
Genetic and Functional Analysis of neuronatinin Mice with Maternal or Paternal Duplication of Distal...
12
DEVELOPMENTAL BIOLOGY 190, 66–77 (1997) ARTICLE NO. DB978681 Genetic and Functional Analysis of neuronatin in Mice with Maternal or Paternal Duplication of Distal Chr 2 Nobuaki Kikyo,* ,1 Christine M. Williamson,² Rosalind M. John,* Sheila C. Barton,* Colin V. Beechey,² Simon T. Ball,² Bruce M. Cattanach,² M. Azim Surani,* and Josephine Peters² ,2 *Wellcome/CRC Institute of Cancer and Developmental Biology, and Physiological Laboratory, University of Cambridge, Tennis Court Road, Cambridge CB2 1QR, United Kingdom; and ²Mammalian Genetics Unit, Medical Research Council, Harwell, Didcot, Oxon OX11 0RD, United Kingdom Functional differences between parental genomes are due to differential expression of parental alleles of imprinted genes. Neuronatin (Nnat) is a recently identified paternally expressed imprinted gene that is initially expressed in the rhombomeres and pituitary gland and later more widely in the central and peripheral nervous system mainly in postmitotic and differenti- ating neuroepithelial cells. Nnat maps to distal chromosome (Chr) 2, which contains an imprinting region that causes morphological abnormalities and early neonatal lethality. More detailed mapping analysis of Nnat showed that it is located between the T26H and T2Wa translocation breakpoints which is, surprisingly, proximal to the reported imprinting region between the T2Wa and T28H translocation breakpoints, suggesting that there may be two distinct imprinting regions on distal chromosome 2. To investigate the potential role of Nnat, we compared normal embryos with those which were PatDp.dist2.T26H (paternal duplication/maternal deficiency of chromosome 2 distal to the translocation breakpoint T26H) and MatDp.dist2.T26H. Expression of Nnat was detected in the PatDp.dist2.T26H embryos, where both copies of Nnat are paternally inherited, and normal embryos but no expression was detected in the MatDp.dist2.T26H embryos with the two maternally inherited copies. The differential expression of Nnat was supported by DNA methylation analysis with the paternally inherited alleles being unmethylated and the maternal alleles fully methylated. Although experimental embryos appeared grossly similar phenotypically in the structures where expression of Nnat was detected, differences in folding of the cerebellum were observed in neonates, and other more subtle developmental or behavioral effects due to gain or loss of Nnat cannot be ruled out. q 1997 Academic Press INTRODUCTION For example, gynogenetic cells contribute substantially to the brain, especially neocortex, while the androgenetic cells Genetic and embryological studies have established the are restricted to the hypothalamus and the dorsal root gan- requirement of both parental genomes for normal develop- glia (Fundele and Surani, 1994; Allen et al., 1995; Keverne ment in the mouse (Cattanach, 1986; Fundele and Surani, et al., 1996). These studies reflect cumulative effects of im- 1994; Gold and Pedersen, 1994; Latham et al., 1995). In printed genes in uniparental cells. chimeric studies with uniparental cells, gynogenetic and Genetic studies have identified mouse autosomal regions androgenetic cells show distinct and reciprocal properties. whose influence on development is dictated by their paren- tal origins (Cattanach, 1986). These regions include chro- mosomes (Chr) 2, 6, 7, 11, 12, and 17 based on detection of 1 Present address: Laboratory of Molecular Embryology, NIH, developmental and behavioral phenotypes (Cattanach and Bethesda, MD 20892-5431. Jones, 1994; Beechey and Cattanach, 1997). To date, 15 im- 2 To whom correspondence should be addressed. Fax: (01235) 834776. E-mail [email protected]. printed genes have been identified in the mouse and most 66 0012-1606/97 $25.00 Copyright q 1997 by Academic Press All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Genetic and Functional Analysis of neuronatinin Mice with Maternal or Paternal Duplication of Distal...

DEVELOPMENTAL BIOLOGY 190, 66–77 (1997)ARTICLE NO. DB978681
Genetic and Functional Analysis of neuronatinin Mice with Maternal or PaternalDuplication of Distal Chr 2
Nobuaki Kikyo,*,1 Christine M. Williamson,† Rosalind M. John,*Sheila C. Barton,* Colin V. Beechey,† Simon T. Ball,†Bruce M. Cattanach,† M. Azim Surani,*and Josephine Peters†,2
*Wellcome/CRC Institute of Cancer and Developmental Biology, and Physiological Laboratory,University of Cambridge, Tennis Court Road, Cambridge CB2 1QR, United Kingdom;and †Mammalian Genetics Unit, Medical Research Council, Harwell, Didcot,Oxon OX11 0RD, United Kingdom
Functional differences between parental genomes are due to differential expression of parental alleles of imprinted genes.Neuronatin (Nnat) is a recently identified paternally expressed imprinted gene that is initially expressed in the rhombomeresand pituitary gland and later more widely in the central and peripheral nervous system mainly in postmitotic and differenti-ating neuroepithelial cells. Nnat maps to distal chromosome (Chr) 2, which contains an imprinting region that causesmorphological abnormalities and early neonatal lethality. More detailed mapping analysis of Nnat showed that it is locatedbetween the T26H and T2Wa translocation breakpoints which is, surprisingly, proximal to the reported imprinting regionbetween the T2Wa and T28H translocation breakpoints, suggesting that there may be two distinct imprinting regions ondistal chromosome 2. To investigate the potential role of Nnat, we compared normal embryos with those which werePatDp.dist2.T26H (paternal duplication/maternal deficiency of chromosome 2 distal to the translocation breakpoint T26H)and MatDp.dist2.T26H. Expression of Nnat was detected in the PatDp.dist2.T26H embryos, where both copies of Nnatare paternally inherited, and normal embryos but no expression was detected in the MatDp.dist2.T26H embryos with thetwo maternally inherited copies. The differential expression of Nnat was supported by DNA methylation analysis withthe paternally inherited alleles being unmethylated and the maternal alleles fully methylated. Although experimentalembryos appeared grossly similar phenotypically in the structures where expression of Nnat was detected, differences infolding of the cerebellum were observed in neonates, and other more subtle developmental or behavioral effects due togain or loss of Nnat cannot be ruled out. q 1997 Academic Press
INTRODUCTION For example, gynogenetic cells contribute substantially tothe brain, especially neocortex, while the androgenetic cells
Genetic and embryological studies have established the are restricted to the hypothalamus and the dorsal root gan-requirement of both parental genomes for normal develop- glia (Fundele and Surani, 1994; Allen et al., 1995; Kevernement in the mouse (Cattanach, 1986; Fundele and Surani, et al., 1996). These studies reflect cumulative effects of im-1994; Gold and Pedersen, 1994; Latham et al., 1995). In printed genes in uniparental cells.chimeric studies with uniparental cells, gynogenetic and Genetic studies have identified mouse autosomal regionsandrogenetic cells show distinct and reciprocal properties. whose influence on development is dictated by their paren-
tal origins (Cattanach, 1986). These regions include chro-mosomes (Chr) 2, 6, 7, 11, 12, and 17 based on detection of1 Present address: Laboratory of Molecular Embryology, NIH,developmental and behavioral phenotypes (Cattanach andBethesda, MD 20892-5431.Jones, 1994; Beechey and Cattanach, 1997). To date, 15 im-2 To whom correspondence should be addressed. Fax: (01235)
834776. E-mail [email protected]. printed genes have been identified in the mouse and most
66
0012-1606/97 $25.00Copyright q 1997 by Academic Press
All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba

67Genetic Imprinting of neuronatin
