
Cleavage of di- and tripeptides by Prevotella ruminicola
8
Anaerobe (1997) 3,3542 P HYS I O LO GY/STR U CTU RAL B I O LO GY/B I OC H E M I STRY Peptidases of the Rumen Bacterium, Prevotella ruminicola R. John Wallace, Nest McKain, Glen A. Broderickl, Lyle M. Rodez, Nicola D. Walker, G. James Newbold and Jan Kopecny3 Rowett Research lnstitute, Bucksburn, Aberdeen AB21 9SB, U,K, (Received L4lune L996, accepted 20 Noaember 1996) Key Words: Preaotella ruminicola, dipeptidyl aminopeptidase, dipeptidyl peptidase, peptidase, rumen Preaotella (formerly Bacteroides) ruminicola is a numerous rumen bacterium which plays a significant role in the metabolism of proteins and peptides in the rumen. Measurement of the hydrolysis of synthetic aminopeptidase substrates by sonicated extracts and whole cells of different species of rumen bacteria indicated that P. ruminicola had the greatest range and specific activity of dipeptidyl peptidases among the species tested. Streptococcus boaishydrolysed some dipeptidyl peptidase substrates to a lesser extent, and several species broke down Ala2-p-nitroanilide, including Ruminobncter amylophilus, Ruminococcus spp. and Veillonella paraula. Dipeptidyl pepti- dases, which cleave dipeptides from the amino-terminus of longer peptides, were much more active than aminopeptidases removing single amino acids in P, ruminicola.Ion-exchange chromatography of sonicated extracts of P. ruminicola M384 revealed at least four distinct activities: one hydrolysed Ala2-p-nitroanilide, ValAla-p-nitroanilide, Alaa and Ala5; anothet was an O2-sensitive activity hydrolysing GlyArg-4-methoxynapthylamide, ArgArg- 4-methoxynaphthylamide, Gly5 and ValGlySerGlu, similar to dipeptidyl peptidase type I DPP-1); a third hydrolysed GlyPro-p-nitroanilide and GlyPro- -methoxynapthylamide and was similar to dipeptidyl peptidase type IV DPP-4); a fourth broke down LysAla-4-methoxynaphthylamide. A11 of the enzymes, and particularly those active against Ala2'p-nitroanilide and GlyPro-p-nitroanilide, were inhibited by serine protease inhibitors, and all except DPP-4 were inhibited by EDTA. Both DPP-1 and the enzyme hydrolysing LysAla-4-methoxynaphthylamide were inhibited strongly by iodoacetate. DPP-4 was inhibited completely by diprotin A. Competitive inhibition experiments suggested that DPP-1 was less important than the other enzymes in the breakdown of peptide mixtures' 1997 Academic press Address colrespondence to: Dr R.j. Wallace. E-mail [email protected] lPresent address: U.S. Dairy Forage Center, Madisory WI 53706-1108, U.S.A. ?Present address: Agriculture Canada Research Station, Lethbridge, Alberta TlI 481., Canada 3Present address: Institute of Animal Physiology and Genetics, Uhrineves, Prague 1.0, Czech Republic. 1"075-9964/ 97l010035 + 08 $25.00/0 / an960065 @ 1997 Academic Press
Transcript of Cleavage of di- and tripeptides by Prevotella ruminicola

Anaerobe (1997) 3,3542
P HYS I O LO GY/STR U CTU RAL B I O LO GY/B I OC H E M I STRY
Peptidases of the Rumen Bacterium, Prevotellaruminicola
R. John Wallace, Nest McKain, Glen A. Broderickl, Lyle M. Rodez,Nicola D. Walker, G. James Newbold and Jan Kopecny3
Rowett Research lnstitute,Bucksburn, Aberdeen AB21 9SB,U,K,
(Received L4lune L996,accepted 20 Noaember 1996)
Key Words: Preaotellaruminicola, dipeptidylaminopeptidase, dipeptidylpeptidase, peptidase, rumen
Preaotella (formerly Bacteroides) ruminicola is a numerous rumen bacteriumwhich plays a significant role in the metabolism of proteins and peptides inthe rumen. Measurement of the hydrolysis of synthetic aminopeptidasesubstrates by sonicated extracts and whole cells of different species of rumenbacteria indicated that P. ruminicola had the greatest range and specificactivity of dipeptidyl peptidases among the species tested. Streptococcusboaishydrolysed some dipeptidyl peptidase substrates to a lesser extent, andseveral species broke down Ala2-p-nitroanilide, including Ruminobncteramylophilus, Ruminococcus spp. and Veillonella paraula. Dipeptidyl pepti-dases, which cleave dipeptides from the amino-terminus of longer peptides,were much more active than aminopeptidases removing single amino acidsin P, ruminicola.Ion-exchange chromatography of sonicated extracts of P.
