
Freeze-fracture immunogold labeling
12
Histochem Cell Biol (1996) 106:19-30 9 Springer-Verlag 1996 Maria Rosaria Torrisi 9 Patrizia Mancini Freeze-fracture immunogold labeling Accepted: 2 May 1996 Abstract Several approaches have been developed to combine immunogold cytochemistry and freeze-fracture techniques. These methods are highly heterogeneous re- garding both the sequence of the procedural steps and the aspect of the resulting images. They imply immuno- labeling either before or after freeze-fracture or even im- munolabeling of platinum/carbon replicas of the freeze- fractured membranes, and have been used alternatively or in parallel to address different questions related to cell membrane structure, composition and dynamics or to in- tracellular membrane traffic. This review will briefly de- scribe these methods and report most of their immuno- gold cytochemical applications, with the aim of facilitat- ing selection of the most appropriate approach. Introduction Freeze-fracture technique was designed for the analysis of membrane structure. During freeze-fracture, cell membranes are split along their hydrophobic interior into two complementary halves, the outer or exoplasmic leaf- M.R. Torrisi ( ~ ) 9 R Mancini Dipartimento di Medicina Sperimentale e Patologia Viale Regina Elena, 324 1-00161 Rome, Italy Tel. +39-6-4468450; Fax +39-6-4454820 e-mail trrsmr @flashnet.it Fig. 1 A schematic view of the freeze-fracture process. During fracture, membranes (left) are split along their hydrophobic interior into two complementa- ry leaflets: the exoplasmic out- er leaflet (E/ace) and the pro- toplasmic inner leaflet (P face) (right). Transmembrane pro- teins may partition with the outer or the inner leaflet of the freeze-fractured membranes let (E face) and the inner or protoplasmic leaflet (P face) (Fig. 1). Although lipids and peripheral proteins partition during fracture with the corresponding monolayer, inte- gral and transmembrane proteins may partition with the outer or the inner membrane half. Platinum/carbon (Pt/C) replicas of freeze-fractured membranes provide views of the split membrane hydrophobic interior, where transmembrane proteins are believed to appear as intra- membrane particles. Different methods have been there- fore developed to analyze in a single image not only the ultrastructural features of the freeze-fractured mem- branes but also their composition and dynamics. The use of gold particles in lectin cytochemistry or in immunocy- tochemistry, because of their high electron density, en- abled easy visualization of the labeling on Pt/C replicas, and the efforts towards development of new methods in- creased considerably. These methods can be separated in two groups: (a) techniques in which the immunolabeling is performed before freeze-fracture and replication, including the la- bel-fracture and the fracture-flip techniques, and (b) dif- ferent approaches in which the immunolabeling is per- formed on the replicas of the freeze-fractured mem- branes. Separate consideration is deserved by the frac- ture-label method, in which cells are labeled after freeze- fracture and samples may be observed in conventional thin sections. We will review here the methods in which freeze-fracture has been combined with immunogold la-
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Freeze-fracture immunogold labeling

Histochem Cell Biol (1996) 106:19-30 �9 Springer-Verlag 1996
M a r i a R o s a r i a Torr is i �9 P a t r i z i a M a n c i n i
Freeze-fracture immunogold labeling
Accepted: 2 May 1996
A b s t r a c t Several approaches have been developed to combine immunogold cytochemistry and freeze-fracture techniques. These methods are highly heterogeneous re- garding both the sequence of the procedural steps and the aspect of the resulting images. They imply immuno- labeling either before or after freeze-fracture or even im- munolabeling of platinum/carbon replicas of the freeze- fractured membranes, and have been used alternatively or in parallel to address different questions related to cell membrane structure, composition and dynamics or to in- tracellular membrane traffic. This review will briefly de- scribe these methods and report most of their immuno- gold cytochemical applications, with the aim of facilitat- ing selection of the most appropriate approach.
Introduction
Freeze-fracture technique was designed for the analysis of membrane structure. During freeze-fracture, cell membranes are split along their hydrophobic interior into two complementary halves, the outer or exoplasmic leaf-
M.R. Torrisi (~) �9 R Mancini Dipartimento di Medicina Sperimentale e Patologia Viale Regina Elena, 324 1-00161 Rome, Italy Tel. +39-6-4468450; Fax +39-6-4454820 e-mail trrsmr @flashnet.it
Fig. 1 A schematic view of the freeze-fracture process. During fracture, membranes (left) are split along their hydrophobic interior into two complementa- ry leaflets: the exoplasmic out- er leaflet (E/ace) and the pro- toplasmic inner leaflet (P face) (right). Transmembrane pro- teins may partition with the outer or the inner leaflet of the freeze-fractured membranes
let (E face) and the inner or protoplasmic leaflet (P face) (Fig. 1). Although lipids and peripheral proteins partition during fracture with the corresponding monolayer, inte- gral and transmembrane proteins may partition with the outer or the inner membrane half. Platinum/carbon (Pt/C) replicas of freeze-fractured membranes provide views of the split membrane hydrophobic interior, where transmembrane proteins are believed to appear as intra- membrane particles. Different methods have been there- fore developed to analyze in a single image not only the ultrastructural features of the freeze-fractured mem- branes but also their composition and dynamics. The use of gold particles in lectin cytochemistry or in immunocy- tochemistry, because of their high electron density, en- abled easy visualization of the labeling on Pt/C replicas, and the efforts towards development of new methods in- creased considerably.
These methods can be separated in two groups: (a) techniques in which the immunolabeling is performed before freeze-fracture and replication, including the la- bel-fracture and the fracture-flip techniques, and (b) dif- ferent approaches in which the immunolabeling is per- formed on the replicas of the freeze-fractured mem- branes. Separate consideration is deserved by the frac- ture-label method, in which cells are labeled after freeze- fracture and samples may be observed in conventional thin sections. We will review here the methods in which freeze-fracture has been combined with immunogold la-

20
beling and we will report studies where these methods have been applied to the analysis of membrane composi- tion and dynamics.
