrio 2001) Our study is thefirst to address relationships among Mexican
Notropis
using mtDNA sequences and to provide a phylogeneticframework with the northern species of the genera
Notropis
and
Hybopsis
The purpose of this work was thus to use the com-
plete sequence of the cytochrome
b
gene to infer phy-logenetic affinities of Mexican endemic species of thegenus
Notropis
with species that represent differentsubgenera or species groups of the genus
Notropis
from the USA and Canada and the genus
Hybopsis
and to test previous hypotheses of paraphyletic lin-eages within Mexican endemic species In particularwe used molecular sequence data from 30 geographi-cally distinct populations of seven Mexican endemicspecies of
Notropis
to test evolutionary hypothesesregarding their patterns of diversification in centraland southern Mexico
MATERIAL AND METHODS
The taxa sequenced in this study were previouslyassigned to four different genera (Mayden 1991) ndash
Hybopsis
Aztecula
Notropis
and
Yuriria
ndash and rep-resent the different clades proposed by Mayden (1989)and Coburn amp Cavender (1992) Due to the controver-sial taxonomy of the Mexican species analysed wehave followed the classification of Eschmeyer (httpwwwfishbaseorg) We sequenced 34 specimens of 30populations representing seven Mexican endemic spe-cies of minnows (
N moralesi
N boucardi
N imeldae
N sallaei
N calientis Y alta
and one undescribedspecies of the genus
Notropis
referred to here as
N
sp 1) distributed over central and southern Mexicoplus three species of the genus
Notropis
from Canada(
N rubellus
N anogenus
and
N heterodon
obtainedfrom the Royal Ontario Museum) Seventeen cyprinidsequences were obtained from GenBank and includedin the phylogenetic analyses
N atherinoides
(AF352272)
N volucellus
(AF352268)
N boops
(AF352261)
N suttkusi
(AF352287)
N jemezanus
(AF352277)
N amabilis
(AF352269)
N stilbius(AF352286) N shumardi (AF117200) N potteri(AF117192) N blennius (AF117171) Hybopsisamblops (AF117153) Hybopsis winchelli (AF117165)Cyprinella spiloptera (L07753) Pteronotropissignipinnis (AF261230) Moapa coriacea (AF452075)Barbus sclateri (AF334083) and Tinca tinca (Y10451)The last three taxa were used as outgroups
DNA SOURCES
Fish samples were collected by electrofishing in thesouthern and central region of Mexico from Sierrade Miahuatlaacuten in Oaxaca at about 16degN latitude tothe Lerma drainage in Guanajuato State some21degN These collections (Table 1) cover the Atlanticand the Pacific slopes and represent five riverbasins Papaloapan Balsas Atoyac Lerma andPaacutenuco (Fig 1) The specimens analysed correspondto populations inhabiting the entire distributionarea known for N moralesi N boucardi N imeldaeand N sp 1 We obtained N sallaei specimens fromthe Paacutenuco and Lerma drainages N calientis andY alta were fished from Lerma river drainage andadjacent lakes
DNA EXTRACTION
Total DNA was extracted from muscle tissue Frozenpieces of tissue were ground in liquid N2 and homog-enized in 600 microL extraction buffer (NaCl 014 M mag-nesium acetate 15 mM KCl 5 mM 1 SDS) Theresulting homogenate was extracted using standardproteinase Kphenolndashchloroform protocols (Sambrook
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 325
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 1
S
peci
men
s u
sed
in t
his
stu
dy a
nd
loca
lity
in
form
atio
n
Pop
ula
tion
de
sign
atio
nS
peci
es
MN
CN
ID
Dra
inag
eR
iver
Loc
alit
yS
tate
Gen
Ban
k R
ef
1 F
ran
cisc
oN
otro
pis
imel
dae
353
Ato
yac
San
Fra
nci
sco
San
Pab
lo C
oatl
an O
axac
aA
F46
9131
2 G
ruta
sN
im
eld
ae 1
095
Ato
yac
De
las
Gru
tas
rive
r (S
prin
g)S
ola
de V
ega
Oax
aca
AF
4691
323
Sab
ino
N i
mel
dae
113
8A
toya
cA
gua
del
Sab
ino
Sol
a de
Veg
a O
axac
aA
F46
9130
4 V
erde
N m
oral
esi
97A
toya
cG
ran
de-V
erde
Noc
hix
tlan
Oax
aca
AF
4691
535
Bra
voN
mor
ales
i 10
67A
toya
cB
ravo
San
Ju
an T
eita
Tla
xiac
o O
axac
aA
F46
9152
6 I
gual
ites
N m
oral
esi
5059
Bal
sas
Igu
alit
esT
lapa
Gu
erre
roA
F46
9151
7 C
oico
yan
N m
oral
esi
1086
Bal
sas
Coi
coya
n d
e la
s F
lore
sC
oico
yan
Ju
xtla
hu
aca
Oax
aca
AF
4691
458
Ch
iqu
ito
N m
oral
esi
133
Bal
sas
Ch
iqu
ito
(Spr
ing)
Tla
xiac
o O
axac
aA
F46
9150
9 S
alad
oN
mor
ales
i 99
0B
alsa
sS
alad
oH
uaj
uap
an O
axac
aA
F46
9156
10 I
sabe
lN
mor
ales
i 19
99B
alsa
sS
ta Isa
bel
Ch
ilap
a G
uer
rero
AF
4691
4811
Mix
teco
N m
oral
esi
5048
Bal
sas
Tec
oloy
anX
och
ihu
ehu
etla
n G
uer
rero
AF
4691
4912
Mix
teco
-1N
mor
ales
i 10
6B
alsa
sM
ixte
coA
sun
cioacuten
Tla
xiac
o O
axac
aA
F46
9144
13 T
elol
oapa
nN
mor
ales
i 20
09B
alsa
sT
elol
oapa
nT
elol
oapa
n G
uer
rero
AF
4691
4714
Hon
doN
mor
ales
i 91
4P
apal
oapa
nH
ondo
Con
cepc
ioacuten
Bu
ena
Vis
ta O
axac
aA
F46
9157
15 G
ran
de
N m
oral
esi
150
154
Pap
aloa
pan
Gra
nde
Tep
eln
eme
de M
orel
os O
axac
aA
F46
9154
ndash55
16 A
mac
uya
cN
mor
ales
i 34
67B
alsa
sA
mac
uya
cG
ruta
s de
Cac
ahoa
mil
pa G
uer
rero
AF
4691
4617
Pol
lo
N b
ouca
rdi
3487
Bal
sas
del
Pol
lo C
ol L
agu
nil
la
Cu
ern
avac
a M
orel
osA
F46
9158
18 H
uel
lapa
nN
bou
card
i 34
74B
alsa
sH
uel
lapa
n l
agoo
nE
l T
exal
Cu
ern
avac
a M
orel
osA
F46
9159
19 T
axin
gu
N s
alla
ei 3
125
Paacuten
uco
Tax
ingu
Dam
Acu
lco
Meacutex
ico
DF
A
F46
9135
20 A
lmoy
aN
sal
laei
351
7L
erm
aA
lmoy
a de
l R
iacuteo l
agoo
nA
lmoy
a de
l R
iacuteo M
eacutexic
o D
F
AF
4691
3621
SM
Cu
evas
N s
p 4
17 4
19B
alsa
sS
an M
igu
el C
uev
as (
Spr
ing)
Juxt
lah
uac
a O
axac
aA
F46
9133
ndash34
22 P
iru
les
N c
alie
nti
s 33
26L
erm
aP
iru
les
Dam
San
Ju
an R
ayas
Qu
eret
aro
AF
4691
3723
Zac
apu
N c
alie
nti
s 36
663
717
Zac
apu
Zac
apu
lag
oon
Zac
apu
Mic
hoa
caacuten
AF
4691
41ndash4
224
Ch
arco
N c
alie
nti
s 39
18L
erm
aC
har
co d
el I
nge
nio
San
Mig
uel
All
ende
Gu
anaj
uat
oA
F46
9143
25 M
inzi
taN
cal
ien
tis
3813
Cu
itze
oL
a M
inzi
ta D
amL
a M
inzi
ta M
ich
oacaacute
nA
F46
9138
26 S
Mig
uel
N c
alie
nti
s 38
29 3
831
Ler
ma
San
Mig
uel
(S
prin
g)S
an M
igu
el 2
033
m M
ich
oacaacute
nA
F46
9139
ndash40
27 C
eja
Yuri
ria
alta
333
4L
erm
aC
eja
de B
ravo
Dam
La
Cej
a de
Bra
vo Q
uer
etar
oA
F46
9161
28 Z
acap
uY
alt
a 34
49L
erm
aZ
acap
u l
agoo
n o
utl
edP
anin
dicu
aro
Mic
hoa
caacuten
AF
4691
6329
Laj
asY
alt
a 33
70L
erm
aL
ajas
tri
buta
ryS
an M
igu
el A
llen
de G
uan
aju
ato
AF
4691
6230
Min
zita
Y a
lta
3809
Cu
itze
oL
a M
inzi
ta D
amL
a M
inzi
ta M
ich
oacaacute
nA
F46
9160
31 C
anad
aN
ru
bell
us
1703
32G
ran
d R
iver
Bra
ntf
ord
On
tari
o C
anad
aA
F46
9164
32 C
anad
aN
het
erod
on 1
7033
1E
ire
Lak
eR
onde
au P
rovi
nci
al P
ark
On
tari
o C
anad
aA
Y14
0697
33 C
anad
aN
an
ogen
us
1703
36S
t C
lair
Lak
eM
itch
ell
Bay
On
tari
o C
anad
aA
Y14
0698
Nu
mbe
rs a
ssig
ned
to
popu
lati
ons
corr
espo
nd
to t
hos
e of
th
e m
ap (
see
Fig
1)
MN
CN
ID
nu
mbe
rs
nu
mbe
rs o
f vo
uch
er s
peci
men
s de
posi
ted
in t
he
ich
thyo
logy
coll
ecti
on o
f th
e M
use
o N
acio
nal
de
Cie
nci
as N
atu
rale
s (S
pain
) G
enB
ank
Ref
acc
essi
on n
um
ber
for
nu
cleo
tide
seq
uen
ces
depo
site
d in
Gen
Ban
kT
opot
ypes
or
spec
imen
s fo
un
d cl
ose
to t
hei
r te
rra
typi
ca
326 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Fritsch amp Maniatis 1989) The DNA was resuspendedin ultrapure water and used for amplification by PCR
AMPLIFICATION AND CLONING
Combinations of two sets of versatile primers (Zardoyaamp Doadrio 1998) were used in PCR amplifications toisolate two contiguous and overlapping fragments (of660 and 521 bp) that covered the entire cytochrome bgene The PCR reactions were conducted in 25 microl vol-umes (67 mM Tris-HCl pH 83 15 mM MgCl2 04 mM
each primer 1 microl DNA extraction and 1 unit Taq DNApolymerase (Promega)) After an initial denaturationstep at 94degC for 2 min 35 cycles were performed as fol-lows denaturation at 94degC (1 min) annealing at 48degC(1 min) and extension at 72degC (145 min) with a finalextension of 7 min at 72degC
After PCR amplification the DNA was precipitatedcloned into p-GEMT vectors and transformed intocompetent E coli (JM109) Positive clones weresequenced using the FS-Taq Dye Deoxy terminatorcycle sequencing kit (Applied Biosystems Inc) usingan automated DNA sequencer (Applied Biosystems3700) following the manufacturerrsquos instructions TheDNA sequences of both strands were obtained using
M13 universal (forward and reverse) sequencing prim-ers The sequences determined here have been depos-ited in GenBank under accession numbers AF469130ndashAF469164 AY140697 and AY140698
CYTOCHROME b SEQUENCE ANALYSES
Forty-nine complete minnow cytochrome b genesequences (1141 bp) were aligned manually withsequences of N atherinoides H amblops Cyprinellaspiloptera Tinca tinca and Barbus sclateri No ambig-uous alignments or gaps were found All codon posi-tions were included in the phylogenetic analyses
All Mexican specimens representing 30 populationsof seven minnow species were used to determine theirpopulation structure (Table 1) The intrapopulationaldivergence levels were corrected for ancestral poly-morphisms to avoid the effect of inflating divergencebetween phylogroups or species We applied the netsequence divergence correction described by Nei(1987) and the index of nucleotide diversity (π) (Nei ampLi 1979) using DnaSP ver 350 (Rozas amp Rozas 1999)Higher level relationships were inferred using 22 spe-cies of Mexican USA and Canadian NotropisHybop-sis and two species of the phylogenetically related
Figure 1 Localities from which specimens of the genera Notropis and Yuriria were sampled Numbers indicate collectionsample (see Table 1) Drainages are indicated by letters A Atoyac B Balsas C Lerma D Paacutenuco E PapaloapanDistribution range of each species is presented in different grey patterns
115degW
30degN
25deg
20deg
15deg
110deg 105deg 100deg 95deg 0 200 km
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 327
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
genera Cyprinella and Pteronotropis included in theNotropin clade (Phoxinini lineage Shiner clade)(Coburn amp Cavender 1992) The genus Moapa fromthe Western clade Phoxinini was selected as a taxo-nomic related outgroup whereas Tinca was chosen asa more distant outgroup
Phylogenetic trees were recovered using maximumparsimony (MP) neighbour joining (NJ) maximumlikelihood (ML) and Bayesian inference (BI) We usedfour different methods for inferring phylogenies to testthe congruence among methods and compare for thebest approximation for our data There has recentlybeen controversy about the overcredibility of molecu-lar phylogenies obtained with Bayesian inference(Suzuki Glazco amp Nei 2002) while others have sug-gested posterior probability as a better indicator ofstatistical confidence than bootstrap probability (Wil-cox et al 2002) Both bootstrap and posterior proba-bility were included It is well known that MP givesgood results in the Felsenstein inverse zone and MLin both zones when more than 1000 nucleotides areanalysed However accuracy decreases when theinternal nodes are short (Swofford et al 2001) Theanalysis of 1141 nucleotides resulted in short internalnodes within trees so we decided to include the fourmethods of analysis to infer the phylogeny
The MP analysis was performed through heuristicsearches with ten random stepwise additions of taxaMULTREE option and TBR branch swapping For theMP analyses equal weight of transversions (Tv) andtransitions (Ts) or Tv five times the weight of Ts basedon empirical evidence were used For ML BI and NJanalyses we used the hierarchical likelihood ratio test(LRT) implemented in Modeltest V304 (Posada ampCrandall 1998) to find the evolutionary model thatbest fit our data The HKY + I + G model (base fre-quencies A 03294 C 02805 G 01433 T 02469Nst 2 TsTv ratio 56889 Γ 09381 proportion ofinvariable sites 04383) was selected when Mexicanminnow populations were taken into account and theGTR + I + G model (base frequencies A 02853 C03302 G 01125 T 02720 Nst 6 Γ 07619 propor-tion of invariable sites 04915) was selected when onespecimen per species was analysed
All phylogenetic analyses were performed usingPAUP version 40b8 (Swofford 2001) and MR BAYES(Huelsenbeck amp Ronquist 2001) Robustness of theinferred trees was tested by bootstrapping (Felsen-stein 1985) with 100 pseudoreplications for MP and1000 for NJ with 1000 quartet puzzling steps forML and 500 000 generations for Bayesian posteriorprobability
Our two alternative phylogenetic hypotheses forN imeldae-N sp 1 clade were tested using the Shimo-dairandashHasegawa test (Shimodaira amp Hasegawa 1999)as implemented in PAUP
The genetic divergences widely discussed throughthe text were based on the uncorrected p-distances Toavoid confusion between derived divergences based onthe different evolutionary models selected (HKY +I + G and GTR + I + G) we considered more appropri-ate the uncorrected p genetic distances which allowscomparisons among our two data sets and could permita broad comparison with other related taxa thatshowed a different model of evolution
RESULTS
The complete cytochrome b gene was successfullyamplified and sequenced in 37 specimens of the generaNotropis and Yuriria Of the 1141 bp aligned for alltaxa 488 sites were variable and 373 were parsimonyinformative when only Mexican minnow populationswere taken into account Informative characters roseto 387 when one specimen per species for all NorthAmerican minnows analysed was considered
