
Expression of the amino acid dimorphism in the small myelin-associated glycoprotein cytoplasmic...
10
Ž . Molecular Brain Research 54 1998 252–261 Research report Expression of the amino acid dimorphism in the small myelin-associated glycoprotein cytoplasmic domain in rat peripheral nerves during postnatal development Petri Kursula a, ) , Veli-Pekka Lehto a , Bertrand Garbay b , Claude Cassagne b , Anthony M. Heape a a Biocenter Oulu and Department of Pathology, UniÕersity of Oulu, Kajaanintie 52 D, FIN-90220 Oulu, Finland b Laboratory of Membrane Biogenesis, C.N.R.S.-UniÕersity of Bordeaux II, Bordeaux, France Accepted 21 October 1997 Abstract Ž . The myelin-associated glycoprotein MAG is one of the proteins expressed during the period of myelin formation and is believed to play a major role in the initiation of myelination. It exists as two differentially expressed isoforms, L- and S-MAG, that are generated by alternative mRNA splicing. A nucleotide dimorphism at the mRNA level resulting in an ArgrPro dimorphism in the cytoplasmic tail of the S-MAG protein has previously been detected in the rat brain. In this study, we show that this dimorphism is detectable in the rat peripheral nervous system. We propose an allelic origin for the dimorphism and demonstrate the differential expression of the S-MAG alleles in the sciatic nerves of heterozygous rats during the period of active myelination. We also present data on the properties of the two S-MAG cytoplasmic domains produced as GST fusion proteins. The importance of this differentially expressed amino acid dimorphism is discussed, taking into account both its probable effect on the S-MAG cytoplasmic domain function and its significance in functional and structural studies concerning the S-MAG protein. q 1998 Elsevier Science B.V. Keywords: Myelin-associated glycoprotein; Myelination; Peripheral nervous system; Schwann cell; Development; Differential expression; Polymorphism; Phosphorylation 1. Introduction In rodent peripheral nerves, active myelination begins at birth, peaks at about 2 weeks, and slows to a low basal w x level by the age of 1 month 36 . The onset of myelination by Schwann cells in the peripheral nerves is signalled by the expression of a subset of proteins that are required for w x the formation of the myelin sheath 23 . The myelin-asso- Ž . ciated glycoprotein MAG is one of the proteins ex- pressed during this period and is believed to play a major w x role in the initiation of myelination 27,32 , although this w x role has recently been questioned 14,26 . MAG is a transmembrane protein that belongs to the immunoglobulin superfamily of cell adhesion molecules w x 3,22,28 . Two isoforms of MAG, L- and S-MAG, whose polypeptide chains differ only by their respective cytoplas- ) Corresponding author. Fax: q 358-8-330687; E-mail: [email protected] mic domains, are produced from the same gene by alterna- w x tive mRNA splicing of exon 12 22,34 . These two iso- Ž . forms are differentially expressed in both the central CNS Ž . and peripheral nervous systems PNS . In the rat CNS, L-MAG mRNA is predominantly expressed during the early active myelination period, while S-MAG mRNA predominates in the adult. In the PNS, on the other hand, S-MAG represents at least 95% of the total MAG through- w x out development 34 . Maximal expression of S-MAG mRNA in the peripheral nerves has been reported to occur w x approximately 2 weeks after birth, in both rats 31 and w x mice 17 . An amino acid dimorphism in the 9-amino acid, S- MAG-specific cytoplasmic sequence encoded by exon 12 w x of the mag gene, has been reported in the rat CNS 34 , but has since been totally ignored. A G-to-C nucleotide substitution at the mRNA level changes the amino acid 579 of S-MAG from a positively charged arginine residue to an uncharged, hydrophobic imino group, proline. This 0169-328Xr98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.
-
Upload
uni-hamburg -
Category
Documents
-
view
0 -
download
0
Transcript of Expression of the amino acid dimorphism in the small myelin-associated glycoprotein cytoplasmic...

Ž .Molecular Brain Research 54 1998 252–261
Research report
Expression of the amino acid dimorphism in the small myelin-associatedglycoprotein cytoplasmic domain in rat peripheral nerves during
postnatal development
Petri Kursula a,), Veli-Pekka Lehto a, Bertrand Garbay b, Claude Cassagne b,Anthony M. Heape a
a Biocenter Oulu and Department of Pathology, UniÕersity of Oulu, Kajaanintie 52 D, FIN-90220 Oulu, Finlandb Laboratory of Membrane Biogenesis, C.N.R.S.-UniÕersity of Bordeaux II, Bordeaux, France
Accepted 21 October 1997
Abstract
Ž .The myelin-associated glycoprotein MAG is one of the proteins expressed during the period of myelin formation and is believed toplay a major role in the initiation of myelination. It exists as two differentially expressed isoforms, L- and S-MAG, that are generated byalternative mRNA splicing. A nucleotide dimorphism at the mRNA level resulting in an ArgrPro dimorphism in the cytoplasmic tail ofthe S-MAG protein has previously been detected in the rat brain. In this study, we show that this dimorphism is detectable in the ratperipheral nervous system. We propose an allelic origin for the dimorphism and demonstrate the differential expression of the S-MAGalleles in the sciatic nerves of heterozygous rats during the period of active myelination. We also present data on the properties of the twoS-MAG cytoplasmic domains produced as GST fusion proteins. The importance of this differentially expressed amino acid dimorphism isdiscussed, taking into account both its probable effect on the S-MAG cytoplasmic domain function and its significance in functional andstructural studies concerning the S-MAG protein. q 1998 Elsevier Science B.V.
Keywords: Myelin-associated glycoprotein; Myelination; Peripheral nervous system; Schwann cell; Development; Differential expression; Polymorphism;Phosphorylation
1. Introduction
In rodent peripheral nerves, active myelination begins atbirth, peaks at about 2 weeks, and slows to a low basal
w xlevel by the age of 1 month 36 . The onset of myelinationby Schwann cells in the peripheral nerves is signalled bythe expression of a subset of proteins that are required for
w xthe formation of the myelin sheath 23 . The myelin-asso-Ž .ciated glycoprotein MAG is one of the proteins ex-
pressed during this period and is believed to play a majorw xrole in the initiation of myelination 27,32 , although this
w xrole has recently been questioned 14,26 .MAG is a transmembrane protein that belongs to the
immunoglobulin superfamily of cell adhesion moleculesw x3,22,28 . Two isoforms of MAG, L- and S-MAG, whosepolypeptide chains differ only by their respective cytoplas-
) Corresponding author. Fax: q 358-8-330687; E-mail:[email protected]
mic domains, are produced from the same gene by alterna-w xtive mRNA splicing of exon 12 22,34 . These two iso-
Ž .forms are differentially expressed in both the central CNSŽ .and peripheral nervous systems PNS . In the rat CNS,
L-MAG mRNA is predominantly expressed during theearly active myelination period, while S-MAG mRNApredominates in the adult. In the PNS, on the other hand,S-MAG represents at least 95% of the total MAG through-
w xout development 34 . Maximal expression of S-MAGmRNA in the peripheral nerves has been reported to occur
w xapproximately 2 weeks after birth, in both rats 31 andw xmice 17 .
