
A new species of Anisopappus (Asteraceae, Inuleae) and its phylogenetic relationships within the...
8
Nordic Journal of Botany A new species of Anisopappus (Asteraceae, Inuleae) and its phylogenetic relationships within the genus S. Ortiz, J. A. R. Paiva and I. Pulgar Ortiz, S., Paiva, J. A. R. & Pulgar, I. 2001. A new species of Anisopappus (Asteraceae, Inuleae) and its phylogenetic relationships within the genus. - Nord. J. Bot. 21: 55-62. Copenhagen. ISSN 0107-055X. A new species of the genus Anisopappus (Asteraceae, Inuleae) from Angola, A. fruticosus, is described. The principal differences with respect to A. chinensis are discussed, and phylogenetic relationships with the other species of the genus are investigated by means of a cladistic analysis using morphological characters. S. Ortiz, Laboratorio de Botanica, Facultade de Farmacia, Universidade de San- tiago, 15706 Santiago de Compostela. Galicia-Spain. - 1 A. R. Paiva, Depar- tamento de Botcinica, Faculdade de Citncias e Tecnologia. Universidade de Coimbra, 3049 Coimbra, Portugal. - I. Pulgar. Laboratorio de Botanica. Facultade de Farmacia. Universidade de Santiago. I5706 Suntiago de Compostela. Galicia- Spain. Introduction Anisopappus Hook. & Am. is a genus traditionally included in the tribe Inuleae (Asteraceae) (Hoffmann 1890; Wild 1964; Anderberg 1991; Eldenas & Anderberg 1996; Eldenas et al. 1998), though recently Eldenas et al. (1999) concluded that this genus is more closely related to the tribe Heliantheae s. 1. The distribution area of Anisopappus covers southern Asia (where only Anisopappus chinensis Hook. & Am. is present), tropical and southern Africa, and Madagascar. In his revision of the genus, Wild (1964) accepted 29 species. More recently, 10 new species have been described (Lawalrke 1974; Lisowski 1986a, 1986b, 1988; Ortiz & Paiva 1995), and Eldenas & Anderberg (1996) accepted about 40 species. However, in our synopsis of the genus (Ortiz et al. 1996) we accepted only 17 species, 10 subspecies and 4 varieties in view of our conclusion that pappus morphology has limited diagnostic value. Many previous workers (Humbert 1923; Wild 1964; Lisowski 1986a, 1986b, 1988) have used pappus morphology as a diagnostic character, and indeed several species of Anisopappus have been described entirely or almost entirely on the basis of pappus morphology alone. Following recent studies on Anisopappus, we describe a new species from Angola and analyse the phylogenetic relationships between this species and the other species of the genus. Materials and methods Description of the new species Anisopappus fmti- cosus is based on the morphological study of ma- terial from the herbaria of COI, K, LISC, LUAI, and PRE (abbreviations based on Holmgren et al. 1990). Styles and cypselas of dried material were moun- ted on aluminium stubs, and coated with a c. 30 nm layer of Au for study with a scanning electron microscope (SEM) (LEO 435VP), operating at 15 kV. The phylogenetic analysis of Anisopappus, for investigation of phylogenetic relationships between Accepted 25-01-2001 Nord. J. Bot. 21(2)2001 55
Transcript of A new species of Anisopappus (Asteraceae, Inuleae) and its phylogenetic relationships within the...

Nordic Journal of Botany
A new species of Anisopappus (Asteraceae, Inuleae) and its phylogenetic relationships within the genus
S. Ortiz, J. A. R. Paiva and I. Pulgar
Ortiz, S., Paiva, J. A. R. & Pulgar, I. 2001. A new species of Anisopappus (Asteraceae, Inuleae) and its phylogenetic relationships within the genus. - Nord. J. Bot. 21: 55-62. Copenhagen. ISSN 0107-055X.
A new species of the genus Anisopappus (Asteraceae, Inuleae) from Angola, A . fruticosus, is described. The principal differences with respect to A . chinensis are discussed, and phylogenetic relationships with the other species of the genus are investigated by means of a cladistic analysis using morphological characters.
