Syntactic comprehension deficits are associated with MRI white matter alterations in dementia
Upload
khangminh22Category
view
0download
0
EXPLORING THE ROLE OF WHITE
MATTER DISCONNECTION IN THE
HETEROGENEITY OF PARKINSON'S
DISEASE
A thesis submitted in fulfilment of the requirements for the degree of
Doctor of Philosophy
by
Julie Mae Hall
2018
School of Social Sciences and Psychology
Western Sydney University

ii
Primary Supervisor Doctor Ahmed A Moustafa
Western Sydney University
Auxiliary Supervisors Professor Simon J G Lewis
The University of Sydney
Professor Peter Keller
Western Sydney University

iii
Voor papa
I did it! Ongeloveloos! I wish we could celebrate this milestone, talk about science
and all of the other beautiful things you showed me in life.

iv
Acknowledgements
Firstly, I want to thank the people affected by Parkinson’s disease who
generously volunteered their time to participate in our research. I hope this
project will ultimately contribute to a real and positive difference in their lives.
This thesis could not exist without the help and support of numerous people. I
would like to thank my supervisors, Professor Simon Lewis and Dr Ahmed
Moustafa for consistently providing valuable feedback to this thesis and
introducing me to many interesting questions. Doc, I’m grateful for your
guidance throughout my PhD as well as your support beyond your duties as a
supervisor. It has been an honour to work in the Parkinson’s Disease Research
Clinic. Ahmed, thank you for giving me the freedom to explore my interest and
your positivity during my candidature.
The PD team, my family away from home that has made this journey so
memorable. Mac, you have been the most influential person in my academic
career so far. Thank you for your guidance throughout the years, explaining the
most abstract things with analogies that are easy to grasp. I don’t think I’ve ever
met someone with such a creative mind. Claire, thank you for being such a patient
and passionate teacher, always offering new clever perspectives. With your
brightness and ever-present banter you know how to make science fun. Kaylena,
you’ve been very influential in my PhD, always enthusiastic and open to provide
valuable feedback. Even more important, your kindness, our passionate
discussions and fun outings have made my time here absolutely amazing. Alana,
you’ve taught me many important life lessons and you never fail to make me
laugh. Moran, thank you for the fun times and encouraging me to go to Sydney in
the first place! Deb, Jennifer, Matthew, Elie, Paul, Joseph, and all the PD team
members, thank you for all your feedback and ideas, but most of all, for creating
such a supportive and enjoyable atmosphere. I’d like to thank my friends at the
Brain and Mind Centre, with a special mention to Anke, Maz, Stacey, Django,
Nathan, the running group, and Cunning Stunts for the fun activities, needed
caffeine breaks or simply cheering me on along the way. Courtney, thank you for
everything, my world is brighter with you in it. You make me cry of laughter and

v
are my rock when I need it most. Our connection is so strong that I know that
even when I am on the other side of the planet, we will still be us.
Lastly, I’d like to thank my family. Mama, you have been incredibly supportive
about my choices in life. You and dad not only made sure I got plenty
opportunities and freedom to explore the world, but also taught me the
importance of education, to be resilient and independent. Thank you for
continuously encouraging me. Ruby, we are really close as sisters. I have missed
not having you around. You are caring and always take the time to listen to me.
You and Jochem are the most generous people I know, nothing is too much
trouble. Jochem and little Morris, the true golden boy, thank you for all your
support, love and laughter.

vi
Statement of Authentication
The work presented in this thesis is, to the best of my knowledge, original and
contains no copy or paraphrase of work published by another person, accept
where duly acknowledged in the text. I hereby declare that I have not submitted
this material, either in full or in part, for a degree at this or any other institution.
Julie Mae Hall
Signature
Date
22 Aug 2018

vii
Table of Contents ACKNOWLEDGEMENTS............................................................................................................................................IVSTATEMENTOFAUTHENTICATION.......................................................................................................................VILISTOFTABLES.........................................................................................................................................................IXLISTOFFIGURES........................................................................................................................................................XBOXINDEX.................................................................................................................................................................XIPUBLICATIONSPRESENTEDFOREXAMINATION...............................................................................................XIIABBREVIATIONS..................................................................................................................................................XVIIIABSTRACT..................................................................................................................................................................1CHAPTER1:INTRODUCTION...................................................................................................................................41.1IntroductiontoParkinson’sdisease...........................................................................................................41.2Whitematteranddiffusiontensorimaging...........................................................................................81.3DiffusiontensorimaginginParkinson’sdisease...............................................................................111.4FreezingofgaitinParkinson’sdisease.................................................................................................121.5VisualhallucinationsinParkinson’sdisease......................................................................................161.6Parkinson’sdiseaseasadisconnectiondisorder...............................................................................191.7Aimsofthethesis.............................................................................................................................................22References..................................................................................................................................................................24CHAPTER2:DIFFUSIONALTERATIONSASSOCIATEDWITHPARKINSON’SDISEASESYMPTOMATOLOGY:AREVIEWOFTHELITERATURE...............................................................................................................................39Abstract......................................................................................................................................................................39Introduction..............................................................................................................................................................40SearchStrategy.......................................................................................................................................................42Results.........................................................................................................................................................................43Conclusionsandfuturedirections...................................................................................................................66References..................................................................................................................................................................70CHAPTER3:FREEZINGOFGAITANDITSASSOCIATIONSINTHEEARLYANDADVANCEDCLINICALMOTORSTAGESOFPARKINSON’SDISEASE:ACROSS-SECTIONALSTUDY...................................................................84Abstract......................................................................................................................................................................84Introduction..............................................................................................................................................................85Methods......................................................................................................................................................................86Results.........................................................................................................................................................................89Discussion..................................................................................................................................................................96Referrences.............................................................................................................................................................101CHAPTER4:ALTERATIONSINWHITEMATTERNETWORKTOPOLOGYCONTRIBUTETOFREEZINGOFGAITINPARKINSON’SDISEASE..........................................................................................................................111Abstract...................................................................................................................................................................111Introduction...........................................................................................................................................................112Materialsandmethods.....................................................................................................................................113Results......................................................................................................................................................................120Discussion...............................................................................................................................................................123References...............................................................................................................................................................132CHAPTER5:DYSFUNCTIONINATTENTIONALPROCESSINGINPATIENTSWITHPARKINSON’SDISEASEANDVISUALHALLUCINATIONS...........................................................................................................................141Abstract...................................................................................................................................................................141Introduction...........................................................................................................................................................142Methods...................................................................................................................................................................143Results......................................................................................................................................................................145

viii
Discussion...............................................................................................................................................................147References...............................................................................................................................................................149CHAPTER6:CHANGESINSTRUCTURALNETWORKTOPOLOGYCORRELATEWITHSEVERITYOFHALLUCINATORYBEHAVIOURINPARKINSON’SDISEASE..............................................................................153Abstract...................................................................................................................................................................153Introduction...........................................................................................................................................................154Methods...................................................................................................................................................................156Results......................................................................................................................................................................162Discussion...............................................................................................................................................................168References...............................................................................................................................................................1757.GENERALDISCUSSION.....................................................................................................................................1887.1Summaryofkeyfindings..........................................................................................................................1887.2Limitationsofthecurrentwork............................................................................................................1977.3Futuredirections..........................................................................................................................................2027.4Concludingremarks....................................................................................................................................2097.5References.......................................................................................................................................................211APPENDICES........................................................................................................................................222

ix
List of tables
TABLE2-1:ANOVERVIEWOFTHEDTILITERATUREINPARKINSON’SDISEASE.......................................50
TABLE3-1:CHARACTERISTICSOFPARKINSON’SDISEASEPATIENTSWITHANDWITHOUTFREEZINGOFGAIT(FOG)INTHEEARLYANDADVANCEDSTAGES........................................................................................92
TABLE3-2:EVALUATIONOFCLINICALCHARACTERISTICSOFPARKINSON’SDISEASEPATIENTSWITHANDWITHOUTFREEZINGOFGAIT(FOG)INTHEEARLYANDADVANCEDSTAGES,ILLUSTRATINGTHESTATISTICALDIFFERENCESBETWEENTHEFOURGROUPS..............................................................................94
TABLE4-1:DEMOGRAPHICANDCLINICALDATAOFPATIENTSWITHANDWITHOUTFREEZINGOFGAIT.................................................................................................................................................................................121
TABLE4-2:NODESWITHSIGNIFICANTLYDIFFERENTPARTICIPATIONCOEFFICIENTSBETWEENGROUPS.................................................................................................................................................................................124
TABLE4-3:NODESWITHSIGNIFICANTLYDIFFERENTMODULEDEGREEZ-SCORESCOEFFICIENTSBETWEENGROUPS................................................................................................................................................125
SupplementaryTable4-1:Pearsoncorrelationcoefficient(p-value)betweendifferent............thresholdsacrossdifferencescoresbetweenfreezersandnon-freezers......................................139
TABLE5-1:DEMOGRAPHICALDATAANDCLINICALCHARACTERISTICS...................................................144TABLE5-2:ACCURACYPERCENTAGESFOREACHCUEANDFLANKERCONDITIONANDREACTIONTIMESOFTHEATTENTIONALNETWORKS(136)......................................................................................................146
TABLE6-1:DEMOGRAPHICSANDCLINICALVARIABLES...............................................................................163TABLE6-2:SPEARMAN’SRHOCORRELATIONBETWEENTHEPARTICIPATIONCOEFFICIENTANDTHEHSS........................................................................................................................................................................166
TABLE6-3:SPEARMAN’SRHOCORRELATIONBETWEENTHEMODULEDEGREEZ-SCOREANDTHEHSS.................................................................................................................................................................................168
Supplementary Table 6-1:Nodesofthediverseclub..........................................................184

x
List of figures
FIGURE1-1:SIMPLIFIEDREPRESENTATIONOFTHEBASALGANGLIA-THALAMOCORTICALCIRCUITINA)HEALTHYINDIVIDUALSANDB)PARKINSON’SDISEASEPATIENTS...................................................................5
FIGURE1-2:SCHEMATICREPRESENTATIONOFDIFFUSIONDISPLACEMENT.THEDIRECTIONOFGREATESTDIFFUSIVITY(Λ1)ISASSUMEDTOBEPARALLELTOTHELOCALDIRECTIONOFWHITEMATTER....................................................................................................................................................................10
FIGURE2-1:FLOWCHARTOFSEARCHSTRATEGY............................................................................................43
FIGURE3-1:THEPERCENTAGEOFFREEZERSANDNON-FREEZERSIDENTIFIEDWITHTHEFREEZINGOFGAITQUESTIONNAIREITEM3ACROSSDIFFERENTHOEHNANDYAHRSTAGES(MEASUREDINTHEON-STATE)OFTHEDISEASE........................................................................................................................................90
FIGURE4-1A:IMAGINGPROCESSINGFLOW.B:REPRESENTATIONOFABRAINNETWORKWITHAMODULARSTRUCTURE.DIFFERENTGRAPHMETRICSAREHIGHLIGHTED.................................................116
FIGURE4-2A:NODESWITHASIGNIFICANTLYBISCOREINFREEZERSCOMPAREDTONON-FREEZERS,B:NODESWITHASIGNIFICANTLYDIFFERENTWISCOREINFREEZERSCOMPAREDTONON-FREEZERS)122
FIGURE4-3:REPRESENTATIONOFTHERICH-CLUBNETWORK..................................................................123
SupplementaryFigure4-1:Averagedegreeoftherichclubnodes..............................................139FIGURE6-1:EXAMPLEOFSINGLEANDHIDDENIMAGESOFTHEBPP.......................................................157
FIGURE6-2: NBSANALYSISREVEALSASUB-NETWORK,COMPRISING183EDGESWITHREDUCEDCONNECTIVITYSTRENGTHCORRELATEDTOINCREASEDHSS....................................................................164
FIGURE6-3:OVERLAPBETWEENTHEIDENTIFIEDSTRUCTURALSUBNETWORKANDFUNCTIONALRESTING-STATENETWORKS...............................................................................................................................165
FIGURE6-4:NODESRANKEDACCORDINGTOTHEBISCORES.....................................................................167
FIGURE6-5:SIGNIFICANTCHANGESINBETWEEN-ANDWITHIN-MODULESCORESASSOCIATEDWITHTHEHSS.................................................................................................................................................................169
Supplementaryfigure6-1:Meanconnectivitymatrix........................................................186Supplementaryfigure6-2:Size of sub-networks correlated with the HSS for different t-statistics ................................................................................................... 186
FIGURE7-1:AHYPOTHETICALOVERVIEWOFHOWPATHOLOGYMAYSPREADIFNODESWITHHIGHCONNECTIVITYSTRENGTHDISPLAYINCREASEDVULNERABILITY...............................................................208

