
Computational Design of Peptides That Target Transmembrane Helices

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 864
Functional expression of CFTR in rat oviduct epitheliumActa Biochim Biophys Sin (2008): 864-872 | © 2008 Institute of Biochemistry and Cell Biology, SIBS, CAS | All Rights Reserved 1672-9145http://www.abbs.info; www.blackwellpublishing.com/abbs | DOI: DOI: 10.1111/j.1745-7270.2008. 00469.x
Functional expression of cystic fibrosis transmembrane conductance regulator inrat oviduct epithelium
Minhui Chen1, Jianyang Du1, Weijian Jiang3, Wulin Zuo1, Fang Wang1, Manhui Li1, Zhongluan Wu1, HsiaochangChan2*, and Wenliang Zhou1*1 School of Life Science, Sun Yat-sen University, Guangzhou 510275, China2 Epithelial Cell Biology Research Center, Department of Physiology, Faculty of Medicine, Chinese University of Hong Kong, Shatin,NT, Hong Kong, SAR3 Department of Pharmacology, Zhongshan School of Medicine, Sun Yat-sen University, Guangzhou 510080, China
Received: May 06, 2008 Accepted: August 06, 2008This work was supported by grants from the National Natural ScienceFoundation of China (No. 30770817), the the National Basic ResearchProgram of China (Nos. 2006CB504002 and 2006CB944002), and theSouth China National Research Center for Integrated Biosciences inCollaboration with Zhongshan University (No. 008)Corresponding authors:We n l i a n g Z h o u : T e l / F a x , 8 6 - 2 0 - 8 4 1 1 0 0 6 0 ; E - m a i l ,[email protected] Chan: Tel, 852-26961105; Fax, 852-26037155; Email,[email protected]
The aim of this study was to investigate the functionalexpression of cystic fibrosis transmembrane conductanceregulator (CFTR) with electrophysiological and moleculartechnique in rat oviduct epithelium. In whole-cell patch clamp,oviduct epithelial cells responded to 100 µM 8-bromo-adenosine 3',5'-cyclic monophosphate (8-Br-cAMP) with arise in inward current in Gap-free mode, which was inhibitedsuccessively by 5 µM CFTR(inh)-172, a CFTR specificinhibitor, and 1 mM diphenylamine-2-carboxylate (DPC), theCl− channel blocker. The cAMP-activated current exhibiteda linear current-voltage (I-V) relationship and time- andvoltage-independent characteristics. The reversal potentialsof the cAMP-activated currents in symmetrical Cl− solutionswere close to the Cl− equilibrium, 0.5±0.2 mV (n=4). WhenCl− concentration in the bath solution was changed from 140mM to 70 mM and a pipette solution containing 140 mM Cl−was used, the reversal potential shifted to a value close to thenew equilibrium for Cl−, 20±0.6 mV (n=4), as compared withthe theoretic value of 18.7 mV. In addition, mRNA expressionof CFTR was also detected in rat oviduct epithelium. Westernblot analysis showed that CFTR protein is found in the oviductthroughout the cycle with maximal expression at estrus, andimmunofluorescence and immunohistochemistry analysisrevealed that CFTR is located at the apical membrane of the
epithelial cells. These results showed that the cAMP-activatedCl− current in the oviduct epithelium was characteristic ofCFTR, which provided direct evidence for the functionalexpression of CFTR in the rat oviduct epithelium. CFTR mayplay a role in modulating fluid transport in the oviduct.
Keywords oviduct; cystic fibrosis transmembraneconductance regulator (CFTR); patch clamp; estrus cycle
Possessing complex physiological function, the oviduct isan important part of the female reproductive system. It iswell known that oviduct epithelial cells secret a variety ofnutrient substances and factors, which provide anappropriate environment for a series of reproductive events,including capacitation of sperm, maturation of oocyte andembryo development [1,2].
The mammalian oviduct is capable of active fluidsecretion from the blood into the lumen that is driven byelectrogenic Cl− secretion across the oviduct epithelium[3]. Cystic fibrosis transmembrane conductance regulator(CFTR) plays a critical role in the regulation of ion transportin a number of exocrine epithelia, including the lungs,intestine, pancreas and sweat gland duct [4−7]. AlthoughCFTR mRNA was detected in murine oviduct and cysticfibrosis (CF) mouse oviduct exhibited defective cAMP-mediated Cl− secretion [8−10], no direct evidence of theelectrophysiological characterization of CFTR as a cAMP-activated Cl− channel in the oviduct has been provided withpatch clamp technique. Therefore, one of the aims of ourpresent study was to investigate the electrophysiologicalproperties of CFTR in rat oviduct epithelium using thewhole-cell patch clamp technique.
