
Self-gelling hydrogels based on oppositely charged dextran microspheres
Upload
independentCategory
view
1download
0
The FASEB Journal • Research Communication
GABAA and GABAB receptors of distinct propertiesaffect oppositely the proliferation of mouseembryonic stem cells through synergistic elevation ofintracellular Ca2�
Marija Schwirtlich,*,1 Zsuzsa Emri,†,1 Karoly Antal,† Zoltan Mate,* Zoya Katarova,*,2 andGabor Szabo*,2,3
*Laboratory of Molecular Biology and Genetics, Institute of Experimental Medicine, and†Department of Neurochemistry, Chemical Research Center, Hungarian Academy of Sciences,Budapest, Hungary
ABSTRACT Gamma-amminobutyric acid (GABA), themajor inhibitory neurotransmitter in the central ner-vous system of vertebrates, serves as an autocrine/paracrine signaling molecule during development,modulating a number of calcium (Ca2�)-dependentprocesses, including proliferation, migration, and dif-ferentiation, acting via 2 types of GABA receptors(GABARs): ionotropic GABAARs and metabotropicGABABRs. Here, we demonstrate that mouse embry-onic stem cells (mESCs), which possess the capacity forvirtually unlimited self-renewal and pluripotency, syn-thesize GABA and express functional GABAARs andGABABRs, as well as voltage-gated calcium channels(VGCCs), ryanodine receptors (RyRs), and inwardlyrectifying potassium (GIRK) channels. On activation,both GABAR types triggered synergistically intracellu-lar calcium rise. Muscimol (a GABAAR agonist) inducedsingle Ca2� transients involving both VGCC-mediatedCa2� influx and intracellular stores, while baclofen (aGABABR agonist) evoked Ca2� transients followed byintercellular Ca2� waves and oscillations that wereresistant to antagonists and entirely dependent on Ca2�
release from intracellular stores. Prolonged treatmentwith muscimol slightly inhibited, while baclofen orSR95531 (a GABAAR antagonist) significantly facili-tated, mESC proliferation. GABAAR-specific ligandsalso induced morphological and gene expressionchanges indicating a differentiation shift. Our datasuggest that the interplay between GABARs and down-stream (coupled) effectors differentially modulatesmESC proliferation/differentiation through selectiveactivation of second messenger signaling cascades.—Schwirtlich, M., Emri, Z., Antal, K., Mate, Z., Katarova,Z., Szabo, G. GABAA and GABAB receptors of distinctproperties affect oppositely the proliferation of mouseembryonic stem cells through synergistic elevation ofintracellular Ca2�. FASEB J. 24, 000–000 (2010). www-.fasebj.org
Key Words: Ca2� imaging � ES cell self-renewal/differentia-tion � connexin 43
Gamma-aminobutyric acid (GABA) serves a dualfunction as the main inhibitory neurotransmitter in thenervous system of vertebrates and as a signaling mole-cule during development and in multiple cell types ofthe body (1–4). In the adult brain, GABA released atinhibitory synapses acts on ionotropic and metabo-tropic GABA receptors (GABAARs and GABABRs, re-spectively) to inhibit the spread of membrane depolar-ization. The ionotropic GABAARs are Cl�-selectiveheteropentameric channels that assemble from distinctsubunit subclasses (�1–6, �1–3, �1–4, and the � substi-tutes �, e, �, and �) and display a remarkable structuralheterogeneity associated with diverse functions (5–7).In the adult nervous system, GABAARs spatially segre-gate into synaptic and extrasynaptic classes mediatingthe so-called phasic and tonic inhibition, respectively(5, 7, 8). During development, because of the reversedchloride gradient, GABAAR activation evokes mem-brane depolarization and rise in intracellular calciumconcentration ([Ca2�]i) levels due to voltage-gatedcalcium channel (VGCC)-mediated Ca2� influx andthereby modulates a variety of [Ca2�]i-dependent cel-lular processes, including proliferation, migration, anddifferentiation (1–3).
The G-protein coupled metabotropic GABABRs func-tion as obligatory heterodimers comprised of 2 distinctsubunits, GABABR1 and GABABR2, respectively (9, 10).In the adult nervous system, GABABRs are functionallysegregated: the presynaptic GABABRs inhibit VGCC-mediated [Ca2�]i elevations and thereby neurotrans-mitter release, whereas postsynaptic GABABRs coupledto G-protein-activated inwardly rectifying K� (GIRK)channels mediate slow postsynaptic inhibition (11, 12).Activation of GABABRs in both mature and developingneurons causes membrane hyperpolarization and de-
1 These authors contributed equally to this work.2 These authors contributed equally to this work.3 Correspondence: Institute of Experimental Medicine, P.O.
Box 67, 1450-Budapest, Hungary. E-mail: [email protected]: 10.1096/fj.09-143586
10892-6638/10/0024-0001 © FASEB
The FASEB Journal article fj.09-143586. Published online December 11, 2009.

crease in [Ca2�]i (2, 3, 9). Consequently, activation ofGABABRs in immature neurons leads to inhibition ofthe GABAAR-evoked [Ca2�]i rise (13, 14). However,GABABRs can exert an opposite effect. For instance,GABABR activation enhances the [Ca2�]i elevation evokedby the metabotropic glutamate receptor mGluR1 (15,16) and triggers Ca2� transients due to Ca2� releasefrom inositol triphosphate (IP3)-sensitive intracellularCa2� stores in chromaffin cells and astrocytes (17–19).
GABA has long been known as a modulator ofneuronal progenitor cell proliferation (reviewed inrefs. 1–3, 20, 21). Pioneer experiments showed thatGABA triggers depolarization and a rise in [Ca2�]i andinhibits cell proliferation of neocortical progenitorcells (22). Similarly, GABA inhibits the proliferation ofadult neural stem cells (23) and cancer cells (24), whileit stimulates the proliferation of ventricular zone cortical(25) or cerebellar (26) progenitors, Schwann cells (27),cultured oligodendrocytes (28), embryonic neural pro-genitor cells (29), and growth plate chondroctes (30).
Here we have studied GABAR -mediated Ca2� signalingand its effect on the proliferation of mouse embryonicstem cells (mESCs). The mESCs are in vitro derivatives ofthe inner cell mass (ICM) of mouse blastocysts (31) andare characterized by a nearly unlimited capacity for self-renewal and pluripotency, the ability to differentiate intovirtually all cell types of the body (32, 33). We show for thefirst time that mESCs express GABAARs and GABABRs ofunique subunit composition and properties that on acti-vation synergistically trigger [Ca2�]i rise but oppositelyaffect mESC proliferation, acting through essentially dif-ferent mechanisms.
MATERIALS AND METHODS
Cell culture
mESCs (line R1) were grown on mitomycin C (100 mg/ml;Sigma, St. Louis, MO, USA)-treated primary embryonic fibro-blasts in the presence of 20% heat-inactivated ESC-qualifiedFCS (Invitrogen, Carlsbad, CA, USA) and 1000 U/ml ofleukemia inhibitory factor (LIF; ESGRO; Chemicon Interna-tional, Temecula, CA, USA; ref. 34).
RT-PCR
Total RNA was isolated from mitogen-treated primary embry-onic fibroblasts, mESCs, and whole brain using TRI reagent(Sigma). First-strand cDNA was synthesized from 5 g of RNAusing SuperScript III reverse transcriptase (Invitrogen). cDNAsynthesized from 1 g RNA was amplified using gene-specificprimers. �-Actin was coamplified for 16–20 cycles.
Primer sequences and cycle parameters specific for eachprimer pair are listed in Supplemental Table 1.
Immunocytochemistry and immunoblotting
One-day-old mESC cultures were fixed for 20 min with 4%paraformaldehyde in PBS, washed in PBS and Tris-bufferedsaline (TBS), and thereafter permealized in 0.2% TritonX-100-TBS and blocked for 1 h at room temperature in 0.5%blocking reagent (Roche, Mannheim, Germany)-TBS. Incu-
bation with primary antibodies was overnight at 4°C in 1%BSA-TBST (TBS�0.05% Tween 20) as follows: goat anti-Oct-3/4 (1:1000) and goat anti-GABAA�3 (1:250; Santa CruzBiotechnology, Santa Cruz, CA, USA); rabbit anti-GABAA�3(1:1000; Novus Biologicals Inc., Littleton, CO, USA); rabbitanti-GABABR2 and mouse anti-Cx43 (both 1:500; BD Bio-sciences, San Jose, CA, USA); and rabbit anti-GABA (1:5000;Sigma). Following TBST washes, Alexa Fluor 488 rabbitanti-goat IgG, TX Red goat anti-rabbit IgG (both fromInvitrogen) or biotinylated anti-rabbit IgG (Vector, Burlingame,CA, USA) were applied for 1 h at room temperature. Tertiaryantibody was ExtrAvidin-HRP (1:1000; Sigma). Cells were ana-lyzed under Zeiss Axioscop-2 (Carl Zeiss, Jena, Germany)equipped with Axiocam-HR and Axiovision software.