Embryos and newborn mice with maternal or paternal duplica-of them map to these imprinting regions. The majority oftion for specific regions of Chr 2 were generated by the standardthese genes are also imprinted in humans where some ge-method of intercrossing reciprocal translocation heterozygotesnetic disorders can be ascribed to their loss or gain of func-(Searle and Beechey, 1978; Cattanach, 1986). The genetically de-tion. (Brenton et al., 1995; Ledbetter and Engel, 1995).fined imprinting region lies between the breakpoints of two recipro-It is important to identify the remaining imprinted genescal translocations T(2;8)2Wa in band 2H3 and T(2;16)28H in band
for a comprehensive understanding of the roles of genetic 2H4 (Cattanach et al., 1992) (Fig. 3B). The T(2;8)26H translocationimprinting. We have established two systematic screens for with a more proximal breakpoint in band 2H1 (de Boer and vanthis purpose. In both types of screen, we have used andro- Gijsen, 1974), hereafter referred to as T26H, was used routinelygenetic, parthenogenetic, and normal mouse embryos at 8.5 because two visible genetic markers, bp and pa, were associated
with the translocation making typing easier, and the imprintingdays postcoitum (dpc) as the source of material for a ge-phenotypes generated using T(2;8)26H and T(2;8)2Wa were similarnome-wide search. In one screen cDNA subtractive hybrid-(Cattanach et al., 1992). Mice of genotype MatDp.dist2.T26H andization and differential screening were used successfully toPatDp.dist2.T26H were generated from reciprocal crosses of /detect novel imprinted genes, Peg1/Mest (Kaneko-Ishino etT26H /// / / 1 / T26H bp/pa / bp or pa T26H bp/pa / bp. Inal., 1995) and Peg3 (Kuroiwa et al., 1996). In a second screeneach mating one of the parents was homozygous for brachypodismdescribed here, we have successfully used mRNA differen-(bp), a mutation causing shortening of the limbs, that maps distal
tial display (Liang et al., 1995) to identify further novel to the T26H breakpoint. Offspring homozygous for bp could beimprinted genes. identified from 15.5 dpc onward and were either MatDp.dist2.T26H
In these studies we identified neuronatin (locus symbol or PatDp.dist2.T26H depending on the genotype of the parents.Nnat) and showed that it is imprinted so that there is only MatDp.dist2.T26H and PatDp.dist2.T26H embryos at all stages
were also identified by typing for the microsatellite markerexpression of the paternal allele. Nnat was simultaneouslyD2Mit226 as described previously (Williamson et al., 1995). Em-detected in the cDNA subtraction screen and named as Peg5bryos of genotypes MatDp.dist2.T26H and PatDp.dist2.T26H are(Kagitani et al., 1997). Nnat is highly expressed in the devel-expected to occur at a frequency of about 16% each of live progenyoping mammalian nervous system (Joseph et al., 1994;from T26H// intercrosses, whereas embryos of genotypes MatDp.-Wijnholds et al., 1995; Dou and Joseph, 1996). We mappedprox2.T26H (maternal duplication proximal to the T26Hthis gene to a region on mouse Chr 2, which contains anbreakpoint) and PatDp.prox2.T26H are much rarer (1–5% of prog-
imprinting region causing reciprocal phenotypic abnormali- eny) (Searle et al., 1971). From crosses of pa T26H bp/pa / bpties depending on parental origin. A detailed analysis of females with / T26H //pa a bp males, two MatDp.prox2.T26Himprinting of Nnat as well as its expression and phenotypic have been identified using the visible marker gene pallid (pa), lo-consequences was carried out by comparing normal em- cated proximal to T26H. One pallid embryo, identified by its pink
eye color, was found in utero at 15.5 dpc, while another was identi-bryos with the PatDp.dist2.T26H (paternal duplication/ma-fied at birth and reared to adulthood.ternal deficiency of Chr 2 distal to the translocation
For some experiments the reciprocal translocation T(2;8)2Wa,breakpoint T26H) and MatDp.dist2.T26H embryos with al-hereafter referred to as T2Wa, was used. Mice of genotype MatDp.-tered dosage of the Nnat gene product. We also suggest thatdist2.T2Wa were generated by intercrossing T2Wa// and, due tothe mRNA differential display (DD) method of comparinga lack of suitable distal genetic markers, were only taken when themouse uniparental embryos is a useful method for detectingwell documented phenotype was clearly visible. Four pa embryos,
novel imprinted genes. aged between 13.5 and 17.5 dpc, of genotype MatDp.prox2.T2Wawere derived by crossing pa / / T2Wa/pa a bp / females with // / T2Wa// / / / males as described previously (Cattanach and
MATERIALS AND METHODS Beechey, 1995), pa being proximal to the breakpoint of T2Wa onChr 2. As with T26H, recovery of embryos with duplications forregions proximal to translocation breakpoints are expected with aMicefrequency of only 1–5% of progeny.
Embryos for DD were derived from (C57BL/6J 1 CBA) F1 (hereaf-ter called F1) and 129/Sv strain mice. Diploid parthenogenetic em-
mRNA Differential Display (DD)bryos were prepared by activating eggs from F1 females (Barton etal., 1987) and diploid androgenetic embryos were made by pronu- Normal, parthenogenetic, and androgenetic mouse embryos withclear transplantation (Barton and Surani, 1993) using F1 1 129/Sv 5–15 somites at 8.5 dpc were pooled separately to isolate totaleggs; the female (F1) pronucleus was removed from a fertilized egg RNA by the acid–guanidinium–phenol method (Chomczynski andand replaced by a second 129/Sv male pronucleus from another egg. Sacchi, 1987). DD was performed as described previously (Liang etBoth parthenogenetic and androgenetic eggs were cultured for 2 al., 1995) with some modifications. We increased total RNA to 1days and transferred at the 4- to 8-cell stage to the oviducts of Day mg/20 ml for reverse transcription and dNTP to 20 mM for polymer-1 (day of plug) pseudopregnant F1 recipient females. Both F2 and ase chain reaction (PCR).homozygous 129/Sv embryos derived from normal fertilization,henceforth called normal, were used as controls during the DD RT–PCR for Verification of Differential Expressionanalysis to exclude loci detected as a result of strain polymor-phisms. Mus spretus, the C57BL/6J and C3H/HeH strains, and the RNA samples prepared independently of those used in DD were
compared by reverse transcription PCR (RT–PCR). Total RNA washybrid (C3H/HeH 1 101/H) (hereafter called 3H1) of M. musculuswere used for the analysis of restriction fragment length variants reverse transcribed with random hexamers (Life Technologies) by
using SuperScript II (Life Technologies). The primers 5*-AGGTAC-in mice from interspecific crosses.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba

68 Kikyo et al.
TCCCTGCAGAAG-3* and 5*-GCTGAGATGGTGCTGTTC-3* fluorescein–avidin with three 41T washes between each layer.Slides were dehydrated through an ethanol series and air dried.were used to detect the expression of Nnat. Three alternatively
spliced forms, neuronatin 1, 2, and 3, have been described and the Chromosomes were counterstained with propidium iodide in Vec-tashield solution (Vector). Spreads were examined using an MRCabove sequences correspond to the region of nucleotides 226–638
of neuronatin 3, which is common to all three forms (Wijnholds 600 confocal microscope.et al., 1995). PCR was performed using AmpliTaq (Perkin Elmer)as described in its instructions. The PCR mixture was denaturedat 947C for 3 min then amplified by 24 cycles at 947C for 30 sec, In Situ Hybridization of Nnat567C for 30 sec, and 727C for 1 min followed by a final extension
A 1.4-kb fragment of Nnat cDNA, containing all three exonsat 727C for 5 min. The PCR products were confirmed by Southernsubcloned into pBluescript SK(0) (Stratagene) (a generous gift fromhybridization with a probe of an internal sequence, 5*-TCGTCA-F. Kagitani and F. Ishino) was used to prepare sense and antisenseGAGGAGCACTTG-3* end labeled with [g-32P]dATP (DuPont)RNA probes by in vitro transcription using the DIG RNA labeling(Sambrook et al., 1989). The mouse b-actin gene was used as akit (Boehringer Mannheim). Parasagittal sections from normal,control for RT–PCR using 5*-GCTGTGCTATGTTGCTCTAG-3*MatDp.dist2.T26H and PatDp.dist2.T26H mouse embryos at 13.5and 5*-ATCGTACTCCTGCTTGCTGA-3* as a primer pair and 5*-and 17.5 dpc were used for in situ hybridization. Furthermore,AAGAGCTATGAGCTGCCTGA-3* as a probe in Southern hybrid-transverse sections of brains from normal embryos at 13.5 dpc,ization.newborn, and 8 months old were compared.