S. Ortiz, Laboratorio de Botanica, Facultade de Farmacia, Universidade de San- tiago, 15706 Santiago de Compostela. Galicia-Spain. - 1 A . R. Paiva, Depar- tamento de Botcinica, Faculdade de Citncias e Tecnologia. Universidade de Coimbra, 3049 Coimbra, Portugal. - I. Pulgar. Laboratorio de Botanica. Facultade de Farmacia. Universidade de Santiago. I5706 Suntiago de Compostela. Galicia- Spain.
Introduction
Anisopappus Hook. & Am. is a genus traditionally included in the tribe Inuleae (Asteraceae) (Hoffmann 1890; Wild 1964; Anderberg 1991; Eldenas & Anderberg 1996; Eldenas et al. 1998), though recently Eldenas et al. (1999) concluded that this genus is more closely related to the tribe Heliantheae s. 1. The distribution area of Anisopappus covers southern Asia (where only Anisopappus chinensis Hook. & Am. is present), tropical and southern Africa, and Madagascar. In his revision of the genus, Wild (1964) accepted 29 species. More recently, 10 new species have been described (Lawalrke 1974; Lisowski 1986a, 1986b, 1988; Ortiz & Paiva 1995), and Eldenas & Anderberg (1996) accepted about 40 species. However, in our synopsis of the genus (Ortiz et al. 1996) we accepted only 17 species, 10 subspecies and 4 varieties in view of our conclusion that pappus morphology has limited diagnostic value. Many previous workers (Humbert 1923; Wild 1964; Lisowski 1986a, 1986b, 1988) have used
pappus morphology as a diagnostic character, and indeed several species of Anisopappus have been described entirely or almost entirely on the basis of pappus morphology alone.
Following recent studies on Anisopappus, we describe a new species from Angola and analyse the phylogenetic relationships between this species and the other species of the genus.
Materials and methods
Description of the new species Anisopappus fmti- cosus is based on the morphological study of ma- terial from the herbaria of COI, K, LISC, LUAI, and PRE (abbreviations based on Holmgren et al. 1990).
Styles and cypselas of dried material were moun- ted on aluminium stubs, and coated with a c. 30 nm layer of Au for study with a scanning electron microscope (SEM) (LEO 435VP), operating at 15 kV.
The phylogenetic analysis of Anisopappus, for investigation of phylogenetic relationships between
Accepted 25-01-2001
Nord. J. Bot. 21(2)2001 5 5

A. fmticosus and the other species of the genus, was (considered a synonym of A. davyi S. Moore), A. based on the morphological analysis of Eldenas & burundensis Lisowski (considered a synonym of A. Anderberg (l996), though with some modifications. ubercornensis G. Taylor), and A. buchwaldii (0. In accordance with our synopsis of the genus (Ortiz Hoffm.) Wild, A. oliverunus Wild, A. discolor Wild, et al. 1996), we removed A. bumpsianus Lisowski and A. scrophularifolius (Baker) Wild (considered
Table 1. Characters and states used in phylogenetic analysis.
1. 2.
3. 4. 5. 6. 7. 8. 9.
10.
11. 12. 13. 14. 15. 16.
17. 18. 19. 20. 21. 22.
23. 24. 25. 26. 27. 28. 29. 30. 31. 32.
33. 34. 35. 36. 37. 38.