ruminicola M384 revealed at least four distinct activities: one hydrolysedAla2-p-nitroanilide, ValAla-p-nitroanilide, Alaa and Ala5; anothet was anO2-sensitive activity hydrolysing GlyArg-4-methoxynapthylamide, ArgArg-4-methoxynaphthylamide, Gly5 and ValGlySerGlu, similar to dipeptidylpeptidase type I DPP-1); a third hydrolysed GlyPro-p-nitroanilide andGlyPro- -methoxynapthylamide and was similar to dipeptidyl peptidasetype IV DPP-4); a fourth broke down LysAla-4-methoxynaphthylamide. A11
of the enzymes, and particularly those active against Ala2'p-nitroanilide andGlyPro-p-nitroanilide, were inhibited by serine protease inhibitors, and allexcept DPP-4 were inhibited by EDTA. Both DPP-1 and the enzymehydrolysing LysAla-4-methoxynaphthylamide were inhibited strongly byiodoacetate. DPP-4 was inhibited completely by diprotin A. Competitiveinhibition experiments suggested that DPP-1 was less important than theother enzymes in the breakdown of peptide mixtures'
1997 Academic press
Address colrespondence to: Dr R.j. Wallace.E-mail [email protected] address: U.S. Dairy Forage Center, Madisory WI53706-1108, U.S.A.?Present address: Agriculture Canada Research Station, Lethbridge,Alberta TlI 481., Canada3Present address: Institute of Animal Physiology and Genetics,Uhrineves, Prague 1.0, Czech Republic.
1"075-9964/ 97l010035 + 08 $25.00/0 / an960065 @ 1997 Academic Press

36
Introduction
R. J. Wallace et al.
Excessive protein breakdown and ammonia produc-tion by rumen microorganisms contribute to theinefficient utilisation of dietary N which occurs inruminants, and which causes wastage of nitrogenousfeed resources and environmental pollution [L]. Aspeptides are intermediates in the breakdown process/the mechanisms by which peptides are broken downare highly relevant to one of the most pressingproblems of ruminant livestock production.
Peptidase activity in the rumen was shown to bemainly bacterial in origio except for dipeptidasewhere ciliate protozoa make a significant contribution["4]. Oligopeptides of alanine, Ala5 and Alaa, werecleaved by the release of Ala2rather than free Ala fromthe N-terminus [3-5], and other peptides appeared tobe broken down by a similar mechanism [3,5],suggesting that dipeptidyl peptidase (DPP; formerlyknown as dipeptidyl aminopeptidase [6]) activity wasinvolved. From the hydrolysis of synthetic substratesby whole cells, the main activity in ruminal fluidappeared to be dipeptidyl peptidase type I (DPP-1;
[7]). Screening of pure cultures of rumen bacteria [8]and selective isolation of DPP-1 positive bacteria [9]showed that only Preaotella ruminicola of the mostcommon rumen bacteria possessed significant DPP-1activity. The involvement of P. ruminicola in ruminalpeptide metabolism [10-13] and the fact that P.
ruminicola can on occasion constitute more than 60% ofthe total eubacteria present in the rumen [14] havebeen known for many years. This study was under-taken to investigate lhe dipeptidyl peptidases of P.
ruminicola with regard to their importance relative toaminopeptidases cleaving single amino acids, theirsensitivity to protease inhibitors, and their likelysignificance in the catabolism of peptides by P.
ruminicola.
Methods
Bacterial strains and growth medium
Preaotella ruminicola M384 was isolated from therumen of a sheep [15], and is maintained in the culturecollection of the Rowett Research Institute. P. rumini-cola and other strains of rumen bacteria, for which thesources were given previously [8], were grown at39"Cin the liquid form of medium M2 of Hobson [16],which is a general purpose medium containing rumi-nal fluid.
Screening of pure cultures of rumen bacteria fordipeptidyl peptidase sctiaities
Overnight, stationary-phase cultures of bacteria wereharvested by centrifuging at27 500 g for 15 min at 4'C.Pellets were washed once with anaerobic 25 mMpotassium phosphate pH7.0, resuspended to hall theoriginal volume in the same bufery and" used inpeptidase assays in multiwell plates with 4-methox-ynaphthylamide (MNA) substrates similar to thosedescribed before [7], except that a fluorimetric platereader with 365 and 450 nm excitation and emissionfilters respectively was used to measure MNA release.AII operations except the incubation of the plates werecarried out under CO2. The assay mixtures contained201tL of 2.5mM substrate dissolved in buffet, exceptLeuVaI-MNA which was dissolved in ethanof 50 pL4mM dithiothreitol (DTT), 809,L water and 50pL of '
cell suspension or extract. The plates were incubated '-u -at 39"C, and the rate of reaction was calculated fromthe initial rate of release of MNA. Activity against .
Ala2-p-anilide (Ala2-pNA) was determined in an assaycontaining 100pll of 4mM substrate dissolved inanaerobic 25mM potassium phosphate buffer pI{7.0,50pL 4mM DTT and 50pL of cell suspension. Theassay was carried out in multiwell plates incubated at39'C in an optical plate reader with a 420nmfi1ter andresults were calculated from pNA release, usually aftert h incubation. Possible metabolism of the products ofthe peptidase assays was checked with each strain inincubations with 0.25mM MNA or 0.5mM pNA inplace of the substrate.
Peptidase actiaities o/ P. ruminicola M384
The activity of P. ruminicola M384 against a range ofdipeptidyl- and amino acyl-pNA substrates was inves- \.-z-tigated using sonicated washed cell suspensions.Overnight cultures (200 mL) were harvested by cen-trifugation as before, washed and resuspended in20 mL of anaerobic buffer, then sonicated. The wholesonicated suspension was used in peptidase assayssimilar to that described above for Ala2-pNA, exceptthat the final concentration of substrates in the assaywas 1mM.
