
Hedgehog Signal Transduction via Smoothened Association with a Cytoplasmic Complex Scaffolded by the...
14
Molecular Cell, Vol. 12, 1261–1274, November, 2003, Copyright 2003 by Cell Press Hedgehog Signal Transduction via Smoothened Association with a Cytoplasmic Complex Scaffolded by the Atypical Kinesin, Costal-2 that are the targets for this information and then intro- duce the cell membrane components that are its source. The ultimate target for Hh pathway action in Drosophi- la, where genetically and biochemically based under- standing of Hh signal transduction is most advanced, Lawrence Lum, 1,3 Chi Zhang, 1,3 Sekyung Oh, 1 Randall K. Mann, 1 Doris P. von Kessler, 1 Jussi Taipale, 1,4 Frances Weis-Garcia, 2 Ruoyu Gong, 1 Baolin Wang, 1,5 and Philip A. Beachy 1, * 1 Howard Hughes Medical Institute is the latent zinc finger transcription factor, Cubitus in- Department of Molecular Biology and Genetics terruptus (Ci). Ci activity is controlled by a multicompo- The Johns Hopkins School of Medicine nent cytoplasmic complex which, in addition to Ci, in- Baltimore, Maryland 21205 cludes the kinesin-like protein Costal-2 (Cos2; Robbins 2 Department of Immunology et al., 1997; Sisson et al., 1997) and the serine/threonine Monoclonal Antibody Core Facility kinase Fused (Fu; Preat et al., 1990); some fraction of Memorial Sloan-Kettering Cancer Center these complexes may also contain Suppressor of fused New York, New York 10021 [Su(fu); Stegman et al., 2000], a protein lacking known functional motifs (Pham et al., 1995). In the stimulated state, this complex promotes or permits steady-state Summary nuclear localization of a minute fraction of Ci protein (Chen et al., 1999), resulting in transcriptional activation The seven-transmembrane protein Smoothened (Smo) of target genes. In the unstimulated state, all full-length transduces extracellular activation of the Hedgehog Ci is localized in the cytoplasm, and a significant propor- (Hh) pathway by an unknown mechanism to increase tion of it is proteolytically processed to produce a trun- transcriptional activity of the latent cytoplasmic tran- cated repressor form, CiR, that actively suppresses tran- scription factor Ci (Cubitus interruptus). Here, we scription of target genes in the nucleus (Aza-Blanc et present evidence that Smo associates directly with a al., 1997; Methot and Basler, 1999). With regard to the Ci-containing complex that is scaffolded and stabi- Ci transcriptional effector, the unstimulated state of the lized by the atypical kinesin, Costal-2 (Cos2). This Hh pathway thus is defined by absence of nuclear Ci complex constitutively suppresses pathway activity, and production of CiR, whereas the stimulated state is but Hh signaling reverses its regulatory effect to pro- distinguished by absence of CiR and nuclear presence mote Ci-mediated transcription. In response to Hh ac- of Ci. tivation of Smo, Cos2 mediates accumulation and Increased nuclear localization of Ci in the stimulated phosphorylation of Smo at the membrane as well as state or proteolytic production of CiR in the unstimulated phosphorylation of the cytoplasmic components Fu state each result from the operation of distinct cellular and Su(fu). Positive response of Cos2 to Hh stimulation processes. The central transductory mechanism thus is requires a portion of the Smo cytoplasmic tail and the not simply switched on or off upon Hh stimulation but Cos2 cargo domain, which interacts directly with Smo. rather alternates between two states that are actively maintained. Early genetic analyses assigned simple Introduction positive or negative roles to pathway components based on mutant phenotypes. Losses of Fu or Ci function thus produce phenotypic effects similar to loss of Hh func- Genetic studies from insects to humans have estab- tion, suggesting reduction of pathway activity and a lished the critical biological roles of the Hedgehog (Hh) positive role in the pathway for these components. Loss signaling pathway in many aspects of embryonic pat- of Cos2 in turn appears to cause a gain in pathway terning (Ingham and McMahon, 2001; Muenke and activation, suggesting a negative regulatory role for Beachy, 2001) and in tumorigenesis (Taipale and Beachy, Cos2. More recent analyses, however, have revealed 2001). Fundamental questions remain, however, regard- greater complexity in the function of some of these com- ing the mechanism by which the extracellular Hh protein ponents. For example, Cos2 appears to be required signal elicits intracellular responses that direct cell pro- positively in cells just anterior to the anterior/posterior liferation and differentiation. Thus, although our current compartment boundary for expression of engrailed, and understanding of Hh signal transduction encompasses other targets associated with the highest levels of Hh several critical components and events, we do not yet pathway activation (Wang et al., 2000; Wang and Holm- have an answer to the fundamental question of how gren, 2000). One additional attribute of Cos2 is its appar- intracellular components first receive information re- ent ability to autonomously effect normal pathway regu- garding pathway activation from the cell membrane. As lation in the combined absence of Fu and Su(fu), as a foundation for addressing this question, we summarize revealed by the striking ability of Su(fu) mutations to what is known of the intracellular pathway components restore normal patterning to fu mutant individuals (Preat, 1992). *Correspondence: [email protected] We now consider the membrane components that 3 These authors contributed equally to this work. function upstream of this cytoplasmic apparatus to me- 4 Present address: Molecular/Cancer Biology Program, University of diate Hh signaling. Activation of the pathway is triggered Helsinki, Helsinki, Finland. by stoichiometric binding of Hh ligand to Patched (Ptc), 5 Present address: Institute of Genetic Medicine, Joan and Sanford I. Weill Medical College, New York, NY 10021. a transporter-like protein that in the absence of Hh sup-
Transcript of Hedgehog Signal Transduction via Smoothened Association with a Cytoplasmic Complex Scaffolded by the...

Molecular Cell, Vol. 12, 1261–1274, November, 2003, Copyright 2003 by Cell Press
Hedgehog Signal Transduction via SmoothenedAssociation with a Cytoplasmic Complex Scaffoldedby the Atypical Kinesin, Costal-2
that are the targets for this information and then intro-duce the cell membrane components that are its source.
The ultimate target for Hh pathway action in Drosophi-la, where genetically and biochemically based under-standing of Hh signal transduction is most advanced,
Lawrence Lum,1,3 Chi Zhang,1,3 Sekyung Oh,1
Randall K. Mann,1 Doris P. von Kessler,1
Jussi Taipale,1,4 Frances Weis-Garcia,2
Ruoyu Gong,1 Baolin Wang,1,5 and Philip A. Beachy1,*1Howard Hughes Medical Institute
is the latent zinc finger transcription factor, Cubitus in-Department of Molecular Biology and Geneticsterruptus (Ci). Ci activity is controlled by a multicompo-The Johns Hopkins School of Medicinenent cytoplasmic complex which, in addition to Ci, in-Baltimore, Maryland 21205cludes the kinesin-like protein Costal-2 (Cos2; Robbins2 Department of Immunologyet al., 1997; Sisson et al., 1997) and the serine/threonineMonoclonal Antibody Core Facilitykinase Fused (Fu; Preat et al., 1990); some fraction ofMemorial Sloan-Kettering Cancer Centerthese complexes may also contain Suppressor of fusedNew York, New York 10021[Su(fu); Stegman et al., 2000], a protein lacking knownfunctional motifs (Pham et al., 1995). In the stimulatedstate, this complex promotes or permits steady-stateSummarynuclear localization of a minute fraction of Ci protein(Chen et al., 1999), resulting in transcriptional activationThe seven-transmembrane protein Smoothened (Smo)of target genes. In the unstimulated state, all full-lengthtransduces extracellular activation of the HedgehogCi is localized in the cytoplasm, and a significant propor-(Hh) pathway by an unknown mechanism to increasetion of it is proteolytically processed to produce a trun-transcriptional activity of the latent cytoplasmic tran-cated repressor form, CiR, that actively suppresses tran-scription factor Ci (Cubitus interruptus). Here, wescription of target genes in the nucleus (Aza-Blanc et
present evidence that Smo associates directly with aal., 1997; Methot and Basler, 1999). With regard to the
Ci-containing complex that is scaffolded and stabi-Ci transcriptional effector, the unstimulated state of the
lized by the atypical kinesin, Costal-2 (Cos2). ThisHh pathway thus is defined by absence of nuclear Ci
complex constitutively suppresses pathway activity,and production of CiR, whereas the stimulated state is
but Hh signaling reverses its regulatory effect to pro- distinguished by absence of CiR and nuclear presencemote Ci-mediated transcription. In response to Hh ac- of Ci.tivation of Smo, Cos2 mediates accumulation and Increased nuclear localization of Ci in the stimulatedphosphorylation of Smo at the membrane as well as state or proteolytic production of CiR in the unstimulatedphosphorylation of the cytoplasmic components Fu state each result from the operation of distinct cellularand Su(fu). Positive response of Cos2 to Hh stimulation processes. The central transductory mechanism thus isrequires a portion of the Smo cytoplasmic tail and the not simply switched on or off upon Hh stimulation butCos2 cargo domain, which interacts directly with Smo. rather alternates between two states that are actively
maintained. Early genetic analyses assigned simpleIntroduction positive or negative roles to pathway components based
on mutant phenotypes. Losses of Fu or Ci function thusproduce phenotypic effects similar to loss of Hh func-Genetic studies from insects to humans have estab-tion, suggesting reduction of pathway activity and alished the critical biological roles of the Hedgehog (Hh)positive role in the pathway for these components. Losssignaling pathway in many aspects of embryonic pat-of Cos2 in turn appears to cause a gain in pathwayterning (Ingham and McMahon, 2001; Muenke andactivation, suggesting a negative regulatory role forBeachy, 2001) and in tumorigenesis (Taipale and Beachy,Cos2. More recent analyses, however, have revealed2001). Fundamental questions remain, however, regard-greater complexity in the function of some of these com-ing the mechanism by which the extracellular Hh proteinponents. For example, Cos2 appears to be requiredsignal elicits intracellular responses that direct cell pro-positively in cells just anterior to the anterior/posteriorliferation and differentiation. Thus, although our currentcompartment boundary for expression of engrailed, andunderstanding of Hh signal transduction encompassesother targets associated with the highest levels of Hhseveral critical components and events, we do not yetpathway activation (Wang et al., 2000; Wang and Holm-have an answer to the fundamental question of howgren, 2000). One additional attribute of Cos2 is its appar-intracellular components first receive information re-ent ability to autonomously effect normal pathway regu-garding pathway activation from the cell membrane. Aslation in the combined absence of Fu and Su(fu), asa foundation for addressing this question, we summarizerevealed by the striking ability of Su(fu) mutations towhat is known of the intracellular pathway componentsrestore normal patterning to fu mutant individuals(Preat, 1992).