Fracture-label
The fracture-label technique permits direct labeling of freeze-fractured plasma and intracellular membranes (Pinto da Silva 1984). Cells or tissues are fixed in glutar- aldehyde, cryoprotected by 30% glycerol and frozen as in conventional freeze-fracture. The frozen samples are fractured in a glass or plastic container filled with liquid nitrogen by crushing with a glass or metal pestle to ob- tain small fragments, or using a scalpel to obtain larger fragments. The freeze-fractured samples are then thawed, gradually deglycerinated and cytochemically or immunocytochemically labeled. Two variants of the method have been developed: thin section fracture-label (Pinto da Silva et al. 1981b) and critical point drying fracture-label (Pinto da Silva et al. 1981a). In the first method, the fracture-labeled small samples are processed for conventional thin section electron microscopy, whereas in the second, larger samples are dried through the critical point and replicated by Pt/C evaporation. Al- though initially both variants were used in combination with lectin cytochemistry and frequently performed in parallel, critical point drying fracture-label has not been widely applied for immunolocalizations, probably be- cause of the difficult interpretation of the fracture face replicas obtained, which appear altered and lacking typi- cal ultrastructural details such as intramembrane parti- cles. In contrast, thin section fracture-label provides full preservation of the cell ultrastructure, similar to that ob- served in conventional thin section images. Identification of the fracture plans and of the intracellular fractured membranes in thin sections is easy and the subsequent interpretation of the results is usually simple.
In fracture-label technique, either isolated cells or tis- sues can be used, in particular for labeling of intracellu- lar components. When the purpose of the experiments re- quires labeling of the freeze-fractured plasma mem- branes, however, cells may be first embedded in a gel matrix of bovine serum albumin (BSA) cross-linked by glutaraldehyde, before freezing and fracturing (Fig. 2B-D). This procedure allows the gel to retain after frac- ture the outer exoplasmic leaflet of the plasma mem- brane and to prevent its loss during thawing (Fig. 2C). The gel matrix also occasionally permits one to maintain in close apposition and to simultaneously label the two complementary exoplasmic and protoplasmic fracture faces from the same plasma membrane (an event which has been called "crack") (Fig. 2B, D). On the other hand, fracture-label of isolated cells allows simultaneous com- parison of the pattern and density of labeling on intracel- lular fractured membranes with those on the unfractured cell surfaces.
The ultrastructural observation of the freeze-fractured membrane leaflets shows interrupted bilayered segments
(Fig. 2B-D), instead of uninterrupted monolayers. This structural feature was interpreted as the result of post- fracture reorganization of the monolayers into bilayered structures occurring during thawing in an aqueous solu- tion. In fact, if the fractured samples are freeze-substitut- ed in osmium/acetone, a procedure which does not allow subsequent labeling, the fractured membrane halves ap- pear as uninterrupted monolayers (Pinto da Silva et al. 1981b). On the other hand, these reorganization events are responsible for labeling on the exoplasmic faces of lectin binding sites or of epitopes originally exposed to the outer surface of the unfractured cells (Fig. 2A) and represent therefore a favorable process in fracture-label technique.
Immunogold thin section fracture-label has been ap- plied mainly to address problems related to (a) the parti- tion during fracture of transmembrane proteins and (b) the intracellular distribution of membrane components.
Partition of transmembrane proteins with the two fractured membrane leaflets may depend on the structur- al properties of the proteins themselves, as initially pro- posed from the fracture-label analysis of the partition on fracture of the two major glycoproteins, glycophorin and band 3, in human erythrocytes (Pinto da Silva and Torrisi 1982). The study showed that glycophorin, which is composed of a large extracellular portion, partitions pref- erentially with the outer exoplasmic face of the freeze- fractured membrane, whereas band 3, which is exposed mainly to the cytoplasmic side of the membrane and is associated with the cytoskeleton, partitions preferentially with the inner protoplasmic face. Subsequent studies on other cell systems, however, revealed that the depen- dence of behavior on fracture on protein structure cannot be considered a general rule and that stable or dynamic interactions of the transmembrane proteins with other membrane components or with membrane-associated el- ements may be determinant in this process. Most of the lymphocyte antigens we studied (CD8, HLA) partitioned during fracture with the exoplasmic face (Fig. 2B, D) (Mancini et al. 1994; Torrisi and Pavan 1993), but CD4, despite its structural similarity to glycophorin, was most- ly associated with the inner protoplasmic membrane half (Pavan et al. 1989a). When CD8 antigens were cross-
Fig. 2A-D Surface immunolabeling (A) and fracture immunola- beling (B, C) of CD8 antigens on human lymphocytes. The immu- nogold labeling of CD8 molecules, performed with anti-CD8 anti- bodies and protein A-colloidal gold, is dense and uniform over the unfractured cell surface (A) of a CD8+ T lymphocyte, whereas platelets and CD8 lymphocytes appear unlabeled. In B, comple- mentary protoplasmic (P) and exoplasmic (E) faces of a fracture- labeled plasma membrane of a CD8 + lymphocyte, where gold par- ticles are confined to the E face and the P face appears unlabeled. E faces from freeze-fractured plasma membranes of CD8 lym- phocytes are unlabeled (C). In D, fracture-immunolabeling of HLA antigens on human lymphocytes: the gold immunolabeling, performed with anti-HLA antibodies and protein A-colloidal gold, is distributed mainly on the E faces. Nu nucleus. A x16000, B x24000, C x33000, D x21000; bars" 1 ]am. B and C reproduced from Mancini et al. (1994); Verlag; D reproduced from Torrisi and Pavan (1993) by copyright permission of John Libbey Eurotext

21

22
linked by incubation with specific antibodies at 37 ~ C to induce patching and capping, a process which is known to induce new interactions of the antigens with the un- derlying cytoskeleton, the partition of CD8 on fracture became protoplasmic (Mancini et al. 1994). In transfec- ted cells, expressing only exogenous CD8 molecules at the cell surface, the capping process was not efficiently induced by cross-linking antibodies and the partition on fracture of CD8 remained exoplasmic, probably as a con- sequence of lack of interaction with the cytoskeleton (Mancini et al. 1994). All these results on fracture-la- beled human lymphocyte plasma membranes seemed to confirm that interactions, more than structural properties, are responsible for partition on fracture with the proto- plasmic faces, and extended the initial observation of heterogeneous expression of transmembrane sialoglyco- proteins on the inner membrane half of freeze-fractured lymphocytes (Torrisi and Pinto da Silva 1982). Consis- tent with the above observations, stable association of the protein dystrophin in skeletal muscle cells with the cytoskeleton appeared responsible for the association of the protein with the protoplasmic face of the fi'acture-la- beled plasma membranes (Squarzoni et al. 1992).
