
Galactosylated dopamine enters into the brain, blocks the mesocorticolimbic system and modulates...
11
GALACTOSYLATED DOPAMINE ENTERS INTO THE BRAIN, BLOCKS THE MESOCORTICOLIMBIC SYSTEM AND MODULATES ACTIVITY AND SCANNING TIME IN NAPLES HIGH EXCITABILITY RATS L. A. RUOCCO, a D. VIGGIANO, a A. VIGGIANO, a E. ABIGNENTE, b M. G. RIMOLI, b D. MELISI, b A. CURCIO, b M. NIEDDU, c G. BOATTO, c E. CARBONI, d U. A. GIRONI CARNEVALE a1 AND A. G. SADILE a * a Department of Experimental Medicine, Faculty of Medicine, II Univer- sity of Naples, Naples, Italy b Department of Pharmaceutical and Toxicological Chemistry, Faculty of Pharmacy, University of Naples “Federico II,” Naples, Italy c Department of Pharmacology, Chemistry and Toxicology, Faculty of Pharmacy, University of Sassari, Sassari, Italy d Department of Toxicology, Faculty of Pharmacy, University of Cagli- ari, Cagliari, Italy Abstract—Pathological conditions, such as Parkinson’s dis- ease and attention deficit hyperactivity disorder, have been linked to alterations of specific dopamine (DA) pathways. However, since exogenous DA does not cross the blood– brain barrier, DA levels can be modulated e.g. by DA precur- sors or DA reuptake blockers. Hereby histochemical, analyt- ical and behavioral evidence shows that a galactosylated form of DA (GAL-DA) carries DA into the brain, thus modu- lating activity and nonselective attention in rats. To this aim adult male rats of the Naples high-excitability (NHE) and random bred controls (NRB) lines were given a single i.p. injection of GAL-DA (10 or 100 mg/kg). Three hours later the behavior was videotaped and analyzed for horizontal activity, orienting frequency and scanning duration. The dose of 100 mg/kg of GAL-DA reduced by 25% the horizontal activity in NHE rats, mainly in the first part of the testing period. No effect was observed on orienting frequency or on scanning duration. However, GAL-DA 100 mg/kg was associated with longer rearing episodes in the second part of the testing period in NHE rats. In parallel experiments histochemistry with a galactose- specific lectin showed 10% increase in galactose residues into the striatum between 0.5 and 3.0 h. To quantify the level of GAL-DA, its metabolite DA-succinate and DA in the prefrontal cortex, neostriatum, and cerebellum, rats were killed 2.0 h after the injection of prodrug. Mass high performance liquid chroma- tography (HPLC) was used for analysis of GAL-DA and DA succinate whereas electrochemical HPLC for DA. Both HPLC techniques demonstrate that GAL-DA carries and releases DA into the brain. Specifically 100 mg/kg of GAL-DA increased DA level in the striatum in the NHE rats only. Moreover, DA in the mesencephalon (MES) was correlated positively with striatal and prefrontal cortex DA in NHE rats. In contrast DA in the MES was negatively correlated with striatal DA in NRB. GAL-DA dis- rupted these correlations in both rat lines. Thus, this new DA prodrug may modify DA neurotransmission and might have a potential clinical application. © 2008 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: Naples rat lines, prodrug, behavior, HPLC, lectin histochemistry, ADHD. The neurotransmitter dopamine (DA) is involved in differ- ent control systems, such as motor activity, attentive pro- cesses, prediction of reward, and prolactin secretion (Mis- sale et al., 1998; Bentivoglio and Morelli, 2005). DA is synthesized by groups of neurons located in the basal brain (Lindvall and Bjorklund, 1974). Extracellular DA level is mainly regulated by a passive diffusion and an active reuptake system, i.e. the dopamine membrane transporter (DAT) (for a review see e.g. (Viggiano et al., 2004)). Several diseases have been linked to alterations of specific DA pathways. For instance, Parkinson’s disease (PD) and some neuropsychiatric disorders, such as atten- tion deficit hyperactivity disorder (ADHD) (Sergeant et al., 2003; Faraone et al., 2003), are characterized by altered levels of brain DA (Fernandez-Espejo, 2004). The thera- peutic approach consists in modulating brain DA levels (So- lanto, 1998; Olanow et al., 2004; Pisani et al., 2005). How- ever, DA does not cross the blood– brain barrier (BBB), mainly because of its hydrophilicity and the absence of spe- cific carrier system (Bonina et al., 2003; Dalpiaz et al., 2007). Therefore the current therapeutic strategy for PD is based on L-DOPA, a DA precursor, that enters the CNS by the amino acid transporter (system L) (Mercuri and Bernardi, 2005) and is enzymatically converted into DA (Djaldetti and Melamed, 2001). However, the L-DOPA treatment is associated with side effects such as motor fluctuations, probably due to fast increase in brain DA level (Mercuri and Bernardi, 2005). Therefore, new therapeutic strategies which increase or mod- ulate brain DA level are necessary. Hydrophilic prodrugs could cross the BBB after glyco- or galactosylation, by carrier- mediated membrane transport mechanisms, such as the glu- cose transporters (GLUT). More specifically, the conjugation of potentially CNS-active drugs with amino acids and hex- oses actively transported across the BBB (Polt et al., 1994; Bonina et al., 1999; Dalpiaz, et al., 2007) represents a prom- ising strategy. In fact, glucose, galactose and other hexoses are transported across the BBB by the glucose carrier Glut-1, that is located on the membrane of endothelial cells (Old- 1 Now at Doctorate in Neuroscience, Department of Neuroscience, Univ. Naples “Federico II,” Italy. *Corresponding author. Tel: 39-081-5665827; fax: 39-081-5665827. E-mail address: [email protected] (A. G. Sadile). Abbreviations: ADHD, attention deficit hyperactivity disorder; ANOVA, analysis of variance; BBB, blood– brain barrier; DA, dopamine; DAT, dopamine membrane transporter; GAL-DA, galactosylated form of dopamine; HPLC, high performance liquid chromatography; NHE, Na- ples high excitability; NRB, random bred control; PD, Parkinson’s disease; ROD, relative optical density. Neuroscience 152 (2008) 234 –244 0306-4522/08$32.000.00 © 2008 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2007.11.021 234
Transcript of Galactosylated dopamine enters into the brain, blocks the mesocorticolimbic system and modulates...

