
Histone Deacetylase 5 Epigenetically Controls Behavioral Adaptations to Chronic Emotional Stimuli

Histone deacetylase inhibitor trichostatin A affectsporcine oocyte maturation in vitro
Yong-Xun JinA, Ming-Hui ZhaoA, Zhong ZhengA,B, Jung-Suk KwonA,Seul-Ki LeeA, Xiang-Shun CuiA,C and Nam-Hyung KimA,C
ADepartment of Animal Sciences, Chungbuk National University, Cheong-Ju,
361-763, South Korea.BCollege of Veterinary Medicine, Chungbuk National University, Cheong-Ju,
361-763 South Korea.CCorresponding authors. Emails: [email protected]; [email protected]
Abstract. Previous studies show that porcine oocyte aging resulting from asynchronised IVM impairs embryodevelopmental competence. In the present study we investigated whether trichostatin A (TSA; an inhibitor of histone
deacetylation) prolongs the maturation time and prevents the aging of oocytes. Porcine oocytes were cultured in mediumcontaining increasing concentrations of TSA (300 nM) for 24, 44 or 64 h. The percentage of oocytes that underwentgerminal vesicle breakdownwas significantly lower in the TSA-treated group (300 nM) than in the control group. TSA did
not affect oocyte quality at MII based on levels of maturation-promoting factor, the phosphorylation status of mitogen-activated protein kinase or histone H3K9 acetylation analysis. We also compared the preimplantation developmentalcompetence and the viability of pathenogenetic embryos treated with 100 nMTSA for 24 h and then continuously cultured
for another 24 h in TSA free condition. No significant differences were observed for either parameter between the TSA-treated and control groups. These results indicate that TSA prolongs the IVMof porcine oocytes but that oocyte quality andaging are not affected. These findings provide a feasible option by which to adjust the initiation time of downstreamexperiments based on porcine matured oocytes.
Additional keywords: acetylation, histone H3K9, oocyte aging, p-MAPK, prolong.
Received 15 January 2013, accepted 13 May 2013, published online 13 June 2013
Introduction
Porcine oocytes obtained from pro-oestrous ovaries have com-pleted the first round of meiosis and are arrested at the germinal
vesicle (GV) stage, which is evidenced by the presence of alarge nucleus. Meiosis resumes in response to species-specificstimuli, and meiotic progression is signified by GV breakdown
(GVBD). Thereafter, meiosis arrests at MII until fertilisation(Voronina and Wessel 2003; Richard 2007). Unfortunately, it isdifficult to synchronise porcine oocytes during IVM, a processdefined as oocyte aging, and misregulated IVM can result in the
formation of poor-quality oocytes that are often unsynchronised(Wilcox et al. 1998; Liu andKeefe 2002). If fertilisation does notoccurwithin a specific time limit, unfertilised oocytes remaining
in the oviduct (in vivo aging) or in culture (in vitro aging)deteriorate progressively with time. This process is defined asoocyte aging (Sun et al. 2012). Considering that the optimal time
for oocyte fertilisation is,24 h (Mailhes et al. 1998;Gordo et al.2002), improved conditions for oocyte IVM are needed.
Histone deacetylation plays a crucial role in inhibiting oocyte
maturation and meiosis (Endo et al. 2005; Akiyama et al. 2006;Ivanovska and Orr-Weaver 2006; Wang et al. 2006a, 2006b).A previous study has shown that immature oocytes are associated
with high levels of histone acetylation and histone deacetylation(HDAC). By anaphase I (AI) and telophase I (TI), high levels ofhistone acetylation are once again evident, followedby high levels
of histone deacetylation at MII. Equally important, inhibition ofHDAC during meiosis can result in oocyte aneuploidy(an abnormal number of chromosomes) and subsequent embryo
death, indicating that HDAC is crucial for normal chromosomefunction during the oocyte cycle (Wang et al. 2006a).
Selective HDAC inhibitors (HDACi) such as trichostatin A(TSA) affect histone acetylation. Previous studies have shown
TSA to affect chromosomal segregation in different cell types,indicating that TSA-induced hyperacetylation can disrupt peri-centromeric heterochromatin and affect interactions between
kinetochores and spindle microtubules (Kuroda et al. 2004;Sugiura et al. 2006). Although HDACi can result in histonehyperacetylation, IVM in porcine oocytes is unaffected. Con-
versely, IVM in mouse oocytes was delayed by TSA (Gottlicheret al. 2001; Kramer et al. 2003; De La Fuente et al. 2004).
Presently, there is little information available on the role
of histone acetylation during the cycling and maturation ofporcine oocytes. Thus, the objective of the present in vitro studywas to investigate whether TSA-induced histone acetylation
CSIRO PUBLISHING
Reproduction, Fertility and Development, 2014, 26, 806–816
http://dx.doi.org/10.1071/RD13013
Journal compilation � CSIRO 2014 www.publish.csiro.au/journals/rfd

regulates the time required for porcine oocytes to transit fromGVBD to MII.
Materials and methods
Oocyte collection and IVM
Prepubertal porcine ovaries were obtained from a localslaughterhouse. Cumulus-oocyte complexes (COCs) were iso-lated and cultured in tissue culturemedium (TCM)-199 (GIBCO
BRL, Grand Island, NY, USA) supplemented with 0.57mMcysteine, 10 ngmL�1 epidermal growth factor (EGF), 10 IUmL�1
pregnant mare’s serum gonadotropin (PMSG) and 10 IUmL�1
human chorionic gonadotropin (hCG) under light mineral oil at38.58C and 5% CO2 (v/v). Thereafter, oocytes were collected atdifferent stages of maturity.
Parthenogenic activation and culture of embryos
Upon maturation, cumulus cells were removed by repeatedpipetting in the presence of 1mgmL�1 hyaluronidase for
2–3min. Oocytes were parthenogenetically activated with thecalcium ionophore A23187 (50mM) for 5min, followed byincubation in PZM-5 medium containing 7.5 mgmL�1 cyto-chalasin B (CB; Sigma-Aldrich, St Louis, MO, USA) for 3 h.
Embryos were cultured in PZM-5 medium supplemented with0.4% bovine serum albumin (BSA; w/v) under light mineral oilfor 7 days at 38.58C and 5% CO2 (v/v) and then harvested.
To assess pronuclear formation, embryos were activated for10 h, fixed in 4% paraformaldehyde (w/v) and stained withHoechst 33342 (4 mgmL�1) for 5min. Embryos were then
washed with polyvinyl alcohol (PVA)–phosphate-bufferedsaline (PBS), mounted and imaged by fluorescence microscopy(Eclipse Ti-U; Nikon, Tokyo, Japan). Oocytes with one nucleus
but unequal 2–4-cell-like cytoplasmic morphologies weredefined as fragmented oocytes.