xi
Box Index
This thesis includes ‘Boxes’ throughout the text that refer to a co-authored publication in
the Appendix. The text boxes complement the content of the thesis.
BOX1:EVIDENCEFORTHEIMPACTOFFREEZINGOFGAITONAPATIENT’SQUALITYOFLIFE................14
BOX2A:EVIDENCEFORTHEASSOCIATIONBETWEENFREEZINGOFGAITANDEXECUTIVEDYSFUNCTION.................................................................................................................................................................................108
BOX2B:EVIDENCEFORTHEASSOCIATIONBETWEENFREEZINGOFGAITANDEXECUTIVEDYSFUNCTION.................................................................................................................................................................................108BOX3:EVIDENCEFORTHEASSOCIATIONBETWEENFREEZINGOFGAITANDMOODDISTURBANCES..109
BOX4:IDENTIFYINGRISKFACTORSFORFREEZINGOFGAIT.......................................................................200
BOX5:EVIDENCEFORIMPAIREDSENSORYEVIDENCEACCUMULATIONINPARKINSON’SDISEASEPATIENTSWITHVISUALHALLUCINATIONS.....................................................................................................203
BOX6:COGNITIVETRAININGINPATIENTSWITHFREEZINGOFGAIT........................................................205

xii
Publications Presented for Examination
This thesis principally represents the work of Ms Julie Mae Hall. As supervisors,
Doctor Ahmed Moustafa, Professor Simon Lewis and Professor Peter Keller
provided support in her studies and in the reviewing the Introduction and Discussion
chapters of this thesis. Each author below contributed to the publication of the
Chapters by reviewing the written work prior to journal submission. Contributions of
all co-authors are listed below for each reference.
The following papers form the basis of this thesis:
Chapter 2: Hall, J. M., Ehgoetz Martens, K. A., Walton, C. C., O’Callaghan, C.,
Keller, P. E., Lewis, S. J. G., Moustafa, A. A. Diffusion alterations
associated with Parkinson's disease symptomatology: A review of the
literature. Parkinsonism and Related Disorders, 2016
Performed literature search and selection: JMH. Wrote the manuscript:
JMH. Critically appraised the manuscript: all authors.
Chapter 3: Hall, J. M., Shine, J. M., Walton, C. C., Gilat, M., O’Callaghan, C.,
Naismith, S. L., Lewis, S. J. G. Freezing of Gait and its associations in the
early and advanced stages of Parkinson’s disease. Journal of Parkinson’s
Disease, 2015
Designed the study: JMH. Collected the data: JMH, JMS, CCW, MG, CO,
SJGL. Designed and performed analyses: JMH. Wrote the manuscript:
JMH. Critically appraised the manuscript: all authors.
Chapter 4: Hall, J. M., Shine, J.M., Ehgoetz Martens, K. A., Gilat, M., Broadhouse,
K. M., Szeto, J. Y. Y., Walton, C. C., Moustafa, A. A., Lewis, S. J. G.
Alterations in white matter network topology contribute to freezing of
gait. Journal of Neurology, 2018

xiii
Designed the study: JMH. Data collection: JMH, JMS, CCW, MG, SJGL.
Designed the analyses: JMH, JMS. Performed analyses: JMH. Processed
and quality controlled data: JMH, JYS. Wrote the manuscript: JMH.
Critically appraised the manuscript: all authors.
Chapter 5: Hall, J. M., O’Callaghan, C., Shine, J. M., Muller, A. J., Phillips, J. R.,
Walton, C. C., Lewis, S. J. G., Moustafa, A. A. Dysfunction in attentional
processing in patients with Parkinson's disease and visual hallucinations.
Journal of Neural Transmission, 2016
Designed the study: JMH. Collected the data: JMH, CO, JMS, AJM.
Designed and performed analyses: JMH. Wrote the manuscript: JMH.
Critically appraised the manuscript: all authors.
Chapter 6: Hall, J. M., O’Callaghan, C., Ehgoetz Martens, K. A., Phillips, J. R.,
Moustafa, A. A., Lewis, S. J. G., Shine J. M. Changes in structural
network architecture correlate with severity of hallucinatory behaviour
in Parkinson’s disease. Network Neuroscience, 2019
Designed the study: JMH, JMS. Collected the data: JMH, MG, SJGL,
JMS. Designed and performed analyses: JMH, JMS. Processed and
quality controlled the data: JMH. Wrote the manuscript: JMH. Critically
appraised the manuscript: all authors.

xiv
Other peer-reviewed publications associated with this thesis
A substantial contribution was made to the following papers that are complementary
to the thesis. The outcomes of these studies helped to inform the theoretical
framework and interpretations of the data reported in the thesis. As such, this work
has been included in the Appendix.
Appendix 1. Walton, C. C., Shine, J. M., Hall, J. M., O’Callaghan, C., Mowszowski,
L., Gilat, M., Szeto, J. Y. Y., Naismith, S. L., Lewis, S. J. G. The major
impact of freezing of gait on quality of life in Parkinson’s disease. Journal
of Neurology, 2015
JMH: Critically appraised the manuscript
Appendix 2. Walton, C. C., Shine, J. M., Mowszowski, L., Gilat, M., Hall, J. M.,
Naismith, S. L., Lewis, S. J. G. Impaired cognitive control in Parkinson’s
disease patients with freezing of gait in response to cognitive load. Journal
of Neurotransmission, 2015
JMH: Statistical analysis and critically appraised the manuscript
Appendix 3. Gilat, M., Shine, J. M., Walton, C. C., O’Callaghan, C., Hall, J. M.,
Lewis, S. J. G. Brain activation underlying turning in Parkinson’s disease
patients with and without Freezing of Gait. NPJ Parkinson’s Disease,
2015
JMH: Critically appraised the manuscript
Appendix 4. Walton C.C., O’Callaghan, C., Hall, J. M., Gilat, M., Mowszowski, L.,
Naismith, S.L., Burrell, J., Shine, J.M., Lewis, S. J. G. Antisaccade errors

xv
reveal cognitive control deficits in Parkinson’s disease patients with
freezing of gait. Journal of Neurology, 2015
JMH: Data collection, statistical analysis and critically appraised the
manuscript
Appendix 5. Ehgoetz Martens, K. A., Hall, J. M., Gilat, M., Georgiades, M. J., Walton,
C. C., Lewis, S. J. G. Anxiety is associated with freezing of gait and
attentional set-shifting in Parkinson's disease: A new perspective for early
intervention. Gait & Posture, 2016
JMH: Data collection and critically appraised the manuscript
Appendix 6. Ehgoetz Martens, K. A., Szeto, J. Y. Y., Muller, A. J., Hall, J. M., Gilat,
M., Walton, C. C., Lewis, S. J. G. Cognitive function in Parkinson's
disease patients with and without anxiety. Neurology Research
International, 2016
JMH: Data collection and critically appraised the manuscript
Appendix 7. Gilat, M., Hall, J. M., Ehgoetz Martens, K. A., Shine, J. M., Walton, C.
C., MacDougall, H. G., Moore, S. T., Lewis, S. J. G. Staircase climbing is
not solely a visual compensation strategy to alleviate Freezing of Gait in
Parkinson's disease. Journal of Neurology, 2016.
JMH: Data analysis and critically appraised the manuscript.
Appendix 8. Gilat, M., Bell, P. T., Ehgoetz Martens, K. A., Georgiades, M. J., Hall, J.
M., Walton, C. C., Lewis, S. J. G., Shine, J. M. Dopamine depletion
impairs gait automaticity by altering cortico-striatal and cerebellar
processing in Parkinson's disease. NeuroImage, 2017

xvi
JMH: Critically appraised the manuscript.
Appendix 9. Ehgoetz Martens, K. A., Lukasik, E., Georgiades, M, J., Gilat, M., Hall, J.
M., Walton, C. C., Lewis, S. J. G. Predicting the onset of freezing of gait:
A Longitudinal study. Movement Disorders, 2017
JMH: Study design and critically appraised the manuscript.
Appendix 10. Ehgoetz Martens, K. A., Hall, J. M., Georgiades, M, J., Gilat, M., Walton,
C. C., Matar, E., Lewis, S. J. G., Shine, J. M. The functional network
signature of heterogeneity in freezing of gait. Brain, 2018
JMH: Critically appraised the manuscript.
Appendix 11. Walton, C. C., Mowszowski, L. Gilat, M., Hall, J. M., O’Callaghan, C.,
Muller, A. J., Georgiades, M. J., Szeto, J. Y. Y., Ehgoetz Martens, K. A.,
Shine, J. M., Naismith, S. L., Lewis, S. J. G. Cognitive training for
freezing of gait in Parkinson’s disease: a randomized controlled trial. NPJ
Parkinson's Disease, 2018
JMH: Data collection and analysis, critically appraised the manuscript.
Appendix 12. O’Callaghan, C. Hall, J. M., Tomassini, A., Muller, A. J., Walpola, I. C.,
Moustafa, A. A., Shine, J. M., Lewis, S. J. G. Visual Hallucinations Are
Characterized by Impaired Sensory Evidence Accumulation: Insights
From Hierarchical Drift Diffusion Modeling in Parkinson's Disease.
Biological Psychiatry: Cognitive Neuroscience and Neuroimaging, 2017
JMH: Data collection and critically appraised the manuscript
Appendix 13. Moustafa, A.A., Chakravarthy, S., Phillips, J. R., Crouse, J. J., Gupta, A.,
Frank, M. J., Hall, J. M., Jahanshahi, M. Interrelations between cognitive

xvii
dysfunction and motor symptoms of Parkinson's disease: behavioural and
neural studies. Reviews in the Neurosciences, 2016
JMH: Edited and critically appraised the manuscript

xviii
Abbreviations
PD Parkinson’s disease
FOG Freezing of gait
VH Visual hallucinations
PIGD Postural instability and gait disorder
MCI Mild cognitive impairment
RBD REM sleep behaviour disorder
SNpc Substantia nigra pars compacta
SNpr Substantia nigra pars reticulata
GPi Globus pallidus internus
GPe Globus pallidus externus
STN Subthalamic nucleus
GABA Gamma-aminobutyric acid
MRI Magnetic resonance imaging
DTI Diffusion tensor imaging
FA Fractional anisotropy
MD Mean diffusivity
TBSS Tract-Based Spatial Statistics
SLF Superior longitudinal fasciculus
IF-OF Inferior fronto-occipital fasciculus