Further support for the hypothesis that dynamic inter- actions can modify the behavior of transmembrane pro- teins on fracture came from a study on epidermal growth factor receptors (EGFRs) (Lotti et al. 1991), which were shown to modify their partition from exoplasmic to pro- toplasmic when activated by the EGF. Since this change in partition was more evident after receptor clustering at 37 ~ C, induced either by the ligand or by anti-receptor antibodies, it may reflect receptor cross-linking and in- teractions among the cytosolic tails.
In infected cells, interaction of the viral envelope pro- teins with the underlying nucleocapsids at the sites of budding was shown to be responsible for their behavior on fracture (Torrisi and Pavan 1995). In cells infected with Sindbis virus, an RNA enveloped virus used as a model system in membrane cell biology, the envelope vi- ral glycoproteins were seen to partition with the exoplas- mic faces of all intracellular membranes during their pro- cessing and transport to the cell surface; they acquired a protoplasmic partition only at the cell plasma membrane, because of interaction with the nucleocapsid during viral budding (Torrisi and Bonatti 1985). If the Sindbis virus proteins were expressed on the cell surface by transfec- tion or following implantation by low-pH-mediated fu- sion, i.e. in the absence of nucleocapsid anchorage, their behavior on fracture was exoplasmic (Torrisi et al. 1990). The role played by nucleocapsid/envelope protein inter- action in determining changes in partition on fracture of the viral transmembrane proteins seems to represent a general rule, followed also when budding occurs at intra- cellular membranes. In cells producing Epstein-Barr vi- rus (EBV), where the envelope of the herpesvirus is ac- quired at the inner nuclear membrane, the major enve- lope glycoprotein was seen to cluster on budding figures and to partition with the protoplasmic face of the freeze- fractured inner nuclear membranes where nucleocapsids
were visible underneath the fractured leaflet (Torrisi et al. 1989b).
The second useful advantage of fracture-label tech- nique is its possible application to the analysis of intra- cellular membrane components, approaching problems related to their distribution, compartmentalization and transport (Torrisi and Pinto da Silva 1984; Torrisi et al. 1989a). The method, in fact, exposes large areas of the intracellular membranes to the immunolabeling, provid- ing full accessibility of the antigens; in the meantime, it preserves well the cellular ultrastructure and permits easy identification of the labeled organelles. In Sindbis virus-infected cells, immunogold fracture label was used to study the intracellular localization of the viral enve- lope glycoproteins, which are synthesized, processed and transported in large amounts to the cells surface, follow- ing the cellular biosynthetic and exocytic pathway. Im- munolabeling with specific antibodies and protein A- gold was found intracellularly on freeze-fractured endo- plasmic reticulum, outer nuclear membranes and Golgi cisternae (Torrisi and Bonatti 1985). The use of fracture- label technique in that study allowed us to observe an in- teresting finding: the presence of viral envelope glyco- proteins on the inner nuclear membranes, a site not sup- posed to be involved in intracellular membrane traffic. With further fracture-label experiments, we were able to demonstrate that the viral glycoproteins may freely dif- fuse to and from the inner nuclear membrane, probably through the nuclear pore complexes (Torrisi et al. 1987).
The possibility, in fracture-label, to expose inner nu- clear membranes to the immunolabeling was helpful in several studies on herpesvirus-infected cells, since these viruses acquire their envelope by budding at the inner nuclear membranes. Results obtained by parallel applica- tion of lectin cytochemistry and immunogold cytochem- istry in fracture-labeled EBV-producing cells (Torrisi et al. 1989b) and herpes simplex type 1 (HSV-1)-infected cells (TolTisi et al. 1992) demonstrated that the two her- pesviruses acquire at the inner nuclear membranes an en- velope similar in composition to that of extracellular vir- ions, but containing immature type of proteins. In addi-
Fig. 3A-F Fracture-label of CD8-transfected epithelial cells (FRT-U10 cells, Mancini et al. 1994), showing examples of CD8 molecules on freeze-fractured intracellular membranes. Positive immunolabeling with anti-CD8 monoclonal antibody and protein A-colloidal gold is found on the exoplasmic faces of the freeze- fractured plasma membranes (A, B, D), and on Golgi cross-frac- tured (E) or freeze-fractured cisternae (F). Protoplasmic (A) and exoplasmic (B) faces of the freeze-fractured inner nuclear mem- branes are unlabeled. Freeze-fractured mitochondrium membranes appear also unlabeled (C). In E, the immunolabeling, penetrated inside the cross fractured cisternae, is dense on the inner surface of the dilatated rims (from E Mancini, M.C. Pascale, S. Bonatti, M.R. Torrisi, unpublished observations). P Protoplasmic face of freeze-fractured plasma membrane, E exoplasmic face of freeze- fractured plasma membrane, INp protoplasmic face of freeze-frac- tured inner nuclear membrane, 1Ne exoplasmic face of freeze-frac- tured inner nuclear membrane, Nu nucleus, M mitochondrium, G Golgi. A x14000, B x20000, C x31000, D x26000, E x37000, F x58000; bars A, B 1 ~tm, C-F 0.5 gm

23
~ il . . . . . . . . . . i ~ ~ ili i i �84184 � 8 4
~' ~i ~ ~'~ i' ~i~ ~' '~i ~i~i ~'~ ~!i,~i ~ ' ~ ~ i ~i~i~i~i~!i~!!~i ~i ~ ~ ~, ~

24
tion, in the herpesvirus-infected cell system, the pathway followed by the viral proteins and the virions for their in- tracellular transport and maturation is still debated. Frac- ture-label experiments demonstrated that, during intracel- lular transport of the viruses, the envelope glycoproteins become glycosylated to intermediate forms and finally to mature forms (Di Lazzaro et al. 1995), implying transport of the virions through the Golgi complex. In Vero cells infected with a mutant HSV-1, deleted in the UL20 pro- tein, the immunogold fracture-label method revealed that the viral glycoproteins are present on the nuclear mem- branes, but are not transported to the cell surface, sug- gesting a role for the UL20 protein in the transport of vi- ral glycoproteins to the cell plasma membranes and in the exit of the virions from the cell (Avitabile et al. 1994).
