
Expression of the IgLON cell adhesion molecules kilon and OBCAM in hypothalamic magnocellular...
12
Expression of the IgLON Cell Adhesion Molecules Kilon and OBCAM in Hypothalamic Magnocellular Neurons SEIJI MIYATA, 1 * NOBUO FUNATSU, 1 WATARU MATSUNAGA, 1 TOSHIKAZU KIYOHARA, 1 YOSHIHIRO SOKAWA, 1 AND SHOHEI MAEKAWA 2 1 Department of Applied Biology, Kyoto Institute of Technology, Kyoto 606-8585, Japan 2 Division of Bioscience, Graduate School of Science and Technology, Kobe University, Kobe 657-8501, Japan ABSTRACT The vasopressin (AVP) and oxytocin (OXT) magnocellular neurons in the hypothalamic supraoptic (SON) and paraventricular nuclei (PVN) display reversible structural plasticity of neurons and glial cells under different conditions of neuropeptide secretion. In the present study, we investigated the expression of two immunoglobulin superfamily (IgSF) proteins, Kilon and OBCAM, in the magnocellular neurons by using monoclonal antibodies. Anti-Kilon antibody reacted specifically with the bacterially expressed recombinant Kilon but not with the recombinant OBCAM, and similarly anti-OBCAM antibody specifically recognized the recombinant OBCAM. Western blotting analysis revealed the specific expression of Kilon and OBCAM in the SON homogenates. Although Kilon and OBCAM of the SON homogenates were present as the insoluble form, most Kilon was present in the Triton-insoluble fraction, and OBCAM was localized mainly in the Triton-soluble fraction. Immunocytochemistry revealed Kilon and OBCAM immunoreactivity in the magnocellular neurons of the SON and PVN of the rat hypothalamus compared with outside of the SON and PVN in the hypothal- amus. The double-labeling study with confocal microscopy further demonstrated that Kilon immunoreactivity was observed mainly in the dendrites of AVP-secreting neurons and also occasionally OXT-secreting neurons. However, OBCAM immunoreactivity was exclusively seen in the dendrites of AVP-secreting magnocellular neurons. Chronic physiological stimu- lation by 2% NaCl had no effect on the expression levels of either IgLON protein in the SON. Our study thus demonstrated specific expression of Kilon and OBCAM in the hypothalamic magnocellular neurons, particularly in dendrites, suggesting that they confer on magno- cellular neurons the ability to rearrange dendritic connectivity. J. Comp. Neurol. 424:74 – 85, 2000. © 2000 Wiley-Liss, Inc. Indexing terms: GPI; oxytocin; vasopressin; plasticity; IgCAM; dendrite; supraoptic nucleus; adhesion molecules The neurohypophysial hormones arginine vasopressin (AVP) and oxytocin (OXT) are synthesized mainly in the magnocellular neurons of the supraoptic (SON) and paraventricular nucleus (PVN) in the hypothalamus (Swaab et al., 1975; Sawchenko and Swanson, 1982). The function of peripheral AVP is chiefly considered to be the regulation of body fluid homeostasis, including extracellu- lar osmolarity and intravascular volume (for review, see Cunningham and Sawchenko, 1991). OXT causes contrac- tion of uterine smooth muscle during parturition and of myoepithelial cells in the mammary glands, producing milk ejection, and is also released by increased plasma osmolarity. In the hypothalamo-neurohypophysial system (HNS), reversible structural reorganization is seen under chronic physiological demand such as lactation and dehy- dration (for reviews, see Hatton, 1990, 1997, 1999). In the Grant sponsor: the Japan Society for the Promotion of Science; Grant number: Scientific Research 11640663; Grant sponsor: the Ministry of Education, Science, Sports and Culture of Japan; Grant number: Scientific Research 10470016. *Correspondence to: Dr. Seiji Miyata, Department of Applied Biology, Kyoto Institute of Technology, Matsugasaki, Sakyo-ku, Kyoto 606-8585, Japan. E-mail: [email protected] Received 20 December 1999; Revised 9 March 2000; Accepted 12 April 2000 THE JOURNAL OF COMPARATIVE NEUROLOGY 424:74 – 85 (2000) © 2000 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Expression of the IgLON cell adhesion molecules kilon and OBCAM in hypothalamic magnocellular...

Expression of the IgLON Cell AdhesionMolecules Kilon and OBCAM in
Hypothalamic Magnocellular Neurons
SEIJI MIYATA,1* NOBUO FUNATSU,1 WATARU MATSUNAGA,1
TOSHIKAZU KIYOHARA,1 YOSHIHIRO SOKAWA,1AND SHOHEI MAEKAWA2
1Department of Applied Biology, Kyoto Institute of Technology, Kyoto 606-8585, Japan2Division of Bioscience, Graduate School of Science and Technology, Kobe University,
Kobe 657-8501, Japan
ABSTRACTThe vasopressin (AVP) and oxytocin (OXT) magnocellular neurons in the hypothalamic
supraoptic (SON) and paraventricular nuclei (PVN) display reversible structural plasticity ofneurons and glial cells under different conditions of neuropeptide secretion. In the presentstudy, we investigated the expression of two immunoglobulin superfamily (IgSF) proteins,Kilon and OBCAM, in the magnocellular neurons by using monoclonal antibodies. Anti-Kilonantibody reacted specifically with the bacterially expressed recombinant Kilon but not withthe recombinant OBCAM, and similarly anti-OBCAM antibody specifically recognized therecombinant OBCAM. Western blotting analysis revealed the specific expression of Kilon andOBCAM in the SON homogenates. Although Kilon and OBCAM of the SON homogenateswere present as the insoluble form, most Kilon was present in the Triton-insoluble fraction,and OBCAM was localized mainly in the Triton-soluble fraction. Immunocytochemistryrevealed Kilon and OBCAM immunoreactivity in the magnocellular neurons of the SON andPVN of the rat hypothalamus compared with outside of the SON and PVN in the hypothal-amus. The double-labeling study with confocal microscopy further demonstrated that Kilonimmunoreactivity was observed mainly in the dendrites of AVP-secreting neurons and alsooccasionally OXT-secreting neurons. However, OBCAM immunoreactivity was exclusivelyseen in the dendrites of AVP-secreting magnocellular neurons. Chronic physiological stimu-lation by 2% NaCl had no effect on the expression levels of either IgLON protein in the SON.Our study thus demonstrated specific expression of Kilon and OBCAM in the hypothalamicmagnocellular neurons, particularly in dendrites, suggesting that they confer on magno-cellular neurons the ability to rearrange dendritic connectivity. J. Comp. Neurol. 424:74–85,2000. © 2000 Wiley-Liss, Inc.
Indexing terms: GPI; oxytocin; vasopressin; plasticity; IgCAM; dendrite; supraoptic nucleus;
adhesion molecules
The neurohypophysial hormones arginine vasopressin(AVP) and oxytocin (OXT) are synthesized mainly inthe magnocellular neurons of the supraoptic (SON) andparaventricular nucleus (PVN) in the hypothalamus(Swaab et al., 1975; Sawchenko and Swanson, 1982). Thefunction of peripheral AVP is chiefly considered to be theregulation of body fluid homeostasis, including extracellu-lar osmolarity and intravascular volume (for review, seeCunningham and Sawchenko, 1991). OXT causes contrac-tion of uterine smooth muscle during parturition and ofmyoepithelial cells in the mammary glands, producingmilk ejection, and is also released by increased plasmaosmolarity. In the hypothalamo-neurohypophysial system
(HNS), reversible structural reorganization is seen underchronic physiological demand such as lactation and dehy-dration (for reviews, see Hatton, 1990, 1997, 1999). In the
Grant sponsor: the Japan Society for the Promotion of Science; Grantnumber: Scientific Research 11640663; Grant sponsor: the Ministry ofEducation, Science, Sports and Culture of Japan; Grant number: ScientificResearch 10470016.
*Correspondence to: Dr. Seiji Miyata, Department of Applied Biology,Kyoto Institute of Technology, Matsugasaki, Sakyo-ku, Kyoto 606-8585,Japan. E-mail: [email protected]
Received 20 December 1999; Revised 9 March 2000; Accepted 12 April2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 424:74–85 (2000)
© 2000 WILEY-LISS, INC.

hypothalamic nuclei, the structural reorganization iscaused by retraction of glial cells from their usual posi-tions between adjacent cell bodies and/or neighboring den-drites, which results in soma-somatic, soma-dendritic, anddendro-dendritic membrane apposition (Perlmutter et al.,1984, 1985; Salm et al., 1985; Bobak and Salm, 1996;Hawrylak et al., 1998).