1
ABSTRACT
Idiopathic Parkinson’s disease is a neurodegenerative condition that presents with a
myriad of motor and non-motor symptoms. The complex and heterogeneous character
of Parkinson’s disease suggests that it is a multisystem disorder with widespread
pathology affecting different anatomical structures. In addition to localized dysfunction,
changes in connections between brain regions and networks may contribute to
Parkinson’s symptomology. This thesis explores this hypothesis in two prevalent
symptoms of Parkinson’s disease: freezing of gait and visual hallucinations.
Freezing of gait is an involuntarily paroxysmal breakdown of an individual’s footstep
pattern whilst walking. Freezing of gait can be triggered by environmental factors or
additional cognitive load. Furthermore, freezing behaviour can also be observed in non-
gait tasks, such as during speech or handwriting. These observations suggest that freezing
cannot solely be attributed to a motor dysfunction. This thesis provides support for the
notion that freezing of gait results from ineffective communication between distinct brain
regions. Specifically, this thesis showed that freezing of gait is associated with deficits in
executive functioning, mood and sleep disturbances as well as with the non-tremor
dominant motor phenotype, confirming its multi-system nature.
To investigate the underlying mechanisms of this multi-system symptom, the structural
network topology in patients with freezing of gait was assessed. Importantly, the brain
displays a modular structure and changes in the diversity of inter-modular connections
(between-module connectivity) and the connections of a node within its own module
(within-module connectivity) can inform about the integration and segregation of
specialized communities. This thesis showed that patients with freezing of gait
showed reduced between-module connectivity in subcortical and frontoparietal
nodes. Several of these nodes were found within the brain’s ‘rich club’, a group of
highly interconnected nodes that form the structural backbone of the connectome
and, therefore, play an important role in global information integration between
different modules in the brain. Freezing of gait could thus arise due to ineffective

2
communication between major cortical and subcortical integration centres.
Furthermore, changes in within-module scores were observed across different
sensory processing modalities. This may reflect a compensatory strategy and could
indicate that compared to non-freezers, freezers rely more heavily on exogenous
stimuli to guide their actions, perhaps due to a loss of automaticity.
Visual hallucinations represent another common, yet poorly understood symptom of
Parkinson’s disease. The pathophysiology of visual hallucinations is likely multi-faceted
with changes across attentional and perceptual brain networks. First, we investigated
attentional processing capability in patients with visual hallucinations using a
computerized flanker task. With increasing task demands, Parkinson’s disease patients
with visual hallucinations showed lower accuracy rates than patients without this
symptom, yet the reaction times were similar between groups for correct trials. These
findings suggest that hallucinators have specific impairments in sustained attention and
conflict monitoring. Furthermore, the episodic nature suggests that visual hallucinations
may arise during periods of ineffective integration of information.
To investigate whether the severity of hallucinatory behaviour was associated with ability
to successfully integrate information, changes in structural network topology were
assessed. Severity of hallucinatory behaviour was associated with reduced connectivity
across a structural bilateral sub-network. Nodes within this network showed higher
between- and within-module scores compared to nodes outside this network. This may
indicate that the system requires to reroute information across less efficient pathways,
affecting the sensory integration process. Furthermore, severity of hallucinations was
associated with changes in between-module scores across top-down visual processing
centres and attentional networks. Thus, impaired integration across visual processing sites
may result in the inefficient transfer of information that gives rise to visual hallucinations
in Parkinson’s disease.
The findings presented in this thesis suggest that freezing of gait and visual hallucinations
may be the result of alterations in the brain’s information processing capability as
indicated by reduced interactions between brain networks. By investigating the inherent

3
complexity of the brain’s interacting subsystems and their connections, we can advance
our understanding of neurodegenerative disorders and ultimately, aid the developments of
new and improved treatment strategies.

CHAPTER1INTRODUCTION

4
Chapter 1: Introduction
1.1 Introduction to Parkinson’s disease
Parkinson’s disease was first described in the Western medical literature in 175 AD as
shaking palsy (van Swieten, 1742). However, it was not until 1817 that James Parkinson
wrote a detailed essay that established this disorder as a medical condition (Parkinson,
1817). Parkinson’s disease has traditionally been viewed as a movement disorder, with
cardinal motor symptoms such as rest tremor, bradykinesia and muscular rigidity. Only
recently, Parkinson’s disease has been reclassified as a far more complex disorder,
affecting non-motor aspects including, but not limited to, cognition, speech, mood and
sleep (Postuma et al., 2015). To date, Parkinson’s disease is amongst the most prevalent
neurodegenerative disorders, yet is still poorly understood.
Age is the greatest risk factor for Parkinson’s disease and with ageing population, the
number of cases worldwide are expected to increase from an estimated 4.1 million in
2005, to between 8.7 and 9.3 million by 2030 (Dorsey et al., 2007). Parkinson’s disease
has a detrimental impact on quality of life and independence, especially with advancing
disease (Schrag et al., 2000). Despite decades of research into this debilitating disorder
there is no cure and whilst symptomatic treatments have been developed, many features
are poorly managed or even untreated due to a lack of understanding of the underlying
mechanisms.
1.1.1 Basal ganglia dysfunction
The main neuropathological hallmarks of Parkinson’s disease are Lewy bodies and Lewy
neuritis (Lewy, 1912; Ohama & Ikuta, 1976). Lewy pathology, discovered in 1912 by the
neurologist Dr Friedrich Lewy, consist of abnormal deposits of protein, with alpha-
synuclein as their main component, that disrupt the brain’s functioning. It is not known
whether these inclusions directly disrupt neuronal functioning or represent some form of
compensatory effort, but their presence is associated with cell death and thus dysfunction.
The substantia nigra pars compacta (SNpc) is considered to be particularly vulnerable to

5
Lewy pathology. The subsequent loss of dopaminergic neurons in SNpc result in
depletion of dopaminergic input to the striatum. The onset of the cardinal symptoms of
Parkinson’s disease correlates with depletion in the dorsal striatum, specifically in the
putamen. The striatum projects gamma-aminobutyric acid (GABAergic) efferents to the
output nuclei of the basal ganglia, the ventrolateral internal pallidal segment (GPi) and
the caudolateral substantia nigra pars reticulata (SNpr), either directly or indirectly via
the intrinsic nuclei of the basal ganglia. The GPi/SNpr consist of inhibitory GABAergic
neurons with a high rate of discharge that tonically fire to inhibit the brainstem nuclei.
Thus, inhibitory projections to the GPi and SNpr, via the direct pathway, are associated
with facilitating behaviour. Conversely, the indirect pathway inhibits the external part of
the globus pallidus (GPe), which in turn results in disinhibition of the subthalamic
nucleus (STN). The STN subsequently sends excitatory glutamatergic projections to
GPi/SNpr neurons, thus activation of this pathway is associated with stopping or halting
movement. A simplified description of the basal ganglia and its projections is illustrated
in Figure 1.
Figure 1. Simplified representation of the basal ganglia-thalamocortical circuit in a) healthy individuals
and b) Parkinson’s disease patients. GPe = globus pallidus externus; GPi = globus pallidus internus; SNc

6
= substantia nigra pars compacta; SNr = substantia nigra pars reticulate; STN = subthalamic nucleus;
PPN = pedunculopontine nucleus; SC = spinal cord; GABA = gamma-aminobutyric acid; BG = basal
ganglia. Thickness of lines and dashed lines represent increased and decreased activity.
The basal ganglia are components of parallel basal ganglia–thalamocortical circuitry that
display a spatial topological organization and are functionally subdivided into a motor,
oculomotor, limbic and associative areas, following a dorsal-ventral gradient (Middleton
et al., 2000). However, the input to the striatum from these segregated loops overlap and
the striatum receives convergent input from multiple, often related, cortical areas. This
enables the striatal nuclei to integrate information from different cortical areas and
modalities (Bell et al., 2016; Chudler et al., 1995; Nagy et al., 2006). Evidently, basal
ganglia dysfunction has major implications in patients with Parkinson’s disease.
Dopamine depletion disrupts the balance of the direct and indirect routes and leads to a
reduced ability of the basal ganglia to control the thalamic output to the cortex.
Dysfunction within these subcortical nuclei has been associated with motor disturbances
such as bradykinesia, impaired locomotion and postural muscle tone (Berardelli et al.,
200; Takakusaki et al., 2003), as well as deficits in executive functioning, working
memory, motivation and learning capacity (Aarts et al., 2011; Frank, 2005; Middleton et
al., 2000; Moustafa et al., 2008).
1.1.2 Lewy pathology
Other regions outside the SNpc are also affected by Lewy bodies, and according to the
Braak hypothesis, Lewy bodies and Lewy neurites evolve according to a topographical
sequence along major fibre pathways, starting with pre-symptomatic Lewy pathology in
the dorsal motor nucleus of the vagal nerve and frequently in the anterior olfactory
nucleus (Braak et al., 2003). This involvement may explain prodromal features such as
the dysregulation of parasympathetic functions such as constipation and the loss of smell,
which are commonly seen prior to the clinical diagnosis of Parkinson’s disease (Goldman
& Postuma, 2014). In Braak Stage 2, the Lewy pathology spreads in a caudal to rostral
direction via the brainstem, affecting the serotonergic raphe nuclei, gigantocellular

7
reticular nucleus, and the noradrenergic coeruleus–subcoeruleus complex. These nuclei
are involved in mood, somatic motor functioning and sleep mechanisms respectively;
functions that can become deregulated before the cardinal motor symptoms of
Parkinson’s disease appear. Indeed, only in Braak Stage 3, when Parkinson’s disease
pathology reaches the SNpc, the pedunculopontine nucleus, the magnocellular nuclei of
the basal forebrain and the amygdala, is it possible to make a diagnosis of Parkinson’s
disease by current criteria (Postuma et al., 2015). In Braak Stage 4, Lewy pathology
progresses to the hypothalamus, thalamus, and the anteromedial temporal mesocortex.
Finally, in Braak Stage 5 and 6, the higher order association areas, including the insula
and cingulate are affected.
Whilst the stages described by Braak and colleagues generally appear to be confirmed
when a clinical diagnosis of Parkinson’s disease has been made, it remains difficult to
predict the course of the disease and the manifestations of the different symptoms in an
individual. Indeed, not all patients will experience the same set of symptoms and even the
chronological order of the manifestation of symptoms differs from one patient to another
(Lewis et al., 2005). Additionally, neurofibrillary tangles, senile plaques, microvascular
disease and argyrophilic inclusions also contribute to Parkinsonian pathology and their
anatomical distribution varies between cases (Dickson et al., 2009; Halliday et al., 2014).
Finally, axonopathy and synaptic dysfunction may impair white matter integrity in
Parkinson’s disease (O’Malley, 2010). This complex and diverse milieu of
neuropathological factors renders it challenging to provide a generalized model
explaining the mechanisms underlying Parkinson’s disease and may cause the great
clinical diversity seen in patients, which makes diagnosis and treatment strategies
difficult. Finally, Parkinson’s heterogeneous manifestation raises the question of whether
this neurodegenerative disorder is currently adequately characterized, or better serves as
an umbrella term for different types of the condition (Fereshtehnejad et al., 2017).
Studies using magnetic resonance imaging (MRI) combined with sensitive and specific
behavioural tests have refined our understanding of Parkinson’s disease’s highly complex
and heterogeneous pathophysiology. Such insights are crucial for facilitating accurate and
timely diagnosis as well as to establish a robust brain-behaviour relationship to inform

