
HGF promotes survival and growth of maturing sympathetic neurons by PI-3 kinase- and MAP...
12
HGF promotes survival and growth of maturing sympathetic neurons by PI-3 kinase- and MAP kinase-dependent mechanisms Jane Thompson, a,b, * Xavier Dolcet, a,c Mark Hilton, a Mary Tolcos, a,d and Alun M. Davies a,e a Department of Preclinical Veterinary Sciences, Royal (Dick) School of Veterinary Studies, University of Edinburgh, Edinburgh EH9 1QH, Scotland, United Kingdom b Fujisawa Institute of Neuroscience, Edinburgh EH8 9JE, Scotland, United Kingdom c Lab Anatomia Patologica-Genetica, Hospital Arnau de Vilanova, 25198 Lleida, Spain d Department of Anatomy and Cell Biology, University of Melbourne, Parkville, VIC, 3010 Australia e School of Biosciences, Cardiff CF10 3US, Wales, United Kingdom Received 26 April 2004; revised 26 July 2004; accepted 27 July 2004 Available online 28 October 2004 Hepatocyte growth factor (HGF) is a pleiotrophic factor whose many functions include promoting neuronal survival and growth. Hitherto, these effects have been observed in the presence of other neurotrophic factors like NGF and CNTF, and this requirement for an accessory factor has made it difficult to elucidate the signaling pathways that mediate its survival and growth-enhancing effects. Here, we show that HGF promotes the survival of mature sympathetic neurons of the superior cervical ganglion (SCG) grown at low density in defined medium lacking other neurotrophic factors. This effect was first clearly observed in cultures established from postnatal day 20 (P20) mice and became maximal by P40. HGF also enhanced the growth of neurite arbors from neurons throughout postnatal development and in the adult. HGF treatment resulted in phosphorylation of Akt and ERK1/ ERK2. Preventing Akt activation with the phosphatidylinositol-3 (PI-3) kinase inhibitor LY294002 blocked the HGF survival response, and inhibition of ERK activation with the MEK inhibitors PD98059 or U0126 reduced the HGF survival response and the neurite growth- promoting effects of HGF. These results indicate that HGF promotes the survival and growth of maturing sympathetic neurons by both PI-3 kinase- and MAP kinase-dependent mechanisms. D 2004 Elsevier Inc. All rights reserved. Introduction Hepatocyte growth factor (HGF), also known as Scatter factor, is a mutlifunctional cytokine that exerts a variety of effects on many cell types. It was first identified as a potent mitogen for hepatocytes (Montesano et al., 1991; Nakamura et al., 1989) and has subsequently been shown to play a role in morphogenesis, angiogenesis, and tumorogenesis. The effects of HGF are mediated via the receptor tyrosine kinase Met. (Bladt et al., 1995; Tsarfaty et al., 1992). Gene-targeting experiments in mice have shown that HGF and Met are required for the development of the placenta, liver, and skeletal muscle of the limbs and trunk (Bladt et al., 1995; Maina et al., 1996; Schmidt et al., 1995; Uehara et al., 1995).HGF promotes the survival and proliferation of several cell types, including Schwann cells, chondrocytes, keratinocytes, and leio- myosarcoma cells, and stimulates the migration and dissociation of epithelial sheets (Brinkmann et al., 1995; Gheradi and Stoker, 1991; Krasnoselsky et al., 1994; Mildner et al., 2002; Takebayashi et al., 1995; Xiao et al., 2000). HGF plays a role in several aspects of neural development (Maina and Klein, 1999). Experiments in chick embryos have suggested that HGF might play a role neural induction (Bronner- Fraser, 1995). Later in embryonic development and in the adult nervous system, a variety of neurons and glial cells express HGF and Met (Andermarcher et al., 1996; Di Renzo et al., 1993; Jung et al., 1994; Krasnoselsky et al., 1994; Maina et al., 1997, 1998; Sonnenberg et al., 1993; Thewke and Seeds, 1996, 1999). HGF promotes the survival of a subset of motoneurons and has been implicated in guiding motor axons to their targets (Caton et al., 2000; Ebens et al., 1996; Novak et al., 2000; Wong et al., 1997; Yamamoto et al., 1997). HGF cooperates with CNTF to promote motoneuron and parasympathetic neuron survival (Davey et al., 2000; Wong et al., 1997) and with NGF to promote sensory neuron survival and outgrowth of sensory and sympathetic axons (Maina et al., 1997, 1998; Yang et al., 1998). HGF enhances the number of tyrosine hydroxylase-positive neurons in midbrain cultures (Hama- noue et al., 1996), protects cerebellar granule neurons from apoptosis following serum withdrawal (Zhang et al., 2000), and promotes neurite growth from a subset of hippocampal neurons in culture (Korhonen et al., 2000). HGF also rescues hippocampal CA1 neurons following transient global ischemia (Miyazawa et al., 1044-7431/$ - see front matter D 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.mcn.2004.07.007 * Corresponding author. Fujisawa Institute of Neuroscience in Edin- burgh, Level 6, Appleton Tower, Crichton Street, Edinburgh, EH8 9JE, Scotland, United Kingdom. Fax: +44 131 667 9381. E-mail address: [email protected] (J. Thompson). Available online on ScienceDirect (www.sciencedirect.com.) www.elsevier.com/locate/ymcne Mol. Cell. Neurosci. 27 (2004) 441 – 452
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of HGF promotes survival and growth of maturing sympathetic neurons by PI-3 kinase- and MAP...

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 27 (2004) 441–452
HGF promotes survival and growth of maturing sympathetic neurons
by PI-3 kinase- and MAP kinase-dependent mechanisms
Jane Thompson,a,b,* Xavier Dolcet,a,c Mark Hilton,a Mary Tolcos,a,d and Alun M. Daviesa,e
aDepartment of Preclinical Veterinary Sciences, Royal (Dick) School of Veterinary Studies, University of Edinburgh, Edinburgh EH9 1QH, Scotland,
United KingdombFujisawa Institute of Neuroscience, Edinburgh EH8 9JE, Scotland, United KingdomcLab Anatomia Patologica-Genetica, Hospital Arnau de Vilanova, 25198 Lleida, SpaindDepartment of Anatomy and Cell Biology, University of Melbourne, Parkville, VIC, 3010 AustraliaeSchool of Biosciences, Cardiff CF10 3US, Wales, United Kingdom
Received 26 April 2004; revised 26 July 2004; accepted 27 July 2004
Available online 28 October 2004
Hepatocyte growth factor (HGF) is a pleiotrophic factor whose many
functions include promoting neuronal survival and growth. Hitherto,
these effects have been observed in the presence of other neurotrophic
factors like NGF and CNTF, and this requirement for an accessory
factor has made it difficult to elucidate the signaling pathways that
mediate its survival and growth-enhancing effects. Here, we show that
HGF promotes the survival of mature sympathetic neurons of the
superior cervical ganglion (SCG) grown at low density in defined
medium lacking other neurotrophic factors. This effect was first clearly
observed in cultures established from postnatal day 20 (P20) mice and
became maximal by P40. HGF also enhanced the growth of neurite
arbors from neurons throughout postnatal development and in the
adult. HGF treatment resulted in phosphorylation of Akt and ERK1/
ERK2. Preventing Akt activation with the phosphatidylinositol-3 (PI-3)
kinase inhibitor LY294002 blocked the HGF survival response, and
inhibition of ERK activation with the MEK inhibitors PD98059 or
U0126 reduced the HGF survival response and the neurite growth-
promoting effects of HGF. These results indicate that HGF promotes
the survival and growth of maturing sympathetic neurons by both PI-3
kinase- and MAP kinase-dependent mechanisms.
D 2004 Elsevier Inc. All rights reserved.
Introduction
Hepatocyte growth factor (HGF), also known as Scatter factor,
is a mutlifunctional cytokine that exerts a variety of effects on
many cell types. It was first identified as a potent mitogen for
hepatocytes (Montesano et al., 1991; Nakamura et al., 1989) and
1044-7431/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.mcn.2004.07.007
* Corresponding author. Fujisawa Institute of Neuroscience in Edin-
burgh, Level 6, Appleton Tower, Crichton Street, Edinburgh, EH8 9JE,
Scotland, United Kingdom. Fax: +44 131 667 9381.
E-mail address: [email protected] (J. Thompson).
Available online on ScienceDirect (www.sciencedirect.com.)
has subsequently been shown to play a role in morphogenesis,
angiogenesis, and tumorogenesis. The effects of HGF are mediated
via the receptor tyrosine kinase Met. (Bladt et al., 1995; Tsarfaty et
al., 1992). Gene-targeting experiments in mice have shown that
HGF and Met are required for the development of the placenta,
liver, and skeletal muscle of the limbs and trunk (Bladt et al., 1995;
Maina et al., 1996; Schmidt et al., 1995; Uehara et al., 1995).HGF
promotes the survival and proliferation of several cell types,
including Schwann cells, chondrocytes, keratinocytes, and leio-
myosarcoma cells, and stimulates the migration and dissociation of
epithelial sheets (Brinkmann et al., 1995; Gheradi and Stoker,
1991; Krasnoselsky et al., 1994; Mildner et al., 2002; Takebayashi
et al., 1995; Xiao et al., 2000).
