
Low-dose ionizing radiation effects on differentiation of HL60 cells
Upload
independentCategory
view
1download
0
The FASEB Journal express article 10.1096/fj.03-0382fje. Published online November 3, 2003. Folate deficiency and ionizing radiation cause DNA breaks in primary human lymphocytes: a comparison Chantal Courtemanche, Arnold C. Huang, Ilan Elson-Schwab, Nicole Kerry, Bernice Y. Ng, and Bruce N. Ames1 Department of Molecular and Cell Biology, University of California at Berkeley, Berkeley, California, and Children�s Hospital Oakland Research Institute, Oakland, California 1Corresponding author: Bruce N. Ames, Children�s Hospital Oakland Research Institute, 5700 Martin Luther King Jr. Way, Oakland, California 94609. E-mail: [email protected] ABSTRACT DNA double-strand breaks, the most serious DNA lesion caused by ionizing radiation, are also caused by several vitamin or mineral deficiencies, such as for folate. Primary human lymphocytes were either irradiated or cultured at different levels of folate deficiency to assess cell proliferation, apoptosis, cell cycle, DNA breaks, and changes in gene expression. Both radiation and folate deficiency decreased cell proliferation and induced DNA breaks, apoptosis, and cell cycle arrest. Levels of folate deficiency commonly found resulted in effects similar to those caused by 1 Gy of radiation, a relatively high dose. Though both radiation and folate deficiency caused DNA breaks, they affected the expression of different genes. Radiation activated excision and DNA double-strand break repair genes and repressed mitochondrially encoded genes. Folate deficiency activated base and nucleotide excision repair genes and repressed folate-related genes. No DNA double-strand break repair gene was activated by folate deficiency. These findings suggest that a diet poor in folate may pose a risk of DNA damage comparable to that of a relatively high dose of radiation. Our results also suggest that research on biological effects of low-dose radiation should take into account the nutritional status of the subjects, because folate deficiency could confound the effects of low-dose radiation.
Key words: DNA damage � nutrition � ionizing radiation � microarray � cancer risk
NA damage can result from exposure to both external and endogenous sources of stress (1). Though a comparison of DNA damage and cancer risk from different stresses is difficult when different kinds of DNA damage are produced, comparison can be made if
a common endpoint is used such as the induction of DNA double-strand breaks (DSB). Both ionizing radiation and cellular deficiencies of vitamins and minerals can cause DSB, which is a primary cause of chromosomal aberrations and cancer (2�4). Although radiation and vitamin deficiency both cause DSB, a different cellular response and a different level of DNA damage may ensue.
Cellular response to radiation has been extensively studied. Linear energy transfer (LET) radiation produces an estimated 25�40 DSB per cell per Gy, following acute exposure (5). Radiation also produces reactive oxygen species, base lesions, and DNA single-strand breaks
D

(SSB), which can be converted to DSB when damages are clustered (6). Ionizing radiation induces arrests in the G1, S, and G2 phases of the cell cycle; the ataxia telangiectasia mutated protein (ATM) appears to be involved in the initiation of these three checkpoint pathways (7). Recently, microarrays have been used to measure global gene expression changes after irradiation, allowing the identification of new genes implicated in the ensuing response (8�12).
Relatively high doses of radiation (between 0.5 and 10 Gy) are usually used in studies measuring the effects of ionizing radiation, mainly because acute biological effects are not easily detected following exposure to low dose of radiation. Typical population exposure, however, from occupational or background radiation (~1×10−4 Gy per year, resulting in 1×10−5 DSB per cell per day) falls in the low-dose range (13). The risks of low dose exposure, such as DNA damage and cancer, have historically been linearly extrapolated from high dose studies. Whether is it meaningful to do this has been debated. New models have been proposed in which DNA damage occurs only above a certain threshold of radiation (14�16). In addition, low doses of radiation have been shown to induce an adaptive response, reducing damage induced by a subsequent irradiation (17�19). In theory, the linear model ensures protection of the populace against hypothetical hazards, although its use could divert resources from more important risks (20).
It seems likely that the amount of DNA damage resulting from background radiation is much less than that occurring from endogenous oxidative metabolism, which results in ~1 × 10−1 DSB per cell per day (21, 22). Nutritional deficiencies in vitamins, minerals, and antioxidants, which are common in the population, could also contribute significantly to DNA damage and cancer risks (4, 23�25).
Folate is an essential vitamin that is involved in one-carbon transfer reactions and in the formation of purines and pyrimidines for DNA and RNA synthesis (26). A deficiency in folate has been linked to increased risk for several types of cancer (4, 27). A folate concentration of 12 nM is in the range found in plasma of individuals having a low consumption of fruits and vegetables (28, 29). Lower plasma folate (from 5 to 12 nM) has also been measured in folate-deficient individuals (30, 31). Folate levels of individuals having a better diet are ~30 nM, although it may not be the optimal folate level, as folic acid supplementation can raise plasma folate to 50 nM (29). Before the recent folate fortification of grains, the percentage of the U. S. population that had a very low intake (<50% of the recommended daily allowance [400 µg]) of folate was 50% for women and 25% for men (4, 32).
The most likely mechanism for folate-related enhancement of carcinogenesis is the misincorporation of uracil during DNA synthesis. Folate is required for the synthesis of deoxythymidylate (dTMP) from deoxyuridylate (dUMP) and hence for maintaining a proper ratio of dTMP/dUMP (23). Removal of uracil by uracil-DNA glycosylase causes a transient SSB; two nearby opposing lesions, such as a uracil opposite an oxidized base, can cause a DSB. Oxidized bases in DNA are common (33�35). Folate-deficient individuals have millions of uracils misincorporated per cell, as well as increased chromosomal breakage (23, 28, 36, 37). Vitamin B12 deficiency, which is common in the population, also causes uracil misincorporation into DNA and DSB by the same mechanism as folate deficiency (4, 38). In addition, DNA excision repair and mismatch repair may be impaired in folate-deficient cells (39�41). Excessive uracil misincorporation and defective DNA repair may explain why folate-deficient cells are more sensitive to the DNA-damaging effects of a second external stress (38, 42, 43).

In the present study, we compared the effects of folate deficiency on DNA damage and the cellular response to such damage with the effects and cellular response caused by ionizing radiation. We have measured DSB formation, apoptosis, cell cycle variation, and gene expression changes after irradiation and during folate deficiency, using normal human T lymphocytes in primary culture. Our results suggest that physiological levels of folate deficiency cause as much DNA damage as relatively high levels of ionizing radiation, but with a different cellular response.
MATERIALS AND METHODS
Cell culture and irradiation
Normal human T lymphocytes were obtained from AllCells (Berkeley, CA) and cultured in folate-free RPMI 1640 medium (Invitrogen, Carlsbad, CA) supplemented with 10% dialyzed FBS (Invitrogen) and antibiotics. Folate (pteroyl-L-glutamic acid, Sigma, St. Louis, MO) was added to the folate-free medium to a final concentration of 0, 6, 12, 30, or 3000 nM folate. Phytohemagglutinin (PHA) (22.5 µg/ml, Sigma) and interleukin-2 (IL2) (10,000 U/ml, Sigma) were added on the first day of culture to ensure proliferation of T lymphocytes. Every 3 days, the cells were spun down and resuspended at 0.5 × 106 cells/ml in fresh medium supplemented with IL2, for a total of 10 days of culture. For radiation, lymphocytes were cultured in 3000 nM folate-containing medium for a week and then irradiated with γ-rays from a 135Cs source (Shepherd) at a dose rate of 0.25 Gy/min, for a total dose of 0, 0.1, 0.25, 1, or 5 Gy.
Proliferation and cell cycle analysis
Proliferation kinetics were measured at 24-h intervals after irradiation or during folate deficiency by counting cell density with a Coulter counter, and cell viability was assessed with Trypan blue exclusion on a hemacytometer. For cell cycle analysis, cells were harvested at 24-h intervals after irradiation or during folate deficiency. Cells (0.5 × 106) were washed with PBS, permeabilized in 0.3% saponin, treated with 100 µg/ml RNase A, and stained with 10 µg/ml propidium iodide. After 20 min incubation at room temperature, the distribution of DNA content was measured using a FACSort flow cytometer (Becton Dickinson, San Jose, CA). The percentage of cells in the G1, S, or G2/M phases of the cell cycle was determined using the ModFit software (Becton Dickinson).
Detection of apoptosis
Apoptosis was measured using the annexin-V binding assay (Roche Molecular Biochemicals, Indianapolis, IN) that allows distinction between viable, apoptotic, and necrotic cells. Cells (0.5×106) were harvested 24 h after irradiation or at day 8 of folate-deficient culture. Cells were washed in PBS and incubated in a binding buffer containing annexin-V-fluorescein and propidium iodide for 15 min at room temperature before analysis using a FACSort flow cytometer.

