
hERG Channel Inhibitors in Extracts of Coptidis Rhizoma
6
Introduction ! In the myocardium, the hERG (human ether-a-go- go-related gene) channel conducts the rapid de- layed rectifier K + current (I Kr ) which accelerates the repolarization of the action potential. Channel inhibition leads to delayed repolarization and, consequently, to a prolonged action potential visi- ble as an extended QT interval on the electrocar- diogram (ECG) [1]. Class III anti-arrhythmic agents block I Kr . Thus, hERG channel inhibition may be associated with pro-arrhythmic and anti- arrhythmic effects. Drug-induced QT prolonga- tion is a risk factor for the development of tor- sades de pointes (TdP), a potentially life-threaten- ing arrhythmia. This has led to several drug with- drawals, restrictions of use, warnings, and rela- bellings [2]. One of the most recent withdrawals due to an arrhythmogenic risk has been the anti- tussive drug clobutinol in 2007 [3]. Since both car- diac and noncardiac drugs have the potential to inhibit the repolarizing current I Kr , screening for hERG channel inhibition is routinely performed in the pharmaceutical industry as part of the early pharmacological profiling of leads [4]. In contrast, virtually nothing is known about hERG channel blockers of natural origin. Naringenin is probably the best studied natural product, both in vitro and in vivo. Intake of naringenin via grapefruit juice has been shown to lead to QT prolongation in healthy humans [5–7]. To assess the risk of hERG channel inhibition by medicinal plants, we screened a library of 91 herbal extracts derived from major officinal herb- al drugs of the European and Chinese Pharmaco- poeias. Extracts were tested by means of an auto- mated two-microelectrode voltage-clamp assay Abstract ! Inhibition of the hERG channel delays repolariza- tion and prolongs the QT interval and cardiac ac- tion potential which can lead to sudden death. Several drugs have been withdrawn from the market due to hERG channel inhibition. In the search of hERG channel inhibitors of natural ori- gin, we established an HPLC-based profiling ap- proach which combines HPLC-microfractionation and bioactivity testing on Xenopus laevis oocytes. The methanolic extract of the TCM herbal drug Coptidis rhizoma (Coptis chinensis Franch., Ra- nunculaceae) reduced the peak tail hERG current by 31.7 ± 2.0% at 100 μg/mL. HPLC-based activity profiling pointed towards berberine as the active constituent. However, hERG inhibition by 100 μM of a reference sample of berberine (16.3 ± 1.6%) was less pronounced than previously reported. Subsequent LC‑PDA‑MS analysis showed that ber- berine collected by microfractionation of the Cop- tis extract had been, in part, transformed to active dihydroberberine. Formic acid added to the HPLC mobile phase to reduce peak tailing of protober- berine alkaloids acted as a reducing reagent ac- cording to the mechanism of the Leuckart-Wal- lach reaction. Among other structurally related protoberberines tested, dihydroberberine (30.1 ± 10.1% at 100 μM) was the most potent hERG inhibitor. Abbreviations ! hERG: human ether-a-go-go-related gene ECG: electrocardiogram TdP: torsades de pointes TCM: traditional Chinese medicine ASE: accelerated solvent extraction * The authors contributed equally to this work. hERG Channel Inhibitors in Extracts of Coptidis Rhizoma Authors Anja Schramm 1 *, Igor Baburin 2 *, Steffen Hering 2 , Matthias Hamburger 1 Affiliations 1 Division of Pharmaceutical Biology, University of Basel, Basel, Switzerland 2 Institute of Pharmacology and Toxicology, University of Vienna, Vienna, Austria Key words l " hERG channel inhibition l " herbal extracts l " Coptis chinensis l " Ranunculaceae l " HPLC‑based activity profiling l " protoberberine alkaloids received Nov. 19, 2010 revised February 11, 2011 accepted February 18, 2011 Bibliography DOI http://dx.doi.org/ 10.1055/s-0030-1270920 Published online March 16, 2011 Planta Med 2011; 77: 692–697 © Georg Thieme Verlag KG Stuttgart · New York · ISSN 0032‑0943 Correspondence Prof. Dr. Matthias Hamburger Department of Pharmaceutical Sciences Division of Pharmaceutical Biology University of Basel Klingelbergstrasse 50 4056 Basel Switzerland Phone: + 41 6 12 67 14 25 Fax: + 41 6 12 67 14 74 Matthias.Hamburger @unibas.ch 692 Schramm A et al. hERG Channel Inhibitors … Planta Med 2011; 77: 692–697 Rapid Communications
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of hERG Channel Inhibitors in Extracts of Coptidis Rhizoma

Abstract!
