
Genetic risk factors for major depression in men and women: similar or different heritabilities and...
12
Psychological Medicine, 2001, 31, 605–616. Printed in the United Kingdom " 2001 Cambridge University Press Genetic risk factors for major depression in men and women : similar or different heritabilities and same or partly distinct genes ? K.S.KENDLER, " C.O. GARDNER, M.C.NEALE C.A. PRESCOTT From the Departments of Psychiatry and Human Genetics, Medical College of Virginia, of Virginia Commonwealth University and Virginia Institute for Psychiatric and Behavioral Genetics, Richmond, VA, USA ABSTRACT Background. Although women are at consistently greater risk for major depression (MD) than men, it is unclear whether sex modifies the aetiological impact of genetic factors on MD. Is the heritability of MD different in men and women ? Do the same genetic risk factors predispose to MD in the two sexes? Methods. We obtained a lifetime history of MD by personal interview on two occasions from 6672 individual twins and 2974 complete twin pairs. Three diagnostic criteria of increasing narrowness were employed : DSM-III-R, DSM-III-R plus impairment and Washington University. To increase power by controlling for unreliability of assessment, we evaluated sex differences on genetic risk for MD using a structural equation measurement model. Results. Using DSM-III-R criteria, but not the two narrower definitions, heritability of MD was significantly greater in women than in men. In the three diagnostic systems, the genetic correlation in liability to MD in men and women was estimated at between ›0–50 and ›0–65. These estimates differed significantly from unity for the two broader definitions. Conclusion. Using broad but not narrower definitions of illness, genetic factors play a greater role in the aetiology of MD in women than in men. The genes that influence risk for MD in the two sexes are correlated but are probably not entirely the same. These results raise the possibility that, in linkage and association studies, the impact of some loci on risk for MD will differ in men and women. INTRODUCTION The differences in population risk for major depression (MD) between men and women is one of the most reliable findings in the epi- demiology of MD (Boyd & Weissman, 1981 ; Weissman et al. 1991 ; Kessler et al. 1993). Extensive research, recently summarized (Bebbington, 1996), has sought to clarify the contribution of social and psychological factors to sex differences in MD. An alternative ap- proach is to clarify the existence and nature of sex differences in the role of genetic factors in the aetiology of MD. Two questions are of " Address for correspondence : Professor Kenneth S. Kendler, Box 980126, Richmond, VA 23298-0126, USA. interest. First, do the relative aetiological roles of genetic and environmental risk factors for MD differ in men and women ? Secondly, are the genetic risk factors for MD operative in men and women the same? Several studies have addressed these questions in twins (Bertelsen et al. 1977 ; Kendler et al. 1995 ; McGuffin et al. 1996; Bierut et al. 1999) and adoptees (Cadoret et al. 1985) with con- flicting results. This pattern of findings may result from the large sample sizes needed to detect reliably such sex differences in genetic risk factors for dichotomous traits like lifetime MD (Prescott & Gottesman, 1993 ; Neale & Kendler, unpublished results). In a previous report, we addressed these two questions in a sample of 605
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Genetic risk factors for major depression in men and women: similar or different heritabilities and...

Psychological Medicine, 2001, 31, 605–616. Printed in the United Kingdom" 2001 Cambridge University Press
Genetic risk factors for major depression in men and
women: similar or different heritabilities and same or
partly distinct genes?
K. S. KENDLER," C. O. GARDNER, M. C. NEALE C. A. PRESCOTT
From the Departments of Psychiatry and Human Genetics, Medical College of Virginia, of VirginiaCommonwealth University and Virginia Institute for Psychiatric and Behavioral Genetics, Richmond, VA,
USA
ABSTRACT
Background. Although women are at consistently greater risk for major depression (MD) thanmen, it is unclear whether sex modifies the aetiological impact of genetic factors on MD. Is theheritability of MD different in men and women? Do the same genetic risk factors predispose to MDin the two sexes?
Methods. We obtained a lifetime history of MD by personal interview on two occasions from 6672individual twins and 2974 complete twin pairs. Three diagnostic criteria of increasing narrownesswere employed: DSM-III-R, DSM-III-R plus impairment and Washington University. To increasepower by controlling for unreliability of assessment, we evaluated sex differences on genetic risk forMD using a structural equation measurement model.
Results. Using DSM-III-R criteria, but not the two narrower definitions, heritability of MD wassignificantly greater in women than in men. In the three diagnostic systems, the genetic correlationin liability to MD in men and women was estimated at between 0±50 and 0±65. These estimatesdiffered significantly from unity for the two broader definitions.
Conclusion. Using broad but not narrower definitions of illness, genetic factors play a greater rolein the aetiology of MD in women than in men. The genes that influence risk for MD in the two sexesare correlated but are probably not entirely the same. These results raise the possibility that, inlinkage and association studies, the impact of some loci on risk for MD will differ in men andwomen.
INTRODUCTION
The differences in population risk for majordepression (MD) between men and women isone of the most reliable findings in the epi-demiology of MD (Boyd & Weissman, 1981;Weissman et al. 1991; Kessler et al. 1993).Extensive research, recently summarized(Bebbington, 1996), has sought to clarify thecontribution of social and psychological factorsto sex differences in MD. An alternative ap-proach is to clarify the existence and nature ofsex differences in the role of genetic factors inthe aetiology of MD. Two questions are of
" Address for correspondence: Professor Kenneth S. Kendler, Box980126, Richmond, VA 23298-0126, USA.
interest. First, do the relative aetiological rolesof genetic and environmental risk factors forMD differ in men and women? Secondly, are thegenetic risk factors for MD operative in menand women the same?
Several studies have addressed these questionsin twins (Bertelsen et al. 1977; Kendler et al.1995; McGuffin et al. 1996; Bierut et al. 1999)and adoptees (Cadoret et al. 1985) with con-flicting results. This pattern of findings mayresult from the large sample sizes needed todetect reliably such sex differences in genetic riskfactors for dichotomous traits like lifetime MD(Prescott & Gottesman, 1993; Neale & Kendler,unpublished results). In a previous report, weaddressed these two questions in a sample of
605

606 K. S. Kendler and others
3790 twin pairs interviewed on one occasionabout their lifetime history of MD (Kendler &Prescott, 1999; Kendler et al. 1999). We foundslightly greater correlations in liability to MD inthe female–female (FF) versus the male–male(MM) pairs, but the best-fit model suggestedequal heritability for MD in the two sexes(Kendler & Prescott, 1999; Kendler et al. 1999).In addition, the best-fit model suggested that thegenetic risk factors for MD, while positivelycorrelated in men and women, were not entirelythe same. However, despite the large samplesize, based on a single occasion of measurementthe statistical superiority of this model over onethat constrained the genetic risk factors to beidentical in the two sexes was very modest.
We have recently completed follow-up inter-views in this twin sample in which we re-assessedthe lifetime history of MD. This permitted us tore-examine the critical question of the impact ofsex on genetic risk factors for MD using a‘measurement model ’ (Kendler, 1993; Foley etal. 1998; Kendler et al. 1998). By incorporatingdata from two waves of assessment, this modelcan estimate the impact of genetic and en-vironmental risk factors controlling for theunreliability of measurement associated with theassessment of lifetime history of psychiatricdisorders in general population samples (Prusoffet al. 1988; Rice et al. 1992; Williams et al.1992). For traits with moderate reliability, thegain in statistical power with this model can besubstantial and even greater than that obtainedfrom interviewing a new sample of comparablesize (Neale et al. 1997). Finally, we have exploredwhether the impact of sex differences on geneticrisk factors for MD is stable across definitionsof MD of varying stringency.
