
Propagation of spreading depression inversely correlates with cortical myelin content
Upload
independentCategory
view
1download
0
~Copyright 1995 by Humana Press Inc. All rights of any nature whatsoever reserved. 1044-7393/95/2601--0053 $06.80
Expression of Myelin-Specific Proteins During Development of Normal
and Hypomyelinated Paralytic Tremor Mutant Rabbits
I. S tud ies on the Brain H o m o g e n a t e s
JOANNA SYPECKA* AND KRYSTYNA DOMAIqSKA-JANIK Department of Neurochemistry, Medical Research Centre,
Polish Academy of Sciences, 3 Dworkowa Str., 00-784, Warsaw, Poland
Received May 6, 1994; Accepted December 12, 1994
ABSTRACT
The paralytic tremor (pt) rabbit is an X-linked recessive mutant characterized by hypomyelination of the CNS. The onset of myelin mutants ' neurological symptoms typically occurs about the tenth postnatal day. A partial recovery is often observed; thus, the life-span of affected animals is almost normal and they can breed successfully. Mutants presenting this phenotype were chosen for our study. Because proteins can serve as excellent markers for the myelin formation pro- cess, we examined the developmental expression of several important myelin proteins (PLP/DM-20, MBP, CNP, MAG, and MOG) in both pt mutant and control rabbit brain homogenates.
Expression of the investigated proteins occurs in rabbits as fol- lows: CNP and MAG are already present at the early postnatal stage; PLP/DM-20 and MBP appear about the 10th postnatal day; MOG, ex- pressed last, has been detected on the 28th postnatal day. Whereas the MBP, CNP, MAG, and MOG content is only slightly reduced in mature pt mutant brain homogenates (80-90% of control values), the amount of PLP corresponds to approximately 30-40% of that present in controls. Expression of all of the examined proteins is substantially retarded in maturing brains, which leads to the conclusion that besides severe PLP deficiency, retardation of oligodendrocyte maturation is another probable feature of pt mutation.
*Author to whom all correspondence and reprint requests should be addressed.
Molecular and Chemical Neuropathology 53 Vol. 26, 1995

and D o m a n s k a - J a m k 54 S y p e c k a ' "
Index Entries: Hypomyelinated mutant; development; oligo- dendrocyte maturation; myelin markers; protein expression.
Abbreviations: Proteolipid protein (PLP); myelin basic protein (MBP); 2'3'-cyclic nucleotide 3'-phosphohydrolase protein (CNP); myelin associated glycoprotein (MAG); and myelin/oligodendrocyte glycoprotein (MOG).
I N T R O D U C T I O N
Myelin, a specialized extension of oligodendroglial (CNS) or Schwann cell (PNS) plasma membrane, plays an essential role in nerve function. Dis turbances in normally multilamellar and well-compacted myelin struc- ture lead to many serious neurological diseases (Quarles et al., 1989). Exploring the mechanisms of myelin formation, structure, and the func= tioning of its components might help to develop an appropriate therapy. Spontaneous myelin mutations have been reported in animals; some are excellent models to investigate demyelinating diseases in humans (Hogan and Greenfield, 1984). Today, numerous mutations are well characterized; their genetic backgrounds are understood, as well as their ways of affecting myelin. Among the existing myelin mutants, we can distinguish a group that inherits the mutation in an X-linked manner (for review, see Duncan, 1990). This group usually consists of very strongly affected animals. The severity of the neurological symptoms is rather surprising considering their cause--typically (except in jimpy mouse [Macklin et al., 1987]) a single amino acid substitution in a proteolipid protein (PLP) (Hudson, 1990).
Paralytic tremor (pt) rabbit is one of the X-linked recessive mutants (Osetowska and Leszowski, 1975), but differs somewhat from the others. A pathology of the myelin sheath, which is thin and uncompacted, has been observed and well described (Taraszewska, 1984; Taraszewska and Zelman, 1985, 1987). Altered myelin composition has been reported (Domanska- Janik et al., 1986, 1988, 1992). Biochemical studies have shown that mye- lination is deficient and delayed. In pt mutation, the first neurological symptoms appear about the 10th postnatal day; usually a coarse body tremor is visible. As the disease progresses, the animal may also develop exaggerated tendon reflexes, increased muscular tone, and spastic limb paresis. After a certain period of time, a partial recovery often occurs that makes the life-span of affected animals almost normal. The recovered animals can also breed successfully.