In situ hybridization was carried out as described previously(Wilkinson and Nieto, 1993). Briefly, embryos were fixed in 4%Restriction Fragment Length Variation of Nnat inparaformaldehyde at 47C overnight and sections were treated withMice from Interspecific Crosses proteinase K (20 mg/ml) and hybridized with the probes overnightat 657C. The sections were washed at 657C and adsorbed with alka-RNA was isolated from appropriate mice (see Fig. 2) and theline phosphatase-conjugated anti-DIG antibody overnight at 47C.Nnat gene was amplified by RT–PCR as described above usingAlkaline phosphatase activity was detected by using BM purple AP5*-GCTCATCATCGGCTGGTA-3* and 5*-ACAACTGCTGCGT-substrate (Boehringer Mannheim) followed by counterstain withGAGAC-3* as primers. The PCR products were digested with HinfI0.5% eosin. Each embryo was also stained with hematoxylin–eosinand analyzed by Southern hybridization with an oligonucleotidefor histological observation.probe of an internal sequence, 5*-CAACTGTGTGCAGCATGG-3*
as described above.
Methylation AnalysisChromosome Mapping Ten micrograms of genomic DNA was digested with the appro-
priate restriction enzymes and analyzed by Southern hybridizationRT–PCR for positioning Nnat. Nnat was initially mapped be-using a cDNA clone of Nnat as a probe.tween D2Mit48 and D2Mit29 on mouse distal Chr 2 by using a
backcross panel (Kagitani et al., 1997). Mapping was carried outfurther by RT–PCR testing Nnat expression in mice which are dupli-cation/deficient for Chr 2 proximal and distal to the T26H and T2Wa RESULTSbreakpoints. PCR consisted of 30 cycles at 947C for 1 min, 557C for1 min, and 727C for 1 min followed by a final 10-min extension at Isolation of Imprinted Genes by DD727C. Primers for Nnat were 5*-CGGACTCCGAGACCAGTAGA-3* and 5*-GCTTCTGCAGGGAGTACCTG-3*, (these sequences cor- Eighty primer pairs were used in this screen and approxi-respond to nucleotides 2–244 of neuronatin 3; (Wijnholds et al., mately 80 to 100 bands per primer pair were observed on1995)), and for control, Hprt, were 5*-GAAATGTCAGTTGCT- DNA sequencing gels. The overall pattern of bands wasGCGTC-3* and 5*-GCCAACACTGCTGAAACATG-3* (Konecki et similar in all six embryonic samples (two pools each ofal., 1982). normal, parthenogenetic, and androgenetic embryos) indi-
Fluorescence in situ hybridization (FISH). A mouse BAC clonecating good reproducibility. So far we have detected 10 dif-of 100 kb containing Nnat was isolated from a gridded 129 mouseferentially expressed bands: 6 were observed only in RNABAC library (Genome Systems), labeled by nick translation usingisolated from normal and parthenogenetic embryos (puta-the Bionick kit (Life Technologies), and used as a probe. FISH wastive maternally expressed imprinted genes) and 4 were de-performed on slides of 12.5 dpc embryonic liver cells from T2Wa
homozygotes with 250 ng of the BAC probe and 5 mg of Cot 1 DNA tected only in RNA from normal and androgenetic embryosin 50% formamide hybridization buffer. Briefly, mitotic chromo- (putative paternally expressed imprinted genes). The first ofsome spreads prepared as described (Evans, 1987) were dried, passed these candidates was seen strongly in one (but not both)through an ethanol series, and placed in acetone for 10 min. Chro- normal and two parthenogenetic embryonic pools; the samemosomes were denatured for 3 min at 707C in 70% formamide/ band was very faint in two androgenetic embryonic pools21 SSC, quenched in ice-cold ethanol, and dehydrated through an (Fig. 1A). This DNA sequence was completely homologousethanol series. Slides were hybridized overnight at 377C. The
to bases 1530–1856 of H19 (EMBL X58196), a known mater-washes were 50% formamide/21 SSC, 3 1 5 min; 21 SSC, 5 min;nally expressed imprinted gene on distal Chr 7 indicating41 SSC/0.05% Tween 20 (41T), 5 min; all at 407C, and then 41T,that the method is capable of detecting imprinted genes. A5 min at room temperature. Fluorescein–avidin and biotinylatedsecond candidate was detected only in normal and andro-anti-avidin were then used to detect the bound probe, by layering
fluorescein–avidin, biotinylated anti-avidin, and a second layer of genetic embryos (Fig. 1B). The DNA sequence of this second
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba

69Genetic Imprinting of neuronatin
by increasing the total RNA for reverse transcription to1.0 mg/20 ml instead of 0.2 mg/20 ml. In addition, bandslonger than 500 bp increased by increasing dNTP from 2to 20 mM in PCR (data not shown). So far we have isolated2 genuine imprinted genes of 6400–8000 transcripts exam-ined in 8.5-dpc embryos giving a conservative estimate of0.025%. However, such an estimate may be flawed becauseof limited sensitivity of DD, temporal or tissue-specificimprinting, and the possibility that a single gene may berepresented more than once during analysis. Hayashizakiet al. estimated the number of imprinted genes to be about100, based on the frequency of differentially methylatedloci in maternal and paternal alleles (Hayashizaki et al.,1994).
The RT–PCR product of Nnat with specific primers wasclearly detected only in RNA isolated from both normaland androgenetic embryos at 8.5 dpc after 24 cycles of PCR(Fig. 1C). The expression of the control gene, b-actin, wassimilar in all RNA samples after 16 cycles of PCR. Thesedata suggested that Nnat was expressed only from the pater-nal allele at 8.5 dpc.
The imprinting of Nnat was further confirmed using arestriction fragment length variant that distinguishes al-leles derived from M. musculus and M. spretus. The RT–PCR analysis after 26 cycles amplified only neuronatin 2FIG. 1. Differential expression of H19 and Nnat. (A, B) mRNAand neuronatin 3, which was confirmed by Southern hybrid-differential display. RNA was isolated from two pools each of par-ization with an oligonucleotide probe described above (datathenogenetic (PG), normal and androgenetic (AG) embryos at 8.5
dpc. PGs and normal 1 are F2 and normal 2 and AGs are 129/Sv not shown). After digestion of the amplified product withstrain. T12VC (where V may be A, C, or G) and 5*-CTGTACCCCC- HinfI, this probe detected a 765-bp fragment (neuronatin 2)3* were used in A and T12VG and 5*-AGTTAGGCAC-3* in B as and a 774-bp fragment (neuronatin 3) in M. musculusprimers. An arrow indicates the band detected strongly in PG and (which were not clearly separated on the gel), and a 260-bpnormal embryos in A (H19) and only in normal and AG embryos fragment (neuronatin 2 and neuronatin 3) in M. spretus. Toin B (Nnat). (C) Verification of differential expression of Nnat by examine the expression of Nnat in mice with a maternallySouthern hybridization of RT–PCR products. //0 RT refers to the
derived M. musculus allele, F1 embryos from the crosspresence and absence, respectively, of reverse transcriptase. PCR(C57BL/6J female 1 M. spretus male), or adult mice fromproducts were detected by hybridization with an oligonucleotidethe cross (3H1 female 1 M. spretus male) were tested. Toprobe of an internal sequence. b-actin was used as a control foranalyze the expression of Nnat in mice with a paternallyRT–PCR. The size of the PCR product is 413 bp for Nnat and 445
bp for b-actin, respectively. derived M. musculus allele, female F1 (C57BL/6J 1 M.spretus) were backcrossed to C57BL/6J males, and femaleF1 (3H1 1M. spretus) were backcrossed to 3H1 males. Prog-eny that inherited the M. spretus allele from the motherwere identified by restriction fragment length variant analy-candidate was 99% homologous to the nucleotides 587–
1139 of neuronatin 3 (EMBL X83570). The remaining eight sis of genomic DNA (data not shown), and then used forRT–PCR analysis. Only the paternal allele of Nnat wasband differences are being investigated.
