
tesis doctoral: decoding p300 evoked potentials for brain ...

Developmental Cell, Vol. 8, 153–166, February, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.devcel.2004.12.015
c-Myb and p300 Regulate HematopoieticStem Cell Proliferation and Differentiation
Mark L. Sandberg, Susan E. Sutton,Mathew T. Pletcher, Tim Wiltshire,Lisa M. Tarantino, John B. Hogenesch,and Michael P. Cooke*Genomics Institute of the Novartis Research
FoundationSan Diego, California 92121
Summary
Precise control of hematopoietic stem cell (HSC) pro-liferation and differentiation is needed to maintain alifetime supply of blood cells. Using genome-wideENU mutagenesis and phenotypic screening, we haveidentified a mouse line that harbors a point mutationin the transactivation (TA) domain of the transcriptionfactor c-Myb (M303V), which reduces c-Myb-depen-dent TA by disrupting its interaction with the tran-scriptional coactivator p300. The biological conse-quences of the c-MybM303V/M303V mutation includethrombocytosis, megakaryocytosis, anemia, lympho-penia, and the absence of eosinophils. Detailed analy-sis of hematopoiesis in c-MybM303V/M303V mice revealsdistinct blocks in T cell, B cell, and red blood cell de-velopment, as well as a remarkable 10-fold increasein the number of HSCs. Cell cycle analyses show thattwice as many HSCs from c-MybM303V/M303V animalsare actively cycling. Thus c-Myb, through interactionwith p300, controls the proliferation and differentia-tion of hematopoietic stem and progenitor cells.
Introduction
Hematopoiesis is a tightly controlled process requiringthe ordered regulation of gene expression throughoutdevelopment, from the hematopoietic stem cell (HSC)to mature and fully differentiated cells (Smith, 2003).The use of traditional gene targeting to identify theroles of genes in hematopoiesis has been invaluable forsome genes, but it has been of limited utility for manyothers due to embryonic lethality that often resultswhen a gene is disrupted so that it does not produceany functional protein. Confounding this problem is thatmany of the genes involved in hematopoiesis are re-quired at multiple stages of development, yet the em-bryonic lethality prevents examination of more matureanimals (Perry and Soreq, 2002). To circumvent thelimitations of traditional gene targeting, we employed aforward genetics approach using the mutagen N-ethyl-N-nitrosourea (ENU) and phenotypic screening of off-spring from mutagenized C57BL/6J (B6) mice for reces-sive traits (Nelms and Goodnow, 2001; Wiltshire et al.,2003).
The Myb family of proteins is a small family of threeclosely related nuclear transcription factors, A-, B-, andc-Myb (Oh and Reddy, 1999). All three members are de-
*Correspondence: [email protected]
fined by their unique DNA binding domain and overalldomain structure. Myb family members contain anamino-terminal, site-specific DNA binding domain, a cen-tral transactivation domain, and a carboxy-terminal regu-latory domain that modifies the effects of the transacti-vation domain (Lipsick, 1996; Oh and Reddy, 1999). Inthe case of A-Myb and C-Myb, deletion of the carboxyterminal regulatory domain increases transactivation,whereas, in B-Myb, deletion of the regulatory domaindecreases transactivation (Dubendorff et al., 1992; Naka-goshi et al., 1993; Oh and Reddy, 1997, 1998; Takahashiet al., 1995).
c-Myb is the founding member of the myb family andis the most intensely studied and best characterized ofthe myb proteins. c-Myb was first identified as the cel-lular homolog of the transforming gene of two avianretroviruses that induce leukemia, avian myeloblastosisvirus (AMV), and E26 (Baluda and Reddy, 1994). Subse-quent searches for proteins with sequence similarity toc-Myb resulted in the cloning and characterization ofA-Myb and B-Myb (Nomura et al., 1988). c-Myb plays acritical role in hematopoietic development, with nullmice dying at day 15 of embryogenesis due to a failureto transition from fetal to adult erythropoiesis (Mucen-ski et al., 1991). Most of what is known about c-Mybinvolves its role in hematopoiesis, during which it is ex-pressed most abundantly in immature and transformedhematopoietic cells, with expression generally declin-ing as hematopoietic cells differentiate (Gonda, 1998;Lipsick and Wang, 1999; Oh and Reddy, 1999). Morerecent work has shown that c-Myb is important in colondevelopment and that its expression, as in hematopoi-etic cells, declines as cells differentiate (Rosenthal etal., 1996; Thompson et al., 1998; Zorbas et al., 1999).
In adult mice, c-Myb is essential at multiple steps ofhematopoiesis (Gonda, 1998; Lipsick and Wang, 1999).However, defining the precise stages of hematopoiesis atwhich c-Myb is required in adult mice by using traditionalgene targeting strategies has been limited because of theembryonic lethality of c-Myb−/− mice. Several groupshave tried to circumvent this by expressing dominant-negative c-Myb or by generating chimeric c-Myb−/−
rag−/− mice; however, both methods have limitations(Allen et al., 1999; Badiani et al., 1994; Pearson andWeston, 2000; Siu, 2001; Taylor et al., 1996). Thus,much of what is known about c-Myb and its role in he-matopoiesis has arisen from in vitro studies, whichhave suggested that c-Myb may regulate the activityand expression of other proteins that are required atvarious stages of hematopoiesis (Ness, 1996; Ness,1999). However, it is difficult to extrapolate from thesein vitro studies to the whole organism; thus, developingan animal model to study c-Myb function in vivo is animportant goal. Using ENU mutagenesis, we have cre-ated a hypomorphic allele of c-Myb and have deter-mined the effect of this allele on hematopoiesis in adultmice. c-Myb mutants display multilineage hematopoi-etic alterations including anemia, lymphopenia, and eo-sinopenia, as well as elevated numbers of megakaryo-cytes, platelets, and hematopoietic stem cells (HSCs).

Developmental Cell154
These results define the stages of hematopoiesis regu- fllated by c-Myb and delineate an important role for c-Myb
in controlling HSC proliferation and differentiation. a
TResultsUtRecessive Thrombocytosis in an ENU-Induced
Mutant Strain miUsing ENU mutagenesis and a breeding strategy to
identify recessive mutations, we identified one mouse- otline, family 17, which had multiple mice with elevated
levels of platelets in the peripheral blood (Figure 1A). teTo determine heritability and identify the underlying ge-
netic mutation responsible for this phenotype, mice Rmfrom family 17 were crossed to 129S1/SvImJ (129S1)
mice. Peripheral blood platelet levels were measured a
Figure 1. Mapping and Identification of an ENU-Induced Mutation in c-Myb in Family 17
(A) Peripheral blood platelet levels in G3 offspring from ENU-mutagenized mice from the first 29 families screened. Mice with peripheral bloodplatelet levels > 3 SD from the mean are labeled in red. Family 17, which had greatly elevated platelet levels, is boxed.(B) Peripheral blood platelet levels in B6/129 F2 animals. Squares and circles represent male and female mice, respectively.(C) Mapping of the high-platelet phenotype. Shown are the results from fine mapping of the region of chromosome 10 linked to the elevatedplatelet phenotype. Markers are listed in the first column and indicate chromosome and megabase position. The genotype for each SNPmarker is represented as: B, homozygous C57BL/6J genotype; S, homozygous 129SvIm/J; H, heterozygous. Italics indicate inferred geno-types. On the right is a diagram of mouse chromosome 10 showing the interval to which the high-platelet phenotype mapped with the relativelocations of 3 of the 125 genes reported within the interval.(D) Sequence from representative unaffected and affected mice in the region encoding amino acids 300–306 of c-Myb. An A-to-G transitionthat was found only in family 17 is boxed.(E) Domain structure of c-Myb: DNA binding, R1, R2, and R3; transactivation/acidic, TA; negative regulatory, NR.
rom F2 mice, and 41 of 149 (28%) had elevated plateletevels, indicating a heritable phenotype consistent with
nonlethal recessive trait (Figure 1B).
hrombocytosis Maps to a Point Mutation in c-Mybsing single nucleotide polymorphism (SNP)-based geno-
yping between B6 and 129S1, the underlying mutationapped to a 9 MB interval on chromosome 10, contain-
ng approximately 125 genes (Figure 1C). We focusedn the proto-oncogene c-Myb, as it is localized withinhis region, expressed in multiple hematopoietic cellypes, and is known to be essential for hematopoi-sis (Gonda, 1998; Lipsick and Wang, 1999; Oh andeddy, 1999). Sequencing of all exons of c-Myb fromultiple affected and unaffected control mice revealed
n A-to-G transition that is present and uniquely homo-
c-Myb Regulates HSC Number and Differentiation155
zygous in mice that display the elevated platelet phe-notype (Figure 1D). The A-to-G mutation in c-Myb infamily 17 results in a methionine-to-valine substitutionat amino acid 303 (M303V) in the transactivation (TA)domain of c-Myb, which mediates the association withthe transcriptional coactivator p300 and is required forproper c-Myb function (Figure 1E) (Parker et al., 1999).