Single cell electrophoresis
Single cell electrophoresis (Comet assay) was performed in alkaline condition to evaluate SSB and DSB. Cells were embedded in agarose on Comet slides (Trevigen, Gaithersburg, MD) before irradiation and kept at 4°C to stop DNA repair, following irradiation. The unirradiated, folate-deficient cells were harvested on day 9 of culture. Cells were washed in PBS and resuspended in PBS and low melting point agarose at a final concentration of 0.6% agarose, and 100,000 cells/ml. Cell suspension was pipetted onto a Comet slide and lysed at 4°C in the dark for 1 h in Trevigen lysing solution. DNA was allowed to unwind for 1 h at room temperature in alkaline buffer (0.3 M NaOH, 1 mM EDTA) and subjected to electrophoresis in TBE buffer (90 mM Tris base, 90 mM boric acid, 2 mM EDTA) for 30 min at 25 V. The slides were removed and stained with SYBR Green (Molecular Probes, Eugene, OR), and comet tails from 50 cells were blindly scored on a scale of 0 (low damage) to 4 (high damage) using a fluorescent microscope.
Statistics
Each experiment was repeated three times using lymphocytes from a different donor. Mean values and SE were calculated. Linear regression and ANOVA were used to compare data within and between datasets. P < 0.05 was considered statistically significant.
Microarray analysis of gene expression
The Stress and Aging array was purchased from Operon Technologies (Alameda, CA) and contains 70mer oligos from 695 known genes implicated in DNA repair, cell cycle regulation, stress response, inflammation, and micronutrient metabolism. The array also contains oligos for 19 control genes, for a total of 704 genes. Each oligo was printed in triplicate, for a total of 2112 spots. For radiation, the treated sample was RNA purified from lymphocytes 1 h after irradiation and the control sample was RNA purified at the same time from unirradiated lymphocytes from the same donor. For folate deficiency, the treated sample was lymphocytes cultured in folate deficiency for 9 days, and the control sample was lymphocytes from the same donor cultured in 3000 nM folate for 9 days.
Total RNA was purified from 6 × 107 lymphocytes using the RNeasy Mini Kit (Qiagen, Valencia, CA). RNA quality and quantity were determined by electrophoresis and spectroscopy. Poly(A)+ RNA was subsequently isolated from total RNA using the Oligotex mRNA Mini Kit (Qiagen). RNA labeling was done using reverse transcription and aminoallyl coupling of RNA. mRNA (4 µg) was mixed with 33 µg/ml oligo dT, heated at 70°C for 10 min, and then snap-cooled on ice. Reverse transcription was carried out for 2 h at 42°C using 13 U/µl of the Superscript II reverse transcriptase (Invitrogen), 500 µM dNTPs, and 200 µM aminoallyl dUTP. RNA was hydrolyzed in 0.2 N NaOH and 0.1 M EDTA at 65°C for 15 min, neutralized with 0.3 M Tris-HCl pH 7.4, and washed three times using a microcon-30 filter unit (Millipore, Bedford, MA).
Coupling of cDNA with Cy3 (or Cy5) monoreactive dye (Amersham Pharmacia Biotech, Piscataway, NJ) was done in 50 mM sodium bicarbonate for 1 h. After quenching of uncoupled dyes with 1.3 M hydroxylamine, the probe was washed using the QIAquick PCR Purification Kit (Qiagen), and labeling efficiency was checked by spectroscopy. The probe, in 22 µl

hybridization solution (4× SSC, 0.3% SDS, 1 mg/ml sheared salmon sperm DNA), was heated at 95°C for 2 min and applied to the array slide under a LifterSlip (Erie Scientific, Portsmouth, NH). Hybridization was carried out for 16 h at 65°C. The slide was washed with wash solution 1 (1× SSC, 0.03% SDS), solution 2 (0.2× SSC), and solution 3 (0.05× SSC), and then air-dried and scanned using GenePix 4000b (Axon Instruments, Union City, CA). Signal intensities of the 2112 spots were quantified for Cy3 and Cy5 channels using GenePix Pro 3.0 (Axon Instruments). Signal intensities for triplicates were averaged, and data for Cy3 and Cy5 were normalized using the control genes. Data that had a signal-to-noise ratio <5 were eliminated from further analysis. The ratio of signal intensity in the treated sample to that in the control sample was calculated. Genes with a ratio higher than 2 for up-regulation (or lower than 0.5 for down-regulation) were selected.
RESULTS
Folate deficiency and high doses, but not low doses, of radiation inhibit growth of primary human lymphocytes
Normal human T lymphocytes can proliferate in culture for a few weeks when stimulated with PHA and IL2. We investigated whether radiation or folate deficiency affects the proliferation. For the radiation studies, lymphocytes were cultured in folate-sufficient medium for 7 days and then irradiated at 0, 0.1, 0.25, 1, or 5 Gy. The total number of cells under each condition was counted for 3 consecutive days (until day 10 of culture) after irradiation (Fig. 1A). The growth curves for lymphocytes exposed to 1 Gy or 5 Gy were significantly lower than for those exposed to 0 Gy, but the growth curves for lymphocytes exposed to the lower doses of 0.1 Gy and 0.25 Gy were not statistically different from the growth curve of unirradiated cells. Cells irradiated with 1 Gy had a proliferation rate of ~50% of that of unirradiated cells (P<0.001); 5 Gy completely abolished lymphocyte proliferation (P<0.001).
For folate deficiency studies, lymphocytes were cultured in 0, 6, 12, 30, or 3000 nM folate for 10 days, and the total number of cells for each folate level was counted at 24-h intervals from day 6 to day 10. Cell culture medium containing 3000 nM folate was used as the control because it is the concentration in standard RPMI 1640 medium. However, maximum proliferation can be achieved using 100 nM folate (data not shown). Physiological concentrations of folate in human plasma are ~30 nM, whereas 12 nM is generally considered to be in the deficient range (28, 29). Cells cultured in 12, 6, or 0 nM folate had a decreased growth curve, relative to the growth curve of cells cultured in 3000 nM folate (Fig. 1B). The proliferation rate of cells cultured in 12 nM folate was 46% of that of control cells (P<0.001) and 35% for cells cultured in 6 nM (P<0.001). Lymphocytes cultured in 0 nM folate did not proliferate (P<0.001). Folate deficiency does not interfere with PHA activation; PHA-activated cells cultured in 0 nM folate for a week resume proliferating when folate is added back (unpublished results).
Apoptosis is increased by both folate deficiency and high doses, but not low doses, of ionizing radiation
Because the growth curves of lymphocytes were lowered by both radiation and folate deficiency, we examined whether there was an increase in apoptosis (programmed cell death) under each of these conditions. Cells were collected 24 h after irradiation and after 8 days of folate deficiency