Immunofluorescence analysis and fluorescenceintensity analysis
Oocytes were washed with PVA-PBS three times and fixed in4% paraformaldehyde (w/v) at room temperature for 30min.
Thereafter, oocytes were treated with 0.1% Trixon-100 at 378Cand blocked with 3% BSA (w/v) for 1 h each. Oocytes wereincubated with anti-histone H3 acetyl lysine 9 (H3-aceK9;Abcam, Cambridge, UK) for 1 h at room temperature, followed
by three washes with PBS-PVA. Oocytes were then incubatedwith Alexa Fluor 488 goat anti-rabbit antibody (Invitrogen,Carlsbad, CA, USA) for 1 h, washed with PVA-PBS for three
times andmountedwith 40,6-diamidino-2-phenylindole (DAPI).Microscopy was performed using an inverted confocal micro-scope with a �40 oil immersion objective (Zeiss, Oberkochen,
Germany). Images were processed with ZEN software (ZeissMicrosystems, Oberkochen, Germany).
Real-time reverse transcription–polymerase chainreaction analysis of apoptosis-related genes
Expression of the apoptosis-related genes Bcl-xL and Caspase 3were analysed by real-time reverse transcription–polymerasechain reaction (RT-PCR). Total RNA was extracted from
blastocysts (n¼ 15) with a Dynabead mRNA DIRECT kit(Invitrogen Dynal, Oslo, Norway). First-strand total cDNAs
were synthesised with a cDNA synthesis kit (Takara, Dalian,China) using an oligo(dT)12–18 primer (Invitrogen). The pri-mers used for the PCR experiments were as follows: Gapdh
(housekeeping gene), 50-GGGCATGAACCATGAGAAGT-30
(sense) and 50-AAGCAGGGATGATGTTCTGG-30 (anti-sense); Bcl-xL, 50-GGAGCTGGTGGTTG ACTTTC-30 (sense)and 50-CTAGGTGGTCATTCAGGTAAGG-30 (antisense); andCaspase 3, 50-GAGGCAGACTTCTTGTATGC-30 (sense) and50-CATGGACACAATACATGGAA-30 (antisense).
Western blot analysis
The protocol for western blot analysis in the present studybasically followed that described previously (Jin et al. 2009).Briefly, oocytes (30 oocytes per sample) were solubilised in
20 mL of 1� sodium dodecyl sulfate (SDS) sample buffer(62.5mM Tris-HCl (pH 6.8 at 258C), 2% w/v SDS, 10% glyc-erol, 50mM dithiothreitol (DTT) and 0.01% w/v bromophenol
blue or phenol red) and heated for 5min at 958C. For westernblotting, proteins were resolved on a 5%–12% Tris–SDS–polyacrylamide gel electrophoresis (PAGE) gel for 1.5 h at80–100V. Samples were then transferred to nitrocellulose
membranes (Hybond-ECL; Amersham, Buckinghamshire, UK)at 300mA for 2 h in transfer buffer (25mM Tris base, 200mMglycine, 20% methanol, pH 8.5). After blocking with 5% skim
milk in PBS for 1 h, membranes were incubated for at least 2 hwith anti-phosphorylated (p-) p44/42 mitogen-activated proteinkinase (MAPK) antibodies (Cell Signaling Technology,
Beverly, MA, USA) diluted 1 : 500 in blocking solution (1�Tris-buffered saline (TBS), 0.1%Tween-20, 5%w/v non-fat drymilk), washed three times in TBST (20mM Tris-HCl, pH 7.5,250mM NaCl, 0.1% Tween-20) and incubated for 1 h with
horseradish peroxidise (HRP)-linked anti-rabbit IgG (Cell Sig-naling Technology) diluted 1 : 2000 in blocking solution. Afterthree washes with TBST, antibody binding was visualised using
a Chemiluminescence Luminol Reagent (Invitrogen). Proteinbands were analysed and the integrated optical density (OD)of each band was determined with Image-Pro Plus software
(version 6.0; Media Cybernetics, Springfield, MA, USA).
p34cdc2 kinase activity
p34cdc2 kinase activity was quantified using the Mesacup cdc2kinase assay (MBL, Nagoya, Japan), according to the methods
described by Shojo et al. (2000). Using this method, thecorrelation coefficient between p34cdc2 kinase activity (asdetermined by the cdc2 kinase assay) and histone H1 kinase
activity (as measured by radioactive detection) can be as highas 0.9961.
Briefly, 5mL oocyte extract (containing 20 oocytes) was
mixed with 45 mL kinase assay buffer (25mM HEPES, pH 7.5,containing 10mMMgCl2, 10%MVpeptide (SLYSSPGGAYC)solution (MBL, Nagoya, Japan) and 0.1mMATP). The mixture
was incubated for 30min at 308C. The reaction was terminatedby the addition of 200mL PBS containing 50mM EGTA.Phosphorylation of the MV peptide was detected by ELISA.The O.D. of p34cdc2 kinase activity in the control group was
TSA delays porcine oocyte maturation and aging Reproduction, Fertility and Development 807

arbitrarily set at 1, with the p34cdc2 kinase activity in theTSA-treated group expressed relative to that in the control
group. This experiment was repeated three times.
Terminal deoxynucleotidyl transferase dUTP nick-endlabelling
Parthenogenetically activated embryos were fixed and per-meabilised with 0.3% Triton X-100 (v/v) at room temperaturefor 1 h. After washing twice with PBS-PVA, embryos wereincubated with fluorescein-conjugated dUTP and terminal
deoxynucleotidyl transferase enzyme (In Situ Cell DeathDetection Kit; Roche, Mannheim, Germany) at 378C for 1 h inthe dark. After counterstaining with Hoechst 33342
(40 mgmL�1) and RNase A (4 mgmL�1) at 378C for 1 h to labelnuclei, embryos were washed with PBS-PVP, mounted andimaged by confocal microscopy. Excitation and emission
wavelengths for Hoechst 33342 were 543 and 585 nm, respec-tively; for terminal deoxynucleotidyl transferase dUTP nick-endlabelling (TUNEL), the excitation and emission wavelengths
were 488 and 515 nm, respectively. This experiment was per-formed at least three times.
Experimental design
Experiment 1 examined the effect of TSA on the reinitiation ofoocyte meiosis. Different TSA concentrations and incubation
times were tested. Specifically, to determine whether inhibitionof histone acetylation arrests oocytes at the GV stage andinduces synchronisation, oocytes were treated with 0, 5, 50, 100,
200 or 300 nM TSA. Nuclear morphology and lysine 9-acetylated histone H3 (acH3K9) activity were assessed 28, 44and 64 h after IVM (Fig. 1a).
