
Elevation of cytoskeletal protein breakdown in aged Wistar rat brain
9
UNCORRECTED PROOF NBA 6255 1–9 Neurobiology of Aging xxx (2005) xxx–xxx Elevation of cytoskeletal protein breakdown in aged Wistar rat brain 3 Eric Bernath b , Nancy Kupina b , Ming Cheng Liu b,c , Ronald L. Hayes a,b,c , Colleen Meegan b,c , Kevin K.W. Wang a,b,c,d,∗ 4 5 a Department of Psychiatry, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida, Gainesville, FL 32610, USA 6 b Department of Neuroscience, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida, Gainesville, FL 32610, USA 7 c Center for Traumatic Brain Injury Studies, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida, Gainesville, FL 32610, USA 8 d Center for Neuroproteomics and Biomarkers Studies, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida, Gainesville, FL 32610, USA 9 10 Received 27 April 2004; received in revised form 4 February 2005; accepted 25 February 2005 11 Abstract 12 Previous studies indicated there is an overall increase of proteolysis in aging rat brains. We monitored the potential degradation of cytoskeletal proteins in neuronal tissue taken from cerebral cortex and cerebellum of young (3 month) and aging (17, 21 and 23.5 month) Wistar rats. We found significant age-dependent proteolysis of cytoskeletal proteins (II-spectrin and microtubule-associated protein MAP-2A/B) in the cerebral cortex and the cerebellum. The pattern of II-spectrin breakdown shows a marked increase in 150- and 145-kDa fragments (SBDP150 and SBDP145, respectively), but we did not detect the caspase-3-mediated 120-kDa fragment (SBDP120) in aged rat brains, suggesting the involvement of the calpain proteases. The pattern of MAP-2A/B breakdown in aged rat brains mirrors that produced by in vitro calpain digestion of 3-month control rat brain MAP-2A/B. In aged rat brains, there is no significant increase in pro-caspase-3 processing; rather, there is a moderate reduction in pro-caspase-3 protein and caspase-3 hydrolytic activity in the cortex. These results point to selective susceptibility of cytoskeletal proteins to calpain-mediated degradation, but not caspase-3 in aging rat brains. 13 14 15 16 17 18 19 20 21 © 2005 Published by Elsevier Inc. 22 Keywords: Aging; Alzheimer’s disease; Calpain; Caspase; MAP-2A/B; II-spectrin 23 24 1. Introduction 25 Dysfunction in the homeostatic regulation of calcium 26 (Ca 2+ ) has been one of the many notable theories pro- 27 posed to account for age-related neuronal degradation 28 [7,12,13,16,30,56,59] and (of more immediate therapeutic 29 potential) neurodegenerative diseases such as stroke and 30 Alzheimer’s disease [6,49]. Our study was designed to focus 31 on experimental evidence supporting the calcium hypothesis 32 for aging [29,54]. Concerning this theory, past investigators 33 have focused on studying the regulation of Ca 2+ in intra- 34 cellular and extracellular areas and its dysfunctional link to 35 degenerative alterations in the physiology of neuronal activ- 36 ities. Of specific interest are the effects of Ca 2+ on synaptic 37 transmissions [36,53] and hypoxic-ischemic cellular death 38 ∗ Corresponding author. Tel.: +1 352 294 0031; fax: +1 352 392 2579. E-mail address: [email protected] (K.K.W. Wang). [9,24]. Many laboratories, including ours, have focused on 39 the secondary dysfunctional role of Ca 2+ on cellular physiol- 40 ogy and the potential role of calpain-overactivation in causing 41 neurodegenerative responses [53,60]. It has been shown in a 42 number of studies that a sustained elevation in intracellu- 43 lar Ca 2+ triggers calpain activation, which leads to cellular 44 damage [5,21,24,51]. Activated calpain exerts its damage by 45 degrading various cellular substrates such as II-spectrin, 46 calmodulin binding proteins, microtubule-associated pro- 47 teins (MAPs) and neurofilaments. These processes are of- 48 ten associated with oncotic necrosis (oncosis). However, cer- 49 tain pathophysiological conditions (such as hypoxia or mod- 50 erate excitotoxicity) can also trigger activation of the pro- 51 apoptotic protease caspase-3. Like calpain, caspase-3 can de- 52 grade a large number of neuronal proteins, leading to apopto- 53 sis [57]. In fact, both caspase-3 and calpain-mediated prote- 54 olytic events have been implicated in neurodegenerative dis- 55 eases such as Alzheimer’s disease [1,4,25,26,44,45,48,50]. 56 1 0197-4580/$ – see front matter © 2005 Published by Elsevier Inc. 2 doi:10.1016/j.neurobiolaging.2005.02.013
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Elevation of cytoskeletal protein breakdown in aged Wistar rat brain

F
Neurobiology of Aging xxx (2005) xxx–xxx
Elevation of cytoskeletal protein breakdown in aged Wistar rat brain3
Eric Bernathb, Nancy Kupinab, Ming Cheng Liub,c, Ronald L. Hayesa,b,c,Colleen Meeganb,c, Kevin K.W. Wanga,b,c,d,∗
4
5
a Department of Psychiatry, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida, Gainesville, FL 32610, USA6b Department of Neuroscience, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida, Gainesville, FL 32610, USA7
c Center for Traumatic Brain Injury Studies, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida, Gainesville, FL 32610, USA8d Center for Neuroproteomics and Biomarkers Studies, McKnight Brian Institute, L4-100, P.O. Box 100256, University of Florida,
Gainesville, FL 32610, USA9
10
Received 27 April 2004; received in revised form 4 February 2005; accepted 25 February 2005
11
Abstract12
Previous studies indicated there is an overall increase of proteolysis in aging rat brains. We monitored the potential degradation of cytoskeletalproteins in neuronal tissue taken from cerebral cortex and cerebellum of young (3 month) and aging (17, 21 and 23.5 month) Wistar rats.W thec DP150a ggesting thei o calpaind ather, therei usceptibilityo
13
14
15
16
17
18
19
20
21
©22
K23
24
125
26
(27
p28
[29
p30
A31
o32
f33
h34
c35
d36
i37
t38
on39
- 40
sing41
a 42
ellu-43
lar44
by45
46
pro-47
re of-48
, cer-49
od-50
ro-51
an de-52
opto-53
rote-54
dis-55
56
1 02 d
COR
RE
CTE
D P
RO
Oe found significant age-dependent proteolysis of cytoskeletal proteins (�II-spectrin and microtubule-associated protein MAP-2A/B) inerebral cortex and the cerebellum. The pattern of�II-spectrin breakdown shows a marked increase in 150- and 145-kDa fragments (SBnd SBDP145, respectively), but we did not detect the caspase-3-mediated 120-kDa fragment (SBDP120) in aged rat brains, su
nvolvement of the calpain proteases. The pattern of MAP-2A/B breakdown in aged rat brains mirrors that produced by in vitrigestion of 3-month control rat brain MAP-2A/B. In aged rat brains, there is no significant increase in pro-caspase-3 processing; r
s a moderate reduction in pro-caspase-3 protein and caspase-3 hydrolytic activity in the cortex. These results point to selective sf cytoskeletal proteins to calpain-mediated degradation, but not caspase-3 in aging rat brains.2005 Published by Elsevier Inc.