METHOD
Subjects
Our twin sample derives from two inter-relatedprojects utilizing the population-based VirginiaTwin Registry (Kendler & Prescott, 1999) –formed from a systematic review of all birthcertificates in the Commonwealth of Virginia.The female–female twin pairs used in this studycome from birth years 1934–1974. Twin pairsbecame eligible to participate if both membershad previously responded to a mailed question-naire, the response rate to which was C 64%.
Eighty-eight per cent of our sample was firstinterviewed face-to-face in 1987–9 and hassubsequently been the subject of three additionaltelephone interview waves. We examine hereresults from the last two of these wavescompleted in 1992–4 and 1995–7, respectively.The mean interval (³..) between these twointerviews was 31±5³7 (range 13–49) months.
The male–male and male–female twin pairs,covering the birth years 1940–1974, were ascer-tained in a separate study beginning in 1993(Kendler & Prescott, 1999) in which wesucceeded in interviewing by telephone 72% ofthose eligible. This sample was followed-up in asecond wave of interviews (79±4% of whom werecompleted face-to-face) with a mean inter-interview interval of 19±0³9 months (range12–65) months.
For the purposes of this study, the time 1interview refers to the 3rd wave interview withthe female–female twins and the 1st waveinterview with the male–male}male–female twinpairs. The time 2 interview refers to the 4th waveinterview with the female–female pairs and the2nd wave interview with the male–male}male–female pairs. As outlined previously (Kendler &Prescott, 1999), zygosity was determined by acombination of self-report measures, photo-graphs and polymorphic DNA markers with avalidated error rate of % 5%. Interviewers wereblind to information about the co-twin andprior interviews with the twin.
Measures
Lifetime MD was assessed, with high inter-raterreliability (κ¯ 0±96³0±04), by a structured psy-chiatric interview based on the SCID (Spitzer &Williams, 1985; Kendler et al. 1992a). However,we assessed last-year history for MD and alifetime history for MD prior to the last year intwo separate sections. Interviewers had at least amaster ’s degree in a mental health-related fieldor a bachelor’s degree in such a field and 2 yearsof clinical experience. Each interview wasreviewed by at least one senior staff member forcompleteness and consistency. Data weredouble-entered to minimize data entry errors.
For this study, we utilized three definitions oflifetime MD all requiring an age at onset at orprior to the time 1 interview. The time 2interviews contained a special item to determine,for individuals reporting recent first onsets of

Genetic risk factors for major depression 607
MD, whether the MD preceded or followed thedate of the time 1 interview. If individuals at thewave 2 interview reported an onset of MD whichoccurred after the time 1 interview, they were,for the purposes of these analyses, counted asunaffected.
The three definitions for lifetime MD utilizedwere: DSM-III-R criteria (American PsychiatricAssociation, 1987), DSM-III-R criteria plussignificant episode-related impairment (DSM-III-RI) and Washington University (WU) (orFeighner) criteria (Feighner et al. 1972). Weutilize DSM-III-RI for MD rather than DSM-IV criteria because the time 1 assessment in thefemale–female pairs lacked an item assessingepisode-related distress required to rate criterionC (American Psychiatric Association, 1994). Byrequiring impairment rather than impairment ordistress, DSM-III-RI criteria for MD shouldbe narrower than DSM-IV criteria. WU criteriarequire 1 month of dysphoric mood plus & 5 of8 criteria : poor appetite}weight loss, sleepdifficulty, energy loss, agitation}retardation,guilt, difficulty concentrating and suicidalideation.
Statistical analysis
We use a liability-threshold model to estimatethe genetic and environmental contributions totwin resemblance. For categorical characteristicslike MD, the estimates are for the resemblanceof twins in a pair for their liability to develop thedisorder (Pearson, 1901; Falconer, 1965). Liab-ility is assumed to be continuous and normallydistributed in the population, with individualswho exceed a theoretical threshold expressingthe disorder. We present the tetrachoric cor-relation, defined as the correlation in membersof twin pairs for the liability to MD (Pearson,1901; Falconer, 1965).
We divide the variation in liability to MD intothree classes : (i) additive genetic (A), whichcontributes twice as much to the correlation inMZ twins as DZ twins; (ii) family or ‘common’environment (those familial factors which maketwins similar in their liability to MD) (C), whichcontributes equally to the correlation in MZand DZ twins; and (iii) individual specific en-vironment (E), which reflects environmentalexperiences not shared by both members of atwin pair and therefore contributes to differencesbetween them in their liability to MD. Because
previous analyses, as well as analyses of themodels used here, provided no evidence forsignificant dominance genetic effects on theliability to MD, it will not be considered further.
Our model for MD utilizes simultaneouslyboth our time 1 and time 2 data from the fivetwin-zygosity groups: female–female MZ,female–female DZ, male–male MZ, male-maleDZ and male–female DZ. As pictured in Fig. 1,the model assumes that there is a true latentliability to a lifetime history of MD. Each of ourtwo assessments of lifetime history are fallibleindices of this liability. The paths λ
f"and λ
f#represent the degree to which the assessments oflifetime history of MD obtained at the two timepoints reflect this liability in females. The pathsλm"
and λm#
represent the corresponding pathsin males. The square of these λ paths is ameasure of the test–retest reliability of theseassessments. The other paths to lifetime historyofMD(κ
f", κ
f#, κ
m", κ
m#) represent the proportion
of variance due to the unreliability of theindividual assessments of lifetime history of MDat times 1 and 2 in females and males,respectively. Within each sex, the sums of λ# andκ# are constrained to unity. The latent liability tolifetime MD is then modelled in a standard sex-specific twin design that consists of two majorfeatures. First, the model permits the estimationof sex-dependent genetic and environmentalparameters (e.g. sex-specific genetic risk factorsin females A
Fand A
M). Secondly, the model
estimates the correlation between the geneticand familial–environmental factors in males andfemales by the parameters r
aand r
crespectively.
Further elaborations of this model are poss-ible. In particular, the errors of measurementmay be correlated in twin pairs. To test thisassumption, the model can be modified to testwhether the error variance is influenced bygenetic, common environmental or individual-specific environmental factors that act only attime 1 or only at time 2.
There are two critical differences between thismodel and standard twin models. First, thismodel provides separate estimates for error ofmeasurement (κ) and true individual specificenvironment (e) that contributes to the liabilityto reliably assessed MD. Secondly, it provides adirect estimate of the degree to which theindividual assessments of lifetime history of MDreflect the latent liability (λ).

