
Genetic Parameters for Common Health Disorders of Holstein Cows
10
GENETICS AND BREEDING Genetic Parameters for Common Health Disorders of Holstein Cows H. A. URIBE,1 B. W. KENNEDY> S. W. MARTINf and D. F. KELTON2 University of Guelph Guelph, ON, Canada N1G 2W1 ABSTRACT Observations on 7416 Canadan Hol- stein cows were examined to estimate genetic parameters for the most common diseases of dairy cows. Mastitis, ovarian cyst, ketosis, milk fever, abomasal dis- placement, and culling that is due to reproductive failure or leg problems were analyzed as binomial traits, assum- ing an underlying threshold model that included fixed and random effects. Sire and residual components of variance were estimated by REML to provide heritability estimates from paternal half- sibs. A multiple-trait mixed model was also used to estimate genetic and en- vironmental correlations between production and disease traits. Heritabili- ties of disease traits were relatively low and ranged from 0 to .15, except for displaced abomasum (h?- = .28). Evidence of genetic antagonism existed between incidence of mastitis and milk produc- tion. Incidence of milk fever was geneti- cally associated with cows of lower genetic potential for production. Genetic associations between displaced aboma- sum and production traits were small, and estimates of genetic correlations be- tween ovarian cyst and milk production were inconsistent across lactations. Keto- sis was antagonistically associated genet- ically with production of milk and fat but was favorably associated with production of protein. The long-term cumulative ef- fect of genetic selection against diseases might be useful to diminish their inci- dence. (Key words: heritability, genetic correla- tion, milk production, disease incidence) Received June 10, 1994. Accepted September 19, 1994. Tentre for Genetic Improvement of Livestock. ZDepartment of Population Medicine. 1995 J Dairy Sci 78:421-430 INTRODUCTION Milk production has increased dramatically over the last two decades. However, this im- provement has not necessarily resulted in proportional increases of profits to dairy farm- ers. Cows with high milk production have relatively greater demands for labor (11) and increased frequency of health disorders (1 1, 25). As a result, some North American (26) and Scandinavian (28) dairy researchers have ques- tioned the selection of cows solely on milk production traits and have suggested that some emphasis be placed on other traits, such as health, that contribute to profitability. Inclu- sion of health traits in selection programs has been limited because of a lack of reliable data concerning health disorders. In addition, the discrete nature of most disease observations makes their statistical analysis and interpreta- tion more difficult (16). Antagonistic genetic correlations between milk production and disease traits indicate that disease incidence increases as a consequence of genetic improvement of milk production (27). The argument that losses from increased disease incidence can be compensated by addi- tional revenue from higher production has not been completely researched (26). Even if losses that were due to disease can be offset by additional production (11, 25, 34), ethical con- siderations might not allow researchers or producers to ignore the impact of selection for increased production on the health status and general welfare of cows (26, 29). Some attempts have been made, principally by Scandinavian researchers (9, 20, 28, 30), to examine genetic aspects of disease traits from large data files recorded in the field. In Canada, the genetic components of diseases in dairy cows, except for SCC, have not been widely investigated. The intent of this work was to estimate heritabilities of some common clinical diseases of Holstein dairy cows and to measure genetic and environmental correlations between dis- ease and production traits. 42 1
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Genetic Parameters for Common Health Disorders of Holstein Cows

GENETICS AND BREEDING
Genetic Parameters for Common Health Disorders of Holstein Cows
H. A. URIBE,1 B. W. KENNEDY> S. W. MARTINf and D. F. KELTON2 University of Guelph
Guelph, ON, Canada N1G 2W1
ABSTRACT
Observations on 7416 Canadan Hol- stein cows were examined to estimate genetic parameters for the most common diseases of dairy cows. Mastitis, ovarian cyst, ketosis, milk fever, abomasal dis- placement, and culling that is due to reproductive failure or leg problems were analyzed as binomial traits, assum- ing an underlying threshold model that included fixed and random effects. Sire and residual components of variance were estimated by REML to provide heritability estimates from paternal half- sibs. A multiple-trait mixed model was also used to estimate genetic and en- vironmental correlations between production and disease traits. Heritabili- ties of disease traits were relatively low and ranged from 0 to .15, except for displaced abomasum (h?- = .28). Evidence of genetic antagonism existed between incidence of mastitis and milk produc- tion. Incidence of milk fever was geneti- cally associated with cows of lower genetic potential for production. Genetic associations between displaced aboma- sum and production traits were small, and estimates of genetic correlations be- tween ovarian cyst and milk production were inconsistent across lactations. Keto- sis was antagonistically associated genet- ically with production of milk and fat but was favorably associated with production of protein. The long-term cumulative ef- fect of genetic selection against diseases might be useful to diminish their inci- dence. (Key words: heritability, genetic correla- tion, milk production, disease incidence)
Received June 10, 1994. Accepted September 19, 1994. Tentre for Genetic Improvement of Livestock. ZDepartment of Population Medicine.
1995 J Dairy Sci 78:421-430
INTRODUCTION
Milk production has increased dramatically over the last two decades. However, this im- provement has not necessarily resulted in proportional increases of profits to dairy farm- ers. Cows with high milk production have relatively greater demands for labor (11) and increased frequency of health disorders (1 1, 25). As a result, some North American (26) and Scandinavian (28) dairy researchers have ques- tioned the selection of cows solely on milk production traits and have suggested that some emphasis be placed on other traits, such as health, that contribute to profitability. Inclu- sion of health traits in selection programs has been limited because of a lack of reliable data concerning health disorders. In addition, the discrete nature of most disease observations makes their statistical analysis and interpreta- tion more difficult (16).