An amino acid dimorphism in the 9-amino acid, S-MAG-specific cytoplasmic sequence encoded by exon 12
w xof the mag gene, has been reported in the rat CNS 34 ,but has since been totally ignored. A G-to-C nucleotidesubstitution at the mRNA level changes the amino acid579 of S-MAG from a positively charged arginine residueto an uncharged, hydrophobic imino group, proline. This
0169-328Xr98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0169-328X 97 00345-8

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261 253
substitution of the fourth carboxy-terminal amino acid mayaffect the intracellular ligand-binding properties of S-MAGand, hence, alter its biological functions.
Using a reverse transcription–polymerase chain reac-Žtion–restriction fragment length polymorphism RT–
.PCR–RFLP approach, we have investigated the existence,w x579frequency and expression of the ArgrPro dimorphism
during postnatal development in the sciatic nerves of indi-vidual rats, in order to determine the origin of the twoS-MAG mRNA subtypes. We also present data on thebiochemical properties of the two S-MAG cytoplasmicdomains.
2. Materials and methods
2.1. Animals
Two groups of Sprague–Dawley rats were used. In theexperiments using the pooled nerves from 5- to 15-day-oldrats, the animals were from the Institute of Cellular Bio-
Ž .chemistry and Genetics IBCG-CNRS, Bordeaux, France .In the developmental study, the rats were from the Univer-
Ž .sity of Oulu Experimental Animal Center Oulu, Finland .
2.2. Enzymes and reagents
Ž .Avian myeloid virus reverse transcriptase AMV RT ,5= AMV RT reaction buffer, SmaI restriction endonucle-ase, 10=Multicore buffer, T4 polynucleotide kinase, 10=kinase buffer, T4 DNA ligase and 10= ligase buffer
Ž .were from Promega Madison, WI, USA . BamHI andXhoI restriction endonucleases were from New England
Ž .Biolabs Beverly, MA, USA . Deoxynucleoside triphos-Ž .phates dNTPs , pGEX-4T-1 vector, glutathione sepharose
4B, thrombin and the T7Sequencing kit were from Pharma-Ž .cia Biotech Uppsala, Sweden . AmpliTaq thermostable
DNA polymerase, 10=PCR buffer II and MgCl were2Ž .from Perkin Elmer Foster City, CA, USA . The pCRII
cloning vector and the Wizard PCR Preps DNA purifica-Ž .tion kit were from Invitrogen San Diego, CA, USA .
w35 x Ž . w 32 x Ža- S -dATP 1000 Cirmmol , g- P -ATP 3000.Cirmmol and the first strand cDNA synthesis kit were
Ž .from Amersham Buckinghamshire, UK . Rat brain proteinŽ .kinase C was from Calbiochem La Jolla, CA, USA . All
other reagents and solvents were of analytical reagentgrade.
2.3. Gene-specific oligonucleotide primers
The oligonucleotide primers were synthesized on anApplied Biosystems DNA Synthesizer at the Biocenter
Ž .Oulu core facility Oulu, Finland . The rat MAG cDNA-w x Ž .specific 3,22,28 oligonucleotide primers see Fig. 1 were:Ž XMAG-1 5 -AAA CGC GGA TCC ACC CAG ACG AGA
X . Ž XAGA-3 , MAG-2 5 -TAC CGG ACC TCG AGT CAG
X. Ž XTGA CAA TCC-3 , MAG-4 5 -CCA GCT TCT CAGX. Ž XCGG GAG AC-3 and MAG-5 5 -CTC CTG GGG CTC
X. XTCA GTG AC-3 . The sense MAG-1 primer contains a 56-nucleotide overhang and a BamHI site, in addition to a15-nucleotide sequence corresponding to the last threeamino acids of the transmembrane domain and the firsttwo amino acids of the MAG cytoplasmic domainŽ . w xnucleotides 1725–1739 28 . The antisense MAG-2primer contains a 5X 8-nucleotide overhang and an XhoIsite, as well as 13 nucleotides complementary to the
ŽS-MAG-specific exon 12 sequence nucleotides 1862–. w x1874 28 . The sense MAG-4 primer is homologous to 20
Ž .nucleotides of the MAG cDNA nucleotides 1762–1781w x28 . The antisense MAG-5 primer, partly overlapping withMAG-2, contains the last 20 nucleotides of exon 12Ž . w xnucleotides 1867–1886 28 . Primers MAG-1 and MAG-2yield a 178-bp cDNA PCR product containing the entirecoding region for the S-MAG cytoplasmic domain, withflanking BamHI and XhoI sites.
The primers used for the control reactions give rise to aPCR product of 250 base pairs and were designed accord-ing to the rat glyceraldehyde-3-phosphate dehydrogenaseŽ . w x Ž XGAPDH mRNA sequence 13 : upstream GAPDH-1 5 -
X.GTC ATC CAT GAC AAC TTT GG-3 and downstreamŽ X X.GAPDH-2 5 -ATC CAC AAC GGA TAC ATT GG-3 .
2.4. Sciatic nerÕe dissection, RNA isolation and reÕersetranscription
Rats were anesthetized with CO and sacrificed by2
decapitation, or cervical dislocation. Sciatic nerves wererapidly dissected out from the vertebral column to theknee, immediately frozen in liquid nitrogen, and stored aty808C until needed.
Total RNA was isolated from the pooled sciatic nervesw xof 5- to 15-day-old rats according to Chirgwin et al. 4 .
For the developmental study, total RNA was preparedfrom the two sciatic nerves of individual rats, aged 5, 15,21 and 32 days and 8 months, as described by Chomczyn-
w xski and Sacchi 5 . An aliquot of each RNA preparationwas analysed for the presence of 28S and 18S rRNA by
w xformaldehyde agarose gel electrophoresis 25 .Two mg of RNA from the 5- to 15-day-old pooled
samples was reverse transcribed using random hexanucleo-tide primers with the first strand cDNA synthesis kit,according to the manufacturer’s protocol.
Sciatic nerve total RNA from individual animals wasŽ .reverse transcribed q428C, 60 min in a 30-ml reaction
mixture containing 100 ng of RNA, 5 U of AMV RT, 1mM of each dNTP, 1= AMV RT reaction buffer and 2.5
Ž .mM 75 pmol of MAG-5 and GAPDH-2 oligonucleotideŽprimers. The reaction was stopped by heating q958C, 5
.min and the samples were stored at y208C until PCR wasperformed. Control reactions without RNA template andwithout reverse transcriptase were performed to check forreagent and sample contamination.