The cDNA subtraction hybridization method (Kaneko- expressed in the brain of embryos at 13 dpc (Fig. 2A) and inyoung and adult mice (Fig. 2B). By increasing the numberIshino et al., 1995; Kuroiwa et al., 1996) and the RLGS
method (Hayashizaki et al., 1994) have proved to be effec- of PCR cycles, however, the transcript from the repressedmaternal allele also appeared weakly (data not shown, seetive means for the identification of imprinted genes. Nnat
was also isolated as Peg5 by cDNA subtraction screen (Kagi- Discussion). Neuronatin 1 was not detectable by this RT–PCR; it may be expressed at a very low level or at differenttani et al., 1997). Here we demonstrate that DD may simi-
larly be a reliable method for detecting imprinted genes. stages of development.This technique is relatively quick and it requires less than10 mg of total RNA to screen more than 10,000 transcripts.
Imprinting in the Distal Region of Chr 2Most important of all, DD enables immediate and simulta-neous comparisons between multiple samples, an advan- Imprinting and positioning of Nnat in the context of
the defined distal Chr 2 imprinting region. The initialtage clearly demonstrated in the detection of H19 gene (seeFig. 1A). The reproducibility of the method was improved mapping of Peg5/Nnat indicated that that this gene is lo-
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba

70 Kikyo et al.
In order to determine whether Nnat lies within the pre-viously defined imprinting region on distal Chr 2, we lookedfor expression/repression of the gene in duplication/defi-cient mice derived from the reciprocal translocation atT2Wa, that defines the proximal boundary of the previouslydefined imprinting region. Surprisingly, Nnat was detectedin the brain of a newborn mouse carrying MatDp.dist2.-T2Wa (lane 10) but was absent in the brain of a 17.5-dpcembryo carrying MatDp.prox2.T2Wa (lane 14). Another em-bryo was found at 15.5 dpc which also showed repression ofNnat (results not shown). These expression results thereforeindicate that Nnat lies proximal to the T2Wa breakpointand thus outside the previously defined imprinting regionbetween the translocation breakpoints of T2Wa and T28Has shown in Fig. 3B.
The position of Nnat in relation to the translocationbreakpoint in T2Wa homozygotes was verified by FISHanalysis. T2Wa is a reciprocal translocation between Chrs2 and 8 with breakpoints at 2H3 and 8A4 giving rise to twoclearly recognizable products, a very long 28 chromosome
FIG. 2. Verification of imprinting of Nnat by analysis of restric- and a very short 82. Clear signal was present in 20 full meta-tion fragment length variation in mice from interspecific crosses. phase spreads in the distal half of the long 28 chromosome(A) Nnat was amplified by RT–PCR using RNA isolated from the and no signal could be detected on the small 82 chromosomebrains of newborn C57BL/6J, newborn M. spretus, (C57BL/6J female as shown in Fig. 3C thus verifying that Nnat lies proximal to1 M. spretus male) F1 at 13 dpc and (C57BL/6J 1 M. spretus) F1 the T2Wa breakpoint on Chr 2 and outside of the previouslyfemale 1 C57BL/6J male at 13 dpc. After digestion with HinfI, the defined imprinting region.PCR products were analyzed by Southern hybridization with an
Expression of Nnat and phenotypic analysis of embryos.oligonucleotide probe of an internal sequence. (B) RNA was isolatedFurther detailed analysis of Nnat was carried out by compar-from the brains of adult C3H/HeH, M. spretus and (3H1 female 1ing normal embryos with PatDp.dist2.T26H and MatDp.-M. spretus male) F1, and 4-week-old (3H1 1 M. spretus) F1 femaledist2.T26H embryos. The level of Nnat transcript was high1 3H1 male. The RT–PCR products were analyzed as in A. Both
C3H/HeH and 3H1 have exactly the same Nnat variant (data not in the brain and spinal cord and low in the dorsal root gan-shown). glia, tongue, gut, and limb mesenchyme in a normal embryo
at 13.5 dpc, which agreed with the previous data (Wijnholdset al., 1995) (Fig. 4B). Nnat was expressed weakly in theesophagus and lung, although variable, in addition to highexpression level in the neural tissues in a normal embryocated on distal Chr 2 (Kagitani et al., 1997). Nnat was fur-
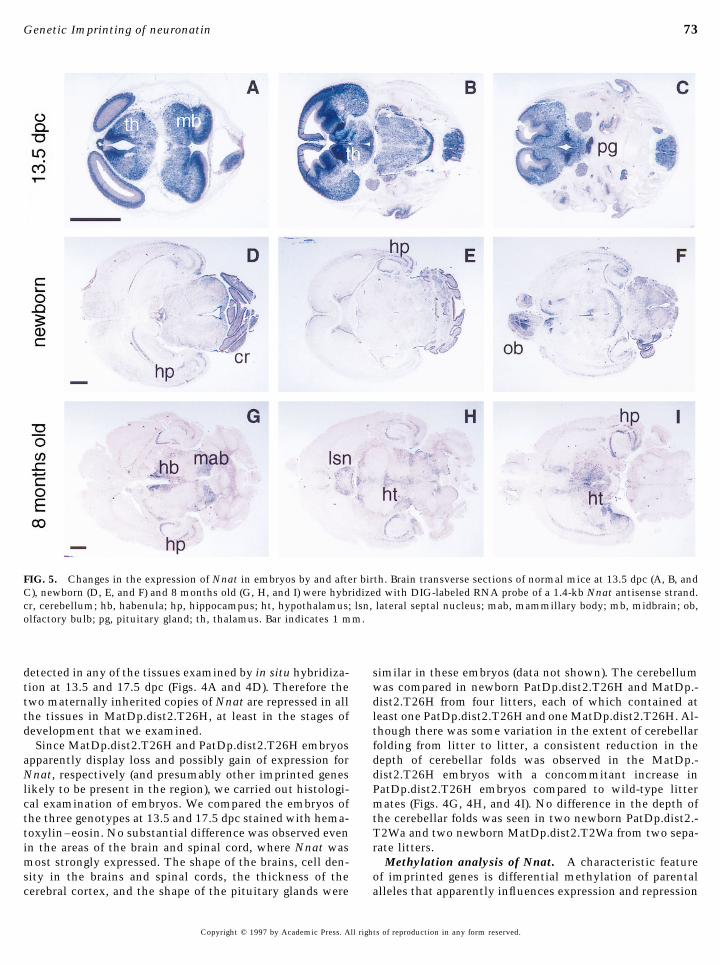
ther positioned in distal Chr 2 in relation to the breakpoints at 17.5 dpc (Fig. 4E). The expression of Nnat in developingand aging brain was studied by comparing transverse sec-of two translocations, T26H and T2Wa, by looking for its
expression/repression in embryos of different genotypes. tions of the brain from 13.5-dpc, newborn, and 8-month-oldmice. At 13.5 dpc, Nnat was widely expressed in the brainRT–PCR was used to test for Nnat expression in the brain
and Hprt primers were included as an amplification control. apart from the periventricular zone (Figs. 5A, 5B, and 5C).The signal was especially strong in the pituitary gland (Fig.The Hprt primers amplified within an exon and produced
a band of 332 bp in only the samples containing reverse 5C) and the areas adjacent to the periventriclar zone in thethalamus and midbrain (Figs. 5A and 5B). However, in thetranscriptase as shown in Fig. 3A, showing that the ampli-
fication was not due to genomic contamination in RNA. newborn mouse it was detected only in the cerebellum,especially the cortex (Figs. 5D and 5E), hippocampus (Figs.The Nnat primers produced a 243-bp band, which corre-
sponds to neuronatin 3, from a phenotypically normal new- 5D, 5E, and 5F), and olfactory bulbs (Fig. 5F). At 8 months,Nnat was detected in the hippocampus (Figs. 5G, 5H, andborn mouse derived from the T26H intercross (lane 2). This
band was absent in a newborn mouse of genotype MatDp.- 5I), habenula, mammillary body (Fig. 5G), lateral septal nu-cleus (Fig. 5H), and hypothalamus (Figs. 5H and 5I) but wasdist2.T26H (lane 4), confirming that Nnat is paternally ex-
pressed, assuming that the translocation does not disrupt absent in the cerebellar cortex (Figs. 5G and 5H).We then went on to examine expression of Nnat in thean important control element. From subsequent analysis
this possibility was ruled out. The band was present in a PatDp.dist2.T26H embryos which contains two potentiallyactive copies of Nnat. There was no detectable difference15.5-dpc embryo of genotype MatDp.prox2.T26H (lane 6).