The M303V Mutation in c-Myb ReducesTransactivation and Association with p300To determine if the phenotype in c-MybM303V/M303V miceis due to a destabilization of c-Myb, we measured thelevels of c-Myb in whole bone marrow from c-Mybwt/wt
and c-MybM303V/M303V mice by Western blot. The levelsof c-Myb in the bone marrow of c-Mybwt/wt mice wereapproximately equal to those found in c-MybM303V/M303V
mice, indicating that the M303V mutation in c-Mybdoes not significantly alter the levels of c-Myb (datanot shown).
To determine if the M303V mutation affects TA byc-Myb, we fused the DNA binding domain of Gal4 toderivatives of c-Myb containing WT, M303V, or 304GPTA domains (Figure 2A). As a control, we included apreviously described mutation, in which a glycine anda proline are inserted after amino acid 304 of c-Myb
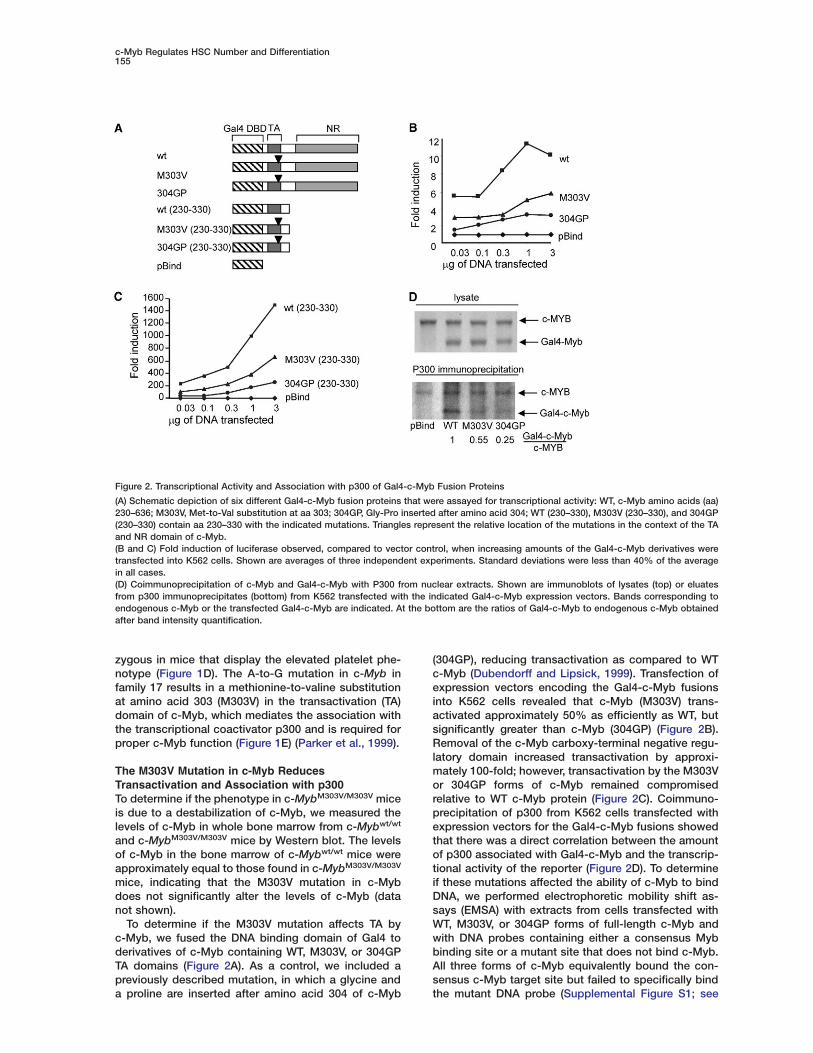
Figure 2. Transcriptional Activity and Association with p300 of Gal4-c-Myb Fusion Proteins
(A) Schematic depiction of six different Gal4-c-Myb fusion proteins that were assayed for transcriptional activity: WT, c-Myb amino acids (aa)230–636; M303V, Met-to-Val substitution at aa 303; 304GP, Gly-Pro inserted after amino acid 304; WT (230–330), M303V (230–330), and 304GP(230–330) contain aa 230–330 with the indicated mutations. Triangles represent the relative location of the mutations in the context of the TAand NR domain of c-Myb.(B and C) Fold induction of luciferase observed, compared to vector control, when increasing amounts of the Gal4-c-Myb derivatives weretransfected into K562 cells. Shown are averages of three independent experiments. Standard deviations were less than 40% of the averagein all cases.(D) Coimmunoprecipitation of c-Myb and Gal4-c-Myb with P300 from nuclear extracts. Shown are immunoblots of lysates (top) or eluatesfrom p300 immunoprecipitates (bottom) from K562 transfected with the indicated Gal4-c-Myb expression vectors. Bands corresponding toendogenous c-Myb or the transfected Gal4-c-Myb are indicated. At the bottom are the ratios of Gal4-c-Myb to endogenous c-Myb obtainedafter band intensity quantification.
(304GP), reducing transactivation as compared to WTc-Myb (Dubendorff and Lipsick, 1999). Transfection ofexpression vectors encoding the Gal4-c-Myb fusionsinto K562 cells revealed that c-Myb (M303V) trans-activated approximately 50% as efficiently as WT, butsignificantly greater than c-Myb (304GP) (Figure 2B).Removal of the c-Myb carboxy-terminal negative regu-latory domain increased transactivation by approxi-mately 100-fold; however, transactivation by the M303Vor 304GP forms of c-Myb remained compromisedrelative to WT c-Myb protein (Figure 2C). Coimmuno-precipitation of p300 from K562 cells transfected withexpression vectors for the Gal4-c-Myb fusions showedthat there was a direct correlation between the amountof p300 associated with Gal4-c-Myb and the transcrip-tional activity of the reporter (Figure 2D). To determineif these mutations affected the ability of c-Myb to bindDNA, we performed electrophoretic mobility shift as-says (EMSA) with extracts from cells transfected withWT, M303V, or 304GP forms of full-length c-Myb andwith DNA probes containing either a consensus Mybbinding site or a mutant site that does not bind c-Myb.All three forms of c-Myb equivalently bound the con-sensus c-Myb target site but failed to specifically bindthe mutant DNA probe (Supplemental Figure S1; see

Developmental Cell156
the Supplemental Data available with this article on- irline), suggesting that these mutations do not change
the ability of c-Myb to bind DNA. Collectively, thesedata demonstrate that the M303V c-Myb mutation al-
clows c-Myb to bind to target sequences but fails to effi-
ociently recruit p300, resulting in suboptimal transactiva-
Ttion of c-Myb target sequences.
iasc-MybM303V/M303V Mice Display Multilineage
Hematopoietic Defects ucTo characterize the effect of reduced c-Myb activity on
hematopoiesis, we performed detailed hematopoietic slanalysis on c-MybM303V/M303V mice. Peripheral blood
analysis showed that the c-MybM303V/M303V mice dis- c5played thrombocytosis, anemia, lymphopenia, and eo-
sinopenia (Figure 3A and Supplemental Table S1). csc-MybM303V/M303V mice had normal levels of most my-
eloid cell populations (neutrophils, monocytes, and ba- tnsophils), but they were devoid of eosinophils (Figure
3A). In addition to alterations in the number of platelets itand red blood cells, both of these cell types are signifi-
cantly larger in c-MybM303V/M303V mice compared to cLnormal littermates, suggesting the possibility of further
developmental alterations (Supplemental Table S1). CbFlow-cytometric analysis of B and T cell populations in
the peripheral blood showed that the numbers of both mtB220+ B cells and CD4+ or CD8+ T cells were also
lower in c-MybM303V/M303V mice (Figure 3B). Aside from gaa small increase in platelet levels and a small decrease
Figure 3. Hematological Analysis of PeripheralBlood from c-Mybwt/wt, c-Mybwt/M303V, andc-MybM303V/M303V Mice
(A) Analysis of peripheral blood fromc-Mybwt/wt (n = 22), c-Mybwt/M303V (n = 35),and c-MybM303V/M303V (n = 19) mice by a he-matology analyzer. Squares and circles repre-sent male and female mice, respectively.(B) Peripheral blood levels of B220+ B cellsand CD4+ and CD8+ single-positive T cellsmeasured by flow cytometry.
n RBCs, heterozygous c-Mybwt/M303V mice appearedelatively normal (Figure 3 and Supplemental Table S1).