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261254
Ž .Fig. 1. A Outline of the method for the detection of the nucleotide dimorphism in the rat S-MAG mRNA region encoded by exon 12 of the mag gene.The region corresponding to the S-MAG cytoplasmic domain in the cDNA is shaded. Locations of the regions of complementarity of the oligonucleotideprimers are shown below the cDNA with arrows. The box contains the two possible nucleotide and amino acid sequences of the rat S-MAG exon12-encoded region. The SmaI restriction site in exon 12 is shown in bold type. The translational stop codon of S-MAG is indicated by an asterisk. WhenRT–PCR is performed with the MAG-4 and MAG-5 primers, followed by restriction analysis with SmaI, a 127-bp product is detected from samplescontaining S-MAG-P579 mRNA, whereas the presence of the S-MAG-R579 mRNA results in products of 100 and 27 base pairs. The primers MAG-1 and
Ž .MAG-2 were used to clone the cDNA encoding the entire cytoplasmic domain of S-MAG for use in recombinant protein production. B The three distinctRFLP patterns obtained from the RT–PCR products from the total sciatic nerve RNA of individual rats are shown. Lane 1; the 127-bp SmaI-resistantcDNA product indicates the presence of only the S-MAG-P579 mRNA in the individual. Lane 2; the presence of the 127-bp and 100-bp cDNA productsshows that the animal expressed both of the S-MAG mRNAs. Lane 3; the 100-bp product resulting from the restriction cleavage by SmaI shows that theindividual expressed only S-MAG-R579 mRNA.
2.5. Polymerase chain reaction and restriction digestion
ŽAll reaction mixtures contained final total concentra-.tions in 20 ml : 5 ml of the reverse transcribed cDNA
sample, 500 mM of each dNTP, 2.8 mM of MgCl ,2
1=PCR buffer II, 1 U of AmpliTaq DNA polymerase andŽ .2.5 mM 50 pmol of MAG-4 and MAG-5 primers
Ž .GAPDH-1 and GAPDH-2 for the control reactions over-layed with mineral oil. The DNA polymerase was added
Ž .after heating q948C, 4 min and cooling the mixture onŽ .ice. The samples were then heated q948C, 5 min , andŽcDNA amplification was performed cycle: q928C, 1 min;
.q538C, 1 min; q728C, 1 min . A final extension stepŽ .q728C, 3 min was performed before cooling the samplesto 08C.
In preliminary tests using 1 mg of total RNA from thenerves of 8-month-old rats, the exponential phase of am-plification continued for at least 48 cycles for both the
Ž .S-MAG and GAPDH cDNA templates Fig. 2 . Thus, 42or 45 cycles were performed when the products were to bequantitated by densitometry.
For the RFLP analysis, 10 U of SmaI, 3 ml 10=Ž .Multicore buffer and H O to a final volume of 30 ml2
were added to the 20-ml RT–PCR mixture, and the diges-tion was performed for 16–18 h at q258C.
2.6. Agarose gel electrophoresis, densitometry and calcu-lation of the expression leÕels
Gel electrophoresis of the reaction products was per-Ž .formed on a 3% agarose gel poured and run 5 Vrcm in
Ž Ž .TBE buffer 89 mM tris hydroxymethyl aminomethaneŽ .Tris , 89 mM boric acid, 2 mM ethylene diamine te-
.traacetic acid, pH 8.3 , containing 1 mgrml ethidiumbromide. SmaI-treated samples and the correspondingGAPDH control samples were loaded into the same wells.The gel was photographed under ultraviolet light usingPolaroid 553 film. Densitometric analysis of the pho-tographs was performed using a Microtek MFS-300QSgrayscale scanner connected to an Apple Macintosh IIcicomputer, and quantitation was carried out with NIH Im-age 1.59 software. To allow the comparison of the relativenumber of DNA molecules present in each band, the bandintensities given by NIH Image were corrected as follows:
Ž .the GAPDH control band 250 bp intensity was divided579 Žby a factor of 2.50 and that of the S-MAG-P band 127
.bp by a factor of 1.27. The intensity of the 100-bp bandwas not corrected.
For each sample, the ratio of total S-MAGexpressionrGAPDH expression was calculated, and mean
Ž .values "standard deviation were determined for each

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261 255
Fig. 2. Determination of the duration of the exponential phase of PCRamplification. One-mg total RNA samples from the sciatic nerves of adultrats were amplified with MAG-4 and MAG-5 or GAPDH-1 and GAPDH-2primers and quantitated as described in Section 2. The band intensities ofthe ethidium bromide-stained products after agarose gel electrophoresisare shown. The intensities were corrected by assigning a value of 100 tothe intensity at cycle number 48. The exponential phase of amplification
Ž .continued for at least 48 cycles of PCR with both MAG open circlesŽ .and GAPDH filled circles primers.
age in the assay series. Two samples from 15-day-old ratsexpressing both S-MAG mRNAs were present in eachassay series in order to correct for differences betweendifferent series. The highest S-MAG expression being
Ž .observed at the age of 5 days see Section 3 , the meanS-MAGrGAPDH ratio from these animals was assigned avalue of 100, to which all other ratios were compared. Theexpression ratios of the individual S-MAG mRNA sub-typesrGAPDH were calculated in a similar fashion.
2.7. Polymerase chain reaction with 32P-end-labelledprimers
One nmol of the MAG-4 primer was end-labelled with32 P in a 20-ml reaction mixture containing 10 U T4polynucleotide kinase, 1=kinase buffer and 10 mCi g-w32 x Ž .P -ATP q378C, 1 h . PCR and SmaI digestion ofcDNA samples from animals expressing both S-MAGmRNA subtypes were carried out as above, except thatonly 30 cycles of amplification were performed.
The reaction products were resolved by non-denaturingŽ . Ž .10% polyacrylamide gel electrophoresis PAGE in 1=
TBE buffer. The gel was then sealed in plastic and auto-radiography was performed with Kodak X-ray film at
room temperature for 120 h. Densitometric analysis of thefilm was carried out as described above for the pho-tographs of agarose gels, except that no correction wasmade for product length and the GAPDH expression val-ues obtained for the samples from the non-radioactiveexperiments were used in the calculations.
2.8. Subcloning and identification of the PCR products
Non-digested PCR products obtained with the MAG-4and MAG-5 primers from the pooled sciatic nerve cDNAand from three individual 32-day-old rats exhibiting differ-
w xent S-MAG genotypes were purified 24 and subclonedinto the pCRII vector using T4 DNA ligase in 1= ligase
w xbuffer. Both strands of the insert were then sequenced 29 ,T7 w35 xusing the Sequencing kit and a- S -dATP.