The nucleotide composition in the cytochrome bgene indicates a bias against guanine (see Modeltestparameters in methods) which is particularly strongin third codon positions (9) Most of the variabilityamong cytochrome b sequences was detected at thirdcodon positions No saturation was detected at anycodon position (Fig 2) However separate plots of AndashG
Figure 2 Relationships between uncorrected meansequence divergence (p-distance) and number of transi-tions () and transversions () at all codon positions (A)and at third codon positions (B) in the cytochrome b genefor all pairwise comparisons for Mexican populationsamples
020406080
100120140160180
005 01 015 02p
Cha
nges
020406080
100120140160
005 01 015 02
p
Cha
nges
0
0
A
B
328 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
and CndashT transitional divergences at third codon posi-tions suggested that AndashG transitions changed fasterthan CndashT ones (data not shown)
Pairwise sequence divergence (uncorrected p-distance) among northern species of the genus Notro-pis analysed ranged from 39 (between N blenniusand N potteri) to 155 (N potteri and N volucellus)Among Mexican species net sequence divergenceranged from 39 (between specimens of N moralesiand N boucardi) to 115 (between specimens ofN sallaei and N boucardi) (Table 2) Within Mexicanspecies the mean divergence among specimensrepresenting different populations was lt1 (amongN imeldae specimens and among specimens of geo-graphically close populations of N moralesi) to 3(among specimens of regionally separated populationsof N moralesi) Nucleotide diversity (π) within mostMexican species ranged from 0005 to 0015 Howeverfour specimens from two different sample sites ofN calientis fell outside this range showing over 7sequence divergence and nucleotide diversity up to0046 (Table 2)
Cytochrome b sequences were analysed using fourcommon methods of phylogenetic inference (MP NJML BI) and recovered congruent topologies (Figs 3and 4) Topologies recovered from unweighted andweighted most parsimonious trees were similar butthe weighted analyses yielded a more highly resolvedtopology congruent to the remaining recovered analy-ses When one specimen per species was taken intoaccount in order to place the Mexican species into alarger phylogenetic framework all cyprinids of thegenera Notropis and Hybopsis were grouped togetherwith Pteronotropis and Cyprinella in a basal positionAccording to MP ML NJ and BI all Mexican speciesof the genera Notropis and Yuriria formed a well-supported clade with respect to the remaining speciesof the genus Notropis from the USA and Canada North-ern species of the genus Notropis formed three differ-ent well-supported clades while basal relationshipsamong clades remained unresolved The phylogeneticposition of the genus Hybopsis remained unresolved(Fig 3) Interestingly the genus Yuriria groupedtogether with all Mexican Notropis in all analysesThe robustness of these results was confirmed by highbootstrap values and Bayesian posterior probability
When populations were taken into account all Mex-ican Notropis plus Yuriria clustered together withrespect to the northern Notropis Hybopsis and Cyp-rinella forming a well-supported clade with bootstrapvalues (MP 94 ML 72 NJ 86) and BI (100)(Fig 4) Within this clade four well-supported lin-eages could be distinguished (Fig 4) (A) grouped allspecimens representing populations of N boucardiand N moralesi (B) grouped N imeldae and N sp 1(C) was formed by all specimens of N sallaei and
N calientis (D) included the Y alta group The genusYuriria was grouped with N calientis-N sallaei as sis-ter species using NJ (85) and ML (74) (Figs 3 4)The relative position of the clade N imeldae-N sp 1was different depending on the phylogenetic analysisaccording to MP (79) ML (58) and BI (100) it wasconsidered a sister group of the N moralesi-N boucardi clade whereas it appeared as a sistergroup of the clade (Yuriria (N calientis N sallaei)) inthe NJ analysis (66) (Fig 4) In order to test the twoalternative hypotheses we performed a ShimodairandashHasegawa test no statistically significant differenceswere observed between the two hypotheses
SOUTHERN MEXICAN NOTROPIS
The populations analysed which have been tradition-ally assigned to N boucardi and N moralesi formedtwo well-supported clades (mean sequence divergenceamong them was 507 ranging from 43 to 60 andnet sequence divergence (dA) was 395) One cladecontained 14 specimens belonging to 13 populations ofN moralesi which inhabit three different drainages(Atoyac Balsas and Papaloapan) and include speci-mens sampled in the type locality of the species Meansequence divergence among specimens of N moralesi(137) ranged from 02 to 30 (nucleotide diversity(π) was 0013) This clade was the sister group of spec-imens of N boucardi (Cuernavaca Balsas drainage)(π 0008) (Fig 4)
The remaining members of the southern MexicanNotropis formed another well-supported clade whichincluded all specimens of N imeldae from differentrivers of the Atoyac drainage and a population of Not-ropis sp 1 from an isolated spring of the Balsas drain-age (Oaxaca State) N sp 1 was identified astaxonomically distinct on the basis of isozyme data(Schoumlnhuth et al 2001) and awaits a formal descrip-tion (Schoumlnhuth 2002) Mean sequence divergencebetween N imeldae and N sp 1 was 7 (dA 67) Netsequence divergence between both species and othersouth Mexican Notropis ranged from 99 to 118 Netsequence divergence between the N imeldae-N sp 1clade and central Mexican cyprinids ranged from 80to 96 (Table 2)
CENTRAL MEXICAN CYPRINIDS
N calientis N sallaei and Y alta are distributed overthe trans-Mexican volcanic belt in central Mexicoinhabiting the Riacuteo Lerma drainage the lagoons of theMesa Central and the Riacuteo Paacutenuco headwaters A cladecomposed of these species was well-supported in theML (74) and NJ (85) analyses Mean sequencedivergence between N sallaei and N calientis (921)ranged from 85 to 101 (dA 608) and mean
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 329
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 2
R
ange
an
d m
ean
(S
D)
of p
erce
nta
ge g
enet
ic d
iver
gen
ce f
or t
he
intr
aspe
cifi
c (d
iago
nal
) an
d in
ters
peci
fic
(abo
ve d
iago
nal
) co
mpa
riso
ns
amon
g sp
ecim
ens
anal
ysed
for
cyt
och
rom
e b
base
d on
un
corr
ecte
d p-
dist
ance
s L
ast
row
in
dia
gon
al (
bold
) sh
ows
nu
cleo
tide
div
ersi
ty (
π) w
ith
in s
peci
es
Net
seq
uen
ce d
iver
gen
ce(d
A)
is b
elow
th
e di
agon
al
Mex
ican
min
now
sM
inn
ows
from
US
A
and
Can
ada
N b
ouca
rdi
N m
oral
esi
N i
mel
dae
N s
p 1
N c
alie
nti
sN
sal
laei
Y a
lta
Not
ropi
sH
ybop
sis
N b
ouca
rdi
08
43ndash
60
111
ndash11
412
1ndash1
23
128
ndash13
712
5ndash1
29
121
ndash12
813
8ndash1
63
142
ndash15
10
875
07 (
04)
112
7 (0
1)
122
7 (0
1)
133
7 (0
3)
127
5 (0
1)
125
2 (0
1)
152
3 (0
6)
147
8 (0
4)
000
8N
mor
ales
i3
950
2ndash3
010
2ndash1
18
113
ndash11
913
1ndash1
52
122
ndash13
511
9ndash1
36
129
ndash16
413
9ndash1
48
137
(0
6)10
95
(03
)11
65
(02
)14
26
(04
)12
86
(03
)12
61
(03
)14
96
(07
)14
30
(03
)0
013
N i
mel
dae
105
59
980
1ndash0
80
58 (
03)
000
5
70ndash
71
706
(0
1)10
8ndash1
17
114
0 (0
2)
99ndash
104
100
9 (0
2)
94ndash
100
984
(0
2)11
7ndash1
46
134
5 (0
9)
123
ndash12
912
65
(02
)
N s
p 1
118
410
97
677
010
3ndash1
11
106
9 (0
3)
95ndash
98
968
(0
1)10
6ndash1
12
109
8 (0
2)
122
ndash15
013
79
(08
)13
7ndash1
39
138
5 (0
1)
N c
alie
nti
s10
59
112
38
068
351
4ndash7
04
69 (
22)
004
6
85ndash
101
921
(0
4)10
4ndash1
23
110
8 (0
5)
121
ndash16
014
04
(09
)12
7ndash1
40
133
8 (0
4)
N s
alla
ei11
53
113
99
69
296
081
51
570
015
90ndash
94
927
(0
1)12
0ndash1
43
128
7 (0
6)
126
13
312
98
(02
)
Y a
lta
113
511
19
859
923
87
750
9ndash1
91
47 (
03)
001
4
113
ndash15
113
25
(09
)13
5ndash1
43
139
6 (0
2)
Not
ropi
s3
9ndash15
512
25
(24
)12
2ndash1
53
137
8 (0
8)
Hyb
opsi
s7
47
45
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 325
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 1
S
peci
men
s u
sed
in t
his
stu
dy a
nd
loca
lity
in
form
atio
n
Pop
ula
tion
de
sign
atio
nS
peci
es
MN
CN
ID
Dra
inag
eR
iver
Loc
alit
yS
tate
Gen
Ban
k R
ef
1 F
ran
cisc
oN
otro
pis
imel
dae
353
Ato
yac
San
Fra
nci
sco
San
Pab
lo C
oatl
an O
axac
aA
F46
9131
2 G
ruta
sN
im
eld
ae 1
095
Ato
yac
De
las
Gru
tas
rive
r (S
prin
g)S
ola
de V
ega
Oax
aca
AF
4691
323
Sab
ino
N i
mel
dae
113
8A
toya
cA
gua
del
Sab
ino
Sol
a de
Veg
a O
axac
aA
F46
9130
4 V
erde
N m
oral
esi
97A
toya
cG
ran
de-V
erde
Noc
hix
tlan
Oax
aca
AF
4691
535
Bra
voN
mor
ales
i 10
67A
toya
cB
ravo
San
Ju
an T
eita
Tla
xiac
o O
axac
aA
F46
9152
6 I
gual
ites
N m
oral
esi
5059
Bal
sas
Igu
alit
esT
lapa
Gu
erre
roA
F46
9151
7 C
oico
yan
N m
oral
esi
1086
Bal
sas
Coi
coya
n d
e la
s F
lore
sC
oico
yan
Ju
xtla
hu
aca
Oax
aca
AF
4691
458
Ch
iqu
ito
N m
oral
esi
133
Bal
sas
Ch
iqu
ito
(Spr
ing)
Tla
xiac
o O
axac
aA
F46
9150
9 S
alad
oN
mor
ales
i 99
0B
alsa
sS
alad
oH
uaj
uap
an O
axac
aA
F46
9156
10 I
sabe
lN
mor
ales
i 19
99B
alsa
sS
ta Isa
bel
Ch
ilap
a G
uer
rero
AF
4691
4811
Mix
teco
N m
oral
esi
5048
Bal
sas
Tec
oloy
anX
och
ihu
ehu
etla
n G
uer
rero
AF
4691
4912
Mix
teco
-1N
mor
ales
i 10
6B
alsa
sM
ixte
coA
sun
cioacuten
Tla
xiac
o O
axac
aA
F46
9144
13 T
elol
oapa
nN
mor
ales
i 20
09B
alsa
sT
elol
oapa
nT
elol
oapa
n G
uer
rero
AF
4691
4714
Hon
doN
mor
ales
i 91
4P
apal
oapa
nH
ondo
Con
cepc
ioacuten
Bu
ena
Vis
ta O
axac
aA
F46
9157
15 G
ran
de
N m
oral
esi
150
154
Pap
aloa
pan
Gra
nde
Tep
eln
eme
de M
orel
os O
axac
aA
F46
9154
ndash55
16 A
mac
uya
cN
mor
ales
i 34
67B
alsa
sA
mac
uya
cG
ruta
s de
Cac
ahoa
mil
pa G
uer
rero
AF
4691
4617
Pol
lo
N b
ouca
rdi
3487
Bal
sas
del
Pol
lo C
ol L
agu
nil
la
Cu
ern
avac
a M
orel
osA
F46
9158
18 H
uel
lapa
nN
bou
card
i 34
74B
alsa
sH
uel
lapa
n l
agoo
nE
l T
exal
Cu
ern
avac
a M
orel
osA
F46
9159
19 T
axin
gu
N s
alla
ei 3
125
Paacuten
uco
Tax
ingu
Dam
Acu
lco
Meacutex
ico
DF
A
F46
9135
20 A
lmoy
aN
sal
laei
351
7L
erm
aA
lmoy
a de
l R
iacuteo l
agoo
nA
lmoy
a de
l R
iacuteo M
eacutexic
o D
F
AF
4691
3621
SM
Cu
evas
N s
p 4
17 4
19B
alsa
sS
an M
igu
el C
uev
as (
Spr
ing)
Juxt
lah
uac
a O
axac
aA
F46
9133
ndash34
22 P
iru
les
N c
alie
nti
s 33
26L
erm
aP
iru
les
Dam
San
Ju
an R
ayas
Qu
eret
aro
AF
4691
3723
Zac
apu
N c
alie
nti
s 36
663
717
Zac
apu
Zac
apu
lag
oon
Zac
apu
Mic
hoa
caacuten
AF
4691
41ndash4
224
Ch
arco
N c
alie
nti
s 39
18L
erm
aC
har
co d
el I
nge
nio
San
Mig
uel
All
ende
Gu
anaj
uat
oA
F46
9143
25 M
inzi
taN
cal
ien
tis
3813
Cu
itze
oL
a M
inzi
ta D
amL
a M
inzi
ta M
ich
oacaacute
nA
F46
9138
26 S
Mig
uel
N c
alie
nti
s 38
29 3
831
Ler
ma
San
Mig
uel
(S
prin
g)S
an M
igu
el 2
033
m M
ich
oacaacute
nA
F46
9139
ndash40
27 C
eja
Yuri
ria
alta
333
4L
erm
aC
eja
de B
ravo
Dam
La
Cej
a de
Bra
vo Q
uer
etar
oA
F46
9161
28 Z
acap
uY
alt
a 34
49L
erm
aZ
acap
u l
agoo
n o
utl
edP
anin
dicu
aro
Mic
hoa
caacuten
AF
4691
6329
Laj
asY
alt
a 33
70L
erm
aL
ajas
tri
buta
ryS
an M
igu
el A
llen
de G
uan
aju
ato
AF
4691
6230
Min
zita
Y a
lta
3809
Cu
itze
oL
a M
inzi
ta D
amL
a M
inzi
ta M
ich
oacaacute
nA
F46
9160
31 C
anad
aN
ru
bell
us
1703
32G
ran
d R
iver
Bra
ntf
ord
On
tari
o C
anad
aA
F46
9164
32 C
anad
aN
het
erod
on 1
7033
1E
ire
Lak
eR
onde
au P
rovi
nci
al P
ark
On
tari
o C
anad
aA
Y14
0697
33 C
anad
aN
an
ogen
us
1703
36S
t C
lair
Lak
eM
itch
ell
Bay
On
tari
o C
anad
aA
Y14
0698
Nu
mbe
rs a
ssig
ned
to
popu
lati
ons
corr
espo
nd
to t
hos
e of
th
e m
ap (
see
Fig
1)
MN
CN
ID
nu
mbe
rs
nu
mbe
rs o
f vo
uch
er s
peci
men
s de
posi
ted
in t
he
ich
thyo
logy
coll
ecti
on o
f th
e M
use
o N
acio
nal
de
Cie
nci
as N
atu
rale
s (S
pain
) G
enB
ank
Ref
acc
essi
on n
um
ber
for
nu
cleo
tide
seq
uen
ces
depo
site
d in
Gen
Ban
kT
opot
ypes
or
spec
imen
s fo
un
d cl
ose
to t
hei
r te
rra
typi
ca
326 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Fritsch amp Maniatis 1989) The DNA was resuspendedin ultrapure water and used for amplification by PCR
AMPLIFICATION AND CLONING
Combinations of two sets of versatile primers (Zardoyaamp Doadrio 1998) were used in PCR amplifications toisolate two contiguous and overlapping fragments (of660 and 521 bp) that covered the entire cytochrome bgene The PCR reactions were conducted in 25 microl vol-umes (67 mM Tris-HCl pH 83 15 mM MgCl2 04 mM
each primer 1 microl DNA extraction and 1 unit Taq DNApolymerase (Promega)) After an initial denaturationstep at 94degC for 2 min 35 cycles were performed as fol-lows denaturation at 94degC (1 min) annealing at 48degC(1 min) and extension at 72degC (145 min) with a finalextension of 7 min at 72degC
After PCR amplification the DNA was precipitatedcloned into p-GEMT vectors and transformed intocompetent E coli (JM109) Positive clones weresequenced using the FS-Taq Dye Deoxy terminatorcycle sequencing kit (Applied Biosystems Inc) usingan automated DNA sequencer (Applied Biosystems3700) following the manufacturerrsquos instructions TheDNA sequences of both strands were obtained using
M13 universal (forward and reverse) sequencing prim-ers The sequences determined here have been depos-ited in GenBank under accession numbers AF469130ndashAF469164 AY140697 and AY140698
CYTOCHROME b SEQUENCE ANALYSES
Forty-nine complete minnow cytochrome b genesequences (1141 bp) were aligned manually withsequences of N atherinoides H amblops Cyprinellaspiloptera Tinca tinca and Barbus sclateri No ambig-uous alignments or gaps were found All codon posi-tions were included in the phylogenetic analyses