eywords:Aging; Alzheimer’s disease; Calpain; Caspase; MAP-2A/B;�II-spectrin
. Introduction
Dysfunction in the homeostatic regulation of calciumCa2+) has been one of the many notable theories pro-osed to account for age-related neuronal degradation
7,12,13,16,30,56,59]and (of more immediate therapeuticotential) neurodegenerative diseases such as stroke andlzheimer’s disease[6,49]. Our study was designed to focusn experimental evidence supporting the calcium hypothesis
or aging[29,54]. Concerning this theory, past investigatorsave focused on studying the regulation of Ca2+ in intra-ellular and extracellular areas and its dysfunctional link toegenerative alterations in the physiology of neuronal activ-
ties. Of specific interest are the effects of Ca2+ on synapticransmissions[36,53] and hypoxic-ischemic cellular death
∗ Corresponding author. Tel.: +1 352 294 0031; fax: +1 352 392 2579.E-mail address:[email protected] (K.K.W. Wang).
[9,24]. Many laboratories, including ours, have focusedthe secondary dysfunctional role of Ca2+ on cellular physiology and the potential role of calpain-overactivation in cauneurodegenerative responses[53,60]. It has been shown innumber of studies that a sustained elevation in intraclar Ca2+ triggers calpain activation, which leads to celludamage[5,21,24,51]. Activated calpain exerts its damagedegrading various cellular substrates such as�II-spectrin,calmodulin binding proteins, microtubule-associatedteins (MAPs) and neurofilaments. These processes aten associated with oncotic necrosis (oncosis). Howevertain pathophysiological conditions (such as hypoxia or merate excitotoxicity) can also trigger activation of the papoptotic protease caspase-3. Like calpain, caspase-3 cgrade a large number of neuronal proteins, leading to apsis[57]. In fact, both caspase-3 and calpain-mediated polytic events have been implicated in neurodegenerativeeases such as Alzheimer’s disease[1,4,25,26,44,45,48,50].
197-4580/$ – see front matter © 2005 Published by Elsevier Inc.oi:10.1016/j.neurobiolaging.2005.02.013
N NBA 6255 1–9
U

DO
F
2 E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx
Therefore, it is conceivable that calpain and caspase-3 pro-57
teases might also be activated in the normal aging process58
[10,61]. In fact, there have been some sporadic reports show-59
ing increased activities of calpains[3,5,21,52]and caspases60
[28,62]in aging brain and/or other organs. Hinman et al.[18]61
also recently showed that there is a significant increased of62
activated calpain-1 immunoreactivity in antigen-DR (HLA-63
DR)-positive microglia in the white matter of the aged rhesus64
monkey.65
In the present study, we evaluated the hypothesis that pro-66
teolysis plays a role in cellular damage due to aging by exam-67
ining calpain- and caspase-mediated proteolysis in neuronal68
tissue during the process of normal aging in the Wistar rat.69
2. Materials and methods70
2.1. Tissue preparation71
All procedures were carried out in strict compliance with72
the Institutional Animal Care and Use Committee of Parke-73
Davis and Pfizer Global Research and Development and the74
Society of Neuroscience. Male Wistar rats (Charles River75
Laboratories), age’s 3, 12, 7, 21 and 23.5 months were anes-76
thetized with 4% isoflurane before being sacrificed via car-77
diac puncture. Animals were first perfused with saline, then78
t sent-79
i ther80
t d fe-81
m on,82
e over83
d84
f ain-85
i ains86
w pes-87
t88
289
using90
a me91
o92
E d93
p to the94
f 2 h95
w ared96
w ere97
d the98
s99
100
t ide,101
N V102
f ia a103
B 2 h.104
T Tris105
b and106
then incubated in primary antibody [beta-actin (1:1000,107
Sigma Chemical), MAP-2A/B (1:1000, Sigma Chemical),108
caspase-3 (1:500, Santa Cruz),�II-spectrin (1:1000, Affin- 109
ity Co., UK) and SBDP150i-specific antibody (Cell Sig-110
naling Technology, #2121)] overnight at 4◦C. Secondary 111
biotinylated antibodies [anti-mouse IgG or anti-rabbit IgG112
(1:500, Caliches)] were exchanged for 1 h, followed by a ter-113
tiary streptavidin–alkaline phosphatase conjugate (1:3000,114
Caliches) for 30 min. The blots were developed in a115
linear range with substrate 5-bromo-4-chloro-3-indolyl-116
phosphate and nitroblue tetrazolium (BCIP/NBT, Kirkegaard117
& Perry). Densitometric analysis of the protein bands118
was quantified using NIH Image software on a Macintosh119
computer. 120
2.3. Assay for caspase-3 activity in neuronal tissue 121
Sample lysate was taken immediately following prepa-122
ration and mixed with a buffer containing 100 mM Hepes,123
10% glycerol, 10 mM DT, 0.5 mM EGTA and 40 mM acetyl-124
Asp-Glu-Val-Asp-7-amido-4-methylcoumarin (Ac-DEVD-125
MCA, Peptide International). After 2 h, the assay was mea-126
sured on a Millipore CytoFluor 2300 plate-reader[31] us- 127
ing a fluorescence of excitation 380± 15 nm and emission 128
460± 15 nm. 129
2 130
sec-131
t es132
w laced133
o e 134
w aline135
( the136
c oge-137
n rum138
( en139
i , 140
S vel-141
o ces142
w ride143
( nd144
E h a145
p 146
3 147
3 148
a 149
time150
c l pro-151
t ic-152
t Gel153
l ata154
UN
CO
RR
EC
TE
he brains were extracted and divided into regions repreng the cerebral cortex, midbrain and cerebellum, while oissues were removed (hearts, liver, kidneys, muscle anur) for inclusion in a parallel study. Following dissectiach brain region was either flash frozen in isopentanery ice (−70◦C) or stored in 4% paraformaldehyde at 4◦C
or 12 h before processing for immunohistochemical stng. Immediately preceding protein extraction, frozen brere crushed into powder using a pre-cooled mortar and
le over dry ice.
.2. Protein extraction and sample analysis
Protein samples were extracted from neuronal tissuemodified Triton extraction method. A proportionate voluf 20 mM Tris–HCl buffer (pH 7.4), 5 mM EDTA, 5 mMGTA, 150 mM NaCl, 1 mM DTT, 1% Triton X-100 anrotease inhibitor (Roche #11 836 153 001) was added
rozen (−70◦C) powdered tissue and stored in wet ice forith intermittent agitation. The homogenate was then cleith centrifugation, protein concentrations of the lysate wetermined using a modified Lowry assay (BioRad) andamples were stored in 50% glycerol at−70◦C.