HGF plays a role in several aspects of neural development
(Maina and Klein, 1999). Experiments in chick embryos have
suggested that HGF might play a role neural induction (Bronner-
Fraser, 1995). Later in embryonic development and in the adult
nervous system, a variety of neurons and glial cells express HGF
and Met (Andermarcher et al., 1996; Di Renzo et al., 1993; Jung et
al., 1994; Krasnoselsky et al., 1994; Maina et al., 1997, 1998;
Sonnenberg et al., 1993; Thewke and Seeds, 1996, 1999). HGF
promotes the survival of a subset of motoneurons and has been
implicated in guiding motor axons to their targets (Caton et al.,
2000; Ebens et al., 1996; Novak et al., 2000; Wong et al., 1997;
Yamamoto et al., 1997). HGF cooperates with CNTF to promote
motoneuron and parasympathetic neuron survival (Davey et al.,
2000; Wong et al., 1997) and with NGF to promote sensory neuron
survival and outgrowth of sensory and sympathetic axons (Maina
et al., 1997, 1998; Yang et al., 1998). HGF enhances the number of
tyrosine hydroxylase-positive neurons in midbrain cultures (Hama-
noue et al., 1996), protects cerebellar granule neurons from
apoptosis following serum withdrawal (Zhang et al., 2000), and
promotes neurite growth from a subset of hippocampal neurons in
culture (Korhonen et al., 2000). HGF also rescues hippocampal
CA1 neurons following transient global ischemia (Miyazawa et al.,

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452442
1998) and cerebellar granule neurons following NMDA excitotoxi-
city (Hossain et al., 2002).
Binding of HGF to Met activates cytoplasmic effectors via
multifunctional docking sites located in its intracellular domain
(Ponzetto et al., 1994). In nonneuronal cells, Met can activate
several signaling cascades, including the Ras/MAP kinase and
JNK/SAP kinase pathways via Grb2 and Gab1, phospholipid
pathways through binding of phosphatidylinositol-3 kinase (PI3K)
and phospholipase C (PLC), phosphotyrosine-mediated pathways
through interaction with Src tyrosine kinase, SHP2 tyrosine
phosphatase, Nck, and the STAT pathway (Boccaccio et al.,
1998; Kochhar and Iyer, 1996; Nguyen et al., 1997; Ponzetto et al.,
1994; Rodrigues et al., 1997; Weidner et al., 1995). Studies of mice
that express Met receptors in which the multifunctional docking
sites have been replaced with specific binding motifs for PI3K, Src,
or Grb2 have shown that signaling through PI3K is sufficient to
promote outgrowth of specific motor axons in vivo and to mediate
the growth-promoting effects of HGF on sympathetic neurites in
vitro (Maina et al., 2001). However, the signaling pathways that are
important for neuronal survival have not been ascertained.
Previous work has shown that HGF enhances the survival and
differentiation of sympathetic neuroblasts via an autocrine mech-
anism and subsequently increases the growth of neurites from
postmitotic embryonic sympathetic neurons grown in the presence
of NGF but does not enhance the survival of these neurons either
alone or in the presence of NGF (Maina et al., 1998). Here, we
have investigated effects of HGF on postnatal and adult sympa-
thetic neurons and found that it promotes their survival and growth
in culture in the absence of other neurotrophic factors. This finding
has in turn permitted us to investigate the signaling pathways that
are important for survival and have found that PI3K/Akt and, to a
lesser extent, MAP kinase signaling mediate the HGF survival
response.
Fig. 1. Bar charts of the numbers of P1, P10, P20, and P40 SCG neurons
surviving after 48-h incubation in defined medium without added factors
(Cont) or medium supplemented with 2 ng/ml NGF, 10 ng/ml HGF, or 10
ng/ml anti-HGF expressed as a percentage of the initial count made 3–6 h
after plating. The means and standard errors are shown (n = 9 for each
condition).
Results
HGF promotes the survival of maturing SCG neurons
To investigate if HGF influences the survival of sympathetic
neurons after birth and in the adult, we studied the effects of HGF
on superior cervical ganglion (SCG) neurons in low-density
dissociated cultures established at intervals from postnatal day 1
(P1) to adulthood (P40). For comparison, the neurons were also
cultured with NGF which is known to promote the survival of
sympathetic neurons in the postnatal period. In P1 cultures,
neurons grown with HGF died as rapidly as neurons in control
medium (defined medium with no added growth factors), so that
by 48-h incubation, almost all of the neurons grown under these
conditions had died. In contrast, NGF promoted the survival of the
majority of P1 neurons (Fig. 1A), and survival was neither
enhanced nor reduced by the concomitant presence of HGF (data
not shown). Between P1 and P40, increasing numbers of neurons
survived in control medium. This is due to many of the neurons
acquiring the capacity to survive independently of neurotrophic
factors with age (Orike et al., 2001a). Throughout this period of
postnatal development, between 60% and 80% of the neurons
survived in NGF-supplemented medium. In P10 cultures, signifi-
cantly more neurons survived in HGF-supplemented medium
compared with control medium (40% vs. 20% survival after 48-h
incubation, P b 0.001, t test), and in P20 and P40 cultures, HGF
enhanced neuronal survival as effectively as NGF. Indeed by P40,
the number of neurons surviving with HGF was consistently higher
in the presence of HGF, although this trend did not approach
statistical significance (Figs. 1 and 2). Fig. 2A shows that there was
no additional survival in P40 cultures supplemented with HGF plus
NGF compared with either factor alone at 24, 48, 72, and 96 h data
points, suggesting that HGF and NGF enhance the survival of
essentially the same subset of neurons in the adult. Fig. 2B plots
the dose response of P40 neurons to HGF, showing that a
concentration of 10 ng/ml was maximally effective in promoting
survival. This concentration of HGF was used in all other
experiments.
Because HGF enhances the survival of sympathetic neuroblasts
by an autocrine mechanism, we investigated whether endogenously
produced HGF plays any role in sustaining the survival of postnatal
and adult neurons by studying the effect of function-blocking anti-
HGF antiserum on the survival of these neurons. Figs. 1 and 2A
show that there were no fewer neurons surviving in the presence of
anti-HGF antibody compared with control cultures. To confirm that
the anti-HGF antibody blocks the function of HGF, we set up a
separate series of experiments to investigate if anti-HGF at the
same concentration as that used in the above experiments blocks
the survival-promoting effect of 10 ng/ml HGF in P40 cultures.
After 48-h incubation, the percent survival in cultures supple-
mented with HGF was 88.0% F 9.4% compared with 47.5% F7.3% in cultures supplemented with HGF + anti-HGF, 49.0% F

Fig. 2. (A) Bar chart of the numbers of P40 SCG neurons surviving after 24-, 48-, 72-, and 96-h incubation in defined medium without added factors (Cont) or
medium supplemented with 2 ng/ml NGF, 10 ng/ml HGF, NGF plus HGF, or 10 ng/ml anti-HGF expressed as a percentage of the initial count made 3–6 h after
plating. The means and standard errors are shown (n = 9 for each condition). (B) Graph showing the percentage survival of P40 SCG neurons grown for 48 h
with a range of HGF concentrations. For comparison, the percent survival in control cultures (bar labeled C) is also shown. The means and standard errors are
shown (n = 9 for each condition). (C) Graph showing the level of Met mRNA relative to GAPDH mRNA in total RNA extracted from P1 to P40 SCG. The
mean and standard error of measurements made from three separate assays for each age are shown.
J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452 443
5.6% in cultures containing anti-HGF alone, and 42.1% F 4.6% in
control cultures (n = 3 for all data points). Taken together, these
results suggest that secreted HGF does not play a role in sustaining
the survival of postnatal and adult SCG neurons. However, because
intracellular HGF is not accessible to the antibody, we cannot
exclude the possibility that endogenously produced HGF could
bind and activate its receptor intracellularly.
Developmental changes in Met expression
To determine if the age-related effects of HGF on sympathetic
neuron survival are correlated with changes in Met expression, we
used competitive (reverse transcription–polymerase chain reaction)
RT–PCR to quantify the levels of Met mRNA in the SCG from P1
to P60. To compensate for any developmental changes in cell
number and size of the ganglia dissected from different animals,
we also quantified the level of GAPDH mRNA which encodes a
housekeeping enzyme. Fig. 2C shows that Met mRNA was
expressed throughout this period and that there was a gradual
increase in expression after P10, which may indicate acquisition of
the HGF survival response.
