
Extracellular matrix stimulates reactive oxygen species production and increases pancreatic cancer...
48
1 EXTRACELLULAR MATRIX STIMULATES REACTIVE OXYGEN SPECIES PRODUCTION AND INCREASES PANCREATIC CANCER CELL SURVIVAL THROUGH 5-LIPOXYGENASE AND NADPH OXIDASE $ . Mouad Edderkaoui * , Peggy Hong * , Eva C. Vaquero * , Jong K. Lee * , Lars Fischer *# , Helmut Friess # , Markus W. Buchler # , Markus M. Lerch & , Stephen J. Pandol * , Anna S. Gukovskaya * . * Department of Medicine, Veterans Affairs Greater Los Angeles Healthcare System and University of California, Los Angeles, California. & Department of Gastroenterology, Endocrinology and Nutrition, Ernst-Moritz-Arndt Universität Greifswald, Germany. # Department of General Surgery, University of Heidelberg, Heidelberg, Germany. Running title : ROS ECM proteins pancreatic cancer cells Anna Gukovskaya, Ph.D VA Greater Los Angeles Healthcare System. West Los Angeles VA Healthcare Center 11301 Wilshire Blvd, Blg 258, Rm 340 Los Angeles, CA 90073, USA. Tel: (310) 478-3711 x41525 Fax: 310-268-4578 E-mail: [email protected] Articles in PresS. Am J Physiol Gastrointest Liver Physiol (July 21, 2005). doi:10.1152/ajpgi.00197.2005 Copyright © 2005 by the American Physiological Society.
Transcript of Extracellular matrix stimulates reactive oxygen species production and increases pancreatic cancer...

1
EXTRACELLULAR MATRIX STIMULATES REACTIVE OXYGEN SPECIES
PRODUCTION AND INCREASES PANCREATIC CANCER CELL SURVIVAL
THROUGH 5-LIPOXYGENASE AND NADPH OXIDASE$.
Mouad Edderkaoui*, Peggy Hong*, Eva C. Vaquero*, Jong K. Lee*, Lars Fischer*#,
Helmut Friess#, Markus W. Buchler#, Markus M. Lerch&, Stephen J. Pandol*, Anna S.
Gukovskaya*.
* Department of Medicine, Veterans Affairs Greater Los Angeles Healthcare System and
University of California, Los Angeles, California. & Department of Gastroenterology,
Endocrinology and Nutrition, Ernst-Moritz-Arndt Universität Greifswald, Germany. #
Department of General Surgery, University of Heidelberg, Heidelberg, Germany.
Running title: ROS ECM proteins pancreatic cancer cells
Anna Gukovskaya, Ph.D
VA Greater Los Angeles Healthcare System.
West Los Angeles VA Healthcare Center
11301 Wilshire Blvd, Blg 258, Rm 340
Los Angeles, CA 90073, USA.
Tel: (310) 478-3711 x41525
Fax: 310-268-4578
E-mail: [email protected]
Articles in PresS. Am J Physiol Gastrointest Liver Physiol (July 21, 2005). doi:10.1152/ajpgi.00197.2005
Copyright © 2005 by the American Physiological Society.

2
ABBREVIATIONS
ROS reactive oxygen species
ECM extracellular matrix
NOX non-phagocytic oxidase
LO lipoxygenase
NDGA nordihydroguaiaretic acid
Ac-DEVD-AMC Ac-Asp-Glu-Val-Asp-7-amino-4-methylcoumarin
AnV Annexin V
DPI diphenylene iodonium
ECL enhanced chemiluminescence
FACS fluorescence-activated cell sorter
FBS fetal bovine serum
FCCP flourocyanide m-chlorophenylhydrazone
MtDNA mitochondrial DNA
PI propidium iodide
PS phosphatidylserine
zVAD-fmk z-Val-Ala-Asp(OMe)-CH2F

3
ABSTRACT
Extracellular matrix (ECM) facilitates pancreatic cancer cells survival, which is of central
importance for pancreatic adenocarcinoma that is highly fibrotic. Here we show that reactive
oxygen species (ROS) mediate the prosurvival effect of ECM in human pancreatic cancer cells.
Fibronectin and laminin stimulated ROS production and NADPH oxidase activation in pancreatic
cancer cells. Both pharmacologic and molecular approaches show that fibronectin stimulated
ROS production through activation of NADPH oxidase and NADPH oxidase-independent
pathways, and that 5-lipoxygenase (5-LO) mediates both these pathways. Analysis of the
mechanisms of ROS production by ECM proteins and growth factors indicate that activation of
NADPH oxidase, Nox4 is a common mechanism employed by both ECM proteins and growth
factors to increase ROS in pancreatic cancer cells. We also found that Nox4 is present in human
pancreatic adenocarcinoma tissues, and that these tissues display membrane NADPH oxidase
activity. ECM proteins and growth factors activate NADPH oxidase through different
mechanisms; by contrast to ECM proteins, growth factors activate NADPH oxidase through 5-
LO-independent mechanisms. Inhibiting 5-LO or NADPH oxidase with pharmacologic inhibitors
of these enzymes and with Nox4 or 5-LO antisense oligonucleotides markedly stimulated
apoptosis in cancer cells cultured on fibronectin. Our results indicate that ROS generation via 5-
LO and downstream NADPH oxidase mediates the prosurvival effect of ECM in pancreatic
cancer cells. These mechanisms may play an important role in pancreatic cancer resistance to
treatments and thus represent novel therapeutic targets.

4
INTRODUCTION
Reactive oxygen species (ROS) are highly reactive O2 metabolites that include
superoxide radical (O2ֹ-), hydrogen peroxide (H2O2), and hydroxyl radical (OH.) (44).
Whereas ROS are classically thought of as cytotoxic and mutagenic (23; 33), recent
evidence suggests that ROS such as superoxide and hydrogen peroxide serve as
regulators in signal transduction pathways (38). ROS are involved in proliferation, cell
death and adhesion pathways (7; 9; 39; 42). The targets of ROS include such key
signaling molecules as transcription factor NF-kB, MAP kinases, and tyrosine
phosphatases (10; 15; 46; 55).
In phagocytes, large amounts of ROS are produced during the oxidative burst by the
plasma membrane NAD(P)H oxidase, a well characterized multicomponent enzyme with
gp91phox and p22phox catalytic subunits that together form an integral complex called
flavocytochrome b558 (3; 4). Non-phagocytic cells also produce significant amounts of
ROS, albeit much smaller than in phagocytes (4; 5; 25). In non-phagocytic cells, ROS are
produced by mitochondria as well as by a number of ROS-generating plasma membrane
and cytosolic enzymes (44). Over the past several years, members of the family of non-
phagocytic NADPH oxidases (NOXes) homologous to gp91phox were shown to mediate
ROS production in non-phagocytic cells (4; 5; 25; 26). Based on our present state of
knowledge the family consists of 7 members. NOXes are classified into 3 main groups
according to their domain structure. In particular, Nox1, -3 and -4 are very similar in size
and structure to gp91phox (now termed Nox2), and they also contain the domains required
to transfer electrons from NADPH to molecular oxygen to form superoxide. NOXes are

5
expressed (at different levels) in various tissues and are thought to be a major mechanism
of ROS production in non-phagocytic cells.
The products of PLA2/LO pathway such as leukotrienes and HETEs are also involved
in ROS generation. They may participate in activation of NADPH oxidase (30; 51-53)
and also produce ROS as by products during oxidation of arachidonic acid by
lipoxygenases (22).
We recently showed that different Nox isoforms are expressed in pancreatic cancer
cells (48). In particular, Nox4 is present in these cells at both mRNA and protein level.
Growth factors activate NADPH oxidase resulting in increase in cellular ROS. We also
showed that ROS produced via NADPH oxidase mediate pro-survival effect of growth
factors in pancreatic cancer cells (48).
The important characteristic of pancreatic adenocarcinoma is high expression of
extracellular matrix protein (16; 29; 49; 54). When deprived of ECM, normal cells
undergo apoptosis. This process, termed anoikis, controls cell proliferation by deleting
misplaced cells. Cancer cells are usually resistant to anoikis. We recently showed that
detachment from ECM stimulates pancreatic cancer cells death, and that fibronectin and
laminin markedly protect pancreatic cancer cells from death (47). The mechanisms
through which ECM inhibit pancreatic cancer cells death are not well understood. In
particular, the role of ROS in the prosurvival effects of ECM proteins has not been
studied. Information on the effects of ECM proteins on cellular ROS, in general, are
limited (20; 32; 50). Nothing is known on the effects of ECM on ROS production in
pancreatic cancer cells.