*Correspondence: [email protected] We now consider the membrane components that3 These authors contributed equally to this work.
function upstream of this cytoplasmic apparatus to me-4 Present address: Molecular/Cancer Biology Program, University ofdiate Hh signaling. Activation of the pathway is triggeredHelsinki, Helsinki, Finland.by stoichiometric binding of Hh ligand to Patched (Ptc),5 Present address: Institute of Genetic Medicine, Joan and Sanford
I. Weill Medical College, New York, NY 10021. a transporter-like protein that in the absence of Hh sup-

Molecular Cell1262
presses activity of the seven transmembrane protein that can be used to study biochemical changes associ-ated with Hh stimulation. Previous studies have demon-Smoothened (Smo; Taipale et al., 2002). Smo activity
indeed can be modulated by direct binding to exoge- strated that wing imaginal disc-derived cl-8 cells canproduce a faithful and physiologically meaningful tran-nously added small molecules (Chen et al., 2002a,
2002b; Frank-Kamenetsky et al., 2002), suggesting that scriptional response to Hh stimulation (Chen et al., 1999;Lum et al., 2003). To further validate this system and asPtc might normally suppress Smo through transport of
an endogenous small molecule effector. Whatever the a baseline for studies of transduction, we examined thekinetics of phosphorylation and accumulation of Hhmechanism of Ptc action, pathway activation results
from reduction or loss of Ptc activity that can be caused pathway components in cl-8 cells. Most pathway com-ponents, including Smo, Cos2, Fu, and Ci, have beeneither by Hh binding or by ptc mutation, and pathway
activation in either circumstance requires the activity of reported to undergo rapid changes in phosphorylationupon pathway activation (Therond et al., 1996; RobbinsSmo. This essential role of Smo suggests that it may be
the critical membrane component that relays informa- et al., 1997; Wang and Holmgren, 1999; Chen et al.,1999; Denef et al., 2000). In addition, Smo, Ci, and Ptction regarding state of pathway activation to cyto-
plasmic components. proteins have been reported to accumulate to higherlevels as a consequence of posttranslational regulationIts sequence and predicted transmembrane structure
clearly identify Smo as a member of the family of seven (Aza-Blanc et al., 1997; Alcedo et al., 2000; Denef etal., 2000) or increased gene transcription (Forbes et al.,transmembrane receptors that in general transduce ex-
tracellular signals by coupling to heterotrimeric GTP 1993; Chen et al., 1999).Phosphorylated forms of Fu and Cos2 (Fu-P andbinding proteins (GPCRs). This similarity, together with
a genetically revealed requirement for PKA in pathway Cos2-P) were first detected within 15 min of addition ofHhN-conditioned medium (HhN medium; Figure 1A). Weregulation, led to early proposals for Smo function in
which activation of G�i by Smo produces a decrease in also noted the appearance of a small fraction of Su(fu)as a slower migrating doublet, in a time scale similar tocAMP levels and PKA activity to activate the pathway
(Alcedo et al., 1996; van den Heuvel and Ingham, 1996). that for the appearance of Fu-P and Cos2-P. Treatmentof cl-8 cell lysates with lambda phosphatase (�PPase)The modulation of PKA activity, however, is not required
for pathway activation, and low levels of unregulated caused the slower-migrating Su(fu) species to collapseinto a single species that comigrates with Su(fu) ob-PKA activity suffice to maintain the regulated state of
the Hh pathway (Ohlmeyer and Kalderon, 1998). Thus, served in the absence of Hh (Figure 1B), indicating thatthe slower migrating Su(fu) species result from phos-it remains unclear how information regarding Smo acti-
vation is transmitted to the cytoplasmic complex. Phar- phorylation. Western blot analysis of extracts from Dro-sophila embryos revealed similar slow-migrating forms,macological and mutational data do suggest, as is
widely believed for other members of the seven trans- suggesting that Su(fu) phosphorylation also occurs innormal development (Figure 1C). Even after prolongedmembrane receptor family (Bourne, 1997), that activa-
tion of Smo may be mediated by a conformational shift Hh stimulation, Su(fu)-P constitutes only a minor fractionof the total Su(fu) protein (Figure 1A). This suggests that(Taipale et al., 2000). Furthermore, in Drosophila, path-
way activation is accompanied by accumulation of in- only a fraction of Su(fu) is affected by the processes thatcause production of Cos2-P and Fu-P in response to Hh.creased levels of a phosphorylated form of Smo at the
plasma membrane (Denef et al., 2000). As described previously, Smo accumulates and be-comes phosphorylated in response to Hh in a similarIn this work, we approach the question of transduction
from membrane to cytoplasm by systematically explor- time scale as accumulation of Fu-P. Ci accumulationlikely reflects changes in proteolytic processing as pre-ing the effects of RNAi-targeting of individual pathway
components on biochemical markers of pathway activ- viously described (Aza-Blanc et al., 1997). Accumulationof Ptc protein is apparent by 4 hr, consistent with tran-ity. The kinesin-like protein Cos2 in particular appears
to play a central role, as it mediates multiple events scriptional activation of the ptc gene in response to Hh.Overall, these observations indicate that cl-8 cells nottriggered by Hh-signaling, including phosphorylation
and accumulation of Smo and Fu, and phosphorylation only have the capacity to produce a physiologicallymeaningful transcriptional response to the Hh signal,of Su(fu). We also demonstrate that the cytoplasmic
components Cos2, Fu, and Ci form a native complex but also that modification and accumulation of pathwaycomponents in response to Hh are faithfully repre-with Smo and that this interaction is mediated by a direct
interaction between the Smo cytotail and the Cos2 stalk sented.and cargo domains. Based on the results presentedhere, we propose a model for Hh signal transduction Cos2 Is Required for Fu Protein Stabilitythat involves direct transmission of information from To examine the roles of individual components in bio-Smo at the plasma membrane to cytoplasmic compo- chemical events associated with pathway activation, wenents via Cos2. utilized the S2-related S2R� embryo-derived cell line,
in which treatment with dsRNA uniformly produces theRNAi effect in all cells of the culture (Clemens et al.,Results2000). Although S2R� cells do not express Ci and lacka transcriptional response (data not shown), they retainHh-Induced Accumulation and Phosphorylation
of Pathway Components in cl-8 Cells the posttranslational changes in most pathway compo-nents in response to Hh (Figure 1D, lanes 1 and 2).Our studies of Hh signal transduction largely are based
in cultured cells, which provide homogeneous material Furthermore, as expected, phosphorylations of Cos2,

Hedgehog Signaling via Cos2 Association with Smo1263
Figure 1. Hh-Induced Protein Stabilization and Modification of Pathway Components
(A) Western blot analysis of pathway components in cl-8 cells upon HhN stimulation. Proteins in the unstimulated state or modified in responseto HhN are noted (black and blue arrows, respectively). Loading control: �-tubulin.(B) Su(fu) is phosphorylated in response to HhN. Lysates from HhN-treated cl-8 cells (24 hr) were incubated in the presence/absence oflambda phosphatase (�PPase, 200 U) and the phosphatase inhibitor Na3VO4 (1 mM), prior to Western blotting.(C) Su(fu) protein in Drosophila embryos. The pattern of slower migrating Su(fu) proteins is similar to that observed in HhN-stimulated cl-8 cells.(D) RNAi of Cos2 reduces levels of Fu expression, and of Hh-induced phosphorylation of Smo, Fu, and Su(fu). RNAi in S2R� cells (2 days)was followed by incubation with HhN (24 hr) and Western blot analysis. Faster-migrating Smo in unstimulated cells can be detected uponlonger exposure, and slower migration of Smo in stimulated cells is due to phosphorylation (Supplemental Figure S1A).(E) RNAi of Fu decreases Hh-dependent Su(fu) phosphorylation (lanes 6 and 2). Increased Su(fu) phosphorylation, resulting from RNAi of Ptc(lanes 3 and 4), is also decreased by RNAi of Fu (lanes 7 and 8).(F) Role of pathway components in biochemical events associated with pathway activity. The blue shading highlights functions of pathwaycomponents that, to the best of our knowledge, have not previously been reported.