Fracture-label was also used by another group to study the intracellular distribution of opsin in rod photo- receptor membranes of Xenopus laevis and mouse reti- nas: immunolabeling revealed localization of opsin on rod outer segments, and its transport from rod inner to outer segments, as a mechanism occurring during disc assembly (Defoe and Besharse 1985; Besharse and Wet- zel 1995). An interesting application of fracture-label, al- though not involving the immunolabeling approach, was reported for the localization and quantitation of phospho- lipids in fractured tissues using enzyme-gold cytochem- istry (Coulombe etal. 1988): phospholipase A2, ad- sorbed to colloidal gold, specifically labeled the glyc- erophospholipids on intracellular membranes and per- mitted comparison of the labeling density over different membrane compartments.
In Fig. 3, examples of immunolabeling of CD8 mole- cules on freeze-fractured Fisher rat thyroid epithelial cells (FRT-UI0 cells) expressing by transfection CD8 protein are shown. As reported above (Mancini et al. 1994), the labeling on the freeze-fractured plasma mem- branes was confined to the exoplasmic face. The intra- cellular freeze-fractured membranes were differentially immunolabeled by the anti-CD8 monoclonal antibody used, which recognizes mainly the mature forms of the protein after processing through the Golgi apparatus (R Mancini, M.C. Pascale, S. Bonatti, M.R. Torrisi, unpub- lished results). The gallery of images shown in Fig. 3 points out how, in fracture-label samples, the intracellu- lar ultrastructure is well preserved and provides exam- ples of different intracellular membrane fractured faces and fractured organelles.
Label-fracture
The label-fracture technique represents an ideal method to study at high resolution the distribution of surface molecules, such as antigens or receptors, on the cell plas- ma membranes (Pinto da Silva and Kan 1984). The tech- nique, although limited to the exoplasmic outer leaflet of the freeze-fractured plasma membrane, permits correla- tion, in a single view, of the distribution of the surface la- beled membrane components and that of typical freeze- fracture morphological features such as the intramem-
Fig. 4A-D Label-fracture. A Isolated cells are gold immunola- beled and frozen. B Freeze-fracture splits the cell plasma mem- branes into exoplasmic outer and protoplasmic inner halves. C Replicas of the freeze-fractured membranes are obtained by plati- num (Pt) and carbon (C) evaporation. D Samples are thawed and washed in distilled water. Exoplasmic membrane halves and entire cells with protoplasmic membrane halves remain attached to the Pt/C replicas. Replicas are then picked up on grids, dried and ob- served at TEM. (Modified from Pinto da Silva and Kan 1984)
brane particles (IMPs). The comparison of the distribu- tion of gold immunolabeling and of IMPs and their pos- sible coincidence can be better visualized by experimen- tally inducing redistribution and clustering of either the surface labeled components or of the IMPs. In this tech- nique (Fig. 4), the cells are first immunolabeled with specific antibodies directed against surface exposed anti- gens, followed by protein A-gold or secondary antibody-

gold conjugated. The size of gold particles should be suf- ficiently large to appear clearly visible behind a Pt/C cast (usually particles of at least 10 nm in diameter are used). A good advantage of this method is that the immunola- beling can be performed also on unfixed cells, preserving the best conditions for optimal antigenicity and labeling efficiency; in addition, double or triple labeling proce- dures can also be easily applied. After immunolabeling, cells are fixed in glutaraldehyde, impregnated in glycerol as a cryoprotectant and frozen. Cells are freeze-fractured in a freeze-fracture device and the fracture faces replicat- ed by Pt/C evaporation as in conventional freeze-fracture technique. Instead of being digested with acids or deter- gents, replicas are floated in distilled water, picked up with Formvar-coated grids and observed in a transmis- sion electron microscope. This procedure allows the la- beled outer exoplasmic leaflet of the plasma membrane to remain attached to the replica. Immunolabeling in the resulting image is underneath the fracture face and, therefore, gold particles are not shadowed.
The label-fracture method was initially applied to map lectin binding sites on human erythrocyte and leu- kocyte membranes (Pinto da Silva and Kan 1984) and on boar sperm cell surfaces (Aguas and Pinto da Silva 1984; Pinto da Silva 1987). An interesting approach, combin- ing autoradiography and label-fracture method, was used to localize radioactive-ligand binding to surface recep- tors (Carpentier et al. 1985). Our group has mostly ap- plied the label-fracture technique to analyze the dynam- ics during patching and capping of lymphocyte trans- membrane proteins. We first analyzed the clustering of CD3 and CD4 human lymphocyte antigens (Pavan et al. 1989a) and the possible aggregation of IMPs. Surface immunolabeling was performed using specific monoclo- hal antibodies directed against the lymphocyte antigens, followed by protein A-colloidal gold conjugates. To demonstrate that the transmembrane proteins bearing the antigens correspond to IMPs, we induced a generalized aggregation of IMPs by treatment of unfixed lympho- cytes with 30% glycerol. In these samples, the immuno- labeling was confined to the particle aggregates, showing that the CD4 and CD3 transmembrane proteins corre- spond to IMPs. Patching and capping of the antigens was then induced by treatment with the corresponding anti- bodies at 37 ~ C, but it was not accompanied by a parallel redistribution of IMPs. These observations have been in- terpreted as the consequence of a highly dynamic pro- cess in which migration to the capping area of IMPs cor- responding to the antigens implies exclusion from that area of IMPs corresponding to other membrane proteins not involved in the capping phenomenon. These dynamic events would be responsible for maintaining a structural equilibrium in the plane of the plasma membrane. These results have been confirmed by further applications of the same method to follow the clustering of other lym- phocyte antigens, such as HLA and LFA-1 molecules (Pavan et al. 1989b, 1992a) (Figs. 5, 6). Both the timing and sequence of antigen migration, first to small patches and then to larger patches and to a final single cap, was comparable for all antigens analyzed. The capping pro-
25
Fig. 5A, B Immunogold label-fracture of human lymphocytes. A Distribution of immunolabeled HLA antigens on the cell surface is uniform in pre-fixed cells. Intramembrane particles (IMPs) on the exoplasmic fracture face appear also uniformly distributed. B Patching of HLA antigens, induced by incubation with specific an- tibodies at 37 ~ C for 10 min, shows large clusters of gold particles on the cell surface. IMPs on the exoplasmic face appear uniformly distributed. A x21000, B xl8000; bars' 1 gm. Reproduced from Torrisi and Pavan (1993) by copyright permission of John Libbey Eurotext
cess was more efficient when LFA-1 molecules were linked to the cytoskeleton upon phorbol ester treatment (Pavan et al. 1992a), a finding in favor of the participa- tion of cytoskeletal elements in the regulation of this process. The label-fracture images in all these conditions

26
Fig. 6 Stereo pair views of an immunogold label-fractured human lymphocyte. Capping of LFA-1 molecules is induced by treatment with anti-CD18 monoclonal antibody at 37~ for 20 rain. The gold particles are concentrated into a single cap. IMPs are not con- centrated in the capping area, but appear uniformly distributed on the exoplasmic fracture face. x30000; bar 0.5 gm. Reproduced from Pavan et al. (1992)
on human erythrocytes revealed a clustered distribution of the receptors and permitted quantitation of the number of clusters per cell and the size of the single cluster (Chevalier and Kazatchkine 1989). Successively, the method allowed comparison of the CR1 clusters ob- served on erythrocytes with those present on the surface of resting or stimulated polymorphonuclear leukocytes, and the results were discussed in light of the role played by CR1 in the immune response (Paccaud et al. 1990). Finally, a more recent application of immunogold label- fracture to locate surface receptors is described in two studies on GABAA receptors. In the first, the distribution of the receptors in various neuronal cell types and in cells transfected with different receptor subunits was compared (Caruncho et al. 1993); in the second, double immunolabeling using gold particles of different sizes was used to analyze the subunit composition of the re- ceptor (Caruncho and Costa 1994).
