
c-Myb and p300 Regulate Hematopoietic Stem Cell Proliferation and Differentiation
Upload
independentCategory
view
7download
0
a PATTERNS & PHENOTYPES
Hematopoietic Prostaglandin D Synthase(H-Pgds) Is Expressed in the Early EmbryonicGonad and Participates to the Initial NuclearTranslocation of the SOX9 ProteinBrigitte Moniot,1 Andalib Farhat,1 Kosuke Aritake,2 Faustine Declosmenil,1 Serge Nef,3
Naomi Eguchi,2 Yoshihiro Urade,2 Francis Poulat,1† and Brigitte Boizet-Bonhoure1†*
In mammals, the Prostaglandin D2 (PGD2) signaling pathway is involved in male gonadal development,regulating Sox9 gene expression and SOX9 protein subcellular localization through lipocalin prostaglan-din D synthase (L-Pgds) activity. Nevertheless, because L-Pgds is downstream of Sox9, its expression can-not explain the initial nuclear translocation of the SOX9 protein. Here, we show that another source ofPGD2, hematopoietic-Pgds (H-Pgds) enzyme is expressed in somatic and germ cells of the embryonicgonad of both sexes, as early as embryonic day (E) 10.5, before the onset of L-Pgds expression. Inhibitionof H-Pgds activity by the specific HQL-79 inhibitor leads to impaired nuclear translocation of SOX9 pro-tein in E11.5 Sertoli cells. Furthermore, analysis of H-Pgds2/2 male embryonic gonads confirms abnormalsubcellular localization of SOX9 protein at the E11.5 early stage of mouse testicular differentiationsuggesting a role for H-Pgds-produced PGD2 in the initial nuclear translocation of SOX9. DevelopmentalDynamics 240:2335–2343, 2011. VC 2011 Wiley-Liss, Inc.
Key words: Pgds; prostaglandin D2; SOX9; Sertoli cell; subcellular localization
Accepted 3 August 2011
INTRODUCTION
Sex-specific regulation of gonad orga-nogenesis is the result of the cell-au-tonomous response of some cells tothe sex-determining switch SRYdirectly activating Sox9 expression(Hacker et al., 1995). In mouse, theexpression of Sry in the male genitalridge occurs only in a very narrowtime window at �11.5 dpc (Hackeret al., 1995) and in order to achievethe threshold level of Sox9 transcript
required to Sertoli cell differentiationand gonad morphogenesis, positive-feedback pathways involving Sox9gene expression combined with eitherFGF9 signaling (Kim et al., 2006) orwith high-glucose metabolic statethrough an extracellular matrix(ECM; Hiramatsu et al., 2009) areinvolved. In addition to these, the lip-ocalin-prostaglandin D synthase (L-Pgds) gene through production ofprostaglandin D2 (PGD2) forms anamplification loop with the Sox9 gene
whereby SOX9 initiates and maintainsL-Pgds gene transcription and PGD2
participates in the maintenance ofSox9 expression during Sertoli cell dif-ferentiation (Wilhelm et al., 2007b;Moniot et al., 2009).The L-Pgds gene is male specifically
expressed at early stages of mousegonadogenesis (Adams and McLaren,2002) and L-Pgds expression isdescribed as a dynamic wave-likeexpression pattern closely resemblingthose of Sry and Sox9 in the
Dev
elop
men
tal D
ynam
ics
1Institut de Genetique Humaine, Department of Genetic and Development, Montpellier, France2Department of Molecular Behavioral Biology, Osaka Bioscience Institute, Osaka, Japan3Department of Genetic Medicine and Development, University of Geneva Medical School, Geneva, SwitzerlandGrant sponsor: CNRS; the Agence Nationale pour la Recherche; Grant number: ANR-07-BLAN-00440; Grant sponsor: thePNRREndocrinology INSERM; the Ligue Regionale contre le Cancer.†Drs. Poulat and Boizet-Bonhoure equally contributed to this work.*Correspondence to: Brigitte Boizet-Bonhoure, IGH CNRS URP1142 Department of Genetic and Development, 141, rue dela Cardonille, 34396 Montpellier Cedex5. E-mail: [email protected]
DOI 10.1002/dvdy.22726Published online 1 September 2011 in Wiley Online Library (wileyonlinelibrary.com).
DEVELOPMENTAL DYNAMICS 240:2335–2343, 2011
VC 2011 Wiley-Liss, Inc.

embryonic testis, starting at 17 tailsomite stage (ts), shortly after theonset of Sox9 expression (Wilhelmet al., 2007a). We recently observedthat L-Pgds expression is abolished inE11.5-E12.0 male Sox9�/� gonads(Ck19-Cre; Sox9flox/flox mice), showingthat SOX9 is required for the initia-tion of L-Pgds gene expression. More-over, ablation of Sox9 after the onsetof L-Pgds expression (E13.5-E14.5Amh-Cre; Sox9flox/flox) also induces astrong down-regulation of L-Pgdstranscript expression, demonstratingthe requirement for SOX9 protein inthe maintenance of L-Pgds geneexpression in embryonic Sertoli cells.In vitro luciferase assays and chroma-tin immunoprecipitation experimentsperformed on embryonic gonads haveshown that SOX9 can directly bindand activate the L-Pgds promoter(Wilhelm et al., 2007a).
L-Pgds�/� mutant XY gonadsexhibited a delay in Sertoli cell differ-entiation up to E13.5 (Moniot et al.,2009) which was associated with areduced level of Sox9 gene expressionand subsequent Amh gene expression,thereby confirming in vivo that PGD2
increases the transcription of theSox9 gene in mouse (Wilhelm et al.,2005). PGD2 also has an effect at theposttranslational level on nucleartranslocation of the SOX9 protein(Malki et al., 2005b) that occurs spe-cifically in the male gonad. Upon sexdetermination, SOX9 nuclear translo-cation is strongly stimulated by PGD2
signaling, transduced by the cAMP/PKA pathway through the PGD2 re-ceptor DP1 (Malki et al., 2005b) and adelay in the nuclear translocation ofSOX9 in the L-Pgds�/� mutant XYgonads confirms the involvement ofPGD2 in this process (Moniot et al.,2009).
All together, these experimentsshowed that the onset and mainte-nance of L-Pgds expression is com-pletely dependent on Sox9 by lateE11.5 and that PGD2, in addition toregulating the Sox9 transcription,regulates the subcellular localizationof SOX9 (Malki et al., 2005a,b; Moniotet al., 2009). This PGD2 pathway,with the FGF9 signaling pathway(Kim et al., 2006) and with a high-glu-cose metabolic state (Hiramatsuet al., 2009), define feedforward path-ways of Sox9 gene expression,
towards the maintenance of Sox9transcription and subsequently forSertoli cell differentiation and testiscord formation.