Habit herbaceous (0). Habit woody (1). Anisopuppus-type septate hair (0). Asteriscus-type septate hair (I) . A. pinnatifdus-type septate hair (2) (see Eldenas & Anderberg, 1996). Wooly hair present (0). Absent (1). Stems without wings (0). Stems with more or less prominent decurrent wings (1). Petioles minute or completely absent (0). Petioles long, distinct (1). Leaves lobed to pinnatipartite (0). Leaves simple, undivided (1). Leaf venation palmate (0). Leaf venation pinnate (1). Veining costate (2). Leaf main veins distintc, veinlets diffuse (0). The entire network of veins is sunken (1). Leaves toothed (0). Leaves entire (1). Capitula solitary on long branches (0). Few capitula together in loose corymbs (1). Capitula numerous, in more or less dense corymbs (2). Capitula campanulate to hemispheric (0). Capitula obconic to cyathiform (1). Receptacle more or less flat (0). Receptacle hemisphaerical or conical (1). Receptacle narrowly conical (2). Receptacle epaleate (0). Receptacle paleate (1). Staminodia present in ray-florets (0). Staminodia absent in ray-florets (1). Disc-floret corolla-lobes short (I 0.5 mm long) (0). Disc-floret corolla-lobes long (> 0.5 mm long) (1). Disc-floret corolla-lobes without sclerenchymatous cells accompanying the vascular strands (0). With more or less narrow, submarginal sclerenchyme (1). With broad, marginal sclerenchymatous bands (2). Disc-floret corolla without bristly hairs (0). Disc-floret corolla with bristly hairs (I). Endothecial tissue with radial thickenings (0). Endothecial tissue with polarized thickenings (1). Filament collar cells longer than wide (0). Filament collar cells wider than long (1). Anther thecae long calcarate (0). Anther thecae shortly calcarate (1). Anther theca ecalcarate (2). Anther tails minute (0). Anther tails long ( I ) . Anther tails completely unbranched (0). Anther tails with short branches (I) . Anther tails with long branches (2). Style with crystals rectangular-prismatic or without crystal deposits (0). Style with crystals star-shaped (1). Style branch apex obtuse, rounded (0). Style branch apex acute (1). Style branch vein narrow (0). Style branch vein thick (1). Sweeping-hairs acute (0). Sweeping-hairs obtuse (1). Cypsela straight (0). Cypsela curved (1). Cypsela yellowish-brown (0). Cypsela dark reddish-brown (1). Cypsela trilocular (0). Cypsela more or less terete (I). Cypsela triquetrous (2). Cypsela with sclerenchymatic tissue (0). Cypsela without sclerenchymatic tissue (1). Cypsela surface smooth (0). Cypsela surface with distinctly raised ribs (1). Cypsela epidermis without crystals (0). Cypsela with one elongated crystal in each epidermal cell (2). Crystals or crystal fragments in parenchyma (2). Epidermal cells smooth (1). Epidermal cells papillose ( I ) . Epidermal cells equal in size to subepidermal cells (0). Epidermal cells enlarged (I). Carpopodium absent (0). Carpopodium present (1). Pappus bristles absent (0). Pappus bristles present (1). Pappus scales present (0). Pappus scales absent (1). Pappus scales larger, regular (0). Pappus scales large, serrate, some with elongate midportion (I). Pappus scales irregular, serrate (2). Pappus scales reduced into a minute rim (3).
56 Nord. J. Bor. 21(2) 2001
synonyms of A . chinensis). However, we kept A. rhombifolius Wild, a species included by us (Ortiz et al. 1996) in the A. chinensis complex, but probably different in accordance with the data hrnished by Eldenas & Anderberg (1 996) principally regarding the type of cypsela hairs, unique in the genus. We also included A . longipes (Comm. ex Cass.) Wild, a prostrate endemic species from Madagascar probably related to A . chinenis, and A . pseudopinnatifdus, a species described by us (Ortiz & Paiva 1995) from Namibia; neither species was included by Eldenas & Anderberg (1996) in their phylogenetic analysis. As regards Anisopappus athanasioides S. Ortiz & Paiva, a species described from somewhat immature material (Ortiz & Paiva 1995), we agree with Eldenas & Anderberg (1996: 174), who consider that the specimen from which it was described is “sig- nificantly different from all other species of Ani- sopappus”. We have subsequently found more com- plete material of this taxon, indicating that besides
its distinct capitula and inflorescences it has mature ray-florets very different to those of Anisopappus. This species was therefore not included in the present analysis.
In relation to these modifications we coded ‘-‘ instead of 2 for character 35 (pappus scale shape) of A . abercornensis and A . chinensis since it is variable in the different synonyms listed above.
