
Fine-structural characterization of plant microbodies
24
Planta (Berl.) 81, 229--252 (1968) Fine-Structural Characterization of Plant Microbodies SUE ELLEN FI~EDERICK*, ELDON I-I.NEWCOMB, EUGENE L. VIGIL** and WILLIAM P. WEtgGIN Department of Botany, University of Wisconsin, Madison Received April 3, 1968 Summary. Morphology and distribution of the relatively less well known organelles of plants have been studied with the electron microscope in tissues fixed in glutaraldehyde and postfixed in osmium tetroxide. An organelle comparable morphologically to the animal microbody and similar to the plant microbody isolated by MOImEN~r~,R et al. (1966) has been encountered in a variety of plant species and tissues, and has been studied particularly in bean and radish roots, oat coleoptiles, and tobacco roots, stems and callus. The organelle has variable shape and is 0.5 to 1.5 tz in the greatest diameter. It has a single bounding mem- brane, a granular to fibrillar matrix of variable electron density, and an intimate association with one or two cisternae of rough endoplasmic reticulum (ER). Microbodies are easily the most common and generally distributed of the less well characterized organelles of plant cells. It seems very probable that they contain the enzymes characteristic of animal lysosomes (containing hydrolases) or animal microbodies (containing catalase and certain oxidases). Spherosomes are also possible sites of enzyme activity but are not as common or as widely distributed as microbodies. For this reason it appears likely that the particles designated as "plant lysosomes', "spherosomes", "peroxisomes", etc., in some of the cyto- chemical and biochemical studies on enzyme localization will prove to be micro- bodies. Variations in the morphology and ER associations of microbodies in tissues of bean and radish are described and discussed. "Crystal-containing bodies" (CCBs) are interpreted as a specialized type of mierobody characteristic of metabolically less active cells. Stages in the formation of CCBs from microbodies of typical appearance are illustrated for Arena. The general occurrence of microbodies in meristematic and differentiating cells and their close association with the Et~ suggest that they may play active roles in cellular metabolism. The alterations in their morphology and numbers that are observed in certain differentiating cells suggest further that the enzyme complements and metabolic roles of microbodies might change during cellular differentiation. If so, microbodies could be the functional equivalent of both microbodies and lysosomes of animal cells. Introduction Considerable uncertainty exists in the literature on plant fine structure and physiology concerning the appearance, properties, and distinctions, if any, between a number of single-membrane-limited organelles variously referred to as plant microbodies, plant lysosoInes, * NASA Predoctoral Trainee. ** Public Health Service Postdoctoral Fellow. 16 Planta (Berl.), Bd. 81
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Fine-structural characterization of plant microbodies

Planta (Berl.) 81, 229--252 (1968)
Fine-Structural Characterization of Plant Microbodies
SUE ELLEN FI~EDERICK*, ELDON I-I. NEWCOMB, EUGENE L. VIGIL** a n d WILLIAM P. WEtgGIN
Department of Botany, University of Wisconsin, Madison
Received April 3, 1968
Summary. Morphology and distribution of the relatively less well known organelles of plants have been studied with the electron microscope in tissues fixed in glutaraldehyde and postfixed in osmium tetroxide. An organelle comparable morphologically to the animal microbody and similar to the plant microbody isolated by MOImEN~r~,R et al. (1966) has been encountered in a variety of plant species and tissues, and has been studied particularly in bean and radish roots, oat coleoptiles, and tobacco roots, stems and callus. The organelle has variable shape and is 0.5 to 1.5 tz in the greatest diameter. I t has a single bounding mem- brane, a granular to fibrillar matrix of variable electron density, and an intimate association with one or two cisternae of rough endoplasmic reticulum (ER). Microbodies are easily the most common and generally distributed of the less well characterized organelles of plant cells. I t seems very probable that they contain the enzymes characteristic of animal lysosomes (containing hydrolases) or animal microbodies (containing catalase and certain oxidases). Spherosomes are also possible sites of enzyme activity but are not as common or as widely distributed as microbodies. For this reason it appears likely that the particles designated as "plant lysosomes' , "spherosomes", "peroxisomes", etc., in some of the cyto- chemical and biochemical studies on enzyme localization will prove to be micro- bodies.
Variations in the morphology and E R associations of microbodies in tissues of bean and radish are described and discussed. "Crystal-containing bodies" (CCBs) are interpreted as a specialized type of mierobody characteristic of metabolically less active cells. Stages in the formation of CCBs from microbodies of typical appearance are illustrated for Arena.
The general occurrence of microbodies in meristematic and differentiating cells and their close association with the Et~ suggest that they may play active roles in cellular metabolism. The alterations in their morphology and numbers that are observed in certain differentiating cells suggest further that the enzyme complements and metabolic roles of microbodies might change during cellular differentiation. If so, microbodies could be the functional equivalent of both microbodies and lysosomes of animal cells.
Introduction
Cons ide rab le u n c e r t a i n t y exis ts in t he l i t e r a tu r e on p l a n t f ine
s t r u c t u r e a n d p h y s i o l o g y conce rn ing t h e appea rance , p roper t i es , a n d
d i s t inc t ions , if any , b e t w e e n a n u m b e r of s i n g l e - m e m b r a n e - l i m i t e d organe l les v a r i o u s l y r e fe r red to as p l a n t mic robodies , p l a n t lysosoInes,
* NASA Predoctoral Trainee. ** Public Health Service Postdoctoral Fellow.
16 Planta (Berl.), Bd. 81

230 S.E. FREDERICK, E. H. I~EWCO~B, E. L. VIGIL and W. P. W~RGI~:
c rys ta l -con ta in ing bodies and spherosomes. Numerous references to these organelles occur, b u t often the f ine-s t ruc tura l ident i t ies are not clear.
The need for clar i f icat ion is he ightened b y the current in te res t in es tabl ishing the funct ions of var ious members of this pa r t i cu la r group of p l an t organelles. I t is an a t t r ac t i ve poss ib i l i ty t h a t one or more of t hem m a y be a source of hydro ly t i c enzymes which could be re leased dur ing cell d i f ferent ia t ion to br ing abou t b reakdown of cy top lasmic components and of localized regions of the cell wall. However , more def ini t ive s tudies of the types of organelles p resen t in p l an t cells are needed before known biochemical funct ions can be assigned to pa r t i cu la r t ypes of cy top lasmic inclusions.
W o r k in this l abo ra to ry has shown the occurrence of a d is t inc t ive t ype of membrane - l imi t ed body in a va r i e t y of p l a n t species and tissues. This organelle appears to be ident ica l to the body isolated, described, and ident i f ied as a " p l a n t m i c r o b o d y " by MOLL:ENtIAUEI~ et al. (1966). I t s des ignat ion as a p l an t mic robody seems app rop r i a t e in view of i ts m a r k e d s t ruc tu ra l s imi la r i ty to the an imal mic robody s tudied exten- s ively in l iver and k idney of ve r t eb ra tes (reviewed b y D~ D v v ~ and BAUDHUIN, 1966).
Our observat ions indicate t h a t p l an t microbodies are widely distr i- b u t e d in mer i s t emat ic and di f ferent ia t ing cells. I n view of this, i t would no t be surpr is ing to f ind t h a t t hey p lay a subs tan t i a l role in cellular metabol i sm. I t is i m p o r t a n t t h a t the i r i den t i t y and morpho logy be c lear ly es tabl ished in order to avoid confusion in subsequent biochemical work. Clarif icat ion of the f ine-s t ruc tura l proper t ies of p l a n t microbodies is the pr inc ipa l objec t ive of this paper .
Material and Methods Radish seeds (Raphanu8 8ativus L., cv. "White Icicle") were surface-sterilized
in a 10% Chlorox solution (Chlorox : 5.25% NaOC1) and then allowed to germinate at 22 to 24 ~ in petri plates containing moist filter paper. Seeds of bean (Phaseolus vulgaris L., cv. "Dwarf Horticulture") were soaked overnight in aerated tap water, then placed in moist sand or vermiculite to complete germination. Roots of intact, 2-day-old radish seedlings and of bean seedlings several days old were immersed in 3% glutaraldehyde and the terminal 2 to 3 mm of the tips removed. Fixation was in 3 % glutaraldehyde in 0.05 M phosphate buffer at pH 6.8 for a period of 1.5 hours. After fixation, the tissue was rinsed several times with phosphate buffer and post-fixed in 2 % osmium tetroxide for 2 hours. Dehydration was carried out in an acetone series followed by propylene oxide. The segments were embedded in Araldite-Epon (MOLLENHAUER, 1964). All steps in the fixation and dehydration processes were carried out at room temperature.
Seeds of Avena sativa L., cv. "Victory", were dehulled, soaked for 2 hours in distilled water, and placed on moist filter paper. After exposure to red light for 24 hours, the seeds were left in the dark at 24 ~ in a relative humidity of 85 % until the primary leaves had penetrated the eoleoptiles. At this time the coleoptiles, 4 to 5 cm in length, were immersed in 3 % glutaraldehyde and divided into 1-mm

