
Melanopsin Contributions to Irradiance Coding in the Thalamo-Cortical Visual System
Upload
independentCategory
view
0download
0
Harmful Algae 17 (2012) 25–34
Growth and toxicity responses of Mediterranean Ostreopsis cf. ovata to seasonalirradiance and temperature conditions
Eleonora Scalco a, Christophe Brunet a, Francesca Marino a, Rachele Rossi b, Vittorio Soprano b,Adriana Zingone a, Marina Montresor a,*a Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italyb Istituto Zooprofilattico Sperimentale del Mezzogiorno, Via Salute 2, 80055 Portici, Italy
A R T I C L E I N F O
Article history:
Received 24 June 2011
Received in revised form 10 February 2012
Accepted 11 February 2012
Available online 10 March 2012
Keywords:
Growth rates
HABs
Mediterranean Sea
Ostreopsis cf. ovata
Palytoxins
Photophysiology
A B S T R A C T
The genus Ostreopsis includes several species capable of producing various palytoxin-like compounds
which have harmful effects on humans and marine fauna. Species in this genus are regular members of
the epiphytic community in tropical seas but their geographic range have shown an apparent expansion
towards temperate regions in recent years. This is the case of the Mediterranean Sea, where Ostreopsis cf.
ovata is responsible for intense blooms in summer and autumn. With the aim of describing the growth
performances of O. cf. ovata, we carried out a multifactorial experiment in which strains isolated from 3
locations along the Italian coast were grown at different combinations of temperature, daylength and
irradiance encompassing different seasonal conditions.
Temperature constrained growth of O. cf. ovata within a window of 18–30 8C, while best
performances were recorded at 22 and 26 8C. Growth was maximum at 12 h daylength, whereas it
was limited by photon flux density at short daylength (9 h) and often showed photosaturation at the
longest daylength (15 h). Cells with anomalous shape were found during the exponential phase at the
lowest and highest temperatures, while cells with a reduced size were produced at all conditions except
18 8C. Total toxin concentration varied between the different environmental conditions and between
exponential and post-exponential growth phases without a clear pattern, whereas toxin composition
was less variable. Our results suggest that O. cf. ovata is adapted to intermediate temperatures and
daylength conditions such as those recorded in the natural environment at the beginning of summer
and/or at the beginning of autumn, when this dinoflagellate builds up its biomass along the coast of the
Mediterranean Sea. Photosaturation instead occurs at the highest temperature and irradiance
conditions, thus supporting the observations of generally lower cell abundances in late July–August.
� 2012 Elsevier B.V. All rights reserved.
Contents lists available at SciVerse ScienceDirect
Harmful Algae
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/ha l
1. Introduction
The benthic dinoflagellate genus Ostreopsis was described bySchmidt (1901) from the Gulf of Siam (Thailand), with Ostreopsis
siamensis Schmidt as the type species. Eight more species weredescribed in the genus from 1981 to 1999 (Faust, 1999). Ostreopsis
species live in close contact with macroalgae and/or submergedrocks or sediments (Faust and Gulledge, 2002), to which theyconnect through threads (Barone and Prisinzano, 2006) which havebeen shown to be trichocysts (Honsell et al., 2011). Cells are oftenembedded in an abundant mucilaginous matrix (Heil et al., 1993),and both free cells and mucous can also be found in the watercolumn. Six Ostreopsis species have been reported as responsible
* Corresponding author. Tel.: +39 081 5833259; fax: +39 081 7641355.
E-mail address: [email protected] (M. Montresor).
1568-9883/$ – see front matter � 2012 Elsevier B.V. All rights reserved.
doi:10.1016/j.hal.2012.02.008
for toxic events (Rhodes, 2010) and 3 of them have been shown toproduce potent toxins analogues of palytoxin (PlTX), which arecomplex, high molecular weight, and water soluble polyalcohols.The production of ostreocin-D was detected in O. siamensis (Usamiet al., 1995; Ukena et al., 2002), while mascarenotoxin-a and -b(McTX-a/-b) were found in Ostreopsis mascarenensis (Lenoir et al.,2004). After the discovery of ovatoxin-a (OvTX-a) in Mediterraneanstrains of Ostreopsis cf. ovata (Ciminiello et al., 2008), four moreOvTX types (Ciminiello et al., 2010a; Rossi et al., 2010), and twoMcTXs (Rossi et al., 2010) were detected in this species. Ostreopsis
blooms have been associated with severe skin irritations (Deedsand Schwartz, 2010) and respiratory problems in humans throughaerosolization of toxins (Ciminiello et al., 2008). In addition, thesetoxins could be transmitted through the food-web to humans thateat bottom-feeding fishes or shellfish (Aligizaki et al., 2008),causing vomiting, difficulty in breathing, kidney problems and, insome cases, death (Deeds and Schwartz, 2010).

E. Scalco et al. / Harmful Algae 17 (2012) 25–3426
Ostreopsis species were initially reported mostly from tropicaland subtropical seas, where they are associated with otherepiphytic dinoflagellates such as Gambierdiscus, Coolia and benthicProrocentrum species (e.g. Besada et al., 1982). In recent years, therange of Ostreopsis species has shown an apparent expansion totemperate regions, where they can form intense blooms (Rhodes,2010). This is the case of New Zealand waters (Shears and Ross,2009), the Japan Sea (Selina and Orlova, 2010), and theMediterranean Sea (Vila et al., 2001; Aligizaki and Nikolaidis,2006; Monti et al., 2007; Mangialajo et al., 2008; Totti et al., 2010).In the Mediterranean Sea, the first Ostreopsis bloom was detectedin 1972, when a species reported as O. siamensis (Taylor, 1979), theonly species known in the genus at that time, occurred at highconcentrations within a mucilaginous matrix covering macroalgaein Villefranche-sur-Mer Bay (Ligurian Sea; Max Taylor, pers. com.).O. ovata was first described as epiphyte of brown and redmacroalgae growing on coral reefs of Ryukyu Islands, FrenchPolynesia and New Caledonia (Fukuyo, 1981). In the Mediterra-nean Sea, it was recorded in Italian (Tognetto et al., 1995) waterssince the 80 s. Sansoni et al. (2003) described for the first time aclear impact on the benthic fauna during a bloom of this species insummers 1998, 2000 and 2001 in coastal north Tyrrhenian waters,where impacts on humans through inhalation were also reported(Simoni et al., 2003). The first massive case of intoxication byinhalation was recorded in summer 2005 along the Genoa coast(Ligurian Sea) (Ciminiello et al., 2006).
Molecular investigations have revealed that Mediterranean andAtlantic populations of O. ovata cluster into the same phylogeneticgroup, while Pacific isolates are genetically distinct (Penna et al.,2010). Molecular information is lacking on the type material of O.
ovata, consequently, the taxonomic status of this species remainsuncertain. It is therefore advisable to use the designation O. cf.ovata when referring to isolates from the Mediterranean Sea. Inaddition to this genotype, some recently isolated strains showother different molecular signatures (Aligizaki et al., 2010), whileanother species, O. cf. siamensis, is reported from coastal waters ofSpain and Sicily (Penna et al., 2010).
Along the Mediterranean coasts, O. cf. ovata has been recordedfrom June through October and at times in November. Blooms (upto 106 cells g�1 fresh weight of macroalgae) occur in early summerin the north-western Mediterranean basin, with a second minorincrease in biomass in late summer–autumn (Mangialajo et al.,2011); in the Adriatic and Aegean Sea blooms are instead onlyrecorded in late summer-beginning of autumn (Aligizaki andNikolaidis, 2006; Totti et al., 2010). Proliferations in the order of105 cells g�1 fresh weight of macroalgae occur over a wide range ofwater temperatures, from about 19 to 28 8C, without any clearcorrelation between cell abundance and local temperature values(Mangialajo et al., 2011). O. cf. ovata grows both in association withmacroalgae and on pebbles and stones on the sea bottom and ittherefore experiences a broad range of irradiance conditions,depending on the depth at which the macroalgae grow as well ason the position of the dinoflagellate on the macroalga. The effect ofirradiance and daylength on the growth capacity of thisdinoflagellate has still not been explored in depth, although thepresence of O. cf. ovata in temperate waters at different times of thesummer–autumn period has raised interest on its ecophysiologicalrequirements and on its potential toxicity under differentenvironmental conditions. Subtle differences in growth perfor-mances have been demonstrated between two strains from theTyrrhenian and the Adriatic Sea, which also differed in cell size andat times produced cells with anomalous morphology (Guerriniet al., 2010). Other studies showed that strains from Tyrrhenianwaters have the highest growth rates at 30 8C (Graneli et al., 2010).
We carried out a multifactorial experiment in which 3 strainsisolated from distant sites of the Italian coasts were grown under
different combinations of temperature; daylength and irradiance,with the aim of describing the growth responses of O. cf. ovata inrelation with the seasonal variations. Growth rate and the relativeabundance of cells with normal shape and size, small-sized cells,and cells with anomalous morphology along the growth curveswere also investigated. To address potential risks related tovariations in toxin production under different environmentalconditions, the quantity and quality of toxin was measured for oneof the strains in the exponential and post exponential phase overthree combinations of temperature and daylength values.