Experiment 2 evaluated the effect of increasing concentra-
tions of TSA on oocyte maturation. Oocytes were incubatedwith 0, 5, 50, 100, 200 or 300 nMTSA for 24 h and then culturedin TSA-free medium for an additional 20 h to initiate maturation
(Fig. 2a). The phosphorylation status of MAPKwas determinedin oocytes after IVM.
Experiment 3 determined the effects of TSA on oocyte
quality and aging. Oocytes were incubated with 100 nM TSAfor 24 h to initiate maturation and then cultured in TSA-freemedium for an additional 20, 32 or 42 h (Fig. 3a). p34cdc2 kinase
activity and histone acH3K9were assessed in oocytes containingan extruded first polar body.
0 h 28 h
Exp 1: Effect of TSA treatment length on oocyte meiosis resumption.
Control
100con50 nM200 nM
5 nM100 nM
300 nM
80
60
40
20
0con
Mei
osis
res
umpt
ion
rate
(%
)
5 nM 50 nM 100 nM 200 nM 300 nM
TSA treatment group
MII-44 h
44 h 64 h
50GVBD-28 h
30
40
20
10
0con
Mei
osis
res
umpt
ion
rate
(%
)
5 nM 50 nM 100 nM 200 nM 300 nM
100
con
50 nM
200 nM
5 nM
100 nM
300 nM
80
60
40
20
0con
Mei
osis
res
umpt
ion
rate
(%
)
5 nM 50 nM 100 nM 200 nM 300 nM
MII-64 h
∗ ∗
∗
∗∗
(a)
(b)
(c) (d)
∗
∗∗
∗ ∗
Fig. 1. Trichostatin A (TSA) arrests porcine oocytes at germinal vesicle breakdown (GVBD). (a) Experimental Design 1. Oocytes at the germinal vesicle
(GV) stage were treated with 0, 5, 50, 100, 200 or 300 nMTSA. Themorphology of the nucleus and the level of lysine 9-acetylated histone H3 (acH3K9) at 28,
44 and 64 h were assessed. (b) The rate of meiotic reinitiation in oocytes at GVBD incubated with increasing concentrations of TSA for 28 h. (c, d ). The rate of
IVM in oocytes incubated with increasing concentrations of TSA for 44 (c) or 64 h (d ). Oocytes were visualised with Hoechst 33342. This experiment was
repeated at least three times and more than 30 oocytes were examined in each experimental group.
808 Reproduction, Fertility and Development Y.-X. Jin et al.

Experiment 4 investigated the effect of TSA on developmen-tal competency. Oocytes at MII were obtained from Experiment
3 and used for this experiment. Oocytes containing the first polarbody were parthenogenetically activated by electrical activa-tion. After CB treatment, oocytes were cultured in PZM-5
medium for 7 days, and developmental rate, total cell number,cell viability (i.e. apoptosis) and apoptotic gene expression wereinvestigated.
Data analysis
At least three replicates were performed for each treatment.Statistical analyses were conducted using analysis of variance(ANOVA) and differences between treatment groups were
determined by Duncan’s multiple comparison test. Data arepresented as the mean� s.e.m., and statistical significance wasset at P, 0.05.
Results
Time-dependent effect of TSA on the reinitiationof oocyte meiosis
High concentrations of TSA increased the synchronisation rate
of oocytes at GVBD 28 h after IVM (37.2� 2.1%, 43.2� 7.3%and 46.0� 5.9% for 100, 200 and 300 nM TSA, respectively)compared with the control (23.3� 9.6%) and low TSA con-
centration groups (28.1� 2.1% and 29.5� 1.9% for 5 and50 nM TSA, respectively; P, 0.05; Fig. 1b). High concentra-tions of TSA decreased the IVM rate (75.0� 5.4%, 58.5� 7.2%
and 62.9� 6.2% for 100, 200 and 300 nM TSA, respectively)44 h after IVMcomparedwith the control (93.5� 5.9%) and lowTSA concentration groups (90.3� 8.7% and 89.6� 7.7% for
5 and 50 nM TSA, respectively; P, 0.05; Fig. 1c). Afterprolonged treatment (i.e. 64 h), a higher rate of oocytes treatedwith 100 nM TSA reached MII than in the 200 and 300 nM
GV
DNA AcH3k9 Merg
GVBD
MI
MII
Control
28 h
44 h
64 h
100 nM TSA 200 nM TSA 300 nM TSA
2.0
1.5
1.0
0.5
028 h 64 h
con
200 nM 300 nM
100 nM
44 h
GVBD stageR
elat
ive
fluor
esce
nce
inte
nsity
(a) (b)
(c)
∗∗
∗∗ ∗
Fig. 2. Trichostatin A (TSA) affects histone deacetylation in porcine oocytes. (a) Porcine oocytes were cultured for 0, 28, 36 or 44 h, cumulus cells were
removed and oocytes were fixed and stained. GV, freshly isolated oocytes at the germinal vesicle (GV) stage; GVBD, 28 h cultured oocytes; MI, 36 h cultured
oocytes at first metaphase; MII, 44 h cultured oocytes at second metaphase. (b) Porcine oocytes were cultured for 28, 44 or 64 h in the absence (control) or
presence of 100, 200 or 300 nM TSA and immunostained with a monospecific lysine 9-acetylated histone H3 (acH3K9) antibody. (c) acH3K9 fluorescence
(green) in oocytes at GVBD. DNA (blue) was visualised with Hoechst 33342 (lower panels). This experiment was repeated at least three times and more than
30 oocytes were examined in each experimental group. Bar¼ 20mm.
