
Expression of the surface antigen A2B7 in adult and developing honeybee olfactory pathway
15
~ Pergamon Int. J. Devl Neuroscience, Vol. 14, No. 7/8, pp. 997-1011, 1996 Copyright © 1996 ISDN Published by Elsevier Science Ltd. Printed in Great Britain 0736-5748/96 $15.00+ 0.00 PII: S0736-5748(96)00065-2 EXPRESSION OF THE SURFACE ANTIGEN A2B7 IN ADULT AND DEVELOPING HONEYBEE OLFACTORY PATHWAY JEAN GASCUEL, FRANCK BROUILLARD, CHRISTIAN PAPIN and CLAUDINE MASSON* Laboratoire de Neurobiologie Compar6e des Invertdbrds, INRA-CNRS (URA 1190), BP 23, F-91440 Cedex, Bures-sur-Yvette, France Abstract--In order to identify molecules involved in the development of the honeybee olfactory pathway, hybridoma technology has been used. Among different cell lines, A2B7 has been selected. It produces a specific antibody for a surface glycoprotein of 91 kDa. This protein is mainly expressed by both the antennal receptor cells and mushroom body neurons. Based on (i) the spatio-temporal pattern of expression during pupal development; (ii) the cell surface location of the antigen; and (iii) the partial molecular characterization of the antigen, a putative role for this protein in axonal fasciculation and guidance is discussed. Copyright © 1996 ISDN. Key words: development, olfactory pathway, sensory nerve, mushroom body, insect, honeybee, cell-cell interactions, axon guidance, axon fasciculation. The construction of the adult nervous system is basically dependent on the ability of individual neurons to navigate in search of the appropriate target, with which synapses are finally formed. During these developmental processes, neurites have to orient in a variety of cellular and molecular environments, which provide the positional information they need. Selective fasciculation of axons is one strategy by which neurons navigate. Since the existence of the growth cone has been dem- onstrated, 39 and its role as sensory and motor unit for the growing neurite has been established, the question of the molecular signals involved in axonal guidance has represented a central challenge in developmental neurobiology (for review, see Ref. 23). Recently, a great step in the understanding of axonal guidance mechanisms has been achieved with the identification of cell surface molecules or of diffusible molecules involved in growth c o n e g u i d a n c e . 4,lv'z°-zz'24,3s'47 Faking advantage of hybridoma technology and molecular genetics, as well as the presence of identified neurons, the molecular basis of fasciculation and axon guidance in insects is being extensively examined during embryogenesis (for review, see Ref. 17). By contrast, the question of how the adult insect CNS, which generates complex behavior, is established in later developmental and post-embryonic stages, is still poorly documented, especially from a molecular point of view. Considering the importance of the architecture of the olfactory neuronal network in the processing of olfactory cues, the understanding of the mechanisms underlying its development appears to be of crucial importance. In such a context, the search for the rules governing the construction of the insect olfactory pathway seems to be relevant. The sensory axon terminals converge onto neuropilar structures, the olfactory glomeruli, where they synapse with antennal lobe neurons (Fig. 1) (for review see Ref. 26). These glomeruli are identifiable and invariant neuropilar structures 2'6'42 thought to represent functional units. This seems to be supported by data collected in Drosophila, 4~ and in vertebrates where olfactory neurons expressing one type of odor receptor protein project to a small and distinct set of glomeruli. 4°'5° 3-he results gained so far in the honeybee 1'25'12concerning the role of sensory axons in glomeruli formation are consistent with the bulk of data accumulated in the moth Manduca s e x t a . 35-37'49 Interestingly, the sensory afferents that have been demonstrated as crucial for glomeruli devel- opment ~'12'37'49 a r e also capable of specifying the type of glomerular organization that is dependent *To whom all correspondence should be addressed. 997
-
Upload
univ-paris5 -
Category
Documents
-
view
0 -
download
0
Transcript of Expression of the surface antigen A2B7 in adult and developing honeybee olfactory pathway

~ Pergamon Int. J. Devl Neuroscience, Vol. 14, No. 7/8, pp. 997-1011, 1996 Copyright © 1996 ISDN
Published by Elsevier Science Ltd. Printed in Great Britain 0736-5748/96 $15.00 + 0.00
PII : S0736-5748(96)00065-2
E X P R E S S I O N O F T H E S U R F A C E A N T I G E N A2B7 I N A D U L T A N D
D E V E L O P I N G H O N E Y B E E O L F A C T O R Y P A T H W A Y
JEAN GASCUEL, FRANCK BROUILLARD, CHRISTIAN PAPIN and CLAUDINE MASSON*
Laboratoire de Neurobiologie Compar6e des Invertdbrds, INRA-CNRS (URA 1190), BP 23, F-91440 Cedex, Bures-sur-Yvette, France
Abstract--In order to identify molecules involved in the development of the honeybee olfactory pathway, hybridoma technology has been used. Among different cell lines, A2B7 has been selected. It produces a specific antibody for a surface glycoprotein of 91 kDa. This protein is mainly expressed by both the antennal receptor cells and mushroom body neurons. Based on (i) the spatio-temporal pattern of expression during pupal development; (ii) the cell surface location of the antigen; and (iii) the partial molecular characterization of the antigen, a putative role for this protein in axonal fasciculation and guidance is discussed. Copyright © 1996 ISDN.
Key words: development, olfactory pathway, sensory nerve, mushroom body, insect, honeybee, cell-cell interactions, axon guidance, axon fasciculation.
The construction of the adult nervous system is basically dependent on the ability of individual neurons to navigate in search of the appropriate target, with which synapses are finally formed. During these developmental processes, neurites have to orient in a variety of cellular and molecular environments, which provide the positional information they need. Selective fasciculation of axons is one strategy by which neurons navigate. Since the existence of the growth cone has been dem- onstrated, 39 and its role as sensory and motor unit for the growing neurite has been established, the question of the molecular signals involved in axonal guidance has represented a central challenge in developmental neurobiology (for review, see Ref. 23). Recently, a great step in the understanding of axonal guidance mechanisms has been achieved with the identification of cell surface molecules or of diffusible molecules involved in growth c o n e g u i d a n c e . 4,lv'z°-zz'24,3s'47
Faking advantage of hybridoma technology and molecular genetics, as well as the presence of identified neurons, the molecular basis of fasciculation and axon guidance in insects is being extensively examined during embryogenesis (for review, see Ref. 17). By contrast, the question of how the adult insect CNS, which generates complex behavior, is established in later developmental and post-embryonic stages, is still poorly documented, especially from a molecular point of view.
Considering the importance of the architecture of the olfactory neuronal network in the processing of olfactory cues, the understanding of the mechanisms underlying its development appears to be of crucial importance. In such a context, the search for the rules governing the construction of the insect olfactory pathway seems to be relevant. The sensory axon terminals converge onto neuropilar structures, the olfactory glomeruli, where they synapse with antennal lobe neurons (Fig. 1) (for review see Ref. 26). These glomeruli are identifiable and invariant neuropilar s t r u c t u r e s 2'6'42 thought to represent functional units. This seems to be supported by data collected in Drosophila, 4~ and in vertebrates where olfactory neurons expressing one type of odor receptor protein project to a small and distinct set of glomeruli. 4°'5°
3-he results gained so far in the honeybee 1'25'12 concerning the role of sensory axons in glomeruli formation are consistent with the bulk of data accumulated in the moth Manduca s e x t a . 35-37'49
Interestingly, the sensory afferents that have been demonstrated as crucial for glomeruli devel- opment ~'12'37'49 a r e also capable of specifying the type of glomerular organization that is dependent
*To whom all correspondence should be addressed.