2. Materials and methods
2.1. Strain maintenance
Three strains of O. cf. ovata were used for the experiments:strain D483 isolated from the Gulf of Naples (Tyrrhenian Sea,408470N, 148110E) in September 2008, strain CBA-T isolated fromPortonovo (Adriatic Sea, 438340N; 138350E) in September 2008 andstrain OS2T isolated from the Gulf of Trieste (Adriatic Sea, 458390N138450E) in September 2006. The ITS region of the strains wassequenced by Antonella Penna (University of Urbino, Italy) and theanalysis of the sequences confirmed that strains belong to theMediterranean ribotype of O. ovata (Penna et al., 2005, 2010). Stockcultures were maintained at 22 8C, 100 mmol photons m�2 s�1, anda 12L:12D h photo-cycle, in K/2 culture medium without theaddition of silicates (Keller et al., 1987) prepared with oligotrophicseawater adjusted to 36. The growth medium was diluted 50:50(v:v) with filtered (through 0.22 mm) and autoclaved seawater.
2.2. Experimental design
A multi-factorial experiment was designed to test the growthresponse of O. cf. ovata over a range of temperatures (14 8C, 18 8C,22 8C, 26 8C, 30 8C and 34 8C), and daylength (9L:15D, 12L:12D and15L:9D h photo-cycle) conditions, which included those recordedin the natural environment over the seasonal cycle. For eachcombination of temperature and daylength, growth performancewas investigated at two photon flux density (PFD) conditions, 50and 200 mmol photons m�2 s�1. These conditions resulted in adaily PFDs ranging between 1.62 and 10.8 mol photons m�2 d�1.
All combinations of temperature and daylength were testedbetween 14 and 30 8C, with the exception of the short daylengthcondition at 30 8C and the long daylength condition at 14 8C,resulting in a total number of 26 experiments. The experimentsplanned at 34 8C were not performed because no strain survivedthose conditions (see below). Strains were gradually acclimated ateach combination of temperature and daylength values. To thisend, strains were grown in glass Erlenmeyer flasks under a PFD of125 mmol photons m�2 s�1 and kept in the exponential growthphase by diluting the cultures with fresh medium. Starting fromthe maintenance conditions of the stock cultures (22 8C, 12L:12D),subsequent acclimation steps were carried out by decreasing orincreasing temperature by 4 8C while keeping the photo-cycleconstant. For each temperature step, conditions were keptconstant at least 7 days before acclimating the strains at adifferent photocycle conditions for at least 7 more days. Strainacclimation to temperature higher than 30 8C was carried out intwo steps, growing them for 10 days at 32 8C and subsequentlytransferring them at 34 8C.
The enumeration of O. cf. ovata cells in culture presentstechnical problems due to production of mucilaginous materialwhich results in agglomerates and causes cell adhesion to theculture vessel walls. When the experiments are performed in batchor semi-continuous cultures, the vigorous mixing needed todisgregate these agglomerates before subsampling for cell

Table 1Relative percentage variation of specific growth rates of Ostreopsis cf. ovata between
12L and 9L daylength conditions (Eq. (1)) at the different temperature and PFD
combinations. Significant differences (p < 0.001) based on Student’s t-test between
growth rates at the two daylength conditions are reported with an asterisk.
18 8C 22 8C 26 8C
50 mmol photons m�2 s�1 0.64 (*) 0.41 (*) 0.61 (*)
200 mmol photons m�2 s�1 0.51 (*) 0.29 (*) 0.61 (*)
Table 2Relative percentage variation of specific growth of Ostreopsis cf. ovata strains
between 12L and 15L daylength conditions (Eq. (2)) at the different temperature
and PFD combinations. Significant differences (p < 0.001) based on Student’s t-test
between growth rates at the two daylength conditions are reported with an asterisk
(n.s.: non significant).
18 8C 22 8C 26 8C 30 8C
50 mmol photons m�2 s�1 0.06 (n.s.) �0.24 (*) 0.09 (n.s.) �0.14 (n.s.)
200 mmol photons m�2 s�1 0.28 (*) �0.07 (n.s.) 0.43 (*) 0.27 (*)
Table 3Relative percentage variation of specific growth of Ostreopsis cf. ovata strains
between the two PFD conditions (50 and 200 mmol photons m�2 s�1) at the different
temperature and daylength combinations (Eq. (3)). Significant differences
(p < 0.001) based on Student’s t-test between growth rates at the two PFD
conditions are reported with an asterisk (n.s.: non significant; n.a.: data not
available).
18 8C 22 8C 26 8C 30 8C
9L 0.25 (*) 0.44 (*) 0.33 (*) n.a.
12L �0.02 (n.s.) 0.32 (*) 0.33 (*) 0.36 (*)
15L �0.35 (*) 0.21 (*) �0.08 (n.s.) 0.002 (n.s.)
E. Scalco et al. / Harmful Algae 17 (2012) 25–34 27
counting may not be effective and can affect cell growth. Toobtain more precise counts without interfering with growth,multiple 6-well culture plates (Corning Incorporated, Euroclones.p.a.) were used in this study, which allowed treating the wholecontent of the wells with a mucous-dissolving substance at eachsampling point. In practice, for each experiment a 200 mLworking stock to be used as inoculum was prepared by dilutingthe exponentially growing cultures of the individual strains withK/2 medium down to a concentration of 200 cells mL�1. Aftercareful mixing, 5 mL of the working stock of each strain weredeployed in 2 wells of 10 culture plates, each containing, at theend, duplicate cultures of the three strains. To limit evaporation,the space among wells was filled with culture medium and theplates were sealed with parafilm. The plates were then incubatedat the selected experimental conditions over a time intervalbetween 15 and 22 days, encompassing the entire period ofexponential growth and part or all of the stationary phase. Forcell enumeration, every 2–3 days, Na-EDTA at a final 0.01 Mconcentration was added into the wells of one culture plate todissolve mucous aggregates, followed by fixation with formal-dehyde at a final concentration of 0.4%. After careful mixing, 1 mLof the fixed sample was placed in a Sedgewick Rafter slide andcells were enumerated at a Zeiss Axiophot light microscope.Separate counts were obtained for normal cells, small cells(dorso-ventral size <40 mm), and cells with anomalous mor-phology. Maximum specific growth d�1 values were obtained byestimating the linear regression of ln transformed cell concen-trations over the exponential portion of the growth curve(Guillard, 1980).
2.3. Statistical analyses
Two-way ANOVA and multiple regression analyses wereperformed on the dataset (N = 132) of maximum specific growthrates obtained from experiments conducted between 18 and 30 8C– i.e. the conditions at which growth was measurable – to test theeffect of PFD, daylength and temperature on growth performance.The relative percentage variation of specific growth ratesmeasured at the different temperatures and PFD combinationswhen decreasing (from 12L to 9L) or increasing (from 12L to 15L)daylength was calculated using the following equations, where GRis the average (N = 6) specific growth rate measured at eachcondition:
GR9�12 ¼ ðGR12 h � GR9 hÞ � ðGR12 hÞ�1 (1)
GR12�15 ¼ ðGR12 h � GR15 hÞ � ðGR12 hÞ�1 (2)
The relative percentage variation of specific growth ratesmeasured at the different temperature and daylength combina-tions when decreasing PFD from 200 to 50 mmol photons m�2 s�1
was calculated using the following equations, where GR is theaverage (N = 6) specific growth measured at each condition.
GRPFD ¼ ðGR200 � GR50Þ � ðGR200Þ�1 (3)
For each comparison, the difference between pairs of datasetswas estimated with Student’s t-test (Tables 1–3).
To assess the role of different experimental factors in theformation of anomalous and small cells, a non parametricSpearman correlation analysis was carried out on the percentageof cell types recorded during the exponential growth phase inrelation with temperature, PFD, daylength conditions, and therespective specific growth rates. The statistical analyses werecarried out using Systat 7.
2.4. Toxin analysis
The quantity and quality of toxins produced by O. cf. ovata
strain D483 were estimated at different experimental conditionsthat simulated those typical for spring (temperature 18 8C,photocycle 12L:12D), early summer (temperature 22 8C, photo-cycle 15L:9D), and late summer (temperature 26 8C, photocycle15L:9D) in Italian seas. Experiments were carried out at a PFD of50 mmol photons m�2 s�1. Cultures were acclimated at the differ-ent experimental conditions for 10 days. For each experimentalcondition, six replicate Erlenmeyer flasks containing 1 L of K/2culture medium were inoculated with a culture in the exponentialgrowth phase to reach an initial cell concentration of2000 cells mL�1. Cell concentration in the flasks was monitoredevery 4–5 days. The flasks were carefully mixed and a subsample of3 mL was treated with Na-EDTA, fixed with formaldehyde andenumerated in a Sedgewick Rafter slide.
