
Fc Gamma Receptor Glycosylation Modulates the Binding of IgG Glycoforms: A Requirement for Stable...
15
Fc Gamma Receptor Glycosylation Modulates the Binding of IgG Glycoforms: A Requirement for Stable Antibody Interactions Jerrard M. Hayes,* ,†,# Asa Frostell, ‡ Eoin F. J. Cosgrave, § Weston B. Struwe, ∥ Oscar Potter, ⊥ Gavin P. Davey, † Robert Karlsson, ‡ Cecilia Anneren, ‡ and Pauline M. Rudd # † School of Biochemistry & Immunology, Trinity Biomedical Sciences Institute, Trinity College, Pearse St. Dublin 2, Dublin, Ireland ‡ GE Healthcare, Rapsgatan 23, SE-75184 Uppsala, Sweden § Pharmaceutical Life Sciences Group, Waters Corporation, 34 Maple Street, Milford, Massachusetts 01757, United States ∥ Chemistry Research Laboratory, Department of Chemistry, University of Oxford, Oxford OX1 3TA, United Kingdom ⊥ Agilent Technologies, Inc., 5301 Stevens Creek Boulevard, Santa Clara, California 95051, United States # NIBRT-Glycoscience Group, NIBRT-The National Institute for Bioprocessing, Research, and Training, Foster Avenue, Blackrock, County Dublin, Ireland ABSTRACT: FcγRs play a critical role in the immune response following recognition of invading particles and tumor associated antigens by circulating antibodies. In the present study we investigated the role of FcγR glycosylation in the IgG interaction and observed a stabilizing role for receptor N-glycans. We performed a complete glycan analysis of the recombinant FcγRs (FcγRIa, FcγRIIa, FcγRIIb, FcγRIIIa Phe158/Val158 , and FcγRIIIb) expressed in human cells and demonstrate that receptor glycosylation is complex and varied between receptors. We used surface plasmon resonance to establish binding patterns between rituximab and all receptors. Complex binding was observed for FcγRIa and FcγRIIIa. The IgG−FcγR interaction was further investigated using a combination of kinetic experiments and enzymatically deglycosylated Fc γ RIa and FcγRIIIa Phe158/Val158 receptors in an attempt to determine the underlying binding mechanism. We observed that antibody binding levels decreased for deglycosylated receptors, and at the same time, binding kinetics were altered and showed a more rapid approach to steady state, followed by an increase in the antibody dissociation rate. Binding of rituximab to deglycosylated FcγRIIIa Phe158 was now consistent with a 1:1 binding mechanism, while binding of rituximab to FcγRIIIa Val158 remained heterogeneous. Kinetic data support a complex binding mechanism, involving heterogeneity in both antibody and receptor, where fucosylated and afucosylated antibody forms compete in receptor binding and in receptor molecules where heterogeneity in receptor glycosylation plays an important role. The exact nature of receptor glycans involved in IgG binding remains unclear and determination of rate and affinity constants are challenging. Here, the use of more extended competition experiments appear promising and suggest that it may be possible to determine dissociation rate constants for high affinity afucosylated antibodies without the need to purify or express such variants. The data described provide further insight into the complexity of the IgG−FcγR interaction and the influence of FcγR glycosylation. KEYWORDS: Glycosylation, antibody, rituximab, Fcγ receptors, surface plasmon resonance, glycan analysis ■ INTRODUCTION Immunoglobulins are an important class of glycoproteins responsible for recognizing foreign material and tumor associated antigens and communicating this information within the innate and adaptive immune systems. Following opsoniza- tion of target antigens, immunoglobulin G (IgG) coated immune complexes interact with effector cells of the innate immune system through cell surface receptors (FcγR) specific for the Fc region of the antibody molecule. FcγRs are present on platelets and a variety of innate immune cells, including monocytes, granulocytes, macrophages, B-cells, and natural killer cells, where they trigger effector responses such as neutrophil activation, macrophage phagocytosis, natural killer cell ADCC, and B-cell inhibition. 1−5 In general, the receptors can be broadly classified into three distinct families, FcγRI (CD64), FcγRII (CD32), and FcγRIII (CD16). FcγRI members bind IgG with high affinity (K D , 10 −9 M), while the FcγRII and FcγRIII members bind with lower affinity (K D , 10 −6 −10 −7 M). Both high and low affinity FcγRs bind IgG immune complexes with high avidity, but only the high affinity FcγRIa can appreciably bind monomeric IgG. 6 The FcγR family can be further divided into activating (FcγRIa, FcγRIIa, and FcγRIIIa) or inhibitory (FcγRIIb) receptors, based on the immune response following the binding of IgG. Significant heterogeneity also exists within FcγRs, Received: April 24, 2014 Published: October 27, 2014 Article pubs.acs.org/jpr © 2014 American Chemical Society 5471 dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−5485
Transcript of Fc Gamma Receptor Glycosylation Modulates the Binding of IgG Glycoforms: A Requirement for Stable...

Fc Gamma Receptor Glycosylation Modulates the Binding of IgGGlycoforms: A Requirement for Stable Antibody InteractionsJerrard M. Hayes,*,†,# Asa Frostell,‡ Eoin F. J. Cosgrave,§ Weston B. Struwe,∥ Oscar Potter,⊥
Gavin P. Davey,† Robert Karlsson,‡ Cecilia Anneren,‡ and Pauline M. Rudd#
†School of Biochemistry & Immunology, Trinity Biomedical Sciences Institute, Trinity College, Pearse St. Dublin 2, Dublin, Ireland‡GE Healthcare, Rapsgatan 23, SE-75184 Uppsala, Sweden§Pharmaceutical Life Sciences Group, Waters Corporation, 34 Maple Street, Milford, Massachusetts 01757, United States∥Chemistry Research Laboratory, Department of Chemistry, University of Oxford, Oxford OX1 3TA, United Kingdom⊥Agilent Technologies, Inc., 5301 Stevens Creek Boulevard, Santa Clara, California 95051, United States#NIBRT-Glycoscience Group, NIBRT-The National Institute for Bioprocessing, Research, and Training, Foster Avenue, Blackrock,County Dublin, Ireland
ABSTRACT: FcγRs play a critical role in the immune response followingrecognition of invading particles and tumor associated antigens by circulatingantibodies. In the present study we investigated the role of FcγRglycosylation in the IgG interaction and observed a stabilizing role forreceptor N-glycans. We performed a complete glycan analysis of therecombinant FcγRs (FcγRIa, FcγRIIa, FcγRIIb, FcγRIIIaPhe158/Val158, andFcγRIIIb) expressed in human cells and demonstrate that receptorglycosylation is complex and varied between receptors. We used surfaceplasmon resonance to establish binding patterns between rituximab and allreceptors. Complex binding was observed for FcγRIa and FcγRIIIa. TheIgG−FcγR interaction was further investigated using a combination of kineticexper iments and enzymat ica l ly deg lycosy la ted FcγRIa andFcγRIIIaPhe158/Val158 receptors in an attempt to determine the underlyingbinding mechanism. We observed that antibody binding levels decreased for deglycosylated receptors, and at the same time,binding kinetics were altered and showed a more rapid approach to steady state, followed by an increase in the antibodydissociation rate. Binding of rituximab to deglycosylated FcγRIIIaPhe158 was now consistent with a 1:1 binding mechanism, whilebinding of rituximab to FcγRIIIaVal158 remained heterogeneous. Kinetic data support a complex binding mechanism, involvingheterogeneity in both antibody and receptor, where fucosylated and afucosylated antibody forms compete in receptor bindingand in receptor molecules where heterogeneity in receptor glycosylation plays an important role. The exact nature of receptorglycans involved in IgG binding remains unclear and determination of rate and affinity constants are challenging. Here, the use ofmore extended competition experiments appear promising and suggest that it may be possible to determine dissociation rateconstants for high affinity afucosylated antibodies without the need to purify or express such variants. The data described providefurther insight into the complexity of the IgG−FcγR interaction and the influence of FcγR glycosylation.
KEYWORDS: Glycosylation, antibody, rituximab, Fcγ receptors, surface plasmon resonance, glycan analysis
■ INTRODUCTION
Immunoglobulins are an important class of glycoproteinsresponsible for recognizing foreign material and tumorassociated antigens and communicating this information withinthe innate and adaptive immune systems. Following opsoniza-tion of target antigens, immunoglobulin G (IgG) coatedimmune complexes interact with effector cells of the innateimmune system through cell surface receptors (FcγR) specificfor the Fc region of the antibody molecule. FcγRs are presenton platelets and a variety of innate immune cells, includingmonocytes, granulocytes, macrophages, B-cells, and naturalkiller cells, where they trigger effector responses such asneutrophil activation, macrophage phagocytosis, natural killercell ADCC, and B-cell inhibition.1−5 In general, the receptors
can be broadly classified into three distinct families, FcγRI(CD64), FcγRII (CD32), and FcγRIII (CD16). FcγRImembers bind IgG with high affinity (KD, 10
−9 M), while theFcγRII and FcγRIII members bind with lower affinity (KD,10−6−10−7 M). Both high and low affinity FcγRs bind IgGimmune complexes with high avidity, but only the high affinityFcγRIa can appreciably bind monomeric IgG.6
The FcγR family can be further divided into activating(FcγRIa, FcγRIIa, and FcγRIIIa) or inhibitory (FcγRIIb)receptors, based on the immune response following the bindingof IgG. Significant heterogeneity also exists within FcγRs,
Received: April 24, 2014Published: October 27, 2014
Article
pubs.acs.org/jpr
© 2014 American Chemical Society 5471 dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−5485

resulting from polymorphisms in their extracellular domains.Two variants of FcγRIIa exist, which differ in the amino acid inposition 131 (Arg131/His131). The Arg131 substitution hasincreased susceptibility to SLE,7 nephropathy,8 and bacterialinfections.9 Polymorphic variants also exist for FcγRIIIa witheither a phenylalanine or valine residue in position 158 in theextracellular region (Phe158/Val158). Val158 results insignificantly increased affinity for IgG, while the Phe158 variantis linked to autoimmune diseases, including SLE andrheumatoid arthritis (RA).10,11 Polymorphic variants also existfor FcγRIIIb (NA1 and NA2), which are believed todemonstrate further heterogeneity in glycosylation.12
The extensive glycosylation of both antibody and FcγRcontribute to the properties of the IgG−FcγR interaction.Removal of the N-linked glycans at Asn297 in the IgG heavychains causes a loss of structural integrity and decreasedcommunication with the FcγR.13,14 Less is known about therole of FcγR glycosylation; however, recent studies have shownthat elimination of the sugar moiety at Asn162 on FcγRIIIareduces the binding affinity of afucosylated IgG by over anorder of magnitude, presumably through destabilization of theIgG−FcγR interaction.15 In addition, glycosylation at Asn45 onFcγRIIIa negatively affects the binding of IgG.16 These findingsimplicate an active role for glycosylation in positively andnegatively influencing antibody binding and downstreamimmune effector functions. Interestingly, the N-linked glycansof FcγRIIIa have recently been shown to form uniquecarbohydrate−carbohydrate interactions with the N-glycans ofafucosylated IgG, explaining the increased binding and ADCCactivity of antibodies lacking core-fucose.17 Furthermore,glycosylation of FcγRs is cell type specific,18 which adds furtherimportance to how these receptors are glycosylated by cells ofthe immune system as this will likely play a key role inregulating the IgG−FcγR interaction and the downstreamimmunological response.Human FcγRs differentially bind IgG of different subclasses
and a number of studies have been performed, which describethe IgG−FcγR interaction and the selectivity of the receptors. Acomprehensive study by Brunhs et al.6 described the binding ofpolyclonal and monoclonal IgG1, IgG2, IgG3, and IgG4, as wellas therapeutic anti-CD20 and anti-RhD to the complete set ofhuman FcγRs, including polymorphic variants. More recentlyVisser et al. reported affinities of rituximab and a biosimilarantibody to a similar set of FcγRs.19 Other investigations havefocused more on specific receptors.20,21 With varying FcγR andIgG sources and with different surface plasmon resonance(SPR) experimental set-ups, the reported binding affinities varyby up to 10-fold but, with the exception of FcγRI, areconsistently in the micromolar range. The affinity for FcγRI ishigher and consistently in the nanomolar range. In the presentstudy an SPR assay format combining histidine capture ofFcγRs expressed in HEK293 cells and single-cycle kinetics25
was used to investigate rituximab binding across a panel ofreceptors. To complement the interaction analysis and producea more complete picture of both glycosylation and its influenceon IgG binding, a detailed analysis of the glycosylation state ofrituximab and each FcγR (FcγRIa, FcγRIIa, FcγRIIb,FcγRIIIaPhe158/Val158, and FcγRIIIb), including monosaccharidesequence and linkage information was performed.The function of FcγR glycosylation and its role in receptor
stability and interaction with IgG was further investigated byexamining the binding of rituximab to enzymatically deglycosy-lated FcγRIa, FcγRIIIaPhe158/Val158, and fully glycosylated forms.