TSA delays porcine oocyte maturation and aging Reproduction, Fertility and Development 809
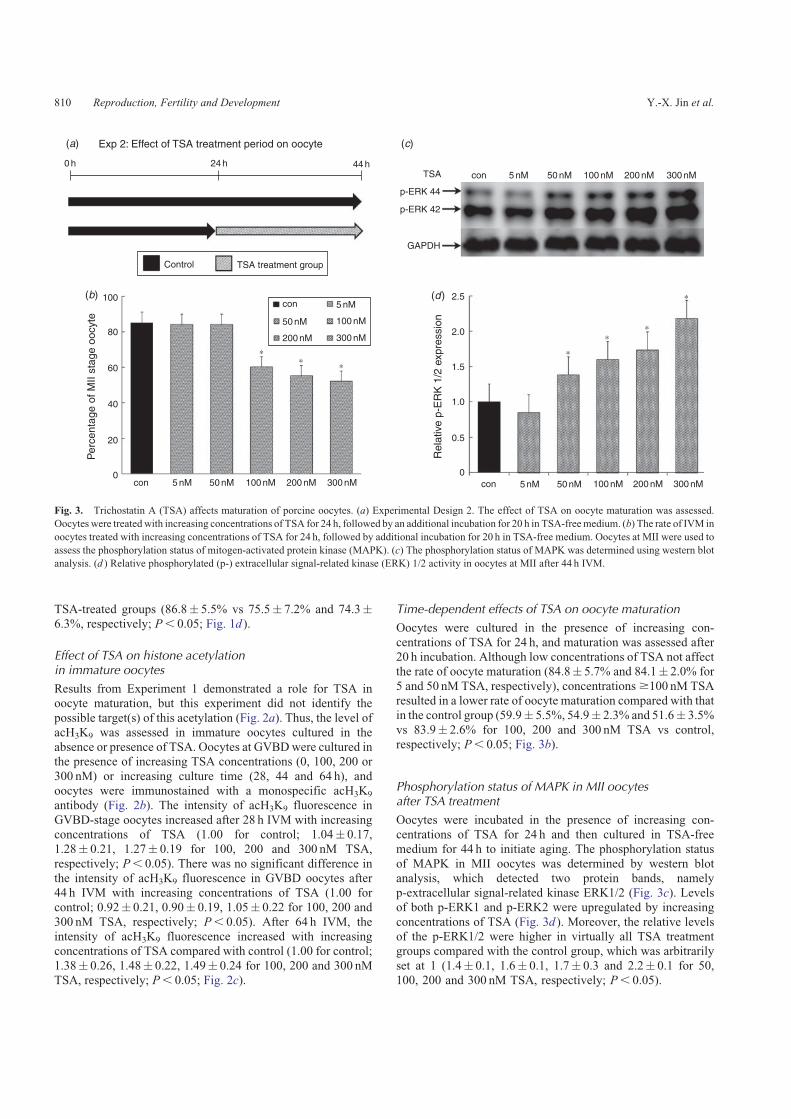
TSA-treated groups (86.8� 5.5% vs 75.5� 7.2% and 74.3�6.3%, respectively; P, 0.05; Fig. 1d ).
Effect of TSA on histone acetylationin immature oocytes
Results from Experiment 1 demonstrated a role for TSA inoocyte maturation, but this experiment did not identify the
possible target(s) of this acetylation (Fig. 2a). Thus, the level ofacH3K9 was assessed in immature oocytes cultured in theabsence or presence of TSA. Oocytes at GVBDwere cultured inthe presence of increasing TSA concentrations (0, 100, 200 or
300 nM) or increasing culture time (28, 44 and 64 h), andoocytes were immunostained with a monospecific acH3K9
antibody (Fig. 2b). The intensity of acH3K9 fluorescence in
GVBD-stage oocytes increased after 28 h IVM with increasingconcentrations of TSA (1.00 for control; 1.04� 0.17,1.28� 0.21, 1.27� 0.19 for 100, 200 and 300 nM TSA,
respectively; P, 0.05). There was no significant difference inthe intensity of acH3K9 fluorescence in GVBD oocytes after44 h IVM with increasing concentrations of TSA (1.00 forcontrol; 0.92� 0.21, 0.90� 0.19, 1.05� 0.22 for 100, 200 and
300 nM TSA, respectively; P, 0.05). After 64 h IVM, theintensity of acH3K9 fluorescence increased with increasingconcentrations of TSA compared with control (1.00 for control;
1.38� 0.26, 1.48� 0.22, 1.49� 0.24 for 100, 200 and 300 nMTSA, respectively; P, 0.05; Fig. 2c).
Time-dependent effects of TSA on oocyte maturation
Oocytes were cultured in the presence of increasing con-centrations of TSA for 24 h, and maturation was assessed after20 h incubation. Although low concentrations of TSA not affectthe rate of oocyte maturation (84.8� 5.7% and 84.1� 2.0% for
5 and 50 nM TSA, respectively), concentrations$100 nM TSAresulted in a lower rate of oocyte maturation compared with thatin the control group (59.9� 5.5%, 54.9� 2.3% and 51.6� 3.5%
vs 83.9� 2.6% for 100, 200 and 300 nM TSA vs control,respectively; P, 0.05; Fig. 3b).
Phosphorylation status of MAPK in MII oocytesafter TSA treatment
Oocytes were incubated in the presence of increasing con-centrations of TSA for 24 h and then cultured in TSA-freemedium for 44 h to initiate aging. The phosphorylation status
of MAPK in MII oocytes was determined by western blotanalysis, which detected two protein bands, namelyp-extracellular signal-related kinase ERK1/2 (Fig. 3c). Levelsof both p-ERK1 and p-ERK2 were upregulated by increasing
concentrations of TSA (Fig. 3d ). Moreover, the relative levelsof the p-ERK1/2 were higher in virtually all TSA treatmentgroups compared with the control group, which was arbitrarily
set at 1 (1.4� 0.1, 1.6� 0.1, 1.7� 0.3 and 2.2� 0.1 for 50,100, 200 and 300 nM TSA, respectively; P, 0.05).
100 2.5
2.0
1.5
1.0
0.5
0
Control TSA treatment group
con
80
60
40
20
0con 5 nM
0 h
(a)
(b) (d)
(c)
24 h
Exp 2: Effect of TSA treatment period on oocyte
44 hTSA
p-ERK 44
p-ERK 42
GAPDH
Per
cent
age
of M
II st
age
oocy
te
Rel
ativ
e p-
ER
K 1
/2 e
xpre
ssio
n
50 nM 100 nM 200 nM 300 nM
50 nM 100 nM
200 nM 300 nM
5 nM
con 5 nM 50 nM 100 nM 200 nM 300 nM
con 5 nM 50 nM 100 nM 200 nM 300 nM
∗∗ ∗
∗
∗∗
∗
Fig. 3. Trichostatin A (TSA) affects maturation of porcine oocytes. (a) Experimental Design 2. The effect of TSA on oocyte maturation was assessed.
Oocytes were treatedwith increasing concentrations of TSA for 24 h, followed by an additional incubation for 20 h in TSA-freemedium. (b) The rate of IVM in
oocytes treated with increasing concentrations of TSA for 24 h, followed by additional incubation for 20 h in TSA-free medium. Oocytes at MII were used to
assess the phosphorylation status of mitogen-activated protein kinase (MAPK). (c) The phosphorylation status of MAPK was determined using western blot
analysis. (d ) Relative phosphorylated (p-) extracellular signal-related kinase (ERK) 1/2 activity in oocytes at MII after 44 h IVM.