Molecular Cell1264
Fu, and Su(fu) are affected by RNAi of Ptc and Smo, (Figure 1E, compare lanes 7 and 3) is also reduced byRNAi directed against Fu. As noted above, RNAi of Cos2,demonstrating that these events are both Hh- and Smo
dependent (Figure 1D, lanes 3,4 and 5,6). Surprisingly, which reduces Fu levels, also results in loss of Su(fu)phosphorylation. These results suggest that Su(fu) mayRNAi targeting of Cos2 (Figure 1D, lanes 7 and 8) caused,
in addition to loss of Cos2 protein, a dramatic reduction be a substrate for Fu kinase activity, but do not ruleout the possibility of an indirect role for Fu in Su(fu)of Fu protein and a reduction of Smo protein in the Hh-
stimulated state (discussed below); no change in Su(fu) phosphorylation.The observations presented thus far are summarizedprotein levels was noted. Similar results were observed
in S2 cells (see Supplemental Figure S1 at http://www. in Figure 1F. Most notably, Cos2 activity is required notonly for phosphorylation of Fu and Su(fu) but also formolecule.org/cgi/content/full/12/5/1261/DC1).
The dramatic dependence of Fu protein levels on Cos2 stabilization and phosphorylation of Smo, suggesting arole for Cos2 in mediating bidirectional exchange be-function suggests that all or most Fu associates with
Cos2. To better understand the stoichiometric relation- tween membrane and cytoplasm.ship between Cos2 and Fu, we generated monoclonalantibodies (mAbs) that are able to IP endogenous Cos2 Interactions between Smo and Componentsand Fu proteins (see Supplemental Table S1). Immuno- of the Cytoplasmic Complexprecipitations using these mAbs revealed similar relative To purify and identify proteins that may mediate transferintensities of bands containing Cos2 and Fu, suggesting of information between Smo and the cytoplasmic com-that Cos2 and Fu are stably associated at a constant plex, we generated additional mAbs able to immunopre-ratio in cl-8 (Figure 2A, lanes 1–6) and S2 cells (Figure cipitate (IP) native Smo and Su(fu) protein (see Supple-2A, lanes 7–10; data not shown), in the presence or mental Table S1). Using silver-stains of complexesabsence of HhN stimulation. To further examine the sta- purified using the �-Smo matrix (see Experimental Pro-bilizing role of Cos2 on Fu in vivo, we monitored Fu cedures) and fractionated by SDS-PAGE, we identifiedlevels by immunostaining wing imaginal discs con- three bands (labeled 1, 2, and 3 in the HhN-stimulatedtaining clones that lacked Cos2 function. We found that sample; Figure 3A, lane 4) that were not present in theclones homozygous for cos2W1 identified by loss of control IP with IgG matrix (lane 1). Proteins in Bands 1�-Myc mAb staining show dramatically reduced levels and 3 were both enriched in the HhN-stimulated sampleof Fu in both the anterior (Figure 2B) and posterior (data (Figure 3A, compare lanes 3 and 4), and Band 3 ap-not shown) compartments of the disc. peared to migrate more diffusely. Tandem mass spec-
trometry coupled to microcapillary-based chromatogra-Hh-Induced Phosphorylation of Smo, Fu, and Su(fu) phy of trypsin-digested peptides from Band 3 identifiedand Accumulation of Smo All Require Cos2 seven Smo peptides (Figure 3B; see also SupplementalIn addition to a marked effect on levels of Fu protein, Table S2, consistent with the expected migration of Smoloss of Cos2 also reduced Hh-induced phosphorylation and with the Hh-induced accumulation and phosphory-of Fu and Su(fu), suggesting a role for Cos2 in mediating lation of Smo protein (see above). The identity of Smoinformation transfer from the membrane to cytoplasmic was confirmed by Western blot analysis (Figure 3A,components. Thus, for example, RNAi-induced loss of lanes 7 and 8; Smo), and no peptides from any otherCos2 not only abolished Hh-induced phosphorylation proteins could be identified, suggesting that Smo is theof Su(fu) but also dramatically reduced phosphorylation most abundant protein migrating within the Band 3of the residual Fu protein present in HhN-stimulated region.S2R� cells (Figure 1D, lanes 7 and 8). Perhaps the most Mass spectrometry analysis of material from Band 1surprising consequence of Cos2 loss was the failure of (Figure 3A, lane 4) identified exclusively Cos2 and Cicells to accumulate Hh-induced Smo-P (Figure 1D, lanes peptides, suggesting that these comigrating proteins7 and 8; Supplemental Figure S1B). We confirmed this associate with Smo and constitute the major specieseffect in vivo, again by immunostaining wing imaginal within Band 1. Using the �-Cos2 matrix and Westerndiscs containing homozygous cos2W1 clones (Figure 2C blotting, we also were able to detect Smo as a proteinand Supplemental Figure S2). We noted a marked reduc- copurified with Cos2 from HhN-stimulated cells (Figuretion of Smo levels in such clones occurring in the anterior 3A, lane 6), although no discrete band was evident bycompartment, particularly near the compartment bound- silver staining (Figure 3A, lane 2), possibly because ofary, where Smo levels normally are elevated due to inac- the diffuse migration of phosphorylated Smo upon HhNtivation of Ptc by Hh stimulation (Denef et al., 2000); no stimulation. As Fu kinase is known to associate withdifference in levels of Smo was noted in cos2W1 clones Cos2, we also analyzed peptides from Bands 4 and 6in the posterior compartment. (Figure 3A, lanes 2 and 4), which are strong apparent
background bands that migrate in the approximaterange expected for Fu (�90 kDa). We found that FuFu Is Required for Su(fu) Phosphorylation
The phenotypic effects of fu mutations are suppressed peptides indeed are the major non-IgG species in thesebands. This identification was confirmed by analysis ofby Su(fu) mutations (Preat, 1992), indicating that the
major role of Fu is to oppose Su(fu) function and sug- Band 7 in the �-Cos2 matrix purification (Figure 3A,lane 2), which was identified as Fu and likely representsgesting the biochemical model that Fu kinase may do
so by phosphorylating Su(fu) protein. Indeed, RNAi of unphosphorylated Fu.These results show that the major non-IgG proteinsFu results in a decrease in Fu levels and a corresponding
loss of Su(fu) phosphorylation (Figure 1E, compare lanes present following immunoaffinity purification with the�-Smo matrix were Cos2 and Ci, and Fu, which co-6 and 2). Su(fu) phosphorylation induced by RNAi of Ptc
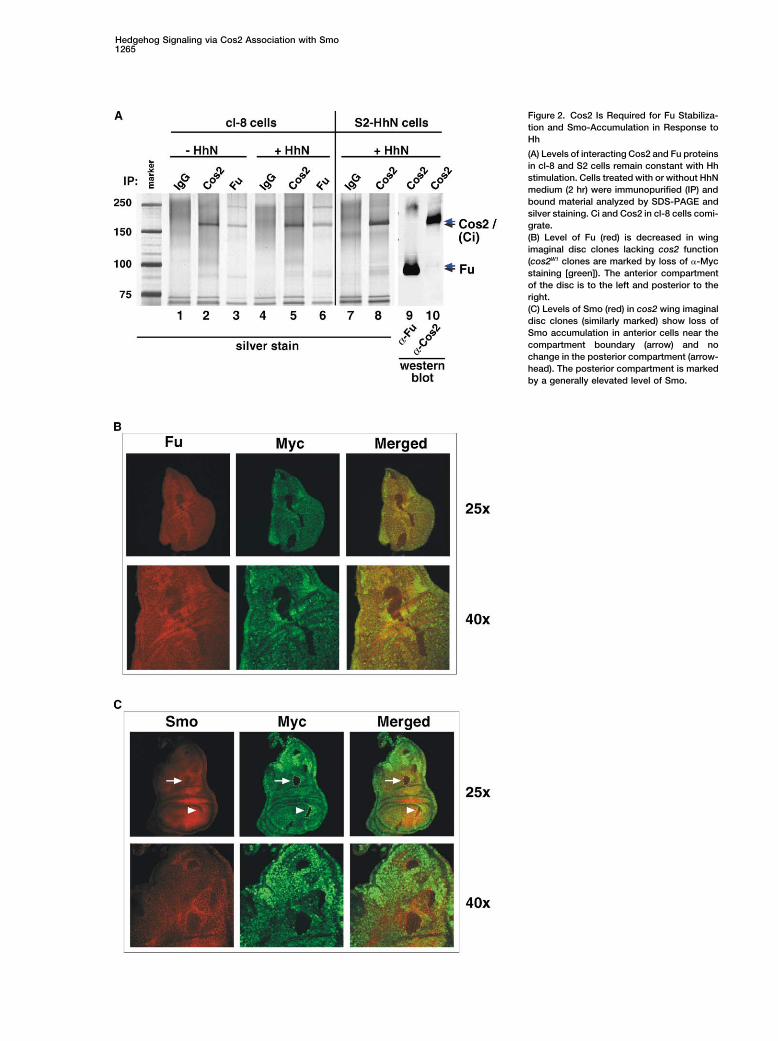
Hedgehog Signaling via Cos2 Association with Smo1265
Figure 2. Cos2 Is Required for Fu Stabiliza-tion and Smo-Accumulation in Response toHh
(A) Levels of interacting Cos2 and Fu proteinsin cl-8 and S2 cells remain constant with Hhstimulation. Cells treated with or without HhNmedium (2 hr) were immunopurified (IP) andbound material analyzed by SDS-PAGE andsilver staining. Ci and Cos2 in cl-8 cells comi-grate.(B) Level of Fu (red) is decreased in wingimaginal disc clones lacking cos2 function(cos2W1 clones are marked by loss of �-Mycstaining [green]). The anterior compartmentof the disc is to the left and posterior to theright.(C) Levels of Smo (red) in cos2 wing imaginaldisc clones (similarly marked) show loss ofSmo accumulation in anterior cells near thecompartment boundary (arrow) and nochange in the posterior compartment (arrow-head). The posterior compartment is markedby a generally elevated level of Smo.