Additionally, we investigated the cyclic variations in the

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 865
Functional expression of CFTR in rat oviduct epithelium
expression of CFTR in rat oviduct, as the number ofspermatozoa migrating through the oviduct is significantlyhigher in estrus than in other stages [11]; the high migrationrates may result from fluid accumulation, which is highestduring estrus [12]. CFTR expression may regulate uterinefluid production to facilitate sperm transport in differentstages. We examined the cyclic variations using Westernblot analysis.
Materials and Methods
AnimalsImmature and mature female Sprague-Dawley rats werepurchased from the Animal Center of Sun Yat-senUniversity (Guangzhou, China). Animals were housed andfed according to the guidelines of the Sun Yat-sen UniversityAnimal Use Committee; all procedures were approved priorto each experiment. Animals were kept in a room with aconstant temperature (20 ºC) with a 12L:12D photoperiodand were allowed to access food and water ad libitum.
Medium and drugsHanks’ balanced salt solution, penicillin/streptomycin,Dulbecco’s modified Eagle’s medium/F12 (DMEM/F12),0.25% trypsin and SuperScript One-Step reversetranscription-polymerase chain reaction (RT-PCR) withPlatinumTaq were purchased from Invitrogen and Gibco(Carlsbad, USA). Fetal bovine serum was from Hyclone(Logan, USA). Collagenase, 8-bromoadenosine 3',5'-cyclicmonophosphate (8-Br-cAMP), CFTR inhibitor(inh)-172and diphenylamine-2-dicarboxylic acid (DPC) werepurchased from Sigma (St. Louis, USA). The primersfor CFTR and glyceraldehyde-3-phosphate dehydrogenase(GAPDH) were synthesized by the Sangong Company(Shanghai, China).
Culture of rat oviduct epitheliumImmature female Sprague-Dawley rats weighing 100−120g were killed by CO2 inhalation. Their lower abdomenswere opened, and the oviducts were isolated andmicrodissected under sterile conditions to remove fat andconnective tissues. The tissues were cut into smallsegments, incubated in 0.25% (W/V) trypsin for 30 min at37 ºC and then in 1 mg/ml collagenase I for 10 min at37 ºC with vigorous shaking (150 strokes/min). The cellswere separated by centrifugation (800 g, 5 min). The pelletswere resuspended in DMEM/F12 containing 10% fetalbovine serum, penicillin (100 IU/ml), and streptomycin(100 µg/ml). The cell suspension was incubated at 37 ºCin 95% O2/5% CO2.
Immunofluorescence of cytokeratinParaformaldehyde-fixed primary rat oviduct epithelial cellswere washed in phosphate-buffered saline (PBS) andincubated with 1% bovine serum albumin (BSA) for 15min before incubation with the primary mouse anti-ratcytokeratin antibody (Cat. No. BM0030; BOSTER, Wuhan,China) at room temperature for 90 min. After three washeswith PBS, cells were incubated for 60 min with thesecondary anti-mouse IgG-fluorescein isothiocyanate(FITC) conjugate (BOSTER) in the dark, followed by threewashes with PBS. Cells were mounted onto glass slidesusing a 1:1 mixture of Vectashield medium (VectorLaboratories, Burlingame, USA) and 0.3 M Tris solution(pH 8.9). Cells were then viewed using TCS SP2 ConfocalImaging System (Leica Microsystems, Wetzlar, Germany).
Whole-cell patch clamp recordingAfter 2 d of culture, the rat oviduct epithelial cells wereused for patch clamping. The cell cultures were incubatedin Ca2+-free Dulbecco’s PBS solution containing 1 mMethylene glycol tetraacetic acid (EGTA) for 10−15 min toseparate the cells. The cells were then moved to a 1 mlchamber that was fixed on a X-Y axis stage of an OlympusBX51WI immersing lens microscopy system (Tokyo,Japan). Recording was performed at room temperatureusing a Multiclamp700A amplifier and the Digidata 1322series interface (AXON Instrument, Foster City, USA).Signals were filtered at 10 kHz. Pclamp 9.0 softwaresystem (AXON Instrument) was used for data recordingand analysis.