997
998 J. Gascuel et al.
on the sexual origin of the receptor cells. 44 These data taken together suggest that signals given by the sensory neurons are necessary for the specific axonal guidance and the induction of specific glomerular organization. Moreover, in the honeybee, different types of sensory axon are differ- entially targeted suggesting a specific mechanism for axon guidance. ~] This feature addresses the question of the mechanisms involved in the guidance of specific axons to specific glomeruli.
Using hybridoma technology we attempted to identify candidate molecules involved in the development of the honeybee brain during metamorphosis. In this paper we report results obtained with A2B7, one of the selected hybridoma cell lines, which produces a specific antibody for a 91 kDa surface glycoprotein. This protein is mainly expressed by both antennal receptor cells and mushroom body neurons whenever and wherever their axons fasciculate. Preliminary results of this work have been presented elsewhere. ~5
A
Antennae
SA ...
T1
Antennal lobe
D(71)
I (8)
~..:...-
i T3
P(~
Mushroom bodies
AGT
G T
1 ~tvc
.... 4: -.'-'::~ .... C N
,@ • . . . 1 - .
- q)
B C

A2B7 expression in honeybee olfactory pathway 999
E X P E R I M E N T A L P R O C E D U R E S
G~ .~,mration of monoclonal antibodies
Fwenty bra ins of adu l t honeybees were dissected in sterile saline and h o m o g e n i z e d in p h o s p h a t e -
buffered saline (PBS) with 5 m M E D T A , 20 m g / m l p h e n y l m e t h y l s u l f o n y l f luor ide and a cockta i l
o t p ro tease inh ib i tors (1 /~g/ml of an t ipa in , chymos ta t in , leupept in , peps ta t in , N-p- tosyl -c- lys ine
c h i o r o m e t h y l ke tone , and N- tosy l - c -pheny la l an ine c h l o r o m e t h y l ketone) . The h o m o g e n a t e was
ce~ltrifuged at 1000 g (4°C) for 10 min. The s u p e r n a t a n t was then cen t r i fuged at 200,000 g (4°C) for
1 hr. The m e m b r a n e pellet was r e suspended in PBS. F o r the first i m m u n i z a t i o n , in ject ion was m a d e
with comple te F r e u n d ' s ad juvant . F o r the next three inject ions, i ncomple t e F r e u n d ' s a d j u v a n t was
used. I n t r a p e r i t o n e a l inject ions were p e r f o r m e d with at least 3 week in tervals using B A L B / c mouse .
The last inject ion was p e r f o r m e d wi thou t ad juvan t , 3 days before the fusion. Spleen cells were fused
wilh Sp2/0 cell line.
Screening of hybridomas and immunostaining
H y b r i d o m a supe rna t an t s were screened by i m m u n o s t a i n i n g on 7 / , m f rozen sect ions of h o n e y b e e
b ra in fixed with ace tone and s tored at - 8 0 ° C . Af ter b lockage with PBS con ta in ing 3% bov ine
se rum a l b u m i n (BSA), sect ions were i ncuba t ed for 2 hr in h y b r i d o m a supe rna t an t , washed three
t imes with 3% BSA in PBS and then i ncuba t ed with 1:100 F I T C - l a b e l e d an t i -mouse I g G an t ibody .
Sect ions were washed in 3% BSA in PBS for 2 hr and viewed with an epi f luorescence mic ro scope
Lcitz (Diaplan) . The p r i m a r y a n t i b o d y ( h y b r i d o m a s u p e r n a t a n t ) was omi t t ed in contro ls .
(_',:~ll culture
The m e t h o d of cell cu l tu r ing has been extensively descr ibed previous ly . 13'14 Briefly, 1-day-old
p~lpal honeybee bra ins were dissected in sterile saline. Calyces of the m u s h r o o m bodies and an t enna l
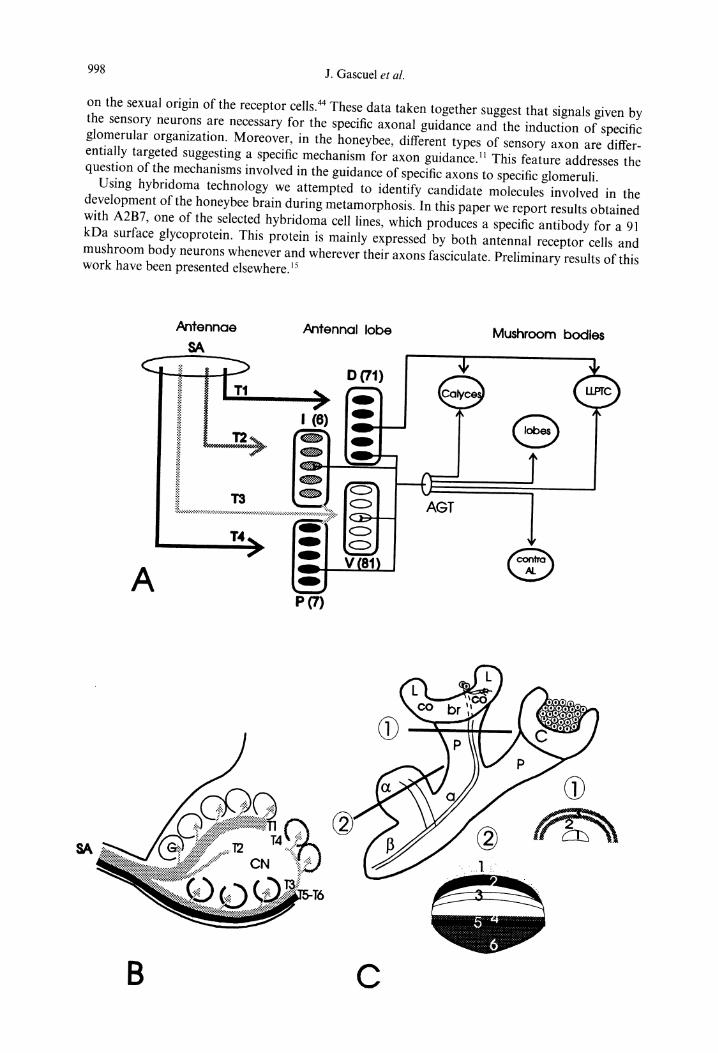
Fig. 1. General organization of the honeybee olfactory pathways. (A, modified from Ref. 27, and B). As in other insects, the soma of the honeybee olfactory receptor cells, are housed in the sensilla along the length of the flagellum. 9 The sensory axons (S.A.) that form the sensory antennal nerve project to the antennal lobe, the first relay station of the insect olfactory pathway (for review see Ref. 26). The spatial organization of the antennal lobe has been studied in detail. 2 At this level, the antennal nerve divides into six main sensory tracts (T1-T6); each of tracts T1-T4 projects to a population of glomeruli: the dorsal (D, to 71 glomeruli), intermediary (I, to 6 glomeruli), ventral (V, to 81 glomeruli), and posterior (P, to 7 glomeruli). T5 projects to the dorsal lobe, T6 divides in three bundles projecting to the protocerebrum and the sub-oesophageal ganglion. Each glomerulus represents an invariant and identifiable morphological subunit where sensory axon terminals synapse with antennal lobe neurons. 2 The glomeruli (G) are located around a coarse neuropile (CN) and are delineated by glial cells that are crucial for their morphogenesis. 1~'~2 Composed of several tracts, the AGT (tractus antenno-globularis) represents the main efferent bundle of the antennal lobe, and connects it with the mushroom bodies. 2 The axons of the two morphological subtypes of the antennal lobe output neurons 2v4~ are distributed in between the different tracts of the AGT, and project to different parts of the protocerebrum, especially to the ipsilateral mushroom body where they synapse with intrinsic neurons, the so-called Kenyon cells l° (lc). In the honeybee brain, the mushroom bodies represent multimodal processing centers that are crucial for memory formation, especially of odors (for review, see Refs. 8 and 28), and its cellular organization has been extensively described 3°'43 (for review, see Ref. 29). Mushroom bodies represent two pairs of large and precisely organized protocerebral structures. Each pair consists of two calyces (C) and two peduncles (p), which fuse and then divide in c~ and fl lobes. Calyces constitute input structures, while ~ and fl lobes have been demonstrated to be both input and output areas. 43 Soma of the Kenyon cells, are grouped into and around a cup-shaped neuropilar structures, the so-called calyces. The latter, are organized in three areas (from the outermost intrinsic neurons, the Kenyon cells to the center: the lips (1), the collar (co) and the basal ring (br) each thought to receive different categories of sensory inputs 3°'2 (for review see Ref. 29). Hence, the Kenyon cells can be divided into three subsets according to the projection area of their dendritic arbors in the calyces. Their respective axons (a) project into the pedunculus at the base of which each is subdivided in two branches, one branch leading into the c~ lobe and the other into the [3 lobe. The three subsets of axons corresponding to the three subsets of Kenyon cells fasciculate and form all along the pedunculus three encased gutter shaped structures. At the junction of the pedunculus and the lobes these fascicles divide and project into layers. Thus, to the three subsets of Kenyon cells correspond, the three encased gutter shaped structures in the pedunculus and the three compartments of the calyces (lip, collar and basal ring) as well as the different bands in the lobes. (SA, sensory axons; D, dorsal population of glomeruli; I, intermediary population of glomeruli; V, ventral population of glomeruli; P, posterior population of glomeruli; LLPTC, lateral lobe of the protocerebrum; G, glomeruli; CN, coarse neuropil; 1, lip of the calyces; co, collar of the calyces; C, calyces; br, basal ring
of the calyces: P, pedunculus; ~ and fl = ~ and fl lobes).