HGF enhances the growth of SCG neurites
Previous work has shown that HGF cooperates with NGF to
enhance the growth of embryonic sensory and sympathetic neurites
in culture and that Met signaling is required for the proper growth
of embryonic sensory nerves in vivo (Maina et al., 1997, 1998). To
determine if HGF also affects the growth of postnatal and adult
sympathetic neurites, we used stereological methodology to
quantify the mean total length of the neurite arbors of SCG
neurons grown with and without HGF. Because different subsets of
sympathetic neurons might possess different neurite growth
characteristics independent of the presence of HGF, it was

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452444
important to compare the effect of the presence and absence of
HGF in the same subset of neurons. To achieve this, we
supplemented all cultures with NGF since, as shown in Fig. 1;
no more neurons survive with HGF plus NGF than NGF alone.
The cultures were set up at stages throughout postnatal sympathetic
neuron development, and neurite arbor length was quantified at 24,
48, 72, and 96 hourly intervals after plating. At all ages and all time
points, neurite arbors were longer in cultures containing NGF plus
HGF compared with cultures containing NGF alone. This differ-
ence was statistically significant in two of the four time points in
P1 cultures, all time points in P10 cultures, three time points in P20
cultures, and one time point in P40 cultures (Fig. 3A).
Analysis of neurite growth from P1 SCG neurons grown over a
broad range of HGF concentrations revealed that 10 ng/ml
produced the greatest increase in length of neurite arbors in
Fig. 3. (A) Bar charts of the total length of neurite arbors growing from P1, P10, P2
supplemented with 2 ng/ml NGF, NGF plus 10 ng/ml HGF, or 10 ng/ml anti-H
*Significant difference ( P b 0.05) in neurite length between NGF supplemente
difference ( P b 0.01) in neurite length between NGF supplemented cultures and cu
of P1 SCG neurons grown for 96-h incubation with a range of HGF concentration
Bar chart showing the neurite arbor lengths of Bax-deficient P1 SCG neurons gro
supplemented with either 10 ng/ml HGF or anti-HGF. (D) Bar chart showing the n
growth factors (Cont) and cultures supplemented with either 10 ng/ml HGF or anti
NGF-supplemented medium (Fig. 3B). This was the same level of
HGF that was most effective in promoting neuronal survival at
later ages. These results also demonstrate that HGF is capable of
enhancing neurite growth in the early postnatal period, although it
has no apparent effect on neuronal survival at this stage.
Although the above studies show that HGF cooperates with
NGF to enhance the growth of postnatal sympathetic neurites
beyond that promoted by NGF alone, they do not show if HGF is
capable of promoting neurite outgrowth alone. To address this
question in the immediate postnatal period when most sympathetic
neurons are dependent on NGF for survival, we established
cultures from Bax-deficient embryos. Bax is required for the death
of NGF-deprived sympathetic neurons (Deckwerth et al., 1996), so
by using Bax-deficient neurons, we were able to compare the effect
of the presence and absence of HGF on neurite growth
0, and P40 SCG neurons after 24-, 48-, 72-, and 96-h incubation in medium
GF. The means and standard errors are shown (n = 9 for each condition).
d cultures and cultures supplemented with NGF plus HGF, **Significant
ltures supplemented with NGF plus HGF. (B) Graph of neurite arbor lengths
s. The means and standard errors are shown (n = 9 for each condition). (C)
wn for 72 h in defined medium without growth factors (Cont) and cultures
eurite arbor lengths of P40 SCG neurons grown in defined medium without
-HGF. The means and standard errors are shown (n = 9 for each condition).

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452 445
independently of NGF. Fig. 3C shows that the neurites emerging
from P1 Bax-deficient sympathetic neurons were significantly
longer in the presence of HGF compared with neurons grown with
no factors (P b 0.05, t test). These studies indicate that HGF is
able to promote neurite outgrowth from postnatal sympathetic
neurons independently of NGF.
To explore the possibility that the neurite outgrowth observed
in control cultures of Bax-deficient neonatal SCG neurons may
have been due in part to the secretion of endogenous HGF, we
treated these cultures with function-blocking anti-HGF anti-
bodies. Fig. 3C shows that this treatment did not significantly
reduce neurite outgrowth compared with control cultures,
suggesting that an HGF autocrine loop is unlikely to play a
significant role in promoting neurite growth in this particular
experimental paradigm. Likewise, because a substantial propor-
tion of adult SCG neurons survive independently of added factors
and because anti-HGF does not impair the survival of these
neurons, it was also possible to determine whether endogenously
secreted HGF plays any role promoting neurite growth from adult
neurons. Fig. 3D shows that there was no significant difference
in the total neurite length between control cultures and cultures
treated with anti-HGF. Thus, despite the fact that the majority of
adult mouse SCG neurons coexpress HGF and Met (Yang et al.,
1998), these results suggest that an HGF autocrine loop is
unlikely to play a significant role in promoting neurite growth
from adult sympathetic neurons. Fig. 4 illustrates the typical
appearance of SCG neurons grown under different experimental
conditions at P40.
Fig. 4. Photomicrographs of representative P40 SCG neurons grown with NGF (A
after 96-h incubation, and the neurite arbors were stained with h-tubulin antibod
photographed with a Zeiss confocal microscope. Scale bar = 30 Am.
Role of PI-3 kinase/Akt signaling in mediating HGF survival
To ascertain which signaling pathways mediate the survival
effects of HGF in sympathetic neurons, we investigated the roles to
two pathways that have previously been implicated to various
extents in the survival-promoting effects of NGF in sympathetic
neurons, namely, the PI-3 kinase/Akt and MAP kinase pathways
(Creedon et al., 1996; Crowder and Freeman, 1998; Kuruvilla et
al., 2000; Mazzoni et al., 1999; Rodriguez-Viciana et al., 1994;
Vaillant et al., 1999; Xue et al., 2000). We begun studying the role
of PI-3 kinase/Akt pathway by using Western blotting to ascertain
whether HGF treatment results in phosphorylation of the serine/
threonine kinase Akt, one of the downstream mediators of PI-3
kinase implicated in cell survival. P40 SCG neurons were plated in
defined medium without growth factors for 6 h, then treated for 10
min with either HGF, NGF, or both factors. Western blots of
protein extracts probed with antibodies to phospho-Akt revealed
that both NGF and HGF caused marked and similar increases in the
level of phospho-Akt compared with untreated cultures. The level
of phospho-Akt in cultures treated with NGF plus HGF was similar
to that observed in cultures treated with either factor alone (Fig.
5A). These results indicate that both NGF and HGF promote
phosphorylation of Akt in mature sympathetic neurons and that
these factors in combination do not appear to have an additive
effect on Akt activation.
To investigate the significance of PI-3 kinase signaling in
mediating the HGF survival response in mature sympathetic
neurons, we treated P40 cultures with the PI-3 kinase inhibitor
), HGF (B), NGF + HGF (C), and no factors (D). The cultures were fixed
y followed by FITC-labeled secondary antibody and were visualized and
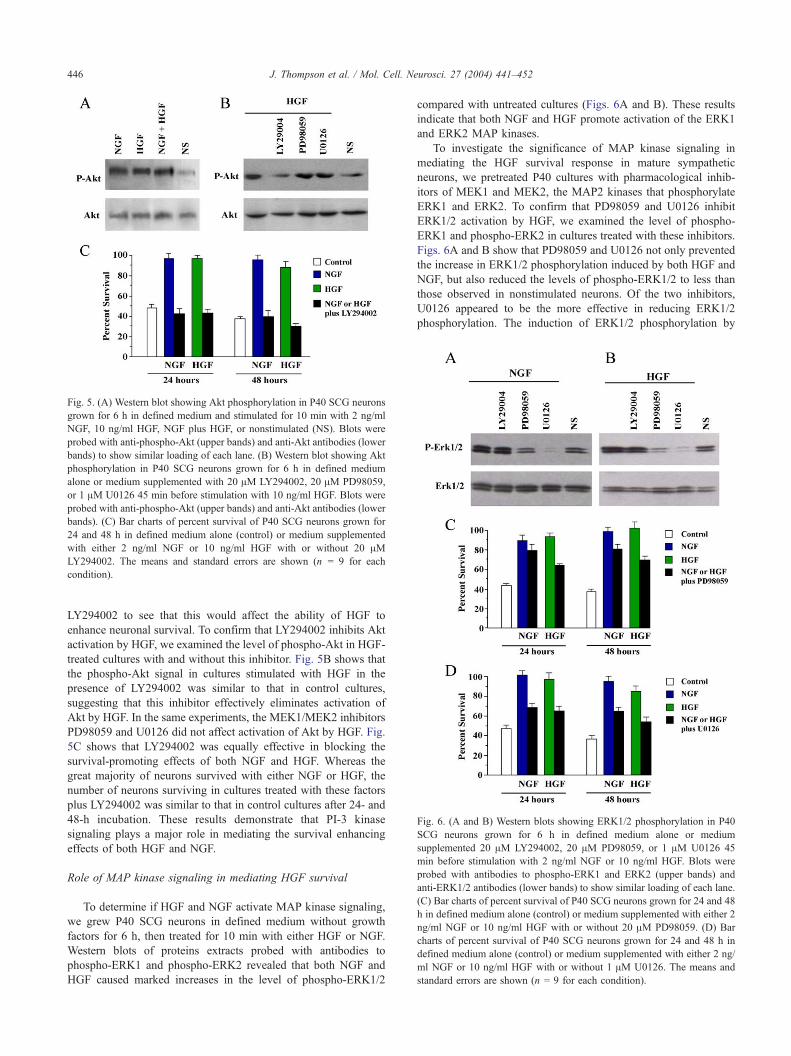
Fig. 5. (A) Western blot showing Akt phosphorylation in P40 SCG neurons
grown for 6 h in defined medium and stimulated for 10 min with 2 ng/ml
NGF, 10 ng/ml HGF, NGF plus HGF, or nonstimulated (NS). Blots were
probed with anti-phospho-Akt (upper bands) and anti-Akt antibodies (lower
bands) to show similar loading of each lane. (B) Western blot showing Akt
phosphorylation in P40 SCG neurons grown for 6 h in defined medium
alone or medium supplemented with 20 AM LY294002, 20 AM PD98059,
or 1 AM U0126 45 min before stimulation with 10 ng/ml HGF. Blots were
probed with anti-phospho-Akt (upper bands) and anti-Akt antibodies (lower
bands). (C) Bar charts of percent survival of P40 SCG neurons grown for
24 and 48 h in defined medium alone (control) or medium supplemented
with either 2 ng/ml NGF or 10 ng/ml HGF with or without 20 AMLY294002. The means and standard errors are shown (n = 9 for each
condition).