Each sample was normalized to contain 25�g of pro-ein and run on SDS-PAGE [4–20% (w/v) polyacrylamovex] with a Tris/glycine running buffer solution at 200
or 2 h. Gels were transferred to a PVDF membrane vioRad semidry electrotransferring unit at 20 mA forhe blots were then placed in Blotto (Tween–TBS:ase, Tris–HCl, Tween-20 and 10% non-fat dry milk),
PR
O
NBA 6255 1–9
.4. Immunohistochemistry in neuronal tissue
Paraformaldehyde-stored brains were transverselyioned at 100�m, using a chilled tissue slicer. The slicere placed in a sucrose solution (20%) before being pn glass microscope slides and stored at 4◦C. Samples werashed in a series, which included phosphate buffered s
PBS), 80% formic acid, 0.5% Triton buffer (to permeateell membrane), 1% hydrogen peroxide (to quench endous peroxidase activity) and blocked in 2% horse seprovided by Vectastain Elite ABC kit). Slides were thncubated over night at 4◦C with anti-MAP-2A/B (1:500igma Chemical) and then washed in PBS. Following depment using the Vectastain Elite ABC Kit, the brain sliere visualized with a diaminobenzidine tetrahydrochlo
DAB) solution and counter-stained with Hematoxylin aosin (H&E) Staining. All sections were examined withase contrast microscope at 400× resolution.
. Results
.1. Immunoblot analysis ofαII-spectrin in young andged rat brains
Western immunoblots were prepared to evaluate aourse of degradation of two protease-sensitive neuronaeins (�II-spectrin and MAP-2A/B) to gauge an overall pure of biochemical functions during the aging process.oading controls with beta-actin were performed for all d

OR
RE
CTE
D P
RO
OF
E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx 3
Fig. 1. Age-related increase of�II-spectrin breakdown in rat cortex. (A) Two sets of representative immunoblots of�II-spectrin breakdown during timecourse are shown (totaln= 5–8). Spectrin intact bands (280 kDa). SBDP150 and SBDP145 are indicated by arrows. (B) Time course of protein extracts ofcerebral cortex tissue at 3, 17, 21 and 23.5 months were assayed in immunoblots against�II-spectrin antibody (Affiniti) at 1:1000 concentration. Dark barsindicate 150-kDa spectrin breakdown products (SBDP150) while shaded bars indicate intact SBDP145. Data are shown as SBDP/intact spectrin ratio. Dataare means± S.E.M. (n= 5 for control and 23.5 month,n= 6 for 21 month,n= 8 for 17 month). Values showing significance different from 3-month controlanimal are indicated by∗p≤ 0.05 or** p< 0.005, with Student’s unpairedt-test. (C) One set of representative immunoblots of caspase-produced SBDP150i(Cell Signaling Technology) was shown (samples as in top panel of A). The far right lane (+) is SDBP150i positive control (lysate from human SH-SY5Y cellssubjected to staurosporin treatment)[33]. (D) For comparison,�II-spectrin was degraded in rat cortex lysate digested with calpain and caspase-3, producingSBDP150/SBDP145 and SBDP150i/SBDP120, respectively. Analysis of variance (ANOVA) was also performed on SBDP levels with respect to age.
(Figs. 1 and 2). Since the loading was relatively even, we155
did not further normalize�II-spectrin and MAP-2A/B data156
to actin. Control Wistar rats (3 months,n= 5) were compared157
with rats of ages 17 months (n= 8), 21 months (n= 6) and158
23.5 months (n= 5). Immunoblot samples were separated by159
brain section, with testing done on the cerebral cortex and160
cerebellum.161
Analysis with�II-spectrin antibody revealed significant162
breakdown of this protein in the cortex and cerebellum in163
aged rats (Figs. 1 and 2). Fig. 1A shows cortical samples sub-164
jected to�II-spectrin immunoblot analysis. The intact�II-165
spectrin appears in all lanes as a band of 280 kDa. However,166
levels of�II-spectrin breakdown products 150 and 145 kDa167
(SBDP150 and SBDP145) increased significantly during the168
time course of aging. To standardize comparison across dif-169
ferent blots, we expressed SBDP level as a ratio of the intact170
�II-spectrin level in the same samples. In the cortex, the high-171
est increase of SBDP150 was observed at 17 months, but172
SBDP150 levels were also significantly elevated at 20 and173
23.5 months, based upon Student’st-test analysis (Fig. 1A174
and B). Similar to the SBDP150 counterpart, SBDP145 is175
also significantly elevated in the cortex of 17-, 20- and 23-176
month-old rats (Fig. 1). On the other hand, in the cerebellum,177
SBDP150 and SBDP145 were also significantly and progres-178
sively elevated in 17, 20 and 23 months, when compared179
to young rat controls (3 months) (Fig. 2A and B). Further, 180
ANOVA (analysis of variance) showed that SBDP145 sig-181
nificantly increases in both cortex and cerebellum with re-182
spect to animal age, while age-related SBDP150 increasing183
did not reach statistical significance. In addition, evenness184
of gel loading was confirmed with beta-actin as the standard185
(Figs. 1A and 2A). 186
To compare the SBDPs observed in the aged rat brains187
to those generated by these proteases (calpain and caspase-188
3), we subjected naıve 3-month rat cortex lysate to in vitro 189
calpain and caspase-3 digestion, respectively (Fig. 1C). Both 190
SBDP150 and SBDP145 were prominently generated by cal-191
pain proteolysis[43,47,57]. In contrast, in vitro caspase-3 di-192
gestion generated and a fragment just below 150-kDa band193
(termed “SBDP150i”) and a characteristic 120-kDa spectrin194
breakdown product (SBDP120)[33,58]. Both SBDP150 and 195
SBDP145 were prominently generated by calpain proteolysis196
[47,57]. In comparison, while both SBDP150 and SBDP145197
were prominently observed in aged rat cortex and cerebel-198
lum, the SBDP120 was noticeably absent (Figs. 1A and 2A). 199
We further examined, if there was also a lack of casapse-200
3-produced SBDP150i. Indeed, using an anti-SBDP150i-201
specific antibody (cell signaling), we confirmed that caspase-202
3-generated SBDP150i was also absent in cortex and cere-203
bellum from all age groups (Fig. 1C).
