
Assessing vulnerability of freshwater minnows in the Gangetic ...
13
Climate Risk Management 33 (2021) 100325 Available online 18 May 2021 2212-0963/© 2021 Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/). Assessing vulnerability of freshwater minnows in the Gangetic floodplains of India for conservation and management: Anthropogenic or climatic change risk? Uttam Kumar Sarkar a, *, 1 , Koushik Roy b, 1 , Malay Naskar a , Gunjan Karnatak a , Mishal Puthiyottil a , Snigdha Baksi a , Suman Kumari a , Lianthuamluia Lianthuamluia a , Basanta Kumar Das a a ICAR-Central Inland Fisheries Research Institute, Barrackpore 700120, West Bengal, India b University of South Bohemia in Ceske Budejovice, Faculty of Fisheries and Protection of Waters, South Bohemian Research Center of Aquaculture and Biodiversity of Hydrocenoses, Institute of Aquaculture and Protection of Waters, ˇ Cesk´ e Budˇ ejovice 370 05, Czech Republic A R T I C L E INFO Keywords: Vulnerability assessment Climate change Small indigenous fishes Mesh size Overfishing threat ABSTRACT Minnows are the most ignored yet indispensable group of freshwater fishes in Asian inland wa- ters. The reproductive resilience of minnows facing climatic variability, using a wetland inhab- iting species Amblypharyngodon mola (Mola carplets) in lower Indo-Gangetic floodplains, was validated. Results revealed that spawning decision in females (threshold gonadosomatic index ≥ 5 units) is neither cued by water temperature nor rainfall. They can maintain pre-spawning fitness (condition factor 1.12–1.25 units) within a broad temperature (22–33 ◦ C) and rainfall (0–800 mm) window by active feeding, thus no risk of skipped spawning decisions while facing future climatic variabilities. Present breeding phenology (May-December) might have prolonged in the recent decade, especially the tail-end, concomitant with increasingly hot and rainy monsoon (May-August) and warmer post-monsoon months (September-December). Minnows are expected to prosper in a future climatic scenario, contributing to ecosystem balance (algal grazers) and regional food security. Female first maturity (♀ puberty) was encountered at 4.7–5.1 cm total length, hinting at a probable increase in the recent decade. Climate-favored prolonged recruit- ment window, in absence of extreme fishing pressure (currently), might have led to such pattern. However, this state might be temporary and labile. Minnows may soon get altered to earlier puberty (=warning sign of stock collapse) if fishing pressure intensifies under a reproductively favoring climate progression. Threshold body girth for spawning females was estimated at 3.2–3.4 cm (+17% than non-breeding ones). Fishing nets having mesh sizes (=total circumfer- ence) at least > 32–34 mm will most likely be the key to minnows’ endurance or survival in the coming decades. * Corresponding author. E-mail address: [email protected] (U. Kumar Sarkar). 1 UKS and KR contributed equally to the work. Contents lists available at ScienceDirect Climate Risk Management journal homepage: www.elsevier.com/locate/crm https://doi.org/10.1016/j.crm.2021.100325 Received 17 February 2021; Received in revised form 28 April 2021; Accepted 13 May 2021
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Assessing vulnerability of freshwater minnows in the Gangetic ...

Climate Risk Management 33 (2021) 100325
Available online 18 May 20212212-0963/© 2021 Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Assessing vulnerability of freshwater minnows in the Gangetic floodplains of India for conservation and management: Anthropogenic or climatic change risk?
Uttam Kumar Sarkar a,*,1, Koushik Roy b,1, Malay Naskar a, Gunjan Karnatak a, Mishal Puthiyottil a, Snigdha Baksi a, Suman Kumari a, Lianthuamluia Lianthuamluia a, Basanta Kumar Das a
a ICAR-Central Inland Fisheries Research Institute, Barrackpore 700120, West Bengal, India b University of South Bohemia in Ceske Budejovice, Faculty of Fisheries and Protection of Waters, South Bohemian Research Center of Aquaculture and Biodiversity of Hydrocenoses, Institute of Aquaculture and Protection of Waters, Ceske Budejovice 370 05, Czech Republic
A R T I C L E I N F O
Keywords: Vulnerability assessment Climate change Small indigenous fishes Mesh size Overfishing threat
A B S T R A C T
Minnows are the most ignored yet indispensable group of freshwater fishes in Asian inland wa-ters. The reproductive resilience of minnows facing climatic variability, using a wetland inhab-iting species Amblypharyngodon mola (Mola carplets) in lower Indo-Gangetic floodplains, was validated. Results revealed that spawning decision in females (threshold gonadosomatic index ≥5 units) is neither cued by water temperature nor rainfall. They can maintain pre-spawning fitness (condition factor 1.12–1.25 units) within a broad temperature (22–33 ◦C) and rainfall (0–800 mm) window by active feeding, thus no risk of skipped spawning decisions while facing future climatic variabilities. Present breeding phenology (May-December) might have prolonged in the recent decade, especially the tail-end, concomitant with increasingly hot and rainy monsoon (May-August) and warmer post-monsoon months (September-December). Minnows are expected to prosper in a future climatic scenario, contributing to ecosystem balance (algal grazers) and regional food security. Female first maturity (♀ puberty) was encountered at 4.7–5.1 cm total length, hinting at a probable increase in the recent decade. Climate-favored prolonged recruit-ment window, in absence of extreme fishing pressure (currently), might have led to such pattern. However, this state might be temporary and labile. Minnows may soon get altered to earlier puberty (=warning sign of stock collapse) if fishing pressure intensifies under a reproductively favoring climate progression. Threshold body girth for spawning females was estimated at 3.2–3.4 cm (+17% than non-breeding ones). Fishing nets having mesh sizes (=total circumfer-ence) at least > 32–34 mm will most likely be the key to minnows’ endurance or survival in the coming decades.
* Corresponding author. E-mail address: [email protected] (U. Kumar Sarkar).
1 UKS and KR contributed equally to the work.
Contents lists available at ScienceDirect
Climate Risk Management
journal homepage: www.elsevier.com/locate/crm
https://doi.org/10.1016/j.crm.2021.100325 Received 17 February 2021; Received in revised form 28 April 2021; Accepted 13 May 2021
Climate Risk Management 33 (2021) 100325
2
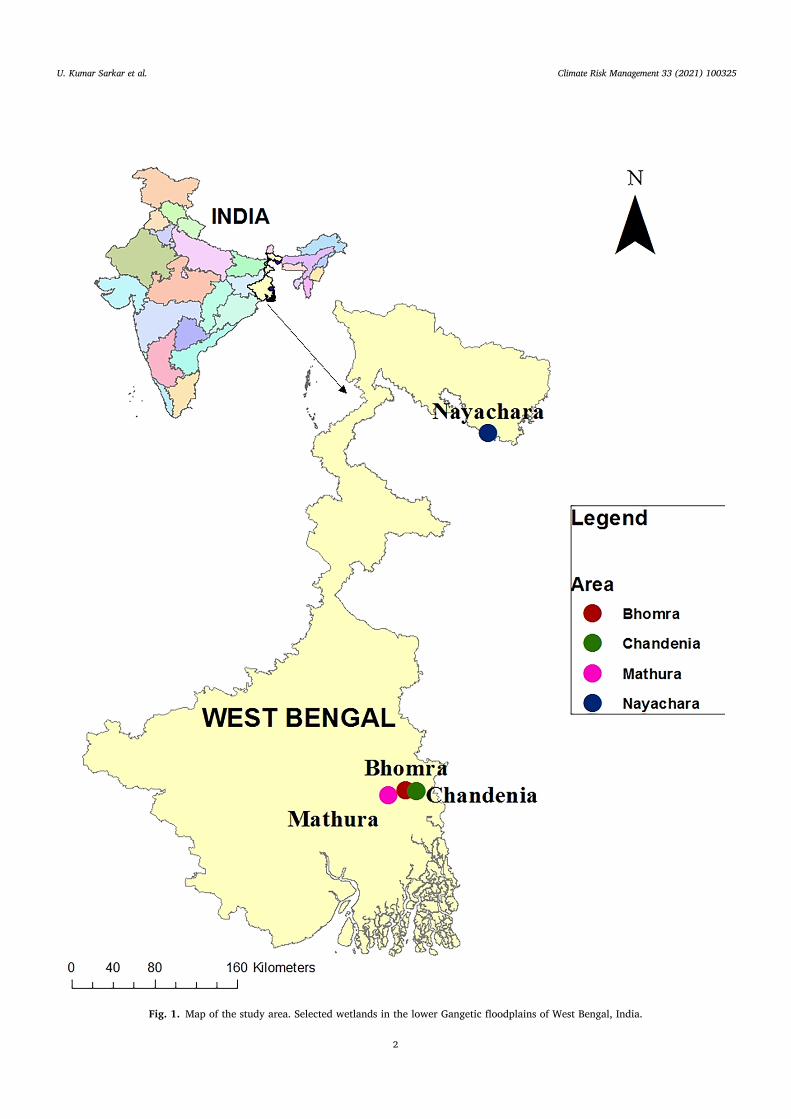
Fig. 1. Map of the study area. Selected wetlands in the lower Gangetic floodplains of West Bengal, India.