Antagonistic genetic correlations between milk production and disease traits indicate that disease incidence increases as a consequence of genetic improvement of milk production (27). The argument that losses from increased disease incidence can be compensated by addi- tional revenue from higher production has not been completely researched (26). Even if losses that were due to disease can be offset by additional production (11, 25, 34), ethical con- siderations might not allow researchers or producers to ignore the impact of selection for increased production on the health status and general welfare of cows (26, 29).
Some attempts have been made, principally by Scandinavian researchers (9, 20, 28, 30), to examine genetic aspects of disease traits from large data files recorded in the field. In Canada, the genetic components of diseases in dairy cows, except for SCC, have not been widely investigated.
The intent of this work was to estimate heritabilities of some common clinical diseases of Holstein dairy cows and to measure genetic and environmental correlations between dis- ease and production traits.
42 1

422 URIBE ET AL.
MATERIALS AND METHODS
Health data were provided by the Depart- ment of Population Medicine, University of Guelph, with cooperation from the Ontario DHI Corporation and the Ontario Milk Mar- keting Board. The information was recorded between 1989 and 1991 from herds in southern Ontario enrolled in a research project known as the Ontario Dairy Monitoring and Analysis Program. Its objective was to study relation- ships among measures for herds of disease, management, production, reproductive perfor- mance, and profitability in Ontario. The data were information on 7416 cows in 98 herds.
A cow was considered to be diseased at the first report of a particular hsease by the veterinarian or the farmer. The same disease shown more than once during a lactation was considered to be one incidence for analysis. Health traits studied were mastitis, ovarian cyst, ketosis, milk fever, displaced abomasum, and culling for reproductive failure or leg problems.
The production data used for analyses cor- responded to the same lactation in which the cow was recorded as having or not having the disease during a given lactation. Production traits were 305-d production of milk, fat, and protein and percentages of fat and protein. Production data were from Agriculture Canada from the data used for evaluation of bulls and cows in Canada and were adjusted for age and month of calving.
Clinical presence or absence of a specific disease generates discrete data with a binomial distribution. Disease observations may be grouped in two ordered categories (having or not having the disease), and susceptibility of cows to a certain disease follows an underlying continuous normal distribution that is not ob- served. Only cows that exceed some unknown threshold of susceptibility show clinical signs of the disease (6, 23). Gianola and Foulley (8) and Harville and Mee (12) proposed a non- linear set of equations that are solved itera- tively for the values of the thresholds and effects in the model for analysis of categorical data.
A FORTRAN program GFCATI (L. R. Schaeffer, 1992, personal communication) was used to compute the elements and to solve the nonlinear equations (8). Estimation of variance components was by REML (21).
A sire model was used to estimate heritabil- ity of each disease trait as follows:
where
Yijklm = Value of observation ijklm, j.4 = populat’ ion mean,
Hi = fixed effect of herd i, Cj = fixed effect of calving season j, Yk = fixed effect of year k, SI = random effect of sire 1 - N(0, 14).
e i j b = residual error - N(0, Id) . and
Because of the low incidence of diseases, inclusion of herd-year interaction in the model would have generated empty cells, and sub- classes containing empty cells must be elimi- nated from the analysis (12).
First lactation cows differ from multiparous cows; they produce less milk and have differ- ent rates of incidence for many diseases (19). Therefore, a separate analysis was performed for primiparous cows if 25% of primiparous cows were recorded with the disease. When the frequency of disease was 4% during first lactation, data for primiparous and multiparous cows were pooled to avoid loss of information by empty cells. For the combined analysis of all cows, the model also included the fixed effect of parity. Cows were only represented once; i.e., cows did not have records repeated over lactations. Only sires having at least five daughters with records in at least two herds were considered for analysis. Relationshps among sires were ignored. Heritability was calculated as the ratio of four times the sire component of the variance to one plus the sire variance (1).
To assess the genetic relationship between each health trait and the production of milk, fat, and protein and percentages of fat and protein, the data were analyzed using a two- trait mixed sire model for one continuous and one discrete trait using the method of Janss and Foulley (13). The method allowed joint analysis of the continuous and the discrete traits with unequal design matrices and could accommodate missing data on some traits. Es- timation of genetic effects was based on
Journal of Dairy Science Vol. 78, No. 2, 1995
GENETIC PARAMETERS FOR HEALTH DISORDERS 423
threshold model methodology (12, 13). The extension of the threshold model to include discrete and continuous variables was exam- ined by Foulley et al. (7).
The computations were performed using a FORTRAN program BIVTHM (L. Janss, 1993, personal communication). The statistical model used for the discrete traits was as indicated previously for estimation of heritability. The model used for the continuous traits was as follows:
where
yiju = production record (production of milk, fat, or protein or percentages of fat or protein) from the same lac- tation as the measure of disease oc- currence,
p = overall mean, HYi = fixed effect of the herd-year of calv-
Cj = fixed effect of the calving season j (i
Sk = random effect of the sire k - N(0, I
qjk = random error associated with obser-
ing i,
= 1 2 3 ,
d), and
vation ijkl - N(0, 1.2).