Fig. 6. (A and B) Western blots showing ERK1/2 phosphorylation in P40
SCG neurons grown for 6 h in defined medium alone or medium
supplemented 20 AM LY294002, 20 AM PD98059, or 1 AM U0126 45
min before stimulation with 2 ng/ml NGF or 10 ng/ml HGF. Blots were
probed with antibodies to phospho-ERK1 and ERK2 (upper bands) and
anti-ERK1/2 antibodies (lower bands) to show similar loading of each lane.
(C) Bar charts of percent survival of P40 SCG neurons grown for 24 and 48
h in defined medium alone (control) or medium supplemented with either 2
ng/ml NGF or 10 ng/ml HGF with or without 20 AM PD98059. (D) Bar
charts of percent survival of P40 SCG neurons grown for 24 and 48 h in
defined medium alone (control) or medium supplemented with either 2 ng/
ml NGF or 10 ng/ml HGF with or without 1 AM U0126. The means and
standard errors are shown (n = 9 for each condition).
J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452446
LY294002 to see that this would affect the ability of HGF to
enhance neuronal survival. To confirm that LY294002 inhibits Akt
activation by HGF, we examined the level of phospho-Akt in HGF-
treated cultures with and without this inhibitor. Fig. 5B shows that
the phospho-Akt signal in cultures stimulated with HGF in the
presence of LY294002 was similar to that in control cultures,
suggesting that this inhibitor effectively eliminates activation of
Akt by HGF. In the same experiments, the MEK1/MEK2 inhibitors
PD98059 and U0126 did not affect activation of Akt by HGF. Fig.
5C shows that LY294002 was equally effective in blocking the
survival-promoting effects of both NGF and HGF. Whereas the
great majority of neurons survived with either NGF or HGF, the
number of neurons surviving in cultures treated with these factors
plus LY294002 was similar to that in control cultures after 24- and
48-h incubation. These results demonstrate that PI-3 kinase
signaling plays a major role in mediating the survival enhancing
effects of both HGF and NGF.
Role of MAP kinase signaling in mediating HGF survival
To determine if HGF and NGF activate MAP kinase signaling,
we grew P40 SCG neurons in defined medium without growth
factors for 6 h, then treated for 10 min with either HGF or NGF.
Western blots of proteins extracts probed with antibodies to
phospho-ERK1 and phospho-ERK2 revealed that both NGF and
HGF caused marked increases in the level of phospho-ERK1/2
compared with untreated cultures (Figs. 6A and B). These results
indicate that both NGF and HGF promote activation of the ERK1
and ERK2 MAP kinases.
To investigate the significance of MAP kinase signaling in
mediating the HGF survival response in mature sympathetic
neurons, we pretreated P40 cultures with pharmacological inhib-
itors of MEK1 and MEK2, the MAP2 kinases that phosphorylate
ERK1 and ERK2. To confirm that PD98059 and U0126 inhibit
ERK1/2 activation by HGF, we examined the level of phospho-
ERK1 and phospho-ERK2 in cultures treated with these inhibitors.
Figs. 6A and B show that PD98059 and U0126 not only prevented
the increase in ERK1/2 phosphorylation induced by both HGF and
NGF, but also reduced the levels of phospho-ERK1/2 to less than
those observed in nonstimulated neurons. Of the two inhibitors,
U0126 appeared to be the more effective in reducing ERK1/2
phosphorylation. The induction of ERK1/2 phosphorylation by
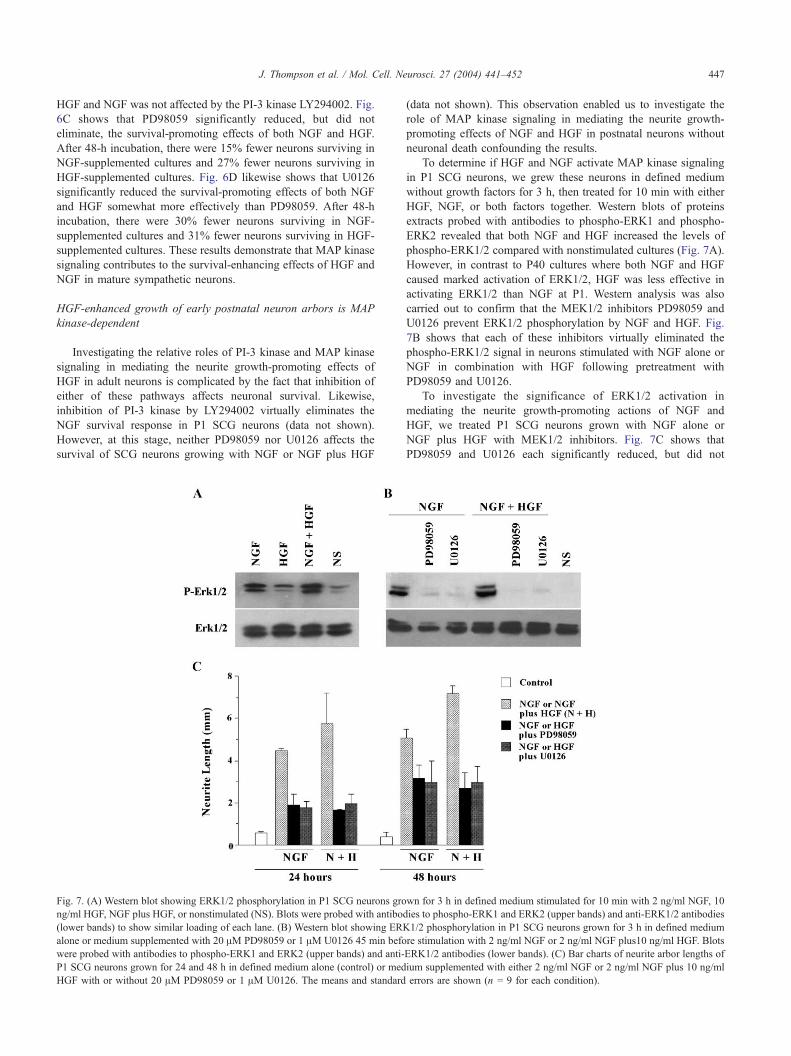
J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452 447
HGF and NGF was not affected by the PI-3 kinase LY294002. Fig.
6C shows that PD98059 significantly reduced, but did not
eliminate, the survival-promoting effects of both NGF and HGF.
After 48-h incubation, there were 15% fewer neurons surviving in
NGF-supplemented cultures and 27% fewer neurons surviving in
HGF-supplemented cultures. Fig. 6D likewise shows that U0126
significantly reduced the survival-promoting effects of both NGF
and HGF somewhat more effectively than PD98059. After 48-h
incubation, there were 30% fewer neurons surviving in NGF-
supplemented cultures and 31% fewer neurons surviving in HGF-
supplemented cultures. These results demonstrate that MAP kinase
signaling contributes to the survival-enhancing effects of HGF and
NGF in mature sympathetic neurons.
HGF-enhanced growth of early postnatal neuron arbors is MAP
kinase-dependent
Investigating the relative roles of PI-3 kinase and MAP kinase
signaling in mediating the neurite growth-promoting effects of
HGF in adult neurons is complicated by the fact that inhibition of
either of these pathways affects neuronal survival. Likewise,
inhibition of PI-3 kinase by LY294002 virtually eliminates the
NGF survival response in P1 SCG neurons (data not shown).
