
Iron Distribution in Geopolymer with Ferromagnetic Rich Precursor

Journal of Neuroscience Research 33:319-329 (1992)
Rapid Communication Evidence for Intracellular Cleavage of the Alzheimer’s Amyloid Precursor in PC12 Cells K. Sambamurti, J. Shioi, J.P. Anderson, M.A. Pappolla, and N.K. Robakis Department of Psychiatry and Fishberg Research Center for Neurobiology, Mount Sinai School of Medicine, New York, New York
The Alzheimer’s amyloid precursor (APP) is cleaved by an unidentified enzyme (APP secretase) to produce soluble APP. Fractionation of PC12 cell homogenates in a detergent-free buffer showed the presence of the Kunitz protease inhibitor (KP1)-containing soluble APP (nexin 11) in the particulate fraction. Digitonin or sodium carbonate treatment of this fraction solu- bilized nexin 11 suggesting that it is contained in the lumen of vesicles. Nexin I1 production was not af- fected by lysosomotropic agents, suggesting that APP secretase is not a lysosomal enzyme. Labelling of cell surface proteins by iodination failed to detect full- length APP on the surface of PC12 cells, suggesting that most of this protein is located intracellularly. Furthermore, pulse-chase experiments showed that nexin I1 is detected in cell extracts before it appears in the culture medium. Cellular nexin I1 was detected at zero time of chase after only 5 min of pulse labelling with 35S-sulfate, indicated that APP secretase cleav- age takes place immediately after APP is sulfated. Temperature block, pulse-chase, and 35S-sulfate-la- belling experiments suggested that APP is cleaved by APP secretase intracellularly in the trans-Golgi net- work (TGN) or in a post-Golgi compartment. 0 1992 Wiley-Liss, Inc.
Key words: amyloid precursor proteins, APP secre- tase, exocytic vesicles, trans-Golgi network, intracel- lular localization
INTRODUCTION Alzheimer’s disease (AD) is characterized by pro-
gressive dementia and brain depositions of amyloid p- protein (APP) in the form of senile plaques and cerebro- vascular amyloid. Similar depositions are found in aged Down’s syndrome patients and to a lesser extent in nor- mal aging. ApP is a self-aggregating peptide that con- sists of 42-43 residues (Glenner and Wong, 1984a,b; Kang et al., 1987). Molecular cloning studies have
shown that APP is part of at least three distinct Alz- heimer’s amyloid precursors (APPs)-APP,,, APP,, I ,
and APP,,,-encoded by a single gene on chromosome 21. APP,5, and APP,,, contain a 56 amino acid insert with high sequence homology to the Kunitz-type serine protease inhibitors (KPI), which is absent from the third APP species (Goldgarber et al., 1987; Kang et al., 1987; Kitaguchi et al., 1988; Ponte et al., 1988; Robakis et al., 1987; Tanzi et al., 1987, 1988). Full-length APPs are integral membrane glycoproteins containing a large ex- tracytoplasmic sequence, a transmembrane region, and a small cytoplasmic domain. The APP sequence spans the junction of the extracytoplasmic and transmembrane re- gions of the full-length APP.
In addition to the membrane-associated forms, con- stitutively secreted APP, lacking the transmembrane and cytoplasmic regions, has been detected in conditioned media of several cell cultures including PC12 cells (Re- folo et al., 1989; Weidemann et al., 1989), and in human plasma and cerebrospinal fluid (Palmert et al., 1989a,b). Secreted APP, containing the KPI domain, was first identified as nexin I1 from the conditioned medium of fibroblast cell cultures (Van Nostrand et al., 1989; Oltersdorf et al., 1989). It is produced after the KPI- containing full-length APP is cleaved by the putative enzyme APP secretase within the ApP sequence between amino acids Lys,, and Leu,, (Anderson et al., 1991). APP,,, is processed similarly to produce the correspond- ing soluble form (Esch et al., 1990). Thus this cleavage event is incompatible with production of the intact amy- loid peptide. It has been suggested that cellular inability to process all APPs through the secretase pathway may
Received May 4, 1992; revised July 7, 1992; accepted July 9, 1992.
Address reprint requests to Dr. Nikolaos K. Robakis, Department of Psychiatry, Box 1229, Mount Sinai Medical Center, One Gustave L. Levy Place, New York, NY 10029.
J.P. Anderson’s present address is Athena Neurosciences, 800F Gate- way Boulevard, South San Francisco, CA 94080.
0 1992 Wiley-Liss, Inc.

320 Sarnbarnurti et al.
result in altered APP catabolism that leads to the produc- tion of APP (Robakis et al., 1991; Sisodia et al., 1990). Presently, the subcellular organelle in which APP is cleaved is not known, although one hypothesis is that secreted APP is derived from plasma membrane-associ- ated full-length APP (Weidemann et al., 1989) and then directly released into the extracellular environment. Here, we report that the KPI-containing APP is cleaved intracellularly either in the trans-Golgi network (TGN) or in a post-Golgi compartment, and the resultant nexin I1 is transported in vesicles and exocytosed. In addition, nexin I1 production was not inhibited by lysosomotropic agents, indicating that APP secretase is not a lysosomal enzyme.
MATERIALS AND METHODS Antisera
Antisera were prepared against synthetic peptides R 1 and R7 as described (Anderson et al., 1989; Refolo et al., 1989). Peptides R1 and R7 correspond to APP,,, amino acid sequences 729-75 1 and 293 -3 15 respec- tively (Ponte et al., 1988). R1 peptide derives from the cytoplasmic domain of APP, while R7 derives from the extra-cytoplasmic domain and is specific to the KPI in- sert. Anti-GP96 was a kind gift from Dr. Pramod K. Srivastava at the Mount Sinai Medical Center. Mono- clonal antibodies against synaptophysin and secretogra- nin I were obtained from Boehringer Mannhein.
Cell Cultures PC12 cells obtained from American Type Culture
Collection were maintained in RPMI medium, contain- ing 10% horse serum and 5% fetal calf serum, on 100- mm Corning dishes coated with 100 p1 of 0.2 mg/ml collagen (Boehringer Mannheim). Experiments with PC12 cell cultures were performed in RPMI medium supplemented with apotransferrin, insulin, putrescine, selenium, and progesterone (supplements; Bottenstein and Sato, 1979).
Cell Homogenization and Fractionation PC12 cells (approximately lo7 per plate) were ho-
mogenized in approximately 200 pl of 10 mM HEPES pH 7.4, 0.32 M sucrose (Buffer A) supplemented with protease inhibitors (1 mM PMSF, 1 pg/ml aprotinin, 5 pg/ml pepstatin, 5 pg/ml leupeptin, and 1 mM 1,lO- phenanthroline) , using a dounce domogenizer. Homog- enized cells were centrifuged at 1,OOOg for 10 min and the post-nuclear supernatant was centrifuged at 16,OOOg for 30 min to yield a pellet (F2P) and a supernatant (F2S). Pellets were resuspended in 200 p1 of Buffer A and an equal volume (40 p1) of each fraction was loaded
on each lane. Samples were analyzed on Western blots as previously described (Anderson et al., 1989; Refolo et al., 1989) except that reacting proteins were detected by the Amersham enhanced chemiluminescence method.
Digitonin and Sodium Carbonate Extraction of Nexin I1 From Pellet F2P
F2P fractions resuspended in 200 pl of Buffer A were treated with various concentrations of digitonin for 15 min on ice and then centrifuged at 16,OOOg for 30 min. Supernatant was collected and pellets were resus- pended in 200 p1 of Buffer A. Equal volumes (40 pl) of each sample were analyzed on Western blots (Diaz and Stahl, 1989). Alternatively, F2P pellets were treated with 0.1 M sodium carbonate, pH 1 1.5, as described by Fujiki et al. (1982). After centrifugation, supernatant and pellet fractions were analyzed on Western blots.