Following SPR analysis we observed decreased antibodybinding to deglycosylated receptors with reduced affinity. Inaddition, we show altered binding kinetics to deglycosylatedFcγRIIIaPhe158/Val158 with a more rapid approach of steady statebinding levels. We also demonstrate an increase in the antibodydissociation rate from deglycosylated receptors, suggesting astabilizing function for glycosylation on FcγRs and theinteraction with IgG. The data provide further links betweenreceptor glycosylation, antibody binding, and carbohydrate-mediated stabilization of the complex IgG−FcγR interaction.
■ EXPERIMENTAL SECTION
N-linked Glycan Release and 2-AB Labeling
Human Fcγ receptors used for glycosylation analysis and IgGbinding studies were purchased from Sino Biological, Inc.(Beijing, China). Catalogue numbers: FcγRIa, 10256-H08H;FcγRIIa, 10374:H08H; FcγRIIb, 10259-H08H;, FcγRIIIa,10389-H08H; FcγRIIIb, 11046-H08H. Three different batchesof recombinant Fcγ receptor produced in human cells wereused for analysis and replication. Each lyophilized receptor (100μg) was reconstituted in PBS to 0.5 mg/mL, according to themanufacturer’s recommendation. N-linked glycans werereleased and labeled by two different methods. The SDS-PAGE immobilization and release method followed theprotocol described by Royle et al.22 The second glycan releasemethod was performed in solution without immobilization ofthe protein. The reconstituted receptor was incubated with 100mM DTT at 70 °C for 10 min followed by incubation with 10mM iodoacetamide for 45 min in the dark. Glycans werereleased by the addition of PNGase F (0.025 U) (Prozyme,Inc.) and incubation overnight at 37 °C. The following dayprotein was removed by filtration through 10 kDa spin columns(Millipore) and glycans were purified by solid phase extractionusing porous graphitized carbon (PGC) prior to labeling with2-AB and purification using Phytips (Phynexus, Inc.).
Hydrophilic Interaction Ultra Performance LiquidChromatography (HILIC UPLC)
2-AB labeled glycans were prepared as 80% (v/v) acetonitrilesolutions for analysis by HILIC UPLC using a Waters BEH-Glycan 1.7 μm (150 mm × 2.1 mm) column and fluorescencedetection (excitation at 420 nm and emission at 330 nm) aspreviously described.23 All separations were performed using aWaters Acquity H-Class UPLC instrument over a 30 minperiod at 40 °C using 50 mM ammonium formate pH 4.4 assolvent A and 100% (v/v) acetonitrile as solvent B. 2-AB-labeled dextran was used as an internal calibration standard andfor the generation of glucose unit (GU) values. Retention timesfor 2-AB labeled dextran peaks were used to fit a fifth orderpolynomial distribution curve using Waters Empower 3software, which provided the means for converting chromato-graphic retention times into standardized glucose unit (GU)values as previously described.22 Glycans were named accordingto the Oxford notation.24
Weak Anion Exchange HPLC (WAX HPLC)
WAX HPLC was used to analyze the extent of terminalsialylation of receptor N-glycans and for fractionation ofcharged structures. A Waters Biosuite DEAE 10 μm AXC (7.5mm × 75 mm) column was used for separation of chargedcarbohydrate structures. Solvent A was 100 mM ammoniumacetate pH 7 in 20% (v/v) methanol and solvent B was 20%(v/v) acetonitrile. All WAX HPLC separations were performed
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855472

on a Waters Alliance 2695 instrument with online Waters 4795fluorescence detector. Carbohydrate samples were separatedover a 30 min gradient. The relative proportions of terminalcharged glycan structures were determined by comparison toN-linked glycans released from bovine fetuin, which containsneutral, mono-, di-, tri-, and tetra-sialylated structures. ForWAX-fractionation experiments glycan pools were separated byWAX HPLC, and peaks from this analysis were manuallycollected. Glycans were dried, reconstituted in Milli-Q water,and analyzed by HILIC UPLC. This 2-dimensional approachwas used to deconvolute the complex FcγR N-glycan pools. FornanoLC/MS/MS analysis, peaks from HILIC UPLC analysis ofWAX fractionated glycan pools were manually collected, dried,and reconstituted in 10 μL of Milli-Q water.
Exoglycosidase Panel Digests
Exoglycosidase arrays were used to sequence the N-glycans ofhuman FcγRs. Using this technique, sequence, composition,and linkage specificity information was obtained. Fluorescentlylabeled glycans were routinely digested in 50 mM sodiumacetate, pH 5.5, at 37 °C overnight in a volume of 10 μL using aselected panel of exoglycosidase enzymes. The following dayenzymes were removed using 10 kDa spin filters (Pall Corp,NY, USA). Digested 2-AB-labeled glycans were analyzed byHILIC UPLC as described previously. The specificities of theexoglycosidase enzymes used are well-established and specificmonosaccharides were removed as follows: terminal sialic acidin all linkages, α(2,6), α(2,3), and α(2,8), is removed with 1mU/μL Arthrobacter ureafaciens sialidase (ABS)(Prozyme);terminal galactose monosaccharides were removed using 0.5mU/μL bovine testes β-galactosidase (BTG)(Prozyme), whichreleases both β(1,3)- and β(1,4)-linked galactose; terminal N-acetylglucosamine (GlcNAc) monosaccharides in β(1,4) link-age were released with 40 mU/μL Streptococcus pneumoniaehexosaminidase (GUH)(Prozyme); core α(1,6)-fucose wasselectively removed using 1 mU/μL bovine kidney α-fucosidase(BKF)(Prozyme); terminal nonreducing end fucose in α(1−3)-and α(1−4)-linkages was removed using 0.004 mU/μL almondmeal α-fucosidase (AMF)(Prozyme); α-linked mannose wasremoved using 150 mU/μL jack bean α-mannosidase (JBM)-(Prozyme); and βGalNAc and βGlcNAc were removed using50 mU/μL jack bean β-N-acetylhexosaminidase (JBH)-(Prozyme).
Mass Spectrometry of N-Glycans
LC/FLD-MS. Glycans were analyzed using a Waters Xevo G2QTof coupled to an Acquity UPLC with a Waters BEH Glycancolumn (1.0 × 150 mm, 1.7 μm particle size). The analyzer wasset to sensitivity mode, and data was acquired in negative modewith the following conditions: 2 kV capillary voltage, 25 V conevoltage, 4 V extraction voltage, 100 °C source temperature, 280°C desolvation temperature, 10L/h cone gas flow, and 600L/hdesolvation gas flow. Solvent A was 50 mM ammonium formatepH 4.4 , and solvent B was 100% acetonitrile. The flow rate was30−47% buffer A for 24 min, 70% A for 3 min, and 30% A for 3min. Injection volume was 10 μL, and samples were diluted in70% acetonitrile prior to analysis. Data acquisition andprocessing were conducted with Waters MassLynx version 4.1.NanoLC/MS/MS. LC-MS(2) analysis of 2-AB labeled
glycans from fractionated samples was performed using anAgilent Technologies 1200 series capillary/nano liquidchromatography system in conjunction with an AgilentChipCube interface and a 6340 ion trap mass spectrometer(Agilent Technologies, Santa Clara, CA). The instrument was
operated in positive ion mode with a spray voltage of −1.8 kV.Samples were run on a custom HPLC-chip, which consisted ofa 500 nL porous graphitized carbon enrichment column and a43 mm separation column packed with Tosoh TSK Gel Amide80 (3 μm). Data-dependent acquisition was used to automati-cally target [M + 2H]2+ or [M + H + NH4]
2+ ions of the labeledglycans for isolation in the ion trap and fragmented therein bycollision induced dissociation to yield MS2 spectra.
