
Glutamate release correlates with brain-derived neurotrophic factor and trkB mRNA expression in the...
11
Ž . Molecular Brain Research 42 1996 317–327 Research report Glutamate release correlates with brain-derived neurotrophic factor and trk B mRNA expression in the CA1 region of rat hippocampus Torkel Falkenberg a,b , Nils Lindefors a, ) , Francesca Camilli b , Madis Metsis c , Urban Ungerstedt b a Department of Clinical Neuroscience, Psychiatry Section, Karolinska Institutet, P.O. Karolinska Hospital, S-171 76 Stockholm, Sweden b Department of Physiology and Pharmacology, Karolinska Institutet, P.O. Karolinska Hospital, S-171 76 Stockholm, Sweden c Department of Medical Biochemistry and Biophysics, Karolinska Institutet, P.O. Karolinska Hospital, S-171 76 Stockholm, Sweden Accepted 21 May 1996 Abstract Ž . Synthesis of the neurotrophic factor brain-derived neurotrophic factor BDNF and its receptor TrkB in the hippocampus have been proposed to be influenced by endogenous glutamate. To test this hypothesis we have investigated if increases in BDNF and trk B mRNAs are associated with changes in the synaptic release of glutamate in the dorsal hippocampus in the conscious rat by combining the technique of in vivo microdialysis with in situ hybridization histochemistry. A 35% and 66% increase in extracellular levels of glutamate in the dorsal CA1 region was detected following injection into the lateral entorhinal cortex of 2.4 and 9.6 mg of the non-NMDA glutamate receptor agonist quisqualate, respectively. The increase in glutamate was attenuated by local administration of tetrodotoxin Ž . TTX indicating neuronal origin. Levels of BDNF and trk B mRNAs were increased in the hippocampus in a dose-dependent fashion following the stimulations. The extracellular levels of glutamate in individual animals correlated to the levels of BDNF and trk B mRNAs in the dorsal CA1 region of the hippocampus. This study provides for the first time evidence of an entorhinal cortex influenced concentration-dependent relationship between the release of endogenous glutamate in vivo and neuronal expression of mRNAs for BDNF and its receptor trk B in the hippocampus. Keywords: Brain-derived neurotrophic factor; TrkB; Glutamate; Quisqualate; Entorhinal cortex; Hippocampus 1. Introduction Ž . Brain-derived neurotrophic factor BDNF , a member of a structurally related protein family including nerve Ž growth factor, neurotrophin-3 and neurotrophin-4r5 for w x. review see 32,42 is expressed in high levels in the w x hippocampus 15,22,44 . Neurotrophins exert their effects by binding to transmembrane receptors containing tyrosine kinase domains. These receptors mediate cell growth and w x differentiation 8,23 . BDNF binds to the tyrosine kinase receptor trk B, a member of the trk gene family of tyrosine wx kinase transmembrane receptors 5 . The trk B gene en- Ž trk B . codes a 145-kDa glycoprotein gp145 which consti- tutes an essential component of the high-affinity receptor w x for BDNF 26,49,52 and is expressed in the hippocampus w x 26 . Transcription of the trk B gene may generate multiple ) Ž. Corresponding author. Fax: q46 8 32-6209; E-mail: [email protected] transcripts encoding the functional gp145 trk B receptor with an intact tyrosine kinase domain or C-terminally truncated Ž trk B . forms of the receptor gp95 which lack intracellular w x kinase domains 27,38 . Glutamate is the major excitatory amino acid transmit- w x ter utilized in the brain 18 . Glutamate activity in the brain is conveyed by ionotropic receptors, the NMDA and non- Ž . NMDA kainate and quisqualaterAMPA subtype recep- w x tors 58 . Additionally, a metabotropic glutamate receptor has been identified, which is activated by glutamate-in- w x duced membrane phosphoinositide turn-over 10 . BDNF mRNA levels in hippocampal neurons have been suggested to be influenced by excitatory glutamatergic w x w x 4,24,63,64 and inhibitory GABAergic 64 neurotransmis- sion as determined by systemic or intrahippocampal injec- tions of glutamate or GABA analogues. However, sys- temic administration of transmitter analogues affects many regions of the brain unspecifically. In this study we have applied local injection of a glutamate receptor agonist to 0169-328Xr96r$15.00 Copyright q 1996 Elsevier Science B.V. All rights reserved. Ž . PII S0169-328X 96 00134-9
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Glutamate release correlates with brain-derived neurotrophic factor and trkB mRNA expression in the...

Ž .Molecular Brain Research 42 1996 317–327
Research report
Glutamate release correlates with brain-derived neurotrophic factor andtrkB mRNA expression in the CA1 region of rat hippocampus
Torkel Falkenberg a,b, Nils Lindefors a,), Francesca Camilli b, Madis Metsis c,Urban Ungerstedt b
a Department of Clinical Neuroscience, Psychiatry Section, Karolinska Institutet, P.O. Karolinska Hospital, S-171 76 Stockholm, Swedenb Department of Physiology and Pharmacology, Karolinska Institutet, P.O. Karolinska Hospital, S-171 76 Stockholm, Sweden
c Department of Medical Biochemistry and Biophysics, Karolinska Institutet, P.O. Karolinska Hospital, S-171 76 Stockholm, Sweden
Accepted 21 May 1996
Abstract
Ž .Synthesis of the neurotrophic factor brain-derived neurotrophic factor BDNF and its receptor TrkB in the hippocampus have beenproposed to be influenced by endogenous glutamate. To test this hypothesis we have investigated if increases in BDNF and trkB mRNAsare associated with changes in the synaptic release of glutamate in the dorsal hippocampus in the conscious rat by combining thetechnique of in vivo microdialysis with in situ hybridization histochemistry. A 35% and 66% increase in extracellular levels of glutamatein the dorsal CA1 region was detected following injection into the lateral entorhinal cortex of 2.4 and 9.6 mg of the non-NMDAglutamate receptor agonist quisqualate, respectively. The increase in glutamate was attenuated by local administration of tetrodotoxinŽ .TTX indicating neuronal origin. Levels of BDNF and trkB mRNAs were increased in the hippocampus in a dose-dependent fashionfollowing the stimulations. The extracellular levels of glutamate in individual animals correlated to the levels of BDNF and trkB mRNAsin the dorsal CA1 region of the hippocampus. This study provides for the first time evidence of an entorhinal cortex influencedconcentration-dependent relationship between the release of endogenous glutamate in vivo and neuronal expression of mRNAs for BDNFand its receptor trkB in the hippocampus.
Keywords: Brain-derived neurotrophic factor; TrkB; Glutamate; Quisqualate; Entorhinal cortex; Hippocampus
1. Introduction
Ž .Brain-derived neurotrophic factor BDNF , a memberof a structurally related protein family including nerve
Žgrowth factor, neurotrophin-3 and neurotrophin-4r5 forw x.review see 32,42 is expressed in high levels in thew xhippocampus 15,22,44 . Neurotrophins exert their effects
by binding to transmembrane receptors containing tyrosinekinase domains. These receptors mediate cell growth and
w xdifferentiation 8,23 . BDNF binds to the tyrosine kinasereceptor trkB, a member of the trk gene family of tyrosine
w xkinase transmembrane receptors 5 . The trkB gene en-Ž t r k B.codes a 145-kDa glycoprotein gp145 which consti-
tutes an essential component of the high-affinity receptorw xfor BDNF 26,49,52 and is expressed in the hippocampus
w x26 . Transcription of the trkB gene may generate multiple
) Ž .Corresponding author. Fax: q46 8 32-6209; E-mail:[email protected]
transcripts encoding the functional gp145t r k B receptor withan intact tyrosine kinase domain or C-terminally truncated
Ž t r k B.forms of the receptor gp95 which lack intracellularw xkinase domains 27,38 .
