
Expression of Adhesion Molecules in Chronic Serum Sickness in Rats
7
Expression of Adhesion Molecules in Chronic Serum Sickness in Rats Lorenzo Benatuil, Gustavo Parra, Jaimar Rinco ´n, Yasmir Quiroz, and Bernardo Rodrı ´guez-Iturbe Department of Immunobiology, Instituto de Investigaciones Biome ´dicas, Fundacite-Zulia, Centro de Cirugı ´a Experimental, Universidad del Zulia and Servicio de Nefrologı ´a, Hospital Universitario, Maracaibo, Venezuela Peripheral blood leukocytes infiltrate the kidney in chronic serum sickness (CSS). We therefore studied the expression of CD54 and its ligands CD18 and CD11b/c in CSS in 10 rats with CSS, 6 rats immunized similarly who did not developed proteinuria (no-CSS group), and 10 normal rats (control group). Intense (6 to 35 times more than controls) leukocyte infiltration was observed in CSS. The CSS group over-expressed CD54 in glomeruli and interstitium in association with increments in CD18- and CD11b/c-positive cells rang- ing 2.5 to 7 times the number found in controls. 75% of infiltrating leukocytes expressed CD18 and 87% ex- pressed CD11b/c. The non-CSS group had leukocyte infiltration and expression of adhesion molecules sim- ilar to control group. Adherence of CD43-positive cells to renal tissues was 4 times higher in renal tissue from CSS rats than to normal kidney. Pretreatment with corresponding Mabs reduced adherence by half. We concluded that over-expression of CD54 and its li- gands CD18 and CD11b/c in infiltrating leukocytes oc- cur in CSS. Binding experiments suggest the func- tional relevance of these molecules. © 1999 Academic Press Key Words: CD54; CD11b/c; CD18; CD45 in chronic serum sickness. INTRODUCTION It is now well recognized that leukocytes have a preeminent role in the pathogenesis of immune com- plex glomerulonephritis in experimental animals (1, 2) and man (3, 4). In fact, the intensity of the cellular infiltrate correlates with the severity of the serum sick- ness nephritis (5) and administration of antimacroph- age serum prevents the disease (2). The infiltration of circulating leukocytes in the kid- ney is made possible by the intercellular adhesion mol- ecules (6), which, therefore, play an important role in the pathogenesis of glomerulonephritis (7). Among these molecules, CD54 (intercellular adhesion mole- cule 1 (ICAM-1)) is expressed normally on the vascular endothelium and may be expressed by renal tubular cells and mesangial cells. CD18 (leukocyte function- associated antigen 1 b chain 5 LFA-1 b chain) is nor- mally expressed in leukocytes and acts as ligand for ICAM-1 (8). CD11b/c is expressed by monocytes and polymorphonuclear leukocytes and participates in the adherence of these cells to the vascular endothelium and mesangium (6, 9). Recent investigations demon- strated an overexpression of ICAM-1 in CSS associated to the monocyte and lymphocyte infiltration (10). The present studies were done to evaluate the expression of these molecules and to further define the leukocyte infiltrate (lymphocytes and monocytes) in the kidney of rats with chronic serum sickness (CSS) and to estab- lish in vitro their potential functional role in this ex- perimental model. MATERIAL AND METHODS Induction of chronic serum sickness. CSS was in- duced in 16 male Sprague–Dawley rats of 200 –250 g of weight, following the protocol described by Sa ´ nchez- Crespo et al. (11). Briefly, rats were immunized with subcutaneous (5 mg) ovoalbumin in complete Freund’s adjuvant, after 3 weeks received a similar dose but in incomplete Freund’s adjuvant and 1 week after the second immunization received daily intraperitoneal in- jections (i.p.) of 10 mg of ovoalbumin in PBS until the day of sacrifice. Before the immunization, and every 3 days after i.p. ovalbumin was started, samples of plasma and 24-h urine were obtained. Immunized rats were divided in two groups, CSS group and no-CSS group, depending on protein excre- tion. The rats of the CSS group (n 5 10) had protein- uria higher than 100 mg/24 h and were sacrificed 5– 6 weeks after the beginning of i.p. OA. The rats from the no-CSS group (n 5 6) with protein excretion lower than 100 mg/24 h were sacrificed at the 7th week. The con- trol group (n 5 10) was represented by rats that re- ceived