The influence of air on peptidase activities wasinvestigated using sonicated suspensions incubated inHungate tubes under CO2, except when air wasdeliberately introduced into the tube. Immediatelybefore incubatiory 2.0 mL of suspension were dilutedin 98mL of either air-saturated 25mM potassiumphosphate buffe1, pH 7.5, or CO2-saturated 25mMpotassium phosphate buffet, pH7.5, containing 2mMcysteine HCI in place of DTT in order to preventinterference in HPLC analysis of alanine peptides.TWelve millilitres of diluted suspension were then

Peptidases ol P. ruminicola 37
added to 4mL of a solution containing 1mM Ala5,ValGlySerGlu, G1y5 LeuGly2, Ala2-pNA or GlyArg-MNA dissolved in the corresponding aerobic oranaerobic buffer. Samples (2mL) were removed peri-odically into 0.5 mL of 1..25M phosphoric acid, centri-fuged and analysed for remaining peptides by HPLCl7l, p-nitroaniline by its absorbance at 420nrr. or4-methoxynaphthylamine by fluorimetry.
Chromatographic characterisation of peptidase actiaities
Anion-exchange chromatography was done usingeither a Pharmacia MonoQ column as describedpreviously l17l or a Pharmacia BioresourcerM acolumn. The procedure used with the latter columnwas as follows. Sonicated P. ruminicola M384 cells werecentrifuged as before to remove cell debris andunbroken cells, then 1pg ml-l DNase (type I, bovinepancreas; Sigma) and RNase (type I-A, bovine pan-creas; Sigma) were added. The mixture was incubatedat 39"C for t h, then centrifuged at 42 000 g for t h. Thesupernatant was washed five times with 50 mM Trisbuffer pH7.0 containing 1 mM DTT and concentratedusing a Centricon-L0 (Amicon; molecular weight cut-off 10 000). Portions (0.2 mL) of extract were applied tothe BioresourcerM Q (1.0mL) strong anion exchangecolumn and eluted with 50mM Tris pH 8.0 buffercontaining 1mM DTT with increasing NaCl concen-trations controlled by a Waters 650E liquid chromatog-raphy system. Fractions were analysed for peptidaseactivities as described before, except that the samplesize was 130 pL and the concentration of pNA sub-strates in the assay mixture was 0.4mM. An incuba-tion time of 2 h was used with all substrates. The effectof air on the activities of each fraction was determinedby omitting DTT from the assay and bubbling themixture with air.
Wuence of protease inhibitors and competing peptides
Cultures were growry concentrated 20-fold in anaero-bic buffel, and sonicated as before. The influence ofprotease inhibitors on the hydrolysis of GlyArg-MNA,LysAla-MNA, Ala2-pNA, and GIyPro-pNA was deter-mined in continuous assays using plate readers as
described above, with final concentrations in the assayof 0.25mM MNA substrates and 1mM pNA sub-strates, and the incubation time was 60min. Theprotease inhibitors were added in 20 pL of appropriatesolvent (see Table 3), followed by substrate and finallythe diluted, sonicated cell suspension.
The influence of peptide mixtures on different DPPactivities was determined using a pancreatic caseinhydrolysate, Trypticase (BBL), and Special Peptone(Oxoid), a mixture of peptides from meat, plant and
yeast hydrolysates. The peptides were added to a finalconcentration of 2.6'Lmg/mL, equivalent to an aver-age peptide concentration of 5mM assuming anaverage molecular weight of 52'LDa [18]. The pepti-dase substrates were added to a final concentration of1mM in continuous assays similar to those describedabove.
Protein determination
Protein in whole cells and sonicated suspensions wasdetermined using the Folin reagent following alkalinedigestion t19l with bovine serum albumin as
standard.
Results
Dipeptidyl peptidase actiuities of pure cultures of rumenbacteria
TWenty strains of 'L4 species of common rumenbacteria were screened for their activities against a
range of dipeptidyl peptidase substrates (Table 1). Inpreliminary experiments, sonicated bacteria werecompared with intact washed cells in screening foractivity. No qualitative differences were observedbetween the types of preparatiory so whole cells wereused for this part of the study. MNA substrates werealso used because they provide a wider range ofsubstrates than is available commercially with p-ni-troanilides. The only exception was Ala2-pNA,because we were unable to find a commercial sourceof Ala2-MNA.
Significant dipeptidyl peptidase activity wasdetected only in six species, P. ruminicola, Ruminobacteramylophilus, Ruminococcus albus, Ruminococcus flaaefa-ciens, Streptococaus boais and Veillonella paraula. P.
ruminicola had the highest activities, particularlytowards Ala2-pNA and GlyArg-MNA. R. arnylophilus,R. albus, R. flauefaciens, S. boais and V. paraula hadsignificant activity against AIa2-pNA. S. boais alsobroke down GlyPro-MNA at a rate only slightly lessthan P. ruminicola. Hydrolysis of MNA or pNA wasminor in most species (Table 1), so major activitieswould not have been obscured.
Peptidase actiz;ities o/ P. ruminicola with dipeptidyl- andamino acyl-pN A substrates
The peptidase activities of sonicated P. ruminicolaM384 against pNA substrates (Table 2) demonstratedthat activity against dipeptidyl-pNA substrates was

3B R.J. Wallace et al.