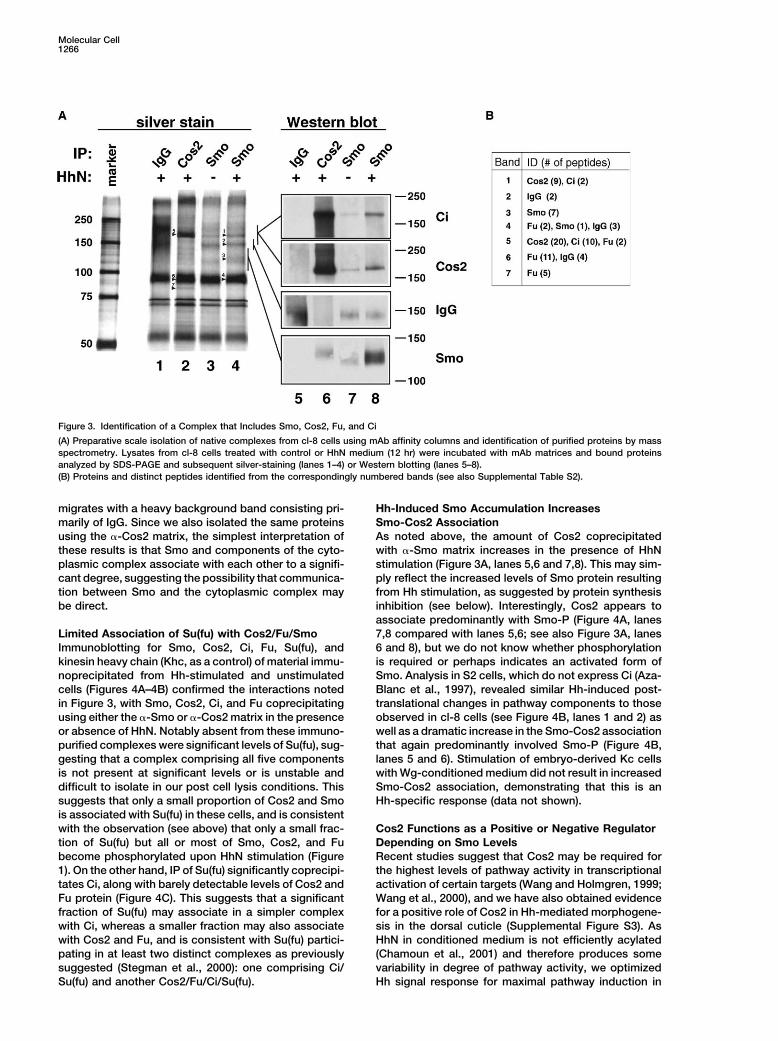
Molecular Cell1266
Figure 3. Identification of a Complex that Includes Smo, Cos2, Fu, and Ci
(A) Preparative scale isolation of native complexes from cl-8 cells using mAb affinity columns and identification of purified proteins by massspectrometry. Lysates from cl-8 cells treated with control or HhN medium (12 hr) were incubated with mAb matrices and bound proteinsanalyzed by SDS-PAGE and subsequent silver-staining (lanes 1–4) or Western blotting (lanes 5–8).(B) Proteins and distinct peptides identified from the correspondingly numbered bands (see also Supplemental Table S2).
migrates with a heavy background band consisting pri- Hh-Induced Smo Accumulation IncreasesSmo-Cos2 Associationmarily of IgG. Since we also isolated the same proteins
using the �-Cos2 matrix, the simplest interpretation of As noted above, the amount of Cos2 coprecipitatedwith �-Smo matrix increases in the presence of HhNthese results is that Smo and components of the cyto-
plasmic complex associate with each other to a signifi- stimulation (Figure 3A, lanes 5,6 and 7,8). This may sim-ply reflect the increased levels of Smo protein resultingcant degree, suggesting the possibility that communica-
tion between Smo and the cytoplasmic complex may from Hh stimulation, as suggested by protein synthesisinhibition (see below). Interestingly, Cos2 appears tobe direct.associate predominantly with Smo-P (Figure 4A, lanes7,8 compared with lanes 5,6; see also Figure 3A, lanesLimited Association of Su(fu) with Cos2/Fu/Smo
Immunoblotting for Smo, Cos2, Ci, Fu, Su(fu), and 6 and 8), but we do not know whether phosphorylationis required or perhaps indicates an activated form ofkinesin heavy chain (Khc, as a control) of material immu-
noprecipitated from Hh-stimulated and unstimulated Smo. Analysis in S2 cells, which do not express Ci (Aza-Blanc et al., 1997), revealed similar Hh-induced post-cells (Figures 4A–4B) confirmed the interactions noted
in Figure 3, with Smo, Cos2, Ci, and Fu coprecipitating translational changes in pathway components to thoseobserved in cl-8 cells (see Figure 4B, lanes 1 and 2) asusing either the �-Smo or �-Cos2 matrix in the presence
or absence of HhN. Notably absent from these immuno- well as a dramatic increase in the Smo-Cos2 associationthat again predominantly involved Smo-P (Figure 4B,purified complexes were significant levels of Su(fu), sug-
gesting that a complex comprising all five components lanes 5 and 6). Stimulation of embryo-derived Kc cellswith Wg-conditioned medium did not result in increasedis not present at significant levels or is unstable and
difficult to isolate in our post cell lysis conditions. This Smo-Cos2 association, demonstrating that this is anHh-specific response (data not shown).suggests that only a small proportion of Cos2 and Smo
is associated with Su(fu) in these cells, and is consistentwith the observation (see above) that only a small frac- Cos2 Functions as a Positive or Negative Regulator
Depending on Smo Levelstion of Su(fu) but all or most of Smo, Cos2, and Fubecome phosphorylated upon HhN stimulation (Figure Recent studies suggest that Cos2 may be required for
the highest levels of pathway activity in transcriptional1). On the other hand, IP of Su(fu) significantly coprecipi-tates Ci, along with barely detectable levels of Cos2 and activation of certain targets (Wang and Holmgren, 1999;
Wang et al., 2000), and we have also obtained evidenceFu protein (Figure 4C). This suggests that a significantfraction of Su(fu) may associate in a simpler complex for a positive role of Cos2 in Hh-mediated morphogene-
sis in the dorsal cuticle (Supplemental Figure S3). Aswith Ci, whereas a smaller fraction may also associatewith Cos2 and Fu, and is consistent with Su(fu) partici- HhN in conditioned medium is not efficiently acylated
(Chamoun et al., 2001) and therefore produces somepating in at least two distinct complexes as previouslysuggested (Stegman et al., 2000): one comprising Ci/ variability in degree of pathway activity, we optimized
Hh signal response for maximal pathway induction inSu(fu) and another Cos2/Fu/Ci/Su(fu).

Hedgehog Signaling via Cos2 Association with Smo1267
into cl-8 cells different ratios of constructs for expres-sion of Cos2 and Smo (Cos2 and Smo constructs). Inthese experiments, we cotransfected full-length Hh to-gether with reporter construct, which produces maximalpathway activity, as indicated by the lack of increasedreporter expression upon addition of HhN medium aloneor in combination with RNAi of Ptc (Figure 5A). Transfec-tion of the Smo construct in these maximally stimulatedcells produced an increase in the level of reporter ex-pression, whereas introduction of the Cos2 constructcaused a decrease in reporter expression (Figure 5B,dashed lines). Cotransfection of these two constructs,however, produced a more complex pattern of changesin reporter activity. At a fixed level of Smo construct, asCos2 construct levels increased, reporter activity firstrose, then decreased to levels like those of control trans-fected cells, and then, at highest levels of Cos2, weresignificantly depressed below control levels (Figure 5B).It is notable that increasing Cos2 expression in maxi-mally stimulated cells actually increased the level ofpathway activity in the presence of higher Smo expres-sion (10 or 100 ng Smo construct). The activity of Cos2thus appears to depend on relative levels of Smo activ-ity, and higher levels of Smo in maximally stimulatedcells apparently can convert the regulatory effect ofCos2 from negative to positive. We note that the Smoand Cos2 constructs produce similar protein levels, asmeasured with the use of Smo- and Cos2-Renilla lucifer-ase fusion proteins (Smo-R and Cos2-R, respectively;inset, Figure 5B).
