
Growth and metabolism in sugarcane are altered by the creation of a new hexose-phosphate sink
14
Plant Biotechnology Journal (2007) 5, pp. 240–253 doi: 10.1111/j.1467-7652.2006.00235.x 240 © 2007 Blackwell Publishing Ltd Summary An efficient in planta sugarcane-based production system may be realized by coupling the synthesis of alternative products to the metabolic intermediates of sucrose metabolism, thus taking advantage of the sucrose-producing capability of the plant. This was evaluated by synthesizing sorbitol in sugarcane (Saccharum hybrids) using the Malus domestica sorbitol- 6-phosphate dehydrogenase gene (mds6pdh). Mature transgenic sugarcane plants were compared with untransformed sugarcane variety Q117 by evaluation of the growth, metabolite levels and extractable activity of relevant enzymes. The average amounts of sorbitol detected in the most productive line were 120 mg/g dry weight (equivalent to 61% of the soluble sugars) in the leaf lamina and 10 mg/g dry weight in the stalk pith. The levels of enzymes involved in sucrose synthesis and cleavage were elevated in the leaves of plants accumulating sorbitol, but this did not affect sucrose accumulation in the culm. The activity of oxidative reactions in the pentose phosphate pathway and the non-reversible glyceraldehyde-3-phosphate dehydrogenase reaction were elevated to replenish the reducing power consumed by sorbitol synthesis. Sorbitol-producing sugarcane generated 30%−40% less aerial biomass and was 10%−30% shorter than control lines. Leaves developed necrosis in a pattern characteristic of early senescence, and the severity was related to the relative quantity of sorbitol accumulated. When the Zymomonas mobilis glucokinase (zmglk) gene was co-expressed with mds6pdh to increase the production of glucose-6-phosphate, the plants were again smaller, indicating that glucose-6-phosphate deficiency was not responsible for the reduced growth. In summary, sorbitol hyperaccumulation affected sugarcane growth and metabolism, but the outcome was not lethal for the plant. This work also demonstrated that impressive yields of alternative products can be generated from the intermediates of sucrose metabolism in Saccharum spp. Blackwell Publishing, Ltd. Oxford, UK PBI Plant Biotechnology Journal 1467-7644 © 2007 Blackwell Publishing Ltd ? 2007 ? Original Article Sorbitol synthesis in sugarcane Barrie Fong Chonget al. Growth and metabolism in sugarcane are altered by the creation of a new hexose-phosphate sink Barrie Fong Chong 1,2, *, Graham D. Bonnett 2,3 , Donna Glassop 2,3 , Michael G. O’Shea 1,2 and Stevens M. Brumbley 1,2, * , † 1 David North Plant Research Centre, BSES Limited, PO Box 86, Indooroopilly, Qld 4068, Australia 2 Cooperative Research Centre for Sugar Industry Innovation Through Biotechnology, The University of Queensland, St Lucia, Qld 4072, Australia 3 CSIRO, Plant Industry, Queensland Bioscience Precinct, 306 Carmody Road, St Lucia, Qld 4067, Australia Received 22 September 2006; revised 12 November 2006; accepted 17 November 2006. *Correspondence (fax +61 7 3871 0383; e-mail [email protected]) or (fax +61 7 3871 0383; e-mail [email protected]) †Present address: Australian Institute for Bioengineering and Nanotechnology, The University of Queensland, St Lucia, Qld 4072, Australia Keywords: glucose-6-phosphate, Saccharum spp., sorbitol, sorbitol-6- phosphate dehydrogenase. Introduction Sugarcane (Saccharum hybrids) is a very efficient biomass producer as a result of its high photosynthetic rate and specialized C 4 pathway for assimilating atmospheric CO 2 . A large proportion of the biomass is stored in the stem as sucrose. This has led to the concept that this grass would be an efficient in planta biofactory for the synthesis of specific industrial chemicals (McQualter et al., 2005; Ming et al., 2006). With the exception of high-value, low-volume speciality products (e.g. pharmaceuticals), the viability of in planta production for numerous chemicals depends on the generation of substantial yields without a deleterious impact on the host. A logical approach is to match the class of product with the type of crop to be used as a biofactory. For example, oilseed crops are ideally suited for the production of designer oils (Alonso and Maroto, 2000) because they efficiently produce large quantities of fatty acids. A similar rationale proposes that sugarcane has a marked advantage over other plants for the production of chemicals that can be derived from sugar
Transcript of Growth and metabolism in sugarcane are altered by the creation of a new hexose-phosphate sink

Plant Biotechnology Journal (2007) 5, pp. 240–253 doi: 10.1111/j.1467-7652.2006.00235.x
240 © 2007 Blackwell Publishing Ltd
SummaryAn efficient in planta sugarcane-based production system may be realized by coupling the
synthesis of alternative products to the metabolic intermediates of sucrose metabolism, thus
taking advantage of the sucrose-producing capability of the plant. This was evaluated by
synthesizing sorbitol in sugarcane (Saccharum hybrids) using the Malus domestica sorbitol-
6-phosphate dehydrogenase gene (mds6pdh). Mature transgenic sugarcane plants were
compared with untransformed sugarcane variety Q117 by evaluation of the growth,
metabolite levels and extractable activity of relevant enzymes. The average amounts of
sorbitol detected in the most productive line were 120 mg/g dry weight (equivalent to 61%
of the soluble sugars) in the leaf lamina and 10 mg/g dry weight in the stalk pith. The levels
of enzymes involved in sucrose synthesis and cleavage were elevated in the leaves of plants
accumulating sorbitol, but this did not affect sucrose accumulation in the culm. The activity
of oxidative reactions in the pentose phosphate pathway and the non-reversible
glyceraldehyde-3-phosphate dehydrogenase reaction were elevated to replenish the
reducing power consumed by sorbitol synthesis. Sorbitol-producing sugarcane generated
30%−40% less aerial biomass and was 10%−30% shorter than control lines. Leaves
developed necrosis in a pattern characteristic of early senescence, and the severity was
related to the relative quantity of sorbitol accumulated. When the Zymomonas mobilis
glucokinase (zmglk) gene was co-expressed with mds6pdh to increase the production of
glucose-6-phosphate, the plants were again smaller, indicating that glucose-6-phosphate
deficiency was not responsible for the reduced growth. In summary, sorbitol
hyperaccumulation affected sugarcane growth and metabolism, but the outcome was not
lethal for the plant. This work also demonstrated that impressive yields of alternative products
can be generated from the intermediates of sucrose metabolism in Saccharum spp.
Blackwell Publishing, Ltd.Oxford, UKPBIPlant Biotechnology Journal1467-7644© 2007 Blackwell Publishing Ltd? 2007??Original ArticleSorbitol synthesis in sugarcaneBarrie Fong Chong et al.
Growth and metabolism in sugarcane are altered by the creation of a new hexose-phosphate sinkBarrie Fong Chong1,2,*, Graham D. Bonnett2,3, Donna Glassop2,3, Michael G. O’Shea1,2 and Stevens M. Brumbley1,2,*,†1David North Plant Research Centre, BSES Limited, PO Box 86, Indooroopilly, Qld 4068, Australia 2Cooperative Research Centre for Sugar Industry Innovation Through Biotechnology, The University of Queensland, St Lucia, Qld 4072, Australia 3CSIRO, Plant Industry, Queensland Bioscience Precinct, 306 Carmody Road, St Lucia, Qld 4067, Australia
Received 22 September 2006;
revised 12 November 2006;
accepted 17 November 2006.
*Correspondence (fax +61 7 3871 0383;
e-mail [email protected]) or
(fax +61 7 3871 0383;
e-mail [email protected])
†Present address: Australian Institute for
Bioengineering and Nanotechnology,
The University of Queensland, St Lucia,
Qld 4072, Australia
Keywords: glucose-6-phosphate,
Saccharum spp., sorbitol, sorbitol-6-
phosphate dehydrogenase.
Introduction
Sugarcane (Saccharum hybrids) is a very efficient biomass
producer as a result of its high photosynthetic rate and
specialized C4 pathway for assimilating atmospheric CO2. A large
proportion of the biomass is stored in the stem as sucrose.
This has led to the concept that this grass would be an
efficient in planta biofactory for the synthesis of specific industrial
chemicals (McQualter et al., 2005; Ming et al., 2006). With
the exception of high-value, low-volume speciality products
(e.g. pharmaceuticals), the viability of in planta production
for numerous chemicals depends on the generation of
substantial yields without a deleterious impact on the host.
A logical approach is to match the class of product with the
type of crop to be used as a biofactory. For example, oilseed
crops are ideally suited for the production of designer oils (Alonso
and Maroto, 2000) because they efficiently produce large
quantities of fatty acids. A similar rationale proposes that
sugarcane has a marked advantage over other plants for the
production of chemicals that can be derived from sugar

Sorbitol synthesis in sugarcane 241
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
building blocks via a few enzymatic steps. This theory has
already been demonstrated in sugar beet (Sevenier et al., 1998),
a major source of the caloric sweetener sucrose. Sugar beet
transformed with the Helianthus tuberosus fructosyltransferase
converts sucrose into low-molecular-weight fructans, a low-
caloric sweetener. The oligosaccharides accumulate to an
astonishing 40% of the tap root dry weight without
detriment to the host. Sugar manipulation in sugarcane may
be similarly productive.
This work examines sugar manipulation in sugarcane by
engineering a new carbon sink into sugarcane, specifically
the six-carbon sugar alcohol sorbitol. Sorbitol is an ideal
candidate for production in sugarcane because its synthesis
requires only a single enzyme that utilizes glucose-6-
phosphate directly. Moreover, high concentrations of this
compound are likely to be tolerated by the plant because it
is a compatible solute. Sorbitol itself has intrinsic value as a
non-caloric sweetener and is also used to manufacture
ascorbic acid and personal care products (Kirschner, 2004).
The sorbitol-6-phosphate dehydrogenase (S6PDH) gene
(mds6pdh), derived from Malus domestica (Kanayama
et al., 1992), was expressed in sugarcane to catalyse the reduced
nicotinamide adenine dinucleotide phosphate (NADPH)-
assisted reduction of glucose-6-phosphate to sorbitol-6-
phosphate (EC 1.1.1.200). This phosphorylated intermediate
is subsequently cleaved by nonspecific endogenous
phosphatases to liberate sorbitol (Tao et al., 1995).
Sorbitol has been produced in tobacco by the expression
of the mds6pdh gene. Transgenic tobacco plants produced
0.2−130 µmol sorbitol/g fresh weight. Although normal
growth was observed in transgenic tobacco plants producing
less than 3 µmol sorbitol/g fresh weight, aberrant growth
and development (lesions, stunted growth, infertility and
no root growth) was observed with increasing sorbitol con-
centrations (Sheveleva et al., 1998). The altered growth char-
acteristics were attributed to altered carbohydrate allocation
and the depletion of glucose-6-phosphate, affecting cell
wall biosynthesis. A working hypothesis in the present inquiry
presumes that non-trivial quantities of sorbitol can be
synthesized in sugarcane without agronomic penalty. This is
a reasonable expectation as glucose-6-phosphate is rapidly
turned over during sucrose synthesis. Furthermore, it is
replenished from sucrose reserves via a futile cycle (Figure 1)
involving sucrose phosphate synthase (SPS), sucrose synthase
(SuSy) and neutral invertase (INV) (Sacher et al., 1963).
