
Population Differences in Responses of Fathead Minnows (Pimephales promelas) to Visual and Chemical...
10
Ethology 93, 3 1 4 0 (1993) 0 1993 Paul Parey Scientific Publishers, Berlin and Hamburg ISSN 0179-1613 Department of Biology, University of Saskatchewan, Saskatoon Population Differences in Responses of Fathead Minnows (Pimephafes promefas) to Visual and Chemical Stimuli from Predators ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITH MATHIS, A., CHIVERS, D. P. & SMITH, R. J. F. 1993: Population differences in responses of fathead minnows (Pimephules promelus) to visual and chemical stimuli from predators. Ethology 93, 31-40. Abstract Fathead minnows (Cyprinidae: Pimephules promefus) from a population that is sympatric with predatory northern pike (Esocidae: Esox lun'us) exhibited a fright reaction to the visual stimulus of a live northern pike significantly more often than minnows from a population that is allopatric with pike. The fright response included increased use of shelter, dashing and freezing. Minnows from the pike-sympatric population also exhibited a significantly greater fright response, measured as a reduction in activity, following exposure to chemical stimuli from pike (i.e. water from a tank that had contained a pike) than did minnows from the pike-allopatric population. There was no significant change in activity by minnows from either population following exposure to chemical stimuli from nonpiscivorous peacock gudgeons (Eleotridae: Tuteurndinu ocefficuudu), suggesting that the differ- ence between the two populations is specific to stimuli from pike rather than a general difference in response to chemical stimuli from heterospecific fishes. Fathead minnows apparently utilize at least a two-tiered predator recognition system that incorporates both visual and chemical cues. Corresponding author: A. MATHIS, Department of Biology, University of Saskatchewan, Saskatoon, Saskatchewan, Canada, S7N OWO. Introduction The intensity of anti-predator behaviour is often correlated with the levels of predation that are experienced by prey populations under natural conditions. Predator-associated inter-population variation in anti-predator responses has been documented for a variety of organisms (e.g. spiders: RIECHERT & HEDRICK 1990; salamanders: DUCEY & BRODIE 1991; deermice: HIRSCH & BOLLES 1980). In these studies, individuals from populations in which predators are common typically exhibit stronger anti-predator responses than individuals from popula- tions that experience lower levels of predation. U.S. Copyright Clearance Center Code Statement: 01 79-1613/93/9301-0031$02.50/0
-
Upload
missouristate -
Category
Documents
-
view
3 -
download
0
Transcript of Population Differences in Responses of Fathead Minnows (Pimephales promelas) to Visual and Chemical...

Ethology 93, 3 1 4 0 (1993) 0 1993 Paul Parey Scientific Publishers, Berlin and Hamburg ISSN 0179-1613
Department of Biology, University of Saskatchewan, Saskatoon
Population Differences in Responses of Fathead Minnows (Pimephafes promefas)
to Visual and Chemical Stimuli from Predators
ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITH
MATHIS, A., CHIVERS, D. P. & SMITH, R. J. F. 1993: Population differences in responses of fathead minnows (Pimephules promelus) to visual and chemical stimuli from predators. Ethology 93, 3 1 - 4 0 .
Abstract
Fathead minnows (Cyprinidae: Pimephules promefus) from a population that is sympatric with predatory northern pike (Esocidae: Esox lun'us) exhibited a fright reaction to the visual stimulus of a live northern pike significantly more often than minnows from a population that is allopatric with pike. The fright response included increased use of shelter, dashing and freezing. Minnows from the pike-sympatric population also exhibited a significantly greater fright response, measured as a reduction in activity, following exposure to chemical stimuli from pike (i.e. water from a tank that had contained a pike) than did minnows from the pike-allopatric population. There was no significant change in activity by minnows from either population following exposure to chemical stimuli from nonpiscivorous peacock gudgeons (Eleotridae: Tuteurndinu ocefficuudu), suggesting that the differ- ence between the two populations is specific to stimuli from pike rather than a general difference in response to chemical stimuli from heterospecific fishes. Fathead minnows apparently utilize at least a two-tiered predator recognition system that incorporates both visual and chemical cues.
Corresponding author: A. MATHIS, Department of Biology, University of Saskatchewan, Saskatoon, Saskatchewan, Canada, S7N OWO.
Introduction
The intensity of anti-predator behaviour is often correlated with the levels of predation that are experienced by prey populations under natural conditions. Predator-associated inter-population variation in anti-predator responses has been documented for a variety of organisms (e.g. spiders: RIECHERT & HEDRICK 1990; salamanders: DUCEY & BRODIE 1991; deermice: HIRSCH & BOLLES 1980). In these studies, individuals from populations in which predators are common typically exhibit stronger anti-predator responses than individuals from popula- tions that experience lower levels of predation.