For toxin extraction, three flasks in exponential growth phaseand three flasks in post-exponential phase were harvested bycentrifugation. PlTXs were extracted as described in Rossi et al.(2010). The PlTX standard was purchased from Wako ChemicalsGmbH (Neuss, Germany). Sample solutions of standard material(50, 20, 10 ppm) were prepared in MeOH:water (1:1, v:v). Massspectral analyses were performed using an Agilent LC/TOF/MS, atime-of-flight mass spectrometer equipped with an ElectroSprayinterface coupled with an Agilent liquid chromatograph model1100. The LC equipment included a binary pump and an autosampler. Separation of palytoxin-like molecules was performedinjecting the samples and the standard material (5 mL sampleinjection volume) in a Phenomenex Luna HILIC 3 m150 mm � 2 mm. The flow rate was 0.3 mL min�1 and gradientelution was performed with two eluents, water (eluent A) andacetonitrile:water 95:5 (v:v) (eluent B), both containing 0.1%formic acid. Initial conditions were: 2 min column equilibration
E. Scalco et al. / Harmful Algae 17 (2012) 25–3428
with 20% eluent B, followed by a linear gradient to 50% eluent B in20 min, a linear gradient to 100% eluent B in 10 min and isocraticelutions for 6 min with 100% eluent B. The LC/TOF/MS analysisworked in positive ion mode, with mass range set at m/z 100–3500 u at a resolving power of 10,000. The conditions of ESI sourcewere as follows: drying gas (N2) flow rate 11 mL min�1, drying gastemperature 300 8C, nebulizer 45 psig, capillary voltage 4000 V,fragmentor 250 V; skimmer voltage 60 V. All acquisitions and dataanalyses were controlled by Agilent LC/TOF/MS Software (Agilent,USA–Germany). Tuning mix (G1969-85003) was used for lockmass calibration in our assay. The new ovatoxins OvTX-b, OvTX-c,and OvTX-d reported in Rossi et al. (2010) have been renamed hereas OvTX-e, OvTX-b, and OvTX-c, to match the nomenclatureproposed by Ciminiello et al. (2010b).
3. Results
3.1. Growth at different temperature and irradiance conditions
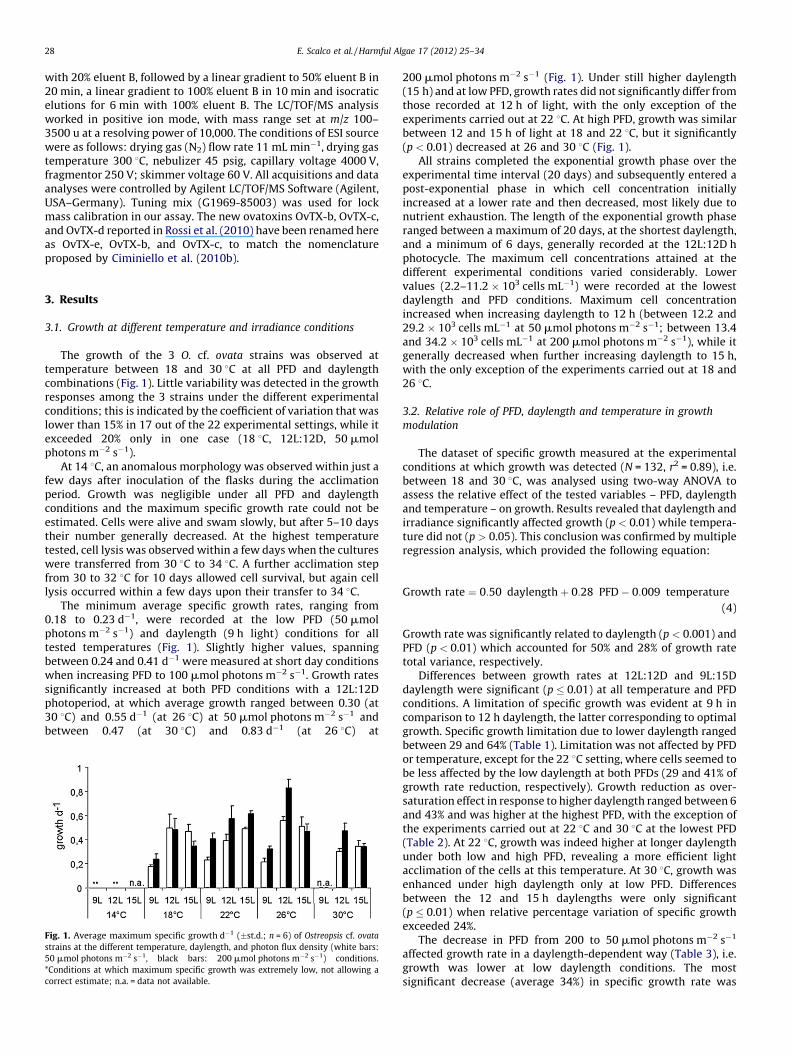
The growth of the 3 O. cf. ovata strains was observed attemperature between 18 and 30 8C at all PFD and daylengthcombinations (Fig. 1). Little variability was detected in the growthresponses among the 3 strains under the different experimentalconditions; this is indicated by the coefficient of variation that waslower than 15% in 17 out of the 22 experimental settings, while itexceeded 20% only in one case (18 8C, 12L:12D, 50 mmolphotons m�2 s�1).
At 14 8C, an anomalous morphology was observed within just afew days after inoculation of the flasks during the acclimationperiod. Growth was negligible under all PFD and daylengthconditions and the maximum specific growth rate could not beestimated. Cells were alive and swam slowly, but after 5–10 daystheir number generally decreased. At the highest temperaturetested, cell lysis was observed within a few days when the cultureswere transferred from 30 8C to 34 8C. A further acclimation stepfrom 30 to 32 8C for 10 days allowed cell survival, but again celllysis occurred within a few days upon their transfer to 34 8C.
The minimum average specific growth rates, ranging from0.18 to 0.23 d�1, were recorded at the low PFD (50 mmolphotons m�2 s�1) and daylength (9 h light) conditions for alltested temperatures (Fig. 1). Slightly higher values, spanningbetween 0.24 and 0.41 d�1 were measured at short day conditionswhen increasing PFD to 100 mmol photons m�2 s�1. Growth ratessignificantly increased at both PFD conditions with a 12L:12Dphotoperiod, at which average growth ranged between 0.30 (at30 8C) and 0.55 d�1 (at 26 8C) at 50 mmol photons m�2 s�1 andbetween 0.47 (at 30 8C) and 0.83 d�1 (at 26 8C) at
Fig. 1. Average maximum specific growth d�1 (�st.d.; n = 6) of Ostreopsis cf. ovata
strains at the different temperature, daylength, and photon flux density (white bars:
50 mmol photons m�2 s�1, black bars: 200 mmol photons m�2 s�1) conditions.
*Conditions at which maximum specific growth was extremely low, not allowing a
correct estimate; n.a. = data not available.
200 mmol photons m�2 s�1 (Fig. 1). Under still higher daylength(15 h) and at low PFD, growth rates did not significantly differ fromthose recorded at 12 h of light, with the only exception of theexperiments carried out at 22 8C. At high PFD, growth was similarbetween 12 and 15 h of light at 18 and 22 8C, but it significantly(p < 0.01) decreased at 26 and 30 8C (Fig. 1).
All strains completed the exponential growth phase over theexperimental time interval (20 days) and subsequently entered apost-exponential phase in which cell concentration initiallyincreased at a lower rate and then decreased, most likely due tonutrient exhaustion. The length of the exponential growth phaseranged between a maximum of 20 days, at the shortest daylength,and a minimum of 6 days, generally recorded at the 12L:12D hphotocycle. The maximum cell concentrations attained at thedifferent experimental conditions varied considerably. Lowervalues (2.2–11.2 � 103 cells mL�1) were recorded at the lowestdaylength and PFD conditions. Maximum cell concentrationincreased when increasing daylength to 12 h (between 12.2 and29.2 � 103 cells mL�1 at 50 mmol photons m�2 s�1; between 13.4and 34.2 � 103 cells mL�1 at 200 mmol photons m�2 s�1), while itgenerally decreased when further increasing daylength to 15 h,with the only exception of the experiments carried out at 18 and26 8C.
3.2. Relative role of PFD, daylength and temperature in growth
modulation
The dataset of specific growth measured at the experimentalconditions at which growth was detected (N = 132, r2 = 0.89), i.e.between 18 and 30 8C, was analysed using two-way ANOVA toassess the relative effect of the tested variables – PFD, daylengthand temperature – on growth. Results revealed that daylength andirradiance significantly affected growth (p < 0.01) while tempera-ture did not (p > 0.05). This conclusion was confirmed by multipleregression analysis, which provided the following equation:
Growth rate ¼ 0:50 daylength þ 0:28 PFD � 0:009 temperature
(4)
Growth rate was significantly related to daylength (p < 0.001) andPFD (p < 0.01) which accounted for 50% and 28% of growth ratetotal variance, respectively.
Differences between growth rates at 12L:12D and 9L:15Ddaylength were significant (p � 0.01) at all temperature and PFDconditions. A limitation of specific growth was evident at 9 h incomparison to 12 h daylength, the latter corresponding to optimalgrowth. Specific growth limitation due to lower daylength rangedbetween 29 and 64% (Table 1). Limitation was not affected by PFDor temperature, except for the 22 8C setting, where cells seemed tobe less affected by the low daylength at both PFDs (29 and 41% ofgrowth rate reduction, respectively). Growth reduction as over-saturation effect in response to higher daylength ranged between 6and 43% and was higher at the highest PFD, with the exception ofthe experiments carried out at 22 8C and 30 8C at the lowest PFD(Table 2). At 22 8C, growth was indeed higher at longer daylengthunder both low and high PFD, revealing a more efficient lightacclimation of the cells at this temperature. At 30 8C, growth wasenhanced under high daylength only at low PFD. Differencesbetween the 12 and 15 h daylengths were only significant(p � 0.01) when relative percentage variation of specific growthexceeded 24%.