All Mexican specimens representing 30 populationsof seven minnow species were used to determine theirpopulation structure (Table 1) The intrapopulationaldivergence levels were corrected for ancestral poly-morphisms to avoid the effect of inflating divergencebetween phylogroups or species We applied the netsequence divergence correction described by Nei(1987) and the index of nucleotide diversity (π) (Nei ampLi 1979) using DnaSP ver 350 (Rozas amp Rozas 1999)Higher level relationships were inferred using 22 spe-cies of Mexican USA and Canadian NotropisHybop-sis and two species of the phylogenetically related
Figure 1 Localities from which specimens of the genera Notropis and Yuriria were sampled Numbers indicate collectionsample (see Table 1) Drainages are indicated by letters A Atoyac B Balsas C Lerma D Paacutenuco E PapaloapanDistribution range of each species is presented in different grey patterns
115degW
30degN
25deg
20deg
15deg
110deg 105deg 100deg 95deg 0 200 km
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 327
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
genera Cyprinella and Pteronotropis included in theNotropin clade (Phoxinini lineage Shiner clade)(Coburn amp Cavender 1992) The genus Moapa fromthe Western clade Phoxinini was selected as a taxo-nomic related outgroup whereas Tinca was chosen asa more distant outgroup
Phylogenetic trees were recovered using maximumparsimony (MP) neighbour joining (NJ) maximumlikelihood (ML) and Bayesian inference (BI) We usedfour different methods for inferring phylogenies to testthe congruence among methods and compare for thebest approximation for our data There has recentlybeen controversy about the overcredibility of molecu-lar phylogenies obtained with Bayesian inference(Suzuki Glazco amp Nei 2002) while others have sug-gested posterior probability as a better indicator ofstatistical confidence than bootstrap probability (Wil-cox et al 2002) Both bootstrap and posterior proba-bility were included It is well known that MP givesgood results in the Felsenstein inverse zone and MLin both zones when more than 1000 nucleotides areanalysed However accuracy decreases when theinternal nodes are short (Swofford et al 2001) Theanalysis of 1141 nucleotides resulted in short internalnodes within trees so we decided to include the fourmethods of analysis to infer the phylogeny
The MP analysis was performed through heuristicsearches with ten random stepwise additions of taxaMULTREE option and TBR branch swapping For theMP analyses equal weight of transversions (Tv) andtransitions (Ts) or Tv five times the weight of Ts basedon empirical evidence were used For ML BI and NJanalyses we used the hierarchical likelihood ratio test(LRT) implemented in Modeltest V304 (Posada ampCrandall 1998) to find the evolutionary model thatbest fit our data The HKY + I + G model (base fre-quencies A 03294 C 02805 G 01433 T 02469Nst 2 TsTv ratio 56889 Γ 09381 proportion ofinvariable sites 04383) was selected when Mexicanminnow populations were taken into account and theGTR + I + G model (base frequencies A 02853 C03302 G 01125 T 02720 Nst 6 Γ 07619 propor-tion of invariable sites 04915) was selected when onespecimen per species was analysed
All phylogenetic analyses were performed usingPAUP version 40b8 (Swofford 2001) and MR BAYES(Huelsenbeck amp Ronquist 2001) Robustness of theinferred trees was tested by bootstrapping (Felsen-stein 1985) with 100 pseudoreplications for MP and1000 for NJ with 1000 quartet puzzling steps forML and 500 000 generations for Bayesian posteriorprobability
Our two alternative phylogenetic hypotheses forN imeldae-N sp 1 clade were tested using the Shimo-dairandashHasegawa test (Shimodaira amp Hasegawa 1999)as implemented in PAUP
The genetic divergences widely discussed throughthe text were based on the uncorrected p-distances Toavoid confusion between derived divergences based onthe different evolutionary models selected (HKY +I + G and GTR + I + G) we considered more appropri-ate the uncorrected p genetic distances which allowscomparisons among our two data sets and could permita broad comparison with other related taxa thatshowed a different model of evolution
RESULTS
The complete cytochrome b gene was successfullyamplified and sequenced in 37 specimens of the generaNotropis and Yuriria Of the 1141 bp aligned for alltaxa 488 sites were variable and 373 were parsimonyinformative when only Mexican minnow populationswere taken into account Informative characters roseto 387 when one specimen per species for all NorthAmerican minnows analysed was considered
The nucleotide composition in the cytochrome bgene indicates a bias against guanine (see Modeltestparameters in methods) which is particularly strongin third codon positions (9) Most of the variabilityamong cytochrome b sequences was detected at thirdcodon positions No saturation was detected at anycodon position (Fig 2) However separate plots of AndashG
Figure 2 Relationships between uncorrected meansequence divergence (p-distance) and number of transi-tions () and transversions () at all codon positions (A)and at third codon positions (B) in the cytochrome b genefor all pairwise comparisons for Mexican populationsamples
020406080
100120140160180
005 01 015 02p
Cha
nges
020406080
100120140160
005 01 015 02
p
Cha
nges
0
0
A
B
328 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
and CndashT transitional divergences at third codon posi-tions suggested that AndashG transitions changed fasterthan CndashT ones (data not shown)
Pairwise sequence divergence (uncorrected p-distance) among northern species of the genus Notro-pis analysed ranged from 39 (between N blenniusand N potteri) to 155 (N potteri and N volucellus)Among Mexican species net sequence divergenceranged from 39 (between specimens of N moralesiand N boucardi) to 115 (between specimens ofN sallaei and N boucardi) (Table 2) Within Mexicanspecies the mean divergence among specimensrepresenting different populations was lt1 (amongN imeldae specimens and among specimens of geo-graphically close populations of N moralesi) to 3(among specimens of regionally separated populationsof N moralesi) Nucleotide diversity (π) within mostMexican species ranged from 0005 to 0015 Howeverfour specimens from two different sample sites ofN calientis fell outside this range showing over 7sequence divergence and nucleotide diversity up to0046 (Table 2)
Cytochrome b sequences were analysed using fourcommon methods of phylogenetic inference (MP NJML BI) and recovered congruent topologies (Figs 3and 4) Topologies recovered from unweighted andweighted most parsimonious trees were similar butthe weighted analyses yielded a more highly resolvedtopology congruent to the remaining recovered analy-ses When one specimen per species was taken intoaccount in order to place the Mexican species into alarger phylogenetic framework all cyprinids of thegenera Notropis and Hybopsis were grouped togetherwith Pteronotropis and Cyprinella in a basal positionAccording to MP ML NJ and BI all Mexican speciesof the genera Notropis and Yuriria formed a well-supported clade with respect to the remaining speciesof the genus Notropis from the USA and Canada North-ern species of the genus Notropis formed three differ-ent well-supported clades while basal relationshipsamong clades remained unresolved The phylogeneticposition of the genus Hybopsis remained unresolved(Fig 3) Interestingly the genus Yuriria groupedtogether with all Mexican Notropis in all analysesThe robustness of these results was confirmed by highbootstrap values and Bayesian posterior probability
When populations were taken into account all Mex-ican Notropis plus Yuriria clustered together withrespect to the northern Notropis Hybopsis and Cyp-rinella forming a well-supported clade with bootstrapvalues (MP 94 ML 72 NJ 86) and BI (100)(Fig 4) Within this clade four well-supported lin-eages could be distinguished (Fig 4) (A) grouped allspecimens representing populations of N boucardiand N moralesi (B) grouped N imeldae and N sp 1(C) was formed by all specimens of N sallaei and
N calientis (D) included the Y alta group The genusYuriria was grouped with N calientis-N sallaei as sis-ter species using NJ (85) and ML (74) (Figs 3 4)The relative position of the clade N imeldae-N sp 1was different depending on the phylogenetic analysisaccording to MP (79) ML (58) and BI (100) it wasconsidered a sister group of the N moralesi-N boucardi clade whereas it appeared as a sistergroup of the clade (Yuriria (N calientis N sallaei)) inthe NJ analysis (66) (Fig 4) In order to test the twoalternative hypotheses we performed a ShimodairandashHasegawa test no statistically significant differenceswere observed between the two hypotheses
SOUTHERN MEXICAN NOTROPIS
The populations analysed which have been tradition-ally assigned to N boucardi and N moralesi formedtwo well-supported clades (mean sequence divergenceamong them was 507 ranging from 43 to 60 andnet sequence divergence (dA) was 395) One cladecontained 14 specimens belonging to 13 populations ofN moralesi which inhabit three different drainages(Atoyac Balsas and Papaloapan) and include speci-mens sampled in the type locality of the species Meansequence divergence among specimens of N moralesi(137) ranged from 02 to 30 (nucleotide diversity(π) was 0013) This clade was the sister group of spec-imens of N boucardi (Cuernavaca Balsas drainage)(π 0008) (Fig 4)
The remaining members of the southern MexicanNotropis formed another well-supported clade whichincluded all specimens of N imeldae from differentrivers of the Atoyac drainage and a population of Not-ropis sp 1 from an isolated spring of the Balsas drain-age (Oaxaca State) N sp 1 was identified astaxonomically distinct on the basis of isozyme data(Schoumlnhuth et al 2001) and awaits a formal descrip-tion (Schoumlnhuth 2002) Mean sequence divergencebetween N imeldae and N sp 1 was 7 (dA 67) Netsequence divergence between both species and othersouth Mexican Notropis ranged from 99 to 118 Netsequence divergence between the N imeldae-N sp 1clade and central Mexican cyprinids ranged from 80to 96 (Table 2)
CENTRAL MEXICAN CYPRINIDS
N calientis N sallaei and Y alta are distributed overthe trans-Mexican volcanic belt in central Mexicoinhabiting the Riacuteo Lerma drainage the lagoons of theMesa Central and the Riacuteo Paacutenuco headwaters A cladecomposed of these species was well-supported in theML (74) and NJ (85) analyses Mean sequencedivergence between N sallaei and N calientis (921)ranged from 85 to 101 (dA 608) and mean
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 329
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 2
R
ange
an
d m
ean
(S
D)
of p
erce
nta
ge g
enet
ic d
iver
gen
ce f
or t
he
intr
aspe
cifi
c (d
iago
nal
) an
d in
ters
peci
fic
(abo
ve d
iago
nal
) co
mpa
riso
ns
amon
g sp
ecim
ens
anal
ysed
for
cyt
och
rom
e b
base
d on
un
corr
ecte
d p-
dist
ance
s L
ast
row
in
dia
gon
al (
bold
) sh
ows
nu
cleo
tide
div
ersi
ty (
π) w
ith
in s
peci
es
Net
seq
uen
ce d
iver
gen
ce(d
A)
is b
elow
th
e di
agon
al
Mex
ican
min
now
sM
inn
ows
from
US
A
and
Can
ada
N b
ouca
rdi
N m
oral
esi
N i
mel
dae
N s
p 1
N c
alie
nti
sN
sal
laei
Y a
lta
Not
ropi
sH
ybop
sis
N b
ouca
rdi
08
43ndash
60
111
ndash11
412
1ndash1
23
128
ndash13
712
5ndash1
29
121
ndash12
813
8ndash1
63
142
ndash15
10
875
07 (
04)
112
7 (0
1)
122
7 (0
1)
133
7 (0
3)
127
5 (0
1)
125
2 (0
1)
152
3 (0
6)
147
8 (0
4)
000
8N
mor
ales
i3
950
2ndash3
010
2ndash1
18
113
ndash11
913
1ndash1
52
122
ndash13
511
9ndash1
36
129
ndash16
413
9ndash1
48
137
(0
6)10
95
(03
)11
65
(02
)14
26
(04
)12
86
(03
)12
61
(03
)14
96
(07
)14
30
(03
)0
013
N i
mel
dae
105
59
980
1ndash0
80
58 (
03)
000
5
70ndash
71
706
(0
1)10
8ndash1
17
114
0 (0
2)
99ndash
104
100
9 (0
2)
94ndash
100
984
(0
2)11
7ndash1
46
134
5 (0
9)
123
ndash12
912
65
(02
)
N s
p 1
118
410
97
677
010
3ndash1
11
106
9 (0
3)
95ndash
98
968
(0
1)10
6ndash1
12
109
8 (0
2)
122
ndash15
013
79
(08
)13
7ndash1
39
138
5 (0
1)
N c
alie
nti
s10
59
112
38
068
351
4ndash7
04
69 (
22)
004
6
85ndash
101
921
(0
4)10
4ndash1
23
110
8 (0
5)
121
ndash16
014
04
(09
)12
7ndash1
40
133
8 (0
4)
N s
alla
ei11
53
113
99
69
296
081
51
570
015
90ndash
94
927
(0
1)12
0ndash1
43
128
7 (0
6)
126
13
312
98
(02
)
Y a
lta
113
511
19
859
923
87
750
9ndash1
91
47 (
03)
001
4
113
ndash15
113
25
(09
)13
5ndash1
43
139
6 (0
2)
Not
ropi
s3
9ndash15
512
25
(24
)12
2ndash1
53
137
8 (0
8)
Hyb
opsi
s7
47
45
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

326 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Fritsch amp Maniatis 1989) The DNA was resuspendedin ultrapure water and used for amplification by PCR
AMPLIFICATION AND CLONING
Combinations of two sets of versatile primers (Zardoyaamp Doadrio 1998) were used in PCR amplifications toisolate two contiguous and overlapping fragments (of660 and 521 bp) that covered the entire cytochrome bgene The PCR reactions were conducted in 25 microl vol-umes (67 mM Tris-HCl pH 83 15 mM MgCl2 04 mM
each primer 1 microl DNA extraction and 1 unit Taq DNApolymerase (Promega)) After an initial denaturationstep at 94degC for 2 min 35 cycles were performed as fol-lows denaturation at 94degC (1 min) annealing at 48degC(1 min) and extension at 72degC (145 min) with a finalextension of 7 min at 72degC