The Smo Cytotail Is Required for Hh ResponseTo identify Smo determinants important for communi-cating with cytoplasmic components, we focused onthe predicted carboxy-terminal tail, as this represents
Figure 4. Hh-Induced Accumulation of a Smo-Cos2-Fu-Ci Complex the largest cytoplasmic segment of Smo that could beLysates from cl-8 cells (A) treated with control or HhN medium involved. We generated a series of Smo constructs with(2 hr), and S2 parental or stable HhN-transfected cell lines (B), were progressively larger C-terminal deletions or with muta-incubated with mAb matrices and bound proteins analyzed by West- tions in conserved potential phosphorylation sites (Fig-ern blotting. Boxed region indicates a shorter exposure. Negative
ure 6A). To test the function of these mutant proteins, wecontrol: kinesin heavy chain (Khc). (C) Su(fu) associates predomi-targeted endogenous Smo mRNA using RNAi directednantly with Ci. Bound proteins from cl-8 cell lysates incubated with
mAb matrices were analyzed by Western blotting. Little Fu, Cos2, against the 5� UTR and cotransfected trimmed and mu-and no Smo (not shown) were detectable in �-Su(fu) IP, and no tated Smo constructs lacking the 5� UTR (Figure 6B).Su(fu) was detected in �-Cos2 IP. Whereas RNAi targeting endogenous Smo strongly sup-
pressed pathway response, cotransfection with wild-type Smo construct lacking the 5�UTR restored full activ-our cl-8 cell reporter assay (Chen et al., 1999) by usingity. We found that mutations in most conserved residuesRNAi to decrease levels of Ptc. In this context of maximaland all of the potential phosphorylation sites tested hadpathway activity, additional RNAi of Cos2 dramaticallylittle or no effect on Smo function, whereas C-terminaldecreased the level of reporter expression (Figure 5A).truncations with deletions more extensive than N686This requirement for Cos2 in maximal pathway activation(RRN�C, containing Smo residues 1–686; see Figure 6A)was apparent even when RNAi of Cos2 was not fullyfailed to restore pathway function. Immunostaining ofeffective, as indicated by a somewhat weaker basal acti-wild-type and truncated Smo proteins expressed in cl-8vation in the experiment shown (Figure 5A; compare tocells revealed similar expression levels and subcellularLum et al., 2003). Combined RNAi of Su(fu) with Cos2localizations, indicating that loss of function likely doesrestored the maximal induced level of pathway activity,not result from altered subcellular localization or mis-consistent with a previously proposed role for Cos2 infolding (data not shown).suppressing Su(fu) function (Wang et al., 2000). A possi-
ble mechanism for Cos2-mediated inactivation Su(fu) issuggested by the requirement of Cos2 for stabilization/ Smo Cytotail Is Required for Direct
Association with Cos2phosphorylation of Fu and for Fu-dependent phosphory-lation of Su(fu) (see above). Cos2-R was coexpressed with Smo variants in S2 cells
and was coprecipitated with �-Smo matrix. By measur-To test whether Smo levels might specify the regula-tory effect of Cos2 in pathway activation, we introduced ing the associated Renilla luciferase activity, we deter-

Molecular Cell1268
Figure 5. A Positive Role for Cos2 in High-Level Hh Signal Response
(A) Cos2 is required for highest levels of Hhpathway activity in cl-8 cells. Activity of aptc-luciferase reporter (normalized to controlRenilla luciferase) was used to measure Hhpathway response. RNAi of Ptc maximizesresponse to HhN medium (dashed line; seetext) in a Smo-dependent manner. CombinedRNAi of Cos2 and Ptc dramatically reducesreporter activity, indicating a requirement forCos2 function in maximal Smo-mediatedpathway activity. RNAi of Ptc alone does notinduce constitutive reporter activity due to acompensatory increase in ptc mRNA expres-sion (Lum et al., 2003). Cotransfection of full-length Hh with reporter construct also resultsin maximal levels of pathway activity. Foldinduction by HhN is noted above.(B) Relative levels of Smo and Cos2 deter-mine effect on Hh pathway activity. In cellscotransfected with Hh expression constructto produce maximal pathway activity, re-porter activity at fixed levels of Smo increasesat lower levels of added Cos2 but decreasesat highest levels. Dashed lines indicate levelsof reporter activity with 0.1 �g of Smo alone(top), Cos2 alone (bottom), or YFP control(middle). Inset: 0.1 �g of Smo- or Cos2-Renilla luciferase fusion proteins (Smo-R andCos2-R, respectively) were transfected intocl-8 cells followed by incubation with controlor HhN medium. Similar levels of each proteinare noted, and HhN stimulation does notcause accumulation of Smo-R, likely due tohigh constitutive levels of expression.
mined that Cos2-R associates with wild-type Smo, less both of these domains lack determinants for Fu and Cibinding (Monnier et al., 2002; C.Z. and P.A.B., unpub-efficiently with the RRN�C protein, and not with the
more extreme Smo truncations (Figure 6C). The Smo lished data), a role for mammalian homologs of theseproteins in COS1 cell lysates seems unlikely.cytotail determinants required for rescue of Smo signal-
ing activity and for association with Cos2 thus are coinci-dent. The determinants of Cos2 required for interaction Cos2 Cargo Domain Is Required
for Smo Responsivenesswith Smo were determined by cotransfecting constructsencoding Smo-R with Myc epitope-tagged Cos2 subdo- To test the role of Cos2 cargo and stalk domains in signal
transduction via Smo, we first measured the effects onmains (Figure 6D, diagram). We observed considerableRenilla luciferase activity in �-Myc immunoprecipitated reporter activity of coexpressing in cl-8 cells Cos2 lack-
ing either the cargo domain alone (�cargo) or Cos2 lack-samples with full-length Cos2, as well as with stalk andcargo domains (Figure 6D, graph). ing both cargo and stalk domains. Whereas the latter
deletion is severely compromised in activity (data notWe tested in vitro the possibility of a direct interactionbetween the Smo cytotail and Cos2 using a purified, shown), �cargo retains full potency in pathway suppres-
sion as compared to wild-type Cos2 (Figure 6F; redbacterially expressed GST fusion protein containingSmo cytotail sequences extending from Ser557 to tracings). Both wild-type Cos2 and �cargo retain deter-
minants for binding of Fu and Ci (Monnier et al., 2002;Asn686 (GST-SmoC-RRN) and a lysate of COS1 cellsexpressing Cos2. We found that the GST-SmoC-RRN C.Z. and P.A.B., unpublished data), but display dramati-
cally different responses to cotransfection with Smo.protein but not GST alone precipitated Cos2 (Figure6E), suggesting that these proteins, each expressed in Transfection of wild-type Cos2 thus yields increased
reporter activity in the presence of a sufficient relativeheterologous systems, bind each other directly. Consis-tent with this possibility, we found that Smo association level of activated Smo, whereas �cargo is unable to
respond (Figure 6F; green tracings).with Cos2 in S2 cells is not perturbed by RNAi targetingof Fu (data not shown), a tightly associated partner ofCos2 (see above). The GST-SmoC-RRN protein was also Hh-Induced Smo Accumulation Is Required
for Strong Responseable to interact in vitro with cargo-Fc and stalk-Fc pro-teins but not Fc protein alone, consistent with our obser- As Cos2 response depends critically on the relative level
of Smo (Figures 5B and 6F), we examined the role ofvations in S2 cells (see Figure 6D). Furthermore, since

Hedgehog Signaling via Cos2 Association with Smo1269
Smo accumulation in response to Hh stimulation. We responses to Hh signaling that do not involve transcrip-tional regulation via Ci/Gli. For example, a recently de-confirmed that cycloheximide (CHX) inhibition of protein
synthesis (Figure 7A) prevented HhN-induced accumu- scribed chemoattractant activity for Shh in axon guid-ance appears to be mediated by Smo, yet proceeds inlation of Smo (Figure 7B, compare lanes 4 and 2; Denef
et al., 2000). Despite a failure to accumulate in response a short time scale and with a local cell polarity thatsuggests a possible nonnuclear mechanism of responseto HhN stimulation, however, the Smo protein in these
cells is nevertheless phosphorylated (Figure 7B, lane 4, (Charron et al., 2003).In searching for other mediators of information trans-Smo), suggesting that these cells retain responsiveness
to Hh stimulation and that Smo may be activated. Nota- fer from membrane to cytoplasm, we were surprised tofind cytoplasmic components copurifying with Smo. Asbly, under conditions of inhibited protein synthesis, a
significantly smaller fraction of Cos2, Fu, and Su(fu) was these were the only Drosophila proteins identified in thebands excised, we conclude that these complexes werephosphorylated, suggesting that a full response to Hh
stimulation requires protein synthesis, and further sug- highly pure, and that Smo associates stably with compo-nents of the cytoplasmic complex in vivo. We furthergesting a role for Smo accumulation in this response.