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
3
1. Introduction
Small indigenous fishes or minnows are perhaps the most ignored, yet, an indispensable assortment of fishes in inland freshwaters. Their adults reaching can reach up to 8.5–18 cm total body length (Sarkar et al. 2019a). Minnows lack full-fledged commercial interest in regional fish markets fetching around or below 3 USD kg−1 and often perceived as ‘by-catch’ sales to other commercially important fish groups. They have a major contribution in inland fish diversity. About 450 out of ~ 765 identified freshwater fish species in Indian inland waters belong to this category, primarily cyprinids (Sarkar and Lakra, 2010). Their importance as cheap and nutrient-rich food fish also cannot be overlooked (Nordhagen et al. 2020, Mohanty et al. 2011). It is proven that consumption of minnows or small indigenous fishes has benefits of higher micronutrient delivery in the human diet, especially pregnant and lactating women, infants, and young children (6–23 months) (Bogard et al. 2015). For example – the subject of our study, Amblypharyngodon mola offers a nutrient profile of 18.5 g high-quality protein (with complete amino acid profile), ~1 g PUFA (polyunsaturated fatty acids; dominated by omega-3 fractions), essential minerals 1.64 g (rich in Ca, Zn, P, and Mg) and very high vitamin-A content (>1500 retinol activity equivalent) per 100 g whole-body consumption (equivalent to ~ 50 mola of ~ 5.9 cm length) (Mohanty et al. 2011).
Taking the example of India (the world’s second-largest inland fish producer; FAO, 2018), inland fisher community and fisheries researchers presume minnows as the first and easy hits of climate change. Therefore, any decline in their population is attributed to changing climate or habitat deterioration (Sarkar et al., 2019a). Despite their status of staple aquatic food for many poor fishing communities worldwide, assessment of their vulnerability in the context of future supply has been overlooked (Sarkar et al., 2019a, Bogard et al., 2015). Presently such assessments are even more critical because of the regional patterns of changing climate. For example, in our present study area, the mean air temperature has increased in the range of 0.18–0.28 ◦C (mean 0.22 ◦C), and total annual rainfall has decreased in the range of 135.6–257 mm (mean 192.5 mm) during last three and half decades (1980–2015) (Karnatak et al. 2018, Naskar et al. 2018). Interestingly, the general biology of minnows has remained of much interest to fishery biologists but scarcely dealt with their vulnerabilities. Morphometrics, population characterization, feeding ecology, and reproductive biology of the minnow A. mola have been adequately studied (reviewed in Gupta and Banerjee, 2014, 2015).
The IUCN (International Union for Conservation of Nature) classifies most minnows, including A. mola, as ‘least concern’ (Chaudhry, 2010). However, a prologue on the issue of climate change and the declining population of minnows in rivers and wetlands can be found in Sarkar et al. (2018b), Naskar et al. (2018), Mandal and Nandi (2015), and ICSF (2010). It can be attributed to the existing loophole in the regional fishing strategies (or lack of legislation). Around/above 20 mm mesh size fishing nets have been operational to harvest minnows and previously reported from lower Gangetic floodplain wetlands (Sandhya et al., 2016, 2019; Sarkar et al., 2018b; Das and Barat, 2014). Floodplain wetlands are essential resources for inland freshwaters in the Indian sub-continent and support inland fisheries (either open-access or managed by fishermen cooperatives) quite strongly (Mamun 2007; Naskar et al. 2018). India has 0.55 million ha of wetlands supporting 85 fish species and have production potential of>1000 kg ha−1 (Sarkar et al., 2021a; Naskar et al., 2018). Following our previous attempt with P. sophore (Sarkar et al., 2019a), we attempted to validate the high reproductive resilience of minnows towards climatic variability, using a second necessary ‘group’ of minnows in the region, i.e., A. mola.
We hypothesized that an artificial threat on recruitment of minnow population exists, rather than the ‘easily presumed’ climate change or habitat deterioration. We further added – if regional climate change is not a critical threat (following a reproduction-climate change vulnerability assessment; Sarkar et al. 2019a), how at least, the threat from anthropogenic fishing pressure could be mitigated (?). This information may reverse the regional perception of declining SIFs germplasm (Sarkar et al., 2019a, 2019b, 2018a, 2018b; Naskar et al., 2018). For this, we propose a novel parameter, ‘pre-spawning girth (Girthspawn50)’. Girthspawn50 is an applied parameter capable of giving responsible ‘minimum’ mesh size recommendations to prevent recruitment overfishing threats (Sarkar et al. 2021b). Here, we classified the act of indiscriminative fishing as the ‘anthropogenic-threat variable.’ By indiscriminative fishing, we imply the irresponsible and data-blind selection of mesh sizes in fishing nets used in regional inland waterbodies – ultimately leading to pop-ulation decline (discussed above).
We envisaged to address the above idea through a systematic data-driven approach that involved identification of – (a) breeding phenology; (b) climato-hydrological influence and threshold gonadosomatic index (GSI) for breeding; (c) pre-spawning fitness and climate preferendum; (d) Sizes at maturity and pre-spawning girth; (e) back-validation of Girthspawn50 with LM50 using length-girth relationship, (f) regional climatic variability and reproductive vulnerability to changing climate, and; (g) mesh sizes of operational
Table 1 Descriptive account of female Amblypharyngodon mola sampled from floodplains in lower-Gangetic plains (West Bengal, India).
Parameters a
Total Length (TL, cm)
Weight (g) Girth (cm) b GSI (units) Condition factor (units)
Growth coefficient (b-value of LWR) c
Length-Girth relationshipd
4.5–8.5 (5.9 ±0.6)
0.88–6 (2.13 ±0.76)
1.6–4.4 (2.1 ±0.8)
0.1–14.5 (3.35 ±3.83)
0.62–1.85 (1.01 ±0.17)
2.56 (0.745, p < 0.01) Girth = 0.060+(0.46*Length)
a Data presented in Range and Mean ± Standard error of mean (in parentheses). b Girth divided by 2 = Body depth. c Calculated using log-transformed length-weight data of females and linear regression of length-weight relationship (LWR). The calculated value of ‘b’ and adjusted R2 and p-value of LWR (in parentheses) were given. d Length-girth regression (LGR) based on female sex data. Adjusted R2-value = 0.457, p < 0.01.
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
4
fishing nets and anthropogenic threat.
2. Materials and methods
2.1. Study area, species selected, sample collection, and analysis
Five Gangetic floodplain wetlands in West Bengal, India, were selected: Bishnupur (33 ha), Bhomra (46 ha), Chandania (48 ha), Mathura (20 ha) and Nayachara (62 ha) (Fig. 1). One minnow species, Amblypharyngodon mola (Hamilton, 1822), commonly known as ‘mola carplets’ or ‘mourola’ (in vernacular language). The criteria for its selection as a model fish species in the present study was due to its dominance (among minnows), consistent availability, and social relevance as a poor man’s food fish. The species has a moderate market price throughout the year around INR 200/kg or USD 2.7/kg (author observations). Altogether 817 specimens (327 female, 490 male, and non-sexed) were collected between June 2015 and December 2018, spanning over two complete breeding seasons. Only females were selected as model sex owing to the ease in observing gonadal maturity status. Sample collection and analysis were done following Sarkar et al. (2019a), Sarkar et al. (2019b). Descriptive data on female mola carplets sampled during the present study is furnished in Table 1. Simultaneously with fish sampling, hydrological parameters were recorded following standard methodologies (APHA, 2012). Location-wise data on monthly rainfall (mm) was procured from Indian Meteorological Department (IMD, Alipore, West Bengal). Habitat parameters recorded during the study period are provided in Table 2.