Glutamate is the major excitatory amino acid transmit-w xter utilized in the brain 18 . Glutamate activity in the brain
is conveyed by ionotropic receptors, the NMDA and non-Ž .NMDA kainate and quisqualaterAMPA subtype recep-
w xtors 58 . Additionally, a metabotropic glutamate receptorhas been identified, which is activated by glutamate-in-
w xduced membrane phosphoinositide turn-over 10 .BDNF mRNA levels in hippocampal neurons have been
suggested to be influenced by excitatory glutamatergicw x w x4,24,63,64 and inhibitory GABAergic 64 neurotransmis-sion as determined by systemic or intrahippocampal injec-tions of glutamate or GABA analogues. However, sys-temic administration of transmitter analogues affects manyregions of the brain unspecifically. In this study we haveapplied local injection of a glutamate receptor agonist to
0169-328Xr96r$15.00 Copyright q 1996 Elsevier Science B.V. All rights reserved.Ž .PII S0169-328X 96 00134-9

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327318
allow for better opportunities to specifically activate affer-ent neuronal pathways to the hippocampus. The lateral
Ž .entorhinal area LEA is a part of the entorhinal cortex thatconstitutes a major input for the hippocampus via the
w xperforant pathway 48,57,61 . We have previously stimu-lated glutamate receptors in the LEA by local injection of
w x2.4 mg quisqualate 16,31 and demonstrated that BDNFand trkB mRNA levels in the dorsal hippocampus weremarkedly increased following this stimulation. We hypoth-esized that the increased hippocampal BDNF mRNA ex-pression was due to an elevated release of glutamate in thiscircuitry since stimulation of non-NMDA glutamate recep-tors in cell cultures of hippocampal neurons increase levels
w xof BDNF mRNA 63 . To test this hypothesis our principalaim in this study was to investigate if extracellular gluta-mate levels could be correlated to the cellular expressionof mRNAs encoding BDNF and its receptor trkB in thedorsal CA1 region of the hippocampus. To investigate thisa microdialysis cannula was chronically implanted in thedorsal hippocampal CA1 region and a guide cannula wasimplanted in the lateral part of LEA of the rat. Thisexperimental setup allowed us to stimulate cell bodies inLEA by local injection of increasing concentrations ofquisqualate and simultaneously measure changes in extra-cellular levels of glutamate in the dorsal CA1 region.Subsequently, levels of mRNAs encoding BDNF and thereceptor trkB were determined in each animal by in situhybridization histochemistry. In addition, behaviour wasmonitored following glutamate receptor stimulation in theentorhinal cortex to evaluate the functional consequencesof activation of the LEA-hippocampal circuitry.
Our results suggest that BDNF and trkB mRNA ex-pression in vivo in the dorsal hippocampal CA1 region isdirectly influenced by glutamate release regulated by affer-ents of LEA.
2. Materials and methods
2.1. Animals and in ÕiÕo microdialysis
ŽMale Sprague–Dawley rats mean b.wt. 300 g; Alab.AB, Sollentuna, Sweden were maintained on a standard
12r12 light–dark cycle and allowed food and water adlibitum. On the day of surgery, rats were anaesthetizedŽ .1.5% halothanerair mixture delivered at 1.4 lrmin and
Žthe dura was exposed through two burr holes 3 mm in.diameter in the skull. A microdialysis cannula of concen-
Žtric design CMA 12, CMA Mikrodialys AB, Stockholm,.Sweden outer diameter 0.5 mm with a 1-mm dialysing
membrane at the tip was permanently implanted in the leftdorsal CA1 region in the hippocampus at the stereotaxiccoordinates from bregma: APsy3.8, Ls2.2, Dsy3.2w x Ž . Ž41 Fig. 1 . A guide cannula CMA Mikrodialys AB,
.Stockholm, Sweden outer diameter 0.7 mm was implantedpermanently in the left LEA 1 mm dorsal from the injec-
Fig. 1. Schematic drawing of the experimental setup showing the anatom-ical location of the chronically implanted microdialysis cannula in thedorsal CA1 region and the modified microdialysis cannula serving as aninjection cannula in the lateral part of LEA.
tion site located at stereotaxic coordinates from bregma:w x Ž .APsy6.3, Ls6.6, Dsy8.0 41 Fig. 1 . The micro-
dialysis cannula and the guide cannula were permanentlyfixed to the skull with stainless steel screws and meth-ylacrylic cement. A new microdialysis cannula was used ineach rat. Local anaesthetic was applied onto the skull areaadjacent to the cement to minimise postoperative pain. Allexperiments were performed in awake freely moving ratson the second day following cannula implantations.
On the day of the experiment, the rat was placed in ahemispheric bowl and the implanted microdialysis cannulawas connected via a liquid swivel to a microinfusion pumpŽCMA 120 system 4, CMA Mikrodialys AB, Stockholm,
.Sweden . The microdialysis cannula was perfused at aflow rate of 2 mlrmin with an artificial cerebrospinal fluidŽ . q qACSF containing 155 mM Na , 2.9 mM K , 1.1 mMCa2q, 0.83 mM Mg2q, 132.8 mM Cly, 0.5 mM PO2y, 0.54
mM SO2y and adjusted to a pH of 7.3 by gassing with4w x95%:5% O :CO . TTX, a sodium channel antagonist 402 2
Ž .Janssen Pharmaceutica, Belgium was dissolved in waterŽ .and stored in 1.0 mM aliquots stock solution at y708C
for up to 6 months. Whenever TTX was used stocksolution was diluted in ACSF to a final concentration of 10mM and perfusion with TTX started at the beginning ofthe experiment. Dialysates were collected every 20 minand started immediately following connection of the mi-crodialysis cannula to the perfusion apparatus and contin-ued for 6 h. The relative level of glutamate in eachdialysate following ACSF or quisqualate injection wasdefined as the ratio between glutamate concentration ineach dialysate following injection and the mean glutamateconcentration in three subsequent samples immediatelypreceding the insertion of the injection cannula. The high-

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327 319
est relative level of glutamate found in any of the dialysatescollected 3 h following injection of ACSF or quisqualatewas defined as the maximum extracellular level of gluta-mate.
2.2. Chemical stimulation
A modified microdialysis cannula without membranefunctioned as an injection cannula with a tip diameter less
Ž .than 0.1 mm and coupled to an Hamilton syringe 2 ml .Following dialysis for approximately 140 min when extra-cellular levels of glutamate were stable the injection can-nula was inserted through the guide cannula into the
Ž .superficial layers of the lateral part of LEA Fig. 1 .Following another 20-min perfusion period, 0.5 ml of
Ž . ŽACSF control or quisqualate Research Biomedical Inter-. Ž .national, Massachusetts, USA either 0.24, 2.4 or 9.6 mg
in ACSF was injected slowly over a period of 3 min. TheHamilton syringe was disconnected 10 min after injectionwhereas the injection cannula remained in locationthroughout the whole experiment terminating 3 h afterinjection. Subsequently, the animal was anaesthetized withhigh concentration of halothane and killed with an over-
Ždose of Mebumal intracardially 120 mg, Nord Vacc,.Sweden and the brain was frozen at y208C.
2.3. BehaÕioural eÕaluation
The animals were visually observed during the experi-ment. The following behavioural rating scale modified
w xfrom Sperk et al. 50 was used for determination ofbehaviour: 1, normal behaviour, occasional locomotion; 2,increased locomotor and rearing activity, occasionally ‘wetdog shakes’; 3, increased rate of ‘wet dog shakes’, tailerection; 4, rare and mild repetitive movements affectingforelimbs and head, indicating seizure-like activity; 5, mildto severe intermittent repetitive movements affecting thewhole body with rearing, falling over, foam coming fromthe mouth; 6, prolonged, generalized, seizure-like activitywith the same symptoms as in 5; 7, generalized seizureswith early death. Animal behaviour was rated every 20min throughout the length of the experiment.
