
Microbial Transformation of Steroids: Current Trends in Cortical Side Chain Cleavage

Molecular Microbiology (2003)
48
(4), 947–958
© 2003 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology 1365-2958Blackwell Publishing Ltd, 200348
4947958
Original Article
Fodrin cleavage by PetA. Canizalez-Roman and F. Navarro-Garcia
Accepted 29 January, 2003. *For correspondence. [email protected]; Tel. (
+
52) 5 5747 3990; Fax (
+
52)5 5747 7081.
Fodrin CaM-binding domain cleavage by Pet from enteroaggregative
Escherichia coli
leads to actin cytoskeletal disruption
Adrián Canizalez-Roman andFernando Navarro-García*
Department of Cell Biology, CINVESTAV-IPN, Ap. Postal 14–740, 07000 México, DF, Mexico.
Summary
We have previously shown that the plasmid-encodedtoxin (Pet) of enteroaggregative
Escherichia coli
produces cytotoxic and enterotoxic effects. Pet-intoxicated epithelial cells reveal contraction of thecytoskeleton and loss of actin stress fibres. Peteffects require its internalization into epithelial cells.We have also shown that Pet degrades erythroidspectrin. Pet delivery within the intestine suggeststhat Pet may degrade epithelial fodrin (non-erythroidspectrin). Here we demonstrate that Pet has affinityfor
aaaa
-fodrin (formally named
aaaa
II spectrin)
in vitro
and
in vivo
and cleaves epithelial fodrin, causing its redis-tribution within the cells. When Pet has produced itscytoskeletal effects, fodrin is found in intracellularaggregates as membrane blebs. Pet cleaves recombi-nant GST-fodrin, generating two breakdown productsof 37 and 72 kDa. Sequencing of the 37 kDa fragmentdemonstrated that the cleavage site occurred withinfodrin’s 11th repetitive unit between M
1198
and V
1199
, inthe calmodulin binding domain. Site-directedmutagenesis of these amino acids prevented fodrindegradation by Pet. Pet also cleaves epithelial fodrinfrom cultured Pet-treated cells. A mutant in the Petserine protease motif was unable to cause fodrinredistribution or to cleave GST-fodrin. This is the firstreport showing cleavage of
aaaa
-fodrin by a bacterialprotease. Cleavage occurs in the middle of the calm-odulin binding domain, which leads to cytoskeletondisruption.
Introduction
Enteroaggregative
Escherichia coli
(EAEC) has beenassociated with persistent infant diarrhoea, especially indeveloping countries (Bhan
et al
., 1989; Cravioto
et al
.,
1991; Smith
et al
., 1997). We have shown that a 104 kDaEAEC protein, termed Pet (plasmid-encoded toxin), isrequired for EAEC-induced damage to human intestinalmucosa (Henderson
et al
., 1999). Plasmid-encoded toxinis a member of the autotransporter class of secreted pro-teins (Eslava
et al
., 1998) and, together with Tsh, EspP,EspC, Pic, SigA, Hbp, Sat and SepA proteins, comprisesthe serine protease autotransporter of
Enterobacteri-aceae
(SPATE) subfamily (Henderson and Nataro, 2001).The defining feature of autotransporters is their self-contained secretion system (Henderson
et al
., 1998). Petcauses raises in Isc (short-circuit current) and decreasesin electrical resistance of rat jejunum mounted in theUssing chamber, an effect that is accompanied bymucosal damage, increased mucus release, exfoliation ofcells and development of crypt abscesses (Navarro-Garcia
et al
., 1998). Pet appears to be a cytoskeleton-altering toxin, because it induces contraction of thecytoskeleton, loss of actin stress fibres and release offocal contacts in HEp-2 and HT29/C1 cell monolayers,followed by complete cell rounding and detachment(Navarro-Garcia
et al
., 1999). We have also shown thatPet cytotoxicity and enterotoxicity depend on Pet’s serineprotease activity, as both effects are inhibited by phenyl-methylsulphonyl fluoride (PMSF) and are not induced byPet S260I, which is mutated in the catalytic serine andthereby lacks
in vitro
protease activity (Navarro-Garcia
et al
., 1999).Recently, we have also shown that Pet enters the
eukaryotic cell and that trafficking through the vesicularsystem appears to be required for the induction of cyto-pathic effects (Navarro-Garcia
et al
., 2001). Moreover, thePet serine protease motif is the main requisite for thecytopathic effects, as the internalization assays haveshown that Pet and mutant S260I are found inside epithe-lial cells, but that only native Pet produces cytopathiceffects (Navarro-Garcia
et al
., 2001). All these data sug-gest an intracellular target for Pet. Moreover, we havefound that Pet produces degradation of erythroid spectrin.Spectrin degradation by Pet depends also upon theserine-protease motif and generates a 120 kDa break-down product after 12 h of incubation (Villaseca
et al
.,2000). Fodrin is a ubiquitous cytoskeletal protein involvedwith linking integral membrane proteins to cortical actinfilaments, as well as organizing receptor domains and

948
A. Canizalez-Roman and F. Navarro-Garcia
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
48
, 947–958
possibly the control of vesicle traffic at the plasma mem-brane (Coleman
et al
., 1989). This linkage appears to bepost-translationally regulated and several mechanismsthat might control this process have been identified(Mische and Morrow, 1988). Two events appear to beimportant for this regulation: binding of calmodulin to the
a
subunit of fodrin (Harris
et al
., 1988, 1989) and theproteolysis of fodrin by calcium-dependent neutral pro-teases (Lynch and Baudry, 1984).
Because Pet is delivered into the intestine during theEAEC–epithelial cell interaction and fodrin is a non-erythroid spectrin that is found in epithelial cells, we haveinvestigated further the interaction between Pet andfodrin. Pet showed affinity to
a
-fodrin (formally named
a
IIspectrin)
in vitro
and
in vivo
. In addition, Pet was able tocleave
a
-fodrin and the specific cleavage site occurredwithin the calmodulin binding domain of fodrin. Fodrindegradation
in vitro
and
in vivo
correlated with fodrin redis-tribution inside of epithelial cells. These events lead tocytoskeleton disruption and cell exfoliation.