1000 J. Gascuel et al.
lobes were dissected and separately pooled in 5 + 4 culture medium. The cells were then treated with trypsin (0.05% in saline without Ca 2+ and Mg 2+ with 1 mM EDTA) for 10 min. Trypsin action was blocked with 5 + 4 culture medium containing 20% fetal calf serum. Cells were mechanically dissociated with a Pasteur pipette in conditioned 5 + 4 culture medium and the dissociated cells were transferred in 1006 Falcon Petri dishes that had been previously coated with poly-D-lysine.
Cell culture immunochemistry
In standard immunocytochemical experiments, cell cultures were fixed in 2% paraformaldehyde in PBS for 20 rain, blocked with 3% BSA in PBS for 15 min and incubated in A2B7 supernatant diluted 1:2 in 3% BSA in PBS for 2 hr. The cultures were washed three times in 3% BSA in PBS and incubated in secondary antibody (anti-mouse IgG FITC diluted 1:100 in 3% BSA in PBS) for 2 hr. The cells were washed three times with 3% BSA in PBS, mounted in citifluor, and viewed either with a confocal microscope, or an epifluorescence Leitz microscope.
In immunostaining of living cells, both A2B7 and rat anti-tubulin antibodies were concentrated by ultrafiltration using Centricon 60 concentrator (Amicon) and then rediluted in 5+4 culture medium at 1:200 for anti-tubulin and at the same dilution as in the original supernatant for A2B7. Three kinds of treatment were applied to the cultures. First, live cell cultures were incubated with A2B7 antibody in culture medium. Second, live cell cultures were incubated with anti-tubulin in culture medium. Third, in order to test if the anti-tubulin antibody diluted in culture media was still efficient, we tested it on cell cultures fixed in 2% paraformaldehyde, in microtubule-stabilizing PIPES buffer, permeabilized for 4 min in 0.1% Triton X-100 in PIPES buffer and then incubated with anti-tubulin (1:200) in 5 + 4 culture medium. All three categories of culture were incubated in different primary antibody solutions for 2 hr and processed in the same way, i.e. washed three times in cell culture medium, incubated in secondary antibody anti-mouse FITC-conjugated (in the case of A2B7 antibody-treated cultures), or anti-rat FITC-conjugated antibody (in the case of anti- tubulin treated cultures). The cells were washed three times in 3 % BSA in PBS, quickly stained with Hoescht, and mounted in citifluor before viewing with an epifluorescence Leitz microscope. All experiments were carried out at room temperature.
Periodate treatment
Frozen sections of honeybee brains were incubated for 2 hr at 4°C in the dark with 50 mM acetate buffer (pH 4.5) containing 75 mM of sodium metaperiodate. Controls were incubated with the same acetate buffer without sodium metaperiodate. Sections were washed and incubated in 3% BSA in PBS for 15 min before processing as described in the screening and immunostaining section.
Immunopre c ip i tat ion
Heads from 450 honeybees were ground in liquid nitrogen and homogenized with a glass homo- genizer and teflon pestle in Tris buffer 50 mM (pH 7.9) containing 1/tg/ml of the following protease inhibitors: antipain, chymostatin, leupeptin, pepstatin, N-p-tosyl-c-lysine chloromethyl ketone and N-tosyl-L-phenylalanine chloromethyl ketone. The homogenate was centrifuged (1200 g, 10 min, 4°C) in order to eliminate large debris, then the supernatant was centrifuged (200,000 g, 45 rain, 4°C). The pellet enriched in cell membranes was resuspended for 45 rain in 2 ml of Tris buffer 50 mM (pH 7.9) containing 1% NP-40 and protease inhibitors, as described above, in order to solubilize membrane proteins. After centrifugation (100,000 g, 1 hr, 4°C) to remove insoluble material, the lysate was incubated (1 hr, 4°C, with shaking) with 20 ml of mouse and goat serum coupled to beaded agarose in order to prevent nonspecific binding to A2B7 antibodies during the immunoprecipitation procedure. The lysate was quickly centrifuged, and the pellet containing serum-agarose and non- specifically bound proteins was discarded. One half of the supernatant was incubated with 140/~1 of A2B7 antibody, while the other was incubated with 140/tl of 216 antibody overnight at 4°C with gentle shaking.
The 216 antibody is another monoclonal antibody that we have raised, which stains the glial cells (Gascuel et al., unpublished). Both antibodies are IgG1. Each fraction was then incubated with 80 ml of secondary antibody goat anti-mouse IgG 1 coupled to beaded agarose. After ! hr of incubation at 4°C with shaking and a quick centrifugation, the pellets containing the immune complexes were washed with: Tris buffer 50 mM (pH 7.9), NaC1 1M, Tris buffer 50 mM (pH 7.9) containing 1%

A2B7 expression in honeybee olfactory pathway 1001
Tween-20 and finally distilled water. Samples were then mixed with 2 x concentrated Laemmli buffer, boiled for 5 min, pelleted by centrifugation, and the supernatants subjected to SDS-PAGE. Proteins separated on a 6% polyacrylamide gel under reducing conditions were visualized by silver staining (Sigma).
RESULTS
Neuroanatomical background dealing with the bee olfactory pathway is provided in Fig. 1.
A2B7 antigen pattern expression in adult honeybee brain
The A2B7 antigen was expressed mainly in three structures of the brain; the antennal nerve, the mtlshroom bodies, and the visual system.