In a similar fashion to the hypothalamic nuclei, theastrocytes in the neurohypophysis (pituicytes) generallysurround or enclose the axonal terminals of magnocellularneurons under basal conditions. However, they release theengulfed axonal terminals with chronic physiologicalstimulation and permit the neurosecretory terminals tomake contact with the basal lamina to a greater extent(Tweedle and Hatton, 1982, 1987; Matsunaga et al.,1999a,b). This structural reorganization can be mimickedby using in vitro studies in which shape conversion ofpituicytes is found in response to the b-adrenergic ago-nists adenosine and endothelin (Bicknell et al., 1989;Luckman and Bicknell, 1990; Smithson et al., 1990; Hat-ton et al., 1991; Miyata et al., 1999).
Cell adhesion molecules (CAMs) appear to be of partic-ular relevance since they intervene in most cell-cell andcell-matrix interactions for neuronal plasticity (Hoffman,1998). The CAMs of the immunoglobulin superfamily(IgSF) are defined by the one or more extracellular Ig-likedomains and have a major role in controlling neurogenesisin the brain (Williams and Barclay, 1988; Hunkapillarand Hood, 1989). Some IgSF molecules also contain sev-eral fibronectin type-III-like repeats in addition to five orsix Ig-like domains. Among the IgSF molecules, MAG,SC1, TLN, NCAM-140, and NCAM-180 are known to bethe transmembrane type, and Thy-1, limbic system-associated protein (LAMP), opioid-binding cell adhesionmolecule (OBCAM), neurotrimin (Ntm), Kilon (kindredof IgLON), NCAM-120, and F3 are of the glycosylphos-phatidylinositol (GPI)-anchored type. They are importantregulators of axonal growth, fasciculation, and synapto-genesis via heterophilic or homophilic interactions(Hunkapillar and Hood, 1989).
Three IgSF molecules, LAMP (Levitt, 1984), OBCAM(Schofield et al., 1989), and Ntm (Struyk et al., 1995), areclassified into a subgroup of IgSF, IgLON (IgSF contain-ing LAMP, OBCAM, and Ntm). Each of the IgLON pro-teins has three Ig-like domains, is attached to the lipidmembrane by the GPI anchor, and is expressed on distinctsubpopulations of brain neurons. The first identifiedIgLON protein, LAMP, is expressed predominantly on thesurface of cortical and subcortical neurons of the limbicsystem, which are related through particular circuits andmediate cognitive behavior, memory, and learning (Levitt,1984; Zacco et al., 1990; Pimenta et al., 1996; Reinoso etal., 1996). LAMP was detected on the surface of cell bod-ies, dendrites, axonal membrane, and growth cones duringearly development (Horton and Levitt, 1988; Keller andLevitt, 1989). However, in mature limbic neurons, its im-munoreactivity is eventually lost on axons postnatally butis maintained in the dendritic and somatic compartments(Zacco et al., 1990). LAMP was shown to bind homophili-cally and mediate cell adhesion and outgrowth by using anassay in which LAMP-positive cells bound to a LAMPsubstrate (Pimenta et al., 1995; Zhukareva and Levitt,1995). A soluble form of LAMP is known to stimulateoutgrowth of limbic neurons, possibly mediated throughL-type Ca21-channel activation (Zhukareva et al., 1997).
Another IgLON protein, Ntm, shows a largely comple-mentary expression pattern to that of LAMP (Struyk etal., 1995). An in vitro experiment demonstrated that Ntmbinds homophilically to dorsal root ganglia and hippocam-pal neurons in promoting outgrowth and neuronal attach-ment and also binds heterophilically to superior cervicalganglia neurons in inhibiting outgrowth and neuronalattachment (Orlando et al., 1998; Gil et al., 1998). OB-CAM has been shown to have a much more restricteddistribution, with the highest expression in the cerebralcortex and hippocampus, compared with LAMP and Ntm(Struyk et al., 1995). Recently, we identified a new IgLONprotein and named it Kilon (Funatsu et al., 1999). A ho-mology search of the coding sequence of this proteinshowed high degrees of similarity to those of LAMP (56%),OBCAM (49%), and Ntm (48%). Kilon is expressed at highlevels in the mature adult brain compared with embryonicand early postnatal brains, and its localization in thecerebral cortex and hippocampus corresponds well to thatof LAMP (Funatsu et al., 1999; Zacco et al., 1990).
The HNS has been shown to express high levels ofseveral kinds of IgSF proteins (for reviews, see Theodo-sis and Poulain, 1993; Theodosis et al., 1994, 1998;Theodosis and MacVicar, 1996). Neural cell adhesionmolecule (NCAM) is expressed by neurons and glialcells, and modulation of adhesion is closely related tothe molecule’s content of polysialic acid (PSA). How-ever, the NCAM molecule containing more than 30%PSA is abundant throughout the late embryonic andearly postnatal brains; it is gradually replaced by sev-eral adult isoforms containing less than 10% PSA(Finne et al., 1983; Chuong and Edelman, 1984). PSA-NCAM is still abundantly expressed in the HNS of theadult brain, and its immunoreactivity was observed indendrites, axons, terminals, and associated astrocytes,but not in neuronal somata (Theodosis et al., 1991;Bonfanti et al., 1992; Kiss et al., 1993). Significantamounts of F3 are expressed in the developing tissuesand extracellular fluids at the time of axon extensionand synaptogenesis (Gennarini et al., 1989).
Moreover, the soluble type of F3 promotes neuriteoutgrowth of cultured neurons, whereas the GPI-anchored form confers adhesive properties to F3-transfected cells (Durbec et al., 1992). F3 was alsoshown to be localized to magnocellular neuron somataas the insoluble form and in neuronal terminals as thesoluble form (Olive et al., 1995; Pierre et al., 1998).However, the molecular species of IgSF proteins, par-ticularly the GPI-anchored type, have not been com-pletely elucidated in the HNS. Here, we produced spe-cific monoclonal antibodies against Kilon and OBCAMand demonstrated with Western blotting and immuno-histochemistry that two IgLON proteins were expressedstrictly in the hypothalamic magnocellular SON andPVN. Moreover, a double-labeling study using confocalmicroscopy showed that both proteins were targetedspecifically to the dendrites of magnocellular neurons.The dendrites of magnocellular neurons show drasticrearrangement including membrane apposition in re-sponse to chronic physiological stimulation. Therefore,there appears to be a close relationship between theexpression of IgLON proteins and dendritic plasticity.
75IgLON IN MAGNOCELLULAR NEURONS

MATERIALS AND METHODS
Animals
Male Wistar rats (8–12 weeks old) from our laboratorycolony were used for biochemical and anatomical studies;they were maintained under the following conditions:23 6 2°C, 14:10-hour light-dark cycle, and food and waterad libitum. Osmotic stimulation was performed by givinga 2% NaCl solution for 2 and 7 days instead of normaldrinking water. All experimental protocols were carriedout in accordance with the guidelines of the PhysiologicalSociety of Japan.
Chemicals
Monoclonal antibodies against OXT- (PS-38) and AVP-neurophysin (PS-41) were generously provided by Dr. H.Gainer, from the National Institutes of Health (Ben-Baraket al., 1985). The polyclonal antibodies against OXT andAVP were purchased from UCB Bioproduct (L’Alleud, Bel-gium) and Chemicon (Temecula, CA), respectively. Biotinyl-ated goat anti-rabbit IgG, biotinylated horse anti-mouse IgG,Texas Red streptavidin, ABC Elite kit, and Vectashield (non-fluorescent mounting medium) were purchased from VectorLaboratories (Burlingame, CA). Fluorescein isothiocyanate(FITC)-labeled goat anti-rabbit, FITC-labeled horse anti-mouse IgG, and horseradish-peroxidase (HRP)-conjugatedanti-mouse IgG were purchased from Kirkegaard and Perry(Gaithersburg, MD). An enhanced chemiluminescence kitand x-ray film were purchased from Amersham. Isopropyl-1-thio-b-D-galactopyranoside (Nakarai, Kyoto, Japan),phosphatidylinositol-specific phospholipase C (PI-PLC; Fu-nakoshi, Tokyo, Japan), peptide N-glycosidase F (PNGF;Boehringer Mannheim, Germany), pGEX-2T (Amersham),and pBluescript II KS(1) (Stratagene, La Jolla, CA) werealso used for biochemical study.