Fine-Structural Characterization of Plant Mierobodies 23I
segments. The segments were subjected to the same fixation and embedment procedures as the root tips of radish and bean. Root and stem segments of young tobacco seedlings (Nicotiana tabacum L., ev. "Wisconsin 38") and cultured callus cells of this species were fixed and embedded in the same manner.
Silver-gray sections of all embedded material were cut on a Sorvall MT-1 or MT-2 Ultramicrotome, mounted on bare 300 • 75 copper grids, and stained for 10 min in ~queous 2% uranyl acetate, followed by lead citrate for 5 rain. Sections were viewed in a Hitachi HU-11A microscope at an accelerating voltage of 50 or 75 kV.
Results
General Observations. The organelles identified as plant microbodies on the basis of observations made in this laboratory are bounded by a single membrane, are approximate ly 0.5 to 1.5 ~ in the greatest diameter, exhibit spherical, elongate, or irregularly-shaped profiles in section, have a granular to fibrillar matr ix of varying electron density, and possess a close relationship to one or more cisternae of the endoplasmic ret iculum (ER). Dense amorphous or crystalline inclusions m a y occur in their matrix, bu t are no t typical features. ~[hese inclusions appear to be associated with certain stages of microbody development, and to be more characteristic of some plant species and cell types than of others.
Our detailed observations on plant microbodies have been made on tissues in root tips of bean and radish, coleoptiles of Arena, and in stems, roots, and callus of tobacco. In addition, we have observed similar organelles in the tissues of a number of other plant species. Although they va ry somewhat in appearance, mierobodies f rom different tissues in the same plant and from different plant species are sufficiently alike tha t they can be distinguished readily fi'om other cellular com- ponents.
I n order to establish a p ro to type tha t can be used to distinguish them f rom other cytoplasmic inclusions with which they m a y be confused, the mierobodies commonly encountered in cells of the bean root tip roll be described in detail.
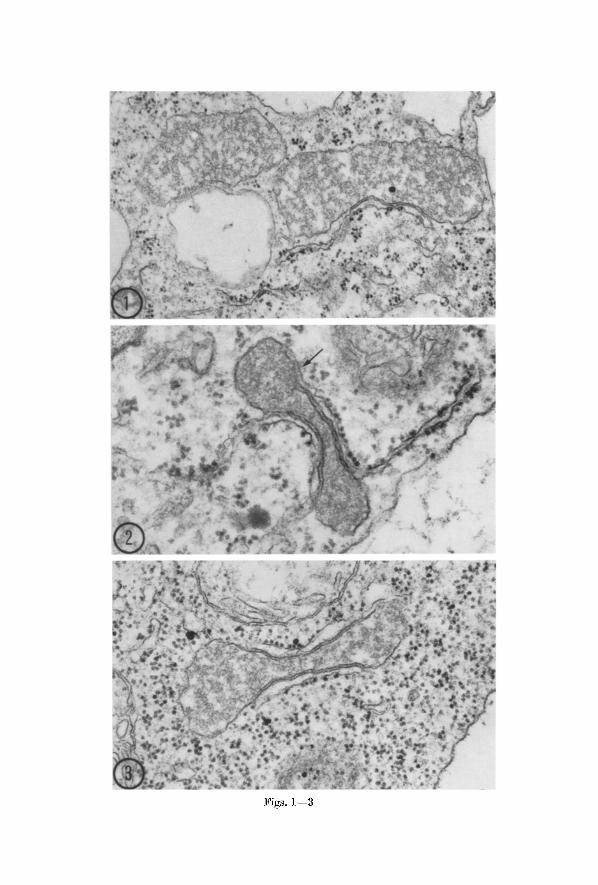
Microbodies o] Phaseolus vulgaris. 0rganelles conforming to the above description of plant mierobodies have been observed in meristematic cells, cells of the cortex, epidermis and root cap, and several types of differentiating eelIs in the xylem and phloem of the root tip of bean (Figs. 1--10). The microbodies exhibit a var ie ty of spherical to elongate shapes in section, and are about 0.5 to 1.5 ~ in the greatest diameter. I n some, the matr ix is composed of a fibrillar material aggregated so as to give random dense and t ransparent regions (Figs. 1, 3, 4 and 5), while in others it is more uniformly electron dense (Figs. 2, 6--10). Generally the matr ix extends to the limiting membrane.
The microbodies in bean root-t ip cells are wi thout fur ther definitive internal structure. Crystalline inclusions are not observed, a l though
16"
~igs. 1--3