A ~ztennal nerve and antennal lobe
Sagittal sections through the antennal nerve and the antennal lobe showed that the antennal nerve was stained by the A2B7 antibody. In the proximal part of the antennal nerve, it was not cl~:,ar if the antibody labeled all fibers or only sub-groups of them. In the antennal lobe, the sensory tract T1 (Fig. 2a) and the group of tracts T3-T6 were also stained at least at their entrance into the a~:~tennal lobe (not shown). It is important to note that the staining of these tracts decreased very abruptly as the tracts progressed into the antennal lobe (Fig. 2a). This decrease in expression along the antero-posterior axis of the tracts could be due to an actual decrease of the expression of the a~;~tigen by the axons themselves, or to a reduction of the number of axons constituting the tracts as they penetrate the antennal lobe and as their axons project into the glomeruli.
Mushroom bodies
Mushroom bodies were the brain structure that displayed the strongest A2B7 staining. In frontal sections of pedunculus of mushroom bodies, A2B7 staining was organized in three main layers, from the center of the pedunculus to the periphery; layers 1 and 3 were marked, while layer 2 was not. In the middle of layer 3 a fine band that was more strongly marked was visible (Fig. 2b). In horizontal sections at the level of the junction between the pedunculus and the calyces these different layers could be recognized (Fig. 2c). In the ~ lobes, the staining revealed a layered organization that superposed the different layers previously described by Mobbs. 3° Layers 1, 3, 4, and 6 were stained by the antibody while layers 2 and 5 were not (Fig. 2d). Layer 3 was characterized by having its central band strongly stained.
Visual system
Insect optic lobes are formed of four layers: the retina, the lamina, the medulla, and the lobula. The photoreceptors are located in the retina, while the visual signals are processed in the three following layers. In the adult honeybee, staining was mainly localized to the retina (not shown) and the lamina, while the medulla and the lobula were only faintly stained (Fig. 2e). A fine tract, which could correspond to the antero-visual tract, was also stained.
EXPRESSION OF A2B7 ANTIGEN IN DEVELOPING ADULT
In the worker honeybee, the pupal stage lasts 9 days, E representing the emergence day for the adult.
Olfactory system
To establish the temporal pattern of expression of the A2B7 antigen during olfactory system development, we carried out immunocytochemical experiments on transverse sections of antennae at the level of the scapus for different developmental stages, i. e. E-9, E-7, and E-3. At E-9, axon bundles were not identifiable, but staining was nevertheless visible on the periphery of the cells, which will give rise to the future antenna: neurons, glial cells, accessory cells and epidermal cells (Fig. 3a). At this stage it was difficult to say which of these cells were stained. As early as the E-7

Fig. 2. A2B7 staining in the adult honeybee. (a). Antennal nerve and antennal lobe. Sagittal sections through the antennal nerve (AN) and antennal lobe (AL and dotted line). Note that staining decreases along tract T1 as it penetrates the coarse neuropil (original magnification, 200 x ). (b) Frontal section of mushroom bodies. A2B7 staining is organized in three layers from the center to the periphery (original magnification, 150 x ). (c) Horizontal section at the level between calyces and pedunculus (original mag- nification, 400 x ). (d) Section of mushroom body ~ lobes. A2B7 (original magnification, 150 x ). (e) A2B7 staining in the visual system (LA, lamina; LO, lobulla; ME, medulla; white arrow, small bundle leaving
the lobula toward protocerebrum (original magnification, 200 x ).
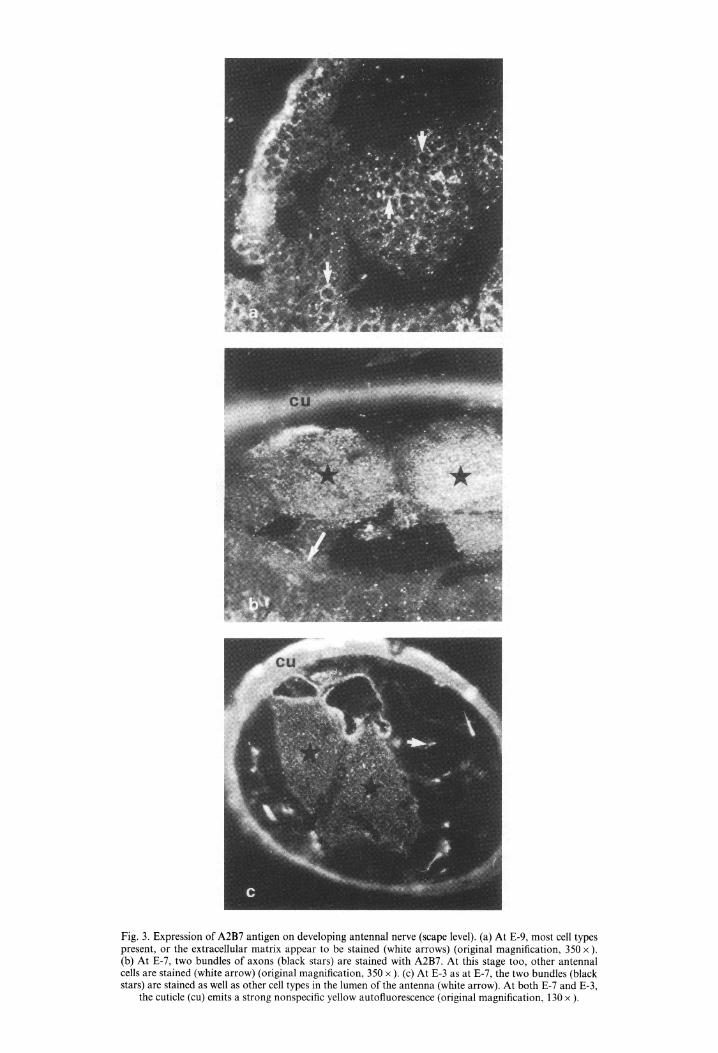
Fig. 3. Expression of A2B7 antigen on developing antennal nerve (scape level). (a) At E-9, most cell types present, or the extracellular matrix appear to be stained (white arrows) (original magnification, 350 x ). (b) At E-7, two bundles of axons (black stars) are stained with A2B7. At this stage too, other antennal cells are stained (white arrow) (original magnification, 350 x ). (c) At E-3 as at E-7, the two bundles (black stars) are stained as well as other cell types in the lumen of the antenna (white arrow). At both E-7 and E-3,
the cuticle (cu) emits a strong nonspecific yellow autofluorescence (original magnification, 130 x ).

1004 J. Gascuel et al.
stage (Fig. 3b), two bundles of axons were easily identifiable and appeared to be intensely stained. At E-3 (Fig. 3c), the pattern of expression was similar to E-7. At both E-7 and E-3, other cells in the lumen of the scape appeared to be marked.
Mushroom bodies
During development, the typical adult layered organization of the mushroom bodies, especially at the level of the lobes, is not clearly established. Therefore, it is difficult to superpose and interpret the staining according to Mobbs. 3° At E-9 (Fig. 4a), on transverse sections two layers were stained in the central part of each pedunculus. At the level of the ~ lobe (Fig. 4a'), two layers appeared to be stained, of which the external one could correspond to adult layer 1. At E-6, the staining in the center of the pedunculus increased and two fine lateral bands appeared in place of the future layer 3 (Fig. 4b). In ~ lobes (Fig. 4b'), the external part of the layer 1 was more heavily stained than the rest of the layer. A fine, bright band was visible, which could correspond to the future fine, bright band described in the adult layer 3. More ventrally, one other fine band appeared clearly and then one, perhaps two, large band(s) could be seen. In the area of the junction between the ~ and/~ lobes bands appeared to be stained (Fig. 4b"). At E-4 (Fig. 4c), staining at the level of the pedunculus appeared to be very similar to the adult staining pattern except that only the fine band was visible in layer 3. In the ~ lobe (Fig. 4c'), layer 1 was uniformly stained and the fine, bright band of layer 3 was still visible; the other fine band appeared at this time in the future location of layer 4, and the putative future layer 6 appeared to be stained.