and were analyzed using annexin-V-fluorescein/propidium iodide staining to distinguish between viable, apoptotic, and necrotic cells. For radiation, a significant increase in apoptosis was observed only at the highest dose, 5 Gy, where 16% of the cells were apoptotic, compared with 8% for unirradiated cells (P<0.001). Cells irradiated with 0.1, 0.25, or 1 Gy had a similar percentage of apoptotic cells as unirradiated cells. Although folate-deficient cells only showed statistically significant increases in apoptosis at 6 nM (P<0.05) and 0 nM (P<0.001), compared with non-folate-deficient cells (Fig. 2), linear regression analysis showed a statistically significant dose-dependent increase in apoptosis in the folate-deficient cells relative to the non-folate-deficient cells (P<0.05). The percentage of apoptotic cells was 7% for lymphocytes cultured in 3000 nM folate, 8% in 30 nM, 13% in 12 nM, 17% in 6 nM, and 31% in 0 nM. In this study, physiological condition of folate deficiency induced a level of apoptosis similar to that induced by 5 Gy of radiation.
Both folate deficiency and ionizing radiation at relatively high doses cause DNA breaks, as measured by the Comet assay
We measured DNA breaks using the alkaline comet assay, which is a method that allows detection of both SSB and DSB (44). DNA damage was increased for radiation doses ≥1 Gy, although statistical significance was achieved only at 5 Gy where the Comet score was 1.87, compared with 0.81 for unirradiated cells (P<0.001) (Fig. 3A). A Comet score of 1.20 was obtained when lymphocytes were irradiated with 1 Gy. Similarly, cells cultured for 8 days in low folate showed an increase in DNA damage. Though complete folate deficiency (0 nM) was required to reach statistical significance (P<0.05) (Fig. 3B), linear regression analysis showed a statistically dose-dependent trend in DNA damage (P<0.05). The Comet score was 0.81 for control cells (3000 nM), 1.33 for 12 nM, and 2.01 for 0 nM. For the conditions we used in this study, physiological concentrations of folate deficiency induced as much DNA damage as did 1 Gy of radiation, a relatively high dose of radiation.
Ionizing radiation and folate deficiency differentially affect the cell cycle of lymphocytes
Cell cycle arrest is an early marker of induction of cell cycle checkpoints, DNA damage, and initiation of repair mechanisms. Flow cytometry of propidium iodine-stained cells was used to study how radiation and folate deficiency affect cell cycle distribution. Radiation doses ≥1 Gy caused a significant arrest in the G2/M phase of the cell cycle (Fig. 4A). Unirradiated lymphocytes had 6.0% of their cells in G2/M, whereas it was 15.3% after 1 Gy of radiation (P<0.001) and 26.8% after 5 Gy (P<0.001). The G2/M arrest was still evident 72 h after irradiation (data not shown). In contrast to radiation, folate deficiency caused an arrest in the S phase of the cell cycle in an apparently dose-dependent manner (Fig. 4B). The percentage of cells into the S phase was 34.5% for control cells, 53.4% for 12 nM (P<0.001), and 69.4% for 0 nM (P<0.001). Folate deficiency leads to decreased thymidine and purine availability, which may inhibit DNA synthesis and consequently cell proliferation (45). When 3H-thymidine was added to folate-deficient cells, the thymidine was rapidly incorporated into the cells, suggesting that DNA repair of uracil misincorporation (or slow DNA synthesis) was occurring in S-arrested cells (data not shown).

Ionizing radiation and folate deficiency induce differential gene expression changes, as measured by DNA microarrays
Radiation and folate deficiency induce DNA breaks, but by different mechanisms; therefore, we investigated how gene expression profiles were affected. We used Operon�s Stress and Aging array, which contains 704 genes. For radiation studies, RNA was extracted 1 h after 1 or 5 Gy of radiation. Changes in gene expression were determined by comparing mRNA from irradiated cells to mRNA from unirradiated cells obtained from the same donor. Genes whose expressions were changed twofold or more by either radiation dose are listed in Table 1. After irradiation, 24 genes were up-regulated, including DNA repair and stress genes, and 23 genes were down-regulated, mostly cell cycle and mitochondrial genes. All genes listed in Table 1 had a ratio higher than twofold after 1 Gy or 5 Gy of radiation; the ratios were generally lower in the case of cells exposed to only 1 Gy.
For folate deficiency studies, lymphocytes were cultured in 0 nM folate (complete deficiency) or 12 nM folate (physiological deficiency) for 10 days before RNA from the samples was extracted. The mRNA levels from the folate-deficient cells were compared with mRNA levels from folate-sufficient (3000 nM) control cells obtained from the same donor. Most of the 24 genes up-regulated in folate deficiency were DNA repair and mitochondrial genes; half of the 14 down-regulated genes were involved in folate metabolism (Table 2). Similar to radiation, there was a dose-dependant effect on gene expression; all genes but one were affected by 0 nM folate, whereas the ratios were often under the threshold of 2 for 12 nM folate.
DISCUSSION
In the current study, we sought to compare the ability to damage DNA of an exogenous source of stress, ionizing radiation, and an endogenous source, folate deficiency. Our results suggest that physiologically relevant levels of folate deficiency cause more DNA breaks, induce more apoptosis, and halt the proliferation of normal human lymphocytes in primary culture more than acute exposure to relatively high doses of radiation.
The population is generally exposed to only very low doses of radiation. Because risk estimates for low doses are hard to assess, they are often extrapolated from effects observed at higher doses. The usefulness of this linear model has been debated because it may overestimate cancer risks (15, 16, 20). Other models have been proposed in which low-dose radiation is harmless under a certain threshold, or may even induce a radioadaptive response (14�16, 18). On the other hand, deficiencies in vitamins and minerals are common in the population and may contribute to much of preventable cancer (4).
Ionizing radiation and folate deficiency can both produce DSB. Using microarray analysis of gene expression, we showed that radiation and folate deficiency activated different sets of genes, suggesting that different cell responses resulted from each type of exposure. This finding was not unexpected, because radiation and folate deficiency lead to DSB by different mechanisms. Ionizing radiation can form DSB either directly or indirectly by forming clusters of oxidative damage, which can be converted to DNA breaks during repair (2, 6). Folate deficiency, however, causes an imbalance in the thymidine/uracil ratio, which results in uracil being incorporated into DNA during synthesis. The repair of two nearby opposing lesions can cause a DSB (23).