U.S. Copyright Clearance Center Code Statement: 01 79-1613/93/9301-0031$02.50/0

32 ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITH
Freshwater systems are especially well suited for inter-population compari- sons of antipredator behaviour because populations often differ in abundance and variety of predatory species. Several instances of differential anti-predator responses among freshwater fish populations have been documented. Guppies (Poecifiu reticufutu) from populations that commonly occurred with predatory fish exhibited significantly greater avoidance of predatory fish (MAGURRAN & SEGHERS 1990), selective avoidance of the “attack cone” around the jaws of the predator (MAGURRAN & SEGHERS 1990), and greater cohesiveness of schools (SEGHERS 1974 a) than individuals from populations where predatory fish were uncommon or absent. Similar responses have been reported for European min- nows (Phoxinus phoxinus), where individuals from populations that were sympat- ric with predatory northern pike (€sox fucius) exhibited reduced foraging (LEVES- LEY & MAGURRAN 1988), increased shoaling (MAGURRAN & PITCHER 1987) and increased numbers of inspection visits to either live or model pike predators (MAGURRAN 1986; MAGURRAN & PITCHER 1987) than minnows from populations that were allopatric with pike. Three-spined sticklebacks (Gusterosteus uculeutus) from populations that presumably experienced high levels of predation also exhibited more intense fright responses following exposure to predators than sticklebacks from populations with fewer natural predators (HUNTINGFORD 1982; GILES & HUNTINGFORD 1984).
All of the above studies included either the visual stimulus of the predator or were based on responses to general disturbance (e.g. chasing with a net). Chemical recognition of predators occurs in a wide variety of vertebrates (review: WELDON 1990), but to our knowledge no studies have considered possible population differences in the ability of individuals to recognize predators on the basis of chemical stimuli. In our study, we evaluated the effectiveness of both visual and chemical stimuli in mediating population differences in predator recognition abilities of fathead minnows (Pimephufes promefus).
In south-central Saskatchewan, fathead minnows occur in large numbers at two sites that differ in terms of the presence of predatory fish. Pike Lake is an oxbow lake of the South Saskatchewan River and contains large numbers of northern pike. Minnow shoals in Pike Lake commonly occupy the same micro- habitats (i.e. aquatic vegetation in shallow water) as are occupied by pike (pers. obs.). The second site, Marshy Creek, drains into Redberry Lake, a large saline lake that is an enclosed evaporation basin with no endemic piscivorous fish species. Therefore, minnows from this system are inexperienced with predatory fish in general and, specifically, have no experience with northern pike.
Our goal in this study was to determine if fathead minnows from Marshy Creek and Pike Lake differ in their ability to recognize northern pike as a potential predator on the basis of either visual cues alone or chemical cues alone. More specifically, we tested the hypotheses that fathead minnows from Pike Lake and Marshy creek differ in their responses to the following stimuli: (1 ) the visual stimulus of a live northern pike when chemical stimuli from the pike are not present, (2) chemical stimuli from a northern pike when visual stimuli from the pike are not present and (3) chemical stimuli from nonpiscivorous gobioid fish (peacock gudgeons: Tateurndinu ocefficuudu). We predicted that fathead min-

Population Differences in Predator Recognition by Fathead Minnows 33
nows from Pike Lake should exhibit a fright response to both visual and chemical stimuli from the pike while those from Marshy Creek should fail to respond to the pike stimuli with a fright reaction. Neither population was predicted to exhibit a fright reaction in response to the gudgeon stimulus.
Methods
Collection and Maintenance
Northern pike were col1eci:ed in the fall of 1990 and the spring of 1991 from Eagle Creek, a tributary of the North Saskatchewan River in south-central Saskatchewan, and from Pike Lake. Pike were maintained in 300-1 holding tanks at approx. 15 “C on a 14L : 10D photoperiod and were fed 1-2 fathead minnows once every 5 d.
Fathead minnows were collected from both Marshy Creek and Pike Lake. Minnows were captured in the fall of 1990 and were maintained in large (approx. 18,000 I) outdoor pools throughout the winter at temperatures ranging from approx. 15 “C (fall months) to 4 “C (winter months). Additional minnows were captured in the spring of 1991 and all minnows were placed in holding tanks in the laboratory. Minnows from the two populations were maintained separately in the laboratory in either 300-1 or 600-1 holding tanks at approx. 15-18 “C on a 14L : 10D photoperiod and were fed daily with commercial fish food.