CELL AND REGION SPECIFICITY OF A2B7 ANTIGEN
We examined the expression of A2B7 antigen by different neuron types cultured separately. Sensory (Fig. 5a) and mushroom body neurons (Fig. 5c) were stained while antennal lobe neurons (Fig. 5b) were not. Interestingly, this cell type specificity is the same as the one observed in vivo since on frozen sections A2B7 mainly stained axons of mushroom bodies and antennal axons but not neurons of antennal lobes.
Moreover, the A2B7 antigen was not expressed in all parts of the neuron. In vivo, the dendritic part of the mushroom body neurons did not express A2B7 antigen, since there was no staining in the calyces; in the cell bodies, the staining was either absent or faintly present. However, in vitro, the cell bodies were generally stained, although occasionally some neurons were observed with no staining in the cell body. However, both in vivo and in vitro, the most highly stained structures were the growth cones and the axons.
Cell surface localization of A2B7 antigen
In order to play a role in cell-cell interactions a molecule has to be expressed at the cell surface. Several lines of evidence suggest that A2B7 is associated with membranes. Immunostaining experiments carried out with A2B7 antibody on mushroom body neurons in cultures and analysed with the confocal microscope (data not shown) suggested that the antibodies bind to the cell membrane. However, with the protocol used for standard immunocytochemistry, even in the absence of detergents that permeabilize the cell, we could not insure that the integrity of the cell membrane was preserved during the entire staining procedure. The possibility that the staining observed could be due to antibodies having artifactually penetrated the cell to bind internal epitope(s) has to be considered.
Therefore, in order to eliminate this possibility we performed immunostaining on living cells by incubating them in culture medium containing A2B7 or anti-tubulin antibody. Our results demonstrated that living cells were stained by A2B7 antibody (Fig. 6c) but not with anti-tubulin (Fig. 6a), indicating that the A2B7 epitope is localized at the surface of the cell. The staining obtained on permeabilized cells with anti-tubulin diluted in 5+4 culture medium is a control demonstrating the efficiency of anti-tubulin antibody when diluted in culture medium (Fig. 6d). The A2B7 antigen could be either a transmembrane glycoprotein or a glycosylphosphatidylinositol (GPI) anchored-protein. To determine which of these two possibilities is correct, we incubated living cells with phospholipase C, which cleaves the glycosyl phosphatidylinositol anchor. Our preliminary

A2B7 expression in honeybee olfactory pa thway 1005
Ci (
Fig. 4. Expression of A2B7 antigen in developing mushroom bodies. (a). Frontal section at E-9 of pedunculus. Two thin tracts are stained in the central part of each pedunculus (arrow). A light staining is visible on the anterior and posterior visual tracts (arrow heads). (a'). In the ~ lobes two layers appear to be stained (arrows) (a and a'; original magnification, 150 x ). (b). At E-6, the staining in the central part of mushroom bodies increase, and two fine lateral bands appear in the future location of layer 3 (arrow). In the ~ lobe (v, ventral; d, dorsal). (b') The setting up of the different layers progresses. (b") A2B7 staining at the junction of ~ and/3 lobes (b, b' and b"; original magnification, 100 x ). (c and c') At E-4 the staining pattern is like the adult pattern except for the thickness of layer 3 on both the pedunculus and ~ lobe (arrows) (original magnification, 150 x ). (d and d') Adult staining of pedunculus (P), calyces (C). Note the
thickness of layer 3 compared to its counterpart at E-4 (arrows) (original magnification, 150 x ).

1006 J. Gascuel et al.
Fig. 5. Cell specificity of A2B7 antigen in culture. (a) A2B7 antennal cells. The staining appears on a group of antennal cells, among which sensory neurons (SN) appear strongly stained. Individual axons and axonal bundles (a) as well are stained. Note the apparent membrane localization of the staining at the level of sensory neuron cell bodies (original magnification, 500 x). (b) Antennal lobe neurons in culture. No staining is visible on antennal lobe neurons (ALN) or on neurites (n) (original magnification, 250 x ). (c) Mushroom body neurons in culture. A2B7 strongly stains axon, growth cone (gc), filopodia (f) and cell bodies of neurons of mushroom bodies (MbN). Note the apparent membrane localization of the staining
(original magnification, 200 x ).
data (not shown) suggest that A2B7 antigen is likely to be a t ransmembrane protein and not a GPI- anchored protein.
Glycosylation of the antigen
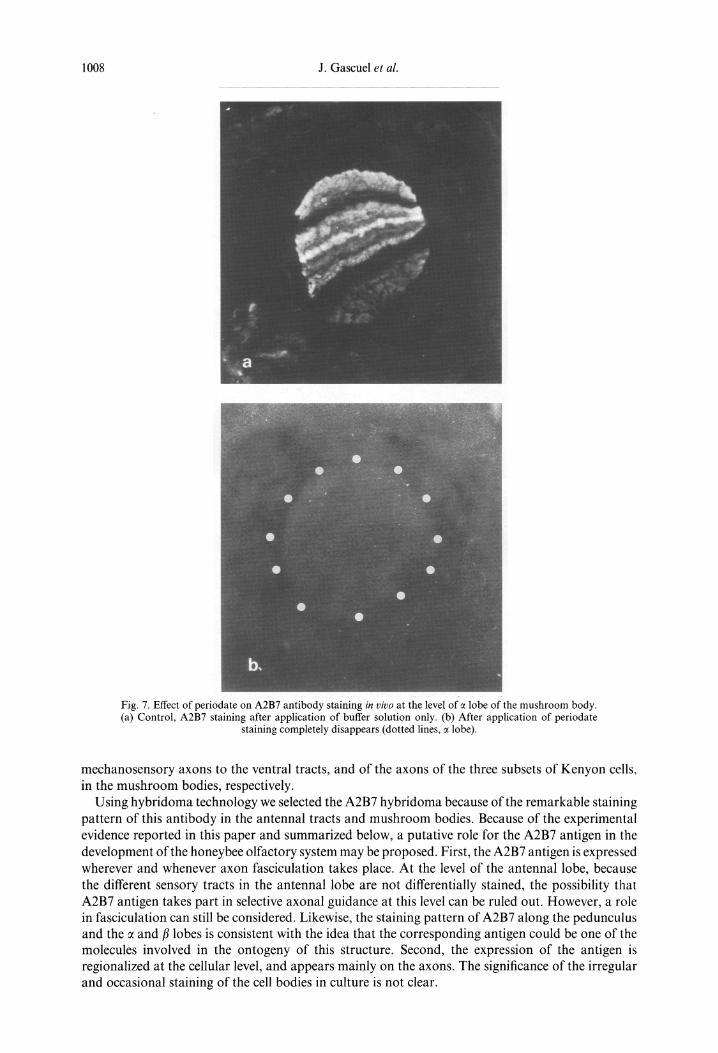
Periodate treatment of the frozen sections resulted in a loss of immunoreactivity, while sections treated with acetate buffer alone were still stained by A2B7 (Fig. 7a), with no change in the staining pattern. This suggests the possibility of glycosylation of the A2B7 epitope (Fig. 7b).
