
Exposure of human leukemic cells to direct electric current
14
ORIGINAL ARTICLE © Copyright 2005 by Humana Press Inc. All rights of any nature whatsoever reserved. 1085-9195/05/42:61–74/$30.00 Cell Biochemistry and Biophysics 61 Volume 42, 2005 INTRODUCTION Direct electric current (DC) can influence the growth and cell biology of several systems (1–5). Because of such properties, DC has been widely used for the treatment of tumors (6–11). However, the use of DC as an alternative therapy for cancer is limited by the poor knowledge of the cellular effects generated in different conditions in which animal cells are exposed to electric fields (2). In this context, different polarities of electric current have been shown to induce diverse effects on animal cells, which is reflected by the vary- ing efficacies of treatments of cancers with DC using positive or negative electrodes (8,12–14). In such experimental systems, cells are exposed to DC as they are to the soluble products generated after oxidation or reduction of physiologic compounds (15–17). Therefore, the products of electrolysis can certainly influence the cells that are exposed to DC in aqueous systems. Parameters involved in DC treatment such as electric field strength, electrode polarity, and period of stimulation also influence the type of cell response to an electric stimulus. For instance, the suppression or Exposure of Human Leukemic Cells to Direct Electric Current Generation of Toxic Compounds Inducing Cell Death by Different Mechanisms Venicio F. Veiga, 1 Leonardo Nimrichter, 1 Cesar A. Teixeira, 3 Marcelo M. Morales, 2 Celuta S. Alviano, 1 Marcio L. Rodrigues, 1 and Carla Holandino 3, * 1 Instituto de Microbiologia Professor Paulo de Góes, 2 Instituto de Biofísica Carlos Chagas Filho, and 3 Departamento de Medicamentos-Faculdade de Farmácia, Centro de Ciências da Saúde, Universidade Federal do Rio de Janeiro, CCS, Bloco K, Segundo Andar, Sala 50, Ilha do Fund ~ ao, 21941–590, Rio de Janeiro, Brazil Abstract Treatment with direct electric current (DC) influences the growth of several cancer cells. In this work, we evaluated the effects of DC treatment on the human leukemic cell line HL60. Human cells were separately treated in the presence of the cathode or the anode or without contact with the electrodes. In all systems, DC- treated cells presented an impaired ability to proliferate. Growth inhibition was dependent on the generation of soluble products of electrolysis. Cathodic treatment of HL60 cells predominantly induced lysis, whereas treatment without contact with electrodes did not induce alterations in cell viability. In contrast, cell stimula- tion by the anode resulted in irreversible membrane damage, as demonstrated by trypan blue and 7- aminoactinomycin staining. Analysis of these cells by transmission electron microscopy indicated that necrosis is a major mechanism inducing cell death. In addition, apoptotic-like cells were observed under light microscopy after anodic treatment. Accordingly, DNAfrom anodic-treated cells presented a typical pattern of apoptosis. Apoptotic cell death was only generated after the treatment of HL60 cells in conditions in which the generation of chloride-derived compounds was favored. These results indicate that the nature of the prod- ucts from cathodic or anodic reactions differently influences the mechanisms of cell death induced by DC- derived toxic compounds. Index Entries: Direct electric current; human leukemic cells; tumor growth; apoptosis; necrosis. *Author to whom all correspondence and reprint requests should be addressed. E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Exposure of human leukemic cells to direct electric current

ORIGINAL ARTICLE
© Copyright 2005 by Humana Press Inc.All rights of any nature whatsoever reserved.1085-9195/05/42:61–74/$30.00
Cell Biochemistry and Biophysics 61 Volume 42, 2005
INTRODUCTION
Direct electric current (DC) can influence thegrowth and cell biology of several systems (1–5).Because of such properties, DC has been widely usedfor the treatment of tumors (6–11). However, the useof DC as an alternative therapy for cancer is limited bythe poor knowledge of the cellular effects generated indifferent conditions in which animal cells are exposedto electric fields (2). In this context, different polarities
of electric current have been shown to induce diverseeffects on animal cells, which is reflected by the vary-ing efficacies of treatments of cancers with DC usingpositive or negative electrodes (8,12–14). In suchexperimental systems, cells are exposed to DC as theyare to the soluble products generated after oxidationor reduction of physiologic compounds (15–17).Therefore, the products of electrolysis can certainlyinfluence the cells that are exposed to DC in aqueoussystems. Parameters involved in DC treatment such aselectric field strength, electrode polarity, and period ofstimulation also influence the type of cell response toan electric stimulus. For instance, the suppression or
Exposure of Human Leukemic Cells to Direct Electric Current
Generation of Toxic Compounds Inducing Cell Death by Different Mechanisms
Venicio F. Veiga,1 Leonardo Nimrichter,1 Cesar A. Teixeira,3 Marcelo M. Morales,2
Celuta S. Alviano,1 Marcio L. Rodrigues,1 and Carla Holandino3,*1Instituto de Microbiologia Professor Paulo de Góes, 2Instituto de Biofísica Carlos Chagas Filho,
and 3Departamento de Medicamentos-Faculdade de Farmácia, Centro de Ciências da Saúde, Universidade Federal do Rio de Janeiro, CCS, Bloco K, Segundo Andar, Sala 50, Ilha do Fund~ao,
21941–590, Rio de Janeiro, Brazil
Abstract
Treatment with direct electric current (DC) influences the growth of several cancer cells. In this work, weevaluated the effects of DC treatment on the human leukemic cell line HL60. Human cells were separatelytreated in the presence of the cathode or the anode or without contact with the electrodes. In all systems, DC-treated cells presented an impaired ability to proliferate. Growth inhibition was dependent on the generationof soluble products of electrolysis. Cathodic treatment of HL60 cells predominantly induced lysis, whereastreatment without contact with electrodes did not induce alterations in cell viability. In contrast, cell stimula-tion by the anode resulted in irreversible membrane damage, as demonstrated by trypan blue and 7-aminoactinomycin staining. Analysis of these cells by transmission electron microscopy indicated thatnecrosis is a major mechanism inducing cell death. In addition, apoptotic-like cells were observed under lightmicroscopy after anodic treatment. Accordingly, DNA from anodic-treated cells presented a typical pattern ofapoptosis. Apoptotic cell death was only generated after the treatment of HL60 cells in conditions in whichthe generation of chloride-derived compounds was favored. These results indicate that the nature of the prod-ucts from cathodic or anodic reactions differently influences the mechanisms of cell death induced by DC-derived toxic compounds.
Index Entries: Direct electric current; human leukemic cells; tumor growth; apoptosis; necrosis.
*Author to whom all correspondence and reprint requestsshould be addressed. E-mail: [email protected]

enhancement of proliferation of mouse lymphomacells varied significantly when the cells were electri-cally stimulated within a narrow range of low-levelDC (5).
Mechanisms of DC-induced cell growth or inhibitionhave been described in different experimental systems.For example, DC treatment of multicellular tumorspheroids results in adenosine triphosphate release,which concomitantly activates purinergic receptors,elicits a Ca2+ wave spreading through the tumor spher-oid tissue, and stimulates tumor growth (18). Calciumrelease seems in fact to be involved in DC-inducedtumor growth. In a previous study by Wartenberg andcoworkers (19), a single electrical field pulse inducesproduction of reactive oxygen species, which mediatedrelease of intracellular Ca2+ and activated cell-cycleactivity in multicellular spheroids tumors. On the otherhand, DC-treated tumor cells can have their growthinhibited, depending on cell type and conditions of elec-tric stimulation. The mechanisms involved in such inhi-bition include changes in membrane conformation,alterations in cell organization, and appearance of mito-chondrial lesions (15,16,20).