Table 1. Hydrolysis of dipeptidyl peptidase substrates by rumen bacteria
Rate of hydrolysis (nmol/mg protein/min)
Species Alar-pNA ArgArg-MNA GlyArg-MNA GlyPro-MNA LysAla-MNA LeuVal-MNA pNA MNA
Anaer ouibrio lip olytica 5SButyriaibrio fibrisola ens SH13Eubacterium ruminantium 2388F ibr ob act er su ccino gene s 585Lachnospir a multipar a D32Me gasphaer a el s denii LClMit suolce lk m ult iacidu s 46 / 5Preootella ruminicola M384Ruminob act er amy lophilu s W P 225Ruminococcus albus SY3Ruminococcus fla,t efaciens FDlf elenomonas ri,miiantium 2108Streat oco ccus boztis ES'J.
Veilion ell a p ar a ul a L59
00.20.20,1000
1.6.9).q1^
5.403.46.0
000000
0
000000
000
0
0
0
06.r0.9000J,J
0
000000
026.0
0
0000.40
0,
0
0
0
0009.1.
00
0
0
0.60
001.302.000 0.8 0,80 0.2. 0000.50 0.4 0.80 0.3 1..2
0.3 1".4 0.10000 1,.4 0000000000000
W9sled cell suspensions of rumen bacteria were incubated at 39 "C with 0.25 mM dipeptidyl peptidase substrates, except for Alar-pNAwhich was 2 mM. Initial rates of release of 4-methoxynaphthylamine (MNA) and p-niiroiniline 1pt'tA) were measured flriorimetriciliy orby.increase in absorbance resPectively in continuous assays. Results are the means of incubations with cell suspensions from three separatecultures.
much greater than against amino acyl-pNA substrates.Activity against Ala2-pNA was highest, but GlyPro-pNA was broken down more rapidly than GlyArg-pNA, in contrast to the MNA substrates (Table 1).Amino acyl peptidases were apparently absent. How-eve4 although no pNA appeared to be released fromamino acyl-pNA substrates under these conditions,there was a background activity 9f p'nitroanilinemetabolism which would obscure activities less than5% ol the activity against Ala2-pNA. Thus, the aminoacyl-pNA activities may have been present but theywere an order of magnitude lower than DPPactivities.
lnfluence of air on peptidase actiaities and peptidebreakdown
The influence of ai-r on peptidase activities wasdetermined by diluting sonicated cells or fractions
Thble 2, Hydrolysis of dipeptidyl and amino acyl p-nitroanilidesby a sonicated suspension of Preaotella ruminialaMSS4
p-Nitroanilide(1 mM)1
p-Nitroaniline released(nmol/mg protein/min)z
Mean SD
generated from liquid chromatography in aerated -buffeX, or in anaerobic buffer containing DT!, andincubating under air or CO2. GlyArg-MNA break-down by both types of extract was abolished by ai4 aswas ArgArg-MNA breakdown in the HPLC fractions.Gly5 and ValGlySerGlu breakdown by the sonicatedcells were decreased by 78 and 94/o respectively by air.In contrast, breakdown of Ala5 by sonicated cellsdecreased by less than2}% in the presence of ai4 andAIa5 continued to be cleaved to Ala3 and Ala2, with noAlaa formed. The rates of breakdown of Ala2-pNA,LysAla-MNA, GlyPro-MNA and LeuVal-MNA inHPLC fractions were unaffected by air (results notshown).
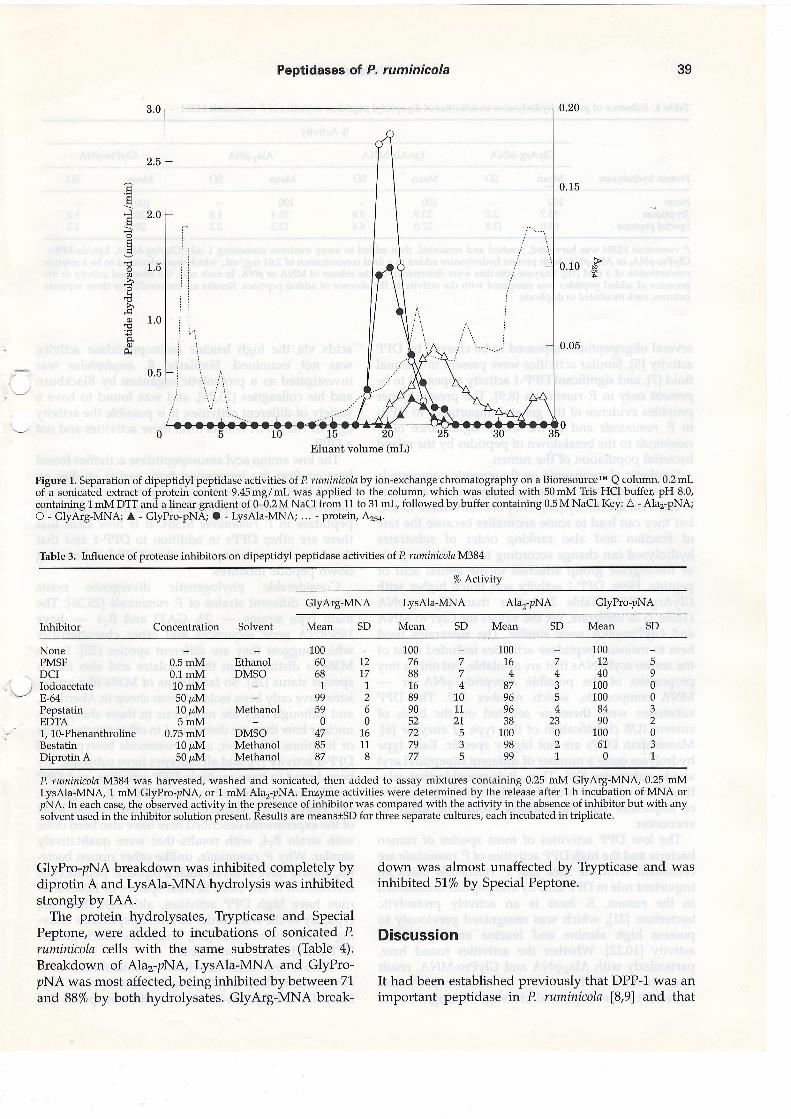
It was shown previously by ion-exchange chroma-tography that P. ruminicolil produces separate cyto-plasmic di, tri- and oligopeptidases U7l. A similar',--.]'anion-exchange chromatographic separatiory with amuch shallower salt gradient,'was done with Pruminicola M384 to separate the dipeptidyl peptidaseactivities (Figure 1). Activities against Alar-pNA,GlyArg-MNA and GlyPro'pNA eluted in separatepeaks. Activity against A1a5, Alaa and ValAla-pNA co-eluted with activity against Alar-pNA. Activity againstArgArg-MNA, GlyArg-pNA, ValGlySerGlu andLysAla-MNA coincided with activity against GlyArg-MNA. Only GlyPro-MNA was broken down by theGlyPro-pNA-hydrolysing peak.