2.4. Biochemical analysis
Glutamate concentration was determined by HPLC us-in g p re c o lu m n d e r iv a t iz a t io n w ith o -
Ž .phthaldialdehydermercaptoethanol OPA reagent fol-lowed by fluorometric detection as described by Meana et
w x Žal. 36 . Briefly, 10 ml of the OPA reagent 0.4 M borate,0.04 M ophthaldialdehyde, 0.4 M 2-mercaptoethanol, pH
.10.4 was added to 10 ml of perfusate and following 60 sreaction time, 15 ml of the derivatized aliquot was injected
Ž .into a column 100=3.2 mm prepacked with 5 mmŽNucleosil-100 C-18 packing material Knauer, Berlin, Ger-
. Žmany . Derivatization was made in an autosampler CMA
.200, CMA Microdialys AB, Stockholm, Sweden at 48C.The eluent was a mixture of 0.1 M sodium acetate, 9%methanol adjusted to pH 6.95 and 1.5% tetrahydrofurane.The eluent was pumped via an online CMA 260 degasserŽ .CMA 260, CMA Microdialys AB, Stockholm, Sweden ,
Žby a Spectra Physics 8800-020 precision pump Spectra-.Physics, San Jose, CA, USA equipped with a two-way
valve to the autosampler, at a flow of 1 mlrmin. AŽmultilinear gradient 100% eluent changing to 100%
methanol over 6 min and returning to 100% eluent 2 min.later was used. The fluorometric detector was an Hitatchi
ŽF1000 fluorescence spectrophotometer Hitachi, Tokyo,.Japan with excitation wavelength set at 370 nm and
emission cut off filter set at 450 nm. An SP 4220Ž .Spectra-Physics, San Jose, CA, USA integrator was usedto determine concentration of glutamate following calibra-tion of the integrator using a 1.875 mM glutamate stan-dard. The limit of detection was 5 pmolrsample. Thesample volume used in this experiment was 10 ml.
2.5. Tissue preparation
Ž .Tissue sections of hippocampus 12 mm were cut usingŽ .a cryostat Leitz, Wetzlar, West Germany and mounted
onto silanised slides. The location of the microdialysiscannula in the dorsal CA1 region and the injection cannulain the lateral part of LEA was verified during cryostatcutting. Animals showing ectopic cannula positions wereexcluded from the study.
2.6. In situ hybridization and image analysis
The in situ hybridization protocol was modified fromw xBunnemann et al. 6 . Coronal tissue sections were fixed in
4% paraformaldehyde for 15 min, rinsed once in phosphatebuffered saline pH 7.5 and twice in distilled water. Thesections were subsequently treated with 0.1 M HCl for 10min and acetylated for 20 min with 0.25% acetic anhydridein 0.1 M ethanolamine, followed by rinsing two times inphosphate buffered saline pH 7.5 and dehydration withethanol and air dried. The sections were hybridizedovernight in a humidified chamber at 428C with 75
Žmlrsection of hybridization buffer 50% formamide, 20mM Tris-HCl pH 7.6, 1 mM EDTA pH 8.0, 0.3 M NaCl,0.1 M dithiotreitol, 0.5 mgrml yeast tRNA, 0.1 mgrml
Ž .poly-A-RNA Sigma, St. Louis, MO , 1=Denhart’s solu-. 6tion, 10% dextrane sulphate containing 2.5 = 10
w35 xc.p.m.rml of the S UTP-labelled cRNA probe. Probeswere synthesized from cDNA fragments cloned into pB-SKS vector and linearized with appropriate restrictionenzymes. The following parts of cDNA were used: BDNF,fragment covering amino acids 75–155 in preproBDNF
w x t r k B Žprotein sequence 33 ; gp95qgp145 probe recogniz-Ž t r k B. Ž t r k B.ing both truncated gp95 and full-length gp145
.receptor mRNAs , spanning bp 1133–1458 in trkB cDNAw x t r k B Žsequence 38 ; gp145 probe that recognizes kinase
( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327320
domain of trkB and thus the full-length gp145t r k B mRNA.only , spanning bp 1647–2057 in trkB cDNA sequence
w x38 . The specificity of all used cRNA probes have beenw xexamined with RNAse protection assay 20 . After hy-
bridization the sections were washed once in 1=SSC atŽ .378C for 40 min, treated with RNase 10 mgrml in 0.5 M
NaCl, 20 mM Tris–HCl pH 7.5, 2 mM EDTA at 378C for30 min followed by three 10-min washes with 0.5=SSCat 488C, 0.2=SSC at 608C and 0.1=SSC at 608C. Theslides were then dehydrated with ethanol and exposed to
Ž .b-max X-ray film Amersham, Buckinghamshire, UK for21 days. The autoradiograms from the in situ hybridiza-tions were analysed with Microcomputer Imaging DeviceŽ .Imaging Research, Canada . Optical density in eachanatomical region was determined over the whole regionin at least two subsequent sections from each animal.Optical density values were converted to relative levels ofradioactivity using external standards exposed to the samefilm. Optical density values were measured in an intervalwith linear correlation to tissue levels of radioactive iso-tope.
BDNF and trkB mRNA levels were not determined inanimals locally perfused with TTX. This was due tomethodological difficulties in reproducibly determiningmRNA levels in tissue adjacent to the lesion produced bythe microdialysis cannula implantation and to uncertaintyof the diffusion volume of TTX in tissue.
2.7. Statistics
Data are presented as mean"S.E.M. except for be-havioural ratings which are presented as median"minrmax. Statistical analyses of levels of glutamate in theipsilateral dorsal CA1 region due to treatment and levels ofBDNF, gp95qgp145t r k B and gp145t r k B mRNAs in thehippocampus and in each region due to treatment wereperformed using ANOVA. Correlation analysis usedANOVA and a simple model regression analysis. Be-havioural ratings were statistically evaluated usingKruskal–Wallis test.
3. Results
3.1. Extracellular leÕels of glutamate in CA1 after injec-tion with increasing concentrations of quisqualate intoLEA
The different treatment groups did not show any signifi-Žcant difference in basal glutamate concentration the mean
of three 20-min dialysates preceding insertion of the injec-. Ž .tion cannula into LEA of glutamate Table 1 . The inser-
tion of the injection cannula and injection of ACSF intoLEA did not produce any significant change in the extra-cellular levels of glutamate in the ipsilateral dorsal CA1
Žregion compared to basal extracellular levels data not
Table 1Ž .Basal concentration of glutamate mean"S.E.M. in perfusates obtained
with a microdialysis cannula implanted into the dorsal left hippocampalCA1 region
Ž .Group Glutamate mM
Ž .All rats 1.08"0.26 ns20Ž .ACSF 0.96"0.23 ns4Ž .0.24 mg 0.35"0.07 ns4Ž .2.4 mg 1.05"0.46 ns5Ž .9.6 mg 0.94"0.64 ns3Ž .9.6 mgqTTX 10 mM 1.67"0.30 ns4
Ž .No difference between groups was found P)0.05, ANOVA in respec-tive concentration of glutamate.