Apoptosis is another biologic process apparently asso-ciated with DC-induced tumor reduction. Kurokawa andcoworkers (21) described that treatment of humanleukemic cell lines with DC intensities varying from 0.2to 5 mA-induced cell shrinkage and internucleosomalDNA fragmentation. In their model, cells were simulta-neously submitted to anodic and cathodic reactions,which impairs the establishment of a direct relationbetween the products of electrolysis and the cellulardamage generated after stimulation with DC.
We have previously shown that the nature of cyto-toxicity generated by DC is dependent on the polarity ofthe electrode used for treatment of tumor cells (16,17).In an experimental model in which cells are separatelyexposed to cathodic or anodic polarities, we observedthat distinct alterations are induced by each indepen-dent electrolytic reaction (15–17). In the present work,we evaluate the occurrence of cell death after DC stim-ulation using electrodes of different polarities. Ourresults show that, besides the cellular effects previouslydescribed in other models, toxic compounds producedduring anodic stimulation induce apoptosis and necro-sis in the human leukemic cell line HL60. The inductionof apoptosis in anodic-treated cells was dependent onthe generation of chlorine-derived electrolysis products.In contrast, cathodic treatment caused cell lysis andnecrosis, but not apoptosis. These results demonstratethe relevance of electrode polarity in the generation ofcytotoxic compounds killing cancer cells and help toelucidate the mechanisms involved in the tumor inhibi-tion observed after electric stimulation.
MATERIAL AND METHODS
Cell CultureHuman promyelocytic leukemia HL60 cells were
obtained from the American Type Culture Collection.Cells were grown at 37°C, in 25 cm2 culture flasks con-taining Dulbecco’s modified Eagle’s medium (DMEM)supplemented with 10% fetal bovine serum (FBS). pHcontrol was warranted by the addition of 3 g/L N-(2-hydroxyethyl)-piperazine-N′-(2-ethanesulfonic acid)and 0.2 g/L NaHCO3 to the medium composition, aspreviously described (22). The initial inoculum was 5 ×104 cells/mL, which were subcultured every 2 d andmaintained in a log-phase growth, as described else-where (17,23).
DC StimulationHuman cells were collected by centrifugation,
washed, and suspended (1.0 × 106 cells/mL) in phos-phate-buffered saline (PBS, g/L: 0.20 KH2PO4, 2.17Na2HPO4-7H20, 8.0 NaCl, 0.20 KCl, pH 7.4) at 308mOsm. The cellular suspensions were then distributedin a system of three acrylic chambers with internal vol-umes of 3 cm3, connected in series by filter-paperbridges, and fitted with platinum electrodes in theirextremities. Chamber conformation and distribution ofelectrodes are diagramed in Fig. 1. In this system, cellsuspensions can be separately exposed to cathodic oranodic reactions, and to electric current without contactwith the electrodes, in the intermediary chamber (IC).
Cell suspensions (2 mL) were treated with a DC valueof 2 mA, which was similar to those used in previousstudies (7,16,17,20,24,25). Human cells were stimulatedfor 0 (control), 2, 4, 6, 8, and 10 min using a DC source(KLD Biossistemas Equipamentos Eletrônicos Ltda,Brazil; model ETM 901). The cross-sectional area of thechamber was 0.6 cm2, as determined by multiplying thedepth of the fluid (0.6 cm) by the width of the chamber (1cm). Because the total current passing through the cham-ber was 2 mA, a current density of 3.3 mA/cm2 was gen-erated, associated with a voltage gradient of 378,38 V/m.In all further experiments, cells were treated for 6 min, anintermediate period that was representative of the alter-ations in morphology and viability. Treatments were per-formed at 25°C, and temperature variation wasmonitored with a thermometer. The higher temperatureachieved was 25.4°C, after 10 min of stimulation, indicat-ing that DC treatment did not significantly increase tem-perature levels. Control systems consisted of exposure ofhuman cells to the same chambers exactly as describedpreviously, except for the presence of electric current(Fig. 1B). Alternatively, an additional experiment wasperformed after reduction of the internal volume of theexposure chamber, allowing treatments using 1.0 and 1.5
62 Veiga et al.
Cell Biochemistry and Biophysics Volume 42, 2005

mL cell suspensions. Variations in electric conductivityduring DC stimulation were monitored with a conduc-tivity meter (Microsiemens).
Cell GrowthTo evaluate the influence of DC treatment on the pro-
liferation of leukemic cells, they were suspended in 2mL PBS (106 cells/mL) and treated for 6 min with DC.Aliquots of 400 µL of the cell suspensions were taken,washed in PBS, counted in a Neubauer chamber, andsuspended in 2 mL DMEM supplemented with FBS.The number of viable cells in the different systems wasdetermined as previously described (15–17) and the sus-pensions containing similar cell numbers cultivated in a24-well plate. Aliquots of 25 µL of the cell culture weretaken after 0, 4, 18, and 24 h, and the total number ofcells determined in a Neubauer chamber.
Cell ViabilityCell viability was evaluated by the trypan blue
method, through the determination of the number ofremaining cells in a Neubauer chamber after DC treat-ment, as previously described (16,17,20,27,28). Non-viable cells were divided in two classes, which were (1)the trypan blue-stained cells that retained their shapeand (2) lysed leukemic cells. The number of nonviable,lysed cells was considered as the difference between theinitial number of cells added to the chamber and thenumber of remaining cells.
Influence of the Electrolytic Medium in Cellular Damage
To establish a relationship between the compositionof the electrolytic medium and cellular damage, HL60cells (106/mL) were treated with DC in 2 mL DMEM(without FBS) for 6 min. After stimulation, HL-60 cellswere cultivated for 24 h in a 24-well plate at 37°C in twodifferent situations: (1) at the same medium of treat-ment or (2) in fresh DMEM after removal of the mediumof stimulation by centrifugation. Alternatively, themedium of treatment of stimulated HL60 cells was usedto cultivate untreated cells after the removal of DC-treated populations by centrifugation.
To study the toxic effects of chlorine-derived com-pounds, which are generated in the reaction of electrol-ysis products with amino acids, HL-60 cells were alsotreated with DC in PBS supplemented with glutamine(0.5, 1.0, and 1.5 mM) for further cultivation in DMEMfor 0, 1, and 3 h. The number of viable cells in the dif-ferent systems was determined as previously described(16,17), and in all experimental situations the suspen-sions containing similar cell numbers were cultivatedin a 24-well plate. Aliquots of 25 µL of the cell culturewere then taken after 0, 1, and 3 h, and the total num-ber of cells determined in a Neubauer chamber. Humancells stimulated in these conditions were also screenedfor the occurrence of apoptosis, as described in a fol-lowing section.
Generation of Toxic Compounds Inducing Cell Death 63
Cell Biochemistry and Biophysics Volume 42, 2005
Fig. 1. Graphic representation of the in vitro system used for direct electric current (DC) treatment of HL60 cells (A).The cell suspensions are distributed over each individual chamber (AC for anodic chamber, IC for intermediary chamber,and CC for cathodic chamber). Platinum electrodes are inserted into the lateral chambers (AC and CC), allowing the sys-tem to be connected to a DC source. This distribution permits the separate exposition of cells to cathodic or anodic reac-tions, and to electric current without contact with the electrodes (IC). Control cells are exposed to the same conditions,except for the use of DC. In this case, they are placed into a control cuvette (B), in which they have direct contact with theplatinum electrode and the filter paper bridge. Cell populations treated in this chamber were called nonstimulated cells(NS). Internal volume: 3 cm3.