( )2.9. Glutathione-S-transferase GST fusion protein pro-duction and analysis
The cDNAs encoding the cytoplasmic domains of S-MAG and the three last amino acids of the transmembranedomain were cloned from the pooled sciatic nerve cDNAof the 5- to 15-day-old rats by PCR using the MAG-1 andMAG-2 primers. The PCR products were purified withWizard PCR Preps kits, digested with BamHI and XhoI,and ligated into the pGEX-4T-1 expression vector. Thecorrect insertion of the cDNA was confirmed by sequenc-
w xing 29 .The vectors containing the S-MAG cDNA inserts were
used to transform Escherichia coli BL 21 cells. Small-scalew xGST fusion protein 30 production and purification were
performed according to instructions of the plasmid manu-facturer, using 0.2 mM isopropyl-b-D-thiogalactosideŽ . Ž .IPTG for the induction 1–16 h, q378C . Aliquots of thepurified fusion proteins were digested with thrombinŽ .q378C, 1 h , in order to cleave the GST from the S-MAGpolypeptides. The proteins were resolved by sodium dode-
Ž . w xcyl sulphate SDS –PAGE 21 and visualised by silverw xstaining 6 . The gels were dried under vacuum and photo-
graphed.
2.10. Phosphorylation in Õitro by protein kinase C
GST and both GST-S-MAG cytoplasmic domain fusionproteins were produced as described above, using 0.1 mMIPTG and an induction time of 1 h. For each reaction, 1mg of the purified protein immobilized on glutathionesepharose was phosphorylated, at q378C, in 50 mM Tris–HCl pH 7.5, 1.2 mM calcium acetate, 30 mgrml L-a-phosphatidyl-L-serine, 2.4 mgrml phorbol 12-myristate13-acetate, 3 mM dithiotreitol, 7.2 mM MgCl , 0.5 mCi2
w32 x Ž .g- P -ATP and 0.08 U protein kinase C PKC . Aliquotswere removed at timed intervals, and the reaction wasstopped by the addition of 10 mM ATP and sample buffer

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261256
Fig. 3. RFLP patterns obtained from two 5-day-old litters. Lanes 1–6; allanimals in the first litter expressed only S-MAG-R579. Lanes 7–11; allexcept one animal in the second litter expressed both of the S-MAGmRNAs.
w x Ž .21 and heating q988C, 5 min . Three independent reac-tions were carried out for each protein. Control reactionswere performed in the absence of PKC.
The proteins were separated by SDS–PAGE on 15%gels and visualized by Coomassie staining. Autoradiogra-phy was performed with Kodak X-ray film for 10 days aty808C, with intensifying screens. Densitometry of thegels and autoradiograms was carried out as describedabove for the PCR products. The intensity of the phospho-rylated fusion protein band on the X-ray film was cor-rected by two independent methods: by dividing it eitherby the intensity of the corresponding Coomassie-stainedprotein band, or by that of the autophosphorylated PKCband observed in autoradiography.
3. Results
3.1. Existence and frequency of the S-MAG mRNA nu-cleotide dimorphism in the rat peripheral nerÕous system
The G-to-C nucleotide substitution in the S-MAGmRNA sequence encoded by exon 12 of the rat mag genesuppresses a unique SmaI restriction site at the cDNAlevel, allowing the use of a simple RT–PCR–RFLP methodto study the presence and expression of the S-MAG dimor-
Ž .phism in rat sciatic nerves Fig. 1 . S-MAG cDNA wasamplified by RT–PCR from the total RNA isolated fromthe sciatic nerves of individual animals with the S-MAG-specific primers MAG-4 and MAG-5, and the productswere digested with SmaI.
After agarose gel electrophoresis, RT–PCR–RFLPproducts of 127 andror 100 bp were observed in all cases.
Fig. 4. Dideoxy sequencing results of plasmids containing either the579 Ž .RT–PCR product originating from the S-MAG-R left panel , or the
579 Ž .S-MAG-P right panel mRNA. Only the region surrounding thenucleotide dimorphism in exon 12 is shown. The position of the nu-cleotide dimorphism is shown by arrows. The premature stop for thesequencing reaction is only detectable in the sequence of the S-MAG-P579
RT–PCR product.
Analysis of the individual samples revealed that eachanimal could have one of three distinct RT–PCR–RFLPproduct patterns: only the 127-bp product, only the 100-bp
Ž .product, or both of the above Fig. 1B . Furthermore, allthree patterns, or only one pattern, could be found within a
Žsingle litter the 32-day-old and one 5-day-old litter, re-. Ž .spectively . Two litters of the same age 5 days also
Ž .showed different pattern distributions Fig. 3 .Among the 29 samples analysed, 11 individuals had
only the 100-bp product, 14 had both the 100- and the127-bp products, and 4 had only the 127-bp product. Table1 shows the frequency of each RFLP pattern in the differ-ent litters at each age studied, and demonstrates that thepresence of a given pattern is not age-dependent. The factthat the observed RFLP resulted from the expected nu-
w xcleotide dimorphism in exon 12 34 was confirmed bysubcloning and sequencing the RT–PCR products fromRNA samples from randomly chosen individuals and from
Ž .the pooled nerves of 5- to 15-day-old animals Fig. 4 .
3.2. DeÕelopmental expression of the S-MAG mRNA nu-cleotide dimorphism in the rat sciatic nerÕe
The total expression of S-MAG mRNA in the rat sciaticnerves relative to the amount of GAPDH expression was
Table 1S-MAG mRNA RT–PCR–RFLP patterns observed during postnatal development in individual litters
RT–PCR–RFLP product sizes 127 bp 100 and 127 bp 100 bp Total579 579 579 579Ž . Ž . Ž . Ž .S-MAG mRNA subtypes P P rR R
Litter ageŽ .Phenotype distribution number of individuals
5 days 0 0 6 65 days 0 4 1 515 days 0 5 1 621 days 3 3 0 632 days 1 2 3 6Total 4 14 11 29

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261 257
determined at different ages by RT–PCR–RFLP and den-Ž .sitometry Figs. 5 and 6 . The relative and total expression
Ž 579.levels of SmaI-sensitive S-MAG-R and SmaI-resistantŽ 579. 32mRNA S-MAG-P were also determined using a P-
Ž .end-labelled primer Fig. 7 .The expression of total S-MAG mRNA relative to the
expression of GAPDH, irrespective of the S-MAG sub-types expressed by the individuals, was highest at the age
Ž .of 5 days, and declined thereafter Fig. 6A . However, themean total S-MAG mRNA expression during developmentwas higher in young animals expressing the S-MAG-P579
579 Žsubtype than in those expressing only S-MAG-R Fig..6B . In the 21-day-old animals expressing only S-MAG-
P579 mRNA, total S-MAG mRNA expression was signifi-cantly higher than in the animals with both S-MAG mRNAsubtypes. These differences were no longer visible at theage of 32 days. The study included no animals under theage of 21 days expressing only the S-MAG-P579 mRNA.