6
In the present study we provide evidence that in pancreatic cancer cells, ECM
proteins, fibronectin and laminin, increase intracellular ROS and stimulate NADPH
oxidase activity. Both pharmacologic and molecular approaches show that fibronectin
stimulated ROS production through activation of NADPH oxidase and NADPH oxidase-
independent pathways. 5-lipoxygenase (5-LO) is a critical regulator of ECM-induced
ROS generation; it mediates both activation of NADPH oxidase and ROS generation
through NADPH oxidase-independent mechanism. Inhibiting ROS with NADPH oxidase
and 5-LO inhibitors, Nox4, or 5-LO antisense stimulated apoptosis in pancreatic cancer
cells. Thus, NADPH oxidase and 5-LO provide mechanisms through which ECM
proteins protect pancreatic cancer cells from death. The pro-survival effect of ROS may
be an important mechanism of pancreatic cancer cell resistance to therapy.
EXPERIMENTAL PROCEDURES
Reagents. The polyclonal Nox4 antibody generated from a His-tagged
recombinant fragment of human Nox4 encoding the C-terminal amino acids 494 to 513
described in (32) was a gift from Dr. Barry Goldstein (Thomas Jefferson University,
Philadelphia, Pennsylvania). Antibody against 5-LO from Cayman Chemical and anti-α5
antibody from Santa Cruz Biotechnology, Inc. 2’7’-dichlorofluorescein diacetate (DCFH-
DA) was from Molecular Probes (Eugene, OR); DEVD-AMC (Ac-Asp-Glu-Val-Asp-
aminomethylcoumarin), from Peptide Institute, Inc (Osaka, Japan); zVAD-fmk (z-Val-
Ala-Asp(OMe)-CH2F), from Enzyme Systems Products (Livermore, CA). AACOCF3,

7
nordihydroguaiaretic acid (NDGA), baicalein, and REV-5901 were from Calbiochem
(San-Diego, CA). All other reagents were from Sigma Chemical (St. Louis, MO).
Cell culture. Human pancreatic adenocarcinoma cell lines, the poorly
differentiated MIA PaCa-2, slightly differentiated PANC-1, and moderately to well
differentiated CAPAN-1 were obtained from the American Type Culture Collection
(Manassas, VA). MIA PaCa-2 and PANC-1 cells were grown in 1/1 D-MEM/F-12
medium (GIBCO Invitrogen Corporation, Grand Island, NY) supplemented with 15%
fetal bovine serum (FBS), 4 mM L-glutamine, and antibiotic/antimicotic solution (Omega
Scientific, Tarzana, CA). CAPAN-1 cells were grown in RPMI 1960 medium (GIBCO
Invitrogen Corporation, Grand Island, NY) supplemented with 10% fetal bovine serum
(FBS), 4 mM L-glutamine, and antibiotic/antimicotic solution (Omega Scientific,
Tarzana, CA). Cells were maintained at 37°C in a humidified atmosphere containing 5%
CO2 and were used between passages 4 and 12.
MIA PaCa-2, CAPAN-1 and PANC-1 cells were plated at a density of 5 x 105/ml
on 100-mm culture dishes, coated with either poly(2-hydroxyethyl metacrylate)
(polyHEMA) or with an ECM protein, cultured for up to 72 h in serum free-D-MEM/F-
12 or RPMI 1640 media, collected, and processed for the specified analyses. PolyHEMA
prevents both cell attachment on plastic and endogenous cell matrix deposition (14).
Plates were coated twice with 3 ml of polyHEMA dissolved in 95% ethanol (10 mg/ml),
allowed to dry, and washed twice with PBS before used. Compared with polyHEMA, we
observed similar but less pronounced responses in cells grown on plastic or on plastic
coated with polylysine (24). Plates coated with human laminin, fibronectin (Sigma
Chemical, St. Louis, MO) or bovine collagen I (Chemicon, Temecula, CA) were prepared

8
by applying 2 µg/cm2 of the specified protein for 1 h at room temperature. Cells were
cultured for 48h or indicated time in the absence of FBS. Inhibitors or vehicle were added
to the culture medium just before plating the cells.
Tissue samples. Pancreatic cancer tissues were obtained from surgical specimens
from 4 different patients with pancreatic ductal cell adenocarcinoma. Tissue samples
were frozen in liquid nitrogen immediately on surgical removal and maintained at –80°C
until use. All studies were approved by the ethics committees of the University of
Heidelberg, Germany, and by the human subjects committee of the West Los Angeles
VAGLAHS, USA.
Generation of Rho0 cells. MIA PaCa-2 Rho0 cells were depleted of mitochondrial
DNA (mtDNA) as we described before (48) by incubating wild type cells (Rho+) for 6-8
weeks with 100 ng/ml ethidium bromide. The medium was supplemented with 4.5 mg/ml
glucose, 50 µg/ml uridine, and 100 µg/ml pyruvate to compensate for the respiratory
metabolism deficit as previously described. After selection, the MIA PaCa-2 Rho0 cells
were cultured in the above-specified medium without ethidium bromide. The mtDNA
depletion was verified with PCR using human mtDNA specific primers and with western
blot analysis for mitochondrial DNA coded cytochrome c oxidase, subunit IV as we
discussed previously (48).
Intracellular ROS levels were measured by flow cytometry in cells loaded with
the redox-sensitive dye DCFH-DA as we described in (48). The non-fluorescent DCFH-
DA readily diffuses into the cells where it is hydrolyzed to the polar derivative DCFH,
which is oxidized in the presence of H2O2 to the highly fluorescent DCF. DCFH in
addition to H2O2 is also oxidized by HOCl, nitric oxide but not by superoxide (19).

9
Approximately 1x106 cells were incubated in the dark for 30 min at 37 oC with 10 µM
DCFH-DA, harvested, and resuspended in the medium without DCFH-DA. Fluorescence
was recorded on FL-1 channel of FACScan (Becton Dickinson) flow cytometer, data
analyzed with Cell Quest program.
Measurement of NADPH oxidase activity. Superoxide production was measured
in total cell or tissue homogenates by using lucigenin-derived chemiluminescence
according to the protocols described previously (37; 40; 48); Cells or tissue samples from
human pancreatic adenocarcinomas were homogenized in a Dounce homogenizer in the
medium containing 50 mM phosphate buffer, 1 mM EGTA and 150 mM sucrose.
Homogenates were centrifuged and pellets (membrane fractions) were stored at -80˚C.
For superoxide measurements, 50 µg of protein was diluted in 500 µl of the same lysis
buffer. Dark-adapted lucigenin was added to the sample, and chemiluminescence
measurement immediately started. Chemiluminescence (in arbitrary units) was measured
at 15-sec intervals for 1 min in a Turner 20/20 luminometer (Turner Designs, Sunnyvale,
CA). NADPH was used as a substrate. The specificity of the measurement was confirmed
by adding either a non-enzymatic superoxide scavenger, tiron (10 mM), or superoxide
dismutase (Sigma; 200 units/ml). In some experiments the homogenate was pre-
incubated for 10 min with DPI as described in figure legends. In this and other assays,
protein concentration was measured by the Bradford assay (BioRad Laboratories,
Hercules, CA).
It has been shown that at high doses lucigenin can by itself stimulate additional
superoxide production, which is especially pronounced with NADH as a substrate (21;
28). As we described previously (48) we measured chemiluminescence in membrane

10
fractions at 5, 25, 50, 100, and 200 µM lucigenin in the presence and absence of NADPH
or NADH. By using both NADH and NADPH as substrates we confirmed that at 5-50
µM lucigenin does not induce an artificial O2ֹ- production in pancreatic cancer cells. The
data presented were obtained with 15 µM lucigenin.
Transfections. Transfection of MIA PaCa-2 cells with Nox4 antisense
oligonucleotides was done as we described previously (48). 1.5 nmoles of the Nox4
antisense phosphorothioate oligonucleotides, Nox4 AS1: 5’-AGC TCC TCC AGG ACA
CAG CC, or Nox4 AS2: 5’-GGA CAC AGC CAT GCC GCC (1), was applied for a 60-
mm dish. The Nox4 scrambled oligonucleotides, 5’-TCG AGG AGG TCC TGT GTC
GG, was used as a control. 6 hours post-transfection, fresh medium was supplied and
cells were cultured for additional 72 hours before specified analyses. Transfection with 5-
LO antisense oligonucleotides was performed using the same technique with 5-LO
antisense phosphorothioate oligonucleotides, 5-LO AS1: 5’CAC AGT CAC GTC GTC
GTA TGA ATC CAC C 3’ (7) or 5-LO AS2: 5’-GTG ACC GTG TAG GAG GGC AT,
and 5-LO scrambled phosphorothioate oligonucleotides: 5’CGT CCA GAC CAA GGA
CGA GCT TGC A 3’ as a control.
The efficiency of transfection assessed by expression of the green-fluorescent
protein containing plasmid was about 50-60% in both cell lines MIA PaCa-2 and PANC-
1. We did not use CAPAN-1 cells in transfection experiments because of their low
transfection efficiency. The specificity of Nox4 and 5-LO antisense oligonucleotides was
checked by BLASTing them against the human genome.
The transfection efficiency for CAPAN-1 cells was low therefore transfection
experiments were performed with Mia PaCa-2 and PANC-1 cells only.