608 K. S. Kendler and others
rc
ra
AF AMCFEF CM EM
emcmamafcfef
Liability to MDin females
Liability to MDin males
km2km1kf1 kf2
Lifetimehistory of MD
in femalesat time 1
Lifetimehistory of MD
in femalesat time 2
Lifetimehistory of MD
in malesat time 1
Lifetimehistory of MD
in malesat time 2
jm2jm1jf2jf1
F. 1. A twin model for the liability to major depression (MD) in females and males including error of measurement. Observedvariables are depicted in boxes and latent variables in circles and ellipses. This model assumes that individual twins have a true latentliability to MD, which is imperfectly indexed by two assessments, at time 1 and time 2. The paths λ
f"and λ
f#and λ
m"and λ
m#represent
the degree to which the assessments of lifetime history of MD obtained at the two time points reflect this true liability in femalesand males, respectively. The other paths to lifetime history of MD (κ
f", κ
f#, κ
m", κ
m#) represent the proportion of variance due to the
unreliability of the individual assessments of lifetime history of MD at times 1 and 2 in females and males, respectively. The latentliability to lifetime MD is then modelled in a standard sex-specific twin design with the sources of variance divided between additivegenetic (A), familial environmental (C) and individual-specific environmental factors (E). The model permits the estimation of sex-dependent genetic and environmental parameters (e.g. sex-specific genetic risk factors in females A
Fand males A
M) and the
correlation between the genetic and familial–environmental factors in males and females by the parameters raand r
c, respectively.
Traditionally, we would have included in thismodelling only twin pairs where both membershad been interviewed at both times. However,information about diagnostic thresholds is pro-vided by twins whose co-twin was not inter-viewed. Furthermore, information about causesof twin resemblance at time 1 is contained inpairs where both have been interviewed at time1 but not time 2. Therefore, using a recentoption in the software package Mx (Neale,1991), we fit models to all pairs including thosewith incomplete information by the method ofmaximum likelihood. During optimization, trialvalues of the parameters are used to generate anoverall predicted covariance matrix and anoverall matrix of thresholds. For any particularobservation, the overall matrix is filtered tocreate a submatrix matching those observationsthat are present. Likewise, the correspondingsubset of the matrix of thresholds is created.These matrices are then used to compute the log-
likelihood of the observation in question. Thelog-likelihoods of all cases are summed to obtainthe log-likelihood of the sample, which ismaximized using numerical optimization soft-ware.
An advantage of this method is that it reducesthe possible impact of cooperation bias. Es-sentially it uses information on potentialdifferences in the prevalence of MD in thosetwins with v. without a co-twin participating toobtain a better estimate of the prevalences ofMD in the population. The method is a binarydata maximum likelihood application of the‘missing at random’ principle expounded byLittle & Rubin (1987).
Maximum likelihood analysis of raw ordinaldata does not directly provide an overall test ofgoodness-of-fit of the model. We comparedrelative fits against a saturated model. Twice thedifference in log-likelihood between this satu-rated model and the model being tested yields a

Genetic risk factors for major depression 609
statistic that is (under certain regularityconditions) asymptotically distributed as χ# withdegrees of freedom equal to the difference in thenumber of parameters in the two models.
Alternative models are evaluated by com-paring the difference in their χ# values relative tothe difference in their degrees of freedom (df),according to the principle of parsimony – modelswith fewer parameters are preferable if they donot provide significantly worse fit. Weoperationalized this balance between explana-tory power and parsimony by the use of Akaike’sinformation criterion (AIC) (Akaike, 1987;Williams & Holahan, 1994), calculated as: χ#®2 df. These analyses were based on a total of14421 individual interviews (7746 time 1 and6675 time 2) including 3789 complete pairs attime 1 and 2974 at time 2.
We have previously examined, from severalperspectives, the equal environment assumptionfor MD in this data (that the exposure toenvironmental risk factors for MD is approxi-mately equal in MZ and DZ twins). Theseanalyses, including examinations of wave 1 dataused in this report, found no evidence to rejectthe equal environment assumption (Kendler etal. 1992a, 1993a, b ; Kendler & Prescott, 1999).Using the wave 2 data, we again examine thispossibility in two ways: by examining standardmeasures of the environmental similarity oftwins in childhood (Loehlin & Nichols, 1976)and the frequency of current contact among themembers of the twin pair in adulthood. Usinglogistic regression and controlling for zygosity,we examined whether the mean level of child-hood or adult environmental similarity reportedby the twin pair interacted with the diagnosis ofMD in one twin in predicting the risk for MD inthe co-twin.
RESULTS
Prevalence of lifetime major depression
Four points about the lifetime prevalence ratesof MD in this sample are noteworthy (Table 1).First, rates for DSM-III-R criteria consistentlyexceeded those found for DSM-III-RI criteria,which in turn exceeded those found using WUcriteria. Secondly, rates in females were con-sistently higher than those in males. Thirdly,lifetime prevalence rates at wave 1 were generallyhigher than those reported at wave 2. Fourthly,
rates in DZ same sex (SS) pairs were modestlygreater than those seen in MZ pairs. Fifthly,rates of MD for both males and females weregreater in the DZ opposite-sex (OS) than in theDZ SS pairs.
Equal environment assumption
In our wave 2 data, we examined whether twinresemblance for lifetime MD could be predictedfrom the similarity of the twins’ childhood oradult environments. We performed these analy-ses for all three diagnostic categories, separatelyin male–male and female–female pairs. Of these12 comparisons, one was significant at the 5%level : using the DSM-III-R definition, morefrequent current contact predicted twin resem-blance for lifetime MD in male–male pairs (χ#¯7±7, df¯ 1, P¯ 0±006). This pattern of findingsis consistent with chance expectations (Feild &Armenakis, 1974).
Tetrachoric correlation matrices
Table 2 shows the tetrachoric correlationmatrices, for the five twin types (female–femaleMZ, female–female DZ, male–male MZ, male–male DZ and male–female DZ), of the cor-relation in liability to lifetime MD between andwithin twins over the two occasions of measure-ment using DSM-III-R criteria. These matricescontain three types of correlations. The within-twin across-time correlations are all quite similarand range from 0±67 to 0±76. There appearto be no systematic differences in the stability ofthe diagnosis of lifetime MD across genders ortwin types.
Four trends are evident for the between-twinwithin-time correlations. First, the correlationsare stable across the two times of measurement.For example, in the male–male pairs, the MZcorrelations at times 1 and 2 were 0±33 and0±27 and the parallel DZ correlations were0±10 and 0±12. Secondly, substantially highercorrelations are observed in the MZ twin pairsthan in the DZ pairs in both males and females.This is the expected pattern if genetic risk factorsinfluenced liability to MD in both males andfemales. Thirdly, the correlations in the male–female DZ pairs are lower than that seen in thesame-sex DZ pairs. This pattern is consistentwith the hypothesis that the familial factors thatinfluence MD are not entirely the same in menand women. Fourthly, in both the MZ and the

610 K. S. Kendler and others
Table 1. Prevalence rates for lifetime major depression prior to the Wave 1 interview with threedefinitions in monozygotic and dizygotic female–female, male–male and opposite-sex twin pairs
Zygosity Gender
Wave 1 Wave 2
N DSM-III-R DSM-III-R-I WUC N DSM-III-R DSM-III-R-I WUC
MZ FF 1088 31±3 22±3 17±5 963 35±7 22±8 24±0DZ-SS FF 774 36±0 25±6 22±4 689 35±6 21±9 23±2MZ MM 1735 27±3 19±6 15±6 1526 24±0 15±9 14±5DZ-SS MM 1303 29±2 22±9 18±1 1098 26±0 16±8 15±9DZ-OS F 1422 40±9 31±7 27±2 1203 34±5 25±4 22±5DZ-OS M 1424 31±7 24±8 20±5 1196 26±3 19±6 17±2
DSM-III-R-I indicates DSM-III-R criteria impairment ; WUC, Washington University Criteria ; MZ, monozygotic ; DZ-SS dizygotic,same sex; DZ-OS, dizygotic, opposite sex; FF, female–female ; MM. male–male.