GTA
LEAUa
sb
oc
Pd
a
AelHbsiflaribomNedlp
siGcttstilm
1
U*EAaddpd
Neuroscience 152 (2008) 234–244
0d
ALACTOSYLATED DOPAMINE ENTERS INTO THE BRAIN, BLOCKSHE MESOCORTICOLIMBIC SYSTEM AND MODULATES ACTIVITY
ND SCANNING TIME IN NAPLES HIGH EXCITABILITY RATSawrppE
Kh
Tecssbir(
s(t2lplemcTL
ai2siTucmcooBi
t
. A. RUOCCO,a D. VIGGIANO,a A. VIGGIANO,a
. ABIGNENTE,b M. G. RIMOLI,b D. MELISI,b
. CURCIO,b M. NIEDDU,c G. BOATTO,c E. CARBONI,d
. A. GIRONI CARNEVALEa1 AND A. G. SADILEa*
Department of Experimental Medicine, Faculty of Medicine, II Univer-ity of Naples, Naples, Italy
Department of Pharmaceutical and Toxicological Chemistry, Facultyf Pharmacy, University of Naples “Federico II,” Naples, Italy
Department of Pharmacology, Chemistry and Toxicology, Faculty ofharmacy, University of Sassari, Sassari, Italy
Department of Toxicology, Faculty of Pharmacy, University of Cagli-ri, Cagliari, Italy
bstract—Pathological conditions, such as Parkinson’s dis-ase and attention deficit hyperactivity disorder, have beeninked to alterations of specific dopamine (DA) pathways.owever, since exogenous DA does not cross the blood–rain barrier, DA levels can be modulated e.g. by DA precur-ors or DA reuptake blockers. Hereby histochemical, analyt-cal and behavioral evidence shows that a galactosylatedorm of DA (GAL-DA) carries DA into the brain, thus modu-ating activity and nonselective attention in rats. To this aimdult male rats of the Naples high-excitability (NHE) andandom bred controls (NRB) lines were given a single i.p.njection of GAL-DA (10 or 100 mg/kg). Three hours later theehavior was videotaped and analyzed for horizontal activity,rienting frequency and scanning duration. The dose of 100g/kg of GAL-DA reduced by 25% the horizontal activity inHE rats, mainly in the first part of the testing period. Noffect was observed on orienting frequency or on scanninguration. However, GAL-DA 100 mg/kg was associated with
onger rearing episodes in the second part of the testingeriod in NHE rats.
In parallel experiments histochemistry with a galactose-pecific lectin showed 10% increase in galactose residuesnto the striatum between 0.5 and 3.0 h. To quantify the level ofAL-DA, its metabolite DA-succinate and DA in the prefrontalortex, neostriatum, and cerebellum, rats were killed 2.0 h afterhe injection of prodrug. Mass high performance liquid chroma-ography (HPLC) was used for analysis of GAL-DA and DAuccinate whereas electrochemical HPLC for DA. Both HPLCechniques demonstrate that GAL-DA carries and releases DAnto the brain. Specifically 100 mg/kg of GAL-DA increased DAevel in the striatum in the NHE rats only. Moreover, DA in the
esencephalon (MES) was correlated positively with striatal
Now at Doctorate in Neuroscience, Department of Neuroscience,niv. Naples “Federico II,” Italy.
Corresponding author. Tel: �39-081-5665827; fax: �39-081-5665827.-mail address: [email protected] (A. G. Sadile).bbreviations: ADHD, attention deficit hyperactivity disorder; ANOVA,nalysis of variance; BBB, blood–brain barrier; DA, dopamine; DAT,opamine membrane transporter; GAL-DA, galactosylated form ofopamine; HPLC, high performance liquid chromatography; NHE, Na-
tles high excitability; NRB, random bred control; PD, Parkinson’sisease; ROD, relative optical density.
306-4522/08$32.00�0.00 © 2008 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2007.11.021
234
nd prefrontal cortex DA in NHE rats. In contrast DA in the MESas negatively correlated with striatal DA in NRB. GAL-DA dis-
upted these correlations in both rat lines. Thus, this new DArodrug may modify DA neurotransmission and might have aotential clinical application. © 2008 IBRO. Published bylsevier Ltd. All rights reserved.
ey words: Naples rat lines, prodrug, behavior, HPLC, lectinistochemistry, ADHD.
he neurotransmitter dopamine (DA) is involved in differ-nt control systems, such as motor activity, attentive pro-esses, prediction of reward, and prolactin secretion (Mis-ale et al., 1998; Bentivoglio and Morelli, 2005). DA isynthesized by groups of neurons located in the basalrain (Lindvall and Bjorklund, 1974). Extracellular DA level
s mainly regulated by a passive diffusion and an activeeuptake system, i.e. the dopamine membrane transporterDAT) (for a review see e.g. (Viggiano et al., 2004)).
Several diseases have been linked to alterations ofpecific DA pathways. For instance, Parkinson’s diseasePD) and some neuropsychiatric disorders, such as atten-ion deficit hyperactivity disorder (ADHD) (Sergeant et al.,003; Faraone et al., 2003), are characterized by altered
evels of brain DA (Fernandez-Espejo, 2004). The thera-eutic approach consists in modulating brain DA levels (So-
anto, 1998; Olanow et al., 2004; Pisani et al., 2005). How-ver, DA does not cross the blood–brain barrier (BBB),ainly because of its hydrophilicity and the absence of spe-
ific carrier system (Bonina et al., 2003; Dalpiaz et al., 2007).herefore the current therapeutic strategy for PD is based on-DOPA, a DA precursor, that enters the CNS by the aminocid transporter (system L) (Mercuri and Bernardi, 2005) and
s enzymatically converted into DA (Djaldetti and Melamed,001). However, the L-DOPA treatment is associated withide effects such as motor fluctuations, probably due to fast
ncrease in brain DA level (Mercuri and Bernardi, 2005).herefore, new therapeutic strategies which increase or mod-late brain DA level are necessary. Hydrophilic prodrugsould cross the BBB after glyco- or galactosylation, by carrier-ediated membrane transport mechanisms, such as the glu-
ose transporters (GLUT). More specifically, the conjugationf potentially CNS-active drugs with amino acids and hex-ses actively transported across the BBB (Polt et al., 1994;onina et al., 1999; Dalpiaz, et al., 2007) represents a prom-
sing strategy.In fact, glucose, galactose and other hexoses are
ransported across the BBB by the glucose carrier Glut-1,
hat is located on the membrane of endothelial cells (Old-ved.
egpgcd
cpdeitia(N((tVSAect(
libp
A
AaroloCHDob
E
Tespehwbc
ttw
ha
E
Tced(
Gadb
ginia
ttobPc
Hi
NsTla4c5airCc
I
SCw(apErlr
Co
N1thEf
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244 235
ndorf, 1971; Mueckler, 1994). Consequently, glycoconju-ated drugs may cross the BBB by carrier-mediated trans-ort (Glut-1). Upon reaching the CNS, the glycosyl conju-ate should be enzymatically cleaved to release the activeompound (Halmos et al., 1996; Uriel et al., 1996; Fernan-ez et al., 2000).
Previously, Bonina et al. (2003) demonstrated that DAovalently linked to galactose (galactosylated form of do-amine, GAL-DA) by a succinic acid spacer, is a stableerivative displaying central effects on standard DA mod-ls, such as morphine-induced locomotion and reserpine-
nduced hypolocomotion. The aim of this study is to inves-igate the entry of prodrug, the release of DA by the vectornto the brain and the behavioral effects of GAL-DA onctivity and non-selective attention in a spatial noveltyAspide et al., 1998, 2000). This was carried out on theaples high excitability (NHE) and random bred control
NRB) rats. The NHE rats feature some aspects of ADHDsee e.g. (Sergeant et al., 2003; Faraone et al., 2003)), ashey are hyperactive and inattentive (Cerbone et al., 1993;iggiano et al., 2002b; Gironi Carnevale et al., 2007).pecifically, they may model the mesocortical variant ofDHD (see e.g. (Viggiano et al., 2003)). Since severalvidences suggest the mesocortical branch of mesocorti-olimbic system to be hyperfunctioning, GAL-DA may be aool to block it by mesencephalic D2-DA autoreceptorsGrace et al., 1997).