We also considered the possibility that activation and demonstrate that Smo interacts directly with Cos2,which scaffolds this complex. Consistent with our find-phosphorylation of cytoplasmic components may de-
pend on an Hh-stimulated change in the efficiency of ings, articles presenting genetic evidence for a role ofthe Smo cytoplasmic tail in Hh signaling (Hooper, 2003)Smo interaction with Cos2. We tested this using the
�-Smo matrix to immunopurify Smo complexes from and evidence suggesting a physical interaction betweenSmo and Cos2 (Ruel et al., 2003; Ogden et al., 2003)cells in which CHX treatment was used to prevent Smo
accumulation and found under these conditions that were published during final review of this work. We havenot ruled out additional direct association of Smo withHhN stimulation did not produce an increase in levels
of copurified Cos2 (Figure 7C, compare lanes 1 and 2 other complex components.to 3 and 4). HhN stimulation thus does not appear to alterthe efficiency of Smo interaction with Cos2, although the A Model for Cos2 Complex Response to Hh Signalingoverall level of this interaction increases with Smo-P The identification of a complex that includes both Ciaccumulation and in concert with increased phosphory- and Smo immediately suggested that recruitment of thelation of cytoplasmic complex components. cytoplasmic complex to Smo upon Hh stimulation might
be critical for pathway activity. We demonstrate thatDiscussion Cos2 plays a central role in mediating this association,
functioning both as a scaffold that brings together cyto-Early studies of Cos2 suggested primarily a negative plasmic components (the Cos2 complex) and as a sen-role for Cos2 in pathway regulation, as manifested by sor that monitors the state of pathway activation byphenotypic analysis of cos2 mutations (Sisson et al., interacting with Smo at the membrane. The drawing in1997) and by a requirement for Cos2 in cytoplasmic Figure 7D highlights several important features of aretention of Ci and in its proteolytic processing to pro- model incorporating our observations and focusing onduce CiR (Chen et al., 1999; Wang and Holmgren, 1999; response of the Cos2 complex to Smo.Wang et al., 2000). More recent studies have suggested In the unstimulated state, Smo levels are low, anda potential positive role based on a requirement for Cos2 most of the Cos2 complex therefore is not associatedin transcriptional activation of gene targets associated with or influenced by Smo; even the small fraction ofwith highest levels of pathway activity (Wang et al., 2000; Cos2 complex associated with Smo may be negativeWang and Holmgren, 2000). We extend here the evi- (indicated by the red color) as Smo may not be in andence for such a positive role by demonstrating: (1) a active state. The negative form of the Cos2 complexrequirement for Cos2 in mediating a series of Hh- presumably mediates production of CiR and preventsinduced biochemical changes in pathway components nuclear accumulation of Ci, resulting in a net suppres-(summarized in Figure 1F); (2) an association between sion of transcriptional targets (red arrow). In the interme-the Cos2/Fu/Ci complex and Smo; (3) a direct interaction diate state (Figure 7D, middle drawing), present afterbetween Smo and Cos2; and (4) a requirement for Cos2 a few minutes of stimulation, Smo protein has beenin highest level Hh pathway response in cl-8 cell reporter activated by Hh stimulation (green color) but has notassays and Hh-induced morphogenesis in the dorsal yet accumulated. Therefore, despite a positive state forcuticle. the specific Cos2 complexes affected by Smo, the low
level of activated Smo protein is insufficient for interac-tion with most of the Cos2 complexes. The net outcomeDirect Association of Smo with the Cytoplasmic Complex
Based on the sequence relationship between Smo and with regard to transcriptional targets thus remains neg-ative.GPCRs, previous speculation and experimental work
focused on the possibility that Smo may interact with In stimulated cells, activated Smo exerts a pervasiveinfluence on the Cos2 complex (green color), eitherheterotrimeric G proteins (Alcedo et al., 1996; van den
Heuvel and Ingham, 1996). We have systematically tar- through a stable association (as shown) or through atransient association with enduring effects. This associ-geted G protein components using RNAi in the cl-8 cell
signaling assay, and failed to find a significant role for ation presumably involves a direct binding interactionbetween a portion of the Smo cytotail and the Cos2G proteins in transcriptional regulation via Ci (see Sup-
plemental Figure S6). We cannot, however, rule out a cargo and stalk domains. The evidence suggests thatboth activation and accumulation of Smo are critical, aspotential role for G proteins or other mediators in cellular

Molecular Cell1270
Figure 6. The Smo Cytotail Physically and Functionally Interacts with Cos2
(A) Alignment of Drosophila, Anopheles, and human Smo cytotail sequences. Conserved sequences extending from transmembrane domain7 (TM7) to Ser707 of Drosophila Smo are shown (a complete alignment is shown in Supplemental Figure S5). Blue letters denote the last threeamino acids of C-terminal truncation mutants and the circles indicate Ala substitutions. Ile586 (boxed) is important in Smo protein stabilityas Ala substitution results in loss of Smo expression and function (data not shown).(B) Effects of deleting Smo cytoplasmic tail sequences on Smo function. Endogenous Smo mRNA was targeted using dsRNA correspondingto the 5� untranslated region (UTR; schematic diagram). Expression constructs encoding Smo but lacking the 5� UTR sequence were usedto test for rescue of Smo function. Graph: ptc-luciferase reporter activity in cells transfected with Smo 5� UTR dsRNA with control YFP, wild-type (wt) Smo, or various Smo C-terminal deletion and point mutation expression constructs (see above). Dashed line: control level of signaling.Luciferase activities were normalized to Renilla luciferase activities. A representative assay is shown.(C) Effects of Smo C-terminal deletions on binding to Cos2. Cos2-R was expressed with either YFP, wt Smo, or mutant Smo proteins in S2cells stimulated with HhN. Levels of Cos2-R binding to Smo were measured by Renilla luciferase activity associated with precipitated proteinsusing the �-Smo mAb matrix (see diagram). Coprecipitations were performed in triplicate.(D) Cos2 stalk and cargo domains interact with Smo. Myc epitope-tagged domains of Cos2 (left diagram) coexpressed with Smo-R in S2 cellswere isolated by IP of protein complexes (in triplicate) using �-Myc mAb (9E10) and associated Renilla luciferase activity was measured. Smo-R

Hedgehog Signaling via Cos2 Association with Smo1271
Figure 7. Accumulation of Smo Is Critical for Strong Hh Signaling
(A) Cycloheximide inhibits new protein synthesis in cl-8 cells. Protein synthesis was monitored by incubation of cl-8 cells in S35-Met- and S35-Cys-containing medium for 2 hr. Medium contained cycloheximide (CHX; 50 �g/ml) or vehicle throughout the labeling period and for a 1.5 hrpretreatment period. Resulting lysates were run on SDS-PAGE and dried gel exposed to film.(B) New protein synthesis is required for strong Hh response in cl-8 cells. Cells pretreated as in (A) were incubated with control or HhNmedium for 2 hr in the continuing presence of CHX or vehicle. Pathway component response was determined by Western blotting. Note thatphosphorylation of cytoplasmic components in the presence of CHX occurs but is dramatically reduced.(C) New protein synthesis is required for an increase in Hh-induced Smo-Cos2 association. Levels of Cos2 that co-IP with Smo were measuredby Western blotting of lysates prepared from control or CHX- or HhN-treated cl-8 cells.(D) Model for response of cytoplasmic complex to Smo at three different stages of Hh stimulation. Pathway activity is also partially activatedin the absence of Cos2 (see Discussion).
moderate Smo overexpression alone is unable to fully targets (green arrow) resulting from pathway stimulationpresumably result from the positive action of Cos2 onactivate the pathway in the absence of Hh stimulation
(Chen et al., 1999; Alcedo et al., 2000; Ingham et al., Fu and from loss of the ability to produce CiR.It is the dual action of Cos2 in promoting formation2000). Similarly, a cycloheximide block of Smo accumu-
lation dramatically limits the biochemical changes nor- of CiR and in stabilization and possible activation of Futhat leads to the apparent dual negative and positivemally induced by Hh stimulation, and our cotransfection
experiments demonstrate that transition of Cos2 from roles of Cos2 in pathway regulation. Pathway activationinduced by loss of Cos2 (right panel) thus results froma pathway suppressor to activator requires adequate
levels of activated Smo. The activation of transcriptional loss of CiR, but this activation is only partial because
were coprecipitated with full-length Cos2, as well as stalk and cargo domain proteins. Deletion mutants were expressed at comparable levels(data not shown).(E) Smo cytotail interacts directly with Cos2 in vitro. Glutathione agarose beads alone, preloaded with GST, or with GST-SmoC-RRN fusionprotein (Smo cytotail from Ser557 to Asn686), were incubated with lysates from COS1 cells expressing Cos2, HA-Ci, Cos2 cargo-Fc, Cos2stalk-Fc, or Fc protein alone. Purified GST proteins were visualized by Coommassie staining. Cos2 and Ci were detected using an �-Cos2polyclonal or �-HA epitope monoclonal (for HA-Ci). Fc proteins were detected using an �-huIgG Fc polyclonal antibody. Input: 3% (Cos2 andHA-Ci) or 10% (Fc proteins) of material used for IP.(F) Cos2 cargo domain is required for Smo response. The effects of transfecting different levels of wt Cos2 or �cargo construct with andwithout 10 ng Smo construct on pathway activity were measured using the ptc-luciferase reporter assay in cl-8 cells. Dashed line: controllevel of signaling.

Molecular Cell1272
overexpression, an Ac5C-Gal4/UAS driver was used to express full-Fu is also destabilized, and the pathway-suppressinglength Hh protein that is processed and presumably modified inaction of Su(fu) is unchecked. Consistent with this inter-transfected cell. Cos2 and HA-Ci constructs used for expression inpretation, a combined loss of Cos2 and Su(fu) resultsCOS1 cells were generated in the pRK5 vector. Fc fusion proteins
in maximal pathway activation irrespective of the pres- were generated as described (Supplemental Experimental Proce-ence of Hh (Figure 5A; Wang et al., 2000; Lefers et al., dures). GST-SmoC-RRN construct was generated by ligating a PCR
product corresponding to AA557-686 into the pGEX 4T-1 vector.2001).