Peacock gudgeons were obtained commercially and were maintained in the laboratory in a 37-1 aquarium at approx. 25 “C on a 12L : 12D photoperiod and were fed daily with frozen brine shrimp (Artemiidae: Arternia frunrisruna). Peacock gudgeons are small tropical fish native to Papua New Guinea.
Green swordtails (Poeciliidae: Xiphophorous helleri) were also obtained commercially and were fed the same diet as the fathead minnows. Swordtails, which were used as an alternative diet for the pike prior to collection of the chemical stimuli, are small, tropical fish native to Guatemala and Mexico.
Experiment 1: Response of Fathead Minnows to Visual Stimulus of Pike
Experimental observation tanks (37 I: 50 x 30 X25 cm) were positioned so that two adjacent tanks were visually separated by a removable barrier. All tanks were filled with dechlorinated tap water and were aerated with single airstones located at the back of the tanks. The tanks contained no filtration system, but the bottom of each tank was covered with a shallow layer of sodium zeolite chips to remove ammonia from the water. O n e tank in each pair contained a centrally-located shelter consisting of a ceramic tile (9.8 x 20.0 cm) mounted on three cylindrical glass legs (5.5 cm long).
20 fathead minnows from each population were arbitrarily selected from the holding tanks and were placed individually into experimental tanks that contained shelters (i.e. one fish per tank). Observations were conducted after the minnows had remained in the tanks undisturbed, except for daily feedings, for 2 d. Each minnow was tested twice, once with the predator absent and once with the predator present, resulting in the following four test conditions: (1) Marshy Creek minnows, predator absent, (2) Pike Lake minnows, predator absent, (3) Marshy Creek minnows, predator present, and (4) Pike Lake minnows, predator present.
During the “predator absent” trials, the tanks adjacent to the minnow tanks did not contain fish. All “predator absent” trials were conducted prior to “predator present” trials because fathead minnows that are exposed to a fright stimulus subsequently respond with a fright reaction to neutral stimuli that are presented in the same manner as the fright stimulus (MATHIS & SMITH 1993). Minnows from Marshy Creek and Pike Lake were tested in alternating order. To decrease the possible incidence of habituation by the pike to the test conditions, 10 different northern pike were used as predators [mean fork length = 26.9 cm k 2.02 (1 SD)]. Each pike was used twice for each population with a minimum of 2 d between trials.
During each trial, an observer was positioned approx. 1.5 m in front of each tank. Each minnow was observed for 8 min and the amount of time that the minnow spent beneath the shelter was recorded. At the end of 8 min, the barrier between the pair of tanks was removed and the minnow
Ethology, Vol. 93 ( I ) 3

34 ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITH
was observed for an additional 8 min. During the post-stimulus treatment, we also recorded presence or absence of two additional behaviour patterns. “Dashing” was a sudden burst of very rapid, apparently disoriented swimming. “Freezing” occurred when minnows sank to the substrate and became immobile for a minimum of 30 s. Both of these behavioural patterns have been interpreted as components of the fright reaction in fathead minnows (e.g. LAWRENCE & SMITH 1989).
The “predator absent” treatment was included as a control to determine if there was a population difference in response to any disturbance (i.e. removal of the barrier between the tanks) even in the absence of a potential predator. At the end of the “predator absent” observation, the barrier was replaced between adjacent tanks and one northern pike was placed into the tank beside the minnow tank. Observations of minnows in the “predator present” condition were conducted a minimum of 3 h (max. = 4.5 h) following introduction of the pike. “Predator present” tests were conducted using procedures identical to that described above for the “predator absent” condition.
Pre- and post-stimulus activities within each of the four conditions were evaluated statistically using a Wilcoxon matched-pairs signed-ranks test (ZAR 1984). The responses (i.e. differences in activity between pre- and post-stimulus conditions) of minnows from the two populations to either “predator absent” or “predator present” stimuli were compared using the Mann-Whitney U test (SIEGEL 1956). For the number of occurrences of dashing and freezing, the two populations were compared using the xz test; when the number of occurrences were too low to allow the use of the xz test, the Fisher exact probability test was used (SIEGEL 1956). One-tailed statistical tests were used when there was a predicted direction of difference (i.e. “predator present” condition, Pike Lake versus Marshy Creek) and two-tailed statistical tests were used when there was no predicted direction of difference (i.e. “predator absent” condition, Pike Lake versus Marshy Creek).