11
Measurements of apoptosis. Internucleosomal DNA fragmentation was measured
by using Cell Death Detection ELISAPlus kit (Roche Molecular Biochemicals, Manheim,
Germany) according to the manufacturer’s instructions.
Phosphatidylserine externalization was analyzed with the Annexin-V-FLUOS
Staining Kit from Roche Biochemicals (Indianapolis, IN) as we described before (47;
48). Cells were collected and resuspended at a density of 1 x 106 cells in 500 µl of
binding buffer containing 2 µl Annexin V (AnV) and 1 µl propidium iodide (PI),
incubated in the dark for 30 min at room temperature, and analyzed by flow cytometry.
Effector caspase (DEVDase) activity was measured by a fluorogenic assay in
whole cell lysates using DEVD-AMC as a substrate, as we described (47; 48). The lysate
(50-100 µg of protein) was incubated with 10 µM substrate in a reaction buffer (25 mM
HEPES (pH 7.5), 10% sucrose, 0.1% CHAPS, 10 mM DTT) at 37 °C. Caspase substrate
cleavage releases AMC, which emits fluorescent signal with 380 nm excitation and 440
nm emission. Fluorescence was calibrated using a standard curve for AMC.
Western blot analysis. Cells were incubated in a lysis buffer A (0.5 mM EDTA,
150 mM NaCl, 50 mM Tris, 0.5% Nonidet P-40. pH 7.5) for 30 min at 40C. Tissue
samples from human pancreatic adenocarcinomas were homogenized in the lysis buffer B
(150 mM NaCl, 50 mM Tris, 1% deoxycholic acid, 1% triton, 0.1% SDS). Both buffers
A & B were supplemented with 1mM PMSF, 5ug/ml each of protease inhibitors
pepstatin, leupeptin, chymostatin, antipain, and aprotinin. Cell and tissue lysates were
centrifuged for 10 min at 13,000 x g. Supernatants were collected and proteins were
separated by SDS-PAGE (Invitrogen) and electrophoretically transferred to nitrocellulose
membranes. Nonspecific binding was blocked with 5% milk in Tris-buffered saline (4

12
mM Tris base, 100 mM NaCl, pH 7.5). Membranes were washed in Tris-buffered saline
containing 0.05% Tween 20 (TTBS) and incubated for 2 h with the indicated primary
antibodies and then for 1 h with horseradish peroxidase-conjugated secondary antibody.
Blots were developed with the Supersignal chemiluminescent Substrate (Pierce).
Immunohistochemisrty. Immunostaining of frozen tissue sections from human
pancreatic adenocarcinomas was performed as we described previously (17). The
sections were incubated in a blocking medium containing PBS/glycine, 1% goat serum,
1% bovine serum albumin (BSA) and 1% gelatin for 30 min. Then slides were incubated
for 10 min in a working medium (PBS supplemented with 0.05 M glycine, 0.1% goat
serum, 0.1% BSA and 0.1% gelatin) followed by a 2-hour incubation with anti Nox4
antibody dissolved in a working medium. Bound antibody was detected with FITC-
conjugated secondary antibody.
Statistical analysis. Results are expressed as means ± SEM from at least 3
independent experiments. Statistical analysis was done using unpaired Student’s t test.
The value of P < 0.05 was considered statistically significant.
RESULTS
Fibronectin, laminin and collagen I stimulate intracellular ROS production in MIA
PaCa-2, CAPAN-1 and PANC-1 pancreatic carcinoma cells.
Using the redox-sensitive fluorescence probe DCFH-DA, we determined that
ECM proteins stimulated ROS production in pancreatic cancer cell lines (Fig. 1). In MIA

13
PaCa-2 (Fig. 1A), CAPAN-1 (Fig. 1B) and PANC-1 cells (Fig. 1C), levels of intracellular
ROS were higher in cells attached to ECM proteins than in detached cells. Fibronectin
and laminin increased intracellular ROS to the same extent in all 3 cell lines tested. (Fig.
1D). Thus, fibronectin produces the same increase in ROS in poorly differentiated (MIA
PaCa-2), slightly differentiated (PANC-1) and well-differentiated (CAPAN-1) cells.
Collagen I only stimulated ROS in PANC-1 cells (Fig. 1A-C). Increases in cellular ROS
by fibronectin and laminin were of the same magnitude as effects of exogenous 10 µM
H2O2.
The effect of ECM proteins on cell signaling is mediated through their receptors,
integrins. Figure 1E shows that blockade of fibronectin receptor, α5β1 integrin (2; 18)
with anti- α5 integrin antibody greatly inhibited fibronectin-induced increase in ROS.
Figure 2 shows that detachment from ECM causes time-dependent decrease in ROS
whereas in attached cells, intracellular ROS levels are maintained for up to 72 h.
ECM proteins increase cellular ROS through NADPH oxidase and 5-LO. 5-LO mediates
both NADPH oxidase-dependent and -independent pathway (s).
To determine the sources of intracellular ROS we first used pharmacologic
approach (Fig. 3). We applied inhibitors of various ROS-producing systems at
concentrations reported to completely inhibit their target enzymes. The superoxide
scavenger Tiron, decreased ROS levels in cells cultured on both polyHEMA and ECM
proteins, indicating the contribution of superoxide in ROS generation in pancreatic cancer
cells (Fig. 3A)

14
In MIA PaCa-2, CAPAN-1 (Fig. 3A) cells cultured on polyHEMA (the substrate
that prevents both cell attachment on plastic and endogenous ECM deposition) (47), ROS
generation was partially but significantly inhibited by DPI, an inhibitor of NADPH-
dependent oxidases (4). Inhibitors of phospholipase A2 (PLA2) /5-LO pathway, in
particular, inhibitor of cytosolic PLA2 AACOCF3 (24), a broad spectrum LO inhibitor
(NDGA, (31; 41), and a selective inhibitor of 5-LO, REV-5901 (43), did not significantly
affect intracellular ROS levels in cells cultured on polyHEMA (Fig. 3A,B). Moreover,
the inhibitory effect of DPI plus REV-5901 on the basal ROS level was the same as with
DPI alone (Fig. 3A). These results suggest that NADPH-oxidase but not PLA2 /5-LO
contributes to the ROS production in cells cultured on polyHEMA.
By contrast, ROS production in pancreatic cancer cells cultured on fibronectin
was inhibited by both DPI and inhibitors of PLA2 /5-LO pathway (Fig.3A, B). These data
(Fig. 3A, B) indicate that in cells cultured on ECM proteins ROS production is mediated
through 5-LO and NADPH oxidase. In cells cultured on either polyHEMA or ECM
proteins, inhibitors of 12-LO, xanthine oxidase, and nitric oxide synthase (6; 35; 36) did
not affect intracellular ROS (Table 1) indicating no involvement of these enzymes.
The experiments on MIA PaCa-2 cells show that the effects of laminin on cellular ROS
are also inhibited by DPI and REV-5901 (Fig.3A). They suggest that laminin employs the
same mechanisms as fibronectin to increase cellular ROS.
DPI and REV-5901 inhibited fibronectin-induced increase in cellular ROS in both
poorly differentiated MIA PaCa-2 (Fig.3A) and well differentiated CAPAN-1 (Fig.3B)
cells.