The interesting problem is whether the myelination is deficient because it is inhibited during the entire development of the animal, or whether it is merely delayed, eventually achieving a normal level that corresponds to the animal's recovery. To solve this problem, we performed a develop- mental study including the following periods: early postnatal (before myelination begins), active myelination, and the period when myelina-
Molecular and Chemical Neuropathology Vol. 26, 1995

Myelin Proteins in Homogenates of pt Mutant Rabbit 55
tion reaches a steady-state level. In our developmental study--to our knowledge, the first one concerning rabbits--we examined the content of five important myelin-specific proteins: proteolipid protein (PLP), myelin basic protein (MBP), 2'3'-cyclic nucleotide 3'-phosphohydrolase (CNP), myelin-associated glycoprotein (MAG), and myelin/oligodendrocyte gly- coprotein (MOG) in pt mutant and control rabbit brain homogenates. Pre- liminary results of this work have appeared in Acta Neurobiologiae Experi- mentalis (Sypecka et al., 1993).
MATERIALS AND METHODS
Chinchilla rabbits aged 1, 5, 10, 14, 18, 22, 28, 42, 56, 80, 100, and 120 d, were supplied by the Department of Comparative Neurology, Polish Academy of Sciences (Minsk Maz., Poland). Mutants (homozygous or hemizygous) were strictly controlled for both the pt trait and the pheno- type representing the typical mild course of the disease. Age-matched control rabbits were obtained from the same Chinchilla rabbit line.
Sacrifice of the animals was by decapitation. Each brain, minus cere- bellum, was divided into halves, immediately frozen in liquid nitrogen, and stored at -70~ until use. Half of each brain was then homogenized in 0.32M sucrose; 20% of the homogenate sample was spun for 15 min, 14 krpm; and the pellet was suspended in water with inhibitors (0.1 mM PMSF, 2 mm EDTA, 2 mM EDTA, 10 #g/mL antipain, 5/~g/mL leupeptin, and 5/~g/mL pepstatin). The remaining homogenate sample was reserved for myelin isolation.
Total protein content of the homogenates was determined by the method of Lowry (Lowry et al., 1951). Samples (100/~g) of proteins were separated on 12% polyacrylamide gel (Laemmli, 1970) and either stained with Coomassie blue or transferred to nitrocellulose membranes (Amer- sham, Beds., UK) using the electroblotting technique (Towbin et al., 1979). The major myelin-specific proteins were detected using the follow- ing antibodies:
1. Polyclonal anti-PLP/DM-20 and anti-MBP antibodies prepared in J.-M. Matthieu's laboratory were used after 1:400 dilution;
2. Monoclonal anti-MOG 8-18C5 was used at a final concentra- tion of 1-2 #g (Linington et al., 1984);
3. Monoclonal anti-MAG, diluted for the experiments 1:500, was a gift from R. H. Quarles (Quarles et al., 1981);
4. Monoclonal anti-CNPase was purchased from SIGMA Immuno- chemicals (St. Louis, MO) and diluted 1:200 before application.
Blots destined for PLP/DM-20 and MBP detection after incubation with the primary antibody were subsequently incubated with 12s-I labeled Protein A (NEN, Boston) and exposed to Kodak X-Omat films for several hours at -70~ The procedure included probing the blots first for PLP!
Molecular and Chemical Neuropathology Vol. 26, 1995

56 a n d D o m a n s k a - J a n & S y p e c k a ' "
DM-20 and, after developing autoradiograms, the same membranes were used for MBP detection. Blots with bound anti-MAG, anti-MOG, or anti- CNPase were incubated with sheep anti-mouse horseradish peroxidase- conjugated IgG (Amersham) and detected by ECL technique (Amersham). Blots were exposed to HyperfilmTM-ECL (Amersham) for 1-20 min. All of the immunoblots were subjected to a densitometric analysis (Pharmacia LKB UltroScan XL), and the results were expressed as a percentage of optical density estimated for a 100-d-old control rabbit.