To verify effects of DA released by GAL-DA on high andow affinity target sites, two extreme doses have been used,.e. 10 and 100 mg/kg. The latter has been demonstrated toe below toxicity threshold because peripheral hydrolysisroduce mainly DA-succinate (Bonina et al., 2003).
EXPERIMENTAL PROCEDURES
nimals
dult male of the NHE (n�80) and NRB (n�80) lines from ournimal colony were used throughout. The experiments were car-ied out on 55 to 60-day-old animals. Rats were housed in groupsf two in Macrolon cages on standard conditions. An artificial 12-h
ight/dark cycle was obtained by lights on from 6 AM to 6 PM. Allther parameters fulfilled the requirements of the “Guide for theare and Use of Laboratory Animals” by the National Institutes ofealth (NIH), integrated with the European Communities Councilirective of Nov. 24, 1986 (86/609/EEC). In particular, the numberf animals and their suffering has been minimized and approvedy Bioethical Committee.
xperimental apparatus
he experimental system was a Làt-maze described in detailslsewhere (Viggiano et al., 2002b). Briefly, it consisted of aquared wooden (60�60�40 cm) box with a smaller plastic trans-arent box (40�40�30 cm) into the center. Rats were allowed toxplore the resulting corridor (60 cm long, 15 cm wide and 40 cmigh) (see Sadile et al., 1988 for details). A set of two such boxesas located in a sound-attenuated room. The box was illuminatedy a white, cold 4 W lamp placed 60 cm above the floor in theenter of the wooden cover, providing a 0.1–0.2 �W/cm2.
Rats were exposed to the Làt-maze during a 30-min test athe beginning of the light phase of LD cycle. In order to minimizehe interference with the arousal state the pair from the same cage
as tested at the same time. Rat behavior was monitored by a cigh resolution CCD camera and stored on a tape recorder to benalyzed off-line.
xperimental procedures
he DA system has been probed using a new-synthesis drugonsisting of a galactosylated form of dopamine (GAL-DA) that isxpected to cross the BBB and release DA in the brain (see foretails (Bonina et al., 2003)). Drug has been dissolved in salineNaCl 0.9%). Two doses have been tested: 10 mg/kg and 100 mg/kg.
In order to test possible effects of galactose released fromAL-DA molecule, two control groups receiving an equimolarmount of galactose, i.e. 5 mg/kg and 50 mg/kg respectively,issolved into saline, were used. Drugs have been injected i.p.etween 9:00 and 11:00 AM.
The animals were randomly assigned to treatment and controlroups (n�5 per group). The behavioral analysis was carried out
n double blind conditions. Animals were tested for activity andon-selective attention, 3 h after drug or control injection. This
nterval was chosen on the basis of pilot experiments (Bonina etl., 2003), showing its maximal effect 3 h after the injection.
The behavioral variables were traveled distance, as moni-ored by the frequency of corner crossings, frequency and dura-ion of rearings on hind limbs or leanings against the walls withne or both forepaws. They were visually measured per 3-minlocks. The reliability index was quite high (r�0.914; df�198;�0.001). At the end of the test, the number of fecal boluses wasounted and the floor was carefully cleaned with a wet sponge.
istochemical detection of galactose residuesn the brain
HE and NRB rats between 9:00 and 11:00 AM were given aingle injection of GAL-DA 100 mg/kg (n�20) or vehicle (n�5).he latter was PBS with no galactose. Different time points fol-
owing injection were chosen (0, 0.5, 1.5, 3 and 6 h, n�5/group)nd rats were perfused with saline, followed by paraformaldehyde% in PBS. Brains were postfixed for 2h in the same fixative,ryoprotected in sucrose 18% and frozen on dry-ice. Free floating0 �m-thick cryosections were stained for galactose residues bygalactose-specific biotinylated lectin (Bandeiraea simplicifolia
solectin B4; 1:100 in PBS overnight). Specific binding sites wereevealed by the ABC system (Vector Laboratories, Burlingame,A, USA) and diaminobenzidine (Sigma-Aldrich, Milano, Italy) ashromogen.
mage analysis
lices were analyzed with a Zeiss Axioscope equipped with aCD camera (Hamamatsu Photonics, Italy, C5405). The imagesere converted by a microcomputer-assisted image analyzer
MCID-Elite; Imaging Res. Inc., Canada) to a 640�512 pixel filend quantitatively analyzed, according to the guidelines of Ca-owski (1989). Measurements were obtained by a 5� objective.ight fields of the striatum were sampled for each slice. The
elative optical density (ROD) [ROD�log10 (256/observed grayevels)] was measured in each field. ROD units are linearly cor-elated to the specimen concentration (Capowski (1989).
apillary depletion study and LC-MS analysisf GAL-DA derivatives
HE and NRB rats between 9:00 and 11:00 AM were injected with00 mg/kg GAL-DA (n�5) or PBS vehicle (n�5). Two hours afterhe GAL-DA or saline injection, the animal was anesthetized, theeart exposed and a blood sample was taken intra-cardially.DTA (0.1 M) was added to the sample, centrifuged at 13,000�g
or 10 min. The supernatant was used for further analysis.The rat was then perfused with 10 ml PB to remove red blood
ells from brain parenchyma. The brain was isolated and treated

bacEsoaim
Lpninpgte4(T1g
H(
A1inw(
icm�5panIsouw
S
Gasof1
dpgamsoifcfbac
wsvct
vm
H
w
B
oigncrsPnGamei
stsAs9w
cafre
T
T
05577
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244236
y the capillary depletion technique (Triguero et al., 1990; Shimura etl., 1992). It was further dissected into prefrontal cortex, striatum anderebellum. Brain samples were homogenized in 1 ml ice-cold PB inppendorf tubes and centrifuged at 4500�g at 4 °C for 15 min. Theupernatant was then used for LC-MS analysis. As previously dem-nstrated, while the homogenate contains material from vascularnd non-vascular parenchymal components and, the supernatant
s 95% devoid of blood vessels, the pellet consists of cerebralicrovessels (Triguero et al., 1990).
An Agilent Technologies (Palo Alto, CA, USA) 1100 seriesC/MSD fitted with Rheodyne injector 20 �l loop was used. Ahotodiode array detector was directly coupled to the sprayereedle, where ions were generated by electrospray ionization ESI
n both positive and negative ionization modes. Nitrogen was used asebulizing and drying gas and different fragmentor voltage was ap-lied. Selected ion monitoring (SIM) analyses at m/z 398 for DA-alactose and m/z 254 for DA-succinate were carried out to obtainhe maximum signal intensity for the protonated molecular ion ofach analysis. The analytical column was a Luna® C18 (250 mm�.6 i.d., 5 �m) from Phenomenex, connected to a guard columncartridge 2.1 mm�12.5 mm) filled with the same packing material.he temperature of the column was at 25 °C. The total run time was0 min. The retention times were: DA-succinate 6.4 min and DA-alactose 7.8 min. The gradient program is shown in Table 1.