daily i.p. PBS without antigen. These rats were studied similarly and were sacrificed at the 7th week. Proteinuria was determined by the sulfosalicylic acid method and results were expressed as excretion in 24 h (mg/24 h). In control rats, baseline proteinuria was 2.7 6 3.4 mg/24 h (Table 1); therefore we considered 10 mg/24 h (2SD above the mean) the upper limit of nor- mal. Anti-ovoalbumin antibody titers were determined twice weekly in plasma samples obtained immediately before administering the corresponding dose of i.p. Clinical Immunology Vol. 90, No. 2, February, pp. 196 –202, 1999 Article ID clim.1998.4642, available online at http://www.idealibrary.com on 1521-6616/99 $30.00 Copyright © 1999 by Academic Press All rights of reproduction in any form reserved. 196
-
Upload
udistrital -
Category
Documents
-
view
2 -
download
0
Transcript of Expression of Adhesion Molecules in Chronic Serum Sickness in Rats

ctCsgtwCiiipiitCccgct
s
ppaina
nettcecam
Clinical ImmunologyVol. 90, No. 2, February, pp. 196–202, 1999Article ID clim.1998.4642, available online at http://www.idealibrary.com on
1CA
Expression of Adhesion Molecules in Chronic Serum Sickness in Rats
Lorenzo Benatuil, Gustavo Parra, Jaimar Rincon, Yasmir Quiroz, and Bernardo Rodrıguez-Iturbe
Department of Immunobiology, Instituto de Investigaciones Biomedicas, Fundacite-Zulia, Centro de Cirugıa Experimental,
Universidad del Zulia and Servicio de Nefrologıa, Hospital Universitario, Maracaibo, VenezuelaIpaastptirlp
dwCsaisjddp
gtuwn1tcs
m(2mmt
Peripheral blood leukocytes infiltrate the kidney inhronic serum sickness (CSS). We therefore studiedhe expression of CD54 and its ligands CD18 andD11b/c in CSS in 10 rats with CSS, 6 rats immunizedimilarly who did not developed proteinuria (no-CSSroup), and 10 normal rats (control group). Intense (6o 35 times more than controls) leukocyte infiltrationas observed in CSS. The CSS group over-expressedD54 in glomeruli and interstitium in association with
ncrements in CD18- and CD11b/c-positive cells rang-ng 2.5 to 7 times the number found in controls. 75% ofnfiltrating leukocytes expressed CD18 and 87% ex-ressed CD11b/c. The non-CSS group had leukocyte
nfiltration and expression of adhesion molecules sim-lar to control group. Adherence of CD43-positive cellso renal tissues was 4 times higher in renal tissue fromSS rats than to normal kidney. Pretreatment withorresponding Mabs reduced adherence by half. Weoncluded that over-expression of CD54 and its li-ands CD18 and CD11b/c in infiltrating leukocytes oc-ur in CSS. Binding experiments suggest the func-ional relevance of these molecules. © 1999 Academic Press
Key Words: CD54; CD11b/c; CD18; CD45 in chronicerum sickness.
INTRODUCTION
It is now well recognized that leukocytes have areeminent role in the pathogenesis of immune com-lex glomerulonephritis in experimental animals (1, 2)nd man (3, 4). In fact, the intensity of the cellularnfiltrate correlates with the severity of the serum sick-ess nephritis (5) and administration of antimacroph-ge serum prevents the disease (2).The infiltration of circulating leukocytes in the kid-
ey is made possible by the intercellular adhesion mol-cules (6), which, therefore, play an important role inhe pathogenesis of glomerulonephritis (7). Amonghese molecules, CD54 (intercellular adhesion mole-ule 1 (ICAM-1)) is expressed normally on the vascularndothelium and may be expressed by renal tubularells and mesangial cells. CD18 (leukocyte function-ssociated antigen 1 b chain 5 LFA-1 b chain) is nor-
ally expressed in leukocytes and acts as ligand for b521-6616/99 $30.00opyright © 1999 by Academic Pressll rights of reproduction in any form reserved.
196
CAM-1 (8). CD11b/c is expressed by monocytes andolymorphonuclear leukocytes and participates in thedherence of these cells to the vascular endotheliumnd mesangium (6, 9). Recent investigations demon-trated an overexpression of ICAM-1 in CSS associatedo the monocyte and lymphocyte infiltration (10). Theresent studies were done to evaluate the expression ofhese molecules and to further define the leukocytenfiltrate (lymphocytes and monocytes) in the kidney ofats with chronic serum sickness (CSS) and to estab-ish in vitro their potential functional role in this ex-erimental model.