Flow Cytometry With 7-AminoactinomycinThe progressive increase in membrane permeability
induced by DC treatment was evaluated by 7-aminoactinomycin (7-AAD) incorporation (29,30).Before 7-AAD staining, a cell suspension (2 mL) con-taining 106 cells/mL was treated with DC and washedtwice in PBS. DC-treated and control cells were imme-diately incubated with 7-AAD or, alternatively, reintro-duced into the culture medium and incubated for 0, 4,18, or 24 h, as described previously. Control cells werealso introduced into the treatment chambers and incu-bated as performed for stimulated cells, but withoutany current flow. After these periods, cells were againwashed and incubated in a solution of 7-AAD at 20µg/mL (PBS) for 20 min at 4°C and protected from light.Cells in this staining solution were then analyzed(10,000 events) in a FACS CALIBUR flow cytometer(Becton Dickinson, Franklin Lakes, NJ). The red fluores-cence from 7-AAD was filtered through a 650-long passfilter. Multiparameter data analysis was performed withWinmdi software (Salk Flow Cytometry).
Electron MicroscopyUltrastructural alterations in leukemic cells after DC
treatment were evaluated by transmission electronmicroscopy. Cells were first centrifuged and resus-pended in a 2.5% glutaraldehyde solution. After 2 h ofincubation in this solution, cells were washed in 0.1 Mcacodylate buffer (pH 7.2) containing 0.2 M sucrose andpostfixed in 1% osmium tetroxide for 45 min. After fix-ation, they were preincluded in 1.5% agar, dehydratedwith ethanol and acetone, and embedded in Epon (31).Ultrathin sections were prepared with a diamond knifein an KLB ultramicrotome, collected in 300 mesh coopergrids, counterstained with uranyl acetate and lead cit-rate, and examined under a Zeiss 900 transmission elec-tron microscope operating at 80 kV.
Assay for Morphological ChangesThe morphological features of DC-treated cells were
assessed by staining cytocentrifuged preparations bythe May-Grunwald-Giemsa method. At least 200 cells ineach preparation were examined with a light micro-scope. For analysis of cellular alterations, three separateexperiments were performed.
Analysis of DNA FragmentationHL60 cells were treated with DC as described previ-
ously (DC Stimulation); control systems consisted ofexposure of human cells to the chambers under thesame conditions, except for the presence of electric cur-rent (Fig. 1B). A total of 107 leukemia cells was obtainedin combined experiments. The cells were washed with
PBS and pelleted by centrifugation. DNA fragmentationwas assayed by a modification of the method of Dukeand Sellins groups (32). Cultured cells were washedtwice with ice-cold PBS and suspended in 100 µL lysisbuffer (10 mM Tris HCl/10 mM EDTA/0.5% Triton X-100, pH 8.0), vortex-mixed, sonicated, and incubated onice for 20 min. After centrifugation for 20 min at 4°C(14,000g), the supernatant containing fragmented (solu-ble) DNA was transferred to another tube. Lysis buffer(100 µL) was added to the pellet containing insolubleDNA. Both samples were treated with Rnase A (0.5mg/mL) for 1 h at 37°C and then with proteinase K(Sigma, 0.4 mg/mL) for 1 h at 37°C. After adding 20 µL5 M NaCl and 120 µL isopropanol, the samples wereincubated overnight at –20°C. The soluble fraction ofDNA was determined by electrophoresis on 1.5%agarose gel and has a ladder-like appearance.
RESULTS
Treatment of HL60 Cells With DC Inhibits Proliferation
Treatment with DC has been shown to significantlyinhibit the growth of several tumor cells(2,7–11,14,17,20,24,33–43), which led us to evaluate theinfluence of DC treatment on the proliferation rates ofHL60 human leukemic cells. They were treated with aDC intensity of 2 mA for 6 min, according with previ-ously described models of electric stimulation of ani-mal cell lines (16,17,20). Control cells were exposed tothe same conditions of stimulation, except for the pres-ence of electric current. DC-treated cells were washedto remove products of electrolysis and reinoculated(105) into fresh media; the number of cells in culturewas then determined after 0, 4, 18, and 24 h. Cells thatwere treated with DC in anodic or cathodic chambers,as well as without contact with electrodes, were unableto proliferate in normal rates (Fig. 2).
Kinetics of Trypan Blue Incorporation by DC-Treated Cells
The mechanisms by which DC treatment inhibits thegrowth of tumor cells are poorly known. Because the cellmembrane is commonly affected by the action of DC(16,20,44,45), we evaluated the immediate effects of elec-tric treatment on cell lysis and membrane integrity ofHL60 cells. Cells were treated for periods varying from 0to 10 min and their viability assayed by the trypan bluemethod. Treatment of HL60 cells in the cathodic cham-ber (CC) predominantly induced lysis, which wasgreater in higher periods of treatment (Fig. 3). In con-trast, cell stimulation in the anodic chamber (AC)resulted in a progressive augment of permeability to the
64 Veiga et al.
Cell Biochemistry and Biophysics Volume 42, 2005

Generation of Toxic Compounds Inducing Cell Death 65
Cell Biochemistry and Biophysics Volume 42, 2005
Fig. 2. Treatment of HL60 cells with direct electric current (DC) inhibits proliferation. Cells were treated with DC andinoculated into fresh media. The number of cells was determined after 4, 18, and 24 h. Cells that were treated in an anodicchamber (�), intermediary chamber (�), or cathodic chamber (�) had their growth inhibited. Proliferation rates of cells thatwere not treated with DC (�) are also shown (for details of control preparation, see Material and Methods). Results of fiveindependent experiments expressed as mean ± standard deviation are shown.
Fig. 3. Kinetics of trypan blue incorporation by direct electric current (DC)-treated cell. Cells were treated for periodsvarying from 0 to 10 min and their viability assayed by the trypan blue method. Black bars represent the number of trypanblue-stained cells, whereas white bars are representative of the number of lysed cells. Indices of stained or lysed cells areshown after their treatment in the cathodic (CC), anodic (AC), intermediary (IC), or control (NS) chambers. Results of fiveindependent experiments expressed as mean ± standard deviation are shown.

trypan blue dye, indicating that membrane integrity wasaffected by these conditions of electric stimulation. Celllysis was also observed, although less intensively thanobserved after cathodic treatment. These results areindicative that the cytoplasmic membrane is indeed akey target for electric stimulation in the presence of elec-trodes. Expressive levels of lysis or trypan blue stainingwere not observed when cells were treated in the IC.Exposure of HL60 cells to the chamber of treatment inthe absence of DC resulted in high levels of viability.
To evaluate the concentration of the products of elec-trolysis in viability, HL60 cells were stimulated in thesame exposure conditions, except for the volume of thecell suspension. In this system, the internal volume ofthe exposure chamber was reduced by the addition ofglass slides at the bottom of the chamber, which madepossible the treatment of cell suspensions in the sameconditions but in reduced volumes. This experimentdemonstrated that the lowest viabilities were generatedin smaller volumes (Table 1), indicating that the con-centration of the products of electrolysis directly influ-ences the occurrence of cell death. Cells that weretreated in IC or in the CC presented very similar levelsof viability, which varied from 80% to 95% of viablecells in the different periods and volumes of treatment(data not shown).