Activities against GlyArg-MNA, LysAla-MNA,Ala2-pNA and GlyPro-pNA were tested for theirsensitivity to protease inhibitors in sonicated cells(Table 3). DCI inhibited all of the activities to somedegree, particularly with Ala2-pNA and GlyPro-pNA,as did the other serine protease inhibitor, PMSF.GtyArg-MNA breakdown was more sensitive to IAAand EDTA, which both inhibited activity completely.
Ala, 7,54Gly'Arg 1.32GlyPro 3:60ValAla 4.54
Assays were carried out with whole sonicated cell suspension.Results are meanstSD for samples from three separate cultures ofP, ruminicolaM384, each carried out in duplicate,1No pNA was detected from Ala-, Arg-, Asp-, GIu-, Gly-, Leu-,Met-, Pro-, Tln- or Val-p-nitroanilide.rp-Nitroaniline was metabolised at 0.4 nmol mg protein min byP, ruminicola when added to the reaction mixture at a concentrationof 0.25 mM. No correetion was made for this activity.
1,.21
0.500.580.67
Peptidases of P. ruminicola
0.20
Figure 1. Separation of dipeptidyl peptidase activities of P, ruminicola by ion-exchange chromatography on a BioresourcerM Q column. 0.2 mLof a sonicated extract of protein content 9.45rr.g/mL was applied to the column, which was eluted with 50mM Tris HCI buffer, pH 8.0,containing 1 mM DTT and a linear gradient of 0-0.2 M NaCl from 11 to 31 mL, followed by buffer containing 0.5 M NaCl. Key: A - Ala2-pNA;O - GlyArg-MNA; A - GlyPro-pNA; O - LysAla-MNA; ... - protein, A25a.
Table 3. Influence of protease inhibitors on dipeptidyl peptidase activities of P rarzlrlcola M384
lo AclivilY
GlyArg-MNA LysAIa-MNA Alar-pNA GlyPro-pNA
Inhibitor Concentration Solvent Mean SD Mean SD Mean SD Mean SD
39
2.5
10F
d
Fl 20
=o r'dah
.dhq 1.0E
o
Eluant volume (mL)
NonePMSFDCIIodoacetateE-64PepstatinEDTA1, 10-PhenanthrolineBestatinDiprotin A
0.5 mM Ethanol0.1 mM DMSO10 mM5o g.M10 pM Methanol5mM
0.75 mM DMSO10 p.M Methanol50 pM Methanol
P. ruminicola M384 was harvested, washed and sonicated, then added to assay mixtures containing 0.25 mM GlyArg-MNA, 0.25 mMLysAla-MNA, 1 mM GlyPro-pNA, or 1 mM Alar-pNA. Enzyme activities were determined by the release after t h incubation of MNA orpNA. In each case, the observed activity in the presence of inhibitor was compared with the activity in the absence of inhibitor but with anysolvent used in the inhibitor solution present. Results are means:tsD for three separate cultures, each incubated in triplicate.
100-100*100-10060127671677256817887444091.L1,648731000
992891096510005969011,9648430052213823902
47L6725100010008511.79398261387877599101.
GlyPro-pNA breakdown was inhibited completely bydiprotin A and LysAla-MNA hydrolysis was inhibitedstrongly by IAA.
The protein hydrolysates, Trypticase and SpecialPeptone, were added to incubations of sonicated P.
ruminicola cells with the same substrates (Table 4).
Breakdown of Ala2-pNA, LysAla-MNA and GlyPro-pNA was most affected, being inhibited by between 71
and 88% by both hydrolysates. GlyArg-MNA break-
down was almost unaffected by Trypticase and wasinhibited 51,% by Special Peptone.