Production of monoclonal antibodiesagainst Kilon and OBCAM
Monoclonal antibodies against Kilon and OBCAM wereproduced as described previously (Funatsu et al., 1999).Briefly, frozen whole rat brains were homogenized in 1volume (w/v) of TME solution (10 mM Tris-HCl, 1 mMMgCl2, and 1 mM EGTA; pH 7.4) containing 2% TritonX-100, 1 mM phenylmethylsulfonyl fluoride (PMSF), 0.1%aprotinin, 10 mg/ml leupeptin, and 10 mg/ml pepstatin. Analiquot of 2.4 M sucrose solution was added, and the finalsucrose concentration was adjusted to 0.8 M. The sampleswere then placed in centrifuge tubes and overlaid withTME solution. After centrifugation for 6 hours at 70,000gby using a swing rotor (HITACHI SW27-2, Hitachi, Ja-pan), the membrane fraction concentrated at the interfaceof TME solution and the original sample solution werecollected. The fraction was diluted with 5 volumes of TMEsolution containing 1% Triton X-100 and centrifuged for 1hour at 100,000g. The resulting pellet was recovered andthen homogenized in 5 volumes of TME solution contain-ing 1 M NaCl. After centrifugation for 1 hour at 100,000g,the pellet fraction was recovered, suspended in 5 volumesof 50 mM Tris-HCl buffer containing 1 mM EDTA, 10%glycerol, 10 mM 2-mercaptoethanol, and 5% Nonidet-40(pH 9.5), and kept for 1 hour at 0°C. After centrifugationfor 1.5 hours at 100,000g, the resulting pellet was recov-ered and suspended in 10 mM Tris-HCl buffer containing5 mM EDTA (pH 7.4). The raft, thus obtained, was frozenat 280°C until use.
The raft fraction was further treated with PI-PLC tosolubilize the protein and digested with PNGF to removethe N-linked sugar chains. Aliquots (0.04) mg of proteinwere injected intraperitoneally after mixing with Freund’scomplete adjuvant. After four injections of antigen (once aday) following four immunizations with the adjuvant, thespleen was dissected and used for cell fusion with my-eloma cells. During screening, clones 89B3 and 84A3 werefound to be specific for Kilon and OBCAM, respectively.Similarly, clones 1F3 and 89D1 were screened as specificantibodies for Thy-1 (at 25–27 kDa) and F3 (at 135 kDa)as described in our previous report (Funatsu et al., 1999).
Production of recombinant Kilonand OBCAM
Constructs of bacterial expression vectors of Kilon andOBCAM were prepared as described previously (Funatsuet al., 1999). Briefly, the DNA fragment containing the317-amino acid sequence of Kilon (32–348) was inserted inframe into the EcoRI site of pGEX-2T. The resulting glu-tathione S-transferase (GST) fusion gene expressed theGST fusion protein with a molecular mass of 60 kDa(GST-Kilon). The cDNA clone of rat OBCAM used lackedthe N-terminal 118 amino acids and encoded theC-terminal 226 amino acids (119–345). The DNA frag-ment containing the C-terminal 226-amino acid sequence(119–345) was inserted in frame into the EcoRI and Xholsites of pBluescript II KS(1). The resulting lacZ fusiongene expressed the b-galactosidase fusion protein with amolecular mass of 30 kDa (bN-OBCAM). These plasmidswere introduced to Escherichia coli JM109, and the pro-duction of recombinant proteins was induced with 1 mMIPTG at 37°C. After a 3-hour incubation, cells were col-lected, washed, suspended in sodium dodecyl sulfate(SDS)-sample buffer, and boiled for 5 minutes.
Immunohistochemistry
Male rats (n 5 20) were used for immunohistochemicalexperiments. Under deep anesthesia with sodium pentobar-bital (70 mg/kg), the animals were transcardially perfusedwith 100 ml of heparinized phosphate-buffered saline (PBS)followed by 250 ml of 4% paraformaldehyde in 0.1 M phos-phate buffer (PB; pH 7.4). The brain blocks that included theSON, PVN, suprachiasmatic nucleus, median eminence, andpituitary glands were postfixed with the same fixative for 24hours at 4°C. After several washes in cold PBS, the blockswere sectioned coronally at 25–30 mm by using a vibratome(DTK-1000, DSK, Kyoto, Japan). Sections were washed re-peatedly with cold PBS and preblocked with 5% normalhorse serum in PBS containing 0.3% Triton X-100 (PBST) formore than 24 hours. Hypothalamic sections were then incu-bated with anti-Kilon (dilution, 1:3,000), anti-OBCAM anti-body (dilution, 1:2,000), anti-F3 (dilution, 1:500), or anti-Thy-1 (dilution, 1:500) in PBST containing 1% normalserum for 48 hours at 4°C. Pituitary sections were im-mersed in either anti-Kilon or anti-OBCAM antibody (di-lution, 1:500). The sections were further incubated withbiotinylated horse anti-mouse IgG (dilution, 1:200), fol-lowed by the ABC Elite complex (dilution, 1:200) for 2hours. Peroxidase activity was visualized with 0.02% 3,39-diaminobenzidine, 0.05% nickel sulfate, and 0.01% hy-drogen peroxide in 0.05 M Tris-HCl buffer (pH 7.4).
In some experiments, the co-localization of both IgLON(Kilon or OBCAM) and neuropeptide (OXT or AVP) wasdetermined by the simultaneous visualization of two an-
76 S. MIYATA ET AL.

tigens in same tissue sections. The sections were incu-bated with either monoclonal anti-Kilon or anti-OBCAMantibody (1:20–40) in PBST containing 1% normal serumfor 72 hours at 4°C. The sections were further incubatedwith biotinylated anti-mouse IgG (dilution 1:200) for 2hours, Texas Red streptavidin (dilution 1:200) for 2 hours,polyclonal AVP or OXT antibody (dilution 1:2, 000) for 24hours at 4°C, and FITC-labeled anti-rabbit IgG (dilution1:50) for 2 hours. The sections were finally mounted ongelatin-coated slides and covered with Vectashield mount-ing medium. The distribution of fluorophores in tissuesections was visualized with an Axioplan 2MOT epifluo-rescence microscope coupled to a Carl Zeiss laser-scanningconfocal imaging system (LSM510). The recording config-uration (sequential scanning) for FITC and rhodaminewas used to avoid bleed-through of FITC. Images (1,024 31,024) were saved as TIF files by using Zeiss LSM510Image-Browser software for Windows and printed onCP700DSA, a MITSUBISHI color printer.
Preparation of tissue homogenates
Tissue samples were made from well-hydrated males (n 515, control), 2-day NaCl-treated rats (n 5 10), and 7-dayNaCl-treated rats (n 5 10). Animals were killed by decapi-tation, and brains and hypophyses were rapidly removed.Small blocks of tissue containing the SON and the neurohy-pophysis were dissected out and quickly frozen on dry ice.The tissue was homogenized with a glass homogenizer withan appropriate amount of PBS (pH 7.4) containing 1 mMPMSF, 0.1% aprotinin, 10 mg/ml leupeptin, and 10 mg/mlpepstatin. The protein content was determined by the Lowrymethod (Lowry, 1951), and the homogenate was dissolved inthe electrophoresis sample buffer (pH 6.8) and boiled for 3minutes. In some experiments, tissue homogenates wereultracentrifuged for 60 minutes at 100,000g at 4°C, and theresultant supernatants and pellets were referred to as thesoluble and insoluble proteins, respectively. The pellets werefurther treated with homogenate buffer containing 2% Tri-ton X-100 at 4°C and ultracentrifuged for 60 minutes at100,000g to obtain the Triton-soluble and Triton-insolublefractions.
Western blotting
The boiled protein sample was separated by using 10%SDS polyacrylamide gel electrophoresis (PAGE). Electroblot-ting was performed on polyvinylidene difluoride membranes(pore size, 0.45 mm; Millipore, Bedford, MA) in 20% metha-nol, 25 mM Tris, 192 mM glycine, and 0.05% SDS. Themembranes were pretreated with 5% nonfat dried milk in 50mM Tris-buffered (pH 7.4) saline containing 0.5% Tween 20for 24 hours at 4°C. The membranes were incubated witheither monoclonal anti-Kilon or anti-OBCAM antibody (di-lution 1:1,000) in Tris-buffered saline/0.5% Tween 20 con-taining 0.2% bovine serum albumin for 2 hours at 37°C.Membranes were further incubated with HRP-conjugatedanti-mouse IgG (0.05mg/ml) in Tris-buffered saline/0.5%Tween 20 for 2 hours at 37°C. The peroxidase was finallyactivated with enhanced chemiluminescence, and the spe-cific bands of Kilon and OBCAM were visualized on x-rayfilm. Western blots were scanned with an Apple ColourOneScanner and saved as TIF files by using Photoshop ver 3.0 ona Macintosh computer. The images were printed on FujiFilm Pictorography 3000.