When data were analyzed across lactations, the fixed effect of parity was included. Sires having records on at least five daughters in
two or more herds were used for analysis. Relationships among sires were ignored.
Estimates of genetic correlations were com- puted from sire (co)variances. Environmental (co)variance was calculated as the residual (co)variance minus three times the sire (co)vari- ance. Environmental correlations between health and production traits were computed from environmental (co)variances.
RESULTS AND DISCUSSION
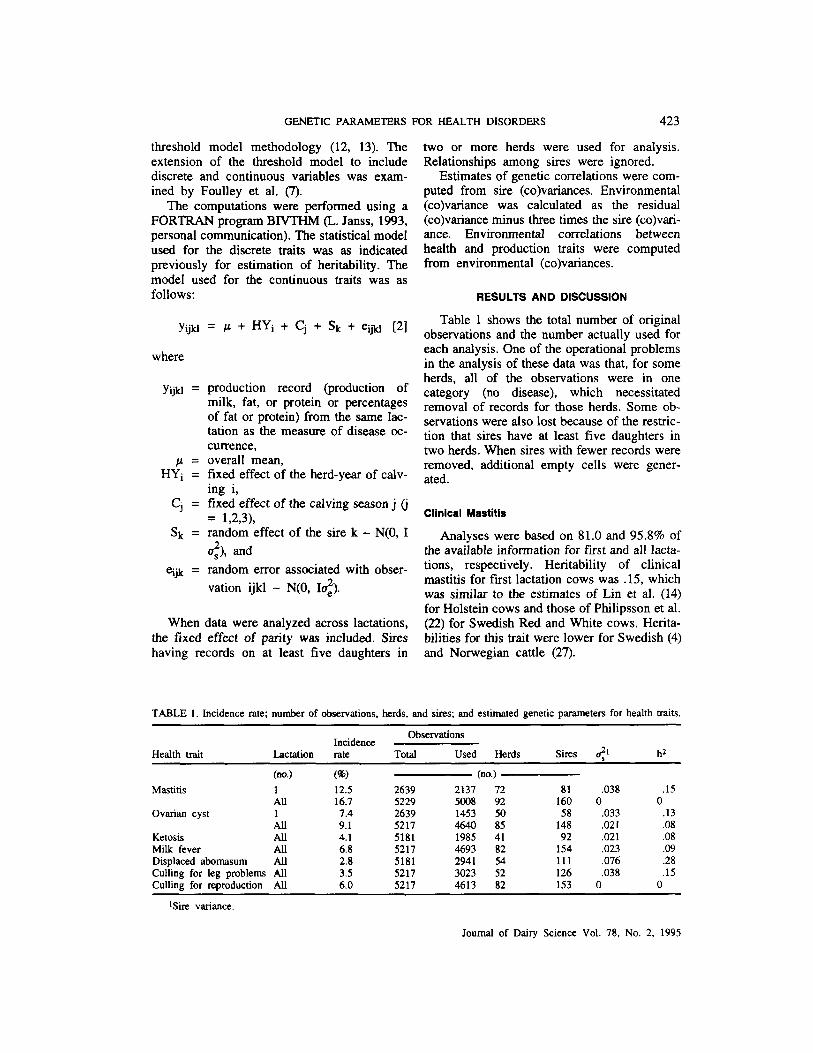
Table 1 shows the total number of original observations and the number actually used for each analysis. One of the operational problems in the analysis of these data was that, for some herds, all of the observations were in one category (no disease), which necessitated removal of records for those herds. Some ob- servations were also lost because of the restric- tion that sires have at least five daughters in two herds. When sires with fewer records were removed, additional empty cells were gener- ated.
Clinical Mastitis
Analyses were based on 81.0 and 95.8% of the available information for first and all lacta- tions, respectively. Heritability of clinical mastitis for first lactation cows was .15, which was similar to the estimates of Lin et al. (14) for Holstein cows and those of Philipsson et al. (22) for Swedish Red and White cows. Herita- bilities for this trait were lower for Swedish (4) and Norwegian cattle (27).
TABLE 1. Incidence rate; number of observations, herds, and sires; and estimated genetic parameters for health traits. ~~~ ~~~~
Observations Incidence
Health trait Lactation rate Total Used Herds sires 21 hz ~ ~~~ ~
(no.) (W (no.) MasUtis 1 12.5 2639 2137 72 81 ,038
All 16.7 5229 5008 92 160 0 ovarian cyst 1 7.4 2639 1453 50 58 .033
All 9.1 5217 4640 85 148 ,021 Ketosis All 4.1 5181 1985 41 92 ,021 Milk fever All 6.8 5217 4693 82 154 ,023 Displaced abomasum All 2.8 5181 2941 54 111 ,076 Culling for leg problems All 3.5 5217 3023 52 126 .038 Culling for reproduction All 6.0 5217 4613 82 153 0
*Sire variance.
.15
.13
.OS
.08
.09
0
.28
.15 0
Journal of Dairy Science Vol. 78, No. 2. 1995

424 URIBE ET AL.
TABLE 2. Genetic and environmental correlations between health and production traits.