Inhibition of the hERG channel delays repolariza-tion and prolongs the QT interval and cardiac ac-tion potential which can lead to sudden death.Several drugs have been withdrawn from themarket due to hERG channel inhibition. In thesearch of hERG channel inhibitors of natural ori-gin, we established an HPLC-based profiling ap-proach which combines HPLC-microfractionationand bioactivity testing on Xenopus laevis oocytes.The methanolic extract of the TCM herbal drugCoptidis rhizoma (Coptis chinensis Franch., Ra-nunculaceae) reduced the peak tail hERG currentby 31.7 ± 2.0% at 100 µg/mL. HPLC-based activityprofiling pointed towards berberine as the activeconstituent. However, hERG inhibition by 100 µMof a reference sample of berberine (16.3 ± 1.6%)was less pronounced than previously reported.Subsequent LC‑PDA‑MS analysis showed that ber-
berine collected by microfractionation of the Cop-tis extract had been, in part, transformed to activedihydroberberine. Formic acid added to the HPLCmobile phase to reduce peak tailing of protober-berine alkaloids acted as a reducing reagent ac-cording to the mechanism of the Leuckart-Wal-lach reaction. Among other structurally relatedprotoberberines tested, dihydroberberine(30.1 ± 10.1% at 100 µM) was the most potenthERG inhibitor.
Abbreviations!
hERG: human ether-a-go-go-related geneECG: electrocardiogramTdP: torsades de pointesTCM: traditional Chinese medicineASE: accelerated solvent extraction
hERG Channel Inhibitors in Extractsof Coptidis Rhizoma
Authors Anja Schramm1*, Igor Baburin2*, Steffen Hering2, Matthias Hamburger1
Affiliations 1 Division of Pharmaceutical Biology, University of Basel, Basel, Switzerland2 Institute of Pharmacology and Toxicology, University of Vienna, Vienna, Austria
Key wordsl" hERG channel inhibitionl" herbal extractsl" Coptis chinensisl" Ranunculaceael" HPLC‑based activity profilingl" protoberberine alkaloids
received Nov. 19, 2010revised February 11, 2011accepted February 18, 2011
BibliographyDOI http://dx.doi.org/10.1055/s-0030-1270920Published online March 16,2011Planta Med 2011; 77: 692–697© Georg Thieme Verlag KGStuttgart · New York ·ISSN 0032‑0943
CorrespondenceProf. Dr. Matthias HamburgerDepartment of PharmaceuticalSciencesDivision of PharmaceuticalBiologyUniversity of BaselKlingelbergstrasse 504056 BaselSwitzerlandPhone: + 41612671425Fax: + [email protected]
692
Schramm A et al. hERG Channel Inhib
Rapid Communications
Introduction!
In the myocardium, the hERG (human ether-a-go-go-related gene) channel conducts the rapid de-layed rectifier K+ current (IKr) which acceleratesthe repolarization of the action potential. Channelinhibition leads to delayed repolarization and,consequently, to a prolonged action potential visi-ble as an extended QT interval on the electrocar-diogram (ECG) [1]. Class III anti-arrhythmicagents block IKr. Thus, hERG channel inhibitionmay be associated with pro-arrhythmic and anti-arrhythmic effects. Drug-induced QT prolonga-tion is a risk factor for the development of tor-sades de pointes (TdP), a potentially life-threaten-ing arrhythmia. This has led to several drug with-drawals, restrictions of use, warnings, and rela-
* The authors contributed equally to this work.
itors… Planta Med 2011; 77: 692–697
bellings [2]. One of the most recent withdrawalsdue to an arrhythmogenic risk has been the anti-tussive drug clobutinol in 2007 [3]. Since both car-diac and noncardiac drugs have the potential toinhibit the repolarizing current IKr, screening forhERG channel inhibition is routinely performedin the pharmaceutical industry as part of the earlypharmacological profiling of leads [4]. In contrast,virtually nothing is known about hERG channelblockers of natural origin. Naringenin is probablythe best studied natural product, both in vitro andin vivo. Intake of naringenin via grapefruit juicehas been shown to lead to QT prolongation inhealthy humans [5–7].To assess the risk of hERG channel inhibition bymedicinal plants, we screened a library of 91herbal extracts derived frommajor officinal herb-al drugs of the European and Chinese Pharmaco-poeias. Extracts were tested by means of an auto-mated two-microelectrode voltage-clamp assay

Fig. 1 Inhibition of potassium currents through hERG channels expressedin Xenopus oocytes by Coptidis rhizoma extract (MeOH, 100 µg/mL).A A mean current inhibition by 31.7 ± 2.0% (n = 6) was observed. B A cur-rent trace recorded in the absence (control) of the drug is superimposedwith the current traces recorded in the presence of Coptidis rhizoma(MeOH; 100 µg/mL) during a 1 Hz pulse train (15 pulses) applied from−80mV. The voltage protocol is shown above the current traces.