Western blots were performed essentially as describedpreviously (35) and reacted first with rabbit Oct-3/4 (1:1000;Cell Signaling Technology, Danvers, MA, USA) or Cx43(1:1000) antibodies followed by alkaline phosphatase (AP)-conjugated anti-rabbit (mouse) antibody and developed byImmobilon Western AP substrate (Millipore, Bedford, MA,USA). Quantitative analysis was performed in Adobe Photo-shop CS (Adobe Systems, San Jose, CA, USA).
[Ca2�]i imaging
mESCs plated on glass coverslips were loaded with thecalcium-sensitive fluorescent dye Fluo-4/AM (2.5 mM; In-vitrogen) in perfusion buffer (composition, in mM: 154 NaCl,5.6 KCl, 2.3 CaCl2, 5.6 glucose, and 10 HEPES, pH 7.4)containing 0.02% pluronic acid (Sigma) for 45 min at 37°C.Coverslips were perfused at room temperature at 1 ml/minunder aeration with 95% O2/5% CO2 initially for 2 min withbuffer to obtain a steady baseline. GABA, muscimol, bicucul-line methiodide, baclofen, SR95531, and cadmium chloride(all from Sigma) and CGP55845, CGP54626, and cyclopia-zonic acid (CPA; Tocris Bioscience, Bristol, UK) were appliedat concentrations as indicated in the figure legends in arandomized order. CPA and Cd2� were applied continuouslyfor 4 min before single-agonist application. The Ca2�-freeperfusion buffer contained 1 mM EDTA. Fluorescence wasmeasured under an Olympus BX61WI laser-scanning confo-cal microscope using FluoView 300 software (Olympus Opti-cal Co. GmbH, Hamburg, Germany). Serial scanning wasperformed at 488/530-nm excitation/emission wavelengthsat 3 s intervals. Fluorescence intensity changes (F%) werecalculated as percentage of baseline fluorescence—the aver-age intensity of the initial 30 frames (2).
Cell proliferation studies
mESCs maintained on fibroblast layers were plated at densityof 1000 cells/well on gelatin-coated 96-well plates (CorningInc., Corning, NY, USA). Ligands were added 12 h later asindicated in the figure legends, and cells were grown foranother 12 or 24 h. Medium was replaced every 12 h.Proliferation was assayed using CyQuant Cell ProliferationAssay Kit (Invitrogen) according to the manufacturer’s in-structions. Fluorescence was measured in a Victor3 V multila-bel bench-top microplate reader (PerkinElmer, Wellesley,MA, USA) at 480/550 nm.
Cells from multiple wells were stained with 0.5% cresylviolet for 15 min at room temperature. Low-magnificationimages of the entire well were obtained, and both colony sizeand total area occupied by mESCs were estimated usingImageJ (http://rsbweb.nih.gov). At least 3 wells were usedper treatment per assay. AP histochemistry was performed asdescribed previously (36).
2 Vol. 24 April 2010 SCHWIRTLICH ET AL.The FASEB Journal � www.fasebj.org

Statistical analysis
Statistical significance was estimated by unpaired 2-tailedStudent’s t test (GraphPad Software, San Diego, CA USA:www.graphpad.com). Values of P � 0.05 were consideredsignificant. Levels of significance were expressed as indicatedin the figure legends.
RESULTS
Undifferentiated mESCs express multiple GABAARand GABABR subunits, GIRK channels, VGCCs, andryanodine receptors (RyRs)
The expression of different GABAAR subunits wasstudied by semiquantitative RT-PCR using specificprimers (Fig. 1A and Supplemental Table 1; ref. 37).We detected GABAAR �1, �3, �4, �5, �3, �1, �2S, �3,and �, as well as ε1 and ε2, but no �2, �6, �1, �2, or theGABACR-specific 1 and 2 subunits in 1-d-old mESCs(Fig. 1A). The most abundant � subunit was �3; �2s(short) was expressed at low levels, and �2L (long)subunit was absent, while �1 was barely detectable (Fig.1A). We also found expression of the major GABABRsubunits (38): both GABABR1 variants R1a and R1bwere present, with a clear prevalence of the sushidomain-containing R1a, as well as GABABR2 (Fig. 1B).GABABR is coupled to and enhances the activity ofpostsynaptic GIRK1–3 channels, generating inhibitorypostsynaptic currents (11). We found expression ofGIRK1 and GIRK2, the most abundant brain subunitsthat account for 90% of the baclofen-induced GIRKcurrent (39; Fig. 1C), and very low levels of GIRK3 (datanot shown).
We also found abundant expression of the functionallyindispensable, pore-forming � subunits of VGCCs of Nand R types, both subunits of T type, but extremely lowlevels of the L-type � subunit in mESCs (Fig. 1D). Theseresults were highly reproducible and are consistent withthe VGCC expression profile of the mouse ICM (40).
The RyRs comprise a group of 3 (RyR1–3) ryanodine-
sensitive Ca2�-permeable channels and can mediateCa2�-induced Ca2� release (CICR) from ER stores,resulting in increase of cytosolic [Ca2�]i (41). OurRT-PCR results clearly show expression of RyR1 andRyR2, but not RyR3 (Fig. 1E), with similar abundancein mESCs and P7 mouse brain.
Cellular distribution of GABAARs, GABABRs, andGABA in undifferentiated mESCs
The cellular localization of the 2 GABAR types wasstudied in 1-d-old cultures of exponentially growingmESCs. Using a GABAAR-�3 subunit-specific antibody,we found that all mESCs, but not the fibroblast layer,were stained (Fig. 2Aa, a�). Small clones displayed amoderate cytoplasmic staining (Fig. 2Ba), which shiftedto and progressively enhanced at cellular borders oflarger clones (Fig. 2Aa; Bb, c). The cellular distributionof GABABRs was examined with GABABR2-specific an-tibodies (Fig. 2Ab). All mESCs, independent of theclone size, displayed a prominent “patchy” staining thatwas enhanced near cellular borders (Fig. 2Bb).
mESC cultures used in this study appeared morpho-logically undifferentiated, with colonies having welldefined borders, staining strongly and uniformly for AP(see below) and 2 specific markers of nondifferentiatedmESCs (32): the nuclear localized Oct-3/4 (Fig. 2Ac)and membrane-bound SSEA-1 (not shown).
mESCs (line R1) also synthesize and accumulateGABA, as demonstrated by the robust staining obtainedwith a highly specific antibody (Fig. 2Ad), in agreementwith the previously reported expression of glutamicacid decarboxylase (GAD) in mESCs (42, 43).
GABAARs evoke Ca2� transients in mESCs:involvement of extracellular Ca2� and Ca2� frominternal stores
The presence of fully assembled functional GABAARsin mESCs was tested by monitoring the [Ca2�]i changes
Figure 1. Representative gel images of RT-PCR products amplified from fibroblasts (F),1-d-old mESC cultures (ES�RT), and P7 mouse brain (MB). Multiple subunits of GABAARs(A) and GABABRs (B); GIRK1 and 2 (C); � subunits of L-, N-, R-, TG-, and TH-type VGCCs(D); and RyR1 and RyR2 subunits (E) are detected in mESCs but not in fibroblasts. Asteriskindicates coamplified �-actin band.