Health wait
Roduction
Lactation Milk Fat Protein Fat Protein
Genetic correlations Mastitis ovarian cyst
Ketosis Milk fever Displaced abomasum Culling for leg problems
Environmental correlation Mastitis ovarian cyst
Ketosis Milk fever Displaced abomasum Culling for leg problems
1 1 All All All All All
I 1 All All All All Au
37 -.14 -.06
.77 -.67 -.a
.27
-.Ol .27 .20 .02 .I5
- . lo -.18
3 9 S O
-.12 .4 1
-.21 .18 .20
.03
.oo .19
-.02 .09
-.07 -.19
.20
. l l -.43 -.30 -.66 .09 .2 1
.02
.20
.25
.09
.I6 -.17 -.23
.21 -.I2
.76 .57 -.13 -.56
.21 -.79
.29 -.24
.24 .12
.20 .07
.03 .07 -.37 -.23 0 .15 -.12 .08 -.OS .04
.05 -.12 -.lo -.25
'First lactation. *All lactations.
Heritability of clinical mastitis for all cows was 0, which is in agreement with results of Dohoo et al. (3), but contrary to those of Lin et al. (14), who obtained a heritability estimate of .18 for cows after their second lactation. Heritability estimates of clinical mastitis across lactations of Holstein cows have ranged from low, .03, to moderate, .14 (15, 31).
Genetic and environmental correlations be- tween clinical mastitis during first lactation and production traits are presented in Table 2. Production of milk, fat, and protein was posi- tively correlated with incidence of mastitis for first lactation cows. The genetic correlation between mastitis and milk production of .37 for first lactation cows was similar to the .38 found by SyvajWi et al. (30) for first lactation Finnish cows. Higher estimates have been reported: Simianer et al. (27) found a correla- tion of -51 for primiparous Norwegan cows, and Syvajilrvi et al. (30) found a correlation of .57 across all lactations for Finnish Ayrshire cows. In contrast, Lyons et al. (15) estimated the genetic correlation between mastitis and milk production to be .18 across lactations, Thompson (31) found a correlation of .26 for Holstein cows across lactations, and Bunch et al. (2) reported a correlation of .21 for first lactation cows.
The estimated genetic correlation between fat production and mastitis for first lactation cows was .39, which is in contrast to results of Lyons et al. (15), who found a genetic correla- tion of 0. Genetic correlation between protein production and mastitis during first lactation in this study was .20 (Table 2).
Genetic correlation between fat percentage and clinical mastitis for f i s t lactation cows was .21, which is contrary to the genetic corre- lation of -.15 reported by Simianer et al. (27) for first lactation Norwegian cows. In contrast to fat percentage, the genetic correlation be- tween protein percentage and mastitis inci- dence was negative, -.12.
Environmental correlations (Table 2) be- tween mastitis and production traits during first lactation were all low. Environmental correlations between mastitis and percentages of fat and protein were low for first lactation Norwegian cows (27).
Heritabilities estimated from the multitrait analyses for first lactation mastitis and produc- tion traits are given in Table 3. Estimates of heritability of mastitis during f is t lactation were similar for each of the five analyses with production traits and were very similar to the .15 obtained from the single-trait analysis (Ta- ble l).
Journal of Davy Science Vol. 78, No. 2, 1995

GENETIC PARAMETERS FOR HEALTH DISORDERS 425
TABLE 3. Average heritabilities for health and production traits from multiple-trait models.
- Trait X
Health Mastitis ovarian cyst Ketosis Milk fever Displaced abomasum Culling for leg problems
Milk Fat Protein Fat, % Protein. %
Production
,146 ,168 . lo4 ,096 .304 ,146
,170 .245 .195 .362 .415
SD - .005 .049 ,031 .011 ,005 ,011
,057 ,065 ,077 ,025 .044
Range
.14-.I5
.12-.22
.09-. 16
.O&. 11
.30-.31
.13-.16
.10-.26
.1&.37
.13-.34
.34-.41
.37-.48
The heritability estimates of .15 for clinical mastitis during first lactation suggest consider- able genetic variability. Also, clinical mastitis for first lactation cows was positively cor- related with the three production traits, which supported the concern of other studies that continued selection for increased production, ignoring information on diseases, might be detrimental to long-term cow health. Despite the small antagonism between production and mastitis, its cumulative effect over several generations might be important.
Ovarian Cyst
For number of herds, no cases of ovarian cysts were reported; when ovarian cysts were reported, they occurred in the daughter of a sire with <5 daughters. Thus, the analyses used only 55.1 and 88.9% of the available informa- tion for first and all lactations. Sire compo- nents of estimates of variance and heritability for cystic ovarian disease are in Table 1.
Heritability of ovarian cyst was .13 for first lactation cows and .OS over all lactations. These estimates were very similar to respective estimates of .12 and .07 reported by Lin et al. (14) for US Holstein cows. The heritability estimate computed herein for ovarian cyst for first lactation cows is larger than that of .05 found by Erb et al. (5) and that of -01 found for ovulatory disorders of primiparous Finnish cows (17). However, in the latter study, the trait also included other infertility problems. Dohoo et al. (3) indicated that heritability of
ovarian cyst for Holstein cows was 0. Lyons et al. (15) found a heritability of .05 for ovarian cysts across lactations of Holstein cows.