-MybM303V/M303V Mice Have Reduced Numbersf Thymocytes and Elevated Megakaryocyteso further characterize the hematopoietic phenotypesn c-MybM303V/M303V mice, we examined lymphocytend megakaryocyte populations in the bone marrow,pleen, and thymus. Total bone marrow cellularity wasnchanged in c-MybM303V/M303V animals; however,-MybM303V/M303V animals contained 2- to 10-fold fewerplenocytes and thymocytes compared to wild-type
ittermates (Figure 4A). The number of CD41+ cells, aell surface marker for megakaryocytes, increased-fold in the bone marrow and 10-fold in the spleen of-MybM303V/M303V animals (Figure 4B). Wright’s-Giemsataining of bone marrow and splenocytes confirmedhat c-MybM303V/M303V mice have greatly increasedumbers of megakaryocytes (Figure 4B). To determine
f the c-MybM303V/M303V phenotype is cell intrinsic, weransplanted bone marrow from Ly5.2/5.2 c-Mybwt/wt or-MybM303V/M303V mice along with an equal dose ofy5.1/5.1 c-Mybwt/wt bone marrow into lethally irradiated57BL/6 Ly5.1/5.1 recipient mice and measured the contri-ution of Ly5.2/5.2 c-MybM303V/M303V or c-Mybwt/wt bonearrow to B, T, or myeloid cells 12 weeks after transplan-
ation. Bone marrow from Ly5.2/5.2 c-MybM303V/M303V miceave rise to reduced numbers of Ly5.2/5.2 peripheral Bnd T cells but normal levels of Ly5.2/5.2 myeloid cells

c-Myb Regulates HSC Number and Differentiation157
Figure 4. Hematological Analysis of Lymphoid Organs and Bone Marrow Transplants from c-Mybwt/wt and c-MybM303V/M303V Mice
(A) Cellularity of thymus, spleen, and bone marrow of c-Mybwt/wt and c-MybM303V/M303V mice. Symbols and color coding are as in Figure 1.(B) Left side: Flow cytometry plots of CD41+ cells in the bone marrow and spleen of a c-Mybwt/wt and a c-MybM303V/M303V mouse. Right side:Wright’s-Giemsa staining of bone marrow and splenocytes from a c-Mybwt/wt and a c-MybM303V/M303V mouse. Arrows point to megakaryocytes.(C) Analysis of peripheral blood populations in lethally irradiated WT Ly5.1/5.1 mice that were transplanted with 5 × 106 bone marrow cells fromLy5.2/5.2 c-Mybwt/wt or c-MybM303V/M303V mice together with 5 × 106 bone marrow cells from Ly5.1/5/1 WT mice. Shown is the percentage of Ly5.2/5.2
cells for each cell population. Individual mice are indicated by symbols, with the horizontal line representing the average of all the mice. Asterisksdenote statistically significant differences between WT and M303V bone marrow (Student’s t test, **p < 0.01, *p < 0.05).
compared to controls that received Ly5.2/5.2 c-Mybwt/wt
bone marrow (Figure 4C). Importantly, cotransplantedLy5.1/5.1 bone marrow was able to reconstitute all lin-
eages, even in the presence of c-MybM303V/M303V bonemarrow, demonstrating that the phenotypes observedin c-MybM303V/M303V mice reflect cell-intrinsic effects of

Developmental Cell158
Figure 5. Altered Lymphocyte Development in c-MybM303V/M303V Mice
(A and B) B and T cell development in the (A) bone marrow and (B) thymus of representative c-Mybwt/wt and c-MybM303V/M303V mice. In parenthesesis the name of the respective population as defined in the text.(C) T cell development in the thymus 12 weeks after transplant with 5 × 106 bone marrow cells from Ly5.2/5.2 c-Mybwt/wt or c-MybM303V/M303V micetogether with an equal number of WT Ly5.1/5.1 bone marrow cells into lethally irradiated WT Ly5.1/5.1 mice. The example shown is representative offour animals of each genotype analyzed.
c-Myb Regulates HSC Number and Differentiation159
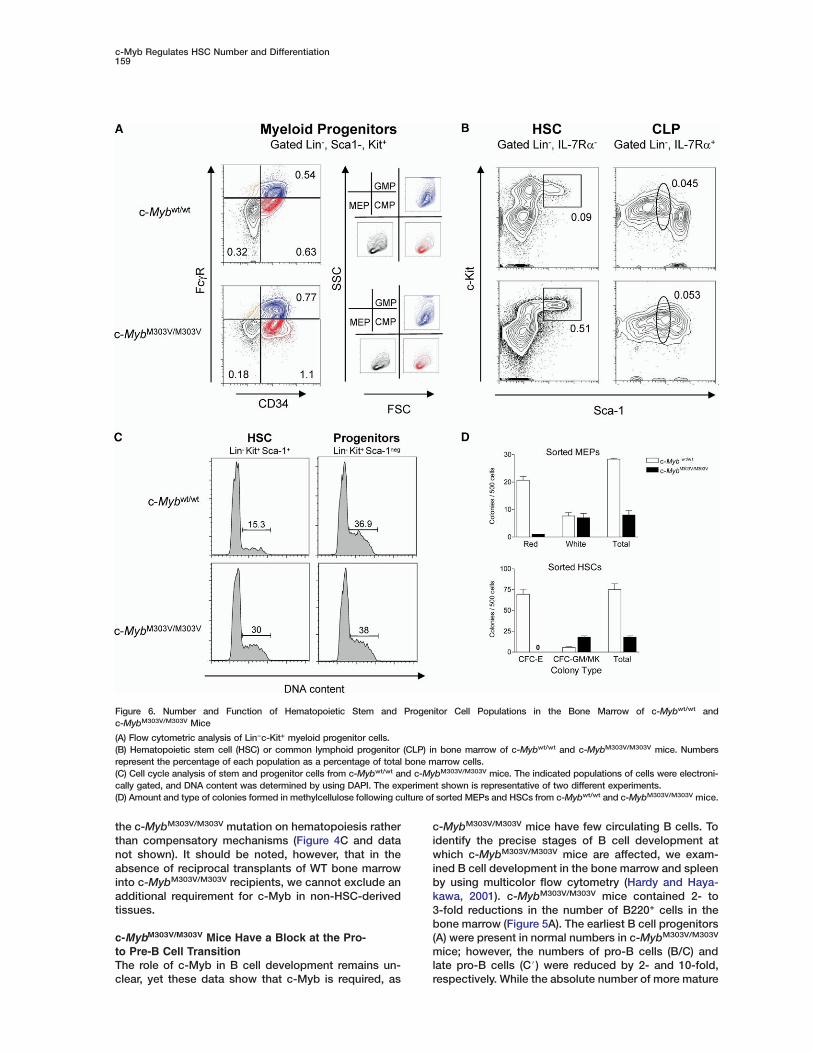
Figure 6. Number and Function of Hematopoietic Stem and Progenitor Cell Populations in the Bone Marrow of c-Mybwt/wt andc-MybM303V/M303V Mice
(A) Flow cytometric analysis of Lin−c-Kit+ myeloid progenitor cells.(B) Hematopoietic stem cell (HSC) or common lymphoid progenitor (CLP) in bone marrow of c-Mybwt/wt and c-MybM303V/M303V mice. Numbersrepresent the percentage of each population as a percentage of total bone marrow cells.(C) Cell cycle analysis of stem and progenitor cells from c-Mybwt/wt and c-MybM303V/M303V mice. The indicated populations of cells were electroni-cally gated, and DNA content was determined by using DAPI. The experiment shown is representative of two different experiments.(D) Amount and type of colonies formed in methylcellulose following culture of sorted MEPs and HSCs from c-Mybwt/wt and c-MybM303V/M303V mice.