Membrane Damage Is Not Reverted by Cultivationof DC-Treated Cells
After the 6-min stimulation, a period of treatment inwhich the most significant alterations in cell numberand viability are observed in this and others systems(16,17,20), cells were reinoculated into fresh media andcultivated for periods of 4, 18, and 24 h. The cells werethen analyzed by flow cytometry, after staining with 7-AAD (29,30). Control cells (Fig. 4), as well as IC- andcathodic-treated populations (not shown), remainedimpermeable to 7-AAD. AC-treated cells, however, pre-
sented an irreversible and progressive augment in theirpermeability to 7-AAD (Fig. 4, R2), which was accom-panied by the generation of cell bodies with a signifi-cantly reduced size (Fig. 4, R3).
Anodic Treatment Induces Necrosis and Apoptosis in HL-60 Cells
The occurrence of 7-AAD–stained cells presentingreduced cell sizes could be indicative of differentprocesses leading to cell death (46). In fact, necroticcells are generated after 24 h of cultivation of anodic-treated cells, as shown in Fig. 5. The cells presentedmembrane discontinuity and matrix rarefaction, asobserved for DC-treated P815 and multidrug-resistantK562 cells (16,17).
Previous studies described that apoptosis is trig-gered in human leukemic cell lines after treatment withelectric current (21). Untreated (control), CC-, and IC-stimulated cells presented similar morphological pro-files (not shown). However, Giemsa staining revealedthe occurrence of morphological changes induced byanodic treatment mainly characterized by cell shrink-age and nuclear chromatin clumping (Fig. 6A). Cellularpresentations suggestive of apoptotic bodies were alsoobserved, mainly when anodic-treated cells were culti-vated for prolonged periods. In fact, biochemical fea-tures of apoptosis were detected after DNA analysis bygel electrophoresis (Fig. 6B). DNA extracted from AC-,but not CC- or IC-stimulated cells, was fragmented intosegments of 180–200 base pairs and presented the typi-cal ladder pattern observed in preparations from apop-totic cells (47).
In anodic reactions performed in the presence ofchloride, hypochlorous acid (HOCl) is generated as aproduct of electrolysis (15). The latter compound canreact with several amino acids to form chloramines,which are potent inducers of apoptosis (48). To evaluateif such products of electrolysis were in fact the inducers
66 Veiga et al.
Cell Biochemistry and Biophysics Volume 42, 2005
Table 1Effect of Chamber Volume on the Viability of HL60 Cells After Different
Periods (min) of DC Treatment
Nonviable Cellsa (%)
Chamber volume 2 min 4 min 6 min 8 min 10 min
1.0 mL 60b 37c 85b 65c 95b 95c 99b 100c 100b 100c
1.5 mL 33b 33c 58b 69c 60b 85c 72b 99c 90b 100c
2.0 mL 33b 37c 53b 65c 57b 65c 58b 83c 85b 94c
aNonviable cells were considered as the addition of the number of lysed and trypan-blue stained cells.bCathodic- and canodic-stimulated cells.

Fig.
4. F
low
cyt
omet
ry o
f dir
ect e
lect
ric
curr
ent (
DC
)-tr
eate
d H
L60
cel
ls a
fter
sta
inin
g w
ith
7-am
inoa
ctin
omyc
in (7
-AA
D).
Cel
ls w
ere
trea
ted
wit
h D
C a
nd in
ocul
ated
into
fre
sh m
edia
. Aft
er c
ulti
vati
on f
or 4
, 18,
and
24
h, c
ells
wer
e ha
rves
ted
and
then
sta
ined
wit
h 7-
AA
D, f
ollo
wed
by
flow
cyt
omet
ry a
naly
sis.
Con
trol
cel
ls re
mai
ned
impe
rmea
ble
to 7
-AA
D, a
s in
ferr
ed b
y th
e hi
gher
con
cent
rati
on o
f cel
ls in
regi
on 1
(R1)
. Ano
dic
-tr
eate
d c
ells
pre
sent
ed a
n ir
reve
rsib
le a
nd p
rogr
essi
ve a
ugm
ent i
n th
eir
perm
eabi
lity
to 7
-AA
D, a
s co
nclu
ded
from
the
incr
ease
d a
mou
nt o
f cel
lsin
reg
ion
2 (R
2). T
hese
eve
nts
wer
e ac
com
pani
ed b
y th
e ge
nera
tion
of
cell
bod
ies
wit
h a
sign
ifica
ntly
red
uced
siz
e, m
ainl
y d
istr
ibut
ed o
ver
regi
on 3
(R
3). D
ata
from
thes
e ex
peri
men
ts a
re r
epre
sent
ativ
e of
one
ana
lysi
s in
five
iden
tica
l ass
ays.
67
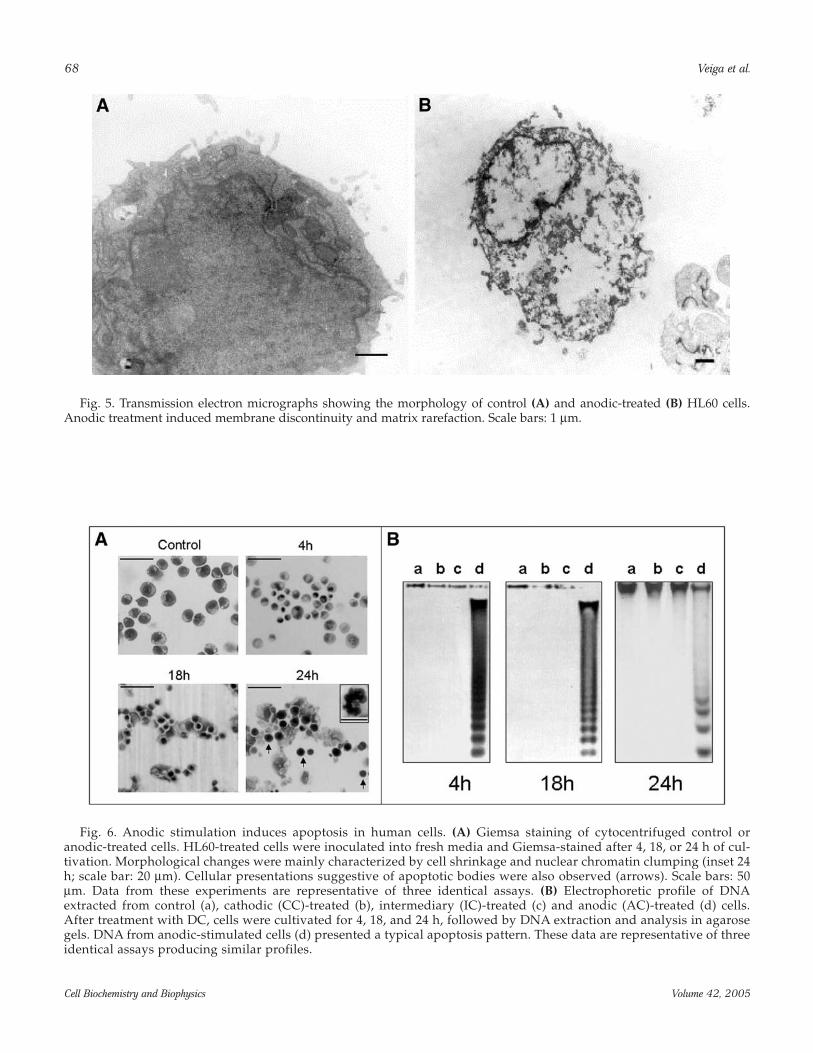
68 Veiga et al.
Cell Biochemistry and Biophysics Volume 42, 2005
Fig. 5. Transmission electron micrographs showing the morphology of control (A) and anodic-treated (B) HL60 cells.Anodic treatment induced membrane discontinuity and matrix rarefaction. Scale bars: 1 µm.