693Rapid Communications
on Xenopus laevis oocytes [8], at a concentration of 100 µg/mL.This concentration has been previously found to be appropriatefor screening of herbal extracts in the oocyte model with otherion channel-type targets such as GABAA receptors [9,10]. Amongthe extracts tested, the methanolic extract of Coptidis rhizoma(Coptis chinensis Franch., Ranunculaceae) induced 31.7 ± 2.0%(n = 6) inhibition of the peak tail hERG current (l" Fig. 1). Coptidisrhizoma (Huanglian) is a well-known herbal drug in traditionalChinese medicine (TCM). According to the Chinese Pharmaco-poeia, it is used as an antipyretic, antimicrobial, and anti-inflam-matory remedy for treating fever, gastrointestinal disorders (e.g.,dysentery, vomiting, and icterus), insomnia, nose bleeding, andtoothache [11,12]. Quaternary protoberberine alkaloids repre-sent the main constituents and are regarded as pharmacologi-cally active principles of the herbal drug [12,13]. On the otherhand, the alkaloids are mainly responsible for the acute toxicityof Coptidis rhizoma extract. Berberine, the most abundant alka-loid [14], also showed the strongest cytotoxicity in different celllines [13]. The broad public considers herbal drugs to be gener-ally safe. However, it is known that some herbs cause drug inter-actions or may display unwanted effects [15]. In the case of Cop-tidis rhizoma, respiratory failure, liver function injury, and severearrhythmia have been reported [13]. Here, we describe the local-ization of hERG channel inhibitory activity of Coptidis rhizomaextract with the aid of an HPLC-based profiling protocol. HPLC-based activity profiling enables rapid identification of pharmaco-logically active natural products in extracts with only milligram-amounts of extracts [16].
Materials and Methods!
General experimental proceduresAnalytical HPLC separations were performed with an Agilent1100 system consisting of an autosampler, degasser, binarypump, column oven, and diode array detector. The HPLC condi-tions were: SunFire C18 (3.5 µm, 3.0 × 150mm) column (Waters),water + 0.1% formic acid (solvent A), methanol + 0.1% formic acid(solvent B), gradient profile: 20%→ 35% B (0–25min), 35%→
100% B (25–26min), 100% B (26–30min), flow rate: 0.4mL/min,DAD 254 nm. For LC‑PDA‑MS analysis, the HPLC was coupled to amicrOTOF ESI‑MS system (Bruker Daltonics). Mass calibrationwas performed with a solution of formic acid 0.1% in 2-propa-nol/water (1 :1) containing 5mM NaOH. Mass spectra were re-corded in the positive ion mode. Data acquisition and analysiswere performed using HyStar 3.2 software. NMR spectra (1H,COSY, HSQC, HMBC, NOESY, NOE) were recorded on a BrukerAvance III 500MHz spectrometer (Bruker Daltonics) operating at500.13MHz for 1H. The instrument was equipped with a 1-mmTXI microprobe. All measurements were carried out at 291.15 K.Topspin 2.1 was used to process and analyse the spectra.
Solvents and chemicalsFormic acid (98.0–100.0%), dimethylsulfoxide (DMSO), andHPLC-grade methanol were from Scharlau. Trifluoroacetic acid(≥ 98%) was purchased from Sigma-Aldrich. Solvents for the ex-traction and isolation process were of analytical grade whereasHPLC-grade solvents were used for HPLC separations. Berberinechloride hydrate (~ 95%), dihydroberberine (≥ 91% by HPLC), and(R,S)-tetrahydroberberine (≥ 95% by TLC andMS) were purchasedfrom Fluka, Phytolab, and Latoxan, respectively.
Plant materialThe drug Coptidis rhizoma (Huanglian) (batch no. 0901503) waspurchased from Yong Quan GmbH. A voucher sample (num-ber 00197) is preserved at the Division of Pharmaceutical Biol-ogy, University of Basel, Switzerland. Prior to extraction, theplant material was frozen with liquid nitrogen and powderedwith a ZM 1 ultracentrifugal mill (sieve size 2.0mm) (Retsch).
ExtractionThe extract for the initial screening and the HPLC-based activityprofiling was prepared by accelerated solvent extraction (ASE)using a Dionex ASE 200 extractor with solvent controller (Dio-nex). Extraction was carried out in 11mL cartridges with the fol-lowing conditions: preheat time, 1min; static extraction cycle,5min; purge, 120 s with nitrogen; temperature, 70°C; pressure,120 bar. Extraction was successively done with petroleum ether,ethyl acetate, and methanol (3 cycles per each solvent). For isola-tion, the ground rhizomes (375 g) were extracted exhaustivelywith methanol by percolation at room temperature. After evapo-ration of the solvent under reduced pressure, a viscous extract(60.8 g) was obtained. All samples, including the extracts, werestored at 4°C until use.