DNA damage after irradiation and during folate deficiency was measured using the Comet assay, an assay that has been widely used for both radiation exposure and nutritional studies (46�48). Because radiation and folate deficiency cause DNA breaks in a different fashion, we used the alkaline Comet assay (44) to simultaneously measure SSB and DSB, therefore allowing one comparison between the two risks. It would also be of interest to measure only DSB. DSB are often considered to the most dangerous DNA lesions, although SSB can be converted to DSB when locations of damage (SSB and/or base lesions) are clustered. Lymphocytes that were irradiated with 5 Gy were virtually all damaged, as were lymphocytes cultured in 0 nM folate, and both Comet scores were increased by ~2.5-fold compared with control cells. These results are similar to those of experiments in which lymphocytes were either irradiated (49, 50) or cultured for 8 days in folate-deficient medium (48). The cells cultured in a physiological level of folate deficiency (12 nM) sustained damage equivalent to that of lymphocytes irradiated with 1 Gy, which is a relatively high dose of radiation.
Similar results were obtained using other techniques to measure DNA breaks. Fenech et al. (31) have also investigated DNA damage resulting from folate deficiency and ionizing radiation in human lymphocytes using the cytokinesis-block micronuclei assay to measure DSB. Crott et al. (28) and Fenech and Morley (51) found that folate deficiency (12 nM) caused a fourfold increase in micronuclei frequency, as did 0.4 Gy of radiation, although the two independent experiments may not be directly comparable. Branda et al. (42) used another cell type and the alkaline filter elution assay to measure SSB and DSB. They found that folate-deficient Chinese hamster ovary cells manifested strand breaks equivalent to a dose of 0.3 Gy.
In addition to increased DNA damage, radiation and folate deficiency had other cellular effects. The proliferation rate was reduced to ~50% by 1 Gy and to nearly 0% by 5 Gy of radiation. Proliferation rate was not reduced by 0.25 Gy and was slightly increased by 0.1 Gy of radiation. Activation of proliferation after low dose of radiation has been observed for some cancer cells during radiotherapy and also, but more rarely, for normal cells (52�54). Similarly for folate deficiency, the proliferation rate was reduced to 46% when cells were cultured in 12 nM folate, whereas cells cultured in the absence of folate totally stopped proliferation.
The lower proliferation rate after irradiation and during folate deficiency was due to both increased apoptosis and cell cycle arrest. Irradiated cells are known to stop in G2/M because DNA damage will activate this checkpoint and prevent cells from undergoing mitosis (55). In this study, a significant arrest in G2/M was observed after 1 and 5 Gy of radiation; the arrest lasted up to 72 h post-irradiation (data not shown). The microarray analysis showed that ATM and GADD45, two genes known to be induced by radiation and to be involved in G2/M arrest (7, 56), were up-regulated by 1 and 5 Gy of radiation. In contrast, folate deficiency induced a cell cycle arrest in the S phase. A prominent arrest in S was observed using a physiological level (12 nM) of folate deficiency. When lymphocytes were cultured in the absence of folate (0 nM), most cells were arrested in the S phase, and no cells were detected in the mitotic phase (G2/M). The S-phase arrest during folate deficiency has been observed with various cell types (45, 57, 58) and is likely due to impaired DNA synthesis, because folate is required for both purine and thymidylate synthesis. The S-phase arrest is reversible, and cell proliferation quickly resumes when folate (or a nucleotide mix) is added back to the folate-deficient cells (45, and unpublished results).

The proportion of lymphocytes undergoing apoptosis increased with the radiation dose and the level of folate deficiency. Cells irradiated with 5 Gy were highly damaged; therefore, the apoptotic pathway was probably activated because the cells were unable to repair their DNA. A similar proportion of apoptotic cells were measured when lymphocytes were cultured in low concentrations (6�12 nM) of folate. In absence of folate (0 nM), almost a third of the lymphocytes were apoptotic. Huang et al. has suggested that folate deficiency may block DNA replication and/or mitosis, which may be involved in apoptosis induction (58). In support of this hypothesis, two antifolates, 5-fluorouracil (inhibitor of thymidylate synthase) and methotrexate (inhibitor of dihydrofolate reductase), have been shown to induce apoptosis in association with DNA synthesis inhibition (59, 60).
Though physiological levels of folate deficiency (12 nM) appear to cause as much DNA damage as relatively high doses (1 Gy) of ionizing radiation, the cellular responses, as measured by DNA microarray, are quite different. We used a microarray designed to measure gene expression changes of DNA repair, cell cycle regulation, stress response, inflammation, and micronutrient metabolism genes. Shortly after lymphocytes were irradiated, many DNA repair and stress genes were up-regulated, including ATM and GADD45, which are genes involved in the G2/M arrest after radiation. This has also been observed in other cell types (8�12). Consistent with the various kinds of DNA damage induced by radiation, many DNA repair genes were up-regulated. RAD51 and XRRC3 interact together in DSB repair (61); Nth-1 and flap structure-specific endonuclease are involved in excision repair (62, 63).
Eight stress-related genes were induced, likely as a result of increased oxidative damage. Interestingly, whereas stress-related genes were up-regulated in a dose-dependent way with radiation, DNA repair genes were up-regulated at 1 Gy, but were less up-regulated at 5 Gy. Irradiation with 5 Gy induced apoptosis and may then be less likely to induce DNA repair as well. Among the other genes that were highly up-regulated by radiation were troponin 1 and T2, although their function in irradiated lymphocytes remains unclear. Down-regulation of cell cycle genes, after irradiation, is consistent with the observed cell cycle arrest. Most of the mitochondrially encoded genes were highly down-regulated by radiation. This suggests that mitochondrial DNA, as compared with nuclear DNA, is more sensitive to DNA damage due to ionizing radiation. After 1 and 5 Gy of radiation, mitochondrial DNA may be too damaged to transcribe new mRNAs. Mitochondrial DNA has been shown to display an increased susceptibility to the radiation-induced loss of integrity compared with nuclear DNA in Hep G2 cells (64). Four nuclear-encoded mitochondria genes were also down-regulated, which may be a consequence of the damage sustained by the mitochondria.
When lymphocytes were cultured in folate-deficient conditions, some DNA repair genes were up-regulated, although they were not the same genes that were activated by radiation. Uracil DNA glycosylase and APEX nuclease are involved in the base excision repair of uracil misincorporation (65). Glyceraldehyde-3-phosphate dehydrogenase is a multifunctional enzyme that has a uracil DNA glycosylase activity (66). Rad23 is part of a DNA damage recognition complex for nucleotide excision repair (67). Genes known to be involved in folate metabolism or to need folate as a cofactor, which were not affected by radiation, were regulated by folate deficiency. Two folate metabolism genes were up-regulated, and seven were down-regulated as a consequence of the lack of folate. In contrast to the effects of radiation, very few cell cycle genes

were affected by folate deficiency, probably because the cell cycle arrest was due to slow DNA synthesis, rather than to an induced arrest such as caused by radiation.
Another observed difference with radiation is that mitochondrially encoded genes were up-regulated during folate deficiency, which is consistent with elevated apoptosis. In dopaminergic neurons, folate deficiency elevates homocysteine levels, which increase oxidative stress, mitochondrial dysfunction, and apoptosis (68). Little has been published on microarray analysis of gene expression during folate deficiency. Jhaveri et al. (69) have found only eight genes whose expression was affected by low folate, but they used a nasopharyngeal epidermoid carcinoma cell line that may be less sensitive to the lack of folate than the activated T-lymphocytes we used.
Our results showed that physiological levels of folate deficiency caused more DNA damage than low-dose radiation (≤0.1 Gy) in primary T lymphocytes. Because our study was done using cultured cells, results cannot be directly extrapolated to estimate cancer risks. Human exposure to chronic low doses of radiation is likely to have a different outcome than acute cell exposure. Interestingly, chronic and acute low doses of radiation can induce a radioadaptive response, therefore protecting the cells from damage induced by a subsequent irradiation (17�19, 70). This adaptive response has not been observed for folate deficiency. Folate deficiency is known to cause DSB (23, 28, 36, 37), but our microarray data showed that DSB recognition and repair genes are not activated. Moreover, folate deficiency has been shown to impair DNA excision repair and mismatch repair (39�41). This further suggests that folate deficiency may be more detrimental than low doses of radiation. In addition, dietary folate deficiency is systemic and also causes damage to sperm DNA (71).
Note that our study did not address the issue of whether low doses of radiation are harmful to human health (for a review of low-dose radiation effects, see ref 72); rather, our study found that low doses of radiation had no measurable effects in comparison with conditions of folate deficiency for the cells we used. Although further investigation is needed, these findings suggest that exposure to very low doses of radiation from background or occupational exposure may pose a smaller cancer risk than consuming a poor diet (4). Many vitamins and minerals are important for DNA integrity. A deficiency in vitamin B12 causes DNA damage by the same mechanism as folate, and antioxidant vitamins and minerals can protect DNA from damage by reactive oxygen species (4). More importantly, our results suggest that research on the biological effects of low-dose radiation, in humans, should take into account the nutritional status of the subjects, because folate (and other vitamin and mineral) deficiency could confound the effects of low-dose radiation or could even have a synergistic effect and increase the sensitivity of cells to radiation (42, 73, 74).
Additional studies comparing the relative effects of ionizing radiation with deficiency in a vitamin such as folate may better elucidate the relative contributions of these two exposures to DNA damage and put both in perspective for radiation exposure standards, and for the determination of optimal vitamin requirements for DNA integrity.