The decrease in PFD from 200 to 50 mmol photons m�2 s�1
affected growth rate in a daylength-dependent way (Table 3), i.e.growth was lower at low daylength conditions. The mostsignificant decrease (average 34%) in specific growth rate was
E. Scalco et al. / Harmful Algae 17 (2012) 25–34 29
recorded at the lowest daylength. Increasing daylength to 12 hunder 50 mmol photons m�2 s�1 reduced growth limitation (aver-age 25%); nevertheless the difference between the two PFDconditions was still significant (p < 0.01) with the exception of the18 8C condition, where growth rate at 200 mmol photons m�2 s�1
was similar to that recorded at 50 mmol photons m�2 s�1 (Table 3).At the longest daylength conditions, higher PFD decreased specificgrowth rates at low (18 8C) and high (26 8C) temperature,suggesting an inhibition of photosynthesis due to PFD oversatura-tion. At 30 8C, cells seemed to cope with the high PFD, growth ratebeing identical to that at low PFD. As for the daylength effect (seeabove), the 22 8C temperature condition at 15L:9D photocycleshowed a peculiar pattern, since growth rate increased under highPFD revealing a very efficient regulation strategy of the cells at thistemperature. The growth rate comparison test was significant(p � 0.01) in all cases when the relative percentage variation ofspecific growth exceeded 21%.
3.3. Morphological changes
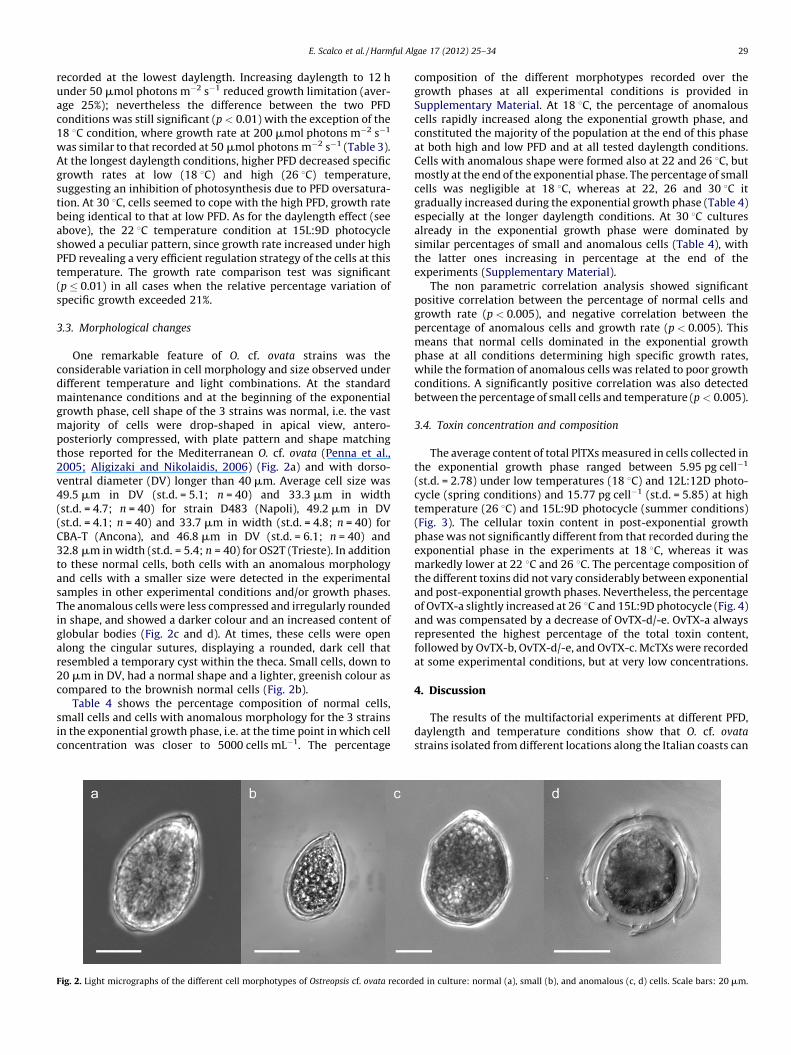
One remarkable feature of O. cf. ovata strains was theconsiderable variation in cell morphology and size observed underdifferent temperature and light combinations. At the standardmaintenance conditions and at the beginning of the exponentialgrowth phase, cell shape of the 3 strains was normal, i.e. the vastmajority of cells were drop-shaped in apical view, antero-posteriorly compressed, with plate pattern and shape matchingthose reported for the Mediterranean O. cf. ovata (Penna et al.,2005; Aligizaki and Nikolaidis, 2006) (Fig. 2a) and with dorso-ventral diameter (DV) longer than 40 mm. Average cell size was49.5 mm in DV (st.d. = 5.1; n = 40) and 33.3 mm in width(st.d. = 4.7; n = 40) for strain D483 (Napoli), 49.2 mm in DV(st.d. = 4.1; n = 40) and 33.7 mm in width (st.d. = 4.8; n = 40) forCBA-T (Ancona), and 46.8 mm in DV (st.d. = 6.1; n = 40) and32.8 mm in width (st.d. = 5.4; n = 40) for OS2T (Trieste). In additionto these normal cells, both cells with an anomalous morphologyand cells with a smaller size were detected in the experimentalsamples in other experimental conditions and/or growth phases.The anomalous cells were less compressed and irregularly roundedin shape, and showed a darker colour and an increased content ofglobular bodies (Fig. 2c and d). At times, these cells were openalong the cingular sutures, displaying a rounded, dark cell thatresembled a temporary cyst within the theca. Small cells, down to20 mm in DV, had a normal shape and a lighter, greenish colour ascompared to the brownish normal cells (Fig. 2b).
Table 4 shows the percentage composition of normal cells,small cells and cells with anomalous morphology for the 3 strainsin the exponential growth phase, i.e. at the time point in which cellconcentration was closer to 5000 cells mL�1. The percentage
Fig. 2. Light micrographs of the different cell morphotypes of Ostreopsis cf. ovata record
composition of the different morphotypes recorded over thegrowth phases at all experimental conditions is provided inSupplementary Material. At 18 8C, the percentage of anomalouscells rapidly increased along the exponential growth phase, andconstituted the majority of the population at the end of this phaseat both high and low PFD and at all tested daylength conditions.Cells with anomalous shape were formed also at 22 and 26 8C, butmostly at the end of the exponential phase. The percentage of smallcells was negligible at 18 8C, whereas at 22, 26 and 30 8C itgradually increased during the exponential growth phase (Table 4)especially at the longer daylength conditions. At 30 8C culturesalready in the exponential growth phase were dominated bysimilar percentages of small and anomalous cells (Table 4), withthe latter ones increasing in percentage at the end of theexperiments (Supplementary Material).
The non parametric correlation analysis showed significantpositive correlation between the percentage of normal cells andgrowth rate (p < 0.005), and negative correlation between thepercentage of anomalous cells and growth rate (p < 0.005). Thismeans that normal cells dominated in the exponential growthphase at all conditions determining high specific growth rates,while the formation of anomalous cells was related to poor growthconditions. A significantly positive correlation was also detectedbetween the percentage of small cells and temperature (p < 0.005).
3.4. Toxin concentration and composition
The average content of total PlTXs measured in cells collected inthe exponential growth phase ranged between 5.95 pg cell�1
(st.d. = 2.78) under low temperatures (18 8C) and 12L:12D photo-cycle (spring conditions) and 15.77 pg cell�1 (st.d. = 5.85) at hightemperature (26 8C) and 15L:9D photocycle (summer conditions)(Fig. 3). The cellular toxin content in post-exponential growthphase was not significantly different from that recorded during theexponential phase in the experiments at 18 8C, whereas it wasmarkedly lower at 22 8C and 26 8C. The percentage composition ofthe different toxins did not vary considerably between exponentialand post-exponential growth phases. Nevertheless, the percentageof OvTX-a slightly increased at 26 8C and 15L:9D photocycle (Fig. 4)and was compensated by a decrease of OvTX-d/-e. OvTX-a alwaysrepresented the highest percentage of the total toxin content,followed by OvTX-b, OvTX-d/-e, and OvTX-c. McTXs were recordedat some experimental conditions, but at very low concentrations.
4. Discussion
The results of the multifactorial experiments at different PFD,daylength and temperature conditions show that O. cf. ovata
strains isolated from different locations along the Italian coasts can
ed in culture: normal (a), small (b), and anomalous (c, d) cells. Scale bars: 20 mm.

Table 4Percentage of normal (N), anomalous (A) and small (S) cells of Ostreopsis cf. ovata recorded for the 3 strains (D483, CBA-T, OS2T) during the exponential growth phase, i.e. at
the day in which cell concentrations was closer to 5000 cells mL�1. Light grey shaded cells mark the cases in which the maximum cell concentration was <5000 cells mL�1.
Bold values indicate the maximum percentage value recorded at each experimental condition.