Treatment With Lysosomal Inhibitors PC12 cell cultures (approximately lo7 cells per
plate) were treated with leupeptin (100 pglml), chloro- quine (50 pM), or ammonium chloride (25 mM), for either 2 or 20 hr at 37°C. Cells were collected and lysed in 200 p l of immunoprecipitate (IP) buffer (50 mM Tris pH 6.8, 150 mM sodium chloride, 5 mM EDTA, 0.5% sodium deoxycholate, 0.5% NP40, 1 mM PMSF, 1 pg/ ml aprotinin, 5 pg/ml pepstatin, 5 pg/ml leupeptin, and 1 mM 1,lO-phenanthroline). Three hundred micrograms of cell extract protein and 50 pg of media protein were analyzed on Western blots as described earlier.
Cycloheximide Treatment PC 12 cell cultures (approximately lo7 cells/plate)
were treated for various times with cycloheximide at 5 pg/ml in 6 ml of RPMI plus supplements with or without added nexin I1 (0.68 pg/ml). Cells were lysed in 300 pl of IP buffer and equal volumes (60 pl) were analyzed on Western blots. Media nexin I1 was concentrated either on heparin agarose or by acetone precipitation and equal volumes corresponding to 4 ml of the starting material were analyzed on Western blots.
Pulse-Labelling PC12 cells were starved for 2 hr in a methionine-
deficient medium, and then labelled in RPMI plus sup- plements in the presence of 80 pCi/ml of 35S-methionine at 15°C for 90 min. After labelling, the medium was removed and replaced with RPMI containing an excess of cold methionine and supplements. Cultures were in cubated for various times at the appropriate temperature,

chilled on ice, and media and cells were collected for immunoprecipitation.
Sulfate Labelling Sulfate labelling was carried out by a modification
of the Tooze and Huttner procedure (1990). Briefly, PC12 cell cultures were incubated at 37°C for 2 hr in RPMI medium lacking sulfate and containing low con- centrations of cysteine (0.5 pg/ml) and methionine (0.15 pg/ml), plus supplements. At the end of the incubation period, the old medium was replaced by fresh medium containing 0.5 mCi/ml of 35S-sulfate (New England Nu- clear) and cells were labelled at 37°C for either 5 min or 2 hr. Samples were chased in RPMI containing sulfate, methionine, cysteine, and supplements and analyzed by immunoprecipitation .
Cell-Surface Labelling Cell-surface iodination was carried out by the
method of Hubbard and Cohn (1976). Briefly, PC12 or C6 cells on culture dishes were washed twice in phos- phate-buffered saline containing 1 mM CaCI, and 0.5 mM MgCl, (PBS) at room temperature and then incu- bated for 30 min at 12-14°C in PBS containing 1 mCi NaI (lZ5I, Amersham), 10 pg/ml leupeptin, 10 pg/ml pepstatin, 1 mM PMSF, 20 pg/ml lactoperoxidase, and 3 pg/ml H,O,. Lactoperoxidase and H,O, were replen- ished at 10 min and 20 min. After labelling, the cells were washed four times in ice-cold PBS containing 5 mM NaI (non-radioactive), scraped off the plates, col- lected by brief centrifugation, and lysed in 400 p1 of IP buffer. Extracts were centrifuged to remove nuclei and cell debris and aliquots containing 2 X lo6 cpm of label were analyzed by immunoprecipitation.
Immunoprecipitation PC12 cells were lysed in IP buffer and then centri-
fuged in a microfuge for 15 min to remove debris and nuclei. Supernatants were made 0.35% in sodium dode- cyl sulfate (SDS) and boiled for 3-5 min. Precipitates were removed by centrifugation and aliquots from the supernatant were TCA-precipitated and counted. Ap- proximately 2 x lo6 cpm of 35S-methionine-labelled cell extract or 5 x lo5 cpm of 35S-sulfate-labelled cell extract were then immunoprecipitated in 200 pl of IP buffer containing 1% Triton X-100 to neutralize the ef- fects of SDS. Secreted APP was immunoprecipitated af- ter conditioned media were made 1 X in IP buffer and boiled in 0.35% SDS. Alternatively, media proteins were concentrated by acetone precipitation (Kitaguchi et al., 1990) and then immunoprecipitated with the appro- priate antisera. Immunoprecipitates were separated on Laemmli gels, soaked in Amplify (Amersham), and au- toradiographed (Laemmli , 1970).
Cleavage of Alzheimer’s Amyloid Precursor 321
F2P F2S CM F2P CM
200 -
97-
68 -
A B Fig. 1. Nexin I1 is found associated with cells. Fractions from PC12 cells homogenates were probed with either anti-R7 (A) or anti-R1 (B) antisera. F2P, high-speed pellet; F2S, high- speed supernatant; CM, conditioned media. Numbers represent kDa. Arrows in A represent 140, 120, and 110 kDa, and in B, arrows represent 140 and 110 kDa.
RESULTS Nexin I1 Is Found in an Intracellular Organelle
PC12 cells were grown as described in Materials and Methods. Under our culture conditions the cells syn- thesized predominantly the KPI-containing APP iso- forms and secreted APP contained predominantly nexin 11, although small amounts of the secreted APPGg5 form were also found in the medium (Refolo et al., 1989; Andersen et al., 199 1). PC 12 cells were homogenized in a detergent-free buffer and fractionated to yield a high speed (16,OOOg) pellet F2P and a supernatant F2S (see Materials and Methods). The F2P fraction reacted with antibodies against secretogranin I and synaptophysin, in- dicating that secretory granules and synaptic vesicles were present in this fraction (data not shown). Figure 1A shows that in agreement with previously published re- sults, the R7 antiserum specific for the KPI-containing APP (Refolo et al., 1989) detects three proteins of 110, 120, and 140 kDa in fraction F2P and a 120 kDa protein in fraction F2S. These signals were abolished in the pres- ence of the R7 peptide (data not shown). Recentrifuga- tion of the high-speed supernatant at 100,OOOg did not sediment the soluble APP (data not shown). On SDS- polyacrylamide gels, the mobility of the 120 kDa protein was identical to that of the secreted APP nexin I1 (Fig. 1A) and it failed to react with the R1 antiserum specific for the cytoplasmic portion of APP, suggesting that this protein is the cell-associated form of nexin I1 (Fig. IB). As expected, both full-length APP species at 110 and 140 kDa, representing the endoplasmic reticulum (ER)