These data place the Nnat locus distal to the T26H in the spatial pattern of expression of Nnat between normaland PatDp.dist2.T26H embryos at these stages (Fig. 4). Bybreakpoint as shown in Fig. 3B, potentially within the pre-
viously defined imprinting region. contrast, in the MatDp.dist2.T26H embryos, Nnat was not
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba
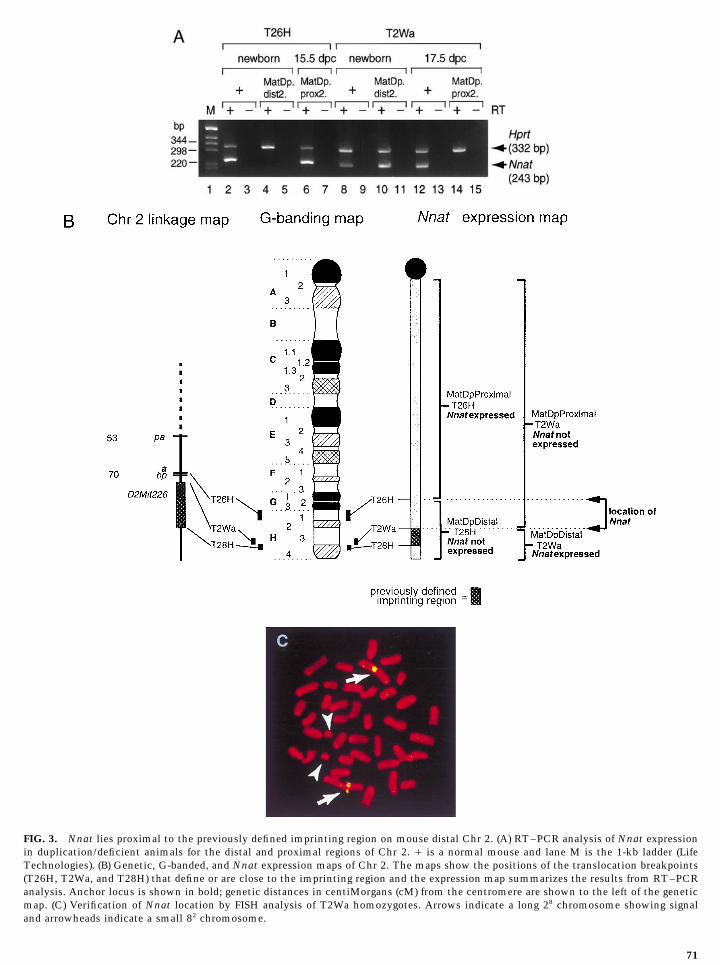
FIG. 3. Nnat lies proximal to the previously defined imprinting region on mouse distal Chr 2. (A) RT–PCR analysis of Nnat expressionin duplication/deficient animals for the distal and proximal regions of Chr 2. / is a normal mouse and lane M is the 1-kb ladder (LifeTechnologies). (B) Genetic, G-banded, and Nnat expression maps of Chr 2. The maps show the positions of the translocation breakpoints(T26H, T2Wa, and T28H) that define or are close to the imprinting region and the expression map summarizes the results from RT–PCRanalysis. Anchor locus is shown in bold; genetic distances in centiMorgans (cM) from the centromere are shown to the left of the geneticmap. (C) Verification of Nnat location by FISH analysis of T2Wa homozygotes. Arrows indicate a long 28 chromosome showing signaland arrowheads indicate a small 82 chromosome.
71
AID DB 8681 / 6x2c$$8681 09-05-97 09:16:43 dba
72 Kikyo et al.
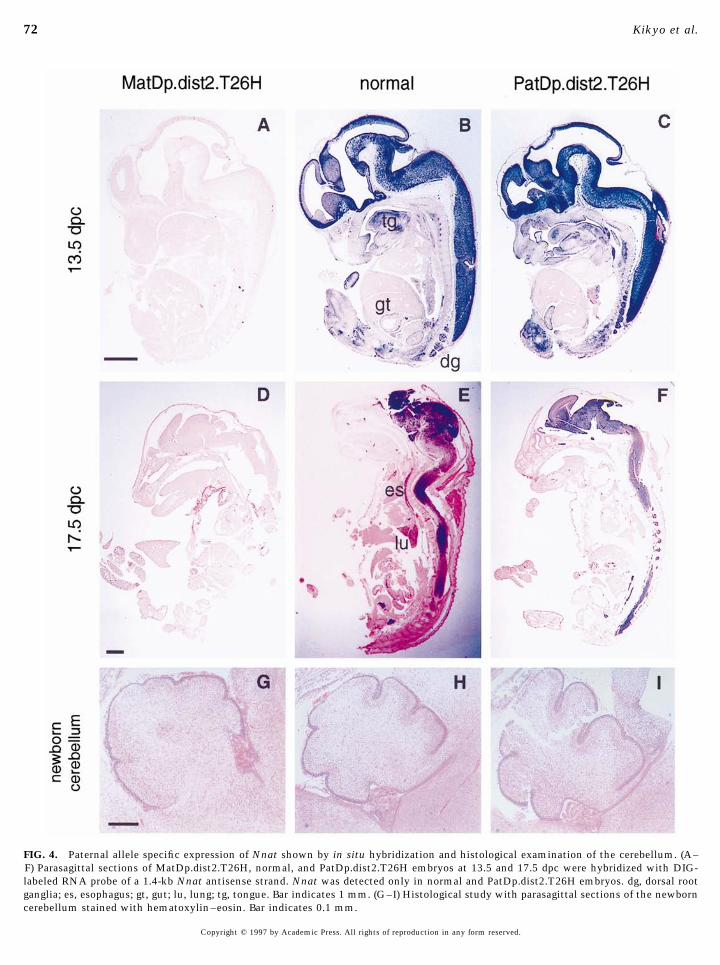
FIG. 4. Paternal allele specific expression of Nnat shown by in situ hybridization and histological examination of the cerebellum. (A–F) Parasagittal sections of MatDp.dist2.T26H, normal, and PatDp.dist2.T26H embryos at 13.5 and 17.5 dpc were hybridized with DIG-labeled RNA probe of a 1.4-kb Nnat antisense strand. Nnat was detected only in normal and PatDp.dist2.T26H embryos. dg, dorsal rootganglia; es, esophagus; gt, gut; lu, lung; tg, tongue. Bar indicates 1 mm. (G–I) Histological study with parasagittal sections of the newborncerebellum stained with hematoxylin–eosin. Bar indicates 0.1 mm.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$8681 09-05-97 09:16:43 dba
73Genetic Imprinting of neuronatin
FIG. 5. Changes in the expression of Nnat in embryos by and after birth. Brain transverse sections of normal mice at 13.5 dpc (A, B, andC), newborn (D, E, and F) and 8 months old (G, H, and I) were hybridized with DIG-labeled RNA probe of a 1.4-kb Nnat antisense strand.cr, cerebellum; hb, habenula; hp, hippocampus; ht, hypothalamus; lsn, lateral septal nucleus; mab, mammillary body; mb, midbrain; ob,olfactory bulb; pg, pituitary gland; th, thalamus. Bar indicates 1 mm.
detected in any of the tissues examined by in situ hybridiza- similar in these embryos (data not shown). The cerebellumwas compared in newborn PatDp.dist2.T26H and MatDp.-tion at 13.5 and 17.5 dpc (Figs. 4A and 4D). Therefore the
two maternally inherited copies of Nnat are repressed in all dist2.T26H from four litters, each of which contained atleast one PatDp.dist2.T26H and one MatDp.dist2.T26H. Al-the tissues in MatDp.dist2.T26H, at least in the stages of
development that we examined. though there was some variation in the extent of cerebellarfolding from litter to litter, a consistent reduction in theSince MatDp.dist2.T26H and PatDp.dist2.T26H embryos
apparently display loss and possibly gain of expression for depth of cerebellar folds was observed in the MatDp.-dist2.T26H embryos with a concommitant increase inNnat, respectively (and presumably other imprinted genes
likely to be present in the region), we carried out histologi- PatDp.dist2.T26H embryos compared to wild-type littermates (Figs. 4G, 4H, and 4I). No difference in the depth ofcal examination of embryos. We compared the embryos of
the three genotypes at 13.5 and 17.5 dpc stained with hema- the cerebellar folds was seen in two newborn PatDp.dist2.-T2Wa and two newborn MatDp.dist2.T2Wa from two sepa-toxylin–eosin. No substantial difference was observed even
in the areas of the brain and spinal cord, where Nnat was rate litters.Methylation analysis of Nnat. A characteristic featuremost strongly expressed. The shape of the brains, cell den-
sity in the brains and spinal cords, the thickness of the of imprinted genes is differential methylation of parentalalleles that apparently influences expression and repressioncerebral cortex, and the shape of the pituitary glands were
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba
74 Kikyo et al.
detail, phenotypic effects and the involvement of differen-tial DNA methylation in regulation of the expression bycomparing PatDp.dist2.T26H and MatDp.dist2.T26H em-bryos.