Microfractionation and activity profilingMicrofractionation for HPLC-based activity profiling was carriedout on an Agilent 1100 system consisting of an autosampler, de-gasser, quaternary pump, column oven, and diode array detector.We separated 2.5mg of the extract by semipreparative RP-HPLCinto 42 one-minute microfractions. Separationwas achieved on aSunFire Prep C18 (5 µm, 10 × 150mm) column (Waters) withwater + 0.1% formic acid (solvent A) and methanol + 0.1% formicacid (solvent B) using the following gradient profile: 15%→ 35%B (0–35min), 35%→ 100% B (35–36min), 100% B (36–42min).The flow rate was 4mL/min, and detection was at 254 nm. 50 µLof a stock solution of the extract (50mg/mL in DMSO) was in-jected. Fractions were collected into glass tubes. After solvent re-moval (Genevac EZ-2 Plus evaporator), the dried films were re-dissolved in methanol, transferred to 4-mL glass vials, and driedagain under N2 gas prior to dissolution in DMSO for bioassay.
Schramm A et al. hERG Channel Inhibitors… Planta Med 2011; 77: 692–697

694 Rapid Communications
Isolation of protoberberines 1–5For isolation of alkaloids, a portion (12.1 g) of the extract was sep-arated by column chromatography on silica gel 60 column (63–200 µm, 9 × 31 cm i.d.). The mobile phase was a mixture of pro-panol, methanol, water, and formic acid (64:30:5 :1 v/v/v/v). Toelute the alkaloids, the polarity of the solvent system was in-creased gradually (64 :30:5:1, 59:30:10:1, 54:30:15:1,49:30:20:1, 39:40:20:1, 29:50:20:1; 2 L each). Fractions of200mL were collected and combined after TLC analysis (silicagel 60 F254 precoated Al sheets, propanol-methanol-water-formicacid 64:30:5:1 v/v/v/v, detection at UV366 nm) into 16 fractions.Final purification of quaternary protoberberines was achieved ona prep HPLC system consisting of a LC-8A preparative pump, SCL-10AVP system controller, and SPD-M10A VP PDA detector andcontrolled by class-VP V6.14 SP2A software (all Shimadzu). Pre-parative HPLC separations were performed on a SunFire PrepC18 OBD (5 µm, 30 × 150mm) column (Waters) usingwater + 0.025% TFA (solvent A) and methanol + 0.025% TFA (sol-vent B) as eluents. The flow rate was 20mL/min, and chromato-grams were recorded at 254 nm. Separation of fraction 10 af-forded 1 (tR 11.1min) and 5 (tR 13.6min) with the following gra-dient: 30% B (0–3min), 30%→ 55% B (3–18min), 55%→ 100% B(18–19min), 100% B (19–23min). Purification of fraction 5yielded 3 (tR 14.1min) and 4 (tR 15.4min), while compound 2 (tR12.4min) was isolated from fraction 16. The gradient was as fol-lows: 30% B (0–4min), 30%→ 55% B (4–24min), 55%→ 100% B(24–26min), 100% B (26–30min). Compound purity was ≥ 95%by 1H‑NMR.
Expression of hERG channels and voltage clamp studiesPreparation of stage V–VI oocytes from Xenopus laevis (NASCO),synthesis of capped run-off complementary RNA (cRNA) tran-scripts from linearized complementary DNA (cDNA) templates,and injection of cRNA were described previously [17]. Currentsthrough hERG channels were studied 1–4 days after microin-jection of the cRNA with the two-microelectrode voltage clampmethodmaking use of a TURBO TEC-03X amplifier (npi electronicGmbH). The bath solution contained: 96mM 2-(N-morpho-lino)ethanesulphonic acid sodium salt, 2mM 2-(N-morpho-
Schramm A et al. hERG Channel Inhibitors… Planta Med 2011; 77: 692–697
lino)ethanesulphonic acid potassium salt, 1mM CaCl2, 5mMHEPES, and 1mM MgCl2 adjusted to pH 7.6 with methanesul-phonic acid. Voltage-recording and current-injecting micro-electrodes were filled with 3mM KCl and pulled to have resis-tances between 0.3 and 2MΩ. Oocytes withmaximal current am-plitudes > 2 µA were discarded to exclude voltage clamp errors.The pClamp software package version 10.0 (Molecular Devices,Inc.) was used for data acquisition.
Voltage protocolThe voltage protocol (see insets in l" Figs. 1B, 2C–D, and l" 5B)was designed to simulate voltage changes during a cardiac actionpotential with a 300ms depolarization to + 20mV (analogous toplateau phase), a repolarization for 300ms to − 40mV (inducing atail current) and a final step to the holding potential. The + 20mVdepolarization rapidly inactivates hERG channels, thereby limit-ing the amount of outward current. During the repolarization to− 40mV, the previously activated channels open due to rapid re-covery from inactivation. The decreases in the resulting tail cur-rent amplitudes were taken as a measure of block developmentduring a pulse train.
Drug application and analysisDrugs were applied by means of a fast ScreeningTool perfusiontechnique (npi electronic GmbH) [8]. ORIGIN software version7.0 (OriginLab Corporation) was used for data analysis. Data aregiven asmean ± S.E. of at least two oocytes and ≥ 2 oocytebatches.
