
Aging, sexual dimorphism, and hemispheric asymmetry of the cerebral cortex: replicability of …
20
Neurobiology of Aging 25 (2004) 377–396 Aging, sexual dimorphism, and hemispheric asymmetry of the cerebral cortex: replicability of regional differences in volume Naftali Raz a,b,∗ , Faith Gunning-Dixon c , Denise Head d , Karen M. Rodrigue a , Adrienne Williamson e , James D. Acker f a Department of Psychology, Institute of Gerontology, Wayne State University, Detroit, MI, USA b Institute of Gerontology, 87 East Ferry St., 226 Knapp Building, Detroit, MI 48202, USA c Department of Psychiatry, University of Pennsylvania Medical Center, Philadelphia, PA, USA d Department of Psychology, Washington University, Saint-Louis, MO, USA e Department of Psychology, University of Memphis, Memphis, TN, USA f Baptist Memorial Hospital-East, Diagnostic Imaging Center, Memphis, TN, USA Received 31 July 2002; received in revised form 13 February 2003; accepted 7 April 2003 Abstract We examined age-, sex-, and hemisphere-related differences in the cerebral cortex. Volumes of the cerebral hemispheres and 13 regions of interest (ROIs) were measured on magnetic resonance images of 200 healthy adults. The strength of association between age and volume differed across ROIs. The lateral prefrontal cortex exhibited the greatest age-related differences, whereas significantly weaker associations were observed in the prefrontal white matter, sensory-motor, and visual association regions. The hippocampal shrinkage was significant in people in their mid-fifties. The primary visual, anterior cingulate, the inferior parietal cortices, and the parietal white matter showed no age-related differences. The pattern of age-related regional differences replicated the findings previously obtained on an independent sample drawn from the same population. Men evidenced larger volumes in all ROIs except the inferior parietal lobule, even after sexual dimorphism in body size was statistically controlled. In some regions (hippocampus and fusiform gyrus) men exhibited steeper negative age-related trends than women. Although a typical pattern of global hemispheric asymmetry was observed, the direction and magnitude of regional volumetric asymmetry was as inconsistent as in the previous reports. Thus, a pattern of age-related shrinkage suggesting increased vulnerability of the lateral prefrontal cortex to aging appears stable and replicable, whereas little consistency exists in sex-related and hemispheric differences in regional cortical volumes. © 2003 Elsevier Inc. All rights reserved. Keywords: MRI; Volumetry; Prefrontal; Hippocampus; White matter; Cingulate; Parietal; Motor; Parahippocampal gyrus 1. Introduction Human brain exhibits substantial individual diversity in size and morphology, and basic biological characteristics such as age and sex contribute a significant share of that variability. To date, quantitative neuroimaging investiga- tions have revealed moderate but consistent shrinkage of the cerebrum and expansion of the cerebro-spinal fluid (CSF) cavities across the adult life-span. The findings regarding the effects of age on separate compartments of brain tissue is less consistent. The preponderance of evidence from in vivo studies suggests that the negative association between age and brain volume is primarily attributable to shrinkage of the gray rather than white matter (for review see [58] ∗ Corresponding author. Fax: +1-508-256-5689. E-mail address: [email protected] (N. Raz). and more recent reports in [5,13,25,34,48]), although there is some evidence to the contrary [36,58]. A general trend in the in vivo volumetric studies of healthy volunteers points to the prefrontal cortex as the coritcal region in which the largest age-related differences are observed, whereas signif- icant but more moderate age effects are evident in posterior association cortices. The primary sensory regions seem to be largely spared by senescence [2,25,35,58]. The prefrontal white matter shows smaller age-related differences than the gray matter in the same region, although greater than those in the posterior white matter, whereas highly discrepant find- ings have been reported in the hippocampus, a region most frequently studied in the volumetric investigation of normal and pathological aging [58]. This general pattern of cortical aging is not observed in all studies and in some samples the volume of parietal lobes appear most strongly related to age [62]. In some cross-sectional studies, a curvilinear 0197-4580/$ – see front matter © 2003 Elsevier Inc. All rights reserved. doi:10.1016/S0197-4580(03)00118-0
-
Upload
ameriphoto -
Category
Documents
-
view
1 -
download
0
Transcript of Aging, sexual dimorphism, and hemispheric asymmetry of the cerebral cortex: replicability of …

Neurobiology of Aging 25 (2004) 377–396
Aging, sexual dimorphism, and hemispheric asymmetry of the cerebralcortex: replicability of regional differences in volume
Naftali Raza,b,∗, Faith Gunning-Dixonc, Denise Headd, Karen M. Rodriguea,Adrienne Williamsone, James D. Ackerf
a Department of Psychology, Institute of Gerontology, Wayne State University, Detroit, MI, USAb Institute of Gerontology, 87 East Ferry St., 226 Knapp Building, Detroit, MI 48202, USA
c Department of Psychiatry, University of Pennsylvania Medical Center, Philadelphia, PA, USAd Department of Psychology, Washington University, Saint-Louis, MO, USA
e Department of Psychology, University of Memphis, Memphis, TN, USAf Baptist Memorial Hospital-East, Diagnostic Imaging Center, Memphis, TN, USA
Received 31 July 2002; received in revised form 13 February 2003; accepted 7 April 2003
Abstract
We examined age-, sex-, and hemisphere-related differences in the cerebral cortex. Volumes of the cerebral hemispheres and 13 regionsof interest (ROIs) were measured on magnetic resonance images of 200 healthy adults. The strength of association between age and volumediffered across ROIs. The lateral prefrontal cortex exhibited the greatest age-related differences, whereas significantly weaker associationswere observed in the prefrontal white matter, sensory-motor, and visual association regions. The hippocampal shrinkage was significantin people in their mid-fifties. The primary visual, anterior cingulate, the inferior parietal cortices, and the parietal white matter showedno age-related differences. The pattern of age-related regional differences replicated the findings previously obtained on an independentsample drawn from the same population. Men evidenced larger volumes in all ROIs except the inferior parietal lobule, even after sexualdimorphism in body size was statistically controlled. In some regions (hippocampus and fusiform gyrus) men exhibited steeper negativeage-related trends than women. Although a typical pattern of global hemispheric asymmetry was observed, the direction and magnitude ofregional volumetric asymmetry was as inconsistent as in the previous reports. Thus, a pattern of age-related shrinkage suggesting increasedvulnerability of the lateral prefrontal cortex to aging appears stable and replicable, whereas little consistency exists in sex-related andhemispheric differences in regional cortical volumes.© 2003 Elsevier Inc. All rights reserved.
Keywords: MRI; Volumetry; Prefrontal; Hippocampus; White matter; Cingulate; Parietal; Motor; Parahippocampal gyrus
1. Introduction
Human brain exhibits substantial individual diversity insize and morphology, and basic biological characteristicssuch as age and sex contribute a significant share of thatvariability. To date, quantitative neuroimaging investiga-tions have revealed moderate but consistent shrinkage of thecerebrum and expansion of the cerebro-spinal fluid (CSF)cavities across the adult life-span. The findings regardingthe effects of age on separate compartments of brain tissueis less consistent. The preponderance of evidence from invivo studies suggests that the negative association betweenage and brain volume is primarily attributable to shrinkageof the gray rather than white matter (for review see[58]
∗ Corresponding author. Fax:+1-508-256-5689.E-mail address: [email protected] (N. Raz).
and more recent reports in[5,13,25,34,48]), although thereis some evidence to the contrary[36,58]. A general trend inthe in vivo volumetric studies of healthy volunteers pointsto the prefrontal cortex as the coritcal region in which thelargest age-related differences are observed, whereas signif-icant but more moderate age effects are evident in posteriorassociation cortices. The primary sensory regions seem tobe largely spared by senescence[2,25,35,58]. The prefrontalwhite matter shows smaller age-related differences than thegray matter in the same region, although greater than thosein the posterior white matter, whereas highly discrepant find-ings have been reported in the hippocampus, a region mostfrequently studied in the volumetric investigation of normaland pathological aging[58]. This general pattern of corticalaging is not observed in all studies and in some samplesthe volume of parietal lobes appear most strongly relatedto age[62]. In some cross-sectional studies, a curvilinear
0197-4580/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/S0197-4580(03)00118-0

378 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
relationship between age and white matter volume has beenobserved, suggesting a possibility that the volume of myeli-nated fibers increases up to the middle age, then levels offand declines in later years[2,13].
Although sex differences are not evident on visual in-spection of brain magnetic resonance imaging (MRI), thesupport for sexual dimorphism in the total cerebral volumeis quite consistent. According to the majority of studies,men possess larger cerebra than women of the same age andhealth status, even if the body size differences are controlledstatistically [4,6,12,13,28,42,47,52]. A rough estimate ofthe magnitude of sex differences reported in those studiesfalls between 1 and 1.5 standard deviations. There is littleagreement, however, on the topography of sex differencesin regional volumes[19,21,46,52,60]. A recent study ofyoung adults suggested that regional sex differences mayfollow a pattern of developmental distribution of estrogenreceptors[24]. There is a distinct possibility that age- andsex-related variations interact in shaping the trajectories ofbrain aging, with men exhibiting steeper age-related de-clines [12,16,41,42,55,83]. However, in some samples, theopposite pattern (i.e. increased age differences in femalebrains) has been observed[46]. Even when sex differencesin brain aging are reported, there is little agreement on theregional distribution of sex-specific age trends.
Understanding of age and sex differences in brain volumesis compounded by the possibility that right and left hemi-spheres may be differentially influenced by aging in men andwomen. In right-handed individuals, the right hemisphere isusually larger than the left[83], and brains display a patternof a counter-clockwise torque, i.e. right hemisphere protrud-ing on the front, and the left hemisphere extending on theback[23,26,28,60,81,84]. Several cortical regions (most no-tably, planum temporale[69]) exhibit asymmetry, and whilethere is no clear evidence of right or left hemi-aging of thebrain (see[56] for a review), sex differences in volumetricasymmetry have been observed[1,21,26].
Inconsistencies in the extant literature on age-, sex-, andhemisphere-related differences in regional brain volumesmay stem from significant heterogeneity of the populationsfrom which study samples are drawn and by the differencesin methods of acquisition and measurement of the MR im-ages. Hence, it is important to establish the convergenceand replicability of the findings across multiple methods andsubject populations, and obtain stable estimates of the ef-fects of age, sex, and hemispheric location on the regionalcortical volumes. To attain these objectives, we estimatedthe volumes of specified cortical regions using in vivo MRIacquired in a large sample of healthy adults drawn from apopulation in which a pattern of differential cortical agingwas previously observed[60]. Although the two samples([60] and the current one) were drawn from the same popu-lation using the same recruiting methods and scanned on thesame scanner, the images were imported from the scannerby different means, preprocessed through different software,presented (and traced) on different monitors with a differ-
ent contrast and sensitivity, and traced by different (althoughpartially overlapping) groups of operators. Thus, the sam-ples could not be combined, and we used a common metricof effect size and correlation coefficients to compare the re-sults of the two studies and to examine across-sample sta-bility of the age-, sex-, and hemisphere-related differences.
2. Subjects and methods
2.1. Subjects
The data for this study were collected in an ongoinginvestigation of neuroanatomical correlates of age-relateddifferences in cognition conducted in a major metropolitanarea in the USA. Subjects were recruited by advertisingin local media and on an urban university campus. Theysigned a consent form approved both by the UniversityCommittee for Protection of Human Subjects in Researchand by the Hospital’s Patients Participation Committee. Allparticipants were screened with a mail-in 66-item healthquestionnaire augmented by telephone and personal in-terviews. Persons who reported history of cardiovascular,neurological or psychiatric conditions, head trauma withloss of consciousness for more than five minutes, thyroidproblems, diabetes, treatment for drug and alcohol abuse ora habit of taking more than three alcoholic drinks per daywere excluded from the study, as were persons who reportedusing anti-seizure medication, anxiolytics, antidepressants,or recreational drugs. Persons who suffered or suspectedthey could suffer from claustrophobia were advised againstparticipation in the study.
A total of 1220 persons responded to advertising, andhealth questionnaires were sent to all of them. Out of theresponders, 288 have never returned the questionnaires.Of the 932 who returned the questionnaires, 505 (54%)were rejected on the basis of the health screening. Themost frequent single reasons for rejection were: high scoreon depression questionnaire (10%), history of neurolog-ical problems (multiple sclerosis, history of meningitis,encephalitis, stroke, brain surgery, head trauma with lossof consciousness—9%), taking medication that can affectcognitive performance (7%), hypo- or hyperthyroid (7%),left-handedness (5%), cardiovascular problems (cardiacby-pass surgery, valve problems, history of heart attack—5%), diabetes mellitus (4%), history of psychiatric hospital-izations (4%), sensory handicaps (blindness, severe hearingimpairment—4%), cancer (3%), insufficient education andhistory of learning disabilities (2%), counter-indication forMRI (e.g. metal implants—2%), alcohol and drug abuse(1%). The remainder (193 persons, 39%) were rejected fora combination of the listed reasons. In addition, 134 (13%)could not be reached for scheduling and 32 (3%) had termi-nated the study prematurely. Thus, 259 persons participatedin the study, and full sets of valid MRI data were available on201 of them—less than 17% of the initial pool of responders.