MATERIAL AND METHODS
Induction of chronic serum sickness. CSS was in-uced in 16 male Sprague–Dawley rats of 200–250 g ofeight, following the protocol described by Sanchez-respo et al. (11). Briefly, rats were immunized withubcutaneous (5 mg) ovoalbumin in complete Freund’sdjuvant, after 3 weeks received a similar dose but inncomplete Freund’s adjuvant and 1 week after theecond immunization received daily intraperitoneal in-ections (i.p.) of 10 mg of ovoalbumin in PBS until theay of sacrifice. Before the immunization, and every 3ays after i.p. ovalbumin was started, samples oflasma and 24-h urine were obtained.Immunized rats were divided in two groups, CSS
roup and no-CSS group, depending on protein excre-ion. The rats of the CSS group (n 5 10) had protein-ria higher than 100 mg/24 h and were sacrificed 5–6eeks after the beginning of i.p. OA. The rats from theo-CSS group (n 5 6) with protein excretion lower than00 mg/24 h were sacrificed at the 7th week. The con-rol group (n 5 10) was represented by rats that re-eived daily i.p. PBS without antigen. These rats weretudied similarly and were sacrificed at the 7th week.Proteinuria was determined by the sulfosalicylic acidethod and results were expressed as excretion in 24 h
mg/24 h). In control rats, baseline proteinuria was.7 6 3.4 mg/24 h (Table 1); therefore we considered 10g/24 h (2SD above the mean) the upper limit of nor-al. Anti-ovoalbumin antibody titers were determined
wice weekly in plasma samples obtained immediately
efore administering the corresponding dose of i.p.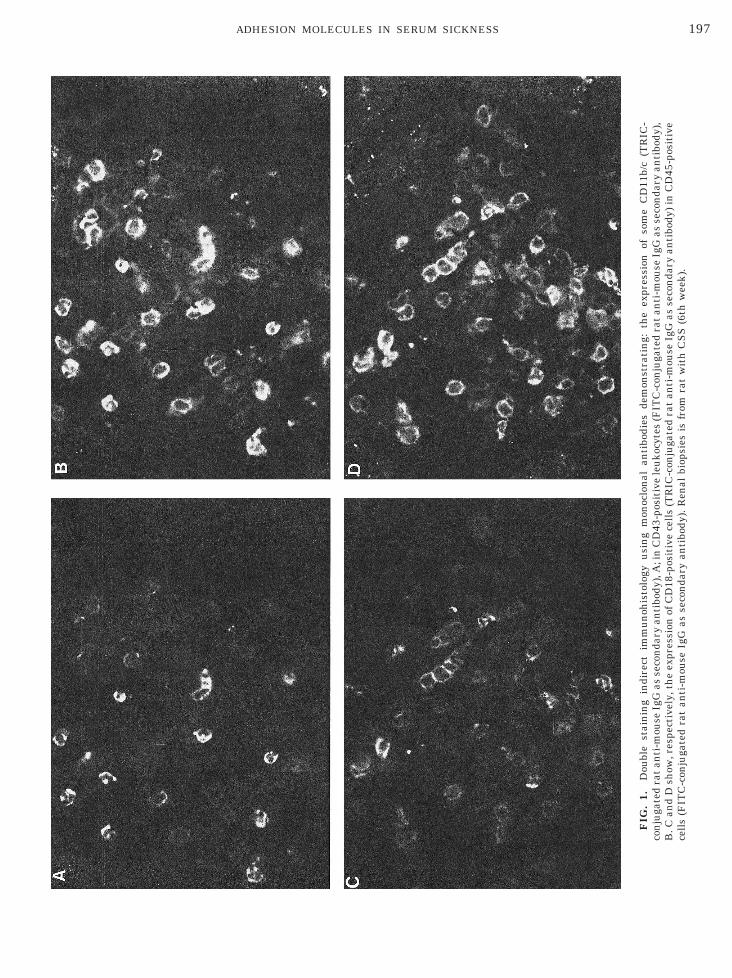
FIG
.1.
Dou
ble
stai
nin
gin
dire
ctim
mu
noh
isto
logy
usi
ng
mon
oclo
nal
anti
bodi
esde
mon
stra
tin
g:th
eex
pres
sion
ofso
me
CD
11b/
c(T
RIC
-co
nju
gate
dra
tan
ti-m
ouse
IgG
asse
con
dary
anti
body
),A
;in
CD
43-p
osit
ive
leu
kocy
tes
(FIT
C-c
onju
gate
dra
tan
ti-m
ouse
IgG
asse
con
dary
anti
body
),B
.Can
dD
show
,res
pect
ivel
y,th
eex
pres
sion
ofC
D18
-pos
itiv
ece
lls
(TR
IC-c
onju
gate
dra
tan
ti-m
ouse
IgG
asse
con
dary
anti
body
)in
CD
45-p
osit
ive
cell
s(F
ITC
-con
juga
ted
rat
anti
-mou
seIg
Gas
seco
nda
ryan
tibo
dy).
Ren
albi
opsi
esis
from
rat
wit
hC
SS
(6th
wee
k).
197ADHESION MOLECULES IN SERUM SICKNESS
oA
aktlclpSc&cmTclCtaLba
TeptwpS
icsflmt
t4McswCwsttCtnrpsmpopba
twtuCttFtCMCtrp
spg
198 BENATUIL ET AL.
valbumin by Ouchterlony double immunodiffusion.ntibody titers were expressed as log 2.