Results and Discussion!
In the search for hERG channel inhibitors of natural origin, weadapted a previously developed protocol for discovery of GABAA
receptor modulators [9]. For this purpose, we separated 2.5mgof the methanolic extract of Coptidis rhizoma by semipreparativeRP-HPLC into 42 one-minutemicrofractions whichwere tested inthe functional bioassay. l" Fig. 2 shows the HPLC chromatogramrecorded at 254 nm and the corresponding activity profile. hERGinhibition was studied during 1Hz pulse trains applied from a
Fig. 2 Activity profiling of Coptidis rhizoma ex-tract (MeOH) for inhibition of the hERG channel.B HPLC chromatogram (254 nm) of a semiprepara-tive separation of 2.5mg extract and (A) inhibition(in %) of hERG currents by 42 one-minute fractionsare shown. C, D The control current (recorded inthe absence of drug) is superimposed with thehERG currents recorded during a 1 Hz pulse train(15 pulses) in the presence of the two most activefractions (24 and 25). Same voltage protocol as inl" Fig. 1.

695Rapid Communications
holding potential of − 80mV (seeMethods for details on the pulseprotocol). Main activity was identified in microfractions 24 and25 (peak tail hERG current inhibition by 63.3 ± 16.7% [n = 2] and59.5 ± 20.9% [n = 2], respectively) which corresponded to thedominant HPLC peak. With the aid of on-line (LC‑PDA‑MS) andoff-line (1mm microprobe NMR of collected peak) spectroscopicdata, the main peak was identified as berberine (4) (l" Fig. 3), apreviously reported hERG channel blocker of natural origin [18,19]. However, inhibition of the hERG current by 100 µM of a ref-erence sample of berberine was, in our hands, less pronouncedthan previously reported (16.3 ± 1.6%, n = 3, but see [18,19]).However, subsequent LC‑PDA‑MS analysis showed that berberine(4) collected bymicrofractionation of the Coptis extract had been,in part, transformed to active dihydroberberine (6). This could beexplained by the mechanism of the Leuckart-Wallach reaction[20], whereby formic acid added to the mobile phase to reducepeak tailing of quaternary alkaloids, acted as a reducing reagent(l" Fig. 4E). The iminium ion structure of protoberberines is sen-sitive to a nucleophilic attack by the hydride ion delivered by for-mic acid. Traces of formic acid remaining in the microfractionsafter evaporation of the mobile phase were apparently sufficientfor the partial reduction of quaternary protoberberines at roomtemperature. The reaction occurred in the aprotic solvent DMSO,which is used to dissolve the fractions for bioassay, but not in theprotic solvent methanol. This differential behaviour can be bestexplained by the solvent capability to solvate ions rather than bythe solvent polarity [21]. The formation of dihydroberberine (6)was confirmed by comparative off-line microprobe NMR andon-line LC‑PDA‑MS analysis in DMSO and methanol of active mi-crofraction 24 (l" Fig. 4A–D). However, dihydroprotoberberinesare also known to auto-oxidize back to the quaternary alkaloids,especially under the influence of light [22]. Given the initialprofiling protocol and the findings described above, microfrac-tions 24 and 25 in the profiling consisted of a mixture of berber-ine (4) and dihydroberberine (6). When tested at 100 µM, dihy-droberberine (6) reduced the peak tail hERG current more effi-
ciently (30.1 ± 10.1% [n = 3, l" Fig. 5]) than the same concentra-tion of the reference sample of berberine (16.3 ± 1.6%). We reporthere on the hERG channel inhibitory property of dihydroberber-ine for the first time.Despite the fact that the activity profile revealed no notable ac-tivity for other peaks in the chromatogram, we isolated structur-ally related quaternary protoberberines for the purpose of pre-liminary information on structure activity relationships for thiscompound class. Compounds 1–5 (l" Fig. 3) were purified bymeans of open column chromatography on silica gel and prepar-ative RP-HPLC. To avoid the above described conversion into di-hydro derivatives in samples used for activity testing, trifluoro-acetic acid (TFA) was used as the mobile phase additive for RP-HPLC. Quaternary alkaloids 1–5, as well as dihydroberberine (6)and (R,S)-tetrahydroberberine (7), were tested at 100 µM fortheir ability to block the human hERG channel (l" Fig. 5). Quater-nary protoberberines were marginally active, and substitutionpattern at positions C(2), C(3), C(9), and C(10) had moderate in-fluence on activity. Among berberine (4) and its derivatives 6 and7, dihydroberberine (6) showed the highest activity (peak tailhERG current inhibition by 30.1 ± 10.1%, n = 3). Berberine (4) dis-played an inhibition (14.9 ± 1.7%, n = 3) similar to that obtainedwith the reference sample of berberine (see above), while tetra-hydroberberine (7) had almost no effect (6.6 ± 5.4%, n = 3,l" Fig. 5A). These observations suggest that the double bond be-tween C(13) and C(13a) and the resulting extended conjugatedπ-electron system are of importance for the inhibitory effect.Although berberine prolongs the action potential duration, theunderlying molecular mechanism remains elusive [23]. The lesspronounced hERG inhibition by berberine, in our hands, may beexplained by reduced channel inhibition during the shorter con-ditioning pre-pulses applied in our study (300ms vs. 4 s in [18,19]). Future studies will show if the different voltage protocolsapplied induce different state-dependent hERG inhibition [17].Coptidis rhizoma preparations are mostly used internally and,therefore, the oral bioavailability of the main alkaloid, berberine,
Fig. 3 Structures of the isolated protoberberinealkaloids 1–5 and the two berberine derivatives di-hydroberberine (6) and (R,S)-tetrahydroberberine(7).