N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 379
One person was removed from the data set because a sig-nificant though asymptomatic infarct in the putamen wasdiscovered after MRI examination. Nineteen participants (7men and 12 women, mean age 58.20±12.20 years) who re-ported having hypertension that was successfully controlledby medication were also included in the sample. None of thesubjects participated in the previously published study ofcortical neuroanatomy of aging conducted in our laboratory[60], although they were drawn from the same population.
All subjects were screened for dementia and depressionusing a modified Blessed Information-Memory-Concentra-tion Test[7] with a cut-off of 30 and Geriatric DepressionQuestionnaire[57] with a cut-off of 15. An experiencedneuroradiologist (J.D.A.) examined the MR scans for signsof space occupying lesions and signs of significant cere-brovascular disease. The final sample consisted of 200 par-ticipants: 119 women (47.28±16.13 years old) and 81 men(46.41± 17.06 years old). The age distribution was approx-imately rectangular, and there were no differences in agebetween the sexes (t < 1). Average education in this samplewas relatively high (15.76± 2.45 years) and correspondedto almost 4 years of college. Men and women did not differin educational attainment (t = 1.26, ns), although older agewas weakly associated with fewer years of formal school-ing (r = −0.22,P < 0.01). All participants were consistentright-handers, as indicated by a score above 75% on the Ed-inburgh Handedness Questionnaire[49].
2.2. MRI protocol
All imaging was performed on a 1.5T Signa scanner (Gen-eral Electric Co., Milwaukee, WI). Volumes were measuredon the images acquired usingT1-weighted 3-D spoiled gra-dient recalled (SPGR) sequence with 124 contiguous ax-ial slices, echo time(TE) = 5 ms, repetition time(TR) =24 ms, field of view(FOV) = 22 cm, acquisition matrix256× 192, slice thickness= 1.3 mm, and flip angle= 30◦.The imaging site was fully staffed by medical physicistsand MRI technologists, and the scanners were routinely cal-ibrated using a standard GE phantom.
2.3. Volumetric image analysis
2.3.1. MR image processing and alignmentAfter acquisition, the images were imported to an Ap-
ple Macintosh G4 (Apple Computer Corp. Cupertino, CA)where they were reformatted to correct for undesirable ef-fects of head tilt (to the left or right shoulder), pitch (for-ward or backward), and rotation (to the right or to the left)using BrainImage 2.3.3, public domain software[61]. There-alignment process consisted of the following steps illus-trated inFig. 1. First, to correct brain position for head pitch,the axial plane was tilted so it passed through the anterior andposterior commissures (incorporating the AC–PC line), asillustrated inFig. 1B. Because an infinite number of planes
could pass though the AC–PC line, an additional landmarkwas necessary to define the axial plane. Therefore, in the nextstep (depicted inFig. 1C), the axial plane was fixed interac-tively by forcing it to pass through the orbits in such a waythat the axial cross-section of the orbits on the right and theleft side of the head was of equal diameter. After correctingfor head tilt, the sagittal plane was moved to pass along thestraight line drawn through the extreme anterior and poste-rior points of the interhemispheric fissure (IHF) as illustratedin Fig. 1D. Finally, the coronal plane was drawn perpendicu-lar to the axial and sagittal planes as defined. The equivalenceof right and left orbital diameters was ascertained through-out the brain volume by inspection of all relevant coronalslices (Fig. 1E). In a few cases, a complete correction of therotation by insuring orbit equivalence was not easy, and theoperator had to check back and forth to maintain the IHF asthe middle anchor of the volume. When such inconsistencyarose, alignment along the IHF (Fig. 1D) took precedenceover the coronal orbital alignment (Fig. 1E). Reformattedimages were partitioned into contiguous 1.5 mm coronal sec-tions. Because of pixel asymmetry, the reformatted imagesappeared shortened along the vertical axis. However, thisdistortion did not affect reliability of volume estimates.
The ROI areas were measured with NIH Image pub-lic domain software (Version 1.60, available at the WorldWide Webhttp://rsb.info.nih.gov/nih-image/). Images weredisplayed on a 21′′ monitor and magnified 4×. Each ROIwas traced manually using a puck and a digitizing tablet.To average the tracing errors across operators and to avoidthe association between a specific operator and a ROI, theslices were divided into two equal groups at random, andeach half-sample was traced by a different operator. Volumeswere computed using the Cavalieri formula[63]. The mea-surements were performed in cm3 using calibration functionof the NIH Image. The initial calibration was performed onimages obtained by scanning a standard GE phantom.
2.4. Reliability of ROI volumes
Reliability of the regional volumetric measures was as-sessed by an intraclass correlation formula that presumesrandom selection of raters, ICC(2)[66]. This is a compre-hensive and conservative index of reliability that not onlytakes into account, like Pearsonr, the order of the mea-sured entities and their values in each set, but is also, liket statistic, sensitive to the differences between the two setsof measures. Thus, to attain high reliability, the operatorsneeded to achieve very good agreement not only on the rel-ative size of the traced structures but also on the absolutesize, for systematic “overtracing” or “undertracing” of a ROIby one operator would result in low ICC. To further pre-vent any operator bias, each set of slices for a given ROIwas split into two equal subsets at random, and each opera-tor traced one subset before the measures were combined tocompute the ROI volume. In applying the rules of region de-marcation, we resolved all questionable cases by consulting

380 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
Fig. 1. The process of brain alignment illustrated step-by-step on actual images as they appear on the computer monitor screen. (A) The initial positionof the brain. Note the head rotation to the right in the axial plane and head pitched up in the sagittal plane. (B) The image is rotated to be re-positionedalong the AC–PC line. Note the anterior and posterior commissures visible on the axial image. (C) The head tilt is adjusted by rotating the image toequalize the orbital diameters in the axial plane. (D) The rightward rotation of the head is adjusted by positioning it along a straight line passing throughthe interhemispheric fissure. (E) The rotation is checked by insuring the equality of orbital diameters in the coronal plane.
N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 381
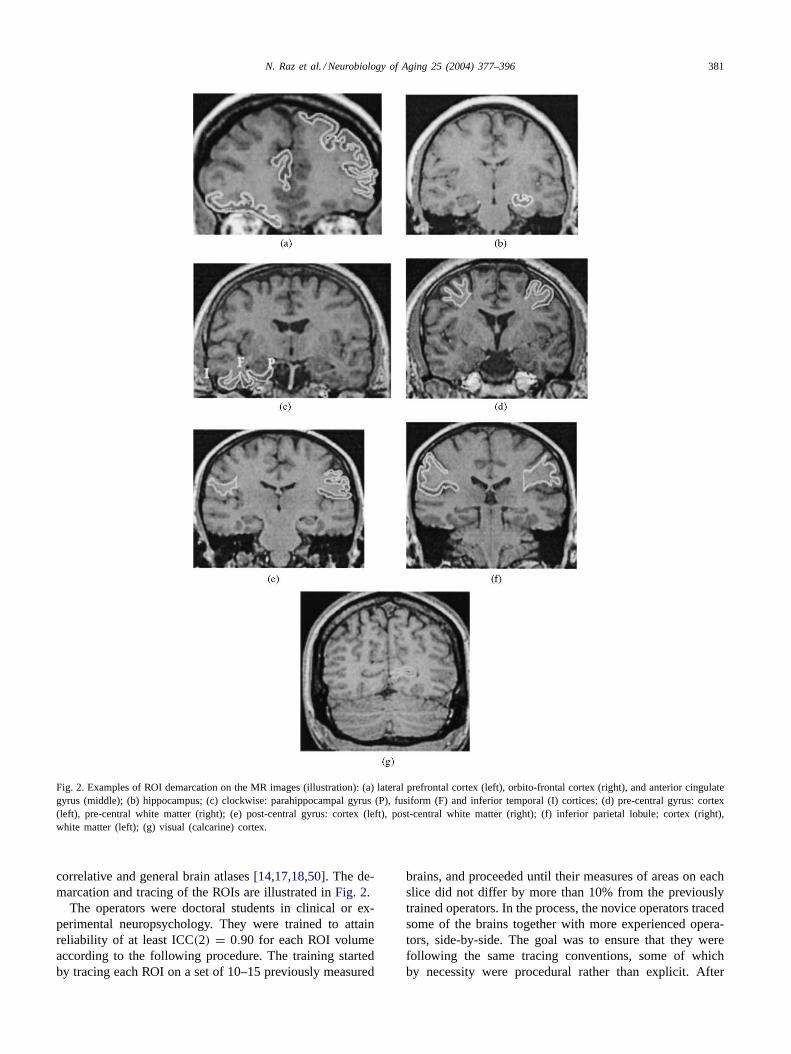
Fig. 2. Examples of ROI demarcation on the MR images (illustration): (a) lateral prefrontal cortex (left), orbito-frontal cortex (right), and anterior cingulategyrus (middle); (b) hippocampus; (c) clockwise: parahippocampal gyrus (P), fusiform (F) and inferior temporal (I) cortices; (d) pre-central gyrus: cortex(left), pre-central white matter (right); (e) post-central gyrus: cortex (left), post-central white matter (right); (f) inferior parietal lobule; cortex (right),white matter (left); (g) visual (calcarine) cortex.
correlative and general brain atlases[14,17,18,50]. The de-marcation and tracing of the ROIs are illustrated inFig. 2.
The operators were doctoral students in clinical or ex-perimental neuropsychology. They were trained to attainreliability of at least ICC(2) = 0.90 for each ROI volumeaccording to the following procedure. The training startedby tracing each ROI on a set of 10–15 previously measured
brains, and proceeded until their measures of areas on eachslice did not differ by more than 10% from the previouslytrained operators. In the process, the novice operators tracedsome of the brains together with more experienced opera-tors, side-by-side. The goal was to ensure that they werefollowing the same tracing conventions, some of whichby necessity were procedural rather than explicit. After