S. E. FRFA)ERIeI~ et al. : Fine-StructurM Characterization of Plant Microbodies 233
frequent ly small, finely granular to amorphous electron-dense areas are present (Fig. 9). These resemble the dense inclusions observed by MOLLE~HAU~ et al. (1966) in mierobodies of the outer root cap cells of maize and vanilla orchid. Their occurrence seems not to be a character- istic of mierobodies in any part icular region of the root t ip; probably they are present in a var ie ty of cell types. The remainder of the matr ix in microbodies with dense inclusions often appears more electron dense and homogeneous than in others. Possibly these variations in the appear- ance of the matr ix reflect a change in content taking place within the mierobody.
The boundaries of some mierobodies are irregular in contour (Figs. 1 and 4). Frequent ly some portions of the limiting membrane are tangent or oblique to the plane of section, causing the membrane to appear indistinct and the boundary diffuse. The matr ix m a y then appear to merge with the surrounding cytoplasm (Figs. 3 and 4).
Rough-surfaced E R is associated with mierobodies in the bean root regardless of cell type (Figs. 1--9). The association appears especially impor tan t in view of its constancy and the absence of a similar relation- ship between the E R and other organelles. I n virtually all eases the E R element bears ribosomes on the outer membrane and lacks them on the membrane facing the mierobody. Surface views of the membranes suggest t ha t the ribosomes are present as polysomes, as in Fig. 6. The appearance in Fig. 6 would be expected if a s tructure like tha t in Fig. 2 or 3 were sectioned normal to the plane shown and included the ribosomal surface of the associated eisterna. Commonly the E R mem- branes tha t run alongside the microbody can be followed for some distance in the cytoplasm where they exhibit the profile of a typical eisterna (Figs. 2 and 4). Small vesiculate E R elements are also seen ra ther frequently adjacent to microbodies (Fig. 5). The diversi ty in the configuration of the E R and in the shape of the mierobodies great ly increases the var ie ty of ER-mierobody associations seen in thin sections.
Some of the mierobodies of bean root tip cells are elongate and somewhat dumbbell-shaped in profile, and are lined with E R eisternae in their eonstrieted regions on one or both sides (Figs. 2 and 3). The frequency with which microbodies of this sort are encountered suggests
Figs. 1--3. Microbodies in cells of the bean root tip. Fig. 1. Two microbodies in meristematic cell. A cistern~ of rough endoplasmic reticulum with ribosomes on
the outer membrane runs alongside the larger mierobody. • 45,000. Fig. 2. Dumbbell-shaped microbody with relatively dense matrix in root cap cell. ER cisternae are seen on both sides of the profile of the microbody. The arrow indicates a region where the membranes are indistinct, suggesting a connection between microbody and ER. • 80,000. Fig. 3. Dumbbell-shaped microbody in a meristematic cell. Rough EI~ is present on both sides of the microbody. The
fibrillar material in the matrix is not uniformly dispersed. • 59,000
Figs. 4 and 5

S. E. FREDERICK et al. : Fine-Structural Characterization of Plant Microbodies 235
that they have the same shape three-dimensionally. We do not consider that these or other amoeboid forms represent microbodies in division.
No clear openings of the cisternae into the interiors of the mierobodies have been observed, although micrographs suggestive of such connec- tions are occasionally obtained. Areas are often seen where the adjacent E g and mic robody membranes are ind is t inc t (arrow, Fig. 2) b u t i t is not clear t h a t these do in fact represent membrane connections. I n sections normal to the membranes the associa ted E R eisternae genera l ly appear to be sepa ra t ed from the microbodies b y a r a the r un i fo rm dis tance of 50 to 80 A. I n some cases i t has been de t e rmined t h a t the E R bears the same spat ia l re la t ionship to a mic robody in several successive sections, indica t ing t h a t the associat ion m a y ex tend for some dis tance a round the organelle.
I n some cells mierobodies bear ing ta i l - l ike extensions have been encountered (Fig. 7). These appendages m a y represent E R eis ternae; if t hey do, t hey cons t i tu te the only cases in which direct connect ions of E R with microbodies can be demons t r a t e d unambiguous ly in our micrographs .
Microbodies in mer i s temat ic cells and in var ious types of differ- en t ia t ing cells in bean m a y exhib i t wide differences in morpho logy and in the i r associat ion with the ER. They also v a r y in numbers and in the ex ten t to which t hey are local ly concen t ra ted in cells. I t is no t known whether changes in the i r appearance and numbers are corre la ted wi th specific cellular changes re la ted to d i f ferent ia t ion; however, such changes have p r e sumab ly a funct ional basis and m a y reflect i m p o r t a n t shifts in the act ivi t ies of the organelles and the i r roles in the cell.
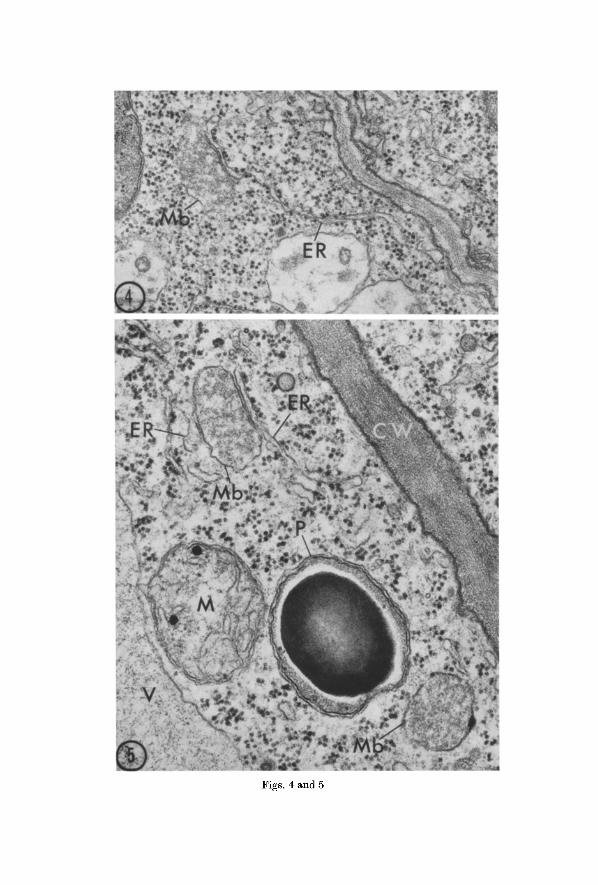
I n mer i s t emat ic cells as well as in cells in ear ly s tages of differentia- t ion, microbodies are re la t ive ly small and possess de l ica te ly s ta ined membranes (Fig. 4). They are often i r regular in outl ine, and the i r contents are th in and are unequa l ly d is t r ibu ted . Associa t ion wi th the E R usua l ly consists of a single f l a t t ened cis terna running alongside for some distance.
Dis ta l ly and la te ra l ly in somewhat older d i f ferent ia t ing cells of the p roeambia l s t rands and in ma tu r ing or ma tu re p a r e n c h y m a cells, the
Fig. 4. Microbody (Mb) in a meristematic cell of a bean root tip. The microbody has an irregular profile and a delicately stained membrane. The associated cisterna of rough endoplasmic reticulum (ER) continues for some distance in the
cytoplasm. • 52,000
Fig. 5. Portion of an elongating cell in a bean root tip showing two microbodies (Mb), a mitochondrion (M), and a plastid (P) with a prominent starch granule lying near the cell wall (CW)~ The microbody in the upper left has a cistema (ER) with ribosomes on the right and a vesicle of endoplasmic retieulum (ER) without
ribosomes on the left. Part of a vacuole (F) at lower left. • 60,000