To explain the onset of SOX9 nu-clear translocation at early E11.5, af-ter initial up-regulation by SRY(Sekido and Lovell-Badge, 2008), wehypothesize that an earlier PGD2 sig-nal could be present within the gonad.Indeed, in addition to L-Pgds, a sec-ond enzyme called hematopoieticPGD synthase (H-Pgds) or GSH-requiring enzyme, mediates the finalregulatory step of PGD2 biosynthesisby catalyzing the conversion of PGH2
into PGD2. H-Pgds is a cytosolic pro-tein responsible for the biosynthesisof PGD2 in immune and inflammatorycells such as mast or Th2 cells and isalso expressed in the spleen, thymus,skin and liver (Kanaoka and Urade,2003) and in the uterine epithelium(Michimata et al., 2002). H-Pgdsexpression, together with the prosta-glandin biosynthetic pathway, wasfound in the human adult testis(Schell et al., 2007). Whereas L-Pgdsexpression in Sertoli and germ cells,has been largely documented (Adamsand McLaren, 2002; Wilhelm et al.,2007b; Moniot et al., 2009), this is notthe case for the precise H-Pgdsexpression profile in the embryonicgonad.
In this study, we found that thePGD2-producing H-Pgds enzyme isexpressed in the embryonic gonad atmid E11.5 (16-17 Ts, tail somites), at atime where L-Pgds remains unex-pressed. Surprisingly, the H-Pgdstranscript is expressed at a similarlevel in both the somatic and germcells of both sexes. Abrogation of H-Pgds activity in HQL-79 treated XYgonads or H-Pgds�/� XY gonadsimpairs nuclear translocation of SOX9protein in E11.5 pre-Sertoli, suggest-ing that an initial H-Pgds-mediatedPGD2 signal could participate to theSOX9 nuclear translocation towardSertoli cell differentiation.
RESULTS AND DISCUSSION
Early Expression of H-Pgds
mRNA in the Embryonic
Gonad
To test whether a PGD2 signalremained in the early gonad in ab-
sence of L-Pgds, we measured levelsof PGD2 in E12.5 L-Pgdsþ/þ and �/�
XY and XX differentiating gonadsusing enzyme-linked immunosorbentassay (ELISA), at a time at which wecan distinguish between L-Pgds andH-Pgds produced PGD2. Significantlevels of PGD2 were found in the L-Pgds�/� XY and þ/þ,�/� XX gonadscompared with the control assay (Fig.1A). Particularly, PGD2 levels in theL-Pgds�/� XY gonads were signifi-cantly reduced by 50% compared withthose of L-Pgdsþ/þ XY gonads (Fig.1A), indicating that, even in the ab-sence of L-Pgds, significant levels ofPGD2 remained in the male gonadand were detected in the femalegonad. Moreover the resulting con-centration of PGD2 in male L-Pgds�/�
gonads is not significantly differentfrom L-Pgdsþ/þ female gonads, sug-gesting that the potential secondsource of PGD2 expressed in the fetalgonads would not be sexually dimor-phic. Indeed, we found that the H-Pgds transcript was expressed at sim-ilar levels in male and female allalong the sex-determining period(E10.5 to E12.5) in SF-1-expressingcells purified as previously described(Nef et al., 2005; Fig. 1B). In addition,the level of H-Pgds mRNA did not sig-nificantly differ between L-Pgds�/�
and L-Pgdsþ/þ E12.5 XY gonads (Fig.1C) and at the same stage L-Pgdsexpression was not modified in H-Pgds�/� XY gonads (data not shown),suggesting that each Pgds expressionis independent on the other.From these results, we conclude
that a significant amount of PGD2 isproduced in gonads of both sexes atearly stages of sexual determination.However, this H-Pgds produced PGD2
is not sufficient to induce Sox9 geneexpression as previously observedwith PGD2 treatment of XX gonadalexplants (Wilhelm et al., 2005). Onehypothesis can explain this observa-tion: in contrast to L-Pgds, H-Pgds isnot a secreted enzyme, suggestingthat PGD2 produced by H-Pgds mightbe less available for the PGD2 recep-tor. Moreover, the Sox9 gene probablyrequired higher concentrations ofPGD2 only available when the L-Pgdsenzyme is expressed.We then performed in situ hybrid-
ization (ISH) experiments for H-PgdsmRNA expression on male and female
Dev
elop
men
tal D
ynam
ics
2336 MONIOT ET AL.

gonads at mid E11.5 (16-17 Ts, [Tailsomites]), E13.5 and E17.5. On thesame slides and following this ISHexperiment, immunofluorescencestaining using SOX9 as the Sertolicell marker in male gonads or using
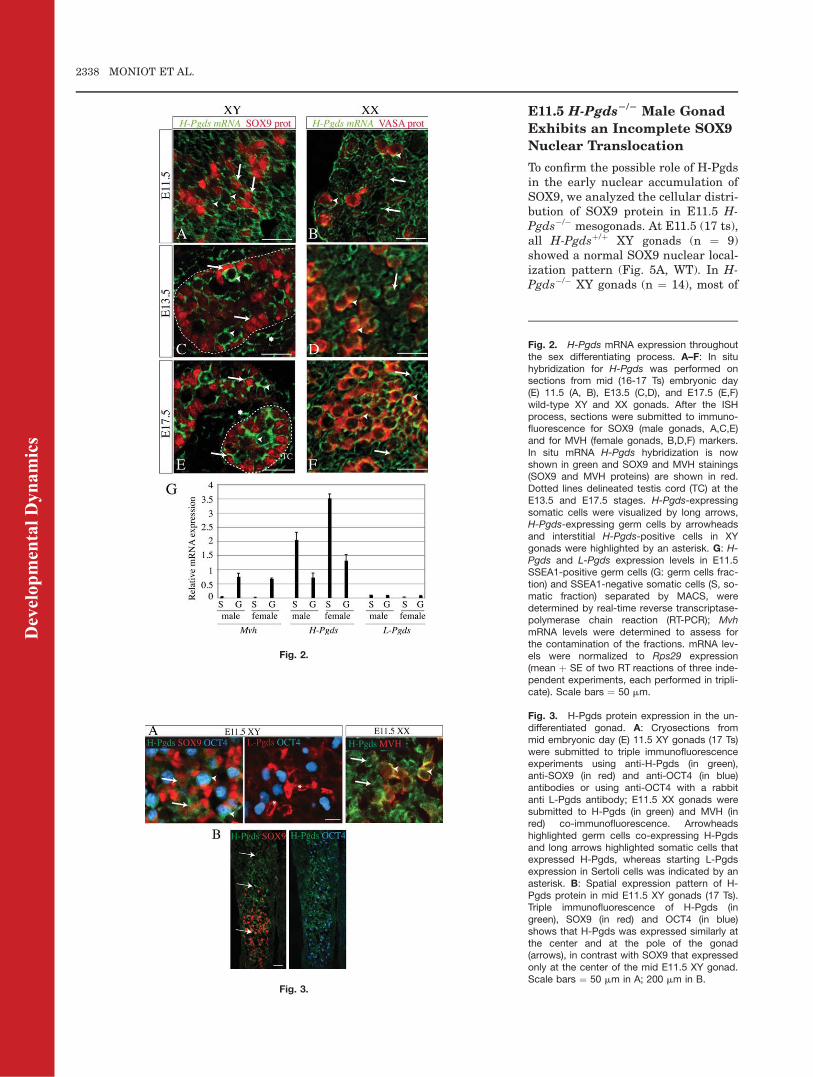
MVH (mouse VASA homolog) as thegerm cell marker in female gonads,was carried out. These experimentsrevealed that H-Pgds mRNA wasexpressed in the somatic and germcells of the early developing gonad ofboth sexes, at the time when thegonad is still in a bipotential and un-differentiated state (at mid E11.5;Fig. 2A,B). Later, at the E13.5 andE17.5 late embryonic stages, H-PgdsmRNA is expressed within both Ser-toli cells and germ cells in the testiscords and in the interstitial compart-ment in XY gonads (Fig. 2C,E) andwithin both the germ cells and so-matic compartment in XX gonads(Fig. 2D,F). In male gonads, not allSOX9-positive cells are expressing H-Pgds, suggesting that this expressiondepends on intrinsic factors of Sertolicells; however, this expression level ishigher than that in interstitial (Ley-dig) cells.