382 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
reaching±10% consistency on the training set, the oper-ators selected a new set of 15 brains, which were evenlydistributed between the young, middle-aged, and older sub-jects. This set was used to test reliability, and if the desiredminimal level of reliability (ICC= 0.90) was not reachedon the selected set at the first attempt, the set was treated asa training set and the whole procedure was repeated untilsuccessful completion. The final reliability estimate wasalways obtained on a newly selected set of brains.
2.5. Region demarcation and tracing rules
2.5.1. Cerebral hemispheresEach cerebral hemisphere was traced separately on coro-
nal slices. Each of the first three slices and the last threeslices was traced to evaluate frontal and occipital petalia;every fourth slice in between was traced, thus creating a4.5-mm inter-slice gap. The traced regions included bothgray and white matter, but excluded the ventricles, the cere-bellum, and the pons.
2.5.2. Prefrontal cortex (PFC) the anterior cingulate gyrus(ACG), and the prefrontal white matter
Eight to 12 coronal slices located within 40% of the dis-tance between the genu of the corpus callosum and thefrontal pole were included in the prefrontal ROI. The lateralprefrontal cortex (LPFC) ROI included superior, middle, andinferior frontal gyri and covered mainly Brodmann areas 9and 46, and parts of areas 8, 10 and 45. The superior andinferior boundaries of the LPFC were the most dorsomedialpoint of the cortex and the lateral orbital sulcus, respectively.
Volume of the orbito-frontal cortex (OFC) was estimatedfrom the same slices, using the most lateral branch of theorbital sulcus as the lateral boundary and the olfactory sulcusas the medial boundary. The OFC included almost the wholearea 47 and a substantial part of area[11].
The anterior cingulate gyrus (ACG) was measured on thecaudal 40% of the slices within the PFC range, i.e. the caudal16% of total frontal slices. The upper and lower boundarieswere defined by the superior and inferior cingulate sulci.After each sulcus had been identified, a horizontal line wasdrawn from the opening of this sulcus through the cortex toits deepest point. The band of cortex between these horizon-tal lines (one at the superior and one at the inferior cingu-late sulci) defined the ACG. The ROI that covered part ofBrodmann areas 24 and 33, consisted of 4–6 slices.
In the prefrontal and other cortical ROIs, only the con-tinuous cortical ribbon was measured; gray matter was ex-cluded if it was completely enclosed by white matter. Thewhite matter adjacent to the prefrontal ROI (FWHITE) wastraced as a ROI complementary to the respective gray matterregions.
Pre-central Gyrus (Motor Cortex, MC) was located be-tween the anterior commissure (AC) and the slice lyingimmediately anterior to the posterior commissure (PC).Between the AC and the mammillary bodies, the inferior
boundary of this ROI was the lateral fissure and the upperboundary was the superior pre-central sulcus. Between thefirst slice through the mammillary bodies and the posteriorboundary, the central sulcus served as the inferior bound-ary, whereas the superior boundary was located dorsally tothe superior pre-central sulcus. Thus, the MC ROI coveredmost of area 4 and a small part of dorsal area 6.
Post-central Gyrus (Somatosensory Cortex, SSC) wasmeasured on the anterior 75% of the slices between thefirst slice that included the mammillary bodies and the PC.Its inferior boundary was the lateral fissure and its superiorboundary was the central sulcus. This ROI covered areas 1,2, and 3, although it is possible that in some brains it alsoincluded small parts of areas 5 and 43.
Inferior Parietal Lobule (IPL) was measured on the coro-nal slices located between the PC and the last slice on whichthe splenium could be observed. The inferior boundary ofthis ROI was the lateral fissure and its superior boundarywas the intraparietal sulcus. Care was taken not to includethe superior temporal gyrus which appears on the rostralslices within the defined boundaries. The described ROIincluded a portion of area 40.
Parietal White Matter (PWHITE). Thewhite matter adja-cent to the parietal (post-central and somatosensory) ROIswas traced as a ROI complementary to the respective graymatter regions.
Parahippocampal Gyrus (PHG) included the entorhinalcortex and the ambient gyrus but not the subiculum. In therostral-caudal direction, this ROI ranged from the limen in-sulae to the most posterior slice on which the pulvinar wasstill visible. The superior border was a horizontal line drawnfrom the medial point of the PHG white matter to the cor-tex. The inferior border of the PHG was a horizontal linedrawn from the most interior point of the collateral sulcusto the white matter. The PHG, as defined above, containedthe entorhinal and perirhinal cortices as well as a part of theuncus.
Inferior Temporal Cortex (IT) tracing began at the mostanterior slice in which the mammillary bodies could beclearly seen. The posterior boundary was the slice followingthe last appearance of the splenium. The lateral boundarywas the inferior temporal sulcus, and the medial boundarywas defined by the occipito-temporal sulcus. In cases inwhich the occipito-temporal sulcus could not be clearlyobserved, the operator demarcated the medial boundary bybisecting the cortex at a 45◦ angle from the most ventro-medial extent of white matter to the surface of the brain.This ROI covered Brodmann area 20 exclusively, withoutits most anterior part.
Fusiform Gyrus (FG). The FG that spans temporal andoccipital lobes was traced starting at the most anterior sliceon which the AC first appeared. The most posterior slice onwhich the temporal portion of the FG was measured wasthe last slice on which the splenium of the corpus callosumcould be identified. The occipital portion of the FG spannedthe anterior 33% of the range between the occipital poles
N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 383
and the slice caudal to the one on which splenium was nolonger present. Between the first slice and the end of thesplenium, the occipito-temporal sulcus served as the lateralboundary of the FG. Starting at the first slice caudal to thesplenium, the most ventrolateral sulcus became the lateralboundary. In most cases, this still was the occipito-temporalsulcus. The collateral sulcus served as the medial boundaryof the FG. In the posterior portion of the FG, the collateralsulcus splits into two sulci. The most lateral sulcus of thetwo was used as the boundary. The medial (lingual) sulcuswas not included in the FG ROI. Thus, Brodmann areas 37and 19 were covered by this ROI.
Visual (Calcarine) Cortex (VC). The volume of the VCwas estimated as the volume of the cortical ribbon lin-ing the calcarine sulcus. This sulcus appeared as the mostventromedial sulcus in the temporal-occipital cortex at thecoronal slice that is mid-vermis or immediately caudal tomid-vermis. It was measured on the anterior 50% of thecoronal slices between the mid-vermis slice and the occip-ital pole. The inferior and superior boundaries of this ROIwere defined as the point at which the opening of the sulcusoccurred. At this point a line was drawn horizontally so thatno cortex (dorsal or ventral) outside of the calcarine sulcuswas included, confining this ROI to area 17.
Hippocampus (HC) volume was measured on a series of19–25 slices aligned perpendicular to the long axis of theright HC. The mammillary bodies defined the rostral bound-ary of the HC, and the slice showing the fornices rising fromthe fimbria marked its caudal boundary. Care was taken notto include the amygdala in this ROI. The resolution of theimages did not allow separate measurement of the HC sub-divisions (e.g. sectors CA1–CA4 and the subiculum). Thisdefinition of the HC used in our previous studies (e.g.[60])was a conservative one. It was designed to avoid confusionin determining the anterior HC boundary with that of theamygdala and confounding the inferior horn enlargementobserved in some older subjects with determination of theextent of the HC.
Intracranial Volume (ICV). The intracranial volume wasestimated from the coronal sections. The operator traced ICVon every eighth slice (range from 11 to 12 slices) betweenthe first slice following the orbits and last slice on whichbrain tissue was visible. Tracing began at the right side ofthe head and proceeded clockwise following the inner tableof the skull until the homologous mark was reached on theleft side of the head, at which point the tracing contourwas allowed to close automatically by drawing a straightline back to the origin. The reliability of this measure wasICC(2) = 0.99.
3. Results
Age, sex, and hemispheric differences in regional corticalvolumes were examined in the framework of a mixed gen-eral linear model. In the model, sex was a two-level cate-
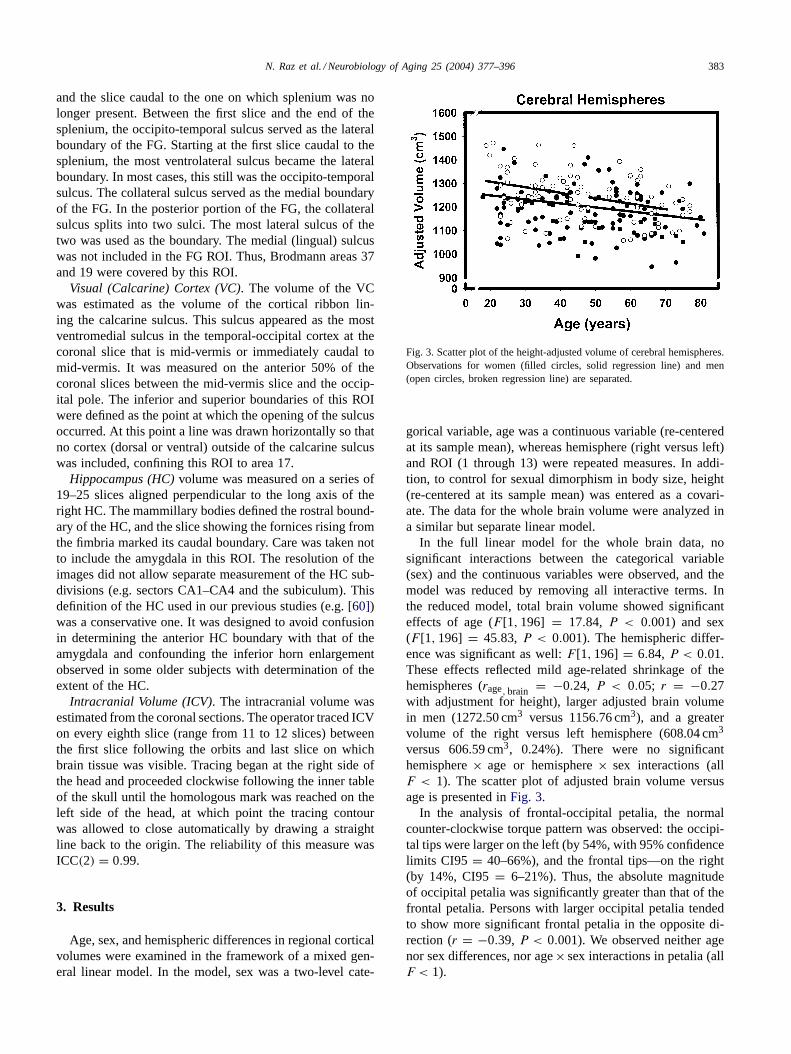
Fig. 3. Scatter plot of the height-adjusted volume of cerebral hemispheres.Observations for women (filled circles, solid regression line) and men(open circles, broken regression line) are separated.
gorical variable, age was a continuous variable (re-centeredat its sample mean), whereas hemisphere (right versus left)and ROI (1 through 13) were repeated measures. In addi-tion, to control for sexual dimorphism in body size, height(re-centered at its sample mean) was entered as a covari-ate. The data for the whole brain volume were analyzed ina similar but separate linear model.
In the full linear model for the whole brain data, nosignificant interactions between the categorical variable(sex) and the continuous variables were observed, and themodel was reduced by removing all interactive terms. Inthe reduced model, total brain volume showed significanteffects of age (F [1, 196] = 17.84, P < 0.001) and sex(F [1, 196] = 45.83, P < 0.001). The hemispheric differ-ence was significant as well:F [1, 196] = 6.84, P < 0.01.These effects reflected mild age-related shrinkage of thehemispheres (rage, brain = −0.24, P < 0.05; r = −0.27with adjustment for height), larger adjusted brain volumein men (1272.50 cm3 versus 1156.76 cm3), and a greatervolume of the right versus left hemisphere (608.04 cm3
versus 606.59 cm3, 0.24%). There were no significanthemisphere× age or hemisphere× sex interactions (allF < 1). The scatter plot of adjusted brain volume versusage is presented inFig. 3.
In the analysis of frontal-occipital petalia, the normalcounter-clockwise torque pattern was observed: the occipi-tal tips were larger on the left (by 54%, with 95% confidencelimits CI95 = 40–66%), and the frontal tips—on the right(by 14%, CI95= 6–21%). Thus, the absolute magnitudeof occipital petalia was significantly greater than that of thefrontal petalia. Persons with larger occipital petalia tendedto show more significant frontal petalia in the opposite di-rection (r = −0.39, P < 0.001). We observed neither agenor sex differences, nor age× sex interactions in petalia (allF < 1).