ACKNOWLEDGMENTS
We thank Eric Winstall and John Nides for critical reading of the manuscript, Jay Tong and Bruce Sun from AllCells for providing the human lymphocytes, and the microarray team at Operon Technologies for their help. C.C. was supported by a fellowship from the Conseil de Recherches Médicales (CRM) du Canada. This work was supported by the U.S. Department of Energy (DE-FG03-00ER62943), the National Foundation for Cancer Research (00-CHORI), and the National Institute of Environmental Health Sciences Center (P30-ES0186).
REFERENCES
1. Ames, B. N., Gold, L. S., and Willett, W. C. (1995) The causes and prevention of cancer. Proc. Natl. Acad. Sci. USA 92, 5258�5265
2. Prise, K. M., Ahnstrom, G., Belli, M., Carlsson, J., Frankenberg, D., Kiefer, J., Lobrich, M., Michael, B. D., Nygren, J., Simone, G., et al. (1998) A review of dsb induction data for varying quality radiations. Int. J. Radiat. Biol. 74, 173�184
3. Fenech, M., and Crott, J. W. (2002) Micronuclei, nucleoplasmic bridges and nuclear buds induced in folic acid deficient human lymphocytes-evidence for breakage-fusion-bridge cycles in the cytokinesis-block micronucleus assay. Mutat. Res. 504, 131�136
4. Ames, B. N., and Wakimoto, P. (2002) Are Vitamin and Mineral Deficiencies a major Cancer Risk? Nat. Rev. Cancer 2, 694�704
5. Ward, J. F. (1988) DNA damage produced by ionizing radiation in mammalian cells: identities, mechanisms of formation, and reparability. Prog. Nucleic Acid Res. Mol. Biol. 35, 95�125
6. Jenner, T. J., Fulford, J., and O�Neill, P. (2001) Contribution of base lesions to radiation-induced clustered DNA damage: implication for models of radiation response. Radiat. Res. 156, 590�593
7. Xu, B., Kim, S. T., Lim, D. S., and Kastan, M. B. (2002) Two molecularly distinct G(2)/M checkpoints are induced by ionizing irradiation. Mol. Cell. Biol. 22, 1049�1059
8. Zhao, R., Gish, K., Murphy, M., Yin, Y., Notterman, D., Hoffman, W. H., Tom, E., Mack, D. H., and Levine, A. J. (2000) Analysis of p53-regulated gene expression patterns using oligonucleotide arrays. Genes Dev. 14, 981�993
9. Ford, B. N., Wilkinson, D., Thorleifson, E. M., and Tracy, B. L. (2001) Gene expression responses in lymphoblastoid cells after radiation exposure. Radiat. Res. 156, 668�671
10. Amundson, S. A., Bittner, M., Meltzer, P., Trent, J., and Fornace, A. J., Jr. (2001) Induction of gene expression as a monitor of exposure to ionizing radiation. Radiat. Res. 156, 657�661
11. Tusher, V. G., Tibshirani, R., and Chu, G. (2001) Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 98, 5116�5121

12. Voehringer, D. W., Hirschberg, D. L., Xiao, J., Lu, Q., Roederer, M., Lock, C. B., Herzenberg, L. A., and Steinman, L. (2000) Gene microarray identification of redox and mitochondrial elements that control resistance or sensitivity to apoptosis. Proc. Natl. Acad. Sci. USA 97, 2680�2685
13. Committee on the Biological Effects of Ionizing Radiation, National Research Council. Health Effects of Exposure to Low Levels of Ionizing Radiation: BEIR V, The National Academies Press, Washington, DC
14. Hoel, D. G., and Li, P. (1998) Threshold models in radiation carcinogenesis. Health Phys.
75, 241�250
15. Pollycove, M., and Feinendegen, L. E. (2001) Biologic responses to low doses of ionizing radiation: Detriment versus hormesis. Part 2. Dose responses of organisms. J. Nucl. Med. 42, 26N�32N, 37N
16. Feinendegen, L. E., and Pollycove, M. (2001) Biologic responses to low doses of ionizing radiation: detriment versus hormesis. Part 1. Dose responses of cells and tissues. J. Nucl. Med. 42, 17N�27N
17. Shadley, J. D., Afzal, V., and Wolff, S. (1987) Characterization of the adaptive response to ionizing radiation induced by low doses of X rays to human lymphocytes. Radiat. Res. 111, 511�517
18. Wolff, S. (1998) The adaptive response in radiobiology: evolving insights and implications. Environ. Health Perspect. 106, Suppl 1, 277�283
19. Iyer, R., and Lehnert, B. E. (2002) Low dose, low-LET ionizing radiation-induced radioadaptation and associated early responses in unirradiated cells. Mutat. Res. 503, 1�9
20. Anderson, L. M. (1998) The cancer risk assessement paradigm: an experimentalist�s viewpoint. Comments Toxicology 6, 283�288
21. Olinski, R., Zastawny, T. H., Foksinski, M., Barecki, A., and Dizdaroglu, M. (1995) DNA base modifications and antioxidant enzyme activities in human benign prostatic hyperplasia. Free Radic. Biol. Med. 18, 807�813
22. Beckman, K. B., and Ames, B. N. (1997) Oxidative decay of DNA. J. Biol. Chem. 272, 19633�19636
23. Blount, B. C., Mack, M. M., Wehr, C. M., MacGregor, J. T., Hiatt, R. A., Wang, G., Wickramasinghe, S. N., Everson, R. B., and Ames, B. N. (1997) Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: implications for cancer and neuronal damage. Proc. Natl. Acad. Sci. USA 94, 3290�3295
24. Ames, B. N. (1998) Micronutrients prevent cancer and delay aging. Toxicol. Lett. 102-103, 5�18