322 Sambamurti et al.
% D i g i t o n i n 0.0 0.025 0.05 0.075 0.1 0.15 0.2 P S P S P S P S P S P S P S 1 2 3
200 -
97 -
68 -
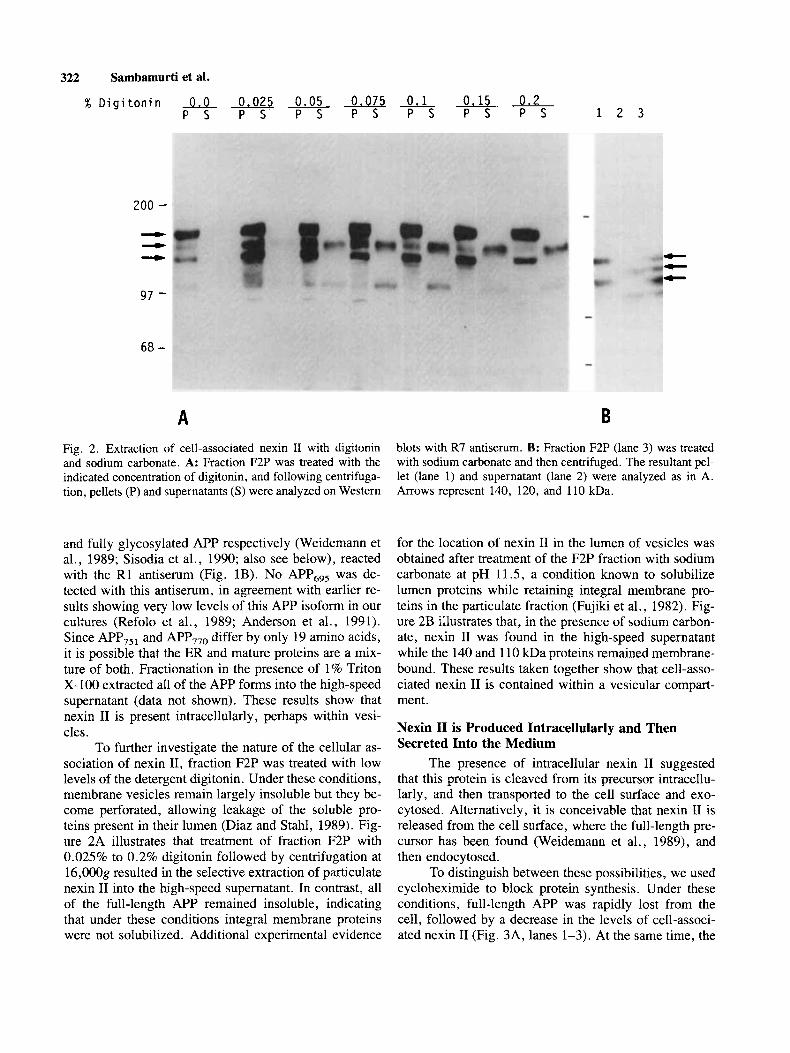
Fig. 2 . Extraction of cell-associated nexin I1 with digitonin and sodium carbonate. A: Fraction F2P was treated with the indicated concentration of digitonin, and following centrifuga- tion, pellets (P) and supernatants ( S ) were analyzed on Western
blots with R7 antiserum. B: Fraction F2P (lane 3) was treated with sodium carbonate and then centrifuged. The resultant pe- let (lane l) and supernatant (lane 2) were analyzed as in A. Arrows represent 140, 120, and 110 kDa.
and fully glycosylated APP respectively (Weidemann et al., 1989; Sisodia et al., 1990; also see below), reacted with the R1 antiserum (Fig. 1B). No APPGg5 was de- tected with this antiserum, in agreement with earlier re- sults showing very low levels of this APP isoform in our cultures (Refolo et al., 1989; Anderson et al., 1991). Since APP,,, and APP,,o differ by only 19 amino acids, it is possible that the ER and mature proteins are a mix- ture of both. Fractionation in the presence of 1% Triton X-100 extracted all of the APP forms into the high-speed supernatant (data not shown). These results show that nexin I1 is present intracellularly, perhaps within vesi- cles.
To further investigate the nature of the cellular as- sociation of nexin 11, fraction F2P was treated with low levels of the detergent digitonin. Under these conditions, membrane vesicles remain largely insoluble but they be- come perforated, allowing leakage of the soluble pro- teins present in their lumen (Diaz and Stahl, 1989). Fig- ure 2A illustrates that treatment of fraction F2P with 0.025% to 0.2% digitonin followed by centrifugation at 16,OOOg resulted in the selective extraction of particulate nexin I1 into the high-speed supernatant. In contrast, all of the full-length APP remained insoluble, indicating that under these conditions integral membrane proteins were not solubilized. Additional experimental evidence
for the location of nexin I1 in the lumen of vesicles was obtained after treatment of the F2P fraction with sodium carbonate at pH 11.5, a condition known to solubilize lumen proteins while retaining integral membrane pro- teins in the particulate fraction (Fujiki et al., 1982). Fig- ure 2B illustrates that, in the presence of sodium carbon- ate, nexin I1 was found in the high-speed supernatant while the 140 and 1 10 kDa proteins remained membrane- bound. These results taken together show that cell-asso- ciated nexin I1 is contained within a vesicular compart- ment.
Nexin I1 is Produced Intracellularly and Then Secreted Into the Medium
The presence of intracellular nexin I1 suggested that this protein is cleaved from its precursor intracellu- larly, and then transported to the cell surface and exo- cytosed. Alternatively, it is conceivable that nexin I1 is released from the cell surface, where the full-length pre- cursor has been found (Weidemann et al., 1989), and then endocytosed.
To distinguish between these possibilities, we used cycloheximide to block protein synthesis. Under these conditions, full-length APP was rapidly lost from the cell, followed by a decrease in the levels of cell-associ- ated nexin I1 (Fig. 3A, lanes 1-3). At the same time, the
Cleavage of Alzheimer’s Amyloid Precursor 323
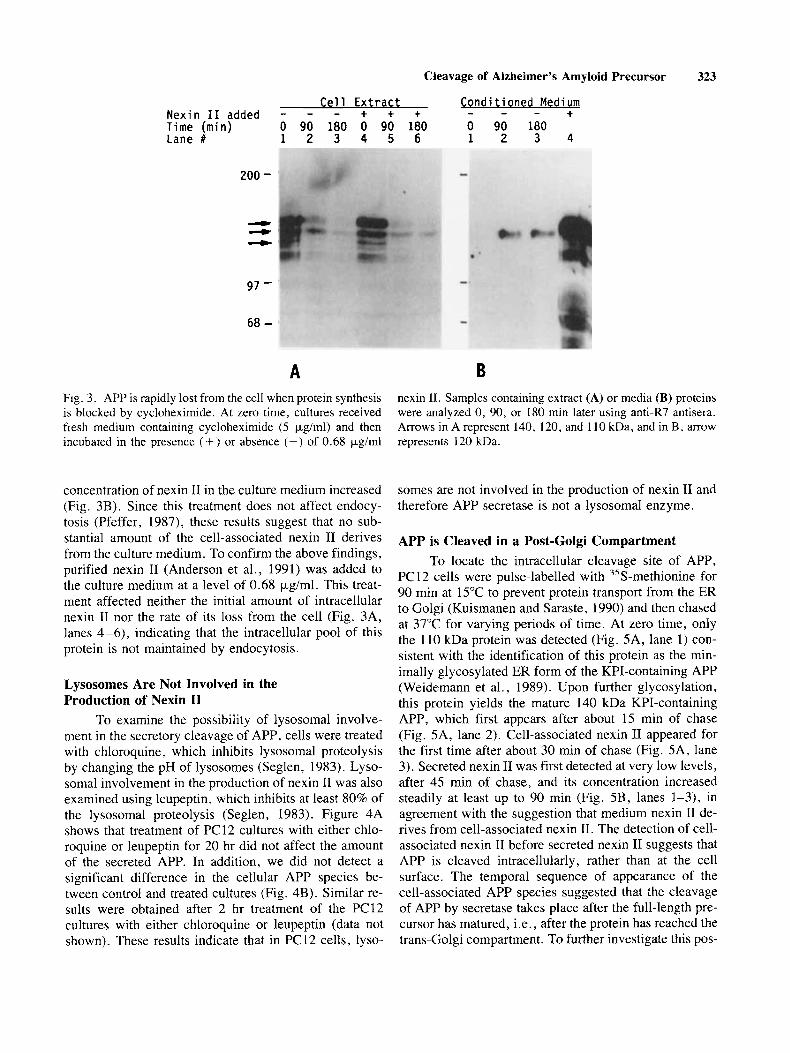
Cell Extract Conditioned Medium + + + - - - + Nexin I 1 added - - - Time (min) 0 90 180 0 90 180 0 90 180 Lane # 1 2 3 4 5 6 1 2 3 4
200 -
97 -
68 -
A Fig. 3 . APP is rapidly lost from the cell when protein synthesis is blocked by cycloheximide. At zero time, cultures received fresh medium containing cycloheximide (5 pg/ml) and then incubated in the presence (+) or absence (-) of 0.68 pglml
concentration of nexin I1 in the culture medium increased (Fig. 3B). Since this treatment does not affect endocy- tosis (Pfeffer, 1987), these results suggest that no sub- stantial amount of the cell-associated nexin I1 derives from the culture medium. To confirm the above findings, purified nexin I1 (Anderson et al., 1991) was added to the culture medium at a level of 0.68 pg/ml. This treat- ment affected neither the initial amount of intracellular nexin I1 nor the rate of its loss from the cell (Fig. 3A, lanes 4-6), indicating that the intracellular pool of this protein is not maintained by endocytosis.