3GABAR-MEDIATED SIGNALING IN MOUSE ES CELLS

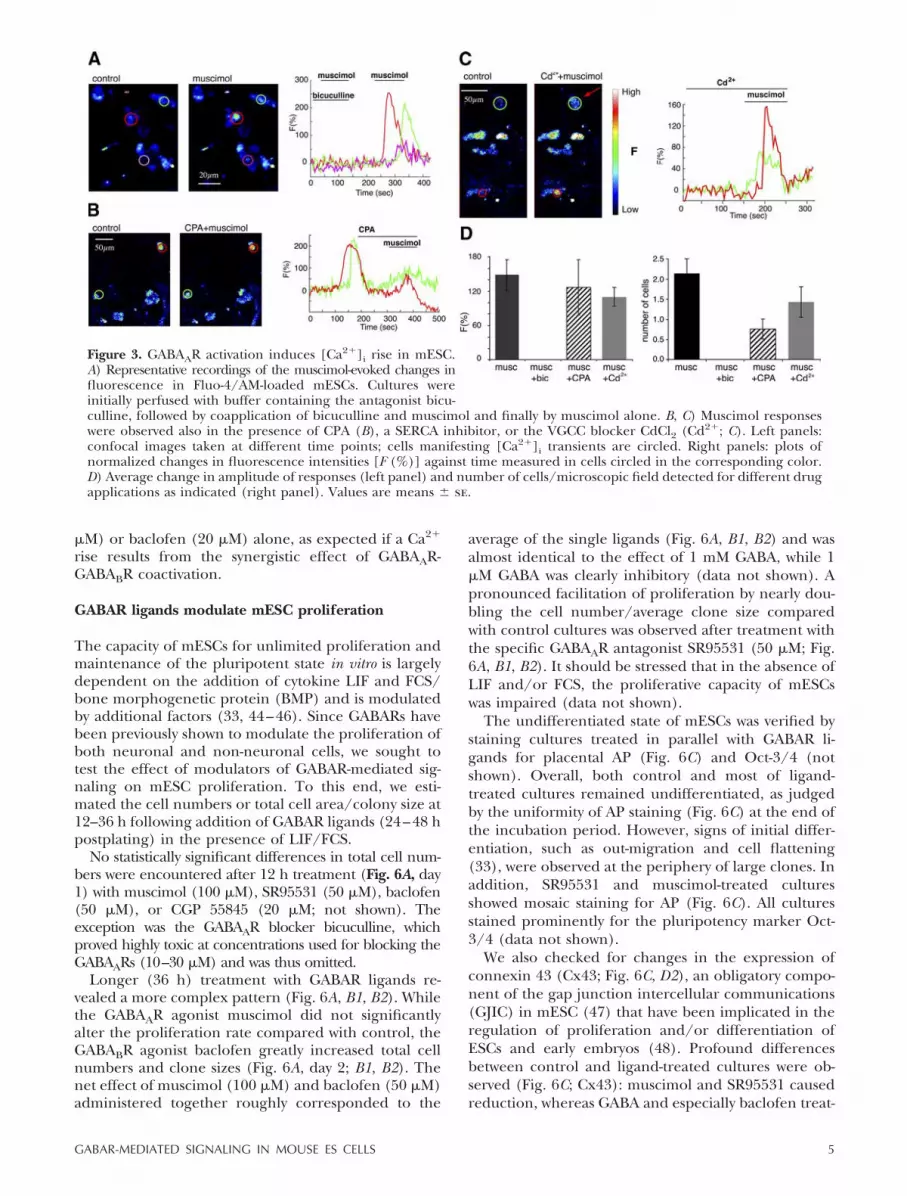
after GABAAR activation. Application of the GABAARagonist muscimol (30 M) induced a sharp increase offluorescence in mESCs (Fig. 3A and SupplementalMovie 1) that could be entirely blocked by the GABAARantagonist bicuculline (20 M; Fig. 3A, D).
The average number of cells per observation fieldresponding to muscimol (2.14�0.36; n�14 fields/31cells), the mean amplitude of their response (F�147.98�26.6%), as well as the full blockade by bicuculline (n�4fields/0 cells) were highly reproducible in similar recordingsfrom different cultures. To test whether [Ca2�]i increaseinduced by GABAAR activation was due to Ca2� release fromintracellular Ca2� stores or Ca2� influx through VGCCs,we applied specific inhibitors to block each pathway.The sarco-endoplasmic reticulum Ca2�-ATPase (SERCA)blocker CPA (10 M) induced quick Ca2� transients(Fig. 3B; n�7 fields/26 cells) followed by a delayedcapacitive Ca2� entry (120.3�4.2 s) triggered by deple-tion of the intracellular stores. Consequently, agonistswere always applied 60 s after CPA addition. In thepresence of CPA, the number of cells showing musci-mol-evoked Ca2� transients was considerably reduced(0.75�0.25; n�4 fields/6 cells; Fig. 3D), with relativelyunchanged but more variable amplitude (126.55�48.11%; Fig. 3B, D). Initial bath application of theVGCC blocker cadmium (Cd2�; 100 M) reduced themean amplitude of muscimol-evoked Ca2� transientsby 37% (F�93.14�13.68; n�6 fields/11 cells; Fig. 3C,D) and only slightly the number of responding cells
(1.83�0.31; n�6 fields/11 cells; Fig. 3C, D). A similareffect was observed when Ca2� was omitted from thebuffer (data not shown).
GABABR agonist baclofen evokes Ca2� transients inmESCs by mobilizing Ca2� from internal stores
Acting on either presynaptic VGCCs or postsynapticGIRK channels, neuronal GABABRs inhibit [Ca2�]ielevation (9, 10). Strikingly, in mESCs the GABABRagonist baclofen (20 M) evoked a sharp and signifi-cant increase of fluorescence (Ca2�; F�177.35�29.88;average responding cells 2.7�0.32; n�23 fields/64cells; Fig. 4A, E and Supplemental Movie 2). In contrastto muscimol, which triggered single transients, thebaclofen-induced fluorescence signal was followed inmost cases by wave propagation to neighboring cellsand delayed oscillations (Supplemental Movie 2 andFig. 4A). Surprisingly, only single transients, but notCa2� waves or oscillations, could be blocked by twopotent and widely used GABABR antagonists, CGP 55845(20 M) and CGP 54626 (10 M; Fig. 4A and Supple-mental Movie 2). Neither antagonist significantly affectedthe number of cells showing [Ca2�]i increase (3.0�0.71;n�5 fields/15 cells) or the mean amplitude of the ba-clofen-evoked transients (210.84�47.73%; n�5 fields/15cells; Fig. 4A, E).
Occasionally we observed a decrease of [Ca2�]i fol-lowing baclofen application, which was reversed byCGP (Fig. 4A), a classical GABABR response. To dis-criminate between the [Ca2�]i rising vs. suppressingeffects of GABABRs in mESCs, we applied KCl (50 mM)before baclofen. Initially, KCl induced a moderate[Ca2�]i rise that was further increased by baclofen (Fig.4B), supporting the notion for a predominantly [Ca2�]i-rising effect of GABABRs in mESCs.
Blockade of the VGCCs with Cd2� (100 M) beforebaclofen reduced the amplitude (91.03�14.92%; n�5fields/16 cells) of the Ca2�transients (presumably dueto a quenching effect) but not the number of respond-ing cells (3.2�0.2; n�5 fields/16 cells; Fig. 4C, E).Furthermore, baclofen-evoked Ca2� transients wereessentially unaffected when Ca2�-free buffer was used(Fig. 4D, E; 2.33�0.67 responding cells; mean ampli-tude F�206.12�53.94%; n�6 fields/14 cells) andnearly completely blocked by 10 M CPA (0.33�0.33;n�3 fields/1 cell; Fig. 4E), indicating an essentiallyinternal Ca2� store-dependent mechanism.
GABA induces fast Ca2� transients inundifferentiated mESCs
Perfusion of mESCs with 1 mM GABA for 90 s resultedin quick induction of Ca2� transients of variable ampli-tudes in single cells or doublets (Fig. 5A1), generallyfollowed by long-lasting oscillations (Fig. 5A2). Theaverage number of responding cells (2.85�0.4; n�13fields/38 cells) was similar (Fig. 5B1), but the meanamplitude was higher (198.99�47.8%; n�13 fields/38cells; Fig. 5B2) compared with either muscimol (30
Figure 2. Cellular distribution of GABAARs, GABABRs, GABA,and Oct-3/4 in mESCs. A) Indirect immunofluorescence of1-d-old cultures using anti-GABAAR-�3 (a), anti-GABABR2(b), anti-Oct-3/4 (c), and anti-GABA (d) antibodies. Insets:enlarged mESC clones from a (a) and b (b), respectively.Arrows (a) and arrowheads (b) mark intensive signal atcellular borders. B) Immunostaining with GABAAR-�3 anti-body reveals cytoplasmic staining in single cells (a), whichgradually shifts to membrane bound in a 2-cell clone (b) andmulticellular clones (c). Scale bars � 20 m.