Experiment 2: Response of Fathead Minnows to Chemical Stimuli from Pike
Stimulus Preparation. Three pike [mean fork length = 19.2 cm 2 3.55 (1 SD)] were used to prepare the stimulus water. For each of three feedings (i.e. once every 5 d) immediately prior to stimulus collection, test pike consumed approx. equal volumes (range = 3 - 4 ml, measured by volumetric displacement in water) of swordtails. MATHIS & SMITH (1993) found that fathead minnows from Marshy Creek (i.e. allopatric with pike) respond with a fright reaction to chemical cues from pike that had eaten conspecific minnows but not to chemical cues from pike that had eaten swordtails. In our experiment, pike were fed swordtails to exclude fright responses based on the predator’s diet. Immediately prior to the last feeding before collection of the stimulus water, pike were moved to a separate holding tank (150 I) where they consumed an equal volume of swordtails as in previous feedings. Approx. 1 h after the prey fish were introduced into the holding tank, each test pike was removed from the tank and rinsed with dechlorinated tap water to remove any residue of the swordtails from the pike’s skin. The pike were then transferred into a new holding tank that was identical to the first holding tank and that contained clean water. After approx. 16 h, test pike were transferred to separate, clear plastic chambers (26 x 8 x 8 cm) that contained 1200 ml of dechlori- nated tap water. The chambers were well-aerated but contained no filtration system. After 3 d, the pike were removed from the chambers and water from the chambers was frozen in separate 5-ml units.
Stimulus water from peacock gudgeons was prepared as a control to determine if the two populations would respond differentially to chemical stimuli from nonpiscivorous fish. Approx. 2 h following feeding with brine shrimp, six gudgeons [mean fork length = 4.83 cm f 0.197 (1 SD)] were removed from the aquarium, rinsed with dechlorinated tap water to remove any residue of the brine shrimp, and placed in 1200 ml of dechlorinated tap water in the same chamber as that described above for collection of stimulus water from the pike. After 3 d the gudgeons were removed from the chamber and the stimulus water was frozen in separate 5-ml units.
Testing Protocol. Test fathead minnows (mean fork length f 1 SD: Marshy Creek = 4.93 cm f 0.571; Pike Lake = 5.10 cm f 0.561) were selected arbitrarily from both laboratory populations and were individually placed into separate plexiglass acclimation tanks (45 x 45 x 20 cm). Acclimation tanks were constantly supplied with an inflow of clean water at a rate of approx. 250 ml/ min and the outflow was regulated so that a constant depth of 4-5 cm was maintained. Acclimation and testing occurred under a photoperiod of 14L : 10D at a mean water temperature of 17 “C. Test minnows were fed daily as before.
After 2 d in the acclimation tanks, minnows were moved into separate testing tanks that were identical to the acclimation tanks except that the flow rate was increased to 500 ml/min. The holding
Population Differences in Predator Recognition by Fathead Minnows 35
tanks and the testing apparatus are described in detail by LEMLY & SMITH (1986, 1987). O u r system differed from that of LEMLY & SMITH (1986, 1987) only in that outflowing water from our tanks was discarded rather than recirculated.
Each testing tank was surrounded by an Opto-Varimex-Aqua tracking meter (Columbus Instruments) which was interfaced to a computer. The activity tracking meter established a grid of light beams in the tank and the computer was preset to “scan” the light beams for breaks at intervals of 0.125 s between scans. The information from the scans was integrated and digitized by the computer to provide an estimate of the position of the fish in the grid and to quantify the fish’s level of activity. Two measures of activity were quantified: (1) total distance travelled (cm) and (2) number of “stereotypic” movements (i.e. activity in which the fish breaks light beams without moving outside of one grid square).
Video equipment allowed an observer in an adjacent room to view the experimental tanks without disturbing the minnows. Trials lasted a total of 16 rnin. At the end of 8 rnin, the observer injected the stimulus water into inflow water lines which passed through the observation room before entering the experimental room. lnjections of vegetable dye into the inflow lines demonstrated that it took approximately 14 s for the dye to reach the experimental tanks following injection. 20 minnows from each population were tested with the order of testing determined randomly. Experimental observations were conducted between 08.30 and 14.45 h.
Responses of the two populations to chemical stimuli from pike and to chemical stimuli from peacock gudgeons were analyzed separately. Because identity of individual pike did not significantly affect responses by minnows of either population (Kruskal-Wallis one-way ANOVA by ranks, P > 0.05 for all comparisons), responses of the minnows to stimulus water from the three different pike were combined for further statistical analyses of data. Statistical analyses were similar to those used for Exp. 1 .