igh performance liquid chromatographyHPLC)-EC analysis
dult male rats of NHE and NRB lines were injected with GAL-DA00 mg/kg i.p. (n�8) or vehicle (n�12). Two hours after the
njection, animals were killed and the brains dissected out ineostriatum and ventral mesencephalon. Brain areas were weighedith a precision of 0.1 mg, homogenized in 1.8% trichloroacetic acid
3 ml/g of tissue) and centrifuged at 13,000�g for 10 min.In order to quantify DA, supernatants (20 �l) were directly
njected into an HPLC apparatus equipped with reverse-phaseolumn (BAS MF-6213) and an electrochemical detector (�650V vs. Ag/AgCl; BAS LC-4B). The working electrode was set at650 mV vs. Ag/AgCl. The eluent consisted of 50 mM NaH2PO4,mM Na2HPO4, 0.1 mM Na2EDTA, 0.5 mM SOS, 15% methanol,H 5.5; flow rate was 0.8 ml/min. The amperometric signal wascquired on a PC through a custom analog-to-digital converter con-ected to the parallel port (based on the ADS-7818 chip, Texasnstruments). Data were acquired and analyzed through customoftware written with LabView (National Instruments). Quantificationf DA in each chromatogram was obtained by calculating the areander the peak (elution time: 3.6 min). This area was then comparedith that corresponding to know DA amount.
tatistics
iven the constant behavioral overactivity of NHE vs. NRB ratscross generation of selection, that accounts for over 120, aample size of five/group was chosen, as it gave statistical powerf about 1. In fact, a mean�S.E.M. of 180�8 for NHE and 130�8or NRB, where the mean is referred to HA�VA score in the first
able 1. The gradient program used for the LC-MS analysis
ime (min) Acetonitrile(mobile phase A, %)
Formic acid 0.1%(mobile phase B, %)
.00 35 65
.00 35 65
.01 40 60
.00 40 60
.01 60 40
0 min of testing on day 1 and the S.E.M. is the maximum b
ispersion level reported during selection, gives a computedower coefficient �0.98 when we sampled five subjects perroup. Behavioral variables were frequency of corner crossingnd frequency and duration of rearings. The fulfillment of require-ents for parametric analysis of data has been assessed by
pecific tests (Levene for univariate analysis, Box test of equalityf covariance matrices for repeated measures analysis) included
n SPSS software. They were submitted to separate two-wayactorial analysis of variance (ANOVA) treatment�time blocks (asovariate). For rearing duration, cumulative frequency histogramsor the different classes of duration were computed and comparedy three-way ANOVA strain�treatment�duration class (as covari-te) followed by two-way ANOVA treatment�duration class (asovariate) for each strain.
The morphometric variable (ROD) was submitted to a one-ay ANOVA. HPLC detected DA levels in brain regions wereubmitted to a three-way factorial ANOVA treatment (GAL-DA vs.ehicle)�line (NHE vs. NRB)�region (mes, striatum, PFC, asovariate). Between-line planned comparisons were made by two-ailed t-test for non-paired data.
In order to extract the information content of within-groupariability of EC-HPLC, covariations were carried out by Spear-an’s rank-order correlation analysis (Edwards, 1967).
The rejection level was set at 2P�0.05 after correction by theölm’s procedure (Hölm, 1979).
All statistical analyses have been performed by SPSS soft-are.
RESULTS
ehavioral effects of GAL-DA
Horizontal activity. As shown in Fig. 1A, only 100 mg/kgf GAL-DA reduced horizontal activity in NHE rats, mainly
n the first part of the testing period, by 25% vs. 50 mg/kgalactose control group, whereas 10 mg/kg of GAL-DA didot produce any significant change vs. 5 mg/kg galactoseontrol group. This was demonstrated by a two-way facto-ial ANOVA treatment�time blocks (as covariate) showingignificant main effects for treatment (F�4.16, df 3/14,�0.027) and for time, (F�45.06, df 9/126, P�0.000) witho interaction effect. In contrast in NRB rats (Fig. 2A)AL-DA produces no effects on this behavioral variable asssessed by a separate two-way factorial ANOVA treat-ent�time blocks (as covariate) showing a significant mainffect for time only (F�32.31, df 9/144, P�0.000) and no
nteraction effect.
Vertical activity. Rearing frequency. GAL-DA did notignificantly affect the orienting frequency, as measured byhe number of rearings in both NHE and NRB rats ashown in Fig. 1B and 2B. In fact separate two-way factorialNOVAs treatment�time blocks (as covariate) showedignificant main effect for time only in NHE (F�13.06, df/126, P�0.000) and NRB (F�19.43, df 9/144, P�0.000),ith no interaction.
Rearing duration. Finally, GAL-DA did not signifi-antly affect the average duration of rearings in both NHEnd NRB rats as shown in Fig. 1C and 2C. Two-wayactorial ANOVAs treatment�time blocks (as covariate),un separately on the two rat lines, gave significant mainffect for time only in NHE (F�3.39, df 9/126, P�0.001)
ut not in NRB rats, with no interaction.
Ftr
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244 237
ig. 1. Behavioral effects of GAL-DA upon activity and non-selective attention in the NHE rats. The frequency of corner crossings, that measuresraveled distance (A), the frequency of rearings (B) and their duration (C) in the Làt maze is plotted over the 30-min exposure per 3-min blocks. Data
epresents mean�S.E.M.; For statistics, see Results section.
Fmb
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244238
ig. 2. Behavioral effects of GAL-DA upon activity and non-selective attention in the Naples NRB rats. The frequency of corner crossings, thateasures traveled distance (A), the frequency of rearings (B) and their duration (C) in the Làt maze is plotted over the 30-min exposure per 3-min
locks. Data represents mean�S.E.M. For statistics, see Results section.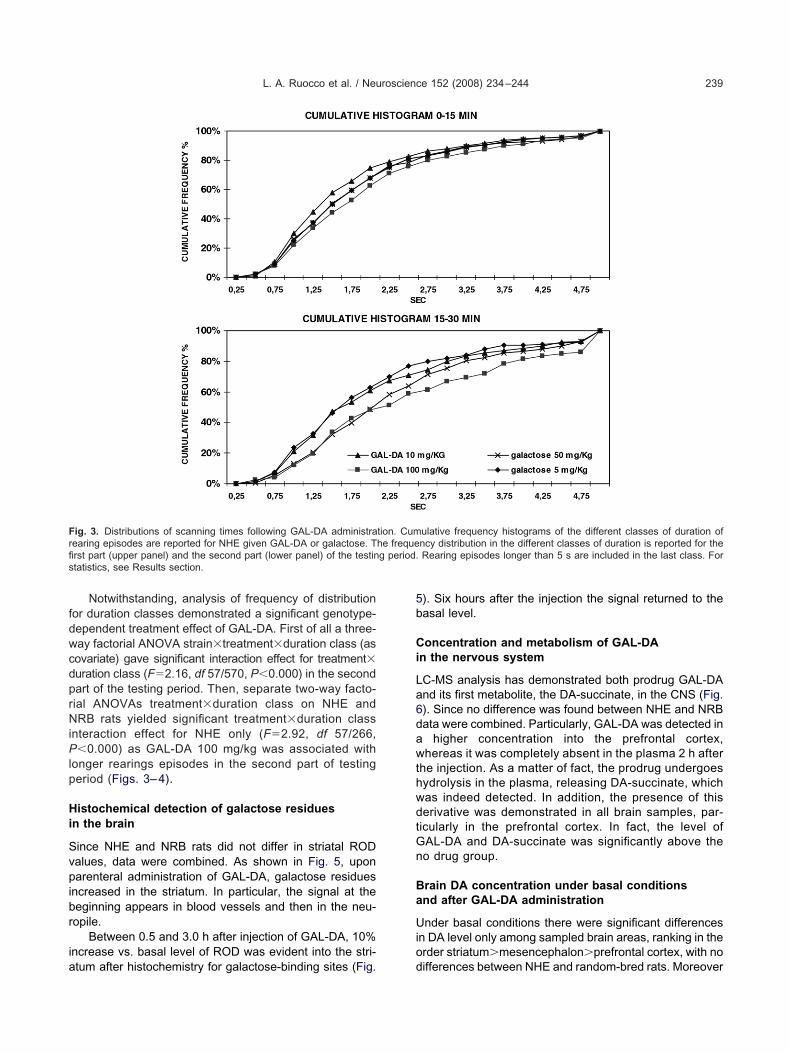
fdwcdprNiPlp
Hi
Svpibr
ia
5b
Ci
La6dawthwdtGn
Ba
Uio
Frfi g period.s
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244 239
Notwithstanding, analysis of frequency of distributionor duration classes demonstrated a significant genotype-ependent treatment effect of GAL-DA. First of all a three-ay factorial ANOVA strain�treatment�duration class (asovariate) gave significant interaction effect for treatment�uration class (F�2.16, df 57/570, P�0.000) in the secondart of the testing period. Then, separate two-way facto-ial ANOVAs treatment�duration class on NHE andRB rats yielded significant treatment�duration class
nteraction effect for NHE only (F�2.92, df 57/266,�0.000) as GAL-DA 100 mg/kg was associated with
onger rearings episodes in the second part of testingeriod (Figs. 3– 4).
