
Floral color changes in Boswellia sacra Flueck. (Burseraceae): A dialogue between plant and...
6
Flora 206 (2011) 821–826 Contents lists available at ScienceDirect Flora j ourna l h omepage: www.elsevier.de/flora Floral color changes in Boswellia sacra Flueck. (Burseraceae): A dialogue between plant and pollinator Marta Mariotti Lippi ∗ , Claudia Giuliani, Tiziana Gonnelli, Laura Maleci Bini Department of Evolutionary Biology – Plant Biology, University of Florence, via La Pira 4, 50121 Florence, Italy a r t i c l e i n f o Article history: Received 1 November 2010 Accepted 12 February 2011 Keywords: Anthocyanins Frankincence Nectary Pollination ecology Reproductive biology Secretion a b s t r a c t The present study provides new information about the reproductive biology of Boswellia sacra (Burser- aceae), focusing on the nectary and its attractiveness for pollinators. The nectary disc changes its color from yellow to orange and red during the flower development. The colors are related to the main period of the stigmatic receptivity, to the dehiscence of anthers with pollen presentation and the nectar secre- tion. Pollinators preferentially visit the flowers in the “yellow” phase and neglect the “red phase”. This suggests a sophisticated dialogue between the plant and its pollinators. The color change from yellow to red occurs in a very short time (less than 24 h) and it is due to the accumulation of anthocyanins. Despite this dialogue between plant and pollinators, the number of fruits is often scanty. © 2011 Elsevier GmbH. All rights reserved. Introduction Boswellia sacra Flueck. (Frankincense tree, Burseraceae) is a well-known plant since it supplies an oleo-gum-resin used for medicinal purposes as well as for its fragrance. In fact, Frankincense is one of the oldest and most precious resins traded throughout the world. Despite the notoriety of the plant, the past great economic importance of its resin, and the amazing history of its commerce and use, until now the biology of B. sacra has been quite neglected, probably because the cultivation of this plant is difficult out of its natural distribution area; on the other hand, research on site is difficult in absence of equipped laboratories. Only a recent paper devoted to the study of the biology of Boswellia (Sunnichan et al., 2005) points to phenology, pollination and breeding system of B. serrata, an Indian species which provides an important gum-resin, the salai guggul. Currently, B. sacra is distributed in South Arabia and in North-East Africa. In Dhofar (Sultanate of Oman), where this research began and was partially carried out, it extends on rocky and gravelly substrates along the coasts, which are dom- inated by monsoon regime, and in the inner semi-desert areas (Raffaelli et al., 2003). Today, B. sacra populations are subjected to a marked decline, due to the excessive exploitation by man and the overgrazing by dromedaries and goats. For this rea- son a Natural Park was instituted at Wadi Dowkha which is one of the most important B. sacra sites in the country. It is ∗ Corresponding author. E-mail address: mariotti@unifi.it (M. Mariotti Lippi). included in the “Frankincense Trail”, an UNESCO World Heritage Site. Winter is the flowering period of B. sacra, although it may extend till April; fruits usually begin to ripe in March. The flowers are grouped in racemes at the end of the branches; they are radial sym- metrical, with a five-toothed green calyx, five free white petals, ten stamens with yellow anthers and one pistil; at the basis of the ovary, a showy nectariferous ring is present which turns from yellow to red and black, as already observed by Thulin and Warfa (1987) and Miller and Morris (1988). In this paper the phenomenon of the color change and its ecological role are investigated, in order to better understand the mechanisms regulating the pollination system. In partic- ular, a micromorphological and histochemical study, focused on the elements which most directly interfere in plant-insects relations and pollination, i.e. nectary, anthers, and pistil, was carried out. Indeed, the color change in flowers is hypothe- sized to direct the pollinators towards the rewarding flowers, assuring the plant to receive an efficient pollination service (Weiss and Lamont, 1997). Materials and methods Flowers of B. sacra were collected from two populations at dif- ferent sites in Dhofar (Sultanate of Oman): along the Dhofar coasts, on the relieves near Al-Mughsail, and in the Wadi Dowkha Natural Park, near Thumrayt, in February and/or March starting from 2006 to 2010 (Fig. 1). 0367-2530/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.flora.2011.04.008
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Floral color changes in Boswellia sacra Flueck. (Burseraceae): A dialogue between plant and...