Results
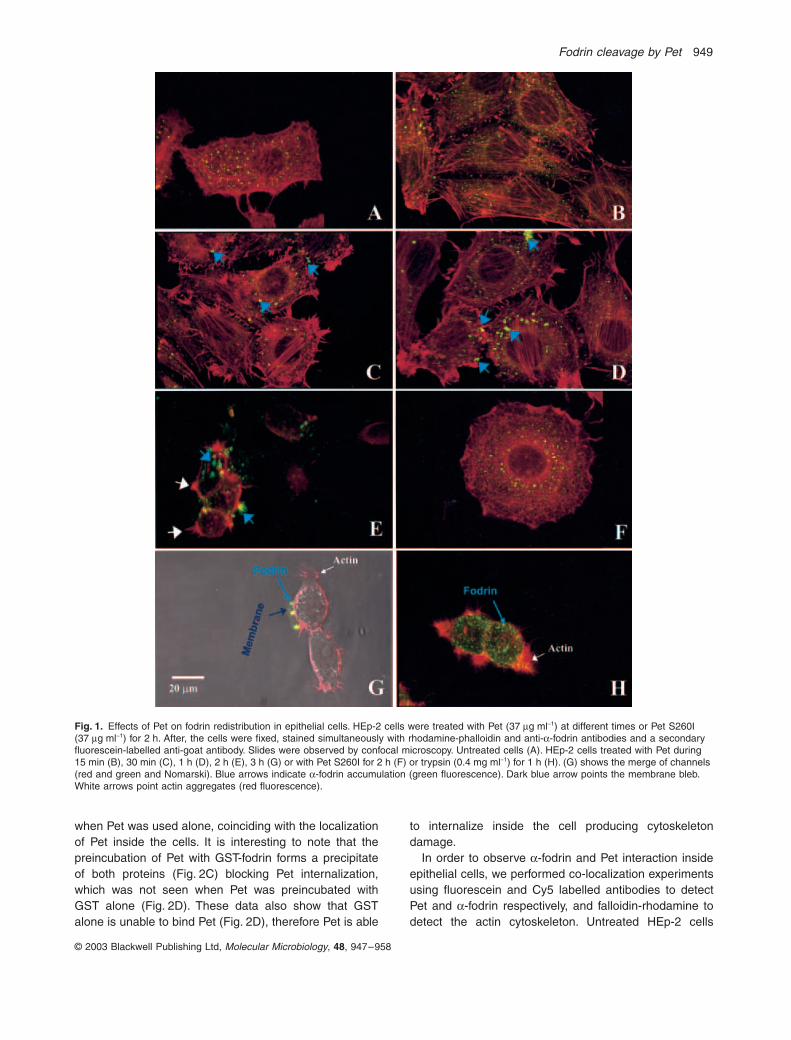
Effects of Pet on fodrin in living epithelial cells
Recently, we have shown that Pet degrades spectrin fromerythrocyte membranes (Villaseca
et al
., 2000). Thereforewe hypothesized that Pet would degrade epithelial fodrin.To address this issue, HEp-2 cells were incubated withPet for various times, and the actin cytoskeleton and
a
-fodrin were stained. Fixed control cells immunofluores-cently stained with anti-
a
II spectrin (anti-fodrin) antibodiesresulted in cytoplasmic punctate fluorescence along theepithelial cell, as reported in other cell types (Younes
et al
., 1989; Vaaraniemi
et al
., 1997; Ziemnicka-Kotula
et al
., 1998), whereas staining with falloidin-rhodamineshowed the classical array of actin-containing stress fibres(Fig. 1A). When the cells were treated with Pet for 15 min,
a
-fodrin and F-actin distribution were not affected andwere similar to the control cells (Fig. 1B). A change in
a
-fodrin distribution was observed after 30 min of Pet–epi-thelial cell interaction, which consisted of an increase inthe size of the
a
-fodrin fluorescent spots (Fig. 1C, bluearrows). After 1 h of interaction, the increase of the
a
-fodrin fluorescent spots was more evident but without aclearly effect on the actin cytoskeleton (Fig. 1D). After 2 hof interaction,
a
-fodrin was redistributed to form intracel-lular aggregates (blue arrows) and the damage to theactin cytoskeleton was observed (Fig. 1E, white arrows),as reported previously (Navarro-Garcia
et al
., 1999). After3 h,
a
-fodrin was focalized in few major aggregates andmost of the remaining small spots were lost, and the cellswere rounded and lost their stress fibres (Fig. 1G). Theintracellular aggregates of
a
-fodrin were found in mem-
brane blebs inside of the cells treated with native Pet(Fig. 1G). It is interesting to note that after 3 h the aggre-gates into blebs contained either actin or
a
-fodrin, there-fore there was no co-localization between them (Fig. 1G),suggesting a disassociation between
a
-fodrin and actin.However, the mutant in the serine protease motif(PetS260I) did not produce this characteristic redistribu-tion of
a
-fodrin nor actin in the HEp-2 cells (Fig. 1F) andtheir distributions were similar to the untreated cells. Onthe other hand, trypsin, which is well known to be unableto degrade fodrin because is unable to enter the cell, hasno effect on the fodrin distribution and there is not intrac-ellular fodrin aggregates, even when the cells are roundedand there is blebs containing F-actin (Fig. 1H).
All these data suggest damage to epithelial
a
-fodrin
invivo
by Pet, which caused a rearrangement of
a
-fodrinfollowed by a disruption of the actin cytoskeleton (Fig. 1).
Interaction of Pet with
a
-fodrin and actin
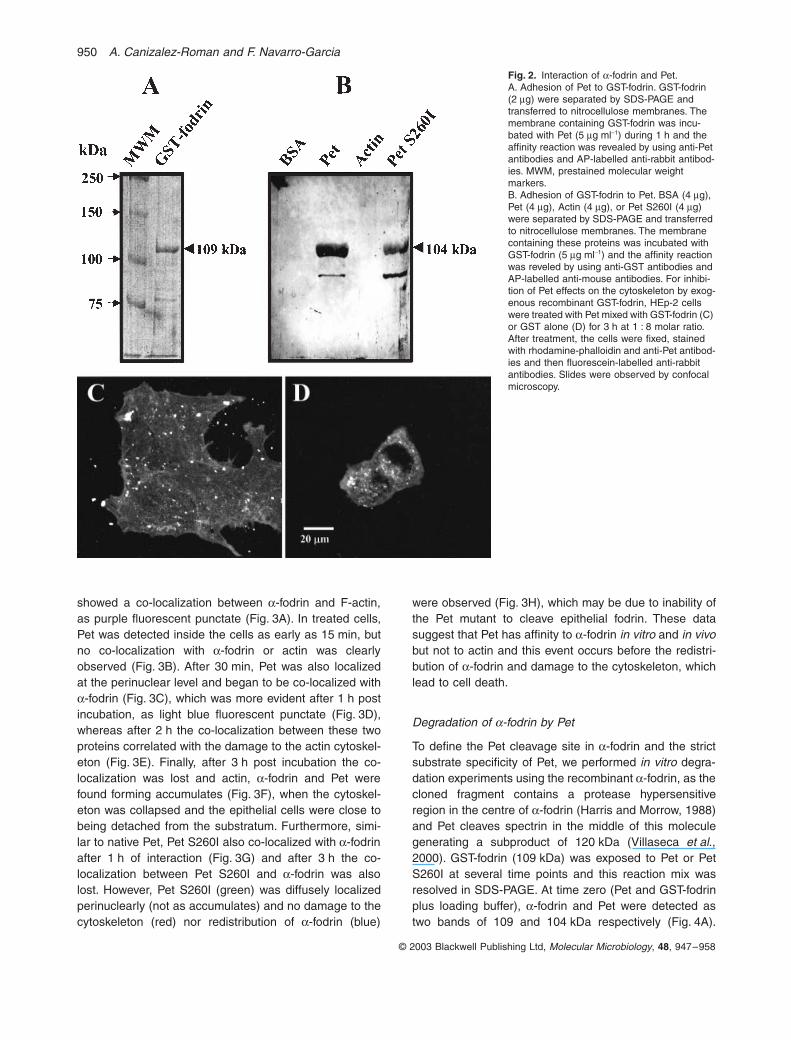
Because Pet disrupts fodrin and actin organization, weexpected an affinity of Pet to
a
-fodrin. To investigate thispossibility, we performed binding experiments betweenPet and either
a
-fodrin or actin by overlay experiments.When a recombinant
a
-fodrin containing repeats 8–14fused to GST (109 kDa) was immobilized in nitrocellulosemembranes, we found adhesion of Pet to the 109 kDaband developed with anti-Pet antibodies (Fig. 2A) but notto GST alone (data not shown), suggesting that Pet hasaffinity to
a
-fodrin. Moreover, when Pet or the mutant PetS260I were immobilized to the membrane,
a
-fodrin wasalso found to bind to the 104 kDa protein (Pet or PetS260I) visualized by anti-GST antibodies (Fig. 2B). Asexpected,
a
-fodrin had not affinity to actin, or to otherirrelevant protein such as BSA (Fig. 2B), because we areusing
a
-fodrin and this chain not posses the actin-bindingdomain.