Surface Plasmon Resonance (SPR)
All IgG−FcγR interaction experiments were performed using aBiacore T200 system (GE Healthcare, Uppsala, Sweden) withanalysis temperature set to 25 °C and sample compartmenttemperature set to 15 °C. Recombinant extracellular domainsof FcγRs expressed with C-terminal His-tags in human/HEK293 cells were from Sino Biological, Inc.: FcγRI(NP_000557.1, Met 1-Pro 288), FcγRIIA (AAA35827, Met1-Ile 218), FcγRIIb (NP_001002274.1, Ala 46-Pro 217),FcγRIIIa F158 (P08637-1, Met 1-Gln 208), FcγRIIIa V158(P08637-1, Met 1-Gln 208), and FcγRIIIb (NP_000561.3, Met1-Ser 200). FcγRs were aliquoted, stored at −70 °C, andthawed only once. Overlayed sensorgrams of rituximab bindingto two different lots and to fresh and 4 month old aliquotsshowed no significant differences in sensorgram appearancesindicating similar binding kinetics (not shown). Rituximab(MabThera) was obtained from pharmacies and storedaccording to the manufacturer’s instructions. Anti-His antibody(from His Capture Kit, GE Healthcare) was amine coupled inthe active and reference flow cell of a CM5 chip, according tothe manufacturer’s instructions. Briefly, the surface wasactivated by injecting a solution containing 0.2 M N-ethyl-N¢-dimethylaminopropylcarbodiimide (EDC) and 50 mM N-hydroxysuccinimide (NHS) for 7 min. Anti-His antibody wasdiluted to 20 μg/mL in 10 mM Na-acetate, pH 4.5, and injectedduring 7 min, and the surface was then blocked by injecting 1M ethanolamine at pH 8.5 for 7 min. Immobilization levels inthe range 6000−8000 RU were used, each run having similarlevels in active and reference flow cell. His-tagged FcγRs at aconcentration of 0.5−1 μg/mL were injected for 60 s using aflow rate of 5 μL/min in active flow cell only, obtaining capturelevels in the range 70−180 RU for kinetic data andapproximately 300 RU for affinity data. A capture stabilizationtime of 1−3 min was applied for some FcγRs. For affinity andkinetic studies rituximab was then injected in order ofincreasing concentration over reference and active flow cellusing five 60 s injections at 30 μL/min, applying a single cyclekinetics procedure.25 Using this procedure all concentrationswere injected in sequence in order from low to highconcentration and a 300 s dissociation time was added afterthe last rituximab injection. Rituximab concentrations in therange 1.2−300 nM were used for FcγRI, 24.7−2000 nM forFcγRIIIa, and 0.5−8 μM for FcγRIIa, FcγRIIb, and FcγRIIIb.Following each experiment both flow cells were regeneratedusing a 30 s injection of 10 mM glycine, pH 1.5. Blank cycles(FcγR capture + buffer injections + regeneration) wereperformed first, last, and upon change of FcγR subtype. Datawere double referenced by first subtraction of reference flowcell and then subtraction of blank cycles. Fitting of data wereperformed using Biacore T200 evaluation software 2.0.
Enzymatic Deglycosylation of FcγRIa andFcγRIIIaPhe158/Val158Fifty micrograms of HEK293 FcγRIa and FcγRIIIaPhe158/Val158were reconstituted in NaHCO3 (100 mM), pH 7, to a
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855473

concentration of 1 mg/mL. For deglycosylation experiments, 5μL PNGase F (2.5 mU/μL) was added, and the volume wasincreased to 100 μL and incubated at 37 °C for 5−6 h. Forcontrol deglycosylation experiments sodium bicarbonate bufferwas used in place of PNGase F. Following PNGase Ftreatment, 1 μL of Endo F1 (17 U/ml)(QA Bio, Inc.) and 1μL of Endo F2 (5 U/ml)(QA Bio, Inc.) were added followedby 5 μL of 250 mM sodium acetate, pH 4.5, and overnightincubation at 37 °C. Five microliters of 250 mM sodiumacetate, pH 4.5, was also added to the control samples in placeof Endo F1 and Endo F2. The next morning deglycosylated andcontrol receptors were purified using 10 kDa spin filters andconcentrated to 1 mg/mL. The extent of receptor deglycosy-lation was determined by SDS-PAGE according to Laemmli26
and by collecting the endoglycosidase released glycans andlabeling with 2-AB followed by HILIC UPLC analysis.
■ RESULTS
Glycosylation of Human Fcγ Receptors Is Complex andDifferential
Glycosylation of human Fcγ receptors expressed in HEK293cells was routinely analyzed by hydrophilic interaction ultra-performance liquid chromatography (HILIC UPLC) followingPNGase F release of N-glycans and fluorescent labeling. Initialg l y c an ana l y s i s o f F c γRIa , F c γRI I a , F c γRI Ib ,FcγRIIIaPhe158/Val158, and FcγRIIIb revealed complex glycosyla-tion with many overlapping peaks (see Figure 1A).
Figure 1. Human FcγRs display complex and differential glycosylation. A. Glycan analysis of Fcγ receptors expressed in Human/HEK293 cells.Glycosylation of receptors was complex and differential with 30−40 unique glycan structures for each receptor. Following enzymatic release andHILIC UPLC analysis common structures (30) were identified and are indicated by dashed vertical lines. Numbered peaks represent the structuresshown in Table 1: (i) FcγRIIIa, (ii) FcγRIa, (iii) FcγRIIa, (iv) FcγRIIb, and (v) FcγRIIIb. GU values were assigned using internal 2-AB labeleddextran standards and integration using Waters Empower 3 software. Glycan structures and relative abundane for each receptor are shown in Table1. Glycan release and analysis experiments were performed in triplicate. B. Weak anion exchange (WAX) HPLC analysis of human FcγRs shows N-glycans are neutral (S0) and monosialylated structures (S1) with minor amounts of disialylated sugars (S2). For WAX fractionation experimentsindividual S0 and S1 peaks were manually collected and dried for nano-LC-MS/MS analysis: (i) Bovine fetuin N-glycan standards, (ii) FcγRIa, (iii)FcγRIIa, (iv) FcγRIIb, (v) FcγRIIIa, (v) FcγRIIIb. WAX HPLC experiments were performed in triplicate. C. FcγRs contain common N-glycans withdifferences in relative abundance. Average peak area percentages were calculated for peaks from individual receptors (FcγRIa, FcγRIIa, FcγRIIb,FcγRIIIa, and FcγRIIIb). Thirty common peaks were identified (see panel A) and values were plotted as the average GU value and average % areafrom three different releases of individual receptors. This value was then used to calculate the average value across the range of receptors for eachpeak to show the distribution in relative abundance between different receptors. Error bars represent the standard deviation for the relative % areas.Large error bars as seen for certain peaks, e.g., GU 6.22 (Man5), signify large differences in abundance for certain N-glycans between receptors. N-glycan structures corresponding to the GU values shown can be seen in Table 1.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855474
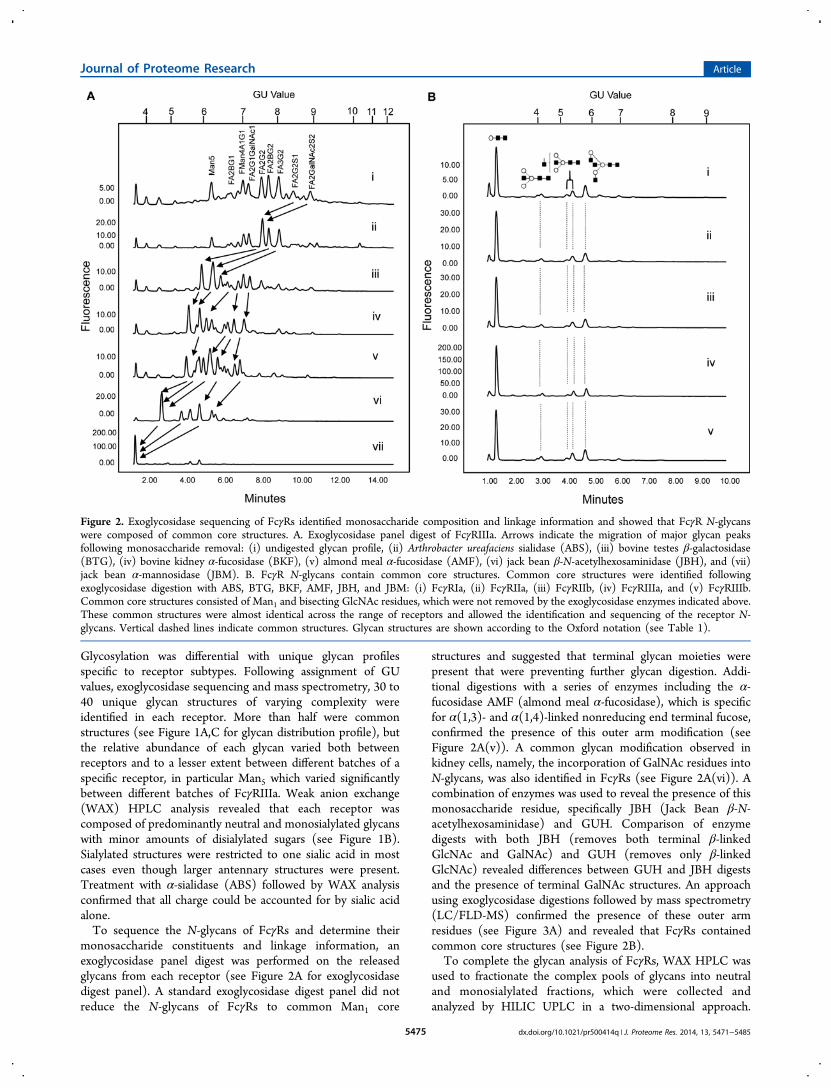
Glycosylation was differential with unique glycan profilesspecific to receptor subtypes. Following assignment of GUvalues, exoglycosidase sequencing and mass spectrometry, 30 to40 unique glycan structures of varying complexity wereidentified in each receptor. More than half were commonstructures (see Figure 1A,C for glycan distribution profile), butthe relative abundance of each glycan varied both betweenreceptors and to a lesser extent between different batches of aspecific receptor, in particular Man5 which varied significantlybetween different batches of FcγRIIIa. Weak anion exchange(WAX) HPLC analysis revealed that each receptor wascomposed of predominantly neutral and monosialylated glycanswith minor amounts of disialylated sugars (see Figure 1B).Sialylated structures were restricted to one sialic acid in mostcases even though larger antennary structures were present.Treatment with α-sialidase (ABS) followed by WAX analysisconfirmed that all charge could be accounted for by sialic acidalone.To sequence the N-glycans of FcγRs and determine their
monosaccharide constituents and linkage information, anexoglycosidase panel digest was performed on the releasedglycans from each receptor (see Figure 2A for exoglycosidasedigest panel). A standard exoglycosidase digest panel did notreduce the N-glycans of FcγRs to common Man1 core
structures and suggested that terminal glycan moieties werepresent that were preventing further glycan digestion. Addi-tional digestions with a series of enzymes including the α-fucosidase AMF (almond meal α-fucosidase), which is specificfor α(1,3)- and α(1,4)-linked nonreducing end terminal fucose,confirmed the presence of this outer arm modification (seeFigure 2A(v)). A common glycan modification observed inkidney cells, namely, the incorporation of GalNAc residues intoN-glycans, was also identified in FcγRs (see Figure 2A(vi)). Acombination of enzymes was used to reveal the presence of thismonosaccharide residue, specifically JBH (Jack Bean β-N-acetylhexosaminidase) and GUH. Comparison of enzymedigests with both JBH (removes both terminal β-linkedGlcNAc and GalNAc) and GUH (removes only β-linkedGlcNAc) revealed differences between GUH and JBH digestsand the presence of terminal GalNAc structures. An approachusing exoglycosidase digestions followed by mass spectrometry(LC/FLD-MS) confirmed the presence of these outer armresidues (see Figure 3A) and revealed that FcγRs containedcommon core structures (see Figure 2B).To complete the glycan analysis of FcγRs, WAX HPLC was
used to fractionate the complex pools of glycans into neutraland monosialylated fractions, which were collected andanalyzed by HILIC UPLC in a two-dimensional approach.