RESULTS
Specificity of anti-Kilon andanti-OBCAM antibodies
Since both Kilon and OBCAM belong to the IgLONsuperfamily and have high degrees of sequence homology(49%; SWISS-PROT data bases, Funatsu et al., 1999), it isvery important to obtain specific antibodies for immuno-logical studies. The 89B3 antibody was found to react witha 46-kDa band in the brain raft fraction and also specifi-cally with the bacterially expressed Kilon fusion protein(GST-Kilon) but not with bN-OBCAM (Fig. 1). After puri-fication by a glutathione-Sepharose column chromatogra-phy, only GST-Kilon at 60 kDa was detected with 89B3antibody. Therefore, the lower band at about 40 kDa wasa GST-free Kilon. This anti-Kilon antibody reacted withone broad band with an apparent molecular mass of 46kDa in the raft fraction. In contrast, 84A3 antibody rec-ognized two broad bands of around 46 and 51 kDa of thebrain raft and also reacted with the bacterially expressedbN-OBCAM but not with GST-Kilon. Two independentgroups have reported that OBCAM in the rat brain hastwo broad bands of around 50 kDa (Hachisuka et al., 1996;Wick et al., 1996). The 89B3 and 84A3 antibodies did not
Fig. 1. Characterization of monoclonal antibodies against Kilon(clone 89B3) and OBCAM (clone 84A3). The rafts from rat brain (3mg/lane) and bacterial cell extracts (2.5 mg/lane) were electrophoresedand processed for Western blotting. Left panel: The 89B3 antibodyrecognized the 46-kDa band of Kilon in the raft and bacterially ex-pressed Kilon (GST-Kilon) but not the bacterially expressed andN-terminal-deleted OBCAM (DOBCAM). Arrows indicate the locationof Kilon (46 kDa) in raft and the GST-Kilon fusion protein (60 kDa).Right panel: The 84A3 antibody recognized the 51- and 46-kDa bandsof OBCAM in the raft and the bacterially expressed OBCAM (DOB-CAM) but not the bacterially expressed Kilon (GST-Kilon). Arrowsindicate the location of OBCAM (51 and 46 kDa) in the raft andDOBCAM (30 kDa).
77IgLON IN MAGNOCELLULAR NEURONS

react with the bacterially expressed GST alone (data notshown). Highly glycosylated proteins generally showbroad bands on SDS-PAGE. Thus, 89B3 and 84A3 anti-bodies specifically recognized Kilon and OBCAM, respec-tively.
Western blotting of the tissue homogenates
To investigate whether Kilon and OBCAM are ex-pressed specifically in the HNS and also whether they areup- or downregulated by chronic osmotic stimulation, wecarried out Western blotting analyses on homogenates ofthe adult rat SON and neurohypophysis (Fig. 2). Identicalamounts of homogenate (5 mg/lane) from the SON andneurohypophysis of well-hydrated and NaCl-treated ani-mals were separated on 10% SDS gel and transferred ontopolyvinylidene difluoride membranes. The SON displayeda specific single Kilon band at 46 kDa (lanes 1–3) and twobroad OBCAM bands at approximately 46 and 51 kDa(lanes 7–9), but the neurohypophysis did not show anydetectable signals for Kilon (lanes 4–6) or OBCAM (lanes10–12). The same results were obtained when more of theneurohypophysial homogenate (20 mg/lane) was loaded.The intensity of Kilon labeling in well-hydrated rats (lane1) was similar to those of 2- and 7-day NaCl-treated rats(lane 1 and 3). Similarly, the intensity of OBCAM labelingin 2- and 7-day NaCl-treated rats (lanes 8 and 9) did notshow any notable changes compared with well-hydratedcontrol animals (lane 7).
To investigate whether each IgLON protein is presentin the insoluble or soluble fraction, or in the Triton-insoluble or Triton-soluble fraction of brain membrane,pellets were subjected to ultracentrifugation and deter-
gent treatment (Fig. 3). IgSF proteins of CAMs such as F3,LAMP, NCAM120, and Ntm have both insoluble and sol-uble forms (He et al., 1987; Durbec et al., 1992; Zhukarevaet al., 1997). Both Kilon (lanes 1–3) and OBCAM (lanes6–8) labeling were observed only in the pellet fractions,demonstrating that these molecules are exclusivelypresent in the insoluble or membrane-bound forms in theSON. Most Kilon was present in the Triton-insoluble frac-tion (lanes 4 and 5), although OBCAM was localized pref-erentially in the Triton-soluble fractions (lane 9), com-pared with the Triton-insoluble fraction (lane 10). Thissuggested that the localization of Kilon in the submicrodo-main within the neuronal membrane is different from thatof OBCAM.
Distribution of Kilon and OBCAMin the HNS
For single labeling, immunohistochemistry for Kilonand OBCAM was performed in well-hydrated and NaCl-treated rats by using the avidin-biotin peroxidase method.Stronger Kilon and OBCAM immunoreactivities were ob-served in the PVN (Fig. 4A,B) and SON (Fig. 5A,B) com-pared with adjacent hypothalamic tissues. In the PVN,Kilon and OBCAM immunoreactivities were seen in mag-nocellular neurons but not in parvocellular neurons, judg-ing by their cell size (Swanson and Sawchenko, 1983).Higher magnification revealed that stronger immunoreac-tivities of Kilon and OBCAM were seen in dendrites com-pared with the somata of magnocellular neurons in thePVN (Fig. 4C,D). Weak OBCAM immunoreactivity wasalso observed in axonal fibers (Fig. 4B,D) and in the in-ternal zone of the median eminence (data not shown).
Fig. 2. Western blotting analysis of Kilon (lanes 1–6) and OBCAM(lanes 7–12) expression in the adult rat SON and neurohypophysisunder well-hydrated and dehydrated (NaCl-treated) conditions. Com-parison of Kilon and OBCAM expression in the SON (5 mg/lane) ofwell-hydrated (lanes 1 and 7), 2-day NaCl-treated (lanes 2 and 8), and7-day NaCl-treated rats (lanes 3 and 9), or in the neurohypophysis (5mg/lane) of well-hydrated animals (lanes 4 and 10), 2-day NaCl-treated (lanes 5 and 11), and 7-day NaCl-treated rats (lanes 6 and 12).The prominent band migrating at 46 kDa, corresponding to the pre-dicted size of Kilon, was detected in the SON but not in the neurohy-pophysis. Two isoforms of OBCAM were detected at 46 and 51 kDa inthe SON but not in the neurohypophysis. Note that chronic osmoticstimulation by 2% NaCl induced no visible differences in the levels ofKilon or OBCAM expression in the SON and neurohypophysis.
Fig. 3. Western blotting analysis of Kilon (lanes 1–5) and OBCAM(lanes 6–10) in the insoluble pellet and Triton-insoluble fractions ofthe adult rat SON. Whole homogenates (lane 1 and 6) prepared fromtissue pieces of the SON were centrifuged at 100,000g for 60 minutesto yield the soluble (supernatant, lanes 2 and 7) and insoluble frac-tions (pellets, lanes 3 and 8). The pellets were then treated withTriton-X 100 and centrifuged once to obtain the Triton-soluble frac-tions (supernatant, lanes 4 and 9) and the Triton-insoluble fraction(pellets, lanes 5 and 10). Note that Kilon was found essentially in theTriton-insoluble fraction, but OBCAM was seen in the Triton-solublefractions at higher levels compared with that in the Triton-insolublefraction.
78 S. MIYATA ET AL.

Fig. 4. Immunocytochemical localization of Kilon (A, C) and OB-CAM (B, D) in coronal sections of the PVN in the rat hypothalamus.A,C: Strong Kilon immunoreactivity was detected in the magnocellu-lar neurons. Higher magnification reveals that the Kilon immunore-activity was stronger in dendrites (arrowheads) compared withsomata (arrow) of the magnocellular neuron. B,D: The immuno-cytochemical localization of OBCAM in the PVN was similar to that of
Kilon. Intense OBCAM immunoreactivity was seen in the PVN, andlabeling in dendrites (large arrowheads) was stronger than that insomata (arrow). Faint immunoreactivity was also seen in the axonalfibers (small arrowheads). Note the absence of prominent immunore-activity of Kilon and OBCAM in the adjacent hypothalamus. v, thirdventricle. Scale bars 5 100 mm.
79IgLON IN MAGNOCELLULAR NEURONS

Kilon and OBCAM immunoreactivies were also observedin the SON of the hypothalamus, and, as seen in the PVN,intense immunoreactivities were observed in the dendriticzone of the SON (Fig. 5A,B). The SON dendrites tend toproject ventrally until they reach the ventral glial lamina,at which point they turn and extend either in a roughlyrostrocaudal direction or ventrolaterally along the peri-amygdaloid cortical surface. Therefore, this dendrite-gathering region is called the dendritic zone (Hatton,1990).