Table 2. Tetrachoric correlation matrix forlifetime DSM-III-R major depression assessed ontwo occasions
Twin type Tw1Tm1 Tw1Tm2 Tw2Tm1 Tw2Tm2
MZ Female–FemaleTw1Tm1 1±000Tw1Tm2 0±671 1±000Tw2Tm1 0±350 0±453 1±000Tw2Tm2 0±310 0±420 0±698 1±000
DZ Female–FemaleTw1Tm1 1±000Tw1Tm2 0±690 1±000Tw2Tm1 0±200 0±200 1±000Tw2Tm2 0±099 0±148 0±751 1±000
MZ Male–MaleTw1Tm1 1±000Tw1Tm2 0±707 1±000Tw2Tm1 0±334 0±350 1±000Tw2Tm2 0±301 0±274 0±721 1±000
DZ Male–MaleTw1Tm1 1±000Tw1Tm2 0±676 1±000Tw2Tm1 0±104 0±234 1±000Tw2Tm2 0±016 0±121 0±726 1±000
DZOS (male¯ 1)Tw1Tm1 1±000Tw1Tm2 0±726 1±000Tw2Tm1 0±114 0±072 1±000Tw2Tm2 0±130 0±078 0±711 1±000
Tw1, twin 1; Tw2, twin 2; Tm1, time 1; Tm2, time 2.
DZ pairs, the correlations are higher in the FFthan in the MM pairs, as expected if MD weremore heritable in women than in men.
Two patterns are noteworthy for the between-twin across-time correlations. First, with theexception of the male–male DZ pairs, the twoparallel correlations (i.e. twin 1–time 1 with twin2–time 2 and twin 1–time 2 with twin 2–time 1)are relatively similar. Secondly, in all five twintypes, the between-twin across-time correlations
are similar in magnitude to the between-twinwithin-time correlations. This is the expectedpattern if the familial factors influencing diseaserisk were the same at the two times of measure-ment.
Model fitting
We describe in detail the model fitting usingDSM-III-R criteria (Table 3) and then morebriefly outline the model fitting using the othercriteria. Model I is the fully saturated basemodel against which the fit of other models canbe judged. In model II, we constrain thethresholds and λ paths to be equal for twin 1and twin 2 within all same-sex pairs. This can bedone with a substantial improvement in theAIC. In models III and IV, we constrainthresholds to be equal in OS and SS female andmale twins, respectively. Both of these attemptsfailed and these models were rejected. Furthermodel-fitting, therefore, incorporated separatethresholds for twins for OS pairs. In model V,we constrained all λ paths to be equal in SS andOS pairs, in males and females and at time 1 andtime 2. This model produced another substantialimprovement in the AIC, indicating that thereliability of MD in this sample does not differas a function of sex or occasion of measurement.
In models VI and VII, we set to zero thecommon environmental and genetic paths, re-spectively. Setting the c paths to zero resulted inno change in the fit of the model and a furtherimprovement in the AIC. By contrast, con-straining to zero the genetic paths produced asubstantial worsening of the fit and the AIC andcould also be rejected by the more rigorous χ#
difference text (model VII v. V χ#¯ 20±4, df¯ 1,P! 0±0001). These results suggest that, within
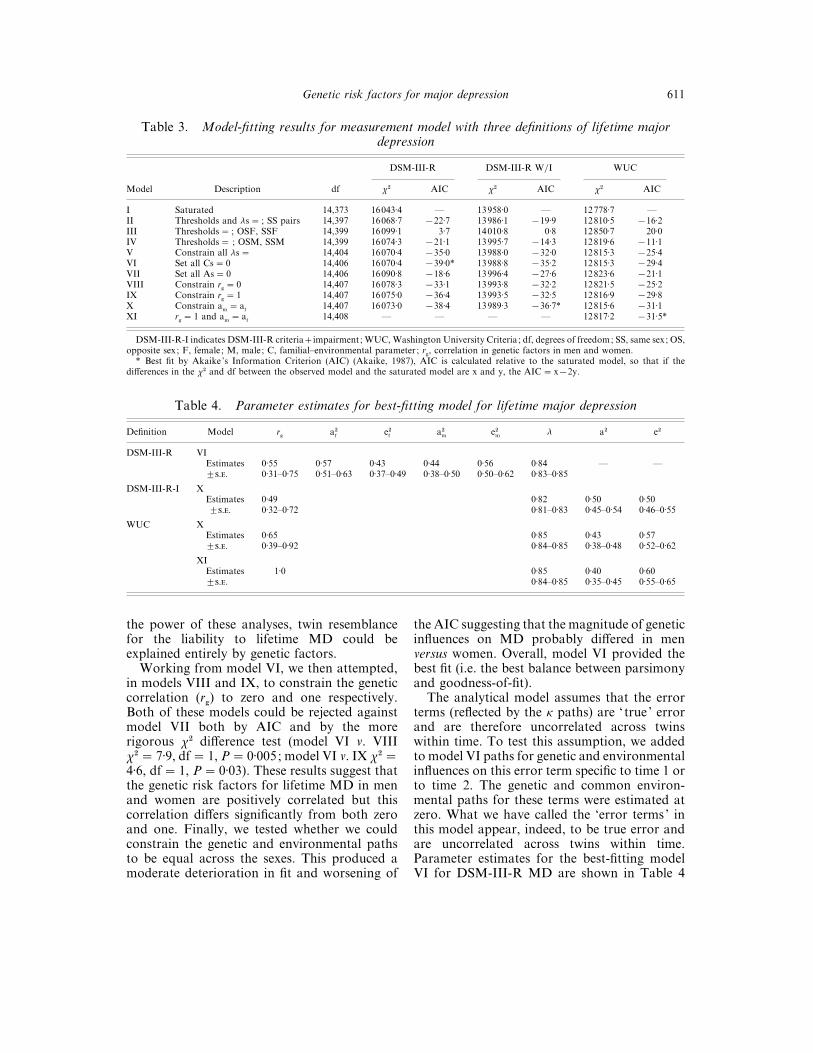
Genetic risk factors for major depression 611
Table 3. Model-fitting results for measurement model with three definitions of lifetime majordepression
Model Description df
DSM-III-R DSM-III-R W}I WUC
χ# AIC χ# AIC χ# AIC
I Saturated 14,373 16043±4 — 13958±0 — 12778±7 —II Thresholds and λs¯ ; SS pairs 14,397 16068±7 ®22±7 13986±1 ®19±9 12810±5 ®16±2III Thresholds¯ ; OSF, SSF 14,399 16099±1 3±7 14010±8 0±8 12850±7 20±0IV Thresholds¯ ; OSM, SSM 14,399 16074±3 ®21±1 13995±7 ®14±3 12819±6 ®11±1V Constrain all λs¯ 14,404 16070±4 ®35±0 13988±0 ®32±0 12815±3 ®25±4VI Set all Cs¯ 0 14,406 16070±4 ®39±0* 13988±8 ®35±2 12815±3 ®29±4VII Set all As¯ 0 14,406 16090±8 ®18±6 13996±4 ®27±6 12823±6 ®21±1VIII Constrain r
g¯ 0 14,407 16078±3 ®33±1 13993±8 ®32±2 12821±5 ®25±2
IX Constrain rg¯ 1 14,407 16075±0 ®36±4 13993±5 ®32±5 12816±9 ®29±8
X Constrain am
¯ af
14,407 16073±0 ®38±4 13989±3 ®36±7* 12815±6 ®31±1XI r
g¯ 1 and a
m¯ a
f14,408 — — — — 12817±2 ®31±5*
DSM-III-R-I indicates DSM-III-R criteriaimpairment ; WUC, Washington University Criteria ; df, degrees of freedom; SS, same sex; OS,opposite sex; F, female; M, male; C, familial–environmental parameter ; r
g, correlation in genetic factors in men and women.