8
pattern and progression of disease pathology. However, the clinical diversity of
Parkinson’s disease has led to discrepancies in MRI studies investigating the underlying
mechanisms of the disorder. This is specifically apparent in studies investigating
structural brain changes. Gross macroscopic atrophy is not a key pathological process in
Parkinson’s disease until the last clinical stages and changes in grey and white matter are
likely to be subtler and symptom related. Indeed, when comparing Parkinson’s disease
patients with healthy controls, studies investigating white matter using diffusion tensor
imaging (DTI) have reported relatively robust subcortical findings but cortical changes
varied, with results not always found to be replicable. This will become evident from the
brief review provided on the current findings of white matter changes in section 1.3 of the
Introduction. First, an overview of DTI theory is provided.
1.2 White matter and diffusion tensor imaging
Over half of the human brain consists of white matter: myelinated axons that connect
neurons from different brain regions into functional circuits (Fields, 2010). Myelin
affects the impulse conduction speed through axons, ensuring the temporal coincidence
of firing from multiple, spatially differently located presynaptic neurons with regards to
their joint postsynaptic firing (Fields, 2010). The summation of the conducted signals in
the postsynaptic neuron creates a larger voltage response, which then can reach a critical
threshold for the recipient neuron to fire. The conduction velocity of axons is therefore
crucial for effective information processing (Fields, 2008) and any changes to the tract
integrity can impair or suppress the transport of neurotransmitters through axons (Braak
et al., 1999). There is some evidence that axonal degeneration is one of the earliest
features of Parkinson’s disease. Specifically, it has been suggested that the loss of striatal
axonal projections and terminals outweigh SN neuronal loss, and is responsible for
progressive clinical deterioration (Burke et al., 2013; Rektor et al., 2018). This
proposition needs further verification, but it warrants the exploration of white matter
integrity in Parkinson’s disease.
One MRI technique that can detect changes in white matter axons is DTI. DTI indirectly
infers white matter integrity by measuring the three-dimensional motion of water

9
molecules (Le Bihan et al., 2001). In the absence of boundaries, water molecule
displacement obeys a Gaussian distribution, described by Einstein’s diffusion equation
(Einstein, 1956). Diffusion in biological tissues however, deviates from this law as
cellular structures and membranes hinder the motion of water molecules. These structures
cause water molecules to take more tortuous paths, thereby decreasing their mean
displacement (Alexander et al., 2007). In fibrous tissues such as homogenous white
matter axons, water moves relatively freely in the direction parallel to the axons.
Conversely, water molecules are hindered in the direction perpendicular of the fibre
orientation. The diffusion tensor can be described as a 3 x 3 covariance matrix that
describes the covariance of the three-dimensional diffusion displacement (normalised by
the diffusion time).
! =!!! !!" !!" !!" !!! !!" !!" !!" !!!
The diagonal elements in the diffusion matrix (Dii) represent the water displacement over
the x, y and z axis (the eigenvalues λ1, λ2 and λ3) and the elements off the diagonal
represent the covariance (the eigenvectors ε1, ε2 and ε3). The tensor can be visualised as
an ellipsoid with the eigenvalues and eigenvectors describing its principal diffusion
direction and apparent diffusivities (see Figure 2).
The size of the eigenvalue is sensitive to change in local tissue integrity, due to axonal
injury or demyelination (Le Bihan et al., 2001). Changes in white matter integrity lead to
less convoluted paths, thereby increasing the mean or apparent displacement (MD) of the
water molecules. Fractional anisotropy (FA) is the most commonly reported measure and
characterizes the orientation distribution of the random movement of water molecules
along the principal axis (Conturo et al., 1999). FA values close to 1 indicate anisotropic
movement whilst values close to 0 indicate isotropic movement. Axial diffusivity (Da =
λ1) is considered to be more specific to fibre degeneration whereas radial diffusivity (Dr
= (λ2 + λ3)/2)) reflects changes in myelin (Le Bihan et al., 2001).

10
Figure 2. Schematic representation of diffusion displacement. The direction of greatest diffusivity (λ1) is
assumed to be parallel to the local direction of white matter.
Since the orientation of the diffusion tensor is thought to be aligned with the axon, white
matter tracts can be reconstructed, in-vivo and non-invasively with a technique called
tractography. Starting at the seed point, a tract can be reconstructed by following its
principal direction, and moving forward in that direction until the tract is terminated, re-
evaluating its direction along the way (e.g. in each voxel). This approach is called
deterministic tractography (Mori et al., 1999). Probabilistic tractography estimates the
‘most likely’ fibre direction in each voxel, accounting for uncertainty (Parker et al.,
2003).
Furthermore, whole brain white matter can be assessed using voxel-based analysis
(originally developed for assessing changes in grey matter) or the more sophisticated
technique Tract-Based Spatial Statistics (TBSS) (Smith et al., 2006). These techniques
are fully automated and do not require a priori tracts or regions of interest. TBSS
estimates a group mean FA skeleton that represents the centres of the fibre bundles that
are common across subjects included in the investigation. The FA data of each subject is
projected on the group mean skeleton, after which voxel-wise statistics can be carried out
(Smith et al., 2006).
λ2 λ1
λ3
λ2 λ1
λ3
Isotropic, unrestricted diffusion
Anisotropic, restricted diffusion
Diffusion trajectory
Diffusion ellipsoid

11
1.3 Diffusion tensor imaging in Parkinson’s disease
Patients in the early stages of the disease already show changes in diffusivity, as observed
by decreased FA values in both the olfactory region and the SN (Rolheiser et al., 2011;
Vaillancourt et al., 2009). A recent meta-analysis including a heterogeneous group of
1087 Parkinson’s disease patients and 769 healthy controls confirmed these changes and
reported both lower FA and higher MD values in the SN (Atkinson-Clement et al., 2017).
Importantly, both FA and MD measures within the SN were found to be good indicators
for Parkinson’s disease identification as well as disease progression (Loane et al., 2016;
Edward Ofori et al., 2015; Zhang et al., 2016). This meta-analysis also showed changes
in the olfactory region, as well as other subcortical structures such as the putamen,
pallidum and internal and external capsules with studies reporting either lower FA or
higher MD values in patients compared to healthy controls. Surprisingly, the opposite
pattern was found in the caudate nucleus and cortico-spinal tract in Parkinson’s disease
patients compared to healthy controls, perhaps reflecting a reorganisation of the
pathways. Increased FA or decreased MD could also reflect gliosis or glial scarring
following axonal or neuronal insult (Budde et al., 2011). However, there is no apparent
explanation of why this opposite pattern only affects the caudate nucleus.
Parkinson’s disease patients in the early clinical stages also show increased axial
diffusivity in the left superior and anterior corona radiata, internal and external capsule.
Furthermore, elevations in radial and mean diffusivity and decreased white matter
volume in the bilateral frontal and temporal areas are observed in patients without
cognitive impairment relative to healthy control subjects (Martin et al., 2009; Rektor et
al., 2018). However, a recent meta-analysis found that diffusivity changes in cortical
areas were vastly heterogeneous: changes in diffusivity were not universal across groups
but correlated to different symptoms (Atkinson-Clement et al., 2017). For example, a
high variability was found in DTI measures of the corpus callosum but its white matter
integrity was found to be associated with gait difficulties and impulse disorders
(Atkinson-Clement et al., 2017). Furthermore, changes in diffusivity in the temporal
cortex were only observed in patients with cognitive decline (Price et al., 2016).

12
Therefore, greater insights into Parkinson’s disease pathology might result from
investigating the underlying mechanisms of isolated symptoms rather than grouping all
Parkinson’s disease patients together. Chapter 2 of this thesis provides an in-depth
review of the current findings on changes in diffusivity associated with specific motor,
cognitive, associative and limbic symptoms.
To get a better understanding of the heterogeneity in Parkinson’s disease, this thesis will
particularly focus on two symptoms, namely freezing of gait and visual hallucinations.
These seemingly disparate symptoms share several common features. Both freezing of
gait and visual hallucinations are more often present in the advanced disease stages and
are associated to the non-tremor dominant phenotype of Parkinson’s disease. Patients
with this pyhenotype typically suffer from a range of cognitive and mood disturbances,
on top of prominent motor symptoms such as gait disturbances and postural instability,
rigidity and bradykinesia (Lewis et al., 2005) compared to patients in the tremor-
dominant group. Furthermore, previous work has suggested that both freezing and
hallucinations are related to attentional network dysfunction (Shine, 2013b).
The next sections will discuss the known associations of freezing of gait and visual
hallucinations as well as the most putative hypotheses regarding their underlying
neurobiology. However, investigations into the white matter changes underlying freezing
of gait have been restricted and contradictory whilst studies evaluating visual
hallucinations are lacking. It is possible that both of these symptoms could be the result
of a broader network dysfunction that arises due to ineffective communication yet
conventional DTI techniques might not be able to adequately capture the related changes.
Therefore, Chapter 4 and 6 of this thesis will investigate whether these symptoms could
be described as failure of effective communication across brain networks.
1.4 Freezing of gait in Parkinson’s disease
Freezing of gait is a common debilitating symptom described as a sudden inability to
generate effective forward stepping, despite the intention to do so (Nutt et al., 2011), and
is often described by patients as though their feet are ‘glued’ to the floor. Freezing of gait

13
has a major impact on a patient’s independence, risk of falls and quality of life (Gray et
al., 2000; Walton et al., 2015; see Box 1). The freezing phenomenon is associated with
impairments in executive functioning (Amboni et al., 2008; Muller et al., 2014; Naismith
et al., 2010a), visuospatial processing (Ehgoetz Martens et al., 2014a; Matar et al., 2013)
and anxiety (Ehgoetz Martens et al., 2014b; Lieberman, 2006). Environmental triggers,
such as obstacles or doorways (Cowie et al., 2012), dual-tasking (de Souza Fortaleza et
al., 2017; Spildooren et al., 2010) and stress (Giladi et al., 2006) can all trigger freezing
of gait, whilst patients can sometimes use cueing strategies to alleviate this symptom
(Rahman et al., 2008). Freezing is not restricted to the lower limbs but can also occur in
the upper limbs (Nieuwboer et al., 2009) and speech (Ackermann et al., 1993), suggesting
that the neural network underlying the phenomenon extends beyond the motor control of
gait (Rahman et al., 2008). These observations have led to the proposal that freezing
cannot be summarized by just a gait disturbance but might be the result of ineffective
information processing across multiple circuits, leading to dysfunction in a common final
pathway (Lewis & Barker, 2009; Lewis & Shine, 2016). In brief, it is proposed that due
to an ineffective segregation of the motor, cognitive and limbic pathways, a variety of

14
stimuli and conditions can cause an overwhelming response when conflict arises. This
overwhelming response leads to an increased firing of the STN, which is part of the
inhibitory hyper-direct pathway of the basal ganglia. In turn, overactivity in the STN
leads to paroxysmal activity in the GPi, subsequently inhibiting the activity of the basal
ganglia output nuclei to the locomotor region in the brainstem, disrupting the effective
motor plans (Lewis & Shine, 2016). Evidence for this hypothesis comes from a number
of functional MRI studies that have shown alterations within frontal and parietal cortical,
cerebellar and brainstem locomotor networks in Parkinson’s disease patients with
freezing of gait. For example, during motor arrests provoked by a virtual reality
paradigm, alterations in BOLD responses were observed within frontoparietal and insular
cortices as well as sensorimotor regions, basal ganglia, thalamus and mesencephalic
locomotor regions (Shine et al., 2013b). Furthermore, a functional decoupling between
the bilateral cognitive control network and the basal ganglia during freezing behaviour
was also evident (Shine, 2013a). Altered task-free (‘resting state’) functional connectivity
has also been shown to be associated with the severity of freezing behaviour, specifically
within the cognitive control network, visual networks and locomotor regions including
Box 1: Evidence for the impact of freezing of gait on a patient’s quality of life
Walton, C. C., Shine, J. M., Hall, J. M., O’Callaghan, C., Mowszowski, L., Gilat, M.,
Szeto, J. Y. Y., Naismith, S. L., Lewis, S. J. G. The major impact of freezing of gait
on quality of life in Parkinson's disease. Journal of Neurology, 2015.
This study performed a regression analysis including factors that are known to impact
a patient’s well-being. The model included cognition, motor severity, sleep and mood
disturbances and freezing of gait. After controlling for disease duration and current
dopaminergic treatment, the significant predictors to a patient’s quality of life were
self-reported sleep-wake and mood disturbances and freezing of gait. Importantly,
freezing of gait accounted for the highest amount of unique variance to a patient’s
quality of life. We hypothesized that this finding is due to a loss of independence and
fear of injury.