the c-MybM303V/M303V mutation on hematopoiesis ratherthan compensatory mechanisms (Figure 4C and datanot shown). It should be noted, however, that in theabsence of reciprocal transplants of WT bone marrowinto c-MybM303V/M303V recipients, we cannot exclude anadditional requirement for c-Myb in non-HSC-derivedtissues.
c-MybM303V/M303V Mice Have a Block at the Pro-to Pre-B Cell TransitionThe role of c-Myb in B cell development remains un-clear, yet these data show that c-Myb is required, as
c-MybM303V/M303V mice have few circulating B cells. Toidentify the precise stages of B cell development atwhich c-MybM303V/M303V mice are affected, we exam-ined B cell development in the bone marrow and spleenby using multicolor flow cytometry (Hardy and Haya-kawa, 2001). c-MybM303V/M303V mice contained 2- to3-fold reductions in the number of B220+ cells in thebone marrow (Figure 5A). The earliest B cell progenitors(A) were present in normal numbers in c-MybM303V/M303V
mice; however, the numbers of pro-B cells (B/C) andlate pro-B cells (C#) were reduced by 2- and 10-fold,respectively. While the absolute number of more mature

Developmental Cell160
Figure 7. Competitive Repopulation Assay Shows an Increased Number of Functional HSCs in Bone Marrow of c-MybM303V/M303V Mice
(A) Mixed bone marrow transplants were conducted by using the indicated ratio of bone marrow from c-MybM303V/M303V or c-Mybwt/wt donor(Ly5.2/5.2) mice along with 5 × 105 competitor bone marrow cells from Ly5.1/5.2 mice into lethally irradiated Ly5.1/5.1 recipient mice.(B) Representative flow cytometric analysis of primary recipient animals receiving Ly5.2/5.2 c-Mybwt/wt (left side) or Ly5.2/5.2 c-MybM303V/M303V
(right side) bone marrow.(C) Peripheral blood analysis of primary recipients. Shown is the percentage of donor-derived (Ly5.2/5.2) cells for each of the recipients 8weeks after transplant (top left panel). Ratio of donor to competitor for each of the indicated cell populations in the peripheral blood 8 weeks

c-Myb Regulates HSC Number and Differentiation161
Kondo et al., 2001). Briefly, hematopoiesis starts with the sis that c-Myb is a key regulator of the erythrocyte/
after transplant. Each point represents the average ± SEM of 3–4 animals for each group.(D) HSC composition in bone marrow of Ly5.1/5/.1 recipients. Bone marrow of mice that received a transplant containing the indicated ratio ofdonor:competitor cells 12 weeks earlier was analyzed for HSC (Lin−Kit+Sca+) content by flow cytometry. Shown is the final ratio of donor:com-petitor cells within the HSC compartment (upper panel) and the absolute percentage of HSCs in each recipient for the wt:wt chimeras (lowerpanel, left half) or M303V:wt chimeras (lower panel, right half).(E) Contribution of Ly5.2/5.2 bone marrow in the peripheral blood of secondary recipients 8 weeks following transplant. Mice showing donorcontribution above 10% were used for tertiary transplants.(F) Limit dilution analysis of HSC numbers in tertiary transplants. The indicated number of bone marrow cells from secondary recipients wastransplanted into lethally irradiated Ly5.1/5.1 recipients, and blood was analyzed 8 weeks later for Ly5.2/5.2 donor cells (left panel). Animalscontaining greater than 1% Ly5.2/5.2 cells in the peripheral blood were scored as positive and were used to calculate the frequency of HSCsin the secondary recipients (right panel).
B cells (D–F) was reduced 2-fold, the relative propor-tions of B220+IgM−, B220+IgM+, or B220+IgM+IgD+ cellswere unchanged in the bone marrow and spleens ofc-MybM303V/M303V mice (Figure 5A and data not shown).Thus, the few c-Myb mutant B cells that progressedbeyond C# are apparently able to mature normally.
c-MybM303V/M303V Mice Have Reduced Numbersof DN1 and DN4 Thymocytesc-Myb is expressed in both early and late stages of Tcell development; however, the embryonic lethality ofc-Myb−/− mice has made it difficult to clearly definethe role of c-Myb in T cell development (Allen et al.,1999, 2001; Pearson and Weston, 2000). To determinethe precise stages of T cell development altered inc-MybM303V/M303V mice, we performed multicolor flowcytometry in the thymus (Godfrey et al., 1993). Despitethe 3- to 10-fold reduction in the number of total thymo-cytes, we observed only minor differences in the pro-portions of various thymocyte subpopulations. Thegreatest difference was an approximate 50% reductionin the percentage and number of early thymocyte DN1progenitors and a reduction in the absolute number ofDN4 progenitors in c-MybM303V/M303V thymi (Figure 5B;see below). Additionally c-MybM303V/M303V mice displayeda slight reduction in the percentage of CD4+CD8+ double-positive thymocytes and a corresponding increase inthe percentage, but not absolute number, of single-positive CD4+ and CD8+ thymocytes.
To determine if c-MybM303V/M303V thymocytes canprogress normally in the presence of WT thymocytes,we performed mixed bone marrow chimera experi-ments with c-MybM303V/M303V and c-Mybwt/wt bone mar-row. Lethally irradiated Ly5.1/5.1 c-Mybwt/wt mice werereconstituted with a mix of bone marrow from Ly5.1/5.1
c-Mybwt/wt mice and Ly5.2/5.2 bone marrow fromc-Mybwt/wt or c-MybM303V/M303V mice. Flow cytometricanalysis of the thymocytes 12 weeks after transplanta-tion revealed a significant reduction in the number ofDN4 cells derived from c-MybM303V/M303V bone marrow(Figure 5C). These data demonstrate that T cells de-rived from c-MybM303V/M303V bone marrow are unable tocompete effectively with WT T cells and exhibit amarked block at the DN3-to-DN4 transition.
c-MybM303V/M303V Mice Have a 5- to 10-FoldIncrease in Phenotypic HSCsBecause c-Myb is highly expressed in HSCs, we nextexamined the numbers of phenotypic HSCs and theirprogeny in c-MybM303V/M303V mice (Denburg et al., 1985;
HSC, which in turn differentiates into the common lym-phoid progenitor (CLP) or common myeloid progenitor(CMP). The CLP gives rise to the NK, B, and T cell lineages,while the CMP gives rise to the granulocyte/macro-phage progenitor (GMP) and the megakaryocyte/erythro-cyte progenitor (MEP). To identify these populations, weperformed flow cytometric staining of the bone marrowand compared the relative amounts of these populationsin WT and c-MybM303V/M303V mice (Akashi et al., 2000;Kondo et al., 1997). The numbers of phenotypic CMPs andGMPs were slightly elevated (<2-fold) in c-MybM303V/M303V
mice, while phenotypic MEPs were slightly decreased (Fig-ure 6A; see below). Strikingly, c-MybM303V/M303V mice con-tained 5- to 10-fold elevated numbers of phenotypic HSCs(Figure 6B; see below). The number of CLPs were un-changed in c-MybM303V/M303V mice compared to their WTlittermates. To determine if the increased numbers ofphenotypic HSCs in c-MybM303V/M303V mice reflectedincreased proliferation of HSCs or reduced differentiation,we performed cell cycle analysis of hematopoietic stemand progenitor cells. This analysis showed that twice asmany Lin−Kit+Sca-1+ HSCs from c-MybM303V/M303V arein the S/G2/M phase of the cell cycle compared to con-trol mice (Figure 6C). Importantly, the DNA content ofmore differentiated progenitor cells of the pheno-type Lin−Kit+Sca-1neg was similar for both WT andc-MybM303V/M303V animals, demonstrating that theincreased number of cycling cells was not a generalproperty of c-MybM303V/M303V animals (Figure 6C).