4 Vol. 24 April 2010 SCHWIRTLICH ET AL.The FASEB Journal � www.fasebj.org
M) or baclofen (20 M) alone, as expected if a Ca2�
rise results from the synergistic effect of GABAAR-GABABR coactivation.
GABAR ligands modulate mESC proliferation
The capacity of mESCs for unlimited proliferation andmaintenance of the pluripotent state in vitro is largelydependent on the addition of cytokine LIF and FCS/bone morphogenetic protein (BMP) and is modulatedby additional factors (33, 44–46). Since GABARs havebeen previously shown to modulate the proliferation ofboth neuronal and non-neuronal cells, we sought totest the effect of modulators of GABAR-mediated sig-naling on mESC proliferation. To this end, we esti-mated the cell numbers or total cell area/colony size at12–36 h following addition of GABAR ligands (24–48 hpostplating) in the presence of LIF/FCS.
No statistically significant differences in total cell num-bers were encountered after 12 h treatment (Fig. 6A, day1) with muscimol (100 M), SR95531 (50 M), baclofen(50 M), or CGP 55845 (20 M; not shown). Theexception was the GABAAR blocker bicuculline, whichproved highly toxic at concentrations used for blocking theGABAARs (10–30 M) and was thus omitted.
Longer (36 h) treatment with GABAR ligands re-vealed a more complex pattern (Fig. 6A, B1, B2). Whilethe GABAAR agonist muscimol did not significantlyalter the proliferation rate compared with control, theGABABR agonist baclofen greatly increased total cellnumbers and clone sizes (Fig. 6A, day 2; B1, B2). Thenet effect of muscimol (100 M) and baclofen (50 M)administered together roughly corresponded to the
average of the single ligands (Fig. 6A, B1, B2) and wasalmost identical to the effect of 1 mM GABA, while 1M GABA was clearly inhibitory (data not shown). Apronounced facilitation of proliferation by nearly dou-bling the cell number/average clone size comparedwith control cultures was observed after treatment withthe specific GABAAR antagonist SR95531 (50 M; Fig.6A, B1, B2). It should be stressed that in the absence ofLIF and/or FCS, the proliferative capacity of mESCswas impaired (data not shown).
The undifferentiated state of mESCs was verified bystaining cultures treated in parallel with GABAR li-gands for placental AP (Fig. 6C) and Oct-3/4 (notshown). Overall, both control and most of ligand-treated cultures remained undifferentiated, as judgedby the uniformity of AP staining (Fig. 6C) at the end ofthe incubation period. However, signs of initial differ-entiation, such as out-migration and cell flattening(33), were observed at the periphery of large clones. Inaddition, SR95531 and muscimol-treated culturesshowed mosaic staining for AP (Fig. 6C). All culturesstained prominently for the pluripotency marker Oct-3/4 (data not shown).
We also checked for changes in the expression ofconnexin 43 (Cx43; Fig. 6C, D2), an obligatory compo-nent of the gap junction intercellular communications(GJIC) in mESC (47) that have been implicated in theregulation of proliferation and/or differentiation ofESCs and early embryos (48). Profound differencesbetween control and ligand-treated cultures were ob-served (Fig. 6C; Cx43): muscimol and SR95531 causedreduction, whereas GABA and especially baclofen treat-
Figure 3. GABAAR activation induces [Ca2�]i rise in mESC.A) Representative recordings of the muscimol-evoked changes influorescence in Fluo-4/AM-loaded mESCs. Cultures wereinitially perfused with buffer containing the antagonist bicu-culline, followed by coapplication of bicuculline and muscimol and finally by muscimol alone. B, C) Muscimol responseswere observed also in the presence of CPA (B), a SERCA inhibitor, or the VGCC blocker CdCl2 (Cd2�; C). Left panels:confocal images taken at different time points; cells manifesting [Ca2�]i transients are circled. Right panels: plots ofnormalized changes in fluorescence intensities [F (%)] against time measured in cells circled in the corresponding color.D) Average change in amplitude of responses (left panel) and number of cells/microscopic field detected for different drugapplications as indicated (right panel). Values are means � se.
5GABAR-MEDIATED SIGNALING IN MOUSE ES CELLS

ment led to an increase in membrane-bound Cx43staining that was stronger along adjacent cell borders.Only small differences at the protein level were de-tected in Western blots (Fig. 6D2).
Western blotting revealed �2-fold up-regulation ofOct-3/4 in samples prepared from GABAR ligand-treated cultures (Fig. 6D1), which is in the range offluctuation of this pluripotent marker in undifferenti-ated mESCs (49). Statistically significant (by 60%)
elevation of Oct-3/4 expression was observed only aftermuscimol treatment (Fig. 6D1).
DISCUSSION
mESC and GABAR expression
As mediators of inhibitory GABAergic neurotransmis-sion, GABARs are major players in controlling brain
Figure 4. GABABR agonist baclofen induces[Ca2�]i rise in mESCs. A) Representative re-cordings of GABABR-evoked change in fluores-cence of Fluo-4/AM-loaded mESCs. Cultureswere initially perfused with baclofen (GABABRagonist) and then CGP (GABABR antagonist),and finally baclofen and CGP were coadminis-tered. Note that CGP does not block the ba-clofen response, and continuous administra-tion of baclofen induces oscillations. B) Timecourse of fluorescence intensity changes mea-sured after addition of KCl followed by ba-clofen: baclofen further increases the KCl-evoked rise of fluorescence. C) Baclofeninduced calcium transients in mESCs after 3min preincubation with the VGCC blocker cad-mium (Cd2�). D) Baclofen evoked [Ca2�]i risein Ca2�-free buffer. Left panel: cells manifest-ing [Ca2�]i transients are circled on imagestaken at different time points. Right panels:normalized fluorescence changes [F (%)] rela-tive to basal level plotted vs. time in the circledcells. E) Average change in amplitude [F (%);bottom panel] and number of cells/micro-scopic field responding to respective treat-ments, as indicated (top panel). *P � 0.05;**P � 0.01
.
Figure 5. GABA is a more potent inducer of [Ca2�]i transients in mESCs compared withmuscimol or baclofen alone. A1, A2) Representative recordings of change in fluorescence inFluo-4/AM-loaded mESCs evoked by GABA. Note that Ca2� transients are followed by
low-amplitude oscillations (A2). A1) Images taken at different time points of the recordings; cells manifesting [Ca2�]itransients are circled. A2) Time course of normalized changes in fluorescence intensity [F (%)] in circled cells. B1,B2) Average number of cells/microscopic field responding to GABA, muscimol, or baclofen (B1) and change in amplitudeof responses [F (%)(B2)]. Values are means � se.
6 Vol. 24 April 2010 SCHWIRTLICH ET AL.The FASEB Journal � www.fasebj.org

excitability, and their assembly and function are beststudied in the adult central nervous system (CNS).However, GABARs of similar composition perform adiversity of functions outside the CNS and during embry-onic development (4, 30, 50–52). Here we show for thefirst time that functional GABAARs and GABABRs areexpressed in mESCs derived from and developmentallyclosely related to the ICM of the mouse blastocyst, areflection of the developmental program of the mousepreimplantation embryo. Remarkably, 11 of the 19GABAAR subunits known to date were moderately orabundantly expressed in undifferentiated mESCs. Wedetected all but 2 � subunits (namely �2 and �6), all �subunits, as well as the � and ε subunits, but only asingle � subunit (�3). Expression of the �3 subunit isvery restricted in the adult brain (6, 7, 53), but it is themost abundant � subunit in mESCs, similar to thedeveloping optic lens (37), pointing to the existence ofcommon features of the embryonic GABAAR composi-tion. The absence of �2 indicates mESCs lack the major“synaptic” GABAAR type composed of 2�1/1�2/2�2 (5,6). The abundant expression of �5, �4, �, and εsubunits that localize mostly or exclusively extrasynap-tically (54, 55) in the brain suggests the prevalence ofextrasynaptic GABAARs that are known to mediate slowand long-lasting “tonic” inhibition (8) and in generalpossess a high affinity for GABA, smaller conductanceswith slower decay and/or desensitization (3, 8, 56).Furthermore, �- and ε-containing extrasynaptic recep-
tors can be potentiated by alcohol, anesthetics, or neuro-steroids (6, 8, 54) or can even form constitutively activereceptors (57, 58) that function in the absence of GABA.This may partly explain the lack of an overt phenotype ofpreimplantation GAD65/67 double knockout embryos(unpublished results). Finally, the cytoplasmic-to-mem-brane shift of GABAAR-�3 indicates a predominant role ofGABAARs in larger clones.