In animals expressing both mRNA subtypes, the totalexpression of the S-MAG-P579 mRNA was three- to four-fold higher than that of S-MAG-R579 during the first 2
Ž .weeks Fig. 7A . Here again, the difference decreased withage and was no longer evident in the 1-month-old rats.Furthermore, S-MAG-P579 expression in the animals ex-pressing both S-MAG mRNAs was highest in the sciaticnerves of the 5-day-old animals, while the expression ofS-MAG-R579 appeared to peak at the age of 15 days. Theethidium bromide-staining and 32 P-end-labelling methods
Ž .gave very similar results Fig. 7B , demonstrating that thedevelopmental profiles were not dependent on the detec-tion and quantitation methods employed.
The absolute amounts of both S-MAG and GAPDHmRNAs increased throughout the period studied. However,the S-MAG mRNA levels increased dramatically alreadybefore postnatal day 15, while those of GAPDH only
Ž .increased rapidly after the age of 15 days not shown .
3.3. Production and analysis of recombinant fusion pro-teins
The cytoplasmic domains of both S-MAG subtypeswere expressed as GST fusion proteins using IPTG induc-
Fig. 5. Agarose gel analysis of the RT–PCR–RFLP products obtainedŽ . Žusing either MAG MAG-4 and MAG-5, upper panel , or GAPDH lower
.panel primers. Three representative individuals from each age group areshown. Lanes 1-3, 5-day-old animals; lanes 4–6, 15-day-old animals;lanes 7–9, 21-day-old animals; lanes 10–12, 32-day-old animals.
Fig. 6. Total expression of S-MAG mRNA in rat sciatic nerves duringpostnatal development. The intensities of the products obtained in theRT–PCR–RFLP analysis were corrected for size and the level of GAPDHmRNA as described in Section 2. In both graphs, mean values"standard
Ž .deviations are shown. A Total expression of S-MAG mRNA in the ratsciatic nerve observed when results from all animals, irrespective of their
Ž .individual RFLP patterns, were considered together. B Total expressionof S-MAG mRNA during postnatal development in the sciatic nerves ofrats exhibiting different RFLP patterns. Open circles represent animalsexpressing only S-MAG-P579; open triangles, animals only expressingS-MAG-R579; filled circles, animals expressing both mRNAs. Only oneor two samples were analysed for the following values: the 15-day valuefor animals expressing only S-MAG-R579 and the 32-day value foranimals expressing only S-MAG-P579, or both S-MAG subtypes.
tion times of 1–16 h. SDS–PAGE and silver staininganalysis of the purified fusion proteins yielded a protein
Ž .band of the expected size 32 kDa for both S-MAGŽ .subtypes Fig. 8 . However, as the induction time was
prolonged, the 32-kDa GST-S-MAG-P579 fusion proteinunderwent increasing degradation to yield a major degra-
Ž .dation product of about 31 kDa Fig. 8; lanes 2, 4 and 6and two or three minor bands. After a 16-h induction, most
579 Žof the GST-S-MAG-R had already been degraded not. 579shown , while the majority of the GST-S-MAG-R fu-
sion protein remained intact, regardless of the inductionŽ .time Fig. 8; lanes 1, 3 and 5 . Thrombin digestion and
( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261258
Ž .Fig. 7. S-MAG mRNA in rat sciatic nerves expressing both subtypes. AŽ 579Expression of the S-MAG mRNA subtypes S-MAG-P -open circles,
579 .S-MAG-R -filled circles in the sciatic nerves. The results were ob-tained by densitometry of X-ray films after RT–PCR–RFLP analysisusing 32 P-end-labelled MAG-4 primer. Mean values"standard deviationsare shown. The values for the age of 32 days were each obtained from the
Ž .analysis of a single animal. B The relative proportions of the S-MAGŽ 579 579 .mRNA subtypes S-MAG-P -circles, S-MAG-R -triangles in the sci-
atic nerve. The results were obtained by densitometry of either ethidiumŽ .bromide-stained agarose gels open symbols , or X-ray films after PCR
32 Ž .with the P-end-labelled MAG-4 primer filled symbols . Mean values"
standard deviations are shown.
Fig. 8. Analysis of GST-S-MAG fusion proteins by SDS–PAGE. The579 Ž .entire cytoplasmic domains of both S-MAG-R lanes 1, 3 and 5 and
579 Ž .S-MAG-P lanes 2, 4 and 6 were produced as GST fusion proteins inE. coli, purified by glutathione affinity chromatography and analysed bySDS–PAGE and silver staining. The induction with 0.2 mM IPTG was
Ž . Ž . Žcarried out for 1 h lanes 1 and 2 , 3 h lanes 3 and 4 , or 5 h lanes 5 and.6 . The locations of the molecular weight markers are indicated on the
right.
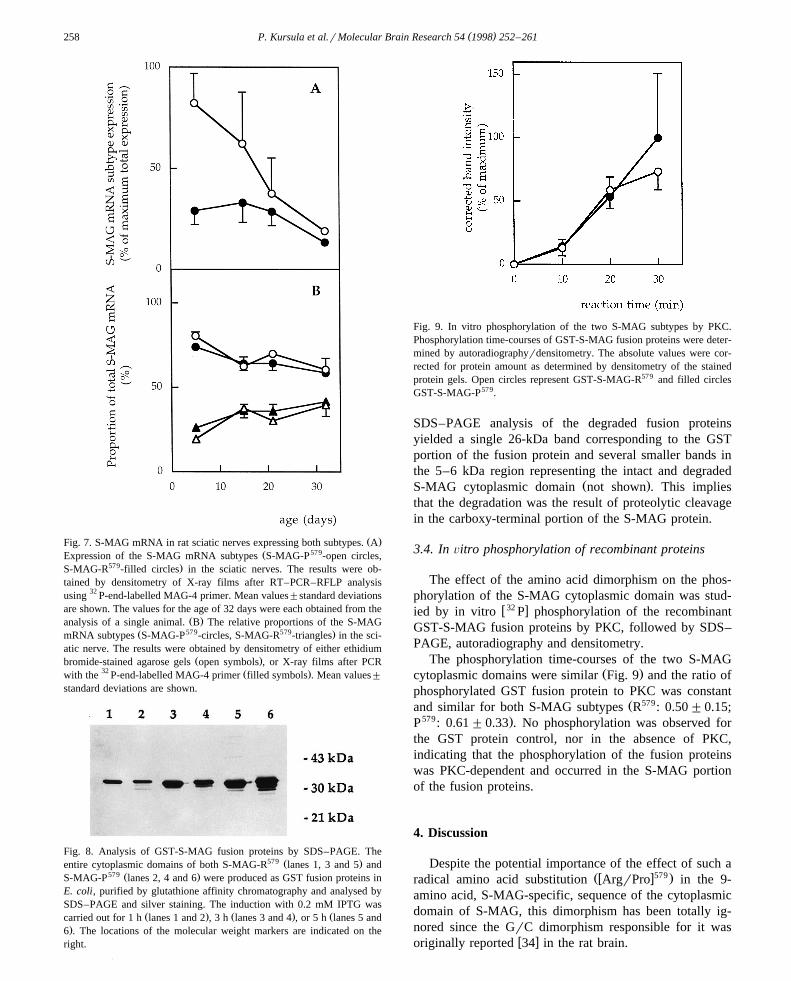
Fig. 9. In vitro phosphorylation of the two S-MAG subtypes by PKC.Phosphorylation time-courses of GST-S-MAG fusion proteins were deter-mined by autoradiographyrdensitometry. The absolute values were cor-rected for protein amount as determined by densitometry of the stainedprotein gels. Open circles represent GST-S-MAG-R579 and filled circlesGST-S-MAG-P579.