2.2. Modeling of environmental cues and breeding thresholds
Modeling climato-hydrological influence on GSI and estimation of threshold GSI for breeding was done in Sarkar et al. (2019a), Sarkar et al. (2019b). The breeding GSI value was used as a reference GSI while identifying thresholds of environmental cues for breeding. All analyses were done in ‘R-software’. Fulton’s condition factor (KF), hereinafter referred to as fitness, was calculated using the formula given in Froese (2006). Pre-spawning fitness (Kspawn50), thermal and precipitation window(s) for the attainment of Kspawn50 (termed as climate preferendum) was estimated following Sarkar et al. (2019a), Sarkar et al. (2019b). Fishes (females) in the ‘mature’ and ‘ripe’ stage were coded as ‘1’ while the rest coded ‘0’. Within the sample distribution of breeding season, the size of the smallest mature or ripe female encountered was deemed as size at first maturity (Ainsley et al. 2011). In comparison, the body size range at which there is a ≥ 50% probability of encountering a mature or ripe female is deemed as size at 50% maturity. Size at 50% maturity (LM50) was estimated using total body length data (binary coded like K spawn50) (Sarkar et al., 2019a, 2019b).
Maximum body depth (in cm) was measured in each fish and multiplied by 2 to give maximum body girth (hereinafter referred to as ‘girth’). Threshold body girth, beyond which>50% of the female fish population may spawn, was coined as pre-spawning girth (Girthspawn50) and estimated following Sarkar et al. (2017). A bivariate linear regression model was developed for total body length (independent variable) and maximum body girth (response variable); length-girth relationship (LGR) (Table 1). The estimated pre- spawning girth (Girthspawn50), to be used for ‘responsible’ minimum mesh size recommendation, was collated with the model extrapolated girth at pre-spawning length (LM50). The coherence between Girthspawn50 and ‘LGR extrapolated girth’ (at LM50) warranted the preciseness of mesh size recommendation. The mean body girth of breeding females (mature and ripe) was also compared with the non-breeding ones (immature and maturing), checking whether there is any need to increase the mesh size demand.
2.3. Climate data analysis and reviewing mesh size of regional fishing nets
Region-specific data on changing patterns of air temperature, annual rainfall, and inter-season rainfall composition over the last three and half decades were taken from Karnatak et al. (2018), Naskar et al. (2018), and Sarkar et al. (2019a), Sarkar et al. (2019b). Data (n = 28) on mesh sizes of different operational fishing nets in the regional floodplain wetlands were collected from the published reviews (Sandhya et al. 2019, Das and Barat, 2014). Minima, maxima, and the inter-quartile range of operational mesh sizes were calculated. Based on Girthspawn50 estimates, a safe or unsafe mesh size range was attributed, keeping in mind the chances of a gravid
Table 2 Descriptive account of habitat parameters recorded in the study area.
Parameters Observed Range
Water temperature (◦C) 20.7–33.4 Dissolved oxygen (mg L−1) 4–8.6 Free carbon dioxide (mg L−1) 4.2–10.3 pH (units) 7.4–8.7 Total Dissolved Solids (mg L−1) 176–593 Conductivity (µS cm−1) 184–797 Total alkalinity (mg L−1) 78–317 Total hardness (mg L−1) 91–265 Salinity (mg L−1) 90.7–268.5 Nitrate (mg L−1) 0.01–0.46 Phosphate (mg L−1) Nil-0.3 Monthly rainfall (mm) 0–928
U. Kumar Sarkar et al.
Climate Risk Management 33 (2021) 100325
5
female with outstretched abdomen to successfully pass through. The girth is supposed to correspond to the complete circumference of a mesh (=mesh size of a fishing net). If mesh size < Girthspawn50, the mesh wraps tightly around the broadest part of the fish body, beyond which a fish cannot escape further. If mesh size > Girthspawn50, gravid females can escape pre-mature fishing mortality.
3. Results
3.1. Breeding phenology and its historical chronology
The breeding phenology of Amblypharyngodon mola females in Gangetic floodplains of West Bengal, India, was observed between May and December (Fig. 2). This eight-month-long breeding season makes this minnow species ‘almost’ a perennial breeder. From a climate perspective, the breeding remains cued within a wide thermal and precipitation window. Historical chronology (1978–2020) of their breeding phenology in the floodplains of lower Indo-Gangetic plains encompassing West Bengal (Eastern India) and Bangladesh have been reviewed (Table 3). A visual analysis of the chronogram reveals that the breeding phenology might have prolonged in the recent decades, especially the tail-end of the breeding season (Fig. 3). The average duration of breeding season re-ported before the year 2008 seems to be 6.3 months. In the recent decade (2008–2020), the average duration of the reported breeding season seems to be 8.3 months (Table 3).
3.2. Environmental cues and breeding thresholds
Mola females showed no significant dependency on water temperature and rainfall as breeding cues. Therefore, no specific breeding thresholds could be estimated. This pattern can also be justified because A. mola breeds through 3 seasons (pre-monsoon, monsoon, and post-monsoon). The threshold GSI for breeding was estimated at ~ 5 units (Deviance explained: 95.6%, GCV: 0.01, p < 0.01) (Fig. 4). Overall, GSI values ranged between 0.1 and 14.5 units. The pre-spawning fitness (Kspawn50) of female mola was estimated at 1.14 units, with a 95% confidence interval (C.I.) of 1.12–1.25 units (Fig. 5a). Our dataset recorded Fulton’s condition factor or body fitness (K) values between 0.62 and 1.85 units. Body fitness levels (K) and sexual maturity (GSI) had a weak, positive, but statistically significant correlation (Pearson’s 2-tailed r = 0.3, p < 0.05) (Fig. 5b). It implies that the females’ spawning decisions are weakly dependent on body fitness levels. However, the gonad weight-adjusted condition factor values (Kadj; from body weight, without gonad weight) showed statistically significant and positive correlations with either of the climatic parameters: (a) Kadj and water temperature (Pearson’s 2-tailed r = 0.6, p < 0.05), or, (b) Kadj and monthly rainfall (Pearson’s 2-tailed r = 0.3, p < 0.05). The correlation coefficient of ‘Kadj-temperature’ is more potent than ‘Kadj-rainfall’ (Fig. 5b). A. mola females seem capable of maintaining fitness levels near Kspawn50 across a broad thermal (water temperature 22–33 ◦C; Fig. 6a) and precipitation window (0–800 mm monthly rainfall; Fig. 6b).
3.3. Size at maturity and its historical chronology
The first maturity of females (♀) was observed at 4.7 cm total body length (TL) in the sampling year 2017, within a size range of 4.5–8.5 cm TL. The second smallest mature female was observed at 5 cm TL. In the sampling year 2018, ♀ first maturity was observed at 5.1 cm. Altogether we encountered ~ 9% of gravid females in the range of 5 ± 0.3 cm TL with the smallest size(s) at maturity, and they mainly were captured in September and October. Whether these are the young-of-the-year (YOY) fish, born in May and matured in September-October, could not be probed. Nonetheless, the present first maturity of females (=puberty) might be happening at ≈5 cm TL. Historical chronology (1978–2020) of reported ♀ first maturity in the floodplains of lower Indo-Gangetic plains encompassing West Bengal, India, and Bangladesh has been reviewed (Table 3). The chronogram hints at a probable delay in attaining puberty in the
Fig. 2. Annual cycle of the gonadosomatic index (GSI, expressed as gonad weight % of total body weight) of Amblypharyngodon mola females in the lower Gangetic floodplains (West Bengal, India). The blue curve is the LOESS fitted trendline of the GSI trajectory observed in our sample. The horizontal green dashed line indicates the benchmark for successful breeding events (=threshold GSI > 4.6 units; Fig. 4). The boxplot heads or outliers emerging above the benchmark indicate breeding phenology. Sufficient samples with conspicuous gonads could not be encountered in March-April. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
6
recent decade, with most females attaining puberty near or above 5 cm TL lately (Fig. 7). Size at 50% maturity (LM50), i.e., size beyond which there is a ≥ 50% chance of encountering a mature female, was much higher than the first maturity. With a 95% confidence interval, ♀ LM50 was estimated to be 6.5–6.7 cm TL (Fig. 8).
Table 3 Chronological information on reported breeding phenology and size at first maturity of Amblypharyngodon mola females in floodplains of lower Indo- Gangetic floodplains (West Bengal, India, and Bangladesh).