The heterodimeric GABABRs assemble from GABABR2and either GABABR1a or GABABR1b, both nearly equallyabundant in mESCs. This strongly suggests the existenceof a heterogeneous receptor population that may segre-gate unequally among mESCs as described for the ner-vous system (12). Further heterogeneity could be predictedfrom putative interactions involving the extracellular “sushidomains” (59) or formation of intracellular complexes be-tween GABAAR-�2S and GABABR1 (60) or RGS-mediatedbinding to coupled GIRK channels (61). The striking “vesic-ular” appearance of the GABABR staining pattern, previouslydescribed in neurons (62) and Schwann cells (27), is con-sistent with the existence of multiprotein GABABR2-con-taining complexes.
GABAARs and GABABRs synergistically elevate [Ca2�]i,acting through distinct Ca2� signaling mechanisms
A novel finding of this study is the abundant expressionof both GABAR types in mESC and their synergistic
Figure 6. GABAR agonists and antagonists differentiallyaffect the proliferation of mESCs. mESCs were treatedwith 100 M muscimol, 50 M SR-95531, 50 M baclofen,1 mM GABA, or 50 M baclofen � 100 M muscimol(bacl/musc). A) Change in fluorescence in multiplytreated and control mESC cultures, measured using thefuorescence-based CyQuant cell proliferation assay underconditions favoring self-renewal (1000 U/ml LIF, 20%FCS). B1) Average size and total area occupied by mESCcolonies as measured in ImageJ, with cresyl violet staining.B2) Representative whole-field images of wells treated withdifferent GABAR ligands, as indicated. Opposite to A, theSR95531-treated cultures in B1 display higher values thanbaclofen-treated cultures, reflecting increase in cell sur-face. Bars represent means � se of 3 experiments. *P �0.05; **P � 0.01. C) Representative images of ligand-treated mESC cultures stained for AP and immunostainedfor Cx43. An apparent decrease in AP and Cx43 staining isseen in some clones after treatment with GABA, musci-
mol, and SR55931 (arrowheads); strong uniform staining for AP and Cx43 is clearly observed in control or baclofen-treatedcultures independent of clone size. D1, D2) Western blots of homogenates from ligand-treated and control cultures stainedwith Oct-3/4 (D1) and Cx43 (D2) antibodies. Bar charts show intensity of Oct-3/4 (45 kDa) and Cx43 (43 kDa) specificbands, normalized to �-actin.
7GABAR-MEDIATED SIGNALING IN MOUSE ES CELLS

effect on the [Ca2�]i level on activation with specificligands. The GABAAR-mediated [Ca2�]i rise in re-sponse to muscimol is clearly of a dual nature, due toCa2� influx through Cd2�-sensitive VGCCs and releasefrom CPA-sensitive (IP3/RyR) internal Ca2� stores(schematically presented in Fig. 7). Accordingly, thecellular machinery required for these responses ispresent in mESCs. The mechanism of [Ca2�]i increaseinduced by paracrine GABAAR activation has beenthoroughly studied in the embryonic nervous system(1–3), and mESCs are no exception from the classicalexamples. Unexpectedly, GABABRs of mESCs also in-duced Ca2� transients, which almost exclusively in-volved release from intracellular Ca2� stores, as they
were independent of externally added Ca2� or thepresence of VGCC inhibitors but could be blockedalmost entirely by the SERCA blocker CPA. This re-sponse is very similar to the recently described ba-clofen-evoked and entirely IP3-mediated [Ca2�]i rise inprimary astroglia cultures (19). Previous data showedthat mESCs or ICM express exclusively IP3Rs, and thereis a later developmental switch from IP3R- to RyR-mediated Ca2� signaling (63, 64). However, the pres-ence of RyRs in mouse (this study) and human (65)ESCs and both mouse and human preimplantationembryos (66, 67) may be functionally relevant. Forinstance, RyRs may mediate CICR in the potentiation of(KCl-induced) [Ca2�]i rise by baclofen, as described
Figure 7. Hypothetical model of GABAR-evoked Ca2�-mediated signaling in mESCs. A) Low concentration of GABA and/orneurosteroids activates GABAARs. This leads to efflux of Cl�, activation of VGCCs, and influx of [Ca2�]i, which activates aCa2�-dependent adenylyl cyclase (AC), resulting in synthesis of cAMP from ATP, activation of MAPK and CREB, and inhibitionof self-renewal. High concentrations of ambient GABA (B) or extracellular Ca2� (A) activate GABABRs coupled to inhibitoryGo/Gi proteins, leading to inhibition of AC and a drop in cAMP levels and cAMP-dependent PKA activity, resulting in reductionof MAPK, CREB phosphorylation, and stimulation of proliferation (A, B). Activation of phospholipase C (PLC) by G�� followingactivation of GABABR and GIRK channels results in production of IP3 [and diacylglycerol (DAG)] which, reacting with IP3Rs,induces Ca2� release from the ER stores (B). A positive feedback activation of PLC (and increased IP3 production) by Ca2�
released from ER and/or store-operated channel (SOC)-mediated influx (73) or by DAG (74) may explain the persistent Ca2�
oscillations that are expected to stimulate self-renewal. Both Ca2� and/or IP3 can enter into neighboring cells throughgap-junction channels (from B to A and C, respectively) inducing short-range intercellular Ca2�, IP3 (and possibly cAMP) waves,and oscillations. Gap-junction hemichannels are permeable for ATP (C, black star), which can activate G-protein-coupled P2Ypurinergic receptors driving PLC activation and IP3 production (C), followed by long-range intercellular Ca2� waves andoscillations. Ca2� is pumped into the ER stores by SERCA and removed from the cytoplasm by the plasma membraneCa2�-ATPase (PMCA; B). SOC (A) mediates the capacitive Ca2� entry following emptying of the intracellular stores afterGABABR activation. Cytosolic Ca2� binds to calmodulin, calcineurin, and many other Ca2�-binding proteins to mediate a widerange of signaling events. Note that RyRs have been omitted since they are functionally similar to IP3Rs but activated by Ca2�
instead.
8 Vol. 24 April 2010 SCHWIRTLICH ET AL.The FASEB Journal � www.fasebj.org

previously for the cerebellar granule neurons (68).Interestingly, the baclofen-triggered [Ca2�]i rise in thelatter experimental paradigm was similarly resistant toCGP35348 (Fig. 4A). Another feature of the GABABR-mediated Ca2� signaling in mESCs is the synchronousinduction of transients in neighboring cells, nearlyalways followed by intercellular wave propagation of thesignal and persistent oscillations. Intercellular Ca2�
waves are thought to arise from the spread of Ca2�
and/or IP3 through gap-junction channels abundantlyexpressed in mESCs or activation of the purinergicreceptors (P2Y) by ATP released though gap-junctionhemichannels (Fig. 7; refs. 69–71). This type of inter-cellular Ca2� signaling becomes functionally importantat the blastocyst stage, concomitant with the increase incell numbers and the demand for coordinated cellularactivity (71). The long-lasting GABABR-evoked Ca2�
oscillations, which may be accompanied by cAMP oscil-lations (Fig. 7; ref. 72) could result from constitutiveactivation of GABABR (or GIRK)-coupled G-proteins(19) or from the persistent activation of a downstreamsecond messenger (i.e., Ca2� and/or IP3) signalingcascade (Fig. 7; refs. 73, 74).
GABAARs and GABABRs oppositely modulate mESCproliferation
Ca2� signaling in the form of Ca2� transients, oscilla-tions, and intracellular Ca2� waves regulates both pro-liferation and differentiation during early embryonicdevelopment (71, 75–77). Furthermore, spontaneous cellcycle-dependent IP3-mediated Ca2� oscillations in mESCsand ICM are required for progression throughthe G1/Sphases (78). In the presence of the cytokine LIF, whichsignals through activation of the transcription factorSTAT3 and FCS/BMP, mESCs undergo unlimited sym-metrical divisions with a short cell cycle that virtuallylacks the G1 and G2 gap phases (32, 44–46, 79). Incontrast, the mitogen-activated protein kinase (MAPK)acting on the early G1 phase stimulates somatic cellproliferation and inhibits mESC self-renewal, whileinhibition of MAPK signaling enhances self-renewal ofmESCs (44).