By purifying somatic and germ cellfractions by immunomagnetic cellsorting, using the anti-SSEA1 anti-body, we confirmed this H-Pgdsexpression pattern in E11.5 mesogo-nads. In both sexes, the SSEA1-posi-tive germ cells fraction expressing thegerm cell marker Mvh, also expressedH-Pgds (Fig. 2G). H-Pgds mRNA wasalso detected in the SSEA1-negativesomatic cells that do not express Mvh(Fig. 2G). At this early stage (E11.5:16-17 Ts), we thus confirm that H-Pgds is expressed in both somaticand germ cells compartments in maleand female, at the time when L-Pgdsis not yet expressed in the male(Fig. 2G).
H-Pgds Protein Is Expressed
in the E11.5 Embryonic
Gonad
However, our experiments cannotruled out that a sex specific transla-tional regulation of H-Pgds could takeplace during gonadal differentiation.To assess the expression of the H-Pgdsprotein, we used an antibody directedagainst H-Pgds whose specificity hasbeen verified previously (Farhat et al.,2011) and performed co-staining withthe OCT4 (in male) or MVH (infemale) germ cell markers and withthe SOX9 Sertoli cell marker. Thisexperiment confirmed that H-Pgds is
expressed in somatic and germ cell lin-eages of both sexes at E11.5, whereasL-Pgds starts to be expressed in themale gonad (Fig. 3A). Furthermore, atmid E11.5 (17 Ts), expression of H-Pgds protein was similar at the centerand at the pole of the male gonad (Fig.3B), suggesting that H-Pgds was notexpressed as a dynamic wave-like asreported for L-Pgds (Wilhelm et al.,2007a) and, thus, was not up-regu-lated by the sex determining factorsSRYand SOX9.
Initial Role of H-Pgds-
Produced PGD2 in SOX9
Subcellular Localization
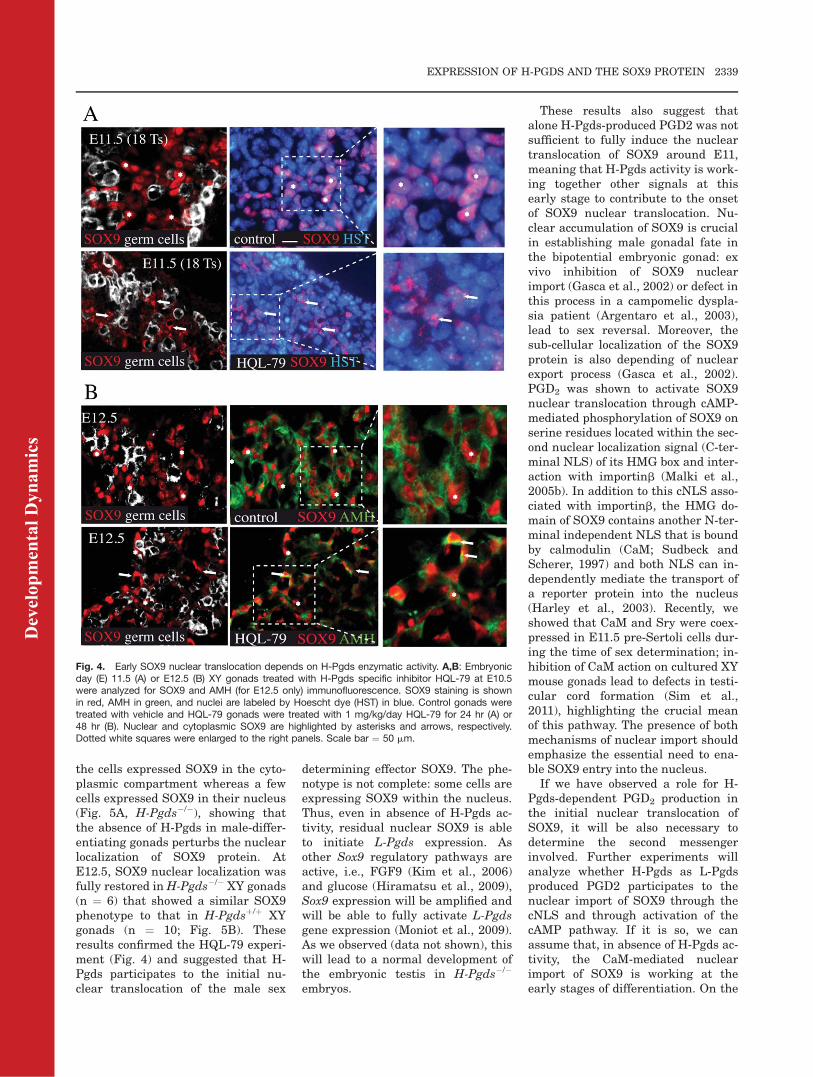
As described in our previous studies(de Santa Barbara et al., 2000; Gascaet al., 2002), the SOX9 protein is cyto-plasmic before testicular determina-tion and is translocated into the nu-cleus while Sry is expressed. L-Pgds-produced PGD2 does not explain thisnuclear translocation because L-Pgdsis downstream of Sox9 in the mousesex determining cascade, suggestingthat another mechanism is requiredto import SOX9 in the nucleus duringearly testicular determination. Tostudy the role of H-Pgds in the nu-clear translocation of SOX9, we orallyadministered the H-Pgds inhibitorHQL-79 (Aritake et al., 2006) to preg-nant mice at the E10.5 developmentalstage, or at E10.5 and E11.5 and ana-lyzed XY gonads at E11.5 (18-20 Ts)or E12.5, respectively. This HQL-79inhibitor has been shown to be spe-cific to H-Pgds and to exactly mimicthe phenotype of H-Pgds depletion(Mohri et al., 2006), without any side-effect except an increase of the PGE2content in the lungs of sensitizedguinea pigs (Matsushita et al., 1998).Daily administration of 0.1, 1 or 10mg/kg/day of HQL-79 led to similarresults: whereas SOX9 was fully nu-clear in the untreated gonads atE11.5, it remained mainly within thecytoplasm of HQL-79 treated gonads(Fig. 4A). At E12.5, while SOX9remained local to the cytoplasm insome cells, the majority of SOX9staining was nuclear (Fig. 4B), sug-gesting that initiation of SOX9 nu-clear translocation was impairedupon inhibition of H-Pgds enzymaticactivity but recovered after E12.5.