Lysosomes Are Not Involved in the Production of Nexin I1
To examine the possibility of lysosomal involve- ment in the secretory cleavage of APP, cells were treated with chloroquine, which inhibits lysosomal proteolysis by changing the pH of lysosomes (Seglen, 1983). Lyso- soma1 involvement in the production of nexin I1 was also examined using leupeptin, which inhibits at least 80% of the lysosomal proteolysis (Seglen, 1983). Figure 4A shows that treatment of PC12 cultures with either chlo- roquine or leupeptin for 20 hr did not affect the amount of the secreted APP. In addition, we did not detect a significant difference in the cellular APP species be- tween control and treated cultures (Fig. 4B). Similar re- sults were obtained after 2 hr treatment of the PC12 cultures with either chloroquine or leupeptin (data not shown). These results indicate that in PC12 cells, lyso-
nexin 11. Samples containing extract (A) or media (B) proteins were analyzed 0, 90, or 180 min later using antiLR7 antisera. Arrows in A represent 140, 120, and 110 kDa, and in B, arrow represents 120 kDa.
somes are not involved in the production of nexin I1 and therefore APP secretase is not a lysosomal enzyme.
APP is Cleaved in a Post-Golgi Compartment To locate the intracellular cleavage site of APP,
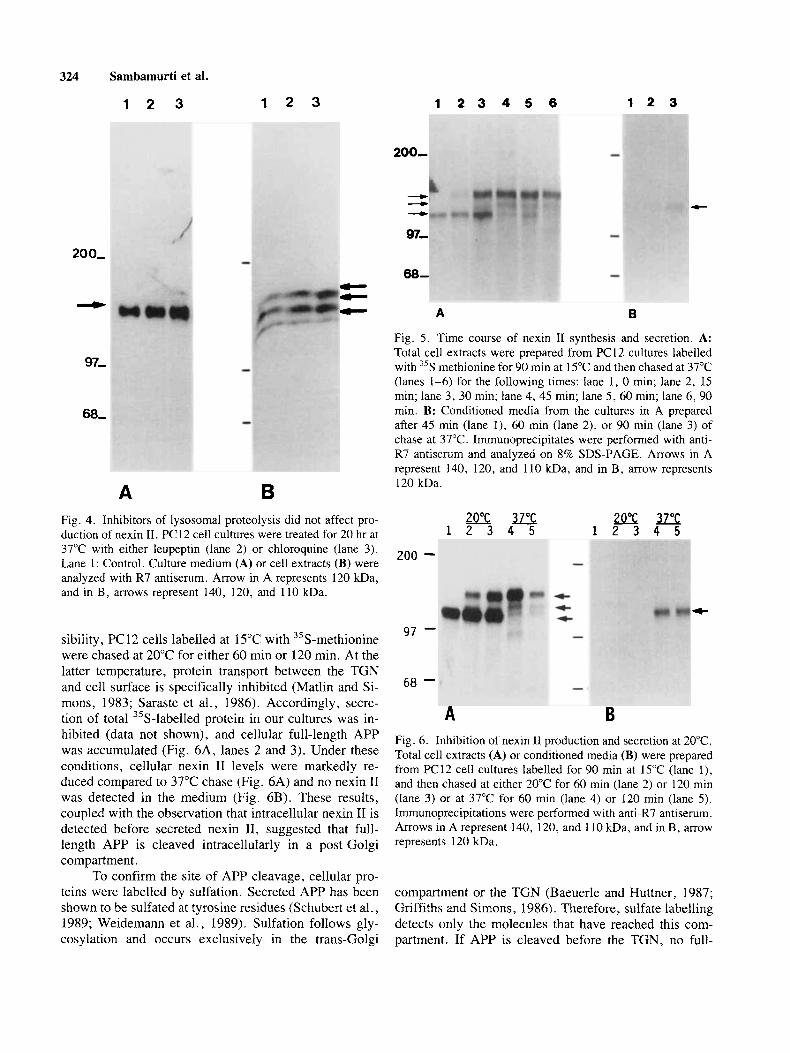
PC 12 cells were pulse-labelled with 35S-methionine for 90 min at 15°C to prevent protein transport from the ER to Golgi (Kuismanen and Saraste, 1990) and then chased at 37°C for varying periods of time. At zero time, only the 110 kDa protein was detected (Fig. 5A, lane 1) con- sistent with the identification of this protein as the min- imally glycosylated ER form of the KPI-containing APP (Weidemann et al., 1989). Upon further glycosylation, this protein yields the mature 140 kDa KPI-containing APP, which first appears after about 15 rnin of chase (Fig. 5A, lane 2). Cell-associated nexin I1 appeared for the first time after about 30 min of chase (Fig. 5A, lane 3). Secreted nexin I1 was first detected at very low levels, after 45 min of chase, and its concentration increased steadily at least up to 90 min (Fig. 5B, lanes 1-3), in agreement with the suggestion that medium nexin I1 de- rives from cell-associated nexin 11. The detection of cell- associated nexin I1 before secreted nexin I1 suggests that APP is cleaved intracellularly, rather than at the cell surface. The temporal sequence of appearance of the cell-associated APP species suggested that the cleavage of APP by secretase takes place after the full-length pre- cursor has matured, i.e., after the protein has reached the trans-Golgi compartment. To further investigate this pos-
324 Sambamurti et al.
1 2 3
200- - 97-
68,
A
1 2 3
B
200-
68- - CI - A
1 2 3 4 5 6 1 2 3
B Fig. 5 . Time course of nexin I1 synthesis and secretion. A: Total cell extracts were prepared from PC12 cultures labelled with "S methionine for 90 rnin at 15°C and then chased at 37°C (lanes 1-6) for the following times: lane 1 , 0 min; lane 2, 15 min; lane 3, 30 min; lane 4, 45 min; lane 5 , 60 min; lane 6, 90 min. B: Conditioned media from the cultures in A prepared after 45 rnin (lane l), 60 min (lane 2), or 90 min (lane 3) of chase at 37°C. Immunoprecipitates were performed with anti- R7 antiserum and analyzed on 8% SDS-PAGE. Arrows in A represent 140, 120, and 110 kDa, and in B, arrow represents 120 kDa.
m x x 1 2 3 4 5
Fig. 4. Inhibitors of lysosomal proteolysis did not affect pro- m37"c 1 2 3 4 5 duction of nexin 11. PC12 cell cultures were treated for 20 hr at
37°C with either leupeptin (lane 2) or chloroquine (lane 3). Lane 1: Control. Culture medium (A) or cell extracts (B) were analyzed with R7 antiserum. Arrow in A represents 120 kDa, and in B, arrows represent 140, 120, and 110 kDa.
200 -
c 97 - sibility , PC12 cells labelled at 15°C with 35S-methionine
were chased at 20°C for either 60 rnin or 120 min. At the latter temperature, protein transport between the TGN
mons, 1983; Saraste et al., 1986). Accordingly, secre- tion of total 35S-labelled protein in our cultures was in-
and cell surface is specifically inhibited (Matlin and Si- 68 - - A B
hibited (data not shown), and cellular full-length APP was accumulated (Fig. 6A, lanes 2 and 3). Under these conditions, cellular nexin I1 levels were markedly re- duced compared to 37°C chase (Fig. 6A) and no nexin I1 was detected in the medium (Fig. 6B). These results, coupled with the observation that intracellular nexin I1 is detected before secreted nexin 11, suggested that full- length APP is cleaved intracellularly in a post-Golgi compartment.