A variation of the label-fracture method has been in- troduced for the analysis of the ultrastructure and com- position of isolated lens fiber junctions (Zampighi et al. 1989). Junctions were immunolabeled, adhered on a glass surface and covered in sandwich with a copper hat before freeze-fracture and replication.
confirmed that uniform distribution of IMPs obtained was a result of re-equilibrium among structural compo- nents of the plasma membranes.
A different situation was found when the label-frac- ture technique was used in Sindbis virus-infected cells to study the dynamics of viral envelope glycoproteins dur- ing the budding of the virus at the cell surface (Pavan et al. 1992b). In fact, IMPs appeared concentrated at the budding sites, and the particle aggregates corresponded to the clustering of the viral glycoproteins, as shown by specific gold immunolabel with antibodies to the enve- lope proteins. This study provided an additional example of the potentiality offered by label-fracture to relate the distribution of membrane components with ultrastructur- al features of the plasma membranes. This approach was also used for another viral system, in a study on measles virus-infected cells (Rutter and Hohenberg 1993), where label fracture was applied to isolated plasma membranes, instead of intact cells, to extend the observation on the protoplasmic fracture faces and to compare their ultra- structure with immunolabeling of viral proteins at the in- ner cytoplasmic surface.
Several cell surface receptors have also been studied by the label fracture technique. The method was used on cell monolayers (Boonstra et al. 1985) to detect gold-im- munolabeled EGFRs on the cell surface of a human epi- thelial cell line: comparison of the results with those ob- tained in parallel with the surface-replication method in- dicates that label-fracture is the ideal technique for quan- titative analysis of the planar distribution of the receptors on the cell surfaces. Later, the same approach was used to follow the redistribution of EGFRs induced by the li- gand binding (Rijken et al. 1993). Analysis by label-frac- ture of the localization of complement receptors (CR1)
Fracture-flip
The fracture-flip method, although requiring a freeze- fracture procedure, permits observation of the real sur- face of the cell, which becomes visible by inversion (flipping) of the exoplasmic fracture face (Fig. 7). In fact, fracture-flip can be considered a variant of the la- bel-fracture approach, since the initial steps are identical to those of label-fracture preparations. At variance with label-fracture, it requires after freeze-fracture coating of the exoplasmic face only with carbon. The replicas are then floated in distilled water as for label-fracture, picked up from the top with a formvar-coated grid, in- verted (flipped) and shadowed at room temperature by platinum evaporation. The resulting replica provides high-resolution views of the cell surface ultrastructure, revealing details such as surface particles, subparticles and rugosities (Andersson-Forsman and Pinto da Silva 1988a). If cells are immunolabeled, the gold particles are on the replicated cell surface and appear therefore shad- owed.
We first applied immunogold fracture-flip to analyze the dynamics of surface particles during patching/cap- ping of lymphocyte antigens (CD4 and HLA molecules) (Pavan et al. 1990), as an extension of our previous stud- ies on capping (Fig. 8). Sequential clustering of the anti- gens obtained by incubation with specific antibodies at 37 ~ C was performed similarity.to the experiments for la- bel-fracture described above. Since gold immunolabeling obscures the ultrastructural details of the bound antigens, we compared cells treated with the antibodies followed, or not, by protein A-gold labeling. In the resulting imag- es, either from gold-labeled or unlabeled cells, the patch- ing and capping areas appeared as platforms different in
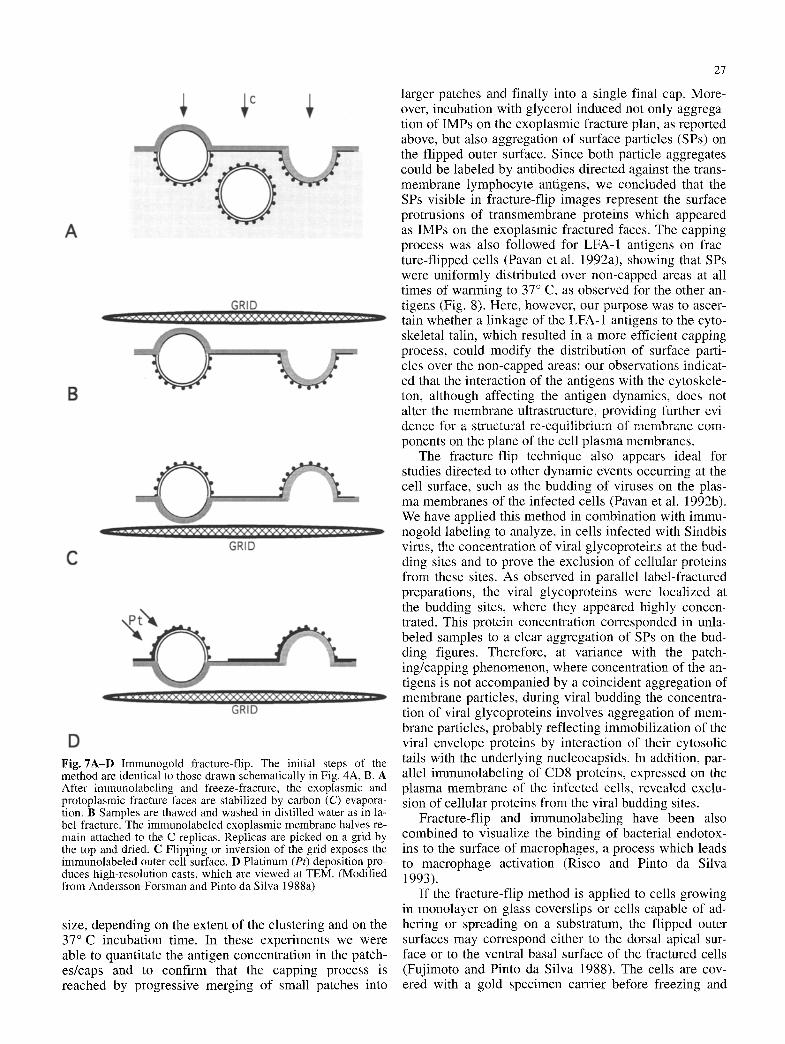
Fig. 7A-D Immunogold fracture-flip. The initial steps of the method are identical to those drawn schematically in Fig. 4A, B. A After immunolabeling and freeze-fracture, the exoplasmic and protoplasmic fracture faces are stabilized by carbon (C) evapora- tion. B Samples are thawed and washed in distilled water as in la- bel-fracture. The immunolabeled exoplasmic membrane halves re- main attached to the C replicas. Replicas are picked on a grid by the top and dried. C Flipping or inversion of the grid exposes the immunolabeled outer cell surface. D Platinum (Pt) deposition pro- duces high-resolution casts, which are viewed at TEM. (Modified from Andersson-Forsman and Pinto da Silva 1988a)