To further understand Pet–fodrin interaction, we per-formed precipitation assays by incubating HEp-2 cells for3 h with Pet previously mixed with exogenous GST-fodrin.If Pet has affinity by GST-fodrin, both proteins must beprecipitated and Pet will be unable to get inside the cellsproducing no cytoskeleton damage. The cells werestained using anti-Pet antibodies and a secondary anti-body labelled with fluorescein and the actin cytoskeletonwas evidenced by using TRITC-falloidin. GST-fodrin wasable to precipitate Pet to 1:8 ratio (Pet:GST-fodrin), pro-ducing a complete inhibition of Pet effects on the actincytoskeleton (Fig. 2C), similar to that observed when Petwas preincubated with PMSF (data not shown). The inhi-bition effect coincided with the lack of fluorescence (Pet)inside of the cells (Fig. 2C). Whereas GST alone (1:8,Pet:GST) was unable to block Pet effects (Fig. 2D) andthe cytoskeletal damage was similar to that observed
Fodrin cleavage by Pet
949
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
48
, 947–958
when Pet was used alone, coinciding with the localizationof Pet inside the cells. It is interesting to note that thepreincubation of Pet with GST-fodrin forms a precipitateof both proteins (Fig. 2C) blocking Pet internalization,which was not seen when Pet was preincubated withGST alone (Fig. 2D). These data also show that GSTalone is unable to bind Pet (Fig. 2D), therefore Pet is able
to internalize inside the cell producing cytoskeletondamage.
In order to observe
a
-fodrin and Pet interaction insideepithelial cells, we performed co-localization experimentsusing fluorescein and Cy5 labelled antibodies to detectPet and
a
-fodrin respectively, and falloidin-rhodamine todetect the actin cytoskeleton. Untreated HEp-2 cells
Fig. 1.
Effects of Pet on fodrin redistribution in epithelial cells. HEp-2 cells were treated with Pet (37
m
g ml
-
1
) at different times or Pet S260I (37
m
g ml
-
1
) for 2 h. After, the cells were fixed, stained simultaneously with rhodamine-phalloidin and anti-
a
-fodrin antibodies and a secondary fluorescein-labelled anti-goat antibody. Slides were observed by confocal microscopy. Untreated cells (A). HEp-2 cells treated with Pet during 15 min (B), 30 min (C), 1 h (D), 2 h (E), 3 h (G) or with Pet S260I for 2 h (F) or trypsin (0.4 mg ml
-
1
) for 1 h (H). (G) shows the merge of channels (red and green and Nomarski). Blue arrows indicate
a
-fodrin accumulation (green fluorescence). Dark blue arrow points the membrane bleb. White arrows point actin aggregates (red fluorescence).
950
A. Canizalez-Roman and F. Navarro-Garcia
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
48
, 947–958
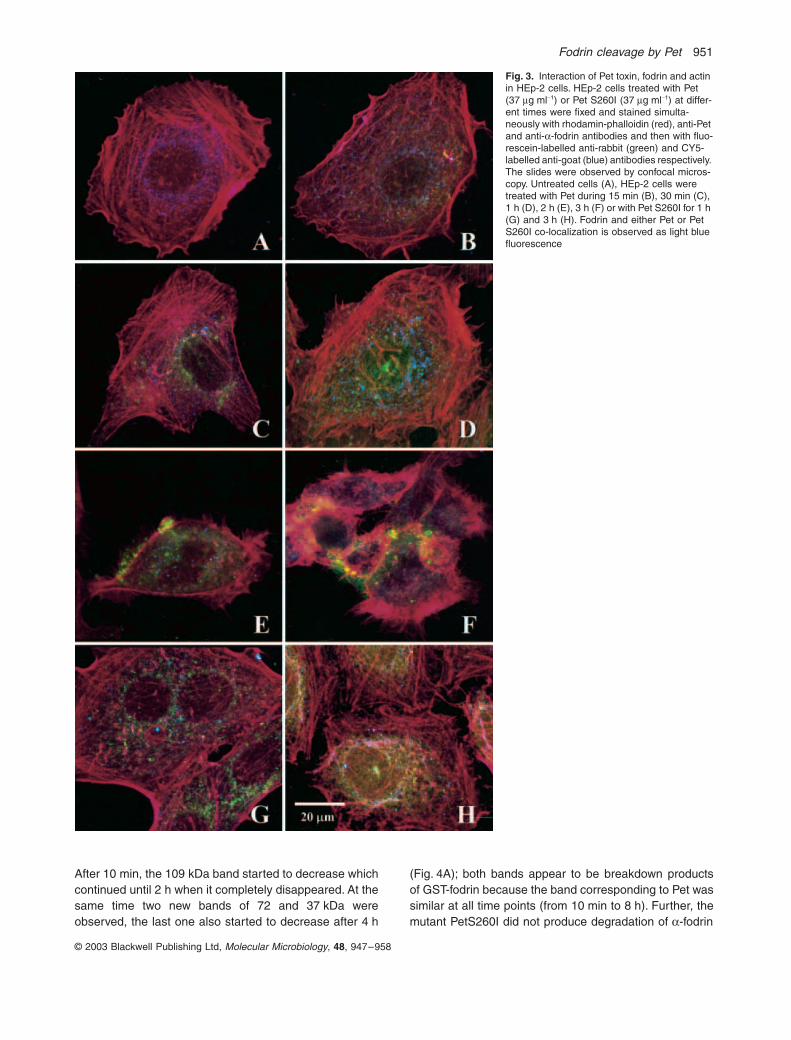
showed a co-localization between
a
-fodrin and F-actin,as purple fluorescent punctate (Fig. 3A). In treated cells,Pet was detected inside the cells as early as 15 min, butno co-localization with
a
-fodrin or actin was clearlyobserved (Fig. 3B). After 30 min, Pet was also localizedat the perinuclear level and began to be co-localized with
a
-fodrin (Fig. 3C), which was more evident after 1 h postincubation, as light blue fluorescent punctate (Fig. 3D),whereas after 2 h the co-localization between these twoproteins correlated with the damage to the actin cytoskel-eton (Fig. 3E). Finally, after 3 h post incubation the co-localization was lost and actin,
a
-fodrin and Pet werefound forming accumulates (Fig. 3F), when the cytoskel-eton was collapsed and the epithelial cells were close tobeing detached from the substratum. Furthermore, simi-lar to native Pet, Pet S260I also co-localized with
a
-fodrinafter 1 h of interaction (Fig. 3G) and after 3 h the co-localization between Pet S260I and
a
-fodrin was alsolost. However, Pet S260I (green) was diffusely localizedperinuclearly (not as accumulates) and no damage to thecytoskeleton (red) nor redistribution of
a
-fodrin (blue)
were observed (Fig. 3H), which may be due to inability ofthe Pet mutant to cleave epithelial fodrin. These datasuggest that Pet has affinity to
a
-fodrin
in vitro
and
in vivo
but not to actin and this event occurs before the redistri-bution of
a
-fodrin and damage to the cytoskeleton, whichlead to cell death.
Degradation of
a
-fodrin by Pet
To define the Pet cleavage site in
a
-fodrin and the strictsubstrate specificity of Pet, we performed
in vitro
degra-dation experiments using the recombinant
a
-fodrin, as thecloned fragment contains a protease hypersensitiveregion in the centre of
a
-fodrin (Harris and Morrow, 1988)and Pet cleaves spectrin in the middle of this moleculegenerating a subproduct of 120 kDa (Villaseca
et al
.,2000). GST-fodrin (109 kDa) was exposed to Pet or PetS260I at several time points and this reaction mix wasresolved in SDS-PAGE. At time zero (Pet and GST-fodrinplus loading buffer),
a
-fodrin and Pet were detected astwo bands of 109 and 104 kDa respectively (Fig. 4A).
Fig. 2.
Interaction of
a
-fodrin and Pet.A. Adhesion of Pet to GST-fodrin. GST-fodrin (2
m
g) were separated by SDS-PAGE and transferred to nitrocellulose membranes. The membrane containing GST-fodrin was incu-bated with Pet (5
m
g ml
-
1
) during 1 h and the affinity reaction was revealed by using anti-Pet antibodies and AP-labelled anti-rabbit antibod-ies. MWM, prestained molecular weight markers.B. Adhesion of GST-fodrin to Pet. BSA (4
m
g), Pet (4
m
g), Actin (4
m
g), or Pet S260I (4
m
g) were separated by SDS-PAGE and transferred to nitrocellulose membranes. The membrane containing these proteins was incubated with GST-fodrin (5
m
g ml
-
1
) and the affinity reaction was reveled by using anti-GST antibodies and AP-labelled anti-mouse antibodies. For inhibi-tion of Pet effects on the cytoskeleton by exog-enous recombinant GST-fodrin, HEp-2 cells were treated with Pet mixed with GST-fodrin (C) or GST alone (D) for 3 h at 1 : 8 molar ratio. After treatment, the cells were fixed, stained with rhodamine-phalloidin and anti-Pet antibod-ies and then fluorescein-labelled anti-rabbit antibodies. Slides were observed by confocal microscopy.