To confirm the site of APP cleavage, cellular pro- teins were labelled by sulfation. Secreted APP has been shown to be sulfated at tyrosine residues (Schubert et al., 1989; Weidemann et al., 1989). Sulfation follows gly- cosylation and occurs exclusively in the trans-Golgi
Fig. 6. Inhibition of nexin 11 production and secretion at 20°C. Total cell extracts (A) or conditioned media (B) were prepared from PC12 cell cultures labelled for 90 min at 15°C (lane l), and then chased at either 20°C for 60 min (lane 2) or 120 rnin (lane 3) or at 37°C for 60 rnin (lane 4) or 120 min (lane 5) . Immunoprecipitations were performed with antiLR7 antiserum. Arrows in A represent 140, 120, and 110 kDa, and in B, arrow represents 120 kDa.
compartment or the TGN (Baeuerle and Huttner, 1987; Griffiths and Simons, 1986). Therefore, sulfate labelling detects only the molecules that have reached this com- partment. If APP is cleaved before the TGN, no full-
1 2 1 2 3
200-
Cleavage of Alzheimer's Amyloid Precursor 325
1 2 1 2
100-
200-
97-
68-
9 7-
68-
4
d
97-
68-
43-
43-
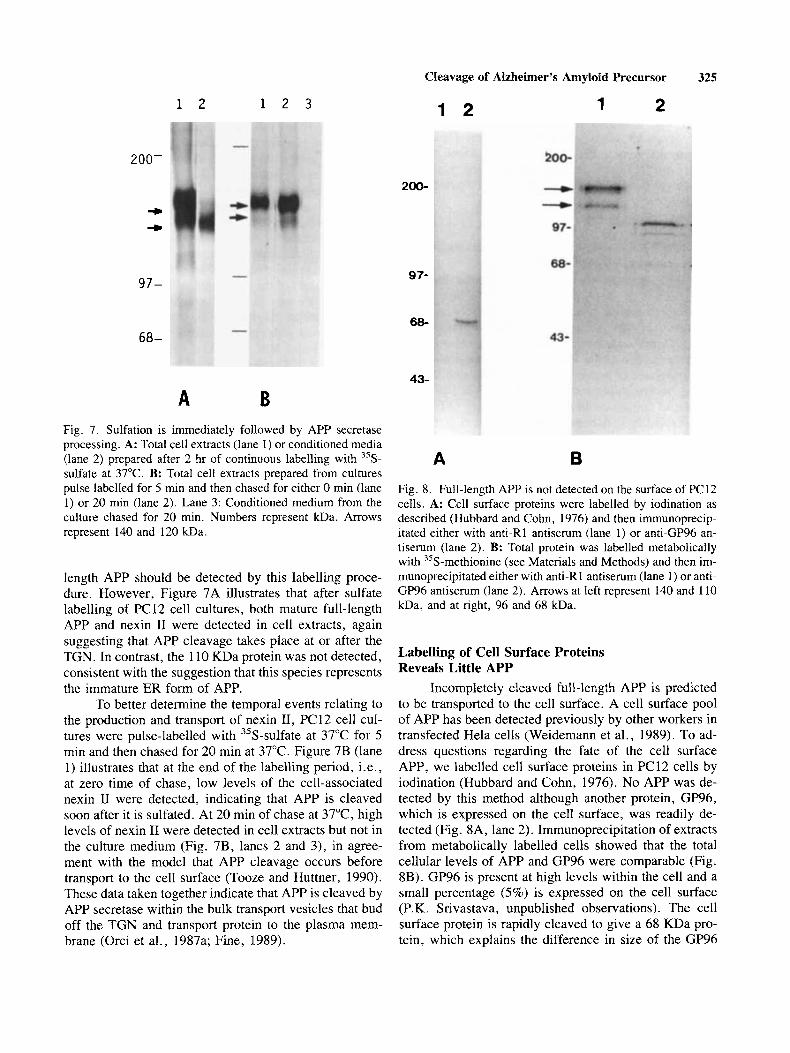
A B Fig. 7. Sulfation is immediately followed by APP secretase processing. A: Total cell extracts (lane 1) or conditioned media (lane 2) prepared after 2 hr of continuous labelling with 35S- sulfate at 37°C. B: Total cell extracts prepared from cultures pulse labelled for 5 min and then chased for either 0 rnin (lane 1) or 20 rnin (lane 2) . Lane 3: Conditioned medium from the culture chased for 20 min. Numbers represent kDa. Arrows represent 140 and 120 kDa.
length APP should be detected by this labelling proce- dure. However, Figure 7A illustrates that after sulfate labelling of PC 12 cell cultures, both mature full-length APP and nexin I1 were detected in cell extracts, again suggesting that APP cleavage takes place at or after the TGN. In contrast, the 110 KDa protein was not detected, consistent with the suggestion that this species represents the immature ER form of APP.
To better determine the temporal events relating to the production and transport of nexin 11, PC12 cell cul- tures were pulse-labelled with 35S-sulfate at 37°C for 5 min and then chased for 20 min at 37°C. Figure 7B (lane 1) illustrates that at the end of the labelling period, i.e., at zero time of chase, low levels of the cell-associated nexin I1 were detected, indicating that APP is cleaved soon after it is sulfated. At 20 min of chase at 37"C, high levels of nexin I1 were detected in cell extracts but not in the culture medium (Fig. 7B, lanes 2 and 3), in agree- ment with the model that APP cleavage occurs before transport to the cell surface (Tooze and Huttner, 1990). These data taken together indicate that APP is cleaved by APP secretase within the bulk transport vesicles that bud off the TGN and transport protein to the plasma mem- brane (Orci et al., 1987a; Fine, 1989).
A B
Fig. 8. Full-length APP is not detected on the surface of PC12 cells. A: Cell surface proteins were labelled by iodination as described (Hubbard and Cohn, 1976) and then immunoprecip- itated either with anti-R1 antiserum (lane 1) or anti-GP96 an- tiserum (lane 2). B: Total protein was labelled metabolically with 35S-methionine (see Materials and Methods) and then im- munoprecipitated either with anti-Rl antiserum (lane 1) or anti- GP96 antiserum (lane 2). Arrows at left represent 140 and 110 kDa, and at right, 96 and 68 kDa.
Labelling of Cell Surface Proteins Reveals Little APP
Incompletely cleaved full-length APP is predicted to be transported to the cell surface. A cell surface pool of APP has been detected previously by other workers in transfected Hela cells (Weidemann et al., 1989). To ad- dress questions regarding the fate of the cell surface APP, we labelled cell surface proteins in PC12 cells by iodination (Hubbard and Cohn, 1976). No APP was de- tected by this method although another protein, GP96, which is expressed on the cell surface, was readily de- tected (Fig. 8A, lane 2). Immunoprecipitation of extracts from metabolically labelled cells showed that the total cellular levels of APP and GP96 were comparable (Fig. 8B). GP96 is present at high levels within the cell and a small percentage (5%) is expressed on the cell surface (P.K. Srivastava, unpublished observations). The cell surface protein is rapidly cleaved to give a 68 KDa pro- tein, which explains the difference in size of the GP96

326 Sambamurti et al.
immunoprecipitates from total and surface-labelled PC 12 cell cultures seen in Figure 8 (Srivastava et al., 1986; P. K. Srivastava, unpublished observations). These data indicate that very little, if any, APP is found on the surface of PC12 cells, lending support to the suggestion that the majority of the full-length APP is located intra- cellularly .
DISCUSSION Normal constitutive processing of full-length APP
involves cleavage within the APP sequence by the pro- tease APP secretase to produce the secreted forms of APP (Esch et al., 1990; Sisodia et al., 1990; Anderson et al., 1991). Since this processing is incompatible with the production of APP, the presence of amyloid depositions in AD, Down's syndrome, or normal aging indicates that at least a fraction of full-length APP escapes cleavage by APP secretase and instead is catabolized through an amy- loidogenic proteolytic pathway (Robakis et al., 1991). A number of lesions could trigger this altered APP catab- olism including mutations of the APP gene (Levy et al., 1990; Goate et al., 1991), abnormally high levels of expression as observed in Down's syndrome (Tanzi, et al., 1987), modifications suppressing APP secretase ac- tivity, or miscompartmentalization of APP resulting in degradation through alternate pathways. Thus, elucida- tion of the factors and subcellular compartments in- volved in the processing of APP is critical to the under- standing of amyloidogenesis. Presently, neither the cellular site of APP secretase cleavage nor the transport of the resulting soluble APP into the extracellular space have been determined.