Cell CultureHow Does Cos2 Function?Cl-8, S2, and S2R� cells were cultured as described (see Supple-Motor proteins recently have been found to play a rolemental Experimental Procedures). Stimulation of cells with HhN-
in regulation of transmembrane receptors, as in the case conditioned medium (HhN medium) was described before (Porter etof rhodopsin and mannose-6-phosphate receptor (Tai al., 1995) with different incubation times noted for each experiment.et al., 1999; Nakagawa et al., 2000). These motor proteins Cells lysis was performed as described (see Supplemental Experi-
mental Procedures).apparently regulate receptor localization but do not playa direct role in receptor function. A recent report sug-
Antibodiesgests that Cos2 overexpression indeed may influenceSmo, Cos2, and Fu mAbs were generated against fusion proteinsSmo localization (Zhu et al., 2003). Our evidence, how-consisting of either the entire coding region of Fu, the N-term extra-
ever, suggests that Cos2 also functions as a primary cellular domain of Smo (1–257), or the motor and neck domain (aasensor of the state of pathway activation by interacting 1–640) of Cos2, fused in-frame with the Fc region of human IgG.with Smo at the membrane and by scaffolding and stabi- �-Su(fu) mAbs and rabbit polyclonal serum were generated using the
entire protein as antigen. �-Cos2 rabbit polyclonal was generatedlizing downstream pathway components.against a purified fusion protein consisting of GST and aa 509–791Kinesins likely share an evolutionary origin with G pro-of Cos2. �-Smo cytotail rat IgG was provided by S. Cohen, �-Ptcteins and myosins (Vale and Milligan, 2000), and all three47H8 mouse mAb was provided by R.L. Johnson, �-Ci 2A1 rat mAb
types of proteins use a conserved mechanism to couple was provided by R. Holmgren. Commercial sources providednucleotide hydrolysis to dramatic conformational changes ���-tubulin mAb E7 (Developmental Studies Hybridoma Bank; IA;in protein structure. Kinesins utilize this mechanism to developed by M. Klymkowsky), �-mKHC rabbit IgG (Cytoskeleton;
CO), �-HA (Zymed; CA), and �-Myc 9E10 mouse mAb and �-Mycgenerate force that allows movement on microtubules.(A14) rabbit IgG (both from Santa Cruz Biotechnology; CA).Sequences essential for microtubule binding, for nucle-
otide binding, and for motor function in other kinesins,Reporter Assays and RNAihowever, are not conserved among Cos2 sequences inFor ptc reporter assays, cl-8 cells were transfected with ptc-lucifer-
the Diptera and are not required for Cos2 function in ase and copia-Renilla luciferase reporter constructs in 24-well platecl-8 cells (see Supplemental Figure S4). It is still possible, format as described (Chen et al., 1999; Lum et al., 2003) along with
either overexpression constructs or dsRNA. Unless indicated, allhowever, that Cos2 retains the capacity for a conforma-experiments were performed in triplicate and luciferase activitiestional shift that may be triggered by Smo activation (Tai-normalize with Renilla luciferase activities. dsRNA was generatedpale et al., 2000). If so, then this conformational changeas described before (Lum et al., 2003; see Supplemental Table S3.may involve the Cos2 cargo domain, which binds toFor RNAi in S2 cells, cells were bathed in 15 �g/ml of dsRNA in
Smo at the cytotail and is likely required for Smo respon- serum free medium for 1 hr followed by addition of 10% FBS andsiveness. transfection with 0.5 �g of dsRNA using Effectene (Qiagen; CA).
Cells were incubated for an additional 2 days to allow protein turn-Whatever its mechanism of activation, our evidenceover prior to treatment with control or HhN medium for an additionalpoints to a role for the atypical kinesin Cos2 as a scaffold24 hr.and sensor that functions as the pivotal component in
transduction of pathway activation from the seven trans-Preparation of Affinity Matrix, Preparative Affinity
membrane receptor Smo to the latent cytoplasmic Chromatography, and Mass Spectrometrytranscription factor Ci. In addition to stabilizing Fu and Preparation of affinity matrix is described in Supplemental Experi-mediating forward signaling events that affect other cy- mental Procedures. For preparative scale affinity chromatography,
lysate from approximately forty 10 cm2 dishes of cl-8 cells (aftertoplasmic components and Ci, Cos2 is also required fortreatment with HhN or control medium overnight) were applied di-the accumulation of activated Smo, a critical aspectrectly (without preclearing) to 100 �l bed volume of matrix andof producing a full response to Hh signal. Cos2 thusincubated batch-wise overnight with rotation. Following extensive
functions not just as a passive sensor for the state of washing with lysis buffer, bound protein was eluted with 0.1 Mpathway activation at the membrane, but is also an ac- glycine (pH 3.0) and neutralized immediately with 1 M Tris (pH 7.4).tive participant in the cellular dynamics of transition from Eluted protein was acetone precipitated at �20C overnight, col-
lected by centrifugation, and resuspended in sample loading bufferthe unstimulated to the stimulated state. These activitieswith DTT (10 mM final) before SDS-PAGE separation and silver-are all the more remarkable in view of the well-recog-staining. Bands of interest were excised, destained, and subjectednized role played by Cos2 in maintaining the unstim-to trypsin digestion and peptide extraction as described (Gharah-
ulated state of the Hh pathway, and together these daghi et al., 1999). Peptides were identified by microcapillary LC-findings suggest that Cos2 dynamics are a critical deter- MS/MS using Thermo Finnigan LCQ Deca ion trap mass spectrome-minant of intracellular Hh pathway response and regu- ter and TurboSEQUEST analyses (CA).lation.
GST Fusion Protein Pull-Down AssaysGST fusion proteins were first purified from COS1 cell or bacteriaExperimental Procedureslysates, respectively, using glutathione agarose by batch incubationat 4C for 4 hr followed by 3 10 min washes in lysis buffer. Protein-Constructs
Expression constructs (Cos2, Cos2-R, Cos2 deletion mutants, Smo, loaded beads were then incubated with lysate from COS1 cellsexpressing Cos2, Ci, cargo-Fc, stalk-Fc, or Fc protein alone, for anSmo-R, and Smo truncation mutants) used in Drosophila cell culture
are described in Supplemental Experimental Procedures. For Hh additional 2 hr followed by 3 10 min washes in lysis buffer. Bound

Hedgehog Signaling via Cos2 Association with Smo1273
protein was analyzed by Western blotting (for Cos2, Ci, and Fc pro- Chen, J.K., Taipale, J., Cooper, M.K., and Beachy, P.A. (2002a).Inhibition of Hedgehog signaling by direct binding of cyclopamineteins).to Smoothened. Genes Dev. 16, 2743–2748.
Smo-R and Cos2-R Coimmunoprecipitation Assays Chen, J.K., Taipale, J., Young, K.E., Maiti, T., and Beachy, P.A.Smo-R was cotransfected with Myc epitope-tagged Cos2 proteins (2002b). Small molecule modulation of Smoothened activity. Proc.or Cos2-R cotransfected with Smo truncation mutants into S2 cells. Natl. Acad. Sci. USA 99, 14071–14076.Transfected cells were lysed as before, and Myc epitope-tagged Clemens, J.C., Worby, C.A., Simonson-Leff, N., Muda, M., Maehama,Cos2 proteins or Smo truncation mutants were immunoprecipitated T., Hemmings, B.A., and Dixon, J.E. (2000). Use of double-strandedwith �-Myc mAb (9E10) matrix or �-Smo mAb, respectively. Associ- RNA interference in Drosophila cell lines to dissect signal transduc-ated Renilla luciferase activity was measured following 3 10 min tion pathways. Proc. Natl. Acad. Sci. USA 97, 6499–6503.washes in lysis buffer.
Denef, N., Neubuser, D., Perez, L., and Cohen, S.M. (2000). Hedge-hog induces opposite changes in turnover and subcellular localiza-Fly Stockstion of patched and smoothened. Cell 102, 521–531.P[w�; FRT]G13 cos2W1/CyO and P[w�; FRT]G13 P[hsp 70-MYC] wereForbes, A.J., Nakano, Y., Taylor, A.M., and Ingham, P.W. (1993).generously provided by M. Scott. yw P[ry�; FLP]12; CyO/Sco fliesGenetic analysis of hedgehog signaling in the Drosophila embryo.were acquired from the Bloomington Drosophila Stock Center (IN).Development (Suppl), 115–124.
Generation of Somatic Clones and Immunostaining of Wing Discs Frank-Kamenetsky, M., Zhang, X.M., Bottega, S., Guicherit, O.,For somatic mutant clone analysis, clones of cos2W1 mutant cells Wichterle, H., Dudek, H., Bumcrot, D., Wang, F.Y., Jones, S., Shulok,were generated by FLP-mediated mitotic recombination (Xu and J., et al. (2002). Small-molecule modulators of Hedgehog signaling:Rubin, 1993) in larvae resulting from crosses as described before identification and characterization of Smoothened agonists and an-(Sisson et al., 1997). Clones were induced by heat-shocking the tagonists. J. Biol. 1, 10.larvae at 37C for 1.5 hr on day 2. Late third instar larvae were heat- Gharahdaghi, F., Weinberg, C.R., Meagher, D.A., Imai, B.S., andshocked again to induce the expression of marker gene before Mische, S.M. (1999). Mass spectrometric identification of proteinswing imaginal discs were dissected, fixed, and stained using the from silver-stained polyacrylamide gel: a method for the removal ofappropriate mAb. Samples were mounted in with DAPI and exam- silver ions to enhance sensitivity. Electrophoresis 20, 601–605.ined by confocal microscopy. Secondary antibodies were pur-
Hooper, J.E. (2003). Smoothened translates Hedgehog levels intochased from Jackson ImmunoResearch Labs (PA).distinct responses. Development 130, 3951–3963.