Figure 2. Exoglycosidase sequencing of FcγRs identified monosaccharide composition and linkage information and showed that FcγR N-glycanswere composed of common core structures. A. Exoglycosidase panel digest of FcγRIIIa. Arrows indicate the migration of major glycan peaksfollowing monosaccharide removal: (i) undigested glycan profile, (ii) Arthrobacter ureafaciens sialidase (ABS), (iii) bovine testes β-galactosidase(BTG), (iv) bovine kidney α-fucosidase (BKF), (v) almond meal α-fucosidase (AMF), (vi) jack bean β-N-acetylhexosaminidase (JBH), and (vii)jack bean α-mannosidase (JBM). B. FcγR N-glycans contain common core structures. Common core structures were identified followingexoglycosidase digestion with ABS, BTG, BKF, AMF, JBH, and JBM: (i) FcγRIa, (ii) FcγRIIa, (iii) FcγRIIb, (iv) FcγRIIIa, and (v) FcγRIIIb.Common core structures consisted of Man1 and bisecting GlcNAc residues, which were not removed by the exoglycosidase enzymes indicated above.These common structures were almost identical across the range of receptors and allowed the identification and sequencing of the receptor N-glycans. Vertical dashed lines indicate common structures. Glycan structures are shown according to the Oxford notation (see Table 1).
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855475

Individual peaks were collected from the HILIC analysis ofneutral and monosialylated fractions and dried for massspectrometry (nanoLC-MS/MS) analysis. Thirty of the mostabundant peaks were collected and mass values andfragmentation data generated for 22 of these structures (seeFigure 3B for nanoLC-MS/MS data). This approach identifiedapproximately 83.5% of the total glycan composition (see Table1 for N-glycans identified from FcγRs). In summary, themajority of N-glycans from FcγRs were complex (70%) withhigh mannose (9%) and hybrids (3%) also present. Corefucosylation was present on 67% of glycans; galactosylation waspresent as mainly mono- and digalactosylated structures withonly a minority being capped with sialic acids. Approximately66% of N-glycans contained no sialic acids with only 12%having one sialic acid and 4% having two sialic acids. Negligibleamounts of higher sialylated structures (tri-sialylated and tetra-sialylated) were found. LacdiNAc (HexNAc-HexNac) struc-
tures containing GalNAc β-linked to antennary GlcNAc werepresent in approximately 19%, outer-arm fucose in approx-imately 10%, and bisecting GlcNAc in 13.5%. Bisecting GlcNAcwas identified based on exoglycosidase digestion and confirmedby mass spectrometry (see Figures 2B and 3). This structurewas shown to persist following full panel digests as there is noknown exoglycosidase that will selectively remove thismonosaccharide. All sialic acids were in α(2,3) linkage togalactose, and all galactose was found in β(1,4) linkages toGlcNAc. The presence of poly-N-lactosamine residues was alsofound in all FcγRs accounting for approximately 2%. It is alsoworth noting that glycosylation of FcγRIIIaPhe158 was found tobe identical to FcγRIIIaVal158, which could be expected as thereis only a single amino acid difference. However, this amino acid(158) is close to the asparagine residue in position 162, whichis known to be glycosylated and important for the IgGinteraction.15,27 The 2-AB labeled N-glycans of rituximab were
Figure 3. Mass spectrometry analysis of FcγR N-glycans identified unusual and complex glycan epitopes. A. Exoglycosidase digestions of FcγRIIIafollowed by mass spectrometry (FLD/LC-MS) reduced the complexity of N-glycans and confirmed the presence of bisecting GlcNAc and GalNAcstructures. Released 2-AB labeled FcγRIIIa N-glycans were digested with ABS, BTG, BKF, AMF, and GUH followed by mass spectrometry, whichidentified common glycan structures consisting of oligomannose, bisecting GlcNAc and GalNAc structures. The top chromatogram represents thefluorescence HILIC profile with the corresponding extracted ion chromatograms of each mass and glycan composition shown below. FLD:Fluorescence detection of N-glycans separated by HILIC UPLC. EIC: Extracted ion chromatograms. B. NanoLC/MS-MS analysis of FcγR N-glycans. Fragmentation and identification of four N-glycan structures were chosen for illustrative purposes to show some of the more complex andunusual carbohydrate structures. The parent m/z shown in the inset spectra on the right-hand side corresponda to [M + NH4 + H]2+ or [M + 2H]2+
ions of glycans present in the respective fractions, and these were used to determine the likely monosaccharide compositions of the N-glycans. Thelarger spectra show MS2 for those parent m/z. These spectra are annotated with cartoons for the most likely B ion structures based on the knowledgethat CID spectra for N-glycans are dominated by B ions resulting from cleavage on the reducing-end side of HexNAc groups. In combination, thesedata were used to deduce likely structures for the glycans. When interpreting this data it is important to be aware that outer-arm fucose residues maymigrate from one position to another during ionization and fragmentation.41 A. F(6)Man4A1F(2)GalNAc1 (peak 16 in Figures 1 and 2 and Table1); B. F(6)A2BG(4)2 (peak 17 in Figures 1 and 2 and Table 1); C. F(6)A2F(2)1G(4)1GalNAc1 (peak 20 in Figures 1 and 2 and Table 1); D.F(6)A2F2(2)G(4)1GalNAc1 (peak 25 in Figures 1 and 2 and Table 1).
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855476

Table 1. N-Glycans of HEK293 FcγRsa
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855477

Table 1. continued
aGlycans were identified and quantified using a combination of HILIC UPLC, exoglycosidase digestions, WAX HPLC, WAX fractionation, and massspectrometry (MS/MS). Average GU (glucose unit) values and % areas are the mean of three individual glycan releases. Peak numbers correspond
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855478

also fully characterized by both HILIC UPLC and by massspectrometry. Rituximab contained five major glycans: A2(3.5%), FA2 (45%), FA2[6]G1 (26.3%), FA2[3]G1 (19.3%),and FA2G2 (1.6%) representing 95% of the total N-glycancomposition.
Kinetic Analysis of Rituximab Binding to FcγRs
Rituximab is a well-characterized monoclonal antibody used fora number of clinical indications including non-Hodgkin’slymphoma, chronic lymphocytic leukemia (CLL), andrheumatoid arthritis (RA). Rituximab was used for all
interaction studies and was heterogeneous with respect tocore fucosylation with approximately 92% of glycans containingthis residue, as determined by glycan analysis. As a result, bothinteraction partners, FcγRs and antibody, display structuralheterogeneity. Binding studies using SPR and histidine-taggedFcγRs captured to an immobilized antihistidine antibody withtarget antibodies injected applying a single cycle kineticsprocedure were performed according to Karlsson et al.25 Inbrief, single cycle kinetics is a further development of the moretraditionally used multicycle kinetics. In the setup of atraditional kinetics assay one antibody concentration is injected
Table 1. continued
to peaks shown in Figure 1. m/z values are for [M + 2H]2+ unless otherwise indicated. Glycans are named according to the Oxford notation asillustrated in the legend below the table. Where linkage information was determined by exoglycosidase sequencing, it is indicated in the glycan name,whereas when the linkage information is unsure, such as for the hybrid structures 10, 13, and 16, it has been omitted from the glycan name and thecartoon figure is a representation with structural composition only. For structures 10, 13, and 16, the mannose on the α(1,6) arm is shown linked tothe mannose in the α(1,6) position for representative purposes only as Jack bean mannosidase is known to remove mannose linked in the α(1,6) andα(1,2) positions more efficiently than α(1,3).42,43
Figure 4. Rituximab binds to FcγRs with different kinetics and exhibits complex binding. SPR sensorgrams of rituximab-FcγR binding kinetics areshown: A. FcγRIa, B. FcγRIIIaVal158, C. FcγRIIIaPhe158, D. FcγRIIa, E. FcγRIIb, and F. FcγRIIIb. In A−F red solid lines represent experimental data; inA−C black dashed lines display one to one fittings. In D−F the steady state affinity fit of the sensorgram data are shown in the inset with the verticalline marking the respective obtained KD values: FcγRIIa 4.2 μM, FcγRIIb 6.9 μM, and FcγRIIIb 3.5 μM. A−C show representative sensorgrams from>10 experiments; D−F show representative sensorgrams from three separate affinity determinations. Five injections of rituximab in increasingconcentration are performed in each curve, resulting in increasing binding level. Dissociation is seen starting from the end of injection of the highestconcentration, at time 550 s, as well as from the end of injection of the lower concentrations.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855479
per interaction cycle, whereas in single cycle kinetics allconcentrations are injected in sequence in the same cycle.Binding of rituximab to the complete set of FcγRs is
presented in Figure 4. Binding profiles varied considerablydepending on the receptor, ranging from stable binding toFcγRIa to rapid dissociation from FcγRIIa, FcγIIb, andFcγRIIIb receptors. For FcγRIIIaPhe158/Val158 dissociation wasbiphasic with rapid initial dissociation followed by slowerdissociation in the late phase. This was more evident for