384 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
In the full linear model for the regional volumes, therewere no significant interactions between the categori-cal variable (sex) and the covariate (height). Hence, themodel was reduced by removing the nonsignificant interac-tive term. However, age× sex interaction was significant(F [1, 195] = 4.45, P < 0.05), and its removal would betantamount to accepting (incorrectly) the null hypothesis ofslope equivalence across the sexes. Thus, it was retainedin the model that revealed three significant interactions:age× ROI (F [12, 2340] = 19.08, P < 0.001), sex× ROI(F [12, 2340] = 5.33, P < 0.01), and hemisphere× ROI(F [12, 2340] = 14.17, P < 0.001). TheF-tests for allinteractions involving more than one degree of freedomin the numerator were adjusted with Huynh–Feldt unbi-ased correction that controls for violation of assumptionof homogeneity of variance–covariance matrix. In addi-tion, significant main effects of age (F [1, 195] = 58.08,P < 0.001), sex (F [1, 195] = 17.54, P < 0.001), andhemisphere (F [1, 195] = 10.23, P < 0.01) were found.Overall, men evidenced larger volumes across the ROIsand the hemispheres, older participants (men and women)had smaller volumes across ROIs, and the combination ofright side ROIs were slightly larger than its left-side ho-mologue. However, the extent of age, sex, and hemisphericdifferences varied across the ROIs.
To examine the pattern of the observed differences, wedecomposed the interactions into simple effects. The analy-
Table 1Correlation matrix of age and regional volumes adjusted for height
Variable Age LPFC OFC ACG FWHITE PrCG PoCG PHG ITC FC VC IPL PWHITE
Lateral prefrontalcortex (LPFC)
−0.61∗
Orbito-frontal cortex(OFC)
−0.43∗ 0.69∗
Anterior cingulategyrus (ACG)
−0.11 0.29∗∗ 0.27∗∗
Prefrontal whitematter (FWHITE)
−0.32∗∗ 0.67∗ 0.63∗ 0.30∗∗
Pre-central gyrus(PrCG)
−0.42∗ 0.52∗ 0.45∗ 0.30∗∗ 0.43∗
Post-central gyrus(PoCG)
−0.42∗ 0.53∗ 0.42∗ 0.19 0.38∗ 0.60∗
Parahippocampalgyrus (PHG)
−0.27∗∗ 0.39∗ 0.44∗ 0.19 0.26∗∗∗ 0.35∗ 0.29∗∗
Inferior temporalcortex (ITC)
−0.28∗∗ 0.26∗∗∗ 0.26∗∗∗ 0.18 0.23 0.39∗∗ 0.34∗∗ 0.33∗
Fusiform cortex (FC) −0.37∗ 0.47∗ 0.36∗ 0.33∗ 0.45∗ 0.50∗ 0.42∗ 0.44∗ 0.16Visual cortex (VC) −0.15 0.28∗∗ 0.29∗∗ 0.20 0.27∗∗ 0.28∗∗ 0.18 0.34∗ 0.27∗∗ 0.28∗∗Inferior parietal
lobule (IPL)−0.01 −0.09 0.05 −0.04 −0.02 −0.01 −0.06 0.08 0.47∗ 0.35∗ 0.15
Parietal white matter(PWHITE)
0.04 0.05 0.17 0.02 0.21 0.17 0.29∗∗ 0.17 0.47∗ 0.38∗ 0.24 0.80∗
Hippocampus (HC) −0.42∗ −0.56∗ 0.39∗ 0.24 0.47∗ 0.45∗ 0.43∗ 0.46∗ 0.37∗ 0.50∗ 0.29∗∗ 0.02 0.14
Pearson correlations, two-tailed; Bonferroni-adjusted for multiple comparisons.∗ P < 0.001.∗∗ P < 0.01.∗∗∗ P < 0.05.
ses of simple effects showed that the strength of associationbetween regional cortical volume and age differed acrossthe examined regions. To control for sexual dimorphism inbody size, all brain volumes were adjusted using covari-ance approach. The adjusted volumes were computed froma linear equation: Volumeadji = Volumerawi − b(Heighti −Mean Height) for each subjecti. In that equation, Volumeadjiis adjusted volume of a given ROI, Volumerawi is raw vol-ume of that ROI,b is a slope of ROI volume regressionon height, and Mean Height is a sample mean of height. Ineach case, the homogeneity of regression slopes (absence ofsex× height interaction) was checked to allow the use ofa common slope for both sexes. The correlations matrix ofheight-adjusted ROI volumes and age is presented inTable 1,and scatter plots and regressions of ROI volumes by age arepresented inFigs. 4–7.
The magnitude of correlations was compared among theregional volumes adjusted for height using Steiger’sZ∗ testthat takes into account the strength of association betweenthe members of all relevant pairs of variables[71]. Com-parisons of the magnitude of correlations with age betweenLPFC volume and the volumes the other 12 ROIs revealedstatistically significant differences, even after a conservativeBonferroni adjustment. The closest in size was the correla-tion between age and the volumes of the HC, orbito-frontalcortex, pre- and post-central gyri (eachr = −0.42; Steiger’sZ∗ = 3.49, 3.91, 3.57, 3.39, respectively, allP < 0.05,

N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 385
Fig. 4. Scatter plots of the height-adjusted volume of the lateral prefrontal cortex, the orbito-frontal cortex, the anterior cingulate gyrus, and the inferiorparietal lobule vs. age. Observations for women (filled circles, solid regression line) and men (open circles, broken regression line) are separated.
adjusted). The greatest difference was found between thecorrelations of age with LPFC and age with parietal whitematter volume (Z∗ = 7.68,P < 0.001, adjusted). Althoughthe prefrontal white matter volume showed a significantlyweaker age-related shrinkage than the LPFC (Z∗ = 5.91,P < 0.01, adjusted), the strength of its association with agewas greater than that of the parietal white matter (Z∗ =4.70, P < 0.01, adjusted). As illustrated on the regressionplots in Figs. 4–6, age-related differences in all ROIs withthe exception of the HC, are best described by linear equa-tions. The HC was the only region in which volume variancewas better explained by the addition of a quadratic age term(F [1, 197] = 9.27, P < 0.001).
We used the regression equations of the total and regionalbrain volumes on age to estimate the ROI shrinkage withinthe age span between 20 and 80 years. The linear shrinkageestimates that are presented inTable 2, range between essen-tially zero for the IPL and 5.36% per decade for the LPFC.The estimate of the hippocampal shrinkage was based ontwo linear regression equations (a bi-linear model fit) be-cause of the significant nonlinearity of its age trend. The
estimate for the age range of 18 through 50 suggested verylittle of a projected change (0.71% per decade), whereas theestimate of shrinkage rate for the age range of 50–80 wasthe highest among all ROIs: 6.38% per decade. To checkwhether the differences among the regions were an artifactof differential variability of the measures, we compared theircoefficients of variation (CV, a ratio of standard deviationto the mean). The comparison of CV values (presented inTable 2) yielded no substantial differences among the ROIs.The only exception was the IPL volume, which evidencedgreater variability than other ROIs (0.23 versus the medianof 0.12). The ICC values are very similar across all ROIs,and differential unreliability cannot explain the differencesin age–volume correlations.
The magnitude of sex differences in cortical volume ad-justed for height was compared across the ROIs using acommon metric of effect size (d [11,31]). The comparisonof effect sizes revealed that all ROI volumes (except IPL)were larger in men with the magnitude of the differencesranging fromd = 0.66 for the calcarine cortex,d = 0.51for the PHG andd = 0.48 for the ACG (moderate effects)

386 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
Fig. 5. Scatter plots of the height-adjusted cortical volume of the pre-central, post-central, and the parahippocampal gyri and primary visual (pericalcarine)cortex vs. age. Observations for women (filled circles, solid regression line) and men (open circles, broken regression line) are separated.
Table 2Descriptive statistics for regional cortical volumes
ROI Mean S.D. Shrinkage per decade (%) CV ICC(2)
Lateral prefrontal cortex 17.70 2.90 5.36 0.17 0.96Orbito-frontal cortex 8.66 1.24 3.35 0.15 0.93Pre-central gyrus 18.84 2.18 2.70 0.12 0.90Anterior cingulate gyrus 2.28 0.23 0.43 0.10 0.90Post-central gyrus 13.76 1.79 3.07 0.13 0.96Inferior temporal gyrus 11.07 1.60 2.29 0.14 0.95Fusiform cortex 18.61 2.24 2.51 0.12 0.93Parahippocampal gyrus 7.01 0.86 1.89 0.12 0.97Hippocampus 6.69 0.87 3.09, 0.711, 6.382 0.13 0.97Inferior parietal lobule 13.66 3.00 +0.14 0.23 0.96Visual (calcarine) cortex 5.60 1.10 1.70 0.19 0.94Prefrontal white matter 38.42 6.44 2.97 0.17 0.95Parietal white matter 22.86 3.61 +0.42 0.16 0.96
Two estimates for the hippocampus: (1) before age 50 and (2) after age 50 (bi-linear fit). Abbreviations: ROI, region of interest; S.D., standard deviation;CV, coefficient of variation; ICC(2), intraclass correlation, a random-raters formula, median reliability for all pairs of operators involved in tracing, butno pair of operators had ICC(2) lower than 0.89. All volumes (in cm3) are adjusted for height. Shrinkage is estimated from regression of each ROIvolume on age in a range of 20–80 years.

N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 387
Fig. 6. Scatter plots of the height-adjusted cortical volumes of the fusiform gyrus, the inferior temporal cortex, and the prefrontal and parietal whitematter vs. age. Observations for women (filled circles, solid regression line) and men (open circles, broken regression line) are separated.
Fig. 7. Scatter plot of the height-adjusted cortical volume of the volumeof the hippocampus vs. age. Observations for women (filled circles,solid quadratic regression line) and men (open circles, broken quadraticregression line) are separated.
to d = −0.00 for the IPL. Sex differences in the volumesof the prefrontal white matter (d = 0.38), and the HC (d =0.41) were somewhat smaller but fell within the 95% confi-dence limit of the PHG difference. Several ROIs (IT, OFC,pre- and post-central gyri) showed small sex differences involume (d ≤ 0.3), which were significantly smaller in com-parison with the calcarine cortex, but not with other ROIs.LPFC cortex and parietal white matter show very small andnon-significant sex differences (d = 0.17 andd = 0.16,respectively).
Significant age× sex interactions observed for someregions indicated that the magnitude of sex differences var-ied across the age span. In those ROIs (HC, FG, and theprefrontal white matter), substantial sex differences wereobserved among people younger than 46 years of age whileno sex-dependent variation in volume was noticed in theirolder counterparts. For the prefrontal white matter the dif-ference dropped fromd = 0.66 for the younger sub-sampleto d = −0.05 for the older group. For the fusiform gyrus,the comparison yieldedd = 0.84 for the young and onlyd = 0.10 for the older participants; in the HC, the difference