.shown . Extracellular levels of glutamate were stablethroughout the collection period after the injection of
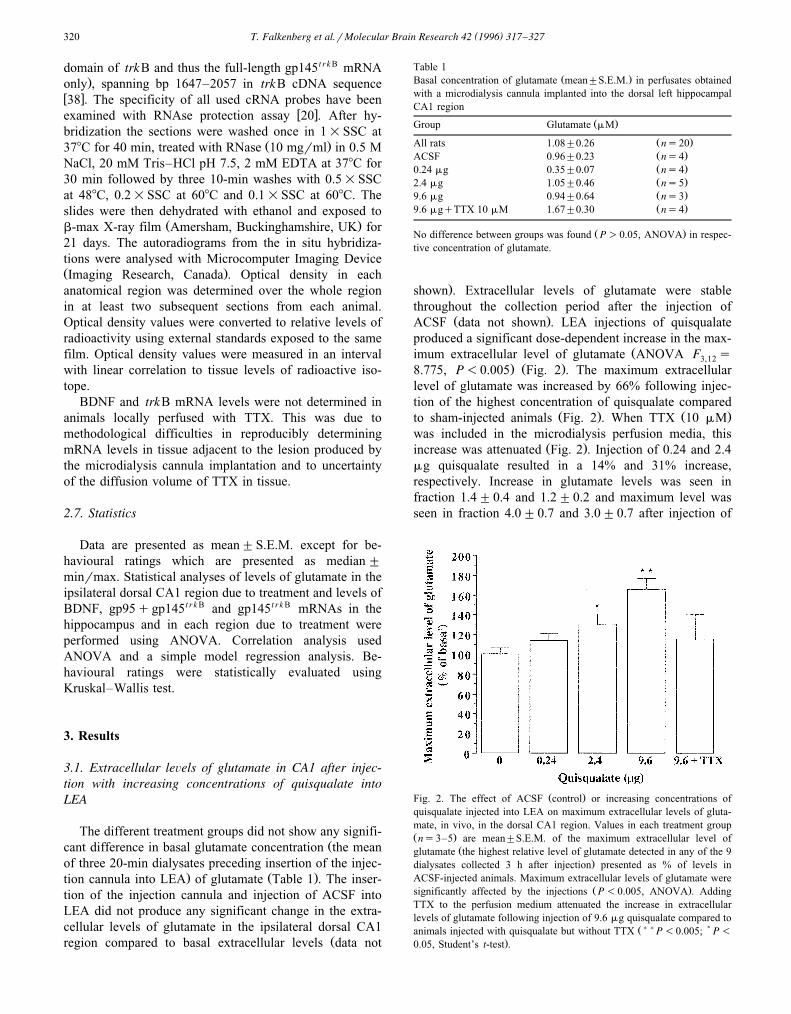
Ž .ACSF data not shown . LEA injections of quisqualateproduced a significant dose-dependent increase in the max-
Žimum extracellular level of glutamate ANOVA F s3,12. Ž .8.775, P-0.005 Fig. 2 . The maximum extracellular
level of glutamate was increased by 66% following injec-tion of the highest concentration of quisqualate compared
Ž . Ž .to sham-injected animals Fig. 2 . When TTX 10 mMwas included in the microdialysis perfusion media, this
Ž .increase was attenuated Fig. 2 . Injection of 0.24 and 2.4mg quisqualate resulted in a 14% and 31% increase,respectively. Increase in glutamate levels was seen infraction 1.4"0.4 and 1.2"0.2 and maximum level wasseen in fraction 4.0"0.7 and 3.0"0.7 after injection of
Ž .Fig. 2. The effect of ACSF control or increasing concentrations ofquisqualate injected into LEA on maximum extracellular levels of gluta-mate, in vivo, in the dorsal CA1 region. Values in each treatment groupŽ .ns3–5 are mean"S.E.M. of the maximum extracellular level of
Žglutamate the highest relative level of glutamate detected in any of the 9.dialysates collected 3 h after injection presented as % of levels in
ACSF-injected animals. Maximum extracellular levels of glutamate wereŽ .significantly affected by the injections P-0.005, ANOVA . Adding
TTX to the perfusion medium attenuated the increase in extracellularlevels of glutamate following injection of 9.6 mg quisqualate compared to
Ž)) )animals injected with quisqualate but without TTX P-0.005; P-.0.05, Student’s t-test .

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327 321
2.4 and 9.6 mg quisqualate respectively. Levels did notcompletely return to control levels derived from ACSF-in-
Žjected animals within the 3-h observation period data not.shown . Total glutamate overflow, collapsed over 3 h
following injection with quisqualate, was similarly dose-dependently affected by the stimulations compared to lev-
Žels in ACSF-injected animals ANOVA F s3.770, P3,11. Ž .-0.05 data not shown .
3.2. BDNF mRNA expression in the hippocampus afterinjection with increasing concentrations of quisqualateinto LEA
BDNF mRNA expressing cells were seen in the granulelayer of the dentate gyrus and throughout the pyramidal
Ž .layer of the hippocampus Fig. 3A . Increasing concentra-tions of quisqualate produced significant increases in BDNF
mRNA levels in all regions of the hippocampus 3 hŽ .subsequent to injection panels A in Figs. 3–5 . Marked
effects were detected in the dentate gyrus where the high-est concentration of quisqualate resulted in a 790% to870% bilateral increase in BDNF mRNA compared tolevels in sham-injected animals. Injection of 2.4 mgquisqualate resulted in a 580% to 630% bilateral increaseand injection of 0.24 mg quisqualate resulted in a 43% and100% increase in the ipsi- and contralateral dentate gyrus,respectively. The effect was less marked in the pyramidalcells compared to the increase obtained in the dentategyrus. BDNF mRNA levels in the ipsilateral CA1 regionincreased with 146% and 212% following injections with2.4 and 9.6 mg quisqualate, respectively. BDNF mRNAlevels in the contralateral CA1 region increased with 93%and 290% following injections with 2.4 and 9.6 mgquisqualate, respectively. In the ipsilateral CA3 region
Ž . t r k B ŽFig. 3. The effect of injections of ACSF or 9.6 mg quisqualate into LEA on cellular levels of mRNAs encoding BDNF A, top panel , gp95q145 B,. t r k B Ž . Ž .middle panel or gp145 C, lower panel in the different ipsilateral hippocampal regions. Representative high magnification =500 bright-field
photomicrographs of in situ hybridization for the different mRNAs in the hippocampus. DG, dentate gyrus.

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327322
injections of 2.4 and 9.6 mg quisqualate resulted in a 86%and 103% increase, respectively whereas a 64% and 130%increase was detected contralaterally. No elevation ofBDNF mRNA levels in the pyramidal cells was detectablefollowing administration of the lowest concentration ofquisqualate.
3.3. TrkB mRNA expression in the hippocampus afterinjection with increasing concentrations of quisqualateinto LEA
In the dorsal hippocampus of control animals, the probedetecting mRNAs encoding gp95t r k B- or gp145t r k B-labelledcells in the granule layer of the dentate gyrus and in the
Ž .pyramidal cell layer Fig. 3B . LEA injections ofquisqualate produced significant increases in mRNAs en-coding both functional and truncated forms of the receptor
Ž .in all regions of the hippocampus panels B in Figs. 3–5 .Injection of 2.4 or 9.6 mg quisqualate resulted in a 105%to 125% increase in the dentate gyrus. The lowest concen-tration produced minor increases bilaterally in the dentategyrus. Minor effects, although significant, were seen ingp95q145t r k B mRNA labelling in the pyramidal cellsbilaterally following quisqualate injections.
Labelling in control animals was observed in the gran-ule cell layer of the dentate gyrus and in the pyramidal celllayer using the probe detecting mRNAs encoding the full
t r k B Ž .length receptor gp145 Fig. 3C . The ipsilateral dentategyrus and CA3 were significantly affected by the stimula-
Ž .tions panels C in Figs. 3–5 . Following injections of 2.4or 9.6 mg quisqualate a roughly 80% increase was seen inthe dentate gyrus. The lowest concentration produced mi-nor effects. Levels of gp145t r k B mRNA in the ipsilateralCA3 region were increased with 38% and 25% following
injections with 2.4 and 9.6 mg quisqualate, respectively.No effect was detected using the lowest concentration.