Monoclonal antibodies. The following monoclonalntibodies (Mab) were used: anti-CD45 (identifies leu-ocyte common antigen (LCA); Biosource Interna-ional, Camarillo, CA, mouse IgG1, distribution 5 alleukocytes) at a concentration of 30 mg/ml; anti CD43,lone W3/13 (identifies mainly T lymphocytes and to aesser degree rat thymocytes, polymorphonuclear cells,lasma cells, and stem cells) (Accurate Chemical &cientific Corp. Westbury, U.S.A., mouse IgG1, at aoncentration of 5 mg/ml; anti-ED1 (Accurate Chemical
Scientific Corp., mouse IgG1, distribution 5 mono-ytes and macrophages) used at a concentration of 30g/ml; anti-CD54 (identifies ICAM-1; Seikagaku Corp.okyo, Japan, mouse IgG1, distribution 5 endothelialells, monocytes, B and T cells, dendrite cells, epithe-ial cells) used at a concentration of 10 mg/ml; anti-D11b/c (Biosource International, mouse IgG2a, dis-
ribution 5 monocytes/macrophages, neutrophils) usedt a concentration of 5 mg/ml; anti-CD18 (identifiesFA-1 b chain; Seikagaku Corp., mouse IgG1, distri-ution 5 lymphocytes, monocytes, neutrophils) used atconcentration of 10 mg/ml.
Histological methods and renal tissue analysis.wo fragments of renal tissue were obtained. One wasmbedded in TissueTek II OTC compound (Lab Tekroducts, Miles Laboratories), frozen in a dry ice-ace-one mixture and stored at 270°C. The other fragmentas fixed in 15% formalin and embedded in Paraplastlus (Monoject, Sherwood Medical, Scientific Division,t. Louis, MO).Adhesion molecules and leukocytes were identified
n frozen 3-mm sections by indirect immunofluores-ence methodology using monoclonal antibodies, as de-cribed before (12), as secondary antibody we useduorescein-conjugated affinity pure F(ab9)2 rat anti-ouse IgG, with minimal cross-reaction with rat pro-
TABGener
Experimentalgroups n
Serum antiovalbumin(log2 titers)
7th day 25th day
Control 10 — —Immunized
-no-CSS 6 0.22 6 1.29 1.58 6 0.13-CSS 10 0.26 6 1.24 3.43 6 2.21
Note. Both immunized groups received similar immunization prignificant differences in proteinuria (P , 0.0001) and serum antiorotein excretion differences between no-CSS and control groups weroups were within normal limits and not significantly different.
eins (Accurate Chemical & Scientific Co.) at a concen- m
ration of 60 mg/ml (1:50). Each antibody was incubated5 min and then, three washes with PBS 0.01 M, 0.15
NaCl, pH 7.4 (3 min each), were accomplished. Nu-lei were stained with ethidium bromide. Tissues weretudied with a Zeiss Axioscope microscope equippedith epifluorescence system and phase contrast.ounts were done with the help of an eyepiece fittedith a grid. Cells with a green cytoplasmatic stain
urrounding an orange nucleus were considered posi-ive. In the glomeruli, results were expressed as posi-ive cells per glomerular cross-section (gcs), except forD54, which was evaluated in a score ranging from 11
o 41 based upon the intensity of fluorescence and theumber of positive cells as follows: grade 0 5 no fluo-escence in glomerular cells; grade 1 5 less than 20%ositive glomerular cells and low fluorescence inten-ity; grade 2 5 20–50% positive glomerular cells withoderated fluorescence brightness; grade 3 5 50–75%
ositive glomerular cells and moderated or intense flu-rescence brightness; grade 4 5 more than 75% ofositive glomerular cells and intense fluorescencerightness. In the interstitium results were expresseds positive cells per mm2.Identification of the nature of the cells expressing
he integrins and its quantitation were determinedith double-staining technique using the concentra-
ions described above as follows: monoclonal antibodiessed to identify the integrins (Mab anti-CD18 or anti-D11b/c) was applied to the tissue using the concen-
ration, incubation time, and washing described above,hereafter; rhodamine (TRIC)-conjugated affinity pure(ab9)2 rat anti-mouse IgG, with minimal cross-reac-ion with rat proteins (Accurate Chemical & Scientifico.) was added, then, tissues were incubated with theab to identify the leukocyte populations (Mab and-D45, Mab anti-CD43, or Mab anti-ED1) at the dilu-
ions described before, and at that time they wereeacted with 50 ml of fluorescein-conjugated affinityure F(ab9)2 rat anti-mouse IgG (20 mg/ml) with mini-
1Data
Urinary protein excretion(mg/24 h)
Glomerular IgG and C3deposits 6–7th weekBaseline 6th week
2.7 6 3.36 3.6 6 1.24 Negative
4.8 6 3.69 14.4 6 12.51 Traces/214.9 6 3.33 322.5 6 122.9 21/31
cols of daily intraperitoneal injections of ovalbumin. There werebumin titers (P , 0.05) between CSS and no-CSS groups. Urinarysignificant only at 6th week (P , 0.05). Baseline proteinuria in all
LEal
otovalre
al cross-reaction with rat proteins (Accurate Chemi-

cZtdppswCc
in
tFwwwtpbawc
r*
199ADHESION MOLECULES IN SERUM SICKNESS
al & Scientific Co.). Tissues were examined with aeiss Axioscope microscope equipped with phase con-rast and with the appropriate filters to identify rho-amine (TRIC) and fluorescein (FITC). In each field,ositive cells for each integrin and for each leukocyteopulation were counted. As control to test the doubletaining procedure, five tissues were double-stainedith CD43 and CD11b as described above. Positivity ofD11b was observed only in 45 6 6% of CD43-positive
ells (Figs. 1A and 1B).IgG and C3 deposits in renal tissues were identified
n frozen sections by indirect immunofluorescence tech-
FIG. 2. Expression of adhesion molecules and leukocyte infiltraespectively) in rats with chronic serum sickness (CSS, F), immunize*P vs control , 0.001.