Schramm A et al. hERG Channel Inhibitors… Planta Med 2011; 77: 692–697
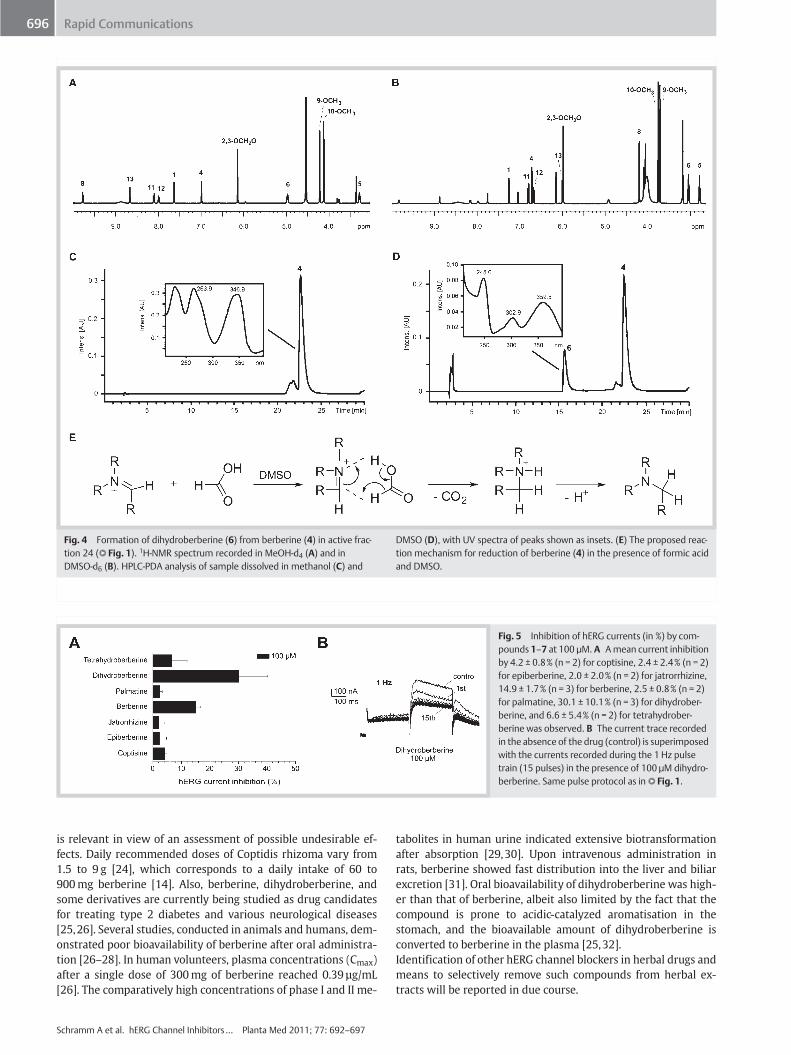
Fig. 4 Formation of dihydroberberine (6) from berberine (4) in active frac-tion 24 (l" Fig. 1). 1H‑NMR spectrum recorded in MeOH‑d4 (A) and inDMSO‑d6 (B). HPLC‑PDA analysis of sample dissolved in methanol (C) and
DMSO (D), with UV spectra of peaks shown as insets. (E) The proposed reac-tion mechanism for reduction of berberine (4) in the presence of formic acidand DMSO.
Fig. 5 Inhibition of hERG currents (in %) by com-pounds 1–7 at 100 µM.A Amean current inhibitionby 4.2 ± 0.8% (n = 2) for coptisine, 2.4 ± 2.4% (n = 2)for epiberberine, 2.0 ± 2.0% (n = 2) for jatrorrhizine,14.9 ± 1.7% (n = 3) for berberine, 2.5 ± 0.8% (n = 2)for palmatine, 30.1 ± 10.1% (n = 3) for dihydrober-berine, and 6.6 ± 5.4% (n = 2) for tetrahydrober-berine was observed. B The current trace recordedin the absence of the drug (control) is superimposedwith the currents recorded during the 1 Hz pulsetrain (15 pulses) in the presence of 100 µM dihydro-berberine. Same pulse protocol as inl" Fig. 1.