UN
C
NBA 6255 1–9

CO
RR
EC
TED
PR
OO
F
4 E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx
Fig. 2. Age-related�II-spectrin breakdown in rat cerebellum. (A) Two sets of representative immunoblots of�II-spectrin breakdown during time course areshown (totaln= 5–8). Spectrin intact bands (280 kDa), SBDP150 and SBDP145 are indicated by arrows. (B) Time course of protein extracts of cerebral cortextissue at 3, 17, 21 and 23.5 months were assayed in immunoblots against�II-spectrin antibody (Affiniti) at 1:1000 concentration. Dark bars indicated 150-kDaspectrin breakdown products (SBDP150) while shaded bars indicate SBDP145. Data are shown as SBDP/intact spectrin ratio. Data are means± S.E.M. (n= 5for control and 23.5 month,n= 6 for 21 month,n= 8 for 17 month). Values showing significance different from 3-month control animal are indicated by∗p≤ 0.02 or** p< 0.05, with Student’s unpairedt-test. In the�II-spectrin intact band (280 kDa), arrows indicate SBDP150 and SBDP145. (C) One set ofrepresentative immunoblots of caspase-produced SBDP150i was shown (samples as in top panel of A). The far right lane (+) is SDBP150i positive controlasin Fig. 1[33]. ANOVA was also performed on SBDP levels with respect to age.
3.2. Immunoblot analysis of MAP-2A/B in young and204
aged rat brains205
Young and aged rat cortical and cerebellar samples were206
also subjected to analysis with Western immunoblotting us-207
ing a microtubule associated protein 2A and 2B (MAP-2A/B)208
antibody. In this case, the intact MAP-2A/B appeared as a209
doublet of about 300 kDa in all lanes (Fig. 3A (cortex) and 4A210
(cerebellum)). A dramatic and decreased intact MAP-2A/B211
density over the time course is seen in the cortex (Fig. 3B)212
and in the cerebellum (Fig. 4B) (based upon Student’st-test213
analysis). The reduction of MAP-2A/B protein levels could214
not be due to uneven gel loading, since equal amounts (20�g)215
were loaded to each lane. Furthermore, results with beta-actin216
showed constant levels between control and aged samples in217
both cortex and cerebellum, indicating sample loadings were218
even (see “actin” panels inFig. 7). In parallel with the reduc-219
tion of intact MAP-2A/B in aged rat brains, multiple break-220
down products of MAP-2A/B were also apparent in cortex221
and cerebellum from aged rats (Figs. 3A and 4A, respec-222
tively). Similar to SBDP analysis, evenness of gel loading223
was confirmed with beta-actin as standard (Figs. 3A and 4A).224
Additional ANOVA post hoc analysis also showed that in-225
tact MAP-2A/B levels significantly decreased with respect226
to animal age (p< 0.05) in both cortex and cerebellum.227
Since MAP-2A/B was reported to be extremely sensitive228
to calpain-mediated proteolysis[15,19,20,51]and no reports 229
have been published on its sensitivity to caspase-3, we sub-230
jected naıve cortex (3 month) lysate to calpain digestion and231
found that the in vitro calpain-proteolyzed MAP-2A/B also232
shows multiple lower molecular weight fragments (Fig. 3C), 233
as was the case observed in aged rat cortex and cerebellum234
(Figs. 3A and 4A). 235
We also performed immunohistochemical analysis with236
MAP-A/B antibody with cortex and cerebellum sam-237
ples from young (3 month) and aged (23.5 month) rats.238
Paraformaldehyde-fixed brains were transversely sectioned239
at 100�m and slices were incubated over night at 4◦C 240
with MAP-2A/B (1:500, Sigma Chemical) and visualized241
with a diaminobenzidine tetrahydrochloride (DAB) (brown)242
(Fig. 5). Slices were also counter-stained with Hematoxylin243
and Eosin Staining (blue/red).Fig. 5 showed representa- 244
tive slices from young cortex have more MAP-2 reactiv-245
ity when compared to aged counterpart (compare panel246
A to B, Fig. 5). Consistent with the Western blot re-247
sults, the decline of MAP-2A/B staining in aged cere-248
bellum appeared more severe (compared panel D to C,249
Fig. 5).
UN
NBA 6255 1–9
CO
RR
EC
TED
PR
OO
F
E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx 5
Fig. 3. Alteration of MAP-2A/B levels in rat cortex during aging. (A) Two representative immunoblots (totaln= 4) of MAP-2A/B breakdown during timecourse are shown. Indicated is the intact MAP-2A/B (280 kDa). Arrows indicate multiple breakdown products (BDPs). (B) Time course of intact MAP-2A/Blevels of cerebellum tissue at 3, 17, 21 and 23.5 months were assayed in immunoblots. Data are ratios of intact MAP-2A/B at time X/MAP-2A/B of 3-monthcontrol animals. Data are means± S.E.M. (n= 4). Values showing significance different from 3-month control animal are indicated by∗p≤ 0.02 or** p< 0.001with Student’s unpairedt-test. (C) For comparison, MAP-2A/B was degraded into multiple BDPs in rat cortex lysate digested with calpain. ANOVA was alsoperformed on MAP-2A/B levels with respect to age.
Fig. 4. Alteration of MAP-2 levels in rat cerebellum during aging. (A) Two representative immunoblots (totaln= 4) of MAP-2 breakdown during time courseare shown. Indicated is the intact MAP-2 (300 kDa). Arrows indicate multiple breakdown products (BDP). (B) Time course of intact MAP-2A/B levels ofcerebellum tissue at 3, 17, 21 and 23.5 months were assayed in immunoblots. Data are ratio of intact MAP-2A/B at time X/MAP-2A/B of 3-month controlanimals. Data are means± S.E.M. (n= 4). Values showing significance different from 3-month control animal are indicated by∗p≤ 0.05 with Student’s unpairedt-test. ANOVA was also performed on MAP-2A/B levels with respect to age.