FcγRIIIaVal158. Data analysis (dotted lines in Figure 4)confirmed this heterogeneity, as antibody binding toFcγRIIIaPhe158/Val158 did not fit well to a one to one bindingmodel. The interaction with FcγRIa also did not adhere to aone to one binding model, but here deviations were significantacross all antibody concentrations. For both FcγRIa andFcγRIIIaVal158 the observed binding at high concentrationsexceeded the binding level predicted by the fitting model. Byfitting data to more complex interaction models, based on
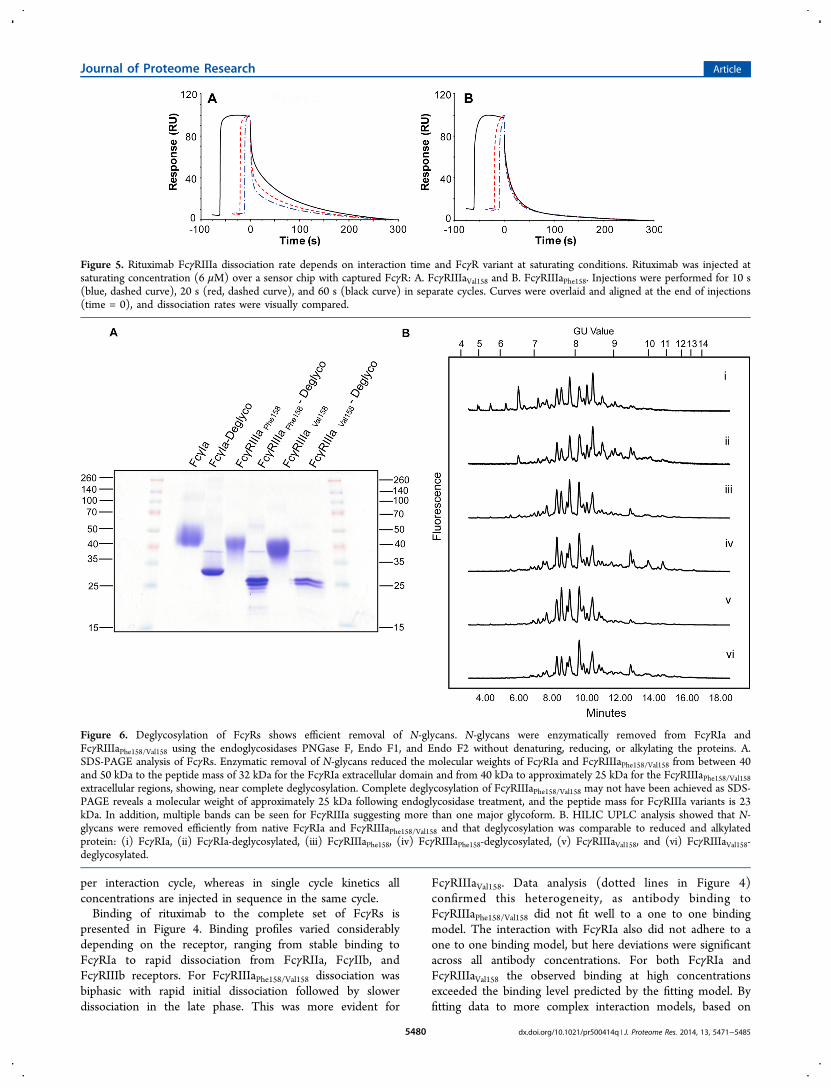
Figure 5. Rituximab FcγRIIIa dissociation rate depends on interaction time and FcγR variant at saturating conditions. Rituximab was injected atsaturating concentration (6 μM) over a sensor chip with captured FcγR: A. FcγRIIIaVal158 and B. FcγRIIIaPhe158. Injections were performed for 10 s(blue, dashed curve), 20 s (red, dashed curve), and 60 s (black curve) in separate cycles. Curves were overlaid and aligned at the end of injections(time = 0), and dissociation rates were visually compared.
Figure 6. Deglycosylation of FcγRs shows efficient removal of N-glycans. N-glycans were enzymatically removed from FcγRIa andFcγRIIIaPhe158/Val158 using the endoglycosidases PNGase F, Endo F1, and Endo F2 without denaturing, reducing, or alkylating the proteins. A.SDS-PAGE analysis of FcγRs. Enzymatic removal of N-glycans reduced the molecular weights of FcγRIa and FcγRIIIaPhe158/Val158 from between 40and 50 kDa to the peptide mass of 32 kDa for the FcγRIa extracellular domain and from 40 kDa to approximately 25 kDa for the FcγRIIIaPhe158/Val158extracellular regions, showing, near complete deglycosylation. Complete deglycosylation of FcγRIIIaPhe158/Val158 may not have been achieved as SDS-PAGE reveals a molecular weight of approximately 25 kDa following endoglycosidase treatment, and the peptide mass for FcγRIIIa variants is 23kDa. In addition, multiple bands can be seen for FcγRIIIa suggesting more than one major glycoform. B. HILIC UPLC analysis showed that N-glycans were removed efficiently from native FcγRIa and FcγRIIIaPhe158/Val158 and that deglycosylation was comparable to reduced and alkylatedprotein: (i) FcγRIa, (ii) FcγRIa-deglycosylated, (iii) FcγRIIIaPhe158, (iv) FcγRIIIaPhe158-deglycosylated, (v) FcγRIIIaVal158, and (vi) FcγRIIIaVal158-deglycosylated.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855480

receptor or antibody heterogeneity or conformational change,fittings were significantly improved, but no definite conclusioncould be made as to the precise binding mechanism. However,this is not surprising, as we would expect the binding to becomplex and differentially modulated by the presence ofdifferently glycosylated variants. For this reason, kinetic andaffinity constants for FcγRIa and FcγRIIIaPhe158/Val158 are notreported, as the definition of these constants depend on thebinding mechanism. Thus, even if steady state was reachedduring injection (as was not the case here) fitting equilibriumdata to a one to one binding model is not appropriate.The binding behavior of rituximab to FcγRIIIaPhe158/Val158 was
further investigated by injecting saturating concentrations (6μM) of the antibody at different injection times. Binding datawere aligned at the end of the injection for analysis. As shownin Figure 5 rituximab dissociated from FcγRIIIaVal158 in aninjection time-dependent manner: the longer the injection timethe slower the off-rate. In contrast, rituximab dissociation fromFcγRIIIaPhe158 was not as clearly affected by the injection time,even though a very minor tendency similar to FcγRIIIaVal158may be seen. The data for FcγRIIIaVal158 indicates either thatthe antibody−receptor complex is stabilized over time due toconformational change or that a variant of the antibody withinherent slower dissociation accumulates on the sensor surfaceover time. Injection time-dependent dissociation was alsoobserved with infliximab and omalizumab (not shown) and wastherefore not unique to rituximab.For FcγRIIa, FcγRIIb, and FcγRIIIb, the shapes of the
binding curves obtained were very similar, as shown in Figure4D−F. For these low affinity receptors sensorgrams werecharacterized by rapid approach to an equilibrium phase(steady state) followed by rapid dissociation. Rituximab affinitydata from three separate determinations was fitted to a steadystate model generating KD values for FcγRIIa of 4.2 μM (±0.95), FcγRIIb 6.9 μM (± 0.67), and FcγRIIIb 3.5 (± 0.15)μM, respectively.
Deglycosylation of FcγRs Influences Binding andInteraction Kinetics of Rituximab
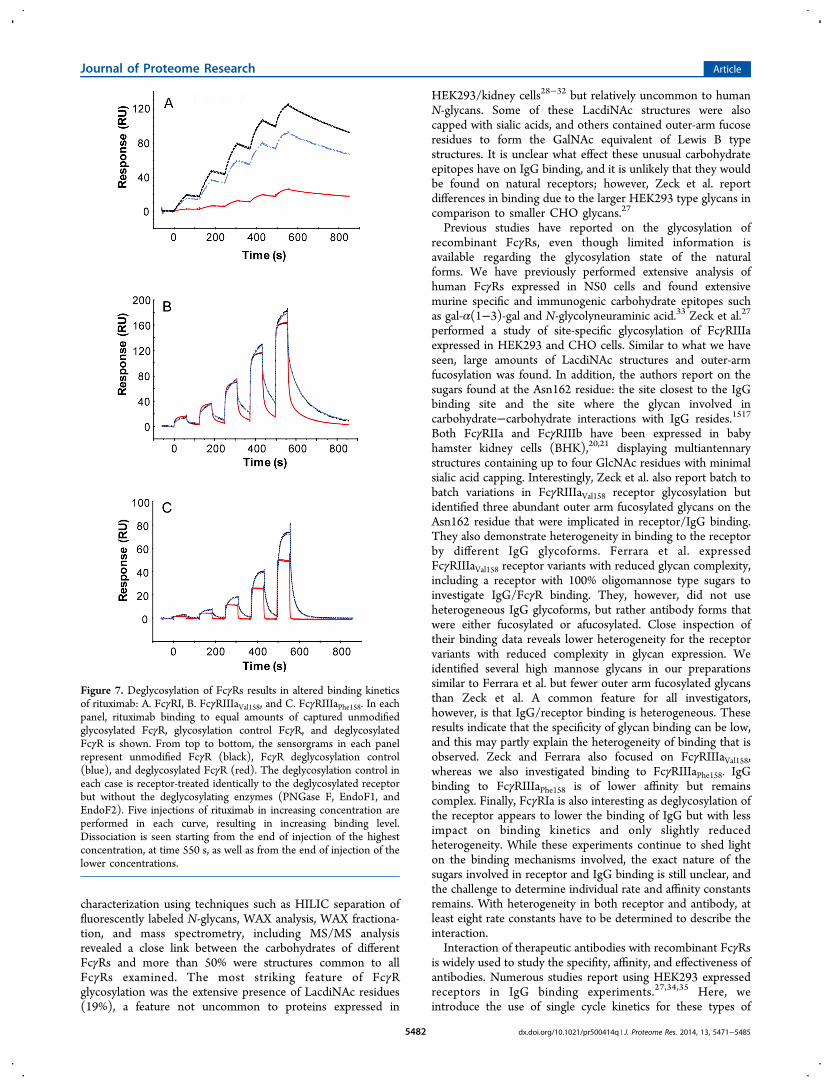
The high affinity FcγRIa and low affinity FcγRIIIaPhe158/Val158were enzymatically deglycosylated without denaturing, reduc-ing, or alkylating the proteins using a combination of theendoglycosidases: PNGase F, Endo F1, and Endo F2.Appropriate deglycosylation controls were included such asFcγRIa and FcγRIIIaPhe158/Val158 treated identically to theirdeglycosylated forms but without the endoglycosidase enzymes.The extent of receptor deglycosylation was monitored by SDS-PAGE analysis of deglycosylated receptors and by labeling theendoglycosidase released receptor N-glycans with 2-ABfollowed by HILIC UPLC analysis. SDS-PAGE showed thatendoglycosidase treatment reduced the molecular weights ofFcγRIa and FcγRIIIaPhe158/Val158 from between 40 and 50 kDa tothe peptide mass of 32 kDa for the FcγRIa extracellular domainand from 40 kDa to approximately 25 kDa for theFcγRIIIaPhe158/Val158 extracellular regions, showing, near com-plete deglycosylation (see Figure 6A). Complete deglycosyla-tion of FcγRIIIaPhe158/Val158 may not have been achieved as SDS-PAGE reveals a molecular weight of approximately 25 kDafollowing endoglycosidase treatment and the peptide mass forFcγRIIIa variants is 23 kDa. In addition, multiple bands can beseen for FcγRIIIa suggesting more than one major glycoform(see Figure 6A). HILIC UPLC analysis also showed that N-glycans were removed efficiently from native FcγRIa and
FcγRIIIaPhe158/Val158 and that deglycosylation was comparable toreduced and alkylated protein (see Figure 6B).Receptors deglycosylated using this approach allowed
subsequent IgG interaction studies to be performed and therole of receptor glycosylation in stability and the binding eventto be assessed. FcγRIa and FcγRIIIaPhe158/Val158 were captured tothe anti-His antibody on the SPR sensor chip, and the bindingof rituximab to deglycosylated receptors was compared withbinding to equal amounts of unmodified and control treatedreceptors (Figure 7). When injecting similar concentrations ofglycosylated and deglycosylated receptors, respectively, over theanti-His antibody, similar levels of receptor were captured,indicating that the His tag was available to the same extent.Furthermore, the binding curves during capture and the capturestability of glycosylated and deglycosylated receptors werecomparable (not shown), indicating similarities between thepreparations and suggesting that deglycosylation had notseriously affected the stability of the proteins, e.g., aggregatedreceptor would display a different curve shape. In the case ofFcγRIa a more than 50% decrease in binding level todeglycosylated receptor was seen, compared to unmodifiedFcγR mainly due to visibly slower association rate (see Figure7A). Binding of rituximab to control treated FcγRIa was alsodecreased, but to a lesser extent. However, binding of rituximabto control treated and unmodified FcγRIIIaPhe158/Val158 wascomparable with overlapping binding curves (Figure 7B,C),indicating that the deglycosylation process itself (withoutenzyme) did not significantly affect binding. Binding levelsobtained for FcγRIIIaVal158 (Figure 7B) were higher than thoseobtained with FcγRIIIaPhe158 (Figure 7C), in agreement withthe documented higher affinity for FcγRIIIaVal158.Interestingly, significant changes in antibody binding to
deglycosylated receptors were observed. Both deglycosylatedFcγRIIIaPhe158 and FcγRIIIaVal158 affinity for rituximab decreasedas shown by decreased binding levels compared to controls.Focusing first on the interaction between antibody andFcγRIIIaPhe158 (Figure 7C), antibody binding to glycosylatedreceptor displayed a biphasic interaction with steep associationin the early association phase and slower in the late associationphase. A biphasic profile is also seen during dissociation, withinitially fast dissociation followed by slower dissociation. Incontrast, binding to deglycosylated receptor displayed a moresimple, monophasic interaction rapidly reaching equilibrium,remaining at equilibrium during the injection and then rapiddissociation back down to baseline. The binding todeglycosylated receptor allowed fitting to the steady statemodel in a similar manner as for FcγRIIa, FcγRIIb, andFcγRIIIb (Figure 4), obtaining an estimated KD of 1.7 μM.Similar behavior was also seen with the interaction betweenantibody and glycosylated/deglycosylated FcγRIIIaVal158 (Figure7B). The difference being that the binding to deglycosylatedreceptor still showed some remaining heterogeneity (biphasicassociation and dissociation), although data almost reachedequilibrium and dissociation was faster than that from theglycosylated receptor.
■ DISCUSSIONThe binding of IgG antibodies with FcγRs is a complexinteraction, further complicated by the significant heterogeneitythat exists in both the antibody and the receptor due to thepresence of differently glycosylated species. The interaction ofrituximab with glyco-analyzed FcγRs was studied to analyze theinfluence of FcγR glycosylation on antibody binding. Structural
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855481