However, at this stage, neither PD98059 nor U0126 affects the
survival of SCG neurons growing with NGF or NGF plus HGF
Fig. 7. (A) Western blot showing ERK1/2 phosphorylation in P1 SCG neurons gro
ng/ml HGF, NGF plus HGF, or nonstimulated (NS). Blots were probed with antibod
(lower bands) to show similar loading of each lane. (B) Western blot showing ERK
alone or medium supplemented with 20 AM PD98059 or 1 AM U0126 45 min befo
were probed with antibodies to phospho-ERK1 and ERK2 (upper bands) and anti-
P1 SCG neurons grown for 24 and 48 h in defined medium alone (control) or med
HGF with or without 20 AM PD98059 or 1 AM U0126. The means and standard
(data not shown). This observation enabled us to investigate the
role of MAP kinase signaling in mediating the neurite growth-
promoting effects of NGF and HGF in postnatal neurons without
neuronal death confounding the results.
To determine if HGF and NGF activate MAP kinase signaling
in P1 SCG neurons, we grew these neurons in defined medium
without growth factors for 3 h, then treated for 10 min with either
HGF, NGF, or both factors together. Western blots of proteins
extracts probed with antibodies to phospho-ERK1 and phospho-
ERK2 revealed that both NGF and HGF increased the levels of
phospho-ERK1/2 compared with nonstimulated cultures (Fig. 7A).
However, in contrast to P40 cultures where both NGF and HGF
caused marked activation of ERK1/2, HGF was less effective in
activating ERK1/2 than NGF at P1. Western analysis was also
carried out to confirm that the MEK1/2 inhibitors PD98059 and
U0126 prevent ERK1/2 phosphorylation by NGF and HGF. Fig.
7B shows that each of these inhibitors virtually eliminated the
phospho-ERK1/2 signal in neurons stimulated with NGF alone or
NGF in combination with HGF following pretreatment with
PD98059 and U0126.
To investigate the significance of ERK1/2 activation in
mediating the neurite growth-promoting actions of NGF and
HGF, we treated P1 SCG neurons grown with NGF alone or
NGF plus HGF with MEK1/2 inhibitors. Fig. 7C shows that
PD98059 and U0126 each significantly reduced, but did not
wn for 3 h in defined medium stimulated for 10 min with 2 ng/ml NGF, 10
ies to phospho-ERK1 and ERK2 (upper bands) and anti-ERK1/2 antibodies
1/2 phosphorylation in P1 SCG neurons grown for 3 h in defined medium
re stimulation with 2 ng/ml NGF or 2 ng/ml NGF plus10 ng/ml HGF. Blots
ERK1/2 antibodies (lower bands). (C) Bar charts of neurite arbor lengths of
ium supplemented with either 2 ng/ml NGF or 2 ng/ml NGF plus 10 ng/ml
errors are shown (n = 9 for each condition).

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452448
eliminate, the neurite growth response of the neurons to NGF alone
and NGF plus HGF. In the absence of inhibitors, neurite arbors
were longer in cultures supplemented with NGF plus HGF
compared with NGF alone, but in cultures treated with inhibitors,
neurite arbors were reduced to the same length in cultures
containing NGF alone and in cultures containing NGF plus HGF.
This suggests that MAP kinase signaling plays a major role in
mediating the neurite growth-enhancing actions of both HGF and
NGF in developing postnatal sympathetic neurons.
Discussion
During embryonic development it has been established that
HGF promotes the survival and differentiation of sympathetic
neuroblasts and enhances the growth of neurites from NGF-
supported, postmitotic sympathetic neurons but does not promote
the survival of these neurons (Maina et al., 1998). To ascertain
whether HGF influences the survival and growth of postnatal and
adult sympathetic neurons, we studied the effects of HGF in
purified cultures of SCG neurons established at stages after birth.
These studies have revealed that, between P10 and P20, HGF
promotes the survival of an increasing proportion of neurons in a
dose-dependent manner. Between P20 and P40, HGF is as
effective as NGF in promoting neuronal survival. The lack of
any additive effect of HGF plus NGF on survival indicates that
these factors promote the survival of the same subset of neurons,
which at P20 and P40 amounts to about a third of the neurons in
the ganglion. The acquisition of the HGF survival response by
sympathetic neurons in the late postnatal period is correlated with a
progressive increase in the levels of transcripts encoding the Met
receptor. Previous work on the roles of HGF in the embryonic
peripheral nervous system has shown that its survival-enhancing
effect on sensory and parasympathetic neurons is only observed in
the presence of another neurotrophic factor (Davey et al., 2000;
Maina et al., 1997). In contrast, in the juvenile and adult
sympathetic nervous system, HGF is capable of promoting
neuronal survival on its own, and this property has facilitated
investigation of the signaling pathways mediating HGF-promoted
survival (below).
Acquisition of the HGF survival response takes place over the
same extended period of postnatal development as the acquisition
of the capacity of many of the neurons to survive independently of
neurotrophic factors. In the present study, up to a third of mouse
SCG neurons are able to survive in serum-free medium without
neurotrophic factors by adulthood. An even larger proportion of
adult rat SCG neurons are able to survive in culture without
neurotrophic factors, and the demonstration that they can do so
when grown as single cells indicates that they require no factors
from other cells to survive (Orike et al., 2001b). These findings
together with the demonstration that the majority of neurons
coexpress HGF and Met in the adult mouse SCG (Yang et al.,
1998) raise the possibility that an HGF autocrine loop may play a
role in sustaining the survival of adult SCG neurons in medium
lacking neurotrophic factors. However, our demonstration that
function-blocking anti-HGF antibody does not affect the survival
of adult SCG neurons suggests that the secretion of HGF from
these neurons does not play a significant role in sustaining their
survival.
In agreement with previous studies of embryonic and neonatal
SCG neurons (Maina et al., 1998; Yang et al., 1998), we have
shown that, throughout the postnatal period and in adult SCG
neurons, HGF enhances neurite growth from neurons incubated in
the presence of NGF. In addition, we have shown that the growth-
enhancing effect of HGF is not dependent on the presence of NGF
because HGF increases neurite length in cultures of Bax-deficient
neonatal SCG neurons surviving without NGF and increases the
length of neurites growing from the subset of adult SCG neurons
that survive independently of NGF. Thus, HGF not only cooperates
with NGF in promoting neurite outgrowth throughout the life of
sympathetic neurons, but also has a direct neurite growth-
promoting effect on these neurons in the postnatal and adult
nervous system.
Although the majority of adult mouse SCG neurons coexpress
HGF and Met (Yang et al., 1998), our results suggest that secreted
HGF acting on its own receptor does not play a significant role in
promoting neurite growth from either neonatal or adult mouse SCG
neurons grown with or without NGF. Function-blocking anti-HGF
antibody that has previously been shown to inhibit the differ-
entiation of mouse sympathetic neuroblasts (Maina et al., 1998)
does not significantly reduce the length of neurites growing from
neurons incubated with NGF at any age studied and does not
reduce the length of neurites from neurons grown without NGF
(either Bax-deficient neonatal neurons or NGF-independent adult
SCG neurons). These findings appear to conflict with the results of
Yang et al. (1998) who showed that anti-HGF antibody reduces
neurite density and neurite elongation in neonatal rat SCG cultures
containing NGF. The conflicting effects of anti-HGF on neurite
growth in these two studies could represent a difference between
the sympathetic neurons of these two species of rodents or could be
related to differences in the experimental paradigms used. For
example, it may be significant that Yang et al. grew SCG neurons
on collagen, whereas we cultured SCG neurons on laminin, which
permits more extensive neurite outgrowth.
The direct survival-promoting effect of HGF on adult SCG
neurons enabled us to investigate which signaling pathways
mediate the antiapoptotic actions of HGF without the complication
of having to supplement the medium with other neurotrophic
factors. We found that pharmacological inhibition of PI-3 kinase
prevented HGF-induced phosphorylation of Akt and eliminated
the HGF survival response, indicating that PI-3 kinase signaling
plays a key role in mediating the antiapoptotic actions of HGF.
Furthermore, pharmacological inhibition of MEK1 and MEK2
prevented HGF-induced phosphorylation of ERK1 and ERK2 and
reduced, but did not eliminate, the HGF survival response,
indicating that MAP kinase signaling also contributes to the
antiapoptotic actions of HGF in adult sympathetic neurons.
Pharmacological inhibition of PI-3 kinase and MAP kinase
signaling in adult SCG had similar effects on the NGF survival
response, suggesting that both HGF and NGF promote the survival
of mature sympathetic neurons by substantially similar intra-
cellular mechanisms. In postnatal rat cerebellar granule cells,
inhibition of PI-3 kinase also prevents the neuroprotective effect of
HGF following treatment with NMDA, quinolinic acid, or low K+-
containing medium (Hossain et al., 2002; Zhang et al., 2000).