Nonetheless, to protect against the possibility of substrate
depletion, transgenic sugarcane co-expressing mds6pdh and
the Zymomonas mobilis glucokinase (GLK) gene (zmglk, EC
2.7.1.2) (Barnell et al., 1990) was produced. NADPH is also
required for the production of sorbitol-6-phosphate (Figure 1).
It is not considered limiting because there are two main
sources of reducing power in the cell. The first is the two
oxidative reactions of the pentose phosphate pathway, and
the second is the non-reversible glyceraldehyde-3-phosphate
dehydrogenase (GAPDH). The operation of the former
consumes glucose-6-phosphate, whereas the latter is linked
to the photosynthetic oxidation of water. Transgenic sugarcane
plants transformed with mds6pdh only, mds6pdh and zmglk
Figure 1 Metabolic reactions in the cytosol that are involved in the synthesis and breakdown of sorbitol and sucrose in sorbitolcane. FBPase, fructose-1,6-bisphosphatase; FK, fructokinase; GAPDH, non-reversible glyceraldehyde-3-phosphate dehydrogenase; GLK, glucokinase; G6PDH, glucose-6-phosphate dehydrogenase; INV, neutral invertase; NADP, oxidized nicotinamide adenine dinucleotide phosphate; NADPH, reduced nicotinamide adenine dinucleotide phosphate; 6PGDH, 6-phosphogluconate dehydrogenase; PGI, phosphoglucoisomerase; PGM, phosphoglucomutase; SDH, sorbitol dehydrogenase; S6PDH, sorbitol-6-phosphate dehydrogenase; SorPP, sorbitol-6-phosphate phosphatase; SPP, sucrose phosphate phosphatase; SPS, sucrose phosphate synthase; SuSy, sucrose synthase; UGPase, UDP-glucose pyrophosphorylase.

242 Barrie Fong Chong et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
and control plants were produced for this study. Throughout
this article, the sorbitol-producing sugarcane plants are called
‘sorbitolcane’.
The objective of this study was to assess the changes that
occur in sucrose metabolism and photoassimilate partitioning
in sugarcane leaf and culm tissue following the introduction
of a new hexose-phosphate sink. This entailed the measurement
of the plant growth characteristics, tissue-specific carbohydrate
distribution and activities of the enzymes and metabolite
levels in key pathways associated with the provision of glucose-
6-phosphate and NADPH (Figure 1). This study reveals numerous
changes in the sugarcane plant resulting from the synthesis
and accumulation of sorbitol, and also provides a new
perspective into the application of sugarcane as an in planta
biofactory.
Results
Five transgenic sugarcane lines were created for this investi-
gation. Three S-lines (S-10, S-34 and S-76) were engineered
to express mds6pdh and the neomycin phosphotransferase
(nptII) selection marker. Two GS-lines (GS-4 and GS-90) expressed
mds6pdh, zmglk and nptII. Two control plants (C-4 and C-
17), expressing only nptII, were also included in the study.
The introduced enzymes were not targeted to a particular
subcellular compartment and therefore probably localized to
the cytosol of sugarcane cells.
Growth and morphological phenotype of sorbitolcane
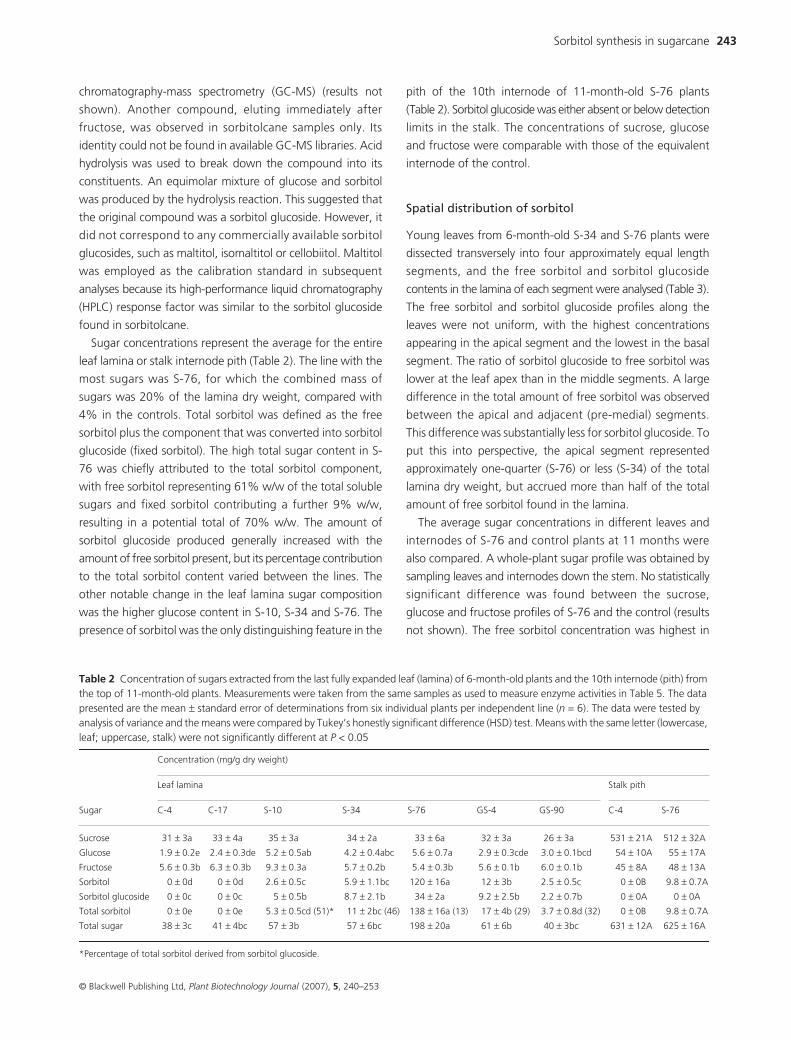
Necrosis was evident in the expanding leaves before the
dewlap was visible. This was most pronounced in S-76, the
plant producing the highest quantity of sorbitol (Figure 2).
Necrosis originated at the leaf apex and advanced towards
the leaf base as the leaf matured. Regions of healthy, bleached
and necrotic tissue were excised from the leaf lamina and
analysed for sorbitol content. The measurements indicated
that the sorbitol concentration (mean ± standard error) was
higher in the necrotic zone (38.5 ± 0.4 mg sorbitol/g dry weight)
than in the bleached (3.7 ± 0.3 mg sorbitol/g dry weight) or
healthy (0.9 ± 0.03 mg sorbitol/g dry weight) zones.
Plant biomass yield and size were evaluated by comparing
two control lines with the sorbitolcane lines S-10, S-34 and
S-76 (Table 1). S-10 and S-34 stalk yield, stalk length and
number of internodes were less than those of the controls. Of
the three S-lines tested, S-76 was larger (in terms of stalk
length), but still produced less stalk biomass than the control
C-4. The plants engineered with enhanced GLK activity
(GS-4 and GS-90) were shorter and yielded less stalk biomass
than the control C-4.
Photosynthate partitioning
Soluble carbohydrates accumulating in the leaf were shown
to be sorbitol, sucrose, glucose and fructose. Sorbitol was
detected in sorbitolcane lines, but was absent in the controls.
The authenticity of the sugars was confirmed by gas
Table 1 Size and biomass yield of 11-month-old sorbitolcane plants. The data presented are the mean ± standard error of determinations from six individual plants per independent line (n = 6). The data were tested by analysis of variance and the means were compared by Tukey’s honestly significant difference (HSD) test. Means with the same letter were not significantly different at P < 0.05
C-4 C-17 S-10 S-34 S-76 GS-4 GS-90
Leaf mass (g FW) 566 ± 29a 590 ± 16a 591 ± 53a 471 ± 30a 550 ± 31a 631 ± 58a 616 ± 53a
Stalk mass (g FW) 1687 ± 63a 1528 ± 138ab 917 ± 126c 788 ± 92c 1016 ± 111bc 1038 ± 136bc 1113 ± 158bc
Stalk length (cm) 301 ± 7a 269 ± 11ab 206 ± 14c 204 ± 12c 260 ± 17abc 241 ± 12bc 230 ± 15bc
No. internodes 34 ± 1a 33 ± 1a 27 ± 1b 25 ± 1b 29 ± 2ab 30 ± 1ab 30 ± 1ab
Tiller mass (g FW) 416 ± 258a 334 ± 185a 149 ± 99a 124 ± 108a 143 ± 84a 47 ± 35a 286 ± 149a
No. tillers 1.5 ± 0.7a 1.2 ± 0.7a 0.3 ± 0.2a 0.3 ± 0.2a 0.5 ± 0.2a 0.7 ± 0.2a 1 ± 0.4a
FW, fresh weight.
Figure 2 Visual appearance of a section of the last fully expanded leaf from an 8-month-old S-76 plant (bottom leaf) and a control plant (top leaf). Necrosis developed at the S-76 leaf apex and margins.
Sorbitol synthesis in sugarcane 243
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
chromatography-mass spectrometry (GC-MS) (results not
shown). Another compound, eluting immediately after
fructose, was observed in sorbitolcane samples only. Its
identity could not be found in available GC-MS libraries. Acid
hydrolysis was used to break down the compound into its
constituents. An equimolar mixture of glucose and sorbitol
was produced by the hydrolysis reaction. This suggested that
the original compound was a sorbitol glucoside. However, it
did not correspond to any commercially available sorbitol
glucosides, such as maltitol, isomaltitol or cellobiitol. Maltitol
was employed as the calibration standard in subsequent
analyses because its high-performance liquid chromatography
(HPLC) response factor was similar to the sorbitol glucoside
found in sorbitolcane.