25. Fenech, M., and Ferguson, L. R. (2001) Vitamins/minerals and genomic stability in humans. Mutat. Res. 475, 1�6
26. Bailey, L. B., Moyers, S., and Gregory, J. F. (2001) Folate. Present Knowledge in Nutrition 8th Edition, ILSI Press
27. Choi, S. W., and Mason, J. B. (2000) Folate and carcinogenesis: an integrated scheme. J. Nutr. 130, 129�132
28. Crott, J. W., Mashiyama, S. T., Ames, B. N., and Fenech, M. F. (2001) Methylenetetrahydrofolate reductase C677T polymorphism does not alter folic acid deficiency-induced uracil incorporation into primary human lymphocyte DNA in vitro. Carcinogenesis 22, 1019�1025
29. Bronstrup, A., Hages, M., Prinz-Langenohl, R., and Pietrzik, K. (1998) Effects of folic acid and combinations of folic acid and vitamin B-12 on plasma homocysteine concentrations in healthy, young women. Am. J. Clin. Nutr. 68, 1104�1110
30. Jacob, R. A., Gretz, D. M., Taylor, P. C., James, S. J., Pogribny, I. P., Miller, B. J., Henning, S. M., and Swendseid, M. E. (1998) Moderate folate depletion increases plasma homocysteine and decreases lymphocyte DNA methylation in postmenopausal women. J. Nutr. 128, 1204�1212
31. Fenech, M. F., Dreosti, I. E., and Rinaldi, J. R. (1997) Folate, vitamin B12, homocysteine status and chromosome damage rate in lymphocytes of older men. Carcinogenesis 18, 1329�1336
32. Jacques, P. F., Selhub, J., Bostom, A. G., Wilson, P. W., and Rosenberg, I. H. (1999) The effect of folic acid fortification on plasma folate and total homocysteine concentrations. N. Engl. J. Med. 340, 1449�1454
33. Ravanat, J. L., Douki, T., Duez, P., Gremaud, E., Herbert, K., Hofer, T., Lasserre, L., Saint-Pierre, C., Favier, A., and Cadet, J. (2002) Cellular background level of 8-oxo-7,8-dihydro-2�-deoxyguanosine: an isotope based method to evaluate artefactual oxidation of DNA during its extraction and subsequent work-up. Carcinogenesis 23, 1911�1918
34. Helbock, H. J., Beckman, K. B., Shigenaga, M. K., Walter, P. B., Woodall, A. A., Yeo, H. C., and Ames, B. N. (1998) DNA oxidation matters: the HPLC-electrochemical detection assay of 8- oxo-deoxyguanosine and 8-oxo-guanine. Proc. Natl. Acad. Sci. USA 95, 288�293
35. Beckman, K. B., and Ames, B. N. (1998) The free radical theory of aging matures. Physiol. Rev. 78, 547�581
36. Chen, A. T., Reidy, J. A., Annest, J. L., Welty, T. K., and Zhou, H. G. (1989) Increased chromosome fragility as a consequence of blood folate levels, smoking status, and coffee consumption. Environ. Mol. Mutagen. 13, 319�324

37. Everson, R. B., Wehr, C. M., Erexson, G. L., and MacGregor, J. T. (1988) Association of marginal folate depletion with increased human chromosomal damage in vivo: demonstration by analysis of micronucleated erythrocytes. J. Natl. Cancer Inst. 80, 525�529
38. Fenech, M. (2001) The role of folic acid and Vitamin B12 in genomic stability of human cells. Mutat. Res. 475, 57�67
39. Duthie, S. J., Narayanan, S., Blum, S., Pirie, L., and Brand, G. M. (2000) Folate deficiency in vitro induces uracil misincorporation and DNA hypomethylation and inhibits DNA excision repair in immortalized normal human colon epithelial cells. Nutr. Cancer 37, 245�251
40. Choi, S. W., Kim, Y. I., Weitzel, J. N., and Mason, J. B. (1998) Folate depletion impairs DNA excision repair in the colon of the rat. Gut 43, 93�99
41. Cravo, M. L., Albuquerque, C. M., Salazar de Sousa, L., Gloria, L. M., Chaves, P., Dias Pereira, A., Nobre Leitao, C., Quina, M. G., and Costa Mira, F. (1998) Microsatellite instability in non-neoplastic mucosa of patients with ulcerative colitis: effect of folate supplementation. Am. J. Gastroenterol. 93, 2060�2064
42. Branda, R. F., and Blickensderfer, D. B. (1993) Folate deficiency increases genetic damage caused by alkylating agents and gamma-irradiation in Chinese hamster ovary cells. Cancer Res. 53, 5401�5408
43. Branda, R. F., O�Neill, J. P., Brooks, E. M., Trombley, L. M., and Nicklas, J. A. (2001) The effect of folate deficiency on the cytotoxic and mutagenic responses to ethyl methanesulfonate in human lymphoblastoid cell lines that differ in p53 status. Mutat. Res. 473, 51�71
44. Olive, P. L., Wlodek, D., Durand, R. E., and Banath, J. P. (1992) Factors influencing DNA migration from individual cells subjected to gel electrophoresis. Exp. Cell Res. 198, 259�267
45. Koury, M. J., Price, J. O., and Hicks, G. G. (2000) Apoptosis in megaloblastic anemia occurs during DNA synthesis by a p53-independent, nucleoside-reversible mechanism. Blood 96, 3249�3255

46. Maluf, S. W., Passos, D. F., Bacelar, A., Speit, G., and Erdtmann, B. (2001) Assessment of DNA damage in lymphocytes of workers exposed to X-radiation using the micronucleus test and the comet assay. Environ. Mol. Mutagen. 38, 311�315
47. Malyapa, R. S., Bi, C., Ahern, E. W., and Roti Roti, J. L. (1998) Detection of DNA damage by the alkaline comet assay after exposure to low-dose gamma radiation. Radiat. Res. 149, 396�400
48. Duthie, S. J., and Hawdon, A. (1998) DNA instability (strand breakage, uracil misincorporation, and defective repair) is increased by folic acid depletion in human lymphocytes in vitro. FASEB J. 12, 1491�1497
49. Trenz, K., Rothfuss, A., Schutz, P., and Speit, G. (2002) Mutagen sensitivity of peripheral blood from women carrying a BRCA1 or BRCA2 mutation. Mutat. Res. 500, 89�96
50. Popanda, O., Ebbeler, R., Twardella, D., Helmbold, I., Gotzes, F., Schmezer, P., Thielmann, H. W., von Fournier, D., Haase, W., Sautter-Bihl, M. L., et al. (2003) Radiation-induced DNA damage and repair in lymphocytes from breast cancer patients and their correlation with acute skin reactions to radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 55, 1216�1225
51. Fenech, M., and Morley, A. A. (1986) Cytokinesis-block micronucleus method in human lymphocytes: effect of in vivo ageing and low dose X-irradiation. Mutat. Res. 161, 193�198
52. Schmidt-Ullrich, R. K., Mikkelsen, R. B., Dent, P., Todd, D. G., Valerie, K., Kavanagh, B. D., Contessa, J. N., Rorrer, W. K., and Chen, P. B. (1997) Radiation-induced proliferation of the human A431 squamous carcinoma cells is dependent on EGFR tyrosine phosphorylation. Oncogene 15, 1191�1197
53. Kavanagh, B. D., Lin, P. S., Chen, P., and Schmidt-Ullrich, R. K. (1995) Radiation-induced enhanced proliferation of human squamous cancer cells in vitro: a release from inhibition by epidermal growth factor. Clin. Cancer Res. 1, 1557�1562
54. Suzuki, K., Kodama, S., and Watanabe, M. (2001) Extremely low-dose ionizing radiation causes activation of mitogen-activated protein kinase pathway and enhances proliferation of normal human diploid cells. Cancer Res. 61, 5396�5401
55. Hartwell, L. H., and Kastan, M. B. (1994) Cell cycle control and cancer. Science 266, 1821�1828