Immunoprecipitation of the A2B7 antigen
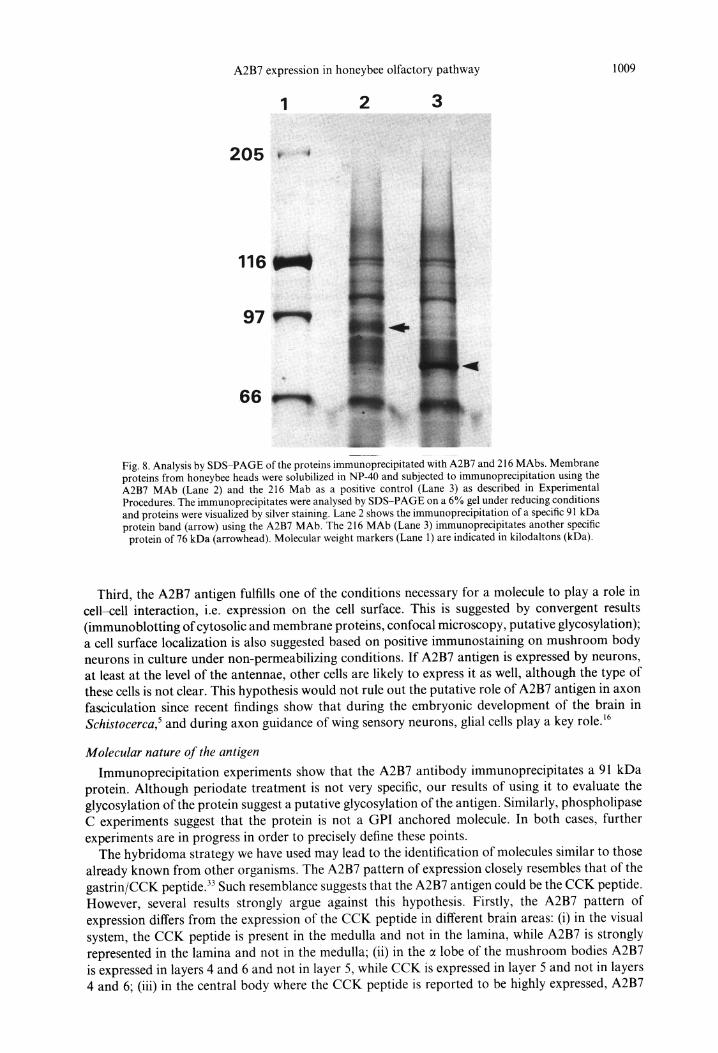
Different conditions for solubilization of membrane protein from honeybee heads has been investigated and the optimal one, considering the rate of extraction and epitope conservation of the antigens, was to use NP-40 in low-ionic strength buffer at pH 7.9. Under these conditions, membrane proteins were solubilized and subjected to an immunoprecipitat ion procedure using A2B7 MAb and 216 MAb, another mouse IgG1, as a positive control. The proteins precipitated by the MAbs were analysed by S D S - P A G E and visualized by silver staining. The result (Fig. 8) indicated that the A2B7 antibody immunoprecipitates a single 91 kDa protein band while the 216 antibody immunoprecipitates a 76 kDa protein.
DISCUSSION
The insect and especially the honeybee olfactory system presents, at least at two levels, interesting features in the context of the search for the cellular and molecular mechanisms involved in axon guidance to specific targets. First, in the antennal lobe the sensory axons are kept precisely organized in a number of tracts as they penetrate the antennal lobe, each tract targeting to one population of identified glomeruli. 2 Interestingly, sensory axons of different diameters (possibly corresponding to different sensory modalities: chemosensory or mechanosensory) are differentially distributed among these tracts. ~ Second, honeybee mushroom bodies are highly compartmentalized structures (Fig. l c) both at the structural and the functional levels, 3°'29'45 in which three subsets of axons, cor- responding to three subsets of Kenyon cells, fasciculate and form the three layers of the pedunculus and the bands in the ~ lobes. This strongly suggests a specific mechanism for guidance of the

A2B7 expression in honeybee olfactory pa thway 1007
. . . . . ~ ; ~ .
,~it, M b ~
Fig. 6. Cell surface localization of A2B7 antigen (original magnification, 200 x ). (a) Application of anti- tubulin on non-permeabilized mushroom body neurons in culture. Note the absence of staining attesting to the absence of penetration of the antibodies into the cells (arrows, cell bodies). (b) Hoechst staining of the same neurons as in (a) (arrows, cell bodies). (c) A2B7 staining of non-permeabilized mushroom body neurons demonstrating the surface localization of the antigen. Note the strong staining on the growth cone (gc), filopodia (arrows) and axon (a). When cell permeabilization does not occur, neuronal cell bodies (MbN) are not stained. (d) Control staining using anti-tubulin diluted in culture medium on permeabilized
cells (MbN) (arrow, axons).
1008 J. Gascuel et al.
Fig. 7. Effect of periodate on A2B7 antibody staining in vivo at the level of e lobe of the mushroom body. (a) Control, A2B7 staining after application of buffer solution only. (b) After application of periodate
staining completely disappears (dotted lines, e lobe).
mechanosensory axons to the ventral tracts, and of the axons of the three subsets of Kenyon cells, in the mushroom bodies, respectively.
Using hybridoma technology we selected the A2B7 hybridoma because of the remarkable staining pattern of this antibody in the antennal tracts and mushroom bodies. Because of the experimental evidence reported in this paper and summarized below, a putative role for the A2B7 antigen in the development of the honeybee olfactory system may be proposed. First, the A2B7 antigen is expressed wherever and whenever axon fasciculation takes place. At the level of the antennal lobe, because the different sensory tracts in the antennal lobe are not differentially stained, the possibility that A2B7 antigen takes part in selective axonal guidance at this level can be ruled out. However, a role in fasciculation can still be considered. Likewise, the staining pattern of A2B7 along the pedunculus and the c~ and/~ lobes is consistent with the idea that the corresponding antigen could be one of the molecules involved in the ontogeny of this structure. Second, the expression of the antigen is regionalized at the cellular level, and appears mainly on the axons. The significance of the irregular and occasional staining of the cell bodies in culture is not clear.
A2B7 expression in honeybee olfactory pathway
1 2 3 . . . . . . . . . . . . . . . . . . . . .
1009
2 0 5 , '
1 1 6
97 ip, q
s e
Fig. 8. Analysis by SDS-PAGE of the proteins immunoprecipitated with A2B7 and 216 MAbs. Membrane proteins from honeybee heads were solubilized in NP-40 and subjected to immunoprecipitation using the A2B7 MAb (Lane 2) and the 216 Mab as a positive control (Lane 3) as described in Experimental Procedures. The immunoprecipitates were analysed by SDS-PAGE on a 6% gel under reducing conditions and proteins were visualized by silver staining. Lane 2 shows the immunoprecipitation of a specific 91 kDa protein band (arrow) using the A2B7 MAb. The 216 MAb (Lane 3) immunoprecipitates another specific
protein of 76 kDa (arrowhead). Molecular weight markers (Lane 1) are indicated in kilodaltons (kDa).