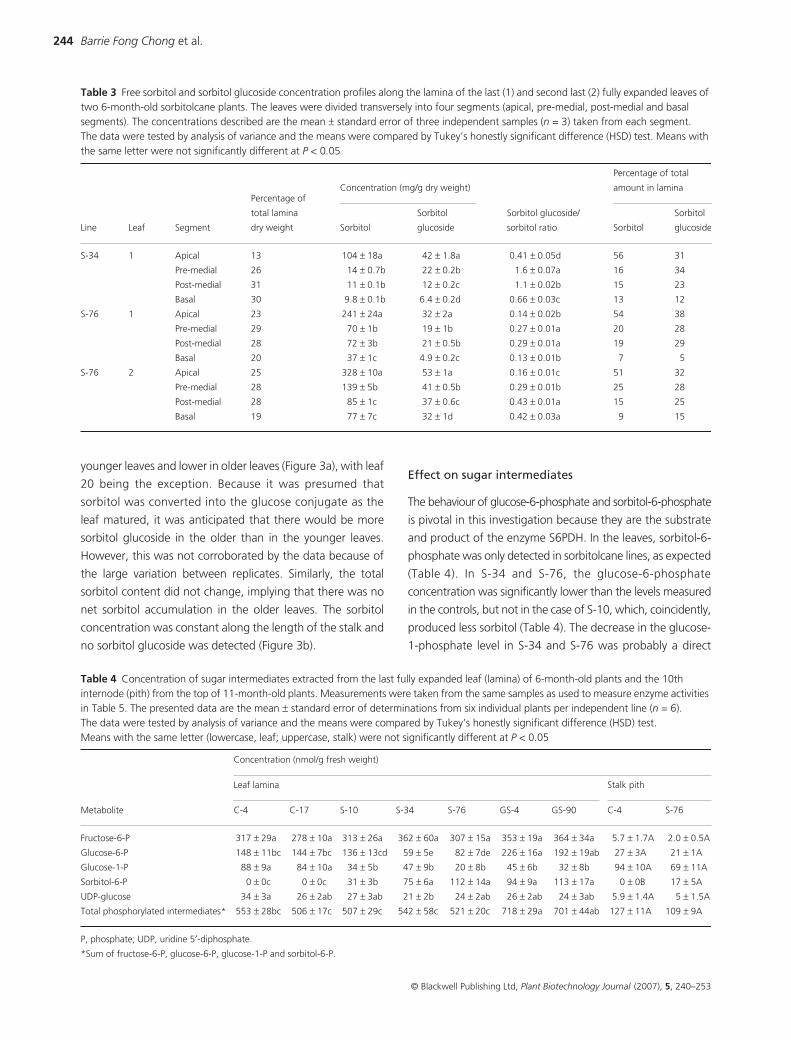
Sugar concentrations represent the average for the entire
leaf lamina or stalk internode pith (Table 2). The line with the
most sugars was S-76, for which the combined mass of
sugars was 20% of the lamina dry weight, compared with
4% in the controls. Total sorbitol was defined as the free
sorbitol plus the component that was converted into sorbitol
glucoside (fixed sorbitol). The high total sugar content in S-
76 was chiefly attributed to the total sorbitol component,
with free sorbitol representing 61% w/w of the total soluble
sugars and fixed sorbitol contributing a further 9% w/w,
resulting in a potential total of 70% w/w. The amount of
sorbitol glucoside produced generally increased with the
amount of free sorbitol present, but its percentage contribution
to the total sorbitol content varied between the lines. The
other notable change in the leaf lamina sugar composition
was the higher glucose content in S-10, S-34 and S-76. The
presence of sorbitol was the only distinguishing feature in the
pith of the 10th internode of 11-month-old S-76 plants
(Table 2). Sorbitol glucoside was either absent or below detection
limits in the stalk. The concentrations of sucrose, glucose
and fructose were comparable with those of the equivalent
internode of the control.
Spatial distribution of sorbitol
Young leaves from 6-month-old S-34 and S-76 plants were
dissected transversely into four approximately equal length
segments, and the free sorbitol and sorbitol glucoside
contents in the lamina of each segment were analysed (Table 3).
The free sorbitol and sorbitol glucoside profiles along the
leaves were not uniform, with the highest concentrations
appearing in the apical segment and the lowest in the basal
segment. The ratio of sorbitol glucoside to free sorbitol was
lower at the leaf apex than in the middle segments. A large
difference in the total amount of free sorbitol was observed
between the apical and adjacent (pre-medial) segments.
This difference was substantially less for sorbitol glucoside. To
put this into perspective, the apical segment represented
approximately one-quarter (S-76) or less (S-34) of the total
lamina dry weight, but accrued more than half of the total
amount of free sorbitol found in the lamina.
The average sugar concentrations in different leaves and
internodes of S-76 and control plants at 11 months were
also compared. A whole-plant sugar profile was obtained by
sampling leaves and internodes down the stem. No statistically
significant difference was found between the sucrose,
glucose and fructose profiles of S-76 and the control (results
not shown). The free sorbitol concentration was highest in
Table 2 Concentration of sugars extracted from the last fully expanded leaf (lamina) of 6-month-old plants and the 10th internode (pith) from the top of 11-month-old plants. Measurements were taken from the same samples as used to measure enzyme activities in Table 5. The data presented are the mean ± standard error of determinations from six individual plants per independent line (n = 6). The data were tested by analysis of variance and the means were compared by Tukey’s honestly significant difference (HSD) test. Means with the same letter (lowercase, leaf; uppercase, stalk) were not significantly different at P < 0.05
Sugar
Concentration (mg/g dry weight)
Leaf lamina Stalk pith
C-4 C-17 S-10 S-34 S-76 GS-4 GS-90 C-4 S-76
Sucrose 31 ± 3a 33 ± 4a 35 ± 3a 34 ± 2a 33 ± 6a 32 ± 3a 26 ± 3a 531 ± 21A 512 ± 32A
Glucose 1.9 ± 0.2e 2.4 ± 0.3de 5.2 ± 0.5ab 4.2 ± 0.4abc 5.6 ± 0.7a 2.9 ± 0.3cde 3.0 ± 0.1bcd 54 ± 10A 55 ± 17A
Fructose 5.6 ± 0.3b 6.3 ± 0.3b 9.3 ± 0.3a 5.7 ± 0.2b 5.4 ± 0.3b 5.6 ± 0.1b 6.0 ± 0.1b 45 ± 8A 48 ± 13A
Sorbitol 0 ± 0d 0 ± 0d 2.6 ± 0.5c 5.9 ± 1.1bc 120 ± 16a 12 ± 3b 2.5 ± 0.5c 0 ± 0B 9.8 ± 0.7A
Sorbitol glucoside 0 ± 0c 0 ± 0c 5 ± 0.5b 8.7 ± 2.1b 34 ± 2a 9.2 ± 2.5b 2.2 ± 0.7b 0 ± 0A 0 ± 0A
Total sorbitol 0 ± 0e 0 ± 0e 5.3 ± 0.5cd (51)* 11 ± 2bc (46) 138 ± 16a (13) 17 ± 4b (29) 3.7 ± 0.8d (32) 0 ± 0B 9.8 ± 0.7A
Total sugar 38 ± 3c 41 ± 4bc 57 ± 3b 57 ± 6bc 198 ± 20a 61 ± 6b 40 ± 3bc 631 ± 12A 625 ± 16A
*Percentage of total sorbitol derived from sorbitol glucoside.
244 Barrie Fong Chong et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
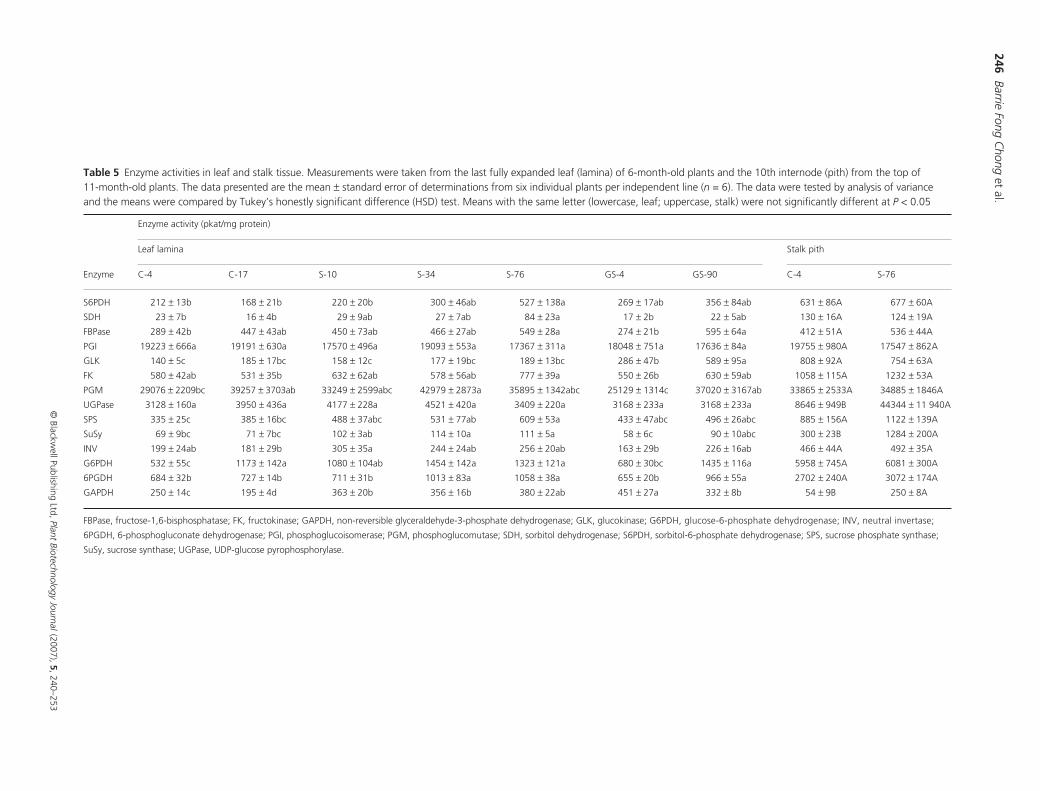
younger leaves and lower in older leaves (Figure 3a), with leaf
20 being the exception. Because it was presumed that
sorbitol was converted into the glucose conjugate as the
leaf matured, it was anticipated that there would be more
sorbitol glucoside in the older than in the younger leaves.
However, this was not corroborated by the data because of
the large variation between replicates. Similarly, the total
sorbitol content did not change, implying that there was no
net sorbitol accumulation in the older leaves. The sorbitol
concentration was constant along the length of the stalk and
no sorbitol glucoside was detected (Figure 3b).