ique. m
Tests of inhibition of adhesion of leukocytes to renalissues by monoclonal antibodies to CD54 and CD18.our experiments were done in duplicate. First, cellsere isolated from lymph nodes of healthy rats. Cellsere centrifuged for 10 min at 1200 rpm, washed twiceith RPMI 1640, and resuspended in RPMI 1640 con-
aining 10% fetal calf serum. Cells were then placed inlastic sterile 25-ml flasks (Corning, USA) and incu-ated overnight at 37°C with 5% CO2 to eliminatedherent cells. Next day, cells in the supernatant wereashed with RPMI 1640, resuspended to obtain a con-
entration of 107 cells/ml, and stimulated with phorbol
n in glomeruli (A and B, respectively) and interstitium (C and D,ats that did not develop proteinuria (no-CSS, E) and controls (C, h).
tiod r
yristate acetate (PMA, Sigma) 50 hg/ml for 30 min
(1tmttat1
fomstwibttmsAawnwa
hts
sdTpwsew
C
taowlTmtw
aI
200 BENATUIL ET AL.
13). Then cells were washed three times with RPMI640 at 1300 rpm for 10 min and afterwards werereated with a mixture of anti-CD54 and anti-CD18onoclonal antibodies (each diluted to a final concen-
ration of 40 mg/ml). After 45 min of incubation at 37°C,he preparation was washed with plain RPMI 1640 andcridine orange 14 mg/ml was added. After three addi-ional washes, the number of cells was adjusted to 2 307 cells/ml.Simultaneously, frozen 4-mm sections from four dif-
erent rats with proteinuria in which we had previ-usly shown overexpression of CD54 and CD18 in glo-erulus and interstitium, were selected, along with
ections from four control rats. These tissues werehawed and incubated for 15 min in Hank’s medium,ashed twice with PBS 0.01 M, pH 7.4, and incubated
n humid chamber with 50 ml of the monoclonal anti-ody mixture described previously at the same concen-ration but diluted in Hank’s medium. At the end ofhis incubation, tissues were washed twice with Hank’sedium, overlaid with 100 ml of the cellular suspen-
ion, and incubated in humid chamber for 1 h at 37°C.fterwards, six gentle washings with PBS were donend fluorescent cells adhered to renal cross-sectionsere counted. The following controls were used: (a)ormal rat kidney (n 5 4) and (b) kidneys from ratsith CSS (n 5 4) both treated with the nonpertinent
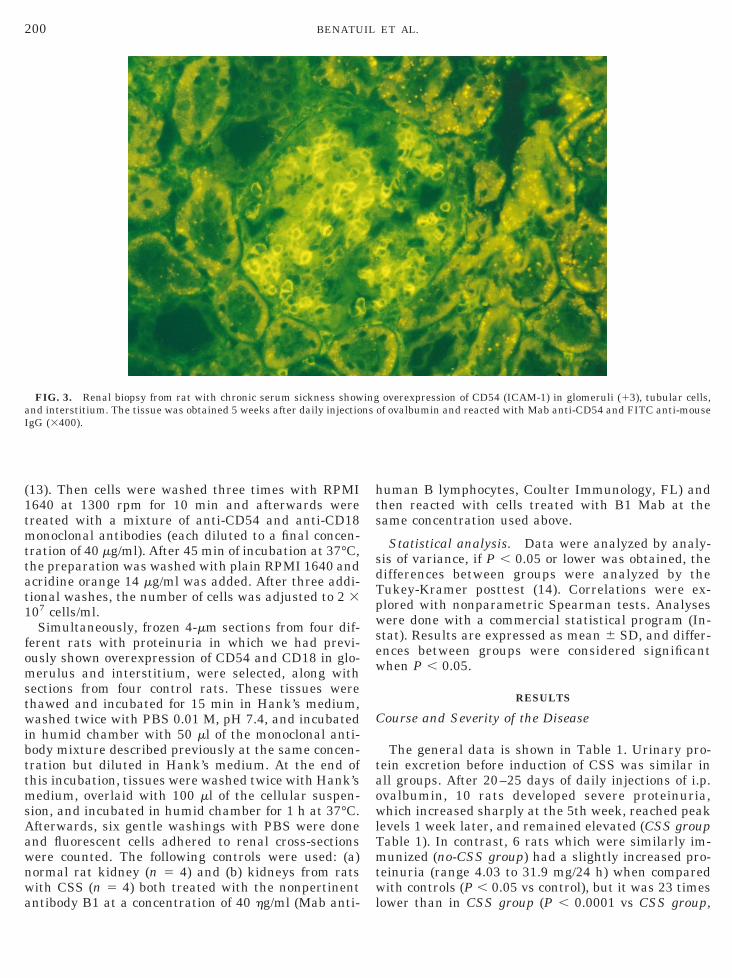
FIG. 3. Renal biopsy from rat with chronic serum sickness shownd interstitium. The tissue was obtained 5 weeks after daily injectiogG (3400).