Year Onset Offset Duration (months) First maturity TL (cm) Reference
1978 March September 7 – Afroze and Hossain (1990) 1983 June October 5 – Parveen (1984) 1988 May October 6 – Rahman (1989) 1990 May September 5 – Mustafa (1991) 1999 May October 6 – Kohinoor et al. (2005) 2000 March October 8 4.7 Azadi and Mamun (2004) 2004 April October 7 4.4 Suresh et al. (2007) 2005 March August 6 4.8 Saha et al. (2009) 2007 April October 7 4.3 Hoque and Rahman (2008) 2008 May October 6 4.6 Alam et al. (2018) 2010 April December 9 5.2 (5–5.5) Gupta and Banerjee (2013) 2011 – – – 4.9 Rahman et al. (2018) 2011 April October 7 4.9 Mondal and Kaviraj (2013) 2013 April December 9 – Mondal et al. (2020) 2014 April December 9 5.2 Pal and Mahapatra (2018) 2015 – – – 5.3 Karnatak et al. unpublished 2016 March October 8 5.3 Ferdous Ahamed et al. (2017) 2017–2018 May December 8 4.7–5.0 Present study 2020 April December 9 – Hossain (2021)
*Converted reported SL (4.1 cm) to TL by adding + 1.2 cm based on standard morphometry of A. mola females (Baishya et al. 2010)
Fig. 3. Chronogram of the breeding phenology of Amblypharyngodon mola females in the lower Indo-Gangetic floodplains. Blue bars indicate the active breeding season. Data source (n = 17): Afroze and Hossain (1990), Parveen (1984), Rahman (1989), Mustafa (1991), Azadi and Mamun (2004), Kohinoor et al. (2005), Suresh et al. (2007), Saha et al. (2009), Hoque and Rahman (2008), Alam et al. (2018), Gupta and Banerjee (2013), Mondal and Kaviraj (2013), Mondal et al. (2020), Pal and Mahapatra (2018), Ferdous Ahamed et al. (2017), Hossain (2021) and present study. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
U. Kumar Sarkar et al.
Climate Risk Management 33 (2021) 100325
7
3.4. Pre-spawning girth and its back-validation
Threshold body girth, beyond which>50% of the female fish population may spawn, i.e., Girthspawn50 was estimated at 3.2–3.4 cm (with 95% C.I.) (Fig. 9). The range of recorded body girth in our sample was 1.6–4.4 cm. The model for length-girth relationship (Adj. R2: 0.457, p < 0.01) was estimated as: Girth = 0.06+ (0.46*Length) (Table 1). The mediocre R2 value of the LGR might be attributed to (a) variations within samples due to the presence of different maturity stages (with different girth) and (b) variations in body shape
Fig. 4. GAM model showing threshold GSI for breeding (4.6 units) of female Amblypharyngodon mola in Gangetic Floodplain wetlands. Breeding event = 1 indicate successful spawning. GSI is expressed as gonad weight % of total body weight. Black dashes on the x-axis indicate data points.
Fig. 5. (a, b): (a) Kaplan-Meier survival estimate of Pre-spawning fitness (Kspawn50: 1.12–1.25) of female Amblypharyngodon mola in Gangetic Floodplain wetlands. (b) correlogram of body fitness (CF; standard and adjusted), gonadal maturation (GSI), and climatic parameters (water temperature; rainfall).
Fig. 6. (a, b): LOESS smoothed thermal (left panel; 22–33 ◦C) and precipitation preferendum (right panel; 0–800 mm) for attainment of Kspawn50 in Amblypharyngodon mola. The unit of condition factor (K) is expressed as density factor or mass deposited per cubic three-dimensional length (length, width, height) with a non-standard unit of g per cm3 %.
U. Kumar Sarkar et al.
Climate Risk Management 33 (2021) 100325
8
Fig. 7. Chronology of reported the first maturity in Amblypharyngodon mola females from the lower Indo-Gangetic floodplains. Data points are indicated in red, and the blue line is GLM fitted trendline. Marginal boxplot indicates the distribution of reported first maturity (♀) across the literature. Data source (n = 12): Azadi and Mamun (2004), Suresh et al. (2007), Saha et al. (2009), Hoque and Rahman (2008), Alam et al. (2018), Karnatak et al. unpublished, Ferdous Ahamed et al. (2017) and the present study. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
Fig. 8. Kaplan-Meier survival estimate for size at 50% maturity (LM50; 6.5–6.7 cm) of female Amblypharyngodon mola in Gangetic Flood-plain wetlands.
Fig. 9. Kaplan-Meier survival estimate for pre-spawning girth (Girthspawn50; 3.2–3.4 cm) of female Amblypharyngodon mola in Gangetic Floodplain wetlands. Corresponds to responsible minimum mesh size recommendations.
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
9
form. Based on this model and using LM50 estimates (6.5–6.7 cm) as input, the projected girth at 50% maturity came out to be ~ 3–3.1 cm. This is in close agreement with our Girthspawn50 estimates. The immature, maturing and spent females had a mean body girth of 5.4 cm. The mature or ripe females, with their abdomen outstretched due to fully formed ovaries, increased girth by + 16.9% (mean girth 6.3 cm). It means gravid females require a bigger mesh to swim through without being stuck, necessitating a concomitant 16.9% increase in the provision of responsible mesh size. A historical chronology in this regard could not be derived due to the lack of historical data in this regard.
3.5. Regional climate change patterns
Over the last three and half decades, the climate in and around lower Gangetic floodplains, of which the present study is a part, has warmed up by 0.18–0.28 ◦C (mean 0.22 ◦C). The increasing trend of mean air temperature was most intense during monsoon months (May to August). During the period 1980–2015, there have been eight instances (years) when the mean air temperature in post- monsoon months (September to December; autumn–winter) remained substantially warm, i.e., ≥26 ◦C. About 50% of such hot post-monsoon months have occurred frequently since 2006. The annual rainfall has decreased by 135.6–257 mm (mean 192.5 mm). Rainfall distribution across pre-monsoon (January-April), monsoon (May to August), and post-monsoon (September-December) have changed too. Pre and post-monsoonal rainfall have decreased by 0.6–2.4% and 3.3–5.8%, respectively. The monsoonal rainfall has increased (3.8–6.2%).
3.6. Mesh sizes of operational fishing nets and anthropogenic threat
Swollen-bellied mature-ripe females are more vulnerable to be obstructed in the fishing net meshes than the slender-bellied immature-maturing females. Presently, the mesh size of operational fishing nets in the regional wetlands ranges from 1 mm to 100 mm, with an inter-quartile range of 5–30 mm. The mesh size is essentially the total perimeter or circumference of individual mesh in fishing nets. Collating the Girthspawn50 of female mola, a major anthropogenic threat on recruitment of A. mola is apparent due to the use of fishing gears having mesh sizes below 32–34 mm. Since gear-specific fish catch data is usually not maintained at these open-water fisheries resources, a retrospective evaluation of the damage done could not be accomplished.
4. Discussions
4.1. Reproductive vulnerability to regional climate change
Over the past three and half decades, the climate in the lower Gangetic floodplains has been warming (mean air temperature +0.05–0.08 ◦C decade−1) and becoming water-stressed (total annual rainfall − 38.74–73.42 mm decade−1) (Sarkar et al., 2019a, 2019b). Presently, the breeding phenology of mola in lower Gangetic floodplains has duration of eight months, i.e., May to December. Review of existing literature on the species revealed a potential prolongation of the breeding phenology (Table 3). The tail end of the breeding season might have prolonged since the last decade (2008–2020). It might be concomitant to the frequent occurrences of unusually warmer post-monsoonal months since 2006 (≥26 ◦C mean air temperature; Karnatak et al. 2018). Globally, such pheno-logical responses to warming climate have been documented in temperate to tropical fish species and presently summarized in an open-access database (https://ficli.shinyapps.io/database/; Krabbenhoft et al., 2020). In the Indian sub-continent, the breeding phenology of several fish species have either prolonged (e.g., Puntius sophore; Sarkar et al. 2019), shortened (Tor putitora; Joshi et al., 2018), unchanged (Channa punctata; Gudusia chapra; Sarkar et al., 2021b, Karnatak et al., 2018) or projected to be altered (Eutro-piichthys vacha; Sarkar et al., 2019a) in response to changing climate.