Effect on sugar intermediates
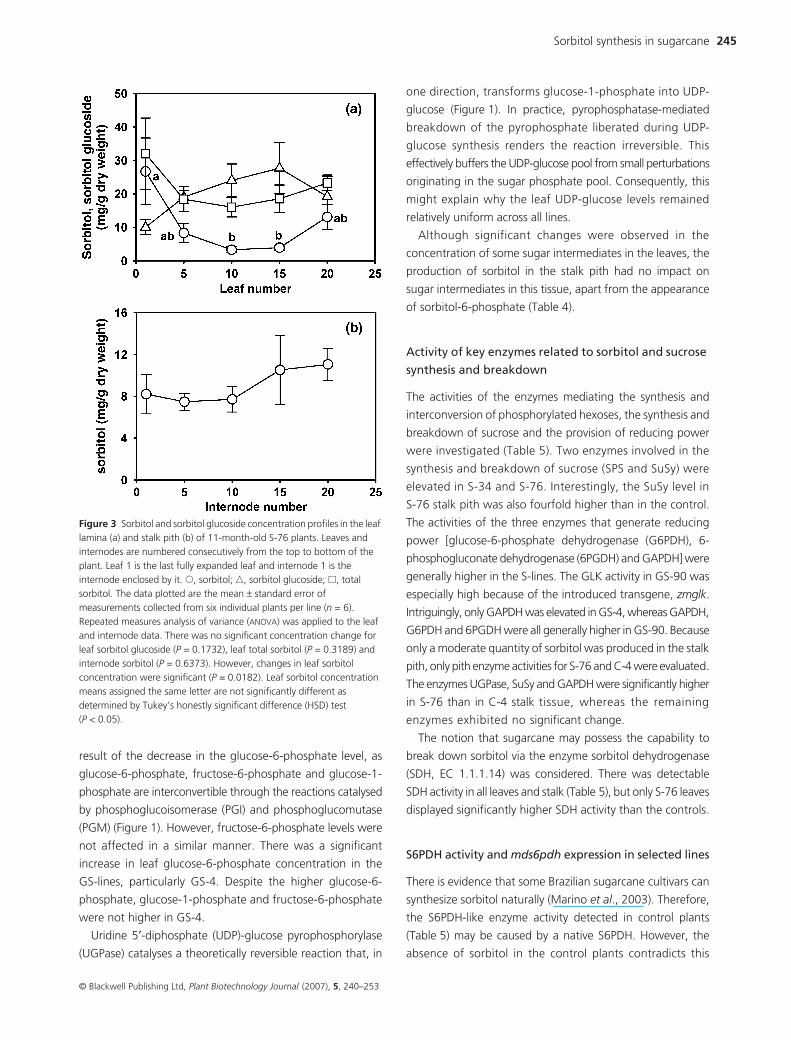
The behaviour of glucose-6-phosphate and sorbitol-6-phosphate
is pivotal in this investigation because they are the substrate
and product of the enzyme S6PDH. In the leaves, sorbitol-6-
phosphate was only detected in sorbitolcane lines, as expected
(Table 4). In S-34 and S-76, the glucose-6-phosphate
concentration was significantly lower than the levels measured
in the controls, but not in the case of S-10, which, coincidently,
produced less sorbitol (Table 4). The decrease in the glucose-
1-phosphate level in S-34 and S-76 was probably a direct
Table 3 Free sorbitol and sorbitol glucoside concentration profiles along the lamina of the last (1) and second last (2) fully expanded leaves of two 6-month-old sorbitolcane plants. The leaves were divided transversely into four segments (apical, pre-medial, post-medial and basal segments). The concentrations described are the mean ± standard error of three independent samples (n = 3) taken from each segment. The data were tested by analysis of variance and the means were compared by Tukey’s honestly significant difference (HSD) test. Means with the same letter were not significantly different at P < 0.05
Line Leaf Segment
Percentage of
total lamina
dry weight
Concentration (mg/g dry weight)
Sorbitol glucoside/
sorbitol ratio
Percentage of total
amount in lamina
Sorbitol
Sorbitol
glucoside Sorbitol
Sorbitol
glucoside
S-34 1 Apical 13 104 ± 18a 42 ± 1.8a 0.41 ± 0.05d 56 31
Pre-medial 26 14 ± 0.7b 22 ± 0.2b 1.6 ± 0.07a 16 34
Post-medial 31 11 ± 0.1b 12 ± 0.2c 1.1 ± 0.02b 15 23
Basal 30 9.8 ± 0.1b 6.4 ± 0.2d 0.66 ± 0.03c 13 12
S-76 1 Apical 23 241 ± 24a 32 ± 2a 0.14 ± 0.02b 54 38
Pre-medial 29 70 ± 1b 19 ± 1b 0.27 ± 0.01a 20 28
Post-medial 28 72 ± 3b 21 ± 0.5b 0.29 ± 0.01a 19 29
Basal 20 37 ± 1c 4.9 ± 0.2c 0.13 ± 0.01b 7 5
S-76 2 Apical 25 328 ± 10a 53 ± 1a 0.16 ± 0.01c 51 32
Pre-medial 28 139 ± 5b 41 ± 0.5b 0.29 ± 0.01b 25 28
Post-medial 28 85 ± 1c 37 ± 0.6c 0.43 ± 0.01a 15 25
Basal 19 77 ± 7c 32 ± 1d 0.42 ± 0.03a 9 15
Table 4 Concentration of sugar intermediates extracted from the last fully expanded leaf (lamina) of 6-month-old plants and the 10th internode (pith) from the top of 11-month-old plants. Measurements were taken from the same samples as used to measure enzyme activities in Table 5. The presented data are the mean ± standard error of determinations from six individual plants per independent line (n = 6). The data were tested by analysis of variance and the means were compared by Tukey’s honestly significant difference (HSD) test. Means with the same letter (lowercase, leaf; uppercase, stalk) were not significantly different at P < 0.05
Concentration (nmol/g fresh weight)
Leaf lamina Stalk pith
Metabolite C-4 C-17 S-10 S-34 S-76 GS-4 GS-90 C-4 S-76
Fructose-6-P 317 ± 29a 278 ± 10a 313 ± 26a 362 ± 60a 307 ± 15a 353 ± 19a 364 ± 34a 5.7 ± 1.7A 2.0 ± 0.5A
Glucose-6-P 148 ± 11bc 144 ± 7bc 136 ± 13cd 59 ± 5e 82 ± 7de 226 ± 16a 192 ± 19ab 27 ± 3A 21 ± 1A
Glucose-1-P 88 ± 9a 84 ± 10a 34 ± 5b 47 ± 9b 20 ± 8b 45 ± 6b 32 ± 8b 94 ± 10A 69 ± 11A
Sorbitol-6-P 0 ± 0c 0 ± 0c 31 ± 3b 75 ± 6a 112 ± 14a 94 ± 9a 113 ± 17a 0 ± 0B 17 ± 5A
UDP-glucose 34 ± 3a 26 ± 2ab 27 ± 3ab 21 ± 2b 24 ± 2ab 26 ± 2ab 24 ± 3ab 5.9 ± 1.4A 5 ± 1.5A
Total phosphorylated intermediates* 553 ± 28bc 506 ± 17c 507 ± 29c 542 ± 58c 521 ± 20c 718 ± 29a 701 ± 44ab 127 ± 11A 109 ± 9A
P, phosphate; UDP, uridine 5′-diphosphate.
*Sum of fructose-6-P, glucose-6-P, glucose-1-P and sorbitol-6-P.
Sorbitol synthesis in sugarcane 245
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
result of the decrease in the glucose-6-phosphate level, as
glucose-6-phosphate, fructose-6-phosphate and glucose-1-
phosphate are interconvertible through the reactions catalysed
by phosphoglucoisomerase (PGI) and phosphoglucomutase
(PGM) (Figure 1). However, fructose-6-phosphate levels were
not affected in a similar manner. There was a significant
increase in leaf glucose-6-phosphate concentration in the
GS-lines, particularly GS-4. Despite the higher glucose-6-
phosphate, glucose-1-phosphate and fructose-6-phosphate
were not higher in GS-4.
Uridine 5′-diphosphate (UDP)-glucose pyrophosphorylase
(UGPase) catalyses a theoretically reversible reaction that, in
one direction, transforms glucose-1-phosphate into UDP-
glucose (Figure 1). In practice, pyrophosphatase-mediated
breakdown of the pyrophosphate liberated during UDP-
glucose synthesis renders the reaction irreversible. This
effectively buffers the UDP-glucose pool from small perturbations
originating in the sugar phosphate pool. Consequently, this
might explain why the leaf UDP-glucose levels remained
relatively uniform across all lines.
Although significant changes were observed in the
concentration of some sugar intermediates in the leaves, the
production of sorbitol in the stalk pith had no impact on
sugar intermediates in this tissue, apart from the appearance
of sorbitol-6-phosphate (Table 4).
Activity of key enzymes related to sorbitol and sucrose
synthesis and breakdown
The activities of the enzymes mediating the synthesis and
interconversion of phosphorylated hexoses, the synthesis and
breakdown of sucrose and the provision of reducing power
were investigated (Table 5). Two enzymes involved in the
synthesis and breakdown of sucrose (SPS and SuSy) were
elevated in S-34 and S-76. Interestingly, the SuSy level in
S-76 stalk pith was also fourfold higher than in the control.
The activities of the three enzymes that generate reducing
power [glucose-6-phosphate dehydrogenase (G6PDH), 6-
phosphogluconate dehydrogenase (6PGDH) and GAPDH] were
generally higher in the S-lines. The GLK activity in GS-90 was
especially high because of the introduced transgene, zmglk.
Intriguingly, only GAPDH was elevated in GS-4, whereas GAPDH,
G6PDH and 6PGDH were all generally higher in GS-90. Because
only a moderate quantity of sorbitol was produced in the stalk
pith, only pith enzyme activities for S-76 and C-4 were evaluated.
The enzymes UGPase, SuSy and GAPDH were significantly higher
in S-76 than in C-4 stalk tissue, whereas the remaining
enzymes exhibited no significant change.
The notion that sugarcane may possess the capability to
break down sorbitol via the enzyme sorbitol dehydrogenase
(SDH, EC 1.1.1.14) was considered. There was detectable
SDH activity in all leaves and stalk (Table 5), but only S-76 leaves
displayed significantly higher SDH activity than the controls.
S6PDH activity and mds6pdh expression in selected lines
There is evidence that some Brazilian sugarcane cultivars can
synthesize sorbitol naturally (Marino et al., 2003). Therefore,
the S6PDH-like enzyme activity detected in control plants
(Table 5) may be caused by a native S6PDH. However, the
absence of sorbitol in the control plants contradicts this
Figure 3 Sorbitol and sorbitol glucoside concentration profiles in the leaf lamina (a) and stalk pith (b) of 11-month-old S-76 plants. Leaves and internodes are numbered consecutively from the top to bottom of the plant. Leaf 1 is the last fully expanded leaf and internode 1 is the internode enclosed by it. �, sorbitol; �, sorbitol glucoside; �, total sorbitol. The data plotted are the mean ± standard error of measurements collected from six individual plants per line (n = 6). Repeated measures analysis of variance (ANOVA) was applied to the leaf and internode data. There was no significant concentration change for leaf sorbitol glucoside (P = 0.1732), leaf total sorbitol (P = 0.3189) and internode sorbitol (P = 0.6373). However, changes in leaf sorbitol concentration were significant (P = 0.0182). Leaf sorbitol concentration means assigned the same letter are not significantly different as determined by Tukey’s honestly significant difference (HSD) test (P < 0.05).
246Barrie Fong C
hong et al.