istochemical detection of galactose residuesn the brain
ince NHE and NRB rats did not differ in striatal RODalues, data were combined. As shown in Fig. 5, uponarenteral administration of GAL-DA, galactose residues
ncreased in the striatum. In particular, the signal at theeginning appears in blood vessels and then in the neu-opile.
Between 0.5 and 3.0 h after injection of GAL-DA, 10%ncrease vs. basal level of ROD was evident into the stri-
ig. 3. Distributions of scanning times following GAL-DA administraearing episodes are reported for NHE given GAL-DA or galactose. Trst part (upper panel) and the second part (lower panel) of the testintatistics, see Results section.
tum after histochemistry for galactose-binding sites (Fig. d
). Six hours after the injection the signal returned to theasal level.
oncentration and metabolism of GAL-DAn the nervous system
C-MS analysis has demonstrated both prodrug GAL-DAnd its first metabolite, the DA-succinate, in the CNS (Fig.). Since no difference was found between NHE and NRBata were combined. Particularly, GAL-DA was detected in
higher concentration into the prefrontal cortex,hereas it was completely absent in the plasma 2 h after
he injection. As a matter of fact, the prodrug undergoesydrolysis in the plasma, releasing DA-succinate, whichas indeed detected. In addition, the presence of thiserivative was demonstrated in all brain samples, par-
icularly in the prefrontal cortex. In fact, the level ofAL-DA and DA-succinate was significantly above theo drug group.
rain DA concentration under basal conditionsnd after GAL-DA administration
nder basal conditions there were significant differencesn DA level only among sampled brain areas, ranking in therder striatum�mesencephalon�prefrontal cortex, with no
ulative frequency histograms of the different classes of duration ofncy distribution in the different classes of duration is reported for theRearing episodes longer than 5 s are included in the last class. For
tion. Cumhe freque
ifferences between NHE and random-bred rats. Moreover

Gl
Nle2atipG1t
Ca
Sbfwmmcw2c(
Frfi g period.s
Fps1fcad
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244240
AL-DA (100 mg/kg) yielded no significant variation in DAevel across regions and lines except for the striatum of
ig. 4. Distributions of scanning times following GAL-DA administraearing episodes are reported for NRB given GAL-DA or galactose. Trst part (upper panel) and the second part (lower panel) of the testintatistics, see Results section.
ig. 5. Histochemical evidence for GAL-DA crossing the BBB. Upperanel: example of lectin-histochemical detection of galactose bindingites in the striatum at different times after injection of GAL-DA00 mg/kg. Lower panel: ROD of striatal slices histochemically stainedor galactose residues. Note that density of galactose residues in-reases at 0.5–3 h. Data represent mean�S.E.M. of combined NHE
tnd NRB rats (n�10 per group; * P�0.05 two-tailed t-test for unpairedata).
HE rats. In fact a three-way factorial ANOVA treatment�ine�brain area (as covariate) showed no significant mainffect for treatment and line but for brain area (F�143.0, df/72, P�0.000) with only a significant line�treatment inter-ction effect (F�6.49, df 1/36, P�0.015). As a matter of facthe DA content after GAL-DA was 138�21% of basal leveln the mes, 75�17% in the striatum and 46�25% in therefrontal cortex of NRB rats. Conversely DA content afterAL-DA was 109�24% in the mes, 173�18% (t�2.80, df8; 2P�0.012) in the striatum and 73�33% in the prefron-al cortex of NHE rats.
orrelation of DA level in mesencephalonnd striatum after GAL-DA treatment
ubtle covariations in within-group variability were extractedy Spearman rank order correlation analysis (Fig. 7). There-ore, under basal conditions DA level in the mesencephalonas significantly correlated to that in the striatum in positiveanner in NHE (RS�0.916, 2P�0.000) and in a negativeanner in NRB (RS��0.629, 2P�0.030). In addition, similar
ovariation between mesencephalon and prefrontal cortexas evident in NHE under basal conditions (RS��0.786,P�0.021). Interestingly only in the NHE DA in the prefrontalortex was positively correlated with that in the striatumRS��0.786, 2P�0.021). Conversely GAL-DA administra-
ulative frequency histograms of the different classes of duration ofncy distribution in the different classes of duration is reported for theRearing episodes longer than 5 s are included in the last class. For
tion. Cumhe freque
ion disrupted any such correlation in NHE and in NRB.

Tmrsa
B
Na2ltr
tSi2
oit
eaios
hoi
aNdd(bt
E
TavDccdabhtt
rs
T
TahisolB
Ea
TaostcDNqasamCe
Fartr
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244 241
DISCUSSION
he aim of this study was to provide evidence for a phar-acological effect of a GAL-DA on behavior in rats. The
esults indicate that GAL-DA is able to cross the BBB, ashown by histochemistry and capillary depletion analyses,nd to interact with the DA system.
ehavioral effects of GAL-DA
HE rats show a hyperactive profile which is associated tohypertrophic mesocortical DA system (Viggiano et al.,
002a,b, 2003, 2004). GAL-DA (100 mg/kg) reduces theocomotor activity in NHE rats, while increasing the dura-ion of rearing episodes in the second part of testing pe-iod.
This is in agreement with the effects of a D1 agonist inhese animals. In fact, similarly to GAL-DA, low doses ofKF81297, a D1R agonist, reduce orienting frequency and
ncrease scanning durations in NHE rats (Viggiano et al.,003).
Moreover, previous findings showed the central effectsf GAL-DA on classical DAergic models, such as morphine
nduced locomotion and reserpine-induced hypolocomo-ion (Bonina et al., 2003).