In contrast to their synergistic elevation of [Ca2�]i,GABAARs and GABABRs impose opposite effects onmESC proliferation operating through distinct Ca2� sig-naling and may affect distinct downstream second-mes-senger pathways (Fig. 7). In immature neurons, GABAARstrigger depolarization, influx of Ca2� through VGCCs,activation of adenylyl cyclase (AC), cAMP and MAPKsignaling, activation of CREB (cAMP response elementbinding) protein, and an increase in brain-derived neu-rotropic factor synthesis (80, 81), ultimately promotingdifferentiation. Based on the present data, it seems plau-sible to suggest that in the presence of LIF/FCS, persis-tent tonic activation of GABAARs acts mainly to prolongthe G1 phase, leading to inhibition of self-renewal andstimulation of somatic cell proliferation, while GABAARblockade causes a shortening of G1 and stimulation ofproliferation. In contrast, GABABRs coupled to pertussis
toxin-sensitive Go/G1 proteins inhibit membrane-boundAC, reduce the level of cAMP, and thus stimulate prolif-eration (44). Another route based on GABABR-mediatedCa2� signaling reported here may involve an increase inIP3 through activation of phospholipase C by the G-protein �� subunits (Fig. 7) and inhibition of MAPKby a protein kinase C-mediated mechanism (82).This route is also expected to enhance proliferation.Finally, the GABABR-induced Ca2� oscillations facili-tate the progression through the G1/S checkpoint (78),stimulating self-renewal (symmetric division), consistent withour observation (Fig. 6) that baclofen enhances proliferationwithout pronounced differentiation.
These diverse proliferation effects correlate with areduced (GABAAR) or enhanced (GABABR) surfaceexpression of connexin 43 (Cx43), implying that GJICsare important mediators of GABAR effects. Consistentwith this, GJICs have been recently implicated in theproliferation and maintenance of pluripotency ofmESCs (83) and in the cell-cell coupling and prolifer-ation of neural progenitors (48, 69).
Natural sources of GABA for proliferating mESCscan be the FCS and/or mESCs, which can accumulateand release GABA via both vesicular and nonvesicularGABA mechanisms (unpublished results).
CONCLUSIONS
Our data suggest that “cross-talk” between the twoGABARs modulates the proliferation rate, dependenton the availability of GABA and/or allosteric modula-tors, expression levels of the 2 receptors, downstreameffectors, and intercellular coupling. The proposedmechanism may partially account for the growth regu-lation of the ICM as well. Finally, GABAR-mediatedsignaling is a potent modulator of the proliferation ofembryonic carcinoma (EC) cell line P19 (unpublishedresults) and may play a similar role in some aggressivecancer cells for which GABARs have been proposed aspossible therapeutical targets (84, 85).
The authors thank Reinhardt Fassler and Attila Aszodi (MaxPlanck Institute of Biochemistry, Martinsried, Germany) and ElenGocza [Agricultural Biotechnology Center (ABC), Godollo, Hun-gary] for providing mESCs and helping with setting up the mESCculture; Zsolt Selmeczy and Gyorgy Hasko (Institute of Experimen-tal Medicine, Budapest, Hungary) for providing equipment; andKatalin Dome for excellent technical assistance. The authors thankMark Eyre for critical reading of the manuscript. Part of this workwas presented in abstract form at the FENS Forum 2006.
REFERENCES
1. Ben-Ari, Y., Gaiarsa, J. L., Tyzio, R., and Khazipov, R. (2007)GABA: a pioneer transmitter that excites immature neurons andgenerates primitive oscillations. Physiol. Rev. 87, 1215–1284
2. Barker, J. L., Behar, T., Li, Y. X., Liu, Q. Y., Ma, W., Maric, D.,Maric, I., Schaffner, A. E., Serafini, R., Smith, S. V., Somogyi, R.,Vautrin, J. Y., Wen, X. L., and Xian, H. (1998) GABAergic cells andsignals in CNS development. Perspect. Dev. Neurobiol. 5, 305–322
3. Owens, D. F., and Kriegstein, A. R. (2002) Is there more toGABA than synaptic inhibition? Nat. Rev. Neurosci. 3, 715–727
9GABAR-MEDIATED SIGNALING IN MOUSE ES CELLS

4. Watanabe, M., Maemura, K., Kanbara, K., Tamayama, T., andHayasaki, H. (2002) GABA and GABA receptors in the centralnervous system and other organs. Int. Rev. Cytol. 213, 1–47
5. Mody, I., and Pearce, R. A. (2004) Diversity of inhibitory neurotrans-mission through GABA(A) receptors. Trends Neurosci. 27, 569–575
6. Mohler, H. (2006) GABA(A) receptor diversity and pharmacol-ogy. Cell Tissue Res. 326, 505–516
7. Sieghart, W., and Sperk, G. (2002) Subunit composition, distri-bution and function of GABA(A) receptor subtypes. Curr. Top.Med. Chem. 2, 795–816
8. Mody, I. (2008) Extrasynaptic GABAA receptors in the cross-hairs of hormones and ethanol. Neurochem. Int. 52, 60–64
9. Bettler, B., Kaupmann, K., Mosbacher, J., and Gassmann, M.(2004) Molecular structure and physiological functions ofGABA(B) receptors. Physiol. Rev. 84, 835–867
10. Bettler, B., and Tiao, J. Y. (2006) Molecular diversity, traffickingand subcellular localization of GABAB receptors. Pharmacol.Ther. 110, 533–543
11. Ulrich, D., and Bettler, B. (2007) GABA(B) receptors: synaptic func-tions and mechanisms of diversity. Curr. Opin. Neurobiol. 17, 298–303
12. Huang, Z. J. (2006) GABAB receptor isoforms caught in actionat the scene. Neuron 50, 521–524
13. Obrietan, K., and van den Pol, A. N. (1998) GABAB receptor-mediated inhibition of GABAA receptor calcium elevations indeveloping hypothalamic neurons. J. Neurophysiol. 79, 1360–1370
14. Obrietan, K., and van den Pol, A. N. (1999) GABAB receptor-mediated regulation of glutamate-activated calcium transientsin hypothalamic and cortical neuron development. J. Neuro-physiol. 82, 94–102
15. Hirono, M., Yoshioka, T., and Konishi, S. (2001) GABA(B)receptor activation enhances mGluR-mediated responses atcerebellar excitatory synapses. Nat. Neurosci. 4, 1207–1216
16. Tabata, T., Araishi, K., Hashimoto, K., Hashimotodani, Y., vander Putten, H., Bettler, B., and Kano, M. (2004) Ca2� activity atGABAB receptors constitutively promotes metabotropic gluta-mate signaling in the absence of GABA. Proc. Natl. Acad. Sci.U. S. A. 101, 16952–16957
17. Nilsson, M., Eriksson, P. S., Ronnback, L., and Hansson, E.(1993) GABA induces Ca2� transients in astrocytes. Neuroscience54, 605–614
18. Parramon, M., Gonzalez, M. P., Herrero, M. T., and Oset-Gasque, M. J. (1995) GABAB receptors increase intracellularcalcium concentrations in chromaffin cells through two differ-ent pathways: their role in catecholamine secretion. J. Neurosci.Res. 41, 65–72
19. Meier, S. D., Kafitz, K. W., and Rose, C. R. (2008) Developmen-tal profile and mechanisms of GABA-induced calcium signalingin hippocampal astrocytes. Glia 56, 1127–1137
20. Varju, P., Katarova, Z., Madarasz, E., and Szabo, G. (2001) GABAsignalling during development: new data and old questions. CellTissue Res. 305, 239–246