15
the supplementary motor cortex, mesencephalic and cerebellar motor regions (Fling et al.,
2014; Tessitore et al., 2012). Together, these results confirm the involvement of both
higher order cortical and locomotor regions in the manifestation of freezing of gait.
Other models of freezing exist that have aimed to describe freezing behaviour in
Parkinson’s disease. Plotnik and colleagues (2012) hypothesized that freezing of gait
results from poor control of one or more gait disturbances associated with freezing:
deterioration in bilateral coordination, gait symmetry and rhythmicity, dynamic postural
control and/or step scaling (Plotnik et al., 2012). When the overall gait performance can
no longer be maintained above a certain threshold, a freezing episode takes place.
Additionally, freezing of gait is often accompanied by a trembling of the knee and it was
found that anticipatory postural adjustments coexist with knee trembling during
automatically triggered stepping responses to platform perturbations. This observation
gave rise to the suggestion that freezing results from a decoupling between pre-planned
motor programs, such as anticipatory postural adjustment during step initiation and the
release of an inherent stepping movement (Jacobs et al., 2009). The mechanisms in both
models indicate involvement of the pedunculopontine nucleus (PPN), a collection of
cholinergic and glutamatergic neurons located in the brainstem that is involved in
voluntary limb movements, the planning of movement, posture as well as attention and
arousal (Garcia-Rill et al., 2015; Tsang et al., 2010). Furthermore, the PPN provides
sensory feedback to the cerebral cortex (Li et al., 2015). These models are supported by
research into deep brain stimulation of the PPN, which can sometimes improve postural
instability and gait disorders (Wang et al., 2016). However, these models do not account
for the symptoms that often accompany freezing, such as executive dysfunction and
anxiety (Hall et al., 2014). In contrast, Vandenbossche et al. (2013) suggest that the
interplay between decreased gait automaticity and impairments in cognitive control play
a key role in the occurrence of freezing of gait. Parkinson’s disease patients suffer from
motor automaticity as a result of subcortical impairments and therefore rely more heavily
on attentional resources. During challenging or ambiguous task requirements, freezing
episodes can occur as a result of a breakdown in stepping motion. This hypothesis also
incorporates the amelioration of freezing by using cueing strategies. Visual or auditory

16
cues force patients to utilize attentional control to maintain a stepping motion,
presumably by circumventing the impaired subcortical structures (Bella et al., 2017; Ghai
et al., 2018; Nieuwboer, 2008).
Evidently, more research into the associations and underlying mechanisms of this
prevalent symptom is highly necessary. Chapter 3 of this thesis aimed to explore the
epidemiology of freezing, as well as associated symptoms in different clinical stages of
Parkinson’s disease. Subsequently, Chapter 4 assessed changes in the structural network
topology of patients with freezing of gait, to explore whether this symptom can be
described as a result of a more general network dysfunction due to ineffective
information processing.
1.5 Visual hallucinations in Parkinson’s disease
Similar to freezing of gait, hallucinations are more common in advanced Parkinson’s
disease. Visual hallucinations are the most frequently observed, with a reported
prevalence of up to 80% in the later clinical stages (Hely et al., 2008; Amar et al., 2014).
In addition, visual hallucinations are more often associated with dementia, sleep disorders
and depression (Sanchez-Ramos et al., 1996). Visual hallucinations range from relatively
benign misperceptions, such as a shadow in the peripheral visual field to well-formed
(often more frightening) hallucinations with a loss of insight, such as seeing people or
animals.
Visual symptoms are often reported in Parkinson’s disease, with 78% of patients
subjectively reporting visual complaints, including double vision whilst reading and
misjudging objections and distances (Weil et al., 2016). Visual hallucinations more often
occur in dim-light environments. Furthermore, it is well known that decreased contrast
sensitivity and colour vision, as well as impaired visual acuity are common features in
Parkinson’s disease (Archibald et al., 2011; Weil et al., 2016). Such visual complaints
more prevalent in Parkinson’s disease patients with hallucinations than in patients
without (Gallagher et al., 2011). In addition to end organ deficits that might be regarded
as arising from the eyes, hallucinators have reduced glucose consumption in the occipital-

17
temporal regions (Boecker et al., 2007) and reduced grey matter volume in higher visual
processing areas of the brain, such as the superior parietal lobe compared to non-
hallucinators (Ramírez-Ruiz et al., 2007; Lenka et al., 2015). Thus there are likely to be
combined ‘peripheral and central’ contributions to the visual hallucinations observed in
Parkinson’s disease. One of the earlier theories about the mechanisms underlying
Parkinsonian hallucinations proposed that they arise from decreased visual input, which
in turn leads to the ‘inappropriate release of stored images’ (ffytche et al., 1999).
Interestingly, significant glucose hypometabolism in visual association areas have been
shown in patients with dementia with Lewy bodies (Higuchi et al., 2000). Whilst the
visual pathway appears to be intact in this patient group, the inferior longitudinal
fasciculus, involved in visuospatial cognition, is degenerated in dementia with Lewy
bodies (Ota et al., 2009). Whilst lack of sensory input adds to the occurrence of simple
misperceptions, loss of visual stimuli due to eye disease or stroke does not always lead to
hallucinations. Indeed, some patients without any visual dysfunction also experience
hallucinations, suggesting that these deficits alone are not sufficient to explain their
occurrence (Muller et al., 2014). Neuropathological studies in dementia with Lewy
bodies have shown aggregated Lewy bodies in the inferior temporal cortex, including the
amygdala and parahippocampus, which have been linked with visual hallucinations
(Harding et al., 2002). The inferior temporal cortex integrates predictive information
from the orbitofrontal cortex with high spatial frequency information from the visual
areas (Trapp and Bar, 2015).
Models in Parkinson’s disease incorporate the interplay between perceptual and
attentional impairments to the occurrence of misperceptions and hallucinations. For
example, The Perception and Attention Deficit Model proposed that a hallucination might
intrude due to a combination of impaired attention and visual perception (Collerton et al.,
2005). Proto-objects are subconscious object representations competing for further
processing and reaching conscious awareness. When an incorrect visual proto-object is
bound to the attentional focus of a scene, due to both impaired attentional binding and
poor sensory activation of the proto-object, a visual hallucination can intrude within a
relatively intact scene representation. Object perception relies primarily on the visual

18
ventral stream, whilst attentional object recognition is processed in the lateral prefrontal
cortex (Collerton et al., 2005). Cholinergic pathology might disrupt communication
between these areas, resulting in inadequate integration of sensory information and
perceptual expectations. While this proposition is promising for recurrent visual
hallucinations, certain aspects need further exploration, including the manifestation of
unfamiliar images, and physiological details of proto-objects. Shine and colleagues
(2011) proposed that a breakdown in communication between exogenous and
endogenous attentional network give rise to visual hallucinations. Specifically, in the
presence of an ambiguous percept, an inability to recruit the dorsal attentional network, a
network responsible for focusing on externally driven percepts, leads to an over-reliance
on attentional networks that are less equipped to interpret visual stimuli, such as the
salience ventral attentional network and the default mode network, involved in the
retrieval and manipulation of episodic memory and semantic knowledge (Raichle et al.,
2001). This hypothesis has gained support from task-free functional neuroimaging studies
showing reduced coherence between the dorsal and ventral attentional network in
individuals prone to hallucinate (Shine et al., 2014). Hallucinators also showed reduced
grey matter volume within anterior insula, a node involved in shifting attention to the
dorsal attentional network when an ambiguous percept is presented. Furthermore, a
functional connectivity analysis showed increased coupling between the default mode
network and the visual networks during misperceptions (Shine et al., 2015).
Despite these advances, the exact mechanisms underlying this phenomenon are still
poorly understood. Their episodic and involuntary nature provide challenges for
investigating this symptom in real time, as it is difficult to elucidate misperceptions or
hallucinations in a clinical setting. This thesis will first investigate the attentional
processes in patients with visual hallucinations using an objective computerized task,
followed by an investigation of structural connectome changes associated with
hallucinatory behaviour. To date, few studies have investigated the contribution of white
matter alterations to this symptom, and assessing the network topology can shine a new
light on the interplay between the major attentional networks that possibly underlie
adequate perceptual processing.

19
1.6 Parkinson’s disease as a disconnection disorder
Considering their episodic nature and myriad of associations, it is anticipated that both
freezing of gait and visual hallucinations are the result of a multisystem communication
failure (Caligiore et al., 2016). By re-framing our understanding of these two symptoms
as impairments of a more general impairment in effective information transfer, we could
improve not only our conceptual understanding of freezing of gait and visual
hallucinations, but perhaps of Parkinson’s disease in general, and ultimately improve
currently available treatments.
To explore the role of connectivity in Parkinson’s disease, a newly emerging toolbox of
analytical techniques is required. Graph theory offers a tool to explore the inherent
complexity of the brain’s interacting subsystems and their connections. White matter
axons in the human brain together form the structural connectome and provide the
foundation for distributed patterns of brain activity. Brain regions strongly influence one
another through their interconnections as part of a larger network, and this organizational
structure plays a crucial role in healthy brain functioning (Bassett et al., 2009; van den
Heuvel et al., 2009). Many biological networks, including the brain, display a modular
structure in which communities of densely interconnected nodes are only sparsely
connected with nodes of other sub-networks (Sporns et al., 2004). This structure
promotes specialized processing and minimizes ‘wiring costs’ (Bullmore et al., 2012).
High degree nodes or ‘connector hubs’ mediate communication between the relatively
segregated modules and thus play a significant role in integrating information. Connector
hubs have a higher number of connections with other nodes than one would expect by
chance and damage to these key nodes significantly changes the balance in community
structure (Guimerà, & Amaral, 2005).
Recently, complex network analysis using graph metrics have led to new insights in
neuropsychiatric and neurodegenerative disorders such as Alzheimer’s disease (Phillips
et al., 2015; Vecchio et al., 2015), schizophrenia (van den Heuvel et al., 2013; van den
Heuvel et al., 2010; Zalesky et al., 2011), multiple sclerosis (Li et al., 2013; Shu et al.,

20
2011) and epilepsy (Bonilha et al., 2012; Liu et al., 2014; Otte et al., 2012). Notably, a
meta-analysis of grey matter lesions across 26 neurological disorders showed that on
average, connections central to the brain's infrastructure were disproportionally affected
(Crossley et al., 2014). This finding was replicated in a recent meta-analysis on macro-
scale white matter pathways, showing that brain hubs were involved in the pathology of
twelve neurological and psychiatric disorders (de Lange, 2018). Connections critical for
integration and global network communication were particularly affected across
disorders. The basal ganglia are part of a fundamental circuit that supports large-scale
network integration. Given their known substantial involvement in Parkinson’s disease,
this condition could also be described as a disconnection disorder. Additionally, recent
work has shown an important role for the noradrenergic locus coeruleus in network
integration (Shine et al., 2018). The locus coeruleus is a small brainstem structure that
has been documented to show significant cell death already in the early stages of
Parkinson’s disease (Gesi et al., 2000).
However, to date, relatively few studies have investigated the structural network topology
in Parkinson’s disease. These studies observed an unaffected modular structure, but a
reduced white matter connectivity in frontoparietal-striatal nodes in Parkinson’s disease
patients compared to healthy controls (Tinaz et al., 2017). Nodes within these superior
frontal and parietal regions, as well as in the anterior and posterior cingulate cortices and
insula are identified as connector hubs, and it is possible that the selective damage to
these regions cause a range of symptoms in Parkinson’s disease. Furthermore, reduced
global and local efficiency was observed when exploring changes in network topology
(Li et al., 2017; Wen et al., 2017), whilst whole brain analysis using TBSS yielded
negative results in the same individuals (Wen et al., 2017). These findings provide
evidence for the notion that rather than a focalised lesion, Parkinson’s disease patients
show a reorganisation of white matter network architecture. Indeed, the multitude of
potentially altered individual connections, especially in a heterogeneous population,
imposes significant methodological and statistical restrains. Utilizing a broader network
dysfunction model could circumvent these constraints and may also benefit the
neuromechanically explanation of Parkinson’s symptomatology.