© Blackw
ell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
Table 5 Enzyme activities in leaf and stalk tissue. Measurements were taken from the last fully expanded leaf (lamina) of 6-month-old plants and the 10th internode (pith) from the top of 11-month-old plants. The data presented are the mean ± standard error of determinations from six individual plants per independent line (n = 6). The data were tested by analysis of variance and the means were compared by Tukey’s honestly significant difference (HSD) test. Means with the same letter (lowercase, leaf; uppercase, stalk) were not significantly different at P < 0.05
Enzyme
Enzyme activity (pkat/mg protein)
Leaf lamina Stalk pith
C-4 C-17 S-10 S-34 S-76 GS-4 GS-90 C-4 S-76
S6PDH 212 ± 13b 168 ± 21b 220 ± 20b 300 ± 46ab 527 ± 138a 269 ± 17ab 356 ± 84ab 631 ± 86A 677 ± 60A
SDH 23 ± 7b 16 ± 4b 29 ± 9ab 27 ± 7ab 84 ± 23a 17 ± 2b 22 ± 5ab 130 ± 16A 124 ± 19A
FBPase 289 ± 42b 447 ± 43ab 450 ± 73ab 466 ± 27ab 549 ± 28a 274 ± 21b 595 ± 64a 412 ± 51A 536 ± 44A
PGI 19223 ± 666a 19191 ± 630a 17570 ± 496a 19093 ± 553a 17367 ± 311a 18048 ± 751a 17636 ± 84a 19755 ± 980A 17547 ± 862A
GLK 140 ± 5c 185 ± 17bc 158 ± 12c 177 ± 19bc 189 ± 13bc 286 ± 47b 589 ± 95a 808 ± 92A 754 ± 63A
FK 580 ± 42ab 531 ± 35b 632 ± 62ab 578 ± 56ab 777 ± 39a 550 ± 26b 630 ± 59ab 1058 ± 115A 1232 ± 53A
PGM 29076 ± 2209bc 39257 ± 3703ab 33249 ± 2599abc 42979 ± 2873a 35895 ± 1342abc 25129 ± 1314c 37020 ± 3167ab 33865 ± 2533A 34885 ± 1846A
UGPase 3128 ± 160a 3950 ± 436a 4177 ± 228a 4521 ± 420a 3409 ± 220a 3168 ± 233a 3168 ± 233a 8646 ± 949B 44344 ± 11 940A
SPS 335 ± 25c 385 ± 16bc 488 ± 37abc 531 ± 77ab 609 ± 53a 433 ± 47abc 496 ± 26abc 885 ± 156A 1122 ± 139A
SuSy 69 ± 9bc 71 ± 7bc 102 ± 3ab 114 ± 10a 111 ± 5a 58 ± 6c 90 ± 10abc 300 ± 23B 1284 ± 200A
INV 199 ± 24ab 181 ± 29b 305 ± 35a 244 ± 24ab 256 ± 20ab 163 ± 29b 226 ± 16ab 466 ± 44A 492 ± 35A
G6PDH 532 ± 55c 1173 ± 142a 1080 ± 104ab 1454 ± 142a 1323 ± 121a 680 ± 30bc 1435 ± 116a 5958 ± 745A 6081 ± 300A
6PGDH 684 ± 32b 727 ± 14b 711 ± 31b 1013 ± 83a 1058 ± 38a 655 ± 20b 966 ± 55a 2702 ± 240A 3072 ± 174A
GAPDH 250 ± 14c 195 ± 4d 363 ± 20b 356 ± 16b 380 ± 22ab 451 ± 27a 332 ± 8b 54 ± 9B 250 ± 8A
FBPase, fructose-1,6-bisphosphatase; FK, fructokinase; GAPDH, non-reversible glyceraldehyde-3-phosphate dehydrogenase; GLK, glucokinase; G6PDH, glucose-6-phosphate dehydrogenase; INV, neutral invertase;
6PGDH, 6-phosphogluconate dehydrogenase; PGI, phosphoglucoisomerase; PGM, phosphoglucomutase; SDH, sorbitol dehydrogenase; S6PDH, sorbitol-6-phosphate dehydrogenase; SPS, sucrose phosphate synthase;
SuSy, sucrose synthase; UGPase, UDP-glucose pyrophosphorylase.
Sorbitol synthesis in sugarcane 247
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
suggestion. The S6PDH assay employed in this study was an
indirect measure of the sorbitol-synthesizing capability in
tissue extracts because it was based on the reverse reaction,
i.e. the oxidation of sorbitol-6-phosphate. Consequently, the
S6PDH-like activity detected in the controls may not be the
result of a legitimate S6PDH enzyme. Thus, a more accurate
representation of the true MdS6PDH activity in the S- and GS-
lines may be the difference between the measured values
and the value obtained for the controls.
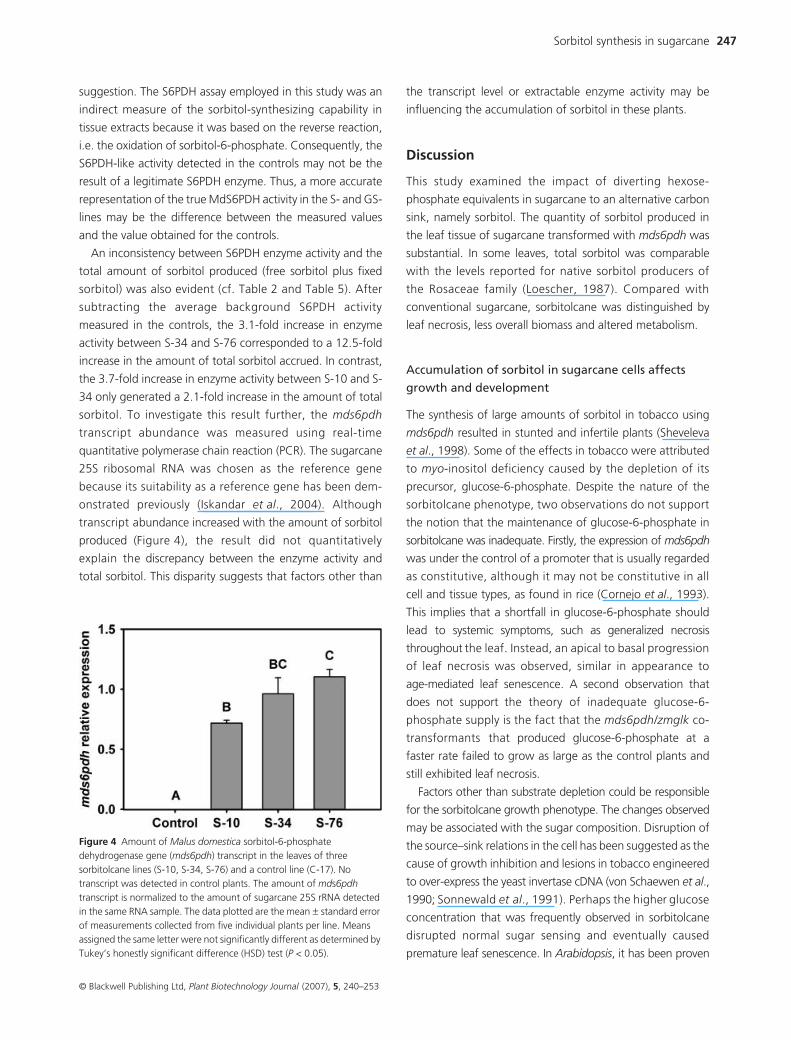
An inconsistency between S6PDH enzyme activity and the
total amount of sorbitol produced (free sorbitol plus fixed
sorbitol) was also evident (cf. Table 2 and Table 5). After
subtracting the average background S6PDH activity
measured in the controls, the 3.1-fold increase in enzyme
activity between S-34 and S-76 corresponded to a 12.5-fold
increase in the amount of total sorbitol accrued. In contrast,
the 3.7-fold increase in enzyme activity between S-10 and S-
34 only generated a 2.1-fold increase in the amount of total
sorbitol. To investigate this result further, the mds6pdh
transcript abundance was measured using real-time
quantitative polymerase chain reaction (PCR). The sugarcane
25S ribosomal RNA was chosen as the reference gene
because its suitability as a reference gene has been dem-
onstrated previously (Iskandar et al., 2004). Although
transcript abundance increased with the amount of sorbitol
produced (Figure 4), the result did not quantitatively
explain the discrepancy between the enzyme activity and
total sorbitol. This disparity suggests that factors other than
the transcript level or extractable enzyme activity may be
influencing the accumulation of sorbitol in these plants.
Discussion
This study examined the impact of diverting hexose-
phosphate equivalents in sugarcane to an alternative carbon
sink, namely sorbitol. The quantity of sorbitol produced in
the leaf tissue of sugarcane transformed with mds6pdh was
substantial. In some leaves, total sorbitol was comparable
with the levels reported for native sorbitol producers of
the Rosaceae family (Loescher, 1987). Compared with
conventional sugarcane, sorbitolcane was distinguished by
leaf necrosis, less overall biomass and altered metabolism.
Accumulation of sorbitol in sugarcane cells affects
growth and development
The synthesis of large amounts of sorbitol in tobacco using
mds6pdh resulted in stunted and infertile plants (Sheveleva
et al., 1998). Some of the effects in tobacco were attributed
to myo-inositol deficiency caused by the depletion of its
precursor, glucose-6-phosphate. Despite the nature of the
sorbitolcane phenotype, two observations do not support
the notion that the maintenance of glucose-6-phosphate in
sorbitolcane was inadequate. Firstly, the expression of mds6pdh
was under the control of a promoter that is usually regarded
as constitutive, although it may not be constitutive in all
cell and tissue types, as found in rice (Cornejo et al., 1993).
This implies that a shortfall in glucose-6-phosphate should
lead to systemic symptoms, such as generalized necrosis
throughout the leaf. Instead, an apical to basal progression
of leaf necrosis was observed, similar in appearance to
age-mediated leaf senescence. A second observation that
does not support the theory of inadequate glucose-6-
phosphate supply is the fact that the mds6pdh/zmglk co-
transformants that produced glucose-6-phosphate at a
faster rate failed to grow as large as the control plants and
still exhibited leaf necrosis.
Factors other than substrate depletion could be responsible
for the sorbitolcane growth phenotype. The changes observed
may be associated with the sugar composition. Disruption of
the source–sink relations in the cell has been suggested as the
cause of growth inhibition and lesions in tobacco engineered
to over-express the yeast invertase cDNA (von Schaewen et al.,
1990; Sonnewald et al., 1991). Perhaps the higher glucose
concentration that was frequently observed in sorbitolcane
disrupted normal sugar sensing and eventually caused
premature leaf senescence. In Arabidopsis, it has been proven
Figure 4 Amount of Malus domestica sorbitol-6-phosphate dehydrogenase gene (mds6pdh) transcript in the leaves of three sorbitolcane lines (S-10, S-34, S-76) and a control line (C-17). No transcript was detected in control plants. The amount of mds6pdh transcript is normalized to the amount of sugarcane 25S rRNA detected in the same RNA sample. The data plotted are the mean ± standard error of measurements collected from five individual plants per line. Means assigned the same letter were not significantly different as determined by Tukey’s honestly significant difference (HSD) test (P < 0.05).

248 Barrie Fong Chong et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
that glucose induces the expression of phospholipase D
(Xiao et al., 2000), which is a promoter of leaf senescence
(Fan et al., 1997). Glucose is also implicated in the dis-
mantling of the photosynthetic apparatus during senescence
(Wingler et al., 1998).
The quantity of sorbitol that accumulated at the leaf apex
may not be entirely age dependent. It is possible that sorbitol
diffused into xylem vessels and was subsequently propelled
to the leaf apex by guttation. A high osmolality resulting from
the extreme concentration of sorbitol may be problematic,
because it can lead to metabolic imbalances between
subcellular compartments, which may cause aberrant growth
(Heineke et al., 1992). The generally lower sorbitol glucoside/
sorbitol ratio at the leaf apex conflicts with the contention
that sorbitol is translocated in the guttation fluid. However,
the result is consistent in the presence of a competing process,
such as the translocation of sorbitol glucoside into vacuoles.
Many plants employ glucose as a tag to target secondary
metabolites and endogenous/exogenous toxins into vacuoles
(Bartholomew et al., 2002).