Genetic and environmental correlations be- tween cystic ovarian disease and production traits are given in Table 2. For first lactation cows, the estimated genetic correlations be- tween ovarian cyst and production of milk, fat, and protein were -.14, S O , and .11, respec- tively. For all lactations, respective genetic correlation estimates were -.06, -.12, and -.43. Lyons et al. (15) estimated genetic correlations between cystic ovaries and production of milk and fat of -.01 and .24, respectively. M k - tysaari et al. (17) found genetic correlations between milk production and ovulatory disord- ers of .65 and .02 for Finnish cows during their first and second lactations, respectively. Genetic correlations (Table 2) between ovarian cyst and percentages of fat and protein for first lactation cows were high and positive, .76 and .57, but were negative over all lactations, -.13
Environmental correlations during first lac- tation were .27, 0, and .20 between cystic ovaries and production of milk, fat, and pro- tein, respectively, and were in a similar range over all lactations (Table 2). Environmental correlations between ovarian cysts and percen- tages of fat and protein were moderate and negative during first lactation, -.37 and -.23, respectively, and close to 0 over all lactations, 0 and .15, respectively.
Heritability estimates from the multitrait analysis for ovarian cyst and production traits are given in Table 3. Estimates ranged from .12 to .22. Mean heritability of the five esti- mates was .17. Heritability of ovarian cyst was similar to that for mastitis (.13). Genetic varia- tion of cystic ovaries might be higher than previously thought for other traits related to fertility.
Genetic correlation estimates between cystic ovaries and production traits were inconsistent except for those with milk production. The small and negative correlation with milk production (Table 2) indicated that selection for high volume of milk was not antagonistic or predisposing to the disease.
and -.56.
Ketosis
The frequency of clinical ketosis was very low for first lactation cows. Loss of informa-
Journal of Dairy Science Vol. 78, No. 2, 1995

426 URIBE ET AL.
tion because of empty cells provided too few data for first lactation cows for meaningful analyses. Therefore, sire components of vari- ance and the heritability of clinical ketosis are across all lactations in Table 1. Nevertheless, only 38.3% of the 5181 observations available were used for analysis.
Heritability of clinical ketosis was .09, which is in agreement with results of Philips- son et al. (22), who found heritabilities of .06 and .09, respectively, for first lactation Swed- ish Red and White and Swedish Friesian, and those of Lyons et al. (15). who reported a heritability for ketosis of .08 for US Holsteins. Heritabilities across lactations were similar for Finnish Ayrshires (9) and first lactation Nor- wegian cows (27). Estimates by Solbu (28) for Norwegian cows were smaller than that of the present study and averaged .02 over subsets of data corresponding to different years. Herita- bilities were smaller, .02, for Finnish Ayrshires (10) than those estimated in the present study. Recently, heritabilities for ketosis were esti- mated to be .02 and .03 for Finnish cows during their first and second lactations, respec- tively (18). Heritability for ketosis estimated in the present study, however, was much lower than estimates of .30 and .31 by Van Vleck (33) and Dohoo et al. (3). respectively.
Genetic and environmental correlations be- tween ketosis and production traits across lac- tations are in Table 2. The highest genetic correlation, .77, was between ketosis and milk production. This correlation was higher than the estimates of Grohn et al. (9) for first lacta- tion Finnish Ayrshire cows, .30, Lyons et al. (15) for US Holsteins, .26, and Mantysaari et al. (16) for primiparous Finnish cows, .17. However, this estimate was close to the .65 of Simianer et al. (27) for first lactation Nor- wegian cows. Genetic correlation between fat production and incidence of ketosis was .41. Protein production was moderately and nega- tively correlated, -.30, with the disease.
Ketosis was moderately and positively cor- related, .2 1 , with fat percentage (Table 2). The genetic correlation between protein percentage and ketosis was high and negative, -.79. Simi- aner et al. (27) reported genetic correlations between ketosis and percentages of fat and protein of -.38 and -.65, respectively. En- vironmental correlations between ketosis and the production traits were all small and ranged from -.02 to .09 (Table 2).
Journal of Dairy Science Vol. 78, No. 2, 1995
Heritability estimates from the multiple-trait analyses are in Table 3. Heritability of clinical ketosis was the same for four of the five analyses, .09, and was very similar to that found by the single-trait analysis. However, heritability from the multiple-trait analyses in- volving protein percentage was .16.
The 2% incidence of this metabolic disease was much lower than that in the literature. The genetic component of variation for ketosis across lactations was low @* = .W). his estimate is consistent with most of the litera- ture and indicates that environmental factors, particularly nutritional management, are more important in the short term for control of keto- sis than is genetics. Nonetheless, genetic varia- bility exists for ketosis.
Increased production of milk and fat were highly positively correlated with incidence of ketosis. This finding indicates that, even if heritability of ketosis is low, selection schemes based solely on production of milk and fat should result in a long-term increase in the frequency of clinical ketosis. However, ketosis and protein production showed a moderately negative genetic correlation, indicating that selection to favor protein production reduces incidence of ketosis.
Mllk Fever
Because of its low frequency, the trait for milk fever was also analyzed only across lacta- tions. Information from 154 sires with records in 82 herds were used for analysis, which represented 90% of the available information (Table 1). Incidence of milk fever in the final data was 319, which represented a rate of 6.1%. The sire variance component was .023, and heritability was .09.