Here, we present evidence that nexin 11, a product of APP secretase activity, is present in intracellular ves- icles. On SDS-polyacrylamide gels, intracellular nexin IT displayed the same mobility as the secreted nexin I1 and on immunoblots it showed the same reactivity toward both the anti-R7 and anti-R1 antisera as the secreted APP. Cell-associated nexin I1 was found in the particu- late fraction of cell homogenates, suggesting that it is contained within vesicles. This hypothesis was con- firmed by the digitionin and sodium carbonate extraction experiments. Both of these compounds have been used extensively for the solubilization of lumen proteins while integral membrane proteins remain insoluble (Fujiki et al. 1982; Diaz and Stahl, 1989). Both agents solubilized cell-associated nexin I1 while full-length APP remained membrane bound, suggesting that this protein is present in a soluble form in the lumen of vesicles. Breakage of vesicles explains the presence of nexin I1 in the cytosolic fraction (Fig. 1). Our experiments with cycloheximide strongly suggest that intracellular nexin I1 does not de- rive through endocytosis of media nexin 11. Additional support for this conclusion was obtained from the pulse
chase experiments where cell-associated nexin I1 was detected before media nexin 11.
Nexin I1 has previously been localized to a-gran- ules in blood platelets (Van Nostrand et al., 1990). How- ever, since a-granule contents derive from both mega- karyocyte and plasma proteins (Handagama et al., 1990), the origin of nexin I1 in a-granules was not clear.
To investigate the possibility that APP secretase is a lysosomal enzyme, we treated PC12 cell cultures with inhibitors of lysosomal proteolysis. Our results show that chloroquine or leupeptin did not affect the levels of either secreted or cell-associated nexin 11. Sim- ilarly, the levels of the full-length APP were not affected by these treatments. A third lysosomotropic agent, am- monium chloride (Seglen, 1983), gave results similar to those obtained with either chloroquine or leupeptin (K. Sambamurti and N .K. Robakis, unpublished observa- tions). Since these lysosomotropic agents have been ex- tensively used as indicators of the lysosomal processing of proteins (Arnara et al., 1989), our results strongly suggest that lysosomal enzymes are not involved in the production of nexin 11. In agreement with this sugges- tion, secreted nexin I1 has been shown to contain sialic acid residues (Refolo et al., 1989). Lysosomal process- ing of APP would have resulted in the removal of the sialic acid residues by the resident sialidase and P-galac- tosidase (Neufeld, 1991). Similar results, suggesting that APP-secretase is not a lysosomal enzyme, have been recently obtained by two independent laboratories (Ca- poraso et al., 1992; Golde et al., 1992). However, these results do not exclude the presence of alternative path- ways for APP turnover, which may include lysosomal processing (Cole et al., 1989).
Our pulse-chase experiments with labelled amino acids suggest that APP is cleaved after it is fully glyco- sylated. Since glycosylation is completed in the trans- Golgi compartment of the cell (Kornfeld and Kornfeld, 1985), APP cleavage occurs at or after this organelle. This conclusion is further supported by the sulfation ex- periments, which indicate that APP is cleaved after it is sulfated. Protein sulfation occurs after both glycosylation and sialylation have been completed and it is thought to take place either in the trans-Golgi compartment or the TGN (Baeuerle and Huttner, 1987), which is responsible for routing proteins to various cellular destinations in- cluding secretory vesicles (Griffiths and Simons, 1986). Protein transport from the TGN to the cell surface is inhibited at 20°C in mammalian cells (Kuismanen and Saraste, 1990) including PC12 cells (Tooze and Huttner, 1990). Accordingly, we observed no protein secretion in our PC12 cultures at 20"C, indicating no protein trans- port to the cell surface. Under these conditions, produc- tion of cellular or medium nexin I1 was inhibited, sug- gesting that APP is cleaved in a post-Golgi compartment.
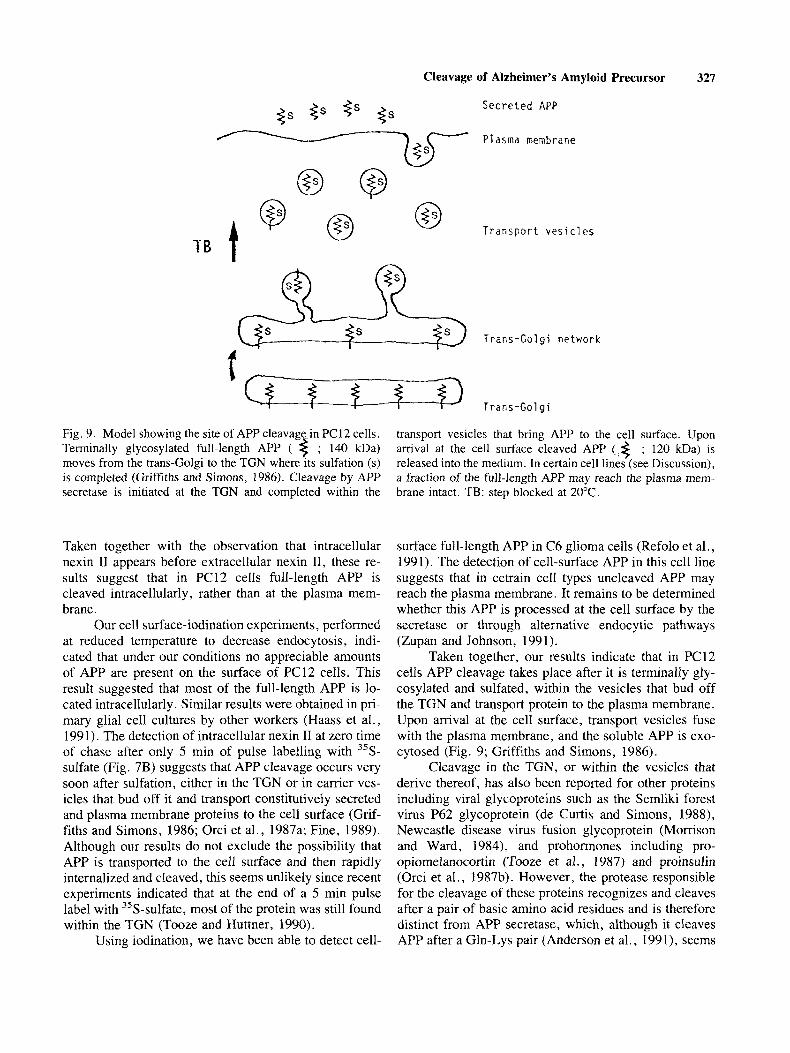
Cleavage of Alzheimer’s Amyloid Precursor 327
Secreted APP
Plasma membrane
o Q T B f @ 0 Transport vesicles
,o.R, Trans-Golgi network
Trans-Golgi
t Fig. 9. Model showing the site of APP cleavage-in PC12 cells. Terminally glycosylated full-length APP ( $ ; 140 kDa) moves from the trans-Golgi to the TGN where its sulfation ( s ) is completed (Griffiths and Simons, 1986). Cleavage by APP secretase is initiated at the TGN and completed within the
Taken together with the observation that intracellular nexin I1 appears before extracellular nexin 11, these re- sults suggest that in PC12 cells full-length APP is cleaved intracellularly, rather than at the plasma mem- brane.