Using a horizontal puller P-97 (Sutter Instrument,Novato, USA) and glass pipettes, we pulled patch pipettes(1.2 mm outside diameter and 0.5 mm inside diameter).Ionic current was recorded using the conventional whole-cell patch clamp technique. The pipettes were filled with asolution containing 120 mM CsCl and 20 mM tetraethyl-ammonium-Cl (pH adjusted to 7.2 with CsOH). The bathsolution contained 135 mM NaCl, 1.2 mM Na2HPO4, 2mM MgCl2, 10 mM HEPES and 10 mM glucose (pH 7.4).The low Cl− (70 mM) bath solution contained 69 mM Na-glutamate, 66 mM NaCl, 1.2 mM Na2HPO4, 2 mM MgCl2,10 mM HEPES and 10 mM glucose (pH 7.4). The resistanceof the patch pipettes was approximately 2−7 MΩ. Positivepressure was applied in the patch pipette before it wasimmersed in the bath solution. When the tip of the pipetteattached to the cell surface, pressure was withdrawn, andthen a giga-seal between the pipette and cell membraneformed normally. After being sucked from the pipette, thewhole cell configuration was confirmed. The oviduct cell

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 866
Functional expression of CFTR in rat oviduct epithelium
was held at its resting potential −30 mV in the episodicrecording mode, and the voltage was clamped from −120mV to +100 mV in 20 mV increments. In the Gap-freerecording mode, the cells were held at −70 mV throughoutthe period of recording. The pipette potential and liquidjunction potential were auto-compensated by theMulticlamp700A amplifier before the electrode touched thecell.
Reverse transcription-PCRTotal RNA was isolated using TRIzol reagent (LifeTechnologies, Gaithersburg, USA), and 2 µg was used toamplify CFTR using Superscript II One-Step RT-PCR withPlatinum Taq (Invitrogen). The primers for CFTR were:sense 5′-GACAACATGGAACACATACCTTCG-3′(corresponding to nucleotides 2514−2537) and antisense5 ′ - TC TC G TTC G TTTC A C AG TC G G TG AG - 3 ′(corresponding to nucleotides 2771−2747), which yieldeda PCR product of 258 bp. The primers for GAPDH weresense 5′-ACTGGCGTCTTCACCACCAT-3′ and antisense5′-TCCACCACCCTGTTGCTGTA-3′, which yielded aPCR product of 300 bp. Reactions were carried out withthe following parameters: denaturation at 94 ºC for 1 min,annealing at 60 ºC for 45 s, and extension at 72 ºC for 1min, for a total of 45 cycles. PCR products were analyzedby agarose gel electrophoresis and visualized by stainingwith ethidium bromide.
Protein extraction and Western blot analysisOviduct tissue from mature female rats at different estrusstages were cut into small pieces and disrupted with aTenbroeck tissue grinder in 3 ml M-PER Mammalian ProteinExtraction Reagent, and complete protease inhibitors(Roche Applied Science, Indianapolis, USA). Homogenateswere centrifuged for 5 min at 14,000 g at 4 ºC. Thesupernatant was collected and the protein concentrationwas determined with BCA protein assay kit (PierceBiotechnology, Rockford, USA). Protein extracts werealiquoted and stored at −80 ºC. Protein was loaded onto8% Tris-glycine polyacrylamide gels (Cambrex, Rockland,USA). After SDS-PAGE separation, proteins weretransferred onto an Immun-Blot polyvinylidene difluoridemembrane (Bio-Rad Laboratories, Hercules, USA).Membranes were blocked in Tris-buffered saline (TBS)with 5% non-fat dry milk and then incubated overnight at4 ºC with rabbit anti-CFTR antibody (Cat. No. ACL-006;Alomone Labs, Jerusalem, Israel) diluted (1:1000) in TBSwith 2.5% non-fat dry milk. After being washed four timesin TBS with 0.1% Tween-20, membranes were incubatedwith donkey anti-rabbit IgG antibody conjugated to
horseradish peroxidase (Cell Signal Technology, Beverly,USA) for 1 h at room temperature. After an additional fourwashes, bindings were detected using Western Lightningchemiluminescence reagent (Perkin Elmer Life Sciences,Boston, USA). The gray scales on the bands of CFTRprotein were normalized to that of β-actin, which wasused as the loading control for the protein sample. Theratio meant the different expression levels of CFTR in otherthree stages as compared with that of proestrus but notsimply CFTR/β-actin.
Immunofluorescence of rat oviduct frozen sectionFresh isolated oviducts were embedded in Tissue-Tek OCTcompound 4583 (Sakura Finetek USA, Torrance, USA),mounted on a cutting block and frozen at −27 ºC. Sections(5 µm) were cut by a Reichert-Jung 1800 Frigocut cryostat(Leica Microsystems, Bannockburn, USA) and stored at4 ºC until use. Sections were rinsed in PBS for 5 min andtreated with 0.05% Triton X-100 for 1 min. Sections weresubsequently washed three times in PBS for 5 min each,incubated for 15 min in 1% (W/V) BSA in PBS with 1%sodium azide to prevent non-specific staining and thenincubated in the anti-CFTR antibody at room temperaturefor 120 min. The sections were incubated with PBS alone,which used as negative control. After three 5 min PBSwashes, the FITC-conjugated secondary antibody wasapplied for 1 h at room temperature, and the slides werethen rinsed again in PBS three times for 5 min. Slides weremounted in a 1:1 mixture of Vectashield medium and 0.3M Tris solution (pH 8.9), and then viewed under the TCSSP2 Confocal Imaging System.