The possibility that the reduced growth observed in
sorbitolcane may be related to a reduced photosynthetic rate
was also explored by measuring the in situ CO2 assimilation
rate from a healthy region of the first fully expanded leaf on
each plant. The average CO2 assimilation rates were not
significantly different under the test conditions (results not
shown). Although the photosynthetic rates on an area basis
were similar, a diminished green leaf area caused by necrosis
may reduce the overall CO2 uptake, thus restricting stalk growth.
Osmolyte tolerance varies between plant species. The
synthesis of low to moderate levels of mannitol (Karakas
et al., 1997) or sorbitol (Sheveleva et al., 1998) in tobacco
had a limited impact, but high sorbitol levels (> 15 µmol sorbitol/
g fresh weight) led to major metabolic imbalances and severely
disrupted growth and development. Persimmon engineered
with high levels of sorbitol (49 µmol sorbitol/g fresh weight)
exhibited severe dwarfism but, surprisingly, no necrosis
(Deguchi et al., 2004). In this respect, sugarcane demonstrates
resilience compared with many other plants. Although the
reduced growth and leaf necrosis observed in sorbitol-
producing sugarcane are undesirable, it is still possible to
propagate sorbitolcane vegetatively, and re-growth also occurs
when the previous shoot is removed.
Enzymes that synthesize and cleave sucrose are more
abundant in sorbitolcane leaves
Sucrose concentration in the leaves was largely unaffected by
the sorbitol sink. Glucose-6-phosphate is an allosteric activator
of SPS in many plants (Huber and Huber, 1996). The lower
glucose-6-phosphate concentration in sorbitolcane did not
seem to impede sucrose synthesis. In addition, fructose-6-
phosphate and UDP-glucose concentrations were unaffected
by the production of sorbitol; hence, these probably did not
affect the underlying catalytic rate of SPS either.
In some sorbitolcane lines, SPS and SuSy levels were higher
in leaf tissue. The activity of these enzymes is often greater in
tissue under osmotic stress. Sweet potato grown on medium
containing a high concentration of sorbitol showed increased
SPS, SuSy and INV enzyme activities and SPS transcription
(Wang et al., 2000). The up-regulation of both sucrose
synthesis and cleavage enzymes may permit the cell to respond
more rapidly to changes in osmotic stress by adjusting the
rate of carbon storage. The sucrose futile cycle in sorbitolcane
leaves did not lead to an increase in sucrose synthesis and,
hence, the sucrose concentration in the stalk was unchanged.
The oxidative pentose phosphate pathway and the
non-reversible GAPDH are more active in sorbitolcane
The reduction of glucose-6-phosphate to sorbitol-6-phosphate
is reliant on NADPH. Two sources of reducing power were
examined in this study: the oxidative reactions of the pentose
phosphate pathway and the non-reversible GAPDH. A mate-
rial disadvantage is associated with the operation of G6PDH
because it competes with S6PDH for glucose-6-phosphate.
The non-reversible version of GAPDH is restricted to plants
(Dennis and Blakeley, 2000) and, in the leaves, it is located in
the cytosol, where it is a component of a redox shuttle that
utilizes triose phosphates to transfer reducing equivalents
from the chloroplast to the cytosol (Kelly and Gibbs, 1973).
In contrast with G6PDH, carbon is conserved in this process
by recycling the triose phosphate redox carrier between the
two compartments.
As expected, the activities of G6PDH, 6PGDH and GAPDH
were generally higher in sorbitolcane leaves that accumulated
appreciable amounts of sorbitol. The exception was GS-4,
which was principally reliant on GAPDH. The higher activity
of these enzymes underscores their importance in the supply
of NADPH for the S6PDH reaction in photosynthetic tissue.
Moreover, there is no evidence of a preferred route for obtaining
the extra NADPH.
It was envisaged that reducing power in the stalk pith
would originate chiefly via G6PDH and 6PGDH. The effectiveness
of GAPDH in the stalk would be diminished, because there is
no photosynthetic avenue to regenerate the electron donor,
glyceraldehyde-3-phosphate. The lower GAPDH activity in
the pith compared with the leaves of the control plants

Sorbitol synthesis in sugarcane 249
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
supports this notion. Surprisingly, however, G6PDH and 6PGDH
activities were not higher in S-76 stalk pith compared with
the control. Instead, the GAPDH activity was almost fivefold
higher. Therefore, it seems that the reducing power used to
synthesize sorbitol in the pith may be derived from the
GAPDH reaction operating in a mode that leads to the net
consumption of glyceraldehyde-3-phosphate.
In conclusion, a number of metabolic changes accompany
the formation of sorbitol in sugarcane. Most notable are the
increased activity of enzymes that can increase the futile
cycling of sucrose and the increased demand on the
machinery that generates reducing power. The synthesis
and accumulation of sorbitol appear to impede growth, but
the plants can still be propagated vegetatively. There is no
evidence linking the reduced growth to a deficit in glucose-
6-phosphate. Reduced growth may be caused by factors
related to sorbitol-mediated stress, the precise nature of
which requires further elucidation. Two items of circumstantial
evidence suggest that the sugarcane plant may possess
measures to lessen the effects of sorbitol hyperaccumulation.
Firstly, sorbitol may be re-assimilated via SDH, and secondly,
conjugation of sorbitol with glucose may diminish its impact.
This study has demonstrated that sugarcane metabolic
intermediates that are normally used to synthesize sucrose
can be readily diverted to an alternative sugar sink.
Experimental procedures
Cloning of mds6pdh and zmglk expression cassettes
Plasmid constructs were cloned into Escherichia coli DH10B
(Invitrogen Corp., Carlsbad, CA, USA) and subsequently grown
on Luria–Bertani (LB) medium containing 100 µg/mL of
ampicillin. Plant expression cassettes were constructed in the
plasmid pU3z-ubi-nos, a version of pAHC20 (Christensen and
Quail, 1996), which contains the constitutive maize ubi-1
promoter and nopaline synthase (nos) terminator. The
M. domestica mds6pdh (GENBANK accession number D11080)
was cloned from cDNA derived from young leaves of the
apple variety Royal Gala. The 933-bp mds6pdh cDNA, with
SpeI-KpnI restriction sites engineered at the ends, was cloned
into the vector pU3z-ubi-nos immediately downstream of the
ubi-1 promoter. The bacterial strain Z. mobilis ATCC 29191
(Australian Collection of Microorganisms, The University of
Queensland) was grown anaerobically on peptone–yeast
extract medium at pH 7.2 and 28 °C. The Z. mobilis zmglk
(GENBANK accession number M60615) was amplified by PCR
from genomic DNA isolated from the bacterial culture. The
984-bp zmglk gene, with SpeI-KpnI sites engineered at the
ends, was cloned downstream of the ubi-1 promoter in the
vector pU3z-ubi-nos. A consensus monocot ribosomal binding
site, CACC (Joshi et al., 1997), was also included immediately
upstream of the initiator codon of the cloned genes.
Sugarcane transformation
Embryogenic callus was prepared from sugarcane cultivar
Q117, as described previously (Franks and Birch, 1991). The
callus was propagated in the dark at 25 ± 1 °C on Murashige–
Skoog (MS) agar (Murashige and Skoog, 1962) supplemented
with 3 mg/L of 2,4-dichlorophenoxy acetic acid (2,4-D). The
antibiotic selection plasmid pUKN (Joyce et al., 1998), containing
the ubi-1 promoter, the nptII gene and the nos terminator, was
used to select transgenic plants. The expression cassette(s)
containing the gene(s) of interest and pUKN was co-
transformed into calli by microprojectile bombardment
(Bower et al., 1996). Following a 10-day post-transformation
recovery period in the dark, the calli were transferred to
MS−2,4-D agar that was supplemented with 150 mg/L of
paramomycin. Two weeks later, antibiotic-resistant calli
were transferred to MS agar that was supplemented with
150 mg/L of paramomycin, and incubated at 25 ± 1 °C with
a 16-h photoperiod provided by cool white fluorescent
tubes (photon flux density, 30 µmol/m2/s). Individual callus
clumps were segregated throughout the selection and
regeneration period and subcultured on to fresh agar every
2 weeks. After 8 weeks of regeneration, healthy shoots and
roots developed and the plants were transferred to a
glasshouse. 100 S-lines, 125 GS-lines and 10 controls were
regenerated from the callus under these conditions.
Plant cultivation and sampling
All sugarcane plants that were transferred to the glasshouse
were propagated individually in pots (diameter, 20 cm)
containing a soil substitute comprising equal parts of perlite
and vermiculite (Chillagoe Perlite, Mareeba, Qld, Australia).
The soil was supplemented at three-monthly intervals with
Osmocote slow-release nitrogen fertilizer granules (Scotts
Australia Pty Ltd, Baulkam Hills, NSW, Australia). Plants were
watered for 20 min at 06.00 h and 15.00 h daily at a rate of
2 L/h by drippers connected to an automatic irrigation
system (Hunter SRC Irrigation Controller, Hunter Industries
Incorporated, San Marcos, CA, USA).
Genomic integration of the desired genes was confirmed
by PCR testing for the presence of mds6pdh and zmglk in
crude DNA extracts prepared from the leaf tissue of 1-month-old
plants. Seventy per cent of the S-lines contained mds6pdh and

250 Barrie Fong Chong et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
47% of the GS-lines contained both mds6pdh and zmglk.
Plants that were positive for the desired genes were sub-
sequently tested for sorbitol synthesis.
Three independent S-lines, two independent GS-lines and
two controls were grown for a further 9 months, when
vegetative cuttings were re-propagated to generate biological
replicates. The re-propagated lines were grown for 11 months
and sampled twice during this period. At 6 months, the last
fully expanded leaf from each plant was collected, immediately
frozen in liquid nitrogen and stored at −80 °C until analysis.
At 11 months, the first, fifth, 10th, 15th and 20th leaves
(leaf 1, last fully expanded leaf) and the internode enclosed by
it were harvested. The pith tissue in the internodes was
collected, frozen in liquid nitrogen and stored at −80 °C until
analysis.
Real-time quantitative PCR
Total RNA was extracted from 80 mg of sugarcane leaf
lamina using the Qiagen RNeasy Plant Kit (Qiagen Pty Ltd,
Doncaster, Vic., Australia). cDNA was generated from 100 ng
of DNase I-treated RNA using random hexamer primers and
the ImProm-II reverse transcription system (Promega Corp.,
Madison, WI, USA).
The real-time quantitative PCR probes carried a 5′ FAM
reporter and 3′ BHQ1 quencher. Primers and probes were
designed to align with target DNA derived from the M. domestica
s6pdh (GENBANK accession number D11080) (forward primer,
TTCAAGATGTCCACCGTCAC; reverse primer, AAGCTCGTC-
CTTCTCCAGAC; probe, CGGTCATCGGTCTCGGCCTT) and
reference DNA from the sugarcane 25S ribosomal RNA gene
(GENBANK accession number BQ536525) (forward primer,
CGGCTCTTCCTATCATTGTG; reverse primer, CCTGTCTCA-
CGACGGTCTAA; probe, TTCACCCACCAATAGGGAACGTGA).