21
Whilst previous research on network changes in Parkinson’s disease have mainly focused
on the burden of the disorder in general, this thesis will focus on two different symptoms,
namely freezing of gait and visual hallucinations. First, Parkinson’s disease is a
heterogeneous disorder and focusing on symptoms separately may give distinctive
insights into their pathophysiology. Freezing of gait and visual hallucinations are likely
part of the same clinical phenotype. Furthermore, as mentioned above, both symptoms
are of episodic nature and can be triggered by physiological and environmental factors.
Therefore both freezing and hallucinations may result from a diffuse, network-level
dysfunction (Lewis & Shine, 2016).

22
1.7 Aims of the thesis
Aim 1: Conduct an extensive literature review to explore the contribution of changes in
diffusivity to Parkinson’s disease.
The first aim of this thesis was to conduct a thorough review of all relevant
literature to provide an overview of the current understanding of white matter
alterations to different symptoms of Parkinson’s disease. Chapter 2 discusses
empirical studies to date using DTI to investigate different motor symptoms such
as bradykinesia, tremor and freezing of gait, cognition, including attention,
executive function, mild cognitive impairment and dementia, mood and sleep
disturbances.
Aim 2: To investigate whether freezing of gait could be a result to ineffective
communication between brain networks (Chapter 3 and Chapter 4).
The pathophysiology of freezing of gait is still poorly understood, but previous
work has shown an association between freezing behaviour and executive
dysfunction. In order to expand our knowledge of this symptom, Chapter 3 aimed
to investigate the prevalence of freezing of gait across the clinical motor stages.
Furthermore, Chapter 3 provides an in-depth comparison of the clinical
phenotype of patients with and without freezing of gait. It was hypothesized that
freezing of gait is a multifaceted symptom rather than a sole gait disturbance. As
such, we expected to show impairments on variables related to frontostriatal
dysfunctions, rather than on variables related to brainstem pathology in
Parkinson’s disease patients with freezing of gait.
Furthermore, the contribution of white matter lesions to freezing of gait is poorly
understood. In Chapter 4 of this thesis, I aimed to investigate the topological
properties of the white matter connectome of Parkinson’s disease patients with
and without freezing of gait. It was hypothesized that Parkinson’s disease patients
exhibit changes in structural network topology, especially in frontoparietal and

23
subcortical networks. Key information processing may be affected in freezers,
which can hamper effective communication between brain networks.
Aim 3: To investigate whether visual hallucinations in Parkinson’s disease arise due to a
reweighting of attentional and perceptual brain networks (Chapter 5 and Chapter
6).
Previous work has associated visual hallucinations with attentional dysfunction.
Chapter 5 aimed to further explore attentional processing deficits in Parkinson’s
disease patients with visual hallucinations using an objective attentional network
test that can entangle different attentional processes. It was hypothesized that a
disruption within and between attentional networks, as previously been shown in
Parkinson’s disease patients with visual hallucinations, would lead to a deficit in
the overall performance of the task.
Finally, Chapter 6 aimed to investigate changes in structural network topology
associated with severity of hallucinatory behaviour in Parkinson’s disease patient.
Specifically, Chapter 6 of this thesis tested the hypothesis that visual
hallucinations arise due to a reweighting of the perceptual processing system.
Changes in connectivity could indicate inefficient integration of different
perceptual sites, which would underlie inadequate interpretation of visual stimuli.

24
References
Aarts, E., van Holstein, M., & Cools, R. (2011). Striatal Dopamine and the Interface
between Motivation and Cognition. Front Psychol, 2(163).
doi:10.3389/fpsyg.2011.00163
Ackermann, H., Gröne, B., Hoch, G., & Schönle, P. (1993). Speech freezing in
Parkinson’s disease: A kinematic analysis of orofacial movements by means of
electromagnetic articulography. Folia Phoniatr Logop, 45(2), 84-89.
Alexander, A. L., Lee, J. E., Lazar, M., & Field, A. S. (2007). Diffusion tensor imaging
of the brain. Neurotherapeutics, 4(3), 316-329. doi:10.1016/j.nurt.2007.05.011
Amar, B. R., Yadav, R., Janardhan Reddy, Y. C., & Pal, P. K. (2014). A clinical profile
of patients with Parkinson’s disease and psychosis. Ann Indian Acad Neurol,
17(2), 187–192. http://doi.org/10.4103/0972-2327.132625
Amboni, M., Cozzolino, A., Longo, K., Picillo, M., & Barone, P. (2008). Freezing of gait
and executive functions in patients with Parkinson's disease. Mov Disord, 23(3),
395-400. doi:10.1002/mds.21850
Archibald, N. K., Clarke, M. P., Mosimann, U. P., & Burn, D. J. (2011). Visual
symptoms in Parkinson's disease and Parkinson's disease dementia. Mov Disord,
26(13), 2387-2395. doi:10.1002/mds.23891
Atkinson-Clement, C., Pinto, S., Eusebio, A., & Coulon, O. (2017). Diffusion tensor
imaging in Parkinson's disease: Review and meta-analysis. Neuroimage Clin, 16,
98-110. doi:10.1016/j.nicl.2017.07.011
Bassett, D. S., Bullmore, E. T., Meyer-Lindenberg, A., Apud, J. A., Weinberger, D. R., &
Coppola, R. (2009). Cognitive fitness of cost-efficient brain functional networks.
Proc Natl Acad Sci USA, 106(28), 11747-11752.
Bell, P. T., & Shine, J. M. (2016). Subcortical contributions to large-scale network
communication. Neurosci Biobehav Rev, 71, 313-322.
doi:10.1016/j.neubiorev.2016.08.036
Bella, S. D., Benoit, C. E., Farrugia, N., Keller, P. E., Obrig, H., Mainka, S., & Kotz, S.
A. (2017). Gait improvement via rhythmic stimulation in Parkinson's disease is
linked to rhythmic skills. Sci Rep, 7, 42005. doi:10.1038/srep42005

25
Berardelli, A., Rothwell, J., Thompson, P., & Hallett, M. (2001). Pathophysiology of
bradykinesia in Parkinson's disease. Brain, 124(11), 2131-2146.
Boecker, H., Ceballos-Baumann, A. O., Volk, D., Conrad, B., Forstl, H., &
Haussermann, P. (2007). Metabolic alterations in patients with Parkinson disease
and visual hallucinations. Arch Neurol, 64(7), 984-988.
doi:10.1001/archneur.64.7.984
Bonilha, L., Nesland, T., Martz, G. U., Joseph, J. E., Spampinato, M. V., Edwards, J. C.,
& Tabesh, A. (2012). Medial temporal lobe epilepsy is associated with neuronal
fibre loss and paradoxical increase in structural connectivity of limbic structures.
J Neurol, Neurosurg Psychiatry, 83(9), 903-909. doi:10.1136/jnnp-2012-302476
Braak, H., Del Tredici, K., Rub, U., de Vos, R. A., Jansen Steur, E. N., & Braak, E.
(2003). Staging of brain pathology related to sporadic Parkinson's disease.
Neurobiol Aging, 24(2), 197-211.
Braak, H., Sandmann-Keil, D., Gai, W. P., & Braak, E. (1999). Extensive axonal Lewy
neurites in Parkinson's disease: a novel pathological feature revealed by alpha-
synuclein immunocytochemistry. Neurosci Let, 265(1), 67-69. doi:Doi
10.1016/S0304-3940(99)00208-6
Budde, M. D., Janes, L., Gold, E., Turtzo, L. C., & Frank, J. A. (2011). The contribution
of gliosis to diffusion tensor anisotropy and tractography following traumatic
brain injury: validation in the rat using Fourier analysis of stained tissue sections.
Brain, 134(8), 2248-2260. doi:10.1093/brain/awr161
Bullmore, E., & Sporns, O. (2012). The economy of brain network organization. Nat Rev
Neurosci, 13(5), 336-349. doi:10.1038/nrn3214
Burke, R. E., & O'Malley, K. (2013). Axon degeneration in Parkinson's disease.
Experimental Neurology, 246, 72-83. doi:10.1016/j.expneurol.2012.01.011
Caligiore, D., Helmich, R. C., Hallett, M., Moustafa, A. A., Timmermann, L., Toni, I., &
Baldassarre, G. (2016). Parkinson’s disease as a system-level disorder. npj
Parkinson's Dis, 2, 16025.
Chudler, E. H., Sugiyama, K., & Dong, W. K. (1995). Multisensory convergence and
integration in the neostriatum and globus pallidus of the rat. Brain Res, 674(1),
33-45.

26
Collerton, D., Perry, E., & McKeith, I. (2005). Why people see things that are not there: a
novel Perception and Attention Deficit model for recurrent complex visual
hallucinations. Behav Brain Sci, 28(6), 737-757; discussion 757-794.
doi:10.1017/S0140525X05000130
Conturo T.E., L. N. F., Cull T.S., Akbudak E., Snyder A.Z., Shimony J.S., McKinstry, R.
C., Burton, H., Raichle M.E. (1999). Tracking neuronal fiber pathways in the
living human brain. Proc Natl Acad Sci USA , 96(18), 10422–10427.
Cowie, D., Limousin, P., Peters, A., Hariz, M., & Day, B. L. (2012). Doorway-provoked
freezing of gait in Parkinson's disease. Mov Disord, 27(4), 492-499.
doi:10.1002/mds.23990
Crossley, N. A., Mechelli, A., Scott, J., Carletti, F., Fox, P. T., McGuire, P., & Bullmore,
E. T. (2014). The hubs of the human connectome are generally implicated in the
anatomy of brain disorders. Brain 137(Pt 8), 2382-95.
de Lange, S., Scholtens, L. H., van den Berg, L. H., Boks, M. P. Bozzali, M., Cahn, W.
…, the Alzheimer's Disease Neuroimaging Initiative (ADNI) (2018). Shared
vulnerability for connectome alterations across psychiatric and neurological brain
disorders. bioRxiv 360586; doi: https://doi.org/10.1101/360586
de Souza Fortaleza, A. C., Mancini, M., Carlson-Kuhta, P., King, L. A., Nutt, J. G.,
Chagas, E. F., . . . Horak, F. B. (2017). Dual task interference on postural sway,
postural transitions and gait in people with Parkinson's disease and freezing of
gait. Gait Posture, 56, 76-81. doi:10.1016/j.gaitpost.2017.05.006
Dickson, D. W., Braak, H., Duda, J. E., Duyckaerts, C., Gasser, T., Halliday, G. M., …
Litvan I. (2009). Neuropathological assessment of Parkinson's disease: refining
the diagnostic criteria. Lancet Neurol, 8(12):1150–1157.
Dorsey, E. R., Constantinescu, R., Thompson, J. P., Biglan, K. M., Holloway, R. G.,
Kieburtz, K., . . . Tanner, C. M. (2007). Projected number of people with
Parkinson disease in the most populous nations, 2005 through 2030. Neurology,
68(5), 384-386. doi:10.1212/01.wnl.0000247740.47667.03
Ehgoetz Martens, K. A., Ellard, C. G., & Almeida, Q. J. (2014a). A closer look at
mechanisms underlying perceptual differences in Parkinson's freezers and non-
freezers. Neuroscience, 274, 162-169. doi:10.1016/j.neuroscience.2014.05.022