Nnat was first identified as a gene strongly expressed inthe rat neonatal brain compared to the expression in brainsfrom young and aged adults, and it was suggested that it isinvolved in neuronal cell differentiation (Joseph et al.,1994). There are stretches of 23 hydrophobic amino acidsat the N terminus and 31 hydrophilic amino acids at the Cterminus in human Nnat, suggesting that the gene mayencode a transmembrane protein (Dou and Joseph, 1996).Recently, Wijnholds et al. (1995) isolated mouse Nnat inde-pendently as a gene that is downregulated in P19 embryonalcarcinoma cells during differentiation. They reported threealternatively spliced forms, neuronatin 1, 2, and 3 whichwere predicted to code for proteins with 55, 54, and 81amino acids, respectively. The expression of Nnat is firstdetected in rhombomere 3 and 5 and Rathke’s pouch at 8.5dpc. In addition to the later expression in paraxial meso-derm, somites, and limb mesenchyme, it is also widely ex-
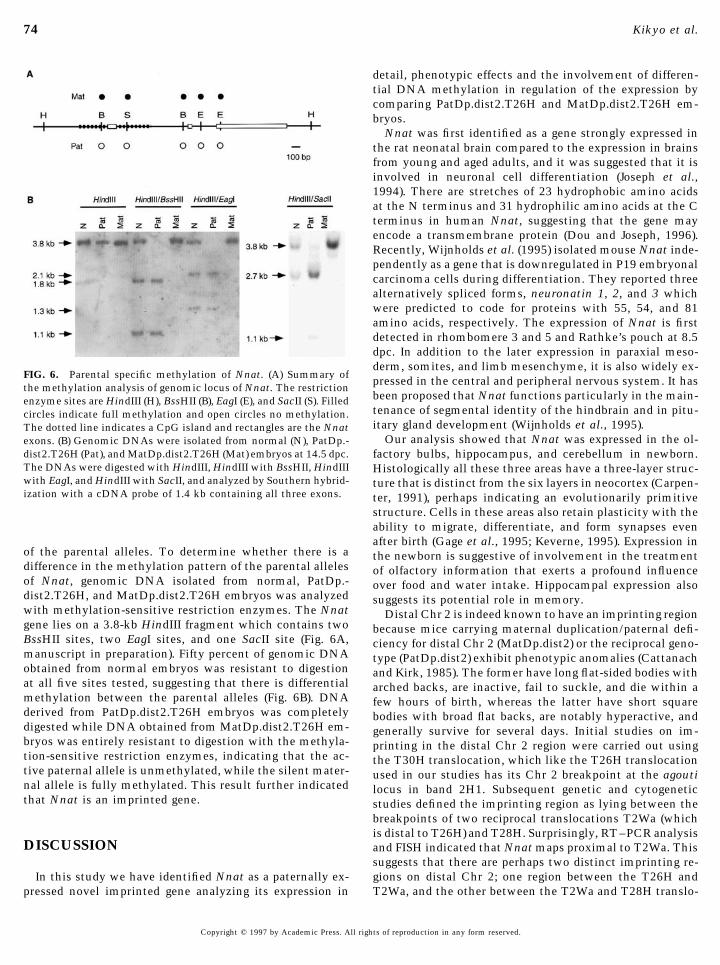
FIG. 6. Parental specific methylation of Nnat. (A) Summary of pressed in the central and peripheral nervous system. It hasthe methylation analysis of genomic locus of Nnat. The restriction
been proposed that Nnat functions particularly in the main-enzyme sites are HindIII (H), BssHII (B), EagI (E), and SacII (S). Filledtenance of segmental identity of the hindbrain and in pitu-circles indicate full methylation and open circles no methylation.itary gland development (Wijnholds et al., 1995).The dotted line indicates a CpG island and rectangles are the Nnat
Our analysis showed that Nnat was expressed in the ol-exons. (B) Genomic DNAs were isolated from normal (N), PatDp.-dist2.T26H (Pat), and MatDp.dist2.T26H (Mat) embryos at 14.5 dpc. factory bulbs, hippocampus, and cerebellum in newborn.The DNAs were digested with HindIII, HindIII with BssHII, HindIII Histologically all these three areas have a three-layer struc-with EagI, and HindIII with SacII, and analyzed by Southern hybrid- ture that is distinct from the six layers in neocortex (Carpen-ization with a cDNA probe of 1.4 kb containing all three exons. ter, 1991), perhaps indicating an evolutionarily primitive
structure. Cells in these areas also retain plasticity with theability to migrate, differentiate, and form synapses evenafter birth (Gage et al., 1995; Keverne, 1995). Expression in
of the parental alleles. To determine whether there is a the newborn is suggestive of involvement in the treatmentdifference in the methylation pattern of the parental alleles of olfactory information that exerts a profound influenceof Nnat, genomic DNA isolated from normal, PatDp.- over food and water intake. Hippocampal expression alsodist2.T26H, and MatDp.dist2.T26H embryos was analyzed suggests its potential role in memory.with methylation-sensitive restriction enzymes. The Nnat Distal Chr 2 is indeed known to have an imprinting regiongene lies on a 3.8-kb HindIII fragment which contains two because mice carrying maternal duplication/paternal defi-BssHII sites, two EagI sites, and one SacII site (Fig. 6A, ciency for distal Chr 2 (MatDp.dist2) or the reciprocal geno-manuscript in preparation). Fifty percent of genomic DNA type (PatDp.dist2) exhibit phenotypic anomalies (Cattanachobtained from normal embryos was resistant to digestion and Kirk, 1985). The former have long flat-sided bodies withat all five sites tested, suggesting that there is differential arched backs, are inactive, fail to suckle, and die within amethylation between the parental alleles (Fig. 6B). DNA few hours of birth, whereas the latter have short squarederived from PatDp.dist2.T26H embryos was completely bodies with broad flat backs, are notably hyperactive, anddigested while DNA obtained from MatDp.dist2.T26H em- generally survive for several days. Initial studies on im-bryos was entirely resistant to digestion with the methyla- printing in the distal Chr 2 region were carried out usingtion-sensitive restriction enzymes, indicating that the ac- the T30H translocation, which like the T26H translocationtive paternal allele is unmethylated, while the silent mater- used in our studies has its Chr 2 breakpoint at the agoutinal allele is fully methylated. This result further indicated locus in band 2H1. Subsequent genetic and cytogeneticthat Nnat is an imprinted gene. studies defined the imprinting region as lying between the
breakpoints of two reciprocal translocations T2Wa (whichis distal to T26H) and T28H. Surprisingly, RT–PCR analysis
DISCUSSION and FISH indicated that Nnat maps proximal to T2Wa. Thissuggests that there are perhaps two distinct imprinting re-gions on distal Chr 2; one region between the T26H andIn this study we have identified Nnat as a paternally ex-
pressed novel imprinted gene analyzing its expression in T2Wa, and the other between the T2Wa and T28H translo-
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba

75Genetic Imprinting of neuronatin
cation breakpoints. It will be important in the future to types of embryos and newborns with maternal or paternalduplication distal to the T2Wa and T26H translocationdetermine whether the two imprinting regions are con-
trolled independently or whether they are linked mechanis- breakpoints were apparently similar even though embryosgenerated using the T2Wa, but not those with the T26Htically, for example, if they share a major regulatory element
such as the imprinting centre defined in the human Chr translocation, will have normal amounts of the Nnat geneproduct. It is possible that the imprinting effects associated15q region (Buiting et al., 1995).