In the neurohypophysial sections stained with a highconcentration of antibodies (1:500), weak OBCAM label-ing was seen (Fig. 5D), but Kilon immunoreactivity wasscarcely observed (Fig. 5C). There were no specific Kilonand OBCAM immunoreactivities in the suprachiasmaticnucleus, where AVP neurons locate, and in other hypotha-lamic areas such as the preoptic area, lateral hypotha-lamic nucleus, and ventromedial hypothalamic nucleus(data not shown). Therefore, Kilon and OBCAM are spe-cifically and strongly expressed in the PVN and SONwithin the hypothalamus. Treatment of animals with 2%NaCl in drinking water for 7 days did not change the
locations or intensities of Kilon and OBCAM immunore-activity in the SON, PVN, and neurohypophysis (data notshown).
Double-labeling study withconfocal microscopy
Confocal image analysis allowed the simultaneous visu-alization of IgLON proteins (Kilon and OBCAM) and neu-ropeptides (AVP and OXT) in the same sections by usingTexas Red streptavidin and FITC (Fig. 6). Strong immu-noreactivities of both Kilon and OBCAM were limited todendrites, compared with somata of magnocellular neu-rons. Kilon immunoreactivity was usually found to over-lap with some dendrites of AVP-secreting magnocellularneurons (Fig. 6A–C), but Kilon-positive dendrites some-times revealed OXT immunoreactivity (Fig. 6D–F). In con-trast, dendrites of AVP-secreting magnocellular neuronsexclusively revealed strong OBCAM labeling (Fig. 6G–I),and no dendrites of OXT-positive neurons contained OB-CAM immunoreactivity (Fig. 6J–L). Some portions ofOXT-positive dendrites were seen as yellow, but this was
Fig. 5. Immunocytochemical localization of Kilon (A, C) and OB-CAM (B, D) in coronal sections of the SON and horizontal sections ofthe neurohypophysis. A,B: In the SON, strong Kilon and OBCAMimmunoreactivities were seen in the dendrites (arrowheads) of themagnocellular neurons, whereas signals for both Kilon and OBCAM
in the magnocellular somata (arrows) were weak. C,D: In the neuro-hypophysis, no Kilon immunoreactivity was seen, but weak labeling ofOBCAM (arrowheads) was observed. Scale bars 5 100 mm in A (ap-plies to A,B); 50 mm in C (applies to C,D).
80 S. MIYATA ET AL.
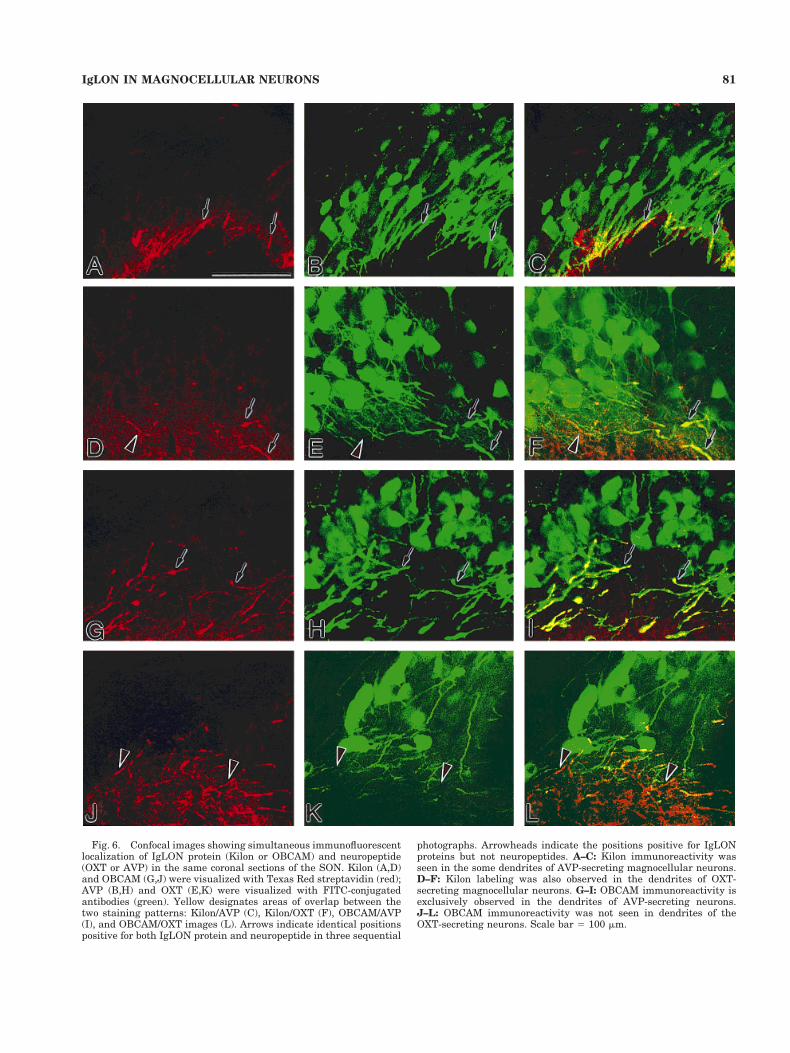
Fig. 6. Confocal images showing simultaneous immunofluorescentlocalization of IgLON protein (Kilon or OBCAM) and neuropeptide(OXT or AVP) in the same coronal sections of the SON. Kilon (A,D)and OBCAM (G,J) were visualized with Texas Red streptavidin (red);AVP (B,H) and OXT (E,K) were visualized with FITC-conjugatedantibodies (green). Yellow designates areas of overlap between thetwo staining patterns: Kilon/AVP (C), Kilon/OXT (F), OBCAM/AVP(I), and OBCAM/OXT images (L). Arrows indicate identical positionspositive for both IgLON protein and neuropeptide in three sequential
photographs. Arrowheads indicate the positions positive for IgLONproteins but not neuropeptides. A–C: Kilon immunoreactivity wasseen in the some dendrites of AVP-secreting magnocellular neurons.D–F: Kilon labeling was also observed in the dendrites of OXT-secreting magnocellular neurons. G–I: OBCAM immunoreactivity isexclusively observed in the dendrites of AVP-secreting neurons.J–L: OBCAM immunoreactivity was not seen in dendrites of theOXT-secreting neurons. Scale bar 5 100 mm.
81IgLON IN MAGNOCELLULAR NEURONS

due to accidental crossing of OBCAM-positive dendritesover those positive for OXT rather than co-localization.
Distribution of F3 and Thy-1 in the SON
F3 and Thy-1 immunohistochemistry were performed inwell-hydrated normal male rats (Fig. 7). F3 immunoreac-tivity was visible mainly in somata of magnocellular neu-rons but not in the dendritic zone (Fig. 7A). F3 immuno-reactivity was also seen in the neurohypophysis aspunctate staining (data not shown). Therefore, F3 waspresent in the terminals of magnocellular neurons. Thy-1immunoreactivity was seen as punctate staining through-out the somatic and dendritic zones of the SON (Fig. 7B).Therefore, Thy-1 immunoreactivity probably representsthe labeling of afferent synapses that come from extra-SON regions. Thus, the other IgSF proteins F3 and Thy-1evidently have different distributions from Kilon and OB-CAM.
DISCUSSION
The major finding of the present study was that twoIgLON proteins, Kilon and OBCAM, are specifically ex-pressed in the hypothalamic magnocellular neurons andare especially targeted to dendrites. Immunocytochemicaland Western blotting experiments demonstrated intenseKilon and OBCAM immunoreactivities in the hypotha-lamic SON and PVN. A double-labeling study using con-focal laser scanning microscopy further demonstrated thatstrong Kilon and OBCAM immunoreactivities werepresent specifically in the dendrites. Kilon was seen to belocalized mainly in dendrites of AVP-secreting magnocel-lular neurons and was occasionally detected in those se-creting OXT. However, OBCAM was observed exclusivelyin dendrites of AVP-secreting neurons. These results sug-gest that Kilon and OBCAM are possibly responsible fordendritic plasticity of the hypothalamic magnocellularneurons.
Technical considerations
Monoclonal antibodies for Kilon (89B3) and OBCAM(84A3) were produced in our previous study (Funatsu etal., 1999). The specificity of anti-Kilon antibody was char-acterized in our previous study (Funatsu et al., 1999). Inthe present study, furthermore, the specificity of bothanti-Kilon and anti-OBCAM antibodies was well docu-mented by using bacterially expressed Kilon and OBCAM.Both monoclonal antibodies were produced by using thePNGF-treated raft fraction as the antigen. Anti-Kilon an-tibody was found to react with a 46-kDa species in thebrain raft and also reacted specifically with the bacteriallyexpressed recombinant Kilon but not with recombinantOBCAM. In contrast, anti-OBCAM antibody recognizedtwo broad bands of around 46 and 51 kDa in the brain raftand also reacted with recombinant OBCAM but not withrecombinant Kilon.