CNPase enzymic activity was assayed as described elsewhere (Domafiska- Janik et al., 1988). The substrate and product, adenosine 2'3'-phosphate and adenosine 2'-phosphate, respectively, were separated on TLC and measured spectrophotometrically at 260 nm. A sample containing sub- strate but no enzyme served as a control. The specific activity of the enzyme was defined as/~mol of product per mg of protein, formed for 1 min at 37~
RESULTS
We examined 12 groups of animals aged 1-420 d. Each group con- sisted of two affected pt mutants and two age-matched controls. All of the mutants presented the same phenotype characteristics for a mild course of the disease. A clear developmental pattern for myelin-specific proteins PLP, MBP, and CNP could be noticed after standard Coomassie blue staining of electrophoreograms of the brain homogenate samples (Fig. 1). In the next experiments, the developmental profile of individual myelin- specific proteins was studied more precisely using immunoblotting and densitometric analysis techniques. We found CNP and MAG to be the first expressed myelin proteins during ontogenesis (Figs. 2, 3, and 4), which could be detected from the first days postpartum, and their con- tents successively increased until the 100th postnatal day. The amounts of CNP calculated from the scanned immunoblots were confirmed by parallel checking of its enzymic activity. Comparing Figs. 2 and 3 shows a high correlation between both methods, thus providing us with an addi- tional control for our Western blotting experiments. All experiments aimed at determining the content of a particular protein by immunoblotting were performed in duplicate at least three times for each protein. A high analogy of scanned immunoblots concerning the content of a given pro- tein was observed; therefore, the most representative immunoblots and their densitometric analysis were chosen for presentation (see figures).
The most abundant and, at the same time, the major myelin consti- tuents, PLP accompanied by its splicing variant DM-20, and MBP, appear comparatively later, about the 10th postnatal day (Figs. 5 and 6, respec- tively). MOG is expressed during ontogenesis as the last of the five inves- tigated proteins; we were able to detect MOG no earlier than the 28th postnatal day (Fig. 7).
Molecular and Chemical Neuropathology Vol. 26, 1995

Myelin Proteins in Homogenates of pt Mutant Rabbit
1 5 10 14 18 ~ t pt co pt co pt co pt co pt co
22 pt co
57
94.0 kDa 67.0 kDa
43.0 kDa
30.0 kDa
20.1 kDa
-PLP
-MBP
14.4 kDa
B 28 42 56 80 100 120
pt co pt co pt co pt co pt co pt co
-PLP
-MBP
Fig. 1. Brain homogenate samples from both pt mutant and control rabbits separated on 12% polyacrylamide gel, stained thereafter with Coomassie blue. A developmental pattern for several proteins was noted. (A) Animals aged 1--22 postnatal days. (B) Animals aged 28--120 postnatal days.
Molecular and Chemical Neuropathology Vol. 26, 1995

58
A
Sypecka and Domahska-Janik
1 5 10 14 18 22 28 4 2 56 80 tO0
B CNP 120
1 O0 c o n t r o l
80 %
6O
o ~ ~
120
1.0 5.0 10,0 14.0 15.0 22,0 28.0 42.( 56,0 50.0 100,0 120,0
A G E ( D A Y S )
Fig. 2. CNP expression in brain homogenates of pt mutant versus control rabbits. (A) The most representative immunoblot, obtained by ECL technique, revealing a slight reduction in CNP level in pt mutant. (B) Relative CNP amounts calculated from a densitometric analysis of the blot.
CNP 2o rapt 1~, con t ro l
1.0 5.0 10,0 1 4 . 0 15.0 22.0 28.0 42.0 56.0 80.0 100.0 120.0
AGE (DAYS)
Fig. 3. Assay of CNP enzymic activity in brain homogenates from both pt mutant and control rabbits. The values are means + SD of four estimations, *statistically significant differences at p < 0.01.