696 Rapid Communications
is relevant in view of an assessment of possible undesirable ef-fects. Daily recommended doses of Coptidis rhizoma vary from1.5 to 9 g [24], which corresponds to a daily intake of 60 to900mg berberine [14]. Also, berberine, dihydroberberine, andsome derivatives are currently being studied as drug candidatesfor treating type 2 diabetes and various neurological diseases[25,26]. Several studies, conducted in animals and humans, dem-onstrated poor bioavailability of berberine after oral administra-tion [26–28]. In human volunteers, plasma concentrations (Cmax)after a single dose of 300mg of berberine reached 0.39 µg/mL[26]. The comparatively high concentrations of phase I and II me-
Schramm A et al. hERG Channel Inhibitors… Planta Med 2011; 77: 692–697
tabolites in human urine indicated extensive biotransformationafter absorption [29,30]. Upon intravenous administration inrats, berberine showed fast distribution into the liver and biliarexcretion [31]. Oral bioavailability of dihydroberberine was high-er than that of berberine, albeit also limited by the fact that thecompound is prone to acidic-catalyzed aromatisation in thestomach, and the bioavailable amount of dihydroberberine isconverted to berberine in the plasma [25,32].Identification of other hERG channel blockers in herbal drugs andmeans to selectively remove such compounds from herbal ex-tracts will be reported in due course.

697Rapid Communications
Acknowledgements!
We thank Dr. Christian Raith and Dr. Melanie Quitschau for valu-able discussions on the reactivity of protoberberines. Financialsupport from the Swiss National Science Foundation (Projects31600-113109 and 205320-126888/1), the Steinegg-Stiftung,Herisau, the Fonds zur Förderung von Lehre und Forschung, Basel(M.H.), and the FWF Project P2239-B11 (S.H.) is gratefully ac-knowledged.
References1 Hancox JC, McPate MJ, El Harchi A, Zhang YH. The hERG potassium chan-nel and hERG screening for drug-induced torsades de pointes. Pharma-col Therapeut 2008; 119: 118–132
2 Raschi E, Vasina V, Poluzzi E, De Ponti F. The hERG K+ channel: targetand antitarget strategies in drug development. Pharmacol Res 2008;57: 181–195
3 Deisemann H, Ahrens N, Schlobohm I, Kirchhoff C, Netzer R, Möller C. Ef-fects of common antitussive drugs on the hERG potassium channel cur-rent. J Cardiovasc Pharm 2008; 52: 494–499
4 Polak S, Wisniowska B, Brandys J. Collation, assessment and analysis ofliterature in vitro data on hERG receptor blocking potency for subse-quent modeling of drugsʼ cardiotoxic properties. J Appl Toxicol 2009;29: 183–206
5 Scholz EP, Zitron E, Kiesecker C, Lück S, Thomas D, Kathöfer S, Kreye VAW,Katus HA, Kiehn J, Schoels W, Karle CA. Inhibition of cardiac HERG chan-nels by grapefruit flavonoid naringenin: implications for the influenceof dietary compounds on cardiac repolarisation. Naunyn Schmiede-bergs Arch Pharmacol 2005; 371: 516–525
6 Lin CR, Ke XG, Ranade V, Somberg J. The additive effects of the activecomponent of grapefruit juice (naringenin) and antiarrhythmic drugson HERG inhibition. Cardiology 2008; 110: 145–152
7 Zitron E, Scholz E, Owen RW, Lück S, Kiesecker C, Thomas D, Kathöfer S,Niroomand F, Kiehn J, Kreye VAW, Katus HA, Schoels W, Karle CA. QTcprolongation by grapefruit juice and its potential pharmacological ba-sis – HERG channel blockade by flavonoids. Circulation 2005; 111:835–838
8 Baburin I, Beyl S, Hering S. Automated fast perfusion of Xenopus oo-cytes for drug screening. Pflugers Arch Eur J Physiol 2006; 453: 117–123
9 Kim HJ, Baburin I, Khom S, Hering S, Hamburger M. HPLC-based activityprofiling approach for the discovery of GABA(A) receptor ligands usingan automated two microelectrode voltage clamp assay on Xenopus oo-cytes. Planta Med 2008; 74: 521–526
10 Zaugg J, Baburin I, Strommer B, Kim HJ, Hering S, Hamburger M. HPLC-based activity profiling: discovery of piperine as a positive GABA(A) re-ceptor modulator targeting a benzodiazepine-independent bindingsite. J Nat Prod 2010; 73: 185–191