size, depending on the extent of the clustering and on the 37~ incubation time. In these experiments we were able to quantitate the antigen concentration in the patch- es/caps and to confirm that the capping process is reached by progressive merging of small patches into
27
larger patches and finally into a single final cap. More- over, incubation with glycerol induced not only aggrega- tion of IMPs on the exoplasmic fracture plan, as reported above, but also aggregation of surface particles (SPs) on the flipped outer surface. Since both particle aggregates could be labeled by antibodies directed against the trans- membrane lymphocyte antigens, we concluded that the SPs visible in fracture-flip images represent the surface protrusions of transmembrane proteins which appeared as IMPs on the exoplasmic fractured faces. The capping process was also followed for LFA-1 antigens on frac- ture-flipped cells (Pavan et al. 1992a), showing that SPs were uniformly distributed over non-capped areas at all times of warming to 37 ~ C, as observed for the other an- tigens (Fig. 8). Here, however, our purpose was to ascer- tain whether a linkage of the LFA-1 antigens to the cyto- skeletal talin, which resulted in a more efficient capping process, could modify the distribution of surface parti- cles over the non-capped areas: our observations indicat- ed that the interaction of the antigens with the cytoskele- ton, although affecting the antigen dynamics, does not alter the membrane ultrastructure, providing further evi- dence for a structural re-equilibrium of membrane com- ponents on the plane of the cell plasma membranes.
The fracture-flip technique also appears ideal for studies directed to other dynamic events occurring at the cell surface, such as the budding of viruses on the plas- ma membranes of the infected cells (Pavan et al. 1992b). We have applied this method in combination with immu- nogold labeling to analyze, in cells infected with Sindbis virus, the concentration of viral glycoproteins at the bud- ding sites and to prove the exclusion of cellular proteins from these sites. As observed in parallel label-fractured preparations, the viral glycoproteins were localized at the budding sites, where they appeared highly concen- trated. This protein concentration corresponded in unla- beled samples to a clear aggregation of SPs on the bud- ding figures. Therefore, at variance with the patch- ing/capping phenomenon, where concentration of the an- tigens is not accompanied by a coincident aggregation of membrane particles, during viral budding the concentra- tion of viral glycoproteins involves aggregation of mem- brane particles, probably reflecting immobilization of the viral envelope proteins by interaction of their cytosolic tails with the underlying nucleocapsids. In addition, par- allel immunolabeling of CD8 proteins, expressed on the plasma membrane of the infected cells, revealed exclu- sion of cellular proteins from the viral budding sites.
Fracture-flip and immunolabeling have been also combined to visualize the binding of bacterial endotox- ins to the surface of macrophages, a process which leads to macrophage activation (Risco and Pinto da Silva 1993).
If the fracture-flip method is applied to cells growing in monolayer on glass coverslips or cells capable of ad- hering or spreading on a substratum, the flipped outer surfaces may correspond either to the dorsal apical sur- face or to the ventral basal surface of the fractured cells (Fujimoto and Pinto da Silva 1988). The cells are cov- ered with a gold specimen carrier before freezing and

28
Fig. 8A-C Stereo views ofim- munogold fracture-flipped hu- man lymphocytes. A Distribu- tion of LFA-I molecules on pre-fixed cells, immunolabeled with anti-CD 18 monoclonal an- tibody and protein A-colloidal gold. The gold particles appear uniformly distributed on the cell surface. Ultrastructural de- tails such as surface particles of the plasma membranes are clearly visible. B, C Capping of HLA antigens, induced by in- cubation with anti-HLA I class antibodies at 37 ~ C for 20 rain, reveals high concentration of the gold particles in a single cap. The distribution of surface particles appears unaltered. Note that in fracture-flip imag- es the immunogold particles are shadowed, unlike those ob- served in label-fracture prepa- rations. A x13000; B x17000; C x22000; bars 1 gin. A Re- produced from Pavan et al. (1992); B reproduced from Pa- van et al. (1990) by copyright permission of The Company of Biologists Limited; C repro- duced from Torrisi and Pavan (1993) by copyright permission of John Libbey Eurotext

fracturing. This method has been used to observe the for- mation of coated pits on the adherent surface of spread- ing macrophages (Fujimoto and Pinto da Silva 1988), but it can be generally applied to compare surface domains of polarized cells ultrastructurally and immunocyto- chemically at high resolution.