Molecular and Chemical Neuropathology Vol. 26, 1995

Myelin Proteins in Homogenates of pt Mutant Rabbit 59
A
B MAG
I 5 i0 14
120
18 22 B8 42 56 80 100 120
I pt [Z~ control
I O0
8o
40 I i
o _ . .
1.0 5.0 10.0 14.0 18.0 22.0 28.0 42.0 56.0 80.0 100,0 120.0
ACE ( days )
Fig. 4. Relative amounts of MAG in brain homogenates from pt mutant versus control rabbits. (A) The most representative immunoblot obtained by ECL technique. There were no significant differences between pt mutant and control animals. (B) A densitometric analysis of the blot.
A comparison of the developmental pattern of each investigated pro- tein in both pt mutants and controls indicates a reduction in all of the myelin-specific proteins in pt mutants. The degree of protein deficiency seems to be higher in young than in mature animals and, in the latter, cor- responds (except for PLP) to 80-90% of control values.
As for PLP, its strong deficiency was observed during the entire in- vestigated development , but was even more severe in young animals. Data obtained from densitometric analysis of the immunoblots (Fig. 5A,B) show a reduction to approximately 30-40% of the amount of PLP present in controls.
D I S C U S S I O N
The process of myelin formation and deposition begins at a strictly defined stage of development and consists of an array of events leading to wrapping axons in a highly specialized and unique multilamellar struc- ture. The developmental biology of myelin is usually studied either in
Molecular and Chemical Neuropathology Vol. 26, 1995

a n d D o m a n s k a . J a m k 60 S y p e c k a ' "
A
. . . . . . . . . . . . . . . . . . ~ . . . . . . . . . . . . . . ,~. ;,:~i: :
I 5 10 14 18 22 22 2,g 28 42 58 80 100 t20
B PLP 1zo pt
F723 ~onbrol 1150
60 ~ 7,
! 0
1,0 5,0 10,0 14-,0 1g, O 22,0 28,0 42.0 56,0 80,0 100,0 120.0
AGE ( days ) Fig. 5. Expression of both proteolipids, PLP (28 kDa) and DM-20 (20 kDa),
in brain homogenate samples from pt mutant versus control rabbits. (A) The most representative immunoblot after detection with usI Protein A. A remarkable deficiency of both proteins is observed in pt mutants. (B) Relative PLP amounts calculated from densitometric analysis of the blot.
vitro in nervous tissue culture or in vivo using rats or mice as model ani- mals. Consequently, myelinogenesis is best known in these two species (Norton, 1984), and little if any data are available on this process in other species--other than the valuable fact that it is not identical for all of them (Quarles et al., 1989).
We examined the sequence of expression of several myelin-specific proteins in rabbit brain. We found CNP to be the first detectable (by two independent methods) myelin protein. Detection of CNP enzymic activity is possible a couple of hours postnatally. This is in agreement with the established opinion that CNP is the earliest appearing myelinogenesis- associated protein (de Vellis, 1990). Although this protein exhibits enzymic activity, no candidate substrate for this enzyme has been discovered in myelin. A hypothesis for CNP function as a cytoskeletal protein has been discussed (Braun et al., 1990).
The second early appearing protein, MAG, is a glycoprotein belong- ing to an immunoglobulin superfamily (Williams and Barclay, 1988). Its localization in periaxonal membranes in CNS and PNS suggests its involve- ment in gl ia-neuron and glia--glia interactions (Brady and Quarles, 1988).
Molecu lar a n d C h e m i c a l N e u r o p a t h o l o g y Vol. 2 6 , 1 9 9 5

Myelin Proteins in Homogenates of pt Mutant Rabbit 61
A
...... . . . . . . . . . . . . . . . . . . . . . . . . ~ ............ , . . . . . . . . :~;: ~: ::::: . . . . . : ~ : ~ J ~ ~ , ~ ......
1 5 i0 14 18 ~ 33 38 38 43 66 BO 100 130
B MBP 120 lira pt F~ oon~ol
100
611
o 1 ~0 5,O ] 0,0 14~0 18.0 22.0 28 .0 4 2 . 0 -56,0 ~(}.O 100~0 ] 20~0
AGE ( days )
Fig. 6. Expression of MBPs in brain homogenates from pt mutant versus control rabbits. (A) The most representative immunoblot, which was first used for PLP/DM-20 detection (Fig. 5A) and thereafter incubated with anti-MBP in order to compare the expression of these proteins. 125I-labeled Protein A was used for detection in both experiments. (B) Relative MBP amounts calculated from densitometric analysis of the blot.