Fp
MD
a
ARA
KAFNPRS
I
wmiwiapndd2st
Nrri(taso
0d
Flora 206 (2011) 821– 826
Contents lists available at ScienceDirect
Flora
j ourna l h omepage: www.elsev ier .de / f lora
loral color changes in Boswellia sacra Flueck. (Burseraceae): A dialogue betweenlant and pollinator
arta Mariotti Lippi ∗, Claudia Giuliani, Tiziana Gonnelli, Laura Maleci Biniepartment of Evolutionary Biology – Plant Biology, University of Florence, via La Pira 4, 50121 Florence, Italy
r t i c l e i n f o
rticle history:eceived 1 November 2010ccepted 12 February 2011
a b s t r a c t
The present study provides new information about the reproductive biology of Boswellia sacra (Burser-aceae), focusing on the nectary and its attractiveness for pollinators. The nectary disc changes its colorfrom yellow to orange and red during the flower development. The colors are related to the main periodof the stigmatic receptivity, to the dehiscence of anthers with pollen presentation and the nectar secre-
eywords:nthocyaninsrankincenceectaryollination ecologyeproductive biology
tion. Pollinators preferentially visit the flowers in the “yellow” phase and neglect the “red phase”. Thissuggests a sophisticated dialogue between the plant and its pollinators. The color change from yellow tored occurs in a very short time (less than 24 h) and it is due to the accumulation of anthocyanins. Despitethis dialogue between plant and pollinators, the number of fruits is often scanty.
© 2011 Elsevier GmbH. All rights reserved.
ecretionntroduction
Boswellia sacra Flueck. (Frankincense tree, Burseraceae) is aell-known plant since it supplies an oleo-gum-resin used foredicinal purposes as well as for its fragrance. In fact, Frankincense
s one of the oldest and most precious resins traded throughout theorld. Despite the notoriety of the plant, the past great economic
mportance of its resin, and the amazing history of its commercend use, until now the biology of B. sacra has been quite neglected,robably because the cultivation of this plant is difficult out of itsatural distribution area; on the other hand, research on site isifficult in absence of equipped laboratories. Only a recent paperevoted to the study of the biology of Boswellia (Sunnichan et al.,005) points to phenology, pollination and breeding system of B.errata, an Indian species which provides an important gum-resin,he salai guggul.
Currently, B. sacra is distributed in South Arabia and inorth-East Africa. In Dhofar (Sultanate of Oman), where this
esearch began and was partially carried out, it extends onocky and gravelly substrates along the coasts, which are dom-nated by monsoon regime, and in the inner semi-desert areasRaffaelli et al., 2003). Today, B. sacra populations are subjectedo a marked decline, due to the excessive exploitation by man
nd the overgrazing by dromedaries and goats. For this rea-on a Natural Park was instituted at Wadi Dowkha which isne of the most important B. sacra sites in the country. It is∗ Corresponding author.E-mail address: [email protected] (M. Mariotti Lippi).
367-2530/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.flora.2011.04.008
included in the “Frankincense Trail”, an UNESCO World HeritageSite.
Winter is the flowering period of B. sacra, although it may extendtill April; fruits usually begin to ripe in March. The flowers aregrouped in racemes at the end of the branches; they are radial sym-metrical, with a five-toothed green calyx, five free white petals, tenstamens with yellow anthers and one pistil; at the basis of the ovary,a showy nectariferous ring is present which turns from yellow tored and black, as already observed by Thulin and Warfa (1987) andMiller and Morris (1988).
In this paper the phenomenon of the color change and itsecological role are investigated, in order to better understandthe mechanisms regulating the pollination system. In partic-ular, a micromorphological and histochemical study, focusedon the elements which most directly interfere in plant-insectsrelations and pollination, i.e. nectary, anthers, and pistil, wascarried out. Indeed, the color change in flowers is hypothe-sized to direct the pollinators towards the rewarding flowers,assuring the plant to receive an efficient pollination service(Weiss and Lamont, 1997).
Materials and methods
Flowers of B. sacra were collected from two populations at dif-
ferent sites in Dhofar (Sultanate of Oman): along the Dhofar coasts,on the relieves near Al-Mughsail, and in the Wadi Dowkha NaturalPark, near Thumrayt, in February and/or March starting from 2006to 2010 (Fig. 1).
822 M.Mariotti Lippi et al. / Flora
Fig. 1. Map of Dhofar (Southern Oman) with indication of the natural populationsof B. sacra (stars), modified from Raffaelli et al. (2003). The white stars indicate thec
F
rosao
rDF
L
fi(e((1
Ae
pdescn
wN
S
hTe
acropetal manner and flowers with nectariferous rings of different
ollection sites.
ield observations
Observations on nectar production and pollination ecology wereegularly carried out on plants growing in the Botanical Gardenf Al-Bahlid (Salalah), early after sunrise (7.30–8 h local time) foreven days. Additional observations were also performed at middaynd sunset (18–18.30 h) in plants growing in the Botanical Gardenf Al-Bahlid and in the National Natural Park of Wadi Dowkah.