Fig. 1. PGD2 levels and H-Pgds expressionin the undifferentiated gonad. A: Levels ofPGD2 were measured in embryonic day (E)12.5 L-Pgds XY and XX þ/þ, �/� gonads (with-out mesonephros) using enzyme-linked immu-nosorbent assay (ELISA). Results wereexpressed in pg PGD2 per gonad; error barsindicate SD of assays (3 pools of 16 gonads)done on two duplicates of two independentdilutions and read five times (at 412 nm). Anasterisk indicates significant differences ofPGD2 levels compared with the XYþ/þ PGD2
level (P-value < 0.01). Control assay (C) wasthe negative control (NBS, nonspecific bindingpoint) of the Elisa kit. B: H-Pgds expressionlevels in E10.5, E11.5, and E12.5 SF-1-posi-tive gonadal cells, were determined by real-time reverse transcriptase-polymerase chainreaction (RT-PCR). H-Pgds mRNA levels werenormalized to Gapdh expression (mean þ SEof two independent experiments, each per-formed in triplicate). m ¼ male; f ¼ female. C:H-Pgds expression levels in E12.5 L-PgdsXYþ/þ, �/� and XXþ/þ, �/� gonads were deter-mined by real-time RT-PCR and normalized toRps29 expression. Individual experimentswere performed on RNA obtained from pooledgonads (without mesonephros) from each lit-termate (n ¼ 3). Errors bars indicate SD ofassays done twice from two RT reactions, intriplicate.
Dev
elop
men
tal D
ynam
ics
EXPRESSION OF H-PGDS AND THE SOX9 PROTEIN 2337
E11.5 H-Pgds2/2 Male Gonad
Exhibits an Incomplete SOX9
Nuclear Translocation
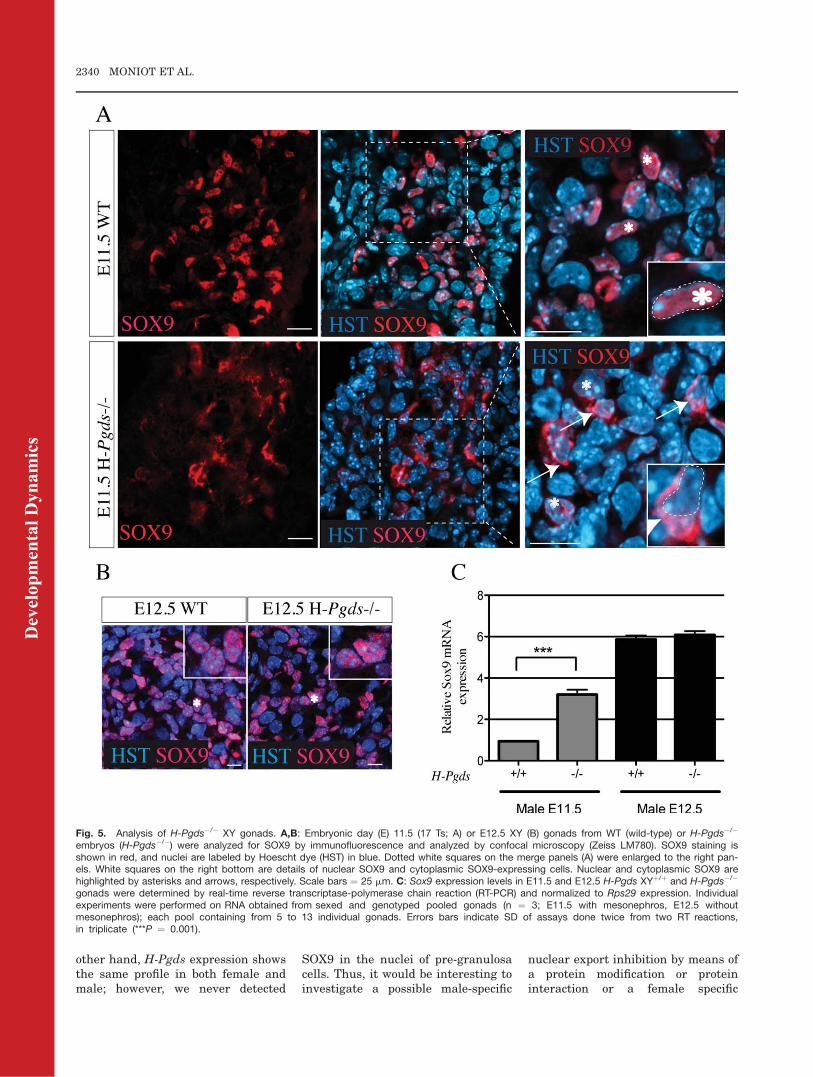
To confirm the possible role of H-Pgdsin the early nuclear accumulation ofSOX9, we analyzed the cellular distri-bution of SOX9 protein in E11.5 H-Pgds�/� mesogonads. At E11.5 (17 ts),all H-Pgdsþ/þ XY gonads (n ¼ 9)showed a normal SOX9 nuclear local-ization pattern (Fig. 5A, WT). In H-Pgds�/� XY gonads (n ¼ 14), most of
Fig. 2.
Fig. 2. H-Pgds mRNA expression throughoutthe sex differentiating process. A–F: In situhybridization for H-Pgds was performed onsections from mid (16-17 Ts) embryonic day(E) 11.5 (A, B), E13.5 (C,D), and E17.5 (E,F)wild-type XY and XX gonads. After the ISHprocess, sections were submitted to immuno-fluorescence for SOX9 (male gonads, A,C,E)and for MVH (female gonads, B,D,F) markers.In situ mRNA H-Pgds hybridization is nowshown in green and SOX9 and MVH stainings(SOX9 and MVH proteins) are shown in red.Dotted lines delineated testis cord (TC) at theE13.5 and E17.5 stages. H-Pgds-expressingsomatic cells were visualized by long arrows,H-Pgds-expressing germ cells by arrowheadsand interstitial H-Pgds-positive cells in XYgonads were highlighted by an asterisk. G: H-Pgds and L-Pgds expression levels in E11.5SSEA1-positive germ cells (G: germ cells frac-tion) and SSEA1-negative somatic cells (S, so-matic fraction) separated by MACS, weredetermined by real-time reverse transcriptase-polymerase chain reaction (RT-PCR); MvhmRNA levels were determined to assess forthe contamination of the fractions. mRNA lev-els were normalized to Rps29 expression(mean þ SE of two RT reactions of three inde-pendent experiments, each performed in tripli-cate). Scale bars ¼ 50 mm.
Fig. 3.
Fig. 3. H-Pgds protein expression in the un-differentiated gonad. A: Cryosections frommid embryonic day (E) 11.5 XY gonads (17 Ts)were submitted to triple immunofluorescenceexperiments using anti-H-Pgds (in green),anti-SOX9 (in red) and anti-OCT4 (in blue)antibodies or using anti-OCT4 with a rabbitanti L-Pgds antibody; E11.5 XX gonads weresubmitted to H-Pgds (in green) and MVH (inred) co-immunofluorescence. Arrowheadshighlighted germ cells co-expressing H-Pgdsand long arrows highlighted somatic cells thatexpressed H-Pgds, whereas starting L-Pgdsexpression in Sertoli cells was indicated by anasterisk. B: Spatial expression pattern of H-Pgds protein in mid E11.5 XY gonads (17 Ts).Triple immunofluorescence of H-Pgds (ingreen), SOX9 (in red) and OCT4 (in blue)shows that H-Pgds was expressed similarly atthe center and at the pole of the gonad(arrows), in contrast with SOX9 that expressedonly at the center of the mid E11.5 XY gonad.Scale bars ¼ 50 mm in A; 200 mm in B.