Like Eldenas & Anderberg (1996), we included six other genera of the Inuleae in the analysis to assess whether Anisopappus is monophyletic. These genera were Asteriscus Tourn. ex Mill., Buphthal- mum L., Calostephane Benth., Geigeria Griess., Inu- la L., and Pulicaria Gaertn. The former four genera are representatives of the clades most closely related to Anisopappus, which is basal within the tribe, whereas Inula and Pulicaria are representatives of the most evolved clade (see Anderberg 1991).
A total of 23 taxa and 38 morphological charac- ters were studied (Tables 1 and 2). Polarization of
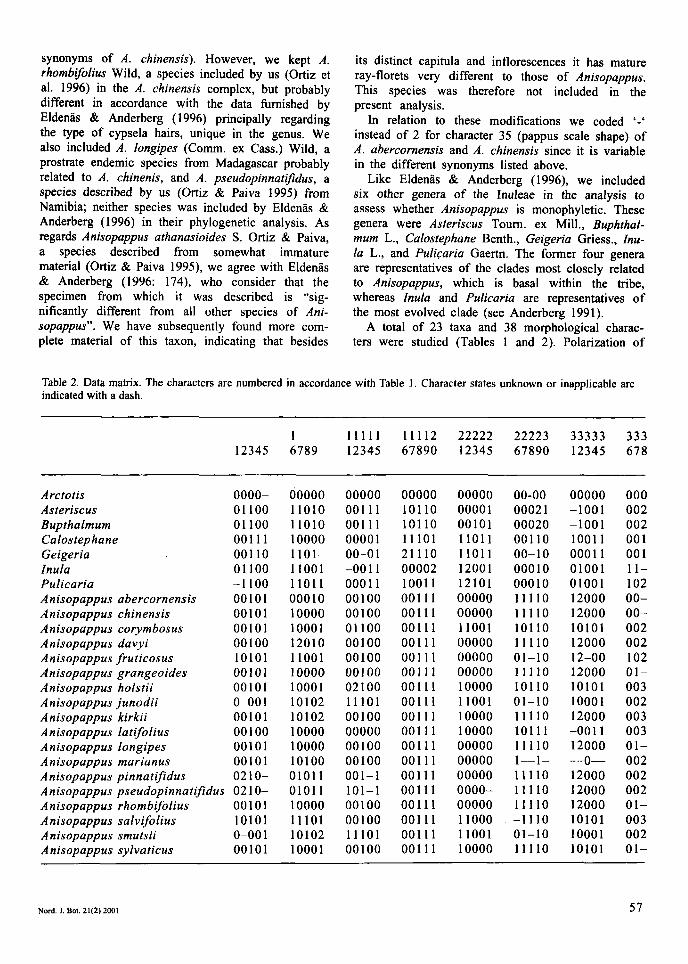
Table 2. Data matrix. The characters are numbered in accordance with Table 1 . Character states unknown or inapplicable are indicated with a dash.