Third, the A2B7 antigen fulfills one of the conditions necessary for a molecule to play a role in cell-cell interaction, i.e. expression on the cell surface. This is suggested by convergent results (immunoblotting ofcytosolic and membrane proteins, confocal microscopy, putative glycosylation); a cell surface localization is also suggested based on positive immunostaining on mushroom body neurons in culture under non-permeabilizing conditions. If A2B7 antigen is expressed by neurons, at least at the level of the antennae, other cells are likely to express it as well, although the type of these cells is not clear. This hypothesis would not rule out the putative role of A2B7 antigen in axon fasciculation since recent findings show that during the embryonic development of the brain in Schistocerca, 5 and during axon guidance of wing sensory neurons, glial cells play a key role. 16
Molecular nature of the antigen
Immunoprecipitation experiments show that the A2B7 antibody immunoprecipitates a 91 kDa protein. Although periodate treatment is not very specific, our results of using it to evaluate the glycosylation of the protein suggest a putative glycosylation of the antigen. Similarly, phospholipase C experiments suggest that the protein is not a GPI anchored molecule. In both cases, further experiments are in progress in order to precisely define these points.
The hybridoma strategy we have used may lead to the identification of molecules similar to those already known from other organisms. The A2B7 pattern of expression closely resembles that of the gastrin/CCK peptide. 33 Such resemblance suggests that the A2B7 antigen could be the CCK peptide. However, several results strongly argue against this hypothesis. Firstly, the A2B7 pattern of expression differs from the expression of the CCK peptide in different brain areas: (i) in the visual system, the CCK peptide is present in the medulla and not in the lamina, while A2B7 is strongly represented in the lamina and not in the medulla; (ii) in the ~ lobe of the mushroom bodies A2B7 is expressed in layers 4 and 6 and not in layer 5, while CCK is expressed in layer 5 and not in layers 4 and 6; (iii) in the central body where the CCK peptide is reported to be highly expressed, A2B7

1010 J. Gascuel et al.
staining is at most very faint. Secondly, the CCK peptide is contained in the cytosolic compartment in large vesicles. By contrast, the A2B7 antigen is a cell surface antigen. Thirdly, the A2B7 antigen is a 91 kDa glycoprotein that does not match the expected molecular weight of a peptide. Finally, the A2B7 antigen is expressed throughout the development of the adult, while the CCK peptide is only expressed at mid-pupal stage. Another possibility, is that the A2B7 antigen could be a receptor for the CCK peptide. However, the expression of the A2B7 antigen in the nonsynaptic neuropile, such as the antennal nerve and mushroom bodies pedunculi, is not consistent with this hypothesis.
In Manduca sexta, antibodies that bind specifically to a subset of sensory axons have been raised. The MPA antibody is reported to stain specifically mechanosensory fibers while the OSA antibody is restricted to the olfactory fibers. 18'19 However, many differences exist between MPA, OSA and A2B7, which suggest that the A2B7 antigen is different from MPA and OSA: (i) even though OSA and A2B7 are membrane-associated antigens, OSA has been reported to faintly stain mushroom bodies; (ii) the molecular weight of A2B7 is 91 kDa while that of the OSA is 66 kDa; (iii) A2B7 is a surface antigen while MPA has been shown to bind to intermediate filaments of the mech- anoreceptor cells.
Another possibility would be that the A2B7 antigen corresponds to one of the membrane molecules already identified in insects as playing roles in cell--cell interaction and especially in axonal fasciculation. In this respect, when compared to the large number of such molecules (fasciclins I and 11, 3 fascicl in 111, 38 neurogl ian , 4 a m a l g a m , 47 chaopt in , 5~ connect in , 34 neurotactin, 7 or other mol- ecules such as the 2B2 antigen), 46 the A2B7 antigen appears to share some characteristics with fasciclin II. Both are transmembrane glycoproteins of similar molecular weight (A2B7, 91 kDa, fasciclin II, 95 kDa). Therefore, the possibility that A2B7 antigen could be a honeybee fasciclin II is strong. However, during post-embryonic development or adulthood, fasciclin II has only been reported to be expressed in Manduca s e x t a wing. 31'32 Only future cloning of the A2B7 gene will definitively determine the molecular identity of the A2B7 antigen.
Acknowled#ements--We would like to thank Drs N. Morel, G. Legal-Lassale, P. Godement, P. Bochet and M. Senguelakis for helpful discussion and E. Genecque for her technical help throughout this work.
REFERENCES
1. Arnold G. and Masson C. (1983) Mise en place des connexions synaptiques de la voie aff6rente antennaire au cours du d6veloppement nymphal de l'ouvri6re d'abeille Apis mellifica ligustica L. C.R. Acad. Sci. III 296, 131-136.
2. Arnold G., Masson C. and Budharugsa S. (1985) Comparative study of the antennal lobes and their afferent pathway in the worker bee and the drone ( Apis mellifica L.). Cell Tiss. Res. 242, 593-605.
3. Bastiani M. J., Harrelson A. L., Snow P. M. and Goodman C. S. (1987) Expression of fasciclin I and II glycoproteins on subset of axons pathways during neuronal development in the grasshopper. Cell 48, 745-755.
4. Bieber A. J., Snow P. M., Hortsch M., Patel N. H., Jacobs J. R., Traquina Z. A., Schilling J. and Goodman C. S. (1989) Drosophila neuroglian: a member of the immunoglobulin superfamily with extensive homology to the vertebrate neural adhesion molecule L 1. Cell 59, 447-460.
5. Boyan G., Therianos S., Williams J. L.D. and Reichert H. (1995) Axogenesis in the embryonic brain of the grasshopper Schistocerca gre#aria: an identified cell analysis of early brain development. Development 121, 75-86.
6. Chambille I., Rospars J. P. and Masson C. (1980) The deutocerebrum of the cockroach Blaberus craniifer Burm. Spatial organization of the sensory glomeruli. J. Neurobiol. 11, 135-254.
7. De La Escalera S. E., Bockamp E. O., Moya F., Piovant M. and Jimenez F. (1990) Characterization and gene cloning of neurotactin, a Drosophila transmembrane protein related to cholinesterases. EMBO J. 9, 3593-3601.
8. Erber J. (1981) Neural correlates of learning in the honey bee. TINS 4, 270--273. 9. Esslen J. and Kaissling K. E. (1976) Zahl und Verteilung antennaler Sensillen bei der Honigbiene. Zoomorphologie 83,
227-251. 10. Fonta C., Sun X. J. and Masson C. (1993) Morphology and spatial distribution of bee antennal lobe interneurons
responsive to odors. Chem. Senses 18, 101-119. 11. Gascuel J. and Masson C. (1991) Quantitative electron microscopic study of the antennal lobe of the honey bee. Tissue
and Cell 23, 341-355. 12. Gascuel J. and Masson C. (1991) Developmental study of afferented and unafferented bee antennal lobe. J. Neurobiol.
22, 795-810. 13. Gascuel J., Masson C. and Beadle D. J. (1991) The morphology and ultrastucture of antennal lobe cells from pupal
honey bees (Apis mellifera) growing in culture. Tissue and Cell 23, 547-559. 14. Gascuel J., Masson C., Bermudez I. and Beadle D. J. (1994) Morphological analysis of honey bee antennal cells growing
in primary cultures. Tissue and Cell 26, 551-558. 15. Gascuel J., Masson C., Brouillard F. and Groome N. (1994) Expression of antigens putatively involved in cell-cell
interactions during bee olfactory system ontogeny. Chem. Senses (abstract) 20, 141. 16. Giangrande A. (1994) Glia in the fly wing are clonally related to epithelial cells and use the nerve as a pathway for
migration. Development 120, 523-534.