15
To further elucidate the role of NADPH oxidase in the effects of ECM proteins on
cellular ROS we measured effects of ECM proteins on NADPH oxidase activity. Both
fibronectin and laminin stimulated NADPH oxidase activity in membrane fractions from
MIA PaCa-2 cells (Fig. 3C). NADPH oxidase activity in cells cultured on fibronectin (as
well as on polyHEMA) was completely prevented by DPI (Fig. 3C). On the other hand,
5-LO inhibitor REV-5901 did not inhibit NADPH oxidase activity in cells cultured on
polyHEMA; but abrogated NADPH oxidase activation by fibronectin (Fig. 3C). These
data suggest that 5-LO mediates fibronectin-induced activation of NADPH oxidase but
not the basal NADPH oxidase activity. Fibronectin stimulated NADPH oxidase activity
in both poorly differentiated MIA PaCa-2 (Fig. 3C) and well differentiated CAPAN-1
(Fig. 3D) cells.
Taken together the results in the Figure 3 show that inhibition of 5-LO prevented
both effects of fibronectin namely activation of NADPH oxidase and increase in cellular
ROS. These data indicate that 5-LO mediates increase in cellular ROS by fibronectin.
They further suggest that activation of NADPH oxidase is one mechanism through which
5-LO increases cellular ROS. DPI, which completely inhibited NADPH oxidase activity
(Fig 3C), only partially decreased fibronectin-induced increase in ROS in MIA PaCa-2
cells (Fig 3A). Furthermore, inhibition of 5-LO with REV-5901 caused additional
inhibition of ROS response to fibronectin in cells treated with DPI, abrogating the effect
of fibronectin on ROS (Fig. 3A). These results suggest that ECM-induced ROS
production is mediated through both NADPH oxidase and NADPH-oxidase-independent
mechanisms. They also suggest that 5-LO mediates both these mechanisms.

16
Lack of the effects of REV-5901 on cellular ROS and NADPH oxidase activity
indicates that 5-LO is not involved in ROS generation in cells cultured on polyHEMA
(Fig. 3A-C). DPI only partially decreased cellular ROS in MIA PaCa-2 cells cultured on
polyHEMA, suggesting contribution of mechanisms other than NADPH oxidase
Inhibiting 5-LO or NADPH oxidase with 5-LO or Nox4 antisense oligonucleotides
decreases ROS production in pancreatic cancer cells cultured on fibronectin.
We showed previously that Nox4 is expressed in pancreatic cancer cells at both
mRNA and protein levels (48). To determine the role of Nox4 in the generation of ROS
by fibronectin we transfected MIA PaCa-2 and PANC-1 cells with Nox4 antisense (as
well as scrambled) oligonucleotides. We used two different antisense (AS)
oligonucleotides targeting the human Nox4 (Fig. 4). Both AS1 (Fig 4A-C) and AS2 (Fig
4D, E) were effective at reducing levels of endogenous Nox4 protein when transfected
into MIA PaCa-2 and PANC-1 cells. Both AS oligonucleotides showed inhibition of
ROS (Fig. 4). In cells cultured on fibronectin, Nox4 antisense oligonucleotides decreased
both intracellular ROS levels (Fig. 4B, E) and NADPH oxidase activity (Fig. 4C)
compared with cells transfected with the scrambled oligonucleotides.
Similarly, to confirm our results obtained with the pharmacologic inhibitors on
the involvement of 5-LO, we transfected MIA PaCa-2 and PANC-1 cells with 5-LO
antisense oligonucleotides. The results show decreased level of intracellular ROS (Fig.
5B, E) and NADPH oxidase activity (Fig. 5C) in cells transfected with 5-LO antisense
oligonucleotides and cultured on fibronectin.

17
The results of transfection experiments confirm the involvement of 5-LO and
Nox4 in fibronectin-induced ROS production.
Involvement of mitochondrial NADH dehydrogenase to the ECM-induced increase in
cellular ROS.
In addition to membrane NADPH oxidase, DPI can also inhibit mitochondrial
NADH dehydrogenase (27).To evaluate the contribution of mitochondrial NADH
dehydrogenase, we used MIA PaCa-2 Rho0 cells depleted of mtDNA that codes for this
enzyme. Using PCR and western blot analysis we showed the absence of mtDNA in Rho0
cells (48).
In Rho0 cells, fibronectin and laminin increased intracellular ROS levels similar to
parental (wild type) Rho+ MIA PaCa-2 cells (Fig. 6A)
To evaluate contribution of mitochondria in fibronectin-induced ROS production,
we also applied mitochondrial complex I inhibitor, rotenone to the wild type MIA PaCa-2
Rho+ cells. In cells cultured with 1µM rotenone for 48 h, fibronectin stimulated
intracellular ROS similar to that in the absence of rotenone (Fig. 6B). These results
indicate that the mitochondrial NADH dehydrogenase is not a major contributor to the
ECM-induced ROS generation in MIA PaCa-2 cells.
Antioxidants as well as Nox4 and 5-LO antisense oligonucleotides stimulate apoptosis in
pancreatic cancer cells cultured on ECM proteins.
We published before that ECM proteins inhibit death responses in pancreatic
cancer cells (47; 47). In particular, we found (47) that attachment to fibronectin or

18
laminin significantly increased survival of MIA PaCa-2 cells cultured for on polyHEMA
without growth factors. We also found that the prosurvival effect of ECM proteins was
similar to that of growth factors (48).
In the following experiments we measured the role of ROS in the prosurvival
effects of ECM proteins. Inhibition of intracellular ROS with antioxidants (DPI, REV-
5901, tiron) stimulated internucleosomal DNA fragmentation, a hallmark of apoptosis, in
the MIA PaCa-2 cells cultured on fibronectin or laminin (Fig. 7A). In contrast, there was
no effect on DNA fragmentation of the inhibitors of enzymes that we found not involved
in ROS production in pancreatic cancer cells, allopurinol (xanthine oxidase), baicalein
(12-LO), and L-NAME (nitric oxide synthase) (not shown). Antioxidants increased DNA
fragmentation in both poor differentiated MIA PaCa-2 and well-differentiated CAPAN-1
cells (Fig.7A, B).
Cell survival was also analyzed by flow cytometry in cells stained with AnV and
PI. AnV was applied to detect phosphatidylserine externalization, a marker of apoptosis;
PI to identify loss of plasma membrane integrity, a hallmark of necrosis. We showed
before (47) that pancreatic cancer cells with apoptotic nuclear morphology are either in
An+/PI- or An+/PI+ group. An+/PI- is a group of cells with early apoptosis, whereas
An+/PI+ group includes cells with late apoptosis associated with the secondary necrosis
(47). Figure 7C shows that DPI increased the percentage of dead cells (An+/PI- together
with An+/PI+) from 35% to 55%. It also decreased survival of MIA PaCa-2 cells from
62% down to 38%.