Fodrin cleavage by Pet
951
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
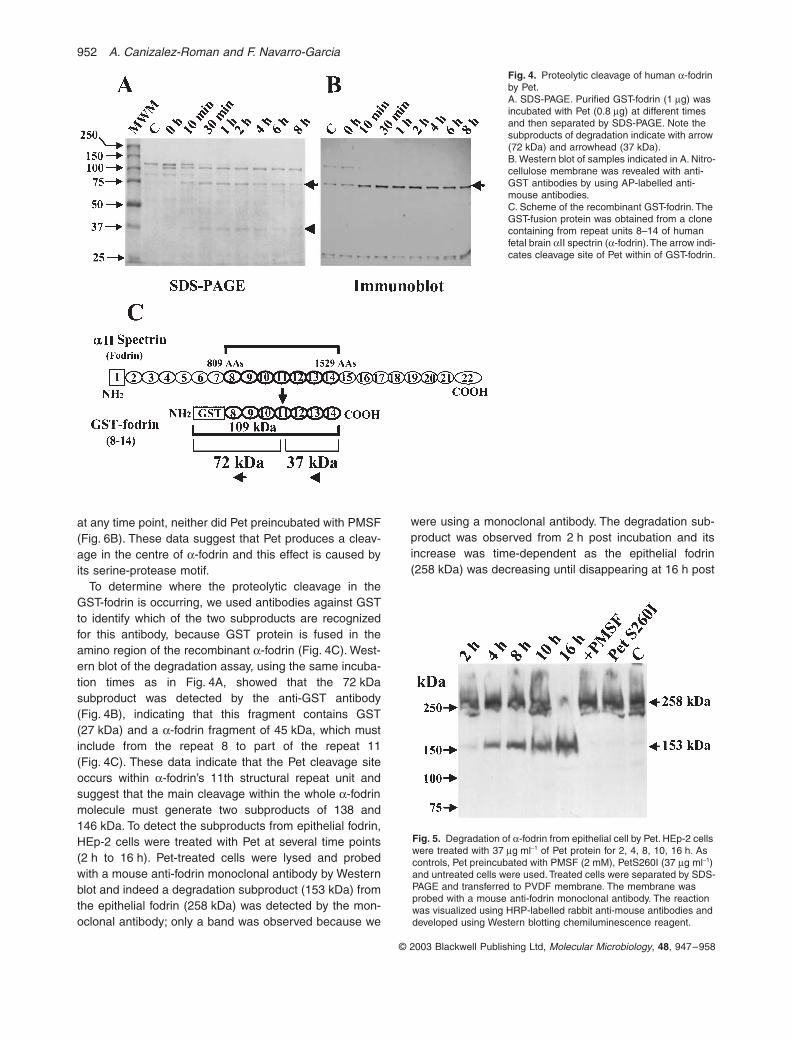
After 10 min, the 109 kDa band started to decrease whichcontinued until 2 h when it completely disappeared. At thesame time two new bands of 72 and 37 kDa wereobserved, the last one also started to decrease after 4 h
(Fig. 4A); both bands appear to be breakdown productsof GST-fodrin because the band corresponding to Pet wassimilar at all time points (from 10 min to 8 h). Further, themutant PetS260I did not produce degradation of a-fodrin
Fig. 3. Interaction of Pet toxin, fodrin and actin in HEp-2 cells. HEp-2 cells treated with Pet (37 mg ml-1) or Pet S260I (37 mg ml-1) at differ-ent times were fixed and stained simulta-neously with rhodamin-phalloidin (red), anti-Pet and anti-a-fodrin antibodies and then with fluo-rescein-labelled anti-rabbit (green) and CY5-labelled anti-goat (blue) antibodies respectively. The slides were observed by confocal micros-copy. Untreated cells (A), HEp-2 cells were treated with Pet during 15 min (B), 30 min (C), 1 h (D), 2 h (E), 3 h (F) or with Pet S260I for 1 h (G) and 3 h (H). Fodrin and either Pet or Pet S260I co-localization is observed as light blue fluorescence
952 A. Canizalez-Roman and F. Navarro-Garcia
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
at any time point, neither did Pet preincubated with PMSF(Fig. 6B). These data suggest that Pet produces a cleav-age in the centre of a-fodrin and this effect is caused byits serine-protease motif.
To determine where the proteolytic cleavage in theGST-fodrin is occurring, we used antibodies against GSTto identify which of the two subproducts are recognizedfor this antibody, because GST protein is fused in theamino region of the recombinant a-fodrin (Fig. 4C). West-ern blot of the degradation assay, using the same incuba-tion times as in Fig. 4A, showed that the 72 kDasubproduct was detected by the anti-GST antibody(Fig. 4B), indicating that this fragment contains GST(27 kDa) and a a-fodrin fragment of 45 kDa, which mustinclude from the repeat 8 to part of the repeat 11(Fig. 4C). These data indicate that the Pet cleavage siteoccurs within a-fodrin’s 11th structural repeat unit andsuggest that the main cleavage within the whole a-fodrinmolecule must generate two subproducts of 138 and146 kDa. To detect the subproducts from epithelial fodrin,HEp-2 cells were treated with Pet at several time points(2 h to 16 h). Pet-treated cells were lysed and probedwith a mouse anti-fodrin monoclonal antibody by Westernblot and indeed a degradation subproduct (153 kDa) fromthe epithelial fodrin (258 kDa) was detected by the mon-oclonal antibody; only a band was observed because we
were using a monoclonal antibody. The degradation sub-product was observed from 2 h post incubation and itsincrease was time-dependent as the epithelial fodrin(258 kDa) was decreasing until disappearing at 16 h post
Fig. 4. Proteolytic cleavage of human a-fodrin by Pet.A. SDS-PAGE. Purified GST-fodrin (1 mg) was incubated with Pet (0.8 mg) at different times and then separated by SDS-PAGE. Note the subproducts of degradation indicate with arrow (72 kDa) and arrowhead (37 kDa).B. Western blot of samples indicated in A. Nitro-cellulose membrane was revealed with anti-GST antibodies by using AP-labelled anti-mouse antibodies.C. Scheme of the recombinant GST-fodrin. The GST-fusion protein was obtained from a clone containing from repeat units 8–14 of human fetal brain aII spectrin (a-fodrin). The arrow indi-cates cleavage site of Pet within of GST-fodrin.
Fig. 5. Degradation of a-fodrin from epithelial cell by Pet. HEp-2 cells were treated with 37 mg ml-1 of Pet protein for 2, 4, 8, 10, 16 h. As controls, Pet preincubated with PMSF (2 mM), PetS260I (37 mg ml-1) and untreated cells were used. Treated cells were separated by SDS-PAGE and transferred to PVDF membrane. The membrane was probed with a mouse anti-fodrin monoclonal antibody. The reaction was visualized using HRP-labelled rabbit anti-mouse antibodies and developed using Western blotting chemiluminescence reagent.

Fodrin cleavage by Pet 953
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
incubation (Fig. 5). Interestingly, when Pet was preincu-bated with PMSF or the serine protease mutant PetS260Iwas used, there was not degradation of the epithelialfodrin even after 16 h post incubation and in both casesthe 258 kDa band was similar to that in untreated cell(Fig. 5). The preservation of the 258 kDa band coincidedwith the preservation of the actin cytoskeleton in theseepithelial cells (data not shown).