Microscopical observations both on fresh and fixed mate-ial were carried out on samples of Al-Mughsail and Wadiowkha brought by air transport in our laboratories oflorence.
ight microscopy
In order to determine the various components of the secretion ofresh isolated pistils and nectariferous discs, different histochem-cal techniques were used: Toluidine Blue (TBO) as general stainO’Brien and McCully, 1981); Mercuric Bromophenol Blue (Maziat al., 1953) for proteins; Acriflavine stain for total polysaccharidesO’Brien and McCully, 1981); Alcian Blue for acidic polysaccharidesBeccari and Mazzi, 1966), Fluorol Yellow-088 (Brundrett et al.,991) for lipids.
To observe pollen germination and pollen tube growth, theniline Blue fluorescence method (Smith and McCully, 1978) wasmployed.
Pistils and nectariferous discs were fixed in 2.5% glutaraldehyderepared in 0.1 M phosphate buffer pH 6.8. The fixed material wasehydrated through an ascending ethanol series. Infiltration andmbedding were carried out in historesin and paraffin. Semi-thinections were stained with PAS reagent for total insoluble polysac-harides (O’Brien and McCully, 1981) and with Blue Nile Sulfate foreutral and acidic lipids (Beccari and Mazzi, 1966).
For testing the presence of anthocyanins, nectariferous ringsere cut off from the flowers and merged in alkaline solution (10%aOH in water) and acid solution (3% HCl in water).
canning electron microscopy (SEM)
Single flowers, or parts of them, were fixed in 2.5% glutaralde-
yde in phosphate buffer (0.1 M, pH 6.8) in the field after gathering.he samples were rinsed in the same buffer, dehydrated in a gradedthanol series, critical point dried and gold coated.206 (2011) 821– 826
Results
Flower organs
Nectariferous ringThe nectary consists in a prominent annular ten-lobed disc
located between the stamens and the ovary, the lobes alternatewith the anther filaments. It is about 3 mm in diameter and repre-sents the most showy part of the flower; color changes from yellowto brilliant red (Fig. 2A).
Very early, before anthesis, the epidermis of the nectariferousring is already well-developed. Near the basis, on the lateral sidesfacing the petals or the ovary, the surface is smooth with juxtaposedquadrangular cells (Fig. 2B), while the distal region of the disc,facing outwards, presents stomata and trichomes that give to thenectary a subtle velvet appearance (Fig. 2C). Trichomes are unicellu-lar or multicellular uniseriate (2–3 cells – Fig. 2D), highly vacuolatedand intercalated with flattened epidermal cells. The external wallsof the trichomes, as well as the epidermal cells are covered bya continuous cuticle layer (Fig. 2E and F). On the apical part ofthe trichomes, nectar drops can be observed, probably secretedtrough micropores (Fig. 2D). The drops display positive response toPAS reagent, Alcian Blue (Fig. 2G), and Mercuric Bromophenol Blue(Fig. 2H), suggesting the presence of polysaccharides and proteins.
The stomatal apparatus is prominent, with outwardly projectingguard cells (Fig. 2F and I); the stomata appeared in the microscopicaspects always open, with thickened material in the pore.
PistilThe pistil (Fig. 2A) is formed by a slightly lobed superior ovary,
with 3–4(–5) locules and a columnar style, with three to four lon-gitudinal furrows, terminating in a capitate stigma (Fig. 3A and B).The receptive surface of the stigma is wet and constituted by unicel-lular papillae that produce a copious amount of exudate, coveringcompletely the stigma and flowing down also on the style (Fig. 3B).The exudate is constituted by acidic polysaccharides and neutrallipids as evidenced by positive response to Alcian Blue and BlueNile Sulfate stains (Fig. 3A).
Transverse sections of the pistil show more than one (3–4) sty-lar canals. The stylar canals are lined with a clearly defined layer ofcells, which show PAS-positive thickenings on the inner tangentialwall and mildly stain with Alcian Blue, indicating their predom-inantly pectic nature. The stylar canals are filled with secretionproducts, staining for acidic polysaccharides, lipids and proteins(Fig. 3C). After fertilization, the ovary, which is 3–4(–5)-locular,begins to become swollen turning in an obovoid fruit, with oneseed in each compartment. At maturity, the fruit looks like a cap-sule. It splits by 3–4 longitudinal sutures releasing each seed witha part of pericarp and leaving the central septum attached at thetip of the peduncle (Fig. 3D).