3.4. Extracellular leÕels of glutamate in relation to BDNFand trkB mRNA expression in the dorsal CA1 regionfollowing LEA injections
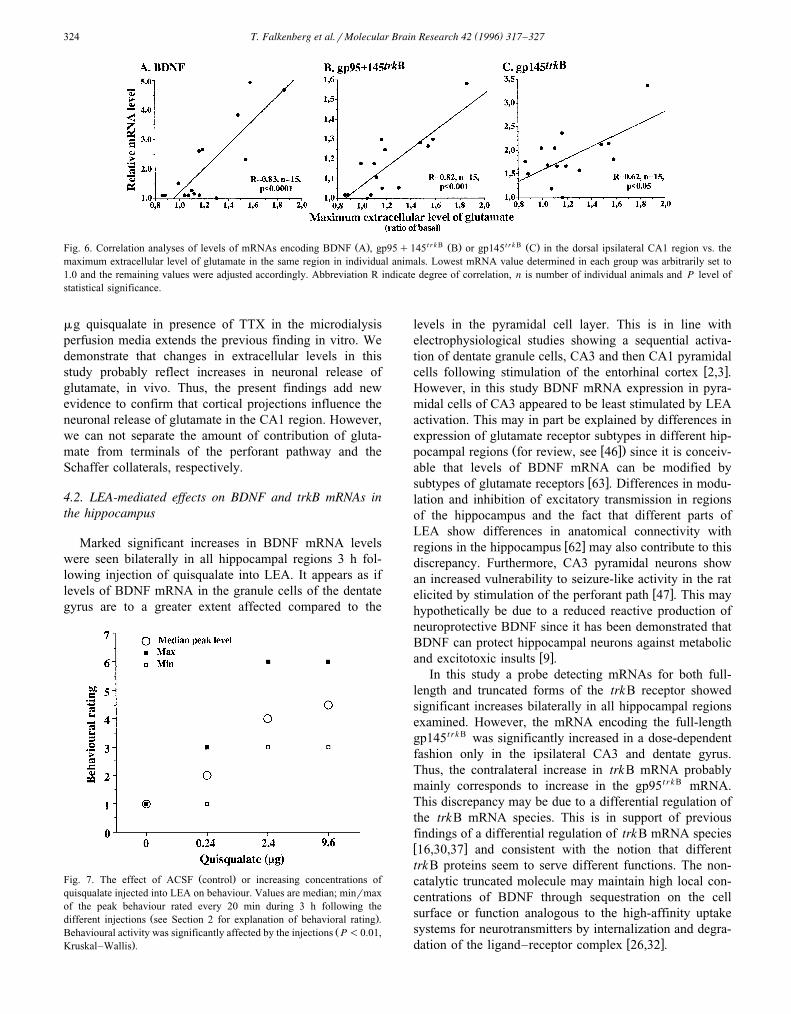
Statistical analysis of glutamate and BDNF data ob-tained from animals receiving injections showed that themaximum extracellular levels of glutamate measured in thedorsal ipsilateral CA1 region correlated significantly withthe relative levels of BDNF mRNA in the same regionŽ .Fig. 6A . Similarly, maximum extracellular levels of glu-tamate correlated significantly with relative levels of gp95
t r k B Ž .q145 mRNA levels Fig. 6B . A weaker but signifi-cant correlation was obtained for gp145t r k B mRNA levelsŽ .Fig. 6C . Accumulated extracellular glutamate measuredover the total 3-h period in the different animal groupswere also significantly correlated with BDNF, gp95q
t r k B t r k B Ž145 and gp145 mRNA, respectively data not.shown .
3.5. BehaÕioural effects of the quisqualate injections intoLEA
In general, the rats were active for approximately 30min following the exposure to the novel experimentalenvironment, thereafter the control rats alternated betweensleep and moderate locomotor activity. No change in be-haviour was observed in the control group of animalsfollowing ACSF injections into LEA. However, there was
Ž .a significant effect on behaviour P-0.01 followingŽ .quisqualate injections Fig. 7 . The 0.24 mg quisqualate
produced an increase in behaviour after injection in two
Ž . t r k BFig. 4. The effect of injections of ACSF or increasing concentrations of quisqualate into LEA on levels of mRNAs encoding BDNF A , gp95q145Ž . t r k B Ž . Ž .B or gp145 C in the dorsal hippocampus 3 h after the injections. Representative low magnification =10 autoradiograms of in situ hybridizationfor the different mRNAs in the hippocampus.

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327 323
animals out of four observed during 20 and 60 min,respectively, appearing as rearing, olfaction and digging.Quisqualate at 2.4 or 9.6 mg resulted in prolonged changesin behavioural activity which was usually seen 30 minfollowing injection of quisqualate with a decline in excita-tion during the last hour of the experiment. Injection of 2.4mg quisqualate produced initially increased locomotor and
Ž . t r k BFig. 5. Relative levels of mRNAs encoding BDNF A , gp95q145Ž . t r k B Ž .B or gp145 C in the different treatment groups. The autoradio-grams from the in situ hybridizations were quantified by computerassisted image analysis. Values are mean"S.E.M. and presented as % ofrelative levels in respective contralateral region in ACSF-injected ani-mals. )P-0.05, ANOVA. DG, dentate gyrus. Bar labelling is explainedin panel A and brain region reference below x-axis in panel C also applyfor panels A and B.
rearing activity, intermittent ‘wet dog shakes’ and tailerection. This stage evolved, usually within 20 min, to a
Ž .long-lasting period 60–120 min with repetitive move-ments affecting forelimbs and head, indicating seizure-likeactivity. Occasionally, circling behaviour either ipsi- orcontralaterally to the injection was observed during declinein seizure behaviour. Injection of 9.6 mg quisqualate pro-duced similar onset and time course, however animalstended to be more affected by the injection and circlingbehaviour was more frequent.
4. Discussion
By combining the techniques of in vivo microdialysisand in situ hybridization we show that stimulation of LEAincreases extracellular levels of glutamate and cellularlevels of mRNAs encoding BDNF and its receptor trkB inthe hippocampus accompanied with changes in behaviour.We also show that glutamate levels can be positivelycorrelated with BDNF and trkB mRNA levels in the CA1region.
4.1. LEA-mediated effects on glutamate release in thehippocampus
Predominantly granule cells but also pyramidal cells inthe dorsal hippocampus receive excitatory afferents fromsuperficial layers of LEA via the perforant pathw x55,59,60,62 . In the present study the excitatory stimulusin the unilateral LEA was found to dose-dependentlyincrease extracellular levels of glutamate in the dorsalipsilateral CA1 region of the hippocampus. This finding issupported by demonstrations showing that electrical kin-dling of the amygdala results in elevated glutamate levels
w xin the lateral ventricle, ipsilaterally to the stimulation 43 .Furthermore, more recent findings show a sustained in-crease in extracellular levels of glutamate in the epilepto-
w xgenic hippocampus in humans 7,13 . Our neurochemicalevidence of a dose-dependent increase in maximum extra-cellular levels of glutamate, in vivo, following stimulation
w xof LEA supports and extends anatomical 62 and electro-w xphysiological 2,3,25 findings of an excitatory influence
on CA1 neurons by the perforant pathway and Schaffercollaterals.
By using a co-culture system consisting of input axonsfrom entorhinal cortex explants and target hippocampalpyramidal cells it has been shown that glutamate can be
2q w xreleased in an activity- and Ca -dependent manner 35 .TTX perfused locally via the microdialysis membraneappears to reach neuron terminals close to the membraneand to have an effect on the extracellular environment due
w xto an effect on neurotransmitter release 12 . Our finding ofan attenuation of the 66% increase in the maximum extra-cellular level of glutamate following stimulation with 9.6
( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327324
Ž . t r k B Ž . t r k B Ž .Fig. 6. Correlation analyses of levels of mRNAs encoding BDNF A , gp95q145 B or gp145 C in the dorsal ipsilateral CA1 region vs. themaximum extracellular level of glutamate in the same region in individual animals. Lowest mRNA value determined in each group was arbitrarily set to1.0 and the remaining values were adjusted accordingly. Abbreviation R indicate degree of correlation, n is number of individual animals and P level ofstatistical significance.
mg quisqualate in presence of TTX in the microdialysisperfusion media extends the previous finding in vitro. Wedemonstrate that changes in extracellular levels in thisstudy probably reflect increases in neuronal release ofglutamate, in vivo. Thus, the present findings add newevidence to confirm that cortical projections influence theneuronal release of glutamate in the CA1 region. However,we can not separate the amount of contribution of gluta-mate from terminals of the perforant pathway and theSchaffer collaterals, respectively.