56. Zhan, Q., Antinore, M. J., Wang, X. W., Carrier, F., Smith, M. L., Harris, C. C., and Fornace, A. J., Jr. (1999) Association with Cdc2 and inhibition of Cdc2/Cyclin B1 kinase activity by the p53-regulated protein Gadd45. Oncogene 18, 2892�2900
57. Zhu, W. Y., and Melera, P. W. (2001) Basal levels of metallothionein I and II expression in mouse embryo fibroblasts enhance growth in low folate through a cell cycle mediated pathway. Cell Biol. Int. 25, 1261�1269
58. Huang, R. F., Ho, Y. H., Lin, H. L., Wei, J. S., and Liu, T. Z. (1999) Folate deficiency induces a cell cycle-specific apoptosis in HepG2 cells. J. Nutr. 129, 25�31
59. Longley, D. B., Boyer, J., Allen, W. L., Latif, T., Ferguson, P. R., Maxwell, P. J., McDermott, U., Lynch, M., Harkin, D. P., and Johnston, P. G. (2002) The role of thymidylate synthase induction in modulating p53-regulated gene expression in response to 5-fluorouracil and antifolates. Cancer Res. 62, 2644�2649
60. Izeradjene, K., Revillard, J. P., and Genestier, L. (2001) Inhibition of thymidine synthesis by folate analogues induces a Fas-Fas ligand-independent deletion of superantigen-reactive peripheral T cells. Int. Immunol. 13, 85�93
61. Liu, N., Lamerdin, J. E., Tebbs, R. S., Schild, D., Tucker, J. D., Shen, M. R., Brookman, K. W., Siciliano, M. J., Walter, C. A., Fan, W., et al. (1998) XRCC2 and XRCC3, new human Rad51-family members, promote chromosome stability and protect against DNA cross-links and other damages. Mol. Cell 1, 783�793
62. Aspinwall, R., Rothwell, D. G., Roldan-Arjona, T., Anselmino, C., Ward, C. J., Cheadle, J. P., Sampson, J. R., Lindahl, T., Harris, P. C., and Hickson, I. D. (1997) Cloning and characterization of a functional human homolog of Escherichia coli endonuclease III. Proc. Natl. Acad. Sci. USA 94, 109�114
63. Rydberg, B., and Game, J. (2002) Excision of misincorporated ribonucleotides in DNA by RNase H (type 2) and FEN-1 in cell-free extracts. Proc. Natl. Acad. Sci. USA 99, 16654�16659
64. Morales, A., Miranda, M., Sanchez-Reyes, A., Biete, A., and Fernandez-Checa, J. C. (1998) Oxidative damage of mitochondrial and nuclear DNA induced by ionizing radiation in human hepatoblastoma cells. Int. J. Radiat. Oncol. Biol. Phys. 42, 191�203
65. Krokan, H. E., Nilsen, H., Skorpen, F., Otterlei, M., and Slupphaug, G. (2000) Base excision repair of DNA in mammalian cells. FEBS Lett. 476, 73�77
66. Meyer-Siegler, K., Mauro, D. J., Seal, G., Wurzer, J., deRiel, J. K., and Sirover, M. A. (1991) A human nuclear uracil DNA glycosylase is the 37-kDa subunit of glyceraldehyde-3-phosphate dehydrogenase. Proc. Natl. Acad. Sci. USA 88, 8460�8464
67. Masutani, C., Sugasawa, K., Yanagisawa, J., Sonoyama, T., Ui, M., Enomoto, T., Takio, K., Tanaka, K., van der Spek, P. J., Bootsma, D., et al. (1994) Purification and cloning of a

nucleotide excision repair complex involving the xeroderma pigmentosum group C protein and a human homologue of yeast RAD23. EMBO J. 13, 1831�1843
68. Duan, W., Ladenheim, B., Cutler, R. G., Kruman, I. I., Cadet, J. L., and Mattson, M. P. (2002) Dietary folate deficiency and elevated homocysteine levels endanger dopaminergic neurons in models of Parkinson�s disease. J. Neurochem. 80, 101�110
69. Jhaveri, M. S., Wagner, C., and Trepel, J. B. (2001) Impact of extracellular folate levels on global gene expression. Mol. Pharmacol. 60, 1288�1295
70. Gourabi, H., and Mozdarani, H. (1998) A cytokinesis-blocked micronucleus study of the radioadaptive response of lymphocytes of individuals occupationally exposed to chronic doses of radiation. Mutagenesis 13, 475�480
71. Wallock, L. M., Tamura, T., Mayr, C. A., Johnston, K. E., Ames, B. N., and Jacob, R. A. (2001) Low seminal plasma folate concentrations are associated with low sperm density and count in male smokers and nonsmokers. Fertil. Steril. 75, 252�259
72. Joiner, M. C., Marples, B., Lambin, P., Short, S. C., and Turesson, I. (2001) Low-dose hypersensitivity: current status and possible mechanisms. Int. J. Radiat. Oncol. Biol. Phys. 49, 379�389
73. Fenech, M. (2001) Recommended dietary allowances (RDAs) for genomic stability. Mutat. Res. 480-481, 51�54
74. Konopacka, M., and Rzeszowska-Wolny, J. (2001) Antioxidant vitamins C, E and beta-carotene reduce DNA damage before as well as after gamma-ray irradiation of human lymphocytes in vitro. Mutat. Res. 491, 1�7
Received July 1, 2003; accepted September 24, 2003.
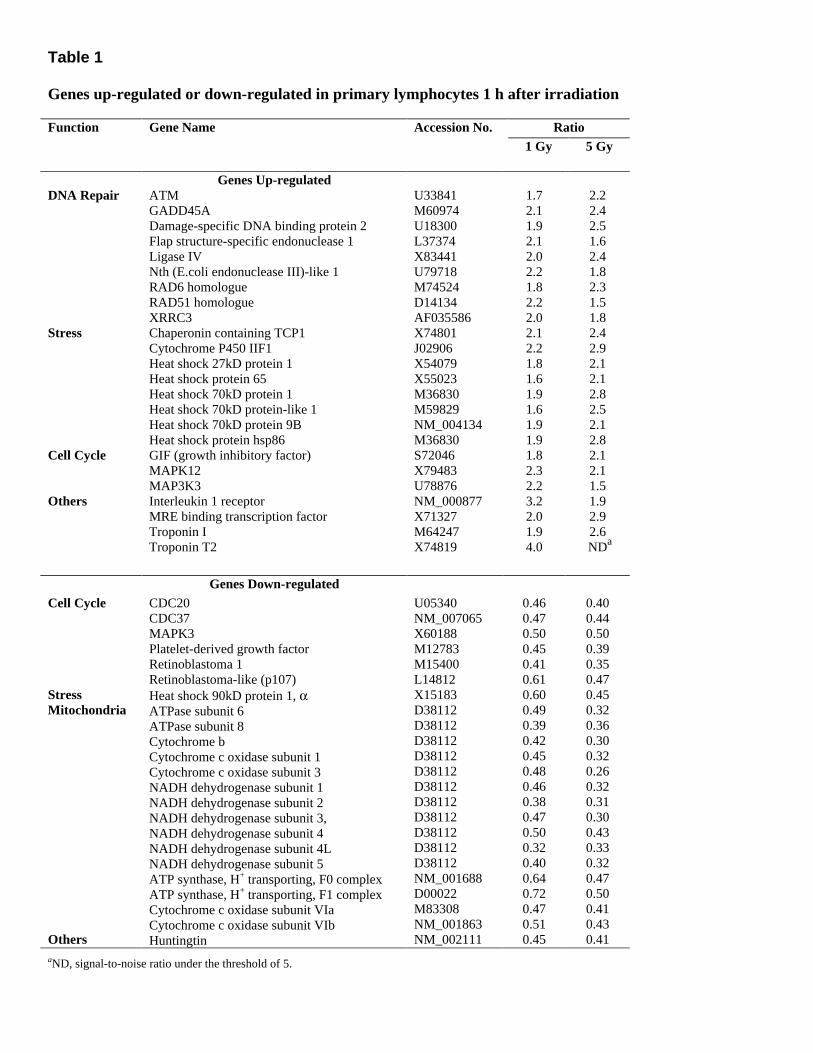
Table 1 Genes up-regulated or down-regulated in primary lymphocytes 1 h after irradiation
Ratio Function Gene Name
Accession No. 1 Gy 5 Gy
DNA Repair Stress Cell Cycle Others
Genes Up-regulated ATM GADD45A Damage-specific DNA binding protein 2 Flap structure-specific endonuclease 1 Ligase IV Nth (E.coli endonuclease III)-like 1 RAD6 homologue RAD51 homologue XRRC3 Chaperonin containing TCP1 Cytochrome P450 IIF1 Heat shock 27kD protein 1 Heat shock protein 65 Heat shock 70kD protein 1 Heat shock 70kD protein-like 1 Heat shock 70kD protein 9B Heat shock protein hsp86 GIF (growth inhibitory factor) MAPK12 MAP3K3 Interleukin 1 receptor MRE binding transcription factor Troponin I Troponin T2
U33841 M60974 U18300 L37374 X83441 U79718 M74524 D14134 AF035586 X74801 J02906 X54079 X55023 M36830 M59829 NM_004134 M36830 S72046 X79483 U78876 NM_000877 X71327 M64247 X74819
1.7 2.1 1.9 2.1 2.0 2.2 1.8 2.2 2.0 2.1 2.2 1.8 1.6 1.9 1.6 1.9 1.9 1.8 2.3 2.2 3.2 2.0 1.9 4.0
2.2 2.4 2.5 1.6 2.4 1.8 2.3 1.5 1.8 2.4 2.9 2.1 2.1 2.8 2.5 2.1 2.8 2.1 2.1 1.5 1.9 2.9 2.6 NDa
Genes Down-regulated Cell Cycle Stress Mitochondria Others
CDC20 CDC37 MAPK3 Platelet-derived growth factor Retinoblastoma 1 Retinoblastoma-like (p107) Heat shock 90kD protein 1, α ATPase subunit 6 ATPase subunit 8 Cytochrome b Cytochrome c oxidase subunit 1 Cytochrome c oxidase subunit 3 NADH dehydrogenase subunit 1 NADH dehydrogenase subunit 2 NADH dehydrogenase subunit 3, NADH dehydrogenase subunit 4 NADH dehydrogenase subunit 4L NADH dehydrogenase subunit 5 ATP synthase, H+ transporting, F0 complex ATP synthase, H+ transporting, F1 complex Cytochrome c oxidase subunit VIa Cytochrome c oxidase subunit VIb Huntingtin
U05340 NM_007065 X60188 M12783 M15400 L14812 X15183 D38112 D38112 D38112 D38112 D38112 D38112 D38112 D38112 D38112 D38112 D38112 NM_001688 D00022 M83308 NM_001863 NM_002111
0.46 0.47 0.50 0.45 0.41 0.61 0.60 0.49 0.39 0.42 0.45 0.48 0.46 0.38 0.47 0.50 0.32 0.40 0.64 0.72 0.47 0.51 0.45
0.40 0.44 0.50 0.39 0.35 0.47 0.45 0.32 0.36 0.30 0.32 0.26 0.32 0.31 0.30 0.43 0.33 0.32 0.47 0.50 0.41 0.43 0.41
aND, signal-to-noise ratio under the threshold of 5.