In order to locate the cleavage site within repeat 11 ofa-fodrin, we determined the N-terminal amino acidsequence of the 37 kDa subproduct from recombinanta-fodrin, since this fragment does not contain the GSTN-terminal amino acid (Fig. 4C). As shown in Fig. 4A,the 37 kDa fragment was more prominent at 1 h postincubation with Pet; thus we sequenced it, and N-termi-nal amino acid sequence from this fragment was VHT-VATFNSIKE (University of California, San Diego,Division of Biology Protein Sequencer Facility), indicat-ing that the Pet cleavage site in human a-fodrin occurswithin a-fodrin’s 11th repetitive unit in the proteasehypersensitivity region between M1198 and V1199, in thehelix C and inside of the calmodulin binding domain(Fig. 6A).
To determine the cleavage site specificity of Pet withina-fodrin, we performed site-directed mutagenesis tochange M1198 and V1199 to arginine and glycine at thesepositions, respectively. This a-fodrin mutant (GST-fodrinM1198R-V1199G) was incubated with Pet in the degrada-tion assay. Pet was unable to cleave GST-fodrin M1198R-V1199G, even after 4 h of interaction, because at this timethe protein band of the a-fodrin mutant (109 kDa) wasunalterable and similar to the zero time or to untreated a-fodrin mutant (Fig. 6B). In contrast, the native GST-fodrinwas clearly degraded by Pet after 1 h of interaction,whereas as we mentioned above neither Pet S260I norPMSF-preincubated Pet were able to cleave native GST-fodrin (Fig. 6B). All these data indicate that Pet cleaves a-fodrin specifically between M1198 and V1199 and there is noalternative cleavage site. On the other hand, the serineprotease motive is the active site to cleave a-fodrin in thisspecific cleavage site.
Discussion
Our previous data have shown that all cell damagecaused by Pet depends upon its serine-protease motif,including the enterotoxic and cytophatic effects as wellas the cytoskeletal damage (Navarro-Garcia et al.,1999). Recently, we have also found that Pet internaliza-tion into epithelial cells is a requisite to produce disrup-tion of the cytoskeleton (Navarro-Garcia et al., 2001). Allthese lead us to suggest that Pet is a cytoskeleton-alter-ing toxin and its protease activity is involved in each ofthe observed phenotypes. Here, we show that the
cytoskeletal damage by Pet on epithelial cells is due toa-fodrin (formally named aII spectrin) degradation. a-fodrin degradation leads to cytoskeleton disruption, asfodrin maintains the connection of the plasma mem-brane to the cytoskeleton (Bennett and Gilligan, 1993).Furthermore, in epithelial cells, the lower portion of theactin filament bundle in the microvillus core is anchoredin the specialized cortex at the apex of the intestinal epi-thelial cell, which contains a dense network of fodrinmolecules that overlies a layer of intermediate filaments(Djabali, 1999).
Fodrin, a multifunctional protein with spectrin-likeproperties, is a major component of the cortical cytosk-eleton of most eukaryotic cells (Morrow, 1989). Fodrin,like spectrin, is a rod-shaped protein consisting of a andb subunits, which form heterodimers aligned in a side-to-side manner. Fodrin heterodimers then further associ-ate end-on-end to form tetramers, which cross-link actinfilaments at their ends (Harris and Morrow, 1990).Fodrin possesses binding sites for many proteins,including actin (Harris and Morrow, 1990), calmodulin(Harris et al., 1988) and ankyrin among others. Proteoly-sis of aII spectrin (a-fodrin) in non-erythroid cells andspectrin in erythrocytes occurs under physiological andpathophysiological conditions. Interestingly, until now, allthe reports about spectrin degradation by pathogenshave been related to erythrocytes not to epithelial cellsor non-erythroid cells, such as Tricomonas vaginalis(Fujino et al., 1998) or the intracellular protozoans Plas-modium falciparum and Plasmodium bergei (Deguercyet al., 1990).
On the other hand, in physiological conditions, pro-teolysis of fodrin by calcium-dependent proteases hasbeen observed during several processes, calpain isactivated in various necrotic and apoptotic conditions,whereas caspase 3 is only activated in neuronal apop-tosis (Wang, 2000). Thus, aII spectrin is degraded to a120 kDa fragment in apoptotic neurons but not innecrotic neurons (Nath et al., 1996). Calpain-mediateda-fodrin breakdown to a 150 kDa and 145 kDa doubletis not only present in necrotic neuronal death, but alsoin most forms of neuronal apoptosis (Nath et al., 1996;Wang et al., 1998). In contrast, caspase-mediatedcleavage of aII spectrin generates a breakdown productof 120 kDa (and also caspase-specific 150 kDa frag-ment) occurring exclusively in neuronal apoptosis(Schulz et al., 1996; Wang, 2000). Using a recombinantaII spectrin (a-fodrin), we are showing here that Petgenerates suproducts of 72 and 37 kDa at the first10 min of interaction, considering that we are using theclone 18531 representing repeats 8–14 of human aIIspectrin, which contains a protease hypersensitiveregion at the centre of aII spectrin, plus GST(109 kDa). The Western blot using anti-GST showed

954 A. Canizalez-Roman and F. Navarro-Garcia
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
that the 72 kDa fragment contains the amino terminalsequence (GST fused to repeat 8), indicating that thecleavage site occurs within the repeat 11, and suggest-ing that the breakdown subproducts in the whole mole-cule are of 138 and 146 kDa. These data wasconfirmed by detecting by a monoclonal antibody asubproduct of 153 kDa from epithelial a-fodrin of Pet-treated HEp-2 cells by using Western blot.
Fodrin distribution, in our cell culture model, wasirregular because we were using the non-polarized epi-thelial cells, HEp-2 cells (laryngeal), which had been
reported in non-polarized MDCK (Younes et al., 1989).However Pet produced a redistribution of this epithelialfodrin characterized by large intracellular aggregates ofa-fodrin, starting about 1 h post inoculation. Further-more, Pet caused plasma membrane blebbing, some ofthem containing either actin or a-fodrin. Interestingly,plasma membrane blebbing, an early apoptotic pheno-type, has been proposed to result as a consequence ofthe loss or cleavage of intact aII spectrin (Martin et al.,1995). Of note, the reported degradation of a-fodrin inapoptotic or necrotic events occur around 8–10 h (Nath
Fig. 6. Specificity of Pet cleavage site on a-fodrin into calmodulin binding domain.A. Pet cleavage site into 11th repeat unit of a-fodrin. Sequencing of the 37 kDa subproduct showed that the cleavage site is localized within 11th repeat unit in the helix C inside calmodulin binding domain, between M1198 and V1199.B. Degradation assay of a-fodrin mutant by Pet. Recombinant GST-fodrin or GST-fodrin mutant (fodrin M1198R-V1199G) (1.8 mg) were incubated with Pet (1.5 mg) at different times and recombinant GST-fodrin was incubated for 1 h with Pet S260I (1.5 mg) or Pet preincubated with PMSF. These reaction mixes were separated by SDS-PAGE. The arrows indicate the degradation subproducts from GST-fodrin.

Fodrin cleavage by Pet 955
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
et al., 1996), as it was observed using Pet; because aWestern blot methodology from unsynchronized lysedcells where Pet internalization and fodrin degradationdid not occur at the same time in all the cells, thus thefodrin degradation is completely observed only after 8–10 h when all the cells were affected. Whereas purifieda-fodrin degradation by Pet occurs at the first 10 min invitro and it is complete degraded after 2 h, because allfodrin was available to Pet. On the other hand, by con-focal microscopy is possible to find the first cells beingaffected and the a-fodrin redistribution in living epithelialcells can be observed between 1 h and 3 h and it ispossible to find plasma membrane blebs containinga-fodrin.