There have been quite a few attempts to quantify environmental cues for breeding and breeding thresholds of some fish species in the Indian sub-continent. For example – in riverine catfishes of River Ganga (Mystus tengara, M. cavasius, E. vacha), there are specific thresholds of water temperature (>27 ◦C, >31 ◦C, ~24 ◦C respectively) and monthly rainfall (>250 mm, >220 mm, ~100 mm respectively) which trigger breeding or spawning (Sarkar et al. 2019b). Whereas in an amphidromous croaker (Johnius coitor), only water temperature (23–25 ◦C) serves such function (Sarkar et al. 2018a). For floodplain dwelling snakehead species (Channa punctata), fulfillment of certain rainfall threshold (800–1400 mm) cue spawning (Karnatak et al., 2018, 2020). On the other hand, minnows like Gangetic pool barb (Puntius sophore) could breed at a low threshold rainfall requirement (>50 mm) and within a wide temperature range (20–30 ◦C) (Sarkar et al. 2019a). In A. mola, neither water temperature nor rainfall significantly triggered spawning. As such, no climatic breeding thresholds (water temperature and rainfall windows) were identified. Due to the lack of specific breeding thresholds of water temperature and rainfall, the present rate of changing climate is highly unlikely to pose any threat to attaining breeding GSI in mola females. The present pattern of changing climate might be favouring their reproduction in the long run (via prolongation of breeding phenology) due to a comparably lower breeding GSI demand (A. mola GSI ~ 5 units versus P. sophore GSI ~ 11 units; Sarkar et al. 2019a).
Even before the environmental cues for breeding, the decision whether a female fish would spawn or skip spawning depends on body fitness levels or condition factors (positively correlated with body calorific value; Myszkowski, 2013). The relationship cascade of climate change → condition factor (energy reserves) → skipped spawning decisions have been adequately described in Sarkar et al. (2017) and Jørgensen et al. (2006). It was revealed that spawning decisions in A. mola females are weakly dependent on body fitness levels. However, the correlations between gonad weight subtracted condition factor and water temperature or rainfall assure that somatic body fitness might remain uncompromised even if the regional rainfall continues to decline under a warming climate. It might
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
10
be true because A. mola can actively feed and keep up its energy reserves even during its breeding season (Mondal and Kaviraj, 2013). In contrast, other small indigenous species cease feeding activities during the breeding season (Gudusia chapra; Mondal and Kaviraj, 2010) and becomes more sensitive towards breeding cues or indiscriminative fishing (Sarkar et al., 2021b). Concerning climate preferenda, for pre-spawning fitness (Kspawn50), P. sophore could attain Kspawn50 within broad rainfall conditions (0–700 mm), while no dependence on temperature existed (Sarkar et al. 2019a). A. mola has equally broad and flexible climate preferenda (22–33 ◦C; 0–800 mm) for the attainment of Kspawn50. Even the Kspawn50 in A. mola (1.12–1.25 units) is lower than the reproductively resilient P. sophore (1.61–1.67 units) (Sarkar et al. 2019a). These insights indicate no chance of skipped spawning decisions in regional minnows (like mola or Gangetic pool barbs) while facing future climatic variabilities. Our finding further validates the high reproductive resilience of minnows in the face of climatic variability (Sarkar et al., 2019a).
4.2. Reproductive vulnerability to anthropogenic threat
In the present study, by ‘anthropogenic threat’, we have studied the possible chances of indiscriminative fishing threats on the native minnow population. Over the last three decades, the size or age at first maturity, whether or not under climate change context, is a well-established proxy marker for assessing indiscriminative fishing threats on exploited, wild fish population (Forestier et al. 2020; Karjalainen et al., 2016; Otero et al., 2012; Kuparinen et al., 2011; Rijnsdorp et al. 2010; Conover et al., 2009; Fukuwaka and Morita, 2008; Edeline et al., 2007; de Roos et al., 2006; Trippel, 1995). Commonly, a decrease in size at first maturity (=advancement of puberty), or rarely, an increase in size at first maturity (delaying of puberty), are both parts of fish species’ response to ‘fishery-induced evolution’ (FIE). Sometimes, the effects of FIE aggravate under an irreversibly changing climate (reviewed in Sarkar et al., 2021b). Although the developed regions (e.g., United States, Canada, United Kingdom) have been indulged in such research for decades, unfortunately, the developing regions with rich fisheries resources (e.g., India, Bangladesh) have lacked such investigations due to data limitations (Sarkar et al., 2021b). Reduction in size at first maturity by ~ 25–36% over four decades of changing climate and indis-criminative fishing was recently documented in clupeid females (Gudusia chapra) (Sarkar et al., 2021b). The reason was attributed to changing climate, which favored gonadal development and breeding, but at the same time, the gravid females were overexploited (indiscriminately fished) due to poor selection of responsible mesh sizes of fishing gears. The FIE, in such case, favored smaller maturing females to avoid such mesh sizes. As a result, the population shifted towards earlier puberty attainment, which regional climate change also facilitated. In contrast, hard-to-catch and bottom-dwelling snakehead fish (Channa punctata) in wetlands did not change their size at first maturity over the same four decades (Karnatak et al., 2020). It might be due to their ability to burrow in the mud and bottom dwelling nature making them less prone to fishing mortality and surface warming, respectively (author observations). Nonetheless, body-size reductions with warming are strongest in aquatic ectotherms than any other taxa on the planet, driven by a temperature-oxygen limitation(s) cascade (reviewed in Verberk et al. 2021).
The present study indicated a probable increase in size at first maturity vis-a-vis probable delay in attaining puberty in female mola minnows; presently at 4.7–5.1 cm or ≈5 cm TL (Table 3; Fig. 7). Even our present ♀ LM50 estimate (6.5–6.7 cm TL) was higher than one previous estimate (5.5–6 cm) from nearby waters earlier this decade (sampling 2009–2010; Gupta and Banerjee, 2013). Although rare and generally perceived to be good, increasing size at maturation has been previously documented in wild Scomber scombrus, sandeels group (Rijnsdorp et al. 2010), Menidia menidia (Conover et al. 2009), Oncorhynchus keta (Fukuwaka and Morita, 2008), Esox lucius (Edeline et al. 2007), and Morone saxatilis, Perca flavescens, Clupea harengus (Trippel, 1995). Such phenomenon has been often attributed to: (a) a release or absence of fishing pressure (Conover et al., 2009; Fukuwaka and Morita, 2008); (b) high-density, inter- species competition, pelagic zooplankton shortages leading to slower growth in the early days of life, due to an over-abundance of young-of-the-year (YOY) fish (Otero et al., 2012; Edeline et al., 2007; Trippel, 1995); (c) cannibalism (in predators) or intense pre-dation pressure which naturally selects individuals growing quickly to a threshold body-size to avoid predation (Forestier et al., 2020; Edeline et al., 2007). We attribute a combination of all these aspects to explain our observation. The fishing pressure and FIE on A. mola might have been less intense compared to other species. It could have been facilitated by a favorably changing climate allowing long breeding season and higher recruitment. The prolonged recruitment might have created a higher density of YOY fish in the habitat. Higher YOY fish availability might have attracted predators in the locality or caused pelagic zooplankton shortages. Presently observed larger sizes at maturity or delayed puberty might be temporary (Conover et al., 2009; de Roos et al., 2006). It may soon get altered to smaller sizes at maturity if climate change intensifies (Kuparinen et al. 2011, Rijnsdorp et al., 2010) or FIE increases (Conover et al., 2009; de Roos et al., 2006).
4.3. Future of minnows under mixed context of climate change and indiscriminative fishing
Even if this increasing trend of the first maturity is ignored, the smallest size at first maturity we observed (4.7 cm TL) matches with ‘one’ earlier estimate from the region, almost two decades ago (4.7 cm based on samples of 1999–2000; Azadi and Mamun, 2004). It indicates, unlike the overexploited clupeids in regional inland waters (Sarkar et al., 2021a, 2021b), any reduction in size at first maturity has ‘definitely’ not happened for this minnow. The present size at first maturity in mola females matches with that of another minnow. P. sophore females also decreased their size at first maturity till 4.7 cm TL compared to earlier (~6 cm TL) (Sarkar et al., 2019a, 2019b). In regional minnow females, we suspect that attaining puberty around 4.7–5 cm TL helps minnow species to avoid indiscriminative fishing mortality (avoiding mesh sizes). This process is also allowed by the regional pattern of changing climate (discussed above). Under the present state of changing climate, minnows seem to be a reproductively resilient species for the future. Under the future climatic scenario, minnows offer high potential towards regional freshwater ecosystem stability (primarily algal feeder, grazing at the base of the aquatic food chain), especially when most regional inland waters are predicted to become eutrophic
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
11
(more algae) under climate change (Das Sarkar et al., 2020; Rolighed et al., 2016). Through successful and prolonged recruitment patterns in future climatic scenarios (present observations), minnows would continue to offer high food and nutritional security to the local and impoverished communities (Nordhagen et al., 2020; Bogard et al., 2015), when livestock production may become limited by future climate variability (Rojas-Downing et al. 2017).