Results
Experiment 1: Response of Fathead Minnows to Visual Stimulus of Pike
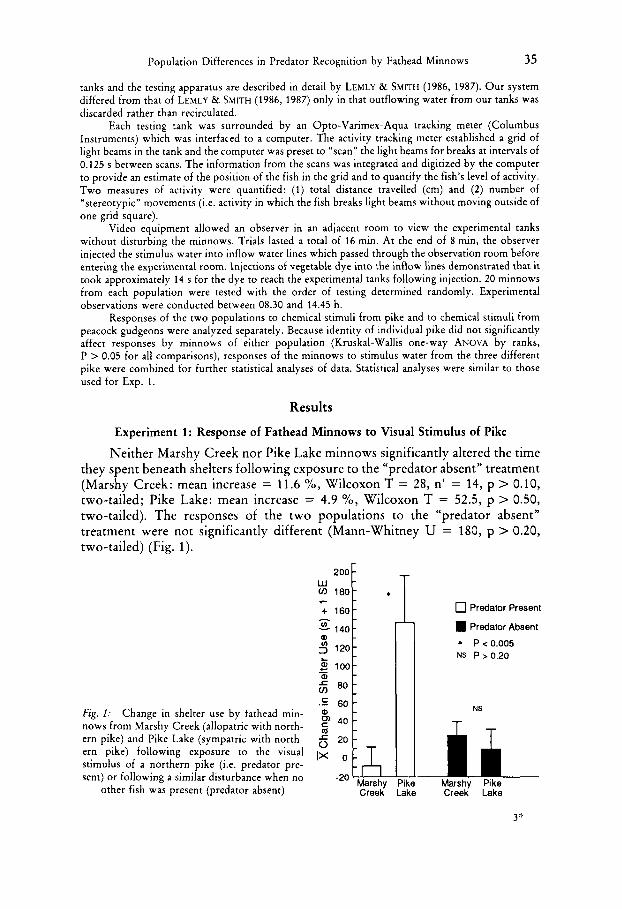
Neither Marshy Creek nor Pike Lake minnows significantly altered the time they spent beneath shelters following exposure to the “predator absent” treatment (Marshy Creek: mean increase = 11.6 %, Wilcoxon T = 28, n’ = 14, p > 0.10, two-tailed; Pike Lake: mean increase = 4.9 %, Wilcoxon T = 52.5, p > 0.50, two-tailed). The responses of the two populations to the “predator absent” treatment were not significantly different (Mann-Whitney U = 180, p > 0.20, two-tailed) (Fig. 1).
200 - v) 180-
+ 160-
-% 140- 0
w r
h -
3 120-
6 8 0 -
L
2 100- - al
- .E 60
Fig. I : Change in shelter use by fathead min- nows from Marshy Creek (allopatric with north- ern pike) and Pike Lake (sympatric with north- ern pike) following exposure to the visual stimulus of a northern pike (i.e. predator pre- sent) or following a similar disturbance when no
(11
6 20 lX
Marshy Pike Marshy Pike other fish was present (predator absent) Creek Lake Creek Lake
-20
0 Predator Present
Predator Absent * P c 0.005 NS P > 0.20
NS
T ,
3”

36 ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITH
Minnows from Marshy Creek also did not significantly alter the time they spent under the shelter following exposure to visual stimuli from northern pike (mean decrease = 5.3 YO, Wilcoxon T = 42, n‘ = 13, p > 0.50, two-tailed). However, minnows from Pike Lake showed a significant overall increase in shelter use following exposure to the visual stimulus of the pike (mean increase = 109 %, T = 16, n’ = 17, p < 0.005, one-tailed). The difference in shelter use by Marshy Creek versus Pike Lake minnows was statistically significant (Mann- Whitney U = 97.5, p < 0.005, one-tailed) (Fig. 1).
In the “predator absent” condition, there was no significant difference between the two populations for either the proportion of trials in which dashing occurred or the proportion of trials in which freezing occurred (Table 1). In the “predator present” condition, significantly greater proportions of Pike Lake minnows exhibited both dashing and freezing in response to the visual stimulus of a pike than did minnows from Marshy Creek (Table 1).
Table I : Number of minnows from Marshy Creek (allopatric with northern pike) and Pike Lake (sympatric with northern pike) exhibiting fright responses (i.e. dashing or freezing) following exposure to the visual stimulus of a northern pike (predator present) or under identical experimental
conditions when no pike were present (predator absent)
Yes No 2 P
A. Predator absent 1. Dashing
0 > 0.99‘) Pike Lake 5 15 Marshy Creek 5 15
Pike Lake 5 15 Marshy Creek 2 18
2. Freezing
- > 0.207
B. Predator present 1 . Dashing
Pike- Lake Marshy Creek
2. Freezing Pike Lake Marshv Creek
13 4
14 5
7 16
6 15
6.55 < 0.02‘)
6.42 < 0.02‘)
a) 2 test, b, Fisher exact probability test.