Dev
elop
men
tal D
ynam
ics
2338 MONIOT ET AL.
the cells expressed SOX9 in the cyto-plasmic compartment whereas a fewcells expressed SOX9 in their nucleus(Fig. 5A, H-Pgds�/�), showing thatthe absence of H-Pgds in male-differ-entiating gonads perturbs the nuclearlocalization of SOX9 protein. AtE12.5, SOX9 nuclear localization wasfully restored in H-Pgds�/� XY gonads(n ¼ 6) that showed a similar SOX9phenotype to that in H-Pgdsþ/þ XYgonads (n ¼ 10; Fig. 5B). Theseresults confirmed the HQL-79 experi-ment (Fig. 4) and suggested that H-Pgds participates to the initial nu-clear translocation of the male sex
determining effector SOX9. The phe-notype is not complete: some cells areexpressing SOX9 within the nucleus.Thus, even in absence of H-Pgds ac-tivity, residual nuclear SOX9 is ableto initiate L-Pgds expression. Asother Sox9 regulatory pathways areactive, i.e., FGF9 (Kim et al., 2006)and glucose (Hiramatsu et al., 2009),Sox9 expression will be amplified andwill be able to fully activate L-Pgdsgene expression (Moniot et al., 2009).As we observed (data not shown), thiswill lead to a normal development ofthe embryonic testis in H-Pgds�/�
embryos.
These results also suggest thatalone H-Pgds-produced PGD2 was notsufficient to fully induce the nucleartranslocation of SOX9 around E11,meaning that H-Pgds activity is work-ing together other signals at thisearly stage to contribute to the onsetof SOX9 nuclear translocation. Nu-clear accumulation of SOX9 is crucialin establishing male gonadal fate inthe bipotential embryonic gonad: exvivo inhibition of SOX9 nuclearimport (Gasca et al., 2002) or defect inthis process in a campomelic dyspla-sia patient (Argentaro et al., 2003),lead to sex reversal. Moreover, thesub-cellular localization of the SOX9protein is also depending of nuclearexport process (Gasca et al., 2002).PGD2 was shown to activate SOX9nuclear translocation through cAMP-mediated phosphorylation of SOX9 onserine residues located within the sec-ond nuclear localization signal (C-ter-minal NLS) of its HMG box and inter-action with importinb (Malki et al.,2005b). In addition to this cNLS asso-ciated with importinb, the HMG do-main of SOX9 contains another N-ter-minal independent NLS that is boundby calmodulin (CaM; Sudbeck andScherer, 1997) and both NLS can in-dependently mediate the transport ofa reporter protein into the nucleus(Harley et al., 2003). Recently, weshowed that CaM and Sry were coex-pressed in E11.5 pre-Sertoli cells dur-ing the time of sex determination; in-hibition of CaM action on cultured XYmouse gonads lead to defects in testi-cular cord formation (Sim et al.,2011), highlighting the crucial meanof this pathway. The presence of bothmechanisms of nuclear import shouldemphasize the essential need to ena-ble SOX9 entry into the nucleus.If we have observed a role for H-
Pgds-dependent PGD2 production inthe initial nuclear translocation ofSOX9, it will be also necessary todetermine the second messengerinvolved. Further experiments willanalyze whether H-Pgds as L-Pgdsproduced PGD2 participates to thenuclear import of SOX9 through thecNLS and through activation of thecAMP pathway. If it is so, we canassume that, in absence of H-Pgds ac-tivity, the CaM-mediated nuclearimport of SOX9 is working at theearly stages of differentiation. On the
Fig. 4. Early SOX9 nuclear translocation depends on H-Pgds enzymatic activity. A,B: Embryonicday (E) 11.5 (A) or E12.5 (B) XY gonads treated with H-Pgds specific inhibitor HQL-79 at E10.5were analyzed for SOX9 and AMH (for E12.5 only) immunofluorescence. SOX9 staining is shownin red, AMH in green, and nuclei are labeled by Hoescht dye (HST) in blue. Control gonads weretreated with vehicle and HQL-79 gonads were treated with 1 mg/kg/day HQL-79 for 24 hr (A) or48 hr (B). Nuclear and cytoplasmic SOX9 are highlighted by asterisks and arrows, respectively.Dotted white squares were enlarged to the right panels. Scale bar ¼ 50 mm.
Dev
elop
men
tal D
ynam
ics
EXPRESSION OF H-PGDS AND THE SOX9 PROTEIN 2339
other hand, H-Pgds expression showsthe same profile in both female andmale; however, we never detected
SOX9 in the nuclei of pre-granulosacells. Thus, it would be interesting toinvestigate a possible male-specific
nuclear export inhibition by means ofa protein modification or proteininteraction or a female specific
Fig. 5. Analysis of H-Pgds�/� XY gonads. A,B: Embryonic day (E) 11.5 (17 Ts; A) or E12.5 XY (B) gonads from WT (wild-type) or H-Pgds�/�
embryos (H-Pgds�/�) were analyzed for SOX9 by immunofluorescence and analyzed by confocal microscopy (Zeiss LM780). SOX9 staining isshown in red, and nuclei are labeled by Hoescht dye (HST) in blue. Dotted white squares on the merge panels (A) were enlarged to the right pan-els. White squares on the right bottom are details of nuclear SOX9 and cytoplasmic SOX9-expressing cells. Nuclear and cytoplasmic SOX9 arehighlighted by asterisks and arrows, respectively. Scale bars ¼ 25 mm. C: Sox9 expression levels in E11.5 and E12.5 H-Pgds XYþ/þ and H-Pgds�/�
gonads were determined by real-time reverse transcriptase-polymerase chain reaction (RT-PCR) and normalized to Rps29 expression. Individualexperiments were performed on RNA obtained from sexed and genotyped pooled gonads (n ¼ 3; E11.5 with mesonephros, E12.5 withoutmesonephros); each pool containing from 5 to 13 individual gonads. Errors bars indicate SD of assays done twice from two RT reactions,in triplicate (***P ¼ 0.001).
Dev
elop
men
tal D
ynam
ics
2340 MONIOT ET AL.

protein that would retain SOX9within the cytoplasm in femalegonads.