Moreover, MAG also expresses the L2/HNK-1 carbohydrate epitope that is claimed to be a marker for cell adhesion molecules (Schachner, 1989) and for the deposition of myelin lamellae.
The major myelin proteins, i.e., PLP accompanied by its splicing variant, DM-20, and MBP appear in the brain homogenates between the fifth and tenth postnatal days. We cannot state whether the expression of these proteins is parallel, but a possible interval could be no longer than 3-4 d. Some published results indicate that MBP appears before PLP (Norton, 1984). Even if their appearance in brain homogenate is parallel in time, there is a diverse localization of both events and different ways of protein transport before their incorporation into forming myelin (Morell et al., 1989). Although DM-20 has been reported to be expressed at the very early stage of development (Ikenaka et al., 1992; Timsit et al., 1992), immunodetect ion with a polyclonal antibody revealed the presence of both proteins at the same moment of development, i.e., on the tenth postnatal day.
Molecular and Chemical Neuropathology Vol. 26, 1995
62 S y p e c k a and D o m a h s k a - J a n i k
A ~ ..... ~ ~ '
~ 2B 42 ~B 80 I~O0 i~O
B MOG 12f1
'tO0
8O %
60
4O
20
13
I Pt
~ c o = t r o l
22 2~i 42 58 BO 100 120
ACE ( aays )
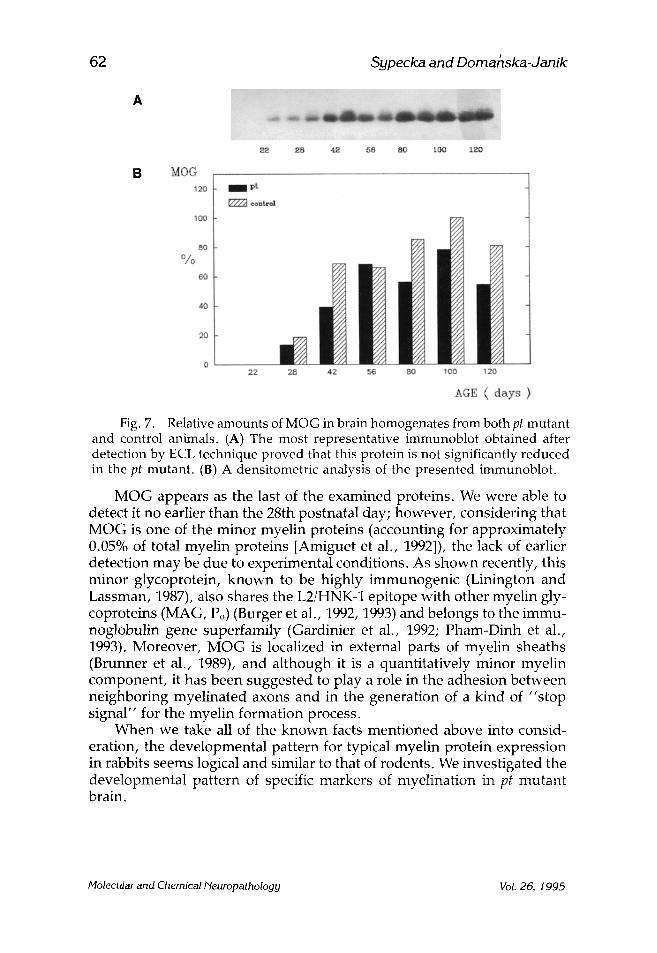
Fig. 7. Relative amounts of MOG in brain homogenates from both pt mutant and control animals. (A) The most representative immunoblot obtained after detection by ECL technique proved that this protein is not significantly reduced in the pt mutant. (13) A densitometric analysis of the presented immunoblot.