Experiment 2: Response of Fathead Minnows to Chemical Stimuli from Pike
Minnows from Marshy Creek did not significantly alter their activity following exposure to chemical stimuli from northern pike in terms of either distance travelled (mean decrease = 22.8 %, Wilcoxon T = 76, n’ = 19, p > 0.20, two-tailed) or stereotypic movements (mean decrease = 14.8 YO, Wilcoxon T = 68, n’ = 19, p > 0.20, two-tailed). In contrast, minnows from Pike Lake significantly decreased their activity following exposure to chemical

Population Differences in Predator Recognition by Fathead Minnows 37
stimuli from pike as measured by both distance travelled (mean decrease = 75.4 %, Wilcoxon T = 7, n’ = 19, p < 0.0005, one-tailed) and stereotypic movements (mean decrease = 52.2 YO, Wilcoxon T = 18, n’ = 18, p < 0.0025, one-tailed). Minnows from Pike Lake exhibited a significantly greater decrease in activity in comparison to minnows from Marshy Creek for both distance travelled (Mann-Whitney U = 115.5, p < 0.025, one-tailed) (Fig. 2A) and stereotypic movements (Mann-Whitney U = 137.5, p < 0.05, one-tailed) (Fig. 2B).
0 Pike Stimulus a, a, > ? 400 I-
Gudgeon Stimulus
$: f, * P<0.025 .s + 0,
NS P > 0.10 -7
- 3
n IX
: - l o o ~ A W I 1 & a,
Marshy Pike Marshy Pike Creek Lake Creek Lake
0 a .-
Fig. 2: Change in distance travelled (A) and number of stereotypic move- ments (B) by fathead minnows from Marshy Creek (allopatric with northern pike) and Pike Lake (sympatric with northern pike) following exposure to chemical stimuli from northern pike or chemical stimuli from nonpiscivarous
peacock gudgeons
B *,r ll& 50
Marshy Pike Creek Lake
* P c 0.05 NS P > 0.10
NS T
Creec Lake
There was no significant change in activity by minnows from either popula- tion following exposure to chemical stimuli from peacock gudgeons as measured by both distance travelled (Marshy Creek: mean decrease = 20.7 %, Wilcoxon T = 75, n’ = 20, p > 0.25, two-tailed; Pike Lake: mean decrease = 51.0 %, T = 65, n’ = 20, p > 0.10, two-tailed) and stereotypic movements (Marshy Creek: mean decrease = 10.6 YO, Wilcoxon T = 71.5, n’ = 20, p > 0.25, two-tailed; Pike Lake: mean decrease == 30.8 YO, T = 73.5, n‘ = 20, p > 0.25, two-tailed). The responses of minnows from Marshy Creek and Pike Lake to chemical stimuli from gudgeons did not differ significantly for either distance travelled (Mann- Whitney U = 187, p > 0.10, two-tailed) (Fig. 2A) or stereotypic movements (Mann-Whitney U = 193.5, p > 0.10, two-tailed) (Fig. 2B).

38 ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITH
Discussion
The results of this study demonstrate that fathead minnows from a popula- tion that is sympatric with northern pike (i.e. Pike Lake) are better able to recognize pike as potential predators than are minnows from a population that is allopatric with pike (i.e. Marshy Creek). These results are consistent with the results of other studies of small fishes that have reported differential predator recognition abilities between individuals from populations that were sympatric versus allopatric with predators (HUNTINGFORD 1982; GILES & HUNTINGFORD 1984; MAGURRAN 1986; MAGURRAN & PITCHER 1987; LEVESLEY & MAGURRAN 1988; MAGURRAN & SEGHERS 1990). Similar results also have been reported for other vertebrates, including pied flycatchers (Ficedulu hypoleucu) (CURIO 1975) and deermice (Peromyscus municulutus) (HIRSCH & BOLLES 1980). Our results further suggest that either visual cues alone or chemical cues alone may be sufficient for predator identification by fathead minnows.
In the experiment demonstrating visual recognition of pike by fathead minnows from Pike Lake, chemical stimuli and other nonvisual cues (e.g. displacement of water by the predator) were not a factor because minnows and pike were kept in a separate tanks with no exchange of water. Furthermore, the difference between the two populations was not merely the result of a differential response to disturbance because identical test conditions in the absence of pike did not elicit a fright response from Pike Lake minnows. The importance of vision as a mechanism for predator recognition for fishes has been suggested by other studies in which prey fishes have exhibited fright responses following exposure to live predators in separate tanks (e.g. MILINSKI 1987; DUGATKIN & ALFIERI 1991) or plastic bags (e.g. COATES 1980) and studies that have used artificial predator models (e.g. SEGHERS 1974 b; MAGURRAN 1986; SMITH & SMITH 1989). In addition, European minnows respond with the most intense fright reaction to predator models whose shape and coloring most realistically resemble that of a live predator (MAGURRAN & GIRLING 1986).