UN
NBA 6255 1–9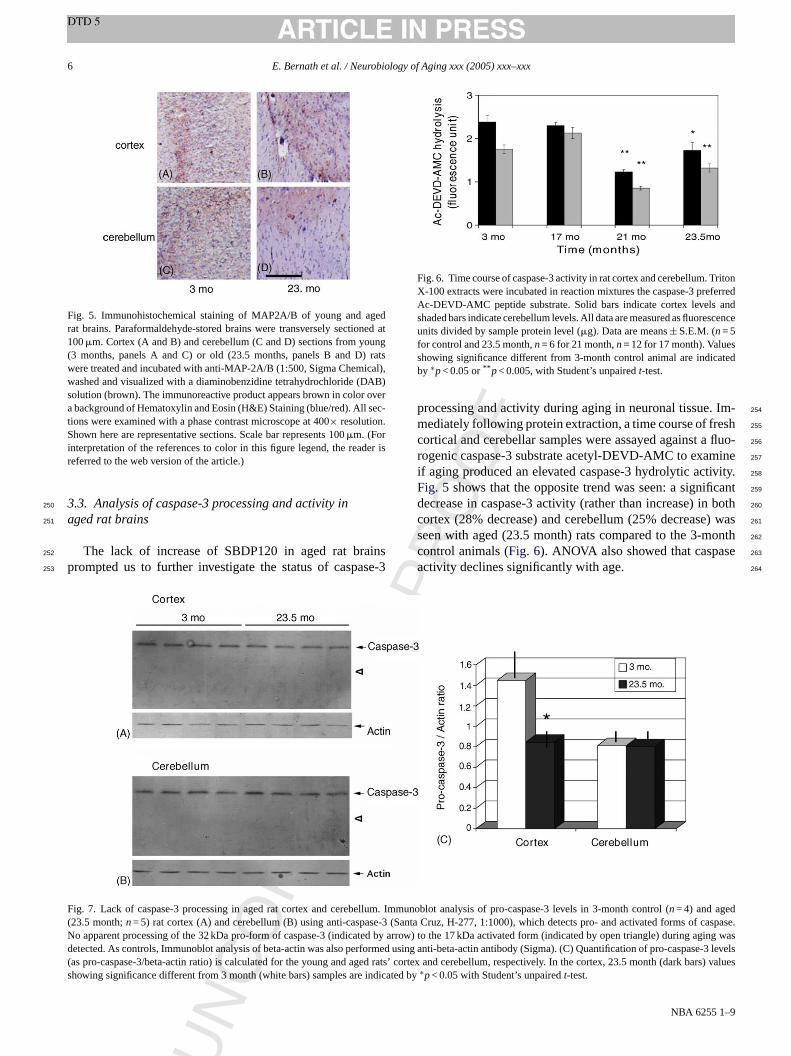
RR
EC
TED
PR
OO
F
6 E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx
Fig. 5. Immunohistochemical staining of MAP2A/B of young and agedrat brains. Paraformaldehyde-stored brains were transversely sectioned at100�m. Cortex (A and B) and cerebellum (C and D) sections from young(3 months, panels A and C) or old (23.5 months, panels B and D) ratswere treated and incubated with anti-MAP-2A/B (1:500, Sigma Chemical),washed and visualized with a diaminobenzidine tetrahydrochloride (DAB)solution (brown). The immunoreactive product appears brown in color overa background of Hematoxylin and Eosin (H&E) Staining (blue/red). All sec-tions were examined with a phase contrast microscope at 400× resolution.Shown here are representative sections. Scale bar represents 100�m. (Forinterpretation of the references to color in this figure legend, the reader isreferred to the web version of the article.)
3.3. Analysis of caspase-3 processing and activity in250
aged rat brains251
The lack of increase of SBDP120 in aged rat brains252
prompted us to further investigate the status of caspase-3253
Fig. 6. Time course of caspase-3 activity in rat cortex and cerebellum. TritonX-100 extracts were incubated in reaction mixtures the caspase-3 preferredAc-DEVD-AMC peptide substrate. Solid bars indicate cortex levels andshaded bars indicate cerebellum levels. All data are measured as fluorescenceunits divided by sample protein level (�g). Data are means± S.E.M. (n= 5for control and 23.5 month,n= 6 for 21 month,n= 12 for 17 month). Valuesshowing significance different from 3-month control animal are indicatedby ∗p< 0.05 or** p< 0.005, with Student’s unpairedt-test.
processing and activity during aging in neuronal tissue. Im-254
mediately following protein extraction, a time course of fresh255
cortical and cerebellar samples were assayed against a fluo-256
rogenic caspase-3 substrate acetyl-DEVD-AMC to examine257
if aging produced an elevated caspase-3 hydrolytic activity.258
Fig. 5 shows that the opposite trend was seen: a significant259
decrease in caspase-3 activity (rather than increase) in both260
cortex (28% decrease) and cerebellum (25% decrease) was261
seen with aged (23.5 month) rats compared to the 3-month262
control animals (Fig. 6). ANOVA also showed that caspase263
activity declines significantly with age. 264
F llum. I d( se-3 (S of caspase.N ed by a g aging wd formed s( ed rats kss indicat
CO
ig. 7. Lack of caspase-3 processing in aged rat cortex and cerebe23.5 month;n= 5) rat cortex (A) and cerebellum (B) using anti-caspao apparent processing of the 32 kDa pro-form of caspase-3 (indicatetected. As controls, Immunoblot analysis of beta-actin was also peras pro-caspase-3/beta-actin ratio) is calculated for the young and aghowing significance different from 3 month (white bars) samples are
UN
mmunoblot analysis of pro-caspase-3 levels in 3-month control (n= 4) and ageanta Cruz, H-277, 1:1000), which detects pro- and activated formsrrow) to the 17 kDa activated form (indicated by open triangle) durinasusing anti-beta-actin antibody (Sigma). (C) Quantification of pro-caspase-3 level’ cortex and cerebellum, respectively. In the cortex, 23.5 month (darbars) valueed by∗p< 0.05 with Student’s unpairedt-test.
NBA 6255 1–9

ED
OF
E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx 7
Parallel immunoblotting analysis of the same samples265
were then performed and probed with an anti-caspase-3 anti-266
body, which detects both pro- and activated forms of caspase-267
3 (Fig. 7A). The 32 kDa pro-caspase-3 protein was observed268
in the cortex and cerebellum of both young and aged rats,269
while the fragment for the 17 kDa activated form was not270
observed, either in young or aged rat brains. Western im-271
munoblots were also performed with anti-beta-actin anti-272
body, which showed relatively constant levels between young273
and aged cortex and cerebellum sample group (Fig. 7A and274
B, respectively). In fact, densitometric analysis showed that,275
with respect to beta-actin levels (as controls), the pro-caspase-276
3 level was actually lower in cortex but remained constant277
in the cerebellum in aged rats with comparison to young rats278
(Fig. 7C). Taken together, these data would suggest that there279
might be an age-related reduction of pro-caspase-3 expres-280
sion and no evidence of increased activation of caspase-3.281
4. Discussion282
The results of this study present evidence of elevated283
proteolysis of�II-spectrin (Figs. 1 and 2) and MAP-2A/B284
(Figs. 3 and 4) in a time course of aging in Wistar rats. Our285
results provide evidence that cytoskeletal protein degrada-286
tion is a common occurrence in aged rat brains. Both of these287
p ele-288
t t of289
t xons290
a291
p hich292
p traf-293
fi294
w eu-295
r egra-296
d se-297
v298
eath299
a have300
s -301
o rone302
t e303
c in the304
p down305
a ellum306
( that307
c gion308
[ rs to309
c urons310
i g311
t any312
s n the313
i in314
n ion315
c ping316
b ins317
had on degeneration. They showed conclusively that MAP-2318
was specifically at risk for being degraded due to the sensi-319
tivity as a calpain substrate. Others highlighted the changes320
in phosphorylation that occurs with the MAP-2 protein[20], 321
which, due to age-related changes, will increase its suscepti-322
bility with time. This happens when the regulation of binding323
changes in the microtubule network, making it more open to324
calpain hydrolysis. It is possible that changes due to spectrin325
breakdown could contribute to this. With the loss of MAP-326
2A/B, dendrites appear to collapse. One of the first studies to327
examine the effects of MAPs in an aged brain[8] saw almost 328
total MAP-2A/B loss in the cortex and hippocampus with329
associated loss in synaptic plasticity. 330
One of the major hypotheses that promoted this study was331
that caspase- and calpain-mediated proteolysis could also be332
identified as a contributor to the protein degeneration seen in333
the aging process. Many studies have identified its increase in334
ischemic events[31,33,35,40]and in head trauma[23,41,46]. 335
The caspase family of proteases has been shown to mediate336
cell suicide[34] in cells either damaged by trauma or having337
undergone irreversible degeneration to the point where pro-338
grammed death is required. Our experiments focused on the339
evaluation of caspase- and calpain-mediated proteolysis in340
aged rat brains. These studies, utilizing anti-caspase-3 West-341
ern immunoblots and an active caspase-3 assay (ac-DEVD-342
AMC) indicated no increase in activated forms of the 32 kDa343
p ase-3344
( s 345
s is to346
S ibed347
c oduct348
( 349
n about350
1 i- 351
b 150352
w bility,353
w here354
w g or355
a 356
w t sig-357
n 358
s nted.359
B ain-360
m , but361
n and362
c DP363
w am-364
p (13365
m lpain-366
m ing in367
b e 368
o use369
v 370
ele-371
t ed372
r age-373
UN
CO
RR
EC
T
roteins play important roles in the stability of the cytoskal system.�II-spectrin is the major structural componenhe cortical membrane cytoskeleton and is abundant in and presynaptic terminals[16,42]. MAP-2A/B is a dendriticrotein that interacts with and stabilizes microtubules, wrovide both structural elements and tracks for organellec [11,27]. The degradation of�II-spectrin and MAP-2A/Bould seriously compromise the normal functions of n
onal tissue. Our results also indicate that age-related dation of both�II-spectrin and MAP-2A/B appears lessere in the cortex than in the cerebellum.