Phenotypic MEPs and HSCs from c-MybM303V/M303V
Mice Form Fewer Erythrocyte Colonies In VitroThe anemia observed in c-MybM303V/M303V mice isconsistent with a corresponding reduction of phe-notypic MEPs but is inconsistent with the elevatedlevels of megakaryocytes. To help address this appar-ent contradiction, methylcellulose colony-formingassays were performed on sorted MEPs and HSCs toexamine the differentiation potential of each into whiteor red blood cell colonies. Both MEPs and HSCs fromc-MybM303V/M303V mice showed dramatic reductionsin the number of red blood cell colonies comparedto cells from WT mice (Figure 6D). Interestingly, therewas a difference between the HSCs and MEPs fromc-MybM303V/M303V mice in their ability to form whiteblood cell colonies. MEPs from c-MybM303V/M303V miceformed white blood cell colonies as efficiently as WTMEPs, yet HSCs from c-MybM303V/M303V mice formed2- to 3-fold more GM/MK colonies compared to WTHSCs. These results are consistent with the hypothe-
Developmental Cell162
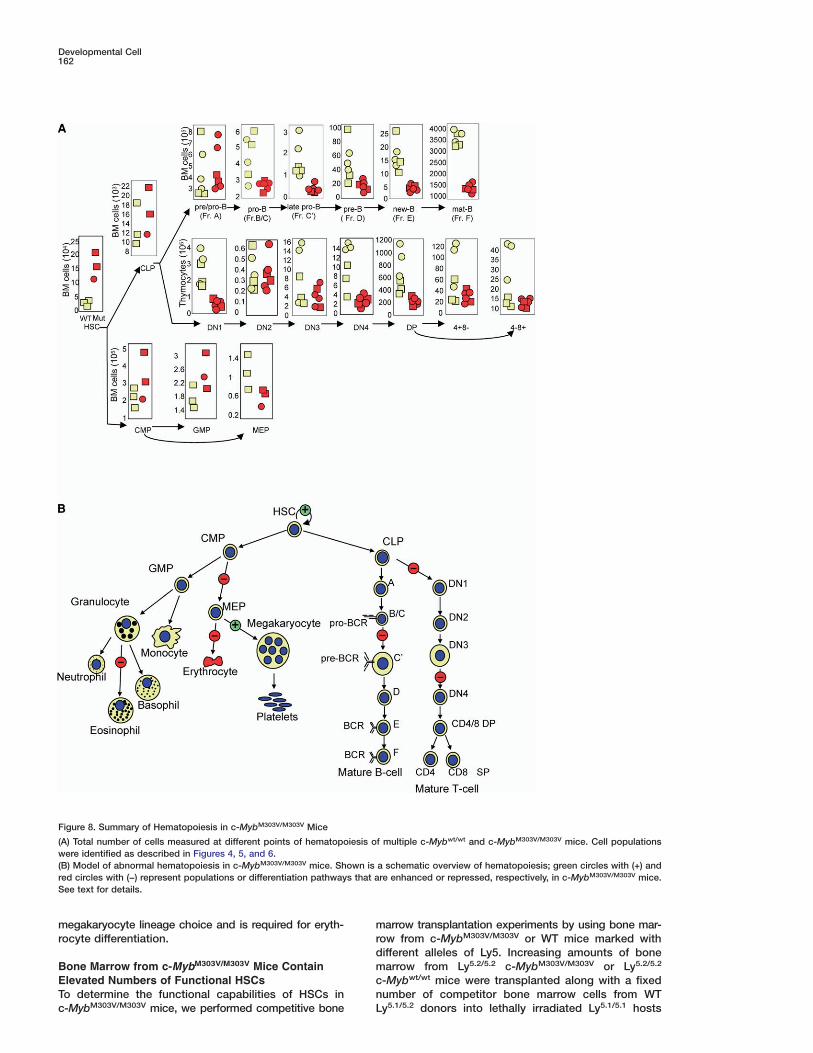
Figure 8. Summary of Hematopoiesis in c-MybM303V/M303V Mice
(A) Total number of cells measured at different points of hematopoiesis of multiple c-Mybwt/wt and c-MybM303V/M303V mice. Cell populationswere identified as described in Figures 4, 5, and 6.(B) Model of abnormal hematopoiesis in c-MybM303V/M303V mice. Shown is a schematic overview of hematopoiesis; green circles with (+) andred circles with (−) represent populations or differentiation pathways that are enhanced or repressed, respectively, in c-MybM303V/M303V mice.See text for details.
megakaryocyte lineage choice and is required for eryth- mrrocyte differentiation.dmBone Marrow from c-MybM303V/M303V Mice Contain
Elevated Numbers of Functional HSCs cnTo determine the functional capabilities of HSCs in
c-MybM303V/M303V mice, we performed competitive bone
arrow transplantation experiments by using bone mar-ow from c-MybM303V/M303V or WT mice marked withifferent alleles of Ly5. Increasing amounts of bonearrow from Ly5.2/5.2 c-MybM303V/M303V or Ly5.2/5.2
-Mybwt/wt mice were transplanted along with a fixedumber of competitor bone marrow cells from WT
Ly5.1/5.2 donors into lethally irradiated Ly5.1/5.1 hosts

c-Myb Regulates HSC Number and Differentiation163
(see Figure 7A for an outline of these experiments).Eight weeks after transplantation, the ratio of Ly5.2/5.2
donor to Ly5.2/5.1 competitor cells was determined formyeloid, T, and B cells by flow cytometry of the periph-eral blood (Figure 7B). As expected, the ratio of donor:competitor cells in the peripheral blood of mice trans-planted with Ly5.2/5.2 c-Mybwt/wt donors was similar tothe original ratio used for the transplant for all threelineages of cells (Figure 7C, open squares). In contrast,mice that received bone marrow from c-MybM303V/M303V
donors had marked differences in the donor:competitorratio for all three cell populations. Specifically, the do-nor:competitor ratio was reduced 5- to 10-fold for bothB and T cells, and the donor:competitor ratio for my-eloid cells was increased 2- to 5-fold in mice that re-ceived bone marrow from c-MybM303V/M303V mice. Theincreased contribution of c-MybM303V/M303V bone mar-row to cells of the myeloid lineage could reflect ei-ther increased differentiation of c-Myb mutant HSCs tothe myeloid lineage or increased engraftment byc-MybM303V/M303V HSCs.
To determine if the increased myeloid chimerism re-flected the level of HSC engraftment, the bone marrowcells of recipient animals were analyzed for the contri-bution of donor and competitor cells to the HSC pool.As expected, control animals that received Ly5.2/5.2
c-Mybwt/wt donor bone marrow had ratios of donor andcompetitor HSCs that reflected the original transplantratio (Figure 7D, upper panel, open squares). In con-trast, the ratio of donor:competitor HSCs in animalstransplanted with c-MybM303V/M303V donors was ele-vated 20-fold (Figure 7D, upper panel, closed circles,p < 0.01, Student’s t test) compared to c-Mybwt/wt chi-meras. Additionally, the percentage of c-Myb mutantHSCs in these recipients was 5-fold higher comparedto cotransplanted competitor cells, or compared to ani-mals that received Ly5.2/5.2 c-Mybwt/wt bone marrow,suggesting a cell autonomous defect in the ability tocontrol HSC pool size (Figure 7D, lower panel).
To determine if the increased number of Lin−Kit+
Sca-1+ cells present in recipient mice that receivedc-MybM303V/M303V bone marrow were long-term HSCs(LT-HSCs) capable of selfrenewal, we performed sec-ondary transplants into irradiated Ly5.1/5.1 recipients byusing bone marrow from mice that originally receivedequivalent numbers of Ly5.2/5.2 c-Mybwt/wt orc-MybM303V/M303V and competitor Ly5.2/5.1 marrow. Inthese secondary recipients, the contribution of Ly5.2/5.2-bearing cells in the peripheral blood was similar wheneither c-Mybwt/wt or c-MybM303V/M303V bone marrow cellswere used as the original bone marrow donor (Figure 7E).As before, the Ly5.2/5.2 c-MybM303V/M303V bone marrowgave robust reconstitution of myeloid lineage cells andlow-level reconstitution of lymphoid cells (data notshown). To determine the absolute number of func-tional HSCs in these secondary recipients, bone mar-row cells were harvested from secondary recipientswith equivalent levels of engraftment and were usedfor tertiary transplants into irradiated Ly5.1/5.1 mice atlimiting dilution. All tertiary mice reconstituted with 1 ×106 cells from Ly5.2/5.2 c-MybM303V/M303V mice containeddetectable levels of multilineage (B, T, and myeloid) en-graftment (Figure 7F, filled circles, and data not shown).In contrast, only 1 of 4 animals transplanted with 1 ×
106 bone marrow cells from Ly5.2/5.2 c-Mybwt/wt animalsgave detectable engraftment (Figure 7F, open squares).Limiting dilution analysis of these data determined thatthe frequency of HSCs was 10-fold higher in mice ori-ginally transplanted with Ly5.2/5.2 c-MybM303V/M303V
bone marrow compared to control Ly5.2/5.2 c-Mybwt/wt
marrow (1/420,000 [SE = 1/257,69–1/684,974] versus1/1,430,000 [SE = 1/1,616,837–1/11,762,168], respec-tively, p < 0.05). The increased frequency of HSCs inthese secondary recipients is consistent with the 5- to10-fold increase in the number of phenotypic HSCs ob-served in c-MybM303V/M303V mice (Figures 6B and 8A)and in primary transplant recipients. Based on thesedata, we conclude that c-MybM303V/M303V mice contain10-fold elevations in the number of LT-HSCs.