Figs. 6--8

S. E. FREDERICK et ~I. : Fine-Structural Characterization of Plant Microbodies 237
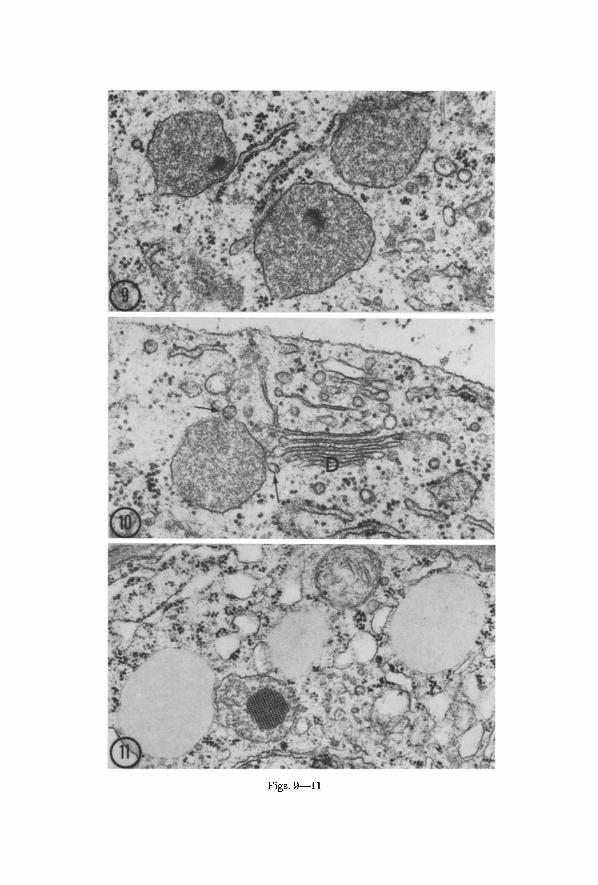
microbodies are commonly larger and have dense contents. They may assume a number of distinctive forms some of which have already been described. The microbodies with dense amorphous cores (Fig. 9) and those with smooth tail-like extensions (Fig. 7) are relatively common in cells of the procambial strands and may be found also in adjacent parenchymatous cells of the cortex. Of interest also are elongate mierobodies whose bulbous terminal portions are joined by a tube-like connection possessing a more prominently stained membrane (Fig. 8). Although the mierobodies of older cells commonly retain an association with the E g , it is usually not as intimate or as extensive as in younger cells.
Some of the microbodies in differentiating cells of the procambial strands show a suggestive association with dietyosomes. In the example shown in Fig. 10, a mierobody with a dense matrix lies adjacent to a dietyosome actively producing vesicles. Several of the vesicles are close to the mierobody and resemble it in content. Their appearance raises the possibility tha t they may fuse with the microbody and contribute to its matrix.
Microbodies arc generally not so numerous as mitochondria or dictyosomes but their numbers are within the same order of magnitude. Essentially all thin sections of living bean root cells show examples of the latter two organelles, while mierobodies are sometimes not re- presented. They appear to vary in numbers with cell type, and are sometimes concentrated locally in the cell. I t is however difficult to make more specific evaluations of their relative numbers in different cell types and of their numbers relative to other cytoplasmic components without statistical studies.
Microbodies of Raphanus sativus. Mierobodies have been found consistently in the different regions of the radish root tip so far examined. Although they are occasionally dumbbell-shaped or irregular in outline like those of bean, they are more frequently round to oval, with a prominently stained bounding membrane. Their matrix is similar to tha t of bean microbodies in electron density and lack of crystalline inclusions, but is usually more homogeneous in appearance (Figs. 12--16).
Figs. 6--8. Microbodies with dense contents and distinctive shapes present in differentiating cells of a procambial strand in bean root. Fig. 6. Mierobody with swollen ends. The appearance of the mid-region suggests that a closely associated ER cisterna has been sectioned tangentially so as to reveal a polysome (Ps) on the outer membrane. • 61,000. Fig. 7. Microbody bearing a "tail" (arrow) in addition to a closely associated ER cisterna (ER). The tail possibly represents a cisterna of ER connected to the mierobody. • 49,000. Fig. 8. A
bi-lobed microbody with dense matrix and a tube-like mid-region, x 60,000
Figs. 9--11

S. E. Fg~D~RICK et al. : Fine-Structural Characterization of Plant Microbodies 239
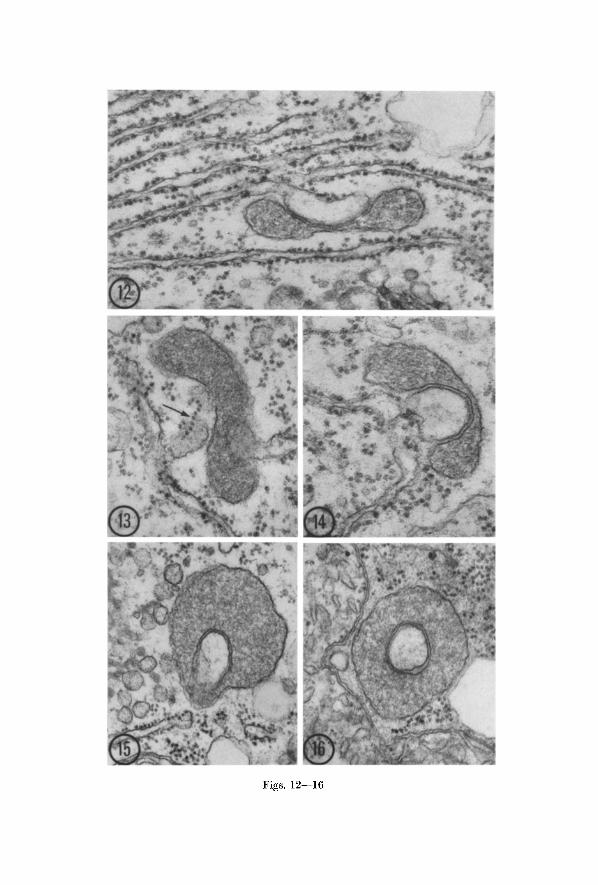
It is in the association with other cytoplasmic membranes (appar- ently the ER) that the radish mierobodies show a marked divergence from those of bean. Although some of the associations are of the familiar sort in which the ER lies alongside the organelle, more typically, at least in young epidermal and cortical cells of the root, vesieulate elements are partially or completely encircled by a mierobody (Figs. 14 to 16). Serial sections would be required to determine whether the sur- rounded components are in fact completely enclosed, or merely lie in pockets.
If the enclosed vesieulate bodies are of Et~ origin, then Figs. 12--16 may represent stages in a process in which microbodies engulf portions of EI~ cisternae. Fig. 12 is particularly suggestive since it clearly shows that the bleb of the eisterna to which the microbody is appressed is a component of an extensive, organized system of EI~. This implies that this particular association between microbody and EI~ may have arisen only a short time previously and may represent a very early stage in the engulfment process. I t is interesting note to that the eisterna has already become swollen and has lost its ribosomes from the surface facing the microbody.
Cry8talloids in Microbodies. Recent fine-structural studies (reviewed in the Discussion) have established that single-membrane-bounded organelles containing crystalline inclusions occur widely in plants. These organelles, which have been termed crystal-containing bodies (CCBs), are similar to the mierobodies found in radish and bean, except for the presence of the highly organized cores.
Observations of eoleoptile tissues of Arena and root and stem tissues of tobacco have shown the presence of these crystal-containing bodies (Figs. 21--240 . However, bodies morphologically indistinguishable from the bean and radish microbodies also occur frequently in the Arena and tobacco tissues (Fig. 17), as do organelles of typical mierobody appear- anee but in the earliest stages of formation of crystalline inclusions (Figs. 18--20). These observations dearly indicate that the CCBs arise as differentiations of microbodies of the non-crystalline type.
Figs. 9 and 10. Microbodies in differentiating cells of a procambial strand in bean root. Fig. 9. Three mierobodies with associated ER eisternae. Two of the micro- bodies exhibit dense inclusions. • 49,000. Fig. 10. Dictyosome (D) adjacent to a mierobody. The arrows point to two dietyosome vesicles with dense contents
resembling the matrix of the mierobody. • 59,000
Fig. 11. Three spherosomes and a CCB in a xylem parenchyma cell in the hypoeotyl of a seedling of Acer platanoides L. Although tile boundaries of the spherosomes are more electron dense than the interiors, they do not appear similar to typical
membranes such as that enclosing the adjacent CCB. • 39,000
Figs. 12--16