After PCR amplification the DNA was precipitatedcloned into p-GEMT vectors and transformed intocompetent E coli (JM109) Positive clones weresequenced using the FS-Taq Dye Deoxy terminatorcycle sequencing kit (Applied Biosystems Inc) usingan automated DNA sequencer (Applied Biosystems3700) following the manufacturerrsquos instructions TheDNA sequences of both strands were obtained using
M13 universal (forward and reverse) sequencing prim-ers The sequences determined here have been depos-ited in GenBank under accession numbers AF469130ndashAF469164 AY140697 and AY140698
CYTOCHROME b SEQUENCE ANALYSES
Forty-nine complete minnow cytochrome b genesequences (1141 bp) were aligned manually withsequences of N atherinoides H amblops Cyprinellaspiloptera Tinca tinca and Barbus sclateri No ambig-uous alignments or gaps were found All codon posi-tions were included in the phylogenetic analyses
All Mexican specimens representing 30 populationsof seven minnow species were used to determine theirpopulation structure (Table 1) The intrapopulationaldivergence levels were corrected for ancestral poly-morphisms to avoid the effect of inflating divergencebetween phylogroups or species We applied the netsequence divergence correction described by Nei(1987) and the index of nucleotide diversity (π) (Nei ampLi 1979) using DnaSP ver 350 (Rozas amp Rozas 1999)Higher level relationships were inferred using 22 spe-cies of Mexican USA and Canadian NotropisHybop-sis and two species of the phylogenetically related
Figure 1 Localities from which specimens of the genera Notropis and Yuriria were sampled Numbers indicate collectionsample (see Table 1) Drainages are indicated by letters A Atoyac B Balsas C Lerma D Paacutenuco E PapaloapanDistribution range of each species is presented in different grey patterns
115degW
30degN
25deg
20deg
15deg
110deg 105deg 100deg 95deg 0 200 km
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 327
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
genera Cyprinella and Pteronotropis included in theNotropin clade (Phoxinini lineage Shiner clade)(Coburn amp Cavender 1992) The genus Moapa fromthe Western clade Phoxinini was selected as a taxo-nomic related outgroup whereas Tinca was chosen asa more distant outgroup
Phylogenetic trees were recovered using maximumparsimony (MP) neighbour joining (NJ) maximumlikelihood (ML) and Bayesian inference (BI) We usedfour different methods for inferring phylogenies to testthe congruence among methods and compare for thebest approximation for our data There has recentlybeen controversy about the overcredibility of molecu-lar phylogenies obtained with Bayesian inference(Suzuki Glazco amp Nei 2002) while others have sug-gested posterior probability as a better indicator ofstatistical confidence than bootstrap probability (Wil-cox et al 2002) Both bootstrap and posterior proba-bility were included It is well known that MP givesgood results in the Felsenstein inverse zone and MLin both zones when more than 1000 nucleotides areanalysed However accuracy decreases when theinternal nodes are short (Swofford et al 2001) Theanalysis of 1141 nucleotides resulted in short internalnodes within trees so we decided to include the fourmethods of analysis to infer the phylogeny
The MP analysis was performed through heuristicsearches with ten random stepwise additions of taxaMULTREE option and TBR branch swapping For theMP analyses equal weight of transversions (Tv) andtransitions (Ts) or Tv five times the weight of Ts basedon empirical evidence were used For ML BI and NJanalyses we used the hierarchical likelihood ratio test(LRT) implemented in Modeltest V304 (Posada ampCrandall 1998) to find the evolutionary model thatbest fit our data The HKY + I + G model (base fre-quencies A 03294 C 02805 G 01433 T 02469Nst 2 TsTv ratio 56889 Γ 09381 proportion ofinvariable sites 04383) was selected when Mexicanminnow populations were taken into account and theGTR + I + G model (base frequencies A 02853 C03302 G 01125 T 02720 Nst 6 Γ 07619 propor-tion of invariable sites 04915) was selected when onespecimen per species was analysed
All phylogenetic analyses were performed usingPAUP version 40b8 (Swofford 2001) and MR BAYES(Huelsenbeck amp Ronquist 2001) Robustness of theinferred trees was tested by bootstrapping (Felsen-stein 1985) with 100 pseudoreplications for MP and1000 for NJ with 1000 quartet puzzling steps forML and 500 000 generations for Bayesian posteriorprobability
Our two alternative phylogenetic hypotheses forN imeldae-N sp 1 clade were tested using the Shimo-dairandashHasegawa test (Shimodaira amp Hasegawa 1999)as implemented in PAUP
The genetic divergences widely discussed throughthe text were based on the uncorrected p-distances Toavoid confusion between derived divergences based onthe different evolutionary models selected (HKY +I + G and GTR + I + G) we considered more appropri-ate the uncorrected p genetic distances which allowscomparisons among our two data sets and could permita broad comparison with other related taxa thatshowed a different model of evolution
RESULTS
The complete cytochrome b gene was successfullyamplified and sequenced in 37 specimens of the generaNotropis and Yuriria Of the 1141 bp aligned for alltaxa 488 sites were variable and 373 were parsimonyinformative when only Mexican minnow populationswere taken into account Informative characters roseto 387 when one specimen per species for all NorthAmerican minnows analysed was considered
The nucleotide composition in the cytochrome bgene indicates a bias against guanine (see Modeltestparameters in methods) which is particularly strongin third codon positions (9) Most of the variabilityamong cytochrome b sequences was detected at thirdcodon positions No saturation was detected at anycodon position (Fig 2) However separate plots of AndashG
Figure 2 Relationships between uncorrected meansequence divergence (p-distance) and number of transi-tions () and transversions () at all codon positions (A)and at third codon positions (B) in the cytochrome b genefor all pairwise comparisons for Mexican populationsamples
020406080
100120140160180
005 01 015 02p
Cha
nges
020406080
100120140160
005 01 015 02
p
Cha
nges
0
0
A
B
328 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
and CndashT transitional divergences at third codon posi-tions suggested that AndashG transitions changed fasterthan CndashT ones (data not shown)
Pairwise sequence divergence (uncorrected p-distance) among northern species of the genus Notro-pis analysed ranged from 39 (between N blenniusand N potteri) to 155 (N potteri and N volucellus)Among Mexican species net sequence divergenceranged from 39 (between specimens of N moralesiand N boucardi) to 115 (between specimens ofN sallaei and N boucardi) (Table 2) Within Mexicanspecies the mean divergence among specimensrepresenting different populations was lt1 (amongN imeldae specimens and among specimens of geo-graphically close populations of N moralesi) to 3(among specimens of regionally separated populationsof N moralesi) Nucleotide diversity (π) within mostMexican species ranged from 0005 to 0015 Howeverfour specimens from two different sample sites ofN calientis fell outside this range showing over 7sequence divergence and nucleotide diversity up to0046 (Table 2)
Cytochrome b sequences were analysed using fourcommon methods of phylogenetic inference (MP NJML BI) and recovered congruent topologies (Figs 3and 4) Topologies recovered from unweighted andweighted most parsimonious trees were similar butthe weighted analyses yielded a more highly resolvedtopology congruent to the remaining recovered analy-ses When one specimen per species was taken intoaccount in order to place the Mexican species into alarger phylogenetic framework all cyprinids of thegenera Notropis and Hybopsis were grouped togetherwith Pteronotropis and Cyprinella in a basal positionAccording to MP ML NJ and BI all Mexican speciesof the genera Notropis and Yuriria formed a well-supported clade with respect to the remaining speciesof the genus Notropis from the USA and Canada North-ern species of the genus Notropis formed three differ-ent well-supported clades while basal relationshipsamong clades remained unresolved The phylogeneticposition of the genus Hybopsis remained unresolved(Fig 3) Interestingly the genus Yuriria groupedtogether with all Mexican Notropis in all analysesThe robustness of these results was confirmed by highbootstrap values and Bayesian posterior probability
When populations were taken into account all Mex-ican Notropis plus Yuriria clustered together withrespect to the northern Notropis Hybopsis and Cyp-rinella forming a well-supported clade with bootstrapvalues (MP 94 ML 72 NJ 86) and BI (100)(Fig 4) Within this clade four well-supported lin-eages could be distinguished (Fig 4) (A) grouped allspecimens representing populations of N boucardiand N moralesi (B) grouped N imeldae and N sp 1(C) was formed by all specimens of N sallaei and
N calientis (D) included the Y alta group The genusYuriria was grouped with N calientis-N sallaei as sis-ter species using NJ (85) and ML (74) (Figs 3 4)The relative position of the clade N imeldae-N sp 1was different depending on the phylogenetic analysisaccording to MP (79) ML (58) and BI (100) it wasconsidered a sister group of the N moralesi-N boucardi clade whereas it appeared as a sistergroup of the clade (Yuriria (N calientis N sallaei)) inthe NJ analysis (66) (Fig 4) In order to test the twoalternative hypotheses we performed a ShimodairandashHasegawa test no statistically significant differenceswere observed between the two hypotheses
SOUTHERN MEXICAN NOTROPIS
The populations analysed which have been tradition-ally assigned to N boucardi and N moralesi formedtwo well-supported clades (mean sequence divergenceamong them was 507 ranging from 43 to 60 andnet sequence divergence (dA) was 395) One cladecontained 14 specimens belonging to 13 populations ofN moralesi which inhabit three different drainages(Atoyac Balsas and Papaloapan) and include speci-mens sampled in the type locality of the species Meansequence divergence among specimens of N moralesi(137) ranged from 02 to 30 (nucleotide diversity(π) was 0013) This clade was the sister group of spec-imens of N boucardi (Cuernavaca Balsas drainage)(π 0008) (Fig 4)
The remaining members of the southern MexicanNotropis formed another well-supported clade whichincluded all specimens of N imeldae from differentrivers of the Atoyac drainage and a population of Not-ropis sp 1 from an isolated spring of the Balsas drain-age (Oaxaca State) N sp 1 was identified astaxonomically distinct on the basis of isozyme data(Schoumlnhuth et al 2001) and awaits a formal descrip-tion (Schoumlnhuth 2002) Mean sequence divergencebetween N imeldae and N sp 1 was 7 (dA 67) Netsequence divergence between both species and othersouth Mexican Notropis ranged from 99 to 118 Netsequence divergence between the N imeldae-N sp 1clade and central Mexican cyprinids ranged from 80to 96 (Table 2)
CENTRAL MEXICAN CYPRINIDS
N calientis N sallaei and Y alta are distributed overthe trans-Mexican volcanic belt in central Mexicoinhabiting the Riacuteo Lerma drainage the lagoons of theMesa Central and the Riacuteo Paacutenuco headwaters A cladecomposed of these species was well-supported in theML (74) and NJ (85) analyses Mean sequencedivergence between N sallaei and N calientis (921)ranged from 85 to 101 (dA 608) and mean
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 329
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 2
R
ange
an
d m
ean
(S
D)
of p
erce
nta
ge g
enet
ic d
iver
gen
ce f
or t
he
intr
aspe
cifi
c (d
iago
nal
) an
d in
ters
peci
fic
(abo
ve d
iago
nal
) co
mpa
riso
ns
amon
g sp
ecim
ens
anal
ysed
for
cyt
och
rom
e b
base
d on
un
corr
ecte
d p-
dist
ance
s L
ast
row
in
dia
gon
al (
bold
) sh
ows
nu
cleo
tide
div
ersi
ty (
π) w
ith
in s
peci
es
Net
seq
uen
ce d
iver
gen
ce(d
A)
is b
elow
th
e di
agon
al
Mex
ican
min
now
sM
inn
ows
from
US
A
and
Can
ada
N b
ouca
rdi
N m
oral
esi
N i
mel
dae
N s
p 1
N c
alie
nti
sN
sal
laei
Y a
lta
Not
ropi
sH
ybop
sis
N b
ouca
rdi
08
43ndash
60
111
ndash11
412
1ndash1
23
128
ndash13
712
5ndash1
29
121
ndash12
813
8ndash1
63
142
ndash15
10
875
07 (
04)
112
7 (0
1)
122
7 (0
1)
133
7 (0
3)
127
5 (0
1)
125
2 (0
1)
152
3 (0
6)
147
8 (0
4)
000
8N
mor
ales
i3
950
2ndash3
010
2ndash1
18
113
ndash11
913
1ndash1
52
122
ndash13
511
9ndash1
36
129
ndash16
413
9ndash1
48
137
(0
6)10
95
(03
)11
65
(02
)14
26
(04
)12
86
(03
)12
61
(03
)14
96
(07
)14
30
(03
)0
013
N i
mel
dae
105
59
980
1ndash0
80
58 (
03)
000
5
70ndash
71
706
(0
1)10
8ndash1
17
114
0 (0
2)
99ndash
104
100
9 (0
2)
94ndash
100
984
(0
2)11
7ndash1
46
134
5 (0
9)
123
ndash12
912
65
(02
)
N s
p 1
118
410
97
677
010
3ndash1
11
106
9 (0
3)
95ndash
98
968
(0
1)10
6ndash1
12
109
8 (0
2)
122
ndash15
013
79
(08
)13
7ndash1
39
138
5 (0
1)
N c
alie
nti
s10
59
112
38
068
351
4ndash7
04
69 (
22)
004
6
85ndash
101
921
(0
4)10
4ndash1
23
110
8 (0
5)
121
ndash16
014
04
(09
)12
7ndash1
40
133
8 (0
4)
N s
alla
ei11
53
113
99
69
296
081
51
570
015
90ndash
94
927
(0
1)12
0ndash1
43
128
7 (0
6)
126
13
312
98
(02
)
Y a
lta
113
511
19
859
923
87
750
9ndash1
91
47 (
03)
001
4
113
ndash15
113
25
(09
)13
5ndash1
43
139
6 (0
2)
Not
ropi
s3
9ndash15
512
25
(24
)12
2ndash1
53
137
8 (0
8)
Hyb
opsi
s7
47
45
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237