ntibody B1 at a concentration of 40 hg/ml (Mab anti- l
uman B lymphocytes, Coulter Immunology, FL) andhen reacted with cells treated with B1 Mab at theame concentration used above.
Statistical analysis. Data were analyzed by analy-is of variance, if P , 0.05 or lower was obtained, theifferences between groups were analyzed by theukey-Kramer posttest (14). Correlations were ex-lored with nonparametric Spearman tests. Analysesere done with a commercial statistical program (In-
tat). Results are expressed as mean 6 SD, and differ-nces between groups were considered significanthen P , 0.05.
RESULTS
ourse and Severity of the Disease
The general data is shown in Table 1. Urinary pro-ein excretion before induction of CSS was similar inll groups. After 20–25 days of daily injections of i.p.valbumin, 10 rats developed severe proteinuria,hich increased sharply at the 5th week, reached peak
evels 1 week later, and remained elevated (CSS groupable 1). In contrast, 6 rats which were similarly im-unized (no-CSS group) had a slightly increased pro-
einuria (range 4.03 to 31.9 mg/24 h) when comparedith controls (P , 0.05 vs control), but it was 23 times
overexpression of CD54 (ICAM-1) in glomeruli (13), tubular cells,of ovalbumin and reacted with Mab anti-CD54 and FITC anti-mouse
ingns
ower than in CSS group (P , 0.0001 vs CSS group,

TbapgIT
E
C0nwcp
ggaCCpatc(
ww1sc2c0
ltpaIt
r16C8Ec
t
I
losTnwr
cpmettomdohoiwCe9i6ppaC
((
(
r
201ADHESION MOLECULES IN SERUM SICKNESS
able 1). In the CSS group, anti-ovalbumin IgG anti-ody levels increased 13 times, starting the first weeknd deposits of immune complexes (12 to 13) ap-eared in mesangial and subepithelial areas in thelomeruli. The proteinuria, serum antibody titers, andgG and C3 deposits in the no CSS-group are shown inable 1.
xpression of Adhesion Molecules and LeukocyteInfiltration
Glomerulus. As shown in Fig. 2A, the expression ofD54 was weak in normal rats (controls mean score.87 6 0.26) as well as in the no-CSS group of immu-ized rats (1.74 6 1.01). In animals with CSS, CD54as increased in mesangium and infiltrating leuko-
ytes (3.04 6 0.44) (Figs. 2A and 3) and correlated withroteinuria (P , 0.05, r 5 0.593).The number of CD18- and CD11b/c-positive cells in
lomeruli (cells/gcs) was comparable in the controlroup (CD18 5 1.82 6 1.11; CD11b/c 5 0.89 6 0.39)nd in the no-CSS group (CD18 5 2.22 6 0.28 andD11b/c 2.22 6 0.30, Fig. 2A). In contrast, rats withSS had increased numbers of CD18- and CD11b/c-ositive (Figs. 1A, 1C, and 2A), cells/gcs (13.5 6 5.47nd 13.74 6 5.47, respectively). Intraglomerular infil-ration of CD11b/c-positive cells (but not CD18-positiveells) correlated with the intensity of CD54 expressionr 5 0.510, P , 0.05).
The number of CD45-, CD43-, and ED1-positive cellsas similar in control and in no-CSS rats, while ratsith CSS showed values 6–35 times higher (Figs. 1B,D, and 2B). The same findings (not shown) were ob-erved for cells expressing la (controls 5 0.82 6 0.26ells/gcs; no-CSS group 5 1.16 6 0.80; CSS 5 6.88 6.43). Cellular infiltration of ED1-positive cells wasorrelated with glomerular CD54 expression (r 5.601, P , 0.05).