AntherAnthers are 10, all of the same size, dorsifixed and versatile, with
longitudinal dehiscence, yellow in color and with few trichomeson their surface (Fig. 3E). Pollen grains are subprolate, tricolporatewith tectate perforate exine (Fig. 3F and G).
Change in color
The inflorescence development does not occur rigorously in an
color are irregularly mixed on the same raceme (Fig. 2A). Concern-ing the color displayed by a nectariferous ring, different phaseswere observed during the flower development (Fig. 4A–D).

M.Mariotti Lippi et al. / Flora 206 (2011) 821– 826 823
Fig. 2. (A) Inflorescence of B. sacra with irregularly mixed flowers at different stages. Note the nectariferous discs of various colors: yellow, orange, and red. (B–H) Nectariferousdisc of B. sacra. (B) SEM micrograph showing smooth, quadrangular epidermal cells on the lateral sides of the nectariferous disc. (C) SEM micrograph showing the distal regionof the nectariferous disc with abundant trichomes and prominent stomata. (D). LM micrograph showing nectary trichomes stained with Toluidine Blue. (E). LM micrographs low-0g (arrow( a and
Y
cssasT(
O
ftipaT(
howing the nectary distal region with abundant trichomes stained with Fluorol Yeluard cells; UV autofluorescence. (G and H) LM micrographs showing nectar drops
H). (I) SEM micrographs showing the nectary distal region with prominent stomat
ellow phaseAt anthesis flowers present the nectariferous ring yellow in
olor. Numerous very minute liquid drops appear on the nectaryurface and rapidly enlarge. They are easily detectable because theyhine to the daylight (Fig. 4A). The stigma is wet, with a smallmount of exudate, and green in color. Its receptivity is demon-trated by the germination of pollen grains on its surface (Fig. 4E).he anthers are swollen of pollen and dehiscence takes placeFig. 4A).
range phaseThe nectariferous ring appears orange in color (Fig. 4B). Only
ew little drops are present on its surface. Nectar is accumulated inhe furrows between the nectary and the basis of the ovary, wheret is re-absorbed. Maximum pollen germination is recorded in this
hase (Fig. 4F), when stigmatic exudate is profuse and pollen tubesre evident in the stylar canals (Fig. 4G). The anthers are empty.he stigma begins drying, its color varying from green to brownFig. 4B).88. (F). LM micrographs showing the prominent stomata with outwardly projectings) displaying positive response to Alcian Blue (G) and Mercuric Bromophenol Blue
abundant trichomes.
Red phaseThe nectariferous ring is brilliant red. Its surface is dry and nectar
does not accumulate any more. Anthers fall down. The stigma is dryand brown (Fig. 4C). All flowers starting with yellow nectaries reachthe red phase. Fifteen hours are sufficient for completing the shiftfrom the yellow to the red phase.
Dark phaseThe nectariferous ring is dark red/brown in color, with black
patches (Fig. 4D); the surface is wrinkly and rough. It can well berecognized for a long time during the development of the fruit. Thestigma and the distal portion of the style are darkening as well.The ovary of some flowers (generally less than 50%) is increasingin size and develops fruits. All the other parts of the flower aresenescent.
The timing of the entire phenomenon, starting from the yellow