Unlike most proteases, Pet toxin displays unusuallystrict substrate specificity, because actin, a-actinin, vin-culin, cadherin, catenin, b1 integrin and FAK amongother proteins related to focal adhesion appear not tobe substrates for Pet (F. Navarro-Garcia, unpubl. data).The substrate specificity appears to be a characteristicof proteases which degrade fodrin; thus, calpainscleave only one or two bonds in proteins with hundredof potential sites. Calpain cleavage site specificity hasbeen hard to study as the affinity of calpain for syn-thetic peptides is typically 10–100 times weaker than forproteins (Stabach et al., 1997); thereby a recombinantprotein as substrate is an excellent model. Calpaincleaves aII spectrin between V1176 and G1177 andcaspase 3 cleavage site is between D1185 and S1186.Calpain subsequently cleaves this region again,between G1230 and S1231, producing a slightly smallerbreakdown subproduct of 145 kDa. In addition, caspase3 cleaves in repeat 13, between D1478 and S1479, produc-ing the apoptosis-specific breakdown subproduct of120 kDa (Wang, 2000). Our finding shows that Petcleavage site is between M1198 and V1199, localized alsowithin a-fodrin’s 11th repetitive unit in the protease(hypersensitive region); specifically in the helix C, insideof the calmodulin binding domain. In fact, mutations inthese two amino acids completely abolished a-fodrindegradation by Pet, even after longer incubation times.Thus, this is the first report showing cleavage of a-fodrin by a foreign bacterial protease (Pet) at the middleof the calmodulin binding domain. The biological mean-ing of this event is unknown, but Harris and Morrow(1990) have shown that calmodulin and calpain I co-ordinately regulate the interaction of fodrin with actin.Moreover, they also showed that calmodulin disruptsfodrin tetramer after calpain I cleavage of the a subunit(Harris and Morrow, 1990). Thus, the cytoskeletal alter-ations produced by Pet, which are characterized by lossof actin stress fibres, contraction of cytoskeleton andcell detachment (Navarro-Garcia et al., 1999), are aconsequence of the disruption of fodrin and actin fila-
ment connection specifically due to a-fodrin cleavage byPet. This is also the first described action mechanismamong members of the SPATE subfamily. Autotransport-ers comprise a special group of virulence-associatedproteins secreted by Gram-negative bacteria that exhibitdiverse biological functions (Henderson et al., 1998;Henderson and Nataro, 2001).
Cytoskeleton contraction due to a-fodrin cleavage byPet may explain our previous observations and frommany other investigators who showed cell damage byEAEC. The enterotoxic effects produced by Pet(Navarro-Garcia et al., 1998) could be caused by thedisruption of the membrane skeleton because a-fodrinwas found to form a macromolecular complex with epi-thelial sodium channels (Zuckerman et al., 1999), andepithelial channels mediate entry of Na from the luminalfluid into cells during the first stage of electrogenic tran-sepithelial Na transport across Na-reabsorbing epithelia(Garty and Palmer, 1997), this explains the diarrhoealpathogenesis caused by EAEC. The damage of epithe-lial cells by EAEC strains in intestinal necropsy of Mexi-can children (Eslava et al., 1993), in vitro organ culturemodel (Hicks et al., 1996), in T84 cultured cells (Nataroet al., 1996), or directly by Pet in HEp-2 and HT29 cul-tured cells (Navarro-Garcia et al., 1999), in vitro organculture (Henderson et al., 1999), or intestinal prepara-tion mounted in Ussing chambers (Navarro-Garcia et al.,1998) are due to cytoskeletal disruption and cell detach-ment produced by a-fodrin disarrangement. Interestingly,both enterotoxic and cytotoxic effects depended uponthe serine protease motif, the active site used for a-fodrin degradation. Furthermore, Pet internalization isneeded to produce the cytotoxic and cytoskeletal dam-age (Navarro-Garcia et al., 2001), as a-fodrin is an intra-cellular protein. Thus, this mechanism appears to be anew system of cellular damage identified in bacterialtoxin, which includes the internalization of the proteaseto allow finally specific a-fodrin degradation to destroythe cell.
Experimental procedures
Strains and plasmids
The minimal Pet clone pCEFN1 described previously wasconstructed by cloning the pet gene of EAEC strain 042 intothe BamHI/KpnI site of pSPORT1 and expressed in E. coliHB101 (Eslava et al., 1998). HB101(pCEFN1) and (pCEFN2)were used to obtain Pet protein and Pet mutant, and super-natant proteins from the HB101(pSPORT1) were used as acontrol (Navarro-Garcia et al., 1999). Clone 18531 represent-ing bp 2531–4689 of human aII spectrin was kindly providedby Paul A. Stabatch and Jon S. Morrow (Stabach et al.,1997), which was cloned into pBluescript using EcoRI adapt-ers and subcloned into the EcoRI site of the inducible bacte-rial expression vector pGEX-3X (Pharmacia, Uppsala,

956 A. Canizalez-Roman and F. Navarro-Garcia
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
Sweden). The strains were maintained on L-agar or L-brothcontaining 100 mg ml-1 ampicillin.
Preparation of recombinant proteins
Broth cultures from pet minimal clone HB101(pCEFN1) orPet serine protease motif mutant HB101(pCEFN2) were incu-bated overnight at 37∞C and then centrifuged at 7000 g for15 min. The culture supernatant was filtered throughout0.22 mm cellulose acetate membrane filters (Corning), con-centrated 100-fold in an ultrafree centrifugal filter device witha 100-kDa cut-off (Millipore), filter-sterilized again and storedat -20∞C for up to 3 months (Navarro-Garcia et al., 1999).
GST-fodrin was prepared as previously described (Stabachet al., 1997). Briefly, overnight bacterial culture from clone18531 expressed in BL21 were diluted 1:10 in fresh media,grown for 1 h, and then induced for 1–3 h with IPTG beforeharvesting by centrifugation. Lysis was achieved by gentlyagitating the bacterial pellet for 30 min at 4∞C in 100 ml of40 mM Tris-HCl, pH 7.5, 100 mM NaCl, DTT 1 mM,Lysozyme 1 mg ml-1 and a cocktail of protease inhibitors(Complete™, Boehringer Mannheim), after which the sam-ples were frozen at -70∞C and rapidly thawed at 30∞C. TritonX-100 was added to a final concentration of 1% (v/v) and thesample was incubated at 4∞C for an additional 30 min andthen sonicated three times for 10 s each. The 15000 g super-natant of the lysate was affinity absorbed on 20 ml of glu-tathione-Sepharose 4B beads (Pharmacia) at 4∞C andwashed five times with 2 ml of PBS with 1 mM DTT. Thebound peptide was eluted during 10 min at room temperaturein 40 ml of the same buffer containing 10 mM reduced glu-tathione.
Cell culture
HEp-2 cells were propagated in humidified 5%CO2-95% airat 37∞C in Dulbecco’s modified Eagle’s medium (DMEM)supplemented with 5% fetal bovine serum (Hyclone), 1%non-essential amino acids, 5 mM L-glutamine, penicillin (100units ml-1), and streptomycin (100 mg ml-1). The subcultureswere serially propagated after harvesting with 10 mM EDTAand 0.25% trypsin (Gibco) in phosphate-buffered saline solu-tion (PBS, pH 7.4). For experimental use, subconfluent HEp-2 cells were resuspended with EDTA-trypsin, plated intoeight-well LabTek slides (VWR), and allowed to grow to c.60% confluence (about 2 days).
Tissue culture assay
For all experiments, Pet or Pet S260I proteins were diluteddirectly into tissue culture medium (without antibiotics andserum) and added to the target cells at a final volume of250 ml per well (for eight-well LabTek slides). Cells incubatedwith trypsin (0.4 mg ml-1; Gibco BRL) for 1 h were used ascontrol. Following the specified incubation times in humidifiedatmosphere of 5%CO2-95% air at 37∞C, the medium wasaspirated, the cells were washed twice with PBS and pro-cessed for both fluorescence-actin staining (FAS) assay andimmunostaining techniques. The cells were fixed with 2%
formalin/PBS, washed, permeabilized by adding 0.2% TritonX-100/PBS and stained with 0.05 mg ml-1 TRITC-phalloidin(Sigma Chemical) and incubated with anti-Pet (Navarro-Garcia et al., 2001) or/and anti-aII spectrin (C-20) antibodies(Santa Cruz Biotech). The antigen-antibody reaction wasdeveloped using fluorescein-labelled goat anti-rabbit IgG(Zymed Laboratory) and/or Cy5-labelled mouse anti-goat IgG(Jackson Immunoresearch) or fluorescein-labelled donkeyanti-goat IgG (Rockland). Slides were mounted on Gelvatol(Sigma Chemical), covered with a glass cover slide andexamined under a Leica TCS SP2 confocal microscope.