Milk fever is a syndrome produced by a sudden decrease in concentration of Ca in plasma at calving. Heritability for milk fever across lactations in ths study was smaller than the estimate of Tveit et al. (32) based on postparturient concentration of Ca in plasma of Norwegian cows, although the traits are not exactly the same. Lyons et al. (15) found a much higher heritability estimate for milk fe- ver across lactations, as did Lin et al. (14), who found heritabilities for Holstein cows from parities 22 of .30 and .42, respectively. In contrast, Philipsson et al. (22) found no genetic

GENETIC PARAMETERS FOR HEALTH DISORDERS 427
variation for incidence of milk fever for two breeds of Swedish cows.
Genetic and environmental correlations be- tween milk fever and production traits are given in Table 2. The three production traits were negatively associated genetically with milk fever. Milk production and milk fever were highly negatively correlated, -.67. Grohn et al. (9) also found negative genetic correla- tion between parturient paresis and milk production, but their estimate was smaller, -.@. However, Tveit et al. (32) found an un- desirable negative correlation, -.5, between postpartum concentration of Ca in blood and milk production. The genetic association be- tween production of fat and protein and milk fever in this study were moderately to largely negative, -.21 and -.66, respectively, and larger than that found by Lyons et al. (15), -.01. These results suggest that cows with superior genetic potential for production of milk and protein might be more resistant to the disease. Genetic correlations between milk fe- ver and fat and protein percentage were .29 and -.24, respectively.
Pooled heritability of milk fever across the five multiple test analyses was .10 (Table 3) and was similar to that from the single-trait analysis (.09) (Table 1). All multiple-trait esti- mates were similar: .08 to .l 1.
Displaced Abomasum
Heritability of displaced abomasum across lactations was .28 (Table I), which was con- siderably higher than the .@ estimated by Lyons et al. (15). Genetic and environmental correlations between displaced abomasum and production traits are in Table 2. The genetic correlation between displaced abomasum and milk production in the present study was low and negative: -.04. Lyons et al. (15) also found a negative but larger genetic correlation, -. 15, between these traits across lactations for US Holsteins. The genetic correlation between fat production and displaced abomasum was .18, which is in disagreement with results of Lyons et al. (15), who found a negative association between these traits, -.28. Protein production and displaced abomasum were positively as- sociated, but the correlation was small, .@. Genetic correlations between displaced aboma- sum and fat and protein percentage were
moderate, .24, and small, .12, respectively. Similar to the analysis of heritability of dis- placed abomasum using the single-trait analy- sis (Table l), multiple-trait estimates of herita- bilities were high and consistent for all five analyses, .30 to .31 (Table 3).
Displaced abomasum has not been exten- sively studied genetically. Its heritability esti- mated from almost 3000 records was moderate (.28) and indicates that, if records were availa- ble, genetic selection could reduce the inci- dence of the disease. Long-term selection for fat production might tend to increase the fre- quency of displaced abomasum because of the small positive correlation, .18. Genetic correla- tions between displaced abomasum and production of milk and protein were very small, and the traits should be independent for selection.
Culling for Leg Problems
Culling for leg problems had a heritability of .15 across lactations (Table 1). No other references of heritability of this trait were found in the literature. Lyons et al. (15) esti- mated heritabilities for leg and foot problems at .08 and .11, respectively. Genetic correla- tions between culling for leg problems and production of milk, fat, and protein were posi- tive and moderate; environmental correlations were negative (Table 2). Genetic correlations between culling for leg problems and percen- tages of fat and protein were moderate, .20, and low, .07, respectively; corresponding en- vironmental correlations were negative.
Culling for leg problems would normally occur near the end of a lactation, and a nega- tive correlation might be expected unless leg problems were very severe, causing cows to be culled regardless of milk production. Genetic correlations between culling for leg problems and production of milk, fat, and protein were positive (Table 2), indicating that long-term selection for these traits might increase culling for impaired legs.
Culling for Reproductive Failure
Cows were culled from the herd for reduced reproductive performance. As with ketosis and milk fever, the incidence of this disorder was very low for first lactation cows; therefore, the analysis was only performed across lactations.
Journal of Dairy Science Vol. 78, No. 2, 1995

42 8 URIBE ET AL.
No genetic variation was found for this trait (Table 1). Although the literature does not contain many reports on this trait, heritabilities for individual reproductive traits were low (15, 17, 18, 33).
Because this trait showed no genetic varia- bility, selection would not have an important role in improvement of general fertility of dairy cattle. This conclusion is consistent with the literature, which indicates very low herita- bility of reproductive disturbances (15, 17).
Production Traits
Multiple-trait heritabilities of production traits are in Table 3. Estimates were similar to those of some previous reports (24, 27). The estimate for milk production tended to be low. Considerable variability occurred among ana- lyses for estimates of heritability. Estimates were obtained using a sire model and ignored relationships among sires.
General D~SCU88lOn
As with many of the other studies of health disorders, in this work the sample size used was small for determination of genetic rela- tionships. As with most other genetic studies related to disease, other shortcomings were that relationships among sires were ignored in the analyses, and standard errors of the esti- mates could not be computed. Ideally, these data should have been analyzed considering relationships among animals; unfortunately, computing packages to analyze binary data under an animal model were not available at the time of the study.