4.2. LEA-mediated effects on BDNF and trkB mRNAs inthe hippocampus
Marked significant increases in BDNF mRNA levelswere seen bilaterally in all hippocampal regions 3 h fol-lowing injection of quisqualate into LEA. It appears as iflevels of BDNF mRNA in the granule cells of the dentategyrus are to a greater extent affected compared to the
Ž .Fig. 7. The effect of ACSF control or increasing concentrations ofquisqualate injected into LEA on behaviour. Values are median; minrmaxof the peak behaviour rated every 20 min during 3 h following the
Ž .different injections see Section 2 for explanation of behavioral rating .ŽBehavioural activity was significantly affected by the injections P-0.01,
.Kruskal–Wallis .
levels in the pyramidal cell layer. This is in line withelectrophysiological studies showing a sequential activa-tion of dentate granule cells, CA3 and then CA1 pyramidal
w xcells following stimulation of the entorhinal cortex 2,3 .However, in this study BDNF mRNA expression in pyra-midal cells of CA3 appeared to be least stimulated by LEAactivation. This may in part be explained by differences inexpression of glutamate receptor subtypes in different hip-
Ž w x.pocampal regions for review, see 46 since it is conceiv-able that levels of BDNF mRNA can be modified by
w xsubtypes of glutamate receptors 63 . Differences in modu-lation and inhibition of excitatory transmission in regionsof the hippocampus and the fact that different parts ofLEA show differences in anatomical connectivity with
w xregions in the hippocampus 62 may also contribute to thisdiscrepancy. Furthermore, CA3 pyramidal neurons showan increased vulnerability to seizure-like activity in the rat
w xelicited by stimulation of the perforant path 47 . This mayhypothetically be due to a reduced reactive production ofneuroprotective BDNF since it has been demonstrated thatBDNF can protect hippocampal neurons against metabolic
w xand excitotoxic insults 9 .In this study a probe detecting mRNAs for both full-
length and truncated forms of the trkB receptor showedsignificant increases bilaterally in all hippocampal regionsexamined. However, the mRNA encoding the full-lengthgp145t r k B was significantly increased in a dose-dependentfashion only in the ipsilateral CA3 and dentate gyrus.Thus, the contralateral increase in trkB mRNA probablymainly corresponds to increase in the gp95t r k B mRNA.This discrepancy may be due to a differential regulation ofthe trkB mRNA species. This is in support of previousfindings of a differential regulation of trkB mRNA speciesw x16,30,37 and consistent with the notion that differenttrkB proteins seem to serve different functions. The non-catalytic truncated molecule may maintain high local con-centrations of BDNF through sequestration on the cellsurface or function analogous to the high-affinity uptakesystems for neurotransmitters by internalization and degra-
w xdation of the ligand–receptor complex 26,32 .

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327 325
BDNF and trkB mRNAs were induced bilaterally in thehippocampus following the unilateral quisqualate injec-tions consistent with the fact that the entorhinal cortex alsogives rise to projections to the contralateral hippocampus
w xin the rat 54 . The present finding of dose-dependentincreases in BDNF and gp95q145t r k B mRNAs in thepyramidal and granule cells may implicate a commonmechanism of activation of BDNF and trkB synthesispossibly related to an autocrine andror paracrine functionfor BDNF in the hippocampus. In support of an autocrinefunction of BDNF, mRNAs for BDNF and trkB areexpressed within the same neurons in the different regions
w xof the hippocampus 29 . Furthermore, in a paracrine fash-ion BDNF can be retrogradely transported, in vivo, byneurons within the hippocampus but also to neurons in the
w xentorhinal cortex from the hippocampus 11 . Our observa-tion of a dose-dependent influence of LEA on the expres-sion of trkB and BDNF mRNAs indicates that corticalafferents may influence BDNF-mediated effects both inthe hippocampus and in the entorhinal cortex by increasingthe expression of both the receptor and ligand in thepyramidal and granule cells in the hippocampus.
4.3. LEA-mediated effects on behaÕioural actiÕity
Much of the extensive cortical input to the hippocampusvia the entorhinal cortex is derived from sensory andmultimodal cortices but also from many subcortical re-gions. In this study we wanted to describe the behaviouralaspects of dose-dependent activation of this input. Noeffect was seen on behaviour following injection of ACSF.However, stimulation of LEA with quisqualate-evokedbehavioural alterations, usually present approximately 30min following administration. These changes were gener-ally preceded by increases in extracellular levels of gluta-mate in the CA1 region. Similar time course in onset ofseizure-like activity has been reported previously in mouseand in cat following intracerebroventricular and intrahip-pocampal injection of comparable concentrations of
w xquisqualate 19,34 , respectively. Injection of 2.4 or 9.6 mgquisqualate-induced behaviour also including seizure-re-lated effects. However, 0.24 mg of quisqualate did notproduce seizure-like activity while changes in behaviourappeared as olfaction, digging and rearing. This suggestthat the stimulated part of LEA-hippocampal circuitry maybe involved in such activities under more physiologicalconditions. The importance of this circuitry for olfaction issupported by findings demonstrating that LEA receives
w xinput from the olfactory bulb 45 and lesions in theentorhinal cortex result in impaired performance of an
w xolfactory discrimination task 1,53 .
4.4. Glutamate release in relation to BDNF and trkBmRNA expression in the dorsal CA1 region
The BDNF gene consists of a multipromotor systemgiving rise to four different mRNAs all encoding the
mature BDNF protein but with different 5X untranslatedw xexons 39,56 . Following stimulation of LEA using the
same concentrations of quisqualate as in this study adifferential and dose-dependent regulation of these differ-ent BDNF promoters in the hippocampus have been
w xdemonstrated 17 . We suggested that physiological activa-tion of the entorhinal cortex may influence, via glutamatecontaining fibres, the BDNF levels in specific hippocampalregions. In support of this notion we provide in vivoevidence of a LEA activity-related glutamate release asso-ciated with an increase of BDNF and trkB mRNA expres-sion in the dorsal CA1 region. Maximum extracellularlevels of glutamate significantly correlated to levels ofmRNAs encoding BDNF and the different forms of thetrkB receptor in the pyramidal cells of CA1. However,levels of mRNA encoding the full-length trkB protein wassomewhat weaker correlated. These results give the im-pression that gp145t r k B mRNA may be more indirectlyinfluenced by glutamate receptor stimulation as comparedto gp95t r k B and BDNF mRNAs. It is likely that theincrease in release of glutamate seen within 20 min follow-ing the stimulation precedes the increase in BDNF mRNAlevels in this study since intraventricular injection ofNMDA have resulted in a detectable increase in hippocam-pal BDNF mRNA levels 1 h but not 15 min following
w xinjection 21 .Observed increases in glutamate release and expression
of different mRNA may to some extent depend on drug-in-duced seizure activity. However, we do not think seizureactivity is a major explanation for observed changes in thepresent study. Accordingly NMDA receptor antagonistsprevent kindling-induced increase in BDNF mRNA levelsw x w x64 but not seizure activity 14 . Furthermore, glutamaterelease in the absence in seizure activity can activate
w xBDNF gene expression 28 . Thus, it appears likely thatthe present observation of increased levels of BDNF andtrkB mRNAs is due to increased glutamatergic neurotrans-mission. This is also consistent with evidence of increasedBDNF mRNA levels in rat dentate gyrus following per-forant pathway stimulation in the absence of seizure activ-
w xity 51 .
5. Conclusion
In conclusion, the present data show that stimulation ofcell bodies in the lateral part of LEA in the awake freelymoving rat in a dose-dependent manner activates a constel-lation of changes in the release of glutamate, in expressionof mRNAs for BDNF and its receptor trkB and in be-haviour. These changes are likely to happen in a sequentialmanner where glutamate release in the hippocampus ispromoting these changes in gene expression and be-haviour.