characterization using techniques such as HILIC separation offluorescently labeled N-glycans, WAX analysis, WAX fractiona-tion, and mass spectrometry, including MS/MS analysisrevealed a close link between the carbohydrates of differentFcγRs and more than 50% were structures common to allFcγRs examined. The most striking feature of FcγRglycosylation was the extensive presence of LacdiNAc residues(19%), a feature not uncommon to proteins expressed in
HEK293/kidney cells28−32 but relatively uncommon to humanN-glycans. Some of these LacdiNAc structures were alsocapped with sialic acids, and others contained outer-arm fucoseresidues to form the GalNAc equivalent of Lewis B typestructures. It is unclear what effect these unusual carbohydrateepitopes have on IgG binding, and it is unlikely that they wouldbe found on natural receptors; however, Zeck et al. reportdifferences in binding due to the larger HEK293 type glycans incomparison to smaller CHO glycans.27
Previous studies have reported on the glycosylation ofrecombinant FcγRs, even though limited information isavailable regarding the glycosylation state of the naturalforms. We have previously performed extensive analysis ofhuman FcγRs expressed in NS0 cells and found extensivemurine specific and immunogenic carbohydrate epitopes suchas gal-α(1−3)-gal and N-glycolyneuraminic acid.33 Zeck et al.27
performed a study of site-specific glycosylation of FcγRIIIaexpressed in HEK293 and CHO cells. Similar to what we haveseen, large amounts of LacdiNAc structures and outer-armfucosylation was found. In addition, the authors report on thesugars found at the Asn162 residue: the site closest to the IgGbinding site and the site where the glycan involved incarbohydrate−carbohydrate interactions with IgG resides.1517
Both FcγRIIa and FcγRIIIb have been expressed in babyhamster kidney cells (BHK),20,21 displaying multiantennarystructures containing up to four GlcNAc residues with minimalsialic acid capping. Interestingly, Zeck et al. also report batch tobatch variations in FcγRIIIaVal158 receptor glycosylation butidentified three abundant outer arm fucosylated glycans on theAsn162 residue that were implicated in receptor/IgG binding.They also demonstrate heterogeneity in binding to the receptorby different IgG glycoforms. Ferrara et al. expressedFcγRIIIaVal158 receptor variants with reduced glycan complexity,including a receptor with 100% oligomannose type sugars toinvestigate IgG/FcγR binding. They, however, did not useheterogeneous IgG glycoforms, but rather antibody forms thatwere either fucosylated or afucosylated. Close inspection oftheir binding data reveals lower heterogeneity for the receptorvariants with reduced complexity in glycan expression. Weidentified several high mannose glycans in our preparationssimilar to Ferrara et al. but fewer outer arm fucosylated glycansthan Zeck et al. A common feature for all investigators,however, is that IgG/receptor binding is heterogeneous. Theseresults indicate that the specificity of glycan binding can be low,and this may partly explain the heterogeneity of binding that isobserved. Zeck and Ferrara also focused on FcγRIIIaVal158,whereas we also investigated binding to FcγRIIIaPhe158. IgGbinding to FcγRIIIaPhe158 is of lower affinity but remainscomplex. Finally, FcγRIa is also interesting as deglycosylation ofthe receptor appears to lower the binding of IgG but with lessimpact on binding kinetics and only slightly reducedheterogeneity. While these experiments continue to shed lighton the binding mechanisms involved, the exact nature of thesugars involved in receptor and IgG binding is still unclear, andthe challenge to determine individual rate and affinity constantsremains. With heterogeneity in both receptor and antibody, atleast eight rate constants have to be determined to describe theinteraction.Interaction of therapeutic antibodies with recombinant FcγRs
is widely used to study the specifity, affinity, and effectiveness ofantibodies. Numerous studies report using HEK293 expressedreceptors in IgG binding experiments.27,34,35 Here, weintroduce the use of single cycle kinetics for these types of
Figure 7. Deglycosylation of FcγRs results in altered binding kineticsof rituximab: A. FcγRI, B. FcγRIIIaVal158, and C. FcγRIIIaPhe158. In eachpanel, rituximab binding to equal amounts of captured unmodifiedglycosylated FcγR, glycosylation control FcγR, and deglycosylatedFcγR is shown. From top to bottom, the sensorgrams in each panelrepresent unmodified FcγR (black), FcγR deglycosylation control(blue), and deglycosylated FcγR (red). The deglycosylation control ineach case is receptor-treated identically to the deglycosylated receptorbut without the deglycosylating enzymes (PNGase F, EndoF1, andEndoF2). Five injections of rituximab in increasing concentration areperformed in each curve, resulting in increasing binding level.Dissociation is seen starting from the end of injection of the highestconcentration, at time 550 s, as well as from the end of injection of thelower concentrations.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855482

binding studies. Observing interactions and complex formationthat does not dissociate completely between injections allowsstudies over an extended period of time, resulting in higherresolution of the data. Furthermore, in combination withcapture of the FcγR, the single cycle kinetic approach savesFcγR reagent and time compared with the more traditionalkinetic assay setup where only one antibody concentration isinjected per interaction cycle. The two kinetic procedures resultin similar kinetic constants.25 We selected the anti-CD20monoclonal antibody rituximab/mabthera as a suitable modelantibody for comparison of binding behavior for differentFcγRs. Consistent with previous reports,6,19,27,36 affinity toFcγRs varied depending on the receptor. The highest affinitywas to FcγRIa and lower affinity to the remaining receptors. KDvalues of 4.2, 6.9, and 3.5 μM were obtained for FcγRIIa,FcγRIIb, and FcγRIIIb, respectively, following fitting to a steadystate model, reflecting the low affinity nature of these receptors.Bruhns et al.6 performed an extensive study presenting data forhuman IgG subclasses, as well as for anti-CD20 antibodiesinteracting with HEK293 expressed FcγRs. Our rituximabaffinity data agree well with their anti-CD20 data for FcγRIIa,FcγRIIb, and FcγRIIIb with data in the 0.5−7 μM range forthese receptors. Visser et al. also present SPR affinity data forrituximab binding to the different FcγRs with reported KDvalues for FcγRIIa, FcγRIIb, and FcγRIIIb all in the range 2.4−12.8 μM.19 Taking into account the different SPR assay setupsused (Bruhns and Visser amine coupled FcγRs, whereas theFcγRs used here were immobilized via the N-terminal His-tag)as well as possible differences in receptor qualities, reportedaffinities for these receptors are consistent. In our study,binding of rituximab to FcγRs was characterized by a rapidapproach to steady state followed by fast dissociation and thesensorgrams contained little kinetic data. To our knowledgesingle cycle kinetics sensorgrams for rituximab binding to FcγRshave not previously been published.The kinetic analysis of rituximab binding to FcγRIa and
FcγRIIIaPhe158/Val158 was complex. When data were fittedassuming a one-to-one binding mechanism, large deviationswere observed, in particular for FcγRIa and FcγRIIIaVal158,whereas rituximab binding to FcγRIIIaPhe158 displayed a fitcloser to a one-to-one binding mechanism. More complexinteraction models (conformational change, FcγR heterogene-ity, antibody heterogeneity, and avidity models) gave betterfitting, but it was not possible to define a precise interactionmechanism. It is likely that both receptor and antibodyheterogeneity is reflected in the observed kinetics, particularlyfucosylated and nonfucosylated antibody glycoforms togetherwith FcγR heterogeneity. As already mentioned, the bindingmechanism is complex, and it therefore cannot be described bya single set of rate constants and by one unique affinityconstant. A minimum of four rate constants and two affinityconstants would be required to describe the data. To avoidoverinterpretation or simplification of these interactions, wedecided not to report values on kinetic and affinity constants.Affinities reported in the literature vary significantly due to thefitting difficulties and differences in experimental setup, as wellin the antibodies and FcγRs used.To investigate the binding mechanism in more detail we
performed an interaction study in which rituximab was allowedto bind to FcγRIIIa at saturating concentrations during 10, 20,or 60 s, after which the dissociation was compared; seeKarlsson and Falt.37 Clear differences between the morphologicvariants of FcγRIIIa were evident (see Figure 5). With
FcγRIIIaVal158, time-dependent dissociation was observed,indicating either that the antibody−receptor complex stabilizedover time due to conformational change or that a variant of theantibody with inherent slower dissociation accumulated on thesensor surface over time. Knowing that approximately 8% of therituximab antibody preparation used here was afucosylated,these results therefore suggest that a large part of theheterogeneity of rituximab binding to FcγRIIIaVal158 is relatedto heterogeneity in the antibody. In contrast, the variation indissociation rate observed for rituximab binding toFcγRIIIaPhe158 with time was minimal, however, with similartendency as for FcγRIIIaVal158. Cardarelli et al. show that there isindeed binding of afucosylated antibody also to FcγRIIIaPhe158but to a far less extent than to FcγRIIIaVal158.