Also, data on multiparous cows are poten- tially subject to selection bias. Only cows with acceptable milk production and health status survive for later lactations, and such selection can influence estimates of genetic parameters. Unfortunately, it is difficult to correct for this selection when data are limited to a relatively short period and historical records on all cows are not available.
Direct comparison of results across studies is difficult with this type of data because the recording systems vary across studies and different traits are often recorded. The diagno- sis of a particular disease is somewhat subjec- tive. The categorization of a cow as diseased depends on the ability and experience of the
veterinarian, the farmer, or both in recognizing the disorder. Also disease recording was on the basis of treatment, particularly for mastitis, milk fever, and ketosis; i.e., treatments for disease were recorded, but the diagnosis of disease was not necessarily recorded. An over recording of milk fever may be implied; some cows in some herds are treated prophylacti- cally with Ca whether or not they show signs of hypocalcemia. However, mastitic cows that were considered by the owner to have mild clinical mastitis and were untreated were not necessarily recorded on the data sheet. This procedure undoubtedly resulted in an under- estimate of clinical mastitis incidence in some herds. An underlying feature of the present study is that all farms were serviced by veterinarians who were enrolled in the same continuing education program given by the Ontario Veterinary College.
A number of antagonistic correlations were suggested between disease traits and measures of milk production. Measurement of the true production potential of cows is also difficult, particularly if the diseases of concern are likely to influence production. In our study, milk fever, ketosis, and displaced abomasum oc- curred early in the lactation and could have had considerable impact on milk production during the rest of the lactation. This impact would be particularly important for cows with displaced abomasum because they would un- dergo a surgical procedure and the essential postsurgical treatments.
In general, most of the parameters estimated herein are within the range of those indicated in previous studies. The heritability estimate was highest, .28, for displaced abomasum. Al- though conclusions vary, genetic variation for most of the diseases was sufficient to be poten- tially useful in programs for genetic selection. With few exceptions, however, heritabilities were small; therefore, short-term management practices focused to provide an adequate en- vironment for the cows should continue to be the most important and logical means to pre- vent these diseases. However, the long-term cumulative effect of genetic selection could significantly affect the permanent improve- ment of disease resistance of dairy cows.
Some diseases are negatively correlated with production traits, indicating that selection schemes might have to trade some genetic
Journal of Dairy Science Vol. 78, No. 2, 1995

GENETIC PARAMETERS FOR HEALTH DISORDERS 429
improvement for production to gain health sta- tus. Because of the relatively low heritability of diseases, the current North American progeny-testing schemes, in which relatively few daughters of each sire are tested, do not allow for much effective emphasis on health- related traits for selection.
Feasibility of including diseases in indexes for dairy selection depends on the economic importance of a specific disease. Economic considerations might differ over countries or regions. Also, ethical considerations could re- quire the dairy industry to improve animal welfare by alleviation of health disturbances, although this might not be the best economic option.
Recently, statistical approaches to analyze discrete data have been developed to the extent that they can be applied to field data for ani- mal breeding, and enhanced computing tech- nology allows the application of these methods. However, in North America, pro- grams to collect data pertaining to diseases have not been as comprehensive as in Scan- dinavian countries.
CONCLUSIONS
Genetic variability, although relatively small, was sufficient for most diseases studied to be potentially useful in breeding programs. Genetic antagonisms between some diseases and production traits suggest the need to con- sider disease implications in long-term im- provement programs and possibly disease measures for selection.
ACKNOWLEDGMENTS
We thank L. R. Schaeffer and L. Janss for providing computer programs. Financial sup- port was provided by the Natural Sciences and Engineering Research Council of Canada and the Ontario Ministry of Agriculture and Food.
REFERENCES
1 Becker, W. A. 1964. Manual of Quantitative Genetics. 4th ed. Academic Enterprises, Pullman, WA.
2 Bunch, K. J.. D.J.S. Heneghan, K. G. Hibitt, and G. J. Rowls. 1984. Genetic influences on clinical mastitis and its relationship with milk yield, season, and stage of lactation. Livest. Prod. Sci. 11:91.
3Doh00, 1. R., S. W. Martin, 1. McMillan, and B. W. Kennedy. 1984. Disease, production, and culling in
Holstein-Friesian cows. 11. Age, season and sire ef- fects. Prev. Vet. Med. 2655.
4Emanuelson. U., B. Danell, and J. Philipsson. 1988. Genetic parameters for clinical mastitis, somatic cell counts, and milk production estimated by multiple- trait restricted maximum likelihood. J. Dairy Sci. 71: 467.
5Erb. R. E., P. M. Hinze, and E. M. Gildow. 1959. Factors influencing prolificacy of cattle. 11. Some evidence that certain reproductive traits are additively inherited. Bull. 30, Washington Agric. Exp. Stn., Cor- vallis.
6Falconer. D. S. 1989. Introduction to Quantitative Genetics. 3rd ed. Longman Sci. Tech., New York, NY.
7Foulley. J. L., D. Gianola, and R. Thompson. 1983. Prediction of genetic merit from data on binary and quantitative variates with an application to calving difficulty, birth weight, and pelvic opening. Genet. Sel. Evol. 15:401.
8 Gianola, D., and J. L. Foulley. 1983. Sire evaluation for ordered categorical data with a threshold model. Genet. Sel. Evol. 15:201.