However, in contrast with adult sympathetic neurons, inhibition of
MEK1/2 in postnatal cerebellar granule cells does not affect the
neuroprotective effects of HGF. The apparent lack of involvement
of MAP kinase signaling in the neuroprotective effects of HGF on
postnatal cerebellar granule cells is similar to the case for the
survival response of neonatal SCG neurons to NGF in our current
study. Thus, whereas PI-3 kinase signaling plays a key role in

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452 449
mediating the survival effects of NGF and HGF at all ages at
which SCG neurons respond to these factors, MAP kinase
signaling only begins to play a role as the neurons become fully
mature. Studies of neonatal rat sympathetic neurons have reported
that MAP kinase activation plays only a minor role or no role at all
in mediating the survival effects of NGF (Creedon et al., 1996;
Klesse and Parada, 1998; Mazzoni et al., 1999; Virdee and
Tolkovsky, 1996; Xue et al., 2000).
The lack of effect of MEK1/2 inhibition on the survival of
neonatal SCG neurons enabled us to investigate role of MAP
kinase signaling in the neurite growth-promoting effects of NGF
and HGF without effects on neuronal viability complicating
interpretation of the results. Pharmacological inhibition of MEK1
and MEK2 blocked growth factor-induced phosphorylation of
ERK1 and ERK2 and significantly reduced neurite arbors to the
same length in cultures treated with NGF alone and NGF plus
HGF. This suggests that MAP kinase signaling plays a role in
mediating the neurite growth-enhancing actions of both HGF and
NGF in neonatal sympathetic neurons, although some component
of neurite growth is not dependent on this signaling pathway.
Activation of Map kinase by growth factors has previously been
shown to play a role in mediating their neurite growth-promoting
effects on PC12 cells and a variety neurons (Aletsee et al., 2001;
Choung et al., 2002; Klinz et al., 1996; Sjogreen et al., 2000).
Although the key role of PI-3 kinase signaling in mediating the
survival-promoting effects of HGF precluded direct assessment of
the role of this signaling pathway in HGF-induced neurite growth,
there is evidence from studies of mice expressing Met receptors
whose multifunctional docking sites have been replaced with
specific binding motifs for PI3K, Src, or Grb2 that signaling
through PI3K is sufficient to promote outgrowth of specific motor
axons in vivo and to mediate the growth-promoting effects of HGF
on sympathetic neurites in vitro (Maina et al., 2001).
In summary, we have provided evidence for a direct survival-
promoting effect of HGF on juvenile and adult sympathetic
neurons and have shown that HGF is capable of regulating the
growth of sympathetic neurites throughout the postnatal period and
in the adult. The antiapoptotic effect of HGF is dependent on PI-3
kinase signaling and to a lesser extent on MAP kinase signaling,
and the neurite growth-promoting effect depends at least in part on
MAP kinase signaling. These findings clarify the signaling
pathways utilized by HGF to mediate its various effects on
neurons and imply that HGF has an ongoing role in modulating
sympathetic neuron plasticity and function in the mature nervous
system.
Experimental methods
Neuronal culture
Superior cervical ganglia were dissected from P1 to P40 CD1
mice. P1 superior cervical ganglia were trypsinized (0.05% trypsin
in Ca2+/Mg2+-free HBSS) for 15 min at 378C and dissociated by
trituration using fire-polished Pasteur pipette. P20 and older
ganglia were first treated with collagenase (2 mg/ml in Ca2+/
Mg2+ containing HBSS) for 30 min at 48C before trypsinization for
20 min at 378C and trituration. Most of the nonneuronal cells were
removed, and the resulting cell suspensions by differential
sedimentation (Davies, 1986) and the neurons (N90% pure) were
plated in defined, serum-free medium in 35-mm culture dishes
coated with poly-l-ornithine and laminin. Purified recombinant
human HGF (R and D Systems, UK), NGF (Genentech), function-
blocking polyclonal anti-HGF antibodies (R and D Systems) or
pharmacological inhibitors of PI-3 kinase (LY294002, Calbio-
chem, UK), and MEK1/MEK2 (PD98059 and U0126, Calbio-
chem) were added to the cultures at the time of plating. The
concentration at which specific inhibition was observed by each
inhibitor was determined using dose–response curves and the
appropriate controls (data not shown).
To study survival under different experimental conditions,
neurons were plated at low density (100 to 300 neurons per 35-mm
dish), and the number of neurons attached within a 12 � 12 mm
grid in the center of each dish was counted 6 h after plating. The
number of neurons surviving in this grid was then counted at 24
hourly intervals, and the number of surviving neurons in the grid at
these time points was expressed as a percentage of the initial count
taken at 6 h. In each experiment, triplicate cultures were set up for
all conditions.
To study neurite outgrowth, cultures were set up following the
same experimental procedure as before, except that the neurons
were plated at a higher density (400 to 800 neurons per 35-mm
dish). Total neurite length was measured for each experimental
condition using digital stereology (Kinetic Imaging Ltd.), and the
average neurite length per neuron was calculated by dividing this
figure by neuron number per unit area.
Protein extraction and Western blotting/ECL detection
Primary cultures of dissociated P1 and P40 SCG neurons were
grown for 3 to 6 h in culture before being stimulated for 10 min
with HGF or NGF. Pharmacological inhibitors of PI-3 kinase or
MEK1/MEK2 were added 45 min before trophic factor stimula-
tion. Cells were harvested and homogenized in lysis buffer (50 mM
Tris pH 7.5, 1% Triton, 150 mM NaCl, 1 mM EDTA), and total
protein (10 Ag per lane) was resolved in 8% SDS–polyacrylamide
gels and transferred onto polyvinylidene difluoride blotting
membrane (BDH Poole, Dorset, UK) using a Mini Trans-Blot
Electrophoretic Transfer Cell (Biorad, Herts, UK) according to the
manufacturer’s instructions. The membranes were blocked for 1 h
in 5% Milk/Tris-buffered saline containing 0.1% Tween 20
(TBST), incubated with rabbit polyclonal antibodies to either
phospho-ERK1/2 or phospho-Akt (Cell Signalling Technology,
UK) overnight in TBST at 48C and incubated with a horseradish
peroxidase-linked donkey antirabbit IgG secondary antibody
(Amersham Biosciences, UK) for 1 h in 5% milk/TBST. The
membranes were then stripped with 100 mM h-mercaptoethanol,
2% SDS, 62.5 mM Tris-HCl, pH 6.8 for 30 min at 558C and
reprobed with rabbit antibodies to either ERK1/2 or Akt (Cell
Signalling Technology). The membranes were developed using the
ECL Western detection kit (Amersham Pharmacia Biotech,
Buckinghamshire, UK).
Imaging of SCG neurons
Primary cultures of dissociated P40 SCG neurons were grown
for 96 h in culture in the presence of NGF (2 ng/ml), HGF (10 ng/
ml), NGF+HGF, or without added factors (control). After 96 h, the
neurons were fixed in 100% methanol for 10 min, and then washed
three times in 1� phosphate-buffered saline (PBS). Fixed neurons
were permeabilized in 1� PBS 0.1% Triton-X for 30 min and then
blocked for 1 h in 10% bovine serum albumin (BSA)/PBS (Sigma).

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452450
Anti h-tubulin II monoclonal antibody (1:1000) (Promega) in 10%
BSA/PBS was added to the fixed neurons and left to incubate
overnight at 48C. The following day primary antibody was
removed, and neurons were washed three times in 1� PBS.
FITC-labeled Ig antimouse secondary antibody (1: 2000) (Sigma)
was added and incubated for 1 h. Neurons were washed three times
in 1� PBS and photomicrographs taken using the confocal
microscope.
Measurement of Met mRNA levels
A quantitative competitive reverse transcription–polymerase
chain reaction (RT–PCR) technique (Wyatt and Davies, 1993) was
used to measure the levels Met mRNA in total RNA extracted from
P1 to P60 SCG. To compare the relative levels of the receptor in
different samples, the cRNA competitor template for the ubiq-
uitous, constitutively expressed GAPDH was also included in the
reverse transcription reactions. Total RNA was extracted (Chomc-
zynski and Sacchi, 1987) treated with DNase, purified using an
RNAID kit (BIO101), and recovered in 40 Al of DEPC-treated
H2O. The RNA was reverse-transcribed for 1 h at 378C with
MMuLV-reverse transcriptase, RNaseH (Biogene Ltd., UK) in a
40-Al reaction containing the manufacturer’s buffer supplemented
with 0.5 mM dNTPs, and 10 AM random hexanucleotides. A 5-Alaliquot of each reverse transcription reaction was then amplified in
a 50-Al PCR reaction containing 1� PC2 buffer (Helena
Biosciences), 0.1 mM dNTPs, 2 U of Taq Supreme (Helena
Biosciences), 40 ng of each primer. The forward and reverse assay
primers for Met cDNA were: 5V-CCAGTCCTATATTGATGTC-3Vand 5V-TTCGAAGGCATGTATGTAC-3V. Met cDNA was amplified
by cycling at 958C for 1 min, followed by 1 min at 488C, followed1 min at 688C, for 37 cycles. The reaction was then completed with
a 10-min extension at 688C. For GAPDH cDNA, cycling
conditions were the same except only 29 cycles were carried out
and the annealing temperature was 558C. The RT–PCR products
were run on 8% nondenaturing polyacrylamide gels and visualized
by staining with syber gold (Molecular Probes, Leiden, Nether-
lands). The intensities of the RT–PCR products of the native
mRNA and competitor cRNA were calculated using Phoretix 1D
Quantifier v4.01 (Biogene Ltd.). The amount of Met or GAPDH
mRNAs in each total RNA sample was calculated from these
values.