The behavioral effects are unlikely to be due to periph-ral actions of GAL-DA. In fact, very little or minimal effectss heart rate or blood pressure have been reported after
.v. injection of this compound (Bonina et al., 2003). On thether hand, the lack of peripheral effects of GAL-DA is
ig. 6. HPLC/mass spectrometry detection of GAL-DA (upper panel)nd its derivative, DA-succinate (lower panel), into plasma and brainegions. The latter have been prepared by the capillary depletionechnique. Data represent mean�S.E.M. of combined NHE and NRBats; n�10.
upported by the plasma level of GAL-DA (see Fig. 6). r
Furthermore, galactose molecule elicits minimal be-avioral effect at the highest dose tested. This effect ispposite to that of GAL-DA, as the latter decreases activ-
ty, whereas the former induces a slight increase.Finally, the behavioral effects of GAL-DA cannot be
ccounted for by different pharmacokinetics (PK) betweenHE and normal NRB, as assessed by histochemicallyetected galactose residues (Fig. 5) as well as by LC-MSetected concentrations of GAL-DA and DA-succinateFig. 6). As matter of fact, this issue is plausible as it haseen raised in a human study with Parkinson’s patientsreated with L-DOPA (Contin et al., 2003).
vidence for GAL-DA crossing BBB
he evidence that GAL-DA crosses the BBB is threefold:) detection of increased brain galactose residues re-ealed by lectin histochemistry, b) direct measurement ofA-succinate and DA-galactose in brain extracts, by theapillary depletion method, and c) altered mesolimbic DAovariation by HPLC-EC. In fact, lectin histochemistryemonstrates an increased density of galactose residuesfter GAL-DA treatment, that is mainly evident aroundlood vessels and in the neuropile (Fig. 5). On the otherand, capillary depletion method removes blood vessels,hus allowing detection of specific biochemical species inhe neuropile.
The conjugated compounds are stable in the blood,eleasing small amounts of DA very slowly, as previouslyuggested (Bonina et al., 2003).
ime course of GAL-DA effects
he histochemical findings indicate a peak brain level 3 hfter the injection. Indeed, modifications of locomotor be-avior can be revealed only 90–120 min after drug admin-
stration (Bonina et al., 2003). Previous studies on drug-tability, indicated that an active plasma level is achievednly after 90–120 min (Bonina et al., 2003). This long
atency in the effects is probably due to the crossing of theBB and to enzymatic cleavage at target sites.
ffects of GAL-DA on the DA systemnd perspectives
here were clear behavioral effects of GAL-DA (seebove). Interestingly, GAL-DA produced differential effectsn DA level in the target sites of the mesocorticolimbicystem in NHE and NRB rats. In fact DA was increased inhe neostriatum and prefrontal cortex of NHE, but de-reased in NRB rats. In the absence of strain differences inA levels under basal conditions, the increased DA level inHE rats induced by GAL-DA may be related to inade-uate DAT functioning induced by GAL-DA itself. Yet, innother model of ADHD, i.e. the spontaneously hyperten-ive rats (SHR (Sagvolden, 2000; Sagvolden et al., 2005)),DAT dysfunction has been demonstrated in synaptoso-al preparations from the dorsal stratum (Leo et al., 2003).onversely, the decrease in NRB rats may be tentativelyxplained as due to blockade of mesocorticolimbic neu-
ons by D2 autoreceptors, and to normal DAT functioning.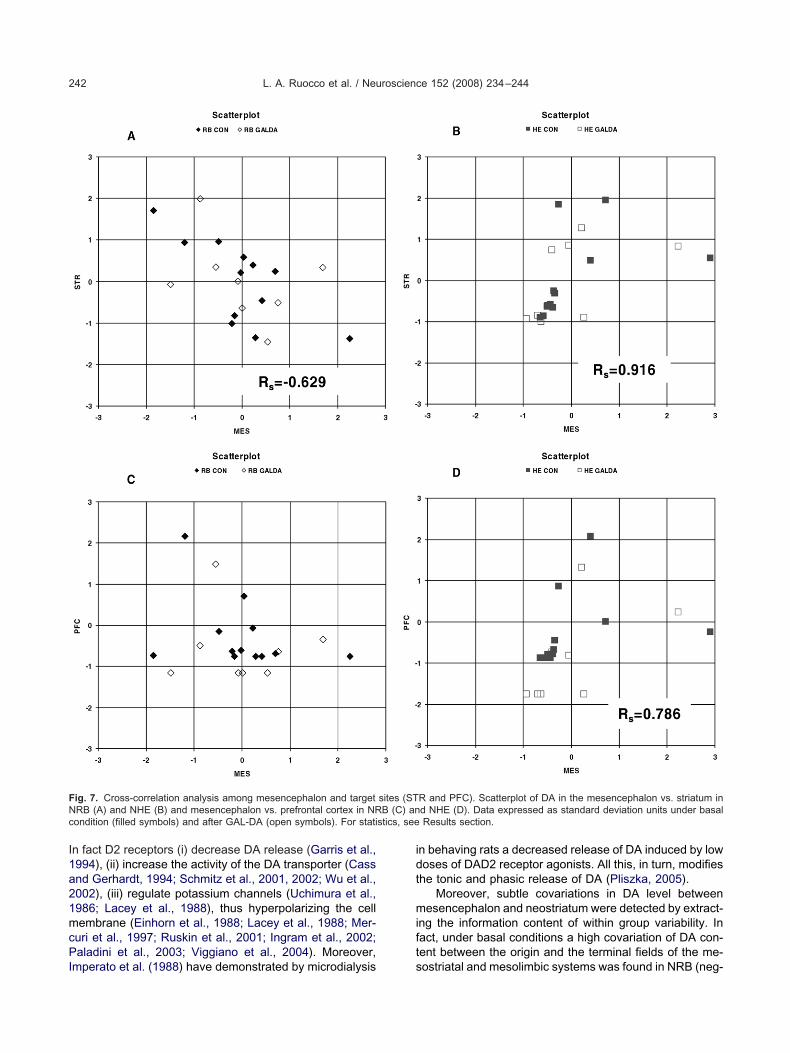
I1a21mcPI
idt
mift
FNc stics, see
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244242
n fact D2 receptors (i) decrease DA release (Garris et al.,994), (ii) increase the activity of the DA transporter (Cassnd Gerhardt, 1994; Schmitz et al., 2001, 2002; Wu et al.,002), (iii) regulate potassium channels (Uchimura et al.,986; Lacey et al., 1988), thus hyperpolarizing the cellembrane (Einhorn et al., 1988; Lacey et al., 1988; Mer-
uri et al., 1997; Ruskin et al., 2001; Ingram et al., 2002;aladini et al., 2003; Viggiano et al., 2004). Moreover,
ig. 7. Cross-correlation analysis among mesencephalon and targetRB (A) and NHE (B) and mesencephalon vs. prefrontal cortex in NRondition (filled symbols) and after GAL-DA (open symbols). For stati
mperato et al. (1988) have demonstrated by microdialysis s
n behaving rats a decreased release of DA induced by lowoses of DAD2 receptor agonists. All this, in turn, modifieshe tonic and phasic release of DA (Pliszka, 2005).
Moreover, subtle covariations in DA level betweenesencephalon and neostriatum were detected by extract-
ng the information content of within group variability. Inact, under basal conditions a high covariation of DA con-ent between the origin and the terminal fields of the me-
R and PFC). Scatterplot of DA in the mesencephalon vs. striatum ind NHE (D). Data expressed as standard deviation units under basalResults section.
sites (STB (C) an
ostriatal and mesolimbic systems was found in NRB (neg-

av
fisc
tfr
D(uscsA
AMCNaE
A
A
B
B
B
C
C
C
C
C
D
D
E
E
F
F
F
G
G
G
H
H
I
I
L
L
L
M
M
M
M
O
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244 243
tive covariation) and NHE rats (positive covariation). Con-ersely, GAL-DA administration disrupted this association.