Overlay assay
Four or two micrograms of the proteins (Pet, PetS260I, GST-fodrin, actin or BSA) were separated by 10% SDS-PAGE.These separated proteins were then transferred to Nitrocel-lulose membrane (Bio-Rad) and the nitrocellulose filter wasblocked overnight in blocking buffer (150 mM NaCl, 8 mMNa2HPO4, 2 mM NaH2PO4 (pH 7.3), 2 mM CaCl2 and 5%non-fat dry milk) at 4∞C. Individual strips were then incubatedfor 1 h in buffer B (20 mM Tris-HCl, 150 mM NaCl, 0.1%Tween 20, 2 mM CaCl2 and 5% bovine serum albumin) witheither 5 mg ml-1 toxin or 5 mg ml-1 GST-fodrin. All wash stepsconsisted of three 10 min incubations with blocking buffer.The strips were washed and then incubated for 1 h in block-ing buffer with mouse polyclonal antiserum raised againstGST (1:1000 dilution) or rabbit polyclonal antiserum raisedagainst Pet (1:120 dilution). Following another wash step, thestrips were incubated for 1 h in blocking buffer with a goatanti-rabbit AP-conjugated antibody (1:2000 dilution) or goatanti-mouse AP-conjugated antibody (1:500 dilution) (bothfrom KPL). Following a final wash, binding was detectedusing 1-Step NBT/BCIP substrate (Pierce).
Degradation assay
One microgram or 2 mg of affinity-purified GST-fodrin or GST-fodrin M1198R-V1199G was mixed with an equal volume of2 ¥ digestion buffer (0.3 mM CaCl2, 10 mM DTT) containingvarious amounts of Pet or PetS260I. Reactions were carriedout at 30∞C at several times and stopped by the addition of4 ¥ SDS sample buffer. All samples were analysed by SDS-PAGE and the gels were stained with brilliant Coomassieblue. The N-terminal amino acid sequence to detect thecleavage site was performed according to Protein Sequenc-ing facility at UCSD. Briefly, the Pet-treated protein was blot-ted on a Immobilon-P polyvinylide difluoride (PVDF)membrane (Bio-Rad) stained with Coomassie blue and thesubproduct of 37 kDa band was cut and sent to UCSD forsequencing the N-terminus. The 12 amino acids were soughtin the fodrin sequence (Accession: AAB60364).
Western immunoblot
The proteins from degradation assays were separated by10% SDS-PAGE, and the protein bands obtained were trans-ferred to nitrocellulose membranes (Towbin et al., 1979).Finally, the membranes were probed with anti-GST mouse

Fodrin cleavage by Pet 957
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
polyclonal antibodies to detect GST-fodrin. Thus, the aminoterminal of GST-fodrin was visualized with goat anti-mouseantibodies conjugated with alkaline phosphatase (KPL). Theantigen-antibody reactions were developed using 1-StepNBT/BCIP substrate (Pierce). To detect the a-fodrin subprod-ucts from Pet-treated HEp-2 cells, we followed the (Brownet al., 1999) method. Briefly, Pet-treated cells (40 mg) werelysed, separated by SDS-PAGE and transferred to PVDFmembrane. The membrane was probed using a mouse anti-fodrin monoclonal antibody (Chemicon). The antigen-antibody reaction was visualized using HRP-labelled rabbitanti-mouse IgG (Zymed Laboratory) and developed using aWestern Blotting Chemiluminescence ‘Luminol’ Reagent(Santa Cruz Biotech).
Inhibition assay by using GST-fodrin
HEp-2 cells were treated during 3 h with Pet (37 mg ml-1)mixed with GST-fodrin at different molar ratio (Pet:GST-fodrin,1:1, 1:2, 1:4 and 1:8). The cells were fixed with 2% formalin/PBS, washed, permeabilized by adding 0.2% Triton X-100/PBS and stained with 0.05 mg ml-1 TRITC-phalloidin (SigmaChemical) and incubated with anti-Pet (Navarro-Garcia et al.,2001) and then with a secondary fluorescein-labelled anti-rabbit IgG (Zymed). As controls the cells were treated withPet alone, Pet mixed with GST (1 : 8 molar ratio), or Petpreincubated with PMSF.
Site-directed mutagenesis
Site-directed mutagenesis was performed using theQuikChange™ Site-Directed Mutagenesis Kit fromStratagene. Briefly, a set of synthetic oligonucleotides5¢-GTCTGCTCGTCTGAGGGGTCACACCGTGGC-3¢ and5¢-GCCACGGTGTGACCCCTCAGACGAGCAGAC-3¢ (acce-ssion U26396) was used to produce GST-fodrin M1198R-V1199G, as we had done previously to produce Pet S260I(Navarro-Garcia et al., 1999). The set of oligonucleotidesencoded for an arginine and glycine to replace methionineand valine at residues 1198 and 1199 of the cleavage site ofa-fodrin by Pet. Directed mutagenesis was performed on theclone 18531 (Stabach et al., 1997) by manufacture’s instruc-tions using PfuTurbo DNA polymerase. After recovering plas-mid DNA from several transformants, we confirmed themutagenized DNA sequences on an Applied Biosystemsmodel 373 A automated sequencer via dye terminator cyclesequencing with Taq polymerase (Perkin-Elmer), accordingto manufacturer’s instructions. Sequencing was performed inthe Biopolymer Laboratory, Department of Microbiology andImmunology, University of Maryland School of Medicine.
Acknowledgements
This work was supported by a grant from Consejo Nacionalde Ciencia y Tecnología de México (CONACYT, 30004 M) toF.N.G. We want to express our gratefulness to James Nataroand Eugenio Frixione for their comments to our manuscript.We also thank to Rocio Huerta and José Luna for theirtechnical help.
References
Bennett, V., and Gilligan, D.M. (1993) The spectrin-basedmembrane skeleton and micron-scale organization of theplasma membrane. Annu Rev Cell Biol 9: 27–66.
Bhan, M.K., Khoshoo, V., Sommerfelt, H., Raj, P., Saza-wal, S., and Srivastava, R. (1989) EnteroaggregativeEscherichia coli and Salmonella associated with nondys-enteric persistent diarrhea. Pediatr Infect Dis J 8: 499–502.
Brown, T.L., Patil, S., Cianci, C.D., Morrow, J.S., and Howe,P.H. (1999) Transforming growth factor beta inducescaspase 3-independent cleavage of alphaII-spectrin(alpha-fodrin) coincident with apoptosis. J Biol Chem 274:23256–23262.
Coleman, T.R., Fishkind, D.J., Mooseker, M.S., and Morrow,J.S. (1989) Contributions of the beta-subunit to spectrinstructure and function. Cell Motil Cytoskeleton 12: 248–263.
Cravioto, A., Tello, A., Navarro, A., Ruiz, J., Villafan, H.,Uribe, F., and Eslava, C. (1991) Association of Escherichiacoli HEp-2 adherence patterns with type and duration ofdiarrhoea. Lancet 337: 262–264.
Deguercy, A., Hommel, M., and Schrevel, J. (1990) Purifica-tion and characterization of 37-kilodalton proteases fromPlasmodium falciparum and Plasmodium berghei whichcleave erythrocyte cytoskeletal components. Mol BiochemParasitol 38: 233–244.
Djabali, K. (1999) Cytoskeletal proteins connecting interme-diate filaments to cytoplasmic and nuclear periphery. HistolHistopathol 14: 501–509.