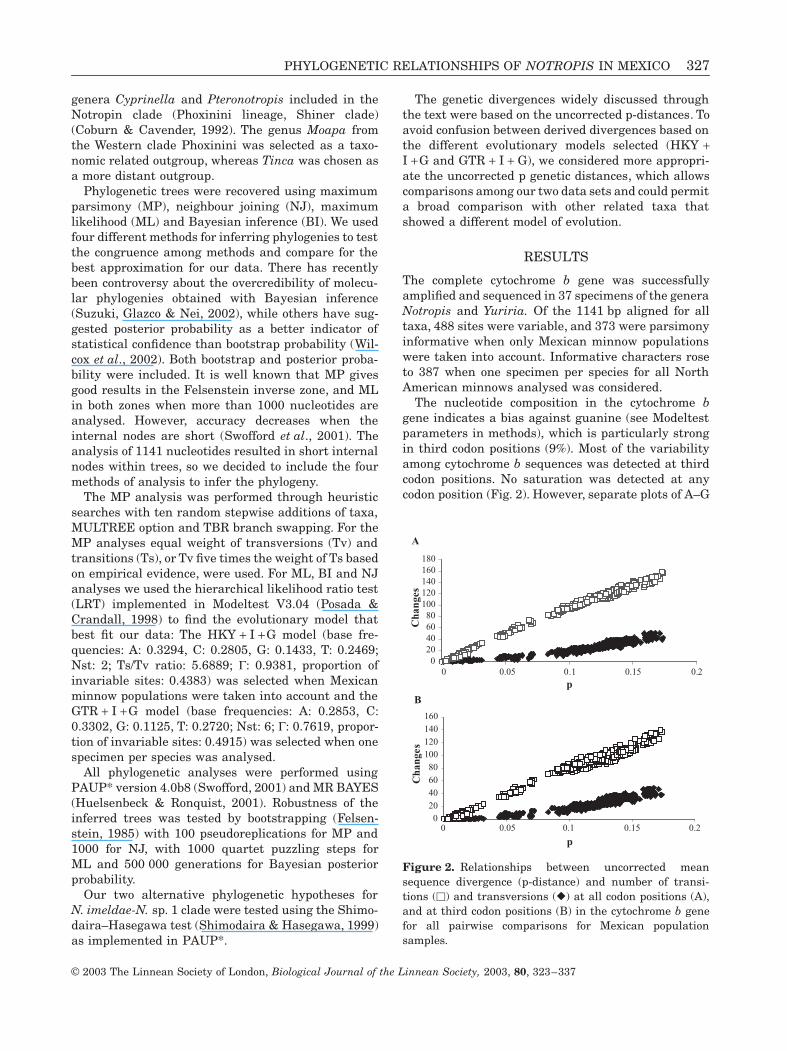
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 327
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
genera Cyprinella and Pteronotropis included in theNotropin clade (Phoxinini lineage Shiner clade)(Coburn amp Cavender 1992) The genus Moapa fromthe Western clade Phoxinini was selected as a taxo-nomic related outgroup whereas Tinca was chosen asa more distant outgroup
Phylogenetic trees were recovered using maximumparsimony (MP) neighbour joining (NJ) maximumlikelihood (ML) and Bayesian inference (BI) We usedfour different methods for inferring phylogenies to testthe congruence among methods and compare for thebest approximation for our data There has recentlybeen controversy about the overcredibility of molecu-lar phylogenies obtained with Bayesian inference(Suzuki Glazco amp Nei 2002) while others have sug-gested posterior probability as a better indicator ofstatistical confidence than bootstrap probability (Wil-cox et al 2002) Both bootstrap and posterior proba-bility were included It is well known that MP givesgood results in the Felsenstein inverse zone and MLin both zones when more than 1000 nucleotides areanalysed However accuracy decreases when theinternal nodes are short (Swofford et al 2001) Theanalysis of 1141 nucleotides resulted in short internalnodes within trees so we decided to include the fourmethods of analysis to infer the phylogeny
The MP analysis was performed through heuristicsearches with ten random stepwise additions of taxaMULTREE option and TBR branch swapping For theMP analyses equal weight of transversions (Tv) andtransitions (Ts) or Tv five times the weight of Ts basedon empirical evidence were used For ML BI and NJanalyses we used the hierarchical likelihood ratio test(LRT) implemented in Modeltest V304 (Posada ampCrandall 1998) to find the evolutionary model thatbest fit our data The HKY + I + G model (base fre-quencies A 03294 C 02805 G 01433 T 02469Nst 2 TsTv ratio 56889 Γ 09381 proportion ofinvariable sites 04383) was selected when Mexicanminnow populations were taken into account and theGTR + I + G model (base frequencies A 02853 C03302 G 01125 T 02720 Nst 6 Γ 07619 propor-tion of invariable sites 04915) was selected when onespecimen per species was analysed
All phylogenetic analyses were performed usingPAUP version 40b8 (Swofford 2001) and MR BAYES(Huelsenbeck amp Ronquist 2001) Robustness of theinferred trees was tested by bootstrapping (Felsen-stein 1985) with 100 pseudoreplications for MP and1000 for NJ with 1000 quartet puzzling steps forML and 500 000 generations for Bayesian posteriorprobability
Our two alternative phylogenetic hypotheses forN imeldae-N sp 1 clade were tested using the Shimo-dairandashHasegawa test (Shimodaira amp Hasegawa 1999)as implemented in PAUP
The genetic divergences widely discussed throughthe text were based on the uncorrected p-distances Toavoid confusion between derived divergences based onthe different evolutionary models selected (HKY +I + G and GTR + I + G) we considered more appropri-ate the uncorrected p genetic distances which allowscomparisons among our two data sets and could permita broad comparison with other related taxa thatshowed a different model of evolution
RESULTS
The complete cytochrome b gene was successfullyamplified and sequenced in 37 specimens of the generaNotropis and Yuriria Of the 1141 bp aligned for alltaxa 488 sites were variable and 373 were parsimonyinformative when only Mexican minnow populationswere taken into account Informative characters roseto 387 when one specimen per species for all NorthAmerican minnows analysed was considered
The nucleotide composition in the cytochrome bgene indicates a bias against guanine (see Modeltestparameters in methods) which is particularly strongin third codon positions (9) Most of the variabilityamong cytochrome b sequences was detected at thirdcodon positions No saturation was detected at anycodon position (Fig 2) However separate plots of AndashG
Figure 2 Relationships between uncorrected meansequence divergence (p-distance) and number of transi-tions () and transversions () at all codon positions (A)and at third codon positions (B) in the cytochrome b genefor all pairwise comparisons for Mexican populationsamples
020406080
100120140160180
005 01 015 02p
Cha
nges
020406080
100120140160
005 01 015 02
p
Cha
nges
0
0
A
B
328 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
and CndashT transitional divergences at third codon posi-tions suggested that AndashG transitions changed fasterthan CndashT ones (data not shown)
Pairwise sequence divergence (uncorrected p-distance) among northern species of the genus Notro-pis analysed ranged from 39 (between N blenniusand N potteri) to 155 (N potteri and N volucellus)Among Mexican species net sequence divergenceranged from 39 (between specimens of N moralesiand N boucardi) to 115 (between specimens ofN sallaei and N boucardi) (Table 2) Within Mexicanspecies the mean divergence among specimensrepresenting different populations was lt1 (amongN imeldae specimens and among specimens of geo-graphically close populations of N moralesi) to 3(among specimens of regionally separated populationsof N moralesi) Nucleotide diversity (π) within mostMexican species ranged from 0005 to 0015 Howeverfour specimens from two different sample sites ofN calientis fell outside this range showing over 7sequence divergence and nucleotide diversity up to0046 (Table 2)
Cytochrome b sequences were analysed using fourcommon methods of phylogenetic inference (MP NJML BI) and recovered congruent topologies (Figs 3and 4) Topologies recovered from unweighted andweighted most parsimonious trees were similar butthe weighted analyses yielded a more highly resolvedtopology congruent to the remaining recovered analy-ses When one specimen per species was taken intoaccount in order to place the Mexican species into alarger phylogenetic framework all cyprinids of thegenera Notropis and Hybopsis were grouped togetherwith Pteronotropis and Cyprinella in a basal positionAccording to MP ML NJ and BI all Mexican speciesof the genera Notropis and Yuriria formed a well-supported clade with respect to the remaining speciesof the genus Notropis from the USA and Canada North-ern species of the genus Notropis formed three differ-ent well-supported clades while basal relationshipsamong clades remained unresolved The phylogeneticposition of the genus Hybopsis remained unresolved(Fig 3) Interestingly the genus Yuriria groupedtogether with all Mexican Notropis in all analysesThe robustness of these results was confirmed by highbootstrap values and Bayesian posterior probability
When populations were taken into account all Mex-ican Notropis plus Yuriria clustered together withrespect to the northern Notropis Hybopsis and Cyp-rinella forming a well-supported clade with bootstrapvalues (MP 94 ML 72 NJ 86) and BI (100)(Fig 4) Within this clade four well-supported lin-eages could be distinguished (Fig 4) (A) grouped allspecimens representing populations of N boucardiand N moralesi (B) grouped N imeldae and N sp 1(C) was formed by all specimens of N sallaei and
N calientis (D) included the Y alta group The genusYuriria was grouped with N calientis-N sallaei as sis-ter species using NJ (85) and ML (74) (Figs 3 4)The relative position of the clade N imeldae-N sp 1was different depending on the phylogenetic analysisaccording to MP (79) ML (58) and BI (100) it wasconsidered a sister group of the N moralesi-N boucardi clade whereas it appeared as a sistergroup of the clade (Yuriria (N calientis N sallaei)) inthe NJ analysis (66) (Fig 4) In order to test the twoalternative hypotheses we performed a ShimodairandashHasegawa test no statistically significant differenceswere observed between the two hypotheses
SOUTHERN MEXICAN NOTROPIS
The populations analysed which have been tradition-ally assigned to N boucardi and N moralesi formedtwo well-supported clades (mean sequence divergenceamong them was 507 ranging from 43 to 60 andnet sequence divergence (dA) was 395) One cladecontained 14 specimens belonging to 13 populations ofN moralesi which inhabit three different drainages(Atoyac Balsas and Papaloapan) and include speci-mens sampled in the type locality of the species Meansequence divergence among specimens of N moralesi(137) ranged from 02 to 30 (nucleotide diversity(π) was 0013) This clade was the sister group of spec-imens of N boucardi (Cuernavaca Balsas drainage)(π 0008) (Fig 4)
The remaining members of the southern MexicanNotropis formed another well-supported clade whichincluded all specimens of N imeldae from differentrivers of the Atoyac drainage and a population of Not-ropis sp 1 from an isolated spring of the Balsas drain-age (Oaxaca State) N sp 1 was identified astaxonomically distinct on the basis of isozyme data(Schoumlnhuth et al 2001) and awaits a formal descrip-tion (Schoumlnhuth 2002) Mean sequence divergencebetween N imeldae and N sp 1 was 7 (dA 67) Netsequence divergence between both species and othersouth Mexican Notropis ranged from 99 to 118 Netsequence divergence between the N imeldae-N sp 1clade and central Mexican cyprinids ranged from 80to 96 (Table 2)
CENTRAL MEXICAN CYPRINIDS
N calientis N sallaei and Y alta are distributed overthe trans-Mexican volcanic belt in central Mexicoinhabiting the Riacuteo Lerma drainage the lagoons of theMesa Central and the Riacuteo Paacutenuco headwaters A cladecomposed of these species was well-supported in theML (74) and NJ (85) analyses Mean sequencedivergence between N sallaei and N calientis (921)ranged from 85 to 101 (dA 608) and mean
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 329
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 2
R
ange
an
d m
ean
(S
D)
of p
erce
nta
ge g
enet
ic d
iver
gen
ce f
or t
he
intr
aspe
cifi
c (d
iago
nal
) an
d in
ters
peci
fic
(abo
ve d
iago
nal
) co
mpa
riso
ns
amon
g sp
ecim
ens
anal
ysed
for
cyt
och
rom
e b
base
d on
un
corr
ecte
d p-
dist
ance
s L
ast
row
in
dia
gon
al (
bold
) sh
ows
nu
cleo
tide
div
ersi
ty (
π) w
ith
in s
peci
es
Net
seq
uen
ce d
iver
gen
ce(d
A)
is b
elow
th
e di
agon
al
Mex
ican
min
now
sM
inn
ows
from
US
A
and
Can
ada
N b
ouca
rdi
N m
oral
esi
N i
mel
dae
N s
p 1
N c
alie
nti
sN
sal
laei
Y a
lta
Not
ropi
sH
ybop
sis
N b
ouca
rdi
08
43ndash
60
111
ndash11
412
1ndash1
23
128
ndash13
712
5ndash1
29
121
ndash12
813
8ndash1
63
142
ndash15
10
875
07 (
04)
112
7 (0
1)
122
7 (0
1)
133
7 (0
3)
127
5 (0
1)
125
2 (0
1)
152
3 (0
6)
147
8 (0
4)
000
8N
mor
ales
i3
950
2ndash3
010
2ndash1
18
113
ndash11
913
1ndash1
52
122
ndash13
511
9ndash1
36
129
ndash16
413
9ndash1
48
137
(0
6)10
95
(03
)11
65
(02
)14
26
(04
)12
86
(03
)12
61
(03
)14
96
(07
)14
30
(03
)0
013
N i
mel
dae
105
59
980
1ndash0
80
58 (
03)
000
5
70ndash
71
706
(0
1)10
8ndash1
17
114
0 (0
2)
99ndash
104
100
9 (0
2)
94ndash
100
984
(0
2)11
7ndash1
46
134
5 (0
9)
123
ndash12
912
65
(02
)
N s
p 1
118
410
97
677
010
3ndash1
11
106
9 (0
3)
95ndash
98
968
(0
1)10
6ndash1
12
109
8 (0
2)
122
ndash15
013
79
(08
)13
7ndash1
39
138
5 (0
1)
N c
alie
nti
s10
59
112
38
068
351
4ndash7
04
69 (
22)
004
6
85ndash
101
921
(0
4)10
4ndash1
23
110
8 (0
5)
121
ndash16
014
04
(09
)12
7ndash1
40
133
8 (0
4)
N s
alla
ei11
53
113
99
69
296
081
51
570
015
90ndash
94
927
(0
1)12
0ndash1
43
128
7 (0
6)
126
13
312
98
(02
)
Y a
lta
113
511
19
859
923
87
750
9ndash1
91
47 (
03)
001
4
113
ndash15
113
25
(09
)13
5ndash1
43
139
6 (0
2)
Not
ropi
s3
9ndash15
512
25
(24
)12
2ndash1
53
137
8 (0
8)
Hyb
opsi
s7
47
45
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

328 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
and CndashT transitional divergences at third codon posi-tions suggested that AndashG transitions changed fasterthan CndashT ones (data not shown)
Pairwise sequence divergence (uncorrected p-distance) among northern species of the genus Notro-pis analysed ranged from 39 (between N blenniusand N potteri) to 155 (N potteri and N volucellus)Among Mexican species net sequence divergenceranged from 39 (between specimens of N moralesiand N boucardi) to 115 (between specimens ofN sallaei and N boucardi) (Table 2) Within Mexicanspecies the mean divergence among specimensrepresenting different populations was lt1 (amongN imeldae specimens and among specimens of geo-graphically close populations of N moralesi) to 3(among specimens of regionally separated populationsof N moralesi) Nucleotide diversity (π) within mostMexican species ranged from 0005 to 0015 Howeverfour specimens from two different sample sites ofN calientis fell outside this range showing over 7sequence divergence and nucleotide diversity up to0046 (Table 2)
Cytochrome b sequences were analysed using fourcommon methods of phylogenetic inference (MP NJML BI) and recovered congruent topologies (Figs 3and 4) Topologies recovered from unweighted andweighted most parsimonious trees were similar butthe weighted analyses yielded a more highly resolvedtopology congruent to the remaining recovered analy-ses When one specimen per species was taken intoaccount in order to place the Mexican species into alarger phylogenetic framework all cyprinids of thegenera Notropis and Hybopsis were grouped togetherwith Pteronotropis and Cyprinella in a basal positionAccording to MP ML NJ and BI all Mexican speciesof the genera Notropis and Yuriria formed a well-supported clade with respect to the remaining speciesof the genus Notropis from the USA and Canada North-ern species of the genus Notropis formed three differ-ent well-supported clades while basal relationshipsamong clades remained unresolved The phylogeneticposition of the genus Hybopsis remained unresolved(Fig 3) Interestingly the genus Yuriria groupedtogether with all Mexican Notropis in all analysesThe robustness of these results was confirmed by highbootstrap values and Bayesian posterior probability