phase, is about two days. Laboratory investigation demonstratedthat isolated nectariferous rings turn from red to green-blue in halfa minute when dipped into an alkaline medium and immediatelyreverse to red, when rinsed in acid solution (Fig. 4H and I). This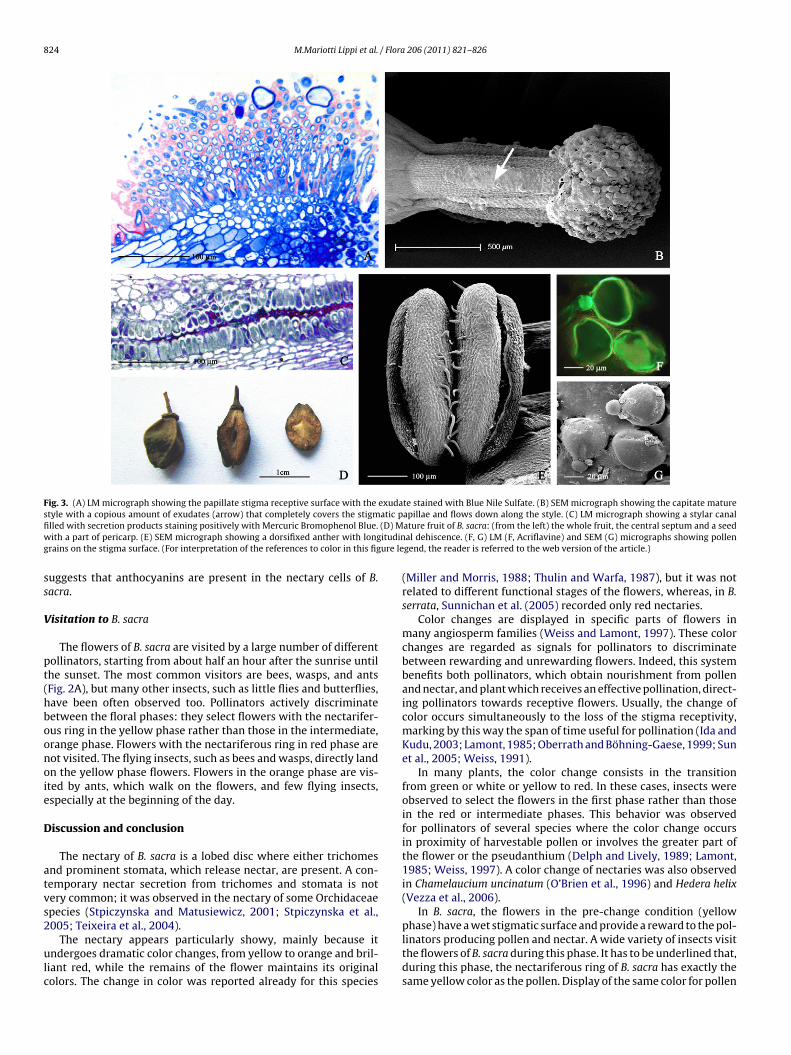
824 M.Mariotti Lippi et al. / Flora 206 (2011) 821– 826
Fig. 3. (A) LM micrograph showing the papillate stigma receptive surface with the exudate stained with Blue Nile Sulfate. (B) SEM micrograph showing the capitate maturestyle with a copious amount of exudates (arrow) that completely covers the stigmatic papillae and flows down along the style. (C) LM micrograph showing a stylar canalfi (D) Mw ituding ure le
ss
V
pt(hboonoie
D
atvs2
ulc
lled with secretion products staining positively with Mercuric Bromophenol Blue.ith a part of pericarp. (E) SEM micrograph showing a dorsifixed anther with long
rains on the stigma surface. (For interpretation of the references to color in this fig
uggests that anthocyanins are present in the nectary cells of B.acra.
isitation to B. sacra
The flowers of B. sacra are visited by a large number of differentollinators, starting from about half an hour after the sunrise untilhe sunset. The most common visitors are bees, wasps, and antsFig. 2A), but many other insects, such as little flies and butterflies,ave been often observed too. Pollinators actively discriminateetween the floral phases: they select flowers with the nectarifer-us ring in the yellow phase rather than those in the intermediate,range phase. Flowers with the nectariferous ring in red phase areot visited. The flying insects, such as bees and wasps, directly landn the yellow phase flowers. Flowers in the orange phase are vis-ted by ants, which walk on the flowers, and few flying insects,specially at the beginning of the day.
iscussion and conclusion
The nectary of B. sacra is a lobed disc where either trichomesnd prominent stomata, which release nectar, are present. A con-emporary nectar secretion from trichomes and stomata is notery common; it was observed in the nectary of some Orchidaceaepecies (Stpiczynska and Matusiewicz, 2001; Stpiczynska et al.,005; Teixeira et al., 2004).
The nectary appears particularly showy, mainly because itndergoes dramatic color changes, from yellow to orange and bril-
iant red, while the remains of the flower maintains its originalolors. The change in color was reported already for this species
ature fruit of B. sacra: (from the left) the whole fruit, the central septum and a seedal dehiscence. (F, G) LM (F, Acriflavine) and SEM (G) micrographs showing pollengend, the reader is referred to the web version of the article.)
(Miller and Morris, 1988; Thulin and Warfa, 1987), but it was notrelated to different functional stages of the flowers, whereas, in B.serrata, Sunnichan et al. (2005) recorded only red nectaries.
Color changes are displayed in specific parts of flowers inmany angiosperm families (Weiss and Lamont, 1997). These colorchanges are regarded as signals for pollinators to discriminatebetween rewarding and unrewarding flowers. Indeed, this systembenefits both pollinators, which obtain nourishment from pollenand nectar, and plant which receives an effective pollination, direct-ing pollinators towards receptive flowers. Usually, the change ofcolor occurs simultaneously to the loss of the stigma receptivity,marking by this way the span of time useful for pollination (Ida andKudu, 2003; Lamont, 1985; Oberrath and Böhning-Gaese, 1999; Sunet al., 2005; Weiss, 1991).