SDS–PAGE analysis of the degraded fusion proteinsyielded a single 26-kDa band corresponding to the GSTportion of the fusion protein and several smaller bands inthe 5–6 kDa region representing the intact and degraded
Ž .S-MAG cytoplasmic domain not shown . This impliesthat the degradation was the result of proteolytic cleavagein the carboxy-terminal portion of the S-MAG protein.
3.4. In Õitro phosphorylation of recombinant proteins
The effect of the amino acid dimorphism on the phos-phorylation of the S-MAG cytoplasmic domain was stud-
w32 xied by in vitro P phosphorylation of the recombinantGST-S-MAG fusion proteins by PKC, followed by SDS–PAGE, autoradiography and densitometry.
The phosphorylation time-courses of the two S-MAGŽ .cytoplasmic domains were similar Fig. 9 and the ratio of
phosphorylated GST fusion protein to PKC was constantŽ 579and similar for both S-MAG subtypes R : 0.50"0.15;
579 .P : 0.61"0.33 . No phosphorylation was observed forthe GST protein control, nor in the absence of PKC,indicating that the phosphorylation of the fusion proteinswas PKC-dependent and occurred in the S-MAG portionof the fusion proteins.
4. Discussion
Despite the potential importance of the effect of such aŽw x579.radical amino acid substitution ArgrPro in the 9-
amino acid, S-MAG-specific, sequence of the cytoplasmicdomain of S-MAG, this dimorphism has been totally ig-nored since the GrC dimorphism responsible for it was
w xoriginally reported 34 in the rat brain.

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261 259
In the present study, we demonstrate the presence of thesame dimorphism in the mRNA isolated from the periph-eral nerves of two completely independent populations ofSprague–Dawley rats. We also demonstrate that the pres-ence of one, or the other, or both of the S-MAG mRNAsubtypes in a given individual is not age-dependent. Fur-
Ž .thermore, in one litter 5-day-old , all individuals ex-Ž 579.pressed only one of the subtypes S-MAG-R , while in
another litter of the same age, most of the animals ex-pressed both subtypes and only one expressed only S-MAG-R579. Taken together with the fact that no post-tran-scriptional editing mechanisms are known that could ac-count for a G-to-C substitution at the mRNA level, our
w x579results demonstrate that the S-MAG ArgrPro dimor-phism in rat peripheral nerves is of allelic origin.
The expressed allele frequencies for the two subtypesare 0.62 for S-MAG-R579 and 0.38 for S-MAG-P579, asdetermined from the 29 individuals included in this study.These values are exactly as expected from the genotypefrequencies observed in this study, according to theHardy–Weinberg equilibrium, suggesting that neither ofthe alleles gives a selective advantage to the individual.
Total S-MAG mRNA expression was higher in younganimals homozygous for S-MAG-P579 than in hetero-zygotes, whose total expression was higher than that in
579 Ž .S-MAG-R homozygotes Fig. 6B . Furthermore, inyoung heterozygotes, the expression of the S-MAG-P579
allele was significantly higher than that of the S-MAG-R579
Ž .allele Fig. 7 . These results indicate that the GrC dimor-phism described here—or another polymorphism locatedelsewhere in the mag gene—may affect the regulatoryregions andror the transcription of the gene, or the pro-cessing, or stability of the mRNA. Evidence for the latterpossibility is provided by the results of the sequencinganalysis of the products. In all the samples analysed, theS-MAG-P579 sequences revealed a premature dideoxy re-action stop just before the dimorphic nucleotide Fig. 4thiswas never observed for the S-MAG-R579, indicating thatthe two messages form different secondary structures inthe region of the nucleotide dimorphism.
The proportion of S-MAG-P579 mRNA in heterozygousanimals decreases from about 80% in 5-day-old rats toapproximately 60% at 1 month. This indicates that, as theanimals mature, either the degradation of this mRNAincreases, or the expression of the S-MAG-P579 allele isdown-regulated more than that of the S-MAG-R579 allele.The clear, progressive developmental change of the pro-portion of the two mRNA species argues against thepossibility that the differences originate from differentefficiencies of the reverse transcription or PCR amplifica-tion with the two templates.
Furthermore, the absolute uncorrected amount of theS-MAG cDNA RT–PCR product was lowest at the age of5 days, while the S-MAG-P579rS-MAG-R579 ratio was atits highest at this age. The results obtained using 30 and 45cycles of PCR and two different detection methods are
Ž .indistinguishable Fig. 7B . These results argue stronglyagainst the possibility that heteroduplex formation duringthe late stages of PCR would cause an inefficiency in therestriction digestion. The unusually long period of expo-nential amplification observed most probably results fromthe sub-optimal reagent concentrations chosen for the PCRto prevent the amplification of nonspecific products.
The expression of total S-MAG mRNA in the rat sciaticnerves observed in this study was highest at the age of 5 to
w x15 days. Using northern blotting 31 and RNase protectionw xassays 34 , S-MAG mRNA expression in the rat sciatic
nerve was found to peak at postnatal days 13 and 6–10,respectively. In both of the above studies, samples frommany animals were pooled and analysed, while we studiedindividual animals and showed that the genotype of theanimal affects the level of S-MAG mRNA expression. Theexpression profile observed for S-MAG-P579 mRNA inheterozygous rats in this study is similar to that observed
w xfor S-MAG expression using RNase protection 34 , whilethe developmental expression of S-MAG-R579 mRNA inthese animals resembles that previously observed by north-
w xern blotting experiments 31 . Thus, ambiguous resultsconcerning the expression of S-MAG mRNA in the PNSmay result from the analysis of randomly chosen animalswith different S-MAG genotypes.