Prior studies have looked at various aspects of cell dnd calcium-related changes in the brain. Experimentshown region-specific changes in calcium[17] and that homestasis in the hippocampus and cortex are especially p
o dysfunction in aged rats, while the Ca2+ changes in therebellum and striatum are less severe. Interestingly,resent study, aging-induced cytoskeleton protein breakppears to affect both the cerebral cortex and the cerebFigs. 1–4). Studies of the human brain have implicatedalpain levels are among the highest in the cerebellar re3]. Also, evaluation of the rat brain has led researcheonclude that aging produces a more drastic loss of nen the cerebellum than in the cortex[37]. When examininhe effects of calpain-mediated spectrin breakdown, mtudies have highlighted the role that proteases play ontegrity of the neuronal structure[24] and the increaseeuronal death[46]. Early studies focused on the relatalpain-mediated proteolysis had on the young develorain [14] and highlighted the susceptibility these prote
PR
O
NBA 6255 1–9
ro-caspase fragment, or any increase in active caspFigs. 5 and 6). Furthermore, anti-�II-spectrin immunoblothowed that there is significant age-related proteolysBDP150 and SBDP145, while the previously descraspase-3 generated 120-kDa spectrin breakdown prSBDP120)[57] was not observed (Figs. 1 and 2). It is worthoting that caspase-3 can also produce a fragment of50 kDa (termed SBDP150i, see[33,58]). It is, thus, possle that SBDP150i can in part contribute to the SBDPe observed in aged rat brain tissues. To test this possie used a SBDP150i-specific antibody and found that tere no detectable signals of SBDP150i in either younged cortical or cerebellar samples (Figs. 1C and 2C). Thus,e conclude that caspase-mediated proteolysis is noificantly elevated in aged rat brains. Calpain-mediated�II-pectrin proteolysis in aging has in fact been documeahr et al.[2] showed a rather selective increase of a calpediated SBDP in both the cortex and telencephalonone in the cerebellum of aged mouse brains. Vicenteolleagues[54] also showed that a calpain-mediated SBas significantly elevated in the neocortex and hippocus but not in thalamus or cerebellum in mid-aged miceonths). These data contrast our observations that caediated SBDP150 and SBDP145 are elevated with agoth cortex and cerebellum (Figs. 1 and 2, respectively). Onf the contributing factors could be species variation (moersus rat).
Our analysis of the extensive proteolysis of the cytoskal protein MAP-2A/B in the cortex and cerebellum of agats supports the importance of calpain activation in

DO
F
8 E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx
related degeneration, since MAP-2A/B is known to be more374
susceptible to calpain- and not caspase-3 activation. MAP-375
2A/B (Figs. 3 and 4). It is worth noting that MAP-2A/B break-376
down products in the cortex are in fact more intense than their377
counterparts in the cerebellum (Fig. 3A versusFig. 4A). Yet,378
the levels of remaining intact MAP-2A/B in aged cerebel-379
lum (23.5 month) were decreased to about 55% of young380
cortex controls, while the MAP-2A/B levels in aged cerebel-381
lum (23.5 months) were further decreased to 25% of young382
cerebellum controls (Fig. 3B versusFig. 4B). We hypothe-383
sized that proteolysis in the cerebellum was even more ac-384
tive, thus, not only reducing the level of intact MAP-1A/B,385
but more efficient in further degrading the fragments into386
smaller peptides, as is the case with digestion with calpain387
in vitro (Fig. 3C). In addition, immunohistochemical analy-388
sis, while not quantitative, showed a more dramatic reduction389
of MAP-2A/B levels in the cerebellum than in cerebral cor-390
tex in the aged (23.5 month) rats, when compared to their391
younger counterpart (3 month) (Fig. 5). Hinman et al.[18]392
recently showed that there is also a significant increase in ac-393
tivated calpain-1 immunoreactivity in microglia and possibly394
myelin-bearing oligodendrocytes in the white matter of the395
aged rhesus monkey. It is, therefore, possible that there is a396
concerted increase of calpain activity in multiple cell-types397
within the aged brain.398
Chronic neurological diseases, such as Parkinson’s and399
A d by400
a in ac-401
t n402
o n403
t ents404
[ lpain405
a rther406
d rod-407
u the408
c d tha409
m ues410
a tion.411
W con-412
c here413
c brain414
a ative415
d416
U417
418
A419
stions420
a nnis421
a ven422
D nts;423
Dr. Stephen Larner (University of Florida) for his statistical424
expertise and Dr. Brian Pike (NIH) concerning aging research425
was greatly appreciated. 426
References 427
[1] AyalaGrosso C, Ng G, Roy S, Robertson GS. Caspase-cleaved428
amyloid precursor protein in Alzheimer’s disease. Brain Pathol429
2002;(12):430–41. 430
[2] Bahr BA, Vanderklish PW, Ha LT, Tin MT, Lynch G. Spectrin break-431
down products increase with age in telencephalon of mouse brain.432
Neurosci Lett 1991;131(2):237–40. 433
[3] Banay-Schwartz M, DeGuzman T, Palkovits M, Lajtha A. Calpain434
activity in adult and aged human brain regions. Neurochem Res435
1994;(19):563–7. 436
[4] Bancher C, Lassmann H, Breitschopf H, Jellinger KA. Mech-437
anisms of cell death in Alzheimer’s disease. J Neural Transm438
1997;(50):141–52. 439
[5] Benuck M, Banay-Schwartz M, DeGuzman TT, Lajtha A. Changes440
in brain protease activity in aging. J Neurochem 1996;(67):2019–29.441
[6] Brinton DR, Yamazaki RS. Advances and challenges in the442
prevention and treatment of Alzheimer’s disease. Pharm Res443
1998;(15):386–98. 444
[7] Carlson JC, Riley JC. A consideration of some notable aging theo-445
ries. Exp Gerontol 1998;(33):127–34. 446
[8] Chauhan N, Siegal G. Age-dependent organotypic expression of447
microtubule-associated proteins (MAP1, MAP2, and MAP5) in rat448
brain. Neurochem Res 1997;(22):713–9. 449
[9] Choi DW. Calcium-mediated neurotoxicity: relationship to spe-450
rosci451
452
[ dur-453
ndent454
. 455
[ lles456
iol457
458
[ ms459
460
[ urol461
462
[ pro-463
2 in464
465
[ ad-466
(De-467
468
[ JE,469
ull470
471
[ ific472
rain.473
474
[ Ac-475
of476
477
[ ule-478
rative479
480
[ tein 2481
rosci482
483
[ in II484
nth-485
486
UN
CO
RR
EC
TE
lzheimer’s disease have both been found to be affecteltered calcium homeostasis resulting in increased calpa
ivity [48], �II-spectrin breakdown[38,43] and degradatiof MAP-2A/B in the cytoskeleton[20]. It has been show
hat neurofibrillary tangles contain broken MAP-2 segm22]. Although these results associate the affects of cas having key roles in the aging process, studies of fuetail are necessary to pinpoint all related breakdown pcts, their roles in the changing physiology of the cell, andascade effects on the system as a whole. It is understooany other effects of aging, from DNA damage to plaqnd toxins, play fundamental parts in neural degeneraith increased examination of these factors, working in
ert with calpain-mediated proteolysis, it is hoped that tan be further understanding of the aging of the humannd its relationship to the pathobiology of neurodegeneriseases[7].