In many plants, the color change consists in the transitionfrom green or white or yellow to red. In these cases, insects wereobserved to select the flowers in the first phase rather than thosein the red or intermediate phases. This behavior was observedfor pollinators of several species where the color change occursin proximity of harvestable pollen or involves the greater part ofthe flower or the pseudanthium (Delph and Lively, 1989; Lamont,1985; Weiss, 1997). A color change of nectaries was also observedin Chamelaucium uncinatum (O’Brien et al., 1996) and Hedera helix(Vezza et al., 2006).
In B. sacra, the flowers in the pre-change condition (yellowphase) have a wet stigmatic surface and provide a reward to the pol-
linators producing pollen and nectar. A wide variety of insects visitthe flowers of B. sacra during this phase. It has to be underlined that,during this phase, the nectariferous ring of B. sacra has exactly thesame yellow color as the pollen. Display of the same color for pollen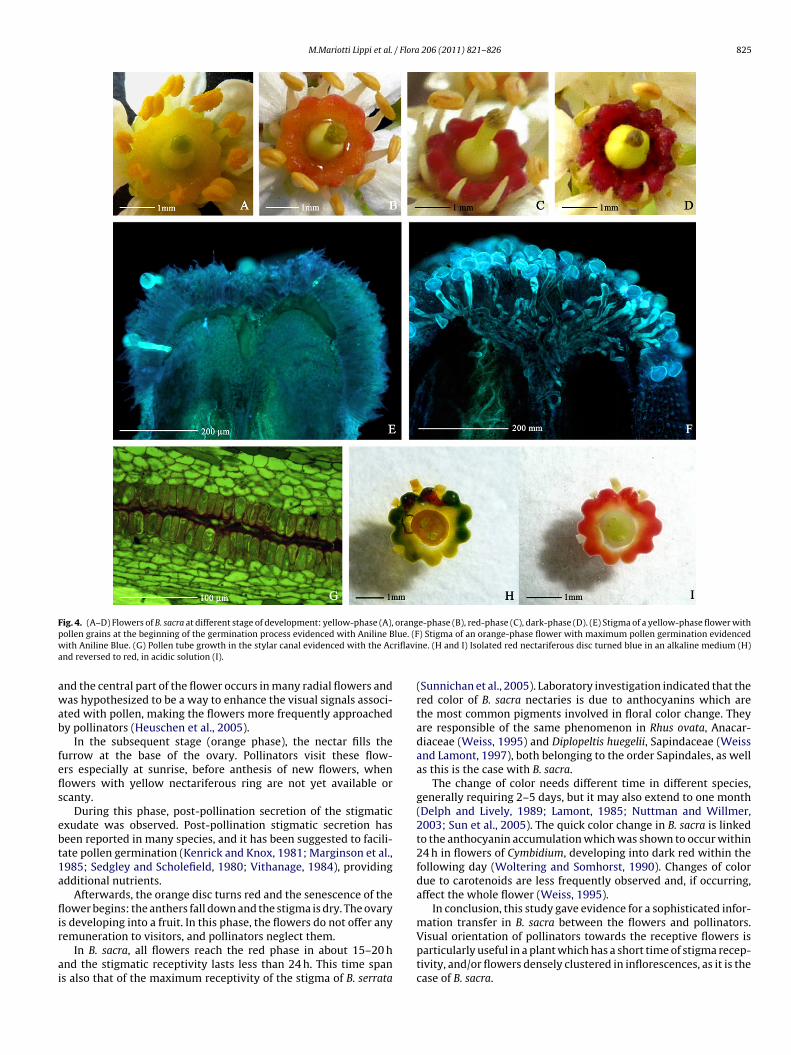
M.Mariotti Lippi et al. / Flora 206 (2011) 821– 826 825
Fig. 4. (A–D) Flowers of B. sacra at different stage of development: yellow-phase (A), orange-phase (B), red-phase (C), dark-phase (D). (E) Stigma of a yellow-phase flower withp lue. (Fw riflavia
awab
fefls
ebt1a
flir
ai
ollen grains at the beginning of the germination process evidenced with Aniline Bith Aniline Blue. (G) Pollen tube growth in the stylar canal evidenced with the Ac
nd reversed to red, in acidic solution (I).
nd the central part of the flower occurs in many radial flowers andas hypothesized to be a way to enhance the visual signals associ-
ted with pollen, making the flowers more frequently approachedy pollinators (Heuschen et al., 2005).
In the subsequent stage (orange phase), the nectar fills theurrow at the base of the ovary. Pollinators visit these flow-rs especially at sunrise, before anthesis of new flowers, whenowers with yellow nectariferous ring are not yet available orcanty.