38 Thus, therelatively smaller complexity in rituximab binding observed forFcγRIIIaPhe158 could be explained by independent binding ofrituximab to different receptor forms, i.e., by receptorheterogeneity and to a lesser extent with heterogeneity in theantibody.The injection time experiments shown in Figure 5
demonstrate slower dissociation for prolonged injection timesconsistent with accumulation of the high affinity afucosylatedrituximab. This phenomenon may be exploited as a means todetermine the relative content of afucosylated antibody indifferent batches, where higher content of afucosylatedantibody may be linked to the higher response at a specifictime after the end of injection. Another extension of thisapproach can be to prolong injection times until no furtherstabilization of binding is observed. At this point the receptorwill have the highest occupancy of the high affinity variant, anda dissociation rate constant for the afucosylated version may beobtained from this data.Data from injection time experiments and experiments using
deglycosylated receptors are clearly supportive of each otherand demonstrate that both receptor and rituximab hetero-geneity is reflected in the observed kinetics. It has previouslybeen demonstrated that nonfucosylated IgG binds much tighterin particular to FcγRIIIaVal158,
15 and more recently Zeck et al.demonstrated that when nonfucosylated IgG1 was used as theanalyte, heterogeneity in binding to FcγRIIIaVal158 waseliminated.27 The observation that rituximab interactions withdeglycosylated receptors become more homogeneous isconsistent with FcγR heterogeneity. When rituximab bindingto deglycosylated FcγRIa was analyzed, we observed asignificant drop in the antibody binding level but no apparentchange in the dissociation rate. With deglycosylated FcγRIIIareceptors, a similar, albeit less pronounced, decrease in bindinglevels was observed, but interaction kinetics were changed andrituximab dissociated more quickly from the receptors.Interestingly, the observed heterogeneity of rituximab bindingto deglycosylated FcγRIIIaPhe158 was eliminated when thereceptor was deglycosylated, and here, steady state was reached.This observation adds further strength to the impact of receptorglycosylation on the heterogeneity observed and its influenceon the IgG−FcγR interaction. However, heterogeneity was stillobserved for rituximab binding to deglycosylated FcγRIIIaVal158.A number of important studies have implicated receptor
glycosylation as an important contributor to the IgG−FcγRinteraction. The vast majority of data comes from FcγRIIIa dueto its role in ADCC and its importance to the biopharmaceut-ical industry. N-linked glycans have been shown to positivelyand negatively regulate IgG binding, and recently, crystal-lization of FcγRIIIa with afucosylated anti-CD20 antibody
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855483

revealed a unique interaction consisting of a carbohydrate−carbohydrate interface, which is weakened when the antibody iscore-fucosylated.15−17,39 We and others believe that Fcγreceptor glycosylation and IgG binding are intricately linked,and therefore, the glycan structural analysis described here andits influence on antibody interactions is necessary to provide amore complete picture of the interaction. We believe that thedata presented provides further compelling evidence for therole of FcγR N-glycans in the interaction with IgG and theimmune response. To our knowledge this is the first report thatshows that the binding interaction with antibody is modulatedand stabilized by receptor glycosylation and that antibody bindswith reduced affinity and dissociates quicker from the receptorin the absence of carbohydrates, making it less potent. Itremains a challenge to fully characterize the impact of FcγRglycosylation and the sugars involved, particularly in theirnatural forms by cells such as monocytes, B-cells, and NK cellsand their role in the immune response. Several importantpapers by Kimberly et al. and Edberg et al. have addressed thisquestion, typically using traditional glycoprotein analysistechniques such as lectin binding assays.12,18 Several criticalattributes were described such as the cell type specificglycosylation of FcγRIIIa by monocytes and NK cells anddifferential ligand binding to these receptors.40 This intriguingdiscovery suggests that different cells of the innate immunesystem will use IgG differently depending on the glycosylationstatus of the receptor. This has implications for activation andcontrol of the immune response based on the cells’ ownglycosylation machinery and its preference to glycosylate in acell-type specific manner. Edberg et al. also comment on thesomewhat ubiquitous presence of high mannose type glycansassociated with FcγRs and suggest that they may play a role inopsonin-dependent receptor ligation, a key feature inphagocytic uptake of bacterial pathogens displaying lectin-type adhesins in fimbriated Escherichia coli, Salmonellatyphimurium, and Pseudomonas aeruginosa. This is particularlyfascinating as it may be the case that FcγRs have unknownfunctions, such as in nonclassical receptor ligation. It is alsohighly likely that glycosylation of receptors other than FcγRswill play a similar role in many other protein−receptor andprotein−ligand interactions.
■ AUTHOR INFORMATION
Corresponding Author
*E-mail: [email protected]. Phone: (353) 1 896 3527. Fax: (353)1 677 2400.
Notes
The authors declare the following competing financialinterest(s): A.F., R.K., and C.A. are employees of GEHealthcare; E.C. is an amployee of Waters Corporation; andO.P. is an employee of Agilent Corporation.
■ ACKNOWLEDGMENTS
This work was supported by the National Institute forBioprocessing, Research, and Training (NIBRT), GE Health-care, and the Industrial Development Authority (IDA) Ireland.
■ REFERENCES(1) Chan, P. L.; Sinclair, N. R. Regulation of the immune response.V. An analysis of the function of the Fc portion of antibody insuppression of an immune response with respect to interaction with
components of the lymphoid system. Immunology 1971, 21 (6), 967−81.(2) Anderson, C. L.; Shen, L.; Eicher, D. M.; Wewers, M. D.; Gill, J.K. Phagocytosis mediated by three distinct Fc gamma receptor classeson human leukocytes. J. Exp. Med. 1990, 171 (4), 1333−45.(3) Young, J. D.; Ko, S. S.; Cohn, Z. A. The increase in intracellularfree calcium associated with IgG gamma 2b/gamma 1 Fc receptor-ligand interactions: role in phagocytosis. Proc. Natl. Acad. Sci. U.S.A.1984, 81 (17), 5430−4.(4) Titus, J. A.; Perez, P.; Kaubisch, A.; Garrido, M. A.; Segal, D. M.Human K-natural killer-cells targeted with hetero-cross-linked anti-bodies specifically lyse tumor-cells invitro and prevent tumor-growthinvivo. J. Immunol. 1987, 139 (9), 3153−3158.(5) Kilchherr, E.; Schumaker, V. N.; Phillips, M. L.; Curtiss, L. K.Activation of the first component of human complement, C1, bymonoclonal antibodies directed against different domains ofsubcomponent C1q. J. Immunol 1986, 137 (1), 255−62.(6) Bruhns, P.; Iannascoli, B.; England, P.; Mancardi, D. A.;Fernandez, N.; Jorieux, S.; Daeron, M. Specificity and affinity ofhuman Fcgamma receptors and their polymorphic variants for humanIgG subclasses. Blood 2009, 113 (16), 3716−25.(7) Duits, A. J.; Bootsma, H.; Derksen, R. H.; Spronk, P. E.; Kater, L.;Kallenberg, C. G.; Capel, P. J.; Westerdaal, N. A.; Spierenburg, G. T.;Gmelig-Meyling, F. H.; et al. Skewed distribution of IgG Fc receptorIIa (CD32) polymorphism is associated with renal disease in systemiclupus erythematosus patients. Arthritis Rheum. 1995, 38 (12), 1832−6.(8) Haseley, L. A.; Wisnieski, J. J.; Denburg, M. R.; Michael-Grossman, A. R.; Ginzler, E. M.; Gourley, M. F.; Hoffman, J. H.;Kimberly, R. P.; Salmon, J. E. Antibodies to C1q in systemic lupuserythematosus: characteristics and relation to Fc gamma RIIA alleles.Kidney Int. 1997, 52 (5), 1375−80.(9) Sanders, L. A.; van de Winkel, J. G.; Rijkers, G. T.; Voorhorst-Ogink, M. M.; de Haas, M.; Capel, P. J.; Zegers, B. J. Fc gammareceptor IIa (CD32) heterogeneity in patients with recurrent bacterialrespiratory tract infections. J. Infect. Dis. 1994, 170 (4), 854−61.