19
Inhibition of ROS with antioxidants also activates effector caspases (Fig. 7D).
Both caspase activation (Fig. 7D) and DNA fragmentation (not illustrated) induced by
antioxidants were inhibited by zVAD-fmk, a broad-spectrum caspase inhibitor.
To further determine the role of NADPH oxidase and 5-LO in the inhibition of
apoptosis by fibronectin we used a molecular approach. Transfection of MIA PaCa-2 and
PANC-1 cells with Nox4 or 5-LO antisense oligonucleotides increased both DNA
fragmentation and the percentage of AnV+ positive cells (Fig. 8A-D). Inhibition of Nox4
or 5-LO by antisense oligonucleotides, also activated effector caspases (Fig. 8E, F). Of
note both AS1 (Fig 8) and AS2 (not shown) Nox4 oligonucleotides, and both AS1 (Fig 8)
and AS2 (not shown) 5-LO oligonucleotides demonstrated similar stimulation of
apoptosis.
These data (Figs 7 & 8) indicate that inhibition of ROS with antioxidants, or
transfection with Nox4 or 5-LO antisense oligonucleotides, all stimulate apoptosis in
MIA PaCa-2 and PANC-1 cells. Apoptosis induced by ROS inhibition is mediated by
caspases.
The effect of the combination of fibronectin and growth factors on cellular ROS and
apoptosis is greater than that of fibronectin or growth factors alone.
We showed previously (48) that growth factors increased cellular ROS in
pancreatic cancer cells through activation of NADPH oxidase. Thus, NADPH oxidase
activation mediates a common pathway for both growth factors and ECM proteins to
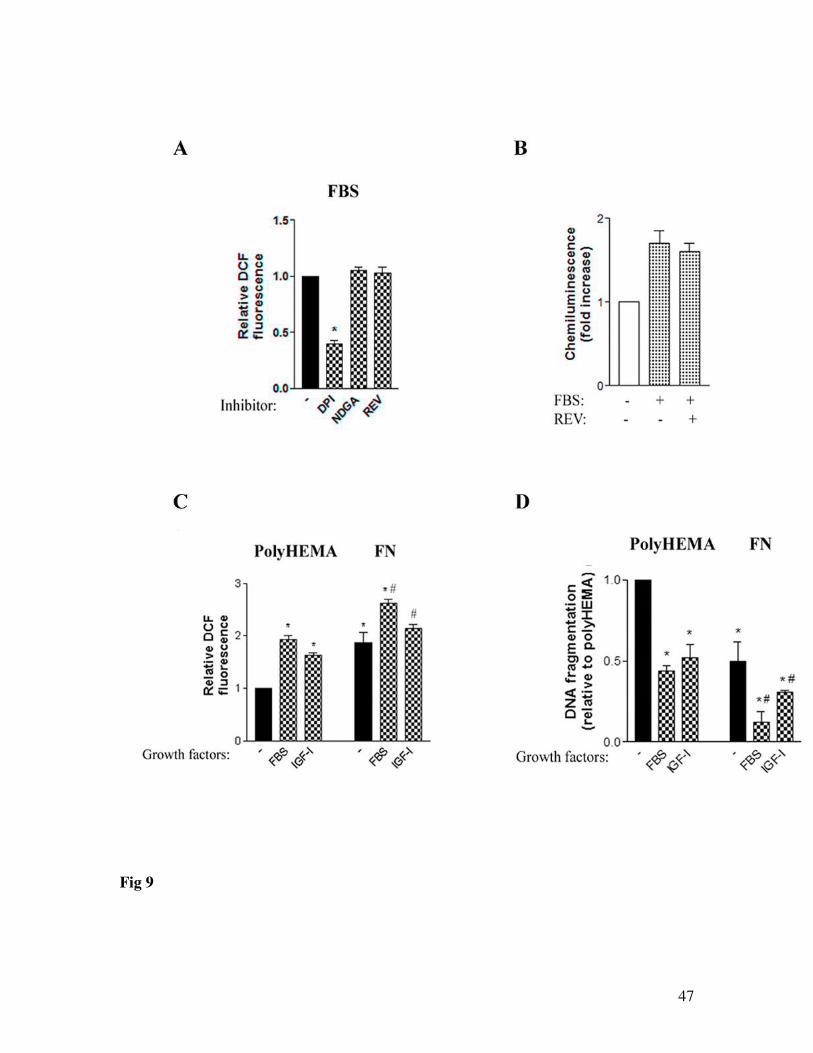
generate ROS in pancreatic cancer cells. As distinct from fibronectin, inhibition of 5-LO
(Fig. 9A, B) or PLA2 (48) had no effect on growth factors-induced cellular ROS or

20
NADPH oxidase activity. Together with the data in Figure 3, these results indicate that 5-
LO is involved in ROS production by ECM proteins but not by growth factors.
We further measured the effects of combined action of fibronectin and growth
factors (FBS or IGF-I) on the cellular ROS and death responses in MIA PaCa-2 cells. In
these experiments we used concentrations of FBS and IGF-1 that provided maximal ROS
production as we showed previously (48). Figure 9C shows that combination of
fibronectin and FBS (or IGF-I) produces greater increase in cellular ROS than growth
factors or fibronectin alone. Similarly, growth factors-induced inhibition of apoptosis was
greater in cells cultured on fibronectin than in those cultured on polyHEMA (Fig 9D).
The effects of fibronectin and growth factors on the cellular ROS and DNA
fragmentation were, however, not additive, indicating that growth factors and ECM
proteins employ overlapping mechanisms to increase ROS and inhibit DNA
fragmentation.
Nox4 is present and active in human pancreatic adenocarcinomas.
Western blot analysis shows that both Nox4 and fibronectin are present in tissue
samples from human pancreatic adenocarcinomas (Fig 10A). Furthermore, NADPH
oxidase activity is present in the membranes fractions of pancreatic cancer (Fig 10B).
Strong Nox4 immunoreactivity is present in the cancer cells (Fig 10C). There was no
staining for Nox4 in stellate, stroma or inflammatory cells (Fig 10C). Of note, mRNA for
Nox4 was detected in normal pancreas (8).

21
DISCUSSION
Our data show that the ECM proteins fibronectin and laminin, and to a lesser
extent collagen I, stimulate ROS generation in pancreatic cancer cells. Preventing cell
adhesion with polyHEMA caused a time-dependent decrease in ROS. The stimulatory
effect of ECM proteins on ROS is mediated through integrin receptors.
Several lines of evidence indicate that the NADPH oxidase Nox4 mediates ECM-
induced increases in ROS in pancreatic cancer cells: (i) fibronectin and laminin stimulate
NADPH oxidase activity; (ii) ECM-induced increases in cellular ROS are inhibited by
DPI, an inhibitor of NADPH oxidase; (iii) mitochondrial NADH dehydrogenase is not
involved in ROS stimulation by ECM proteins; (iv) Nox4 is present in pancreatic cancer
cells; and (v) Nox4 antisense oligonucleotides inhibited ROS production and NADPH
oxidase activity in cells cultured on ECM proteins.
Our results show that 5-LO is another critical mediator of ECM-induced
stimulation of ROS in pancreatic cancer cells. ECM-induced increases in cellular ROS
are inhibited by the specific inhibitors of 5-LO and by transfection with 5-LO antisense
oligonucleotides. Inhibition of 5-LO completely prevented activation of NADPH oxidase
by fibronectin, indicating that 5-LO mediates NADPH oxidase activation by this ECM
protein. Thus, one mechanism of 5-LO-induced ROS generation is through activation of
NADPH oxidase. The pharmacologic analysis indicated that in addition to NADPH
oxidase activation, 5-LO also mediates NADPH oxidase-independent pathways of ROS
generation activated by fibronectin. The nature of NADPH oxidase-independent
mechanisms of the ROS generation by 5-LO remains to be elucidated. Lipoxygenase, for
example, could generate ROS as byproducts of arachidonic acid oxidation (22). The

22
detailed signaling mechanisms linking 5-LO and NADPH oxidase are also not known,
although lipoxygenases have been found to be involved in NADPH oxidase activation in
several cell types (30; 34; 51-53).
We recently showed that detachment from ECM stimulates pancreatic cancer cells
death, and that fibronectin and laminin markedly protect pancreatic cancer cells from
death (47). We also demonstrated that the pro-survival effects of ECM proteins in
pancreatic cancer cells are mediated through mechanisms different from those operating
in normal cells. The present study demonstrates that this pro-survival effect of ECM
proteins is mediated through ROS. Indeed, inhibitors of 5-LO and NADPH oxidase, or
the superoxide scavenger tiron, all inhibited ROS and stimulated caspase activity and
DNA fragmentation in cells cultured on fibronectin or laminin. Furthermore, as shown by
antisense transfections, 5-LO and Nox4 are key mediators of the anti-apoptotic effects of
ECM proteins in pancreatic cancer cells.
5-LO has been shown to mediate proliferation and survival of pancreatic cancer
cells (11-13; 45). Our data suggest that the pro-survival effect of 5-LO is mediated, at
least in part, through ROS stimulation by ECM proteins.
Our results in the present study and in Ref. (48) show that the two major
environmental pro-survival factors, ECM proteins and growth factors, protect pancreatic
cancer cells from death through a common pathway, namely, by stimulating cellular
ROS. Activation of NADPH oxidase, in particular Nox4, is a common mechanism
through which both ECM proteins and growth factors stimulate ROS production and
inhibit apoptosis in pancreatic cancer cells. The mechanisms utilized by ECM proteins
and growth factors to stimulate NADPH oxidase, however, differ. Our results indicate

23
that NADPH oxidase activation by fibronectin is fully mediated by 5-LO, whereas
growth factors activate NADPH oxidase independently of 5-LO. The results of this study
and Ref. (48) show that 5-LO is not involved in ROS stimulation by growth factors.
Thus, ECM proteins and growth factors generate ROS through overlapping but not
identical mechanisms. Combination of both growth factors and ECM proteins, produce
greater increase in ROS and inhibition of apoptosis than growth factors or ECM proteins
alone. However, the effects of growth factors and fibronectin on ROS and apoptosis were
not additive because the pathways they activated overlap.
We found that Nox4 is present in tissues from human pancreatic
adenocarcinomas; furthermore, the membrane fractions of pancreatic adenocarcinomas
display NADPH oxidase activity. Pancreatic adenocarcinomas are notoriously
desmophilic (16; 29; 49; 54); thus, the ECM proteins produced by the stromal cells
surrounding the cancer cells may facilitate the survival of cancer cells through their
effects on Nox4 in the cancer cells.
In conclusion, we have shown that fibronectin and laminin stimulate intracellular
ROS in pancreatic cancer cells through activation of NADPH oxidase, as well as through
NADPH-oxidase independent pathways. 5-LO is an upstream mediator of both pathways.
While growth factors also stimulate ROS, they activate NADPH oxidase independent of
5-LO. Nox4 is the common mediator through which both ECM proteins and growth
factors stimulate ROS. ECM-induced ROS stimulation is an important mechanism of the
pro-survival, anti-apoptotic effect of ECM proteins in pancreatic cancer cells. The results
indicate that 5-LO and Nox4 are key molecules mediating pancreatic cancer cells’

24
resistance to apoptosis and thus represent potential targets for therapeutic strategies to
treat pancreatic adenocarcinoma.
$ This work was supported by the Department of Veterans Affairs Merit Review and the
NIH grant DK-59926 to A. S. G. The authors thank Dr. Ilya Gukovsky for advice and
discussion, and Mohammad Shahsahebi and Yoon Jung for their help in the preparation
of the manuscript.