We carried out detailed expression and phenotypic inves- with the region distal to the T2Wa breakpoint may obscureany additional effects associated with the more proximaltigations in PatDp.dist2.T26H and MatDp.dist2.T26H em-
bryos. Expression of Nnat in the former embryos was identi- region lying between the T2Wa and T26H breakpoints (i.e.,where Nnat lies). Further careful analysis is necessary tocal to that in the corresponding control embryos. However,
we predict that unlike in the control embryos, both copies clarify this. Another approach would be to compare micecarrying maternal and paternal duplications proximal to theof the paternally inherited Nnat gene are expressed in the
PatDp.dist2.T26H, although we cannot confirm this di- T2Wa and T26H breakpoints, but these arise at low frequen-cies (1–5%) so that few have been seen. Thus far four em-rectly in the absence of polymorphic variants of the two
alleles (but the DNA methylation analysis supports this bryos of genotype MatDp.prox2.T2Wa, presumed to havean absence of Nnat expression, have been examined be-notion as described below). By contrast, no expression was
detected in the MatDp.dist2.T26H embryos in any of the tween 13.5 and 19.5 dpc, and none have shown detectablephenotypic abnormalities (Cattanach and Beechey, 1995,tissues examined during development. This is proof that
both the maternally inherited copies of the gene are re- and unpublished). However, there is some indication thatthey may be smaller than their wild-type sibs (60–80%),pressed. Although leaky expression from the repressed ma-
ternal allele was detected by increasing the PCR cycles as although the 19.5-dpc embryo was successfully fostered tosurvive for 8 days. Two MatDp.prox2.T26H mice, presumeddescribed above, this is not surprising because other im-
printed genes such as Igf2 (Sasaki et al., 1992) and H19 to have normal Nnat expression, have been observed, oneexamined at 15.5 dpc and another that survived to adult-(Leighton et al., 1995) also show leaky expression from the
repressed allele. hood; intriguingly both of these appeared to be of normalsize and have a normal phenotype (Cattanach and Beechey,The methylation status of Nnat was also examined in the
PatDp.dist2.T26H and MatDp.dist2.T26H embryos. Both unpublished data). This leads to the speculation that theimprinted gene, Nnat (and possibly other imprinted genes),paternal copies of the gene were unmethylated in the former
and both maternal copies were methylated in the latter, which lies between the T26H and T2Wa breakpoints couldbe responsible for the small size observed in MatDp.prox2.-while in the control, only one allele (presumably maternal)
was methylated. These findings are consistent with the T2Wa mice. The complexity of genotype and low frequencyof occurrence of maternal and paternal duplication embryoscomplete repression of Nnat in the MatDp.dist2.T26H em-
bryos. The data also strongly suggest that both copies of does not allow us to exclude late effects, more subtle abnor-malities, or functional/behavioral effects caused by defi-Nnat are probably expressed in the PatDp.dist2.T26H em-
bryos. The differential methylation observed for Nnat is a ciency of Nnat. We are now undertaking a direct approachto understand the functions of Nnat more precisely.characteristic feature of imprinted genes. For instance, the
active maternal allele of H19 is unmethylated in the CpGisland promoter and in the 5* flanking region, while the ACKNOWLEDGMENTSinactive paternal allele is hypermethylated in these regions
We thank F. Ishino for valuable information and materials prior(Ferguson-Smith et al., 1993; Bartolomei et al., 1993). Theto publication and all the members of our laboratories for theirregion 1 of Igf2r gene containing the promoter with a CpGhelp and advice. We are especially grateful to W. Rideout III for helpisland is unmethylated and the region 2 in an intron iswith molecular studies; K. J. Hilton for preparation of embryos; M.
methylated only in the transcriptionally active maternal Narasimha for help with in situ hybridization; S. Aparicio and E. B.allele (Stoger et al., 1993). Keverne for useful suggestions, the histology department at MRC
The PatDp.dist2.T26H and the MatDp.dist2.T26H will for sections; L. Cobb for advice on histological sections; K. M. S.probably have excess and deficiency of Nnat functional gene Townsend for assistance with analysis of FISH results, H. J. Miller
and S. F. Wroe for Nnat and Hprt primers, respectively; K. Gloverproduct, respectively, making it possible to look for theirfor photographs; and D. Brooker for looking after the mice. Thisphenotypic consequences. There were no overt structuralwork was supported by the Wellcome Trust (Grant 036481), thedefects detectable in areas where the gene is known to beInternational Human Frontier Science Program Organizationexpressed. The cerebellum is changing rapidly around the(Grant RG-516/94M), and the Medical Research Council. N.K. has atime of birth and the decreased cerebellar folding in new-postdoctoral fellowship from the Sankyo Foundation of Life Scienceborn MatDp.dist2.T26H compared to PatDp.dist2.T26Hand is also supported by the Nakayama Foundation for Human
may reflect differences in the rate of development of the Science and the Daiwa Anglo–Japanese Foundation. The animaltwo genotypes or, more interestingly, may be related to studies described in this paper were carried out under the guidanceexpression differences in Nnat particularly because differ- issued by the Medical Research Council in ‘‘Responsibility in theences in cerebellar folding were not seen in MatDp.dist2.- Use of Animals for Medical Research’’ (July 1993) and Home Office
Project Licence No. 30/00875.T2Wa and PatDp.dist2.T2Wa. In other respects, the pheno-
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba

76 Kikyo et al.
cal analysis of genetic imprinting in mouse development. Dev.REFERENCESGenet. 15, 515–522.
Gage, F. H., Ray, J., and Fisher, L. J. (1995). Isolation, characteriza-Allen, N. D., Logan, K., Lally, G., Drage, D. J., Norris, M. L., and tion, and use of stem cells from the CNS. Annu. Rev. Neurosci.
Keverne, E. B. (1995). Distribution of parthenogenetic cells in 18, 159–192.the mouse brain and their influence on brain development and Gold, J. D., and Pedersen, R. A. (1994). Mechanisms of genomicbehavior. Proc. Natl. Acad. Sci. USA 92, 10782–10786. imprinting in mammals. Curr. Top. Dev. Biol. 29, 227–280.
Bartolomei, M. S., Webber, A. L., Brunkow, M. E., and Tilghman, Hayashizaki, Y., Shibata, H., Hirotsune, S., Sugino, H., Okazaki,S. M. (1993). Epigenetic mechanisms underlying the imprinting Y., Sasaki, N., Hirose, K., Imoto, H., Okuizumi, H., Muramatsu,of the mouse H19 gene. Genes Dev. 7, 1663–1673. M., Komatsubara, H., Shiroishi, T., Moriwaki, K., Katsuki, M.,
Barton, S. C., Norris, M. L., and Surani, M. A. H. (1987). Nuclear Hatano, N., Sasaki, H., Ueda, T., Mise, N., Takagi, N., Plass, C.,transplantation in fertilized and parthenogenetically activated and Chapman, V. M. (1994). Identification of an imprinted U2afeggs. In ‘‘Mammalian Development, A Practical Approach’’ (M. binding protein related sequence on mouse chromosome 11 usingMonk, Ed.), pp. 235–253. IRL Press, Oxford. the RLGS method. Nat. Genet. 6, 33–40.
Barton, S. C., and Surani, M. A. (1993). Manipulations of genetic Joseph, R., Dou, D., and Tsang, W. (1994). Molecular cloning of aconstitution by nuclear transplantation. In ‘‘Methods in Enzy- novel mRNA (neuronatin) that is highly expressed in neonatalmology’’ (P. M. Wassarman and M. L. DePamphilis, Eds.), Vol. mammalian brain. Biochem. Biophys. Res. Commun. 201, 1227–225, pp. 732–744. Academic Press, San Diego. 1234.