Temperature (8C) Daylength (h light) D483 CBA-T OS2T
%N %A %S %N %A %S %N %A %S
Low PFD (50 mmol photons m�2 s�1)
18 9 14 86 0 15 80 6 44 55 1
12 72 28 0 57 33 10 72 26 2
15 35 65 0 29 69 2 48 40 12
22 9 53 42 5 60 24 16 26 24 5012 63 24 13 40 19 41 25 14 6115 76 18 6 57 17 27 68 17 15
26 9 22 54 24 36 36 28 20 26 5412 53 32 16 38 14 48 28 9 6315 73 24 3 33 32 34 44 24 32
30 9 – – – – – – – – –
12 25 37 37 8 60 32 35 22 4315 20 54 26 6 61 33 8 45 48
High PFD 200 mmol photons m�2 s�1
18 9 27 73 0 3 95 2 2 90 8
12 65 35 0 44 55 1 27 73 0
15 15 85 0 8 88 5 13 82 6
22 9 79 19 2 56 25 19 56 14 29
12 75 16 9 51 31 18 49 14 36
15 76 23 1 57 29 14 67 25 8
26 9 42 31 28 25 41 34 33 23 4312 76 21 3 62 12 25 48 7 45
15 57 41 3 34 39 27 55 20 25
30 9 – – – – – – – – –
12 15 40 44 5 28 66 10 16 7415 11 65 23 2 64 33 2 45 53
E. Scalco et al. / Harmful Algae 17 (2012) 25–3430
survive within a quite wide range of temperature values (>14 8Cand �32 8C). However, the little growth and the immediate shift tocells with anomalous morphology observed at 18 and 30 8C suggestthat O. cf. ovata is well adapted to a more restricted window, withonly temporary adaptation to higher and lower temperatures.While temperature represents the primary factor for defining theseasonal interval that permits growth, it does not directly affectgrowth response at the different daylength and PFD conditions, butrather modulates it in a rather unpredictable way, thus confirmingthe complex and non linear relationships between these majorenvironmental forcing factors and microalgal growth (Geider,1987; Foy and Gibson, 1993). By contrast, daylength plays a majorrole in driving growth performances of this dinoflagellate. Growthis light limited at the lowest daylength (9 h), while photosaturationgenerally occurs when O. cf. ovata is exposed at longer daylength(15 h), especially at higher PFD.
Darker cells with anomalous morphology were produced inconsiderable amounts at the end of the experiments, i.e. whennutrients were presumably depleted, under all conditions and forall strains. By contrast, in previous studies on Tyrrhenian and
Fig. 3. Average concentration (pg cell�1 � st.d.; n = 3) of palytoxin-like toxins
produced by Ostreopsis cf. ovata strain D483 at three temperature and daylength
combinations, in exponential (white bars) and post-exponential (black bars) growth
phase.
Adriatic strains of O. cf. ovata (Guerrini et al., 2010), a shift in cellshape from normal to irregularly spherical during the progressionfrom the exponential to the stationary growth phase was onlynoticed for Tyrrhenian strains. In our study, cells with anomalousshape were also found during the exponential phase at the lowestand highest temperatures. In these cases, anomalous cells wereformed starting in the first few days of the experiments, whennutrients were certainly available to cell growth. The formation of
Fig. 4. Average (n = 3) percentage composition of the different toxins produced
by Ostreopsis cf. ovata strain D483 at three temperature and daylength
combinations, in exponential and post-exponential growth phase. OvTX-
a = ovatoxin-a, OvTX-b = ovatoxin-b, OvTX-c = ovatoxin-c, OvTX-d/e = ovatoxin-d/e,
McTX-a = mascarenotoxin-a, McTX-c = mascarenotoxin-c.

E. Scalco et al. / Harmful Algae 17 (2012) 25–34 31
cell with an aberrant shape has been reported in O. cf. ovata strainsisolated from Brazilian waters grown in full L1 medium; cells re-gained their normal shape when transferred into diluted L1/2medium (Nascimento et al., 2012). We have also used a dilutedmedium (K/2) in which the concentration of the major nutrients iscomparable to that in L1/2 medium; nevertheless, cells with anaberrant shape were observed towards the end of the experimentsand at the highest and lowest temperature. In general, it seems thatthe production of anomalous cells is a response to unfavourableconditions determined by either limiting or excess nutrients, or bytoo high or too low temperatures. At the end of the experiments,most anomalous cells were motile and dividing couplets wereobserved at times. However, we cannot exclude that division ratesof anomalous cells were lower than those of normal cells, thuscontributing to the establishment of the post-exponential phaseand the final decline of the cultures. Darker cells with anomalousmorphology and globular content might be also interpreted asprecursors of pellicle cysts, because – especially towards the end ofthe growth phase – stages with a granular, darker content wereobserved inside broken thecae. Pellicle cysts are found indinoflagellate cultures exposed to stressful conditions such aschanges in temperature, salinity, or in the late post-exponentialgrowth phase, or as a reaction to the presence of grazers, parasitesor allelochemical compounds produced by other phytoplanktonspecies (reviewed in Bravo et al., 2010a). The formation of cyst-likestages has been reported in cultures of O. cf. ovata (Bravo et al.,2010b) and hyaline cysts, which can be interpreted as pelliclecysts, have also been reported in nature at the end of the bloomperiod of this species (Aligizaki and Nikolaidis, 2006). Furtherstudies should address the vitality, division rates, and fate of thesecells with anomalous morphology in order to assess their effectiverole in natural and experimental conditions.
The formation of smaller cells with DV smaller than 40 mm wasrecorded already during the exponential growth phase at allexperimental conditions, with the exception of 18 8C. The presenceof small cells in the life cycle is known for several dinoflagellates,for which different mechanisms of formation (i.e. depauperatingcell division and budding) have been described from culturematerial (Silva and Faust, 1995). Different hypotheses have beenput forward with regard to their function: they have beeninterpreted as gametes, i.e. cells capable of conjugating andproducing the diploid stage (planozygote), or as a means toincrease population size under non-optimal nutrient concentra-tions. Two cell-size classes have been reported in field samples forboth O. cf. ovata (DV < 45 mm) and O. cf. siamensis (DV < 47 mm)(Aligizaki and Nikolaidis, 2006), while cells with an average DV of34 mm have at times been recorded in natural samples fromBrazilian waters (Nascimento et al., 2012). More information isneeded on cell size distribution during the different phases of thebloom in natural populations, as well as on their actual biologicalsignificance, in order to shed light on the role of these small sized-cells in the life strategy of Ostreopsis species.
4.1. Temperature and irradiance as factors regulating growth
Temperature and irradiance (PFD and daylength) are majordrivers of the seasonal cycle in temperate coastal regions. Theireffects on phytoplankton growth are often considered indepen-dently, while their interaction has seldom been taken into account.Even less attention has been placed on the role of the photocycle inmodulating growth responses of microalgae (but see Sunda andHuntsman, 2004; Rost et al., 2006). Laboratory investigations andmodeling simulations have shown that the interactions betweentemperature, daylength and growth are complex and non-linear(Hagiwara et al., 2002; Sunda and Huntsman, 2004). The resultsobtained in the present study confirm this scenario and highlight
the interactive effect of these 3 variables in conditioning growthrate and photophysiological adjustments in O. cf. ovata. Daylengthand PFD resulted as the major drivers of variability for the growthresponses recorded in our experiments for this dinoflagellate.Growth was limited by PFD at short but not at intermediatedaylength, where it showed the highest values, while over-saturation was often detected at the longest daylength. Photon fluxdensity influences this general pattern: when grown at high PFD,specific growth rates were in fact slightly higher, but their increasewas not proportional to the increased daily irradiance even at thelowest daylength, further supporting the evidence that this speciesperforms better, in terms of growth, at relatively low PFD. This isreasonable when considering that light availability amidstmacroalgal vegetation is generally low (Raniello et al., 2004),but it is in apparent contrast with the proliferation of the speciesalso on stones and pebbles at relatively low depth (Totti et al.,2010), although the latter cases are only reported for the latesummer–early autumn period, when daylength and PFD arereduced as compared to early summer.
Our results also suggest that O. cf. ovata has differentacclimation strategies over an environmental gradient rangingbetween low and high temperature and irradiance conditions. Atintermediate temperature values (22–26 8C), cells are able toregulate efficiently their photosynthetic machinery and maximizespecific growth under the two tested PFD conditions and/or underthe three photo-cycle regimes. This means that, considering theseasonal environmental variations at our latitudes, O. cf. ovata
populations seem to be better adapted to early summer andautumn conditions, when cells can efficiently acclimate to changesin light availability. At the intermediate temperature values ofthese seasons, energy can be invested in both photoregulation andgrowth, while at lower (winter–spring) and higher (mid summer)temperatures, photoregulation capacity is apparently limited andgrowth rate decreases when daylength deviates from the 12 h:12 hphotocycle or when PFD increases. As revealed by the highestspecific growth, the 22–26 8C temperature appears to be optimalfor the cells to transfer energy into photoregulation. Indeed, PFDdoes not negatively affect growth as much, under these conditions.It is noteworthy that this temperature range matches the one of theperiod when O. cf. ovata builds up its biomass in the naturalenvironment, at the beginning of summer and/or at the beginningof autumn.