While visual detection of predators is undoubtedly an important component in an individual’s anti-predator arsenal, reliance solely on visual cues may be of limited utility under conditions in which visibility is poor (e.g. in muddy streams) or when predators are cryptic. Northern pike are primarily ambush foragers that spend most of their time in the cover provided by aquatic vegetation (SAVINO & STEIN 1989) and may therefore be difficult to detect on the basis of visual cues alone. Because of the cryptic nature of pike under natural conditions, predator detection by nonvisual cues may be of critical importance to the survival of potential prey. Our study demonstrates that chemical stimuli from pike elicit a fright response from pike-sympatric fathead minnows even when the predator is not visible. The fright response exhibited by minnows from Pike Lake is apparently a specific response to chemical stimuli from pike rather than a general disturbance response because the minnows did not exhibit a fright response to chemical stimuli from an unfamiliar, nonpiscivorous fish (i.e. peacock gudgeons). MATHIS & SMITH (1993) demonstrated that pike-naive minnows from Marshy Creek did not respond with a fright reaction to chemical stimuli from pike unless

Population Differences in Predator Recognition by Fathead Minnows 39
the pike had recently eaten conspecific minnows. In contrast, the results of our current study indicate that minnows from a population that is sympatric with pike (i.e. Pike Lake) are able to recognize the potential danger represented by the presence of the pike’s chemical stimuli regardless of the pike’s recent diet.
Experience has been shown to alter the predator recognition abilities of both predator-naive European minnows (MAGURRAN 1989) and fathead minnows (MATHIS & SMITH 1993). However, recent experience with predators does not appear to be a requisite for predator recognition because all minnows in our study were maintained in predator-free tanks in the laboratory for a minimum of 6 mo prior to testing. Studies of laboratory-reared guppies (SEGHERS 1974 a, b; BREDEN et al. 1987) and European minnows (MAGURRAN 1990) have shown that popula- tion differences in fright responses may be present in predator-naive individuals, and MAGURRAN (1990) suggested that although anti-predator behaviour in Euro- pean minnows appears to be inherited, it may be modified by early experience with predators. Experiments with individuals reared in predator-free environ- ments are required to determine if there is also a genetic component to the differential predator recognition abilities exhibited by fathead minnows from Pike Lake and Marshy Creek.
O u r study demonstrates that fathead minnows from two populations that vary in exposure to pike predation under natural conditions show corresponding variation in their ability to recognize pike as a potential predator. Furthermore, fathead minnows apparently utilize at least a two-tiered predator recognition system that incorporates both visual and chemical cues. Chemical cues may be of particular importance in the identification of a predator such as the northern pike that is often either behaviorally (SAVINO & STEIN 1989) or morphologically (WEBB 1984) cryptic. The ability to recognize and respond to predators may have important implications in terms of the survival (HIRSCH & BOLLES 1980) and, ultimately, the fitness of prey individuals.
Acknowledgements We thank Susan MATHIEU for sharing her ideas concerning population differences in predator
recognition behaviour by fathead minnows and Elizabeth HINKS for assistance with the set-up of experimental tanks. Funding was provided by the University of Saskatchewan and the Natural Science and Engineering Research Council of Canada.
Literature Cited BREDEN, F., SCOT, M., & MICHEL, E. 1987: Genetic differentiation for anti-predator behaviour in
the Trinidad guppy, Poecilza reticulata. Anim. Behav. 35, 61 8-620. COATES, D. 1980: The discrimination of and reaction towards predatory and nonpredatory species of
fish by humbug damselfish Dascyllus aruanus (Pisces, Pomacentridae). Z. Tierpsychol. 52, 347-354.
CURIO, E. 1975: The functional organization of anti-predator behaviour in the pied flycatcher: a study of avian visual perception. ,4nim. Behav. 23, 1-1 15.
DUCEY, P. K. & BRODIE, E. D., JR. 1991: Evolution of antipredator behaviour: individual and population variation in a neotropical salamander. Herpetologica 47, 89-95.

40 ALICIA MATHIS et al., Population Differences in Predator Recognition by Fathead Minnows
DUGATKIN, L. A. & ALFIERI, M. 1991: Tit-for-tat in guppies (Poeciliu reticuluta): the relative nature of cooperation and defection during predator inspection. Evol. Ecol. 5, 3OC-309.