Interstitium. CD54 was over expressed in endothe-ial and tubular cells. As shown in Fig. 1c, there is a 2.5o 5 times increment in CD54-, CD18-, and CD11b/c-ositive cells in CSS. The increment in CD45-, ED1-,nd CD43-positive cells in CSS is evident in Fig. 2D.nterstitial CD54 expression was correlated with pro-einuria (r 5 0.592, P , 0.05).
As demonstrated by double-staining procedure (rep-esentative example of CD18 and CD45 shown in Figs.C and 1D), 86% 6 3.5 of the CD45-positive cells, 95%
1.82 of the ED1-positive cells, and 98% 6 2.6 of theD43-positive cells were also positive for CD18. Also,7% 6 6.5 of the LCA-positive cells, 75% 6 6.1 of theD1-positive cells, and 85% 6 7.2 of the CD43-positive
ells were simultaneously positive for CD11b/c.Evaluation was done in the glomeruli and tubuloin-
erstitial areas separately. i
nhibition of Leukocyte Adhesion to CSS Kidneywith Monoclonal Antibodies
Table 2 shows the results of blocking the adhesion ofymphocytes to the kidney of rats with CSS. Adherencef overlaid cells was two times higher in the renalections of rats with CSS than normal kidney slices.he use of a nonpertinent Mab (anti-human B cells) didot modify the adherence to CSS kidney. Pretreatmentith Mab anti-ICAM and anti-CD18 induced a 50%
eduction in lymphocyte adhesion in CSS kidney slices.
DISCUSSION
CSS is initiated by immune complex deposition butellular mechanisms come to play a critical role in theathogenesis of the disease (15, 16). Neutrophils,onocytes, and lymphocytes are known to infiltrate
arly the kidney in CSS and play an important role inhe development of proteinuria (10, 15, 17). In long-erm studies in CSS, Yang (10) demonstrated thatver-expression of ICAM-1 correlate with monocytes/acrophages accumulation and with immune complex
eposition. In the present study we have confirmed thisbservation, in fact, ICAM-1 (CD54) was two timesigher in rats with proteinuria than in rats with littler no proteinuria. In addition, we found that leukocytesnfiltrating the glomeruli and the interstitium of ratsith CSS also express CD18 (LFA-1 b chain) andD11b/c, which are ligands for CD54. In fact, CD18 isxpressed by 98% of the infiltrating lymphocytes and5% of the monocytes and neutrophils, while CD11b/cs expressed by 71% of monocytes and neutrophils and6% of lymphocytes. In concert with the increased ex-ression of CD54, CD18-positive cells and CD11b/c-ositive cells are increased 7–14 times in the glomerulind 2–5 times in the renal interstitium in the rats withSS. Interestingly, there is a correlation between the
TABLE 2Inhibition of Leukocyte Adhesion to the Kidney
in CSS by Monoclonal Antibodies
Number of adherentcells/mm2 in renal
cross section
A) Normal kidney no treated 24.5 6 9.76B) CSS kidney 1 cells treated with
B1 Maba90.9 6 11.20b
C) CSS kidney 1 cells treated withanti-CD54 1 anti-CD18 Mab**
42.2 6 21.28
a Renal tissues and cells were both treated with B1 Mab, whicheacts with human B lymphocytes.
b P , 0.01 vs A and C.
ntensity of CD54 expression (glomeruli and intersti-

tattc
flhakatmh1rdd
occpitmu
CDNa1
cC
1
1
1
1
1
1
1
1
1
1
R
202 BENATUIL ET AL.
ium) and the glomerular monocyte infiltration andlso with the severity of proteinuria. These associa-ions also suggest a role for the adhesion molecules inhe glomerular localization of monocytes and lympho-ytes.
In vitro adhesion experiments show that renal tissuerom rats with CSS (over-expressing ICAM-1) bind aarger number of mononuclear cells than tissues fromealthy rats. In addition, a mixture of Mab anti-CD54,nti-CD18, prevents the binding of leukocytes to theidney tissue, suggesting that there is likely in vivoctivity of these molecules. Clearly a definite proof ofheir in vivo relevance would require intravenous ad-inistration of these monoclonal antibodies, such asas been demonstrated in nephrotoxic nephritis (18,9); however, their cost and the long period of timeequired for the development of the disease (21–25ays) of daily intraperitoneal ovoalbumin injectionsiscourage this experiment in the CSS model.In summary, in chronic serum sickness, ICAM-1 is
verexpressed in glomerular cells along with LFA-1 bhain and CD11b/c in infiltrating cells. This incrementorrelates with the severity of the disease and is tem-orarily associated with the lymphocyte and monocytenfiltration in the kidney. The functional relevance ofhe expression of these adhesion molecules in the CSSodel is suggested by in vitro blocking experimentssing monoclonal antibodies.Parts of this work were presented in the XXXIIIongress of the European Renal Association/Europeanialysis Transplant Association, in Amsterdam, Theetherlands, June 18–21, 1996, and have appeared inbstract form (Nephrol. Dial. Transplant. 8, 524A,997).