When populations were taken into account all Mex-ican Notropis plus Yuriria clustered together withrespect to the northern Notropis Hybopsis and Cyp-rinella forming a well-supported clade with bootstrapvalues (MP 94 ML 72 NJ 86) and BI (100)(Fig 4) Within this clade four well-supported lin-eages could be distinguished (Fig 4) (A) grouped allspecimens representing populations of N boucardiand N moralesi (B) grouped N imeldae and N sp 1(C) was formed by all specimens of N sallaei and
N calientis (D) included the Y alta group The genusYuriria was grouped with N calientis-N sallaei as sis-ter species using NJ (85) and ML (74) (Figs 3 4)The relative position of the clade N imeldae-N sp 1was different depending on the phylogenetic analysisaccording to MP (79) ML (58) and BI (100) it wasconsidered a sister group of the N moralesi-N boucardi clade whereas it appeared as a sistergroup of the clade (Yuriria (N calientis N sallaei)) inthe NJ analysis (66) (Fig 4) In order to test the twoalternative hypotheses we performed a ShimodairandashHasegawa test no statistically significant differenceswere observed between the two hypotheses
SOUTHERN MEXICAN NOTROPIS
The populations analysed which have been tradition-ally assigned to N boucardi and N moralesi formedtwo well-supported clades (mean sequence divergenceamong them was 507 ranging from 43 to 60 andnet sequence divergence (dA) was 395) One cladecontained 14 specimens belonging to 13 populations ofN moralesi which inhabit three different drainages(Atoyac Balsas and Papaloapan) and include speci-mens sampled in the type locality of the species Meansequence divergence among specimens of N moralesi(137) ranged from 02 to 30 (nucleotide diversity(π) was 0013) This clade was the sister group of spec-imens of N boucardi (Cuernavaca Balsas drainage)(π 0008) (Fig 4)
The remaining members of the southern MexicanNotropis formed another well-supported clade whichincluded all specimens of N imeldae from differentrivers of the Atoyac drainage and a population of Not-ropis sp 1 from an isolated spring of the Balsas drain-age (Oaxaca State) N sp 1 was identified astaxonomically distinct on the basis of isozyme data(Schoumlnhuth et al 2001) and awaits a formal descrip-tion (Schoumlnhuth 2002) Mean sequence divergencebetween N imeldae and N sp 1 was 7 (dA 67) Netsequence divergence between both species and othersouth Mexican Notropis ranged from 99 to 118 Netsequence divergence between the N imeldae-N sp 1clade and central Mexican cyprinids ranged from 80to 96 (Table 2)
CENTRAL MEXICAN CYPRINIDS
N calientis N sallaei and Y alta are distributed overthe trans-Mexican volcanic belt in central Mexicoinhabiting the Riacuteo Lerma drainage the lagoons of theMesa Central and the Riacuteo Paacutenuco headwaters A cladecomposed of these species was well-supported in theML (74) and NJ (85) analyses Mean sequencedivergence between N sallaei and N calientis (921)ranged from 85 to 101 (dA 608) and mean
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 329
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 2
R
ange
an
d m
ean
(S
D)
of p
erce
nta
ge g
enet
ic d
iver
gen
ce f
or t
he
intr
aspe
cifi
c (d
iago
nal
) an
d in
ters
peci
fic
(abo
ve d
iago
nal
) co
mpa
riso
ns
amon
g sp
ecim
ens
anal
ysed
for
cyt
och
rom
e b
base
d on
un
corr
ecte
d p-
dist
ance
s L
ast
row
in
dia
gon
al (
bold
) sh
ows
nu
cleo
tide
div
ersi
ty (
π) w
ith
in s
peci
es
Net
seq
uen
ce d
iver
gen
ce(d
A)
is b
elow
th
e di
agon
al
Mex
ican
min
now
sM
inn
ows
from
US
A
and
Can
ada
N b
ouca
rdi
N m
oral
esi
N i
mel
dae
N s
p 1
N c
alie
nti
sN
sal
laei
Y a
lta
Not
ropi
sH
ybop
sis
N b
ouca
rdi
08
43ndash
60
111
ndash11
412
1ndash1
23
128
ndash13
712
5ndash1
29
121
ndash12
813
8ndash1
63
142
ndash15
10
875
07 (
04)
112
7 (0
1)
122
7 (0
1)
133
7 (0
3)
127
5 (0
1)
125
2 (0
1)
152
3 (0
6)
147
8 (0
4)
000
8N
mor
ales
i3
950
2ndash3
010
2ndash1
18
113
ndash11
913
1ndash1
52
122
ndash13
511
9ndash1
36
129
ndash16
413
9ndash1
48
137
(0
6)10
95
(03
)11
65
(02
)14
26
(04
)12
86
(03
)12
61
(03
)14
96
(07
)14
30
(03
)0
013
N i
mel
dae
105
59
980
1ndash0
80
58 (
03)
000
5
70ndash
71
706
(0
1)10
8ndash1
17
114
0 (0
2)
99ndash
104
100
9 (0
2)
94ndash
100
984
(0
2)11
7ndash1
46
134
5 (0
9)
123
ndash12
912
65
(02
)
N s
p 1
118
410
97
677
010
3ndash1
11
106
9 (0
3)
95ndash
98
968
(0
1)10
6ndash1
12
109
8 (0
2)
122
ndash15
013
79
(08
)13
7ndash1
39
138
5 (0
1)
N c
alie
nti
s10
59
112
38
068
351
4ndash7
04
69 (
22)
004
6
85ndash
101
921
(0
4)10
4ndash1
23
110
8 (0
5)
121
ndash16
014
04
(09
)12
7ndash1
40
133
8 (0
4)
N s
alla
ei11
53
113
99
69
296
081
51
570
015
90ndash
94
927
(0
1)12
0ndash1
43
128
7 (0
6)
126
13
312
98
(02
)
Y a
lta
113
511
19
859
923
87
750
9ndash1
91
47 (
03)
001
4
113
ndash15
113
25
(09
)13
5ndash1
43
139
6 (0
2)
Not
ropi
s3
9ndash15
512
25
(24
)12
2ndash1
53
137
8 (0
8)
Hyb
opsi
s7
47
45
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237
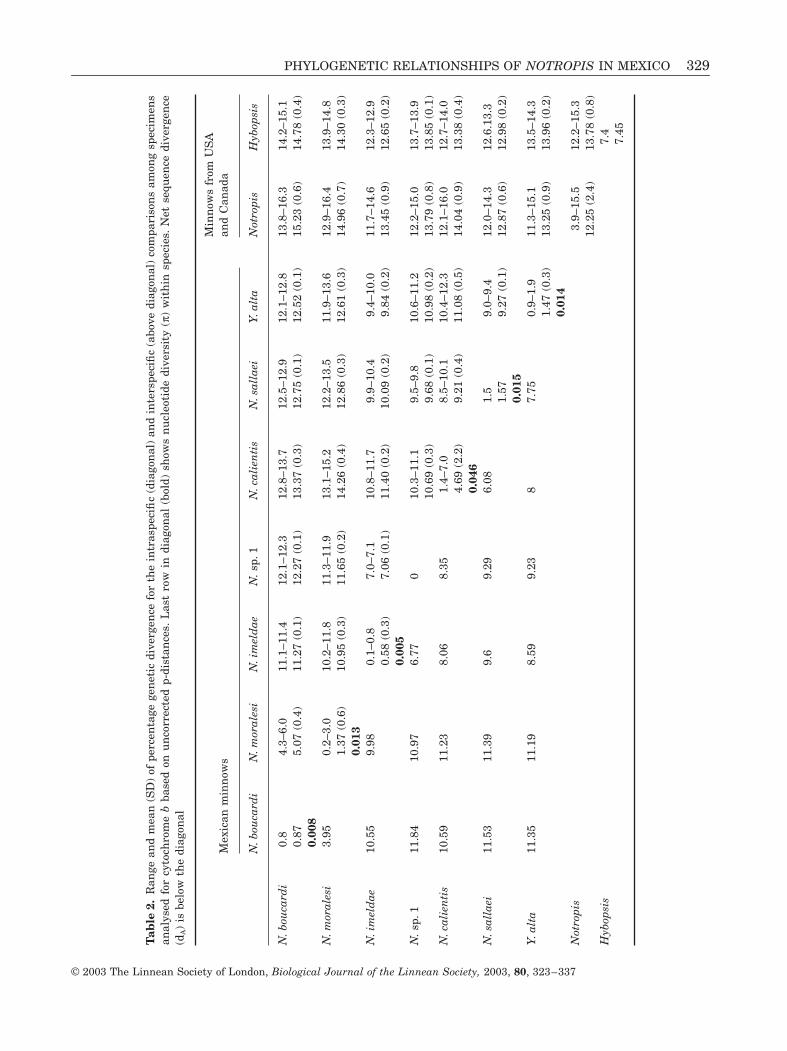
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 329
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Tab
le 2
R
ange
an
d m
ean
(S
D)
of p
erce
nta
ge g
enet
ic d
iver
gen
ce f
or t
he
intr
aspe
cifi
c (d
iago
nal
) an
d in
ters
peci
fic
(abo
ve d
iago
nal
) co
mpa
riso
ns
amon
g sp
ecim
ens
anal
ysed
for
cyt
och
rom
e b
base
d on
un
corr
ecte
d p-
dist
ance
s L
ast
row
in
dia
gon
al (
bold
) sh
ows
nu
cleo
tide
div
ersi
ty (
π) w
ith
in s
peci
es
Net
seq
uen
ce d
iver
gen
ce(d
A)
is b
elow
th
e di
agon
al
Mex
ican
min
now
sM
inn
ows
from
US
A
and
Can
ada
N b
ouca
rdi
N m
oral
esi
N i
mel
dae
N s
p 1
N c
alie
nti
sN
sal
laei
Y a
lta
Not
ropi
sH
ybop
sis
N b
ouca
rdi
08
43ndash
60
111
ndash11
412
1ndash1
23
128
ndash13
712
5ndash1
29
121
ndash12
813
8ndash1
63
142
ndash15
10
875
07 (
04)
112
7 (0
1)
122
7 (0
1)
133
7 (0
3)
127
5 (0
1)
125
2 (0
1)
152
3 (0
6)
147
8 (0
4)
000
8N
mor
ales
i3
950
2ndash3
010
2ndash1
18
113
ndash11
913
1ndash1
52
122
ndash13
511
9ndash1
36
129
ndash16
413
9ndash1
48
137
(0
6)10
95
(03
)11
65
(02
)14
26
(04
)12
86
(03
)12
61
(03
)14
96
(07
)14
30
(03
)0
013
N i
mel
dae
105
59
980
1ndash0
80
58 (
03)
000
5
70ndash
71
706
(0
1)10
8ndash1
17
114
0 (0
2)
99ndash
104
100
9 (0
2)
94ndash
100
984
(0
2)11
7ndash1
46
134
5 (0
9)
123
ndash12
912
65
(02
)
N s
p 1
118
410
97
677
010
3ndash1
11
106
9 (0
3)
95ndash
98
968
(0
1)10
6ndash1
12
109
8 (0
2)
122
ndash15
013
79
(08
)13
7ndash1
39
138
5 (0
1)
N c
alie
nti
s10
59
112
38
068
351
4ndash7
04
69 (
22)
004
6
85ndash
101
921
(0
4)10
4ndash1
23
110
8 (0
5)
121
ndash16
014
04
(09
)12
7ndash1
40
133
8 (0
4)
N s
alla
ei11
53
113
99
69
296
081
51
570
015
90ndash
94
927
(0
1)12
0ndash1
43
128
7 (0
6)
126
13
312
98
(02
)
Y a
lta
113
511
19
859
923
87
750
9ndash1
91
47 (
03)
001
4
113
ndash15
113
25
(09
)13
5ndash1
43
139
6 (0
2)
Not
ropi
s3
9ndash15
512
25
(24
)12
2ndash1
53
137
8 (0
8)
Hyb
opsi
s7
47
45
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237
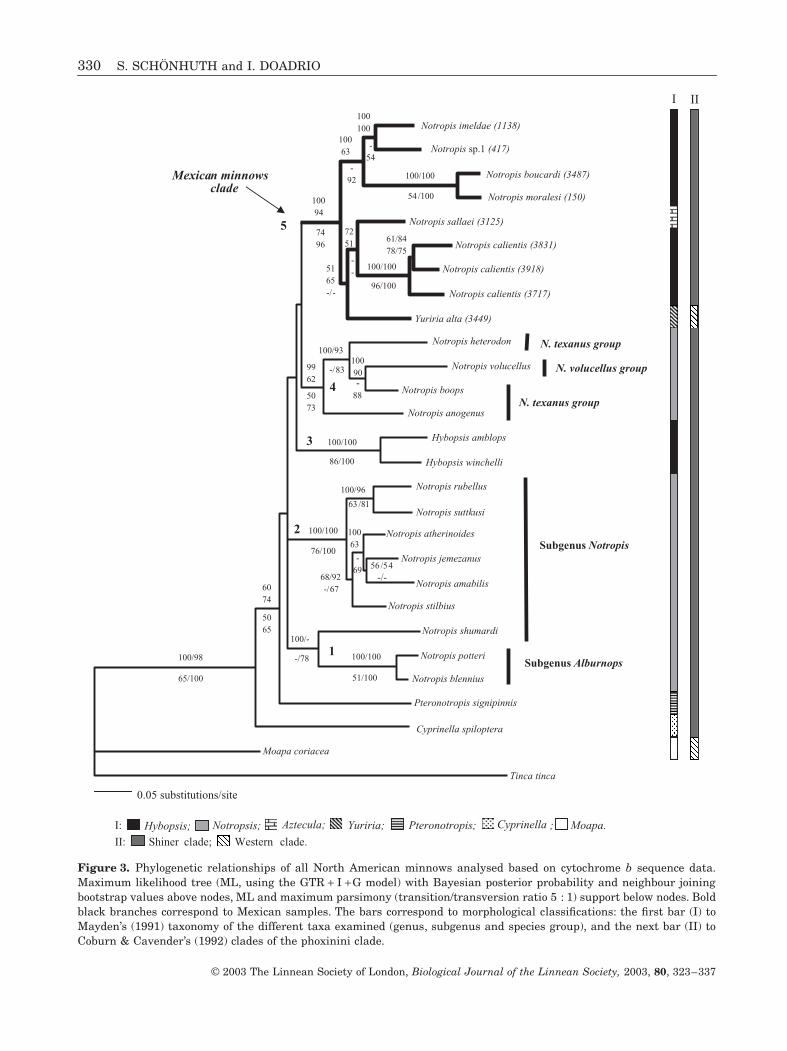
330 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 3 Phylogenetic relationships of all North American minnows analysed based on cytochrome b sequence dataMaximum likelihood tree (ML using the GTR + I + G model) with Bayesian posterior probability and neighbour joiningbootstrap values above nodes ML and maximum parsimony (transitiontransversion ratio 5 1) support below nodes Boldblack branches correspond to Mexican samples The bars correspond to morphological classifications the first bar (I) toMaydenrsquos (1991) taxonomy of the different taxa examined (genus subgenus and species group) and the next bar (II) toCoburn amp Cavenderrsquos (1992) clades of the phoxinini clade
N volucellus group
N texanus group
Cyprinella spiloptera
005 substitutionssite
Notropis imeldae (1138)
Notropis sp1 (417)
Notropis boucardi (3487)
Notropis moralesi (150)
Notropis sallaei (3125)
Notropis calientis (3831)
Notropis calientis (3918)
Notropis calientis (3717)
Yuriria alta (3449)
Notropis heterodon
Notropis volucellus
Notropis boops
Notropis anogenus
Hybopsis amblops
Hybopsis winchelli
Notropis rubellus
Notropis suttkusi
Notropis atherinoides
Notropis jemezanus
Notropis amabilis
Notropis stilbius
Notropis shumardi
Notropis potteri
Notropis blennius
Pteronotropis signipinnis
Moapa coriacea
Tinca tinca
Subgenus Alburnops
Subgenus Notropis
N texanus group
Mexican minnowsclade
5
4
3
2
1
10094
7496
10063
100100
-92
-54
100100
54 100
7251
5165--
--
100100
96100
61847875
10093
-8310090-
88
9962
5073
100100
86100
10096
63 81
100100
76100
10063
-69 56 54
--6892-67
100-
-78 100100
51100
6074
5065
10098
65100
I II
I Yuriria Pteronotropis Cyprinella Moapa
II Shiner clade Western cladeHybopsis Notropsis Aztecula
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237
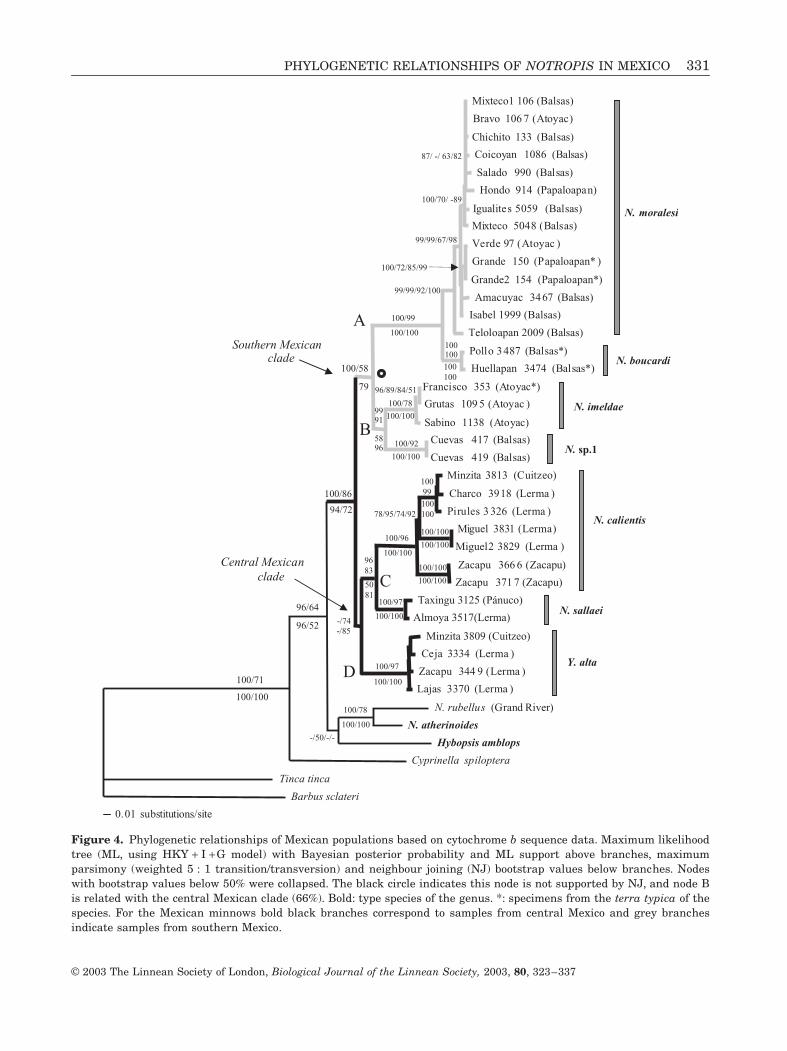
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 331
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Figure 4 Phylogenetic relationships of Mexican populations based on cytochrome b sequence data Maximum likelihoodtree (ML using HKY + I + G model) with Bayesian posterior probability and ML support above branches maximumparsimony (weighted 5 1 transitiontransversion) and neighbour joining (NJ) bootstrap values below branches Nodeswith bootstrap values below 50 were collapsed The black circle indicates this node is not supported by NJ and node Bis related with the central Mexican clade (66) Bold type species of the genus specimens from the terra typica of thespecies For the Mexican minnows bold black branches correspond to samples from central Mexico and grey branchesindicate samples from southern Mexico
Mixteco1 106 (Balsas)
Bravo 106 7 (Atoyac)
Chichito 133 (Balsas)
Coicoyan 1086 (Balsas)
Salado 990 (Balsas)
Hondo 914 (Papaloapan)
Igualites 5059 (Balsas)
Mixteco 5048 (Balsas)
Verde 97 (Atoyac )
Grande 150 (Papaloapan )
Grande2 154 (Papaloapan)
Amacuyac 3467 (Balsas)
Isabel 1999 (Balsas)
Teloloapan 2009 (Balsas)
Pollo 3487 (Balsas)
Huellapan 3474 (Balsas)
Francisco 353 (Atoyac)
Grutas 109 5 (Atoyac )
Sabino 1138 (Atoyac)
Cuevas 417 (Balsas)
Cuevas 419 (Balsas)
Minzita 3813 (Cuitzeo)
Charco 3918 (Lerma )
Pirules 3 326 (Lerma )
Miguel 3831 (Lerma)
Miguel2 3829 (Lerma )
Zacapu 366 6 (Zacapu)
Zacapu 371 7 (Zacapu)
Taxingu 3125 (Paacutenuco)
Almoya 3517(Lerma)
Minzita 3809 (Cuitzeo)
Ceja 3334 (Lerma )
Zacapu 344 9 (Lerma )
Lajas 3370 (Lerma )
N rubellus (Grand River)
N atherinoides
Hybopsis amblops
Cyprinella spiloptera
Tinca tinca
Barbus sclateri
001 substitutionssite
N moralesi
N imeldae
N boucardi
N sp1
N calientis
N sallaei
Y alta
10071
100100
9664
9652
-50--
10078
100100
10086
9472
-74-85
9683
5081
10097
100100
10097
100100
10096
100100
100100
100100
100100
100100
78957492100100
10099
10058
79
10099
100100
100100
100100
100100
10092
100100
10078
96898451
999992100
100728599
99996798
10070 -89
87 - 6382
9991
5896
Southern Mexicanclade
Central Mexicanclade
A
B
D
C
332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

332 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
sequence divergence between these and Y alta rangedfrom 90 to 123 (dA 78) N sallaei from the RiacuteoPaacutenuco showed a genetic divergence of 15 fromN sallaei from the Riacuteo Lerma (π 0015) Populationsof N calientis showed the highest intraspecific geneticdivergences ranging from 14 to 70 (π 0046)N calientis specimens were sampled in the Lermadrainage in the Cuitzeo lagoon and in the Zacapulagoon Most related populations were those fromLerma drainage (Pirules and Charco) and the Cuitzeolagoon (from Minzita) and the most divergent werethose from an isolate spring of the Lerma drainage(San Miguel) and from the Zacapu lagoon
Specimens of Y alta formed a well-supported cladeThese specimens inhabit the same areas asN calientis in the Cuitzeo lagoon and the Lermadrainage except in Zacapu where the distributionrange of Yuriria only reaches the outlet of the lagoonand no specimens were collected from this lagoon norfrom the San Miguel spring Intraspecific sequencedivergences ranged from 09 to 19 (π 0014) Min-imum sequence divergence were found between thisgenus and N sallaei or with N imeldae while maxi-mum distances were found between this species andN boucardi or N moralesi (Table 2)
DISCUSSION
Mean sequence divergence between species of Mexi-can minnows for the entire cytochrome b was 934rising to 1314 if northern species of the genus Not-ropis were included This divergence is similar to thatfound in 16 species of the subgenus Notropis from theUSA (11 plusmn 2) based on the same gene and 13 plusmn 2among subgenera of the genus Notropis (Bielawski ampGold 2001) Mean divergence among Mexican andNorthern Notropis was 138 (128ndash152) This issimilar to that found in both clades separately Thepresent findings indicate high genetic divergenceamong most of the Mexican Notropis species Meangenetic divergence between N calientis and theN moralesi-N boucardi clade (138 net sequencedivergence 109) was similar to divergences betweenthe type species of Notropis (N atherinoides) andHybopsis (H amblops) (124)