Immunohistochemistry of rat oviduct paraffin sectionParaffin-embedded 5 µm sections of paraformaldehyde-fixed oviducts were dewaxed and hydrated. Antigen wasretrieved by treatment in 0.01 M citrate buffer (pH 6.0)for 20 min in a sub-boiling water bath. After rinsing withphosphate buffered saline Tween-20 (PBST), sections wereincubated in normal blocking serum for 30 min and thenwith the rabbit anti-rat CFTR antibodies diluted 1:100 withdiluting buffer (PBS with 1% BSA, 0.1% gelatine and0.05% sodium azide) at 4 ºC overnight. Sections werewashed three times with PBST and incubated withbiotinylated secondary antibody at 37 ºC for 30 min. Afterbeing washed three times with PBST, the sections wereincubated with the horseradish peroxidase conjugateddonkey anti-goat IgG antibody for 30 min and finallywashed three times with PBST. Visualization was achievedby immersing sections in a peroxidase substrate solution3 ,3 ′ -di aminob enz id ine- te t r ach lor ide (K40 11;

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 867
Functional expression of CFTR in rat oviduct epithelium
DakoCytomation, Glostrup, Denmark) until desired stainintensity was reached. Slides were rinsed with pure waterfor 5 min, counterstained with Harris hematoxylin (Sigma),dehydrated, and mounted for observation. Negativecontrols were incubated with antigen of primary antibody.
DNA sequencingThree 258 bp CFTR PCR products (from separate RNAisolations) were cloned into the pCR 2.1 vector (Invitrogen)using T4 DNA ligase, and they were then used to transform“super competent” Escherichia coli cells, which weregrown for 1 d. The DNA was isolated using an isolationkit (Promega, Madison, USA), and the plasmid wassequenced on the ABI PRISM sequencer model 2.1.1(Foster City, USA) and analyzed by BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi). All three sequences obtainedwere 100% homologous to region 2514−2771 of the ratCFTR gene.
Statistical analysisResults were expressed as mean±SEM, and n indicatedthe number of experiments. Data were analyzed usingStudent’s t-test. A P value less than 0.05 was consideredto be statistically significant.
Results
cAMP elicited whole-cell currents in cultured oviductepithelial cellsPath clamp recordings were applied on rat oviduct epithelialcells after two days of primary culture, which were
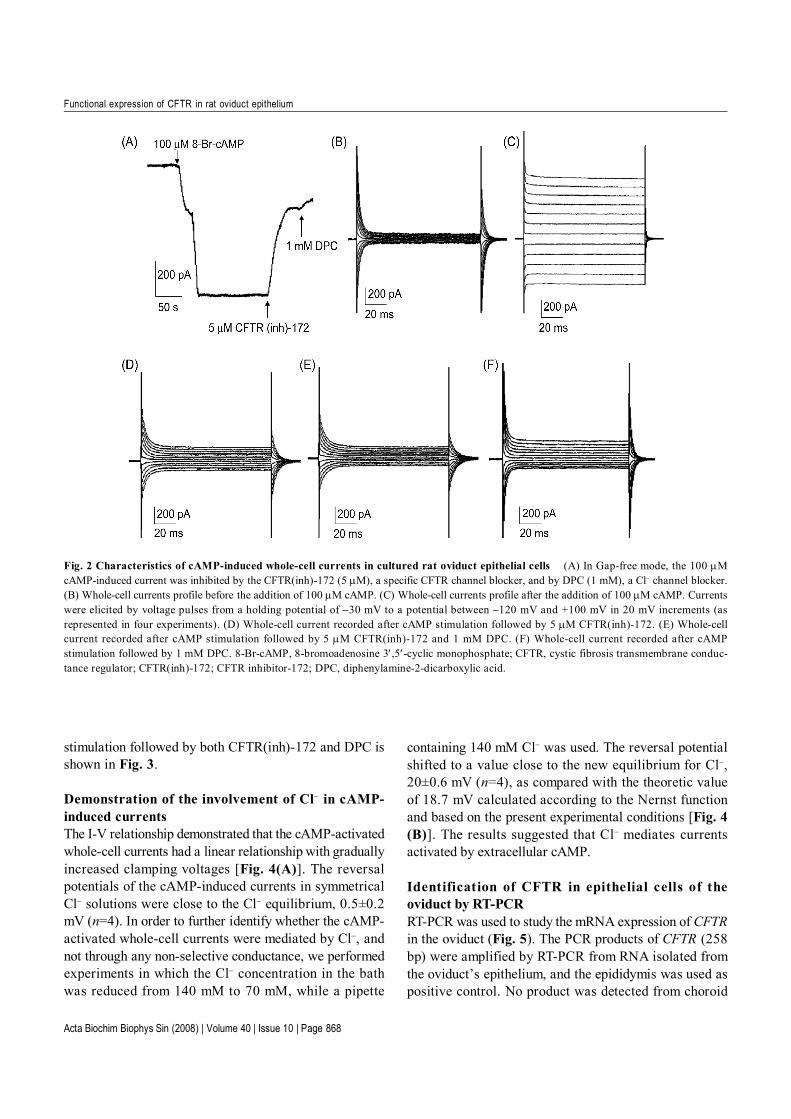