4.2. Differences among strains from different locations
Differences in growth performance were detected amongst O.
cf. ovata strains from different geographic locations along theItalian coast, but they were generally not significant and did notshow any consistent pattern that could indicate preferences ofindividual strains for specific temperature or irradiance conditions.Nevertheless, results from these strains cannot be considered asconclusive with regard to the characterization of populations ofdifferent provenance, which should rather be assessed through theanalysis of physiological responses of a large number of freshlyestablished strains and the consideration of both intra- and inter-population variability (Brand, 1989). Guerrini et al. (2010) reporteddifferences in cell size between 2 O. cf. ovata strains isolated fromthe Adriatic and Tyrrhenian seas, respectively, the former strainbeing larger (average DV 57.2 mm) and the latter smaller (averageDV 40.2 mm). In fact, the strains that we used, when measured atthe beginning of the exponential growth phase, did not showconsiderable differences, the average DV being 49.5 mm in theTyrrhenian strain from Napoli and 46.8 mm for the North-Adriaticstrain from Trieste. In addition, we also observed dramatic changesin cell size and morphology along the exponential phase of growth,which indicates considerable variations in cell size for this species

Ta
ble
5C
om
pa
riso
nb
etw
ee
nto
xin
con
ten
ta
nd
pe
rce
nta
ge
com
po
siti
on
of
Ost
reo
psi
scf
.ov
ata
stra
ins
rep
ort
ed
inG
ue
rrin
ie
ta
l.(2
01
0),
Cim
inie
llo
et
al.
(20
10
b),
Ro
ssi
et
al.
(20
10
)a
nd
Na
scim
en
toe
ta
l.(2
01
2).
Ex
pe
rim
en
tal
con
dit
ion
s:
Gu
err
ini
et
al.
(20
10
)2
08C
,1
6L:
8D
,9
0m
mo
lph
oto
ns
m�
2s�
1,
en
de
xp
on
en
tia
la
nd
sta
tio
na
ry(t
he
sela
tte
rv
alu
es
are
giv
en
inp
are
nth
esi
s)p
ha
ses;
Cim
inie
llo
et
al.
(20
10
a,b
):2
08C
,1
2L:
8D
,9
0m
mo
lph
oto
ns
m�
2s�
1,
sta
tio
na
ry
ph
ase
;R
oss
ie
ta
l.(2
01
0):
188C
,1
2L:
12
D,
50
mm
olp
ho
ton
sm�
2s�
1,
po
st-e
xp
on
en
tia
lp
ha
se;
Na
scim
en
toe
ta
l.(2
01
2):
248C
,1
2L:
12
D,
60
mm
olp
ho
ton
sm�
2s�
1,
ex
po
ne
nti
al
an
dst
ati
on
ary
(th
ese
latt
er
va
lue
sa
reg
ive
nin
pa
ren
the
sis)
ph
ase
s.N
D=
no
td
ete
cte
d.
To
xin
Ad
ria
tic
(pg
cell�
1)
#O
OA
N0
60
1
Gu
err
ini
et
al.
(20
10
)
Ty
rrh
en
ian
(pg
cell�
1)
#O
OT
L06
02
Gu
err
ini
et
al.
(20
10
)
Ad
ria
tic
(pg
cell�
1)
#O
OA
N0
60
1
Cim
inie
llo
et
al.
(20
10
b)
Ad
ria
tic
(%)
#O
OA
N0
60
1
Cim
inie
llo
et
al.
(20
10
b)
Ty
rrh
en
ian
(pg
cell�
1)
#D
48
3
Ro
ssi
et
al.
(20
10
)
Ty
rrh
en
ian
(%)
#D
48
3
Ro
ssi
et
al.
(20
10
)
Bra
zil
(pg
cell�
1)
#LC
A-E
7
Na
scim
en
to
et
al.
(20
12
)
Bra
zil
(%)
#LC
A-E
7
Na
scim
en
to
et
al.
(20
12
)
Bra
zil
(pg
cell�
1)
#LC
A-B
7
Na
scim
en
to
et
al.
(20
12
)
Bra
zil
(%)
#LC
A-B
7
Na
scim
en
to
et
al.
(20
12
)
Ty
rrh
en
ian
(pg
cell�
1)
#D
48
3
Th
isin
ve
stig
ati
on
Ty
rrh
en
ian
(%)
#D
48
3
Th
isin
ve
stig
ati
on
P-P
lTX
1.2
5(2
.5)
0.7
(1.1
)0
.20
.6N
DN
DN
D–
0.6
21
ND
ND
ND
Ov
TX
-a1
8(3
1)
14
(25
)1
85
43
.67
–9
.41
48
.4–
62
.82
0–
40
(71
)2
0–
33
(39
)(7
8–
17
1)
(36
–4
5)
2.1
–9
.81
45
.02
–6
7.9
0
Ov
TX
-b9
27
2.5
1–
4.1
21
9.5
–2
5.0
23
–5
0(7
7)
25
–5
1(4
2)
(87
–2
05
)(4
4–
50
)0
.7–
5.1
17
.53
–3
1.0
3
Ov
TX
-c2
60
.08
–0
.74
0.8
–5
.74
–3
0(1
0)
4–
18
(6)
(3–
37
)(2
–8
)0
.00
5–
1.2
0.1
5–
7.1
1
Ov
TX
-d/e
41
21
.69
–3
.43
14
.6–
25
.93
–8
0(2
3)
3–
40
(13
)(5
–5
5)
(3–
12
)0
.22
–6
.82
.06
–3
1.2
3
McT
X-a
ND
ND
ND
–0
.47
0.4
–2
.0N
DN
D0
.00
6–
0.4
70
.05
–3
.98
McT
X-c
ND
ND
ND
–0
.32
ND
–0
.5N
DN
DN
D–
0.3
2N
D–
4.0
5
E. Scalco et al. / Harmful Algae 17 (2012) 25–3432
under different growth conditions. Indeed, difference in cell sizebetween populations of different geographic origin should betterbe assessed through measurements taken on a large number ofstrains in the same growth phase and under comparableexperimental settings, as well as on natural populations.
Growth estimates for Mediterranean strains of O. cf. ovata wereobtained in previous investigations based on different protocolsand/or experimental conditions (Graneli et al., 2010; Guerrini et al.,2010). The published data are comparable to those recorded in thepresent study, and confirm our finding that O. cf. ovata is notcapable of active growth at temperatures below 18 8C (Graneliet al., 2010). A major difference between our results and those ofGraneli et al. (2010) concerns the growth performance of O. cf.ovata at the highest temperature. While the 3 strains that we testedall showed a decrease in specific growth at 30 8C (average 0.34 d�1,st.d. 0.02; 15 h daylength and 200 mmol photons m�2 s�1) ascompared to 26 8C (average 0.47 d�1, st.d. 0.06), an average specificgrowth of 0.74 d�1 was reported for a Tyrrhenian strain used byGraneli et al. (2010) at 30 8C, under similar light conditions (16 hdaylength and 140 mmol photons m�2 s�1). Either variabilitywithin Tyrrhenian populations or differences in the experimentalsettings and procedures could account for these divergent resultson the performance of O. cf. ovata at high temperature, whichcertainly requires further investigation due to its pivotal role inestablishing the warm-water affinity of the species and thepossibility of its expansion in view of climate change. Interestingly,strains of O. cf. ovata isolated from tropical waters alongthe Brazilian coasts and grown at 24 8C, 12L:12D h and60 mmol photons m�2 s�1 showed extremely low growth ratesof 0.10–0.15 div d�1, which correspond to specific growth of 0.07and 0.1, respectively (Nascimento et al., 2012).
4.3. Toxin content
A considerable variability for toxin content in Mediterranean O.
cf. ovata strains has been recorded in different studies. The first LC/MS quantitative estimates were obtained from natural samplescollected along the Ligurian coasts (Ciminiello et al., 2008), wheretoxin concentrations varied between 1.51 and 3.11 pg cell�1 forOvTX-a and between 0.09 and 0.40 pg cell�1 for putative-palytoxin(P-PlTX). Further analyses on Adriatic and Tyrrhenian Sea isolates(Guerrini et al., 2010) (Table 5) showed much higher totalconcentrations and highlighted higher cellular and dissolvedconcentration of OvTX-a and P-PlTX in the stationary phase ascompared to the exponential phase, with slightly higher valuesmeasured in the Adriatic as compared to the Tyrrhenian strain.
Numerous new PlTX-like compounds have been recentlycharacterized using liquid chromatography coupled to massspectrometry (Ciminiello et al., 2010b; Rossi et al., 2010). In thefirst study, the analyses were carried out on an Agilent 1100 LCbinary system coupled to a hybrid linear ion trap LTQ OrbitrapXLTM Fourier transform mass spectrometer (FTMS) equipped withan ESI ION MAXTM source. A slow gradient elution was used toobtain a chromatographic separation that allowed to identify fournew toxins, OvTX-b, -c, -d and -e, in addition to confirming thepresence of OvTX-a and P-PlTX. In the second study, the analyseswere carried out on an Agilent 1100 LC binary system coupled to anAgilent Time Of Flight (TOF) mass spectrometer. The gradient useddid not provide a chromatographic separation that was sufficientenough to identify OvTX-d and P-PlTX, but did allow theidentification of McTX-a and a new PlTX-like compound namedmascarenotoxin-c (McTX-c). The percentage composition of thetoxin profile matches relatively well in the two publications(Ciminiello et al., 2010b; Rossi et al., 2010), whereas the amount ofthe single toxins differs considerably (Table 5). This might well beattributed to differences amongst strains. Much higher toxin

E. Scalco et al. / Harmful Algae 17 (2012) 25–34 33
concentrations (up to 171 pg cell�1 of OvTX-a and 205 pg cell�1 ofOvTX-b in stationary growth phase) were recently recorded inO. cf. ovata strains – genetically identical to the Mediterraneanones – isolated along the Brazilian coasts (Nascimento et al., 2012)(Table 5).