Immunocytochemical analysis showed strong OBCAMlabeling in the cerebral and hippocampal neurons by using84A3 anti-OBCAM antibody (data not shown). This obser-vation is in good agreement with in situ hybridizationexperiments showing that OBCAM mRNA is strong in thecerebral cortex and hippocampus (Struyk et al., 1995).Two independent groups reported that OBCAM in the ratbrain has two broad bands of around 50 kDa (Hachisukaet al., 1996; Wick et al., 1996). Since the molecular mass ofLAMP is 65 kDa and that of Ntm is 64–68 kDa, muchlarger than Kilon and OBCAM, 89B3 and 84A3 antibodieswere presumed to recognize specifically Kilon and OB-CAM, respectively.
Expression of IgSF proteins in the HNS
The present immunohistochemical study demonstratedintense Kilon and OBCAM immunoreactivity in the mag-nocellular neurons of the hypothalamus, but no prominentimmunoreactivity was observed in the parvocellular neu-rons in the PVN and AVP neurons in the suprachiasmaticnucleus. There was also no specific immunoreactivity inthe outside of the SON and PVN within the hypothala-mus. IgLON proteins such as LAMP, OBCAM, Ntm, andKilon, are known to have a restricted distribution and areexpressed at high levels principally in the cortical plateand hippocampus. LAMP and Ntm have a largely comple-mentary expression pattern, but LAMP is confined to thelimbic cortex, and Ntm is expressed mostly in the senso-
Fig. 7. Immunocytochemical localization of F3 and Thy-1 in theSON of the rat hypothalamus. A: Intense F3 immunoreactivity wasseen in the somata (arrow) of the magnocellular neurons, but noimmunoreactivity was seen in the dendritic zone (arrowheads).B: Thy-1 immunoreactivity was observed as punctate labeling in thesomatic (arrow) and dendritic zones (arrowheads), but not in thesomata itself. ot, optic tract. Scale bars 5 100 mm.
82 S. MIYATA ET AL.

rimotor cortex (Struyk et al., 1995). OBCAM has a muchmore restricted distribution, with the highest expressionin the cortical cortex and hippocampus (Struyk et al.,1995). Kilon also shows a distribution pattern similar tothose of LAMP and Ntm in the cerebral cortex and hip-pocampus (Funatsu et al., 1999).
To date, studies of the IgLON proteins have focused onthe classically defined limbic system, and the hypotha-lamic regions have been mostly ignored. There has beenonly one preliminary report concerning the hypothalamus,in which in situ hybridization experiments indicated ex-pression of LAMP mRNA predominantly in the magnocel-lular neurons of the hypothalamic SON and PVN, but notin the parvocellular neurons (Reinoso et al., 1996). In thisstudy, expression of LAMP mRNA in magnocellular neu-rons was the most intense within the hypothalamus, andthe expression level of LAMP mRNA was the same as thatseen in the limbic areas such as the hippocampus andamygdala. From these results, we concluded that threeIgLON proteins, LAMP, Kilon, and OBCAM, are abun-dantly expressed in the hypothalamic magnocellular neu-rons at the same level as that seen in the limbic system.
The HNS shows structural reorganization, or plasticity,of neurons and glial cells. However, the cellular mecha-nisms and molecules underlying such structural plasticityare not completely understood. Possible candidates for themechanism of structural plasticity are CAMs of IgSF pro-teins and extracellular matrix glycoproteins. PSA-NCAMand F3 are composed of Ig- and fibronectin-III-like do-mains and are principally expressed by neuronal sub-groups in the developing brain, compared with the matureadult brain. However, the HNS continues to express sub-stantial levels of PSA-NCAM (Theodosis et al., 1991; Bon-fanti et al., 1992; Kiss et al., 1993) and F3 (Olive et al.,1995; Pierre et al., 1998) even in adults. PSA-NCAM im-munoreactivity was observed in the magnocellular nucleiand neurohypophysis, in glial cells, dendrites, and axonsterminals but not in somata (Theodosis et al., 1991). Theexpression of PSA-NCAM is not affected by physiologicalconditions such as dehydration and lactation. F3 is notvisible on any cell surface in the magnocellular nuclei butis present in secretory granules and the surface of termi-nals and glial cells (pituicytes) in the neurohypophysis,suggesting that F3 released into extracellular space actsas bioactive component (Pierre et al., 1998). Tenascin-C isan extracellular matrix glycoprotein and a member of thetenascin gene family. Its immunoreactivity was detectedin the extracellular spaces and astrocytic surface of mag-nocellular nuclei and in the perivascular space and glialand axonal surface (Singleton and Salm, 1996; Theodosiset al., 1997). F3, PSA-NCAM, and tenascin-C are known tobe highly expressed in embryonic and early postnatalbrains; Kilon and OBCAM were observed mainly in adultbrains (Funatsu et al., 1999). The high levels of Kilon andOBCAM expression suggest that they may be concernedwith structural plasticity in the hypothalamic magnocel-lular nuclei in addition to the above molecules.
Functional implications of IgLONs inmagnocellular dendrites
The present double-labeling study using confocal laserscanning microscopy demonstrated that both Kilon andOBCAM were present in the dendrites of magnocellularneurons. Weak immunoreactivity of OBCAM was also ob-served at axonal fibers, the internal zone of the median
eminence, and the neurohypophysis, only when a highconcentration of antibody was used. Western blots of Kilonand OBCAM were under detection level in the neurohy-pophysis. Therefore, dendrites of magnocellular neuronscontain much abundant OBCAM, compared with tracelevels of OBCAM in the axons and terminals. Althoughthe reason why Kilon and OBCAM are targeted to mag-nocellular dendrites is not yet clear, a possible explana-tion was suggested by the previous morphological studiesindicating that physiological conditions such as dehydra-tion and lactation alter the states of dendritic organization(Perlmutter et al., 1984, 1985).
Dendrites of the SON descend to the ventral glial lam-inar area near the pial surface, and chronic physiologicalstimulation results in increased membrane apposition andthe formation of dendritic bundles (two or more dendritesapposed without interposing glial processes). Membraneapposition of dendritic bundles was seen within the sameneuronal populations—immunologically identified AVP orOXT neurons (Theodosis et al., 1986). Dye transfer couldbe observed within the same neuronal populations, possi-bly via membrane gap junctions between dendrites (Cob-bett et al., 1985; Hatton and Yang, 1996; Yang and Hat-ton, 1999). Studies in vitro demonstrated homophilicbinding between LAMP molecules, thereby selectivelypromoting neurite outgrowth of the limbic neurons(Zhukareva and Levitt, 1995). Neonatal administration ofanti-LAMP in vivo resulted in a disruption of normalgrowth patterns of the septohippocampal fiber tract (Pi-menta et al., 1995). Functional analyses thus indicatedthat LAMP acts as a selective adhesion molecule, servingas a guidance cue for specific patterns of connectivity. Ntmis also known to promote the outgrowth and neuronalattachment of Ntm-expressing primary neurons in thedorsal root ganglion and hippocampal neurons but not ofsuperior cervical ganglion neurons, which do not expressNtm (Gil et al., 1998). In this experiment, most of theKilon and OBCAM molecules were present as themembrane-bound forms, suggesting that they might par-ticipate in dendritic connectivity via target recognition. IfKilon and OBCAM are similar to Ntm, they might con-tribute to promotion of neuronal attachment withinIgLON-positive dendrites and inhibit that betweenIgLON-negative dendrites. It is likely that IgLON pro-teins are responsible for recognition of dendrites and pos-sibly structural remodeling of dendrites.
Kilon immunoreactivity was also observed, mainly inthe dendrites of AVP-secreting magnocellular neurons; itwas also seen in some of those secreting OXT. As OBCAMwas observed exclusively in dendrites of AVP-secretingmagnocellular neurons, AVP dendrites should containboth Kilon and OBCAM. The reason why two IgLON pro-teins are expressed in the same magnocellular dendrites isnot known. However, the present biochemical study sug-gested one possible explanation; most Kilon was presentin the Triton-insoluble fraction, but OBCAM was localizedmainly in the Triton-soluble fractions. In a previous study,OBCAM was shown to be localized mainly in the Triton-insoluble fraction by using whole brain homogenates (Ha-chisuka et al., 1996). The raft is a membranous subdomainthat is insoluble in nonionic detergents such as TritonX-100. This domain contains many signal-transducingmolecules such as trimeric G proteins, protein tyrosinekinase, cytoskeletal proteins, and calmodulin-binding pro-teins (Maekawa et al., 1997; Simons and Ikonen, 1997).