21. Wang, D. D., Kriegstein, A. R., and Ben-Ari, Y. (2008) GABAregulates stem cell proliferation before nervous system forma-tion. Epilepsy Curr. 8, 137–139
22. LoTurco, J. J., Owens, D. F., Heath, M. J., Davis, M. B., andKriegstein, A. R. (1995) GABA and glutamate depolarize corti-cal progenitor cells and inhibit DNA synthesis. Neuron 15,1287–1298
23. Liu, X., Wang, Q., Haydar, T. F., and Bordey, A. (2005)Nonsynaptic GABA signaling in postnatal subventricular zonecontrols proliferation of GFAP-expressing progenitors. Nat.Neurosci. 8, 1179–1187
24. Watanabe, M., Maemura, K., Oki, K., Shiraishi, N., Shibayama,Y., and Katsu, K. (2006) Gamma-aminobutyric acid (GABA) andcell proliferation: focus on cancer cells. Histol. Histopathol. 21,1135–1141
25. Haydar, T. F., Wang, F., Schwartz, M. L., and Rakic, P. (2000)Differential modulation of proliferation in the neocorticalventricular and subventricular zones. J. Neurosci. 20, 5764–5774
26. Fiszman, M. L., Borodinsky, L. N., and Neale, J. H. (1999) GABAinduces proliferation of immature cerebellar granule cellsgrown in vitro. Brain. Res. Dev. Brain Res. 115, 1–8
27. Magnaghi, V., Ballabio, M., Cavarretta, I. T., Froestl, W., Lam-bert, J. J., Zucchi, I., and Melcangi, R. C. (2004) GABABreceptors in Schwann cells influence proliferation and myelinprotein expression. Eur. J. Neurosci. 19, 2641–2649
28. Luyt, K., Slade, T. P., Dorward, J. J., Durant, C. F., Wu, Y.,Shigemoto, R., Mundell, S. J., Varadi, A., and Molnar, E. (2007)Developing oligodendrocytes express functional GABA(B) re-ceptors that stimulate cell proliferation and migration. J. Neuro-chem. 100, 822–840
29. Fukui, M., Nakamichi, N., Yoneyama, M., Ozawa, S., Fujimori, S.,Takahata, Y., Nakamura, N., Taniura, H., and Yoneda, Y. (2008)Modulation of cellular proliferation and differentiationthrough GABA(B) receptors expressed by undifferentiated neu-ral progenitor cells isolated from fetal mouse brain. J. Cell.Physiol. 216, 507–519
30. Tamayama, T., Maemura, K., Kanbara, K., Hayasaki, H., Yabu-moto, Y., Yuasa, M., and Watanabe, M. (2005) Expression ofGABA(A) and GABA(B) receptors in rat growth plate chondro-cytes: activation of the GABA receptors promotes proliferationof mouse chondrogenic ATDC5 cells. Mol. Cell. Biochem. 273,117–126
31. Martin, G. R. (1981) Isolation of a pluripotent cell line fromearly mouse embryos cultured in medium conditioned byteratocarcinoma stem cells. Proc. Natl. Acad. Sci. U. S. A. 78,7634–7638
32. Niwa, H. (2007) How is pluripotency determined and main-tained? Development 134, 635–646
33. Silva, J., and Smith, A. (2008) Capturing pluripotency. Cell 132,532–536
34. Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W., andRoder, J. C. (1993) Derivation of completely cell culture-derivedmice from early-passage embryonic stem cells. Proc. Natl. Acad.Sci. U. S. A. 90, 8424–8428
35. Kwakowsky, A., Schwirtlich, M., Zhang, Q., Eisenstat, D. D.,Erdelyi, F., Baranyi, M., Katarova, Z. D., and Szabo, G. (2007)GAD isoforms exhibit distinct spatiotemporal expression pat-terns in the developing mouse lens: correlation with Dlx2 andDlx5. Dev. Dyn. 236, 3532–3544
36. Tielens, S., Wymeersch, F., Declercq, H., and Cornelissen, M.(2008) Effect of 17beta-estradiol on the in vitro differentiationof murine embryonic stem cells into the osteogenic lineage. InVitro Cell. Dev. Biol. 44, 368–378
37. Kwakowsky, A., Schwirtlich, M., Kooy, F., Abraham, I., Mate, Z.,Katarova, Z., and Szabo, G. (2008) GABA neurotransmittersignaling in the developing mouse lens: dynamic regulation ofcomponents and functionality. Dev. Dyn. 237, 3830–3841
38. Kaupmann, K., Malitschek, B., Schuler, V., Heid, J., Froestl, W.,Beck, P., Mosbacher, J., Bischoff, S., Kulik, A., Shigemoto, R.,Karschin, A., and Bettler, B. (1998) GABA(B)-receptor subtypesassemble into functional heteromeric complexes. Nature 396,683–687
39. Koyrakh, L., Lujan, R., Colon, J., Karschin, C., Kurachi, Y.,Karschin, A., and Wickman, K. (2005) Molecular and cellulardiversity of neuronal G-protein-gated potassium channels.J. Neurosci. 25, 11468–11478
40. Wang, H., Matsumoto, H., Guo, Y., Paria, B. C., Roberts, R. L.,and Dey, S. K. (2003) Differential G protein-coupled cannabi-noid receptor signaling by anandamide directs blastocyst activa-tion for implantation. Proc. Natl. Acad. Sci. U. S. A. 100, 14914–14919
41. Rossi, D., and Sorrentino, V. (2002) Molecular genetics ofryanodine receptors Ca2�-release channels. Cell Calcium 32,307–319
42. Bain, G., Ramkumar, T. P., Cheng, J. M., and Gottlieb, D. I.(1993) Expression of the genes coding for glutamic acid decar-boxylase in pluripotent cell lines. Brain Res. Mol. Brain Res. 17,23–30
43. Bain, G., Kitchens, D., Yao, M., Huettner, J. E., and Gottlieb,D. I. (1995) Embryonic stem cells express neuronal propertiesin vitro. Dev. Biol. 168, 342–357
44. Orford, K. W., and Scadden, D. T. (2008) Deconstructing stemcell self-renewal: genetic insights into cell-cycle regulation. Nat.Rev. Genet. 9, 115–128
45. Chambers, I., and Smith, A. (2004) Self-renewal of teratocarci-noma and embryonic stem cells. Oncogene 23, 7150–7160
46. Ying, Q. L., Wray, J., Nichols, J., Batlle-Morera, L., Doble, B.,Woodgett, J., Cohen, P., and Smith, A. (2008) The ground stateof embryonic stem cell self-renewal. Nature 453, 519–523
47. Worsdorfer, P., Maxeiner, S., Markopoulos, C., Kirfel, G., Wulf,V., Auth, T., Urschel, S., von Maltzahn, J., and Willecke, K.(2008) Connexin expression and functional analysis of gap
10 Vol. 24 April 2010 SCHWIRTLICH ET AL.The FASEB Journal � www.fasebj.org

junctional communication in mouse embryonic stem cells. StemCells 26, 431–439
48. Wong, R. C., Pera, M. F., and Pebay, A. (2008) Role of gapjunctions in embryonic and somatic stem cells. Stem Cell Rev. 4,283–292
49. Niwa, H., Miyazaki, J., and Smith, A. G. (2000) Quantitativeexpression of Oct-3/4 defines differentiation, dedifferentiationor self-renewal of ES cells. Nat. Genet. 24, 372–376
50. Erdo, S. L., and Wolff, J. R. (1990) Gamma-aminobutyric acidoutside the mammalian brain. J. Neurochem. 54, 363–372
51. Kanbara, K., Okamoto, K., Nomura, S., Kaneko, T., Shigemoto,R., Azuma, H., Katsuoka, Y., and Watanabe, M. (2005) Cellularlocalization of GABA and GABAB receptor subunit proteinsduring spermiogenesis in rat testis. J. Androl. 26, 485–493
52. Xiang, Y. Y., Wang, S., Liu, M., Hirota, J. A., Li, J., Ju, W., Fan, Y.,Kelly, M. M., Ye, B., Orser, B., O’Byrne, P. M., Inman, M. D.,Yang, X., and Lu, W. Y. (2007) A GABAergic system in airwayepithelium is essential for mucus overproduction in asthma.Nat. Med. 13, 862–867
53. Wisden, W., Laurie, D. J., Monyer, H., and Seeburg, P. H. (1992)The distribution of 13 GABAA receptor subunit mRNAs in therat brain. I. Telencephalon, diencephalon, mesencephalon.J. Neurosci. 12, 1040–1062