Further indiscriminative fishing pressure will most likely be the key to minnows’ endurance or survival in the coming decades. If puberty is advanced or smaller size-at-maturity is achieved, it could be an early warning sign of stock collapse (Sarkar et al., 2021b; Conover et al. 2009; Olsen et al., 2004). At this moment, the key to unleashing such detrimental effects depends on the presence or absence of indiscriminative fishing pressure under a favorably changing climate (=warmer monsoon, post-monsoon/ winter; present observations). Our data exhibited a ~ 17% increment in body girth of female minnows when they are ready to breed compared to the non-breeding ones. It is the essence of our parameter ‘pre-spawning girth’ (♀ Girthspawn50), which indirectly estimates if a mesh size is safe enough for gravid females to swim through. We highlight the need to use fishing nets having mesh sizes (=total circumference) above 32–34 mm (or longest mesh opening gap of > 16–17 mm) to abate artificial threats to A. mola or minnow population. Presently 5–30 mm mesh size fishing nets are operational (Sarkar et al., 2018b; Sandhya et al., 2016, 2019; Das and Barat, 2014), despite a general ban on gill nets below 25 mm mesh size in regional waters (Thomas, 2019). It is unclear how most authors have measured and reported mesh sizes of fishing nets, for example, total circumference versus bar length or longest mesh opening (ICES, 2004). Only if the longest mesh opening (gap) of operational fishing nets are > 16–17 mm, the gravid minnow females are safe, otherwise not.
To implement mesh size regulation for multi-species based wetland fishery in the tropical region, it is imperative to develop a first- hand database on ♀ Girthspawn50 and/or length-girth regressions (LGR, Table 1) of dominant minnows in the region (Sarkar et al., 2021a, 2021b). Then, revision of obsolete mesh size limits (unamended since last three decades) are necessary (sensu Kelkar and Dey, 2020). Looking at the similarity of female first maturities (≈5 cm TL) in two minnow species (P. sophore and A. mola; Sarkar et al. 2019, present study), we postulate that minimum responsible mesh size requirements might be interchangeable for most minnows. The key to sustenance of the minnow population in regional inland waters depends on managing anthropogenic (indiscriminative fishing) threats, as the changing climate is even favoring recruitment.
5. Conclusion
Small indigenous fishes or minnows are perhaps the most ignored, yet, an indispensable group of fishes in inland freshwaters worldwide. The disentangling of direct man-made threat from any climate-induced threat on the rich minnow population of South Asian inland waters was done for the first time. We validated the high reproductive resilience of minnows facing climatic variability. The key to sustenance of the minnow population in regional inland waters depends on managing anthropogenic (indiscriminative fishing) threats, as the changing climate is even favoring recruitment. Further indiscriminative fishing pressure will most likely be the key to minnows’ endurance or survival in the coming decades. We recommend responsible compliance to minimum mesh size limits that would allow a sizeable portion of gravid females to breed with statistical certainty without being pre-maturely sieved out of water. We proposed a novel parameter, ‘pre-spawning girth’ - an applied parameter capable of checking anthropogenic (indiscriminative fishing) threats and giving responsible ‘minimum mesh size’ recommendations.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
Authors are thankful to the Director, ICAR-Central Inland Fisheries Research Institute, Barrackpore. The financial help of Indian Council of Agricultural Research, New Delhi (ICAR) for funding in the project National Innovations in Climate Resilient Agriculture (NICRA) is also gratefully acknowledged for this study. The second author separately acknowledges funds from CENAKVA (LM2018099), Biodiversity (CZ.02.1.01/0.0/0.0/16_025/0007370) for supporting him during manuscript preparation. The authors gratefully acknowledge the help of anonymous reviewers for improving this article.
References
Afroze, S., Hossain, M.A., 1990. The reproductive cycle of the freshwater fish Amblypharyngodon mola (Ham.) (Cypriniformes: Cyprinidae). Univ. J. Zool. Rajshahi Univ. 9, 17–21.
Ainsley, S.M., Ebert, D.A., Cailliet, G.M., 2011. Age, growth, and maturity of the whitebrow skate, Bathyraja minispinosa, from the eastern Bering Sea. ICES J. Mar. Sci. 68, 1426–1434. https://doi.org/10.1093/icesjms/fsr072.
Alam, M.T., Hussain, M.A., Sultana, S., Hasan, M.T., Mazlan, A.G., Simon, K.D., Mazumder, S.K., 2018. Population growth and reproductive potential of five important fishes from the freshwater bodies of Bangladesh. Iran. J. Fish. Sci. 17, 657–674. https://doi.org/10.22092/ijfs.2018.116824.
APHA, 2012. Standard methods for examination of water and waste water, 22nd edn. American Public Health Association, Washington, DC. Azadi, M.A., Mamun, A., 2004. Reproductive biology of the Cyprinid, Amblypharyngodon mola (Hamilton) from the Kaptai Reservoir, Bangladesh. Pakistan J. Biol.
Sci. 7, 1727–1729. https://doi.org/10.3923/pjbs.2004.1727.1729. Baishya, A., Dutta, A., Bordoloi, S., 2010. Morphometry and length-weight relationship of Amblypharyngodon mola (Hamilton-Buchanan, 1822). Indian J. Fish 57,
87–91. Bogard, Jessica R., Hother, Anne-Louise, Saha, Manika, Bose, Sanjoy, Kabir, Humayun, Marks, Geoffrey C., Thilsted, Shakuntala Haraksingh, 2015. Inclusion of small
indigenous fish improves nutritional quality during the first 1000 days. Food Nutr. Bull. 36 (3), 276–289. https://doi.org/10.1177/0379572115598885.
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
12
Chaudhry, S., 2010. Amblypharyngodon mola. The IUCN Red List of Threatened Species 2010: e.T166521A6227212. DOI:10.2305/IUCN.UK.2010-4.RLTS. T166521A6227212.en. Downloaded on 20 November 2019.
Conover, David O., Munch, Stephan B., Arnott, Stephen A., 2009. Reversal of evolutionary downsizing caused by selective harvest of large fish. Proc. R. Soc. B Biol. Sci. 276 (1664), 2015–2020. https://doi.org/10.1098/rspb.2009.0003.
Das, R.K., Barat, S., 2014. Fishing gears operated in lentic and lotic water bodies of Cooch Behar District, West Bengal, India. Indian J. Traditional knowledge 13, 619–625.
Das Sarkar, S., Sarkar, U.K., Lianthuamluaia, L., Ghosh, B. Das, Roy, K., Mishal, P., Das, B.K., 2020. Pattern of the state of eutrophication in the floodplain wetlands of eastern India in context of climate change: a comparative evaluation of 27 wetlands. Environ. Monit. Assess. 192, 183. https://doi.org/10.1007/s10661-020- 8114-8.
de Roos, Andre M, Boukal, David S, Persson, Lennart, 2006. Evolutionary regime shifts in age and size at maturation of exploited fish stocks. Proc. R. Soc. B Biol. Sci. 273 (1596), 1873–1880. https://doi.org/10.1098/rspb.2006.3518.
Edeline, E., Carlson, S.M., Stige, L.C., Winfield, I.J., Fletcher, J.M., James, J. Ben, Haugen, T.O., Vollestad, L.A., Stenseth, N.C., 2007. Trait changes in a harvested population are driven by a dynamic tug-of-war between natural and harvest selection. Proc. Natl. Acad. Sci. 104, 15799–15804. DOI:10.1073/pnas.0705908104.
FAO, 2018. The State of World Fisheries and Aquaculture 2018 - Meeting the sustainable development goals. Rome. Licence: CC BY-NC-SA 3.0 IGO. Ferdous Ahamed, M., Rasel, N.S., Motia Gulshan Ara, Z., Ahmed, F., Hossain, M.Y., 2017. Biological aspects of the mola carplet Amblypharyngodon mola (Cyprinidae)
in the Payra River, Southern Bangladesh. Int. J. Fish. Aquat. Stud. 5, 336–339. Forestier, Romain, Blanchard, Julia L., Nash, Kirsty L., Fulton, Elizabeth A., Johnson, Craig, Audzijonyte, Asta, 2020. Interacting forces of predation and fishing affect
species’ maturation size. Ecol. Evol. 10 (24), 14033–14051. https://doi.org/10.1002/ece3.v10.2410.1002/ece3.6995. Fukuwaka, M., Morita, K., 2008. Increase in maturation size after the closure of a high seas gillnet fishery on hatchery-reared chum salmon Oncorhynchus keta. Evol.