S. E. FREDERICK et al. : Fine-Structural Characterization of Plant Mierobodies 241
Mierobodies which lack crystalline inclusions or appear to be in incipient stages of crystal format ion are found predominant ly in the younger cells of the eoleoptile and tobacco tissues. When such structures are seen in older, more mature cells where the CCBs predominate, serial sections indicate t ha t they frequently represent CCBs sectioned in planes including little or none of the crystal. These observations suggest t ha t formation of crystalline inclusions represents a normal pa t te rn of development tha t takes place in mierobodies, especially in mature parenehyma cells.
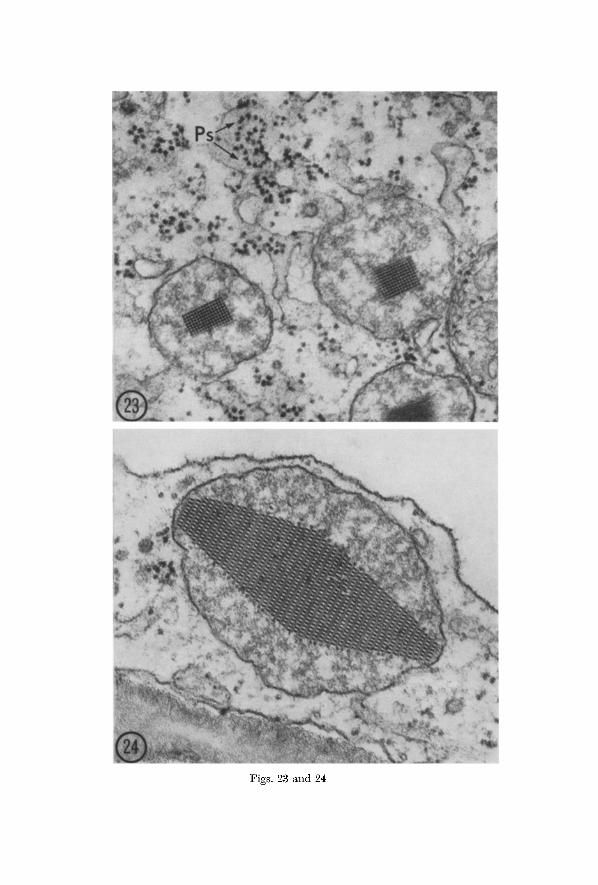
Rough-surfaced E R is associated with mierobodies in all stages of crystal formation in Arena and tobacco al though the relationship is usually not as close nor as extensive as in ordinary microbodies. Specific pat terns in the topographical relationships between E R and mierobodies with crystalline content are not usually discernible. Crystallization within the microbody often starts in a region near the limiting membrane adjacent to the closely associated E R eisterna (Figs. 19--22). Surface views of E R in close association with mierobodies with crystalline inclusions show tha t the ribosomes are present in the polysomal configurations characteristic of rough EI~ generally (Fig. 23). As in the bean and radish root tip, ribosomes are always absent along the ER- membrane surfaces adjacent to the CCBs of Avena and tobacco.
Discussion
General. Organelles similar to the microbodies described in this paper have been observed previously in plant tissues by a number of investi- gators, but have generally received little at tention. Consequently they have not ye t received the thorough morphological characterization necessary to establish them as a separate type of organelle distinguish- able f rom other cytoplasmic bodies. MOLL~HAUE~ et al. (1966) have described some of their morphological characteristics as well as their occurrence in a wide var ie ty of plant species, including algae and fungi, and their isolation in intact form from maize roots and onion stems.
Figs. 12--16. Microbodies in epidermal and cortical cells of radish root tips presumably illustrating successive stages in the enfoldment of ER cisternae within microbodies. Fig. 12. An early stage of association in which the swollen cisternal element is still part of an organized system of rough endoplasmic reticulum but no longer bears ribosomes on the membrane next to the microbody. • 59,000. Fig. 13. The cisterna associated with the microbody is sectioned tangentially revealing a polysome (arrow) on the surface. • 62,000. Fig. 14. A microbody enfolds a greatly swollen ER cisterna. • 71,000. Fig. 15. A microbody encloses a vesicle assmned to have been pinched off an ER cisterna. Note profile of ER nearby. • 48,000. Fig. 16. Microbody containing a vesicle presumably derived from a oisterna of ER. The vesicle may be completely isolated within the microbody or
may lie in a pocket. • 52,000

Figs. 17--22

S. E. FREDERICK et al. : Fine-StrueturM Characterization of Plant Mierobodies 243
However, most of the illustrations in their paper are based on material fixed in permanganate and do not show the structural details tha t are obtMned with glutaraldehyde fixation.
Lack of thorough fine-strueturM characterization of these bodies has been accompanied by a confused terminology. I t is likely tha t the organelles termed "ph ragmosomes" (Po~T~R and CAVLFIELD, 1958; POTTER and MACUADO, 1960 ; MA~TOX, 1961), " lysosomes" or "dense bodies" (ScuN~eF, 1964), and "lysosome-like bodies" (BoucK and CROSSgAW, 1965) are the same as those described in this s tudy. MOLnENI~AU~R et al. (1966) improved the si tuation by noting tha t the bodies t hey were investigating bore a marked structural similarity to the mierobodies of animal cells, and by proposing tha t the plant or- ganelles be called "mierobodies" also. I n the present state of knowledge, this te rm seems preferable to others since it does not imply tha t these bodies have any part icular function, and conforms to historical precedent in using structural appearance as the basis for naming a cytoplasmic component .
Comparison o/Plant and Animal Microbodies. The term " m i e r o b o d y " was first applied to membrane-bounded organelles in convoluted tubule cells of mouse kidney (R~oDI>', 1954). I t is now known tha t mierobodies occur consistently in certain cells of kidney and liver. However, in contras t to plant microbodies, animal mierobodies do not occur in cells generally.
The funetion of mierobodies in animal cells is not ye t clear, despite extensive biochemical s tudy. I t has been shown tha t these organelles contain eatalase and a var ie ty of hydrogen-peroxide-producing oxidases. Several metabolic roles suggested by this part icular combinat ion of enzymes have been discussed by Dn Duv~ and BAUDHUI~ (1966).
The most characteristic morphological features of both animal and plant mierobodies are single limiting membranes, finely granular
Figs. 17--22. Stages in the development of crystM-contMning bodies from ordinary microbodies in parenchyma cells of Avenc~ coleoptiles. The arrows in Figs. 19 22 indicate the ER associated with the microbody. Fig. 17. Microbody shewing little or no crystallization of the matrix. At least one element of the ER is closely associated with the mierobody. • 71,000. Fig. 18. Mierobody with changes in the appearance of the matrix suggestive of incipient erystMlization. Two or more vesieulate elements of the ER lie close to the right side of the mierobody. • 56,000. Fig. 19. Mierobody with erystMlization beginning in the matrix next to the associated ER. • 68,060. Fig. 20. Early stage of crystal formation in a mierobody. The erystMlization oceurs along the margin next to a vesicle of ER. • 68,000. Fig. 21. More extensive erystMlization of the microbody matrix. The crystal lies near the associated ER. • 69,000. Fig. 22. Mierobody showing a somewhat more advanced stage in which crystallization is taking place in the matrix next to a
cisterns of ER. • 63,000
Figs. 23 ~nd 24