Our results show that toxin concentration varied betweenexponential and post-exponential growth phase as well as amongdifferent temperature and daylength conditions, but without aclear pattern. We did not always detect higher cellular toxinconcentrations in post-exponential phase as reported by Guerriniet al. (2010) and Nascimento et al. (2012). In addition, at hightemperature and daylength conditions, toxin concentration in theexponential growth phase doubled that recorded in the stationaryphase. The variability in toxin concentration within a single strainmight be attributed to environmental conditions as well as todifferences in cell type composition. Indeed, considerable differ-ences in toxin content between vegetative cells and cysts havebeen reported for Alexandrium species (Oshima et al., 1992). Interms of toxin composition, we did not detect marked differencesbetween exponential and post-exponential phase, but the per-centage of OvTX-a increased and that of OvTX-d/-e decreased atthe high temperature and photoperiod conditions, correspondingto the summer peak conditions.
4.4. O. cf. ovata growth properties in the lab and blooms in the sea
The growth and ecophysiological properties of Mediterraneanstrains of O. cf. ovata depicted by the results of our experimentsindicate that this dinoflagellate is adapted to summer–autumnconditions, in terms of temperature and daylength, but also showthat cell growth slows down under high PFD for >12 h at very hightemperature. This latter evidence, together with the observationthat growth photosaturation already occurs at relatively low PFD(�200 mmol photons m�2 s�1), might explain the generally lowercell abundances recorded in many places in late July–August. Inthese months in fact, seawater temperature and PFD reach theirmaximum annual values (up to 30 8C, Mangialajo et al., 2011).Nevertheless, this dinoflagellate is capable of photoadaptation atthese extreme conditions, but its capability to invest in biomassbuild-up seems to be impaired during constantly clear and calmweather conditions, where the species might need to invest moreenergy in photo-protective defense. At lower temperatures andlower daily PFDs – as those occurring at shorter daylength and/orunder more variable weather conditions – energy can instead beallocated to growth. Indeed our experiments show that specificgrowth reached maximum values at intermediate temperatureand/or daylength values, under which cells seem to regulate veryefficiently their photosynthetic machinery. These conditionsindeed correspond to those recorded either at the beginning ofsummer and/or at the beginning of autumn, i.e. when O. cf. ovata
builds up its biomass along the coasts of the Mediterranean Sea(Mangialajo et al., 2011).
The timing and intensity of O. cf. ovata blooms depend ondifferent factors such as (i) size of the inoculum present at thebeginning of the growth season, (ii) intrinsic specific growth underdifferent environmental conditions, and (iii) loss factors such asgrazing, parasite attacks or natural cell death. Whether this speciesis capable of overwintering as a benthic resting stage or as rare cellsinterspersed in the perennial macroalgal thalli is still an openquestion. Our experiments show that it can barely survive at 14 8C,but we cannot rule out the possibility that cell survival in naturemay occur at these temperatures, which can be recorded in surfacecoastal waters during the winter–early spring season (Mangialajoet al., 2011).
Overall, specific growth rates measured in the laboratory for O.
cf. ovata are not extremely high, which implies that a relatively
long time might be required until biomass accumulates todetectable levels. If we assume an overwintering stock as low as10 cells (100 g ww)�1 of macroalgae, corresponding to the winterundetectability of the species, and an average specific growth of0.20 d�1, it would take about 2 months to reach a detectable levelof 5000 cells (g ww)�1. Specific growth rates increase as daylengthincreases and cell concentration would increase as well, but it willtake 20 days more to reach 1,500,000 cells (g ww)�1 with a growthrate of 0.4 d�1. These relatively long times for biomass build-upcould explain why the species is not found at detectableconcentrations in late spring, when environmental conditionsshould already be suitable for its growth, at least based on ourexperiments. In situ growth rates might be higher, and they shouldbe investigated along with loss factors in natural populations inorder to better explain and possibly predict the seasonal pattern ofabundance of O. cf. ovata along the Mediterranean coasts.
Acknowledgements
The authors thank Antonella Penna (University of Urbino, Italy)for the genetic characterization of the strains and for providingstrain CBA-T; Marina Monti (National Institute of Oceanographyand Applied Geophysics, Trieste, Italy) for providing strain OS2T;Carmen Minucci and Gandi Forlani (SZN) for help in culturemaintenance. E.S. was supported by the project ‘Ostreopsis ovata eOstreopsis spp.: nuovi rischi di tossicita microalgale nei mariitaliani’ funded by Ministero dell’Ambiente e della Tutela delTerritorio e del Mare; F.M. and R.R. were supported by the project‘Monitoraggio di Ostreopsis ovata lungo le coste della regioneCampania’ funded by Regione Campania; V.S. by the project IZS-ME03/07 ‘Una nuova tossina nelle acque della Campania e delMediterraneo, l’ovatossina: strategie analitiche e di protezionedella salute e della sicurezza alimentare’ funded by Ministero dellaSalute.[SS]
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.hal.2012.02.008.
References
Aligizaki, K., Battocchi, C., Penna, A., Hernandez, F.R., Arsenakis, M., Fraga, F., 2010.Diversity of potentially toxic benthic dinoflagellates in Southern Europe. In: 14thInternational Conference on Harmful Algae, Hersonissos-Crete (Greece), p. 25.
Aligizaki, K., Katikou, P., Nikolaidis, G., Panou, A., 2008. First episode of shellfishcontamination by palytoxin-like compounds from Ostreopsis species (AegeanSea, Greece). Toxicon 51 (3), 418–427.
Aligizaki, K., Nikolaidis, G., 2006. The presence of the potentially toxic generaOstreopsis and Coolia (Dinophyceae) in the North Aegean Sea, Greece. HarmfulAlgae 5 (6), 717–730.
Barone, R., Prisinzano, A., 2006. Peculiarita comportamentale del dinoflagellatoOstreopsis ovata Fukujo (Dinophyceae): la strategia del ragno. NaturalistaSiciliano IV 401–418.
Besada, E.G., Loeblich, L.A., Loeblich, A.R.III, 1982. Observations on tropical, benthicdinoflagellates from ciguatera-endemic areas: Coolia, Gambierdiscus, andOstreopsis. Bull. Mar. Sci. 32 (3), 723–735.
Brand, L.E., 1989. Review of genetic variation in marine phytoplankton species andthe ecological implications. Biol. Oceanogr. 6 (5–6), 397–409.
Bravo, I., Figueroa, R.I., Garces, E., Fraga, S., Massanet, A., 2010a. The intricacies ofdinoflagellate pellicle cysts: The example of Alexandrium minutum cysts from abloom-recurrent area (Bay of Baiona, NW Spain). Deep-Sea Res. Pt II 57 (3–4),166–174.
Bravo, I., Vila, M., Magadan, S., Rial, P., Rodriguez, F., Fraga, S., Franco, J.M., Riobo, P.,Sala, M.M., 2010b. Studies on cultures of Ostreopsis cf. ovata: life cycle obser-vations.In: 14th International Conference on Harmful Algae, Hersonissos-Crete,Greece, p. 29.
Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso, E., Forino, M., Tarta-glione, L., 2010a. LC–MS of palytoxin and its analogues: state of the art andfuture perspectives. Toxicon 57 (3), 376–389.
Ciminiello, P., Dell’Aversano, C., Fattorusso, E., Forino, M., Magno, G.S., Tartaglione,L., Grillo, C., Melchiorre, N., 2006. The Genoa 2005 outbreak. Determination of

E. Scalco et al. / Harmful Algae 17 (2012) 25–3434
putative palytoxin in Mediterranean Ostreopsis ovata by a new liquid chroma-tography tandem mass spectrometry method. Anal. Chem. 78 (17), 6153–6159.
Ciminiello, P., Dell’Aversano, E., Fattorusso, E., Fiorino, M., Tartaglione, L., Grillo, C.,Melchiorre, N., 2008. Putative palytoxin and its new analogue, Ovatoxin-a, inOstreopsis ovata collected along the Ligurian coasts during the 2006 toxicoutbreak. J. Am. Soc. Mass Spectrom. 19 (1), 111–120.
Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso, E., Forino, M., Grauso, L.,Tartaglione, L., Guerrini, F., Pistocchi, R., 2010b. Complex palytoxin-like profileof Ostreopsis ovata. Identification of four new ovatoxins by high-resolutionliquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 24(18), 2735–2744.
Deeds, J.R., Schwartz, M.D., 2010. Human risk associated with palytoxin exposure.Toxicon 56 (2), 150–162.
Faust, M.A., 1999. Three new Ostreopsis species (Dinophyceae): O. marinus sp. nov.,O. belizeanus sp. nov., and O. caribbeanus sp. nov. Phycologia 38 (2), 92–99.
Faust, M.A., Gulledge, R.A., 2002. Identifying Harmful Marine Dinoflagellates.National Museum of Natural History, Washington, DC.
Foy, R.H., Gibson, C.E., 1993. The influence of irradiance, photoperiod and tempera-ture on the growth kinetics of three planktonic diatoms. Eur. J. Phycol. 28 (4),203–212.
Fukuyo, Y., 1981. Taxonomical study on benthic dinoflagellates collected in coralreefs. Bull. Japan Soc. Sci. Fish. 47 (8), 967–978.