Appl. 1, 376–387. https://doi.org/10.1111/j.1752-4571.2008.00029.x. Gupta, S., Banerjee, S., 2015. Length–weight relationship of Amblypharyngodon mola (Ham.-Buch., 1822), a freshwater cyprinid fish from West Bengal, India. Zool.
Ecol. 1–5. DOI:10.1080/21658005.2014.991513. Gupta, S., Banerjee, S., 2013. Studies on some aspects of Reproductive biology of Amblypharyngodon mola. Int. Res. J. Biol. Sci. 2, 69–77. Gupta, S., Banerjee, S., 2014. Feeding and breeding biology of Amblypharyngodon mola–A Review. Int. J. Aquat. Biol. 2, 85–90. Hoque, A.S.M.M., Rahman, M.R., 2008. Reproductive ecology of Mola (Amblypharyngodon mola). J. Agric. Rural Dev. 6, 165–174. Hossain, M.A., 2021. Some Aspects of Reproduction in Amblypharyngodon mola from Sylhet, Northeast Bangladesh. Aquat. Sci. Eng. 36, 140–145. DOI:10.26650/
ASE2020858534. ICES, International Council for the Exploration of the Sea., 2004. Mesh Size Measurement Revisited. ICES Cooperative Research Report, No. 266. 56pp. ICSF, 2010. Workshop report. In: Kumar, K.G., (Ed.), Workshop on Small Indigenous Freshwater Fish Species: Their Role in Poverty Alleviation, Food Security and
Conservation of Biodiversity. Organized by Central Inland Fisheries Research Institute (CIFRI), Barrackpore and International Collective in Support of Fishworkers (ICSF). Available online at http://aquaticcommons.org/4136/1/SIFFS_report.pdf.
Jørgensen, Christian, Ernande, Bruno, Fiksen, Øyvind, Dieckmann, Ulf, 2006. The logic of skipped spawning in fish. Can. J. Fish. Aquat. Sci. 63 (1), 200–211. https:// doi.org/10.1139/f05-210.
Joshi, K.D., Das, S.C.S., Pathak, R.K., Khan, A., Sarkar, U.K., Roy, K., 2018. Pattern of reproductive biology of the endangered golden mahseer Tor putitora (Hamilton 1822) with special reference to regional climate change implications on breeding phenology from lesser Himalayan region. India. J. Appl. Anim. Res. 46, 1289–1295. https://doi.org/10.1080/09712119.2018.1497493.
Karjalainen, J., Urpanen, O., Keskinen, T., Huuskonen, H., Sarvala, J., Valkeajarvi, P., Marjomaki, T.J., 2016. Phenotypic plasticity in growth and fecundity induced by strong population fluctuations affects reproductive traits of female fish. Ecol. Evol. 6, 779–790. https://doi.org/10.1002/ece3.1936.
Karnatak, Gunjan, Sarkar, Uttam Kumar, Naskar, Malay, Roy, Koushik, Gupta, Sandipan, Nandy, Saurav Kumar, Srivastava, Pankaj Kumar, Sarkar, Soma Das, Sudheesan, Deepa, Bose, Arun Kumar, Verma, Vinod Kumar, 2018. Understanding the role of climatic and environmental variables in gonadal maturation and spawning periodicity of spotted snakehead, Channa punctata (Bloch, 1793) in a tropical floodplain wetland, India. Environ. Biol. Fishes 101 (4), 595–607. https://doi.org/10.1007/s10641-018-0722-6.
Karnatak, Gunjan, Sarkar, Uttam Kumar, Naskar, Malay, Roy, Koushik, Nandi, Saurav, Mishal, Puthiyottil, Lianthuamluaia, Lianthuamluaia, Kumari, Suman, Das, Basanta Kumar, 2020. Modeling pre-spawning fitness and optimal climate of spotted snakehead Channa punctata (Bloch, 1793) from a Gangetic floodplain wetland of West Bengal, India. Int. J. Biometeorol. 64 (11), 1889–1898. https://doi.org/10.1007/s00484-020-01976-z.
Kelkar, Nachiket, Dey, Subhasis, 2020. Mesh mash: legal fishing nets cause most bycatch mortality of endangered South Asian river dolphins. Biol. Conserv. 252, 108844. https://doi.org/10.1016/j.biocon.2020.108844.
Kohinoor, A.H.M., Islam, M.S., Thilsted, S.H., Wahab, M.A., 2005. Reproductive biology of three important indigenous small fish viz., mola (Amblypharyngodon mola), chela (Chela cachius) and punti (Puntius sophore). Iranian J. Fish. Sci. 5, 29–48.
Krabbenhoft, T.J., Myers, B.J.E., Wong, J.P., Chu, C., Tingley, R.W., Falke, J.A., Kwak, T.J., Paukert, C.P., Lynch, A.J., 2020. FiCli, the Fish and Climate Change Database, informs climate adaptation and management for freshwater fishes. Sci. Data 7, 124. https://doi.org/10.1038/s41597-020-0465-z.
Kuparinen, Anna, Cano, Jose M., Loehr, John, Herczeg, Gabor, Gonda, Abigel, Merila, Juha, 2011. Fish age at maturation is influenced by temperature independently of growth. Oecologia 167 (2), 435–443. https://doi.org/10.1007/s00442-011-1989-x.
Mandal, F.B., Nandi, N.C., 2015. Loss of Small Indigenous Fish Species in India: A Case of Concern. J. Environ. Sociobiol. 12, 35–45. Mohanty, B.P., Behera, B.K., Sharma, A.P., 2011. Nutritional significance of small in-digenousfishes in human health. Bulletin No 162. CIFRI, Barrackpore. http://
www.cifri.res.in/Bulletins/Bulletin%20No.185.pdf. Mondal, D.K., Kaviraj, A., 2010. Feeding and reproductive biology of Indian shad Gudusia chapra in two floodplain lakes of India. Electron J Biol 6, 98–102. Mondal, D.K., Kaviraj, A., 2013. Feeding and reproductive biology of Amblypharyngodon mola (Cyrpiniformes: Cyprinidae) from two floodplain lakes of India. Int. J.
Aquat. Biol. 1 (3), 125–131. Mondal, S., Wahab, A., Kumar Barm, B., Al-Asif, A., 2020. Breeding Biology of Mola Carplet, (Amblypharyngodon mola, Hamilton, 1822) in Semi-Natural Condition.
Asian J. Anim. Sci. 14, 111–120. https://doi.org/10.3923/ajas.2020.111.120. Mustafa, G., 1991. Composite Culture and Biology of Some Indigenous Fishes of Bangladesh. Unpublished [PhD. Thesis]. Faculty of Biological Science, Dhaka
University, p. 299. Myszkowski, L., 2013. Compensatory growth, condition and food utilization in barbel Barbus barbus juveniles reared at different feeding periodicities with a dry diet.
J. Fish Biol. 82, 347–353. https://doi.org/10.1111/j.1095-8649.2012.03482.x. Naskar, Malay, Roy, Koushik, Karnatak, Gunjan, Nandy, Saurav Kumar, Roy, Aparna, 2018. Quantifying climate change induced threats to wetland fisheries: a
stakeholder-driven approach. Environ. Dev. Sustain. 20 (6), 2811–2830. https://doi.org/10.1007/s10668-017-0018-6. Nordhagen, A., Rizwan, A.A.M., Aakre, I., Moxness Reksten, A., Pincus, L.M., Bøkevoll, A., Mamun, A., Haraksingh Thilsted, S., Htut, T., Somasundaram, T.,
Kjellevold, M., 2020. Nutrient Composition of Demersal, Pelagic, and Mesopelagic Fish Species Sampled Off the Coast of Bangladesh and Their Potential Contribution to Food and Nutrition Security—The EAF-Nansen Programme. Foods 9, 730. https://doi.org/10.3390/foods9060730.
Olsen, E.M., Heino, M., Lilly, G.R., Morgan, M.J., Brattey, J., Ernande, B., Dieckmann, U., 2004. Maturation trends indicative of rapid evolution preceded the collapse of northern cod. Nature 428, 932–935. https://doi.org/10.1038/nature02430.