Phylogenetic analyses including one specimen perspecies in order to place the Mexican species into alarger phylogenetic framework showed strong sup-port for 5 clades (Fig 3) one clade include N potteriand N blennius sister group of N shumardi a secondclade include specimens of the subgenus Notropis(N rubellus-N suttkusi) (N stilbius (N atherinoides(N jemezanus-N amabilis))) a third clade includingboth species of the genus Hybopsis (H amblops andH winchelli) a fourth clade including 4 species of theN texanus + N volucellus species group (N anogenus
(N heterodon (N volucellus-N boops))) and a fifthclade including all Mexican species analysed(N imeldae-N sp 1) (N boucardi-N moralesi) (Y alta(N sallaei-N calientis))
Our results agree with previous analyses (Snelson1968) indicating that N shumardi is more closelyrelated to members of the subgenus Alburnops(N potteri and N blennius) than to the subgenus Not-ropis A later molecular studies proposed the removalof N shumardi from the subgenus Notropis (Bielawskiamp Gold 2001) Relationships among species analysedof the subgenus Notropis are in agreement with thislater study while the fourth clade is conformed by spe-cies of the N texanus species group which includeN volucellus from the N volucellus species group(Mayden 1991) (Fig 3)
Highest divergences among northern species of thegenus Notropis were found among N volucellus(N volucellus species group) and N potteri (subgenusAlburnops) N amabilis or N jemezanus (subgenusNotropis) (155 154 and 153 respectively) Thesedivergences were similar to those found among speciesfrom the genus Hybopsis with the genus Notropis(Table 2) or to those found among some Canadianand USA species of Notropis with the Mexican ones(117ndash163)
Mexican minnows clade show high genetic diver-gence among species which is also associated with ahigh degree of morphological differentiation For thisreason different authors have assigned these speciesto several genera (Aztecula Notropis Hybopsis andYuriria Chernoff amp Miller 1986 Mayden 1991Coburn amp Cavender 1992 Espinosa et al 1993)However the use of non-congruent criteria by theseauthors has led to the current controversial taxonomySeveral authors have considered the species examinedhere as members of distinct phylogenetic groups May-den (1989) ascribed N sallaei in a different clade withrespect to the other Mexican Notropis based on themorphology of the posterior myodome and considerN boucardi and N calientis (included in the genusHybopsis) in a different clade than the genus YuririaCoburn amp Cavender (1992) also based on morpholog-ical characters placed the genus Notropis (sl includ-ing genus Hybopsis) in the Shiner clade and the genusYuriria in the Western clade In contradiction withthese works our genetic analyses yielded topologies inwhich all Notropis species of central and southernMexico studied plus Yuriria form a monophyleticclade regardless of method of analyses with respectto the northern species of the genus Notropis and thegenus Hybopsis (including the type species of bothgenera N atherinoides and H amblops)
This strongly supported clade for the Mexican spe-cies containing different putative genera was unex-pected and suggest the need for a taxonomic revision
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 333
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
of this group taking into account their monophyleticorigin and high genetic divergences
Based on this study and pending a more extensivetaxonomic revision of the genus Notropis we adopt theconservative criterion of considering all Notropis spe-cies from southern and central Mexico examinedincluding Y alta being part of the genus Notropis(Table 3) Hence based on the phylogenetic relation-ships of the genus Yuriria which is included in allanalyses into the Mexican Notropis clade we proposeto change the taxonomic name Y alta to N altus(Jordan 1880) comb nov
SOUTHERN MEXICAN NOTROPIS CLADE
Four species of Notropis restricted to southern Mexi-can rivers represent the southern-most limits for Cyp-rinidae in the western hemisphere These speciesN boucardi N moralesi N imeldae and N sp 1 aredistributed across the Balsas Papaloapan and Atoyacdrainages Some authors (Miller amp Smith 1986 May-den 1989 1991 Espinosa et al 1993) included someor all the species of this clade in the genus HybopsisHowever Mayden (1989) cautioned that this groupingmay be the result of convergent evolution of morpho-logical characteristics arising from a similar benthiclifestyle Our genetic results agree with Maydenrsquoswarning on morphological convergence of N boucardiand N calientis with the genus Hybopsis since thefour southern Mexican species of Notropis(N moralesi N boucardi N imeldae and N sp 1)were resolved in the same clade as Yuriria N calientisand N sallaei suggesting a common ancestry Thisclade was equally distant to the type species of thegenus Hybopsis (H amblops) than it was to the typespecies of the genus Notropis (N atherinoides) How-ever our results disagree with Maydenrsquos (1989) con-clusion which placed these Mexican species indifferent clades
Analysis showed the populations of N moralesi and
N boucardi to be sister taxa (node A) N moralesi hasbeen considered restricted to the upper Papaloapanbasin and N boucardi was considered to have a widegeographical range including the Armeria Atoyac andBalsas basins (De Buen 1955 Espinosa et al 1993Schoumlnhuth et al 2001) However our results show ahigh level of differentiation (43ndash60) for the two pop-ulations from the high Balsas basin in Cuernavacathe type locality of N boucardi This divergence wasbigger than that found among N blennius andN potteri (39) and similar to that found amongN atherinoides with N jemezanus (63) or withN stilbius (62) Our study supports restriction ofN boucardi to upper balsas basing in the Cuernavacaarea whereas populations inhabiting the remainingBalsas Atoyac and upper Papaloapan drainages wereassignable to N moralesi
Notropis moralesi has a broad distribution thatspans three river drainages (Balsas Atoyac and Papal-oapan) but there was low sequence divergence amongpopulations Interestingly the most closely relatedpopulations of N moralesi inhabit the headwaters ofthese three different drainages of the Mixteca RegionThis species shows an intraspecific phylogeographicalpattern (Avise amp Walker 1999) rather than a drainagepattern where the specimens more closely related arethose which inhabit the Mixteca region independentlyof the river drainage in agreement with previous alloz-yme analyses (Schoumlnhuth et al 2001) that suggesteddispersal across drainages through headwater capturein the Mixteca region of Mexico
The N imeldae and N sp 1 clade (node B) showedmore similar genetic divergences with the centralMexican clade than they did with the remaining southMexican Notropis This clade (B) appeared as a mono-phyletic group with the N boucardi-N moralesi cladein MP ML and Bayesian analyses but seemed to bemore related to the central Mexican clade according toNJ analysis (Fig 4) However these two alternativetopologies were not statistically significantly different
Table 3 Traditional and recommended classification of Mexican species analysed
Original description Traditional classificationRecommended based oncyt b analyses
Notropis moralesi (De Buen 1955) Notropis moralesi1 or Hybopsis moralesi2 Notropis moralesiLeuciscus boucardi (Guumlnther 1868) Hybopsis boucardi12 N boucardiN imeldae (Corteacutes 1966) N imeldae1 or Hybopsis imeldae2 N imeldaeCeratichthys sallaei (Guumlnther 1868) N sallei1 or Aztecula sallei2 N sallaeiN calientis (Jordan amp Snyder 1900) N calientis1 or Hybopsis calientis2 N calientisHudsonius altus (Jordan 1880) Hybopsis altus3 or Yuriria alta12 N altus
1Espinosa et al (1993)2Mayden (1991)3Bailey (1951)
334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

334 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
from each other We consider the first hypotheses to bemore plausible taking into account that N imeldaeN boucardi and N moralesi have been traditionallyconsidered to be morphologically closely related(Corteacutes 1966) Furthermore previous allozyme dataalso support N imeldae and N sp 1 to be more closelyrelated to N boucardi-N moralesi than to N sallaei(Schoumlnhuth et al 2001)
CENTRAL MEXICAN MINNOW CLADE
This clade was formed by N sallaei N calientis andY alta all three species inhabiting rivers of centralMexico over the trans-Mexican volcanic belt In ourstudy all the analyses showed N sallaei andN calientis to be sister species This finding contrastswith Maydenrsquos (1989) phylogenetic study of NorthAmerican minnows In this work based on osteologi-cal variation of a large proportion of cyprinidsN sallaei was excluded from a large monophyleticgroup that shared the open posterior myodome (OPM)as a derived character However Mayden includedother Mexican cyprinids (N boucardi N calientis andY alta) in the OPM group but in different clades In arecent mitochondrial DNA study some taxa which donot exhibit the OPM and previously were not includedin the OPM clade defined by Mayden (1989) wereincluded (Simons amp Mayden 1999) These authorssuggest that the OPM may have evolved from a com-mon ancestor of the OPM clade being subsequentlylost in certain taxa Our results resolving N sallaei inthis clade also provide evidence for the presence of ahigh level of homoplasy in this character and thereforeits limited use for phylogenetic analyses
Our results resolving N calientis and N sallaei assister species (node C) contradict previous taxonomichypotheses that assigned N calientis to the genusHybopsis and N sallaei to the monotypic genus Aztec-ula (Mayden 1989 and 1991 Coburn amp Cavender1992)
Notropis sallaei did not show high genetic diver-gence among populations from the Paacutenuco and Lermadrainages These populations were previously consid-ered as distinct species (Aztecula mexicana (Meek1902) and N lermae (Everman amp Goldsborough1902) respectively) until Chernoff amp Miller (1981) rec-ognized N sallaei as a single highly variable taxonOur genetic analyses agree with this morphologicalwork The little sequence divergence among popula-tions from this species in the Paacutenuco (Atlantic coast)and Lerma (Pacific coast) drainages suggest possibleheadwater stream captures among both headwaterdrainages previously hypothesized for other fish dis-tributions (Barbour 1973 Miller amp Smith 1986Weeb 1998 Mateos Sanjur amp Vrijenhoek 2002)
Within the N calientis clade we identified three
genetically distinct lineages The most related speci-mens were those from the Lerma drainage andCuitzeo lagoon populations and the most divergentpopulations of the clade corresponded to those fromthe Zacapu lagoon and the San Miguel spring Inter-estingly sequence divergence between these three lin-eages (56ndash70) was similar to that between otherspecies of genus Notropis examined here (N boucardi-N moralesi 43ndash60 (dA 39) N imeldae-N sp 171 (dA 67) N atherinoides-N amabilis 73) Dif-ferentiation between N calientis populations wasrecognized by Chernoff amp Miller (1986) who describedthem as a complex containing three speciesN calientis (Paacutenuco headwaters and Lerma-Santiagodrainage) N aulidion (Tunal-Mesquital headwater)and N amecae (Ameca drainage) These last two spe-cies are currently though to be extinct (MillerWilliams amp Williams 1989 Mayden 1991) All popu-lations analysed here corresponded to the distributionrange of the species N calientis of the complex asdefined by Chernoff amp Miller (1986) not showing diag-nostic morphological characters of N aulidion orN amecae and being biogeographically distant fromtheir distribution area
Despite the fact that 90 of the putative sister spe-cies of vertebrates show mtDNA sequence divergencesgreater than 2 in the cytochrome b gene (Johns ampAvise 1998) 56 of the species could be subdividedinto at least two major intraspecific phylogroups(Avise amp Walker 1999) These phylogroups are envis-aged as independent evolving historical lineagesequivalent to taxonomic species except for the magni-tude of divergence In our case the specimens of thethree lineages of N calientis analysed occur in geo-graphically proximate areas Hendry et al (2000) indi-cated that there is no clear separation for mtDNAdivergence that distinguishes species from phylo-groups and that phylogroups identified in genetic sur-veys are often later interpreted as cryptic species Thehigh genetic divergence found among N calientis lin-eages may reflect the long isolation of the Zacapulagoon and San Miguel spring or a high rate of evo-lution (Schoumlnhuth 2002) In both places N calientis iscurrently the only cyprinid that could have evolved invicariance as distinct genetic types Our analyses sug-gest that the three lineages analysed of N calientisinhabiting Riacuteo Lerma Zacapu Lagoon and San MiguelSpring (all from the Lerma drainage) may form threedifferent species Chernoff amp Miller (1986) alsodetected small meristic and morphological differencesamong populations of N calientis from the Lerma-Santiago drainage but they suggest intraspecificvariation due to geographical differentiation To clar-ify this point further studies are required