S. E. FREDERICK et al. : Fine-Structural Characterization of Plant Microbodies 245
matrices that may include dense nucleoids of amorphous or crystalline nature, and close association with the endoplasmic reticulnm. The Et~ associated with animal microbodies usually consists of short membranous elements without ribosomes, although in some cases it is composed of rough-surfaced cisternae (DAE~S, 1966). In plant microbodies, according to our findings, the associated ER is, on the other hand, usually in the form of rough-surfaced cisternae. In both animals and plants, the cisternM membranes adjacent to the microbodies are always devoid of ribosomes.
Whether a continuity exists between the interior of the EI~ and that of the microbody is not clear for either animals (DAEMS, 1966; Ess~TER, 1967) or plants. In plants, profiles can be seen in which the ER running alongside appears to merge with the microbody, but the membrane continuity between the two structures is never clearly defined. I t is instructive to contrast this ambiguity with the clear evidence of continuity between the ER and the cisternal swellings in the case of protein accumulation in radish root cells (BONNETT and NEWCOMB, 1965).
There is evidence that both plant and animal mierobodies may arise from the ER. This evidence is more convincing for animal microbodies, and it seems likely that they originate as dilated regions of ER which may or may not become detached (H]cuBAN et M., 1963; NOVI~:OFF and Sn~N, 1964; ESS~ER, 1967). In the infrequent examples in which a plant microbody is interpretable as an enlargement of a cisterna of the ER, there is no unequivocal evidence for continuity between the two structures. Nevertheless, the close association with the El% generally shown by microbodies, together with the rarity of suggestive relation- ships with other cellular structures, makes an origin from the ER seem likely.
Apart from the possible involvement in microbody ontogeny, the significance of the associations of ER with plant microbodies is obscure. These associations are far too common to be coincidental, however, and it seems likely that they are of some functional importance. A possible influence of the cisterna on the microbody can be observed in young CCBs in which crystallization of the matrix frequently starts in the margin next to the adjacent ER.
In view of their ultrastructural similarities, the possibility is attrac- tive that plant microbodies contain enzymes similar to those of animal
Figs. 23 and 24. Crystal-containing microbodies in parcnchyma cells of the Avena coleoptile. Fig. 23. Two CCBs apparently in intimate association with a ramifying ER system. At least two polysomes (Ps) are visible on the surface of the ER. • 74.000, Fig. 24. A microbody sectioned so as to reveal a striking and commonly encountered pattern in a large crystal. Note the imperfection in the crystal as well as the addition of subunits apparently taking place along its margins. • 93,000
17a Planta (Berl.), Bd. 81

246 S.E. FREDEI~ICK, E. H. NEWCO~B, E. L. VIGIL and W. P. WERGIN:
microbodies. However, plant microbodies have not yet been charac- terized biochemieally. Although recent biochemical and eytochemical studies have suggested strongly tha t certain plant enzymes are largely confined to particulate fractions of the cytoplasm, the particles in these fractions have not been adequately characterized ultrastructurally. The resulting confusion in terminology will be considered in a later section.
Microbodies vs. Spherosomes. Most cytoplasmic organelles in both bean and radish are easily distinguished from microbodies. Mitochondria and plastids are generally larger and more numerous, and bear little structural resemblance to them. Young plastids frequently are similar to microbodies in both size and shape, but arc limited by an envelope rather than a single membrane. The only organelles we have observed with which microbodics might be confused are the bodies shown in Fig. l l , which we consider to be spherosomes. We have encountered bodies similar to these in a wide variety of plants. They do not appear to have nearly so general a distribution within tissues as microbodies, although in certain cells they may be quite numerous. In size they arc comparable to microbodies. They are most readily distinguished from microbodies according to the following characteristics: 1. They are smoothly spherical or nearly so, in contrast to an irregularity of margin and shape in microbodies; 2. they are not clearly bounded by a mem- brane; 3. their contents are quite homogeneous, whereas the matrix of microbodies is granular or fibrillar; 4. the electron density of their contents is much less than tha t of microbodics; and 5. they lack the close association with ElZ shown by microbodies.
The fine structure of spherosomes has not been well established, and it is not yet clear whether all published electron-microscopic investiga- tions of "sphcrosomes" refer to the same body (•lCEY-WYssLING et al., 1963 ; Sol~oI~I~ and SOl~OKI~, 1966 ; JACKS et al., 1967). The spherosomes we have observed have certain features in common with those described by SOl~OKI~ and Sol~o~I~ (1966) but commonly lack the electron- transparent central areas described by these authors. We have on occasion observed bodies with such a transparent center; we believe this appearance results from ineomplete penetration of the fixative into spherosomcs.
Spherosomes are commonly considered to be membrane-bounded. In our micrographs of these bodies from a variety of plant species, the electron density is greater at the periphery than in the matrix, but does not resemble a well-defined bounding membrane similar to that sur- rounding other cellular organelles.
Relationship between Microbodies and Crystal-containing Bodies. Single membrane-bounded organelles containing crystalloids have received considerable attention in recent fine-structurM literature on

Fine-Structural Characterization of Plant Mierobodies 247
plants. These bodies, referred to as "crystal-containing bodies" (CCBs), were first described by THORSTO~ and T~IMAC<X (1964) in subepidermal cells of the oat coleoptile. They have been investigated further in Avena (C~oNsHAw, 1964; C~O~SHAW and BoucK, 1965; O'B~I~N and THIMANN, 1967), and have been observed in a wide variety of plant tissues (G~oLA and Bassi, 1964; BoucK, 1965; KOL~HMAI>-]SN et al., 1965; Maei~os, 1965; WALLES, 1965; P~IC~, 1966; AasOTT and S~IT~, 1967; Js.~-s]~N and VALDOW~OS, 1967; PETZOLD, 1967; VILLI~S, 1967). The CCBs contain crystalline nueleoids consisting of highly organized units of macromolecular dimensions and repeat distances which vary according to the species. The nature of the nueleoids has not been determined, but they are generally believed to consist of protein or lipoprotein (CI~oNsI~AW, 1964; O'BeI~N and TmMA~, 1967; PETZOLD, 1967). They are surrounded by a granular to fibrillar matrix. While the CCBs are sometimes associated with ER, the association is not as prominent or frequent as that between ER and the mierobodies described above.
The relationship of the CCBs to typical microbodies has not been clearly established, although several investigators have concluded that the two organelles may be closely related or identical (WALLES, 1965; O ' B R I ~ and T~IMANX, 1967; P~TZOLD, 1967; VILLIS,~S, 1967). No studies have demonstrated the ontogenetic relationships between micro- bodies and CCBs, however, nor have papers devoted to the CCBs included more than casual observations on microbodies.
The numerous transitional forms between microbodies and CCBs observed in the Avena coleoptile and other plant tissues studied in this laboratory indicate that the CCBs represent merely a specialized type of mierobody. The likelihood that they constitute an inactive form of this organelle is suggested by 1. their predominant localization in storage cells and other metabolically less active cells, 2. their less intimate association or complete lack of association with ER, 3. their round to ovoid conformations and smooth contours in contrast to the variety of amoeboid forms found among the microbodies of growing cells, and finally 4. their crystalline content suggesting functional inactivity.
Several proposals of possible functional roles have been made for the CCBs (CRo~sHAw, 1964; MARINOS, 1965; O'B~I~N and TttI~ANN, 1967), while the inconspicuous non-erystalloid mierobodies have received virtually no attention. I t seems to us, however, that active metabolic roles are more likely to be found in the non-crystalloid microbodies. The observation that some species and tissues contain abundant microbodies none of which have crysta]loids appears to support this view.
Enzyme Localization in Plant Particles and the Problem o/ Particle Identification. In recent years it has been demonstrated that certain enzymes in plant cells are not localized in mitoehondria or the other
17"