Discussion
It had been established previously that DPP-1 was animportant peptidase in P. ruminicola 18,91 and that

40 R.J. Wallace et al.
Table 4' Influence of protein hydrolysates on activities of dipeptidyl peptidase activities of P. ruminicolaMsS4
% Activity
GlyArg-MNA LysAla-MNA Alar-pNA GIyPro-pNA
Protein hydrolysate Mean
NonetypticaseSpecial peptone
10095.769.0
2.011.8
1.82.2
10025.412.5
1..2
1.3
s.a8.4
10023.927.0
10020.329.4
P. ruminicola M3B4 was harvested, washed and sonicated, then added to assay mixtures containing 1 mM GlyArg-MNA, LysAla-MNA,GlyPro-pNA, or Alar-p-NA. with proteinhydrolysates-added to a final concentration ot2.6L mg/rr.L, which wai caliulated to-be a peptideconcentration of 5 mM [18]. Enzyme activities were determined by the release of MNA or pNA. In each case, the observed activit] iir thepresence of added- peptides- wq9 compared with the activity in the absence of added peptides. Results are meanstsD for three ieparatecultures, each incubated in duplicate.
several oligopeptides appeared to be cleaved by DPPactivity [5]. Similar activities were present in ruminalfIuidl7l, and significant DPP-L activity appeared to bepresent only in P. ruminicola [8,9]. The present paperprovides evidence of the general importance to DPPsto P. ruminicola and of the likely significance of Pruminicola to the breakdown of peptides by the mixedbacterial population of the rumen.
Synthetic chromogenic or fluorogenic compoundsare used frequently for measuring peptidase activitiesof different specificity [20]. They are convenient to use,but they can lead to some anomalies because the rateof reaction and also ranking order of substrateshydrolysed can change according to the chromogenicor fluorogenic group attached to the amino acid orpeptide. Here, DPP-1 activity was much higher withGlyArg-MNA (Table 2) rather than GlyArg-pNA(Table 1) as substrate, yet the values for GlyPro-MNAand GlyPro-pNA were similar. The substrates usedhere to measure peptidase activities included most ofthe amino acyl-pNAs that are available, but only a tinyproportion of the possible dipeptidyl-pNA or -MNA compounds, which number 400. The DPPsubstrates were therefore selected on the basis ofcurrent IUB classification of this type of enzyme [6].Mammalian DPPs are not highly specific. Each typehydrolyses quite a number of different dipeptidyl arylamide substrates [20]. Thus, it is possible that rela-tively few enzymes could hydrolyse the large numberof sequences that the bacteria would be likely toencounter.
The low DPP activities of most species- of rumenbacteria and the high DPP activities of P. ruminicola arcconsistent with the idea that P. ruminicola plays animportant role in DPP activity and peptide breakdownin the rumen. S. boais is an activelv proteolvticbacterium [21], which was recognized prwiously topossess high alanine and leucine aminopeptidaseactivity [10,22]. Whether the activities found here,particularly with Ala2-pNA and GlyPro-MNA, resultfrom genuine DPP activity, or whether pNA wasreleased by the sequential cleavage of single amino
acids via the high leucine aminopeptidase activity, :
was not examined. Similarly, R. amyLophilus wasinvestigated as a proteolytic organism by Blackburn, iand his colleagues [23,24], and was found to have 3 v.variety of different activities; it is possible the activityagainst Ala2-pNA results from these activities and not vA DPP.
The low amino acyl aminopeptidase activities foundhere confirm the impression from earlier studies onDPP-1 [8,9] and the pattern of breakdown of oligopep-tides by P. ruminicola [45] that DPP is the main type ofpeptidase in P. ruminicola M384. It is now clear thatthere are other DPPs in addition to DPP-1 and thatthese may be more important than DPP-1 in breakingdown peptide mixtures.
Considerable phylogenetic divergence existsbetween different strains of P. ruminicola 125,261. Themain type strains - 23, GA33 and B14 - havel6SrRNA gene sequences and other characteristicswhich suggest they are different species [25]. StrainM384 is distinct from these isolates and also meritsspecies status [26]. So fal strains of M384like organ- \isms have only been isolated from sheep in Aberdeery vand although they are numerous in these sheep, it isunclear how they are distributed in other species, dietsor locations. Howevel, all P. ruminicola biotypes hadDPP-1 activity [9], and all biotypes have subsequentlybeen found to have multiple DPP activities, and DPPprofiles are a useful aid to identification (G. Avgu5tin,H.f. Flint and R.J. Wallace, unpublished results). Mostof the experiments described here have also been donewith strain Br4, with results that were qualitativelysimilar. Why P. ruminicola, unlike other rumen bacte-ria, should have evolved such a biphasic peptidolyticmechanism is not clear. Several species of Bifidobacte-rium have high DPP activities, although these aretypically of the same order as aminopeptidases cleav-ing single amino acids 1271, unlike the much greaterDPP activities in P ruminicola.
At least four main activities were identified in Pruminicola, hydrolysing GlyArg-MNA, Ala2-pNA, Gly-Pro-pNA and LysAla-MNA. The different peaks did

Peptidases oI P. ruminicola 41
not hydrolyse more than one of the four main DPPsubstrates, and they were distinct from dipeptidaseand tripeptidase activities studied before [17]. Preciseidentification of the enzyme types would requirecharacterization of purified enzymes. Howevel, incu-bations of mixed extracts with protease inhibitorsprovided useful information about each of theactivities.