Fig. 6. Anodic stimulation induces apoptosis in human cells. (A) Giemsa staining of cytocentrifuged control oranodic-treated cells. HL60-treated cells were inoculated into fresh media and Giemsa-stained after 4, 18, or 24 h of cul-tivation. Morphological changes were mainly characterized by cell shrinkage and nuclear chromatin clumping (inset 24h; scale bar: 20 µm). Cellular presentations suggestive of apoptotic bodies were also observed (arrows). Scale bars: 50µm. Data from these experiments are representative of three identical assays. (B) Electrophoretic profile of DNAextracted from control (a), cathodic (CC)-treated (b), intermediary (IC)-treated (c) and anodic (AC)-treated (d) cells.After treatment with DC, cells were cultivated for 4, 18, and 24 h, followed by DNA extraction and analysis in agarosegels. DNA from anodic-stimulated cells (d) presented a typical apoptosis pattern. These data are representative of threeidentical assays producing similar profiles.

of cell death after anodic stimulation, the percentage ofnonviable cells was determined by the trypan bluemethod after cultivation of DC-treated cells in differentconditions (Fig. 7). When the cells were cultivated for 24h in the same medium in which they were electricallystimulated, a very low index of viability was observed.In contrast, a lesser number of nonviable cells wasobserved when they were treated with DC and furthercultivated in a fresh medium. In addition, cultivation ofnonstimulated cells in a medium in which HL60 cellshave been previously stimulated resulted in a largepopulation of nonviable cells, suggesting that toxic mol-ecules generated during electric treatment were theactive agents generating cell death.
The influence of chlorine-derived compounds in thegeneration of apoptotic cell death in our system wasevaluated by the anodic treatment of HL-60 cells in PBSsupplemented with glutamine, followed by incubationin culture medium for 1 and 3 h. During DC treatments,no significant variations in electric conductivity wereobserved, as determined in a conductivity meter (datanot shown). In control systems, which were treated inphosphate buffer (identical to PBS except for the pres-ence of sodium chloride), no cellular alterations wereobserved (not shown). In this system, electric conduc-tivity also remained constant during DC treatment inthe different conditions. However, leukemic cells thatwere treated in PBS plus glutamine presented a pro-
gressive augment in trypan blue permeability, indicat-ing loss of viability (Fig. 8A). Exposure of HL60 cells tothe same conditions, except for the presence of DC, didnot affect viability. HL60 cells were also evaluatedmicroscopically for the presence of signs of apoptosis,which were in fact observed. Cell shrinkage and nuclearchromatin clumping, as well as cellular presentationssuggestive of apoptotic bodies, were detected (Fig. 8B).
DISCUSSION
DC can differently influence the growth of manytumoral cell types (49–53). In the present work, wedescribe an inhibitory effect of DC-derived compoundson the proliferation of human leukemic cells. Necrosisand apoptosis were the biologic events associated withcell death in our experimental conditions. In addition,we demonstrate that the mechanism of cell death isdependent on the products of electrolysis and the polar-ity of the electrode, showing for the first time that pro-grammed cell death is induced by toxic compoundsproduced during anodic stimulation.
Electrode polarity and the products generated dur-ing anodic or cathodic stimulations can be hallmarkedas an important issue influencing tumor inhibition(7,8,12,37,51). Morris and coworkers (52) demon-strated that treatment of tumor in mice yields differentresults depending on whether AC or CC stimulation isused. In their model, cell death in the region of contactwith the anodic electrode is not accompanied by celllysis, whereas cathodic-stimulated cells are disruptedin a great extent. In addition, anodic stimulation hasbeen shown to be more effective than the cathodictreatment in the clinical use of DC for the treatment ofcancer (7,8,24,53). Our studies on the in vitro stimula-tion of tumor cells with DC confirm that differentmechanisms are involved in the growth inhibitioncaused by DC (15–17). This and previous studiesdemonstrate that tumor cells can have their growthability impaired even when their morphologies areunaltered, which is a typical feature of IC-treated cells.The mechanisms causing tumor inhibition in theabsence of electrodes are still undefined, but free-float-ing electrons could inactivate ribonucleotide reductase(54), which is responsible by the conversion of build-ing blocks of RNA into DNA during cell division.However, evident cellular alterations are observed incathodic- and anodic-stimulated cells (13–17,37),which confirms that different mechanisms should beinvolved in DC-induced tumor inhibition and, addi-tionally, that they are influenced by the products gen-erated by electrodes with different polarity.
In the present work, an extensive index of cell lysiswas observed when the cells were treated in the cathodic
Generation of Toxic Compounds Inducing Cell Death 69
Cell Biochemistry and Biophysics Volume 42, 2005
Fig. 7. Death of HL60 cells after anodic stimulation isinduced by the products of electrolysis. The percentage ofnonviable cells was determined after cultivation of directelectric current-stimulated cells for 24 h in the samemedium of electric treatment (A) or in fresh medium (B).Alternatively, the cells were removed by centrifugationand the medium of treatment was used to cultivate non-stimulated cells (C). The viability of nonstimulated cellscultivated in fresh medium for 24 h is shown in (D).

chamber, which was accompanied by a progressiveincrease of pH in the medium used for electric treatment.Lysis of cathodic-treated cells in PBS may be explainedby a direct attack of the oxidants generated during DCtreatment on the cell membrane, as previously described(13,55). Superoxide, hydroxyl groups, molecular hydro-gen, and oxygen are major products of cathodic electro-chemical reactions occurring in aqueous solution (14,55).The production of hydroxyl groups in these reactions isprobably a major contributing factor in the pH increase,which, in turn, causes cell lysis (37). Superoxide radicalscan also influence the occurrence of cell lysis, becausethey are generated by cathodic reactions (55) and are effi-cient agents in the destruction of target cells (56,57).
Electrolysis products actually seem to be involved incell death in our experimental conditions, because theirincreased concentrations resulted in higher levels of celldeath (Table 1). Microscopic analysis of Giemsa-stainedcells revealed that, besides the usual morphologic fea-tures of DC-inhibited cells, an alternative mechanism ofdeath was detected. Although leukemic cells treated inthe presence of the cathode or in the absence of elec-trodes presented morphologies that were similar to con-trol systems, several cells stimulated in the presence ofthe anode resembled apoptotic bodies. This observationwas confirmed by DNA electrophoresis and indicatedthat apoptosis is in fact an alternative pathway of celldeath in leukemic cells exposed to DC. The induction of
70 Veiga et al.
Cell Biochemistry and Biophysics Volume 42, 2005
Fig. 8. Stimulation of HL60 cells in the presence of glutamine. (A) Human cells were suspended in phosphate-bufferedsaline supplemented with 0.5 mM (squares), 1.0 (circles), or 1.5 (triangles) glutamine and treated with direct electric current(DC) (open symbols). Cell viability was measured immediately after electric stimulation (0 h) or after incubation in freshmedium supplemented with fetal bovine serum for 1 or 3 h. The viability of HL60 cells incubated in the presence of gluta-mine is also shown (closed symbols). (B) Nonstimulated or DC-treated cells that were incubated in the presence of gluta-mine (0.5 mM) for 3 h were also analyzed microscopically after Giemsa staining. Control cells presented their typicalmorphology, whereas DC-stimulated cells resembled apoptotic bodies. Scale bars: 10 µm.