9Grahn. Y. T., H. Saloniemi, and J. Syvajkkvi. 1986. An epidemiological and genetic study on registered diseases in Finnish Ayrshire cattle. 111. Metabolic diseases. Acta Vet. Scand. 27:209.
10Grahn. Y. T., J. R. Thompson, and M. L. Bruss. 1984. Epidemiology and genetic basis of ketosis in Finnish Ayrshire cattle. Rev. Vet. Med. 3:65.
11 Hansen. L. B., C. W. Young, K. P. Miller, and R. W. Touchberry. 1979. Health care requirements of dairy cattle. I. Response to milk yield selection. J. Dairy Sci. 62:1922.
12 Harville, D. A., and R. W. Mee. 1984. A mixed model procedure for analyzing ordered categorical data. Bio- metrics 40:393.
13 Janss, L., and J. Foulley. 1993. Bivariate analysis for one continuous and one threshold dichotomous trait with unequal design matrices: an application to birth weight and calving difficulty. Livest. Prod. Sci. 33: 183.
14Lin. H. K., P. A. Oltenacu, L. V. Van Vleck, H. N. Erb, and R. D. Smith. 1989. Heritabilities of and genetic correlations among six health problems in Holstein cows. J. Dairy Sci. 72:180.
15Lyons, D. T., A. E. Freeman, and A. L. Kuck. 1991. Genetics of health traits in Holstein cattle. J. Dairy Sci. 74:1092.
16Mintysaari. E. A., Y. T. Grohn, and R. L. Quaas. 1991. Clinical ketosis: phenotypic and genetic correla- tions between occurrences and with milk yield. J. Dairy Sci. 74:3985.
17Mintysaari, E. A., Y. T. Grohn, and R. L. Quaas. 1993. Repeatability and heritability of lactational oc- currence of reproductive disorder in dairy cows. Prev. Vet. Med. 17:111.
18Mintysaari. E. A., and L. D. Van Vleck. 1989. Esti- mation of genetic parameters for production and reproduction in Finnish Ayrshire cattle. J. Dairy Sci. 7212375.
19 Oltenacu, P. A., A. Frick, and B. Lindhe. 1990. Evalu- ation of production, fertility, and health traits in progeny of Swedish Friesian, North American Hol- stein, and Friesian x Holstein crossbred sires. Proc.
Journal of Dairy Science Vol. 78, No. 2, 1995

430 URIBE ET AL.
4th World Congr. Genet. Appl. Livest. Prod., Edin- burgh, Scotland XIV:95.
20Oltenacu. P. A., A. Frick, and B. Lindhe. 1990. Epidemiological study of several clinical diseases, reproductive performance, and culling in primiparous Swedish cattle. F’rev. Vet. Med. 959.
21 Patterson, H. D., and R. Thompson. 1971. Recovery of inter-block information when the block sizes are unequal. Biometrika 58545.
22 Philipsson, I., B. Thafvelin, and I . Hedebro-Veler. 1980. Genetics studies on disease recording in first lactation cows of Swedish dairy breeds. Acta Agric. Scand. 30327.
23 Pirchner, F. 1969. Population Genetics in Animal Breeding. W. H. Freeman and Co., San Francisco, CA.
24 Schutz, M. M.. L. B. Hansen. G. R. Steuernagel, J. K. Reneau, and A. L. Kuck. 1990. Genetic parameters for somatic cells, protein, and fat in milk of Holsteins. J. Dairy Sci. 73494.
25 Shanks, R. D., A. E. Freeman, P. J. Berger, and D. H. Kelley. 1978. Effect of selection for milk production on reproductive and general health of the dauy cow. J. Dairy Sci. 61:1765.
26 Shook, G. E. 1989. Selection for disease resistance. J. Dairy Sci. 72:1349.
27 Simianer, H., H. Solbu, and L. R. Schaeffer. 1991. Estimated genetic correlation between disease and yield traits in dairy cattle. J. Dairy Sci. 74:4358.
28 Solbu, H. 1984. Disease recording in Norwegian dairy cattle. 11. Heritability estimates and progeny testing for mastitis, ketosis, and “all diseases”. Z. Tierz. Zuechtungsbiol. 10151.
29 Solbu. H., and 0. Lie. 1990. Selection for disease resistance in dauy cattle. Roc. 4th World Congr. Genet. Appl. Livest. Prod., Edinburgh, Scotland XVI: 445.
30Syviijiirvi. J., H. Saloniemi, and Y. Grohn. 1986. An epidemiological and genetic study on registered dis- eases in Finnish Ayrshire cattle. IV. Clinical mastitis. Acta Vet. Scand. 27:223.
31 Thompson, J. R. 1984. Genetic interrelationships of parturition problems and production. J. Dairy Sci. 67: 628.
32 Tveit, B., M. Svendsen, and K. Hove. 1991. Heritabil- ity of hypocalcemia at first parturition in Nonvegian cattle: genetic correlations with yield and weight. J. Dairy Sci. 74:3561.
33Van Vleck, L. D. 1964. Variation in type appraisal scores due to sire and herd effects. J. Dairy Sci. 47: 1249.
34 Young, C. W.. V. R. Eidman, and J. K. Reneau. 1985. Animal health and management and their impact on economic efficiency. J. Dairy Sci. 68:1593.
Journal of Dairy Science Vol. 78, No. 2, 1995




