Discussion
The present study supports a critical role for c-Myb-p300 interaction throughout the hematopoietic hierar-chy. Importantly, c-Myb is not required at every stepand for every lineage; rather, there are distinct stepsand lineages that are critically dependent on c-Mybfunction. These include negative regulation of HSC pro-liferation, megakaryocyte numbers, and essential rolesin erythropoiesis, eosinophil, T, and B cell development(Figure 8). Although previous studies have providedclues that c-Myb may play a role in some of these pro-cesses, the severe phenotype of c-Myb−/− animals hasprecluded a detailed examination of c-Myb function inadult animals.
c-Myb Limits Megakaryocyte Developmentand Promotes RBC DevelopmentSeveral lines of evidence support a critical role forc-Myb in directing megakaryocyte and erythrocyte de-velopment. First, c-Myb−/− mice die of anemia in uteroat day 15 due to a failure to initiate fetal liver hemato-poiesis (Mucenski et al., 1991). MEPs are likely formedin c-Myb−/− mice, as megakaryocytes are present in thefetal liver of c-Myb−/− animals (Mucenski et al., 1991).Subsequent studies, using ES cell chimeras, haveshown that c-Myb−/− animals contain low numbers ofdefinitive stem/progenitor cells that can give rise toearly thymic precursors and initiate myeloid cell de-velopment (Allen et al., 1999; Parker et al., 1999). Thus,the failure to initiate fetal erythropoiesis is likely to re-sult from an inability of later progenitor cells to dif-ferentiate into RBCs. Secondly, neonatal mice contain-ing w20% the normal amount of c-Myb also haveanemia and elevated numbers of megakaryocytesand platelets (Emambokus et al., 2003). Finally, thec-MybM303V/M303V mice described herein and otherENU-induced alleles of c-Myb recently published con-tain elevated numbers of platelets and CD41+ mega-karyocytes and reduced numbers of RBCs (Carpinelliet al., 2004). Thus, multiple independent alleles ofc-Myb give an identical phenotype: reduced numbersof RBCs and elevated numbers of megakaryocytes.Since these two cell types arise from a common precur-sor, the simplest explanation is that c-Myb is requiredfor the MEP to initiate the RBC program and also par-ticipates in turning off the megakaryocyte program. Insupport of this view, we find only modest reductions in
the number of MEPs in c-MybM303V/M303V animals com-
Developmental Cell164
pared to normal littermates; however, these cells are nunable to make RBCs following culture. Thus, these bi- spotential cells are present, but their differentiation po- 1tential is limited. Collectively, these data suggest that Hc-Myb normally limits the ability of the HSC to dif- 5ferentiate toward the megakaryocyte lineage while pro- mmoting the RBC program. t
eDistinct Steps in Lymphoid Development tAre Dependent on c-Myb tThe low numbers of B and T cells in the blood of tc-Myb−/− mice suggest that c-Myb may be regulat- Ting both B and T cell development. c-MybM303V/M303V mmice contained normal numbers of phenotypic Lin−Kit+ cSca1+IL7R+ CLPs, suggesting that these early precur- nsor cells are present in normal numbers. Consistent bwith this observation, we found normal numbers of the 7earliest pre-pro B cells. However, the numbers of pro- sB cells were reduced 3-fold, and the numbers of pre-B dcells were down more than 10-fold in c-MybM303V/M303V omice. Importantly, the percentage of B cells at sub- psequent stages of development was not different, palthough the absolute numbers were reduced in cc-MybM303V/M303V mice, and their functional capacity nremains to be tested. Reduced numbers of pro-B and npre-B cells in c-MybM303V/M303V mice suggest that Tc-Myb may play a role in directing IgH chain rearrange- ament or in the expansion of those cells that have suc- fcessfully rearranged IgH.
The ability of c-Myb to regulate T cell development is gwell established; however, identification of the precise Epoints that c-Myb regulates has been controversial. Early astudies demonstrated a requirement for c-Myb at the pre- sDN1 stage of T cell development (Allen et al., 1999), while llater studies with a dominant-negative form of c-Myb pshowed a role for c-Myb in the regulation of DN3 and iDN4 cell expansion post-TCR β chain selection (Pearson gand Weston, 2000). A more recent study with neonatal omice with partial reductions in c-Myb levels showed that min fetal thymus development c-Myb is required for DN1 pto DN2 maturation and the transition from DN2 to DN3, cwith the former step requiring higher threshold levels of tc-Myb (Emambokus et al., 2003). Very recent data gener- nated with conditional null alleles in which Myb is deleted aearly or late in T cell development in the thymus support ma roll for c-Myb in directing the DN3-to-DN4 transition, ssurvival of preselection double-positive cells, and differ- tentiation of CD4 SP cells (Bender et al., 2004, Lieu et al.,2004). In agreement with these data, the c-MybM303V/M303V tmice described here show 2- to 10-fold reductions in bthymocyte number with 3-fold reductions in the number eof both DN1 and DN4 cells in mature animals. Interest- oingly, competitive bone marrow reconstitution experi- tments showed that cells harboring the c-MybM303V/M303V
sallele contained normal numbers of DN1 cells and more pthan 4-fold reductions in DN4 cells (Figure 5). Based on tthe current work and previous studies discussed aabove, it seems that c-Myb can regulate T cell devel-
Eopment at many points; however, the mechanistic de-tails remain an area of active research.
PAIncreased Hematopoietic Stem Cellsl
in c-MybM303V/M303V Mice dOne of the surprising findings of this study was that
c-MybM303V/M303V mice contain 5- to 10-fold more phe-otypic HSCs (Figures 6B and 8A). Reconstitutiontudies with mixed bone marrow chimeras showed a0- to 20-fold increase in the number of functionalSCs in bone marrow from c-MybM303V/M303V mice, and-fold elevations in the absolute number of HSCs in pri-ary recipients; thus, these phenotypic HSCs are func-
ional (Figure 7C). Secondary and tertiary transplantxperiments of these bone marrow chimeras showedhat these HSCs are capable of selfrenewal and long-erm multilineage reconstitution, thus confirming thathese represent bona fide LT-HSCs (Figures 7D and 7E).hese experiments also revealed that the defects in he-atopoiesis in c-MybM303V/M303V mice are cell intrinsic;
-MybM303V/M303V bone marrow gave rise to normalumbers of myeloid and granulocytes, while the num-ers of T and B cells were decreased (Figures 4C andC). Importantly, WT cells cotransplanted into theseame animals differentiated normally, showing that theefects in hematopoiesis resulted from direct effectsf c-MybM303V/M303V on the development of HSC androgenitors and not from secondary effects of the lym-hopenia, anemia, or megakaryocytosis. HSCs from-MybM303V/M303V mice contain an increase in theumber of actively cycling cells, providing a mecha-istic framework for the elevated numbers of HSCs.hus, c-Myb-p300 normally acts to repress the prolifer-tion of HSCs or links proliferation with subsequent dif-erentiation.