A2B7 expression in honeybee olfactory pathway 1011
17. Goodman C. S. and Shatz C. (1993) Developmental mechanisms that generate precise pattern of neuronal connectivity. Cell 72, 77-98.
18. Hishinuma A., Hockfield S., McKay R. and Hildebrand J. G. (1988) Monoclonal antibodies reveal cell-type specific antigens in the sexually dimorphic olfactory system of Manduca sexta. I. Generation of monoclonal antibodies and partial characterization of the antigens. J. Neurosci. 8, 296-307.
19. Hishinuma A., Hockfield S., McKay R. and Hildebrand J. G. (1988) Monoclonal antibodies reveal cell-type specific antigens in the sexually dimorphic olfactory system of Manduca sexta. II. Expression of antigens during postembryonic development. J. Neurosci. 8, 308-315.
20. Kennedy T. E., Serafini T., de la Torre J. R. and Tessier-Lavigne M. (1994) Netrins are diffusible chemotropic factors for commissural axons in the embryonic spinal cord. Cell 78, 425-435.
21. Kolodkin A. L., Matthes D. J., O'Connor T. P., Patel N. H., Bentley D. and Goodman C. S. (1992) Fasciclin IV: sequence, expression and function during growth cone guidance in the grasshopper embryo. Neuron 9, 831-845.
22. Kolodkin A. L., Matthes D. J. and Goodman C. S. (1993) The semaphorin gene encode for a family of transmembrane and secreted growth cone guidance molecules. Cell 75, 1389-1399.
23. Letourneau P. C. and Cypher C. (1991) Regulation of growth cone motility. Cell Motil. Cytoskel. 20, 267-271. 24. Luo Y., Raible D. and Raper J. A. (1993) Collapsin: a protein in brain that induces the collapse and paralysis of neuronal
growth cones. Cell 75, 217-227. 25. Masson C. and Arnold G. (1984) Ontogeny, maturation and plasticity of the olfactory system in the worker bee. J.
Insect. Physiol. 30, 7-14. 26. Masson C. and Mustaparta H. (1990) Chemical information processing in the olfactory system of insects. Physiol. Rev.
70, 199-245. 27. Masson C., Pham-Del6gue M. H., Fonta C., Gascuel J., Arnold G., Nicolas G. and Kerszberg M. (1993) Recent advance
in the concept of adaptation to natural odour signals in the honey bee Apis mellifera L. Apidologie 24, 169-194. 28. Menzel R. (1990). Learning, memory and cognition in the honey bee. In Neurobiology of Comparative Cognition (ed.
Kesner R. P. and Olten D. S.), pp. 237-292. Erlbaum Inc., Hillsdale, NJ, U.S.A. 29. Menzel R., Durst C., Erber J., Eichmtiller S., Hammer M., Hildebrandt H., Mauelshagen J., Mtiller U., Rosenboom
H., Rybak J., Sch~iffer S. and Scheidler A. (1994) The mushroom bodies in the honeybee: from molecules to behaviour. In Neural Basis of Behavioural Adaptation (ed. Schildberger L. and Eisner), pp. 81-102. Fortschritte der Zoologie 39.
30. Mobbs P. G. (1982) The brain of the honey bee Apis mellifera. I. The connections and spatial organization of the mushroom bodies. Phil. Trans. R. Soc. London, B 298, 309-354.
3 !. Nardi J. B. (1990) Expression of a surface epitope on cells that link branches in the tracheal network of Manduca sexta. Development 110, 681-688.
32. Nardi J. B. (1992) Dynamic expression of a cell surface protein during rearrangement of epithelial cells in the Manduca wing monolayer. Developmental Biol. 152, 161-171.
33. Noble M. J. and Goodman L. J. (1987) Immunohistochemical localization of a Gastrin/CCK-like peptide in the brain of the honey bee. In Neurobiology and Behavior of the Honey Bee (eds Menzel R. and Mercer A.), pp. 235-243. Springer Verlag, Berlin.
34. Nose A. and Goodman C. S. (1992) Connectin: a homophilic cell adhesion molecule expressed on a subset ofmotorneuron axons and the target muscles they innervate in Drosophila. Cell 70, 553-567.
35. Oland A. and Tolbert L. P. (1987) Glial pattern during early development of the antennal lobes of Manduca sexta: a comparison between normal lobes and lobes deprived of antennal axons. J. Comp. Neurol. 255, 196-207.
36. Oland L. A. and Tolbert L. P. (1988) Effect of hydroxyurea parallel the effects of radiation in developing olfactory glomeruli in insects. J. Comp. Neurol. 278, 377-387.
37. Oland L. A., Tolbert L. P. and Mossman K. L. (1988) Radiation-induced reduction of the glial population during development disrupts the formation of olfactory glomeruli in an insect. J. Neurobiol. 8, 353-367.
38. Patel N. H., Snow P. M. and Goodman C. S. (1987) Characterisation and cloning of fasciclin III: a glycoprotein expressed on a subset of neurons and axons pathways in Drosophila. Cell 48, 975-998.
39. Ramon y Cajal S. A. (1980) A quelle 6poque apparaissent les expansions des cellules nerveuses de la moelle 6pini6re du poulet. Anat. Anzerger 5, 609-613.
4(I. Ressler K. J., Sullivan S. L. and Buck L. B. (1994) Information coding in the olfactory system: evidence for a stereotyped and highly organized epitope map in the olfactory bulb. Cell 79, 1245-1255.
41. Rodrigues V. and Buchner E. (1984) (3H)-2-deoxyglucose mapping of odor induced neuronal activity in the antennal lobe of Drosophila melanogaster. Brain Res. 324, 374-378.
42. Rospars J. P. and Hildebrand J. G. (1992) Anatomical identification of glomeruli in the antennal lobes of the male sphinx moth Manduca sexta. Cell Tissue Res. 270, 205-227.
43. Rybak J. and Menzel R. (1993) Anatomy of the mushroom bodies in the honey bee brain: the neuronal connections of the ct lobe. J. Comp. Neurol. 334, 444-465.
44. Schneiderman A. M., Matsumoto S. G. and Hildebrand J. G. (1982) Trans-sexually grafted antennae influence devel- opment of sexually dimorphic neurons in moth brain. Nature 298, 844-846.
45. Schfirmann F. W. and Erber J. (1990) FMRfamide-like immunoreactivity in the brain of the honey bee (Apis mellifera). A light and electron microscopical study. Neurosci. 38, 797-807.
46. Seaver E. C., Karlstrom R. O. and Bastiani M. J. (1991) The restricted spatial and temporal expression of a nervous- system-specific antigen involved in axon outgrowth during development of the grasshopper. Development 111, 881-893.
47. Seeger M. A., Haffley L. and Kaufman T. C. (1988) Characterization of amalgam: a member of the immunoglobulin super family from Drosophila. Cell 55, 589-600.
48. Sun X. J., Fonta C. and Masson C. (1993) Odour quality processing by antennal lobe interneurons. Chem. Senses 18, 355-377.
49. Tolbert L. P., Matsumoto S. G. and Hildebrand J. G. (1983) Development of synapses in the antennal lobe of the moth Manduca sexta during metamorphosis. J. Neurosci. 3, 1158-1175.
50. Vassar R., Chao S. K., Sicheran R., Nunez J. M., Vosshall J. B. and Axel R. (1994) Topographic organization of sensory projections to the olfactory bulb. Cell 79, 981-991.
51. Zipursky S. L., Venkatesh T. R. and Benzer S. (1985) From monoclonal antibody to gene for a neuron-specific glycoprotein in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 82, 1855-1859.




