Sox9 Expression in E11.5
H-Pgds2/2 Male Gonad
We then analyzed the level of Sox9transcripts in E11.5 H-Pgds�/� andwt male gonads and surprisingly, wefound that Sox9 mRNA levels weresignificantly increased in the male H-Pgds�/� gonads (Fig. 5C). However, atE12.5, Sox9 (Fig. 5C) and L-Pgds(data not shown) expression levelswere similar in both mutant and wild-type XY gonads. This observation isin discrepancy with previouslyreported results showing that PGD2
activates Sox9 gene expression (Wil-helm et al., 2007a), which in turn willactivate L-Pgds expression (Moniotet al., 2009); one explanation could bethat H-Pgds produced intra-cytoplas-mic PGD2 that could have cellulartargets different from the DP1 or DP2receptors activated by the secreted L-Pgds enzyme. Because at E11.5, Sox9is directly activated by Sry (Sekidoand Lovell-Badge, 2008), we can alsohypothesize that the loss of H-Pgdsexpression in male embryonic gonadsup-regulates the level of Sry tran-scripts; in this case, H-Pgds producedPGD2 would be a negative modulatorof the Sry gene expression. As a mat-ter of fact, Sry gene expression isdown-regulated by cAMP signalingpathway in the human NT2D1 cellline (de Santa Barbara et al., 2001)and in mouse cultured Sertoli cells(Rossi et al., 1993). PKA phosphoryla-tion of the Steroidogenic factor SF-1was shown to switch off Sry geneexpression through the loss of itsbinding to the Sry promoter and of itsinteraction with activator Sp1 (deSanta Barbara et al., 2001). The pri-mary effector of this cAMP stimula-tion that remained to be determinedmight be H-Pgds-produced PGD2.The loss of H-Pgds activity in HQL-79treated or in H-Pgds�/� gonads willkeep the Sry activation on and willindirectly activate Sox9. Furtherexperiments will address the geneticrelationships of H-Pgds with Sox9and/or Sry. As Sox9 is expressed inthe mesonephros of C57BL/6J E11.5XY and XX gonads (17 Ts; Bullejosand Koopman, 2005), the gonadal
Sox9 signal might be hidden by thissignal from the mesonephros. Intro-duction of the Sf-1-Gfp transgenewithin the H-Pgds KO strain willovercome the contamination of theSox9-expressing cells from the meso-nephros and will allow to elucidatethese relationships at the transcrip-tional level.
All together, these data demon-strate that H-Pgds the second enzymeproducing PGD2 is expressed in theundifferentiated gonad in both sexesand in both somatic and germ cellscompartments, at the time when L-Pgds is not yet expressed in the male.They also suggest a role for H-Pgds-produced PGD2 in the initial nucleartranslocation of SOX9. The fact thatSox9 expression levels in E12.5 H-Pgds�/� XY gonads were similar tothose in wild-type gonads explainedthat normal testis developmentoccurred in the H-Pgds mutantembryos (data not shown), that nofemale-to-male sex reversal has beenobserved in H-Pgds�/� XX gonads(our unpublished observations andTrivedi et al., 2006), and that adultanimals are normally fertile. On theother hand, more studies are alsoneeded to elucidate the role of H-Pgdsproduced PGD2 in germ cells. Asgerm cells also express L-Pgds (Mon-iot et al., 2009), the impact of PGD2
signaling on the germ cell biology willbe study in the double knocked downmouse strain L- and H-Pgds.
EXPERIMENTAL
PROCEDURES
Animals and Treatments
Female C57BL/6J mice were pur-chased from Charles River Laborato-ries and H-Pgds KO mice in C57BL/6J genetic background were gener-ated at Osaka Bioscience Institute(Osaka, Japan; Mohri et al., 2006;Trivedi et al., 2006). Mice werehoused at the IGH animal care facil-ity under controlled environmentalconditions (12 hr light/12 hr darkness,temperature 21�C). Animal care andhandling conformed to the Reseau desAnimaleries de Montpellier (RAM)and all procedures were approved bythe Regional Ethic committee.
HQL-79 (4-benzhydryloxy-1-[3-(1H-tetrazol-5-yl)-propylpiperidine; Ari-
take et al., 2006), an inhibitor of H-Pgds activity, was purchased fromCayman Chemical (SpiBio, France).Daily oral administration of HQL-79(0.1, 1, or 10 mg/kg/day; Aritakeet al., 2006) was performed on preg-nant mice at E10.5 and E11.5 as men-tioned in the text. As a control, thesame volume of vehicle (0.5% methyl-cellulose) was orally administrated tomice during the same period.
In Situ Hybridization
The template for production of theantisense H-Pgds RNA probe and cov-ering the whole H-Pgds transcriptwas amplified by polymerase chainreaction (PCR) from embryonic mousecDNAs, cloned in a pCRII Topo vector(Invitrogen) and sequenced using anABI automatic sequencer. Digoxige-nin-labeled riboprobes were synthe-sized using a digoxigenin RNA label-ing kit, following the manufacturer’sinstructions (Roche Diagnostics) andused for in situ hybridization experi-ments as described earlier (Moniotet al., 2008, 2009). The specificity ofthe antisense H-Pgds RNA probe wasverified by hybridization with the con-trol sense H-Pgds RNA probe thatshowed no significant signal (Farhatet al., 2011).
Histochemical and
Immunofluorescence Staining
and Antibodies
Gonads were dissected from stagedembryos, fixed in 4% paraformalde-hyde and processed and cut into 10-mm-thick cryo-sections, as previouslydescribed (Gasca et al., 2002; Malkiet al., 2005b). For immunolabeling,slides were dried, rehydrated, rinsedin PBS, and incubated with differentantibodies (Gasca et al., 2002). SOX9was detected using a 1:200 dilution ofa rabbit polyclonal antibody raisedagainst the N-terminal part of thehuman protein (Notarnicola et al.,2006), L-Pgds using a rabbit anti-L-Pgds antibody (dilution 1/200; Moniotet al., 2009), and H-Pgds using a ratanti-H-Pgds antibody (Cayman10004349, dilution 1/200). Germ cellswere detected using a rabbit anti-MVH (mouse VASA homolog; Abcamab13840, dilution 1/300) or a mouseanti-OCT4 (BD Transduction
Dev
elop
men
tal D
ynam
ics
EXPRESSION OF H-PGDS AND THE SOX9 PROTEIN 2341

Laboratories, dilution 1/100). Appro-priate secondary antibodies (Alexa-Ig,Molecular Probe) were used (dilution1/800). For the immunofluorescenceexperiment consecutive to ISH experi-ment, ISH slides were rinsed in PBSafter the final step of the ISH process,were refixed in 4% paraformaldehydefor 2 hr and processed following theclassical immunofluorescence protocol(Gasca et al., 2002). Slides weremounted in FluorSave reagent andobserved with a fluorescent micro-scope (Leica DM6000) or with a confo-cal microscope (Laser ScanningMicroscope LSM780, Carl Zeiss).
Measurement of PGD2 Levels
To distinguish between L-Pgds and H-Pgds- produced PGD2, E12.5 gonadswere collected without mesonephroifrom L-Pgdsþ/þ, L-Pgds�/� XX andXY embryos, immediately frozen ondry ice and then stored at �80�C.Three sets of 16 gonads were pooledfor each condition, lysed, and proteinswere extracted with cold acetone onice. PGD2 levels were determinedusing the Prostaglandin D2– MOXExpress EIA Kit (Cayman Chemical500151) and normalized to total pro-tein levels using Micro BCA ProteinAssay Kit (Pierce 23235). Controlassay was the negative control (NSB,nonspecific binding assay) of the Elisakit. EIA was conducted once on twoduplicates of two independent dilu-tions of each sample. The assay wasread five times when the absorbanceof B0 (maximum binding of thetracer) wells was in the range of 0.3 to1. Statistical analysis was performedusing Student’s t-test, and resultswere considered statistically signifi-cant at a P < 0.05.