Eslava, C., Navarro-Garcia, F., Czeczulin, J.R., Henderson,I.R., Cravioto, A., and Nataro, J.P. (1998) Pet, anautotransporter enterotoxin from enteroaggregativeEscherichia coli. Infect Immun 66: 3155–3163.
Eslava, C., Villaseca, J., Morales, R., Navarro, A., and Crav-ioto, A. (1993) Identification of a protein with toxigenicactivity produced by enteroaggregative Escherichia coli.Abstracts of the 93rd General Meeting of the AmericanSociety for Microbiology. Volume Abstract. B-105. Wash-ington, D.C.: American Society for Microbiology, p. 44.
Fujino, T., Ishikawa, T., Inoue, M., Beppu, M., and Kikugawa,K. (1998) Characterization of membrane-bound serine pro-tease related to degradation of oxidatively damaged eryth-rocyte membrane proteins. Biochim Biophys Acta 1374:47–55.
Garty, H., and Palmer, L.G. (1997) Epithelial sodium chan-nels: function, structure, and regulation. Physiol Rev 77:359–396.
Harris, A.S., and Morrow, J.S. (1988) Proteolytic processingof human brain alpha spectrin (fodrin): identification of ahypersensitive site. J Neurosci 8: 2640–2651.
Harris, A.S., and Morrow, J.S. (1990) Calmodulin and cal-cium-dependent protease I coordinately regulate the inter-action of fodrin with actin. Proc Natl Acad Sci USA 87:3009–3013.
Harris, A.S., Croall, D.E., and Morrow, J.S. (1988) The calm-odulin-binding site in alpha-fodrin is near the calcium-dependent protease-I cleavage site. J Biol Chem 263:15754–15761.
Harris, A.S., Croall, D.E., and Morrow, J.S. (1989)Calmodulin regulates fodrin susceptibility to cleavage

958 A. Canizalez-Roman and F. Navarro-Garcia
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 48, 947–958
by calcium-dependent protease I. J Biol Chem 264:17401–17408.
Henderson, I.R., and Nataro, J.P. (2001) Virulence functionsof autotransporter proteins. Infect Immun 69: 1231–1243.
Henderson, I.R., Navarro-Garcia, F., and Nataro, J.P. (1998)The great escape: structure and function of the autotrans-porter proteins. Trends Microbiol 6: 370–378.
Henderson, I.R., Hicks, S., Navarro-Garcia, F., Elias, W.P.,Philips, A.D., and Nataro, J.P. (1999) Involvement of theenteroaggregative Escherichia coli plasmid-encoded toxinin causing human intestinal damage. Infect Immun 67:5338–5344.
Hicks, S., Candy, D.C., and Phillips, A.D. (1996) Adhesion ofenteroaggregative Escherichia coli to pediatric intestinalmucosa in vitro. Infect Immun 64: 4751–4760.
Lynch, G., and Baudry, M. (1984) The biochemistry of mem-ory: a new and specific hypothesis. Science 224: 1057–1063.
Martin, S.J., O’Brien, G.A., Nishioka, W.K., McGahon, A.J.,Mahboubi, A., Saido, T.C., and Green, D.R. (1995) Pro-teolysis of fodrin (non-erythroid spectrin) during apoptosis.J Biol Chem 270: 6425–6428.
Mische, S.M., and Morrow, J.S. (1988) Post-translational reg-ulation of the erythrocyte cortical cytoskeleton. Proto-plasma 145: 167–175.
Morrow, J.S. (1989) The spectrin membrane skeleton:emerging concepts. Curr Opin Cell Biol 1: 23–29.
Nataro, J.P., Hicks, S., Phillips, A.D., Vial, P.A., and Sears,C.L. (1996) T84 cells in culture as a model for enteroag-gregative Escherichia coli pathogenesis. Infect Immun 64:4761–4768.
Nath, R., Raser, K.J., Stafford, D., Hajimohammadreza, I.,Posner, A., Allen, H., et al. (1996) Non-erythroid alpha-spectrin breakdown by calpain and interleukin 1 beta-converting-enzyme-like protease(s) in apoptotic cells:contributory roles of both protease families in neuronalapoptosis. Biochem J 319: 683–690.
Navarro-Garcia, F., Canizalez-Roman, A., Luna, J., Sears,C., and Nataro, J.P. (2001) Plasmid-encoded toxin ofenteroaggregative Escherichia coli is internalized by epi-thelial cells. Infect Immun 69: 1053–1060.
Navarro-Garcia, F., Eslava, C., Villaseca, J.M.,Lopez-Revilla, R., Czeczulin, J.R., Srinivas, S., et al.(1998) In vitro effects of a high-molecular-weight heat-labile enterotoxin from enteroaggregative Escherichia coli.Infect Immun 66: 3149–3154.
Navarro-Garcia, F., Sears, C., Eslava, C., Cravioto, A., andNataro, J.P. (1999) Cytoskeletal effects induced by pet, the
serine protease enterotoxin of enteroaggregative Escheri-chia coli. Infect Immun 67: 2184–2192.
Schulz, J.B., Weller, M., and Klockgether, T. (1996) Potas-sium deprivation-induced apoptosis of cerebellar granuleneurons: a sequential requirement for new mRNA andprotein synthesis, ICE-like protease activity, and reactiveoxygen species. J Neurosci 16: 4696–4706.
Smith, H.R., Cheasty, T., and Rowe, B. (1997) Enteroaggre-gative Escherichia coll and outbreaks of gastroenteritis inUK [letter; comment]. Lancet 350: 814–815.
Stabach, P.R., Cianci, C.D., Glantz, S.B., Zhang, Z., andMorrow, J.S. (1997) Site-directed mutagenesis of alpha IIspectrin at codon 1175 modulates its mu-calpain suscep-tibility. Biochemistry 36: 57–65.
Towbin, H., Staehelin, T., and Gordon, J. (1979) Electro-phoretic transfer of proteins from polyacrylamide gels tonitrocellulose sheets: procedure and some applications.Proc Natl Acad Sci USA 76: 4350–4354.
Vaaraniemi, J., Huotari, V., Lehto, V.P., and Eskelinen, S.(1997) Effect of PMA on the integrity of the membraneskeleton and morphology of epithelial MDCK cells isdependent on the activity of amiloride- sensitive ion trans-porters and membrane potential. Eur J Cell Biol 74: 262–272.
Villaseca, J.M., Navarro-Garcia, F., Mendoza-Hernandez, G.,Nataro, J.P., Cravioto, A., and Eslava, C. (2000) Pet toxinfrom enteroaggregative Escherichia coli produces cellulardamage associated with fodrin disruption. Infect Immun 68:5920–5927.
Wang, K.K. (2000) Calpain and caspase: can you tell thedifference? Trends Neurosci 23: 20–26.
Wang, K.K., Posmantur, R., Nath, R., McGinnis, K., Whitton,M., Talanian, R.V., et al. (1998) Simultaneous degradationof alphaII- and betaII-spectrin by caspase 3 (CPP32) inapoptotic cells. J Biol Chem 273: 22490–22497.
Younes, M., Harris, A.S., and Morrow, J.S. (1989) Fodrin asa differentiation marker. Redistributions in colonic neopla-sia. Am J Pathol 135: 1197–1212.
Ziemnicka-Kotula, D., Xu, J., Gu, H., Potempska, A., Kim,K.S., and Jenkins, E.C. (1998) Identification of a candidatehuman spectrin Src homology 3 domain- binding proteinsuggests a general mechanism of association of tyrosinekinases with the spectrin-based membrane skeleton. J BiolChem 273: 13681–13692.
Zuckerman, J.B., Chen, X., Jacobs, J.D., Hu, B., Kleyman,T.R., and Smith, P.R. (1999) Association of the epithelialsodium channel with Apx and alpha- spectrin in A6 renalepithelial cells. J Biol Chem 274: 23286–23295.
Copyright © 2022 FDOKUMEN




