GlyArg-MNA breakdown was inhibited to someextent by serine protease inhibitors, but, as with DPP-1 enzymes of mammalian origin for which GlyArg-MNA is a diagnostic substrate [20]' lt was mainlysensitive to iodoacetate and EDTA, suggesting thatsulfhydryl groups and metal ions are important forcatalysis. The cysteine protease inhibitor, E-64, had no
. effect on GlyArg-MNA breakdown/ however, suggest-
- ing that iodoacetate inhibited GlyArg-MNA break-
- . down by reacting with DTT, which protects the- '-' enzyme from the effects of 02. An enzyme of the DPP-
1 type does not appear to have been reported in any'- bacterial species other than P. ruminicola. This enzyme
differs significantly from DPP-1 of mammalian originin its 02 sensitivity and that ArgArg-MNA was alsohydrolysed by the P. ruminicola DPP-I, whereas it isnot a substrate for mammalian DPP-1 [20].
Activity against Ala2-pNA was much more sensitiveto serine protease inhibitors and seemed to be a typicalserine protease, similar to mammalian DPPs otherthan DPP-1 [20]. Ala2-pNA is not a recognised sub-strate for any of the mammalian DPP enzymes/although it is broken down slowly by DPP-1 and DPP-2 1201. Unlike DPP-? 1281, it had no activity towardstripeptides 1171. At the present time, we consider thisenzyme to be different to the recognised DPPs, anduse the abbreviation Ala-DPP because of its affinity forX-Ala-peptides.
The third activity, hydrolysing GlyPro-pNA and':-\-'GlyPro-MNA, is fairly iypicat otllpp-+ enzymes, in
that it had a restricted range of substrates, it wasinhibited by serine protease inhibitors, and it was- inhibited by diprotin A [20,29]. DPP-4 is fairly wide-spread among microorganisms, and is found forexample, in several genera of lactic acid bacteria
t301.LysAla-MNA is a diagnostic substrate for DPP-2
[20]. Howevel, the mammalian DDP-2 is a serineprotease, unLike the IAA-sensitive enzyme of P.
ruminicola. Like DPP-I, it may have a dependence onmetal ions for activity, as it was sensitive to EDTA.
The DPPs of P, ruminicola are therefore significantlydifferent in their properties to the correspondingenzymes from marnmalian sources. As yet, it is notclear if any of these enzymes is more important thanthe others. Competitive inhibition suggested thatDPP-L was inhibited least by a peptide mixture, whilethe other enzymes were inhibited to similar extents.The peptide mixtures used were Trypticase, a pancre-
atic hydrolysate of casein with an average molecularweight of about 500 [18,31] and Special Peptone, amixture of pancreatic digests of meat, plant and yeastwith a similar wide molecular weight spectrum toTrypticase [18]. Thus, a range of potentially competingpeptides is present in both hydrolysates. Analysis ofpeptidase mutants will be necessary to establish
-tri"t'r, if any, of these DPPs plays a gteater rol-e thanothers in dissimilatory peptide breakdown by Pruminicola. It will also be important to examine howDPPs are regulated, the effects of growth rate andphase on DPP activity and the influence of peptideand amino acid concentration on their expression. Thebacterial cultures examined here were all in stationaryphase, which is probably typical of the slow growthduring most of the daily cycle in meal-fed sheep.However, they were atypical in that the medium wasmuch richer in nutrients than would occur duringmost of the daily cycle in aiao.
Acknowledgements
This work was supported by Royal Gist-brocades nv Delft, TheNetherlands, and the Scottish Office Agriculture and FisheriesDepartment. Glen Broderick was supported by the U.S' Departmentof Agriculture. Lyle Rode was funded by Agriculture Canada. JanKopecny received a Fellowship from the Scottish Office Agricultureand Fisheries Department.
References
1. Leng R.A. and Nolan j.V. (1984). Nitrogen metabolism in therumen. / Dairy Sci 70:1072-1089
2. Newbold C.J., Wallace R.J. and McKain N. (1989) The role ofprotozoa in ruminal peptide metabolism. In Lloyd D., CoombsG.H. and Paget T.A. (eds) Biochemistry and Molecular Biology of'Anaerobic' Protozoa, pp. 42-55. Harwood Academic Press,London
3. Wallace R.J., McKain N, and Newbold C.l. (1990) Metabolism ofsmall peptides in rumen fluid. Accumulation of intermediatesduring hydrolysis of alanine oligomers, and comparison ofpeptidolytic activities of bacteria and protozoa. J Sci Food Agric50: L9l-T99
4. Wallace R.J., Newbold C.J. and McKain N. (1990) Patterns ofpeptide metabolism by rumen microorganisms. In Hoshino S.,
Onodera R., Minato H. and Itabashi H. (eds) The RumenEcosystem. The Microbial Metabolism and its Control, pp. 43-50.