29
Acknowledgements We thank Dr Angelo Del Nero and Giuseppe Lucania for the excellent graphic and photographic work and edi- torial assistance. The funding support of grants from AIRC and CNR (PF ACRO), Italy, is acknowledged.
References
Freeze-fracture replica immunolabeling
This heterogeneous group of methods has in common the ability 1o label, after freeze-fracture, the fracture face replicas. However, it is important to point out that, dur- ing fracture, t ransmembrane proteins partition with the two leaflets and, therefore, the resulting fracture faces do not contain the entire set of proteins present on the un- fractured inner or outer surfaces. Therefore, labeling af- ter fracture does not warrant, as a general rule, staining of the designated membrane component.
The first technique of this group was proposed as a variant of label-fracture, based on the assumption that in label-fracture preparations the exoplasmic faces remain undigested and attached to the replica and that therefore the outer surface of the plasma membrane is exposed for possible immunolabeling (Andersson-Forsman and Pinto da Silva 1988b). Briefly, the replicas of the exoplasmic faces, after washing in distilled water, are picked up with a grid and labeled onto drops of the immunocytochemi- cal reagents, like for staining of thin sections. Due to non-specific adsorption of colloidal gold to the Pt/C cast, the resulting images show a very high density of gold particles between the exoplasinic faces. Specificity of the immunolabeling must therefore be judged carefully, comparing the label on the cell surfaces in experiment and control samples.
The method of immunolabeling freeze-fracture repli- cas has been used to localize MP70 lens membrane pro- tein on the junctional plaques of lens fibers (Gruijters et al. 1987), where the method could be easily applied be- cause of the thinness of this type of samples. Recently a new approach has been proposed for the simultaneous analysis of structural organization and composition of in- tercellular junctions (Fujimoto 1995): tissue slices are freeze-fractured, replicated and digested in sodium dode- cylsulfate (SDS) before immunolabeling. Interestingly, SDS treatment dissolves all unfractured cell components but it cannot digest the fractured faces stabilized by the Pt/C cast; this makes it possible to label not only outer cell surfaces behind the exoplasmic faces but also cyto- plasmic surfaces behind the protoplasmic faces. The method can also be applied to isolated cells, but it may be limited by the partition during fracture of integral membrane proteins, as discussed above.
In conclusion, the capacity to immunolabel intracellu- lar and/or cell surface components specifically and effi- ciently renders freeze-fracture a powerful approach with general application in electron microscopy.
Aguas AP, Pinto da Silva P (1984) High density of transmembrane glycoproteins on the flagellar surface of boar sperm cells. J Cell Biol 99:655-660
Andersson-Forsman C, Pinto da Silva P (1988a) Fracture-flip: new high-resolution images of cell surfaces after carbon stabiliza- tion of freeze-fractured membranes. J Cell Sci 90:531-541
Andersson-Forsman C, Pinto da Silva P (1988b) Label-fracture of cell surfaces by replica staining. J Histochem Cytochem 36: 1413-1418
Avitabile E, Ward PL, Di Lazzaro C, Torrisi MR, Roizman B, Campadelli-Fiume G (1994) The herpes simplex virus UL20 protein compensates for the differential disruption of exocyto- sis of virions and viral membrane glycoproteins associated with the fragmentation of the Golgi apparatus. J Virol 68: 7397-7405
Besharse JC, Wetzel MG (1995) Immunocytochemical localiza- tion of opsin in rod photoreceptors during periods of rapid disc assembly. J Neurocytol 24:371-388
Boonstra J, Belzen N van, Maurik P van, Hage WJ, Blok FJ, Wie- gant FAC, Verkleij AJ (1985) Immunocytochemical demon- strations of cytoplasmic and cell-surface EGF-receptors in A431 cells using cryo-ultramicrotomy, surface replication, freeze etching and label fracture. J Microsc 140:119-129
Carpentier JL, Brown B, Iacopetta B, Orci L (1985) Detection of surface bound ligands by freeze-fracture autoradiography. J Cell Biol 101:887-890
Caruncho HJ, Costa E (1994) Double-immunolabelling analysis of GABAA receptor subunits in label-fracture replicas of cul- tured rat cerebellar granule cells. Receptors Channels 2:143- 153
Caruncho HJ, Puia G, Slobodyansky E, Pinto da Silva P, Costa E (1993) Freeze-fracture immunocytochemical study of the ex- pression of native and recombinant GABAA receptors. Brain Res 603:234-242
Chevalier J, Kazatchkine MD (1989) Distribution in clusters of complement receptor type one (CR 1) on human erythrocytes. J Immunol 142:2031-2036
Coulombe PA, Kan FW, Bendayan M (1988) Introduction of a high resolution cytochemical method for studying the distribu- tion of phospholipids in biological tissues. Eur J Cell Biol 46:564-576
Defoe DM, Besharse JC (1985) Membrane assembly in retinal photoreceptors. II. Immunocytochemical analysis of freeze- fractured rod photoreceptor membrane using anti-opsin anti- bodies. J Neurosci 5:1023-1034
Di Lazzaro C, Campadelli-Fiume G, Torrisi MR (1995) Intermedi- ate forms of glycoconjugates are present in the envelope of herpes simplex virions during their transport along the exocy- tic pathway. Virology 214:619-623
Fujimoto K (1995) Freeze-fracture replica electron microscopy combined with SDS digestion for cytochemical labeling of in- tegral membrane proteins. J Cell Sci I08:3443-3449
Fujimoto K, Pinto da Silva P (1988) Macromolecular dynamics of the cell surface during the formation of coated pits is revealed by fracture-flip. J Cell Sci 91:161-173