810 Reproduction, Fertility and Development Y.-X. Jin et al.

Effect of TSA on oocyte maturation and acetylationof histone H3K9
Results from Experiment 4 demonstrated that TSA affects thesynchronisation of oocytes at MII. To assess the maturation of
oocytes at MII, oocytes were treated with 100 nM TSA for 24 hand then cultured with or without TSA for 44, 56 or 64 h. Feweroocytes reached MII after 44 h IVM in the TSA versus without
TSA group (61� 3% vs 69� 3%, respectively; P, 0.05;Fig. 4b), with both groups having decreased rates comparedwiththe control group (88� 3%). Similarly, after 56 h IVM, both the
TSA andwithout TSA groups had fewer oocytes at theMII stagethan in the control group (72� 7% and 75� 5% vs 94� 7%,respectively; P, 0.05; Fig. 4b). However, there were no sig-
nificant differences in the rate at which oocytes reached MII
100
80
60
40
20
424 390 223
∗
∗
74 114 138 452 275 305
044 h 56 h
MII stage
64 h
44 h 56 h 64 h
44 h24 h0 h 56 h 64 h
Control TSA treatment group
44 h
con
TSA
1.2
0.8
0.4
0
56 h 64 h Per
cent
age
of M
II st
age
oocy
tes
(%)
Rel
ativ
e flu
ores
cenc
e in
tens
ity
Exp 3: Effect of TSA on oocyte aging and it’s quality
44 h 56 h 64 h
con TSA
con3
2
1
0
TSA
con TSA Full-TSA
MPF activity
Rel
ativ
e p3
4cdc2
act
ivity
∗∗
∗
∗
(a) (b)
(d)
(c)
(e)
Fig. 4. Trichostatin A (TSA) affects oocyte quality and aging. (a) Experimental Design 3. Oocytes were matured in the presence of 100 nM TSA for 24 h,
followed by an additional incubation for 20, 32 or 44 h in TSA-free medium, which initiated aging. p34cdc2 kinase activity and lysine 9-acetylated histone H3
(acH3K9) were assessed in oocytes containing an extruded first polar body. (b) Percentage of MII oocytes after 44, 56 or 64 h IVMwith or without TSA. Data
from at least three independent experiments are presented as mean percentages� s.e.m. *P, 0.05. (c) acH3K9 staining and (d ) quantification of fluorescence
intensity were used to detect oocytes at MII. DNA (blue) was visualised with Hoechst 33342 (lower panels). This experiment was repeated at least three times
andmore than 30 oocyteswere examined in each experimental group. Bar¼ 20mm. (e) Relative p34cdc2 kinase activity in aged oocytes cultured under different
conditions. After 44, 56 and 64 h IVM, oocytes containing a first polar body were used for parthenogenesis and subsequently cultured for 10 h. MPF,
maturation-promoting factor.
TSA delays porcine oocyte maturation and aging Reproduction, Fertility and Development 811

after 64 h IVMamong the control, without TSA and TSA groups
(88� 3%, 87� 3% and 85� 5%, respectively; P, 0.05;Fig. 4b). These data suggest TSA delays the IVM time, alsodelaying oocyte aging.
The acetylation of histone H3K9was assessed in TSA-treated
oocytes (Fig. 4c). TSA significantly increased acH3K9 fluores-cence in oocytes after 56 and 64 h IVM compared with thecontrol group. There was no significant difference in acH3K9
fluorescence at 44 h (Fig. 4d ). These results indicate that TSAaffects the maturation of aged oocytes.
We also quantified p34cdc2 kinase activity in without TSA
treated oocytes at MII. There were no significant differences inp34cdc2 kinase activity at 44, 56 or 64 h in either the control orwithout TSA treated groups (Fig. 4e).
Effect of TSA on the developmental capacity of oocytes
The effect of TSA on the developmental capacity of oocytes wasinvestigated. Oocytes were incubated with 100 nMTSA for 24 h
and then cultured in TSA-free medium for 44, 56 or 64 h.
The control group consisted of oocytes cultured in the absence
of TSA for 44, 56 or 64 h. All oocytes at MII were artificiallyactivated and subsequently cultured for 10 h and fixed. Afterfixation and staining with Hoechst 33342, pronuclear formationand fragmented oocytes were confirmed and categorised.
Activated oocytes are shown as a percentage of oocytes exam-ined in Fig. 5b, whereas fragmented oocytes as a percentage ofactivated oocytes are shown in Fig. 5c. In three independent
experiments, TSA did not significantly affect the rate of pro-nuclear formation in the TSA group (81� 3%, 93� 4% and89� 3% after 44, 56 and 64 h, respectively) compared with the
control group (94� 3%, 91� 3% and 80� 4% after 44, 56 and64 h, respectively; P, 0.05; Fig. 5b).
There was no difference in the rate of embryo fragmentationat 44 h between the TSA-treated and control groups (Fig. 5c);
however, the rate of embryo fragmentation at 56 and 64 h wassignificantly decreased in the TSA group (15.2� 0.3% and20.4� 0.3%, respectively) compared with the control group
(4.1� 0.5% and 6.3� 0.3%, respectively; P, 0.05).
44 h
PA
24 h0 h 64 h D 7
Exp 4: Effect of TSA on developmental capacity(a)
Control TSA treatment group
100
80
60
40
20
044 h 56 h 64 h
44 h 56 h 64 h
Per
cent
age
of p
ronu
clea
r fo
rmat
ion
(%)
(b)
con
TSA
con
TSA
25
20
15
10
5
0
Per
cent
age
of fr
agm
ente
d oo
cyte
(%
)
(c)
∗
∗
Fig. 5. Trichostatin A (TSA) affects pronuclear formation and cytoplasmic fragmentation. (a) Experimental Design 4. Oocytes at MII were obtained from
Experiment 3 and used for this experiment. Oocytes containing the first polar were parthenogenetically activated. After cytochalasin B (CB) treatment, oocytes
were cultured in PZM-5 medium for 10 h before being fixed and stained. Pronuclear formed and fragmented oocytes were categorised as activated. Activated
oocytes as a percentage of examined oocytes (b) and fragmented oocytes as a percentage of activated oocytes (c) are shown. Data from three independent
experiments are presented as mean percentages� s.e.m. *P, 0.05.