MOG appears as the last of the examined proteins. We were able to detect it no earlier than the 28th postnatal day; however, considering that MOG is one of the minor myelin proteins (accounting for approximately 0.05% of total myelin proteins [Amiguet et al., 1992]), the lack of earlier detection may be due to experimental conditions. As shown recently, this minor glycoprotein, known to be highly immunogenic (Linington and Lassman, 1987), also shares the L2/HNK-1 epitope with other myelin gly- coproteins (MAG, Po) (Burger et al., 1992, 1993) and belongs to the immu- noglobulin gene superfamily (Gardinier et al., 1992; Pham-Dinh et al., 1993). Moreover, MOG is localized in external parts of myelin sheaths (Brunner et al., 1989), and although it is a quantitatively minor myelin component, it has been suggested to play a role in the adhesion between neighboring myelinated axons and in the generation of a kind of "stop signal" for the myelin formation process.
When we take all of the known facts mentioned above into consid- eration, the developmental pattern for typical myelin protein expression in rabbits seems logical and similar to that of rodents. We investigated the developmental pattern of specific markers of myelination in pt mutant brain.
Molecular and Chemical Neuropathology Vol. 26, 1995

Myelin Proteins in Homogenates of pt Mutant Rabbit 63
A decreased protein content in all markers of myelination--particularly PLP and DM-20--was observed in pt rabbits during the entire investigated development. There is a consensus that this deficiency is more remarkable in young animals than in mature ones. Considering the strong PLP defici- ency attributed to the mutation localized on the X-chromosome, together with a widespread opinion about PLP as an established marker of mature oligodendrocytes (Norton, 1984), we concluded that oligo maturation must be significantly retarted in the pt mutant. Prolonged oligo maturation in pt mutants has already been observed on electronograms in morphological studies of brains of 4-wk-old pt rabbits (Taraszewska and Zelman, 1987).
The existance of myelin mutants such as the pt rabbit offers a great opportunity to investigate the mechanisms of myelinogenesis and the possible functions of particular myelin proteins. In the CNS, myelin is generated by oligodendrocytes that differentiate from progenitor cells and undergo the process called oligo maturation. During this process, oligos change their appearance (Norton, 1984).
The mature oligos express specific proteins that are incorporated into myelin and play different roles in myelin constitution and/or function. When oligos fail to mature at an appropriate stage of development, a retardation in myelination is observed, as in pt rabbits. Our results show that myelination in pt rabbit CNS is retarded and deficient during the entire development because there is an obvious deficit of myelin-specific proteins in pt mutant brain homogenates. This observed deficiency is more severe in young animals and is significantly compensated for at later stages of development, clearly indicating prolonged oligo matura- tion. Although, at the later stages of development, the content of myelin- specific proteins is only reduced to 80-90% of control values (except for PLP and DM-20), it never achieves a normal level.
In spite of an unanswered question concerning the reason for such prolonged oligo maturation, the present studies enable us to formulate a conclusion that the PLP hypoexpression caused by an X-linked mutation affects not only myelin formation and structure, but also (if not primarily) oligodendrocyte maturation. Recently, it has been suggested that PLP is not only the major structural protein but also plays a role as a trans- ducer for extracellular signals through the oligodendrocyte membrane (Popot et al., 1991). In our recent study, we observed an augmentation of calcium-signaling enzymes: calcium-activated neutral protease (CANP- ase) (Domar~ska-Janik et al., 1992) and protein kinase C (PKC) (unpub- lished data) in the mutant brain. A hypothetical link among enhanced calcium signals, PLP deficiency, and a retardation of oligo maturation in pt rabbits will be the subject of our further study. This extension of our developmental studies on purified myelin of pt rabbit brain gives a clearer insight into the nature of myelin pathology in pt mutation.
Molecular and Chemical Neuropathology Vol. 26, 1995

64 S y p e c k a a n d D o m a h s k a - J a n i k
ACKNOWLEDGMENTS
The authors wish to express special thanks to J.-M. Matthieu and M. Tosic for very helpful scientific and technical consultation. The authors are very grateful to R. H. Quarles for providing us with anti-MAG. The technical assistance of Halina Zajac and Teresa Czechmaflska is also very much appreciated. This work was supported by the Medical Research Centre, Polish Academy of Sciences, and the Swiss National Science Foundation, grant 70PP-031712.