Embryo Dissection,
Immunomagnetic Germ Cell
Isolation, Reverse
Transcription, and Real-Time
PCR
During dissection, embryonic gonadswere separated from mesonephroi(E12.5 gonads) or not (E11.5 mesogo-nads) and conserved in RNAlater So-lution (Qiagen) until sex genotyping.Gonads were then pooled and RNAextraction was performed using the
RNeasy Mini Kit (Qiagen). Reversetranscription experiments were car-ried out using 500 ng of total RNApreviously treated with DNAse I(Promega), 250 ng of random primers(Invitrogen) and SuperScript III(Invitrogen), following manufacturerrecommendations.
On the other hand, amplified RNAsfrom E10.5, E11.5, and E12.5 Sf1-BAC-eGFP transgene-positive urogen-ital ridges (Stallings et al., 2002; Nefet al., 2005) were obtained and con-verted into double-stranded cDNA aspreviously described (Malki et al.,2005b). In summary, urogenital ridgesfrom individual embryos were dis-sected, digested with trypsin/ethylene-diaminetetraacetic acid (EDTA) andfiltered through a 40-mm cell strainerto generate single-cell suspensions.Green fluorescent protein (GFP) -posi-tive cells were sorted using a fluores-cence-activated cell sorting VantageSE with a purity of exceeding 97%.Cells were collected in 96-well platescontaining 50 ml of RNA lysis bufferfrom Qiagen RNeasy kit.
Sexed E11.5 mesogonads (n ¼ 100)from 9 littermates were pooled inthree different sets (n ¼ 3) and weredissociated as previously described(Guerquin et al., 2010). Germ cellswere isolated using magnetic sorting(column MSþ MACS, Miltenyi Bio-tech, Germany) and microbeads-linked Stage-Specific Embryonic Anti-gen-1 antibody (SSEA1, a marker ofundifferentiated germ cells; CD15),accordingly to the manufacturer (Mil-tenyi Biotech). The SSEA1-negativecell fraction containing the somaticcells was also recovered. Cells fromboth fractions were recovered by cen-trifugation, total mRNA wasextracted using the RNeasy Mini Kit(Qiagen) and submitted to RT reac-tion. The potential contamination ofSSEA1-negative cell fraction by germcells was assessed by real-time-quan-titative PCR (RT-qPCR) for Mvh; how-ever due to the early E11.5 stage ofthe dissected mesogonads, and due tothe fact that Sox9 is not sex-specifi-cally expressed in the mesonephros,we were unable to test the potentialcontamination of SSEA1-positive cellsby somatic cells.
Resulting cDNAs were then sub-jected to quantitative real-time PCRin a LightCycler 480 (Roche) appara-
tus using the QuantiTect SYBR GreenPCR Kit (Qiagen). Gene expressionlevels of Sox9, H-Pgds, and L-Pgdswere investigated using the followingpairs of primers (from 50 to 30): Sox9Forward, TCGGACACGGAGAACACC;Sox9 Reverse, GCACACGGGGAACTTATCTT; H-Pgds Forward: CACGCTGGATGACTTCATGT; H-Pgds Reverse:AATTCATTGAACATCCGCTCTT; L-Pgds Forward, GGCTCCTGGACACTACACCT; L-Pgds Reverse, ATAGTTGGCCTCCACCACTG. Quantificationwas performed using Gapdh orRps29 (Svingen et al., 2009) as refer-ence genes with the following primers:Gapdh Forward, TGGCAAAGTGGAGATTGTTGCC; Gapdh Reverse, AAGATGGTGATGGGCTTCCCG; Rps29Forward: TGAAGGCAAGATGGGTCAC;Rps29 Reverse: GCACATGTTCAGCCCGTATT. Each sample was measuredin triplicate in three separate experi-ments and quantification was per-formed using a second derivative cal-culation method with LC480 softwareversion 1.5 (Roche). Statistical analy-sis with PRISM5 software (GraphPadSoftware) was performed usingStudent’s t-test, and results wereconsidered statistically significant ata P < 0.05.
ACKNOWLEDGMENTSWe thank Dr. Julien Cau and the Im-agery platform of IGH for their help inimagery and analysis. We thank Dr.Massilva Rahmoun and Emily Kitty(AngloScribe) for critical reading ofthe manuscript. B.B.B. was funded bythe CNRS, by the Agence Nationalepour la Recherche, the PNRREndocri-nology INSERM, and the LigueRegionale contre le Cancer (BBB).
REFERENCES
Adams IR, McLaren A. 2002. Sexuallydimorphic development of mouse pri-mordial germ cells: switching fromoogenesis to spermatogenesis. Develop-ment 129:1155–1164.
Argentaro A, Sim H, Kelly S, Preiss S,Clayton A, Jans DA, Harley VR. 2003.A SOX9 defect of calmodulin-dependentnuclear import in campomelic dyspla-sia/autosomal sex reversal. J Biol Chem278:33839–33847.
Aritake K, Kado Y, Inoue T, Miyano M,Urade Y. 2006. Structural and func-tional characterization of HQL-79, anorally selective inhibitor of human he-matopoietic prostaglandin D synthase.
Dev
elop
men
tal D
ynam
ics
2342 MONIOT ET AL.

J Biol Chem 281:15277–15286.Bullejos M, Koopman P. 2005. Delayed
Sry and Sox9 expression in developingmouse gonads underlies B6-Y(DOM)sex reversal. Dev Biol 278:473–481.
de Santa Barbara P, Moniot B, Poulat F,Berta P. 2000. Expression and subcellu-lar localization of SF-1, SOX9, WT1,and AMH proteins during early humantesticular development. Dev Dyn 217:293–298.
de Santa Barbara P, Mejean C, Moniot B,Malcles MH, Berta P, Boizet-BonhoureB. 2001. Steroidogenic factor-1 contrib-utes to the cyclic-adenosine monophos-phate down-regulation of human SRYgene expression. Biol Reprod 64:775–783.
Farhat A, Philibert P, Sultan C, Poulat F,Boizet-Bonhoure B. 2011. Hematopoi-etic-Prostaglandin D2 synthase throughPGD2 production is involved in theadult ovarian physiology. J Ovarian Res4:3.
Gasca S, Canizares J, De Santa BarbaraP, Mejean C, Poulat F, Berta P, Boizet-Bonhoure B. 2002. A nuclear export sig-nal within the high mobility group do-main regulates the nucleocytoplasmictranslocation of SOX9 during sexualdetermination. Proc Natl Acad Sci U SA 99:11199–11204.
Guerquin MJ, Duquenne C, Lahaye JB,Tourpin S, Habert R, Livera G. 2010.New testicular mechanisms involved inthe prevention of fetal meiotic initiationin mice. Dev Biol 346:320–330.
Hacker A, Capel B, Goodfellow P, Lovell-Badge R. 1995. Expression of Sry, themouse sex determining gene. Develop-ment 121:1603–1614.