Previous data have shown that c-Myb is required toenerate definitive HSCs, that c-Myb−/− animals die at14.5 as a result of a lack of definitive erythropoiesis,nd that few HSCs are found in early embryos (Mucen-ki et al., 1991; Sumner et al., 2000). In contrast, partial
oss of c-Myb results in increased numbers of fetal liverrogenitor cells expressing CD34+ or Sca-1+, support-
ng a negative role for c-Myb in the control of early pro-enitors (Emambokus et al., 2003). Interestingly, manyf the phenotypes described in the c-MybM303V/M303V
ice are also found in mice homozygous for a tripleoint mutation in the KIX domain of the transcriptionaloactivator p300 (p300KIX/KIX) that disrupts the associa-ion of p300 with c-Myb (Kasper et al., 2002). As theumbers of HSCs were not examined in either of thesebove-mentioned studies, it will interesting to deter-ine if the p300KIX/KIX and other c-Myb mutants have
imilar alterations in the number of HSCs and progeni-or cells.
The current study shows that the c-Myb-p300 in-eraction controls hematopoiesis at many distinct points,oth promoting and repressing proliferation and differ-ntiation and thus highlighting c-Myb as a key regulatorf hematopoiesis. Determining how a single transcrip-ion factor is able to control the diverse processes ofelfrenewal, proliferation, and differentiation at distinctoints in hematopoiesis and defining the molecular in-eractions that control c-Myb activity remain importantreas of future research.
xperimental Procedures
lasmids and Cell Culturefull-length cDNA for murine c-Myb was obtained from Mamma-
ian Gene Collection (clone 18531). Mutations in c-Myb were intro-uced by using site-directed mutagenesis by PCR (Cormack,
1999). c-Myb-Gal4 DNA binding domain fusions were made inpBind (Promega).

c-Myb Regulates HSC Number and Differentiation165
MiceC57BL/6 N-ethyl-N-nitrosourea-mutagenized mice were generatedas described elsewhere (Miosge et al., 2002). Hematology mea-surements were taken by running whole blood through a haematol-ogy analyzer (HemaVet 3700, CDC Technologies; or a Cell Dyne3700, Abbott Diagnostics). %CD4, %CD8, and %B220 cells inwhole blood were measured by flow cytometry as described below.SNP mapping of the affected mice was performed as describedelsewhere (Wiltshire et al., 2003).
Affected F2 mice were backcrossed twice to B6 mice to remove129S1 genome contaminations and were used for subsequentanalysis.
Mixed Bone Marrow ChimerasFor bone marrow transplant experiments, the indicated number ofbone marrow cells from Ly5.2/5.2, c-Mybwt/wt, or c-MybM303V/M303V
mice, together with wild-type competitor cells Ly5.1/5.1 or Ly5.1/5.2,were injected intravenously (IV) into the tail veins of lethally irradi-ated Ly5.1/5.1 B6 mice (1200 rads). Mice were analyzed at 8 or 12weeks posttransplant and were stained for CD3, B220, Mac1/Gr1,Ly5.1, and Ly5.2 as described below. For competitive repopulationassays, the donor to competitor engraftment ratio was calculatedas %Ly5.2/5.2/%Ly5.1/5.2. Mice that failed to engraft with either donoror competitor cells (%Ly5.2/5.2 + %Ly5.1/5.2) < 1% were not scored.For secondary transplant experiments, 5 × 106 BM cells fromanimals originally transplanted with 0.5 × 106 Ly5.2/5.2 WT orc-MybM303V/M303V bone marrow were injected IV into lethally irradi-ated Ly5.1/5.1 mice. Twelve weeks later, animals were scored forLy5.2/5.2 cells in the peripheral blood by flow cytometry. For tertiarytransplants, bone marrow cells were harvested from successfullyengrafted (>1% Ly5.2/5.2) secondary recipients and 106, 105, or 104
cells were injected IV into lethally irritated Ly5.1/5.1 recipients alongwith 1 × 105 Ly5.1/5.2 bone marrow cells for radioprotection. Eightweeks later, blood was stained for the percentage of donor cells.Mice that contained >1% Ly5.2/5.2 donor cells were scored as posi-tive and were used to calculate the frequency of HSC by usingL-CALC software v. 1.1 from StemSoft, Inc. (Vancouver, Canada).
AntibodiesUnless indicated otherwise, all antibodies were purchased fromPharmingen. c-Myb proteins were detected by using an antibodyspecific to the carboxy terminus of c-Myb (Santa Cruz Biotech, catsc-516). Anti-human p300 (NM11) was used in the immunoprecipi-tations.
Flow CytometryHSC/CLP lineage cocktail antibodies consisted of CD3, CD4, CD5,CD8, CD19, B220, IgM, Gr-1, and TER119. The lineage cocktail forthe CMP stain was the same as that described above, except IL-7Rα and SCA-1 were also included. For sorting HSCs, bone mar-row cells were stained first with the lineage cocktail and were thendepleted by using Anti-Rat Kappa Microbeads (Miltneyi Biotec,cat474-01) on an AutoMACS magnetic cell separator. Remainingcells were then stained with other antibodies used in the HSC stain.For DNA analysis, bone marrow cells were stained as describedabove for HSC/CLP stain, fixed in 1% formaldehyde for 30 min onice, and resuspended in 1 �g/ml DAPI (Sigma) dissolved in stainingbuffer for 45 min at 4°C.
Methylcellulose AssaysThe indicated populations of cells were purified by flow cytometryas indicated above, and 500 cells were plated into triplicate dishescontaining Methocult GF M3434 (StemCell Technologies, cat03434) supplemented with Flk, TPO, and IL11, all at 100 ng/ml.Colonies were scored 10 days after plating, and morphology wasdetermined by microscopic examination and cytospins, followedby Wright’s-Giemsa staining of pooled or individual colonies assuggested by the manufacturer.
Generation of Nuclear Extracts and ImmunoprecipitationsAapproximately 107 transfected K562 cells were resuspended in150 �l cytoplasmic lysis buffer and were incubated on ice for ap-
proximately 20 min. Nuclei were pelleted and resuspended in 75 �lNuclei extraction buffer for 30 min at 4°C. Debris was spun out,and lysate was stored at −70°C. Immunoprecipitations were per-formed as recommended by Pharmingen. EMSA assays were per-formed by transfecting 293 HEK cells with the expression vectorsfor c-Myb. EMSA was performed exactly as described previously(Monticelli et al., 2002).
Supplemental DataSupplemental Data including a more detailed Experimental Pro-cedures section, two figures, and a table are available at http://www.developmentalcell.com/cgi/content/full/8/2/153/DC1/.
Acknowledgments
We would like to thank Lacey Kischassey and The Genomics Insti-tute of the Novartis Research Foundation vivarium staff for mainte-nance of the mouse colony; Carie Dubard-Jackson, Michael Young,and Samantha Zaharevitz for phenotyping; Christopher Trussell forflow cytometry experiments; and Michael Christiansen, KarstenSauer, Ben Wen, and Andrew Miller for helpful comments on themanuscript.
Received: May 14, 2004Revised: November 18, 2004Accepted: December 16, 2004Published: February 1, 2005
References
Akashi, K., Traver, D., Miyamoto, T., and Weissman, I.L. (2000). Aclonogenic common myeloid progenitor that gives rise to all my-eloid lineages. Nature 404, 193–197.
Allen, R.D., 3rd, Bender, T.P., and Siu, G. (1999). c-Myb is essentialfor early T cell development. Genes Dev. 13, 1073–1078.
Allen, R.D., 3rd, Kim, H.K., Sarafova, S.D., and Siu, G. (2001). Nega-tive regulation of CD4 gene expression by a HES-1-c-Myb complex.Mol. Cell. Biol. 21, 3071–3082.
Badiani, P., Corbella, P., Kioussis, D., Marvel, J., and Weston, K.(1994). Dominant interfering alleles define a role for c-Myb in T-celldevelopment. Genes Dev. 8, 770–782.
Baluda, M.A., and Reddy, E.P. (1994). Anatomy of an integratedavian myeloblastosis provirus: structure and function. Oncogene 9,2761–2774.
Bender, T.P., Kremer, C.S., Kraus, M., Buch, T., and Rajewsky, K.(2004). Critical functions for c-Myb at three checkpoints during thy-mocyte development. Nat. Immunol. 5, 721–729.
Carpinelli, M.R., Hilton, D.J., Metcalf, D., Antonchuk, J.L., Hyland,C.D., Mifsud, S.L., Di Rago, L., Hilton, A.A., Willson, T.A., Roberts,A.W., et al. (2004). Suppressor screen in Mpl−/− mice: c-Myb muta-tion causes supraphysiological production of platelets in the ab-sence of thrombopoietin signaling. Proc. Natl. Acad. Sci. USA 101,6553–6558.