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327326
Acknowledgements
This work was supported by grants from The SwedishŽ .Medical Research Council Grant 8653 , Magnus Berg-
walls Stiftelse, Cancerfonden, The Swedish Society ofMedicine, Alzheimerfonden, The Swedish Society forMedical Research. We thank Dr. W.T. O’Connor and Dr.Michel Goiny for expert technical advice.
References
w x1 Allen, W.F., Effect of ablating the pyriform-amygdaloid areas andhippocampi on positive and negative olfactory conditioned reflexesand on conditioned olfactory differentation, Am. J. Physiol., 132Ž .1941 81–92.
w x2 Andersen, P., Holmqvist, B. and Voorhoeve, P.E., Entorhinal activa-Ž .tion of dentate granule cells, Acta Physiol. Scand., 66 1966
448–460.w x3 Andersen, P., Holmqvist, B. and Voorhoeve, P.E., Excitatory
synapses on hippocampal apical dendrites activated by entorhinalŽ .stimulation, Acta Physiol. Scand., 66 1966 461–472.
w x4 Balların, M., Ernfors, P., Lindefors, N. and Persson, H., Hippocam-´pal damage and kainic acid injection induce a rapid increase inmRNA for BDNF and NGF in the rat brain, Exp. Neurol., 114Ž .1991 35–43.
w x5 Barbacid, M., Lamballe, F., Pulido, D. and Klein, R., The trk familyof tyrosine protein kinase receptors, Biochim. Biophys. Acta ReÕ.
Ž .Cancer, 10 1991 115–127.w x6 Bunnemann, B., Fuxe, K., Bjelke, B. and Ganten, D., The Brain
Renin-Angiotensin System and its Possible InÕolÕment in VolumeTransmission, Raven Press, New York, 1990.
w x7 Carlson, H., Ronne-Engstrom, E., Ungerstedt, U. and Hillered, L.,¨Seizure related elevations of extracellular amino acids in human
Ž .focal epilepsy, Neurosci. Lett., 140 1992 30–32.w x8 Carpenter, G., Receptors for epidermal growth factor and other
Ž .polypeptide mitogens, Annu. ReÕ. Biochem., 56 1987 881–914.w x9 Cheng, B. and Mattson, M.P., NT-3 and BDNF protect CNS neurons
Ž .against metabolicrexcitotoxic insults, Brain Res., 640 1994 56–67.w x10 Collingridge, G.L. and Lester, R.A., Excitatory amino acid receptors
in the vertebrate central nervous system, Pharmacol. ReÕ., 40Ž .1989 43–210.
w x11 DiStefano, P.S., Friedman, B., Radziejewski, C., Alexander, C.,Boland, P., Schick, C.M., Lindsay, R.M. and Wiegand, J., Theneurotrophins: BDNF, NT-3, and NGF display distinct patterns ofretrograde axonal transport in the peripheral and central neurons,
Ž .Neuron, 8 1992 983–993.w x12 Drew, K.L., O’Connor, W.T., Kehr, J. and Ungerstedt, U., Charac-
terization of g-aminobuturic acid and dopamine overflow followingŽ .acute implantation of a microdialysis probe, Life Sci., 45 1989
1307–1317.w x13 During, M.J. and Spencer, D.D., Extracellular hippocampal gluta-
mate and spontaneous seizures in the conscious human brain, Lancet,Ž .341 1993 1607–1610.
w x14 Ernfors, P., Bengzon, J., Persson, H. and Lindvall, O., Increasedlevels of messenger RNA for neurotrophic factors in the brain
Ž .during kindling epileptogenesis, Neuron, 7 1991 165–176.w x15 Ernfors, P., Wetmore, C., Olson, L. and Persson, H., Identification
of cells in rat brain and peripheral tissues expressing mRNA forŽ .members of the nerve growth factor family, Neuron, 5 1990
511–526.w x16 Falkenberg, T., Ernfors, P., Persson, H. and Lindefors, N., Cortical
transynaptic activation of tyrosine kinase receptor trkB messenger
Ž .RNA expression in rat hippocampus, Neuroscience, 51 1992 883–889.
w x17 Falkenberg, T., Metsis, M., Timmusk, T. and Lindefors, N., Entorhi-nal cortex regulation of multiple brain-derived neurotrophic factor
Ž .promoters in the rat hippocampus, Neuroscience, 57 1993 891–896.w x18 Fonnum, F., Glutamate: a neurotransmitter in the mammalian brain,
Ž .J. Neurochem., 42 1984 1–11.w x19 Fukuda, H., Tanaka, T., Kaijima, M., Nakai, H. and Yonemasu, Y.,
Quisqualic acid-induced hippocampal seizures in unanesthetized cats,Ž .Neurosci. Lett., 59 1985 53–59.
w x20 Funakoshi, H., Frisen, J., Barbany, G., Timmusk, T., Zachrisson, O.,´Verge, V.M.K. and Persson, H., Differential expression of mRNAsfor neurotrophins and their receptors after axotomy of the sciatic
Ž .nerve, J. Cell Biol., 123 1993 455–465.w x21 Gwag, B.J. and Springer, J.E., Activation of NMDA receptors
Ž .increase brain-derived neurotrophic factor BDNF mRNA expres-Ž .sion in the hippocampal formation, NeuroReport, 5 1993 125–128.
w x22 Hofer, M., Pagliusi, S.R., Hohn, A., Leibrock, J. and Barde, Y.A.,Regional distribution of brain-derived neurotrophic factor mRNA in
Ž .the adult mouse brain, EMBO J., 9 1990 2459–2464.w x23 Hunter, T. and Cooper, J.A., Protein-tyrosine kinases, Annu. ReÕ.
Ž .Biochem., 54 1985 897–930.w x24 Isackson, P.J., Huntsman, M.M., Murray, K.D. and Gall, C.M.,
BDNF mRNA expression is increased in adult forebrain after limbicseizures: temporal patterns of induction distinct from NGF, Neuron,Ž .1991 937–948.
w x25 Jones, R.S.G., Entorhinal-hippocampal connections: a speculativeŽ .view of their function, Trends Neurosci., 16 1993 58–64.
w x26 Klein, R., Conway, D., Parada, L.F. and Barbacid, M., The trkBtyrosine protein kinase gene codes for a second neurogenic receptor
Ž .that lacks the catalytic kinase domain, Cell, 61 1990 647–656.w x27 Klein, R., Parada, L.F., Coulier, F. and Barbacid, M., trkB, a novel
tyrosine protein kinase receptor expressed during mouse neuralŽ .development, EMBO J., 8 1989 3701–3709.
w x28 Kokaia, Z., Gido, G., Ringstedt, T., Bengzon, J., Kokaia, M., Siesjo,¨ ¨B.K., Persson, H. and Lindvall, O., Rapid increase of BDNF mRNAlevels in cortical neurons following spreading depression: regulationby glutamtergic mechansims independent of seizure activity, Mol.