confirmed by immunofluorescence staining of cytokeratin(Fig. 1), to see whether oviduct epithelial cells functionallyexhibit CFTR conductance. The current recordings wereobtained using pipette and bath solutions containingsymmetrical Cl− concentrations (containing intracellular 140mM Cl− and extracellular 140 mM Cl−, respectively). Undera continuous recording with a −70 mV holding potential,bath application of 8-Br-cAMP (100 µM) elicited inwardwhole-cell currents [Fig. 2(A)]. Series steps of voltage-clamp stimulation from −120 mV to +100 mV at 20 mVincrements were applied to the cell before application of8-Br-cAMP [Fig. 2(B)]. The cAMP-activated whole-cellcurrents demonstrated that they were independent of thestimulation time [Fig. 2(C)].
Effect of CFTR(inh)-172 and DPC on the cAMP-stimulated Cl− conductanceThe currents were maximally activated by cAMP and werethen suppressed by subsequent application of 5 µM CFTR(inh)-172, a specific CFTR channel blocker, and 1mMDPC, the Cl− channel blocker [Fig. 2(A)]. The whole-cellmembrane current after cAMP stimulation followed by5 µM CFTR(inh)-172, 5 µM CFTR(inh)-172 and 1 mMDPC, and 1 mM DPC alone, respectively, in response tosquare voltage pulses from a holding potential at −30 mVto a potential between −120 mV and +100 mV in 20 mVincrements [Fig. 2(D–F)]. The mean percent inhibitionsof CFTR(inh)-172 and DPC were 71.5% (n=4) and 75.2%(n=4), respectively. The current-voltage (I-V) relationshipobtained from the cell at rest, cAMP stimulation alone,cAMP stimulation followed by CFTR(inh)-172 or cAMP
Fig. 1 The primary rat oviduct epithelial cells (A) Primary rat oviduct epithelial cells after 3 d of culture (200×). (B) Primary rat oviductepithelial cells confirmed by immunofluorescence staining of cytokeratin. Bar=40 µm.
Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 868
Functional expression of CFTR in rat oviduct epithelium
stimulation followed by both CFTR(inh)-172 and DPC isshown in Fig. 3.
Demonstration of the involvement of Cl− in cAMP-induced currentsThe I-V relationship demonstrated that the cAMP-activatedwhole-cell currents had a linear relationship with graduallyincreased clamping voltages [Fig. 4(A)]. The reversalpotentials of the cAMP-induced currents in symmetricalCl− solutions were close to the Cl− equilibrium, 0.5±0.2mV (n=4). In order to further identify whether the cAMP-activated whole-cell currents were mediated by Cl−, andnot through any non-selective conductance, we performedexperiments in which the Cl− concentration in the bathwas reduced from 140 mM to 70 mM, while a pipette
containing 140 mM Cl− was used. The reversal potentialshifted to a value close to the new equilibrium for Cl−,20±0.6 mV (n=4), as compared with the theoretic valueof 18.7 mV calculated according to the Nernst functionand based on the present experimental conditions [Fig. 4(B)]. The results suggested that Cl− mediates currentsactivated by extracellular cAMP.