388 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
Table 3Hemispheric asymmetry of regional cortical volumes
ROI Volumes (cm3) unadjusted Direction and percentdifference
Right–left volumecorrelation
Magnitude of right–leftdifference (d)
R L
Lateral prefrontal cortex 8.89 8.82 R0.89 0.91 0.10Orbito-frontal cortex 4.95 4.31 L0.92 0.85 0.11Pre-central gyrus 9.40 9.44 R0.88 0.82 0.06Anterior cingulate gyrus 1.13 1.13 – 0.41 0.01Post-central gyrus 7.08 6.68 R5.81 0.81 0.66Inferior temporal gyrus 5.45 5.62 L3.07 0.82 0.34Fusiform cortex 9.44 9.18 R2.80 0.85 0.40Parahippocampal gyrus 3.39 3.61 L6.28 0.77 0.70Hippocampus 3.35 3.36 L0.59 0.85 0.08Inferior parietal lobule 6.92 6.72 R2.93 0.90 0.31Visual (calcarine) cortex 2.82 2.72 R3.61 0.85 0.11Prefrontal white matter 19.25 19.17 R0.42 0.93 0.06Parietal white matter 11.58 11.28 R2.62 0.89 0.34
Abbreviations: ROI, region of interest; R, right; L, left;d, unbiased effect size estimate[31].
of d = 0.77 for the younger subjects was reduced tod =0.04 for the older group. Another expression of the signif-icant age× sex interactions was sex differences in strengthof age–volume association that were detected the same twoof the examined ROIs. The HC evidenced a significantlystronger negative linear age trend in men than in womenwith regression slopes of−0.30 (±0.005) cm3 per year formen versus−0.016 (±0.004) cm3 per year for women (nooverlap in CI95). In the FG, the slope for men was 0.075(±0.014) cm3 per year versus 0.030 (±0.011) cm3 per yearfor women, with no overlap of CI95.
The magnitude and the direction of hemispheric asymme-try varied across the ROIs. Six ROIs (half of the examinedset) revealed significant hemispheric differences in volume(see Table 3). Some of the differences were substantial(larger PHG on the left and larger post-central gyrus onthe right). Others (the fusiform and the inferior parietalcortices, the parietal white matter and the inferior tem-poral cortex) showed mild asymmetry, whereas virtuallynone was found in the HC, VC and prefrontal regions andthe ACG.
Two health-related characteristics of the subjects—hyper-tension and postmenopausal hormone replacement therapy(HRT)—could have influenced the results. A history of hy-pertension can affect the appearance of the brain[54] andenhance age-related differences in regional cortical volumes[72]. Therefore, we repeated all analyses after removingfrom the sample 19 participants (7 men and 13 women,age range 39–74 years) who reported a history of medicallycontrolled hypertension. As the results remained virtuallyunchanged, we decided to retain that small subgroup ofparticipants in the sample.
Sex differences in the brain volumes and brain agingmay be affected by the hormonal status of the participants[77]. We, therefore, examined whether participation in HRTexplains part of variability to brain volumes. Twenty-one
women who reported receiving estrogen replacement ther-apy were closely matched on age (±1 year), hypertensionstatus, and race to 21 women who received no supplementaryestrogen. The average age of the groups was 59.45± 9.94years for the controls and 60.04±9.79 years for the estrogentherapy group (t < 1, ns). There were two African-Americanwomen in the estrogen group and three in the no-estrogengroup. Each group contained five women who were takinganti-hypertensive medication. The comparison of regionalvolumes and regional proportional measures revealed no dif-ferences related to the estrogen supplementation status—allF < 1.
3.1. The role of correction factor: subsidiary analyses
Because age differences in gross ROI volumes may reflectprocesses that are not necessarily related to the organism’ssize, raw volumes unadjusted for body size may consti-tute the most appropriate dependent variables. However, tocontrol for sexual dimorphism in body size we used heightas a body size index. Our reasoning was[60] that whenone is concerned with sex differences in brain volumes, anextra-cerebral measure of body size may be preferred toICV or head size. The disadvantage of using the ICV toadjust for body size is that ICV growth is influenced bygrowth of the brain. Thus, ICV variance contains varianceshared with the brain volume, which may be undesirableto remove from the brain measures. A valid counterpointmay be made in the case of aging, a process that may beassociated with longitudinal decline in stature and con-founded by secular trends in height[70]. However, in thissample, no associations between age and height were ob-served (r = −0.06 for men, andr = −0.01 for women,r =0.01 overall, all ns). Nonetheless, we re-analyzed the datawhile employing both raw volumes and volumes correctedfor ICV.

N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 389
Men had significantly larger crania than women:1992.47 ± 142.42 cm3 versus 1780.13 ± 125.78 cm3, t =11.11, P < 0.001. The correlation between the intracranialvolume and age wasr = 0.1, ns, whereas its correlationwith height wasr = 0.49, P < 0.001. Not surprisingly,there was a strong association between the total cerebralvolume and ICV:r = 0.73, P < 0.001. When intracranialvolume was used for correction (via covariance equationspecified above for height), no substantive differences in thepattern of results were observed. Some correlations betweencorrected volume and age increased by 0.01–0.09 correla-tion units, others decrease by 0.01–0.03 correlation units.The shape of the regression trends was preserved as well,including the nonlinearity of the HC–age relationship. Theonly exception was the correlation between age and the totalbrain size which increased fromr = −0.24 to r = −0.47.When raw volumes were analyzed and the results comparedto the findings with height-corrected data, the correlationsbetween age and brain volumes were slightly lower butno differences greater than 0.009 correlation points werenoted. Regardless of the correction method, the LPFC ex-hibited the strongest association with age as was the case forheight-corrected volumes. To preserve the continuity withour previous work and to allow comparison across the twosamples we present here the analyses with height-correctedmeasures.
4. Discussion
4.1. Differential brain aging
The answer to the main question of this study—whethera pattern of differential age-related shrinkage of the cerebralcortex is stable and replicable across samples—is affirma-tive, at least for the selected population of healthy, educatedadults from a major North American metropolitan center.Moreover, the results of this study strengthen the belief thatof all examined regions, the LPFC may be the most vulnera-ble to the effects of aging. On that account, our results agreewith the findings of several cross-sectional in vivo studiesof the aging brain (see[58] for a review of the literature and[2,36] for more recent publications). To gauge the stabilityof the observed pattern of differential aging, we compare themagnitude of the negative age–volume associations observedin two samples drawn from the same population: the first—described in a previous publication[60] and the secondone—reported here. The comparison presented inTable 4reveals that not only were the estimates of effect magnitudestatistically equivalent across the two samples, but their rankorder was also closely reproduced, with the LPFC show-ing the largest age-related difference and the IPL and VCrevealing the smallest correlations with age. Although thesomatosensory cortex (post-central gyrus), evidenced some-what stronger association with age than in the 1997 sample,the difference in the effect size was not significant.
The only significant exception in a well-replicated pat-tern of age differences in the two samples was found inthe pre-central gyrus. The volume of that ROI showed noage-related differences in the 1997 study but exhibited asignificant negative association with age in the current sam-ple. A possible reason for this discrepancy may be the in-clusion of a considerably larger volume of the pre-centralgyrus in this study (range 14–25 cm3) than in the 1997 re-port (3–8 cm3). Thus, in the 1997 sample, the definition ofthe primary MC might have been too restrictive, whereas inthe current sample they may have been too broad and possi-bly resulted in inclusion of the premotor and supplementarymotor areas.
Increased vulnerability of the prefrontal regions is con-sistent with the volumetric literature (reviewed in[58]) andthe studies of age-related differences in neurochemical andhemodynamic properties of the cerebral cortex. Amongmultiple examined regions, the prefrontal cortex exhibits thegreatest age-related alteration of GABA and glutamate[27],age differences in dopamine pharmacokinetics and glucosemetabolism[37,79] and age-related declines in regionalcerebral blood flow[3]. The estimated decline in LPFCvolume is, however, less dramatic than a linear age-relatedloss of dopamine receptors, which was estimated at 10–13%per decade[37,77]. Although both LPFC shrinkage andloss of functionality in the dopaminergic system have beentied to age-related cognitive declines[8,39], in the ab-sence of longitudinal studies that examine the relationshipbetween neurochemical and structural changes within thesame sample, it is impossible to establish which of the fac-tors or their interaction are primary in age-related cognitivedeficits.
The mechanisms underlying the predilection of the pre-frontal cortex for age-related shrinkage are unclear. Thefactors contributing to the emergence of such a patternmay include age-related declines in the catecholaminergicsystem that affects PFC neurons via projections, subcorti-cal ischemia at the periphery of the irrigation area of themiddle cerebral artery, and increased plasticity of the pre-frontal circuits (for a review see[58]). Another explanationmay be based on the fact that prefrontal areas contain agreater number of large neurons than occipital cortex, andincreased vulnerability of the large neurons creates the ob-served gradient as it does in some degenerative conditionssuch as AIDS-related encephalopathy[20]. Statistical fac-tors may play a role in generating some of the observeddifferences. Although all measures of the ROI volumeswere equally reliable, and almost all of them exhibitedequivalent variability, a notable example of the inferiorparietal lobule stands out. The volumes of IPL gray andwhite matter showed almost twice as great variability asthe other ROIs, and they did not correlate with any otherregional volumes. In principle, as phylogenetically recentassociation cortex, the IPL is expected to be as vulnerableto aging as the PFC to which it is connected by longitudinalfibers.

390 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
Table 4Comparison of differential aging patterns observed in two independent samples
ROI Sample 1 (1997) Sample 2 (2001) Different?
d CI95 d CI95
Lateral prefrontal cortex −1.18 −0.93 to−1.43 −1.53 −1.31 to−1.75 NoOrbito-frontal cortex −1.09 −0.84 to−1.33 −0.95 −0.74 to−1.16 NoPre-central gyrus 0.18 0.41 to−0.05 −0.92 −0.72 to−1.13 YesPost-central gyrus −0.54 −0.30 to−0.77 −0.92 −0.72 to−1.13 NoAnterior cingulate gyrus −0.36 −0.13 to−0.59 −0.22 0.02 to−0.42 NoInferior temporal gyrus −0.74 −0.51 to−0.98 −0.58 −0.38 to−0.78 NoFusiform cortex −0.74 −0.51 to−0.98 −0.79 −0.59 to−1.00 NoParahippocampal gyrus −0.36 −0.13 to−0.59 −0.56 −0.36 to−0.76 NoHippocampus −0.58 −0.35 to−0.81 −0.95 −0.74 to−1.16 NoInferior parietal lobule −0.36 −0.13 to−0.59 0.07 0.25 to−0.13 NoVisual (calcarine) cortex −0.39 −0.16 to−0.62 −0.30 −0.11 to−0.50 NoPrefrontal white matter −0.45 −0.22 to−0.68 −0.67 −0.47 to−0.87 NoParietal white matter −0.12 0.11 to−0.35 0.28 0.48 to−0.08 No
In the first study, the parietal white matter was reported separately for three regions: two of them were measured in the second study. These regionsshowed no age-related differences; they were combined into parietal white matter ROI. The compared result for study 1 is the mean of two correlations.Abbreviations: ROI, region of interest;d, unbiased effect size estimate; CI95, 95% confidence limits of the effect size.
The trajectories of age differences observed in this studysuggest linear declines for almost all examined ROIs. Wewere unable to replicate the reported nonlinear relation-ship between the white matter volume and age[2,13] orbetween the orbito-frontal cortex volume and age[60]. Anotable exception was the HC, which in contrast to mostof the previous reports (but in agreement with[5,44]) ex-hibited a nonlinear relationship with age. What factors canaccount for such a discrepancy? Because in both studies theimages were acquired on the same scanner and measuredaccording to the same set of rules by a significantly over-lapping group of operators, the sample variations appearto be the most likely culprit. Because subjects are almostnever recruited at random in in vivo volumetry studies,the samples may vary in the proportion of pre-clinically illparticipants. Recent estimates suggest that up to 20% ofnominally healthy participants in cognitive aging studiesmay actually suffer from early dementia[68]. Such contam-ination is possible in samples that, like ours, are composedof highly educated participants, who are more likely to passcognitive screening than their less educated counterpartsof equivalent neurological and cognitive status. Geneticvariability may also play a role in determining the discrep-ancies among the samples in the results concerning theHC volume. For instance, inheritance of an apolipoproteinE4/4 genotype (ApoE-ε4) was associated with longitudinalchanges in hippocampal volume, even at a short follow-up[43].
Relatively low statistical power could have pre-cludeddiscovery of nonlinear nature of age-related shrinkage ofthe HC in earlier studies. Examination of the relationshipbetween age and the hippocampal volume in our 1997sample reveals that a similar non-linear trend was hintedthere as well, but did not reach a conventional (P < 0.05)level of statistical significance (F [1, 145] = 1.74, ns). A
quadratic component is tested for significance only afterthe variance accounted by the linear component has beenremoved making the probability of detecting a significanthigher-order addition relatively low. The nonlinearity ofage–volume relationship in the HC may, however, reflect anonlinear (accelerating) trend in brain aging similar to thatobserved in aging of the fundamental cognitive resources[78].
The true nature of age-related processes underlying thedifferences in regional volumes that are observed in vivo isunclear. After initial reports of massive age-related neuronalloss [9], the pendulum of scientific opinion swung the op-posite direction, and the belief that in mammals, virtuallyno neurons are lost with age took hold (see[40,53] for re-views). A more nuanced view is suggested by findings thatalthough not massive and highly localized in specific gyri,laminae, sectors and types of cells, age-related neuronal lossis significant[67,82]. Postmortem studies of human brainsalso indicate that the number of neurons is associated withcortical volume, which is negatively associated with age[28–30]. In addition, postmortem investigations in humanand nonhuman primates[15,51] as well as spectroscopicstudies[10,64]suggest that some loss of neurons or at leasttheir shrinkage and loss of dendritic arborization may play asignificant role in regional (and specifically prefrontal) vol-ume reduction.
A fundamental limitation to the interpretation of ourresults is the fact that the age trends were estimated from re-gression models based on cross-sectional data. In using suchmodels we assume that time trends can be inferred from asnapshot of age-related differences thus ignoring possiblecohort effects and differences in developmental historiesprior to the time of measurement. Although it is likely thatafter adjustment of regional volumes for height (or cranialvolumes) the impact of secular increase in body size on age