apoptosis by electric and magnetic fields has beendescribed in other systems (21,58–60), although this isthe first report demonstrating that toxic compoundswhose generation is dependent on electrode polarityregulate the induction of this biologic process. UsingDC as the source of electric stimulation, Kurokawa andcoworkers (21) described that treatment of humanleukemic cell lines had undergone apoptotic cell death,but, in their model, cells were simultaneously treatedwith anodic and cathodic polarities, which makes ananalysis of the products of electrolysis and their influ-ence on the generation of cellular damage difficult.
The occurrence of apoptosis in cells exposed to DCwas apparently derived from electrolysis products fromanodic reactions. Cisplatinum is a well-known DNA-damaging agent and its presence in a DC-stimulatedmedium is expected around the anodic region (61), butnot in the cathode compartment. Therefore, it is possiblethat the generation of cisplatinum complexes con-tributes to the induction of apoptosis in the anodicchamber. However, experiments using steel electrodesinstead of platinum probes demonstrated that apoptosisstill occurs, which indicates that cisplatinum complexesare not the major inducers of cell death in our system(Holandino et al., unpublished data). We thereforeinvestigated the influence of chlorine species producedduring anodic stimulations possibly involved in theinduction of apoptosis.
HOCl, which is generated during electric stimulationof aqueous solutions (13–15), is known to modify mem-brane lipids (62–64) as well as protein and DNA. Itsreaction with extracellular amino acids results in thegeneration of chloramines, which are less potent oxi-dants (65–67). The ability of these reactive oxygenspecies in the generation of apoptosis has been recentlydescribed (48,68).
Most studies of HOCl-induced cytotoxicity havebeen carried out on cells incubated in buffer solutionsinstead of cell culture media (67–69). Although themechanism of cell death was not specifically deter-mined in these studies, results were generally reflectiveof necrosis. However, additional studies revealed thattreatment of endothelial cells with HOCl for 15 min inbuffered saline followed by transfer of cells to completemedia resulted in the occurrence of apoptotic cell deathin approx 20% of the population (70). Necrosis was themajor mechanism involved in cell death in the presenceof higher levels of HOCl, which is consistent with ourresults. In our model, leukemic cells were initiallytreated in a buffered system, which means a low avail-ability of amino acids and a consequent low concentra-tion of chloramines. If this hypothesis is true, necrosisand, in lower levels, apoptosis should occur after treat-ment of cells with DC, which is indeed observed in our
experiments. However, treatment of HL-60 cells in PBSsupplemented with glutamine resulted in a predomi-nant occurrence of apoptosis, in accordance with thegeneration of chloramines derived from the reactionbetween HOCl and glutamine. When chloride ions wereremoved from the medium of treatment, no cellulardamage occurred (data not shown). This result could beexplained by the facilitated generation of chloraminesfrom the reaction between the amino acid and HOCl,creating an environment more favorable to apoptoticthan necrotic cell death. In this context, the critical taskwill be to identify the specific molecular targets that ini-tiate the apoptotic cascade.
The influence of DC and its soluble products in thebiology of animal cells and microorganisms is alargely recognized phenomenon (1,3,7,12,24,25,45).The observation of different cellular DC-inducedeffects contributes with the comprehension of severalphysiologic events, and also raises the possibility ofthe use of electric stimulation in the treatment of can-cer (33–35,37,41–43,52,53,71), wound healing (4,72,73),and fracture repair (74). However, the biologic mecha-nisms whereby DC and their derived compoundsinhibit or stimulate cell growth still require clarifica-tion. In this context, the elucidation of the cellularevents triggered after electric stimulation under con-trolled conditions (e.g., electrical field strength, periodof treatment, electrode polarity, and toxic compoundsgenerated during stimulation) will support the use ofin vivo or in vitro models applying DC as a cellgrowth–interfering agent.
ACKNOWLEDGMENTS
We thank Dr. Geraldo A. G. Cidade for helping us inthe physical determinations after DC treatment in theacrylic chamber and Dr. Damijan Miklavcic for helpfuldiscussions. This work was supported by Financiadorade Estudos e Projetos (FINEP), Conselho Nacional deDesenvolvimento Científico e Tecnológico (CNPq),Fundaç~ao de Amparo a Pesquisa no Estado do Rio deJaneiro (FAPERJ), Fundaç~ao José Bonifácio (FUJB), andPrograma de Apoio a Núcleos de Excelência (PRONEX).
REFERENCES
1. Bolton, L., Foleno, B., Means, B., and Petrucelli, S. (1980)Direct-current bactericidal effect on intact skin. Antimicrob.Agents Chemother. 18, 137–141.
2. Chou, C. K., Mc Dougall, J. A, Ahn, C., and Vora, N. (1997)Electrochemical treatment of mouse and rat fibrosarcomaswith direct current. Bioelectromagnetics 18, 14–24.
3. Chu, C. S., McManus, A. T., Pruitt, B. A. Jr., and Mason, A.D. (1988) Therapeutic effects of silver nylon dressings with
Generation of Toxic Compounds Inducing Cell Death 71
Cell Biochemistry and Biophysics Volume 42, 2005

weak direct current on Pseudomonas aeruginosa-infectedburn wounds. J. Trauma 28, 1488–1492.
4. Lee, C. R., Canaday, D. J., and Doong, H. (1993) A review ofthe biophysical basis for the clinical application of electricfields in soft-tissue repair. J. Burn Care Rehabil. 14, 319–335.
5. Lyte, M., Gannon, J. E., and O’Clock, G. D. Jr. (1991) Effectsof in vivo electrical stimulation on enhancement and sup-pression of malignant lymphoma cell proliferation. J. Natl.Cancer Inst. 83, 116–119.
6. Miklavcic, D., Jarm, T., Cemazar, M., Sersa, G., An, D. J.,Belehradek, J. Jr., et al. (1997) Tumor treatment by directelectric current. Tumor perfusion changes. Bioelectrochem.Bioenergetics 43, 253–256.
7. David, S. L., Absolom, D. R., Smith, C. R., Gams, J., andHerbert, M. A. (1985) Effect of low level direct current on invivo tumor growth in hamsters. Cancer Res. 45, 5625–5631.
8. Griffin, D. T., Dodd, N. J., Moore, J. V., Pullan, B. R., andTaylor, T. V. (1994) The effects of low-level direct-currenttherapy on a preclinical mammary carcinoma: tumorregression and systemic biochemical sequel. Br. J. Cancer69, 875–878.
9. Xin, Y., Xue, F., and Zhao, F. (1997). Effectiveness of elec-trochemical therapy in the treatment of lung cancers ofmiddle and late stage. Chin. Med. J. 110, 379–383.
10. Xin, Y. L. (1993) Traditional and Western medical treat-ment of 211 cases of late stage lung cancer. Chung KuChung Hsi I Chieh Ho Tsa Chih 13, 135–113.
11. Xin, Y., Xue, F-Z., Ge, B-S., Zhao, F-R., Shi, B., and Zhang,W. (1997) Electrochemical treatment of lung cancer.Bioelectromagnetics 18, 8–13.
12. Li, K., Xin, Y., Gu, Y., Xu, B., Fan, D., and Ni, B. (1997) Effectsof direct current on dog liver: possible mechanisms fortumor electrochemical treatment. Bioelectromagnetics 18, 2–7.