1 11111 11112 22222 22223 33333 333 12345 6789 12345 67890 1234.5 67890 12345 678
Arctotis Asteriscus Bupthalmum Calostephane Geigeria Inula Pulicaria Anisopappus abercornensis Anisopappus chinensis Anisopappus corymbosus Anisopappus davyi Anisopappus fruticosus Anisopappus grangeoides Anisopappus holstii Anisopappus junodii Anisopappus kirkii Anisopappus latifolius Anisopappus longipes Anisopappus marianus Anisopappus pinnati’dus Anisopappus pseudopinnatifdus Anisopappus rhombifolius Anisopappus salvifolius Anisopappus smutsii Anisopappus sylvaticus
0000- 01 100 01 100 001 11 001 10 01 100 -1 100 00101 00101 00101 00 100 10101 00101 00101 o-oo 1 00101 00 100 00101 00101 02 10- 02 10- 00101 10101 o-oo 1 00101
00000 11010 11010 10000 1101- 11001 11011 000 10 10000 1000 1 12010 11001 10000 1000 1 10102 10102 10000 10000 10100 0101 1 0101 1 10000 1 1 101 10102 1000 1
00000 00000 00111 10110 00111 10110 00001 11101 00-01 21110 -0011 00002 00011 10011 00100 00111 00100 00111 01100 00111 00100 00111 00100 00111 00100 00111 02100 00111 11101 00111 00100 00111 00000 00111 00100 00111 00100 00111 001-1 00111 101-1 00111 00100 00111 00100 00111 11101 00111 00100 00111
00000 00001 00101 11011 1101 1 12001 12101 00000 00000 11001 00000 00000 00000 10000 11001 10000 10000 00000 00000 00000 0000- 00000 11000 11001 10000
00-00 0002 1 00020 00110 00- 10 000 10 000 10 11110 11110 101 10 11110 01-10 11110 101 10 01-10 11110 101 11 11110 1--1- 11110 11110 11110 -1 110 01-10 11110
00000 -1001 -1001 1001 1 0001 1 01001 01001 12000 12000 10101 12000 12-00 12000 10101 1000 1 12000 -001 1 12000 --o-- 12000 12000 12000 10101 1000 1 10101
000 002 002 00 1 00 1 11- 102 00- 00- 002 002 102 01- 003 002 003 003 01- 002 002 002 01- 003 002 01-
Nord. 1. Bot. 21(2) 2001 57

Fig. 1. Anisopappus fruticosus. - A. habit. - B. capitulum. - C. disc (left) and ray floret (right). - D. leaf. - E. phyllary (left) and receptacular palea (right). - F. cypsela. - Based on holotype (Santos & Henriques 1146, LISC).
58 Nord. J . BOI. ZI(2) 2001

characters was determined by the outgroup compa- rison method with Arctotis L. as outgroup, as in the cladistic analysis of Eldenas & Anderberg (1996).
For microscopic examination of floral parts, these were first boiled in water with a surfactant, then mounted in Hoyer’s solution (Anderson 1954).
Wagner parsimony analysis of the data matrix (Table 2) was performed on a PC with the aid of the program HENNIG86 (Farris 1988). We used the multiple hennig option (mh*, for constructing seve- ral initial cladograms by adding taxa in several diffe- rent sequences, then retaining the shortest) followed by the branch-breaker option (bb*, for generating multiple equally parsimonious cladograms). A suc- cessive weighting procedure (Farris 1969) was used to generate cladograms in which the relative weight of homoplasious characters was reduced. All mul- tistate characters were treated as nonadditive. Cha- racter state unknown or inapplicable was indicated with a dash (-) in the matrix.
Besides the addition of the 2 species, we added 3 new characters to the 35 used by Eldenas & Anderberg (1996): characteres 1, 11, and 15 of Table 1. As regards character I , it should be pointed out that on the label of the type of A. fruticosus it is described as suffrutex, but from the texture of its branches there is no doubt that it should be con- sidered woody.
Results and discussion
Anisopappus fruticosus S. Ortiz & Paiva sp. nov.
Frutex ut plurimum 80 cm altus. Folia elliptica, margine serrata, petiolis (5-)lo- 13 mm longis. Capitula dense corymbosa, pedunculis brevibus, 15- 25(-35) mm longis. Flores radii ut plurimum 14 x 3 mm. Cypsela squamosa et setis 3-5(-7) praedita, barbellatis, (0.5-)0.8-1(-1.5) mm longis. - Fig. 1.
Typus: Angola, Huila, Lubango (Sa da Bandeira), Fenda de Tunda-Vala, high plateau. Rocky outcrops. 4.vii.1965. Santos & Henriques 1146 (holotype: LISC, isotypes: K, LUAI, PRE).