Evidence for this hypothesized mechanism comesrom behavioral data showing that GAL-DA administrationncreases activity in C57 mice (Ruocco et al., 2007), whichhow low expression of D2 autoreceptor in the mesen-ephalon (Cabib et al., 2002).
It remains to be ascertained whether and to what ex-ent DA conveyed by GAL-DA may be eventually trans-ormed in other molecules such as norepinephrine aftereuptake by NET.
Other strategies have been previously followed to carryA into the brain, e.g. by peripheral or intranasal L-DOPA
Souza Silva et al., 1997; Silva et al., 1997). Howevernlike L-DOPA, GAL-DA after crossing the BBB, releaseslowly DA. Therefore GAL-DA might be a useful pharma-ological tool for the treatment of neurological diseases,uch as Parkinson’s, or neuropsychiatric disorders such asDHD based on modulation of DA neurotransmission.
cknowledgments—This research has been supported by aIUR-COFIN 2006–2007 grant to A.G.S. We thank R. Conte and. Treno for technical assistance in behavioral experiments, A.astro, A. Nivuoro and G. Spaziano for their skillful technicalssistance on HPLC. The critical revision of the manuscript by Dr.. Vitullo and Prof. M. T. Minelli is gratefully appreciated.
REFERENCES
spide R, Fresiello A, De Filippis G, Gironi Carnevale UA, Sadile AG(2000) Nonselective attention in a rat model of hyperactivity andattention deficit: subchronic methylphenidate and nitric oxide syn-thesis inhibitor treatment. Neurosci Biobehav Rev 24:59–71.
spide R, Gironi Carnevale UA, Sergeant JA, Sadile AG (1998) Non-selective attention and nitric oxide in putative animal models of atten-tion-deficit hyperactivity disorder. Behav Brain Res 95:123–133.
entivoglio M, Morelli M (2005) The organization and circuits of mes-encephalic dopaminergic neurons and the distribution of dopaminereceptors in the brain. In: Handbook of chemical neuroanatomy(Dunnett SB, Bentivoglio M, Bjorklund A, Hokfelt T, eds), pp 1–107.Amsterdam: Elsevier.
onina F, Puglia C, Rimoli MG, Melisi D, Boatto G, Nieddu M, Calig-nano A, La Rana G, De Caprariis P (2003) Glycosyl derivatives ofdopamine and L-dopa as anti-Parkinson prodrugs: synthesis, phar-macological activity and in vitro stability studies. J Drug Target11:25–36.
onina FP, Arenare L, Palagiano F, Saija A, Nava F, Trombetta D, DeCaprariis P (1999) Synthesis, stability, and pharmacological eval-uation of nipecotic acid prodrugs. J Pharm Sci 88:561–567.
abib S, Puglisi-Allegra S, Ventura R (2002) The contribution ofcomparative studies in inbred strains of mice to the understandingof the hyperactive phenotype. Behav Brain Res 130:103–109.
apowski JJ (1989) Computer techniques in neuroanatomy, pp1–483. New York: Plenum Press.
ass WA, Gerhardt GA (1994) Direct in vivo evidence that D2 dopa-mine receptors can modulate dopamine uptake. Neurosci Lett176:259–263.
erbone A, Pellicano MP, Sadile AG (1993) Evidence for and againstthe Naples high- and low-excitability rats as genetic model to studyhippocampal functions. Neurosci Biobehav Rev 17:295–303.
ontin M, Martinelli P, Riva R, Dondi M, Fanti S, Pettinato C, ScaglioneC, Albani F, Baruzzi A (2003) Assessing dopaminergic function inParkinson’s disease: levodopa kinetic-dynamic modeling and
SPECT. J Neurol 250:1475–1481.alpiaz A, Filosa R, De Caprariis P, Conte G, Bortolotti F, Biondi C,Scatturin A, Prasad PD, Pavan B (2007) Molecular mechanisminvolved in the transport of a prodrug dopamine glycosyl conjugate.Int J Pharm 336:133–139.
jaldetti R, Melamed E (2001) New therapies for Parkinson’s disease.J Neurol 248:357–362.
dwards AL (1967) Statistical methods for the behavioral sciences.New York: Holt, Rinehart and Winston.
inhorn LC, Johansen PA, White FJ (1988) Electrophysiological ef-fects of cocaine in the mesoaccumbens dopamine system: studiesin the ventral tegmental area. J Neurosci 8:100–112.
araone SV, Sergeant J, Gillberg C, Biederman J (2003) The world-wide prevalence of ADHD: is it an American condition? WorldPsychiatry 2:104–113.
ernandez C, Nieto O, Rivas E, Montenegro G, Fontenla JA, Fernan-dez-Mayoralas A (2000) Synthesis and biological studies of glyco-syl dopamine derivatives as potential antiparkinsonian agents.Carbohydr Res 327:353–365.
ernandez-Espejo E (2004) Pathogenesis of Parkinson’s disease:prospects of neuroprotective and restorative therapies. Mol Neu-robiol 29:15–30.
arris PA, Ciolkowski EL, Pastore P, Wightman RM (1994) Efflux ofdopamine from the synaptic cleft in the nucleus accumbens of therat brain. J Neurosci 14:6084–6093.
ironi Carnevale UA, Vitullo E, Varriale B, Ruocco LA, Sadile AG(2007) A classical Mendelian cross-breeding study of the Napleshigh and low excitability rat lines. Behav Brain Res 183:130–140.
race AA, Bunnney BS, Moore H, Todd CL (1997) Dopamine-celldepolarization block as a model for the therapeutic actions ofantipsychotic drugs. Trends Neurosci 20:31–37.
almos T, Santarromana M, Antonakis K, Scherman D (1996) Syn-thesis of glucose-chlorambucil derivatives and their recognition bythe human GLUT1 glucose transporter. Eur J Pharmacol 318:477–484.
ölm S (1979) A simple sequentially rejective multiple test procedure.Scand J Stat 6:65–70.
mperato A, Tanda G, Frau R, Di Chiara G (1988) Pharmacologicalprofile of dopamine receptor agonists as studied by brain dialysis inbehaving rats. J Pharmacol Exp Ther 245:257–264.
ngram SL, Prasad BM, Amara SG (2002) Dopamine transporter-mediated conductances increase excitability of midbrain dopamineneurons. Nat Neurosci 5:971–978.
acey MG, Mercuri NB, North RA (1988) On the potassium conduc-tance increase activated by GABAB and dopamine D2 receptors inrat substantia nigra neurones. J Physiol 401:437–453.
eo D, Sorrentino E, Volpicelli F, Eyman M, Greco D, Viggiano D, diPorzio U, Perrone-Capano C (2003) Altered midbrain dopaminer-gic neurotransmission during development in an animal model ofADHD. Neurosci Biobehav Rev 27:661–669.
indvall O, Bjorklund A (1974) The organization of ascending cate-cholamine neuron systems in the rat brain as revealed by glyoxylicacid fluorescence method. Histochemistry 39:97–127.
ercuri NB, Bernardi G (2005) The ‘magic’ of L-dopa: why is it the goldstandard Parkinson’s disease therapy? Trends Pharmacol Sci26:341–344.