In all analyses N altus (Y alta) was resolved in theMexican Notropis clade Net sequence divergence
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 335
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
between N altus and the remainder of the MexicanNotropis examined (dA 775ndash1135) was within thenet sequence divergence range observed among theMexican Notropis species studied (dA 395ndash1184)and within the sequence divergence observed amongthe northern species of the genus Notropis (39ndash155) Coburn amp Cavender (1992) hypothesized thegenus Yuriria in a large polytomy at the base of thelsquowesternrsquo clade of North American cyprinids comprisedof another 20 genera being plesiomorphic for mostskeletal and scale characters to other cyprinids of theMexican Mesa Central whereas the genus Notropissl was included in a different clade (lsquoShinerrsquo) Thephylogenetic position of N altus (Y alta) with respectto the remaining taxa analysed placing it within theMexican Notropis clade and suggesting close relation-ships to Notropis of central Mexico has interestingimplications with regard to taxonomy and contrastswith previous proposed relationships (Mayden 1989Coburn amp Cavender 1992) Phylogenetic relationshipswith other Mexican cyprinids such as the genusAlgansea or species of the genus Cyprinella and espe-cially the genus Dionda must be tested since previouswork suggests a close relationship of the Paacutenucodrainage species of the genus Dionda with the Mexi-can species of the genus Notropis (Mayden 1989Coburn amp Cavender 1992) However our preliminaryresults (S Schoumlnhuth amp I Doadrio unpubl data)seem to indicate that the genus Dionda is not closelyrelated with these Mexican species of the genus Not-ropis Our results suggest that the NotropisHybopsisclade is more closely related to Pteronotropis than it isto Cyprinella as has been previously supported bymorphological studies in which NotropisHybopsisplus Pteronotropis and Cyprinella are included in twodifferent large lineages in the Notropin clade of theShiner clade (Coburn amp Cavender 1992)
We hypothesize that ancestors of the genera Notro-pis and Hybopsis probably underwent rapid speciationin North America resulting in several species groupsand subgenera with high divergences among them Arapid diversification in the evolution of the NotropisHybopsis clade is suggested by the weak support forthe deep branches of the five clades The lack of basalresolution in the NotropisHybopsis phylogeny sug-gests this rapid speciation event in the past or as afunction of sampling The basal polytomy reflects alack of resolution among major groups of the genusNotropis but indicates well-supported clade groupingendemic Mexican species previously placed in distinctclades The Mexican Notropis analysed constitute thesouthern limit of the entire family Cyprinidae in thewestern hemisphere with all relatives living to thenorth (Schoumlnhuth 2002) Our finding of a monophyl-etic clade grouping central and southern Mexican Not-ropis with respect to northern species suggests a
single colonization event with southward dispersaland posterior processes of speciation Furthermoreour results indicate the existence of more species thanpreviously recognized in central and southern Mexico
ACKNOWLEDGEMENTS
We thank Adolfo De-Sostoa Emilio Martiacutenez RaulPineda Humberto Mojica Topiltzin Contreras andPaloma Garzoacuten for their help with the sampling ofspecies Specimens from Canada and N calientis fromGuanajuato (Mexico) were kindly provided by ErlingHolm curator of The Royal Ontario Museum (Can-ada) and Omar Dominguez from the Universidad deMorelia (Mexico) respectively We also thank AnnieMachordom Anabel Perdices Mario Garciacutea-ParisLukas Ruumlber Soraya Villalva and Rafael Zardoya forinsightful comments on the manuscript We especiallyacknowledge Rafael Zardoya for his help with the lab-oratory work Lourdes Alcaraacutez and Antonia Montillaprovided laboratory assistance This study was fundedby the project REN 2001ndash0662GLO
REFERENCES
Avise JC Walker D 1999 Species realities and numbers insexual vertebrates Perspectives from an asexual transmit-ted genome Proceedings of the National Academy of Sci-ences USA 96 992ndash995
Bailey RM 1951 A check list of the fishes of Iowa with keysfor identification In Harlan JR Speaker EB eds Iowa fishand fishing 4th edn Iowa Iowa Conservation Commission1969 327ndash377
Barbour CD 1973 A biogeographical history of Chirostoma(Pisces Atherinidae) a species flock from the Mexican Pla-teau Copeia 3 533ndash556
Bermingham E Martin AP 1998 Comparative mtDNAphylogeography of neotropical freshwater fishes testingshared history to infer the evolutionary landscape of lowerCentral America Molecular Ecology 7 499ndash457
Bielawski JP Gold JR 2001 Phylogenetic relationships ofcyprinid fishes in subgenus Notropis inferred from nucle-otide sequences of the mitochondrially encoded cytochrome bgene Copeia 3 656ndash667
Burr BM Mayden RL 1981 Systematic distribution andlife history notes on Notropis chihuahua (Pisces Cyprin-idae) Copeia 2 255ndash265
Buth DG 1979 Biochemical systematic of the cyprinid genusNotropis-I The subgenus Luxilus Biochemical Systematicsand Ecology 7 69ndash79
Chernoff B Miller RR 1981 Systematics and variation ofthe Aztec shiner Notropis sallei a cyprinid fish from CentralMexico Proceedings of the Biological Society of Washington94 18ndash36
Chernoff B Miller RR 1986 Fishes of the Notropis calientiscomplex with a key to the Southern Shiners of MexicoCopeia 1 170ndash183
336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

336 S SCHOumlNHUTH and I DOADRIO
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Coburn MM Cavender TM 1992 Interrelationships ofNorth American cyprinid fishes In Mayden RL ed Sys-tematics historical ecology and North American freshwa-ter fishes Stanford CA Stanford University Press 328ndash373
Corteacutes MT 1966 Consideraciones sobre el geacutenero Notropis ydescripcioacuten de una especie nueva procedente del riacuteo Atoyacen Juchatengo Oaxaca Meacutexico [Considerations of the genusNotropis and description of a new species from the Atoyacriver in Juchatengo Oaxaca Mexico] Anales de la EscuelaNacional de cencias Biologicas 15 185ndash192 (In Spanish)
De Buen F 1955 El primer representante (Notropismoralesi nov sp) de la familia Cyprinidae en la fauna neo-tropical [First record (Notropis moralesi nov sp) of theCyprinidae family in the neotropical fauna] Annales delInstituto de Biologia Mexico XXVI 527ndash541 (In Spanish)
Dimmick WW 1987 Phylogenetic relationships of Notropishubbsi N welaka and N emiliae (Cypriniformes Cyprin-idae) Copeia 2 316ndash325
Dowling TE Naylor GJP 1997 Evolutionary relationshipsof minnows in the genus Luxilus (Teleostei Cyprinidae) asdetermined from cytochrome b sequences Copeia 1997 758ndash765
Espinosa H Gaspar MT Fuentes P 1993 Listados fauniacutes-ticos de Meacutexico III Los peces dulceacuiacutecolas mexicanos[Catalogue of fauna from Mexico III Mexican freshwaterfishes] Argumedo RL ed Mexico Instituto de Biologiacutea Uni-versidad Nacional Autoacutenoma de Meacutexico (In Spanish)
Felsenstein J 1985 Confidence limits on phylogenies Anapproach using the bootstrap Evolution 39 783ndash791
Hendry PA Vamosi SM Latham SJ Heilbuth JC Day T2000 Questioning species realities Conservation Genetics 167ndash76
Huelsenbeck JP Ronquist FR 2001 MR BAYES Bayesianinference of phylogeny Bioinformatics 17 754ndash755
Johns GC Avise JC 1998 A comparative summary ofgenetic distances in the vertebrates from the mitochondrialcytochrome b gene Molecular Biology and Evolution 151481ndash1490
Machordom A Doadrio I 2001 Evidence of a CenozoicBetic-Kabilian connection based on freshwater fish phylo-geography (Luciobarbus Cyprinidae) Molecular Phylogenet-ics and Evolution 18 252ndash263
Martin AP Bermingham E 1998 Systematics and evolu-tion of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences Molecular Phylo-genetics and Evolution 9 192ndash203
Mateos M Sanjur OI Vrijenhoek RC 2002 Historical bio-geography of the livebearing fish genus Poeciliopsis (Poecil-iidae Cyprinodontiformes) Evolution 56 972ndash984
Mayden RL 1989 Phylogenetics studies of North Americanminnows with emphasis on the genus Cyprinella (TeleosteiCypriniformes) In Mengel RM Johnston RF eds KansasThe University of Kansas Publications Museum of NaturalHistory Miscellaneous publication 80 1ndash189
Mayden RL 1991 Cyprinids of the new world In Winfield IJNelson JS eds Cyprinid fishes systematics biology andexploitation London Chapman amp Hall 240ndash263
Miller RS Smith ML 1986 Origin and geography of thefishes of central Mexico In Hocutt CH Wiley EO eds Thezoogeography of North American freshwater fishes NewYork Wiley-Interscience Publications 487ndash519
Miller RR Williams DJ Williams JE 1989 Extinctions ofNorth American fishes during the past century Fisheries 1422ndash37
Nei M 1987 DNA polymorphism within and between popula-tions In Molecular evolutionary genetics NY ColumbiaUniversity Press 254ndash286
Nei M Li W-H 1979 Mathematical model for studying geneticvariation in terms of restriction endonucleases Proceedingsof the National Academy of Sciences USA 76 5269ndash5273
Posada D Crandall KA 1998 Modeltest Testing the modelsof DNA substitution Bioinformatics 14 817ndash818
Raley ME Wood RM 2001 Molecular systematics of mem-bers of the Notropis dorsalis species group (ActinopterygiiCyprinidae) Copeia 3 683ndash645
Rozas J Rozas R 1999 DnaSP Vers 3 an integrated pro-gram for molecular population genetics and molecular evo-lution analysis Bioinformatics 15 174ndash175
Sambrook J Fritsch EF Maniatis T 1989 Molecular clon-ing a laboratory manual 2nd edn Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Schoumlnhuth S 2002 Sistemaacutetica filogenia e implicacionesbiogeograacuteficas de las poblaciones del geacutenero NotropisRafinesque 1817 (Actinopterygii Cyprinidae) en el centro ysur de Meacutexico [Systematics phylogenetic relationships andbiogeographical implications of the Mexican minnowrsquos popu-lations of the genus Notropis Rafinesque 1817 (Actinoptery-gii Cyprinidae) in central and south Mexico] PhD ThesisUniversidad Complutense de Madrid Spain (In Spanish)
Schoumlnhuth S De-Sostoa A Martiacutenez E Doadrio I 2001Southern Mexican minnows of the genus Notropis (Actinop-terygii Cyprinidae) Genetic variation phylogenetic rela-tionships and biogeographical implications BiochemicalSystematics and Ecology 29 359ndash377
Shimodaira AM Hasegawa M 1999 CONSEL For assess-ing the confidence of phylogenetic tree selection Bioinfor-matics 17 1246ndash1247
Simons AM Mayden RL 1997 Phylogenetic relationships ofthe creek chubs and the spine-fins an enigmatic group ofNorth American Cyprinid fishes (Actinopterygii Cyprin-idae) Cladistics 13 187ndash205
Simons AM Mayden RL 1998 Phylogenetic relationships ofthe western North American Phoxinins (Actinopterygii Cyp-rinidae) as inferred from mitochondrial 12S and 16S riboso-mal RNA sequences Molecular Phylogenetics and Evolution9 308ndash329
Simons AM Mayden RL 1999 Phylogenetic relationships ofNorth American Cyprinids and assessment of homology ofthe Open Posterior Myodome Copeia 1 13ndash21
Snelson FF 1968 Systematics of the cyprinid fish Notropisamoenus with comments on the subgenus Notropis Ibid1968 440ndash442
Stein DW Rogers JS Cashner C 1985 Biochemical sys-tematics of the Notropis roseinipis Complex (Cyprinidaesubgenus Lythrurus) Copeia 1 154ndash163
PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237

PHYLOGENETIC RELATIONSHIPS OF NOTROPIS IN MEXICO 337
copy 2003 The Linnean Society of London Biological Journal of the Linnean Society 2003 80 323ndash337
Suzuki Y Glazco G Nei M 2002 Overcredibility of molec-ular phylogenies obtained by Bayesian phylogenetics Pro-ceedings of the National Academy of Sciences USA 9916138ndash16143
Swofford DL 2001 PAUP phylogenetic analysis using par-simony (and other methods) Version 4 0b8 SunderlandMA Sinauer Associates
Swofford DL Waddell PJ Huesenbeck JP Foster PGLewis PO Rogers JS 2001 Bias in phylogenetic estima-tion and its relevance to choice between parsimony and like-lihood methods Systematic Biology 50 525ndash539
Vallejo de Aquino P 1988 Anaacutelisis inmuno-taxonoacutemico deN sallaei y N lermae (Pisces Cyprinidae) [Immunotaxo-nomical analyses of Notropis sallaei and N lermae (PiscesCyprinidae)] Thesis Instituto Politeacutecnico Nacional MeacutexicoDF (In Spanish)
Warren ML Burr BM Grady JM 1994 Notropis albizona-
tus a new cyprinid fish endemic to the Tennessee and Clu-berland river drainages with a phylogeny of the Notropisprocne species group Copeia 4 868ndash886
Weeb SA 1998 A phylogenetic analysis of the Goodeidae(Teleostei Cyprinodontiformes) Unpublished PhD ThesisUniversity of Michigan Ann Arbor
Wilcox TP Zwickl DJ Heath TA Hillis DM 2002Phylogenetic relationships of the boas and a comparison ofBayesian and bootstrap measures of phylogenetic supportMolecular Phylogenetics and Evolution 25 361ndash371
Zardoya R Doadrio I 1998 Phylogenetic relationships ofIberian cyprinids systematic and biogeographical implica-tions Proceedings of the Royal Society of London 265 1365ndash1372
Zardoya R Doadrio I 1999 Molecular evidence on the evo-lutionary and biogeographical patterns of European cyprin-ids Journal of Molecular Evolution 49 227ndash237





