N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 391
differences could be greatly reduced, only a within-subjectlongitudinal design allows determination of true age-relatedchanges. However, longitudinal studies are vulnerable todifferential attrition of participants and the ensuing positiveselection bias. Also, only a small part of the six-decadeage span sampled in this cross-sectional investigation couldbe covered in a longitudinal study. To date, longitudinalstudies of brain volume changes covered only short periods[62], were based on samples with a restricted age range atthe upper end of the life span[45,62,74,80], and employedcoarse measures such as grossly defined lobes combiningwhite and gray matter, cerebral ventricles, or white matterhyperintensities[62,73,80]. When more refined regionalmeasures were employed, they usually were restricted tothe HC and the PHG[33,45]. Thus, both cross-sectionalstudies and longitudinal studies of regional brain morphom-etry are necessary to gauge the true age effects on brainvolume.
Image analysis methods may influence the results ofin vivo study of the aging brain. Recently, in additionto highly reliable manual methods, semi-automated ap-proaches such as high-dimensional warping into normal-ized anatomical space became available[75]. The promiseof these methods lies in perfect repeatability of the mea-sures and reduction of labor demands on trained operators.However, as demonstrated by a recent study[76], man-ual, landmark-based volume measures and semi-automaticassessment based on local MR signal intensity addressdifferent aspects of neuroanatomy. Additional problem inmeasuring age differences in brain volumes stems fromthe fact that age is associated with changes inT1 relax-ation time [59] that determines image contrast. This con-found is of concern especially when automated methodsof image segmentation are used. Nonetheless, when theage-related differences are not uniform but vary acrossthe regions, the method-related variability becomes aless likely confound, unless a region-specific bias can bedemonstrated.
4.2. Sex differences
In accord with previous reports[12], men in the ex-amined sample evidenced larger cerebral hemispheres andlarger volumes of several ROIs than women did. Thesedifferences were not eliminated by adjustment for sex dif-ferences in body size. However, unlike age-related differ-ences, the pattern of localized sexual dimorphism observedin this study differed from the one found in several previousstudies, including our own sample drawn from the samepopulation.
Several factors could influence the magnitude of ob-served sex differences in regional volumes as well as sexdifferences in aging. The first concern is with adjustmentof volumes by regressing them on height. It is plausiblethat older women who are at risk for osteoporosis exhibitdisproportional shrinkage of stature. However, in this sam-
ple, no correlation between age and stature was observed ineither sex. The second concern is that differential exposureto estrogen could have alleviated some of the “naturally”occurring sex differences in older adults. However, thefact that some of the post-menopausal participants receivedestrogen replacement therapy was inconsequential to sexdifferences or lack thereof. Nonetheless, in the absenceof detailed assessment of the hormonal levels and thera-peutic doses used by our subjects, we cannot claim thathormonal status of the subjects is generally irrelevant tothe findings. Indeed, it has been reported that the dose ofendogenous estradiol is moderately related to cognitiveand behavioral performance of post-menopausal women[65], although studies on cognitive benefits of HRT yieldedinconsistent findings and revealed methodological flaws[32,38]. In future studies, assessment of hormonal levelsand rigorous control of the therapy regimen may clarifythe issue.
In two regions, age trends differed significantly betweenmen and women: The volumes of the HC and the FG showedstronger association with age in men. A strong trend wasobserved in the third ROI—prefrontal white matter, and innone of the examined regions was the opposite trend noted.In general, the tendency for men to show greater age-relateddifferences in comparison to women is more common thanthe opposite and in that sense our findings are consistentwith the literature[55,80]. However, the finding in the HCis in contrast to our previous study[60], in which no sexdifferences were observed in age-related shrinkage of thatregion. The lack of significant sex differences in age–volumeassociations in other ROIs may reflect stringency of subjectselection criteria. By selecting only people without majorage-related medical problems we could have created a biasagainst finding sex differences in age trends. If men aremore prone than women to age-related diseases, selection of“super-healthy” elderly would create a sample in which menwould be more dissimilar to the typical man than women tothe typical woman.
4.3. Cerebral asymmetry
Asymmetry of the petalia and the hemispheric volumesobserved in this sample is consistent with the literature.The pattern of global hemispheric asymmetry (right greaterthan left hemisphere and counter-clockwise torque) ap-peared throughout the age range. Nevertheless, this studydid not improve the consistency of the cumulative recordon regional cortical asymmetry. The findings in the currentsample were mostly at odds with our previous report: only5 out of 13 ROIs showed the same direction, not to men-tion the same magnitude, of volume difference. Thus, evenwithin the same population and under very similar measure-ment conditions, regional volume asymmetry is not a stableand reliable phenomenon. Although regional volumetricasymmetry may be a stable property of individual brains,it may be an unreliable phenomenon at the population

392 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
level, with a prominent exception of the planum temporale[82]. For example, even in a carefully demarcated primarymotor cortex of brains harvested from right-handed indi-viduals no asymmetry was found[56]. It is possible, thatstudies focusing on specific features of sulcal morphologyrather than regional volumes[1] may yield more consistentresults.
5. Summary and conclusions
The results of this study confirm the differential vulnera-bility of the LPFC to aging. The estimated linear decline ofLPFC volume exceeds age-related differences in the otherexamined cortical regions. However, the HC also exhibitsa comparably significant decline after the middle of thefifth decade of life. This pattern of results may indicate thatcortical aging represents a confluence of multiple processesthat exert their influence within at least two time windows.One, spanning the whole adult life-span, is characterizedby steady decline of cortical volume, with different cor-tical regions varying in the rate of decline. The secondwindow is limited to late adulthood (past approximately
Appendix A. Correlation matrix of age and regional volumes adjusted for the intracranial volume (ICV)
Variable Age LPFC OFC ACG FWHITE PrCG PoCG PHG ITC FC VC IPL PWHITE Meanvolume(cm3)
Lateral prefrontalcortex (LPFC)
−0.63∗ 17.70
Orbito-frontalcortex (OFC)
−0.49∗ 0.68∗ 8.66
Anterior cingulategyrus (ACG)
−0.14 0.26∗ 0.20 2.28
Prefrontal whitematter(FWHITE)
−0.39∗ 0.67∗ 0.58∗ 0.25∗∗ 38.42
Pre-central gyrus(PrCG)
−0.47∗ 0.51∗ 0.39∗ 0.25∗∗ 0.37∗ 18.85
Post-central gyrus(PoCG)
−0.47∗ 0.51∗ 0.37∗ 0.15 0.32∗ 0.57∗ 13.76
Parahippocampalgyrus (PHG)
−0.33∗∗∗ 0.37∗ 0.37∗ 0.12 0.16 0.28∗∗∗ 0.23 7.01
Inferior temporalcortex (ITC)
−0.33∗ 0.23 0.14 0.15 0.24 0.35∗ 0.29∗∗∗ 0.27∗∗∗ 11.74
Fusiform cortex(FC)
−0.44∗ 0.45∗ 0.28∗∗∗ 0.26∗∗ 0.37∗ 0.45∗ 0.37∗ 0.37∗ 0.63∗ 18.61
Visual cortex (VC) −0.21 0.25∗∗ 0.22 0.13 0.18 0.22 0.12 0.27∗∗∗ 0.21 0.19 5.60Inferior parietal
lobule (IPL)0.04 −0.12 −0.02 −0.11 −0.13 −0.08 −0.13 −0.00 0.43∗ 0.29∗∗∗ 0.07 13.66
Parietal whitematter (PWHITE)
0.01 0.02 0.04 −0.11 0.05 0.04 0.20 0.04 0.39∗ 0.27∗∗∗ 0.12 0.80∗ 22.86
Hippocampus (HC) −0.46∗ −0.54∗ 0.34∗ 0.19 0.43∗ 0.41∗ 0.39∗ 0.42∗ 0.32∗ 0.53∗ 0.24∗∗ 0.02 0.04 6.69
Pearson correlations, two-tailed; Bonferroni-adjusted for multiple comparisons.∗ P < 0.001.∗∗ P < 0.05.∗∗∗ P < 0.01.
45 years of age), and is exemplified by accelerated declinein a specific region—the HC. A similar two-componentview of brain aging has been recently proposed by Gabrieli[22], who suggested that normal brain aging may exhibit,in mild form, the patterns of differential shrinkage thatare evident in two age-related diseases: Parkinson disease(prefrontal cortex and striatum) and Alzheimer’s disease(HC). This intriguing hypothesis suggests that longitudinalstudies on subjects of a broad age range may show equiv-alent shrinkage of the prefrontal cortices at all ages, whilerevealing hippocampal volume reduction only in olderindividuals.
Acknowledgments
This study was supported in part by National Insti-tute of Health grant AG-11230. Some of the findingsreported here were presented at the Annual Meeting ofthe Society for Neuroscience, San Diego, November 14,2001. We thank Jimmy Dupuis, Shannon Edmonds, SteveLloyd, and Johnny McQuain for assistance in imageanalysis.