Acknowledgments
This work was supported by a grant from the Wellcome Trust.
Mary Tolcos was supported by a Howard Florey Research
Fellowship. Xavier Dolcet was supported by a Spanish Ministry
of Education Fellowship.
References
Aletsee, C., Beros, A., Mullen, L., Pak, K., Dazert, S., Ryan, F., 2001. Ras/
MEK but not p38 signaling mediates NT-3 induced neurite extension
from spiral ganglion neurons. J. Assoc. Res. Otolaryngol. 2, 377–387.
Andermarcher, E., Surani, M.A., Gherardi, E., 1996. Co-expression of the
HGF/SF and c-Met genes during early mouse embryogenesis precedes
reciprocal expression in adjacent tissues during organogenesis. Dev.
Genet. 18, 254–266.
Bladt, F., Riethmacher, D., Isenmann, S., Aguzzi, A., Birchmeier, C., 1995.
Essential role for the Met receptor in the migration of myogenic
precursor cells into the limb bud. Nature 376, 768–771.
Boccaccio, C., Ando, M., Tamagnone, L., Bardelli, A., Michieli, P.,
Battistini, C., Comoglio, P.M., 1998. Induction of epithelial tubules
by growth factor HGF depends on the STAT pathway. Nature 391,
285–288.
Brinkmann, V., Foroutan, H., Sachs, M., Weidner, K.M., Birchmeier, W.,
1995. Hepatocyte growth factor/scatter factor induces a variety of tissue
specific morphogenic programs in epithelial cells. J. Cell Biol. 131,
1573–1586.
Bronner-Fraser, M., 1995. Hepatocyte growth factor/scatter factor (HGF/
SF) in early development: evidence for a role in neural induction.
Trends Genet. 11, 423–425.
Caton, A., Hacker, A., Naeem, A., Livet, J., Maina, F., Bladt, F., Klein, R.,
Birchmeier, C., Guthrie, S., 2000. The branchial arches and HGF are
growth-promoting and chemoattractant for cranial motor axons.
Development 127, 1751–1766.
Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA isolation
by acid guanidinium thiocyanate–phenol–chloroform extraction. Anal.
Biochem. 162, 156–159.
Choung, P.H., Seo, B.M., Chung, C.P., Yamada, K.M., Jang, J.H., 2002.
Synergistic activity of fibronectin and fibroblast growth factor receptors
on neuronal adhesion and neurite extension through extracellular signal
related kinase pathway. Biochem. Biophys. Res. 295, 898–902.
Creedon, D.J., Johnson, E.M., Lawrence, J.C., 1996. Mitogen activated
protein kinase-independent pathways mediate the effects of nerve
growth factor and cAMP on neuronal survival. J. Biol. Chem. 271,
20713–20718.
Crowder, R.J., Freeman, R.S., 1998. Phosphatidylinositol-3 kinase and
Akt protein kinase are necessary and sufficient for the survival of
nerve growth factor-dependent sympathetic neurons. J. Neurosci. 18,
2933–2943.
Davey, F., Hilton, M.C., Davies, A.M., 2000. Cooperation between HGF
and CNTF in promoting the survival and growth of sensory and
parasympathetic neurons. Mol. Cell. Neurosci. 15, 79–87.
Davies, A.M., 1986. The survival and growth of embryonic proprioceptive
neurons is promoted by a factor present in skeletal muscle. Dev. Biol.
115, 56–67.
Deckwerth, T.L., Elliott, J.L., Knudson, C.M., Johnson Jr., E.M., Snider,
W.D., Korsmeyer, S.J., 1996. Bax is required for neuronal death
after trophic factor deprivation and during development. Neuron 17,
401–411.
Di Renzo, M.F., Bertolotto, A., Olivero, M., Putzolu, P., Crepaldi, T.,
Schiffer, D., Pagni, C.A., Comoglio, P.M., 1993. Selective expression
of the Met/HGF receptor in human central nervous system microglia.
Oncogene 8, 219–222.
Ebens, A., Brose, K., Leonardo, E.D., Hanson, M.G.J., Bladt, F.,
Birchmeier, C., Barres, B.A., Tessier-Lavigne, M., 1996. Hepatocyte
growth factor/ scatter factor is an axonal chemoattractant and a
neurotrophic factor for spinal motor neurons. Neuron 17, 1157–1172.
Gheradi, E., Stoker, M., 1991. Hepatocyte growth factor-scatter factor:
mitogen, motogen and met. Cancer Cells 3, 227–232.
Hamanoue, M., Takemoto, N., Matsumoto, K., Nakamura, T., Nakajima, K.,
Kohsaka, S., 1996. Neurotrophic effect of hepatocyte growth factor on
central nervous system neurons in vitro. J. Neurosci. Res. 43, 554–564.
Hossain, M.A., Russell, J.C., Gomes, R., Laterra, J., 2002. Neuroprotection
by scatter factor/hepatocyte growth factor and FGF-1 in cerebellar
granule neurons is phosphatidylinositol-3 kinase/Akt dependent and
MAPK/CREB-independent. J. Neurochem. 81, 365–378.
Jung, W., Castren, E., Odenthal, M., Vande Woude, G.F., Ishii, T., Dienes,
H.P., Lindholm, D., Schirmacher, P., 1994. Expression and functional
interaction of hepatocyte growth factor-scatter factor and its receptor c-
Met in mammalian brain. J. Cell Biol. 126, 485–494.
Klesse, L.J., Parada, L.F., 1998. p21 Ras and phosphatidylinositol-3 kinase
are required for survival of wild-type and NF1 mutant sensory neurons.
J. Neurosci. 18, 10420–10428.

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452 451
Klinz, F.J., Wolff, P., Heumann, R., 1996. Nerve growth factor stimulated
mitogen activated protein kinase activity is not necessary for neurite
outgrowth of chick dorsal root ganglion sensory and sympathetic
neurons. J. Neurosci. Res. 46, 720–726.
Kochhar, K.S., Iyer, A.P., 1996. Hepatocyte growth factor induces
activation of Nck and phospholipase C-gamma in lung carcinoma cells.
Cancer Lett. 104, 163–169.
Korhonen, L., Sjoholm, U., Takei, N., Kern, M.A., Schirmacher, P.,
Castren, E., Lindholm, D., 2000. Expression of c-Met in developing rat
hippocampus: evidence for HGF as a neurotrophic factor for calbindin
d-expressing neurons. Eur. J. Neurosci. 12, 3453–3461.
Krasnoselsky, A., Massay, M.J., DeFrances, M.C., Michalopoulos, G.,
Zarnegar, R., Ratner, N., 1994. Hepatocyte growth factor is a mitogen
for Schwann cells and is present in neurofibromas. J. Neurosci. 14,
7284–7290.
Kuruvilla, R., Ye, H., Ginty, D.D., 2000. Spatially and functionally distinct
roles of the PI3-K effector pathway during NGF signaling in
sympathetic neurons. Neuron 27, 499–512.
Maina, F., Klein, R., 1999. Hepatocyte growth factor, a versatile signal for
developing neurons. Nat. Neurosci. 2, 213–217.
Maina, F., Casagranda, F., Audero, E., Simeone, A., Comologlio, P.M.,
Klein, R., Ponzetto, C., 1996. Uncoupling of Grb2 from the Met
receptor in vivo reveals complex roles in muscle development. Cell 87,
531–542.
Maina, F., Hilton, M.C., Ponzetto, C., Davies, A.M., Klein, R., 1997. Met
receptor signalling is required for sensory nerve development. Genes
Dev. 11, 3341–3350.
Maina, F., Hilton, M.C., Andres, R., Wyatt, S., Klein, R., Davies, A.M.,
1998. Multiple roles for hepatocyte growth factor in sympathetic neuron
development. Neuron 20, 835–846.
Maina, F., Pante, G., Helmbacher, F., Andres, R., Porthin, A., Davies, A.M.,
Ponzetto, C., Klein, R., 2001. Coupling Met to specific pathways results
in distinct developmental outcomes. Mol. Cell 7, 1293–1306.