* Best fit by Akaike’s Information Criterion (AIC) (Akaike, 1987), AIC is calculated relative to the saturated model, so that if thedifferences in the χ# and df between the observed model and the saturated model are x and y, the AIC¯ x®2y.
Table 4. Parameter estimates for best-fitting model for lifetime major depression
Definition Model rg
a#f
e#f
a#m
e#m
λ a# e#
DSM-III-R VIEstimates 0±55 0±57 0±43 0±44 0±56 0±84 — —³.. 0±31–0±75 0±51–0±63 0±37–0±49 0±38–0±50 0±50–0±62 0±83–0±85
DSM-III-R-I XEstimates 0±49 0±82 0±50 0±50³.. 0±32–0±72 0±81–0±83 0±45–0±54 0±46–0±55
WUC XEstimates 0±65 0±85 0±43 0±57³.. 0±39–0±92 0±84–0±85 0±38–0±48 0±52–0±62
XIEstimates 1±0 0±85 0±40 0±60³.. 0±84–0±85 0±35–0±45 0±55–0±65
the power of these analyses, twin resemblancefor the liability to lifetime MD could beexplained entirely by genetic factors.
Working from model VI, we then attempted,in models VIII and IX, to constrain the geneticcorrelation (r
g) to zero and one respectively.
Both of these models could be rejected againstmodel VII both by AIC and by the morerigorous χ# difference test (model VI v. VIIIχ#¯ 7±9, df¯ 1, P¯ 0±005; model VI v. IX χ#¯4±6, df¯ 1, P¯ 0±03). These results suggest thatthe genetic risk factors for lifetime MD in menand women are positively correlated but thiscorrelation differs significantly from both zeroand one. Finally, we tested whether we couldconstrain the genetic and environmental pathsto be equal across the sexes. This produced amoderate deterioration in fit and worsening of
the AIC suggesting that the magnitude of geneticinfluences on MD probably differed in menversus women. Overall, model VI provided thebest fit (i.e. the best balance between parsimonyand goodness-of-fit).
The analytical model assumes that the errorterms (reflected by the κ paths) are ‘ true’ errorand are therefore uncorrelated across twinswithin time. To test this assumption, we addedto model VI paths for genetic and environmentalinfluences on this error term specific to time 1 orto time 2. The genetic and common environ-mental paths for these terms were estimated atzero. What we have called the ‘error terms’ inthis model appear, indeed, to be true error andare uncorrelated across twins within time.Parameter estimates for the best-fitting modelVI for DSM-III-R MD are shown in Table 4

612 K. S. Kendler and others
AF AMEF EM
Liability to MDin females
Liability to MDin males
Lifetimehistory of MD
in femalesat time 1
Lifetimehistory of MD
in femalesat time 2
Lifetimehistory of MD
in malesat time 1
Lifetimehistory of MD
in malesat time 2
+0·55
0·750·660·750·66
0·840·84 0·840·84
0·540·540·540·54
F. 2. Parameter estimates for the best-fitting model (model VI) for lifetime major depression diagnosed using DSM-III-R criteria.
and Fig. 2. The heritability of liability to MD,corrected for unreliability of measurement, wasestimated to be 0±57 in females versus 0±43 inmales. The genetic correlation (r
g) in liability to
MD in men and women was estimated at 0±55.The λ path was estimated at 0±84, indicating atest–retest correlation in liability of 0±71.
The results of model-fitting for DSM-III-RI were similar to those found for DSM-III-R criteria. In particular, using these narrowercriteria, it remained possible to reject the modelthat constrained r
gto unity (model VI v. IX
χ#¯ 4±7, df¯ 1, P¯ 0±03). However, using theDSM-III-RI criteria, model X which con-strained the genetic and environmental paths tobe equal across genders provided an improve-ment in fit over model VI. Parameter estimatesfrom the best-fit model for these narrowercriteria (Table 4) produced a very similarestimate for r
g(0±49), but now the heritability
was estimated to be the same in both sexes andequal to 0±50.
Model-fitting results using WU criteria weresimilar to those found with DSM-III-RIcriteria with one exception. Constraining r
gto
unity now produced a deterioration in modelfitting of only 1±6 χ# units (model VI v. IX). So,by AIC criteria, model IX was slightly superiorto model VI. We therefore fitted an additional
model (model XI) in which both rg
was con-strained to unity and heritabilities were set equalacross the genders. This was, by a slight margin,the overall best fit model. We present parameterestimates, using the WU criteria, for both modelsX and XI. In model X, heritability was slightlylower and r
gslightly higher than that found with
the broader diagnostic criteria.
DISCUSSION
Using the increased power that comes with twooccasions of measurement of lifetime MD, weattempted to clarify whether the role of geneticfactors in the aetiology of MD was of similar ordiffering magnitudes in men and women andwhether the same genetic risk factors for MDwere operative in the two sexes. We attempted toanswer this question for three definitions of MDof varying stringency. We examine each of thosequestions in turn.
Heritability of lifetime major depression in menand women
We found that, when estimated from twooccasions of measurement, the heritability ofliability to lifetime MD using DSM-III-R criteriawas over 30% greater in women than in men. Inour previous data from this sample based on a

Genetic risk factors for major depression 613
single occasion of measurement, we found atrend in the same direction (Kendler & Prescott,1999; Kendler et al. 1999), but the best-fit modelconstrained heritability to be equal in the twosexes. In a combined clinical and population-based twin sample from Sweden, we also foundno evidence for differences in heritability ofaffective illness in men versus women (Kendler etal. 1993c, 1995). McGuffin et al. obtained similarresults for lifetime MD in a British large hospital-based twin sample (McGuffin et al. 1996).However, our results are consistent with a recentreport from the large Australian volunteer twinregistry where the best-fit model estimated theheritability of lifetime MD by DSM-III-Rcriteria to be 0±44 in women and 0±24 in men(Bierut et al. 1999). One model-fitting study offamily data found substantially higher ‘ trans-missibility ’ for MD in women than in men(Reich et al. 1987).