812 Reproduction, Fertility and Development Y.-X. Jin et al.

Developmental rate, total cell number and apoptosis werealso assessed in blastocysts. There was no significant change inthe rate of blastocyst formation after 44 h IVM (34� 3% and
37� 6% in the control and TSA groups, respectively;P, 0.05),
but there was an increase after 64 h of IVM in the TSA comparedwith control group (34� 4% vs 27� 4%, respectively;P, 0.05; Fig. 6a). There was no significant difference in the
total cell number in blastocysts in the control and TSA groups
50
40
30
20
5
4
3
2
1
0
1.2
1.0
0.8
0.6
0.4
0.2
0
1091 93
0
Per
cent
age
of b
last
ocyt
es (
%)
Tota
l cel
l num
ber
50
40
30
20
10
0
con TSA
44 h
44 h
64 h
64 h
44 h 64 h
con TSA
con TSA
con
Bcl-xL
TSAcon TSA
44 h
Apo
ptos
is
64 h
44 h
Fol
d ch
ange
in m
RN
A e
xpre
ssio
n
1.2
1.0
0.8
0.6
0.4
0.2
0
Fol
d ch
ange
in m
RN
A e
xpre
ssio
n
64 h 44 h 64 h
con 24 h-TSA
93 63
∗
∗
∗
(a) (b)
(c) (d )
(e) (f )Caspase 3
Fig. 6. Trichostatin A (TSA) affects developmental competency. Development rate, total cell number, apoptosis and apoptosis-related gene expression was
evaluated. (a) Percentage of oocytes at MII that developed into blastocysts in PZM-5 medium supplemented with TSA. (b) Total and (c) apoptotic
(no. apoptotic cells/total no. cells) cell numbers in blastocysts onDay 7. (d ) Apoptotic cells were detected by terminal deoxynucleotidyl transferase dUTPnick-
end labelling (TUNEL) staining. Yellow, TUNEL; red, chromatin. Bar¼ 20mm. (e, f )Bcl-xL and caspase 3mRNA expression in blastocyst embryos. Data are
the mean� s.e.m. Three replicate trials were performed. *P, 0.05.
TSA delays porcine oocyte maturation and aging Reproduction, Fertility and Development 813

after 44 h IVM (44� 5 vs 46� 3, respectively) and 64 h IVM(41� 3 vs 43� 2, respectively; P, 0.05; Fig. 6b). Apoptotic
cells in blastocysts were detected by TUNEL staining after TSAtreatment (Fig. 6c, d ). There was no significant difference in thedegree of apoptosis in blastocysts in the control and TSA groups
after 44 h IVM (4.0� 0.3 vs 3.7� 0.3 10 nM TSA vs control,respectively). However, the degree of apoptosis was reduced inthe TSA compared with control group after 64 h IVM (3.1� 0.3
vs 5.4� 0.3 10 nM TSA vs control, respectively; P, 0.05;Fig. 6c).
Real-time RT-PCR was used to quantify the mRNA expres-sion of Bcl-xL and Caspase 3 in blastocysts following TSA
treatment. Porcine Gapdhwas used as an internal control. Morethan 10 embryos were used per treatment group, and eachexperiment was repeated three times. After 44 or 64 h IVM,
there was no significant difference in Bcl-xL expression inblastocysts from the TSA and control groups (Fig. 6e). However,there was a significant decrease inCaspase 3mRNA expression
(a member of the pro-apoptotic gene family) after 64 h IVM inthe TSA compared with control group. There was no significantdifferences in Caspase 3 mRNA expression between the TSAand control groups after 44 h IVM (Fig. 6f ).
Discussion
cAMP is a key regulator of oocyte meiosis (Bornslaeger et al.
1986) because cAMP and its analogues synchronise develop-ment of the GV in porcine oocytes by inhibiting GVBD(Funahashi et al. 1997). In the present study, TSA treatment
prolonged the time required to reach oocyte maturation withoutaffecting the maturation rate, indicating that its effects werereversible in vitro. The time required to reach MII was also
significantly prolonged. These results extend previous findingsthat TSA is a new type of HDACi that can maintain oocytes atGVBD by inhibiting HDAC, which essentially arrests theacetylation of Histone 3 and Histone 4 (Endo et al. 2008).
When oocytes remain at MII in the absence of activation orfertilisation, maturation-promoting factor (MPF) and MAPKactivities decline, pre-MPF accumulates and aging increases.
MPF and MAPK are important M-phase-regulating enzymeswhose activities change throughout meiotic progression, withthe activity of both enzymes highest at MII (Kim et al. 2008).
Moreover, MAPK, also known as ERK1/2, is critical for meioticreinitiation and maturation in mammalian oocytes (Meineckeand Krischek 2003; Fan and Sun 2004). In the present study, weconfirmed oocyte quality at MII after 44, 56 and 64 h IVM by
investigating the phosphorylation status of MAPK. We foundthat ERK1/2 levels increased with increasing concentrations ofTSA compared with control at 44 h. During GVBD, histone was
deactylated and it remained acetylated by TSA. Although ourresults showed acetylation to affect the maturation of porcineoocytes, the mechanism remains unclear. Moreover, TSA did
not affect oocyte maturation up to 64 h.Mammalian oocytes are fertilised shortly after ovulation or
IVM. If fertilisation does not occur within a specific time limit,
oocytes remaining in the oviduct or in culture age (Kikuchi et al.1995, 2000; Kim et al. 1996). Unsuccessful and abnormalfertilisation is observed in aged oocytes after transfer and oftenresults in increased abnormal embryo development and early
pregnancy loss (Mailhes et al. 1998; Schramm et al. 2002).Aneuploidy is believed to be the main reason for poor embryo
development and early pregnancy loss (Fujino et al. 1996;Schramm et al. 2002). In addition, HDAC inhibition duringporcine oocyte aging significantly reduces the percentage of
fragmented oocytes. Interestingly, oocytes that mature and agefor 1 day in the presence of TSA retain their developmentalcapacity (Jeseta et al. 2008). Our results show that histone
acetylation delays maturation and prevents aging after long-term incubation (i.e. 64 h): at this time, blastocyst formation washigher in the TSA-treated groups than in the control group.Conversely, there was no significant change in blastocyst cell
number after 44 or 64 h IVM in both groups. Our results showthat TSA significantly delays IVM and aging of oocytes.
Embryonic cell apoptosis has received increasing attention in
recent years, mostly because of its proposed role in cellularresponses that lead to suboptimal developmental conditions(Betts and King 2001). Increased apoptosis is an important
indicator of poor in vitro egg quality. Therefore, changes inapoptosis in porcine blastocysts that occur during maturationmay affect blastocyst quality and embryo survival (Kim et al.
2008). TSA also decreased apoptosis in somatic cells (Wetzel
et al. 2005). The different effects of HDACi on somatic cells andaged pig oocytes may be caused by the upregulation of differentapoptotic pathways. Conversely, oocyte fragmentation is an
end-result of apoptosis (Perez et al. 1999). In the present study,TSA decreased the fragmentation of oocytes at MII and thenumber of apoptotic cells in blastocysts. Our findings show that
TSA inhibits apoptosis in blastocysts that develop from TSA-treated oocytes. Similar results have been observed in somaticcell nuclear transfer (SCNT)-derived blastocysts, although the
mean number of apoptotic nuclei in SCNT blastocysts washigher than that in IVF-derived blastocysts (Hao et al. 2004).Moreover, the incidence of cell death in mammalian blastocystsseems to be associated with cell number and embryo quality
(Levy et al. 2001; Hao et al. 2004), which is in agreement withour results.