Shrub 70-80 cm high. Branches striate, with dense pilosity of spreading, multicellular, Anisopappus- type hairs (see Eldenas & Anderberg, 1996), and sessile to subsessile yellow glands; internodes 4-8 mm long. Leaves 40-55 x 13-20 mm, elliptic to elliptic-lanceolate; lamina (30-)35-45 mm long, ve- nation pinnate; petiole (5) 10-13 mm long; margins serrate, with subobtuse teeth 1.3-1 3 - 3 ) mm long;
base cuneate; apex acute; both surfaces greenish, with a moderate to dense pilosity of pluricellular hairs like those of the stem, and sessile yellow glands, these conspicuously more abundant on the adaxial surface. Capitula numerous, (3-)6-12 arran- ged in corymbiform synflorescences, on erect-patent peduncles 15-25(-35) mm long, radiate, heteroga- mous. Involucre 10-13 x 4-5 mm, campanulate- hemispheric, with c. 40-60 phyllaries arranged in 3(- 4) rows, greeenish, with multicellular hairs like those of the stem, the margins entire; outermost phyllaries 3-4 x 0.5-1 mm, linear; middle phyllaries 4-5 x 1.3- 1.5 mm, lanceolate, with apex acute to acuminate, slightly scarious; innermost phyllaries (4-)5-5.5 x 1.3-1.5 mm, lanceolate, with apex acute to acu- minate; receptacle flat; receptacular paleae 3.5-4(- 4.5) x 0.5-0.8 mm, somewhat folded, with acute and fimbriate apex. Florets c. 500-800 per capitulum. Ray florets c. 40, female; corolla yellow; tube 1.3- 1.8 mm long; limb (8-)lo-12 x ( 1 - ) I 5 3 mm, with short glandular hairs, 3 apical lobes c. 0.3 mm long; style 2.5-3 mm long, style branches c. 0.5 mm long; immature cypselas 1-1.5 x 0.3 mm, slightly obconic, slightly 4-5 angled, longitudinally striate, with erect-patent hairs c. 0.2 mm long, carpopodium absent; pappus of several short scales 0.3-0.4 x 0.3 mm, irregularly dentate-fimbriate, and 3(-4) short bristles 0.8-1 mm long, about half or slightly more than half the length of the corolla tube, barbellate. Disc florets hermaphrodite, actinomorphic; corolla 3-4.5 mm long, apical lobes 0.3-0.5 mm long; anthers 3-3.5 mm long, apical appendage c. 0.5 mm long, lanceolate, endothecial tissue polarized, en- dothecial cells longer than wide, anther base minutely caudate, tails 0.3 mm long, calcarate, filament collar 0.2-0.4 mm long; style 3-4.5 mm long, style branches c. 0.5 mm long, with sweeping hairs acute (Fig. 2A); immature cypselas (0.8-)1-1.2 x 0.2-0.3 mm, slightly obconic, like those of the ray florets; pappus of several short scales 0.2-0.4 x 0.2- 0.3 mm, irregulary dentate-fimbriate, and 3-5(-7) short bristles (0.5-)0.8-1(-1.5) mm long, about one third or slightly more than one third of the length of the corolla, barbellate. - Fig. 1.
Note, The species most morphologically similar to Anisopappus fruticosus is probably A . chinensis Hook. & Am., which can be readly differentiated from the new species by its herbaceous habit, its stems rarely more than 50 cm tall, its ovate- lanceolate leaves with palmate venation, its often solitary capitula on peduncles generally 50-120 mm long, its ray florets less than 8 mm long, its obtuse style sweeping hairs (Fig. 2B) and its pappus without bristles conspicuously longer than the scales.
Nord. 1. BOI. 21(2) 2001 59

Fig. 2. SEM micrographs of the of style branch sweeping hairs. - A. acute sweeping hairs in Anisopappus fruricosus (Santos & Henriques 1146, LISC). - B. obtuse sweeping hairs in Anisopappus chinensis subsp. chinensis (Exell & Mendonca 1168, COI).
The specimen Gossweiler 11 137 from Angola, Huila, Ungueria, near Chaento River, 1340 m alt., 2.vi.1937 (COI, PRE) has similar leaves and a somewhat similar pappus to A. fruticosus, but shows certain differences from A . fruticosus as regards the remaining characters cited above, so we have not included it in A . fruticosus (though this option cannot be ruled out).
Phylogenetic relationships
The cladistic analysis yielded 106 equally parsimonious cladograms, each 97 steps long. The consistency index (CI) was 0.50, and the retention index (RI) 0.70.