Identification of CFTR in epithelial cells of theoviduct by RT-PCRRT-PCR was used to study the mRNA expression of CFTRin the oviduct (Fig. 5). The PCR products of CFTR (258bp) were amplified by RT-PCR from RNA isolated fromthe oviduct’s epithelium, and the epididymis was used aspositive control. No product was detected from choroid
Fig. 2 Characteristics of cAMP-induced whole-cell currents in cultured rat oviduct epithelial cells (A) In Gap-free mode, the 100 µMcAMP-induced current was inhibited by the CFTR(inh)-172 (5 µM), a specific CFTR channel blocker, and by DPC (1 mM), a Cl− channel blocker.(B) Whole-cell currents profile before the addition of 100 µM cAMP. (C) Whole-cell currents profile after the addition of 100 µM cAMP. Currentswere elicited by voltage pulses from a holding potential of −30 mV to a potential between −120 mV and +100 mV in 20 mV increments (asrepresented in four experiments). (D) Whole-cell current recorded after cAMP stimulation followed by 5 µM CFTR(inh)-172. (E) Whole-cellcurrent recorded after cAMP stimulation followed by 5 µM CFTR(inh)-172 and 1 mM DPC. (F) Whole-cell current recorded after cAMPstimulation followed by 1 mM DPC. 8-Br-cAMP, 8-bromoadenosine 3′,5′-cyclic monophosphate; CFTR, cystic fibrosis transmembrane conduc-tance regulator; CFTR(inh)-172; CFTR inhibitor-172; DPC, diphenylamine-2-dicarboxylic acid.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 869
Functional expression of CFTR in rat oviduct epithelium
plexus, which was used as a negative control. GAPDHstandard product was expressed in all the tissues. Theseproducts were confirmed by sequencing (data notshown).
Detection of oviduct CFTR protein expression byWestern blot analysisThe protein expression of CFTR in the oviduct at different
Fig. 3 Current-voltage (I-V) curve Corresponding I-V relationship obtained from the cell at rest (circle), cAMP stimulation alone (filled circle),cAMP stimulation followed by CFTR(inh)-172 (square), and cAMP stimulation followed by CFTR(inh)-172 and DPC (filled square). The recordsrepresent four experiments. 8-Br-cAMP, 8-bromoadenosine 3′,5′-cyclic monophosphate; CFTR, cystic fibrosis transmembrane conductanceregulator; DPC, diphenylamine-2-dicarboxylic acid.
Fig. 4 Demonstration of the involvement of Cl− in cAMP-induced currents Whole-cell recordings obtained 6 min after cAMP stimulationfrom a cell patched with pipette containing 140 mM Cl− and bathed in (A) 140 mM Cl− or (B) 70 mM Cl− with corresponding current-voltagerelationships. Note that reversal potential was shifted from 0 to 20 mV (20±0.6 mV, n=4), close to the new ECl=18.7 mV. The records arerepresentative of four experiments.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 870
Functional expression of CFTR in rat oviduct epithelium
estrus stages was also examined by Western blot analysis.The CFTR antibody raised against a peptide (C)KEETEEEVQDTRL, corresponding to amino acid residues1468−1480 of cytoplasm, the C-terminal part of humanCFTR. The major immunoreactive band of CFTR andβ-actin displayed a molecular mass of about 170 kDa inrat oviduct and 42 kDa in epididymis as positive control[Fig. 6(A,B)]. In addition, β-actin was detected inchoroid plexus as a negative control, but CFTR was notdetected. CFTR protein was detected in the oviductthroughout the estrus cycle and in the expression of estrusand metestrus. However, it was not detected in diestrus,and it was significantly lower in the expression of proestrus[Fig. 6(C)].
Immunolocalization of CFTR in oviductTo investigate the location of CFTR in oviduct, we stainedsections from rat oviduct using immunofluorescence andimmunohistochemistry technique. The apical surface andcytoplasm of the epithelial cells of rat oviduct were highlystained with CFTR antibody [Fig. 7(A), Fig. 8(A)], andthere was no immunoreactivity in negative control [Fig. 7(D), Fig. 8(B)]. Bright field and converged images of CFTRimmunofluorescent stained rat oviduct sections wereshown in [Fig. 7(B)] and [Fig. 7(C)], respectively.
Discussion
The oviduct provides an electrolyte environment for ovumpickup, sperm transport, ovum capacitation, spermcapacitation, ovum fertilization, zygote development andzygote transport [13−15], but the regulation and mechanismof fluid movement across the epithelium remain poorlyunderstood. The present study demonstrated that cAMPstimulated an inward current in cultured rat oviduct epitheliain whole-cell patch clamp. Several lines of evidencesuggested that the cAMP-activated Cl− current in the oviductepithelial cell was characteristic of CFTR [6,16−20]. First,elevating cellular cAMP stimulates whole-cell Cl−-selectiveconductance and exhibited time- and voltage-independentcharacteristics. Second, this conductance has a linear I-Vrelationship. Third, the cAMP-activated Cl− current issuppressed in a voltage-independent manner by 5 µMCFTR(inh)-172, a specific CFTR channel blocker [21,22].In support of these functional data, the detection of CFTRmRNA and protein immunolocalization in rat oviductconfirm earlier reports [8,23], which suggested thepresence of this channel in the oviduct epithelium.