Two major apoptotic protein families exist: (1) the Bcl2
family, which is comprised of anti-apoptotic proteins; and(2) the Caspase family, which is comprised of pro-apoptoticproteins. Members from both families are involved in embryo
preimplantation (Boise et al. 1993). In the present study, real-time RT-PCR was used to quantify the mRNA expression ofBcl-xL and Caspase 3 in blastocysts after TSA treatment. Inblastocysts, Bcl-xL expression did not differ significantly in the
control and TSA-treated oocytes after 44 or 64 h IVM.However,Caspase3 mRNA expression in blastocyst was significantlydecreased in the TSA-treated compared with control group.
Our results illustrate that TSA decreases Caspase 3 expressionand the number of apoptotic cells in blastocysts.
In conclusion, we have shown that TSA plays a critical role in
meiotic progression in porcine oocytes by arresting GVBD,which affects IVM and subsequent preimplantation embryodevelopment. Future studies should investigate developmental
potential and embryonic qualities, including structural integrityand apoptotic patterns, in the presence of TSA. Moreover, TSAwas shown to affect the time required to reach oocytematurationand to significantly delay oocyte aging.
814 Reproduction, Fertility and Development Y.-X. Jin et al.

Acknowledgement
This study was supported by grants from the Next-Generation BioGreen 21
Program (No. PJ00956302, PJ0090802013 and PJ0095942013), Rural
Development Administration, Republic of Korea.
References
Akiyama, T., Nagata, M., and Aoki, F. (2006). Inadequate histone deacety-
lation during oocyte meiosis causes aneuploidy and embryo death in
mice. Proc. Natl Acad. Sci. USA 103, 7339–7344. doi:10.1073/PNAS.
0510946103
Betts, D.H., andKing,W.A. (2001).Genetic regulation of embryo death and
senescence.Theriogenology 55, 171–191. doi:10.1016/S0093-691X(00)
00453-2
Boise, L. H., Gonzales-Garcia,M., Postema, C. E., Ding, L. A., Lindsten, T.,
Turka, L., Mao, X., Nunerz, G., and Thompson, C. B. (1993). Bcl-x, a
bcl-2 related gene that functions as a dominant regulator of apoptotic cell
death. Cell 74, 597–608. doi:10.1016/0092-8674(93)90508-N
Bornslaeger, E. A., Mattei, P., and Schultz, R. M. (1986). Involvement of
cAMP-dependent protein kinase and protein phosphorylation in regula-
tion of mouse oocyte maturation.Dev. Biol. 114, 453–462. doi:10.1016/
0012-1606(86)90209-5
DeLaFuente,R.,Viveiros,M.M.,Burns,K.H.,Adashi,E.Y.,Matzuk,M.M.,
and Eppig, J. J. (2004). Major chromatin remodeling in the germinal
vesicle of mammalian oocytes is dispensable for global transcriptional
silencing but required for centromeric heterochromatin function. Dev.
Biol. 275, 447–458. doi:10.1016/J.YDBIO.2004.08.028
Endo, T., Naito, K., Aoki, F., Kume, S., and Tojo, H. (2005). Changes in
histone modications during in vitromaturation of porcine oocytes.Mol.
Reprod. Dev. 71, 123–128. doi:10.1002/MRD.20288
Endo, T., Kano, K., and Naito, K. (2008). Nuclear histone deacetylases are
not required for global histone deacetylation during meiotic maturation
in porcine oocytes. Biol. Reprod. 78, 1073–1080. doi:10.1095/BIOLRE
PROD.107.067397
Fan, H. Y., and Sun, Q. Y. (2004). Involvement of mitogen-activated protein
kinase cascade during oocyte maturation and fertilization in mammals.
Biol. Reprod. 70, 535–547. doi:10.1095/BIOLREPROD.103.022830
Fujino, Y., Ozaki, K., Yamamasu, S., Ito, F., Matsuoka, I., Hayashi, E.,
Nakamura, H., Ogita, S., Sato, E., and Inoue, M. (1996). DNA fragmen-
tation of oocytes in aged mice. Hum. Reprod. 11, 1480–1483.
doi:10.1093/OXFORDJOURNALS.HUMREP.A019421
Funahashi, H., Cantley, T. C., and Day, B. N. (1997). Synchronization
of meiosis in porcine oocytes by exposure to dibutyryl cyclic
adenosinemonophosphate improves developmental competence follow-
ing in vitro fertilization. Biol. Reprod. 57, 49–53. doi:10.1095/BIOLRE
PROD57.1.49
Gordo, A. C., Rodrigues, P., Kurokawa, M., Jellerette, T., Exley, G. E.,
Warner, C., and Fissore, R. (2002). Intracellular calcium oscillations
signal apoptosis rather than activation in in vitro aged mouse eggs. Biol.
Reprod. 66, 1828–1837. doi:10.1095/BIOLREPROD66.6.1828
Gottlicher,M.,Minucci, S., Zhu, P., Kramer,O.H., Schimpf,A., Giavara, S.,
Sleeman, J. P., Lo Coco, F., Nervi, C., Pelicci, P. G., and Heinzel, T.
(2001). Valproic acid defines a novel class of HDAC inhibitors inducing
differentiation of transformed cells. EMBO J. 20, 6969–6978.
doi:10.1093/EMBOJ/20.24.6969
Hao, Y., Lai, L., Mao, J., Im, G. S., Bonk, A., and Prather, R. S. (2004).
Apoptosis in parthenogenetic preimplantation porcine embryos.
Biol. Reprod. 70, 1644–1649. doi:10.1095/BIOLREPROD.103.
026005
Ivanovska, I., and Orr-Weaver, T. L. (2006). Histone modications and the
chromatin scaffold for meiotic chromosome architecture. Cell Cycle 5,
2064–2071. doi:10.4161/CC.5.18.3253
Jeseta, M., Petr, J., Krejcova, T., Chmelikova, E., and Jilek, F. (2008).
In vitro ageing of pig oocytes: effects of the histone deacetylase inhibitor
trichostatin A. Zygote 16, 145–152. doi:10.1017/S0967199408004668
Jin, Y. X., Cui, X. S., Han, Y. J., and Kim, N. H. (2009). Leptin accelerates
pronuclear formation following intracytoplasmic sperm injection of
porcine oocytes: possible role for MAP kinase inactivation.
Anim. Reprod. Sci. 115, 137–148. doi:10.1016/J.ANIREPROSCI.