248 S.E. FEEDERI~CK, E. H. NEweo~B, E. L. VIGIL and W. P. W~RoI~:
more familiar organelles of the cell, but in other, as yet poorly defined particles. However, a t tempts to identify these particulate structures have been unsatisfactory both because the fine-structural characteriza- tions are either lacking or inadequate, and because the terms applied to the particles ("spherosomcs ", "plant lysosomes ", etc.) have themselves never been adequately defined at the fine structural level.
As a result, the conclusion is almost inescapable that the particles now bearing other names in some of the enzyme localization studies will prove to be identical with microbodies. Examination of a variety of tissues in a number of different plant species has convinced us tha t there are only one or two kinds of poorly defined organelles present in suffi- cient numbers in cells to serve as the sites of any appreciable enzyme activity. Among these, microbodies appear to be the most common and generally distributed. Spherosomes, as previously defined in this paper, are also possible sites of activity although they are neither as common nor as widely distributed as microbodies. No other single-membrane- bounded organelles which could be envisaged as ubiquitous "p lan t lysosomes" have been seen in our studies.
Recently it has been found tha t certain hydrolytic enzymes are localized in cytoplasmic particles in plant cells. Unfortunately, the fine- structural identity of these particles is not yet clear. HARlCINGTON and ALTSCHVL (1963) found tha t several hydrolases are sedimentable with a particulate fraction in onion seedlings. MATILE et al. (1965), BALz (1966) and SEMADENI (1967) have shown similar associations of four acid hydrolases with cytoplasmic particles in seedlings of corn and tobacco; they consider the particles to be "spherosomes" and "prospherosomes", and equate them to animal lysosomes. However, it is not possible to determine from the micrographs whether the particles are identical either to microbodies or spherosomes.
Numerous cytochemieal studies with the light microscope have confirmed the biochemical evidence indicating that acid phosphatase and various other hydrolases are localized in particles in plant cells (AVERS, 1962; OLSZEWSKA and GA]3ARA, 1964; GAHA~r 1965; RASC~ et al., 1965; WALEK-CzERNECKA, 1965; GAHAN and MAPLE, 1966; HOLCO~B et al., 1967). In some of these studies, the particles are identified as "spherosomes". Since the fine structural identity of these bodies is controversial it cannot be determined whether any of them correspond to those observed in our work.
There is however reason to believe that some of the particles iden- tified as "spherosomes" or "phragmosomes" in both light- and electron-microscopic studies are plant microbodies. 0LSZEWSKA and GABARA (1964) and WALEK-CzEx~ECKA (1965) have reported that several acid hydrolases, including aryl sulfatase, are localized in "spherosomes"

Fine-Structural Characterization of Plant Microbodies 249
of plant cells. The bodies described by WALEK-CzEENECKA sometimes contained "lipid (?) crystals", an observation that suggests they may be identical to crystal-containing microbodies. In eytoehemical studies at the electron-microscope level, Pol lx (1966) has localized aryl sulfatase activity in the ER and associated cytoplasmic particles considered to be "phragmosomes". We are not able to identify these particles from the published micrographs, but the association with the ER is sug- gestive of microbodies.
Enzymes other than hydrolases may also be localized in cytoplasmic particles. PLESNIOAI~ et al. (1967) found that an apparent mitochondrial peroxidase activity in higher plants was due to contamination of the mitochondrial fraction by a denser particulate fraction. Recently TOL~EI~T et al. (1968) have reported the isolation of "peroxisomes" from green leaves of spinach, wheat and tobacco. The particles, whose microscopic appearance was not reported, contained glyeolate oxidase, glyoxylate reductase and catalase. In view of our own fine-structural evidence for the presence of numerous microbodies in tobacco leaf cells we suggest that these "peroxisomes" are identical to microbodies. These findings suggest the presence in plant cells of an organelle which might be biochemically slmilar to animal microbodies.
With the exception of the reports just discussed, most of the enzymes which have been localized in the particles under consideration are hydrolases. Biochemically this relates these plant particles to animal lysesomes, organelles which contain a variety of hydrolases and presum- ably play a role in various intracellular digestion processes (DE DI~VE and WATTIAITX, 1966). I t has long been suggested that plants have such particles, but as pointed out above, there are not many likely candidates for the role. In the fine-structural literature, the term "plant lysosome" has been applied to bodies clearly identical with microbodies (ScI~NEPF, 1964), as well as to structures whose nature is less certain (SIEVEI~S, 1966, 1967). If it is found that mierobodies do indeed contain hydrolases rather than the oxidases found in animal microbodies, then it may be necessary to decide whether to continue calling them "microbodies" in deference to their structural resemblance to animal microbodies, or to call them "lysosomes" because of their biochemical resemblance to animal lysosomes.
I t is, however, not at all unreasonable to assume that plant micro- bodies will be found to contain a changing enzyme complement that includes both hydrolases and oxidases at one stage or another during cellular differentiation, and that they will therefore prove to be the functional equivalent of both microbodies and lysosomes of animal cells. Such flexibility in enzyme content may be correlated with the general occurrence of microbodies in plant cells in contrast to the sharply limited
17b P lan t a (Berl.), ]3d. 81

250 S.E. FREDERICK, E. H. NEWCO~R, E. L. VIGm and W. P. W~RGI~:
occurrence of their an imal counterparts , and m a y also explain the considerable var ia t ions observed in the mat r ix of microbodies in dif- ferent tissues. There is clearly a pressing need for the biochemical s tudy of fractions carefully moni tored with the electron microscope and correlated with the s tudy of cellular fine structure.
This work was supported in part by Grant GB-6161 from the National Science Foundation. We thank Mr. B~RRu A. PALEWTZ for the micrograph used for Fig. 11.
References ARGOTS, H. J., and K. M. SMITH: Electron microscopy of virus-infected sunflower
leaves. J. Ultrastruet. Res. 19, 173--195 (1967). AVERS, C. J. : Fine structure of Phleum root meristem cells. I. Mitochondria. Amer.
J. Bet. 49, 996--1003 (1962). BALZ, It. P.: Intrazellul/ire Lokalisation und Funktion yon hydrolytischen
Enzymen bei Tabak. Planta (Berl.) 70, 207--236 (1966). BONI'IETT, H. T., and E. H. NEWCOMB : Polyribosomes and cisternal accumulations
in root cells of radish. J. Cell Biol. 27, 423--432 (1965). Bovcx, G. B. : Fine structure and organelle associations in brown algae. J. Cell Biol.
26, 523--537 (1965). - - , and J. C~oNs~w: The fine structure of differentiathlg sieve tube elements.
J. Cell Biol. 25, 79--96 (1965). CRONS~W, J. : Crystal containing bodies of plant cells. Protoplasma (Wien) 59,
318--325 (1964). - - , and G. B. BoucK: The fine structure of differentiating xylem elements. J. Cell
Biol. 24, 4 1 5 ~ 3 1 (1965). D~EMS, W. T.: The fine structure of mouse-liver mierobodies. J. Microscopic 5,
295--304 (1966). DuvE, C. DE, and P. BAUD~UIN: Peroxisomes (Mierobodies and related particles).
Physiol. Rev. 46, 323--357 (1966). - - , and R. W~TT~VX: Functions of lysosomes. Ann. Rev. Physiol. 28, 435~492
(1966). ESSNER, E. : Endoplasmic retieulum and the origin of microbodies in fetal mouse
liver. Lab. Invest. 17, 71--87 (1967). FREY-WYssLING, A., E. GRIESItABER, and K. M~n%ET~L~LER: Origin of spherosomes
in plant cells. J. Ultrastruct. Res. 8, 506---516 (1963). G~A~, P. B. : Histoehemical evidence for the presence of lysosome-like particles
in root meristem cells of Vicla/aba. J. exp. Bet. 16, 350--355 (1965). - - , and A. J. l~IAPnE: The behavior of lysosome-like particles during cell dif-
ferentiation. J. exp. Bet. 17, 151--155 (1966). GEROLA, P.M., and M. BAssI: Sui eristalloidi proteioi delle cellule vegetali.
Caryologia 17, 399--407 (1964). I-IAR~I~OTO~, J. F., and A. M. A L T s c ~ : Lysosome-like behavior in germinating
onion seeds. (Abstr.) Fed. Prec. 22, 475 (1963). HoncoMB, G. E., A. C. HILDEBRANDT, and R. F. EVERT: Staining and acid phos-
phatase reactions of spherosomes in plant tissue culture cells. Amer. J. Bet. 54, 1204---1209 (1967).
HR~BAN, Z., H. SWIFT, and R. W. WISSL~R: Alterations in the fine structure of hepatocytes produced by fl-3-thienylalanine. J. Ultrastruct. Res. 8, 236--250 (1963).
JACKS, T. J., L. Y. YATSU, and A. M. ALTSO]tUL: Isolation and characterization of peanut spherosomes. Plant Physiol. 42, 585--597 (1967).