Cormack, B. (1999). Site-directed mutagenesis by PCR. In ShortProtocols in Molecular Biology, Fourth Edition,, F.M. Ausubel, R.Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K.Struhl, eds. (New York, NY: John Wiley and Sons), pp. 8.16–8.21.
Denburg, J.A., Messner, H., Lim, B., Jamal, N., Telizyn, S., and Bie-nenstock, J. (1985). Clonal origin of human basophil/mast cellsfrom circulating multipotent hemopoietic progenitors. Exp. Hema-tol. 13, 185–188.
Dubendorff, J.W., and Lipsick, J.S. (1999). Transcriptional regula-tion by the carboxyl terminus of c-Myb depends upon both theMyb DNA-binding domain and the DNA recognition site. Oncogene18, 3452–3460.
Dubendorff, J.W., Whittaker, L.J., Eltman, J.T., and Lipsick, J.S.(1992). Carboxy-terminal elements of c-Myb negatively regulatetranscriptional activation in cis and in trans. Genes Dev. 6, 2524–2535.
Emambokus, N., Vegiopoulos, A., Harman, B., Jenkinson, E., An-derson, G., and Frampton, J. (2003). Progression through key

Developmental Cell166
stages of haemopoiesis is dependent on distinct threshold levels t6of c-Myb. EMBO J. 22, 4478–4488.
PGodfrey, D.I., Kennedy, J., Suda, T., and Zlotnik, A. (1993). A de-rvelopmental pathway involving four phenotypically and functionallyJdistinct subsets of CD3–CD4-CD8- triple-negative adult mouse thy-
mocytes defined by CD44 and CD25 expression. J. Immunol. 150, R4244–4252. R
uGonda, T.J. (1998). The c-Myb oncoprotein. Int. J. Biochem. Cell9Biol. 30, 547–551.SHardy, R.R., and Hayakawa, K. (2001). B cell development path-oways. Annu. Rev. Immunol. 19, 595–621.SKasper, L.H., Boussouar, F., Ney, P.A., Jackson, C.W., Rehg, J., vanCDeursen, J.M., and Brindle, P.K. (2002). A transcription-factor-bind-Sing surface of coactivator p300 is required for haematopoiesis. Na-Iture 419, 738–743.wKondo, M., Scherer, D.C., King, A.G., Manz, M.G., and Weissman,tI.L. (2001). Lymphocyte development from hematopoietic stemTcells. Curr. Opin. Genet. Dev. 11, 520–526.a
Kondo, M., Weissman, I.L., and Akashi, K. (1997). Identification ofa
clonogenic common lymphoid progenitors in mouse bone marrow.3
Cell 91, 661–672.T
Lieu, Y.K., Kumar, A., Pajerowski, A.G., Rogers, T.J., and Reddy, E.P. M(2004). Requirement of c-myb in T cell development and in mature 2T cell function. Proc. Natl. Acad. Sci. USA 101, 14853–14858.
TLipsick, J.S. (1996). One billion years of Myb. Oncogene 13, 223– M235. lLipsick, J.S., and Wang, D.M. (1999). Transformation by v-Myb. On- scogene 18, 3047–3055. W
LMiosge, L.A., Blasioli, J., Blery, M., and Goodnow, C.C. (2002).Analysis of an ethylnitrosourea-generated mouse mutation defines N
aa cell intrinsic role of nuclear factor kappB2 in regulating circulatingB cell numbers. J. Exp. Med. 196, 1113–1119. S
ZMonticelli, S., Ghittoni, R., Kabesch, M., and Vercelli, D. (2002). Mybsproteins repress human Ig epsilon germline transcription by inhib-liting STAT6-dependent promoter activation. Mol. Immunol. 38,
1129–1138.
Mucenski, M.L., McLain, K., Kier, A.B., Swerdlow, S.H., Schreiner,C.M., Miller, T.A., Pietryga, D.W., Scott, W.J., Jr., and Potter, S.S.(1991). A functional c-myb gene is required for normal murine fetalhepatic hematopoiesis. Cell 65, 677–689.
Nakagoshi, H., Takemoto, Y., and Ishii, S. (1993). Functional do-mains of the human B-myb gene product. J. Biol. Chem. 268,14161–14167.
Nelms, K.A., and Goodnow, C.C. (2001). Genome-wide ENU muta-genesis to reveal immune regulators. Immunity 15, 409–418.
Ness, S.A. (1996). The Myb oncoprotein: regulating a regulator. Bio-chim. Biophys. Acta 1288, F123–F139.
Ness, S.A. (1999). Myb binding proteins: regulators and cohorts intransformation. Oncogene 18, 3039–3046.
Nomura, N., Takahashi, M., Matsui, M., Ishii, S., Date, T., Sasamoto,S., and Ishizaki, R. (1988). Isolation of human cDNA clones of myb-related genes, A-myb and B-myb. Nucleic Acids Res. 16, 11075–11089.
Oh, I.H., and Reddy, E.P. (1997). Murine A-myb gene encodes atranscription factor, which cooperates with Ets-2 and exhibits dis-tinctive biochemical and biological activities from c-myb. J. Biol.Chem. 272, 21432–21443.
Oh, I.H., and Reddy, E.P. (1998). The C-terminal domain of B-Mybacts as a positive regulator of transcription and modulates its bio-logical functions. Mol. Cell. Biol. 18, 499–511.
Oh, I.H., and Reddy, E.P. (1999). The myb gene family in cell growth,differentiation and apoptosis. Oncogene 18, 3017–3033.
Parker, D., Rivera, M., Zor, T., Henrion-Caude, A., Radhakrishnan,I., Kumar, A., Shapiro, L.H., Wright, P.E., Montminy, M., and Brindle,P.K. (1999). Role of secondary structure in discrimination betweenconstitutive and inducible activators. Mol. Cell. Biol. 19, 5601–5607.
Pearson, R., and Weston, K. (2000). c-Myb regulates the prolifera-
ion of immature thymocytes following beta-selection. EMBO J. 19,112–6120.
erry, C., and Soreq, H. (2002). Transcriptional regulation of eryth-opoiesis. Fine tuning of combinatorial multi-domain elements. Eur.. Biochem. 269, 3607–3618.
osenthal, M.A., Thompson, M.A., Ellis, S., Whitehead, R.H., andamsay, R.G. (1996). Colonic expression of c-myb is initiated intero and continues throughout adult life. Cell Growth Differ. 7,61–967.
iu, G. (2001). Linking CD4 gene expression and T cell devel-pment. Curr. Mol. Med. 1, 523–532.
mith, C. (2003). Hematopoietic stem cells and hematopoiesis.ancer Control 10, 9–16.
umner, R., Crawford, A., Mucenski, M., and Frampton, J. (2000).nitiation of adult myelopoiesis can occur in the absence of c-Mybhereas subsequent development is strictly dependent on the
ranscription factor. Oncogene 19, 3335–3342.
akahashi, T., Nakagoshi, H., Sarai, A., Nomura, N., Yamamoto, T.,nd Ishii, S. (1995). Human A-myb gene encodes a transcriptionalctivator containing the negative regulatory domains. FEBS Lett.58, 89–96.
aylor, D., Badiani, P., and Weston, K. (1996). A dominant interferingyb mutant causes apoptosis in T cells. Genes Dev. 10, 2732–
744.
hompson, M.A., Rosenthal, M.A., Ellis, S.L., Friend, A.J., Zorbas,.I., Whitehead, R.H., and Ramsay, R.G. (1998). c-Myb down-regu-
ation is associated with human colon cell differentiation, apopto-is, and decreased Bcl-2 expression. Cancer Res. 58, 5168–5175.
iltshire, T., Pletcher, M.T., Batalov, S., Barnes, S.W., Tarantino,.M., Cooke, M.P., Wu, H., Smylie, K., Santrosyan, A., Copeland,.G., et al. (2003). Genome-wide single-nucleotide polymorphismnalysis defines haplotype patterns in mouse. Proc. Natl. Acad.ci. USA 100, 3380–3385.
orbas, M., Sicurella, C., Bertoncello, I., Venter, D., Ellis, S., Mucen-ki, M.L., and Ramsay, R.G. (1999). c-Myb is critical for murine co-
on development. Oncogene 18, 5821–5830.
Copyright © 2022 FDOKUMEN




