Our cell surface-iodination experiments, performed at reduced temperature to decrease endocytosis, indi- cated that under our conditions no appreciable amounts of APP are present on the surface of PC12 cells. This result suggested that most of the full-length APP is lo- cated intracellularly . Similar results were obtained in pri- mary glial cell cultures by other workers (Haass et al., 1991). The detection of intracellular nexin I1 at zero time of chase after only 5 min of pulse labelling with 35S- sulfate (Fig. 7B) suggests that APP cleavage occurs very soon after sulfation, either in the TGN or in carrier ves- icles that bud off it and transport constitutively secreted and plasma membrane proteins to the cell surface (Grif- fiths and Simons, 1986; Orci et al., 1987a; Fine, 1989). Although our results do not exclude the possibility that APP is transported to the cell surface and then rapidly internalized and cleaved, this seems unlikely since recent experiments indicated that at the end of a 5 min pulse label with 35S-sulfate, most of the protein was still found within the TGN (Tooze and Huttner, 1990).
Using iodination, we have been able to detect cell-
transport vesicles that bring APP to the cell surface. Upon arrival at the cell surface cleaved APP (,< ; 120 kDa) is released into the medium. In certain cell lines (see Discussion), a fraction of the full-length APP may reach the plasma mem- brane intact. TB: step blocked at 20°C.
surface full-length APP in C6 glioma cells (Refolo et al., 1991). The detection of cell-surface APP in this cell line suggests that in cetrain cell types uncleaved APP may reach the plasma membrane. It remains to be determined whether this APP is processed at the cell surface by the secretase or through alternative endocytic pathways (Zupan and Johnson, 1991).
Taken together, our results indicate that in PC12 cells APP cleavage takes place after it is terminally gly- cosylated and sulfated, within the vesicles that bud off the TGN and transport protein to the plasma membrane. Upon arrival at the cell surface, transport vesicles fuse with the plasma membrane, and the soluble APP is exo- cytosed (Fig. 9; Griffiths and Simons, 1986).
Cleavage in the TGN, or within the vesicles that derive thereof, has also been reported for other proteins including viral glycoproteins such as the Semliki forest virus P62 glycoprotein (de Curtis and Simons, 1988), Newcastle disease virus fusion glycoprotein (Morrison and Ward, 1984), and prohormones including pro- opiomelanocortin (Tooze et al., 1987) and proinsulin (Orci et al., 1987b). However, the protease responsible for the cleavage of these proteins recognizes and cleaves after a pair of basic amino acid residues and is therefore distinct from APP secretase, which, although it cleaves APP after a Gln-Lys pair (Anderson et al., 1991), seems

328 Sambamurti et al.
to have no specific sequence requirements for cleavage (Maruyama et al., 1991).
The results presented here suggest that APP secre- tase is localized in the vesicles that derive from the TGN. This will facilitate the isolation and cloning of this en- zyme as well as determination of its activity in normal and AD brains.
ACKNOWLEDGMENTS We are grateful to Drs. Lawrence M. Refolo and
Brian Storrie for useful discussions and to Dr. Pramod K. Srivastava for providing the anti-GP96 antiserum and permission to use his unpublished observations. This work was supported by grants AGO8200 and AGO5 138.
NOTE ADDED IN PROOF Kuenczel et al. have recently obtained similar re-
sults using the H4 human neuroglioma cell line (T.J. Raub, personal communication).
REFERENCES Amara JF, Lederkremer G, Lodish HF (1989): lntracellular degrada-
tion of unassembled asialoglycoprotein receptor subunits: A pre-Golgi, nonlysosomal endoproteolytic cleavage. J Cell Biol
Anderson JP, Refolo LM, Wallace W, Mehta P, Krishnamurthi M, Gotlib J, Bierer L, Haroutunian V, Per1 D, Robakis NK (1989): Differential brain expression of the Alzheimer’s amyloid pre- cursor protein. EMBO J 8:3627-3632.
Anderson JP, Esch FS, Keim PS, Sambamurti K, Lieberburg I, Robakis NK (1991): Exact cleavage site of Alzheimer amyloid precursor in neuronal PC-12 cells. Neurosci Lett 128: 126-128.
Baeuerle PA, Huttner WB (1987): Tyrosine sulfation is a trans-Golgi- specific protein modification. J Cell Biol 105:2655-2664.
Bottenstein JE, Sat0 GH (1979): Growth of a rat neuroblastoma cell line in serum-free supplemented medium. Proc Natl Acad Sci USA 76514-517,
Caporaso GL, Gandy S, Buxbaum JD, Greengard P (1992): Chloro- quine inhibits intracellular degradation but not secretion of Alz- heimer P/A4 amyloid precursor protein. Proc Natl Acad Sci
Cole GM, Huynh TV, Saitoh T (1989): Evidence for lysosomal pro- cessing of amyloid-p-protein precursor in cultured cells. Neu- rochem Res 14:933-939.
de Curtis I, Simons K (1988): Dissection of Semlike forest virus glycoprotein delivery from the trans-Golgi network to the cell surface in permeabilized BHK cells. Proc Natl Acad Sci USA 853052-8056.
Diaz R, Stahl PD (1989): Digitonin permeabilization procedures for the study of endosome acidification and function. Methods Cell Biol 3 1 2 - 4 3 .
Esch FS, Keim PS, Beattie EC, Blacher RW, Culwell AR, Oltersdorf T, McClure D, Ward PJ (1990): Cleavage of amylod p peptide during constitutive processing of its precursor. Science 248: 1122-1 124.
109:33 15 -3324.
USA 8912252-2256.
Fine RE (1989): Vesicles without clathrin: Intermediates in bulk flow exocytosis. Cell 58:609 -6 10.
Fujiki Y, Hubbard AL, Fowler S , Lazarow PB (1982): Isolation of intracellular membranes by means of sodium carbonate treat- ment: Application to endoplasmic reticulum. J Cell Biol 93:
Glenner GG, Wong CW (1984a): Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular arnyloid protein. Biochem Biophys Res Commun 120:885- 890.
Glenner GG, Wong CW (1984b): Alzheimer’s disease and Down’s syndrome: Sharing of a unique cerebrovascular amyloid fibril protein. Biochem Biophys Res Commun 122:1131-1135.
Goate A, Chartier-Harlin M-C, Mullan M, Brown J, Crawford F, Fidani L, Giuffra L, Haynes A, Irving N, James L, Mant R, Newton P, Rooke K, Roques P, Talbot C, Pericak-Vance M, Roses A, Williamson R, Rossen M, Owen M, Hardy J (1991): Segregation of a missense mutation in the amyloid precursor protein gene with familial Alzheimer’s disease. Nature 349: 704-706.
Golde TE, Estus S , Younkin LH, Selkoe DJ, Younkin SG (1992): Processing of the amyloid protein precursor to potentially amy- oidogenic derivatives. Science 255:728-730.
Goldgaber D, Lerman MI, McBride WO, Saffiotti U, Gajdusek DC ( 1987): Characterization and chromosomal localization of a DNA encoding brain amyloid of Alzeimer’s disease. Science
Griffiths G, Simons K (1986): The trans-Golgi network: Sorting at the exit site of the Golgi complex. Science 234:438-443.
Handagama P, Rappolee DA, Werb Z, Levin J, Bainton DF (1990): Platelet alpha-granule fibrinogen, albumin and immunoglobin G are not synthesized by rat and mouse megakaryocytes. J Clin Invest 86:1364-1368.
Haass C, Hung AY, Selkoe DJ (1991): Processing of p-amyloid pre- cursor protein in microglia and astrocytes favours an internal localization over constitutive secretion. J Neurosci 11:3783- 3793.
Hubbard SC, Cohn ZA (1976): Specific labels for cell surfaces. In Maddy AH (ed): “Biochemical Analysis of Membranes.” London: Clappman and Hall, pp 427-501.
Kang J, Lemaire H-G, Unterbeck A, Salbaum JM, Masters CL, Grzeschik K-H, Multhaup G, Beyreuther K, Muller-Hill B (1987): The precursor of Alzheimer’s disease amyloid A4 pro- tein resembles a cell-surface receptor. Nature 325:733-736.
Kitaguchi N, Takahashi Y, Tokushima Y, Shiojiri S, Ito H (1988): Novel precursor of Alzheimer’s disease amyloid protein shows protease inhibitory activity. Nature 331530-532.