Mazzoni, I.E., Said, F.A., Aloyz, R., Miller, F.D., Kaplan, D., 1999. Ras
regulates sympathetic neuron survival by suppressing the p53-mediated
cell death pathway. J. Neurosci. 19, 9716–9727.
Mildner, M., Eckhart, L., Lengauer, B., Tschachler, E., 2002. Hepatocyte
growth factor inhibits UVB induced apoptosis of human keratinocytes
but not of keratinocyte derived cell lines via the PI3-kinase/Akt
pathway. J. Biol. Chem. 277, 14146–14152.
Miyazawa, T., Matsumoto, K., Ohmichi, H., Katoh, H., Yamashima, T.,
Nakamura, T., 1998. Protection of hippocampal neurons from ischemia-
induced delayed neuronal death by hepatocyte growth factor: a novel
neurotrophic factor. J. Cereb. Blood Flow Metab. 18, 345–348.
Montesano, R., Matsumoto, K., Nakamura, T., Orci, L., 1991. Identification
of a fibroblast derived epithelial morphogen as hepatocyte growth
factor. Cell 67, 901–908.
Nakamura, T., Nishizawa, T., Hagiya, M., Seki, T., Shimonishi, M.,
Tashiro, K., Shimizu, S., 1989. Molecular cloning and expression of
human hepatocyte growth factor. Nature 342, 440–444.
Nguyen, L., Holgado-Madruga, M., Maroun, C., Fixman, E.D.,
Kamikura, D., Fournier, T., Charest, A., Tremblay, M.L., Wong,
A.J., Park, M., 1997. Association of the multisubstrate docking
protein Gab1 with the hepatocyte growth factor receptor requires a
functional Grb2 binding site involving tyrosine 1356. J. Biol. Chem.
272, 20811–20819.
Novak, K.D., Prevette, D., Wang, S., Gould, T.W., Oppenheim, R.W., 2000.
Hepatocyte growth factor/scatter factor is a neurotrophic survival factor
for lumbar but not for other somatic motoneurons in the chick embryo.
J. Neurosci. 20, 326–337.
Orike, N., Thrasivoulou, C., Wrigley, A., Cowen, T., 2001a. Differential
regulation of survival and growth in adult sympathetic neurons: an in
vitro study of neurotrophin responsiveness. J. Neurobiol. 47, 295–305.
Orike, N., Middleton, G., Borthwick, E., Buchman, V., Cowen, T., Davies,
A.M., 2001b. Role of PI 3-kinase, Akt and Bcl-2-related proteins in
sustaining the survival of neurotrophic factor-independent adult
sympathetic neurons. J. Cell Biol. 154, 995–1005.
Ponzetto, C., Bardelli, A., Zhen, Z., Maina, F., dalla Zonca, P., Giordano, S.,
Graziani, A., Panayotou, G., Comoglio, P.M., 1994. A multifunctional
docking site mediates signaling and transformation by the hepatocyte
growth factor/scatter factor receptor family. Cell 77, 261–271.
Rodrigues, G.A., Park, M., Schlessinger, J., 1997. Activation of the JNK
pathway is essential for transformation by the Met oncogene. EMBO J.
16, 2634–2645.
Rodriguez-Viciana, P., Warne, P.H., Dhand, R., Vanhaesebroeck, B., Gout,
I., Fry, M.J., Waterfield, M.D., Downward, J., 1994. Phosphatidylino-
sitol-3-OH kinase as a direct target of Ras. Nature. 370, 527–532.
Schmidt, C., Bladt, F., Goedecke, S., Brinkmann, V., Zschiesche, W.,
Sharpe, M., Gherardi, E., Birchmeier, C., 1995. Scatter factor/
hepatocyte growth factor is essential for liver development. Nature
373, 699–702.
Sjogreen, B., Wiklund, P., Ekstrom, P.A., 2000. Mitogen activated protein
kinase inhibition by PD98059 blocks nerve growth factor stimulated
axonal outgrowth from adult mouse dorsal root ganglion. Neuroscience
100, 407–416.
Sonnenberg, E., Meyer, D., Weidner, K.M., Birchmeier, C., 1993. Scatter
factor/hepatocyte growth factor and its receptor, the c-Met tyrosine
kinase, can mediate a signal exchange between mesenchyme and
epithelia during mouse development. J. Cell Biol. 123, 223–235.
Takebayashi, T., Iwamoto, M., Jikko, A., Matsumura, T., Enomoto-
Iwamoto, M., Myoukai, F., Koyama, E., Yamaai, T., Matsumoto, K.,
Nakamura, T., 1995. Hepatocyte growth factor/scatter factor modulates
cell motility, proliferation, and proteoglycan synthesis of chondrocytes.
J. Cell Biol. 129, 1411–1419.
Thewke, D.P., Seeds, N.W., 1996. Expression of hepatocyte growth factor/
scatter factor, its receptor, c-Met, and tissue-type plasminogen activator
during development of the murine olfactory system. J. Neurosci. 16,
6933–6944.
Thewke, D.P., Seeds, N.W., 1999. The expression of mRNAs for
hepatocyte growth factor/scatter factor, its receptor c-Met, and one of
its activators tissue-type plasminogen activator show a systematic
relationship in the developing and adult cerebral cortex and hippo-
campus. Brain Res. 821, 356–367.
Tsarfaty, I., Resau, J.H., Rulong, S., Keydar, I., Faletto, D.L., VandeWoude,
G.F., 1992. The Met proto-oncogene receptor and lumen formation.
Science 257, 1258–1261.
Uehara, Y., Minowa, O., Mori, C., Shiota, K., Kuno, J., Noda, T., Kitamura,
N., 1995. Placental defect and embryonic lethality in mice lacking
hepatocyte growth factor/Scatter factor. Nature 373, 702–705.
Vaillant, A.R., Mazzoni, I., Tudan, C., Boudreau, M., Kaplan, D.R., Miller,
F.D., 1999. Depolarization and neurotrophins converge on the
phosphatidylinositol 3-kinase-Akt pathway to synergistically regulate
neuronal survival. J. Cell Biol. 146, 955–966.
Virdee, K., Tolkovsky, A.M., 1996. Inhibition of p42 and p44 mitogen-
activated protein kinase activity by PD98059 does not suppress nerve
growth factor-induced survival of sympathetic neurones. J. Neurochem.
67, 1801–1805.
Weidner, K.M., Sachs, M., Riethmacher, D., Birchmeier, W., 1995.
Mutation of juxtamembrane tyrosine residue 1001 suppresses loss-of-
function mutations of the Met receptor in epithelial cells. Proc. Natl.
Acad. Sci. 92, 2597–2601.
Wong, V., Glass, D.J., Arriaga, R., Yancopoulos, G.D., Lindsay, R.M.,
Conn, G., 1997. Hepatocyte growth factor promotes motor neuron
survival and synergises with ciliary neurotrophic factor. J. Biol. Chem.
272, 5187–5191.
Wyatt, S., Davies, A.M., 1993. Regulation of expression of mRNAs
encoding the nerve growth factor receptors p75 and trkA in developing
sensory neurons. Development 119, 635–648.
Xiao, G.H., Jeffers, M., Bellacosa, A., Mitsuuchi, Y., Vande Woude, G.F.,
Testa, J.R., 2000. Anti-apoptotic signaling by hepatocyte growth factor/
Met via the phosphatidylinositol-kinase pathways. Proc. Natl. Acad.
Sci. 98, 247–252.
Xue, L., Murray, J.H., Tolkovsky, A.M., 2000. The Ras/phosphatidylino-
sitol 3-kinase and Ras/ERK pathways function as independent survival

J. Thompson et al. / Mol. Cell. Neurosci. 27 (2004) 441–452452
modules each of which inhibits a distinct apoptotic signaling pathway in
sympathetic neurons. J. Biol. Chem. 275, 8817–8824.
Yamamoto, Y., Livet, J., Vesjsada, R., Pollock, R.A., Arce, V.,
deLapeyiere, O., Kato, A.C., Henderson, C.E., 1997. Hepatocyte
growth factor (HGF/SF) is an essential muscle derived survival factor
for a subpopulation of embryonic motoneurons. Development 124,
2903–2913.
Yang, X.M., Toma, J.G., Bamji, S.X., Belliveau, D.J., Kohn, J., Park, M.,
Miller, F.D., 1998. Autocrine hepatocyte growth factor provides a local
mechanism for promoting axonal growth. J. Neurosci. 18, 8369–8381.
Zhang, L., Himi, T., Morita, I., Murota, S., 2000. Hepatocyte growth
factor protects cultured rat cerebellar granule neurons via the phos-
phatidylinositol-3 kinase pathway/Akt pathway. J. Neurosci. Res. 59,
489–496.




