13. Samuelsson, L. and Jönsson, L. (1980) Electrolyte destruc-tion of lung tissue. Electrochemical aspects. Acta Radiol.Diagnosis 21, 711–714.
14. Beredson, J. and Simonsson, D. (1994) Electrochemicalaspects of treatment of tissue with direct current. Eur. J.Surg. Suppl. 574, 111–115.
15. Veiga, V. F. (1996) Master’s thesis, Federal University ofRio de Janeiro, Rio de Janeiro, Brazil.
16. Veiga, V. F., Holandino, C., Rodrigues, M. L., Capella, M. A.M., Menezes, S., and Alviano, C. S. (2000) Cellular damageand altered carbohydrate expression in P815 tumor cellsinduced by direct electric current: an in vitro analysis.Bioelectromagnetics 21, 597–607.
17. Holandino C., Veiga, V. F., Rodrigues, M. L., Morales, M.M., Capella, M. A. M., and Alviano, C. S. (2001) Direct cur-rent decreases cell viability but not P-glycoprotein expres-sion and function in human multidrug resistant leukaemiccell. Biolectromagnetics 22, 470–478.
18. Sauer, H. Stanelle, R., Hescheler, J., and Wartenberg, M.(2002) The DC electrical-field-induced Ca+2 response andgrowth stimulation of multicellular tumor spheroids aremediated by ATP release and purinergic receptor stimula-tion. J. Cell Sci. 115, 3265–3273.
19. Wartenberg, M., Hescheler, J., and Sauer, H. (1997)Electrical fields enhance growth of cancer spheroids by
reactive oxygen species and intracellular Ca+2. Am. J.Physiol. 272, R1677–R1683.
20. Holandino, C. Q., Veiga, V. F., Capella, M. A. M., Menezes,S., and Alviano, C. S. (2000) Damage induction by directelectric current in tumoral target cells (P815). Ind. J. Exp.Biol. 38, 554–558.
21. Kurokawa, M., Sakagami, H., Kokubu, F., Noda, H.,Takeda, M., and Adachi, M. (1997) Induction of apoptoticcell death by direct-current treatment in human leukemiccell lines. J. Cancer Res. Clin. Oncol. 123, 370–376.
22. Freshney, R. I. (1994) Culture of Animal Cells: A Manual ofBasic Technique, Wiley-Liss, Inc., New York.
23. Trindade, G. S., Farias, S. L. A., Rumjanek, V. M., andCapella, M. A. M. (2000) Methylene blue reverts mul-tidrug resistance: sensitivity of multidrug resistant cells tothis dye and its photodynamic action. Cancer Lett. 151,161–167.
24. Schauble, M. K., Habal, M. B., and Gullick, H. D. (1977)Inhibition of experimental tumour growth in hamsters bysmall direct currents. Arch. Pathol. Lab. Med. 101, 249–297.
25. Marino, A. A., Morris, D., and Arnold, T. (1986) Electricaltreatment of Lewis lung carcinoma in mice. J. Surg. Res. 41,198–201.
26. Patterson, M. K. Jr. (1979) Measurement of growth andviability of cells in culture, in Methods in Enzymology(Jakoby, W. B., Pastan, I. H., ed.), Academic Press, NewYork, pp. 141–152.
27. Trindade, G. S., Capella, M. A. M., Capella, L. S., Affonso-Mitidieri, O. R., and Rumjanek, V. M. (1999) Differences insensitivity to UVC, UVB and UVA radiation of a mul-tidrug-resistant cell line overexpressing P-glycoprotein.Photochem. Photobiol. 69, 694–699.
28. Capella, L. S., Alcantara, J. S., Moura-Neto, V., Lopes, A. G.,and Capella, M. A. M. (2000) Vanadate is toxic to adherent-growing multidrug resistant cells. Tumor Biol. 21, 54–62.
29. Schmid, I., Uittenbogaart, C. H., Keld, B., and Giorgi, J. V.(1994) A rapid method for measuring apoptosis and dual-color immunofluorescence by single laser flow cytometry.J. Immunol. Methods 170, 145–157.
30. Lecouer, H., Ledru, E., Prévos, M. C., and Gougeon, M. L.(1997) Strategies for phenotyping apoptotic peripheralhuman lymphocytes comparing ISNT, annexin-V and 7-AAD cytofluorimetric staining methods. J. Immunol.Methods 209, 111–123.
31. Lewis, P. R. and Knight, D. P. (1991) Cytological stainingmethods in electron microscopy. In Staining Methods forSectioned Material. (Glauert A. M., ed.), North-hollandPublishing Company, Amsterdam, pp. 2–47.
32. Duke, R. C. and Sellins, C. B. (1989) Cellular Basis ofImmune Modulation. (Kaplan, J. G. ed.) New York.
33. Yi-Hong, L., Ting-Giu, G., Xiang-Liang, Z., Jian-Zhe, Z.,Ya-Wei, H., Shu-Ming, M., et al. (1994) Electrochemicaltherapy for intermediate and advance liver cancer: areport of 50 cases. Eur. J. Suppl. 574, 51–53.
34. Berry, D. P., Dennison A. R., Ward, R., and Maddern, G. J.(2000) Electrolytic ablation of colorectal liver metastases:1-year histological patient follow-up. Dig. Surg. 17,518–519.
72 Veiga et al.
Cell Biochemistry and Biophysics Volume 42, 2005

35. Wemyss-Holden, S. A., Hall, P. M., Robertson, G. S. M.,Dennison, A. R., Vanderzon, P. S., and Maddern, G. J.(2000) A new treatment for unresectable liver tumours:long-term studies of electrolytic lesions in the pig liver.Clin. Sci. 98, 561–567.
36. Humphrey, C. E. and Seal, E. H. (1959) Biophysicalapproach toward tumor regression in mice. Science 130,388–389.
37. Turler, A., Schaefer, H., Schaefer, N., Wagner, M., Maintz,D., Qiao, J. C., et al. (2000) Experimental low-level directcurrent therapy in liver metastases: influence of polarityand current dose. Bioelectromagnetics 21, 395–401.
38. Nordeström, B. W. E. (1994) Electrostatic field interferencewith cellular and tissue function, leading to dissolution ofmetastases that enhances the effect of chemotherapy. Eur.J. Surg. Suppl. 574, 93–109.
39. Jaroszeski, M. J., Gilbert, R. A., and Heller, R. (1997) Invivo antitumor effects of electrochemotherapy in ahepatoma model. Biochim. Biophys. Acta 1334, 15–18.
40. Lie-Cai, S., Cun-Yn, L., Bao-Ping, Z., Tao, W., Yun-Qing, S.,and Wa-Wei, L. (1994) Electrochemical therapy (ECT) forthyroid adenoma during acupuncture anaesthesia: analy-sis of 46 patients. Eur. J. Surg. Suppl. 574, 79–81.
41. Wang, H. L (1994) Electrochemical therapy of 74 cases ofliver cancer. Eur. J. Surg. Suppl. 574, 55–58.
42. Fosh, B. G., Finch, J. G., Anthony, A. A., Texler, M., andMaddern, G. J. (2001) Electrolytic ablation of the rat pan-creas: a feasibility trial. BMC Gastroenterol. 1, 1–11.
43. Teague, B. D., Wemyss-Holden, S. A., Fosh, B. G.,Dennison A. R., and Maddern, G. J. (2002) Electrolysis andother local ablative treatments for non-resectable colorec-tal liver metastases. ANZ J. Surg. 72, 137–141.