Examination of the strict consensus tree of the 106 most parsimonious trees (Fig 3A) reveals poor resolution, as in Eldenas & Anderberg’s (1996) analysis, though the number of unresolved polytomies is even higher in our analysis.
Of particular relevance with respect to Eldenas & Anderberg’s analysis is the fact that Anisopappus latifolius ( S . Moore) B.L. Burtt (a species with somewhat deviant characters in comparison to the rest of the genus, notably the lack of receptacular paleae, lack of sclerenchyma in the cypselae, and presence of enlarged cypsela epidermis cells) forms a polytomy both with the Geigeria-Buphthalmum group and the remaining species of Anisopappus.
Indeed, although Anisopappus latifolius was positioned as a sister group of the rest of the genus Anisopappus in almost all of the 106 most parsimonious trees (e.g. Fig 3B), as in Eldenas & Anderberg’s analysis, in one tree (Fig. 3C) it was positioned as a sister group of the Geigeria- Buphthalmum group, and thus separated from the genus Anisopappus, which would make this genus paraphyletic. However, in the more recent phylogenetic analysis of Eldenas et al. (1999) based on nucleotide sequences of the chloroplast gene ndhF, the two species of Anisopappus analysed, A . latifolius and A. smutsii Hutch., form a monophyletic group with 100 % jackknife support .
As expected, and except in the single tree in which A. latifolius was positioned as a sister group of the Geigeria-Buphthalmum group, the topology of this latter group was identical to that of Eldenas & Anderberg’s strict consensus tree.
In our strict consensus tree (Fig. 3A), the remaining Anisopappus species formed an 1 1 -clade polytomy. However, most of the most parsimonious trees contained two major clades.
The first of these clades (monophyletic in 75% of the 106 trees) comprised A. holstii (0. Hoffm.) Wild, A . sylvaticus (Humbert) Wild, A . salvifolius (DC.) Wild, A . corymbosus Wild, A. smutsii and A. junodii Hutch. (as for example in Fig. 3B). Within this clade, A. holstii was a sister group to the rest of the species in almost all trees, while A . smutsii and A . junodii
Fig. 3. A. strict consensus tree derived from the 106 equally most parsimonious trees. - B. one of the 106 equally most parsimonious trees. - C. the single most parsimonious tree in which A . lurifolius was positioned as sister group to the Geigeriu- Buphrhulmum group. - D. one of the two cladograms resulting from the successive weighting analysis.
60 Nord. J . Bot. ZI(2) 2001

II
C D
Nard. 1. Bat. 21(2) 2001 61

were the most advanced species. Note that topology within the group comprising A . salvifolius, A . corymbosus, A . smutsii and A. junodii was invariable in the 106 most parsimonious trees, and is thus reflected in the strict consensus tree (Fig. 3A).
The other major clade comprised the remaining species of the genus, which formed a monophyletic group in all trees except that in which A . latifolius was positioned as a sister group of the Geigeria- Buphthalmum group. Within this clade, A . kirkii (Oliv.) Brenan was sister group to the remaining species (Fig. 3B), as in Eldenas & Anderberg’s analysis. Within the clade, A. pinnutifidus (Klatt) 0. Hoffm. ex Hutch. and A. pseudopinnatifidus were the most advanced species. This latter species, which was not included in Eldenas & Anderberg’s analysis, forms a clade with A . pinnatijidus, with which it shows great similarity (Ortiz & Paiva 1995). Again as in Eldenas & Anderberg’s analysis, in our strict consensus tree A . rhombifolius and A . grangeoides (Vatke & Hopfner ex Klatt) Merxm. formed a clade, since these species are identical as regards the characters considered in the analysis; notably, both species lack a pappus.