Beechey, C. V., and Cattanach, B. M. (1997). Genetic and physical Kagitani, F., Kuroiwa, Y., Wakana, S., Shiroishi, T., Miyoshi, N.,imprinting map of the mouse. Mouse Genome 95, 100–105. Kobayashi, S., Nishida, M., Kohda, T., Kaneko-Ishino, T., and
Brenton, J. D., Viville, S., and Surani, M. A. (1995). Genomic im- Ishino, F. (1997). Peg5/Neuronatin is an imprinted gene locatedprinting and cancer. In ‘‘Cancer Surveys, Genetics and Cancer: on sub-distal chromosome 2 in the mouse. Nucleic Acids Res.A Second Look’’ (B. A. J. Ponder, W. K. Cavenee, and E. Solomon, (in press).Eds.), Vol. 25, pp. 161–171. Cold Spring Harbor Laboratory Press, Kaneko-Ishino, T., Kuroiwa, Y., Miyoshi, N., Kohda, T., Suzuki,Cold Spring Harbor, NY. R., Yokoyama, M., Viville, S., Barton, S. C., Ishino, F., and Surani,
Buiting, K., Saitoh, S., Gross, S., Dittrich, B., Schwartz, S., Nicholls, M. A. (1995). Peg1/Mest imprinted gene on chromosome 6 identi-R. D., and Horsthemke, B. (1995). Inherited microdeletions in fied by cDNA subtraction hybridization. Nat. Genet. 11, 52–59.the Angelman and Prader-Willi syndromes define an imprinting Keverne, E. B. (1995). Olfactory learning. Curr. Opin. Neurobiol. 5,centre on human chromosome 15. Nat. Genet. 9, 395–400. 482–488.
Carpenter, M. B. (1991). ‘‘Core Text of Neuroanatomy,’’ 4th ed., Keverne, E. B., Fundele, R., Narasimha, M., Barton, S. C., and Sur-Williams and Wilkins, Baltimore. ani, M. A. (1996). Genomic imprinting and the differential roles
Cattanach, B. M. (1986). Parental origin effects in mice. J. Embryol. of parental genomes in brain development. Dev. Brain Res. 92,Exp. Morphol. 97(Suppl.), 137–150. 91–100.
Cattanach, B. M., and Beechey, C. V. (1995). Reassessment of the Konecki, D. S., Brennand, J., Fuscoe, J. C., Caskey, C. T., and Chi-proximal Chr 2 imprinting lethality. Mouse Genome 93, 858– nault, A. C. (1982). Hypoxanthine-guanine phosphoribosyltrans-859. ferase genes of the mouse and Chinese hamster: Construction
Cattanach, B. M., Evans, E. P., Burtenshaw, M., and Beechey, C. V. and sequence analysis of cDNA recombinants. Nucleic Acids(1992). Further delimitation of the distal chromosome 2 im- Res. 10, 6763–6775.printing region. Mouse Genome 90, 82. Kuroiwa, Y., Kaneko-Ishino, T., Kagitani, F., Kohda, T., Li, L.-L.,
Cattanach, B. M., and Jones, J. (1994). Genetic imprinting in the Tada, M., Suzuki, R., Yokoyama, M., Shiroishi, T., Wakana, S.,mouse; implications for gene regulation. J. Inher. Metab. Dis. 17, Barton, S. C., Ishino, F., and Surani, M. A. (1996). Peg3 imprinted403–420. gene on proximal chromosome 7 encodes for a zinc finger protein.
Cattanach, B. M., and Kirk, M. (1985). Differential activity of ma- Nat. Genet. 12, 186–190.ternally and paternally derived chromosome regions in mice. Na- Latham, K. E., McGrath, J., and Solter, D. (1995). Mechanistic andture 315, 496–498. developmental aspects of genetic imprinting in mammals. Int.
Chomczynski, P., and Sacchi, N. (1987). Single-step method of RNA Rev. Cytol. 160, 53–98.isolation by acid guanidinium thiocyanate–phenol–chloroform Ledbetter, D. H., and Engel, E. (1995). Uniparental disomy in hu-extraction. Anal. Biochem. 162, 156–159. mans: Development of an imprinting map and its implications
de Boer, P., and van Gijsen, M. (1974). The location of the positions for prenatal diagnosis. Hum. Mol. Genet. 4, 1757–1764.of the breakpoints involved in the T26H and T70H mouse trans- Leighton, P. A., Ingram, R. S., Eggenschwiler, J., Efstratiadis, A.,locations with the aid of Giemsa-banding. Can. J. Genet. Cytol. and Tilghman, S. M. (1995). Disruption of imprinting caused by16, 783–788. deletion of the H19 gene region in mice. Nature 375, 34–39.
Dou, D., and Joseph, R. (1996). Cloning of human neuronatin gene Liang, P., Bauer, D., Averboukh, L., Warthoe, P., Rohrwild, M.,and its localization to chromosome-20q11.2-12: The deduced pro- Muller, H., Strauss, M., and Pardee, A. B. (1995). Analysis of al-tein is a novel ‘proteolipid’. Brain Res. 723, 8–22. tered gene expression by differential display. In ‘‘Methods in En-
Evans, E. P. (1987). Karyotyping and sexing of gametes, embryos zymology’’ (P. K. Vogt and I. M. Verma, Eds.), Vol. 254, pp. 304–and fetuses and in situ hybridization to chromosomes. In ‘‘Mam- 321. Academic Press, San Diego.malian Development: A Practical Approach’’ (M. Monk, Ed.), pp. Plumb, M., Stein, J., and Stein, G. (1983). Coordinate regulation of93–114. IRL Press, Oxford. multiple histone mRNAs during the cell cycle in HeLa cells.
Ferguson-Smith, A. C., Sasaki, H., Cattanach, B. M., and Surani, Nucleic Acids Res. 11, 2391–2410.M. A. (1993). Parental-origin-specific epigenetic modification of Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). ‘‘Molecularthe mouse H19 gene. Nature 362, 751–755. Cloning: A Laboratory Manual.’’ Cold Spring Harbor Laboratory
Press, New York.Fundele, R. H., and Surani, M. A. (1994). Experimental embryologi-
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba

77Genetic Imprinting of neuronatin
Sasaki, H., Jones, P. A., Chaillet, J. R., Ferguson-Smith, A. C., Bar- Wijnholds, J., Chowdhury, K., Wehr, R., and Gruss, P. (1995). Seg-ment-specific expression of the neuronatin gene during earlyton, S. C., Reik, W., and Surani, M. A. (1992) Parental imprinting:hindbrain development. Dev. Biol. 171, 73–84.Potentially active chromatin of the repressed maternal allele of
Wilkinson, D. G., and Nieto, M. A. (1993). Detection of messengerthe mouse insulin-like growth factor II (Igf2) gene. Genes Dev.RNA by in situ hybridization to tissue sections and whole6, 1843–1856.mounts. In ‘‘Methods in Enzymology’’ (P. M. Wassarman andSearle, A. G., and Beechey, C. V. (1978). Complementation studiesM. L. DePamphilis, Eds.), Vol. 225, pp. 361–373. Academic Press,with mouse translocations. Cytogenet. Cell. Genet. 20, 282–303.San Diego.Searle, A. G., Ford, C. E., and Beechey, C. V. (1971). Meiotic disjunc-
Williamson, C. M., Miller, H. J., Beechey, C. V., and Peters, J.tion in mouse translocations and the determination of centro-(1995). Microsatellite marker D2Mit226 for the identification ofmere position. Genet. Res. 18, 215–235.duplication/deficient mice for the distal region of Chr 2. MouseStoger, R., Kubicka, P., Liu, C.-G., Kafri, T., Razin, A., Ceder, H.,Genome 93, 860.and Barlow, D. P. (1993). Maternal-specific methylation of the
imprinted mouse Igf2r locus identifies the expressed locus as Received for publication February 13, 1997Accepted June 16, 1997carrying the imprinting signal. Cell 73, 61–71.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8681 / 6x2c$$$181 09-05-97 09:16:43 dba




