Table 2 Genes up-regulated or down-regulated in primary lymphocytes during folate deficiency
Ratio Function Gene name
Accession no. 12 nM 0 nM
DNA repair Folate-Related Mitochondria Others
Genes Up-regulated APEX nuclease DNA (cytosine-5-)-methyltransferase 1 Glyceraldehyde-3-phosphate dehydrogenase RAD23 homologue B Single-strand selective uracil DNA glycosylase Choline kinase Serine hydroxymethyl transferase ATPase subunit 6 Cytochrome c oxidase subunit 1 Cytochrome c oxidase subunit 3 NADH dehydrogenase subunit 1 NADH dehydrogenase subunit 2 NADH dehydrogenase subunit 3, NADH dehydrogenase subunit 5 ATP binding cassette sub-family C (CFTR/MRP) Interleukin 1, α Interleukin 1, β Major histocompatibility complex, class I, C Ribosomal protein S9 Ribosomal protein L13a Vimentin
U79268 X63692 X01677 D21090 AF125182 D10704 NM_004169 D38112 D38112 D38112 D38112 D38112 D38112 D38112 L05628 M15329 M15330 M11886 U14971 X56932 X56134
1.4 1.3 2.1 0.8
NDa 0.9 2.3 1.8 2.7 2.0 1.7 2.0 1.7 1.8 1.2 1.3 1.4 2.0 2.3 1.9 3.3
2.3 2.4 2.1 2.2 2.1 2.0 2.2 5.7 3.8 3.5 3.1 3.9 3.4 3.6 2.6 2.3 2.6 2.5 2.4 2.4 3.2
Genes Down-regulated Cell cycle Folate-Related Others
Cyclin-dependent kinase inhibitor 2B MAPK12 Aldehyde dehydrogenase 9 Choline dehydrogenase Folate transporter Folylpolyglutamate synthase Methylene tetrahydrofolate dehydrogenase Phosphate cytidylyltransferase 1, choline, alpha Thymine-DNA-glycosylase Metal-regulatory transcription factor 1 MRE-binding transcription factor Nicotinamide nucleotide transhydrogenase Nucleolar phosphoprotein P130 Thrombospondin 2
U17075 X79483 X75425 AJ272267 U19720 M98045 X16396 NM_005017 U51166 X78710 X71327 Z50101 Z34289 L12350
0.87 0.44 ND 0.28 ND 0.56 0.45 0.56 0.41 0.59 0.37 ND 0.67 0.65
0.38 0.41 0.42 0.62 0.42 0.33 0.44 0.50 0.47 0.25 0.39 0.25 0.33 0.33
aND, signal-to-noise ratio under the threshold of 5.

Fig. 1
Figure 1. Growth curves for primary human lymphocytes subjected to ionizing radiation (A) or cultured in varying folate concentrations (B). Culture and irradiation were carried out as described in Materials and Methods; viable cells were counted at 24 h intervals. Data are mean ± SE of three separate experiments. *P < 0.05 compared with control. ***P < 0.001 compared with control; control is 0 Gy (A) and 3000 nM folate (B).

Fig. 2
Figure 2. Induction of apoptosis 24 h after irradiation (A) or after 8 days of culture in varying folate concentrations (B). The percentage of apoptotic cells was determined using the annexin-V binding assay and flow cytometry. Data are mean ± SE of three separate experiments. *P < 0.05 compared with control. ***P < 0.001 compared with control; control is 0 Gy (A) and 3000 nM folate (B).

Fig. 3
Figure 3. DNA damage score from the alkaline Comet assay as described in Materials and Methods. A) Cells were embedded in agarose, and then irradiated. B) Cells were harvested after 8 days of culture in varying folate concentrations. Data are mean ± SE of three separate experiments. *P < 0.05 compared with control. ***P < 0.001 compared with control; control is 0 Gy (A) and 3000 nM folate (B).

Fig. 4
Figure 4. Cell cycle distribution was measured by propidium iodine staining and flow cytometry. A) Cells were harvested 24 h after irradiation. ***P < 0.001 for the G2/M phase compared with 0 Gy. B) Cells were harvested after 8 days of culture in varying folate concentrations. ***P < 0.001 for the S phase compared with 3000 nM. Data are mean ± SE of three separate experiments.
Copyright © 2022 FDOKUMEN




