Fig. 5 The mRNA expression of CFTR in the oviduct epitheliumPositive control with rat epididymidis complementary DNA indicatedthe expected size of the amplified fragment (258 bp). Lane 1, choroidplexus CFTR (negative control); lane 2, choroid plexus GAPDH (300bp); lane 3, epididymis CFTR (258 bp); lane 4, epididymis GAPDH (300bp); lane 5, oviduct GAPDH (300 bp); lane 6, oviduct CFTR (258 bp).M, DNA marker. CFTR, cystic fibrosis transmembrane conductanceregulator.
Fig. 6 Western blot analysis of the expression of oviduct CFTR protein throughout the estrus cycle Western blot results of CFTR (170kDa) (A) and β-actin (42 kDa) (B) in rat oviduct. Lane 1, choroid plexus as negative control; lane 2, proestrus; lane 3, estrus; lane 4, metestrus; lane5, diestrus; lane 6, epididymis as positive control. Experiments were repeated three times, and the first experiment was shown. (C) Relativeexpression of oviduct CFTR protein normalized with the internal marker β-actin. Values are expressed as mean±SEM (n=3). **P<0.001 comparedwith proestrus; *P<0.01 compared with proestrus; #P>0.05 compared with proestrus. CFTR, cystic fibrosis transmembrane conductance regulator.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 871
Functional expression of CFTR in rat oviduct epithelium
Fig. 7 Immunofluorescence of CFTR in rat oviduct (A) Fluorescence, (B) bright field and (C) converged images of CFTR immunofluorescent-stained rat oviduct sections are shown. Arrows indicate that immunoreactivity of CFTR was detected in the apical membrane of the epithelial cells.(D) Negative control. CFTR, cystic fibrosis transmembrane conductance regulator; L, tubular lumen. Bar=50 µm.
Fig. 8 Immunohistochemistry of CFTR in rat oviduct (A) Immunoreactivity of CFTR was detected in the apical membrane of the epithelialcells. (B) Control oviduct section with primary antibodies pre-absorbed with CFTR antigen. CFTR, cystic fibrosis transmembrane conductanceregulator; L, tubular lumen. Bar=50 µm.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 10 | Page 872
Functional expression of CFTR in rat oviduct epithelium
As a cAMP-activated Cl− channel, CFTR plays a criticalrole in electrolyte and fluid secretion. When it wasstimulated by intracellular cAMP, the channel opened andallowed Cl− efflux. In addition, the estrus cycle-dependentprotein expression of CFTR in the present study wasconsistent with observed cyclic changes in CFTR mRNAexpression [8]. Moreover, estrogen may function as aphysiological regulator of CFTR in the oviduct [23].Therefore, enhanced expression of CFTR at estrus mayresult in a higher rate of chloride secretion and thus greaterfluid accumulation.
The presence of functional CFTR in rat oviduct epithelialcells revealed in our study should help us better understandits role in oviduct secretion and in the pathophysiology ofcystic fibrosis. Previous reports indicated that females withCF have reduced fertility probably due to thick, densecervical mucus that presents a barrier for sperm penetration[24]. However, CFTR mutant in endometrium and oviduct,which results in abnormal electrolyte secretion [25,26],may be an important cause of infertility in CF female patient.
In summary, this study showed that the cAMP-activatedCl− current in the oviduct epithelium was characteristic ofCFTR, which provided direct physiological evidence forthe expression of CFTR in the rat oviduct epithelium. Thepresent results also suggested the capability of the oviductto secrete chloride ion at different stages of the estruscycle. Cyclic variations in the expression of CFTR likelyalter the composition of oviductal fluid to permit successfulreproductive events at different times. The functionalexpression of CFTR indicates that CFTR may play a rolein modulating fluid transport in the oviduct.