REFERENCES
Amiguet P., Gardinier M. V., Zanetta J.-M., and Matthieu J.-M. (1992) Purifica- tion and partial structural and functional characterization of mouse myelin/ oligodendrocyte glycoprotein. J. Neurochem. 58, 1676-1682.
Brady R. O. and Quarles R. H. (1988) Developmental and pathophysiological aspects of the myelin associated glycoprotein. Ceil. MoL Neurobiol. 8, 139--148.
Braun P. E., Bambric L. L., Edwards A. M., and Bernier L. (1990) 2',3' cyclic nucleotide 3'-phosphodiesterase has characteristic of cytoskeletal proteins. A hypothesis for its function. Ann. N.Y. Acad. Sci. 605, 55-65.
Brunner C., Lassman H., Waehneldt T. V., Matthieu J.-M., and Linington C. (1989) Differential ultrastructural localization of myelin basic protein, myelin/ oligodendroglial glycoprotein, and 2',3'-cyclic nucleotide 3'-phosphodi- esterase in the CNS of adult rats. J. Neurochem. 52, 296--304.
Burger D., Perruisseau, G., Simon M., and Steck A. J. (1992) Comparison of the N-linked oligosaccharide structures of the two major human myelin glyco- proteins MAG and Po: Assessment and relative occurrence of oligosaccharide structure by serial lectin affinity chromatography of 14C-glycolipides. J. Neurochem. 58, 845-853.
Burger D., Steck A. J., Bernard C. C. A., Kerlero de Rosbo N. (1993) Human myelin/oligodendrocyte glycoprotein: a new member of the L2/HNH-1 family. J. Neurochem. 61, 1822-1827.
De Vellis J. (1990) Developmental and hormonal regulation of gene expression in oligodendrocytes. Ann. N Y Acad. Sci. 605, 81-89.
Domafiska-Janik K., Wikiel H., and Strosznajder J. (1986) Brain lipids of myelin deficient rabbit during development. Neurochem. Pathol. 4, 135-145.
Domafiska-Janik K., Gajkowska B., de Nechaud B., and Bourre J. M. (1988) Myelin composition and activities of CNPase and Na § K+-ATPase in hypomyelin- ated pt mutant rabbit. J. Neurochem. 50, 122-130.
Domafiska-Janik K., de Nechaud B., Inomata M., Kawashima S., and Zalewska T. (1992) Calcium-activated neutral protease (CANP) in normal and dys- myelinating paralytic tremor mutant rabbit myelin. Mol. Chem. Neuropathol. 16, 283-288.
Duncan I. D. (1990) Dissection of phenotype and genotype of the X-linked myelin mutants. Ann. New York Acad. Sci. 605, 110--121.
Molecular and Chemical Neuropathology Vol. 26, 1995

Myelin Proteins in Homogena te s o f pt Mutant Rabbit 65
Gardinier M. V., Amiguet P., Linington C., and Matthieu, J.-M. (1992) Myelin/ oligodendrocyte glycoprotein cDNA analysis reveals a unique member of the immunoglobulin gene superfamily. J. Neurosci. Res. 33, 177--187.
Hogan E. and Greenfield S. (1984) Animal models of genetic disorders of myelin, in Myelin (Morell P., ed.), pp. 489--534, Plenum Press, New York.
Hudson L. D. (1990) Molecular genetics of X-linked mutants. Ann. N.Y. Acad. Sci. 605, 155--165.
Ikenaka K., Kagawa T., and Mikoshiba K. (1992) Selective expression of DM-20, an alternatively spliced myelin proteolipid protein gene product, in devel- oping nervous system and in nonglial cells. ]. Neurochem. 58, 2248--2253.
Laemmli U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680--685.
Linington C. and Lassmann H. (1987) Antibody responses in chronic relapsing experimental allergic encephalomyelitis: correlation of serum demyelinating activity with antibody titre to the myelin/oligodendrocyte glycoprotein (MOG). ]. Neuroimmunol. 17, 61--69.