54. Collinson, N., Kuenzi, F. M., Jarolimek, W., Maubach, K. A.,Cothliff, R., Sur, C., Smith, A., Otu, F. M., Howell, O., Atack,J. R., McKernan, R. M., Seabrook, G. R., Dawson, G. R., Whiting,P. J., and Rosahl, T. W. (2002) Enhanced learning and memoryand altered GABAergic synaptic transmission in mice lackingthe alpha 5 subunit of the GABAA receptor. J. Neurosci. 22,5572–5580
55. Glykys, J., Mann, E. O., and Mody, I. (2008) Which GABA(A)receptor subunits are necessary for tonic inhibition in thehippocampus? J. Neurosci. 28, 1421–1426
56. Muller, E., Le-Corronc, H., and Legendre, P. (2008) Extrasyn-aptic and postsynaptic receptors in glycinergic and GABAergicneurotransmission: a division of labor? Front. Mol. Neurosci. 1, 3
57. Neelands, T. R., Fisher, J. L., Bianchi, M., and Macdonald, R. L.(1999) Spontaneous and gamma-aminobutyric acid (GABA)-activated GABA(A) receptor channels formed by epsilon sub-unit-containing isoforms. Mol. Pharmacol. 55, 168–178
58. Bollan, K. A., Baur, R., Hales, T. G., Sigel, E., and Connolly,C. N. (2008) The promiscuous role of the epsilon subunit inGABAA receptor biogenesis. Mol. Cell. Neurosci. 37, 610–621
59. Tiao, J. Y., Bradaia, A., Biermann, B., Kaupmann, K., Metz, M.,Haller, C., Rolink, A. G., Pless, E., Barlow, P. N., Gassmann, M.,and Bettler, B. (2008) The sushi domains of secreted GABA(B1)isoforms selectively impair GABA(B) heteroreceptor function.J. Biol. Chem. 283, 31005–31011
60. Balasubramanian, S., Teissere, J. A., Raju, D. V., and Hall, R. A.(2004) Hetero-oligomerization between GABAA and GABABreceptors regulates GABAB receptor trafficking. J. Biol. Chem.279, 18840–18850
61. Lomazzi, M., Slesinger, P. A., and Luscher, C. (2008) Addictivedrugs modulate GIRK-channel signaling by regulating RGSproteins. Trends Pharmacol. Sci. 29, 544–549
62. Ritter, B., Ochojski, M., Kuhn, T., Schwarzacher, S. W., andZhang, W. (2005) Subcellular vesicular aggregations of GABABR1a and R1b receptors increase with age in neurons of thedeveloping mouse brain. Cell Tissue Res. 319, 181–189
63. Rosemblit, N., Moschella, M. C., Ondriasa, E., Gutstein, D. E.,Ondrias, K., and Marks, A. R. (1999) Intracellular calciumrelease channel expression during embryogenesis. Dev. Biol.206, 163–177
64. Yanagida, E., Shoji, S., Hirayama, Y., Yoshikawa, F., Otsu, K.,Uematsu, H., Hiraoka, M., Furuichi, T., and Kawano, S. (2004)Functional expression of Ca2� signaling pathways in mouseembryonic stem cells. Cell Calcium 36, 135–146
65. Goh, G., Self, T., Barbadillo Munoz, M. D., Hall, I. P., Young, L.,and Denning, C. (2005) Molecular and phenotypic analyses ofhuman embryonic stem cell-derived cardiomyocytes: opportuni-ties and challenges for clinical translation. Thromb. Haemost. 94,728–737
66. Stachecki, J. J., and Armant, D. R. (1996) Transient release ofcalcium from inositol 1,4,5-trisphosphate-specific stores regu-
lates mouse preimplantation development. Development 122,2485–2496
67. Balakier, H., Dziak, E., Sojecki, A., Librach, C., Michalak, M., andOpas, M. (2002) Calcium-binding proteins and calcium-releasechannels in human maturing oocytes, pronuclear zygotes and earlypreimplantation embryos. Hum. Reprod. 17, 2938–2947
68. De Erausquin, G., Brooker, G., Costa, E., and Wojcik, W. J.(1992) Stimulation of high affinity gamma-aminobutyric acidBreceptors potentiates the depolarization-induced increase ofintraneuronal ionized calcium content in cerebellar granuleneurons. Mol. Pharmacol. 42, 407–414
69. Elias, L. A., and Kriegstein, A. R. (2008) Gap junctions: multi-faceted regulators of embryonic cortical development. TrendsNeurosci. 31, 243–250
70. Verkhratsky, A., Orkand, R. K., and Kettenmann, H. (1998)Glial calcium: homeostasis and signaling function. Physiol. Rev.78, 99–141
71. Webb, S. E., and Miller, A. L. (2003) Calcium signalling duringembryonic development. Nat. Rev. Mol. Cell. Biol. 4, 539–551
72. Willoughby, D., and Cooper, D. M. (2007) Organization andCa2� regulation of adenylyl cyclases in cAMP microdomains.Physiol. Rev. 87, 965–1010
73. Thore, S., Dyachok, O., Gylfe, E., and Tengholm, A. (2005)Feedback activation of phospholipase C via intracellular mobi-lization and store-operated influx of Ca2� in insulin-secretingbeta-cells. J. Cell. Sci. 118, 4463–4471
74. Hisatsune, C., Nakamura, K., Kuroda, Y., Nakamura, T., andMikoshiba, K. (2005) Amplification of Ca2� signaling by diacyl-glycerol-mediated inositol 1,4,5-trisphosphate production. J. Biol.Chem. 280, 11723–11730
75. Ducibella, T., Huneau, D., Angelichio, E., Xu, Z., Schultz, R. M.,Kopf, G. S., Fissore, R., Madoux, S., and Ozil, J. P. (2002)Egg-to-embryo transition is driven by differential responses toCa(2�) oscillation number. Dev. Biol. 250, 280–291
76. Lu, D. P., Li, Y., Bathgate, R., Day, M., and O’Neill, C. (2003)Ligand-activated signal transduction in the 2-cell embryo. Biol.Reprod. 69, 106–116
77. Jimenez-Gonzalez, C., Michelangeli, F., Harper, C. V., Barratt,C. L., and Publicover, S. J. (2006) Calcium signalling in humanspermatozoa: a specialized “toolkit” of channels, transportersand stores. Hum. Reprod. Update 12, 253–267
78. Kapur, N., Mignery, G. A., and Banach, K. (2007) Cell cycle-dependent calcium oscillations in mouse embryonic stem cells.Am. J. Physiol. Cell Physiol. 292, C1510–C1518
79. White, J., and Dalton, S. (2005) Cell cycle control of embryonicstem cells. Stem Cell Rev. 1, 131–138
80. Obrietan, K., and van den Pol, A. N. (1997) GABA activitymediating cytosolic Ca2� rises in developing neurons is modu-lated by cAMP-dependent signal transduction. J. Neurosci. 17,4785–4799
81. Obrietan, K., Gao, X. B., and Van Den Pol, A. N. (2002)Excitatory actions of GABA increase BDNF expression via aMAPK-CREB-dependent mechanism–a positive feedback circuitin developing neurons. J. Neurophysiol. 88, 1005–1015
82. Shah, B. H., Farshori, M. P., and Catt, K. J. (2004) Neuropep-tide-induced transactivation of a neuronal epidermal growthfactor receptor is mediated by metalloprotease-dependent for-mation of heparin-binding epidermal growth factor. J. Biol.Chem. 279, 414–420
83. Todorova, M. G., Soria, B., and Quesada, I. (2008) Gap junc-tional intercellular communication is required to maintainembryonic stem cells in a non-differentiated and proliferativestate. J. Cell Physiol. 214, 354–362
84. Takehara, A., Hosokawa, M., Eguchi, H., Ohigashi, H., Ishikawa,O., Nakamura, Y., and Nakagawa, H. (2007) Gamma-aminobutyricacid (GABA) stimulates pancreatic cancer growth through overex-pressing GABAA receptor pi subunit. Cancer Res. 67, 9704–9712
85. Schuller, H. M., Al-Wadei, H. A., and Majidi, M. (2008) Gamma-aminobutyric acid, a potential tumor suppressor for small airway-derived lung adenocarcinoma. Carcinogenesis 29, 1979–1985
Received for publication October 14, 2009.Accepted for publication November 5, 2009.
11GABAR-MEDIATED SIGNALING IN MOUSE ES CELLS
Copyright © 2022 FDOKUMEN




