During this phase, post-pollination secretion of the stigmaticxudate was observed. Post-pollination stigmatic secretion haseen reported in many species, and it has been suggested to facili-ate pollen germination (Kenrick and Knox, 1981; Marginson et al.,985; Sedgley and Scholefield, 1980; Vithanage, 1984), providingdditional nutrients.
Afterwards, the orange disc turns red and the senescence of theower begins: the anthers fall down and the stigma is dry. The ovary
s developing into a fruit. In this phase, the flowers do not offer any
emuneration to visitors, and pollinators neglect them.In B. sacra, all flowers reach the red phase in about 15–20 hnd the stigmatic receptivity lasts less than 24 h. This time spans also that of the maximum receptivity of the stigma of B. serrata
) Stigma of an orange-phase flower with maximum pollen germination evidencedne. (H and I) Isolated red nectariferous disc turned blue in an alkaline medium (H)
(Sunnichan et al., 2005). Laboratory investigation indicated that thered color of B. sacra nectaries is due to anthocyanins which arethe most common pigments involved in floral color change. Theyare responsible of the same phenomenon in Rhus ovata, Anacar-diaceae (Weiss, 1995) and Diplopeltis huegelii, Sapindaceae (Weissand Lamont, 1997), both belonging to the order Sapindales, as wellas this is the case with B. sacra.
The change of color needs different time in different species,generally requiring 2–5 days, but it may also extend to one month(Delph and Lively, 1989; Lamont, 1985; Nuttman and Willmer,2003; Sun et al., 2005). The quick color change in B. sacra is linkedto the anthocyanin accumulation which was shown to occur within24 h in flowers of Cymbidium, developing into dark red within thefollowing day (Woltering and Somhorst, 1990). Changes of colordue to carotenoids are less frequently observed and, if occurring,affect the whole flower (Weiss, 1995).
In conclusion, this study gave evidence for a sophisticated infor-mation transfer in B. sacra between the flowers and pollinators.
Visual orientation of pollinators towards the receptive flowers isparticularly useful in a plant which has a short time of stigma recep-tivity, and/or flowers densely clustered in inflorescences, as it is thecase of B. sacra.
8 / Flora
oTtcn(Bitmot
A
2
fisLa
R
B
B
D
H
I
K
L
26 M.Mariotti Lippi et al.
Despite the sophisticated strategy to attract pollinators devel-ped in B. sacra, the number of fruits of this plant is often scanty.his is consistent with observations carried out on B. serrata, wherehe low fruit set is considered not to be the result of inadequateross-pollination but caused by a possible limitation of availableutrients during the flowering period when leaves are absentSunnichan et al., 2005). This latter hypothesis is not convincing for. sacra that at the same time bears flowers and leaves when grow-
ng in the semi-desert environment of Wadi Dowkha. Concerninghe plants growing in the coastal areas, which are influenced by
onsoons, they may present contemporary flowers and leavesr bear only flowers, presumably in dependence of the seasonalrend.
cknowledgments
This research was carried out with financial aid of MURST (PRIN007R95LT4).
Author contribution: Marta Mariotti Lippi was responsible foreld observations and SEM analysis; Claudia Giuliani was respon-ible for LM analysis; Marta Mariotti Lippi, Claudia Giuliani andaura Maleci Bini drafted the discussion and conclusions. Technicalssistance by Tiziana Gonnelli.
eferences
eccari, N., Mazzi, V., 1966. Manuale di Tecnica Microscopica. Società EditriceLibraria, Como.
rundrett, M.C., Kendrick, B., Peterson, C.A., 1991. Efficient lipid staining in plantmaterial with sudan red 7B or fluorol [correction of fluoral] yellow 088 inpolyethylene glycol–glycerol. Biotech. Histochem. 66, 111–116.
elph, L.F., Lively, C.M., 1989. The evolution of floral color change: pollinatorattraction versus physiological constraints in Fuchsia excorticata. Evolution 43,1252–1262.
euschen, B., Gumbert, A., Lunau, K., 2005. A generalised mimicry system involvingangiosperm flower colour, pollen and bumblebees’ innate colour preferences.Plant Syst. Evol. 252, 121–137.
da, T.Y., Kudu, G., 2003. Floral color change in Weigela middendorffiana (Caprifoli-aceae): reduction of geitonogamous pollination by bumble bees. Am. J. Bot. 90,
1751–1757.enrick, J., Knox, R.B., 1981. Post-pollination exudates from stigmas of Acacia(Mimosaceae). Ann. Bot. 48, 103–106.
amont, B., 1985. The significance of flower colour change in eight co-occurringshrub species. Bot. J. Linn. Soc. 90, 145–155.