Geider, R.J., 1987. Light and temperature dependence of the carbon to chlorophyll aratio in microalagae and cyanobacteria: imoplication for physiology and growthof phytoplankton. New Phytol. 106 (1), 1–34.
Graneli, E., Vidyarathna, N.K., Funari, E., Cumaranatunga, P.R.T., Scenati, R., 2010.Can increases in temperature stimulate blooms of the toxic benthic dinoflagel-late Ostreopsis ovata? Harmful Algae 10 (2), 165–172.
Guerrini, F., Pezzolesi, L., Feller, A., Riccardi, M., Ciminiello, P., Dell’Aversano, C.,Tartaglione, L., Dello Iacovo, E., Fattorusso, E., Forino, M., Pistocchi, R., 2010.Comparative growth and toxin profile of cultured Ostreopsis ovata from theTyrrhenian and Adriatic Seas. Toxicon 55 (2–3), 211–220.
Guillard, R.R.L., 1980. Division rates. In: Stein, J.E. (Ed.), Handbook of PhycologicalMethods. Cambridge University Press, Cambridge, pp. 300–311.
Hagiwara, S.-Y., Bolige, A., Zhang, Y., Takahashi, M., Yamagishi, A., Goto, K., 2002.Circadian gating of photoinduction of commitment to cell-cycle transitions inrelation to photoperiodic control to cell reproduction in Euglena. Photochem.Photobiol. 76 (1), 105–115.
Heil, C.A., Maranda, L., Shimizu, Y., 1993. Mucous-associated dinoflagellates: largescale culturing and estimation of growth rate. In: Smayda, T.J., Shimizu, Y.(Eds.), Toxic Phytoplankton Blooms in the Sea. Elsevier Science Publishers, pp.501–505.
Honsell, G., De Bortoli, M., Boscolo, S., Dell’Aversano, C., Tartaglione, L., Battocchi, C.,Penna, A., Berti, F., Fontanive, G., Poli, M., Sosa, S., Yasumoto, T., Tubaro, A., 2011.Ostreopsis cf. ovata (Dinophyceae): toxin content, immunocytochemistry andultrastructure.In: 5th European Phycological Congress, Rhodes Island (Greece),p. 70.
Keller, M.D., Selvin, R.C., Claus, W., Guillard, R.R.L., 1987. Media for the culture ofoceanic ultraphytoplankton. J. Phycol. 23 (4), 633–638.
Lenoir, S., Ten-Hage, L., Turquet, J., Quod, J.P., Bernard, C., Hennion, M.C., 2004. Firstevidence of palytoxin analogues from an Ostreopsis mascarenensis (Dinophy-ceae) benthic bloom in the southwestern Indian Ocean. J. Phycol. 40 (6), 1042–1051.
Mangialajo, L., Bertolotto, R., Cattaneo-Vietti, R., Chiantore, M., Grillo, C., Lemee, R.,Melchiorre, N., Moretto, P., Povero, P., Ruggieri, N., 2008. The toxic benthicdinoflagellate Ostreopsis ovata: quantification of proliferation along the coast-line of Genoa, Italy. Mar. Pollut. Bull. 56 (6), 1209–1214.
Mangialajo, L., Ganzin, N., Accoroni, S., Asnaghi, V., Blanfune, A., Cabrini, M.,Cattaneo-Vietti, R., Chavanon, F., Chiantore, M., Cohu, S., Costa, E., Fornasaro,D., Grossel, H., Marco-Miralles, F., Maso, M., Rene, A., Rossi, A.M., Sala, M.M.,Thibaut, T., Totti, C., Vila, M., Lemee, R., 2011. Trends in Ostreopsis proliferationalong the Northern Mediterranean coasts. Toxicon 57 (3), 408–420.
Monti, M., Minocci, M., Beran, A., Ivesa, L., 2007. First record of Ostreopsis cfr. ovataon macroalgae in the Northern Adriatic Sea. Mar. Pollut. Bull. 54 (5), 598–601.
Nascimento, S.M., Correa, E.V., Menezes, M., Varela, D., Paredes, J., Morris, S., 2012.Growth and toxin profile of Ostreopsis cf. ovata (Dinophyta) from Rio de Janeiro,Brazil. Harmful Algae 13 (1), 1–9.
Oshima, Y., Bolch, C.J., Hallegraeff, G.M., 1992. Toxin composition of resting cysts ofAlexandrium tamarense (Dinophyceae). Toxicon 30 (12), 1539–1544.
Penna, A., Fraga, S., Battocchi, C., Casabianca, C., Giacobbe, M.G., Riobo, P., Vernesi, C.,2010. A phylogeographical study of the toxic benthic dinoflagellate genusOstreopsis Schmidt. J. Biogeogr. 37 (5), 830–841.
Penna, A., Vila, M., Fraga, S., Giacobbe, M.G., Andreoni, F., Riobo, P., Vernesi, C., 2005.Characterization of Ostreopsis and Coolia (Dinophyceae) isolates in the WesternMediterranean Sea based on morphology, toxicity and internal transcribedspacer 5.8S rDNA sequences. J. Phycol. 41 (1), 212–225.
Raniello, R., Lorenti, M., Brunet, C., Buia, M.C., 2004. Photosynthetic plasticity of aninvasive variety Caulerpa racemosa in a costal Mediterranean area: light har-vesting capacity and seasonal acclimation. Mar. Ecol. – Prog. Ser. 271, 113–120.
Rhodes, L., 2010. World-wide occurrence of the toxic dinoflagellate genus OstreopsisSchmidt. Toxicon 57 (3), 400–407.
Rossi, R., Castellano, V., Scalco, E., Serpe, L., Zingone, A., Soprano, V., 2010. Newpalytoxin-like molecules in Mediterranean Ostreopsis cf. ovata (dinoflagellates)and in Palythoa tuberculosa detected by liquid chromatography–electrosprayionization time-of-flight mass spectrometry. Toxicon 56 (8), 1381–1387.
Rost, B., Riebesell, U., Sultemeyer, D., 2006. Carbon acquisition of marine phyto-plankton: effect of photoperiod length. Limnol. Oceanogr. 51 (1), 12–20.
Sansoni, G., Borghini, B., Camici, G., Casotti, M., Righini, P., Rustighi, C., 2003.Fioriture algali di Ostreopsis ovata (Gonyaulacales: Dinophyceae): un problemaemergente. Biologia Ambientale 17 (1), 17–23.
Schmidt, J., 1901. Flora of Koh Chang. Contributions to the knowledge of thevegetation in the Gulf of Siam. Peridiniales. Botanisk Tidskrift 24, 212–221.
Selina, M.S., Orlova, T.Y., 2010. First occurrence of the genus Ostreopsis (Dinophy-ceae) in the Sea of Japan. Bot. Mar. 53 (3), 243–249.
Shears, N.T., Ross, P.M., 2009. Blooms of benthic dinoflagellates of the genusOstreopsis; an increasing and ecologically important phenomenon on temperatereefs in New Zealand and worldwide. Harmful Algae 8 (6), 916–925.
Silva, E.S., Faust, M.A., 1995. Small cells in the life history of dinoflagellates(Dinophyceae): a review. Phycologia 34 (5), 396–408.
Simoni, F., Gaddi, A., Di Paolo, C., Lepri, L., 2003. Harmful epiphytic dinoflagellate onTyrrhenian Sea reef. Harmful Algae News 24, 13–14.
Sunda, W.G., Huntsman, S.A., 2004. Relationships among photoperiod, carbonfixation, growth, chlorophyll a and cellular iron and zinc in a coastal diatom.Limnol. Oceanogr. 48 (5), 1742–1753.
Taylor, F.J.R., 1979. A description of the benthic dinoflagellate associated withmaitotoxin and ciguatoxin, including observations on Hawaiian material. In:Taylor, D.L., Seliger, H.H. (Eds.), Toxic Dinoflagellates Blooms. Elsevier NorthHolland Inc., New York, Amsterdam, Oxford, pp. 71–76.
Tognetto, L., Bellato, S., Moro, I., Andreoli, C., 1995. Occurrence of Ostreopsis ovata(Dinophyceae) in the Tyrrhenian Sea during summer 1994. Bot. Mar. 38 (4),291–295.
Totti, C., Accoroni, S., Cerino, F., Cucchiari, E., Romagnoli, T., 2010. Ostreopsis ovatabloom along the Conero Riviera (northern Adriatic Sea): relationships withenvironmental conditions and substrata. Harmful Algae 9 (2), 233–239.
Ukena, T., Satake, M., Usami, M., Oshima, Y., Fujita, T., Naoki, H., Yasumoto, T., 2002.Structural confirmation of ostreocin-D by application of negative-ion fast-atombombardment collision-induced dissociation tandem mass spectrometricmethods. Rapid Commun. Mass Spectrom. 16 (24), 2387–2393.
Usami, M., Satake, M., Ishida, S., Inoue, A., Kan, Y., Yasumoto, T., 1995. Palytoxinanalogs from the dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 117 (19),5389–5390.
Vila, M., Garces, E., Maso, M., 2001. Potentially toxic epiphytic dinoflagellatesassemblages on macroalgae in the NW Mediterranean. Aquat. Microb. Ecol.26 (1), 51–60.
Copyright © 2022 FDOKUMEN




