ercuri NB, Scarponi M, Bonci A, Siniscalchi A, Bernardi G (1997)Monoamine oxidase inhibition causes a long-term prolongation ofthe dopamine-induced responses in rat midbrain dopaminergiccells. J Neurosci 17:2267–2272.
issale C, Nash SR, Robinson SW, Jaber M, Caron MG (1998)Dopamine receptors: from structure to function. Physiol Rev78:189–225.
ueckler M (1994) Facilitative glucose transporters. Eur J Biochem219:713–725.
lanow CW, Agid Y, Mizuno Y, Albanese A, Bonuccelli U, Damier P,De Yebenes J, Gershanik O, Guttman M, Grandas F, Hallett M,Hornykiewicz O, Jenner P, Katzenschlager R, Langston WJ, Le-
Witt P, Melamed E, Mena MA, Michel PP, Mytilineou C, Obeso JA,
O
P
P
P
P
R
R
S
S
S
S
S
S
S
S
S
S
T
U
U
V
V
V
V
W
L. A. Ruocco et al. / Neuroscience 152 (2008) 234–244244
Poewe W, Quinn N, Raisman-Vozari R, Rajput AH, Rascol O,Sampaio C, Stocchi F (2004) Levodopa in the treatment ofParkinson’s disease: current controversies. Mov Disord 19:997–1005.
ldendorf WH (1971) Brain uptake of radiolabeled amino acids,amines, and hexoses after arterial injection. Am J Physiol221:1629–1639.
aladini CA, Robinson S, Morikawa H, Williams JT, Palmiter RD(2003) Dopamine controls the firing pattern of dopamine neuronsvia a network feedback mechanism. Proc Natl Acad Sci U S A100:2866–2871.
isani A, Centonze D, Bernardi G, Calabresi P (2005) Striatal synapticplasticity: implications for motor learning and Parkinson’s disease.Mov Disord 20:395–402.
liszka SR (2005) The neuropsychopharmacology of attention-deficit/hyperactivity disorder. Biol Psychiatry 57:1385–1390.
olt R, Porreca F, Szabo LZ, Bilsky EJ, Davis P, Abbruscato TJ, DavisTP, Harvath R, Yamamura HI, Hruby VJ (1994) Glycopeptideenkephalin analogues produce analgesia in mice: evidence forpenetration of the blood-brain barrier. Proc Natl Acad Sci U S A91:7114–7118.
uocco LA, Viggiano D, Pignatelli M, Iannaccone T, Rimoli MG, MelisiD, Curcio A, De Lucia S, Carboni E, Gironi Carnevale UA, deCaprariis P, Sadile AG (2007) Galactosylated dopamine increasesattention without reducing activity in c57bl/6 mice. Behav BrainRes, in press.
uskin DN, Bergstrom DA, Shenker A, Freeman LE, Baek D, WaltersJR (2001) Drugs used in the treatment of attention-deficit/hyper-activity disorder affect postsynaptic firing rate and oscillation with-out preferential dopamine autoreceptor action. Biol Psychiatry49:340–350.
adile AG, Gironi Carnevale UA, Vitullo E, Cioffi LA, Welzl H, Bättig K(1988) Maze learning of the Naples high- and low-excitability ratlines. Adv Biosci 70:177–180.
agvolden T (2000) Behavioral validation of the spontaneously hyper-tensive rat (SHR) as an animal model of attention-deficit hyperac-tivity disorder (AD/HD). Neurosci Biobehav Rev 24:31–40.
agvolden T, Johansen EB, Aase H, Russell VA (2005) A dynamicdevelopmental theory of attention-deficit/hyperactivity disorder(ADHD) predominantly hyperactive/impulsive and combined sub-types. Behav Brain Sci 28:397–419.
chmitz Y, Lee CJ, Schmauss C, Gonon F, Sulzer D (2001) Amphet-amine distorts stimulation-dependent dopamine overflow: effectson D2 autoreceptors, transporters, and synaptic vesicle stores.J Neurosci 21:5916–5924.
chmitz Y, Schmauss C, Sulzer D (2002) Altered dopamine releaseand uptake kinetics in mice lacking D2 receptors. J Neurosci
22:8002–8009.ergeant JA, Geurts H, Huijbregts S, Scheres A, Oosterlaan J (2003)The top and the bottom of ADHD: a neuropsychological perspec-tive. Neurosci Biobehav Rev 27:583–592.
himura T, Tabata S, Terasaki T, Deguchi Y, Tsuji A (1992) In-vivoblood-brain barrier transport of a novel adrenocorticotropic hor-mone analogue, ebiratide, demonstrated by brain microdialysisand capillary depletion methods. J Pharm Pharmacol 44:583–588.
ilva MA, Mattern C, Hacker R, Tomaz C, Huston JP, Schwarting RK(1997) Increased neostriatal dopamine activity after intraperitonealor intranasal administration of L-DOPA: on the role of benserazidepretreatment. Synapse 27:294–302.
olanto MV (1998) Neuropsychopharmacological mechanisms ofstimulant drug action in attention-deficit hyperactivity disorder: areview and integration. Behav Brain Res 94:127–152.
ouza Silva MA, Mattern C, Hacker R, Nogueira PJ, Huston JP,Schwarting RK (1997) Intranasal administration of the dopaminer-gic agonists L-DOPA, amphetamine, and cocaine increases dopa-mine activity in the neostriatum: a microdialysis study in the rat.J Neurochem 68:233–239.
riguero D, Buciak J, Pardridge WM (1990) Capillary depletion methodfor quantification of blood-brain barrier transport of circulating pep-tides and plasma proteins. J Neurochem 54:1882–1888.
chimura N, Higashi H, Nishi S (1986) Hyperpolarizing and depolar-izing actions of dopamine via D-1 and D-2 receptors on nucleusaccumbens neurons. Brain Res 375:368–372.
riel C, Egron MJ, Santarromana M, Scherman D, Antonakis K, Her-scovici J (1996) Hexose keto-C-glycoside conjugates: design, syn-thesis, cytotoxicity, and evaluation of their affinity for the glucosetransporter Glut-1. Bioorg Med Chem 4:2081–2090.
iggiano D, Grammatikopoulos G, Sadile AG (2002a) A morphometricevidence for a hyperfunctioning mesolimbic system in an animalmodel of ADHD. Behav Brain Res 130:181–189.
iggiano D, Vallone D, Welzl H, Sadile AG (2002b) The Napleshigh-and low excitability rats: selective breeding, behavioral profile,morphometry and molecular biology of the mesocortical dopaminesystem. Behav Genet 32:315–333.
iggiano D, Vallone D, Ruocco LA, Sadile AG (2003) Behavioural,pharmacological, morpho-functional molecular studies reveal a hy-perfunctioning mesocortical dopamine system in an animal modelof attention deficit and hyperactivity disorder. Neurosci BiobehavRev 27:683–689.
iggiano D, Vallone D, Sadile AG (2004) Dysfunctions in the dopa-mine systems and ADHD: evidences from animals and modeling.Neural Plast 11:93–116.
u Q, Reith ME, Walker QD, Kuhn CM, Carroll FI, Garris PA (2002)Concurrent autoreceptor-mediated control of dopamine releaseand uptake during neurotransmission: an in vivo voltammetric
study. J Neurosci 22:6272–6281.(Accepted 8 November 2007)(Available online 28 November 2007)




