GILES, N . & HUMINGFORD, F. A. 1984: Predation risk and inter-population variation in anti- predator behaviour in the three-spined stickleback, Gasterosteus uculeatus L. Anim. Behav. 32, 264-275.
HIRSCH, S. M. & BOLLES, R. C. 1980: O n the ability of prey to recognize predators. Z. Tierpsychol.
HUNTINGFORD, F. A. 1982: Do inter- and intraspecific aggression vary in relation to predation pressure in sticklebacks? Anim. Behav. 30, 909-916.
LAWRENCE, B. J. & SMITH, R. J. F. 1989: Behavioral response of solitary fathead minnows, Pimephules promefas, to alarm substance. J. Chem. Ecol. 15, 209-219.
LEMLY, A. D. & SMITH, R. J. F. 1986: A behavioral assay for assessing effects of pollutants on fish chemoreception. Ecotoxicol. Environ. Saf. 11, 21C-218.
_ _ & -- 1987: Effects of chronic exposure to acidified water on chemoreception of feeding stimuli in fathead minnows (Pimephules promelus): mechanisms and ecological implications. Environ. Toxicol. Chem. 6, 2 2 S - 2 3 8 .
LEVESLEY, P. B. & MAGURRAN, A. E. 1988: Population differences in the reaction of minnows to alarm substance. J. Fish Biol. 32, 699-706.
MAGURRAN, A. E. 1986: Predator inspection behaviour in minnow shoals: differences between populations and individuals. Behav. Ecol. Sociobiol. 19, 267-273.
-- 1989: Acquired recognition of predator odour in the European minnow (Phoxinus phoxinus). Ethology 82, 216-223.
-- 1990: The inheritance and development of minnow anti-predator behaviour. Anim. Behav. 39, 834-842.
-- & GIRLING, S. L. 1986: Predator model recognition and response habituation in shoaling minnows. Anim. Behav. 34, 51&518.
-- & PITCHER, T. J. 1987: Provenance, shoal size and the sociobiology of predator-evasion behaviour in minnow shoals. Proc. R. SOC. Lond. B 229, 4 3 9 4 6 5 .
- _ & SEGHERS, B. H. 1990: Population differences in predator recognition and attack cone avoidance in the guppy Poecifiu reticulutu. Anim. Behav. 40, 4 4 3 4 5 2 .
MATHIS, A. & SMITH, R. J. F. 1993: Fathead minnows, Pimephulespromefus, learn to recognize pike, Esox lucius, as predators on the basis of chemical stimuli from minnows in the pike's diet. Anim. Behav., in press.
MILINSKI, M. 1987: Tit for tat and the evolution of cooperation in sticklebacks. Nature 3 2 5 , 4 3 3 4 3 5 . RIECHERT, S. E. & HEDRICK, A. V. 1990: Levels of predation and genetically based anti-predator
SAVINO, J. F. & STEIN, R. A. 1989: Behavior of fish predators and their prey: habitat choice between
SEGHERS, B. H . 1974a: Schooling behavior in the guppy (Poeciliu reticulatu): an evolutionary
_ - 1974 b: Geographic variation in the responses of guppies (Poecilia reticuluta) to aerial pre-
SIEGEL, S. 1956: Nonparametric Statistics for the Behavioral Sciences. McGraw-Hill, New York. SMITH, R. J. F. & SMITH, M. J. 1989: Predator-recognition behaviour in two species of gobiid fishes,
WEBB, P. B. 1984: Form and function in fish swimming. Sci. Am. 251, 72-82. WELDON, P. J. 1990: Responses by vertebrates to chemicals from predators. In: Chemical Signals in
Vertebrates V. (MACDONALD, D. W., MULLER-SCHWARZE, C. & SILVERSTEIN, R. M., eds.) Plenum Press, New York, pp. 5OC-521.
54, 71-84.
behaviour in the spider, Agelenopsis upertu. Anim. Behav. 40, 6 7 9 4 8 7 .
open water and dense vegetation. Env. Biol. Fish. 24, 287-293.
response to predation. Evolution 28, 486-489.
dators. Oecologia 14, 93-98.
Asterropteyx semipunctatus and Gnatholepis anjerensis. Ethology 83, 19-30.
ZAR, J. H. 1984: Biostatistical Analysis. 2nd ed. Prentice-Hall, Englewood Cliffs.
Received: Februa y 24, 1992
Accepted: July 30, 1992 f'J. Brockmunn)




