27
Ehgoetz Martens, K. A., Ellard, C. G., & Almeida, Q. J. (2014b). Does anxiety cause
freezing of gait in Parkinson's disease? PLoS One, 9(9), e106561.
doi:10.1371/journal.pone.0106561
Einstein, A. (1956). Investigations on the Theory of the Brownian Movement: Courier
Corporation.
Fereshtehnejad, S. M., Zeighami, Y., Dagher, A., & Postuma, R. B. (2017). Clinical
criteria for subtyping Parkinson’s disease: biomarkers and longitudinal
progression. Brain, 140(7), 1959-1976. doi:10.1093/brain/awx118
ffytche, D. H., & Howard, R. J. (1999). The perceptual consequences of visual loss:
'positive' pathologies of vision. Brain, 122 ( Pt 7), 1247-1260.
Fields, R. D. (2008). White matter in learning, cognition and psychiatric disorders.
Trends Neurosci, 31(7), 361-370. doi:10.1016/j.tins.2008.04.001
Fields, R. D. (2010). Neuroscience. Change in the brain's white matter. Science,
330(6005), 768-769. doi:10.1126/science.1199139
Fling, B. W., Cohen, R. G., Mancini, M., Carpenter, S. D., Fair, D. A., Nutt, J. G., &
Horak, F. B. (2014). Functional Reorganization of the Locomotor Network in
Parkinson Patients with Freezing of Gait. PLoS One, 9(6). doi:ARTN e100291
10.1371/journal.pone.0100291
Frank, M. J. (2005). Dynamic dopamine modulation in the basal ganglia: a
neurocomputational account of cognitive deficits in medicated and nonmedicated
Parkinsonism. J Cogn Neurosci, 17(1), 51-72.
Gallagher, D. A., Parkkinen, L., O'Sullivan, S. S., Spratt, A., Shah, A., Davey, C. C., . . .
Schrag, A. (2011). Testing an aetiological model of visual hallucinations in
Parkinson's disease. Brain, 134(Pt 11), 3299-3309. doi:10.1093/brain/awr225
Garcia-Rill, E., Luster, B., D’Onofrio, S., Mahaffey, S., Bisagno, V., & Urbano, F. J.
(2015). Pedunculopontine arousal system physiology – Deep brain stimulation
(DBS). Sleep Sci, 8(3), 153-161. doi:10.1016/j.slsci.2015.09.001
Ghai, S., Ghai, I., Schmitz, G., & Effenberg, A. O. (2018). Effect of rhythmic auditory
cueing on parkinsonian gait: A systematic review and meta-analysis. Sci Rep,
8(1), 506. doi:10.1038/s41598-017-16232-5

28
Giladi, N., & Hausdorff, J. M. (2006). The role of mental function in the pathogenesis of
freezing of gait in Parkinson's disease. J Neurol Sci, 248(1-2), 173-176.
doi:10.1016/j.jns.2006.05.015
Goldman, J. G., & Postuma, R. (2014). Premotor and non-motor features of Parkinson’s
disease. Current Opinion in Neurology, 27(4), 434–441.
doi:10.1097/WCO.0000000000000112
Grabli, D., Karachi, C., Welter, M. L., Lau, B., Hirsch, E. C., Vidailhet, M., & Francois,
C. (2012). Normal and pathological gait: what we learn from Parkinson's disease.
J Neurol Neurosurg Psychiatry, 83(10), 979-985. doi:10.1136/jnnp-2012-302263
Gratwicke, J. P., & Foltynie, T. (2018). Early nucleus basalis of Meynert degeneration
predicts cognitive decline in Parkinson’s disease. Brain, 141(1), 7-10.
doi:10.1093/brain/awx333
Gray, P., & Hildebrand, K. (2000). Fall risk factors in Parkinson's disease. J Neurosci
Nurs, 32(4), 222-228.
Guimerà, R. & Amaral, L. A. (2005). Functional cartography of complex metabolic
networks. Nature, 433:895–900Hall, J. M., Shine, J. M., Walton, C. C., Gilat, M., Kamsma, Y. P., Naismith, S. L., &
Lewis, S. J. (2014). Early phenotypic differences between Parkinson's disease
patients with and without freezing of gait. Parkinsonism Relat Disord, 20(6), 604-
607. doi:10.1016/j.parkreldis.2014.02.028
Halliday, G. M., Leverenz, J. B., Schneider, J. S., & Adler, C. H. (2014). The
Neurobiological Basis of Cognitive Impairment in Parkinson’S Disease. Mov
Disord, 29(5), 634–650. http://doi.org/10.1002/mds.25857
Hely, M. A., Reid, W. G., Adena, M. A., Halliday, G. M., & Morris, J. G. (2008). The
Sydney multicenter study of Parkinson's disease: the inevitability of dementia at
20 years. Mov Disord, 23(6), 837-844. doi:10.1002/mds.21956
Iansek, R., & Danoudis, M. (2016). Freezing of Gait in Parkinson's Disease: Its
Pathophysiology and Pragmatic Approaches to Management. Mov Disord Clin
Pract, n/a-n/a. doi:10.1002/mdc3.12463

29
Jacobs, J. V., Nutt, J. G., Carlson-Kuhta, P., Stephens, M., & Horak, F. B. (2009). Knee
trembling during freezing of gait represents multiple anticipatory postural
adjustments. Exp Neurol, 215(2), 334-341. doi:10.1016/j.expneurol.2008.10.019
Le Bihan, D., Mangin, J. F., Poupon, C., Clark, C. A., Pappata, S., Molko N., Chabriat,
H. (2001). Diffusion tensor imaging: concepts and applications. J. Magn. Reson.
Imaging, 13, 534–546 doi:10.1002/jmri.1076
Lenka, A., Jhunjhunwala, K. R., Saini, J. Pal, P. K. (2015). Structural and functional
neuroimaging in patients with Parkinson's disease and visual hallucinations: A
critical review. Parkinsonism Rel Disord, 21(7): 683-691.
Lewis, S. J., & Barker, R. A. (2009). A pathophysiological model of freezing of gait in
Parkinson's disease. Parkinsonism Relat Disord, 15(5), 333-338.
doi:10.1016/j.parkreldis.2008.08.006
Lewis, S. J. G., Foltynie, T., Blackwell, A. D., Robbins, T. W., Owen, A. M., & Barker,
R. A. (2005). Heterogeneity of Parkinson's disease in the early clinical stages
using a data driven approach. J Neurol Neurosurg Psychiatry, 76(3), 343-348.
doi:10.1136/jnnp.2003.033530
Lewis, S. J., & Shine, J. M. (2016). The Next Step: A Common Neural Mechanism for
Freezing of Gait. Neuroscientist, 22(1), 72-82. doi:10.1177/1073858414559101
Lewis, S. J. G., & Barker, R. A. (2009). A pathophysiological model of freezing of gait in
Parkinson's disease. Parkinsonism Relat Disord, 15(5), 333-338.
doi:10.1016/j.parkreldis.2008.08.006
Lewy, F. H. (1912). Paralysis Agitans. I. Pathologische Anatomie. In Handbuch der
Neurologie. Edited by M. Lewandowski Berlin, Springer, pp 920-933Li, C., Huang, B., Zhang, R., Ma, Q., Yang, W., Wang, L., . . . Liu, L. (2017). Impaired
topological architecture of brain structural networks in idiopathic Parkinson’s
disease: a DTI study. Brain Imaging Behav, 11(1), 113-128.
Li, M., & Zhang, W. (2015). Oscillations in pedunculopontine nucleus in Parkinson’s
disease and its relationship with deep brain stimulation. Front Neural Circuits, 9,
47. doi:10.3389/fncir.2015.00047
Li, Y., Jewells, V., Kim, M., Chen, Y., Moon, A., Armao, D., . . . Shen, D. (2013).
Diffusion tensor imaging based network analysis detects alterations of

30
neuroconnectivity in patients with clinically early relapsing�remitting multiple
sclerosis. Hum Brain Mapp, 34(12), 3376-3391.
Lieberman, A. (2006). Are freezing of gait (FOG) and panic related? J Neurol Sci, 248(1-
2), 219-222. doi:10.1016/j.jns.2006.05.023
Liu, M., Chen, Z., Beaulieu, C., & Gross, D. W. (2014). Disrupted anatomic white matter
network in left mesial temporal lobe epilepsy. Epilepsia, 55(5), 674-682.
doi:10.1111/epi.12581
Loane, C., Politis, M., Kefalopoulou, Z., Valle-Guzman, N., Paul, G., Widner, H., . . .
Piccini, P. (2016). Aberrant nigral diffusion in Parkinson's disease: A longitudinal
diffusion tensor imaging study. Mov Disord, 31(7), 1020-1026.
doi:10.1002/mds.26606
Martin, W. R. W., Wieler, M., Gee, M., & Camicioli, R. (2009). Temporal lobe changes
in early, untreated Parkinson's disease. Mov Disord, 24(13), 1949-1954.
doi:10.1002/mds.22680
Matar, E., Shine, J. M., Naismith, S. L., & Lewis, S. J. (2013). Using virtual reality to
explore the role of conflict resolution and environmental salience in freezing of
gait in Parkinson's disease. Parkinsonism Relat Disord, 19(11), 937-942.
doi:10.1016/j.parkreldis.2013.06.002
Mazilu, S., Blanke, U., Calatroni, A., Gazit, E., Hausdorff, J. M., & Tröster, G. (2016).
The role of wrist-mounted inertial sensors in detecting gait freeze episodes in
Parkinson’s disease. Pervasive Mob Comput, 33, 1-16.
doi:10.1016/j.pmcj.2015.12.007
Middleton, F. A., & Strick, P. L. (2000). Basal ganglia and cerebellar loops: motor and
cognitive circuits. Brain Res Rev, 31(2), 236-250. doi:10.1016/S0165-
0173(99)00040-5
Middleton, F. A., & Strick, P. L. (2000). Basal ganglia output and cognition: evidence
from anatomical, behavioral, and clinical studies. Brain Cogn, 42(2), 183-200.
Mori S., Crain B. J., Chacko V. P., van Zijl P. C. (1999). Three-dimensional tracking of
axonal projections in the brain by magnetic resonance imaging. Ann. Neurol. 45,
265–269

31
Moustafa, A. A., Sherman, S. J., & Frank, M. J. (2008). A dopaminergic basis for
working memory, learning and attentional shifting in Parkinsonism.
Neuropsychologia, 46(13), 3144-3156.
doi:10.1016/j.neuropsychologia.2008.07.011
Muller, A. J., Shine, J. M., Halliday, G. M., Lewis, S. J. G. (2014). Visual hallucinations
in Parkinson's disease: theoretical models. Mov Disord, 29(13); 1591-8. doi:
10.1002/mds.26004
Nagy, A., Eordegh, G., Paroczy, Z., Markus, Z., & Benedek, G. (2006). Multisensory
integration in the basal ganglia. Eur J Neurosci, 24(3), 917-924.
doi:10.1111/j.1460-9568.2006.04942.x
Naismith, S. L., Shine, J. M., & Lewis, S. J. (2010). The specific contributions of set-
shifting to freezing of gait in Parkinson's disease. Mov Disord, 25(8), 1000-1004.
doi:10.1002/mds.23005
Nieuwboer, A. (2008). Cueing for freezing of gait in patients with Parkinson's disease: a
rehabilitation perspective. Mov Disord, 23 Suppl 2, S475-481.
doi:10.1002/mds.21978
Nieuwboer, A., & Giladi, N. (2013). Characterizing freezing of gait in Parkinson's
disease: models of an episodic phenomenon. Mov Disord, 28(11), 1509-1519.
doi:10.1002/mds.25683
Nieuwboer, A., Vercruysse, S., Feys, P., Levin, O., Spildooren, J., & Swinnen, S. (2009).
Upper limb movement interruptions are correlated to freezing of gait in
Parkinson’s disease. Eur J Neurosci, 29(7), 1422-1430.
Nutt, J. G., Bloem, B. R., Giladi, N., Hallett, M., Horak, F. B., & Nieuwboer, A. (2011).
Freezing of gait: moving forward on a mysterious clinical phenomenon. Lancet
Neurol, 10(8), 734-744. doi:10.1016/S1474-4422(11)70143-0
Ofori, E., Pasternak, O., Planetta, P. J., Li, H., Burciu, R. G., Snyder, A. F., . . .
Vaillancourt, D. E. (2015). Longitudinal changes in free-water within the
substantia nigra of Parkinson’s disease. Brain, 138(8), 2322-2331.
doi:10.1093/brain/awv136
Ohama E. & Ikuta F. (1976). Parkinson's disease. Distribution of Lewy bodies and
monoamine neurons system. Acta Neuropathol 34: 311–319.