Linington C., Webb M., and Woodhams P. (1984) A novel myelin-associated glycoprotein defined by a mouse monoclonal antibody. J. Neuroimmunol. 6, 387-396.
Lowry O. H., Rosebrough N. J., Farr A. L., and Randall R. J. (1951) Protein measurement with Folin phenol reagent. J. Biol. Chem. 193, 265-275.
Macklin W. B., Gardinier M. V., King K. D., and Kampf K. (1987) An AG-AC transition at a splice site in the myelin proteolipid protein gene in jimpy mice results in the removal of an exon. FEBS Lett. 223, 417421.
Macklin W. B., Gardinier M. V., and Obeso Z. O. (1990) Structure and expression of the mouse myelin proteolipid protein gene. Ann. NYAcad. Sci. 605, 183--193.
Morell P., Quarles R. H., and Norton W. T. (1989) Formation, structure, and biochemistry of myelin, in Basic Neurochemist~: Molecular, Cellular and Medical Aspects, 4th ed. (Siegel G. J., Agranoff B. W., Albers R. W., and Molinoff P. B., eds.) pp. 109--136, Raven Press, New York.
Norton W. T. (1984) Oligodendroglia. Plenum, New York. Osetowska E. and Leszowski F. (1975) Introduction to investigation of hereditary
disease of the nervous system on pt rabbit model (in Polish). Neuropathol. Pol. 13, 61--70.
Popot J.-L., Pham-Dinh D., and Dautigny A. (1991) Major myelin proteolipid: the 4-topology. J. Membrane Biol. 120, 233--246.
Pham-Dinh D., Mattei M.-G., Nussbaum J.-L., Roussel G., Pantarotti P., Roeckel N., Mather I. H., Artzt K., Lindhal K. F., and Dautigny A. (1993) Myelin/ oligodendrocyte glycoprotein is a member of the immunoglobulin super- family encoded within the major histocompatibility complex. Proc. Natl. Acad. Sci. USA 90, 7990--7994.
Quarles R. H., Johnson D., Brady R. O., and Sternberger N. H. (1981) Prepara- tion and characterization of antisera to the myelin-associated glycoprotein. Neurochem. Res. 6, 115-q27.
Quarles R. H., Morell P., and McFarlin D. E. (1989) Disease involving myelin, in Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 4th ed. (Siegel
Molecular and Chemical Neuropathology Vol. 26, 1995

66 Sypecka and Domahska-Janik
G. J., Agranoff B. W., Albers R. W., and Molinoff P. B., eds.), pp. 697--713, Raven, New York.
Schachner M. (1989) Families of neural adhesion molecules, in Carbohydrate Recog- nition in Cellular Function. Ciba Foundation Symposium 145, pp. 156--168. Wiley, Chichester.
Sypecka J., Tosic M., Dolivo M., Doma•ska-Janik K., and Matthieu J.-M. (1993) Developmental expression of major myelin specific proteins in hypomyelin- ated pt mutant rabbit. Acta Neurobiol. Exp. 53, 281--284.
Taraszewska A. (1984) Pathology of myelin in pt rabbit (in Polish). Neuropatol. Pol. 22, 205--218.
Taraszewska A. and Zelman I. (1985) Morphometric studies on myelin develop- ment in pt rabbit (in Polish). Neuropathol. Pol. 24, 443--454.
Taraszewska A. and Zelman I. (1987) Electron microscopic study of glia in pt rabbits during myelination. Neuropathol. Pol. 25, 351--368.
Timsit S. G., Bally-Cuif L., Colman D. R., and Zalc B. (1992) DM-20 mRNA is expressed during the embryonic development of the nervous system of the mouse. J. Neurochem. 58, 1172--1175.
Towbin H., Staehelin T., and Gordon J. (1979) Electrophoretic transfer of pro- teins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. USA 76, 4350--4354.
Williams A. F. and Barclay A. N. (1988) The immunoglobulin superfamily- domains for cell surface recognition. Annu. Rev. Immunol. 6, 381-405.
Molecular and Chemical Neuropathology Vol. 26, 1995
Copyright © 2022 FDOKUMEN




