N.
Raz
etal./N
eurobiologyof
Aging
25(2004)
377–396393
Appendix B. Correlation matrix of age and raw (unadjusted) regional volumes
Variable Age LPFC OFC ACG FWHITE PrCG PoCG PHG ITC FC VC IPL PWHITE Mean volume(cm3)
Lateral prefrontalcortex (LPFC)
−60∗ 17.70
Orbito-frontalcortex (OFC)
−42∗ 0.69∗ 8.66
Anterior cingulategyrus (ACG)
−0.10 0.31∗∗ 0.30∗∗ 2.28
Prefrontal whitematter(FWHITE)
−0.31∗∗ 0.68∗ 0.64∗ 0.34∗ 38.42
Pre-central gyrus(PrCG)
−41∗ 0.53∗ 0.46∗ 0.34∗ 0.47∗ 18.85
Post-central gyrus(PoCG)
0.41∗ 0.54∗ 0.44∗ 0.23 0.41∗ 0.62∗ 13.76
Parahippocampalgyrus (PHG)
−0.27∗∗ 0.41∗ 0.45∗ 0.23 0.30∗∗ 0.38∗ 0.33∗∗ 7.01
Inferior temporalcortex (ITC)
−0.27∗∗ 0.28∗∗ 0.24∗∗∗ 0.23 0.27∗∗ 0.42∗ 0.40∗ 0.36∗ 11.74
Fusiform cortex(FC)
−0.36∗ 0.48∗ 0.38∗ 0.36∗ 0.48∗ 0.33∗ 0.44∗ 0.46∗ 0.68∗ 18.61
Visual cortex(VC)
−0.15 0.30∗∗ 0.32∗∗ 0.25∗∗∗ 0.32∗∗ 0.32∗∗ 0.23 0.37∗ 0.32∗ 0.32∗∗ 5.60
Inferior parietallobule (IPL)
0.01 −0.07 0.06 −0.03 −0.01 −0.01 −0.05 0.09 0.47∗ 0.35∗ 0.15 13.66
Parietal whitematter(PWHITE)
0.04 0.08 0.20 0.06 0.25∗∗∗ 0.20 0.32∗∗ 0.20 0.49∗ 0.41∗ 0.28∗∗ 0.79∗ 22.86
Hippocampus(HC)
−0.42∗ −0.57∗ 0.42∗ 0.27∗∗∗ 0.49∗ 0.47∗ 0.44∗ 0.48∗ 0.41∗ 0.56∗ 0.32∗∗ 0.08 0.17 6.69
Pearson correlations, two-tailed; Bonferroni-adjusted for multiple comparisons.∗ P < 0.001.∗∗ P < 0.01.∗∗∗ P < 0.05.

394N
.R
azet
al./Neurobiology
ofA
ging25
(2004)377–396
Appendix C. Raw ROI volumes by age and sex
Age bin 18–30 31–40 41–50 51–60 61–70 71–81
Sex M F M F M F M F M F M FN 17 21 15 23 18 24 8 22 15 19 8 10
Lateral prefrontal cortex 20.76± 2.95 19.79± 2.71 19.37± 2.15 18.38± 2.16 18.44± 3.39 17.67± 2.00 18.38± 2.84 16.62± 2.20 16.04± 1.93 15.16± 2.25 15.40± 2.26 14.17± 1.72Orbito-frontal cortex 9.87± 0.88 9.42± 1.46 9.04± 0.85 8.67± 1.19 9.03± 1.57 8.30± 1.00 8.67± 1.19 8.15± 1.00 8.70± 0.82 7.70± 0.97 8.23± 1.00 7.71± 1.13Anterior cingulate gyrus 2.37± 0.88 2.26± 0.22 2.48± 0.23 2.19± 0.17 2.39± 0.25 2.23± 0.21 2.19± 0.17 2.20± 0.20 2.44± 0.18 2.20± 0.23 2.30± 0.16 2.17± 0.25Prefrontal white matter 43.72± 6.88 38.12± 5.46 42.46± 6.05 37.17± 4.72 43.02± 9.18 36.96± 4.60 37.17± 4.73 37.55± 4.63 38.09± 6.47 33.65± 6.66 35.28± 5.38 33.83± 4.87Pre-central gyrus 20.50± 2.68 20.04± 1.52 21.17± 2.40 18.42± 1.83 9.57± 2.40 18.17± 1.95 18.42± 1.63 17.64± 1.70 18.81± 1.38 17.56± 1.88 18.12± 1.81 17.37± 2.02Post-central gyrus 15.61± 1.98 14.56± 1.67 14.93± 1.93 13.80± 1.92 14.15± 1.76 13.14± 1.64 13.80± 1.92 12.90± 1.44 13.51± 1.31 12.77± 1.79 13.38± 1.97 12.85± 1.56Parahippocampal gyrus 7.79± 0.73 7.08± 0.66 7.56± 0.58 6.69± 0.77 7.15± 0.60 6.99± 1.05 6.67± 0.76 6.72± 0.91 7.15± 0.83 6.68± 0.72 6.89± 0.92 5.99± 0.52Inferior temporal cortex 12.66± 1.43 11.33± 1.44 12.21± 2.40 10.70± 1.63 11.08± 1.13 10.84± 1.22 10.46± 2.00 10.45± 1.34 11.22± 1.58 10.72± 1.73 11.08± 1.08 9.86± 0.83Fusiform cortex 21.03± 2.49 18.64± 2.29 21.02± 2.12 17.85± 1.72 19.14± 2.06 18.55± 1.54 17.85± 1.72 17.84± 1.49 18.69± 1.67 17.56± 1.96 17.16± 2.40 16.52± 2.25Visual cortex 6.02± 1.15 5.52± 1.10 6.28± 1.33 5.39± 0.90 6.07± 0.94 5.40± 0.98 5.39± 0.90 5.19± 0.96 5.77± 0.96 5.24± 1.08 6.09± 1.38 4.88± 0.83Inferior parietal lobule 15.25± 3.74 13.23± 2.72 12.93± 3.39 13.21± 2.91 13.74± 2.02 13.47± 3.47 13.21± 2.91 13.69± 2.96 13.54± 3.22 14.45± 3.86 13.35± 2.37 13.53± 2.68Parietal white matter 24.89± 4.08 21.55± 2.40 22.89± 4.81 21.88± 3.10 23.74± 2.30 21.65± 4.06 21.88± 2.91 22.61± 3.39 23.91± 3.67 23.74± 3.89 23.07± 4.30 22.49± 3.43Hippocampus 7.38± 0.84 6.64± 0.51 7.52± 0.90 6.77± 0.80 7.25± 0.79 6.50± 0.56 6.77± 0.79 6.61± 0.90 6.32± 0.56 6.17± 0.56 5.99± 1.00 5.69± 0.89
The volumes (means± standard deviation) are in cm3.

N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396 395
References
[1] Amunts K, Jäncke L, Mohlberg H, Steinmetz H, Zilles K.Interhemispheric asymmetry of the human motor cortex related tohandedness and gender. Neuropsychologia 2000;38:304–12.
[2] Bartzokis G, Beckson M, Lu PH, Nuechterlein KH, Edwards N,Mintz J. Age-related changes in frontal and temporal volume inmen: a magnetic resonance imaging study. Arch Gen Psychiatry2001;58:461–5.
[3] Bentourkia M, Bol A, Ivanoiu A, Labar D, Sibomana M, CoppensA, et al. Comparison of regional cerebral blood flow and glucosemetabolism in the normal brain: effect of aging. J Neurol Sci2000;181:19–28.
[4] Bhatia S, Bookheimer SY, Gaillard WD, Theodore WH. Measurementof whole temporal lobe and hippocampus for MR volumetry:normative data. Neurology 1993;43:2006–10.
[5] Bigler ED, Andersob CV, Blatter D. Temporal lobe morphologyin normal aging and traumatic brain injury. Am J Neuroradiol2002;23:255–66.
[6] Blatter DD, Bigler ED, Gale SD, Johnson SC, Anderson CV, BurnettBM, et al. Quantitative volumetric analysis of brain MR: normativedatabase spanning 5 decades of life. Am J Neuroradi 1995;16:241–51.
[7] Blessed G, Tomlinson BE, Roth M. The association betweenquantitative measures of dementia and senile change in the cerebralgray matter of elderly subjects. Br J Psychiatry 1968;114:797–811.
[8] Braver TS, Barch DM, Keys BA, Carter CS, Cohen JD, Kaye JA, etal. Context processing in older adults: evidence for a theory relatingcognitive control to neurobiology in healthy aging. J Exp PsycholGen 2001;130:746–63.
[9] Brody H. Organization of the cerebral cortex. III. J Comp Neurol1955;102:511–56.
[10] Brooks R, Kemp G, Lie W. A proton magnetic resonance spectro-scopy study of age-related changes in frontal lobe metaboliteconcentrations. Cereb Cortex 2001;11:598–605.
[11] Cohen J. Power analysis in behavioral sciences. Hillsdale, NJ:Lawrence Erlbaum; 1988.
[12] Coffey CE, Lucke JF, Saxton JA, Ratcliff G, Unitas LJ, Billig B, etal. Sex differences in brain aging: a quantitative magnetic resonanceimaging study. Arch Neurol 1998;55:169–79.
[13] Courchesne E, Chisum HJ, Townsend J, Cowles A, Covington J,Egaas B, et al. Normal brain development and aging: quantitativeanalysis at in vivo MR imaging in healthy volunteers. Radiology2000;216:672–82.
[14] DeArmond SJ, Fusco MM, Dewey MM. Structure of the humanbrain: a photographic atlas. New York: Oxford University Press;1976.
[15] de Brabander JM, Kramers RJK, Uylings HBM. Layer-specificdendritic regression of pyramidal cells with ageing in the humanprefrontal cortex. Eur J Neurosci 1998;10:1261–9.
[16] de Toledo-Morrell L, Sullivan MP, Morrell F, Spanovic C, SpencerS. Gender differences in vulnerability of the hippocampal formationduring aging. Soc Neurosci 1995;21:1708.
[17] Duvernoy HM. In: Bergmann JF, editor. The human hippocampus:an atlas of applied anatomy. Munchen: Verlag; 1988.
[18] Duvernoy H. The human brain: surface, three-dimensional sectionalanatomy, and MRI. Vienna: Springer; 1991.
[19] Filipek R, Richelme C, Kennedy D, Gaviness V. The young adulthuman brain: an MRI based morphometric analysis. Cereb Cortex1994;4:344–60.
[20] Fischer CP, Jorgen G, Gundersen H, Pakkenberg B. Preferential lossof large neocortical neurons during HIV infection: a study of thesize distribution of neocortical neurons in the human brain. BrainRes 1999;828:119–26.
[21] Frederikse ME, Lu A, Aylward E, Barta P, Pearlson G. Sex differencesin the inferior parietal lobule. Cereb Cortex 1999;9:896–901.
[22] Gabrieli J. Distinguishing healthy and pathological aging. In:Proceedings of the Symposium on Neuroscience, Aging, andCognition (SNAC). San Francisco, CA, April 14, 2002.
[23] Galaburda A. Anatomic basis of cerebral dominance. In: DavidsonRJ, Hughdal K, editors. Brain asymmetry. Cambridge, MA: MITPress; 1995. p. 51–73.
[24] Goldstein JM, Seidman LJ, Horton NJ, Makris N, Kennedy DN,Caviness VS, et al. Normal sexual dimorphism of the adult humanbrain assessed by in vivo magnetic resonance imaging. Cereb Cortex2001;11:490–7.
[25] Good C, Johnsrude IS, Ashburner J, Henson RNA, Friston KJ,Frackowiak RSJ. A voxel-based morphometric study of ageing in465 normal adult human brains. NeuroImage 2001a;14:21–36.
[26] Good C, Johnsrude IS, Ashburner J, Henson RNA, Friston KJ,Frackowiak RSJ. Cerebral asymmetry and the effects of sex andhandedness on brain structure: a voxel-based morphometric analysisof 465 normal adult human brains. NeuroImage 2001b;14:685–700.
[27] Grachev ID, Swarnkar A, Szeverenyi NM, Ramachandran TS,Apkarian AV. Aging alters the multichemical networking profileof the human brain: an in vivo1H-MRS study of young versusmiddle-aged subjects. J Neurochem 2001;77:292–303.
[28] Gur RC, Mozley D, Resnick SM, Gottlieb GL, Kohn M, ZimmermanR, et al. Gender differences in age effect on brain atrophymeasured by magnetic resonance imaging. Proc Natl Acad Sci USA1991;88:2845–9.
[29] Gundersen B, Pakkenberg HJG. Neocortical neuron number inhumans: effects of sex and age. J Comp Neurol 1997;384:312–20.
[30] Harding AJ, Halliday GM, Kril JJ. Variation in hippocampal neuronnumber with age and brain volume. Cereb Cortex 1998;8:710–8.
[31] Hedges L, Olkin I. Statistical methods for meta-analysis. Orlando:Academic Press; 1985.
[32] Hogervorst E, Williams J, Dudge M, Riedel W, Jolles J. Thenature of the effects of female gonadal hormone replacement therapyon cognitive function in post-menopausal women: a meta-analysis.Neuroscience 2000;101:485–512.
[33] Jack CR, Petersen RC, Xu Y, O’Brien PC, Smith GE, Ivnik RJ,et al. Rate of medial temporal lobe atrophy in typical aging andAlzheimer’s disease. Neurology 1998;51:993–9.
[34] Jernigan TL, Press GA, Hesselink JR. Methods for measuring brainmorphological features on magnetic resonance images: validation ofand normal aging. Arch Neurol 1990;47:27–32.
[35] Jernigan TL, Archibald SL, Berhow MT, Sowell ER, Foster DS,Hesselink JR. Cerebral structure on MRI. Part I: Localization ofage-related changes. Biol Psychiatry 1991;29:55–67.
[36] Jernigan TL, Archibald SL, Fenema-Notestine C, Gamst AC, StoutJC, Bonner J, et al. Effects of age on tissues and regions of thecerebrum and cerebellum. Neurobiol Aging 2001;22:581–94.
[37] Kaasinen V, Vilkman H, Hietala J, Nagren K, Helenius H, OlssonH, et al. Age-related dopamine D2/D3 receptor loss in extrastriatalregions of the human brain. Neurobiol Aging 2000;21:683–8.
[38] LeBlanc ES, Janowsky J, Chan BKS, Nelson HD. Hormonereplacement therapy and cognition: systematic review and meta-ana-lysis. JAMA 2001;285:1489–99.
[39] Li S-C, Lindenberger U, Frensch PA. Unifying cognitive aging: fromneuromodulation to representation of cognition. Neurocomputing2000;32-33:879–90.
[40] Long JM, Mouton PR, Jucker M, Ingram D. What counts inbrain aging? Design-based stereological analysis of cell number. JGerontol: Biol Sci 1999;54A:B407–17.
[41] Magnotta VA, Andreassen NC, Schultz SK, Harris G, Cizadlo T,Heckel D, et al. Quantitative in vivo measurement of gyrificationin the human brain: changes associated with aging. Cereb Cortex1999;9:151–60.
[42] Matsumae M, Kikinis R, Mórocz IA, Lorenzo AV, Sándor T,Albert MS, et al. Age-related changes in intracranial compartmentvolumes in normal adults assessed by magnetic resonance imaging.J Neurosurg 1996;84:982–91.