25
TABLE 1: Inhibitors of xanthine oxidase, 12-lipoxygenase (12-LO) and nitric oxide synthase did not affect cellular ROS in MIA PaCa-2 cells.
Changes in DCF, %Inhibitor Target polyHEMA Fibronectin Laminin
- - 100 100 100Allopurinol, 1 mM xanthine oxidase 101±2 113±4 102±7Baicalein, 20 µM 12-LO 96±5 100±4 97±2L-NAME, 100 µM Nitric oxide
synthase107±7 96±2 101±5
MIA PaCa-2 cells were cultured for 48 h on polyHEMA, fibronectin or laminin without serum and in the presence or absence of the inhibitors of ROS-generating systems. Changes in intracellular ROS were measured by FACS analysis in cells labeled with DCFH-DA and normalized on that in cells cultured on polyHEMA or ECM proteins without inhibitors. Values are means ± SEM from 3 independent experiments.

26
Reference List
1. Ago T, Kitazono T, Ooboshi H, Iyama T, Han YH, Takada J, Wakisaka M,
Ibayashi S, Utsumi H and Iida M. Nox4 as the major catalytic component of an
endothelial NAD(P)H oxidase. Circulation 109: 227-233, 2004.
2. Arao S, Masumoto A and Otsuki M. Beta1 integrins play an essential role in
adhesion and invasion of pancreatic carcinoma cells. Pancreas 20: 129-137, 2000.
3. Babior BM. NADPH oxidase: an update. Blood 93: 1464-1476, 1999.
4. Babior BM, Lambeth JD and Nauseef W. The neutrophil NADPH oxidase. Arch
Biochem Biophys 397: 342-344, 2002.
5. Bokoch GM and Knaus UG. NADPH oxidases: not just for leukocytes anymore!
Trends Biochem Sci 28: 502-508, 2003.
6. Borges F, Fernandes E and Roleira F. Progress towards the discovery of xanthine
oxidase inhibitors. Curr Med Chem 9: 195-217, 2002.
7. Brar SS, Corbin Z, Kennedy TP, Hemendinger R, Thornton L, Bommarius B,
Arnold RS, Whorton AR, Sturrock AB, Huecksteadt TP, Quinn MT,
Krenitsky K, Ardie KG, Lambeth JD and Hoidal JR. NOX5 NAD(P)H oxidase

27
regulates growth and apoptosis in DU 145 prostate cancer cells. Am J Physiol Cell
Physiol 285: C353-C369, 2003.
8. Cheng G, Cao Z, Xu X, van Meir EG and Lambeth JD. Homologs of gp91phox:
cloning and tissue expression of Nox3, Nox4, and Nox5. Gene 269: 131-140, 2001.
9. Chiarugi P. Reactive oxygen species as mediators of cell adhesion. Ital J Biochem
52: 28-32, 2003.
10. Chiarugi P and Cirri P. Redox regulation of protein tyrosine phosphatases during
receptor tyrosine kinase signal transduction. Trends Biochem Sci 28: 509-514, 2003.
11. Ding XZ, Hennig R and Adrian TE. Lipoxygenase and cyclooxygenase
metabolism: new insights in treatment and chemoprevention of pancreatic cancer.
Mol Cancer 2: 10, 2003.
12. Ding XZ, Iversen P, Cluck MW, Knezetic JA and Adrian TE. Lipoxygenase
inhibitors abolish proliferation of human pancreatic cancer cells. Biochem Biophys
Res Commun 261: 218-223, 1999.
13. Ding XZ, Kuszynski CA, El Metwally TH and Adrian TE. Lipoxygenase
inhibition induced apoptosis, morphological changes, and carbonic anhydrase
expression in human pancreatic cancer cells. Biochem Biophys Res Commun 266:
392-399, 1999.

28
14. Folkman J and Moscona A. Role of cell shape in growth control. Nature 273:
345-349, 1978.
15. Green DR. Death and NF-kappaB in T cell activation: life at the edge. Mol Cell 11:
551-552, 2003.
16. Gress TM, Muller-Pillasch F, Lerch MM, Friess H, Buchler M and Adler G.
Expression and in-situ localization of genes coding for extracellular matrix proteins
and extracellular matrix degrading proteases in pancreatic cancer. Int J Cancer 62:
407-413, 1995.
17. Gukovskaya AS, Vaquero E, Zaninovic V, Gorelick FS, Lusis AJ, Brennan
ML, Holland S and Pandol SJ. Neutrophils and NADPH oxidase mediate
intrapancreatic trypsin activation in murine experimental acute pancreatitis.
Gastroenterology 122: 974-984, 2002.
18. Hall PA, Coates P, Lemoine NR and Horton MA. Characterization of integrin
chains in normal and neoplastic human pancreas. J Pathol 165: 33-41, 1991.
19. Hempel SL, Buettner GR, O'Malley YQ, Wessels DA and Flaherty DM.
Dihydrofluorescein diacetate is superior for detecting intracellular oxidants:
comparison with 2',7'-dichlorodihydrofluorescein diacetate, 5(and 6)-carboxy-2',7'-
dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic Biol
Med 27: 146-159, 1999.

29
20. Honore S, Kovacic H, Pichard V, Briand C and Rognoni JB. Alpha2beta1-
integrin signaling by itself controls G1/S transition in a human adenocarcinoma cell
line (Caco-2): implication of NADPH oxidase-dependent production of ROS. Exp
Cell Res 285: 59-71, 2003.
21. Janiszewski M, Souza HP, Liu X, Pedro MA, Zweier JL and Laurindo FR.
Overestimation of NADH-driven vascular oxidase activity due to lucigenin
artifacts. Free Radic Biol Med 32: 446-453, 2002.
22. Katsuki H and Okuda S. Arachidonic acid as a neurotoxic and neurotrophic
substance. Prog Neurobiol 46: 607-636, 1995.
23. Kensler T, Guyton K, Egner P, McCarthy T, Lesko S and Akman S. Role of
reactive intermediates in tumor promotion and progression. Prog Clin Biol Res 391:
103-116, 1995.
24. Kolko M, Rodriguez de Turco EB, Diemer NH and Bazan NG. Neuronal
damage by secretory phospholipase A2: modulation by cytosolic phospholipase A2,
platelet-activating factor, and cyclooxygenase-2 in neuronal cells in culture.
Neurosci Lett 338: 164-168, 2003.
25. Lambeth JD. NOX enzymes and the biology of reactive oxygen. Nat Rev Immunol
4: 181-189, 2004.

30
26. Lambeth JD, Cheng G, Arnold RS and Edens WA. Novel homologs of
gp91phox. Trends Biochem Sci 25: 459-461, 2000.
27. Li Y and Trush MA. Diphenyleneiodonium, an NAD(P)H oxidase inhibitor, also
potently inhibits mitochondrial reactive oxygen species production. Biochem
Biophys Res Commun 253: 295-299, 1998.
28. Li Y, Zhu H, Kuppusamy P, Roubaud V, Zweier JL and Trush MA. Validation
of lucigenin (bis-N-methylacridinium) as a chemilumigenic probe for detecting
superoxide anion radical production by enzymatic and cellular systems. J Biol
Chem 273: 2015-2023, 1998.
29. Lohr M, Trautmann B, Gottler M, Peters S, Zauner I, Maillet B and Kloppel
G. Human ductal adenocarcinomas of the pancreas express extracellular matrix
proteins. Br J Cancer 69: 144-151, 1994.
30. Luchtefeld M, Drexler H and Schieffer B. 5-Lipoxygenase is involved in the
angiotensin II-induced NAD(P)H-oxidase activation. Biochem Biophys Res
Commun 308: 668-672, 2003.
31. Maccarrone M, Melino G and Finazzi-Agro A. Lipoxygenases and their
involvement in programmed cell death. Cell Death Differ 8: 776-784, 2001.