The production of both S-MAG cytoplasmic domains asGST fusion proteins revealed that the protein products ofthe two alleles have different properties. The S-MAGportion of the GST-S-MAG-P579 fusion protein was de-graded already during the early stages of induction, whereasthere was relatively little degradation of GST-S-MAG-R579.The major degradation product of GST-S-MAG-P579, thesize of which corresponds to a loss of approximately tenamino acids, was not seen in preparations of the GST-S-MAG-R579 protein. This suggests that the proline residuerenders the carboxy terminus of S-MAG unstable, whichmay result in the loss of the entire S-MAG-specific se-quence. Such differential stability of proteins produced by
w xdifferent alleles has been observed previously 12 . Thestrong upregulation of the S-MAG-P579 mRNA expressionduring early myelination in the sciatic nerve may be amechanism to compensate for the accelerated proteolyticdegradation or impaired function of important regions inthe corresponding protein product. That such a mechanismhas evolved and can be so clearly seen at a defined age,strongly suggests that the fully functional cytoplasmicdomain of S-MAG plays an important role during the earlystages of peripheral nerve myelination.
Interactions of cell adhesion molecules with intra-cellular proteins and the cytoskeleton have profound ef-
w xfects on their function 16 . Nervous system cell adhesionw xmolecules can bind to cytoskeletal proteins 7,8 and, upon
binding to extracellular ligands, the immunoglobulin su-perfamily cell adhesion molecules can regulate the distri-
w xbution of cytoskeletal structures within the cell 9 . MAGhas been shown to co-localize with spectrin and F-actin in

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261260
w xSchwann cells 32,33 , suggesting that it may interact withcytoskeletal proteins via its intracellular domain. In this
w x579case, the ArgrPro dimorphism of S-MAG might resultin different affinities of the two forms of S-MAG for thecytoskeletal proteins and, hence, in an alteration of thestability of the myelin sheath.
MAG also interacts with several proteins involved inw xintracellular signal transduction 18,19,35 , suggesting a
role for MAG in the mediation of the signals involved inmyelination. However, studies on the phosphorylation ofS-MAG have yielded somewhat controversial results. Earlystudies suggested that S-MAG is not phosphorylatedw x10,11 . Later, it was shown to be phosphorylated on serine
w x 579residues 1,2,19 . S-MAG-R has a potential PKC phos-phorylation site at serine 577, and this residue is most
w xlikely phosphorylated in vivo and in vitro by PKC 19 .Replacement of arginine-579 with a proline residue abol-ishes the typical consensus sequence for PKC-dependent
Ž w x.phosphorylation SrT-X-RrK 20 at this site. In thisstudy, we show that the phosphorylation of the S-MAGcytoplasmic domain by PKC in vitro is not affected by thedimorphism, indicating that either Ser-577 is not a majorphosphorylation target of PKC, or its phosphorylation doesnot require a basic amino acid residue at position q2.However, it is possible that the dimorphism affects S-MAGphosphorylation by PKC, or other kinases, in vivo.
A developmental RT–PCR–RFLP study of S-MAGcDNA from normal and Trembler mouse sciatic nerves,performed under conditions similar to those employed inthe present study, failed to reveal the existence of theG-to-C dimorphism; all animals only expressed the S-
579 Ž .MAG-R mRNA B. Garbay, unpublished observations .On the other hand, the sequence of the human S-MAG
Žcytoplasmic domain G.C. Miescher, R. Luetzelschwab, S.Huber, F. Ferracin, B. Erne and A.J. Stack, 1996, Gen-
.Bank accession number X98405 reveals that all exceptone of the nine amino acids are either identical, or highly
w xhomologous to the rat and mouse 15 sequences. Theexception concerns the dimorphic amino acid discussed inthe present study, which is a leucine residue in the humanprotein. The difference is due to the changing of the samedimorphic nucleotide to a thymidine.
Acknowledgements
This work was supported by the Finnish AcademyŽ . ŽFinland , the Maud Kuistila Memorial Foundation Fin-
. Ž .land , the Oulu University Central Hospital Finland andŽ .Biocenter Oulu Finland .
References
w x1 D.E.H. Afar, J.L. Salzer, J. Roder, P.E. Braun, J.C. Bell, Differentialphosphorylation of myelin-associated glycoprotein isoforms in cell
Ž .culture, J. Neurochem. 55 1990 1418–1426.
w x2 H.C. Agrawal, A.B. Noronha, D. Agrawal, R.H. Quarles, Themyelin-associated glycoprotein is phosphorylated in the peripheral
Ž .nervous system, Biochem. Biophys. Res. Commun. 169 1990953–958.
w x3 M. Arquint, J. Roder, L.-S. Chia, J. Down, D. Wilkinson, H. Baylet,P. Braun, R. Dunn, Molecular cloning and primary structure ofmyelin-associated glycoprotein, Proc. Natl. Acad. Sci. U.S.A. 84Ž .1987 600–604.
w x4 J.M. Chirgwin, A.E. Przybyla, R.J. McDonald, W.J. Rutter, Isolationof biologically active ribonucleic acid from sources enriched in
Ž .ribonucleases, Biochemistry 18 1979 5294–5299.w x5 P. Chomczynski, N. Sacchi, Single-step method of RNA isolation by
acid guanidium thiocyanate–phenol–chloroform extraction, Anal.Ž .Biochem. 162 1987 156–159.
w x6 C. Damerval, M. le Guilloux, J. Blaisonneau, D. de Vienne, Asimplification of Heukeshoven and Dernick’s silver staining of
Ž .proteins, Electrophoresis 8 1987 158–159.w x7 J.Q. Davis, V. Bennett, Ankyrin-binding activity shared by the
neurofascinrL1rNrCAM family of nervous system cell adhesionŽ .molecules, J. Biol. Chem. 269 1994 27163–27166.
w x8 J.Q. Davis, T. McLaughlin, V. Bennett, Ankyrin-binding proteinsrelated to nervous system cell adhesion molecules: candidates toprovide transmembrane and intercellular connections in adult brain,
Ž .J. Cell Biol. 121 1993 121–133.w x9 R.R. Dubreuil, G. MacVicar, S. Dissanayake, C. Liu, D. Homer, M.
Hortsch, Neuroglian-mediated cell adhesion induces assembly of theŽ .membrane skeleton at cell contact sites, J. Cell Biol. 133 1996
647–655.w x10 A.M. Edwards, M. Arquint, P.E. Braun, J.C. Roder, R.J. Dunn, T.
Pawson, J.C. Bell, Myelin-associated glycoprotein, a cell adhesionmolecule of oligodendrocytes, is phosphorylated in brain, Mol. Cell.