fapan Scientific Societies press, Tokyo5. Wallace R.J., McKain N. and Broderick G.A. (1993) A compar-
ison of the breakdown of pure peptides by Bacteroides ruminicolaand mixed microorganisms from the sheep rumen. CurrMicrobiol 26:333-i36
6. Webb E.C. (1992) Enzyme Nomenclature L992. Academic Press,London
7. Wallace R.J. and McKain N. (1989) Analysis of peptide metabo-lism by ruminal microorganisms. Appl Enoiron Microbiol 552
2372-23768. Wallace R.J. and McKain N. (1991) A survey of peptidase
activity in rumen bacteria. ] Gen Microbiol 737:2259-22649. McKain N., Wallace R.J, and Watt N.D. (1992) Selective isolation
of bacteria with dipeptidyl peptidase type I activity from thesheep rumen. FEMS Microbiol Lett 95:1'69-L74

42 R.J, Wallace et al.
10. Bladen H.A., Bryant M.P. and Doetsch R.N. (1961) A study ofbacterial species from the rumen which produce ammonia fromprotein hydrolyzate. AppI Microbiol 9: 1i5-180
11. Pittman K.A. and Bryant M.P. (1964) Peptides and othernitrogen sources for growth oI Bacieroides ruininicota, J Bacteriol88: 401-410
12, Russell J.B. (1983) Fermentation of peptides by Bacteroidesruminicola B$. Appl Enoiron Microbiol 45: 156G1.574
13. Ling J.R. and Armstead LP. (1995) The in vitro uptake andmetabolism of peptides and amino acids by five species ofrumen bacteria, J Appl Bacteriol 7& 1'J.6-124
14. Van Gylswyk N.O. (1990) Enumeration and presumptiveidentification of some functional groups of bacteria in the rumenof dairy cows fed grass silage-based diets. FEMS Microbiol Ecol73t 243-254
15. Wallace R.J. and Brammall M,L. (1985) The role of differentspecies of bacteria in the hydrolysis of protein in the rumen. /Gen Microbiol 13'L: 821-832
16. Hobson P.N. (1969) Rumen bacteria. In Norris J.R. and RibbonsD.W (eds) Methods in Microbiology, vol 38, pp. 133-145.Academic Press, London
17, Wallace R,j., KopecnyJ., Broderick G.A., Walker N.D., Sichao L.,Newbold C.j, and McKain N. (1995) Cleavage of di- andtripeptides by Preaotella ruminicola, Anaerobe l: 335-343
18. Wallace R.J. (1992) Gel filtration studies of peptide metabolismby rumen microorganisms. J Sci Food Agric 58:177-TB4
19. Herbert D., Phipps P.J. and Strange R.E. (1971) Chemicalanalysis of microbial cells. In Norris f.R. and Ribbons D.W. (eds)Methods in Microbiology, vol 58, pp. 209-444. Academic Press,London
20. McDonald j.K. and Barrett A.J. (1986) Mammalian Proteases: AGlossary and Bibliography, Vol. 2, Exopeptidases. Academic Press,London
21. Russell J.B,, Bottje, W.G. and Cotta M.A. (1981) Degradation ofprotein by mixed cultures of rumen bacteria: identification ofStreptococcus boais as an actively proteolytic rumen bacteria. /Anim Sci 53t 242-252
22. Russell |.B. and Robinson P.H. (1984) Compositions andcharacteristics of strains oI Streptococcus boztis, J Dairy Sci 67:L525-I531,
23. Blackburn T.H. (1968) The protease liberated fuom Bacteroidesamylophilus strain HlB by mechanised disintegration. ,l GenMicrobiol 53:37-5L
24. Blackburn T.H. & Hullah W. A. (L974) The cell-bound protease ofBacteroides amylophilus H18. Can J Microbiol 20l. 43544I
25. Mannarelli B.M., Ericsson L.D., Lee D. and Stack R.J. (1991)Taxonomic relationships among strains of the anaerobic bacte-
tium Bacteroides ruminicola determined bv DNA and extrac-ellular polysaccharide analysis. Appl Enairon Microbiol 57:2975-2980
26. Avgudtin G., Wright F. and FLint H.l. (1994) Genetic diversityand phylogenetic relationships among strains of Prevotella(Bacteroides) ruminicola from the rumen, Inl I Sust Bacteriol 44:.
246-25527. El-Soda M, Macedo A. and Olson N.F. (1992) The peptide
hydrolase system ol Bit'idobacterium species. MilchwissenschaJt 47:87-90
28. McDonald J.K., Leibach F.H., Grindeland R.E. and Ellis S. (1968)Purification of dipeptidyl peptidase II (dipeptidyl arylamidaseII) of the anterior pituitary glan{ peptidase and dipeptideesterase. J Biol Chem 243 41,4341,50
29. Umezawa H., and Aoyagi T. (1983) Trends in research of lowmolecular weight protease inhibitors of microbial origin. InKatunuma N., Umezawa H. and Holzer H. (eds) ProteinaseInhibitors, pp. 3-15. Japanese Scientific Societies Press, Tokyo
30. Casey M.G. and Meyer J. (1985) Presence of X-prolyl dipeptidylpeptidase in lactic acid bacteria. I Dairy Sci 68;3212-32L5
3L. Chen G., Russell J.B. and Sniffen C.j. (1987) A procedure formeasuring peptides rn rumen fluid and data suggesting thatpeptide uptake is a rate-limiting step in ruminal proteindegradation. I. Dairy Sci70: I2LL-L219
Appendix
Abbreaiations
DPP - dipeptidyl peptidaseDCI - 3, -dichloroisocoumarinDTT - dithiothreitolE-64 - L-trans-epoxysuccinyl-leucylamido(4-guanidino)butaneIAA - iodoacetic acidMNA - 4-methoxynaphthylamidePMSF - phenylmethylsulfonyl fluoridepNA - p-nitroanilide




