Kitaguchi N, Takahashi Y, Oishi K, Shiojiri S , Tokushima Y, Ut- sunomiya T, It0 H (1990): Enzyme specificity of proteinase inhibitor region in amyloid precursor protein of Alzheimer’s disease: Different properties compared with protease nexin I. Biochim Biophys Acta 1038:105-113.
Kornfeld R, Kornfeld S (1985): Assembly of asparagine-linked olighosaccharides. Ann Rev Biochem 54:63 1-664.
Kuismanen E, Saraste J (1990): Low temperature-induced transport blocks as tools to manipulate membrane traffic. Methods Cell Biol 32:257-274.
Laemmli UK (1970): Cleavage of structural proteins during the as- sembly of the head of bacteriophage T4. Nature 227:680-685.
Levy E, Carman MD, Fernandez-Madrid IJ, Power MD, Lieberburg I, van Duinen SG, Bots GThAM, Luyendijk W, Frangione B (1990): Mutation of the Alzheimer’s disease amyloid gene in hereditary cerebral hemorrhage, Dutch type. Science 248:
97-1 02.
235:877-880.
1124-1 126.

Cleavage of Alzheimer’s Amyloid Precursor 329
cerebrovascular and the neuritic plaque core amyloid peptides. Proc Natl Acad Sci USA 84:4190-4194.
Robakis NK, Sambamurti K, Anderson JP, Mehta P, Refolo LM (1991): Effects of neurotrophic factors on the secretion and metabolism of the Alzheimer’s amyloid precursor. In Bracket H, Christen W (eds): “Growth factors and Alzheimer’s Disease.” Heidelberg: Spinger-Verlag, pp 208-215.
Saraste J , Palade GE, Farquhar MG (1986): Temperature-sensitive steps in the transport of secretory proteins through the Golgi complex in exocrine pancreatic cells. Proc Natl Acad Sci USA
Schubert D, Lacorbiere M, Saitoh T, Cole G (1989): Characterization of an amyloid @-precursor protein which binds heparin and contains tyrosine sulfate. Proc Natl Acad Sci 86:2066-2069.
Seglen PO (1983): Inhibitors of lysosomal function. Methods Enzy- mol 96:737-764.
Sisodia SS, Koo EH, Beyreuther K, Unterbeck A, Price DL (1990): Evidence that beta-amyloid protein in Alzheimer’s Disease is not derived by normal processing. Science 248:492-495.
Srivastava PK, DeLeo AB, Old LJ (1986): Tumor rejection antigens of chemically induced sarcomas of inbred mice. Proc Natl Acad Sci USA 83:3407-3411.
Tanzi RE, Gusella JF, Watkins PC, Bruns GAP, St. George-Hyslop P, van Keuren ML, Patterson D, Pagan S, Kurnit DM, Neve RL (1987): Amyloid P protein gene: cDNA mRNA distribution, and genetic linkage near the Alzheimer’s locus. Science 235:
Tanzi RE, McClatchey AI, Lamperti ED, Villa-Komaroff L, Gusella JF, Neve RL (1988): Protease inhibitor domain encoded by an amyloid precursor mRNA associated with Alzheimer’s disease. Nature 331528-530.
Tooze J , Hollinshead M, Frank R, Burke B (1987): An antibody specific for an endoproteolytic cleavage site provides evidence that pro-opiomelanocortin is packaged into secretory granules in AtT20 cells before its cleavage. J Cell Biol 105:155-162.
Tooze SA, Huttner WB (1990): Cell-free protein sorting to the regu- lated and constitutive secretory pathways. Cell 60337-847.
Van Nostrand WE, Wagner SL, Suzuki M, Choi BH, Farrow JS, Geddes JW, Cotman CW, Cunningham DD (1989): Protease nexin 11, a potent antichymotrypsin, shows identity to amyloid beta-protein precursor. Nature 341 546-549.
Van Nostrand WE, Schmaier AH, Farrow JS, Cunningham DD (1990): Protease nexin-I1 (Amyloid P-protein precursor): A platelet a-granule protein. Science 248:745-748.
Weidemann A, Konig G, Bunke D, Fischer P, Salbaum JM, Masters CL, Beyreuther K (1989): Identification, biogenesis, and lo- calization of precursors of Alzheimer’s disease A4 amyloid protein. Cell 57:115-126.
Zupan AA, Johnson Jr EM (1991): Evidence for endocytosis-depen- dent proteolysis in the generation of soluble nerve growth fac- tor receptors by A875 human melanoma cells. J Biol Chem 266: 15384-1 5390.
83~6425 -6429.
880-884.
Maruyama K, Kametani F, Usami M, Yamao-Harigaya W, Tanaka K (1991): “Secretase,” Alzheimer amyloid protein precursor se- creting enzyme is not sequence-specific. Biochem Biophys Res Commun 179: 1670-1676.
Matlin KS, Simons K (1983): Reduced temperature prevents transfer of a membrane glycoprotein to the cell surface but does not prevent terminal glycosylation. Cell 34:233-243.
Momson TG, Ward LJ (1984): Intracellular processing of the vesic- ular stomatitis virus glycoprotein and the Newcastle disease virus hemagglutin-neuraminidase glycoprotein. Virus Res 1 : 225-239.
Neufeld EF (1991): Lysosomal storage disease. Ann Rev Biochem
Oltersdorf T, Fritz LC, Schenk DB, Lieberburg I, Johnson-Wood KL, Beattie EC, Ward PJ, Blacher RW, Dovey HF, Sinha S (1989): The secreted form of the Alzheimer’s amyloid precursor pro- tein with the Kunitz domain is protease nexin-11. Nature 341: 144- 147.
Orci L, Ravazzola M, Amherdt M, Perrelet A, Powell SK, Quinn DL, Moore HH (1987a): The trans-most cristernae of the Golgi complex: A compartment for sorting of secretory and plasma membrane proteins. Cell 51:1039-1051.
Orci L, Ravazzola M, Storch MJ, Anderson RG, Vassalli JD, Perrelet A (1987b): Proteolytic maturation of insulin in a postGolgi event which occurs in acidifying clathrin-coated secretory ves- icles. Cell 49365-868.
Palmert MR, Podlisny MB, Witker DS, Oltersdorf T, Younkin LH, Selkoe DJ, Younkin SG (1989a): The beta-amyloid protein precursor of Alzheimer disease has soluble derivatives found in human brain and cerebrospinal fluid. Proc Natl Acad Sci USA 86:6338-6342.
Palmert MR, Siedlak SL, Podlisny MB, Greenberg B, Shelton ER, Chan HW, Usiak M, Selkoe DJ, Perry G, Younkin SG (1989b): Soluble derivatives of the @ amyloid protein precursor of Alzheimer’s disease are labeled by antisera to the @ amyloid protein. Biochem Biophys Res Commun 165:182-188.
Pfeffer SR (1987): The endosomal concentration of a mannose G- phosphate receptor is unchanged in the absence of ligand syn- thesis. J Cell Biol 105:229-234.
Ponte P, Gonzalez-DeWhitt P, Schilling J , Miller J , Hsu D, Green- berg B, Davis K, Wallace W, Lieberberg I, Fuller F, Cordell B (1988): A new A4 Amyloid mRNA contains a domain homol- ogous to serine protease inhibitors. Nature 33 1525-527.
Refolo LM, Salton SRJ, Anderson JP, Mehta P, Robakis, NK (1989): Nerve and epidermal growth factors induce the release of the Alzheimer amyloid precursor from PC-12 cell cultures. Bio- chem Biophys Res Commun 164:664-670.
Refolo LM, Wittenberg I, Friedrich Jr VL, Robakis NK (1991): The Alzheimer amyloid precursor is associated with the detergent- insoluble cytoskeleton. J Neurosci 11:3888-3897.
Robakis NK, Ramakrishna N, Wolfe G, Wisniewski HM (1987): Mo- lecular cloning and characterization of a cDNA encoding the
601257-280.
Copyright © 2022 FDOKUMEN




