Transcript levels were calculated from real-time PCR curves
using the relative standard curve method (Applied Biosystems
Sequence Detection Systems Chemistry Guide, 2003). A
reference gene and target gene PCR was set up for each sample
and comprised 125 pg cDNA, 0.9 µM of each primer, 0.2 µM
of probe and Taqman Universal PCR Master Mix (Applied
Biosystems, Foster City, CA, USA). Relative standards prepared
by serial dilution of the samples were also included in each
run. A CAS-1200 liquid handling robot (Corbett Research Pty
Ltd, Mortlake, NSW, Australia) was employed to assemble
reaction mixtures and perform serial dilutions. The PCRs were
run on a Rotagene 3000 (Corbett Research Pty Ltd) using the
universal PCR cycling conditions recommended by Applied
Biosystems (95 °C for 10 min, followed by 50 cycles of 95 °Cfor 15 s and 60 °C for 1 min).
Determination of enzyme activities
Cell-free leaf extracts were prepared from approximately 1 g
of leaf lamina; 5% w/w polyvinylpolypyrrolidone (PVPP) was
added to the tissue before homogenization in liquid nitrogen.
The homogenate was re-suspended in 2 mL of extraction
buffer comprising 50 mM N-2-hydroxyethylpiperazine-N′-2-
ethanesulphonic acid (HEPES)–KOH (pH 7.5), 10 mM MgCl2, 1 mM
ethylenediaminetetraacetic acid (EDTA), 1 mM ethylene glycol-
bis(β-aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA), 5%
v/v glycerol, 0.1% v/v Triton X-100, 10 mM dithiothreitol, 1 mM
benzamidine–HCl, 1 mM benzamide, 1 mM phenylmethyl-
sulphonyl fluoride, 5 mM aminocapronate, 2 µM antipain and
2 µM leupeptin. The slurry was then centrifuged for 15 min at
3100 g and 4 °C, and the supernatant was desalted using a
3-mL Sephadex G-25-M column (Amersham Biosciences,
Uppsala, Sweden). The column was pre-equilibrated with
extraction buffer, loaded with 900 µL of supernatant and
then eluted with 1.2 mL of extraction buffer. Extracts were
prepared and maintained at 4 °C until assay. Cell-free stem
extracts were prepared analogously, except for the following
modifications. Approximately 4 g of pith tissue was used and
the resultant homogenate was re-suspended in 8 mL of
extraction buffer. The volume of supernatant recovered from
the slurry was reduced to approximately 1 mL using Amicon
Ultra-4, 10 000 NMWL centrifugal filters (Millipore, Bedford,
MA, USA). This was then desalted as described for the leaf
extracts and subsequently assayed.
Fructokinase (EC 2.7.1.4) (Renz et al., 1993), cytosolic fructose
1,6-bisphosphatase (EC 3.1.3.11) (Hurry et al., 1995), GAPDH
(EC 1.2.1.9) (Gao and Loescher, 2000), GLK (EC 2.7.1.2) (Scopes
et al., 1985), G6PDH (EC 1.1.1.49) (Wright et al., 1997), 6PGDH
(EC 1.1.1.43) (Magel et al., 2001), PGI (EC 5.3.1.9) (Burrell
et al., 1994), PGM (EC 5.4.2.2) (Manjunath et al., 1998), SDH
(EC 1.1.1.14) (Yamaguchi et al., 1994), S6PDH (EC 1.1.1.200)
(Tao et al., 1995) and UGPase (EC 2.7.7.9) (Ciereszko et al.,
2001) assays were assembled as described, with the exception
that the reaction buffer employed was Tris-HCl and the
optimum pH was determined for each reaction. The assays
were conducted at 30 °C and quantified by tracking the
change in absorbance in a spectrophotometer (Helios γ,Thermo Electron Corporation, Cambridge, Cambridgeshire,
UK). Neutral INV (EC 3.2.1.26) (Albertson et al., 2001), SPS
(EC 2.4.1.14) (Grof et al., 1998) and SuSy (EC 2.4.1.13) (Zhu
et al., 1997) assays were assembled as described. The assays
were conducted at 37 °C and quantified by measuring the
reaction products using the HPLC method outlined in the
next section. The cell-free extract protein concentration was
determined by the Bradford method (Bradford, 1976) using

Sorbitol synthesis in sugarcane 251
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
bovine serum albumin as the protein standard. Specific enzyme
activities were expressed in units of katals per milligram of
total protein.
Carbohydrate analysis
Approximately 120 mg of plant tissue was vacuum dried and
subsequently homogenized in an FP120 FastPrep cell disruptor
(Savant Instruments, Inc., Holbrook, NY, USA); 750 µL of
0.02% w/v sodium azide was added to the tissue powder.
After 3 h of incubation at 80 °C, the supernatant was collected
and the extraction step was repeated on the pellet. The
pooled extracts were filtered through a 0.2-µm syringe filter
and diluted as required with 0.02% w/v sodium azide.
The sugars in the extract were analysed by isocratic HPLC.
The instrument system consisted of an LC-10AT VP pump
(Shimadzu Corporation, Kyoto, Japan), DGU-12 A inline degasser
(Shimadzu Corporation), Sugar-Pak Guard-Pak guard column
(Waters, Milford, MA, USA), 7.8 × 300-mm Aminex HPX-87P
analytical column (Bio-Rad, Hercules, CA, USA) and RID-10 A
VP refractive index detector (Shimadzu Corporation). The
aqueous mobile phase was pumped at a flow rate of 0.8 mL/
min and the column was operated at 80 °C (CTO-10 A VP
column oven, Shimadzu Corporation); 20 µL of sample was
injected per run (SIL-10AD VP autosampler, Shimadzu Corpo-
ration). The resolved sugars were quantified by refractive
index measurement, and converted to concentration units
by comparison with calibration standards prepared from
analytical grade reagents obtained from Sigma-Aldrich
(St. Louis, MO, USA).
Measurement of sugar intermediates
Fructose-6-phosphate, glucose-6-phosphate, glucose-1-
phosphate, sorbitol-6-phosphate and UDP-glucose were
extracted as described in McLaughlin (2005), with the
following modifications. Approximately 100 mg of tissue was
homogenized in liquid nitrogen; 1 mL of chilled extraction
buffer (91% v/v methanol, 6.1 mM HEPES, pH 7) was then
added to the tissue homogenate. After incubating for 10 min
at 4 °C, the supernatant was recovered by centrifugation.
The pellet was washed four times by repeating the previous
step using 1 mL of wash buffer (75% v/v methanol, 5 mM
HEPES, pH 7) in each wash step. The sugar phosphates and
UDP-glucose were isolated from the methanol extract on a
strong anion exchange (SAX) solid phase extraction (SPE)
column (Isolute, International Sorbent Technology, Hengoed,
Mid-Glamorgan, UK). The SPE steps were carried out as
described in McLaughlin (2005).
The analytes were separated on a Dionex DX500 ion
chromatography system (Dionex Corporation, Sunnyvale,
CA, USA) consisting of a GP50 microbore pump, Dionex
AminoTrap guard column (2 × 50 mm), Dionex CarboPac
PA10 anion exchange column (2 × 250 mm) and ED40
electrochemical detector. The column was operated at 30 °C(Dionex AS50 thermal compartment). The electrochemical
detector was equipped with a gold working electrode and
Ag/AgCl reference electrode. Eluents were prepared from
helium-degassed water and carbonate-free liquid sodium
hydroxide, and stored under a nitrogen atmosphere. The
mobile phase, comprising a gradient mixture of sodium
acetate and sodium hydroxide, was pumped at a flow rate of
0.25 mL/min; 25 µL of sample was injected per run (Dionex
AS50 autosampler). The resolved analytes were quantified by
integrated amperometry, and converted to concentration
units by comparison with calibration standards prepared
from analytical grade reagents obtained from Sigma-Aldrich.
Three different gradient conditions were needed to
separate glucose-6-phosphate/fructose-6-phosphate, UDP-
glucose and glucose-1-phosphate/sorbitol-6-phosphate. The
gradient composition and profiles used to separate glucose-6-
phosphate/fructose-6-phosphate and UDP-glucose are described
in McLaughlin (2005). The eluents used to separate glucose-
1-phosphate and sorbitol-6-phosphate were as follows: A,
1 mM sodium acetate in 75 mM sodium hydroxide; B, 75 mM
sodium hydroxide. The gradient profile applied during the
run was as follows: a linear gradient from 5% A and 95% B
to 15% A and 85% B between 0 and 30 min; a concave gra-
dient to 50% A and 50% B at 35 min; a linear gradient to
100% A at 35.1 min; a linear gradient to 5% A and 95% B
from 40 to 40.1 min; finally, 5% A and 95% B maintained to
50 min.
The recovery of this method was determined by extracting
sugarcane tissue that had been spiked with analytical
standards. The recoveries obtained for glucose-6-phosphate,
glucose-1-phosphate, fructose-6-phosphate, sorbitol-6-
phosphate and UDP-glucose were 95%, 88%, 75%, 79%
and 85%, respectively.
Photosynthetic rate
The photosynthetic rate was determined by measuring the
CO2 exchange in sugarcane leaves in situ using an LI-6400
portable photosynthesis system (LI-COR Biosciences, Inc.,
Lincoln, NE, USA). Measurements were taken from a leaf area
of 6 cm2 at a photon flux of 2000 µmol/m2/s, which was set
by the LED light source in the leaf chamber. The air flow
rate through the leaf chamber was set to 400 µmol/s and

252 Barrie Fong Chong et al.
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
reference CO2 was controlled at 400 µmol CO2/mol by the
CO2 mixer.
Statistical analysis of the data
All statistical analysis was performed using the software
Statistix 8 for Windows (Analytical Software, Tallahassee, FL,
USA) or SAS/STAT (SAS Institute Inc., Cary, NC, USA).
Acknowledgements
We are grateful to Niall Masel (BSES Limited, Indooroopilly,
Qld, Australia) and Peter Abeydeera (Department of Chemical
Engineering, The University of Queensland, St Lucia, Qld,
Australia) for their assistance with the sugar and sugar inter-
mediate chromatography analysis. We are also indebted to
Sooknam Patterson who performed the tissue culture. This
work was funded through the Cooperative Research Centre
for Sugar Industry Innovation Through Biotechnology
(CRCSIIB).
References
Albertson, P.L., Peters, K.F. and Grof, C.P.L. (2001) An improvedmethod for the measurement of cell wall invertase activity in sug-arcane tissue. Aust. J. Plant Physiol. 28, 323–328.
Alonso, D.L. and Maroto, F.G. (2000) Plants as ‘chemical factories’for the production of polyunsaturated fatty acids. Biotechnol.Adv. 18, 481–497.
Barnell, W.O., Yi, K.C. and Conway, T. (1990) Sequence and geneticorganization of a Zymomonas mobilis gene cluster that encodesseveral enzymes of glucose metabolism. J. Bacteriol. 172, 7227–7240.
Bartholomew, D.M., Van Dyk, D.E., Lau, S.-M.C., O’Keefe, D.P.,Rea, P.A. and Viitanen, P.V. (2002) Alternate energy-dependentpathways for the vacuolar uptake of glucose and glutathione con-jugates. Plant Physiol. 130, 1562–1572.