44. Capella, M. A. M., Fonseca, M. F., and Menezes, S. (1991)Synergism between electricity and ionizing radiation. J.Photochem. Photobiol. 8, 371–383.
45. Holandino, C. Q., Capella, M. A. M., Angluster, J., Silva-Filho, F. C., Menezes, S., and Alviano, C. S. (1998) Cell sur-face alterations induced by methylene blue and directelectric current in Escherichia coli. Ind. J. Biochem. Biophys.35, 284–290.
46. Schmid, I., Hausner, M. A., Cole, S. W., Uittenbogaart, C.H., Giorgi, J. V., and Jamieson, B. D. (2001) Simultaneousflow cytometric measurement of viability and lymphocytesubset proliferation. J. Immunol. Methods 247, 175–186.
47. McDonald, E. R. and El-Deiry, W. S. (2000) Cell cycle con-trol as a basis for cancer drug development (review). Int. J.Oncol. 16, 871–886.
48. Englert, R. P. and Shacter, E. (2002) Distinct modes of celldeath induced by different reactive oxygen species. J. Biol.Chem. 277, 20518–20526.
49. Chudomel, V., Soucek, J., Hruba, A., Jerabek, J., Schwarz,J., and Smetana, K. (1989) Positive effect of direct currenton cytotoxicity of human lymphocytes. Neoplasma 36,573–582.
50. Batista, U., Miklavcic, D., and Sersa, G. (1994) Low leveldirect current – cell culture fibroblast model. Bioelectroch.Bioener. 35, 99–101.
51. Yen, Y., Li, J. R., Zou, B. S., Rojas, F., Yu, J., and Chou, C. K.(1999) Electrochemical treatment of human KB cells invitro. Bioelectromagnetics 20, 34–41.
52. Morris, D. M., Marino, A. A., and Gonzales, E. (1992)Electrochemical modification of tumor growth in mice. J.Surg. Res. 5, 306–309.
53. Taylor, T. V., Engler, P., Pullan, B. R., and Holt, S. (1994)Ablation of neoplasia by direct current. Br. J. Cancer 70,342–345.
54. Kulsh, J. (1997) Targeting a key enzyme in cell growth: anovel therapy for cancer. Med. Hypoth. 49, 297–300.
55. Forman, H.J. and Fridovich, I. (1972) Electrolytic univalentreduction of oxygen in aqueous solution demonstratedwith superoxide dismutase. Science 175, 339.
56. Saran, M., Miche, C., and Bors, W. (1998) Radical functionsin vivo: a critical review of current concepts and hypothe-ses. Z. Naturforsch. 53, 210–227.
57. Zhou, M. J. and Petty, H. R. (1993) Superoxide-mediatedlysis of erythrocytes: the role of colloid-osmotic forces. J.Cell. Physiol. 157, 555–561.
58. Hisamitsu, T., Narita, K., Kasahara, T., Seto, A., Yu, Y., andAsano, K. J. (1997) Induction of apoptosis in humanleukemic cells by magnetic fields. Japan J. Physol. 47,307–310.
59. Tofani, S., Barone, D., Cintorino, M., Santi, M. M., Ferrar,A., Orlassino, R., et al. (2001) Static and ELF magneticfields induce tumor growth inhibition and apoptosis.Bioelectromagnetics 22, 419–428.
60. Narita, K., Hankawa, K., Kasahara, T., Hisamitsu, T., andAsano, K. (1997) Induction of apoptotic cell death inhuman leukemic cell line, HL-60, by extremely low fre-quency electric magnetic fields: analysis of the possiblemechanisms in vitro. In Vivo 11, 329–336.
61. Miklavcic, D., Sersa, G., Kryzanovski, M., Novakovi, S.,Bobanovi, F., Golouh, R., et al. (1993) Tumor treatment bydirect electric current–tumor temperature and pH.Bioelectrochem. Bioenerg. 30, 209–220.
62. van den Berg, J. J. Winterbourn, C. C., and Kuypers, F. A.(1993) Hypochlorous acid-mediated modification of cho-lesterol and phospholipid: analysis of reaction productsby gas chromatography-mass spectrometry. J. Lip. Res. 34,2005–2012.
63. Heinecke, J. W., Li, W., Mueller, D. M., Bohrer, A., andTurk, J. (1994) Cholesterol chlorohydrin synthesis by themyeloperoxidase-hydrogen peroxide-chloride system:potential markers for lipoproteins oxidatively damagedby phagocytes. Biochemistry 33, 10127–10136.
64. Carr, A. C., van den Berg, J. J. M., and Winterbounr, C. C.(1996) Chlorination of cholesterol in cell membranes byhypochlorous acid. Arch. Biochem. Biophys. 332, 63–69.
65. Zgliczynski, J. M., Stelmaszynska, T., Ostrowski, W.,Naskalski, J., and Sznajd, J. (1968) Myeloperoxidase ofhuman leukaemic leucocytes. Oxidation of amino acids inthe presence of hydrogen peroxide. Eur. J. Biochem. 4,540–547.
66. Test, S. T., Lampert, M. B., Ossana, P. J., Thoene, J. G., andWeiss, S. J. (1989) Generation of nitrogen-chlorine oxidantsby human phagocytes. J. Clin. Invest. 74, 1341–134.
Generation of Toxic Compounds Inducing Cell Death 73
Cell Biochemistry and Biophysics Volume 42, 2005

67. Thomas, E. L., Grisham, M. B., Melton, D. F., and Jefferson,M. M. (1985) Evidence for a role of taurine in the in vitrooxidative toxicity of neutrophils toward erythrocytes. J.Biol. Chem. 260, 3321–3329.
68. Wagner, B. A., Britigan, B. E., Reszka, K. J., McCormick, M.L., and Burns, C. P. (2002) Hydrogen peroxide-inducedapoptosis of HL-60 human leukemia cells is mediated bythe oxidants hypochlorous acid and chloramines. Arch.Biochem. Biophys. 401, 223–234.
69. Vile, G. F., Rothwell, L. A., and Kettle, A. J. (2000) Initiationof rapid, P53-dependent growth arrest in cultured humanskin fibroblasts by reactive chlorine species. Arch. Biochem.Biophys. 377, 122–128.
70. Vissers, M. C., Pullar, J. M., and Hampton, M. B. (1999)Hypochlorous acid causes caspase activation and apopto-sis or growth arrest in human endothelial cells. Biochem. J.344, 443–449.
71. Xin, Y. L. (1994) Organisation and spread of electrochemi-cal therapy (ECT) in China. Eur. J. Surg. Suppl. 577, 25–30.
72. Karba, R. Semrov, D. Vodovnik, L., Benko, H., andSavrin, R. (1997) DC electrical stimulation for chronicwound healing enhancement. Part 1. Clinical study anddetermination of electrical field distribution in thenumerical wound model. Bioelectrochem. Bioenergetics 43,265–270.
73. Baker, L. L., DeMuth, S. K., Chambers, R., and Villar, F.(1997) Effects of electrical stimulation on wound healingin patients with diabetic ulcers. Diabetes Care 20,405–412.
74. Brighton, C. T., Friedenberg, Z. B., Mitchell, E. I., andBooth, R. E. (1977). Treatment of non-union with constantdirect current. Clin. Orthop. Relat. Res. 124, 106–123.
74 Veiga et al.
Cell Biochemistry and Biophysics Volume 42, 2005




