Ž .Biol. 8 1988 2655–2658.w x11 A.M. Edwards, P.E. Braun, J.C. Bell, Phosphorylation of myelin-as-
sociated glycoprotein in vivo and in vitro occurs only in the cyto-Ž .plasmic domain of the large isoform, J. Neurochem. 52 1989
317–320.w x12 M.B. Faletto, P. Linko, J.A. Goldstein, A single amino acid muta-
Ž .tion Ser180—Cys determines the polymorphism in cytochromeŽ .P450g P4502C13 by altering protein stability, J. Biol. Chem. 267
Ž .1992 2032–2037.w x13 P. Fort, L. Marty, M. Piechaczyk, S. el Sabrouty, C. Dani, P.
Jeanteur, J.M. Blanchard, Various rat tissues express only one majormRNA species from the glyceraldehyde-3-phosphate-dehydrogenase
Ž .multigenic family, Nucleic Acids Res. 13 1985 1431–1442.w x14 M. Fruttiger, D. Montag, M. Schachner, R. Martini, Crucial role for
the myelin-associated glycoprotein in the maintenance of axon-Ž .myelin integrity, Eur. J. Neurosci. 7 1995 511–515.
w x15 N. Fujita, S. Sata, T. Kurihara, R. Kuwano, K. Sakimura, T.Inuzuka, Y. Takahashi, T. Miyatake, cDNA cloning of mousemyelin-associated glycoprotein: a novel alternative splicing pattern,
Ž .Biochem. Biophys. Res. Commun. 165 1989 1162–1169.w x16 B.M. Gumbiner, Proteins associated with the cytoplasmic surface of
Ž .adhesion molecules, Neuron 11 1993 551–564.w x17 T. Inuzuka, N. Fujita, S. Sato, H. Baba, R. Nakano, H. Ishiguro, T.
Miyatake, Expression of the large myelin-associated glycoproteinisoform during the development in the mouse peripheral nervous
Ž .system, Brain Res. 562 1991 173–175.w x18 M.L. Jaramillo, D.E.H. Afar, G. Almazan, J.C. Bell, Identification
of tyrosine 620 as the major phosphorylation site of myelin-associ-ated glycoprotein and its implication in interacting with signaling
Ž .molecules, J. Biol. Chem. 269 1994 27240–27245.w x19 F. Kirchhoff, H.-W. Hofer, M. Schachner, Myelin-associated glyco-
protein is phosphorylated by protein kinase C, J. Neurosci. Res. 36Ž .1993 368–381.
w x20 A. Kishimoto, K. Nishiyama, H. Nakanishi, Y. Uratsuji, H. Nomura,Y. Takeyama, Y. Nishizuka, Studies on the phosphorylation ofmyelin basic protein by protein kinase C and adenosine 3X :5X-mono-

( )P. Kursula et al.rMolecular Brain Research 54 1998 252–261 261
Ž .phosphate-dependent protein kinase, J. Biol. Chem. 260 198512492–12499.
w x21 U.K. Laemmli, Cleavage of structural proteins during the assemblyŽ .of the head of bacteriophage T4, Nature 227 1970 680–685.
w x22 C. Lai, M.A. Brow, K.-A. Nave, A.B. Noronha, R.H. Quarles, F.E.Bloom, R.J. Milner, J.G. Sutcliffe, Two forms of 1B236rmyelin-as-
Ž .sociated glycoprotein MAG , a cell adhesion molecule for postnatalneural development, are produced by alternative splicing, Proc. Natl.
Ž .Acad. Sci. U.S.A. 84 1987 4337–4341.w x Ž .23 M.B. Lees, S.W. Brostoff, Proteins of myelin. In: P. Morell Ed. ,
Myelin, 2nd edn., Plenum, New York, 1983, pp. 197–224.w x24 Q. Li, C.L. Ownby, A rapid method for extraction of DNA from
Ž .agarose gels using a syringe, BioTechniques 15 1993 976–978.w x25 J. Meinkoth, G. Wahl, Hybridization of nucleic acids immobilized
Ž .on solid supports, Anal. Biochem. 138 1984 267–284.w x26 D. Montag, K.P. Giese, U. Bartsch, R. Martini, Y. Lang, H.
Bluthmann, J. Karthigasan, D.A. Kirschner, E.S. Wintergerst, K.-A.¨Nave, J. Zielasek, K.V. Toyka, H.-P. Lipp, M. Schachner, Micedeficient for the myelin-associated glycoprotein show subtle abnor-
Ž .malities in myelin, Neuron 13 1994 229–246.w x27 G.C. Owens, R.P. Bunge, Schwann cells infected with a recombi-
nant retrovirus expressing myelin-associated glycoprotein antisenseŽ .RNA do not form myelin, Neuron 7 1991 565–575.
w x28 J.L. Salzer, W.P. Holmes, D.R. Colman, The amino acid sequencesof the myelin-associated glycoproteins: homology to the immuno-
Ž .globulin superfamily, J. Cell Biol. 104 1987 957–965.
w x29 F. Sanger, S. Nicklen, A.R. Coulson, DNA sequencing with chain-Ž .terminating inhibitors, Proc. Natl. Acad. Sci. U.S.A. 74 1977
5463–5467.w x30 D.B. Smith, K.S. Johnson, Single-step purification of polypeptides
expressed in Escherichia coli as fusions with glutathione-S-trans-Ž .ferase, Gene 67 1988 31–40.
w x31 N. Stahl, J. Harry, B. Popko, Quantitative analysis of myelin proteingene expression during development in the rat sciatic nerve, Mol.
Ž .Brain Res. 8 1990 209–212.w x32 B.D. Trapp, Myelin-associated glycoprotein: location and potential
Ž .functions, Ann. NY Acad. Sci. 605 1990 29–43.w x33 B.D. Trapp, S.B. Andrews, A. Wong, M. O’Connell, W. Griffin,
Co-localization of the myelin-associated glycoprotein and the micro-filament components F-actin and spectrin in Schwann cells of
Ž .myelinated fibers, J. Neurocytol. 18 1989 47–60.w x34 M.B. Tropak, P.W. Johnson, R.J. Dunn, J.C. Roder, Differential
splicing of MAG transcripts during CNS and PNS development,Ž .Mol. Brain Res. 4 1988 143–155.
w x35 H. Umemori, S. Sato, T. Yagi, S. Aizawa, T. Yamamoto, Initialevents of myelination involve Fyn tyrosine kinase signalling, Nature
Ž .367 1994 572–576.w x36 H.deF. Webster, The geometry of peripheral myelin sheaths during
their formation and growth in rat sciatic nerves, J. Cell Biol. 48Ž .1971 348–367.




