However, when we applied more stringentdiagnostic criteria for lifetimeMD toour sample,the best-fit model constrained heritability to beequal across the sexes. Such a change in the best-fit model could be due to a reduction in powerresulting from the increasing rarity of thedisorder and}or increasingly similar estimatesfor the heritability of MD in men and women inthe unconstrained model. Using DSM-III-Rcriteria, estimates for a# in men and women inmodel VI were 0±57 and 0±43, respectively. In thesame model, the estimates using DSM-III-RIand WU criteria in men and women were,respectively, 0±54}0±47 and 0±46}0±40. In both ofthe more restrictive criteria, there remainedsome evidence of greater heritability of MD inwomen. However, consistent with power studiesshowing that differences in heritability of under0±10 across genders will be very difficult to detectwith realistic sample sizes (Prescott &Gottesman, 1993), these differences were notsignificant. The most plausible explanation ofthese findings is that a true difference inheritability of MD across genders exists and it isgreater with broader criteria. However, wecannot rule out the hypothesis that no sucheffect exists and it is detected statistically withone out of three criteria because of chanceeffects.
In both this and previous studies (Kendler etal. 1992a, 1993a, b ; Kendler & Prescott, 1999),we found that heritability estimates for lifetime
MD are unlikely to be biased by excess sharingof etiologically relevant environmental variablesin MZ v. DZ twins. We have, in analysing earlierwaves of assessment in our female–femalesample, obtained two previous estimates usingDSM-III-R criteria for this ‘error-correctedheritability ’ that were slightly higher than thatobtained here : 0±71 (Kendler et al. 1993d ) and0±66 (Foley et al. 1998). Our results are notconsistent with the clinical intuition that morenarrowly defined MD is more heritable. Con-sistent with our prior results in females (Kendleret al. 1992a), heritability ofMDactually declinedmodestly with more stringent definitions ofillness.
The relationship between the genetic riskfactors for MD in men and women
An examination of the similarity of genetic riskfactors for MD in the sexes requires comparingthe degree of resemblance in opposite-sex versussame-sex relatives. If the same familial riskfactors are operative in men and women, thenthe resemblance among same-sex versusopposite-sex relative pairs should be the same.However, if some of these familial risk factorsdiffer, then the resemblance would be system-atically greater in same-sex compared toopposite-sex twin pairs.
A number of family studies have examinedthis question and most, but not all, foundgreater resemblance for lifetime MD in same-sexversus opposite-sex sibling or parent–offspringpairs (Merikangas et al. 1985; Faraone et al.1987; Reich et al. 1987). The Swedish twin studyreferred to above examined opposite-sex DZpairs (Kendler et al. 1993c) and could not rejecta model for the same genetic risk factors for MDin men and women (Kendler et al. 1995).
In our earlier study based on one time ofmeasurement (Kendler & Prescott, 1999;Kendler et al. 1999) and using only DSM-III-Rcriteria, our best-fit model estimated the geneticcorrelation (r
g) for MD between men and women
to equal 0±52. However, when we constrainedrg
to equal unity, the change in model fit wasonly 2±4 χ# units. While such a change is enoughto produce a deterioration in the AIC, it wouldnot be significant by more traditional χ# criteria.It is therefore noteworthy that with the additionof a second occasion of measurement, the samemodel fitting step now resulted in a change that

614 K. S. Kendler and others
was approximately twice as great (4±6 χ# units)and about which it is possible to be considerablymore confident. It is also reassuring that ourestimate of the r
gfor lifetime MD in men and
women (0±55) is similar to what we foundpreviously. Despite the large sample studied andthe use of two times of assessment, however, it ishumbling to note that the accuracy of ourestimate of this critical correlation is quitemodest.
Adding impairment to the DSM-III-R criteriafor MD produced little change in relationshipbetween genetic risk factors in men and women.The initial estimate of r
gwas a bit lower (0±49)
and constraining it to unity resulted in a similardeterioration in fit (4±7 χ# units). These resultssuggest that the magnitude of sharing of geneticrisk factors for MD across genders is not highlydependent on diagnostic stringency.
However, the results using the WU criteriawere different. The initial estimate of r
gwas
higher (0±65) and constraining it to unityproduced such a modest change in fit (1±6 χ#
units) that by AIC criteria the constrained modelwas the preferred one. The difference in resultsbetween the DSM-III-RI and WU criteria inthis regard is somewhat puzzling. It is possiblethat the results are ‘real ’ and derive fromdifferences in the diagnostic criteria. However,given the widely over-lapping confidenceintervals for the estimates of r
gacross the three
criteria sets (Table 4), it is more likely that theseresults reflect stochastic variation and decreasingstatistical power with more restrictive diagnosticsystems. We would conclude that, consistentwith our earlier results with DSM-III-R criteriaand one occasion of measurement (Kendler &Prescott, 1999; Kendler et al. 1999), thesefindings using two times of measurement furthersuggest that the genetic risk factors for MD inmen and women – while positively correlated (inthe range of 0±5 to 0±6) – are not identical.
If correct, this conclusion leads to a furtherintriguing question: what is the basis for thegender difference in genetic risk factors for MD?As recently reviewed by Bebbington (1996), menand women are exposed to a range of differentsocial and psychological factors of aetiologicalrelevance for MD. Might these differentexposures bring forth distinct genetic variationin the two sexes? Alternatively, biologicalfactors, including levels of gonadal hormones
during early development (Collaer & Hines,1995) and the variable hormonal environment ofthe menstrual cycle and pregnancy (Kendler etal. 1992b ; Seeman, 1997) could elicit discretegenetic variation in women and men (Garcia etal. 1989). Finally, the ‘psychopathologic path-way’ to MD in men and women may differ inways, which could explain the observed geneticcorrelation. Men may often develop MDthrough an ‘externalizing pathway’ via anti-social personality, alcoholism and drug abuse,all of which are heritable (Mednick et al. 1984;Begleiter & Kissin, 1995; Lyons et al. 1995;Tsuang et al. 1996). In women, by contrast,internalizing syndromes such as anxiety dis-orders, which are also heritable (Kendler et al.1992c, d, 1993d ), may be important precursorsto MD (Breslau et al. 1995).
Limitations
This study should be interpreted in the contextof four potential methodological limitations.First, the mode of assessment of this sample washeterogeneous including both face-to-face andtelephone interviews. However, both we(Kendler et al. 1992a) and others (Aneshensel etal. 1982; Sobin et al. 1993; Rohde et al. 1997)have found little difference in the assessment ofpsychiatric disorders by these two modes. Ourability to equate the reliability (λ) paths in themale–male twin sample across wave 1 (done byphone) and wave 2 (done mostly face-to-face)provides further evidence for the equivalence ofthese two assessment methods.
Secondly, these analyses did not excludeindividuals who experienced manic episodes. Inour female–female twins, removing cases whomet criteria for bipolar illness produced verysmall effects on prevalence and heritability ofMD (Karkowski & Kendler, 1997). In thissample, we repeated the analyses with the DSM-III-R criteria eliminating pairs with a history ofmania. No change was seen in the best-fittingmodel and the parameter estimates were verysimilar.
Thirdly, our estimates of lifetime prevalencefor DSM-III-R MD were substantially greaterthan those reported in the National ComorbiditySurvey (NCS) (Kessler et al. 1994) although theywere lower than that reported in at least oneother population based study (Rorsman et al.1990). In our interviews, the history of MD is

Genetic risk factors for major depression 615
assessed separately for the last-year and lifetimeprior to last year MD thus providing two‘chances ’ for an individual to meet criteria.While the NCS utilized lay-interviewers anda highly structured psychiatric interview, anapproach which may underestimate the popu-lation rates of illness (Helzer et al. 1985), weemployed experienced clinicians and a semi-structured interview. Like the NCS, we utilizedmethods to encourage ‘effortful responding’.Our subjects were somewhat younger than thoseassessed in the NCS which may also contributeto prevalence differences (Klerman et al. 1985).Recent studies have raised the question ofwhether, compared to clinician assessments, layinterviewers using highly structured instrumentsmay underestimate the rates of depressive illnessin community samples (Eaton et al. 2000;Murphy et al. 2000).