11 Stöger EA, Friedl F. Arzneibuch der Chinesischen Medizin, German edi-tion. Stuttgart: Deutscher Apotheker Verlag; 2009
12 Tang W, Eisenbrand G. Chinese drugs of plant origin. Berlin: Springer-Verlag; 1992: 361–371
13 Ma BL, Ma YM, Shi R, Wang TM, Zhang N, Wang CH, Yang Y. Identifica-tion of the toxic constituents in Rhizoma Coptidis. J Ethnopharmacol2010; 128: 357–364
14 KongWJ, Zhao YL, Xiao XH, Jin C, Li ZL.Quantitative and chemical finger-print analysis for quality control of rhizoma Coptidis chinensis basedon UPLC‑PAD combined with chemometrics methods. Phytomedicine2009; 16: 950–959
15 Ernst E. Harmless herbs? A review of the recent literature. Am J Med1998; 104: 170–178
16 Potterat O, Hamburger M. Natural products in drug discovery – Con-cepts and approaches for tracking bioactivity. Curr Org Chem 2006;10: 899–920
17 Stork D, Timin EN, Berjukow S, Huber C, Hohaus A, Auer M, Hering S.State dependent dissociation of HERG channel inhibitors. Br J Pharma-col 2007; 151: 1368–1376
18 Rodriguez-Menchaca A, Ferrer-Villada T, Lara J, Fernandez D, Navarro-Polanco RA, Sanchez-Chapula JA. Block of hERG channels by berberine:mechanisms of voltage- and state-dependence probed with site-di-rected mutant channels. J Cardiovasc Pharm 2006; 47: 21–29
19 Li BX, Yang BF, Zhou J, Xu CQ, Li YR. Inhibitory effects of berberine on I-K1, I-K, and HERG channels of cardiac myocytes. Acta Pharmacol Sin2001; 22: 125–131
20 Brewer ARE. Leuckart-Wallach reaction. In: Li JJ, Corey EJ, editors. Namereactions of functional group transformations. Hoboken, New Jersey:John Wiley & Sons, Inc.; 2007: 451–455
21 Lukasiewicz A. A study of the mechanism of certain chemical reactions—I: The mechanism of the leuckart-wallach reaction and of the reduc-tion of schiff bases by formic acid. Tetrahedron 1963; 19: 1789–1799
22 Kondo Y, Imai J, Inoue H. Reaction of protoberberine-type alkaloids.Part 12: a facile method for regiospecific oxygenation and excited oxi-dative ring-cleavage of berberine alkaloids. J Chem Soc [Perkin I] 1980:911–918
23 Wang YX, Zheng YM. Ionic mechanism responsible for prolongation ofcardiac action-potential duration by berberine. J Cardiovasc Pharm1997; 30: 214–222
24 Bensky D, Clavey S, Stöger E. Chinese herbal medicine –materia medica,3rd edition. Seattle: Eastland Press, Inc.; 2004: 134–138
25 Turner N, Li JY, Gosby A, To SWC, Cheng Z, Miyoshi H, Taketo MM, CooneyGJ, Kraegen EW, James DE, Hu LH, Li J, Ye JM. Berberine and its more bio-logically available derivative, dihydroberberine, inhibit mitochondrialrespiratory complex I – A mechanism for the action of berberine to ac-tivate AMP-activated protein kinase and improve insulin action. Diabe-tes 2008; 57: 1414–1418
26 Ye MZ, Fu S, Pi RB, He F. Neuropharmacological and pharmacokineticproperties of berberine: a review of recent research. J Pharm Pharma-col 2009; 61: 831–837
27 Zuo F, Nakamura N, Akao T, Hattori M. Pharmacokinetics of berberineand its main metabolites in conventional and pseudo germ-free ratsdetermined by liquid chromatography/ion trap mass spectrometry.Drug Metab Dispos 2006; 34: 2064–2072
28 Hua WY, Ding L, Chen Y, Gong B, He JC, Xu GL. Determination of berber-ine in human plasma by liquid chromatography-electrospray ioniza-tion-mass spectrometry. J Pharm Biomed Anal 2007; 44: 931–937
29 Yang YH, Kang N, Xia HJ, Li J, Chen LX, Qiu F.Metabolites of protoberber-ine alkaloids in human urine following oral administration of coptidisRhizoma decoction. Planta Med 2010; 76: 1859–1863
30 Qiu F, Zhu ZY, Kang N, Piao SJ, Qin GY, Yao XS. Isolation and identifica-tion of urinary metabolites of berberine in rats and humans. DrugMetab Dispos 2008; 36: 2159–2165
31 Tsai PL, Tsai TH. Hepatobiliary excretion of berberine. Drug Metab Dis-pos 2004; 32: 405–412
32 Cheng Z, Chen AF, Wu F, Sheng L, Zhang HK, GuM, Li YY, Zhang LN, Hu LH,Li JY, Li J. 8,8-Dimethyldihydroberberine with improved bioavailabilityand oral efficacy on obese and diabetic mouse models. Bioorg MedChem 2010; 18: 5915–5924
Schramm A et al. hERG Channel Inhibitors… Planta Med 2011; 77: 692–697




