31
32. Mahadev K, Motoshima H, Wu X, Ruddy JM, Arnold RS, Cheng G, Lambeth
JD and Goldstein BJ. The NAD(P)H oxidase homolog Nox4 modulates insulin-
stimulated generation of H2O2 and plays an integral role in insulin signal
transduction. Mol Cell Biol 24: 1844-1854, 2004.
33. Martinez GR, Loureiro AP, Marques SA, Miyamoto S, Yamaguchi LF, Onuki
J, Almeida EA, Garcia CC, Barbosa LF, Medeiros MH and Di Mascio P.
Oxidative and alkylating damage in DNA. Mutat Res 544: 115-127, 2003.
34. Nardi M, Feinmark SJ, Hu L, Li Z and Karpatkin S. Complement-independent
Ab-induced peroxide lysis of platelets requires 12-lipoxygenase and a platelet
NADPH oxidase pathway. J Clin Invest 113: 973-980, 2004.
35. Ogden JE and Moore PK. Inhibition of nitric oxide synthase--potential for a novel
class of therapeutic agent? Trends Biotechnol 13: 70-78, 1995.
36. Pidgeon GP, Tang K, Cai YL, Piasentin E and Honn KV. Overexpression of
platelet-type 12-lipoxygenase promotes tumor cell survival by enhancing
alpha(v)beta(3) and alpha(v)beta(5) integrin expression. Cancer Res 63: 4258-4267,
2003.
37. Rajagopalan S, Kurz S, Munzel T, Tarpey M, Freeman BA, Griendling KK
and Harrison DG. Angiotensin II-mediated hypertension in the rat increases

32
vascular superoxide production via membrane NADH/NADPH oxidase activation.
Contribution to alterations of vasomotor tone. J Clin Invest 97: 1916-1923, 1996.
38. Sauer H, Wartenberg M and Hescheler J. Reactive oxygen species as
intracellular messengers during cell growth and differentiation. Cell Physiol
Biochem 11: 173-186, 2001.
39. Simon HU, Haj-Yehia A and Levi-Schaffer F. Role of reactive oxygen species
(ROS) in apoptosis induction. Apoptosis 5: 415-418, 2000.
40. Sorescu D, Somers MJ, Lassegue B, Grant S, Harrison DG and Griendling
KK. Electron spin resonance characterization of the NAD(P)H oxidase in vascular
smooth muscle cells. Free Radic Biol Med 30: 603-612, 2001.
41. Steele VE, Holmes CA, Hawk ET, Kopelovich L, Lubet RA, Crowell JA,
Sigman CC and Kelloff GJ. Lipoxygenase inhibitors as potential cancer
chemopreventives. Cancer Epidemiol Biomarkers Prev 8: 467-483, 1999.
42. Suh YA, Arnold RS, Lassegue B, Shi J, Xu X, Sorescu D, Chung AB,
Griendling KK and Lambeth JD. Cell transformation by the superoxide-
generating oxidase Mox1. Nature 401: 79-82, 1999.

33
43. Tennant CM, Seale JP and Temple DM. Effects of a 5-lipoxygenase inhibitor,
REV-5901, on leukotriene and histamine release from human lung tissue in-vitro. J
Pharm Pharmacol 39: 309-311, 1987.
44. Thannickal VJ and Fanburg BL. Reactive oxygen species in cell signaling. Am J
Physiol Lung Cell Mol Physiol 279: L1005-L1028, 2000.
45. Tong WG, Ding XZ, Witt RC and Adrian TE. Lipoxygenase inhibitors attenuate
growth of human pancreatic cancer xenografts and induce apoptosis through the
mitochondrial pathway. Mol Cancer Ther 1: 929-935, 2002.
46. Torres M and Forman HJ. Redox signaling and the MAP kinase pathways.
Biofactors 17: 287-296, 2003.
47. Vaquero EC, Edderkaoui M, Nam KJ, Gukovsky I, Pandol SJ and
Gukovskaya AS. Extracellular matrix proteins protect pancreatic cancer cells from
death via mitochondrial and nonmitochondrial pathways. Gastroenterology 125:
1188-1202, 2003.
48. Vaquero EC, Edderkaoui M, Pandol SJ, Gukovsky I and Gukovskaya AS.
Reactive oxygen species produced by NAD(P)H oxidase inhibit apoptosis in
pancreatic cancer cells. J Biol Chem 279: 34643-34654, 2004.

34
49. Watanabe I, Hasebe T, Sasaki S, Konishi M, Inoue K, Nakagohri T, Oda T,
Mukai K and Kinoshita T. Advanced pancreatic ductal cancer: fibrotic focus and
beta-catenin expression correlate with outcome. Pancreas 26: 326-333, 2003.
50. Werner E and Werb Z. Integrins engage mitochondrial function for signal
transduction by a mechanism dependent on Rho GTPases. J Cell Biol 158: 357-368,
2002.
51. Woo CH, Kim BC, Kim KW, Yoo MH, Eom YW, Choi EJ, Na DS and Kim
JH. Role of cytosolic phospholipase A(2) as a downstream mediator of Rac in the
signaling pathway to JNK stimulation. Biochem Biophys Res Commun 268: 231-
236, 2000.
52. Woo CH, Yoo MH, You HJ, Cho SH, Mun YC, Seong CM and Kim JH.
Transepithelial migration of neutrophils in response to leukotriene B4 is mediated
by a reactive oxygen species-extracellular signal-regulated kinase-linked cascade. J
Immunol 170: 6273-6279, 2003.
53. Woo CH, You HJ, Cho SH, Eom YW, Chun JS, Yoo YJ and Kim JH.
Leukotriene B(4) stimulates Rac-ERK cascade to generate reactive oxygen species
that mediates chemotaxis. J Biol Chem 277: 8572-8578, 2002.

35
54. Yen TW, Aardal NP, Bronner MP, Thorning DR, Savard CE, Lee SP and Bell
RH, Jr. Myofibroblasts are responsible for the desmoplastic reaction surrounding
human pancreatic carcinomas. Surgery 131: 129-134, 2002.
55. Zhang Y and Chen F. Reactive oxygen species (ROS), troublemakers between
nuclear factor-kappaB (NF-kappaB) and c-Jun NH(2)-terminal kinase (JNK).
Cancer Res 64: 1902-1905, 2004.

36
Figure legends:
Figure 1. ECM proteins increase intracellular ROS in pancreatic cancer cells.MIA PaCa-2 (A), CAPAN-1 (B) and PANC-1 (C) cells were cultured without serum for 48 h on polyHEMA or ECM proteins: fibronectin, laminin, collagen I. Changes in intracellular ROS were measured by FACS analysis in cells labeled with redox-sensitive dye DCFH-DA. Histograms in (A-C) are representative of at least 3 independent experiments. For quantitative analysis (D) DCF fluorescence was normalized on that in cells cultured on polyHEMA. Values are means ± SEM (n=3). * p < 0.05 vs cells cultured on polyHEMA. In (E) MIA PaCa-2 cells were pre-incubated for 1 hour at 4°C with or without monoclonal anti-α5 integrin antibody (10 µg/ml). DCF fluorescence was normalized on that in cells preincubated without antibody. Values are means ±SEM (n=3).* p < 0.05 vs. cells without pretreatment with antibody
Figure 2. Cells detached from ECM lose the ability to maintain intracellular ROS. MIA PaCa-2 cells were cultured for indicated time without serum on polyHEMA or fibronectin. DCF fluorescence was normalized on that in cells cultured for 3 hours. At 3 hours, there was no difference in DCF fluorescence in cells cultured on polyHEMA and fibronectin. Values are means ± SEM from at least 3 independent experiments for each time point.
Figure 3. Fibronectin and laminin induce ROS production through 5-LO and NAD(P)H oxidase mediated pathways.MIA PaCa-2 (A, C) and CAPAN-1 (B, D) cells were cultured without serum for 48 h on polyHEMA, fibronectin (FN) or laminin (LN) and in the presence or absence of the inhibitors of ROS-generating systems: NADPH oxidase inhibitor DPI (15 µM); PLA2 inhibitor AACOCF3 (10µM); 5-LO inhibitors nordihydroguaiaretic acid (NDGA, 10 µM) and REV-5901 (REV, 15 µM), and superoxide scavenger tiron (10 mM). In (A, B) Changes in intracellular ROS were measured by FACS analysis in cells labeled with DCFH-DA. In (C, D) superoxide production in the absence and presence of 100 µM NADPH was measured by lucigenin derived chemiluminescence in total cell homogenate. Both DCF fluorescence (A, B) and luminescence (C, D) were normalized on that in cells cultured on polyHEMA without inhibitors. Values are means ± SEM from at least 3 independent experiments. * p < 0.05 vs. cells on polyHEMA without inhibitors; # p < 0.05 vs. cells on the same ECM protein without inhibitors.
Figure 4. Transfection with NOX4 antisense oligonucleotides inhibits cellular ROS and NADPH oxidase activity in cells cultured on fibronectin.MIA PaCa-2 (A-C) or PANC-1 (D, E) were transfected with antisense oligonucleotides 1 (AS1), antisense oligonucleotides 2 (AS2), or scrambled (S) Nox4 oligonucleotides, and cultured for 72 hours without serum on fibronectin. In (A, D) Nox4 protein levels were measured in transfected cells by immunoblotting. Blots were stripped and reprobed for actin to confirm equal loading. The densitometric ratios of the intensity of Nox4 band (normalized to actin) between cells transfected with AS and scrambled oligonucleotides were 0.3 in MIA PaCa-2 cells transfected with AS1 (A), and 0.5 and 0.6 in PANC-1 cells transfected, respectively, with AS1 and AS2 (D). Changes in intracellular ROS (B, E) were measured by FACS analysis in cells labeled with DCFH-DA. Histograms (B) are representative of 3 independent experiments. Superoxide production (C) was measured by lucigenin derived chemiluminescence in membrane fractions in the presence of 100 µM NADPH. The chemiluminescence values were normalized on those in