Fourthly, the mean inter-interview intervalwas over 50% greater in the female–female thanin the male–male sample. However, we wereable to constrain our λ estimates across genderswith little change in the model indicating thatthe overall reliability did not differ substantiallybetween the two samples.
This work was supported by NIH grants MH}AA-49492 and AA}DA-09095, Research Scientist Awardsto K.S.K. (MH-01277) and M.C.N. (MH-01458)and the Rachel Banks endowment fund. We ac-knowledge the contribution of the Virginia TwinRegistry, now part of the Mid-Atlantic Twin registry(MATR), to ascertainment of subjects for this study.The MATR, directed by Drs L. Corey and L.Murrelle, has received support from the NationalInstitutes of Health, the Carman Trust and the W. M.Keck, John Templeton and Robert Wood JohnsonFoundations.
REFERENCES
Akaike, H. (1987). Factor analysis and AIC. Psychometrika 52,317–332.
American Psychiatric Association (1987). Diagnostic and StatisticalManual of MentalDisorders, 3rd edn.Revised. AmericanPsychiatricAssociation: Washington, DC.
American Psychiatric Association (1994). Diagnostic and StatisticalManual of Mental Disorders, 4th edn. American PsychiatricAssociation: Washington, DC.
Aneshensel, C. S., Frerichs, R. R., Clark, V. A. & Yokopenic, P. A.(1982). Measuring depression in the community: a comparison oftelephone and personal interviews. Public Opinion Quarterly 46,110–121.
Bebbington, P. (1996). The origins of sex differences in depressivedisorder : bridging the gap. International Review of Psychiatry 8,295–332.
Begleiter, H. & Kissin, B. (eds.) (1995). The Genetics of Alcoholism.
Oxford University Press : New York.
Bertelsen, A., Harvald, B. & Hauge, M. (1977). A Danish twin study
of manic-depressive disorders. British Journal of Psychiatry 130,
330–351.
Bierut, L. J., Heath, A. C., Phil, D., Bucholz, K. K., Dinwiddie,
S. H., Madden, P. A. F., Statham, D. J., Dunne, M. P. & Martin,
N. G. (1999). Major depressive disorder in a community-based
twin sample. Archives of General Psychiatry 56, 557–563.
Boyd, J. H. & Weissman, M. M. (1981). Epidemiology of affective
disorders : reexamination and future directions. Archives of General
Psychiatry 38, 1039–1046.
Bresliau, N., Schultz, L. & Peterson, E. (1995). Sex differences in
depression: a role for preexisting anxiety. Psychiatry Research 58,
1–12.
Cadoret, R. J., O’Gorman, T. W., Heywood, E. & Troughton, E.
(1985). Genetic and environmental factors in major depression.
Journal of Affective Disorders 9, 155–164.
Collaer, M. & Hines, M. (1995). Human behavioral sex differences :
a role for gonadal hormones during early development. Psycho-
logical Bulletin 118, 55–107.
Eaton, W. W., Neufeld, K., Chen, L.-S. & Cai, G. (2000). A
comparison of self-report and clinical diagnostic interviews for
depression: Diagnostic Interview Schedule and Schedules for
ClinicalAssessment inNeuropsychiatry in theBaltimore Epidemio-
logic Catchment Area follow-up. Archives of General Psychiatry
57, 217–222.
Falconer, D. S. (1965). The inheritance of liability to certain diseases,
estimated from the incidence among relatives. Annals of Human
Genetics 29, 51–76.
Faraone, S. V., Lyons, M. J. & Tsuang, M. T. (1987). Sex differences
in affective disorder : genetic transmission. Genetic Epidemiology 4,
331–343.
Feighner, J. P., Robins, E., Guze, S. B., Woodruff, R. A., Winokur,
G. & Munoz, R (1972). Diagnostic criteria for use in psychiatric
research. Archives of General Psychiatry 26, 57–63.
Feild, H. S. & Armenakis, A. A. (1974). On use of multiple tests of
significance in psychological research. Psychological Reports 35,
427–431.
Foley, D. L., Neale, M. C. & Kendler, K. S. (1998). Reliability of a
lifetime history of major depression: implications for heritability
and co-morbidity. Psychological Medicine 28, 857–870.
Garcia, T., Sanchez, M., Cox, J. L., Shaw, P. A., Ross, J. B. A.,
Lehrer, S. & Schachter, B. (1989). Identification of a variant form
of the human estrogen receptor with an amino acid replacement.
Nucleic Acids Research 20, 8364.
Helzer, J. E., Robins, L. N., McEvoy, L. T. & Spitznagel, E. (1985).
A comparison of clinical and diagnostic interview schedule
diagnoses. Archives of General Psychiatry 42, 657–666.
Karkowski, L. M. & Kendler, K. S. (1997). An examination of the
genetic relationship between bipolar and unipolar illness in an
epidemiologic sample. Psychiatric Genetics 7, 159–163.
Kendler, K. S. (1993). Twin studies of psychiatric illness : current
status and future directions. Archives of General Psychiatry 50,
905–915.
Kendler, K. S. & Prescott, C. A. (1999). A population-based twin
study of lifetime major depression in men and women. Archives of
General Psychiatry 56, 39–44.
Kendler, K. S., Neale, M. C., Kessler, R. C., Heath, A. C. & Eaves,
L. J. (1992a). A population-based twin study of major depression
in women: the impact of varying definitions of illness. Archives of
General Psychiatry 49, 257–266.
Kendler, K. S., Silberg, J. L., Neale, M. C., Kessler, R. C., Heath,
A. C. & Eaves, L. J. (1992b). Genetic and environmental factors in
the aetiology of menstrual, premenstrual and neurotic symptoms:
a population-based twin study. Psychological Medicine 22, 85–100.
Kendler, K. S., Neale, M. C., Kessler, R. C., Heath, A. C. & Eaves,
L. J. (1992c). Generalized anxiety disorder in women: a population
based with study. Archives of General Psychiatry 49, 267–272.

616 K. S. Kendler and others
Kendler, K. S., Neale, M. C., Kessler, R. C., Heath, A. C. & Eaves,
L. J. (1992d ). The genetic epidemiology of phobias in women: the
inter-relationship of agoraphobia, social phobia, situational phobia
and simple phobia. Archives of General Psychiatry 49, 273–281.
Kendler, K. S., Neale, M. C., Kessler, R. C., Heath, A. C. & Eaves,
L. J. (1983a). A longitudinal twin study of 1-year prevalence of
major depression in women. Archives of General Psychiatry 50,
843–852.
Kendler, K. S., Neale, M. C., Kessler, R. C., Heath, A. C. & Eaves,
L. J. (1993b). A test of the equal environment assumption in twin
studies of psychiatric illness. Behavior Genetics 23, 21–27.
Kendler, K. S., Pedersen, N. L., Johnson, L., Neale, M. C. & Mathe,
A. A. (1993c). A pilot Swedish twin study of affective illness,
including hospital-and population-ascertained subsamples.