ncited references
[32,39,55].
cknowledgments
The authors sincerely appreciate the support, suggend technical assistance of Rathna Nath, Dr. Kim McGind Michelle Field. Also the help of Drs. Feng Bian, Steuddy and Larry Walker in the execution of experime
PR
O
NBA 6255 1–9
t
cific channel types and role in ischemic damage. Trends Neu1988;(11):465–9.
10] Chondrogianni N, Fragoulis EG, Gonos ES. Protein degradationing aging: the lysosome-, the calpain- and the proteasome-depecellular proteolytic systems. Biogerontology 2002;3(1–2):121–3
11] Cole NB, Lippincott-Schwartz J. Organization of organeand membrane traffic by microtubules. Curr Opin Cell B1995;7(February (1)):55–64.
12] Cuervo AM, Dice FJ. How do intracellular proteolytic systechange with age? Front Biosci 1997;(3):25–43.
13] Drachman DA. Aging and the brain: a new frontier. Ann Ne1997;(42):819–28.
14] Fischer I, Romano-Clarke G, Grynspan F. Calpain-mediatedteolysis of microtubule associated proteins MAP1B and MAPdeveloping brain. Neurochem Res 1991;(16):891–8.
15] Friedrich P, Aszodi A. MAP2: a sensitive cross-linker andjustable spacer in dendritic architecture. FEBS Lett 1991;295cember (1–3)):5–9.
16] Goodman SR, Zimmer WE, Clark MB, Zagon IS, BarkerBloom ML. Brain spectrin: of mice and men. Brain Res B1995;36(6):593–606.
17] Hartmann H, Velbinger K, Eckert A, Muller W. Region-specdownregulation of free intracellular calcium in the aged rat bNeurobiol Aging 1995;(17):557–63.
18] Hinman JD, Duce JA, Siman RA, Hollander W, Abraham CR.tivation of calpain-1 in myelin and microglia in the white matterthe aged rhesus monkey. J Neurochem 2004;289(2):430–41.
19] Johnson GV, Litersky JM, Jope RS. Degradation of microtubassociated protein 2 and brain spectrin by calpain: a compastudy. J Neurochem 1991;56(May (5)):1630–8.
20] Johnson GVW, Jope RS. The role of microtubule-associated pro(MAP-2) in neuronal growth, plasticity, and degeneration. J NeuRes 1992;(333):505–12.
21] Kenessey A, Banay-Schwartz M, DeGuzman T, Lajtha A. Calpaactivity and calpastatin content in brain regions of 3- and 24-moold rats. Neurochem Res 1990;(15):243–9.

ED
OF
E. Bernath et al. / Neurobiology of Aging xxx (2005) xxx–xxx 9
[22] Kosik KS, Orecchio LD, Bakalis S, Duffy L, Neve RL. Partial se-487
quence of MAP-2 in the region of a shared epitope with Alzheimer488
neurofibrillary tangles. J Neurochem 1988;(51):587–98.489
[23] Kupina NC, Detloff MR, Bobrowski WF, Snyder BJ, Hall Ed. Cy-490
toskeletal protein degradation and neurodegradation evolve differ-491
ently in males and females following experimental head injury. Exp492
Neurobiol 2003;180(March (1)):55–73.493
[24] Lee K, Frank S, Vanderklish P, Arai A, Lynch G. Inhibition of494
proteolysis protects hippocampal neurons from ischemia. Proc Natl495
Acad Sci USA 1991;(88):7233–7.496
[25] Lee MS, Kwon YT, Li M, Peng J, Friedlander RM, Tsai LH.497
Neurotoxicity induces cleavage of p35 to p25 by calpain. Nature498
2000;(405):360–4.499
[26] Lee MS, Tsai LH. Cdk5: one of the links between senile plaques500
and neurofibrillary tangles? J Alzheimers Dis 2003;(5):127–37.501
[27] Lippincott-Schwartz J, Cole NB. Roles for microtubules and ki-502
nesin in membrane traffic between the endoplasmic reticulum and503
the Golgi complex. Biochem Soc Trans 1995;(23):544–8.504
[28] Lynch AM, Lynch MA. The age-related increase in IL-1 type I505
receptor in rat hippocampus is coupled with an increase in caspase-506
3 activation. Eur J Neurosci 2002;(15):1779–88.507
[29] Lynch MS. Analysis of the mechanisms underlying the age-related508
impairment in long-term potentiation in the rat. Rev Neurosci509
1998;(July–December (9)):169–201.510
[30] Morrison JH, Hof PR. Life and death of neurons in the aging brain.511
Science 1997;(278):412–24.512
[31] Namura S, Zhu J, Fink K, Endres M, Srinivasan A, Tomaselli KJ,513
et al. Activation and cleavage of caspase-3 in apoptosis induced by514
experimental cerebral ischemia. J Neurosci 1998;(18):3659–68.515
[32] Nath R, Probert A, McGinnis KM, Wang KKW. Evidence516
for activation of caspase-3-like protease in excitotoxin-517
hem518
519
[ ng520
onal521
522
[ IBS523
524
[ is of525
526
[ la-527
aged528
stasis529
530
[ rain531
532
[ . In-533
imer534
535
[ , et536
loss537
nce538
539
[ ang540
ing541
chem542
543
[ yes544
, cal-545
enta-546
i Res547
[42] Riederer BM, Zagon IS, Goodman SR. Brain spectrin (240/235)548
and brain spectrin (240/235E): two distinct subtypes with different549
locations within mammalian neural cells. J Cell Biol 1986;102(June550
(6)):2088–97. 551
[43] Roberts-Lewis JM, Savage MJ, Marcy VR, Pinsker LR, Siman552
R. Immunolocalization of calpain I-mediated spectrin degradation553
to vulnerable neurons in the ischemic gerbil brain. J Neurosci554
1994;14(June (6)):3934–44. 555