396 N. Raz et al. / Neurobiology of Aging 25 (2004) 377–396
[43] Moffat SD, Szekely CA, Zonderman AB, Kabani NJ, ResnickSM. Longitudinal change in hippocampal volume as a function ofapolipoprotein E genotype. Neurology 2000;55:134–6.
[44] Mu Q, Xie J, Wen Z, Weng Y, Shayun Z. A quantitative study ofthe hippocampal formation the amygdala, and the temporal horn ofthe lateral ventricle in healthy subjects 40–90 years of age. Am JNeuroradiol 1999;20:207–11.
[45] Mueller EA, Moore MM, Kerr DCR, Sexton G, Camicioli RM,Howieson DB, et al. Brain volume preserved in healthy elderlythrough the eleventh decade. Neurology 1998;51:1555–62.
[46] Murphy DG, DeCarli C, McIntosh AR, Daly E, Mentis MJ, Pietrini P,et al. Sex differences in human brain morphometry and metabolism:an in vivo quantitative magnetic resonance imaging and positronemission tomography study on the effect of aging. Arch GenPsychiatry 1996;53:585–94.
[47] Noupoulos P, Flaum M, O’Leary D, Andreassen N. Sexualdimorphism in human brain: evaluation of tissue volume, tissuecomposition and surface anatomy using magnetic resonance imaging.Psychiatry Res: Neuroimaging 2000;98:1–13.
[48] Ohnishi T, Matsuda H, Tabira T, Asada T, Uno M. Changes in brainmorphology in Alzheimer disease and normal aging: is Alzheimerdisease an exaggerated aging process? AJNR Am J Neuroradiol2001;22:1680–5.
[49] Oldfield RC. The assessment and analysis of handedness. Neuropsy-chologia 1971;9:97–113.
[50] Ono M, Kubik S, Abernathey CD. Atlas of cerebral sulci. Stuttgart:Thieme; 1990.
[51] Page TL, Einstein M, Duan H, He Y, Flores T, Rolshud D, etal. Morphological alterations in neurons forming corticocorticalprojections in the neocortex of aged Patas monkeys. Neurosci Lett2002;317:37–41.
[52] Passe TJ, Rajagopalan P, Tupler LA, Byrum CE, MacFall JR,Krishnan KR. Age and sex effects on brain morphology. ProgNeuropsychopharmacol Biol Psychiatry 1997;21:1231–7.
[53] Peters A, Morrison JH, Rosene DL, Hyman BT. Are neurons lostfrom the primate cerebral cortex during normal aging? Cereb Cortex1998;8:295–300.
[54] Petrovitch H, White LR, Izmirilian G, et al. Midlife blood pressureand neuritic plaques, neurofibrillary tangles, and brain weight indeath: HAAS. Neurobiol Aging 2001;21:57–62.
[55] Pruessner JC, Collins DL, Pruessner M, Evans AC. Age and genderpredict volume decline in the anterior and posterior hippocampus inearly adulthood. J Neurosci 2001;21:194–200.
[56] Rademacher J, Bürgel U, Geyer S, Schormann T, Schleichner A,Freund HJ, et al. Variability and asymmetry in the human precentralmotor system: a cytoarchitechtonic and myeologenic brain study.Brain 2001;124:2232–58.
[57] Radloff LS. The CES-D scale: a self-report depression scale forresearch in the general population. Appl Psych Measure 1977;1:385–401.
[58] Raz N. Aging of the brain and its impact on cognitive performance:integration of structural and functional findings. In: Craik FIM,Salthouse TA, editors. Handbook of aging and cognition—II.Mahwah, NJ: Erlbaum; 2000. p. 1–90.
[59] Raz N, Millman D, Sarpel G. Cerebral correlates of cognitive aging:grey-white matter differentiation in the medial temporal lobes, andfluid versus crystallized abilities. Psychobiology 1990;18:475–81.
[60] Raz N, Gunning FM, Head D, Dupuis JH, McQuain JD, Briggs SD,et al. Selective aging of the human cerebral cortex observed in vivo:differential vulnerability of the prefrontal gray matter. Cereb Cortex1997;7:268–82.
[61] Reiss A, Hennessey J, Rubin M, Beach L, Subramaniam B.BrainImage, v. 2.3.3. Behavioral Neurogenetics and NeuroimagingResearch Center, Division of the Kennedy Krieger Institute,Baltimore, MD, 1995.
[62] Resnick SM, Goldszal AF, Davatzikos C, Golski S, Kraut MA,Metter EJ, et al. One-year age changes in MRI brain volumes inolder adults. Cereb Cortex 2000;10:464–72.
[63] Rosen GD, Harry JD. Brain volume estimation from serial sectionmeasurements: a comparison of methodologies. J Neurosci Methods1990;35:115–24.
[64] Schuff N, Amend DL, Knowlton R, Norman D, Fein G, Weiner MW.Age-related metabolite changes and volume loss in the hippocampusby magnetic resonance spectroscopy and imaging. Neurobiol Aging1999;20:279–85.
[65] Senanorong V, Vannasaeng S, Poungavarin N, Ploybutrs S,Undompunthurak S, Jamjwamas P, et al. Endogenous estradiol inelderly individuals. Arch Neurol 2002;59:385–9.
[66] Shrout PE, Fleiss JL. Intraclass correlations: uses in assessing ratersreliability. Psychol Bull 1979;86:420–8.
[67] Simic G, Kotovic I, Windlad B, Bogdanovic N. Volume and numberof neurons in human hippocampal formation in normal aging andAlzheimer’s disease. J Comp Neurol 1997;379:482–94.
[68] Sliwinski M, Buschke H. Cross-sectional and longitudinal relation-ships among age, cognition, and processing speed. Psychol Aging1999;14:8–33.
[69] Shapleske J, Rossell SL, Woodruff PWR, David AS. The planumtemporale: a systematic, quantitative of its structural, functional andclinical significance. Brain Res Rev 1999;29:26–49.
[70] Sorkin JD, Muller DC, Reubin A. Longitudinal changes in the heightof men and women: consequential effects on body mass index.Epidemiol Rev 1999;21:247–60.
[71] Steiger JH. Tests for comparing elements of a correlation matrix.Psychol Bull 1980;87:245–51.
[72] Strassburger TL, Lee HC, Daly EM, Szczepanik J, Krasuski JS,Mentis MJ, et al. Interactive effects of age and hypertension onvolumes of brain structures. Stroke 1997;28:1410–7.
[73] Sullivan EV, Pfefferbaum A, Adalsteinsson E, Swan G, Carmelli D.Differential rates of regional brain change in callosal and ventricularsize: a 4-year longitudinal MRI study of elderly men. Cereb Cortex2002;12:438–45.
[74] Tang Y, Whitman GT, Lopez I, Baloh RW. Brain volume changeson longitudinal magnetic resonance imaging in normal older people.J Neuroimaging 2001;11:393–400.
[75] Thompson PM, MacDonald D, Mega MS, Holmes CJ, Evans AC,Toga AW. Detection and mapping of abnormal brain structure witha probabilistic atlas of cortical surfaces. J Comput Assist Tomogr1997;21:567–81.
[76] Tisserand D, Pruessner J, Sanz Arigita E, van Boxtel M, Evans A,Jolles J, et al. Regional frontal cortical volumes decrease differentiallyin aging: an MRI study to compare volumetric approaches andvoxel-based morphometry. Neuroimage 2002;17:657–69.
[77] Van Amelsvoort T, Compton J, Murphy D. In vivo assessment of theeffects of estrogen on the human brain. Trends Endocrinol Metab2001;12:273–6.
[78] Verhaeghen P, Salthouse TA. Meta-analyses of age–cognitionrelations in adulthood estimates of linear and nonlinear age effectsand structural models. Psychol Bull 1997;122:231–49.
[79] Volkow ND, Logan J, Fowler JS, Wang GJ, Gur RC, Wong C, et al.Association between age-related decline in brain dopamine activityand impairment in frontal and cingulate metabolism. Am J Psychiatry2000;157:75–80.
[80] Wahlund LO, Almkvist O, Basun H, Julin P. MRI in successfulaging, a 5-year follow-up study from the eighth to ninth decade oflife. Magn Reson Imaging 1996;14:601–8.
[81] Evans AC. Structural asymmetries in the human brain: a voxel-basedstatistical analysis of 142 MRI scans. Cereb Cortex 2001;11:868–77.
[82] West MJ. Regionally specific loss of neurons in the aging humanhippocampus. Neurobiol Aging 1993;14:287–93.
[83] Xu J, Kobayashi S, Yamaguchi S, Iijima K, Okada K, YamashitaK. Gender effects on age-related changes in brain structure. Am JNeuroradiol 2000;21:112–8.
[84] Zilles K, Dabringhaus A, Geyer S, Amunts K, Qü M, SchleicherA, et al. Structural asymmetries in the human forebrain and theforebrain of non-human primates and rats. Neurosci Biobehav Rev1996;20:593–605.


![Theta burst stimulation-induced inhibition of dorsolateral prefrontal cortex reveals hemispheric asymmetry in striatal dopamine release during a set-shifting task - a TMS-[ 11 C]raclopride](https://static.fdokumen.com/doc/165x107/634562de596bdb97a908e8a4/theta-burst-stimulation-induced-inhibition-of-dorsolateral-prefrontal-cortex-reveals.jpg)

