83IgLON IN MAGNOCELLULAR NEURONS

The detergent insolubility of this fraction is attributed tothe enrichment of cholesterol and sphingomyelin (Schroe-der et al., 1998). Extracellular proteins anchored in thelipid layer by GPI are generally considered to be localizedin the raft (Varma and Mayor, 1998). They reveal differentlocalizations within the membranous subdomain of AVP-secreting dendrites and thus have different functional sig-nificance.
Other IgSF proteins in the HNS
The present observation that F3 was located in themagnocellular somata and terminals is consistent withthe previous results (Olive et al., 1995; Theodosis et al.,1998). F3 was not observed on any cell surface in themagnocellular nuclei but was present not only in secretorygranules but also on the surfaces of axon terminals andglia and in the extracellular spaces. Thy-1 is known as anaxonal protein, and therefore, in the present study, Thy-1immunoreactivity was also observed as punctate stainingin many afferent synapses on magnocellular somatic anddendritic zones. Another IgSF protein, PSA-NCAM, is ex-pressed in the dendrites, axons, terminals, and associatedastrocytes (Theodosis et al., 1991; Bonfanti et al., 1992;Kiss et al., 1993).
Thus, the observations that each GPI-anchored IgSFshows a different subcellular localization within magno-cellular neurons may explain why there are many GPI-anchored IgSF proteins in the brain. In addition, in thisexperiment, no major differences were observed by West-ern blotting or immunohistochemistry in the quantity ordistribution in animals under elevated hormonal demandwith dehydration. In comparison with other IgSF andextracellular matrix components, tenascin-C and PSA-NCAM expression were not affected by age, sex, or phys-iological conditions such as dehydration or lactation(Theodosis et al., 1991; Bonfanti et al., 1992; Singletonand Salm, 1996; Theodosis et al., 1997), although F3 ex-pression is upregulated with dehydration and lactation(Olive et al., 1995; Pierre et al., 1998). Similar totenascin-C and PSA-NCAM, Kilon and OBCAM are con-stitutively produced in the magnocellular neurons andmay serve as permissive factors to allow magnocellulardendrites to undergo morphological changes in response toappropriate stimuli.
In conclusion, Kilon and OBCAM, IgLON proteins,which have been shown to be abundant in the cortical andsubcortical neurons of the limbic system, are also ex-pressed specifically in the magnocellular neurons of thehypothalamus. Moreover, they are targeted to magnocel-lular dendrites, suggesting specific relationships to den-dritic function. Thus, high levels of IgLON protein expres-sion may be correlated with the structural plasticity ofmagnocellular neurons.
ACKNOWLEDGMENTS
S. Miyata received a Scientific Research grant from theJapan Society for the Promotion of Science (no. 11640663),and T. Kiyohara received a Scientific Research grant fromthe Ministry of Education, Science, Sports and Culture ofJapan (no. 10470016).
LITERATURE CITED
Ben-Barak Y, Russel JT, Whitnell MH, Ozato K, Gainer H. 1985. Neuro-physin in the hypothalamo-neurohypophysial system. I. Productionand characterization of monoclonal antibodies. J Neurosci 5:81–97.
Bicknell RJ, Luckman SM, Inenaga K, Mason WT, Hatton GI. 1989.b-Adrenergic and opioid receptors on pituicytes cultured from adult ratneurohypophysis: regulation of cell morphology. Brain Res Bull 22:379–388.
Bobak JB, Salm AK. 1996. Plasticity of astrocytes of the ventral gliallimitants subjacent to the supraoptic nucleus. J Comp Neurol 376:188–197.
Bonfanti L, Olive S, Poulain DA, Theodosis DT. 1992. Mapping of thedistribution of plysialylated neural cell adhesion molecule throughoutthe central nervous system of the adult rat: an immunohistochemicalstudy. Neuroscience 49:419–436.
Chuong C-M, Edelman GM. 1984. Alterations in neural cell adhesionmolecules during development of different regions of the nervous sys-tem. J Neurosci 4:2354–2368.
Cobbett P, Smithson KG, Hatton GI. 1985. Dye-coupled magnocellularpeptidergic neurons of the rat paraventricular nucleus show homotypicimmunoreactivity. Neuroscience 16:885–895.
Cunningham ET, Sawchenko PE. 1991. Reflex control of magnocellularvasopressin and oxytocin secretion. Trends Neurosci 14:406–411.
Durbec P, Gennarini G, Goridis C, Rougan G. 1992. A soluble form of theF3 neuronal cell adhesion molecule promotes neurite outgrowth. J CellBiol 117:877–887.
Finne J, Finne U, Deagostini-Bazin H, Goridis C. 1983. Occurrence ofalpha 2-8-linked polysialosyl units in a neural cell adhesion molecule.Biochem Biophys Res Commun 112:482–487.
Funatsu N, Miyata S, Kumanogoh H, Shigeta M, Hamada K, Endo Y,Sokawa Y, Maekawa S. 1999. Characterization of a novel rat brainglycosylphosphatidylinositol-anchored protein (Kilon), a member of theIgLON cell adhesion molecule family. J Biol Chem 274:8224–8230.
Gennarini G, Cibelli G, Rougan G, Mattei M, Goridis C. 1989. The mouseneuronal cell surface protein F3: a phosphatidyl-inositol-anchoredmember of the immunoglobulin superfamily related to chicken contac-tin. J Cell Biol 109:775–788.
Gil OD, Zanazzi G, Struyk AF, Salzer JL. 1998. Neurotrimin mediatesbifunctional effects on neurite outgrowth via homophilic and hetero-philic interactions. J Neurosci 18:9312–9325.
Hachisuka A, Yamazaki T, Sawada J, Terao T. 1996. Characterization andtissue distribution of opioid-binding cell adhesion molecule (OBCAM)using monoclonal antibodies. Neurochem Int 28:373–379.
Hatton GI. 1990. Emerging concepts of structural-function dynamics inadult brain: the hypothalamo-neurohypophysial system. Prog Neuro-biol 34:437–504.
Hatton GI. 1997. Function-related plasticity in the hypothalamus. AnnuRev Neurosci 20:375–397.
Hatton GI. 1999. Astroglial modulation of neurotransmitter/peptide re-lease from the neurohypophysis:present status. J Chem Neuroanat16:203–222.
Hatton GI, Yang QZ. 1996. Synaptically released histamine increases dyecoupling among vasopressinergic neurons of the supraoptic nucleus:mediation by H1 receptors and cyclic nucleotides. J Neurosci 16:123–129.
Hatton GI, Luckman SM, Bicknell RJ. 1991. Adrenaline activation ofb2-adrenoceptors stimulates morphological changes in astrocytes (pi-tuicytes) cultured from adult rat neurohypophyses. Brain Res Bull26:765–769.
Hawrylak N, Fleming JC, Salm AK. 1998. Dehydration and rehydrationselectively and reversibly alter glial fibrillary acidic protein immuno-reactivity in the rat supraoptic nucleus and subjacent glial limitants.Glia 22:260–271.
He HT, Finne J, Goridis C. 1987. Biosynthesis, membrane association andrelease of NCAM 120, a phosphatidylinositol-linked form of the neuralcell adhesion molecule. J Cell Biol 105:2489–2500.
Hoffman KB. 1998. The relationship between adhesion molecules andneuronal plasticity. Cell Mol Neurobiol 18:461–475.
Horton HL, Levitt P. 1988. A unique membrane protein is expressed onearly development of the limbic system axons and cortical targets.J Neurosci 8:4653–4661.
Hunkapillar T, Hood L. 1989. Diversity of the immunoglobulin gene su-perfamily. Adv Immunol 44:1–63.
Keller F, Levitt P. 1989. Developmental and regeneration-associated reg-
84 S. MIYATA ET AL.

ulation of the limbic system associated membrane protein in explantcultures of the rat brain. Neuroscience 28:455–474.
Kiss JZ, Wang C, Rougan G. 1993. Nerve-dependent expression of highpolysialic acid neural cell adhesion molecule in neurohypophysial as-trocytes of adult rats. Neuroscience 53:213–221.
Levitt P. 1984. A monoclonal antibody to limbic system neurons. Science223:299–301.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Protein measure-ment with the folin phenol reagent. J Biol Chem 93:265–275.
Luckman SM, Bicknell RJ. 1990. Morphological plasticity that occurs inthe neurohypophysis following activation of the magnocellular neuro-secretory system can be mimicked in vitro by beta adrenergic stimula-tion. Neuroscience 39:701–709.