Bower, R., Elliott, A.R., Potier, B.A.M. and Birch, R.G. (1996) High-efficiency, microprojectile-mediated cotransformation of sugar-cane using visible or selectable markers. Mol. Breed. 2, 239–249.
Bradford, M. (1976) A rapid and sensitive method for the quantita-tion of microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal. Biochem. 72, 248–254.
Burrell, M.M., Mooney, P.J., Blundy, M., Carter, D., Wilson, F.,Green, J., Blundy, K.S. and ap Rees, T. (1994) Genetic manipulationof 6-phosphofructokinase in potato tubers. Planta, 194, 95–101.
Christensen, A.H. and Quail, P.H. (1996) Ubiquitin promoter-basedvectors for high-level expression of selectable and/or screenablemarker genes in monocotyledonous plants. Transgenic Res. 5,213–218.
Ciereszko, I., Johansson, H., Hurry, V. and Kleczkowski, L.A. (2001)Phosphate status affects the gene expression, protein content andenzymatic activity of UDP-glucose pyrophosphorylase in wild-typeand pho mutants of Arabidopsis. Planta, 212, 598–605.
Cornejo, M.-J., Luth, D., Blankenship, K.M., Anderson, O.D. andBlechl, A.E. (1993) Activity of a maize ubiquitin promoter intransgenic rice. Plant Mol. Biol. 23, 567–581.
Deguchi, M., Koshita, Y., Gao, M., Tao, R., Tetsumura, T., Yamaki,S. and Kanayama, Y. (2004) Engineered sorbitol accumulationinduces dwarfism in Japanese persimmon. J. Plant Physiol. 161,1177–1184.
Dennis, D.T. and Blakeley, S.D. (2000) Carbohydrate metabolism. InBiochemistry and Molecular Biology of Plants (Buchanan, B.B.,Gruissem, W. and Jones, R.L., eds), pp. 630–675. Rockville, MD:American Society of Plant Biologists.
Fan, L., Zheng, S. and Wang, X. (1997) Antisense suppression ofphospholipase Dα retards abscisic acid- and ethylene-promotedsenescence of postharvest Arabidopsis leaves. Plant Cell, 9, 2183–2196.
Franks, T. and Birch, R.G. (1991) Gene transfer into intact sugarcanecells using microprojectile bombardment. Aust. J. Plant Physiol.18, 471–480.
Gao, Z. and Loescher, W.H. (2000) NADPH supply and mannitolbiosynthesis. Characterization, cloning, and regulation of the non-reversible glyceraldehyde-3-phosphate dehydrogenase in celeryleaves. Plant Physiol. 124, 321–330.
Grof, C.P.L., Knight, D.P., McNeil, S.D., Lunn, J.E. and Campbell, J.A.(1998) A modified assay method shows leaf sucrose-phosphatesynthase activity is correlated with leaf sucrose content across arange of sugarcane varieties. Aust. J. Plant Physiol. 25, 499–502.
Heineke, D., Sonnewald, U., Bussis, D., Gunter, G., Leidreiter, K.,Wilke, I., Raschke, K., Willmitzer, L. and Heldt, H.W. (1992)Apoplastic expression of yeast-derived invertase in potato. Effectson photosynthesis, leaf solute composition, water relations, andtuber composition. Plant Physiol. 100, 301–308.
Huber, S.C. and Huber, J.L. (1996) Role and regulation of sucrose-phosphate synthase in higher plants. Annu. Rev. Plant Physiol.Plant Mol. Biol. 47, 431–444.
Hurry, V.M., Strand, A., Tobiaeson, M., Gardestrom, P. and Oquist,G. (1995) Cold hardening of spring and winter wheat and raperesults in differential effects on growth, carbon metabolism, andcarbohydrate content. Plant Physiol. 109, 697–706.
Iskandar, H.M., Simpson, R.S., Casu, R.E., Bonnett, G.D., Maclean,D.J. and Manners, J.M. (2004) Comparison of reference genes forquantitative real-time polymerase chain reaction analysis of geneexpression in sugarcane. Plant Mol. Rep. 22, 325–337.
Joshi, C.P., Zhou, H., Huang, X. and Chiang, V.L. (1997) Contextsequences of translation initiation codons in plants. Plant Mol.Biol. 35, 993–1001.
Joyce, P.A., McQualter, R.B., Handley, J.A., Dale, J.L., Harding, R.M.and Smith, G.R. (1998) Transgenic sugarcane resistant to sugarcanemosaic virus. Proc. Aust. Soc. Sugar Cane Technol. 20, 204–210.
Kanayama, Y., Mori, H., Imaseki, H. and Yamaki, S. (1992) Nucleo-tide sequence of a cDNA encoding NADP-sorbitol-6-phosphatedehydrogenase from apple. Plant Physiol. 100, 1607–1608.
Karakas, B., Ozias-Akins, P., Stushnoff, C., Suefferheld, M. and Rieger,M. (1997) Salinity and drought tolerance of mannitol-accumulatingtransgenic tobacco. Plant Cell Environ. 20, 609–616.
Kelly, G.J. and Gibbs, M. (1973) A mechanism for the indirecttransfer of photosynthetically reduced nicotinamide adeninedinucleotide phosphate from chloroplast to the cytoplasm. PlantPhysiol. 52, 674–676.
Kirschner, M. (2004) Chemical profile: sorbitol. Chem. Market Rep.266, 43.

Sorbitol synthesis in sugarcane 253
© Blackwell Publishing Ltd, Plant Biotechnology Journal (2007), 5, 240–253
Loescher, W.H. (1987) Physiology and metabolism of sugar alcoholsin higher plants. Physiol. Plantarum, 70, 553–557.
Magel, E.A., Hillinger, C., Wagner, T. and Holl, W. (2001) Oxidativepentose phosphate pathway and pyridine nucleotides in relation toheartwood formation in Robinia pseudoacacia L. Phytochemistry,57, 1061–1068.
Manjunath, S., Lee, C.-H.K., VanWinkle, P. and Bailey-Serres, J.(1998) Molecular and biochemical characterization of cytosolicphosphoglucomutase in maize. Expression during developmentand in response to oxygen deprivation. Plant Physiol. 117, 997–1006.
Marino, C.L., Leite, S.M.M., Farro, A.P.C., Sassaki, F.T., de Campos,H.L.V. and Coscrato, V.E. (2003) Putative metabolic pathway ofmannitol and sorbitol in sugarcane. Sci. Agric. 60, 723–728.
McLaughlin, R.L. (2005) Hyaluronic acid production in continuouscultures of Streptococcus zooepidemicus. PhD Thesis. The Univer-sity of Queensland, Brisbane.
McQualter, R.B., Fong Chong, B., Meyer, K., Van Dyk, D.E., O’Shea,M.G., Walton, N.J., Viitanen, P.V. and Brumbley, S.M. (2005)Initial evaluation of sugarcane as a production platform forp-hydroxybenzoic acid. Plant Biotechnol. J. 3, 29–41.
Ming, R., Moore, P.H., Wu, K.-K., D’Hont, A., Glaszmann, J.C., Tew,T.L., Mirkov, T.E., da Silva, J., Jifon, J., Rai, M., Schnell, R.J., Brum-bley, S.M., Lakshmanan, P., Comstock, J. and Paterson, A.H.(2006) Sugarcane improvement through breeding and biotech-nology. In Plant Breeding Reviews (Janick, J., ed.), 27, 18–118.New York: John Wiley.
Murashige, T. and Skoog, F. (1962) A revised medium for rapidgrowth and bioassays with tobacco tissue cultures. Physiol.Plantarum, 15, 473–497.
Renz, A., Merlo, L. and Stitt, M. (1993) Partial purification frompotato tubers of three fructokinases and three hexokinases whichshow differing organ and developmental specificity. Planta, 190,156–165.
Sacher, J.A., Hatch, M.D. and Glasziou, K.T. (1963) Sugaraccumulation cycle in sugarcane. III. Physical and metabolicaspects of cycle in immature storage tissues. Plant Physiol. 38,348–354.
von Schaewen, A., Stitt, M., Schmidt, R., Sonnewald, U. andWillmitzer, L. (1990) Expression of a yeast-derived invertase in thecell wall of tobacco and Arabidopsis plants leads to accumulationof carbohydrate and inhibition of photosynthesis and strongly
influences growth and phenotype of transgenic tobacco plants.EMBO J. 9, 3033–3044.
Scopes, R.K., Testolin, V., Stoter, A., Griffiths-Smith, K. and Algar,E.M. (1985) Simultaneous purification and characterization ofglucokinase, fructokinase and glucose-6-phosphate dehydro-genase from Zymomonas mobilis. Biochem. J. 228, 627–634.
Sevenier, R., Hall, R.D., van der Meer, I.M., Hakkert, H.J.C., vanTunen, A.J. and Koops, A.J. (1998) High level fructan accumulationin a transgenic sugar beet. Nat. Biotechnol. 16, 843–846.
Sheveleva, E.V., Marquez, S., Chmara, W., Zegeer, A., Jensen, R.G.and Bohnert, H.J. (1998) Sorbitol-6-phosphate dehydrogenaseexpression in transgenic tobacco. High amounts of sorbitol lead tonecrotic lesions. Plant Physiol. 117, 831–839.
Sonnewald, U., Brauer, M., von Schaewen, A., Stitt, M. andWillmitzer, L. (1991) Transgenic tobacco plants expressing yeast-derived invertase in either the cytosol, vacuole or apoplast: apowerful tool for studying sucrose metabolism and sink/sourceinteractions. Plant J. 1, 95–106.
Tao, R., Uratsu, S.L. and Dandekar, A.M. (1995) Sorbitol synthesis intransgenic tobacco with apple cDNA encoding NADP-dependentsorbitol-6-phosphate dehydrogenase. Plant Cell Physiol. 36, 525–532.
Wang, H.-L., Lee, P.-D., Chen, W.-L., Huang, D.-J. and Su, J.-C.(2000) Osmotic stress-induced changes of sucrose metabolism incultured sweet potato cells. J. Exp. Bot. 51, 1991–1999.
Wingler, A., von Schaewen, A., Leegood, R.C., Lea, P.J. and Quick,W.P. (1998) Regulation of leaf senescence by cytokinin, sugars,and light. Plant Physiol. 116, 329–335.
Wright, D.P., Huppe, H.C. and Turpin, D.H. (1997) In vivo and invitro studies of glucose-6-phosphate dehydrogenase from barleyroot plastids in relation to reductant supply for assimilation.Plant Physiol. 114, 1413–1419.
Xiao, W., Sheen, J. and Jang, J.-C. (2000) The role of hexokinase inplant sugar signal transduction and growth and development.Plant Mol. Biol. 44, 451–461.
Yamaguchi, H., Kanayama, Y. and Yamaki, S. (1994) Purificationand properties of NAD-dependent sorbitol dehydrogenase fromapple fruit. Plant Cell Physiol. 35, 887–892.
Zhu, Y.J., Komor, E. and Moore, P.H. (1997) Sucrose accumulationin the sugarcane stem is regulated by the difference between theactivities of soluble acid invertase and sucrose phosphate syn-thase. Plant Physiol. 115, 609–616.
NO2−




