References
1 Buhi WC, Alvarez IM, Kouba AJ. Oviductal regulation of fertiliza-tion and early embryonic development. J Reprod Fertil Suppl 1997,52: 285−300
2 Menezo Y, Guerin P. The mammalian oviduct: biochemistry andphysiology. Eur J Obstet Gynecol Reprod Biol 1997, 73: 99−104
3 Leese HJ. The formation and function of oviduct fluid. J ReprodFertil 1988, 82: 843−856
4 Ameen N, Alexis J, Salas P. Cellular localization of the cystic fibro-sis transmembrane conductance regulator in mouse intestinal tract.Histochem Cell Biol 2000, 114: 69−75
5 Foulkes AG, Harris A. Localization of expression of the cystic fibro-sis gene in human pancreatic development. Pancreas 1993, 8: 3−6
6 Sheppard DN, Welsh MJ. Structure and function of the CFTR chlo-ride channel. Physiol Rev 1999, 79: S23−S45
7 Tizzano EF, O’Brodovich H, Chitayat D, Benichou JC, BuchwaldM. Regional expression of CFTR in developing human respiratorytissues. Am J Respir Cell Mol Biol 1994, 10: 355−362
8 Chan LN, Tsang LL, Rowlands DK, Rochelle LG, Boucher RC, LiuCQ, Chan HC. Distribution and regulation of ENaC subunit andCFTR mRNA expression in murine female reproductive tract. J
Membr Biol 2002, 185: 165−1769 Leung AY, Wong PY, Gabriel SE, Yankaskas JR, Boucher RC. cAMP-
but not Ca(2+)-regulated Cl− conductance in the oviduct is defectivein mouse model of cystic fibrosis. Am J Physiol 1995, 268: C708−C712
1 0 Keating N, Quinlan LR. Effect of basolateral adenosine triphos-phate on chloride secretion by bovine oviductal epithelium. BiolReprod 2008, 78: 1119−1126
1 1 Orihuela PA, Ortiz ME, Croxatto HB. Sperm migration into andthrough the oviduct following artificial insemination at differentstages of the estrous cycle in the rat. Biol Reprod 1999, 60: 908−913
1 2 Chan HC, He Q, Ajonuma LC, Wang XF. Epithelial ion channels inthe regulation of female reproductive tract fluid microenvironment:implications in fertility and infertility. Sheng Li Xue Bao 2007, 59:495−504
1 3 Hunter RH. Vital aspects of fallopian tube physiology in pigs. ReprodDomest Anim 2002, 37: 186−190
1 4 Rodriguez-Martinez H, Tienthai P, Suzuki K, Funahashi H, EkwallH, Johannisson A. Involvement of oviduct in sperm capacitationand oocyte development in pigs. Reprod Suppl 2001, 58: 129−145
1 5 Yee B. The fallopian tube and in vitro fertilization. Clin ObstetGynecol 2006, 49: 34−43
1 6 Anderson MP, Welsh MJ. Calcium and cAMP activate differentchloride channels in the apical membrane of normal and cysticfibrosis epithelia. Proc Natl Acad Sci U S A 1991, 88: 6003−6007
1 7 Cliff WH, Frizzell RA. Separate Cl− conductances activated by cAMPand Ca2+ in Cl−-secreting epithelial cells. Proc Natl Acad Sci U S A1990, 87: 4956−4960
1 8 Huang SJ, Fu WO, Chung YW, Zhou TS, Wong PY. Properties ofcAMP-dependent and Ca(2+)-dependent whole cell Cl− conductancesin rat epididymal cells. Am J Physiol 1993, 264: C794−C802
1 9 Boockfor FR, Morris RA, DeSimone DC, Hunt DM, Walsh KB.Sertoli cell expression of the cystic fibrosis transmembrane con-ductance regulator. Am J Physiol 1998, 274: C922−C930
2 0 Schwiebert EM, Flotte T, Cutting GR, Guggino WB. Both CFTRand outwardly rectifying chloride channels contribute to cAMP-stimulated whole cell chloride currents. Am J Physiol 1994, 266:C1464−C1477
2 1 Ma T, Thiagarajah JR, Yang H, Sonawane ND, Folli C, Galietta LJ,Verkman AS. Thiazolidinone CFTR inhibitor identified by high-throughput screening blocks cholera toxin-induced intestinal fluidsecretion. J Clin Invest 2002, 110: 1651−1658
2 2 Yamamoto S, Ichishima K, Ehara T. Regulation of extracellularUTP-activated Cl− current by P2Y-PLC-PKC signaling and ATPhydrolysis in mouse ventricular myocytes. J Physiol Sci 2007, 57:85−94
2 3 Rochwerger L, Buchwald M. Stimulation of the cystic fibrosis trans-membrane regulator expression by estrogen in vivo. Endocrinology1993, 133: 921−930
2 4 Tizzano EF, Silver MM, Chitayat D, Benichou JC, Buchwald M.Differential cellular expression of cystic fibrosis transmembraneregulator in human reproductive tissues. Clues for the infertility inpatients with cystic fibrosis. Am J Pathol 1994, 144: 906−914
2 5 Chan HC, Shi QX, Zhou CX, Wang XF, Xu WM, Chen WY, ChenAJ et al. Critical role of CFTR in uterine bicarbonate secretion andthe fertilizing capacity of sperm. Mol Cell Endocrinol 2006, 250:106−113
2 6 French PJ, van Doorninck JH, Peters RH, Verbeek E, Ameen NA,Marino CR, de Jonge HR et al. A delta F508 mutation in mousecystic fibrosis transmembrane conductance regulator results in atemperature-sensitive processing defect in vivo. J Clin Invest 1996,98: 1304−1312
Copyright © 2022 FDOKUMEN




