Fine-Structural Characterization of Plant Microbodies 251
JE~SEN, T. E., and J. G. VALDOVlNOS: Fine structure of abscission zones. I. Abs- cission zones of the pedicels of tobacco and tomato flowers at anthesis. Planta (Berl.) 77, 298--318 (1967).
KOL~n~AI~E~, L., H. ZEc~, and D. vo~ WETTSTEIN: The structure of cells during tobacco mosaic virus reproduction. II. iYIesophyll cells containing virus crystals. J. Ceil Biol. 25, 77--98 (1965).
M~NTON, I. : Observations on phragmosomes. J. exp. Bet. 12, 108--113 (1961). MA~I~os, N. G. : Comments on the nature of a crystal-containing body in plant
cells. Protoplasma (u 60, 31--33 (1965). M~TILE, P., J. P. BALZ, E. SEMADE~XI, and M. JesT: Isolation of spherosomes with
lysosome characteristics from seedlings. Z. Naturforsch. 20b, 693--698 (1965). MOLLENHAVEIr H. H. : Plastic embedding mixtures for use in electron microscopy.
Stain Teclmo]. 39, 111--114 (1964). - - D. J. MORRIS, and A. G. KELLEY : The widespread occurrence of plant cytosomes
resembling animal microbodies. Protoplasma (Wien) 62, 44--52 (1966). NOVIKOFF, A. B., and W. S~I~: The endoplasmic reticulum in the Golgi zone and
its relation to microbodies, Golgi apparatus and autophagic vacuoles in rat liver cells. J. Microscopic 3, 187--206 (1964).
O'BRIEN, T. P., and K. V. T ~ N N : Observations on the fine structure of the oat coleoptile. II. The parenehyma cells of the apex. Protop]asma (Wien) 63, 417--442 (1967).
OLSZEWSKA, 1~. J., et B. GABA~A: ltecherches cytochimiques sur la presence de certaines hydl'olases au cours de la cytocin~se chez les plantes sup6rieures. Proteplasma (Wien) 59, 164--179 (1964).
PETZOLD, H. : Kristalloide Einschliisse in Zytoplasma pflanzlicher Zellen. Proto- plasma (Wicn) 64, 120--133 (1967).
PLES~CA~, M., W. D. BONNER, J~., and B. T. STO]CnY: Peroxidase associated with higher plant mitochondria. Plant Physiol. 42, 366--370 (1967).
Po~TE~, K. 1~., and J. B. CAULFIELD: The formation of the cell plate during cyto- kinesis in Allium cepa L. Prec. IVth Internat. Conf. Electron Microscopy (Berlin) 2 , 5O3--509 (1958).
- - , and R. D. MAC~DO: Studies on the endoplasmic reticulum. IV. Its form and distribution during mitosis in cells of onion root tip. J. biophys, biochem. Cytol. 7, 167--180 (1960).
Povx, N.: Ultrastructural localization of aryl sulfatase activity in plant meriste- marie cells. J. Histochem. Cytoehem. 14, 932--933 (1966).
PRICE, W. C.: Flexulous rods in phloem cells of lime plants infected with citrus tristeza virus. Virology 29, 285--294 (1966).
I~Asc~, E. M., C. K~NJIR~PA~A~BAN, and W. F. MILLINGTON: Histechemical local- ization of acid phosphatases in differentiating and necrotic plant cells. J. Cell :Biol. 27, 142A (1965).
l~on:~N, J. : Correlation of ultrastructural organization and iunction in normal and experimentally changed proximal convoluted tubule cells of the mouse kidney. Stockholm: A. B. Godvie 1954.
SC~NEPF, E.: Zur Cytologic und Physiologic der pflanzlichen Driisen. IV. Licht- und elektrenenmikroskopische Untersuchungen an Septalnektarien. Proto- plasma (Wien) 58, 137--171 (1964).
SE~])ENI, E.G.: Enzymatische Charakterisierung der Lysosomen~quivalente (Spharosomen) yon Maiskeimlingen. Planta (Berl.) 72, 91--118 (1967).
SIEVER8, A. : Lysosomen-Ehnliche Kompartimente in Pflanzenzellen. Naturwissen- schaften 53, 334~335 (1966).
- - E]ektronenmikroskopische Untersuchungen zur geotropischen Reaktion. II. Die polare Organisation des normal wachsenden l~hizoids yon Churn/oetida. Proto- plasma (Wien) 64, 225--253 (1967).

252 S.E. FREDERICK eb al. : Fine-Structural Characterization of Plan~ Mierobodies
SO~OKI~, H. P., and S. So~oKI~: The spherosomes of Campanula persici/olia L. Protoplasma (Wien) 62, 216--236 (1966).
T~o~To~, R. M., and K. V. Tm~AN~: On a crystal-containing body in cells of the oat coleoptile. J. Cell Biol. 20, 345--350 (1964).
TOLBERT, N.E., A. OESER, T. KIS~KI, R.H. HAGEMA~, and R.K. Y~wAzAxI: Pcroxisomes from leaves with enzymes related to glycolate metabolism. (Abstr.) Fed. Proc. 27, 344 (1968).
VILLIERS, T.A.: Crystalloid structure in the microbodies of plant embryo cells. Life Sci. 6, 2151--2156 (1967).
W~LEK-CzER~.C~A, A. : Histochemical demonstration of some hydrolytic enzymes in the spherosomes of plant cells. Acta Soc. Bot. Polon. 34, 573--588 (1965).
WALLES, B.: Plastid structures of carotenoid-deficient mutants of sunflower (Helianthus annuus L.). I. The white mutant. Hereditas (Lurid.) 53, 247--256 (1965).
Professor E. H. NEWCOMB Department of Botany, University of Wisconsin Madison, Wisconsin 53706, USA




