37
cells transfected with scrambled oligonucleotides. Values are means ± SEM from 3 independent experiments. * p < 0.05 vs. cells transfected with scrambled oligonucleotides.
Figure 5. Transfection with 5-LO antisense oligonucleotides inhibits cellular ROS and NADPH oxidase activity in cells cultured on fibronectin.MIA PaCa-2 (A-C) or PANC-1 (D, E) were transfected with antisense oligonucleotides 1 (AS1), antisense oligonucleotides 2 (AS2), or scrambled (S) 5-LO oligonucleotides, and cultured for 72 hours without serum on fibronectin. Protein levels of 5-LO (A, D) were measured in transfected cells by immunoblotting. Blots were stripped and reprobed for actin to confirm equal loading. The densitometric ratios of the intensity of 5-LO band (normalized to actin) between cells transfected with AS and scrambled oligonucleotides were 0.3 in MIA PaCa-2 cells transfected with AS1 (A), and 0.4 in PANC-1 cells transfected with either AS1 or AS2 (D). Changes in intracellular ROS (B, E) were measured by FACS analysis in cells labeled with DCFH-DA. Histograms (B) are representative of 3 independent experiments. Superoxide production (C) was measured by lucigenin derived chemiluminescence in membrane fractions in the presence of 100 µM NADPH. The chemiluminescence values were normalized on those in cells transfected with scrambled oligonucleotides. Values are means ± SEM from 3 independent experiments. * p < 0.05 vs. cells transfected with scrambled oligonucleotides.
Figure 6. Fibronectin and laminin stimulate ROS generation in MIA PaCa-2 Rho0 cells depleted of mtDNA.MIA PaCa-2 Rho0 cells depleted of mtDNA were generated by incubating wild type cells for >6 weeks with 100 ng/ml of ethidium bromide. The absence of mtDNA was confirmed by PCR and western blot as described in Methods and in (48). MIA PaCa-2 Rho0 (A) or parental MIA PaCa-2 (B) cells were cultured for 48 h without serum on polyHEMA, fibronectin, or laminin in the presence or absence of the mitochondrial complex inhibitor rotenone (1 µM) (B). Changes in intracellular ROS were measured by FACS analysis in cells labeled with DCFH-DA. Histogram is representative of at least 3 independent experiments. Values were normalized on those in cells cultured in the same conditions on polyHEMA. Values are means ± SEM from 3 independent experiments.
Figure 7. Inhibition of ROS stimulates internucleosomal DNA fragmentation and decreases survival in pancreatic cancer cells.MIA PaCa-2 (A, C, D) and CAPAN-1 (B) cells were cultured without serum for 48h on polyHEMA, fibronectin or laminin in the presence or absence of NADPH oxidase inhibitor DPI (15 µM); 5-LO inhibitor REV 5901(REV, 15 µM), superoxide scavenger tiron (10 mM) or the broad-spectrum caspase inhibitor zVAD-fmk (100 µM). Internucleosomal DNA fragmentation (A, B) was measured using Cell Death Detection ELISA kit. Phosphatidylserine externalization (C) was measured in cells labeled with AnV/PI and analyzed by FACS. An-/PI-
cells were considered as live cells; An+/PI- and An+/PI+ cells, as cells dying through apoptosis and apoptosis associated with secondary necrosis (dead cells). Caspase-3 activity was measured in cell lysates by fluorogenic assay (D). Values were normalized on that in cells cultured on polyHEMA without inhibitors. Values are means ± SEM from at least 4 independent experiments. In (A-C) * p < 0.05 vs. cells cultured on the same condition without inhibitors. In (D) * p < 0.05 vs. cells cultured on polyHEMA, # p < 0.05 vs. cells cultured on fibronectin protein without inhibitors.

38
Figure 8. Transfection with antisense Nox4 or 5-LO stimulates apoptosis in pancreatic cancer cells.MIA PaCa-2 (A, C-E) and PANC-1 (B) were transfected with antisenses 1 or scrambled Nox4 and 5-LO oligonucleotides, and cultured for 72 hours without serum on fibronectin. Internucleosomal DNA fragmentation (A-C) was measured using Cell Death Detection ELISA kit. Phosphatidylserine externalization (D) was measured in cells labeled with AnV/PI and analyzed by FACS. An+/PI- and An+/PI+ cells were considered as dead cells. Caspase-3 activity (E, F) was measured in cell lysates by fluorogenic assay. Values are means ± SEM from at least 3 independent experiments. In (A-C, E, F), values were normalized to those in cells transfected with scrambled oligonucleotides. * p < 0.05 vs. cells transfected with scrambled oligonucleotides.
Figure 9. Mechanisms of ROS production and inhibition of apoptosis by growth factors and ECM proteins partially overlap.MIA PaCa-2 cells were cultured for 48 h on polyHEMA (A-D) or fibronectin (C, D) with or without 15% FBS or 100 ng/ml of IGF-I and in the presence or absence of the inhibitors of ROS-generating systems: NADPH oxidase inhibitor DPI (15 µM); 5-LO inhibitors nordihydroguaiaretic acid (NDGA, 10 µM) and REV-5901 (REV, 15 µM). Changes in intracellular ROS (A, C) were measured by FACS analysis in cells labeled with DCFH-DA.Superoxide production (B) was measured in the presence of 100 µM NADPH by lucigenin derived chemiluminescence in total cell homogenate. Internucleosomal DNA fragmentation (D) was measured using Cell Death Detection ELISA kit. DCF fluorescence (A, C), luminescence (B) and DNA fragmentation (D) were normalized on that in cells cultured on polyHEMA without inhibitors and without growth factors. Values are means ± SEM from at least 3 independent experiments. * p < 0.05 vs. cells cultured on polyHEMA with (A) or without (B-D) growth factors)and without inhibitors; # p < 0.05 vs. cells cultured in the same condition on polyHEMA.
Figure 10. Nox4 and NADPH oxidase activity are present in pancreatic cancer tissues.Western blot analysis of Nox4 and fibronectin (A); superoxide production (B) and immunofluorescence of Nox4 (C) were measured in the tissue samples of pancreatic adenocarcinoma of individual patients. Superoxide production was measured in the membrane fractions in the absence and presence of 100 µM NADPH by lucigenin-derived chemiluminescence. Values are means ± SEM, n=4. (C): Pancreatic cancer tissue sections were stained with primary antibody against Nox4 and FITC-conjugated secondary antibody (left and middle panels) or with secondary antibody only (right panel). In the middle panel non-FITC fluorescence was subtracted from the image in the left panel. (A) and (C): Western blot and images are representative of samples obtained from 3 patients.

39
Fig 1

40
Fig 2

41
Fig 3

42
Fig 4

43
Fig 5

44
Fig 6

45
Fig 7

46
Fig 8
47
Fig 9

48
Fig 10




