2008.10.021
Kikuchi, K., Izaike, Y., Noguchi, J., Furukawa, T., Daen, F. P., Naito, K., and
Toyoda, Y. (1995). Decrease of histone H1 kinase activity in relation
to parthenogenetic activation of pig follicular oocytes matured and
aged in vitro. J. Reprod. Fertil. 105, 325–330. doi:10.1530/JRF.0.
1050325
Kikuchi, K., Naito, K., Noguchi, J., Shimada, A., Kaneko, H., Yamashita, M.,
Aoki, F., Tojo, H., and Toyoda, Y. (2000). Maturation/M-phase promot-
ing factor: a regulator of aging in porcine oocytes. Biol. Reprod. 63,
715–722. doi:10.1095/BIOLREPROD63.3.715
Kim, J. S., Cho, Y. S., Song, B. S., Wee, G., Park, J. S., Choo, Y. K., Yu, K.,
Lee, K. K., Han, Y. M., and Koo, D. B. (2008). Exogenous dibutyryl
cAMP affects meiotic maturation via protein kinase A activation; it
stimulates further embryonic development including blastocyst quality
in pigs. Theriogenology 69, 290–301. doi:10.1016/J.THERIOGENOL
OGY.2007.09.024
Kim, N. H., Moon, S. J., Prather, R. S., and Day, B. N. (1996). Cytoskeletal
alteration in aged porcine oocytes and parthenogenesis. Mol. Reprod.
Dev. 43, 513–518. doi:10.1002/(SICI)1098-2795(199604)43:4,513::
AID-MRD14.3.0.CO;2-#
Kramer, O. H., Zhu, P., Ostendorff, H. P., Golebiewski, M., Tiefenbach, J.,
Peters,M.A., Brill, B., Groner,B., Bach, I., Heinzel, T., andGottlicher,M.
(2003). The histone deacetylase inhibitor valproic acid selectively
induces proteasomal degradation of HDAC2. EMBO J. 22, 3411–
3420. doi:10.1093/EMBOJ/CDG315
Kuroda, T., Naito, K., Sugiura, K., Yamashita, M., Takakura, I., and
Tojo, H. (2004). Analysis of the roles of cyclin B1 and cyclin B2 in
porcine oocyte maturation by inhibiting synthesis with antisense RNA
injection. Biol. Reprod. 70, 154–159. doi:10.1095/BIOLREPROD.103.
021519
Levy, R. R., Cordonier, H., Czyba, J. C., and Guerin, J. F. (2001). Apoptosis
in preimplantation mammalian embryo and genetics. Ital. J. Anat.
Embryol. 106, 101–108.
Liu, L., and Keefe, D. L. (2002). Ageing-associated aberration in meiosis
of oocytes from senescence-accelerated mice. Hum. Reprod. 17,
2678–2685. doi:10.1093/HUMREP/17.10.2678
Mailhes, J. B., Young, D., and London, S. N. (1998). Postovulatory ageing
of mouse oocytes in vitro and premature centromere separation and
aneuploidy. Biol. Reprod. 58, 1206–1210. doi:10.1095/BIOLRE
PROD58.5.1206
Meinecke, B., and Krischek, C. (2003). MAPK/ERK kinase (MEK) signal-
ling is required for resumption of meiosis in cultured cumulus-enclosed
pig oocytes. Zygote 11, 7–16. doi:10.1017/S0967199403001023
Perez, G. I., Tao, X. J., and Tilly, J. L. (1999). Fragmentation and death
(a.k.a. apoptosis) of ovulated oocytes. Mol. Hum. Reprod. 5, 414–420.
doi:10.1093/MOLEHR/5.5.414
Richard, F. J. (2007). Regulation of meiotic maturation. J. Anim. Sci.
85(Suppl.), E4–E6. doi:10.2527/JAS.2006-475
Schramm, R. D., Paprocki, A. M., and Bavister, B. D. (2002). Features
associated with reproductive ageing in female rhesus monkeys. Hum.
Reprod. 17, 1597–1603. doi:10.1093/HUMREP/17.6.1597
Shojo, A., Tatemoto, H., and Terada, T. (2000). Effect of aging on
parthenogenetic activation with cycloheximide and alteration of the
activity of maturation promoting factor during aging of bovine oocytes.
J. Mamm. Ova Res. 17, 35–41.
TSA delays porcine oocyte maturation and aging Reproduction, Fertility and Development 815

Sugiura, K., Naito, K., Endo, T., and Tojo, H. (2006). Study of germinal
vesicle requirement for the normal kinetics of maturation/M-phase-
promoting factor activity during porcine oocyte maturation. Biol.
Reprod. 74, 593–600. doi:10.1095/BIOLREPROD.105.046375
Sun, S.C., Gao,W.W.,Xu,Y.N., Jin,Y.X.,Wang,Q. L.,Yin,X. J., Cui, X. S.,
and Kim, N. H. (2012). Degradation of actin nucleators affects cortical
polarity of aged mouse oocytes.Fertil. Steril. 97, 984–990. doi:10.1016/
J.FERTNSTERT.2012.01.101
Voronina, E., and Wessel, G. M. (2003). The regulation of oocyte matura-
tion. Curr. Top. Dev. Biol. 58, 53–110. doi:10.1016/S0070-2153(03)
58003-6
Wang, Q., Wang, C. M., Ai, J. S., Xiong, B., Yin, S., Hou, Y., Chen, D. Y.,
Schatten, H., and Sun, Q. Y. (2006a). Histone phosphorylation and
pericentromeric histone modications in oocyte meiosis. Cell Cycle 5,
1974–1982. doi:10.4161/CC.5.17.3183
Wang, Q., Yin, S., Ai, J. S., Liang, C. G., Hou, Y., Chen, D. Y., Schatten, H.,
and Sun, Q. Y. (2006b). Histone deacetylation is required for orderly
meiosis. Cell Cycle 5, 766–774. doi:10.4161/CC.5.7.2627
Wetzel, M., Premkumar, D. R. D., Arnold, B., and Pollack, I. F. (2005).
Effect of trichostatin A, a histone deacetylase inhibitor, on glioma
proliferation in vitro by inducing cell cycle arrest and apoptosis.
J. Neurosurg. 103, 549–556.
Wilcox, A. J., Weinberg, C. R., and Baird, D. D. (1998). Post-ovulatory
ageing of the human oocyte and embryo failure. Hum. Reprod. 13,
394–397. doi:10.1093/HUMREP/13.2.394
www.publish.csiro.au/journals/rfd
816 Reproduction, Fertility and Development Y.-X. Jin et al.
Copyright © 2022 FDOKUMEN




