Ž .Brain Res., 4 1993 277–286.w x29 Kokaia, Z., Bengzon, J., Metsis, M., Kokaia, M., Persson, H. and
Lindvall, O., Coexpression of neurotrophins and their receptors inneurons of the central nervous system, Proc. Natl. Acad. Sci. USA,
Ž .90 1993 6711–6715.w x30 Lindefors, N., Brodin, E. and Metsis, M., Spatiotemporal selective
effects on brain-derived neurotrophic factor and trkB messengerRNA in rat hippocampus by electroconvulsive shock, Neuroscience,
Ž .65 1995 661–670.w x31 Lindefors, N., Ernfors, P., Falkenberg, T. and Persson, H., Septal
cholinergic afferents regulate expression of brain-derived neu-rotrophic factor and b-nerve growth factor mRNAin rat hippocam-
Ž .pus, Exp. Brain Res., 88 1992 78–90.w x32 Lindsay, R.M., Wiegand, S.J., Altar, C.A. and DiStefano, P.S.,
Neurotrophic factors: from molecule to man, Trends Neurosci., 17Ž .1994 182–190.
w x33 Maisonpierre, P.C., Le Beau, M.M., Espinosa, R.I., Ip, N.Y., Bellus-cio, L., de la Monte, S.M., Squinto, S., Furth, M.E., and Yancopou-los, G.D., Human and rat brain-derived neurotrophic factor andneurotrophin-3: gene structures, distributions, and chromosomal lo-
Ž .calizations, Genomics, 10 1991 558–568.w x34 Mathis, C. and Ungerer, A., Comparative analysis of seizures in-
duced by intracerebroventricular administration of NMDA, kainateŽ .and quisqualate in mice, Exp. Brain Res., 88 1992 277–282.
w x35 Mattson, M.P., Lee, R.E., Adams, M.E., Guthrie, P.B. and Kater,S.B., Interactions between entorhinal axons and target hippocampalneurons: a role for glutamate in the development of hippocampal
Ž .circuitry, Neuron, 1 1988 865–876.w x36 Meana, J.J., Herrera-Marschitz, M., Brodin, E., Hokfelt, T. and¨

( )T. Falkenberg et al.rMolecular Brain Research 42 1996 317–327 327
Ungerstedt, U., Simultaneous determination of cholecystokinin,dopamine, glutamate and aspartate in cortex and striatum of the rat
Ž .using in vivo microdialysis, Amino Acids, 1 1991 365–373.w x37 Merlio, J.-P., Ernfors, P., Bengzon, J., Kokaia, Z., Kokaia, M.,
Lindvall, O., Smith, M.-J., Siesjo, B.-K., Middlemas, D.S., Hunter,¨T. and Persson, H., Increased production and activation of the
Ž .tyrosine kinase receptor trkB after brain insults, Neuron, 10 1993151–164.
w x38 Middlemas, D.S., Lindberg, R.A. and Hunter, T., trkB, a neuralreceptor protein-tyrosine kinase: evidence for a full-length and two
Ž .truncated receptors, Mol. Cell Biol., 11 1991 143–153.w x39 Nakayama, M., Gahara, Y., Kitamura, T. and Ohara, O., Distinctive
four promoters collectively direct expression of brain-derived neu-Ž .rotrophic factor gene, Mol. Brain Res., 21 1994 206–218.
w x40 Narahashi, T., Chemical tools in the study of excitable membranes,Ž .Physiol. ReÕ., 54 1974 813–889.
w x41 Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic Coordi-nates, Academic Press, San Diego, 1986.
w x42 Persson, H., Neurotrophin production in the brain, Semin. Neurosci.,Ž .5 1993 227–237.
w x43 Peterson, D.W., Collins, J.F. and Bradford, H.F., The kindled amyg-dala model of epilepsy: anticonvulsant action of amino acid antago-
Ž .nists, Brain Res., 275 1983 169–172.w x44 Phillips, H.S., Hains, J.M., Laramee, G.R., Rosenthal, A. and
Winslow, J.W., Widespread expression of BDNF but not NT-3 bytarget areas of basal forebrain cholinergic neurons, Science, 250Ž .1990 290–294.
w x45 Powell, T.P.S., Cowan, W.M. and Raisman, G., The central olfac-Ž .tory connections, J. Anat., 99 1965 791–813.
w x46 Schroder, H., Cellular and subcellular distribution of receptors in the¨entorhinal-hippocampal system: morphologic and biochemical as-
Ž .pects, Hippocampus, 3 1993 139–148.w x47 Sloviter, R.S., Permanently altered hippocampal structure, excitabil-
ity, and inhibition after experimental status epilepticus in the rat:‘the dormant basket cell’ hypothesis and its possible relevance to
Ž .temporal lobe epilepsy, Hippocampus, 1 1991 41–66.w x48 Soerensen, K.E., Projections of the entorhinal area to the striatum,
nucleus accumbens, and cerebral cortex in the guinea pig, J. Comp.Ž .Neurol., 238 1985 308–322.
w x49 Soppet, D., Escandon, E., Maragos, J., Middlemas, D.S., Reid,S.W., Blair, J., Burton, L.E., Stanton, B.R., Kaplan, D.R., Hunter,T., Nikolics, K. and Parada, L.F., The neurotrophic factors brain-de-rived neurotrophic factor and neurotrophin-3 are ligands for the trkB
Ž .tyrosine kinase receptor, Cell, 65 1991 895–903.w x50 Sperk, G., Lassmann, H., Baran, H., Seitelberger, F. and
Hornykiewicz, O., Kainic acid-induced seizures: dose-relationship ofŽ .behavioural and histopathological changes, Brain Res., 338 1985
289–295.
w x51 Springer, J.E., Gwag, B.J. and Sessler, F.M., Neurotrophic factormRNA expression in dentate gyrus is increased following in vivo
Ž .stimulation of the angular bundle, Mol. Brain Res., 23 1994135–143.
w x52 Squinto, S.P., Stitt, T.N., Aldrich, T.H., Davis, S., Bianco, S.M.,Radziejewski, C., Glass, D.J., Masiakowski, P., Furth, M.E., Valen-zuela, D.M., DiStefano, P.S. and Yancopoulos, G.D., trkB encodesa functional receptor for brain-derived neurotrophic factor and neu-
Ž .rotrophin-3 but not nerve growth factor, Cell, 65 1991 885–893.w x53 Staubli, U., Ivy, G. and Lynch, G., Hippocampal denervation causes
rapid forgetting of olfactory information in rats, Proc. Natl. Acad.Ž .Sci. USA, 81 1984 5885–5887.
w x54 Steward, O., Topographic organization of the projections from theentorhinal area to the hippocampal formation of the rat, J. Comp.
Ž .Neurol., 167 1976 285–314.w x55 Swanson, L.W., Kohler, C. and Bjorklund, A., The Limbic Region. I.¨ ¨
The Septohippocampal System, Elsevier, Amsterdam, 1987.w x56 Timmusk, T., Palm, K., Metsis, M., Reintam, T., Paalme, V.,
Saarma, M. and Persson, H., Multiple promoters direct tissue-specificŽ .expression of the rat BDNF gene, Neuron, 10 1993 475–489.
w x57 Van Hoesen, G.W., The primate parahippocampal gyrus: New in-Ž .sights regarding its cortical connection, Trends Neurosci., 5 1982
345–350.w x58 Watkins, J.C. and Evans, R.H., Excitatory amino acid transmitters,
Ž .Annu. ReÕ. Pharmacol. Toxicol., 21 1981 165–204.w x59 White, W.F., Nadler, J.V., Hamberger, A., Cotman, C.W. and
Cummings, J.T., Glutamate as transmitter of hippocampal perforantŽ .path, Nature, 270 1977 356–357.
w x60 Witter, M.P., Organization of the entorhinal-hippocampal system: aŽ .review of current anatomical data, Hippocampus, 3 1993 33–44.
w x61 Witter, M.P. and Groenewegen, H.J., Connections of the parahip-pocampal cortex in the cat. III. Cortical and thalamic efferents, J.
Ž .Comp. Neurol., 252 1986 1–31.w x62 Witter, M.P., Groenewegen, H.J., Lopes da Silva, F.H. and Lohman,
A.H.M., Functional organization of the extrinsic and intrinsic cir-Ž .cuitry of the parahippocampal region, Prog. Neurobiol., 33 1989
161–253.w x63 Zafra, F., Castren, E., Thoenen, H. and Lindholm, D., Interplay´
between glutamate and g-aminobutyric acid transmittor systems inthe physiological regulation of brain-derived neurotrophic factor andnerve growth factor synthesis in hippocampal neurons, Proc. Natl.
Ž .Acad. Sci. USA, 88 1991 10037–10041.w x64 Zafra, F., Hengerer, B., Leibrock, J., Thoenen, H. and Lindholm, D.,
Activity dependent regulation of BDNF and NGF mRNAs in the rathippocampus is mediated by non-NMDA glutamate receptors, EMBO
Ž .J., 9 1990 3545–3550.




