Gruijters WTM, Kistler J, Bullivant S, Goodenough DA (1987) Immunolocalization of MP70 in lens fiber 16-17 nm intercel- lular junctions. J Cell Biol 104:565-572
Lotti LV, Pavan A, Zompetta C, Mancini P, Faggioni A, Frati L, Torrisi MR (1991) Partition of Epidermal Growth Factor re- ceptors on freeze-fractured Plasma membranes of A431 cells is affected by the ligand. J Histochem Cytochem 39:1227- 1235 '

30
Mancini P, Lotti LV, Pascale MC, Bonatti S, Torrisi MR (1994) Surface distribution and partition during freeze-fracture of CD8 antigens on human lymphocytes and on epithelial trans- fected cells. Histochemistry 102:51-57
Paccaud JR Carpentier JL, Schifferli JA (1990) Difference in the clustering of complement receptor type 1 (CR1) on polymor- phonuclear leukocytes and erythrocytes: effect on immune ad- herence. Eur J Immunol 20:283-289
Pavan A, Mancini P, Frati L, Torrisi MR, Pinto da Silva P (1989a) Molecular cytochemistry of CD3 and CD4 antigens in human lymphocytes as studied by label-fracture and fracture-label. Biochim Biophys Acta 978:158-168
Pavan A, Mancini P, Cirone M, Frati L, Torrisi MR, Pinto da Silva P (1989b) Capping of HLA antigens in human lymphocytes as followed by immunogold label-fracture. J Histochem Cyto- chem 37:1489-1496
Pavan A, Mancini P, Lucania G, Frati L, Torrisi MR, Pinto da Si- lva P (1990) High resolution surface views of human lympho- cytes during capping of CD4 and HLA antigens as revealed by immunogold fracture-flip. J Cell Sci 96:151-157
Pavan A, Lucania G, Sansolini T, Frati L, Torrisi MR (1992a) Patching and capping of LFA-1 molecules on human lympho- cytes. Histochemistry 98:253-258
Pavan A, Covelli E, Pascale MC, Lucania G, Bonatti S, Pinto da Silva P, Torrisi MR (1992b) Dynamics of transmembrane gly- coproteins during Sindbis virus budding. J Cell Sci 102:149- 155
Pinto da Silva P (1984) Freeze-fracture cytochemistry. In: Polak JM, Varndell IM (eds) Immunolabelling for electron microsco- py. Elsevier, New York, pp 179-188
Pinto da Silva P (1987) Molecular cytochemistry on freeze-frac- tured cells: freeze-etching, fracture-label, fracture-permeation, and label-fracture. In Miller KR (ed) Advances in cell biology, vol 1. Jai Press, Greenwich, Conn, pp 157-190
Pinto da Silva R Kan FW (1984) Label-fracture: a method for high resolution labeling of cell surfaces. J Cell Biol 99:1156-1161
Pinto da Silva P, Torrisi MR (1982) Freeze-fracture cytochemistry: partition of glycophorin in freeze-fractured human erythrocyte membranes. J Cell Biol 93:463-469
Pinto da Silva P, Kachar B, Torrisi MR, Brown C, Parkison C (1981a) Freeze-fracture cytochemistry: replicas of critical point dried cells and tissues after "fracture-label". Science 213:230-233
Pinto da Silva R Parkison C, Dwyer N (1981b) Cytochemistry of freeze-fracture faces in the erythrocyte membrane. Proc Natl Acad Sci USA 78:343-347
Rijken PJ, Groot RP de, Belzen N van, Laat SW de, Boonstra J, Verkleij AJ (1993) Inhibition of EGF-induced signal transduc- tion by microgravity is indipendent of EGF receptor redistri- bution in the plasma membrane of human A431 cells. Exp Cell Res 204:373-377
Risco C, Pinto da Silva P (1993) Binding of bacterial endotoxins to the macrophage surface: visualization by fracture-flip and immunocytochemistry. J Histochem Cytochem 41:601-608
Rutter G, Hohenberg H (1993) Label-fracture of plasma mem- branes isolated from measles virus-infected cells. J Histochem Cytochem 41:1085-1091
Squarzoni S, Sabatelli R Maltarello MC, Cataldi A, Primio R di, Maraldi NM (1992) Localization of dystrophin COOH-termi- nal domain by the fracture-label technique. J Cell Biol 118:1401-1409
Torrisi MR, Bonatti S (1985) Immunocytochemical study of the partition and distribution of Sindbis virus glycoproteins in freeze-fractured membranes of infected baby hamster kidney cells. J Cell Biol 101:1300-1306
Torrisi MR, Pavan A (1993) Freeze-fracture immunocytochemical approaches for the analysis of membrane structure and dynam- ics. Eur J Dermatol 3:342-346
Torrisi MR, Pavan A (1995) Viral infection studied by freeze-frac- ture cytochemistry. In: Severs NJ, Shotton DM (eds) Rapid freezing, freeze fracture and deep etching. Wiley-Liss, New York, pp 347-359
Torrisi MR, Pinto da Silva P (1982) T-lymphocyte heterogeneity: wheat germ agglutinin labeling of transmembrane glycopro- teins. Proc Natl Acad Sci USA 79:5671-5674
Torrisi MR, Pinto da Silva P (1984) Compartmentalization of in- tracellular membrane glycocomponents is revealed by frac- ture-label. J Cell Biol 98:29-34
Torrisi MR, Lotti LV, Pavan A, Migliaccio G, Bonatti S (1987) Free diffusion to and from the inner nuclear membrane of newly synthesized plasma membrane glycoproteins. J Cell Biol 104:733-737
Torrisi MR, Mancini R Pinto da Silva P (1989a) Cytochemical ac- cess to plasma and intracellular membranes of freeze-fractured hepatocytes and salivary gland cells. In: Motta PM and Riva A (eds) Ultrastructure of the extraparietal glands of the digestive tract. Kluwer, Dordrecht, 177-182
Torrisi MR, Cirone M, Pavan A, Zompetta C, Barile G, Frati L, Faggioni A (1989b) Localization of Epstein-Barr virus enve- lope glycoproteins on the inner nuclear membrane of virus producing cells. J Virol 63:828-832
Torrisi MR, Pavan A, Lotti LV, Migliaccio G, Pascale MC, Covelli E, Leone A, Bonatti S (1990) Anchorage-dependent surface distribution and partition during freeze-fracture of viral trans- membrane glycoproteins. J Histochem Cytochem 38:1421- 1426
Torrisi MR, Di Lazzaro C, Pavan A, Pereira L, Campadelli-Fiume G (1992) Herpes simplex virus envelopment and maturation studied by fracture-label. J Virol 66:554-561
Zampighi GA, Hall JE, Ehring GR, Simon SA (1989) The struc- tural organization and protein composition of lens fiber junc- tions. J Cell Biol 108:2255-2275




