Maekawa S, Kumanogoh H, Funatsu N, Takei N, Inoue K, Endo Y,Hamada K, Sokawa Y. 1997. Identification of NAP-22 and GAP-43(neuromodulin) as major protein components in a Triton insoluble lowdensity fraction of rat brain. Biochim Biophys Acta 1323:1–5.
Matsunaga W, Miyata S, Hashimoto Y, Lin S-H, Nakashima T, KiyoharaT, Matsumoto T. 1999a. Microtubule-associated protein2 (MAP2) inhypothalamo-neurohypophysial system: low molecular weight MAP2 inpituitary astrocytes. Neuroscience 88:1429–1437.
Matsunaga W, Miyata S, Kiyohara T. 1999b. Redistribution of MAP-2immunoreactivity in the neurohypophysial astrocytes of adult ratsduring dehydration. Brain Res 820:7–17.
Miyata S, Furuya K, Nakai S, Bun H, Kiyohara T. 1999. Morphologicalplasticity and rearrangement of cytoskeletons in pituicytes culturedfrom adult rat neurohypophysis. Neurosci Res 33:299–306 .
Olive S, Rougon G, Pierre K, Theodosis DT. 1995. Expression of a glycosylphosphatidylinositol-anchored adhesion molecule, the glycoprotein F3,in the adult rat hypothalamo-neurohypophysial system. Brain Res689:271–280.
Orlando DG, Zanazzi G, Struyk AF, Salzer J. 1998. Neurotrimin mediatesbifunctional effects on neurite outgrowth via homophilic and hetero-philic interactions. J Neurosci 18:9312–9325.
Perlmutter LS, Tweedle CD, Hatton GI. 1984. Neuronal/glial plasticity inthe supraoptic dendritic zone: dendritic bundling and double synapseformation at parturition. Neuroscience 13:769–779.
Perlmutter LS, Tweedle CD, Hatton GI. 1985. Neuronal/glial plasticity inthe supraoptic dendritic zone in response to acute and chronic dehy-dration. Brain Res 361:225–232.
Pierre K, Gennariri G, Poulain DA, Theodosis DT. 1998. Regulated expres-sion of the cell adhesion glycoprotein F3 in adult hypothalamus. J Neu-rosci 18:5333–5343.
Pimenta AF, Zhukareva V, Barbe MF, Reinoso BS, Grimley C, Henzel W,Fischer I, Levitt P. 1995. The limbic system-associated membraneproteins is an Ig superfamily member that mediates selective neuronalgrowth and axon targetting. Neuron 15:287–297.
Pimenta AF, Reinoso BS, Levitt P. 1996. Expression of the mRNAs encod-ing the limbic system-associated membrane protein (LAMP). I. Fetalrat brains. J Comp Neurol 375:289–302.
Reinoso BS, Pimenta AF, Levitt P. 1996. Expression of the mRNAs encod-ing the limbic system-associated membrane protein (LAMP). II. Adultrat brain. J Comp Neurol 375:274–288.
Salm AK, Smithson KG, Hatton GI. 1985. Lactation-associated redistribu-tion of the glial fibrillary acidic protein within the supraoptic nucleus.Cell Tissue Res 242:9–15.
Sawchenko PE, Swanson LW. 1982. Immunohistochemical identification ofneurons in the paraventricular nucleus of the hypothalamus thatproject to the medulla or to the spinal cord in the rat. J Comp Neurol205:260–272.
Schofield PR, McFarland KC, Hayflick JS, Wilcox JN, Cho TM, Roy S, LeeNM, Loh HN, Seeburg PH. 1989. Molecular characterization of a newimmunoglobulin superfamily protein with potential roles in opioidebinding and cell control. EMBO J 8:489–495.
Schroeder RJ, Ahmed SN, Zhu Y, London E, Brown DA. 1998. Cholesteroland sphingolipid enhance the Triton X-100 insolubility of glycosyl-phosphatidylinositol-anchored proteins by promoting the formation ofdetergent-insoluble ordered membrane domains. J Biol Chem 273:1150–1157.
Simons K, Ikonen E. 1997. Functional rafts in cell membrane. Nature387:569–572.
Singleton PA, Salm AK. 1996. Differential expression of tenascin by astro-cytes associated with the supraoptic nucleus (SON) of hydrated anddehydrated rats. J Comp Neurol 373:186–199.
Smithson KG, Suarez I, Hatton GI. 1990. Beta-adrenergic stimulationdecreases glial and increases neural contact with the basal lamina inrat neurointermediate lobes incubated in vitro. J Neuroendocrinol2:693–699.
Struyk AF, Canoll PD, Wolfgang MJ, Rosen CL, D’Eustachio P, Salzer JL.1995. Cloning of neurotrimin defines a new subfamily of differen-tially expressed neural cell adhesion molecules. J Neurosci 15:2141–2156.
Swaab DF, Nijveldt F, Pool CW. 1975. Distribution of oxytocin and vaso-pressin in the rat supraoptic and paraventricular nucleus. J Endocrinol67:461–462.
Swanson LW, Sawchenko PE. 1983. Hypothalamic integration: organiza-tion of the paraventricular and supraoptic nuclei. Annu Rev Neurosci6:269–324.
Theodosis DT, MacVicar B. 1996. Neuron-glia interactions in the hypothal-amus and pituitary. Trends Neurosci 19:363–367.
Theodosis DT, Poulain DA. 1993. Activity-dependent neuron-glial andsynaptic plasticity in the adult mammalian hypothalamus. Neuro-science 57:501–535.
Theodosis DT, Chapman DB, Montagnese C, Poulain DA, Morris JF. 1986.Structural plasticity in the hypothalamic supraoptic nucleus at lacta-tion affects oxytocin—but not vasopressin-secreting neurons. Neuro-science 17:661–678.
Theodosis DT, Rougon G, Poulain DA. 1991. Retention of embryonic fea-tures by an adult neuronal system capable of plasticity: polysialylatedneuroal cell adhesion molecule in the hypothalamo-neurohypophysialsystem. Proc Natl Acad Sci USA 88:5494–5498.
Theodosis DT, Bonfanti S, Olive S, Rougon G, Poulain DA. 1994. Adhesionmolecules and structural plasticity of the adult hypothalamo-neurohypophysial system. Psychoneuroendocrinology 19:455–462.
Theodosis DT, Pierre K, Cadoret MA, Allard M, Faissner A, Poulain DA.1997. Expression of high levels of the extracellular matrix glycoprotein,tenascin-C, in the normal adult hypothalamoneurohypophysial system.J Comp Neurol 379:386–398.
Theodosis DT, El Majdoubi M, Pierre K, Poulain DA. 1998. Factors gover-ing activity-dependent structural plasticity of the hypothalamoneuro-hypophysial system. Cell Mol Neurobiol 18:285–298.
Tweedle CD, Hatton GI. 1982. Magnocellular neuropeptidergic terminalsin neurohypophysis: rapid glial release of enclosed axons during par-turition. Brain Res Bull 8:205– 209.
Tweedle CD, Hatton GI. 1987. Morphological adaptability at neurosecre-tory axonal endings on the neurovascular contact zone of the rat neu-rohypophysis. Neuroscience 20:241–246.
Varma R, Mayor S. 1998. GPI-anchored proteins are organized in submi-cron domains at cell surface. Nature 394:798–801.
Wick MJ, Fan G, Loh HH. 1996. Expression of OBCAM-related cDNAclones in Cos 1 cells: evidence for a phosphatidylinositol linkage to thecell membrane. Brain Res Mol Brain Res 36:322–328.
Williams AF, Barclay AN. 1988. The immunoglobulin superfamily; do-mains for cell surface recognition. Annu Rev Immunol 6:381–405.
Yang QZ, Hatton GI. 1999. Nitric oxide via cGMP-dependent mechanismsincreases dye coupling and excitability of rat supraoptic nucleus neu-rons. J Neurosci 10:4270–4279.
Zacco A, Cooper V, Chantler P, Fisher-Hyland S, Horton HL, Levitt P.1990. Isolation, biochemical characterization and ultrastructural anal-ysis of the limbic system-associated membrane protein (LAMP), aprotein expressed by neurons comprising functional neural circuits.J Neurosci 10:73–90.
Zhukareva V, Levitt P. 1995. The limbic system-associated membraneprotein (LAMP) selectively mediates interactions with specific centralneuron populations. Development 121:1161–1172.
Zhukareva V, Chernevskaya N, Pimenta A, Nowycky M, Levitt P. 1997.Limbic system-associated membrane protein (LAMP) induces neuriteoutgrowth and intracellular Ca21 increase in primary fetal neurons.Mol Cell Neurosci 10:43–55.
85IgLON IN MAGNOCELLULAR NEURONS




