Otero, Jaime, Jensen, Arne J., L’Abee-Lund, Jan Henning, Stenseth, Nils Chr., Storvik, Geir O., Vøllestad, Leif Asbjørn, 2012. Contemporary ocean warming and freshwater conditions are related to later sea age at maturity in Atlantic salmon spawning in Norwegian rivers. Ecol. Evol. 2 (9), 2192–2203. https://doi.org/ 10.1002/ece3.2012.2.issue-910.1002/ece3.337.
Pal, S., Mahapatra, B.K., 2018. Biology of Indigenous Ornamental fish, Amblypharyngodon mola (Hamilton-Buchanan, 1822). In: Mahapatra, B.K., Roy, A.K., Pramanik, N.C. (Eds.), Sustainable Management of Aquatic Resources, 329-351p.
Parveen, S., 1984. Studies on the Culture Methods and Some Aspects of the Biology of Amblypharyngodon Mola. M.Sc Thesis. Department of Zoology, Rajshahi University, p. 137.
U. Kumar Sarkar et al.

Climate Risk Management 33 (2021) 100325
13
Rahman, A.K.A., 1989. Freshwater Fishes of Bangladesh. The Zoological Society of Bangladesh, Department of Zoology, University of Dhaka, Dhaka, Bangladesh. Rahman, Md. Mosaddequr, Hossain, Md. Yeamin, Tumpa, Anannaya Sen, Hossain, Md. Istiaque, Billah, Md. Masum, Ohtomi, Jun, 2018. Size at sexual maturity and
fecundity of the mola carplet Amblypharyngodon mola (Hamilton 1822) (Cyprinidae) in the Ganges River, Bangladesh. Zool. Ecol. 28 (4), 429–436. https://doi. org/10.1080/21658005.2018.1537906.
Rijnsdorp, A., Peck, M., Engelhard, G., Mollmann, C., Pinnegar, J., 2010. Resolving climate impacts on fish stocks. ICES Cooperative Research Report No. 301. 371 pp. Rojas-Downing, M.M., Nejadhashemi, A.P., Harrigan, T., Woznicki, S.A., 2017. Climate change and livestock: impacts, adaptation, and mitigation. Clim. Risk Manag.
16, 145–163. https://doi.org/10.1016/j.crm.2017.02.001. Rolighed, J., Jeppesen, E., Søndergaard, M., Bjerring, R., Janse, J., Mooij, W., Trolle, D., 2016. Climate Change Will Make Recovery from Eutrophication More
Difficult in Shallow Danish Lake Søbygaard. Water 8, 459. https://doi.org/10.3390/w8100459. Saha, B.K., Islam, M.R., Saha, A., Hossain, M.A., 2009. Reproductive Biology of the Mola Carplet Amblypharyngodon mola (Hamilton)(Cypriniformes: Cyprinidae)
from Netrakona Water. Bangladesh J. Sci. Ind. Res. 44, 377–379. Sandhya, K.M., Hassan, M.A., Kumari, S., Mishal, P., Lianthuamluaia, L., Kumar, V., Aftabuddin, M., Meena, D.K., 2016. Length-weight relationships of four
indigenous freshwater fish species from Khalsi wetland in lower Ganga basin, West Bengal, India. J. Appl. Ichthyol. 32, 505-506. DOI:10.1111/jai.13005. Sandhya, K.M., Roy, A., Hassan, M.A., Kumari, S., Mishal, P., Kumar, L.L.V., Aftabuddin, M., Bhattacharjya, B.K., Meena, D.K., Ali, Y., Naskar, B., 2019. Traditional
Fishing Gears, Fish Catch and Species Composition of Selected Floodplain Wetlands of Lower Gangetic Plains, West Bengal, India. Fish. Technol. 56, 101–109. Sarkar, U.K., Roy, K., Karnatak, G., Nandy, S.K., 2018b. Adaptive climate change resilient indigenous fisheries strategies in the floodplain wetlands of West Bengal,
India. J. Water Clim. Chang. jwc2018271. DOI:10.2166/wcc.2018.271. Sarkar, U.K., Roy, K., Karnatak, G., Naskar, M., Puthiyottil, M., Baksi, S., Lianthuamluaia, L., Kumari, S., Ghosh, B. Das, Das, B.K., 2021b. Reproductive environment of
the decreasing Indian river shad in Asian inland waters: disentangling the climate change and indiscriminative fishing threats. Environ. Sci. Pollut. Res. DOI: 10.1007/s11356-021-12852-7.
Sarkar, U.K., Lakra, W.S., 2010. Small indigenous freshwater fish species of India: significance, conservation and utilization. Aquacult. Asia 15, 34–35. Sarkar, U.K., Naskar, M., Roy, K., Sudeeshan, D., Srivastava, P., Gupta, S., Bose, A.K., Verma, V.K., Sarkar, S. Das, Karnatak, G., Nandy, S.K., 2017. Benchmarking pre-
spawning fitness, climate preferendum of some catfishes from river Ganga and its proposed utility in climate research. Environ. Monit. Assess. 189, 491. https:// doi.org/10.1007/s10661-017-6201-2.
Sarkar, U.K., Naskar, M., Roy, K., Sudheesan, D., Gupta, S., Bose, A.K., Srivastava, P.K., Nandy, S.K., Verma, V.K., Sarkar, S. Das, Karnatak, G., 2018a. Baseline information of reproduction parameters of an amphidromous croaker Johnius coitor (Hamilton, 1822) from Ganga river basin, India with special reference to potential influence of climatic variability. Aquat. Living Resour. 31, 4. https://doi.org/10.1051/alr/2017042.
Sarkar, Uttam Kumar, Naskar, Malay, Srivastava, Pankaj Kumar, Roy, Koushik, Das Sarkar, Soma, Gupta, Sandipan, Bose, Arun Kumar, Nandy, Saurav Kumar, Verma, Vinod Kumar, Sudheesan, Deepa, Karnatak, Gunjan, 2019a. Climato-environmental influence on breeding phenology of native catfishes in River Ganga and modeling species response to climatic variability for their conservation. Int. J. Biometeorol. 63 (8), 991–1004. https://doi.org/10.1007/s00484-019-01703-3.
Sarkar, U.K., Roy, K., Naskar, M., Srivastava, P.K., Bose, A.K., Verma, V.K., Gupta, S., Nandy, S.K., Sarkar, S. Das, Karnatak, G., Sudheesan, D., Das, B.K., 2019b. Minnows may be more reproductively resilient to climatic variability than anticipated: synthesis from a reproductive vulnerability assessment of Gangetic pool barbs (Puntius sophore). Ecol. Indic. 105, 727–736. https://doi.org/10.1016/j.ecolind.2019.03.037.
Sarkar, U.K., Mishal, P., Borah, S., Karnatak, G., Chandra, G., Kumari, S., Meena, D.K., Debnath, D., Yengkokpam, S., Das, P., DebRoy, P., Yadav, A.K., Aftabuddin, Md., Gogoi, P., Pandit, A., Bhattacharjya, B.K., Tayung, T., Lianthuamluai, L., Das, B.K., 2021a. Status, potential, prospects, and issues of floodplain wetland fisheries in India: synthesis and review for sustainable management. Rev. Fish Sci. Aquacult. 29 (1), 1–32. https://doi.org/10.1080/ 23308249.2020.1779650.
Suresh, V.R., Biswas, B.K., Vinci, G.K., Mitra, K., Mukherjee, A., 2007. Biology of Amblypharyngodon mola (Hamilton) from a floodplain wetland, West Bengal. Indian J. Fish. 54, 155–161.
Thomas, S.N., 2019. Sustainable Gillnet Fishing. In: ICAR Winter School: Responsible Fishing: Recent Advances in Resource and Energy Conservation 21 November – 11 December 2019. ICAR-CIFT, Kochi, pp. 239–250.
Trippel, Edward A., 1995. Age at maturity as a stress indicator in fisheries. Bioscience 45 (11), 759–771. https://doi.org/10.2307/1312628. Verberk, Wilco C.E.P., Atkinson, David, Hoefnagel, K. Natan, Hirst, Andrew G., Horne, Curtis R., Siepel, Henk, 2021. Shrinking body sizes in response to warming:
explanations for the temperature–size rule with special emphasis on the role of oxygen. Biol. Rev. 96 (1), 247–268. https://doi.org/10.1111/brv.v96.110.1111/ brv.12653.
U. Kumar Sarkar et al.




