206 (2011) 821– 826
Marginson, R., Sedgley, M., Knox, R.B., 1985. Physiology of post-pollination exudatesproduction in Acacia. J. Exp. Bot. 36, 1660–1668.
Mazia, D., Brewer, P.A., Alfert, M., 1953. The cytochemical staining and measurementof protein with Mercuric Bromophenol blue. Biol. Bull. 104, 57–67.
Miller, A.G., Morris, M., 1988. Plants of Dhofar. The Office of the Adviser for Conser-vation of the Environment, Diwan of Royal Court, Sultanate of Oman. Edinburg.Holmes McDougal Limited, Scotland.
Nuttman, C., Willmer, P., 2003. How does insect visitation trigger floral colourchange? Ecol. Entomol. 28, 467–474.
O’Brien, S.P., McCully, M.E., 1981. The Study of Plant Structure Principles and SelectedMethods. Termarcarphi, Melbourne.
O’Brien, S.P., Loveys, B.R., Grant, W.J.R., 1996. Ultrastructure and function of floralnectaries of Chamelaucium uncinatum (Myrtaceae). Ann. Bot. 78, 189–196.
Oberrath, R., Böhning-Gaese, K., 1999. Floral color change and the attraction of insectpollinators in lungwort (Pulmonaria collina). Oecologia 121, 383–391.
Raffaelli, M., Mosti, S., Tardelli, M., 2003. The Frankincense Tree (Boswellia sacraFlueck., Burseraceae) in Dhofar, southern Oman: field-investigations on the nat-ural populations. Webbia 58, 133–149.
Sedgley, M., Scholefield, P.B., 1980. Stigma secretion in the watermelon before andafter pollination. Bot. Gaz. 141, 428–434.
Stpiczynska, M., Matusiewicz, J., 2001. Anatomy and ultrastructure of the spurnectary of Gymnadenia conopsea L. (Orchidaceae). Acta Soc. Botan. Polon. 70,267–272.
Stpiczynska, M., Milanesi, C., Faleri, C., Cresti, M., 2005. Ultrastructure of the nectaryspur of Platanthera chlorantha (Custer) Rchb. (Orchidaceae) during successivestages of nectar secretion. Acta Biol. Cracov., Ser. Botan. 47, 111–119.
Smith, M.M., McCully, M.E., 1978. A critical evaluation of the specificity of anilineblue induced fluorescence. Protoplasma 95, 229–254.
Sun, S.-G., Liao, K., Xia, J., Guo, Y.H., 2005. Floral colour change in Pedicularis mon-beigiana (Orobanchaceae). Plant Syst. Evol. 255, 77–85.
Sunnichan, V.G., Mohan Ram, H.Y., Shivanna,.K.R., 2005. Reproductive biology ofBoswellia serrata, the source of salai guggul, an important gum-resin. Bot. J. Linn.Soc. 147, 73–82.
Teixeira, S.P., Borba, E.L., Semir, J., 2004. Lip anatomy and its implications for thepollination mechanism of Bulbophyllum species (Orchidaceae). Ann. Bot. 93,499–505.
Thulin, M., Warfa, A.M., 1987. The frankincense trees (Boswellia spp., Burseraceae)of northern Somalia and southern Arabia. Kew Bull. 42, 487–500.
Vezza, M., Nepi, M., Guarnieri, M., Artese, D., Rascio, N., Pacini, E., 2006. Ivy (Hederahelix L.) flower nectar and nectary ecophysiology. Int. J. Plant Sci. 167, 519–527.
Vithanage, H.I.M.V., 1984. Pollen-stigma interactions: development and cytochem-istry of stigma papillae and their secretions in Annona squamosa L. (Annonaceae).Ann. Bot. 54, 153–167.
Weiss, M.R., 1991. Floral color changes as cues for pollinators. Nature 354, 227–229.Weiss, M.R., 1995. Floral color change: a widespread functional convergence. Am. J.
Bot. 82, 167–185.Weiss, M.R., 1997. Innate colour preferences and flexible colour learning in the
pipevine swallowtail. Anim. Behav. 53, 1043–1052.Weiss, M.R., Lamont, B., 1997. Floral color change and insect pollination: dynamic
relationship. Isr. J. Plant Sci. 45, 185–199.Woltering, E.J., Somhorst, D., 1990. Regulation of anthocyanin synthesis in Cymbid-
ium flowers: effects of emasculation and ethylene. J. Plant Phys. 136, 295–299.




