ACKNOWLEDGMENTS
Financial support from Asociacion de Amigos del Rinon, Mara-aibo, Consejo de Desarrollo (CONDES), Universidad del Zulia andONICIT.
REFERENCES
1. Hunsicker, L. G., Shearer, T. P., Plattner, S. B., et al., The role ofmonocytes in serum sickness nephritis. J. Exp. Med. 150, 413–425, 1979.
2. Holdsworth, S. R., Nearle, T. J., Wilson, C. B., Abrogation ofmacrophage dependent injury in experimental glomerulonephri-
tis in rabbits. J. Clin. Invest. 68, 686–698, 1985.3. Nolasco, E. B., Cameron, J. S., Hartley, B., et al., Intraglomeru-lar T cells and monocytes in nephritis: Study with monoclonalantibodies. Kidney Int. 31, 1160–1166, 1987.
4. Parra, G., Platt, J. L., Falk, R., Rodrıguez-Iturbe, B., and Mi-chael, A. F., Cell populations and membrane attack complex inglomeruli of patients with post-streptococcal glomerulonephritis:Identification using monoclonal antibodies by indirect immuno-fluorescence. Clin. Immunol. Immunopathol. 33, 324–332, 1984.
5. Parra, G., Mosquera, J., and Rodrıguez-Iturbe, B., Migrationinhibition factor in acute serum sickness nephritis. Kidney Int.38, 1118–1124, 1990.
6. Springer, T. A., Adhesion receptors of the immune system. Na-ture 346, 425–434, 1990.
7. Briscoe, D. M., and Cotran, R. S., Role of leukocyte-endothelialcell adhesion molecules in renal inflammation: in vitro and invivo studies. Kidney Int. 44, (Suppl. 42), S27–S34, 1993.
8. Dal Canton, A., Adhesion molecules in renal disease. Kidney Int.48, 1687–1696, 1995.
9. Brady, H., Mark, D., Jimenez, W. et al., Chemoattractans pro-voke monocyte adhesion to human mesangial cells and mesan-gial cell injury. Kidney Int. 42, 480–487, 1992.
0. Yang, X., Glomerular sequence of events concerning monocyte/macrophage accumulation and ICAM-1 expression during exper-imental serum sickness nephritis in the rat. Pathol. Int. 47,238–249, 1997.
1. Sanchez-Crespo, M., Alonso, F., Barat, A., et al., Rat serumsickness: possible role of inflammatory mediators allowing dep-osition of immune complex in the glomerular basement mem-brane. Clin. Exp. Immunol. 49, 631–638, 1992.
2. Parra, G., Romero, M., Henriquez-La Roche, C., et al., Expres-sion of adhesion molecules in poststreptococcal glomerulonephri-tis. Nephrol. Dial. Transplant. 9, 1412–1417, 1992.
3. Dustin, M. L., and Springer, T. A., T cell receptor cross-linkingtransiently stimulates adhesiveness through LFA-1. Nature 341,619–624, 1989.
4. Godfrey, K., Statistical practice: Comparing the means of severalgroups. N. Engl. J. Med. 313, 1450–1456, 1985.
5. Ren, K., Van Liew, J., and Noble, B., The effect of cyclosporin A ondisease progression of proliferative immune complex glomerulo-nephritis. Clin. Immunol. Immunopathol. 166, 107–113, 1993.
6. Brady, H. R., Leukocyte adhesion molecules and kidney dis-eases. Kidney Int. 45, 1285–1300, 1994.
7. Ruiz-Ortega, M., Gonzalez, S., Seron, D., et al., ACE inhibitionreduces proteinuria, glomerular lesion and extracellular matrixproduction in a normotensive rat model of immune complexnephritis. Kidney Int. 48, 1778–1791, 1995.
8. Kawasaki, K., Yoita E, Yamamoto, T., et al., Antibodies againstintercellular adhesion molecule-1 and lymphocyte function-asso-ciated antigen-1 prevent crescentic glomerulonephritis. J. Im-munol. 150, 1074–1083, 1993.
9. Mulligan, M. S., Johnson, K. J., Todd, R. F., et al., Requirementsfor leukocyte adhesion molecules in nephrotoxic nephritis.
J. Clin. Invest. 91, 577–587, 1993.eceived July 20, 1998; accepted with revision October 7, 1998




