Harley VR, Layfield S, Mitchell CL, For-wood JK, John AP, Briggs LJ, McDo-wall SG, Jans DA. 2003. Defectiveimportin beta recognition and nuclearimport of the sex-determining factorSRY are associated with XY sex-revers-ing mutations. Proc Natl Acad Sci U SA 100:7045–7050.
Hiramatsu R, Matoba S, Kanai-Azuma M,Tsunekawa N, Katoh-Fukui Y, Kuroh-maru M, Morohashi K, Wilhelm D,Koopman P, Kanai Y. 2009. A criticaltime window of Sry action in gonadalsex determination in mice. Develop-ment 136:129–138.
Kanaoka Y, Urade Y. 2003. Hematopoieticprostaglandin D synthase. Prostaglan-dins Leukot Essent Fatty Acids 69:163–167.
Kim Y, Kobayashi A, Sekido R, DinapoliL, Brennan J, Chaboissier MC, PoulatF, Behringer RR, Lovell-Badge R, CapelB. 2006. Fgf9 and wnt4 act as antago-nistic signals to regulate Mammaliansex determination. PLoS Biol 4:e187.
Malki S, Berta P, Poulat F, Boizet-Bon-houre B. 2005a. Cytoplasmic retentionof the sex-determining factor SOX9 viathe microtubule network. Exp Cell Res309:468–475.
Malki S, Nef S, Notarnicola C, ThevenetL, Gasca S, Mejean C, Berta P, PoulatF, Boizet-Bonhoure B. 2005b. Prosta-glandin D2 induces nuclear import ofthe sex-determining factor SOX9 via itscAMP-PKA phosphorylation. EMBO J24:1798–1809.
Matsushita N, Aritake K, Takada A,Hizue M, Hayashi K, Mitsui K, HayashiM, Hirotsu I, Kimura Y, Tani T, Naka-jima H. 1998. Pharmacological studieson the novel antiallergic drug HQL-79:II. Elucidation of mechanisms for anti-allergic and antiasthmatic effects. JpnJ Pharmacol 78:11–22.
Michimata T, Tsuda H, Sakai M, Fuji-mura M, Nagata K, Nakamura M, SaitoS. 2002. Accumulation of CRTH2-posi-tive T-helper 2 and T-cytotoxic 2 cells atimplantation sites of human decidua ina prostaglandin D(2)-mediated manner.Mol Hum Reprod 8:181–187.
Mohri I, Taniike M, Taniguchi H, Kane-kiyo T, Aritake K, Inui T, Fukumoto N,Eguchi N, Kushi A, Sasai H, KanaokaY, Ozono K, Narumiya S, Suzuki K,Urade Y. 2006. Prostaglandin D2-medi-ated microglia/astrocyte interactionenhances astrogliosis and demyelin-ation in twitcher. J Neurosci 26:4383–4393.
Moniot B, Boizet-Bonhoure B, Poulat F.2008. Male specific expression of lipoca-lin-type prostaglandin D synthase(cPTGDS) during chicken gonadal dif-ferentiation: relationship with cSOX9.Sex Dev 2:96–103.
Moniot B, Declosmenil F, Barrionuevo F,Scherer G, Aritake K, Malki S, MarziL, Cohen-Solal A, Georg I, Klattig J,Englert C, Kim Y, Capel B, Eguchi N,Urade Y, Boizet-Bonhoure B, Poulat F.2009. The PGD2 pathway, independ-ently of FGF9, amplifies SOX9 activityin Sertoli cells during male sexual dif-ferentiation. Development 136:1813–1821.
Nef S, Schaad O, Stallings NR, CederrothCR, Pitetti JL, Schaer G, Malki S,Dubois-Dauphin M, Boizet-Bonhoure B,Descombes P, Parker KL, Vassalli JD.2005. Gene expression during sex deter-mination reveals a robust femalegenetic program at the onset of ovariandevelopment. Dev Biol 287:361–377.
Notarnicola C, Malki S, Berta P, PoulatF, Boizet-Bonhoure B. 2006. Transientexpression of SOX9 protein duringfollicular development in the adultmouse ovary. Gene Expr Patterns 6:695–702.
Rossi P, Dolci S, Albanesi C, Grimaldi P,Geremia R. 1993. Direct evidence thatthe mouse sex-determining gene Sry isexpressed in the somatic cells of malefetal gonads and in the germ cell line inthe adult testis. Mol Reprod Dev 34:369–373.
Schell C, Frungieri MB, Albrecht M, Gon-zalez-Calvar SI, Kohn FM, CalandraRS, Mayerhofer A. 2007. A Prostaglan-din D2 system in the human testis. Fer-til Steril 88:233–236.
Sekido R, Lovell-Badge R. 2008. Sexdetermination involves synergisticaction of SRY and SF1 on a specificSox9 enhancer. Nature 453:930–934.
Sim H, Argentaro A, Czech DP, Bagheri-Fam S, Sinclair AH, Koopman P,Boizet-Bonhoure B, Poulat F, HarleyVR. 2011. Inhibition of SRY-calmodulincomplex formation induces ectopicexpression of ovarian cell markers indeveloping XY gonads. Endocrinology152:2883–2893.
Stallings NR, Hanley NA, Majdic G, ZhaoL, Bakke M, Parker KL. 2002. Develop-ment of a transgenic green fluorescentprotein lineage marker for steroidogenicfactor 1. Endocr Res 28:497–504.
Sudbeck P, Scherer G. 1997. Two inde-pendent nuclear localization signals arepresent in the DNA-binding high-mobil-ity group domains of SRY and SOX9. JBiol Chem 272:27848–27852.
Svingen T, Spiller CM, Kashimada K,Harley VR, Koopman P. 2009. Identifi-cation of suitable normalizing genes forquantitative real-time RT-PCR analysisof gene expression in fetal mousegonads. Sex Dev 3:194–204.
Trivedi SG, Newson J, Rajakariar R,Jacques TS, Hannon R, Kanaoka Y,Eguchi N, Colville-Nash P, Gilroy DW.2006. Essential role for hematopoieticprostaglandin D2 synthase in thecontrol of delayed type hypersensitiv-ity. Proc Natl Acad Sci U S A 103:5179–5184.
Wilhelm D, Martinson F, Bradford S, Wil-son MJ, Combes AN, Beverdam A,Bowles J, Mizusaki H, Koopman P.2005. Sertoli cell differentiation isinduced both cell-autonomously andthrough prostaglandin signaling duringmammalian sex determination. DevBiol 287:111–124.
Wilhelm D, Hiramatsu R, Mizusaki H,Widjaja L, Combes AN, Kanai Y, Koop-man P. 2007a. SOX9 regulates prosta-glandin D synthase gene transcriptionin vivo to ensure testis development. JBiol Chem 282:10553–10560.
Wilhelm D, Palmer S, Koopman P. 2007b.Sex determination and gonadal develop-ment in mammals. Physiol Rev 87:1–28.
Dev
elop
men
tal D
ynam
ics
EXPRESSION OF H-PGDS AND THE SOX9 PROTEIN 2343
Copyright © 2022 FDOKUMEN




