The position of A . fruticosus was not well defined in the strict consensus tree (Fig 3A). In all individual most parsimonious trees, however - except in the tree in which A . latifolius was positioned as a sister group of the Geigeria-Buphthalmum group (Fig. 3C), in which A . fruticosus was positioned as a sister group to the remaining species of Anisopappus - A . fruticosus was included in the A . kirkii / A . pseudopinnatijidus group (Fig. 3B). Its position within this clade was variable, although in 65% of the 106 trees it was positioned as a sister group of the most advanced subclade, comprising A . pinnutifidus and A . pseudopinnatijidus, which like A . fruticosus have capitula arranged in loose corymbs (though this character is not as clear in A . pseudopinnatijidus as in the other two taxa).
Analysis using a successive weighting process gave two cladograms (one of which is illustrated in Fig. 3D) with a somewhat similar topology to the most equally parsimonious cladograms in which A . latifolius was placed as the sister group of the remaining species of Anisopappus. However, these two cladograms contained two major clades some- what different to those cited above.
Acknowledgements - Our thanks go to Manuel Lainz for the Latin diagnosis, to Alfredo Lbpez “Tokio” for the illustrations, to Guy Norman for the English translation, and to the keepers of the herbaria mentioned for the loan of study material.
References Anderberg, A. A. 1991. Taxomony and phylogeny of the tri-
be lnuleae (Asteraceae). - Plant Syst. Evol. 176: 75-123. Anderson, L. E. 1954. Hoyer’s solution as a rapid mounting
medium for bryophytes. - Bryologist 57: 242-247. Eldenas, P. & Anderberg, A. A. 1996. A cladistic analysis of
Anisopappus (Asteraceae: Inuleae). - Plant Syst. Evol.
- , Anderberg, A. A & Kallersjo, M. 1998. Molecular phylogenetics of the tribe Inuleae s. str. (Asteraceae), based on ITS sequences of nuclear ribosomal DNA. - Plant Syst. Evol. 210: 159-173.
- , Kallersjo, M. & Anderberg, A. A. 1999. Phylogenetic placement and circunscription of tribes Inuleae s. str. and Plucheae (Asteraceae): evidence from sequences of chloroplast gene ndhF. - Molecular Phylogenetics and Evolution 13: 50-58.
Farris, J. S. 1969. A successive approximation aproach to character weighting. - Syst. Zool. 18: 374-385.
- 1988. Hennig86, ver. 1.5, users reference and software. - Stony Brook, New York.
Hoffmann, 0. 1890. Compositae. - In: Engler, H. G. A. & Prantl. K. A. E. (eds), Die Naturlichen Pflanzenfamilien. Englemann. Leipzig, pp. 87-387.
Holmgren, P. K, Holmgren, N. H. & Bamett, L. C. 1990. Index Herbariorum. I. The Herbaria of the World, 8th ed. - Regnum Veg. 120. New York Botanical Garden.
Humbert, H. 1923. Les ComposCes de Madagascar. - Mem. Linn. SOC. Normandie, Bot. 25: 1-334.
Lawalrke, A. 1974. Anisopappus marianus LawalrCe sp. nov. (Asteraceae) du Za’ire. - Bull. Jard. Bot. Belg. 44: 187- 190.
Lisowski, S. 1986a. Taxons nouveaux d’Inuleae (Asteraceae) du Zaire. - Bull. Jard. Bot. Belg. 56: 199-204.
- 1986b. Inuleae (Asteraceae) nouvelles d’Afrique centrale. - Bull. Jard. Bot. Belg. 56: 349-372.
- 1988. Asteraceae nouvelles d’Afrique centrale. - Bull. Jard. Bot. Belg. 58: 259-268.
Ortiz, S. & Paiva, J. A. R. 1995. Notes on African species of the genus Anisopappus Hook. & Am. (Asteraceae). - Bot. J. Linn. SOC. 117: 39-46.
- , Paiva, J. A. R. & Rodriguez-Oubiiia, J. 1996. An outline of the genus Anisopappus Hook. & Am. (Compositae). - Anales Jard. Bot. Madrid 54: 378-391.
Wild, H. 1964. A revision of the genus Anisopappus Hook. & Am. (Compositae). - Kirkia 4: 45-73.
199: 167-192.
62 Nord. J . BOI. 21(2) 2001




