Archives of General Psychiatry 50, 699–706.
Kendler, K. S., Neale, M. C., Kessler, R. C., Heath, A. C. & Eaves,
L. J. (1993d ). Panic disorder in women: a population based twin
study. Psychological Medicine 23, 397–406.
Kendler, K. S., Neale, M. C., Kessler, R. C., Heath, A. C. & Eaves,
L. J. (1993e). The lifetime history of major depression in women:
reliability of diagnosis and heritability. Archives of General
Psychiatry 50, 863–870.
Kendler, K. S., Pedersen, N. C., Neale, M. C. & Mathe, A. A. (1995).
A pilot Swedish twin study of affective illness including hospital-
and population-ascertained subsamples : results of model fitting.
Behavior Genetics 25, 217–232.
Kendler, K. S., Karkowski, L. M. & Neale, M. C. (1998). A
longitudinal population-based twin study of retrospectively
reported premenstrual symptoms and lifetime major depression
American Journal of Psychiatry 155, 1234–1240.
Kendler, K. S., Gardner, C. O. & Prescott, C. A. (1999). Corrections
to 2 prior published articles. Archives of General Psychiatry 57,
94–95.
Kessler, R. C., McGonagle, K. A., Swartz, M., Blazer, D. & Nelson,
C. B. (1993). Sex and depression in the National Comorbidity
Survey: 1. Lifetime prevalence, chronicity, and recurrence. Journal
of Affective Disorders 29, 85–96.
Kessler, R. C., McGonagle, K. A., Zhao, S., Nelson, C. B., Hughes,
M., Eshleman, S., Wittchen, H.-U. & Kendler, K. S. (1994).
Lifetime and 12-month prevalence of DSM-III-R psychiatric
disorders in the United States : results from the National
Comorbidity Survey. Archives of General Psychiatry 51, 8–19.
Klerman, G. L., Lavori, P. W., Rice, J., Reich, T., Endicott, J. &
Andreasen, N. C. (1985). Birth-cohort trends in rates of major
depressive disorder among relatives of patients with affective
disorder. Archives of General Psychiatry 42, 689–693.
Little, R. J. A. & Rubin, D. B. (1987). Statistical Analysis with
Missing Data. Wiley and Son: New York.
Loehlin, J. C. & Nichols, R. C. (1976). Heredity, Environment and
Personality: A Study of 850 Sets of Twins. University of Texas
Press : Austin, TX.
Lyons, M. J., True, W. R., Eisen, S. A., Goldberg, J., Meyer, J. M.,
Faraone, S. V., Eaves, L. J. & Tsuang, M. T. (1995). Differential
heritability of adult and juvenile antisocial traits. Archives of
General Psychiatry 52, 906–915.
McGuffin, P., Katz, R., Watkins, S. & Rutherford, J. (1996). A
hospital-based twin register of the heritability of DSM-IV unipolar
depression. Archives of General Psychiatry 53, 129–136.
Mednick, S. A., Gabrielli, W. F. Jr. & Hutchings, B. (1984). Genetic
influences in criminal convictions: evidence from an adoption
cohort. Science 224, 891–894.
Merikangas, K. R., Weissman, M. M. & Pauls, D. L. (1985). Geneticfactors in the sex ratio of major depression. Psychological Medicine15, 63–69.
Murphy, J. M., Monson, R. R., Laird, N. M., Sobol, A. M. &Leighton, A. H. (2000). A comparison of diagnostic interviews fordepression in the Stirling County Study. Archives of GeneralPsychiatry 57, 230–236.
Neale, M. C. (1991). Statistical Modelling with Mx. Box 980126,Richmond VA 23298: Department of Psychiatry. Richmond: VA.
Neale, M. C., Sullivan, P. & Kendler, K. S. (1997). Multiplemeasurements increase the statistical power of genetic studies.Poster presented at the International Society for PsychiatricGenetics Annual Meeting.
Pearson, K. (1901). Mathematical contributions to the theory ofevolution. VIII. On the correlation of characters not quantitativelymeasurable. Proceedings of the Royal Society 66, 241–244.
Prescott, C. A. & Gottesman, I. I. (1993). Power limitations indetecting heterogeneity of genetic effects : the case of sex differencesin alcoholism. Presented at the annual meeting of the Society forResearch on Psychopathology, Chicago. October, 1993.
Prusoff, B. A., Merikangas, K. R. & Weissman, M. M. (1988).Lifetime prevalence and age of onset of psychiatric disorders :recall 4 years later. Journal of Psychiatric Research 22, 107–117.
Reich, T., Van Eerdewegh, P., Rice, J., Mullaney, J., Endicott, J. &Klerman, G. L. (1987). The familial transmission of primary majordepressive disorder. Journal of Psychiatric Research 21, 613–624.
Rice, J. P., Rochberg, M., Endicott, J., Lavori, P. W. & Miller, C.(1992). Stability of psychiatric diagnoses : an application to theaffective disorders. Archives of General Psychiatry 49, 824–830.
Rohde, P., Lewinsohn, P. M. & Seeley, J. R. (1997). Comparabilityof telephone and face-to-face interviews in assessing Axis I and IIdisorders. American Journal of Psychiatry 154, 1593–1598.
Rorsman, B., Grasbeck, A., Hagnell, O., Lanke, J., Ohman, R.,Ojesjo, L. & Otterbeck, L. (1990). A prospective study of first-incidence depression: the Lundby study, 1957–1972. British Journalof Psychiatry 156, 336–342.
Seeman, M. V. (1997). Psychopathology in women and men; focuson female hormones. American Journal of Psychiatry 154,1641–1647.
Sobin, C., Weissman, M. M., Goldstein, R. B., Adams, P.,Wickramaratne, P., Warner, V. & Lish, J. (1993). Diagnosticinterviewing for family studies : comparing telephone and face-to-face interview methods for the diagnosis of lifetime psychiatricdisorders. Psychiatric Genetics 3, 227–233.
Spitzer, R. L. & Williams, J. B. W. (1985). Structured ClinicalInterview for DSM-III-R (SCID). Biometrics Research Depart-ment, New York State Psychiatric Institute : New York.
Tsuang, M. T., Lyons, M. J., Eisen, S. A., Goldberg, J., True, W.,Meyer, J. M. & Eaves, L. J. (1996). Genetic influences on abuse ofillicit drugs: a study of 3,297 twin pairs. American Journal ofMedical Genetics 67, 473–477.
Weissman, M. M., Bruce, M. L., Leaf, P. J., Florio, L. P. & Holzer,C. (1991). Affective disorders. In Psychiatric Disorders in America:The Epidemiologic Catchment Area Study (ed. L. N. Robins andD. A. Regier), pp. 53–80. The Free Press : New York.
Williams, J. B. W., Gibbon, M., First, M. B., Spitzer, R. L., Davies,M., Borus, J., Howes, M. J., Kane, J., Pope, H. G., Jr., Rounsaville,B. & Wittchen, H.-U. (1992). The Structured Clinical Interview forDSM-III-R (SCID). II. multisite test–retest reliability. Archives ofGeneral Psychiatry 49, 630–636.
Williams, L. J. & Holahan, P. J. (1994). Parsimony-based fit indicesfor multiple-indicator models : do they work? Structural EquationModeling 1, 161–189.



