(10) Wu, J.; Edberg, J. C.; Redecha, P. B.; Bansal, V.; Guyre, P. M.;Coleman, K.; Salmon, J. E.; Kimberly, R. P. A novel polymorphism ofFcgammaRIIIa (CD16) alters receptor function and predisposes toautoimmune disease. J. Clin. Invest. 1997, 100 (5), 1059−70.(11) Nieto, A.; Caliz, R.; Pascual, M.; Mataran, L.; Garcia, S.; Martin,J. Involvement of Fcgamma receptor IIIA genotypes in susceptibility torheumatoid arthritis. Arthritis Rheum. 2000, 43 (4), 735−9.(12) Kimberly, R. P.; Tappe, N. J.; Merriam, L. T.; Redecha, P. B.;Edberg, J. C.; Schwartzman, S.; Valinsky, J. E. Carbohydrates onhuman Fc gamma receptors. Interdependence of the classical IgG andnonclassical lectin-binding sites on human Fc gamma RIII expressedon neutrophils. J. Immunol 1989, 142 (11), 3923−30.(13) Jefferis, R.; Lund, J.; Pound, J. D. IgG-Fc-mediated effectorfunctions: molecular definition of interaction sites for effector ligandsand the role of glycosylation. Immunol Rev. 1998, 163, 59−76.(14) Allhorn, M.; Olin, A. I.; Nimmerjahn, F.; Collin, M. HumanIgG/Fc gamma R interactions are modulated by streptococcal IgGglycan hydrolysis. PLoS One 2008, 3 (1), e1413.(15) Ferrara, C.; Stuart, F.; Sondermann, P.; Brunker, P.; Umana, P.The carbohydrate at FcgammaRIIIa Asn-162. An element required forhigh affinity binding to non-fucosylated IgG glycoforms. J. Biol. Chem.2006, 281 (8), 5032−6.(16) Shibata-Koyama, M.; Iida, S.; Okazaki, A.; Mori, K.; Kitajima-Miyama, K.; Saitou, S.; Kakita, S.; Kanda, Y.; Shitara, K.; Kato, K.;Satoh, M. The N-linked oligosaccharide at Fc gamma RIIIa Asn-45: aninhibitory element for high Fc gamma RIIIa binding affinity to IgGglycoforms lacking core fucosylation. Glycobiology 2009, 19 (2), 126−34.(17) Ferrara, C.; Grau, S.; Jager, C.; Sondermann, P.; Brunker, P.;Waldhauer, I.; Hennig, M.; Ruf, A.; Rufer, A. C.; Stihle, M.; Umana, P.;Benz, J. Unique carbohydrate-carbohydrate interactions are requiredfor high affinity binding between FcgammaRIII and antibodies lackingcore fucose. Proc. Natl. Acad. Sci. U.S.A. 2011, 108 (31), 12669−74.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855484

(18) Edberg, J. C.; Kimberly, R. P. Cell type-specific glycoforms of Fcgamma RIIIa (CD16): differential ligand binding. J. Immunol. 1997,159 (8), 3849−57.(19) Visser, J.; Feuerstein, I.; Stangler, T.; Schmiederer, T.; Fritsch,C.; Schiestl, M. Physicochemical and functional comparability betweenthe proposed biosimilar rituximab GP2013 and originator rituximab.BioDrugs 2013, 27 (5), 495−507.(20) Takahashi, N.; Yamada, W.; Masuda, K.; Araki, H.; Tsukamoto,Y.; Galinha, A.; Sautes, C.; Kato, K.; Shimada, I. N-glycan structures ofa recombinant mouse soluble Fcgamma receptor II. Glycoconjugate J.1998, 15 (9), 905−14.(21) Takahashi, N.; Cohen-Solal, J.; Galinha, A.; Fridman, W. H.;Sautes-Fridman, C.; Kato, K. N-glycosylation profile of recombinanthuman soluble Fcgamma receptor III. Glycobiology 2002, 12 (8), 507−15.(22) Royle, L.; Radcliffe, C. M.; Dwek, R. A.; Rudd, P. M. Detailedstructural analysis of N-glycans released from glycoproteins in SDS-PAGE gel bands using HPLC combined with exoglycosidase arraydigestions. Methods Mol. Biol. 2006, 347, 125−43.(23) Ahn, J.; Bones, J.; Yu, Y. Q.; Rudd, P. M.; Gilar, M. Separationof 2-aminobenzamide labeled glycans using hydrophilic interactionchromatography columns packed with 1.7 microm sorbent. J.Chromatogr. B: Anal. Technol. Biomed. Life Sci. 2010, 878 (3−4),403−8.(24) Harvey, D. J.; Merry, A. H.; Royle, L.; Campbell, M. P.; Dwek,R. A.; Rudd, P. M. Proposal for a standard system for drawingstructural diagrams of N- and O-linked carbohydrates and relatedcompounds. Proteomics 2009, 9 (15), 3796−801.(25) Karlsson, R.; Katsamba, P. S.; Nordin, H.; Pol, E.; Myszka, D. G.Analyzing a kinetic titration series using affinity biosensors. Anal.Biochem. 2006, 349 (1), 136−47.(26) Laemmli, U. K. Cleavage of structural proteins during assemblyof head of bacteriophage-T4. Nature 1970, 227 (5259), 680−&.(27) Zeck, A.; Pohlentz, G.; Schlothauer, T.; Peter-Katalinic, J.;Regula, J. T. Cell type-specific and site directed N-glycosylationpattern of FcgammaRIIIa. J. Proteome Res. 2011, 10 (7), 3031−9.(28) Van den Eijnden, D. H.; Neeleman, A. P.; Van der Knaap, W. P.;Bakker, H.; Agterberg, M.; Van Die, I. Novel glycosylation routes forglycoproteins: the lacdiNAc pathway. Biochem. Soc. Trans. 1995, 23(1), 175−9.(29) Do, K. Y.; Do, S. I.; Cummings, R. D. Differential expression ofLacdiNAc sequences (GalNAc beta 1−4GlcNAc-R) in glycoproteinssynthesized by Chinese hamster ovary and human 293 cells.Glycobiology 1997, 7 (2), 183−94.(30) Smith, P. L.; Skelton, T. P.; Fiete, D.; Dharmesh, S. M.; Beranek,M. C.; MacPhail, L.; Broze, G. J., Jr.; Baenziger, J. U. The asparagine-linked oligosaccharides on tissue factor pathway inhibitor terminatewith SO4−4GalNAc beta 1, 4GlcNAc beta 1,2 Mana alpha. J. Biol.Chem. 1992, 267 (27), 19140−6.(31) Hiyama, J.; Weisshaar, G.; Renwick, A. G. The asparagine-linkedoligosaccharides at individual glycosylation sites in human thyro-trophin. Glycobiology 1992, 2 (5), 401−9.(32) Sato, T.; Gotoh, M.; Kiyohara, K.; Kameyama, A.; Kubota, T.;Kikuchi, N.; Ishizuka, Y.; Iwasaki, H.; Togayachi, A.; Kudo, T.;Ohkura, T.; Nakanishi, H.; Narimatsu, H. Molecular cloning andcharacterization of a novel human beta 1,4-N-acetylgalactosaminyl-transferase, beta 4GalNAc-T3, responsible for the synthesis of N,N′-diacetyllactosediamine, galNAc beta 1−4GlcNAc. J. Biol. Chem. 2003,278 (48), 47534−44.(33) Cosgrave, E. F.; Struwe, W. B.; Hayes, J. M.; Harvey, D. J.;Wormald, M. R.; Rudd, P. M. N-linked glycan structures of the humanFcgamma receptors produced in NS0 cells. J. Proteome Res. 2013, 12(8), 3721−37.(34) Ha, S.; Ou, Y.; Vlasak, J.; Li, Y.; Wang, S.; Vo, K.; Du, Y.; Mach,A.; Fang, Y.; Zhang, N. Isolation and characterization of IgG1 withasymmetrical Fc glycosylation. Glycobiology 2011, 21 (8), 1087−96.(35) Kainer, M.; Antes, B.; Wiederkum, S.; Wozniak-Knopp, G.;Bauer, A.; Ruker, F.; Woisetschlager, M. Correlation between CD16abinding and immuno effector functionality of an antigen specific
immunoglobulin Fc fragment (Fcab). Arch. Biochem. Biophys. 2012,526 (2), 154−8.(36) Maenaka, K.; van der Merwe, P. A.; Stuart, D. I.; Jones, E. Y.;Sondermann, P. The human low affinity Fcgamma receptors IIa, IIb,and III bind IgG with fast kinetics and distinct thermodynamicproperties. J. Biol. Chem. 2001, 276 (48), 44898−904.(37) Karlsson, R.; Falt, A. Experimental design for kinetic analysis ofprotein-protein interactions with surface plasmon resonance bio-sensors. J. Immunol. Methods 1997, 200 (1−2), 121−33.(38) Cardarelli, P. M.; Rao-Naik, C.; Chen, S.; Huang, H.; Pham, A.;Moldovan-Loomis, M. C.; Pan, C.; Preston, B.; Passmore, D.; Liu, J.;Kuhne, M. R.; Witte, A.; Blanset, D.; King, D. J. A nonfucosylatedhuman antibody to CD19 with potent B-cell depletive activity fortherapy of B-cell malignancies. Cancer Immunol. Immunother. 2010, 59(2), 257−65.(39) Mizushima, T.; Yagi, H.; Takemoto, E.; Shibata-Koyama, M.;Isoda, Y.; Iida, S.; Masuda, K.; Satoh, M.; Kato, K. Structural basis forimproved efficacy of therapeutic antibodies on defucosylation of theirFc glycans. Genes Cells 2011, 16 (11), 1071−80.(40) Edberg, J. C.; Barinsky, M.; Redecha, P. B.; Salmon, J. E.;Kimberly, R. P. Fc gamma RIII expressed on cultured monocytes is aN-glycosylated transmembrane protein distinct from Fc gamma RIIIexpressed on natural killer cells. J. Immunol. 1990, 144 (12), 4729−34.(41) Harvey, D. J.; Mattu, T. S.; Wormald, M. R.; Royle, L.; Dwek, R.A.; Rudd, P. M. ″Internal residue loss″: rearrangements occurringduring the fragmentation of carbohydrates derivatized at the reducingterminus. Anal. Chem. 2002, 74 (4), 734−40.(42) Tomiya, N.; Lee, Y. C.; Yoshida, T.; Wada, Y.; Awaya, J.;Kurono, M.; Takahashi, N. Calculated two-dimensional sugar map ofpyridylaminated oligosaccharides: elucidation of the jack bean alpha-mannosidase digestion pathway of Man9GlcNAc2. Anal. Biochem.1991, 193 (1), 90−100.(43) Dohi, K.; Isoyama-Tanaka, J.; Misaki, R.; Fujiyama, K. Jack beanalpha-mannosidase digestion profile of hybrid-type N-glycans: effect ofreaction pH on substrate preference. Biochimie 2011, 93 (4), 766−71.
Journal of Proteome Research Article
dx.doi.org/10.1021/pr500414q | J. Proteome Res. 2014, 13, 5471−54855485




