[44] Rohn TT, Head E, Nesse WH, Cotman CW, Cribbs DH. Activa-556
tion of caspase-8 in the Alzheimer’s disease brain. Neurobiol Dis557
2001;(8):1006–16. 558
[45] Rohn TT, Rissman RA, Davis MC, Kim YE, Cotman CW, Head E.559
Caspase-9 activation and caspase cleavage of tau in the Alzheimer’s560
disease brain. Neurobiol Dis 2002;(11):341–54. 561
[46] Saatman KE, Bozyczko-Coyne D, Marcy V, Siman R, McIntosh TK.562
Prolonged Calpain-mediated spectrin breakdown occurs regionally563
following experimental brain injury in the rat. J Neuropathol Exp564
Neurol 1996;(55):852–62. 565
[47] Saido T, Yokota M, Nagao S, Yamaura I, Tani E, Tsuchiya T, et al.566
Spacial resolution of fodrin proteolysis in postischemic brain. J Bio567
Chem 1993;(268):25239–43. 568
[48] Saito K, Elce JS, Hamos JE, Nixon RA. Widespread activation569
of calcium-activated neutral proteinase (calpain) in the brain of570
Alzheimer’s disease: a potential molecular basis for neuronal de-571
generation. Proc Natl Acad Sci USA 1993;(90):2628–32. 572
[49] Sandson FO. The neurobiology and pharmacotherapy of Alzheimer’s573
disease. J Neuropsychiatry Clin Neurosci 1999;(11):19–31. 574
[50] Selznick LA, Holtzman DM, Han BH, Gokden M, Srinivasan575
AN, Johnson Jr EM, et al. In situ immunodetection of neuronal576
caspase-3 activation in Alzheimer disease. J Neuropathol Exp Neu-577
rol 1999;(58):1020–6. 578
[ lpain579
ron580
581
[ CR.582
ndro-583
the584
585
[ lls.586
587
[ INS588
589
[ ctrin590
Mol591
592
[ ing593
594
[ rends595
596
[ nia597
in by598
ugust599
600
[ m-601
602
[ cal-603
604
[ Mol605
606
[ e ac-607
ntol608
609
UN
CO
RR
EC
T
and hypoxia/hypoglycemia-injured neurons. J Neuroc1988;(71):186–95.
33] Nath R, Raser KJ, McGinnis K, Nadimpalli R, Stafford D, WaKKW. Effects of ICE-like protease and calpain inhibitors on neurapoptosis. Neuroreport 1996;(8):249–55.
34] Nicholson DW, Thornberry NA. Caspases: killer proteases. T1997;(22):299–306.
35] Nixon RA. A “protease activation cascade” in the pathogenesAlzheimer’s disease. Ann N Y Acad Sci 2000;(924):117–31.
36] Ouanounou A, Zhang L, Charlton M, Carlin P. Differential modution of synaptic transmission by calcium chelators in young andhippocampal CA1 neurons: evidence for altered calcium homeoin aging. J Neurosci 1999;19(February (3)):906–15.
37] Peng MT, Lee LR. Regional differences of neuron loss of rat bin old age. Gerontology 1979;(25):205–11.
38] Peterson C, Vanderklish P, Schubert P, Cotman C, Lynch Gcreased spectrin proteolysis in fibroblasts from aged and Alzhedonors. Neurosci Lett 1991;(121):239–43.
39] Posmantur R, Kampfl A, Siman R, Liu SJ, Zhao X, Clifton GLal. A calpain inhibitor attenuates cortical cytoskeletal proteinafter experimental traumatic brain injury in the rat. Neuroscie1997;(77):875–88.
40] Posmantur R, McGinnis K, Nadimpalli R, Gilbertsen RB, WKKW. Characterization of CPP32-like protease activity followapoptotic challenge in SH-SY5Y neuroblastoma cells. J Neuro1997;(68):2328–37.
41] Pike BR, Zhao X, Newcomb JK, Wang KK, Posmantur RM, HaRL. Temporal relationships between de novo protein synthesispain and caspase 3-like protease activation,and DNA fragmtion during apoptosis in septo-hippocampal cultures. J Neurosc1998;52(June (5)):505–20.
PR
O
NBA 6255 1–9
51] Siman R, Noszek JC. Excitatory amino acids activate caI and induce structural protein breakdown in vivo. Neu1988;(1):279–87.
52] Sloane JA, Hinman JD, Lubonia M, Hollander W, AbrahamAge-dependent myelin degeneration and proteolysis of oligodecyte proteins is associated with the activation of calpain-1 inrhesus monkey. J Neurochem 2003;84(January (1)):157–68.
53] Verkhratsky A, Kettenmann H. Calcium signaling in glial ceTrends Neurosci 1996;19(August (8)):346–52.
54] Verkhratsky A, Toescu EC. Calcium and neuronal ageing. T1998;(21):2–7.
55] Vicente JS, Munirathinam S, Bahr BA. Calpain-mediated spebreakdown identifies vulnerable brain regions at middle age. JBiol Biotech 2002;3:1–22.
56] Walker LC, Cork LC. Alzheimer’s disease: the neurobiology of agin nonhuman primates 1999; 233–243 [chapter 15].
57] Wang KKW. Calpain and caspase: can you tell the difference? TNeurosci 2000;(23):20–6.
58] Wang KKW, Posmantur R, Nath R, McGinnis K, Whitton M, TalaRV, et al. Simultaneous degradation of alphaII- and betaII-spectrcaspase 3 (CPP32) in apoptotic cells. J Biol Chem 1998;273(A(35)):22490–7.
59] Wang KKW. Principles of medical biology intracellular calciubinding proteins 1996;4:255–74.
60] Wang KKW, Yuen P. Development and therapeutic potential ofpain inhibitors. Adv Pharmacol 1997;(37):117–51.
61] Ward WF. Protein degradation in the aging organism. ProgSubcell Biol 2002;(29):35–42.
62] Zhang Y, Chong E, Herman B. Age-associated increases in thtivity of multiple caspases in Fisher 344 rat organs. Exp Gero2000;(37):777–89.




