Ingham, P.W., and McMahon, A.P. (2001). Hedgehog signaling inAcknowledgmentsanimal development: paradigms and principles. Genes Dev. 15,3059–3087.We would like to thank B. Hamaoka for production of recombinant
Su(fu) protein, Y. Ma for the Hh expression construct, R.L. Johnson, Ingham, P.W., Nystedt, S., Nakano, Y., Brown, W., Stark, D., vanR. Holmgren, and S. Cohen for antibodies, J. Sisson and M.P. Scott den Heuvel, M., and Taylor, A.M. (2000). Patched represses thefor fly strains, K. Basler for useful discussion, and members of the Hedgehog signalling pathway by promoting modification of thehybridoma core facility at Memorial Sloan-Kettering for help in gen- Smoothened protein. Curr. Biol. 10, 1315–1318.erating mAbs. Supported by a fellowship from the Life Sciences Lefers, M.A., Wang, Q.T., and Holmgren, R.A. (2001). Genetic dissec-Research Foundation (L.L.) and a grant from NIH. P.A.B. is an investi- tion of the Drosophila Cubitus interruptus signaling complex. Dev.gator of the Howard Hughes Medical Institute. Biol. 236, 411–420.
Lum, L., Yao, S., Mozer, B., Rovescalli, A., Von Kessler, D., Nirenberg,Received: June 5, 2003
M., and Beachy, P.A. (2003). Identification of Hedgehog pathwayRevised: October 14, 2003
components by RNAi in Drosophila cultured cells. Science 299,Accepted: October 15, 2003
2039–2045.Published: October 27, 2003
Methot, N., and Basler, K. (1999). Hedgehog controls limb develop-ment by regulating the activities of distinct transcriptional activatorReferencesand repressor forms of Cubitus interruptus. Cell 96, 819–831.
Monnier, V., Ho, K.S., Sanial, M., Scott, M.P., and Plessis, A. (2002).Alcedo, J., Zou, Y., and Noll, M. (2000). Posttranscriptional regulationHedgehog signal transduction proteins: contacts of the Fused ki-of smoothened is part of a self- correcting mechanism in the Hedge-nase and Ci transcription factor with the Kinesin-related proteinhog signaling system. Mol. Cell 6, 457–465.Costal2. BMC Dev. Biol. 2, 4.Alcedo, J., Ayzenzon, M., Von Ohlen, T., Noll, M., and Hooper, J.E.Muenke, M., and Beachy, P.A. (2001). Holoprosencephaly. In The(1996). The Drosophila smoothened gene encodes a seven-passMetabolic and Molecular Bases of Inherited Disease, C.R. Scriver,membrane protein, a putative receptor for the hedgehog signal. CellA.L. Beaudet, W.S. Sly, D. Valle, B. Childs, K.W. Kinzler, and B.86, 221–232.Vogelstein, eds. (New York: McGraw-Hill), pp. 6203–6230.Aza-Blanc, P., Ramirez-Weber, F.A., Laget, M.P., Schwartz, C., andNakagawa, T., Setou, M., Seog, D., Ogasawara, K., Dohmae, N.,Kornberg, T.B. (1997). Proteolysis that is inhibited by hedgehogTakio, K., and Hirokawa, N. (2000). A novel motor, KIF13A, transportstargets Cubitus interruptus protein to the nucleus and converts itmannose-6-phosphate receptor to plasma membrane through di-to a repressor. Cell 89, 1043–1053.rect interaction with AP-1 complex. Cell 103, 569–581.Bourne, H.R. (1997). How receptors talk to trimeric G proteins. Curr.Ogden, S.K., Ascano, M., Jr., Stegman, M.A., Suber, L.M., Hooper,Opin. Cell Biol. 9, 134–142.J.E., and Robbins, D.J. (2003). Identification of a functional interac-Chamoun, Z., Mann, R.K., Nellen, D., von Kessler, D.P., Bellotto, M.,tion between the transmembrane protein Smoothened and theBeachy, P.A., and Basler, K. (2001). Skinny hedgehog, an acyltransf-kinesin-related protein Costal2. Curr. Biol. 13, 1998–2003.erase required for palmitoylation and activity of the hedgehog signal.Ohlmeyer, J.T., and Kalderon, D. (1998). Hedgehog stimulates matu-Science 293, 2080–2084.ration of Cubitus interruptus into a labile transcriptional activator.Charron, F., Stein, E., Jeong, J., McMahon, A., and Tessier-Lavigne,Nature 396, 749–753.M. (2003). The morphogen Sonic hedgehog is an axonal chemo-Pham, A., Therond, P., Alves, G., Tournier, F.B., Busson, D., Lamour-attractant that collaborates with Netrin-1 in midline axon guidance.Isnard, C., Bouchon, B.L., Preat, T., and Tricoire, H. (1995). TheCell 113, 11–23.Suppressor of fused gene encodes a novel PEST protein involved inChen, C.H., von Kessler, D.P., Park, W., Wang, B., Ma, Y., andDrosophila segment polarity establishment. Genetics 140, 587–598.Beachy, P.A. (1999). Nuclear trafficking of Cubitus interruptus in the
transcriptional regulation of Hedgehog target gene expression. Cell Porter, J.A., von Kessler, D.P., Ekker, S.C., Young, K.E., Lee, J.J.,Moses, K., and Beachy, P.A. (1995). The product of hedgehog auto-98, 305–316.

Molecular Cell1274
proteolytic cleavage active in local and long-range signalling. Nature374, 363–366.
Preat, T. (1992). Characterization of Suppressor of fused, a completesuppressor of the fused segment polarity gene of Drosophila mela-nogaster. Genetics 132, 725–736.
Preat, T., Therond, P., Lamour-Isnard, C., Limbourg-Bouchon, B.,Tricoire, H., Erk, I., Mariol, M.C., and Busson, D. (1990). A putativeserine/threonine protein kinase encoded by the segment-polarityfused gene of Drosophila. Nature 347, 87–89.
Robbins, D.J., Nybakken, K.E., Kobayashi, R., Sisson, J.C., Bishop,J.M., and Therond, P.P. (1997). Hedgehog elicits signal transductionby means of a large complex containing the kinesin-related proteincostal2. Cell 90, 225–234.
Ruel, L., Rodriguez, R., Gallet, A., Lavenant-Staccini, L., and The-rond, P.P. (2003). Stability and association of Smoothened, Costal2and Fused with Cubitus interruptus are regulated by Hedgehog.Nat. Cell Biol. 5, 907–913.
Sisson, J.C., Ho, K.S., Suyama, K., and Scott, M.P. (1997). Costal2,a novel kinesin-related protein in the Hedgehog signaling pathway.Cell 90, 235–245.
Stegman, M.A., Vallance, J.E., Elangovan, G., Sosinski, J., Cheng,Y., and Robbins, D.J. (2000). Identification of a tetrameric hedgehogsignaling complex. J. Biol. Chem. 275, 21809–21812.
Tai, A.W., Chuang, J.Z., Bode, C., Wolfrum, U., and Sung, C.H. (1999).Rhodopsin’s carboxy-terminal cytoplasmic tail acts as a membranereceptor for cytoplasmic dynein by binding to the dynein light chainTctex-1. Cell 97, 877–887.
Taipale, J., and Beachy, P.A. (2001). The Hedgehog and Wnt signal-ling pathways in cancer. Nature 411, 349–354.
Taipale, J., Chen, J.K., Cooper, M.K., Wang, B., Mann, R.K., Milen-kovic, L., Scott, M.P., and Beachy, P.A. (2000). Effects of oncogenicmutations in Smoothened and Patched can be reversed by cyclo-pamine. Nature 406, 1005–1009.
Taipale, J., Cooper, M.K., Maiti, T., and Beachy, P.A. (2002). Patchedacts catalytically to suppress the activity of Smoothened. Nature418, 892–896.
Therond, P.P., Knight, J.D., Kornberg, T.B., and Bishop, J.M. (1996).Phosphorylation of the fused protein kinase in response to signalingfrom hedgehog. Proc. Natl. Acad. Sci. USA 93, 4224–4228.
Vale, R.D., and Milligan, R.A. (2000). The way things move: lookingunder the hood of molecular motor proteins. Science 288, 88–95.
van den Heuvel, M., and Ingham, P.W. (1996). smoothened encodesa receptor-like serpentine protein required for hedgehog signalling.Nature 382, 547–551.
Wang, G., Amanai, K., Wang, B., and Jiang, J. (2000). Interactionswith Costal2 and suppressor of fused regulate nuclear translocationand activity of cubitus interruptus. Genes Dev. 14, 2893–2905.
Wang, Q.T., and Holmgren, R.A. (1999). The subcellular localizationand activity of Drosophila cubitus interruptus are regulated at multi-ple levels. Development 126, 5097–5106.
Wang, Q.T., and Holmgren, R.A. (2000). Nuclear import of cubitusinterruptus is regulated by hedgehog via a mechanism distinct fromCi stabilization and Ci activation. Development 127, 3131–3139.
Xu, T., and Rubin, G.M. (1993). Analysis of genetic mosaics in devel-oping and adult Drosophila tissues. Development 117, 1223–1237.
Zhu, A.J., Zheng, L., Suyama, K., and Scott, M.P. (2003). Alteredlocalization of Drosophila Smoothened protein activates Hedgehogsignal transduction. Genes Dev. 17, 1240–1252.




