32
O'Malley, K.L. (2010). The role of axonopathy in Parkinson's disease. Exp. Neurobiol,
19(3):115–119. DOI: 10.5607/en.2010.19.3.115
Otte, W. M., Dijkhuizen, R. M., van Meer, M. P. A., van der Hel, W. S., Verlinde, S. A.
M. W., van Nieuwenhuizen, O., . . . Braun, K. P. J. (2012). Characterization of
Functional and Structural Integrity in Experimental Focal Epilepsy: Reduced
Network Efficiency Coincides with White Matter Changes. PLoS One, 7(7),
e39078. doi:10.1371/journal.pone.0039078
Parker, G. J., Haroon, H. A., Wheeler-Kingshott, C. A. (2003). A framework for a
streamline-based probabilistic index of connectivity (PICo) using a structural
interpretation of MRI diffusion measurements. J Magn Reson Imaging, 18:242–
54
Parkinson, J. (1817). An essay on the shaking palsy. London, United Kingdom:
Whittingham and Rowland
Phillips, D. J., McGlaughlin, A., Ruth, D., Jager, L. R., & Soldan, A. (2015). Graph
theoretic analysis of structural connectivity across the spectrum of Alzheimer's
disease: The importance of graph creation methods. NeuroImage: Clinical, 7,
377-390. doi:10.1016/j.nicl.2015.01.007
Plotnik, M., Giladi, N., & Hausdorff, J. M. (2012). Is Freezing of Gait in Parkinson's
Disease a Result of Multiple Gait Impairments? Implications for Treatment.
Parkinson’s Disease, 2012, 8. doi:10.1155/2012/459321
Postuma, R. B., Berg, D., Stern, M., Poewe, W., Olanow, C. W., Oertel, W., . . . Deuschl,
G. (2015). MDS clinical diagnostic criteria for Parkinson's disease. Movement
Disorders, 30(12), 1591-1601. doi:10.1002/mds.26424
Price, C. C., Tanner, J., Nguyen, P. T., Schwab, N. A., Mitchell, S., Slonena, E., . . .
Bowers, D. (2016). Gray and White Matter Contributions to Cognitive
Frontostriatal Deficits in Non-Demented Parkinson's Disease. PLoS One, 11(1),
e0147332. doi:10.1371/journal.pone.0147332
Rahman, S., Griffin, H. J., Quinn, N. P., & Jahanshahi, M. (2008). The factors that induce
or overcome freezing of gait in Parkinson's disease. Behav Neurol, 19(3), 127-
136.

33
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J., Gusnard, D. A., Shulman,
G. L. (2001). A default mode of brain function. Proc Natl Acad Sci U S A, 98(2)
676-682; doi:10.1073/pnas.98.2.676
Ramírez-Ruiz, B., Martí, M. J., Tolosa, E., Giménez, M., Bargalló, N, Valldeoriola, F.,
Junqué, C. (2007). Cerebral atrophy in Parkinson's disease patients with visual
hallucinations. Eur J Neurol, 14; 750-756.
Rektor, I., Svátková, A., Vojtíšek, L., Zikmundová, I., Vaníček, J., Király, A., & Szabó,
N. (2018). White matter alterations in Parkinson’s disease with normal cognition
precede grey matter atrophy. PLoS One, 13(1), e0187939.
doi:10.1371/journal.pone.0187939
Rolheiser, T. M., Fulton, H. G., Good, K. P., Fisk, J. D., McKelvey, J. R., Scherfler, C., .
. . Robertson, H. A. (2011). Diffusion tensor imaging and olfactory identification
testing in early-stage Parkinson's disease. J Neurol, 258(7), 1254-1260.
doi:10.1007/s00415-011-5915-2
Sanchez-Ramos, J. R., Ortoll, R., & Paulson, G. W. (1996). Visual hallucinations
associated with Parkinson disease. Arch Neurol, 53(12), 1265-1268.
Schrag, A., Jahanshahi, M., & Quinn, N. (2000). What contributes to quality of life in
patients with Parkinson's disease? J Neurol Neurosurg Psychiatry, 69(3),
308.
Shine, J. M. (2013). Attentional Network Dysfunction in Parkinson’s Disease. (PhD), The
University of Sydney, Sydney.
Shine, J. M., van den Brink, R. L., Hernaus, D., Nieuwenhuis, S., & Poldrack, R. A.
(2018). Catecholaminergic manipulation alters dynamic network topology across
cognitive states. Network neuroscience 2(3), 381-396. doi:10.1162/netn_a_00042
Shine, J. M., Halliday, G. M., Gilat, M., Matar, E., Bolitho, S. J., Carlos, M., . . . Lewis,
S. J. (2014). The role of dysfunctional attentional control networks in visual
misperceptions in Parkinson's disease. Hum Brain Mapp, 35(5), 2206-2219.
doi:10.1002/hbm.22321
Shine, J. M., Halliday, G. M., Naismith, S. L., & Lewis, S. J. G. (2011). Visual
Misperceptions and Hallucinations in Parkinson's Disease: Dysfunction of

34
Attentional Control Networks? Mov Disord, 26(12), 2154-2159.
doi:10.1002/mds.23896
Shine, J. M., Matar, E., Ward, P. B., Bolitho, S. J., Gilat, M., Pearson, M., . . . Lewis, S.
J. (2013a). Exploring the cortical and subcortical functional magnetic resonance
imaging changes associated with freezing in Parkinson's disease. Brain, 136(Pt 4),
1204-1215. doi:10.1093/brain/awt049
Shine, J. M., Matar, E., Ward, P. B., Bolitho, S. J., Gilat, M., Pearson, M., . . . Lewis, S.
J. G. (2013b). Exploring the cortical and subcortical fMRI changes associated
with freezing in Parkinson's disease. Brain, 136(4), 1204-1215.
Shine, J. M., Matar, E., Ward, P. B., Frank, M. J., Moustafa, A. A., Pearson, M., . . .
Lewis, S. J. (2013). Freezing of gait in Parkinson's disease is associated with
functional decoupling between the cognitive control network and the basal
ganglia. Brain, 136(Pt 12), 3671-3681. doi:10.1093/brain/awt272
Shine, J. M., Moore, S. T., Bolitho, S. J., Morris, T. R., Dilda, V., Naismith, S. L., &
Lewis, S. J. (2012). Assessing the utility of Freezing of Gait Questionnaires in
Parkinson's Disease. Parkinsonism Relat Disord, 18(1), 25-29.
doi:10.1016/j.parkreldis.2011.08.002
Shine, J. M., Muller, A. J., O'Callaghan, C., Hornberger, M., Halliday, G. M., & Lewis,
S. J. (2015). Abnormal connectivity between the default mode and the visual
system underlies the manifestation of visual hallucinations in Parkinson's disease:
a task-based fMRI study. NPJ Parkinsons Dis, 1, 15003.
doi:10.1038/npjparkd.2015.3
Shu, N., Liu, Y., Li, K., Duan, Y., Wang, J., Yu, C., . . . He, Y. (2011). Diffusion tensor
tractography reveals disrupted topological efficiency in white matter structural
networks in multiple sclerosis. Cereb Cortex, 21(11), 2565-2577.
Smith, S. M., Jenkinson, M., Johansen-Berg, H., Rueckert, D., Nichols, T. E., Mackay, C.
E., . . . Behrens, T. E. (2006). Tract-based spatial statistics: voxelwise analysis of
multi-subject diffusion data. Neuroimage, 31(4), 1487-1505.
doi:10.1016/j.neuroimage.2006.02.024

35
Snijders, A. H., Takakusaki, K., Debu, B., Lozano, A. M., Krishna, V., Fasano, A., . . .
Hallett, M. (2016). Physiology of freezing of gait. Ann Neurol, 80(5), 644-659.
doi:10.1002/ana.24778
Spildooren, J., Vercruysse, S., Desloovere, K., Vandenberghe, W., Kerckhofs, E., &
Nieuwboer, A. (2010). Freezing of Gait in Parkinson's Disease: The Impact of
Dual-Tasking and Turning. Mov Disord, 25(15), 2563-2570.
doi:10.1002/mds.23327
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C. (2004). Organization,
development and function of complex brain networks. Trends Cogn Sci, 8(9),
418-425. doi:10.1016/j.tics.2004.07.008
Squire, L. R., & Zola-Morgan, S. (1991). The medial temporal lobe memory system.
Science, 253(5026), 1380-1386.
Takakusaki, K., Habaguchi, T., Ohtinata-Sugimoto, J., Saitoh, K., & Sakamoto, T.
(2003). Basal ganglia efferents to the brainstem centers controlling postural
muscle tone and locomotion: a new concept for understanding motor disorders in
basal ganglia dysfunction. Neurosci, 119(1), 293-308.
doi:https://doi.org/10.1016/S0306-4522(03)00095-2
Tessitore, A., Amboni, M., Esposito, F., Russo, A., Picillo, M., Marcuccio, L., . . .
Barone, P. (2012). Resting-state brain connectivity in patients with Parkinson's
disease and freezing of gait. Parkinsonism Relat Disord, 18(6), 781-787.
doi:10.1016/j.parkreldis.2012.03.018
Tinaz, S., Lauro, P. M., Ghosh, P., Lungu, C., & Horovitz, S. G. (2017). Changes in
functional organization and white matter integrity in the connectome in
Parkinson's disease. NeuroImage Clin, 13, 395-404.
doi:10.1016/j.nicl.2016.12.019
Trapp, S. and Bar, M. (2015), Prediction, context, and competition in visual recognition.
Ann. N.Y. Acad. Sci., 1339: 190-198. doi:10.1111/nyas.12680
Tsang, E. W., Hamani, C., Moro, E., Mazzella, F., Poon, Y. Y., Lozano, A. M., & Chen,
R. (2010). Involvement of the human pedunculopontine nucleus region in
voluntary movements(e–Pub ahead of print). Neurology, 75(11), 950-959.
doi:10.1212/WNL.0b013e3181f25b35

36
Vaillancourt, D. E., Spraker, M. B., Prodoehl, J., Abraham, I., Corcos, D. M., Zhou, X. J.,
. . . Little, D. M. (2009). High-resolution diffusion tensor imaging in the
substantia nigra of de novo Parkinson disease. Neurology, 72(16), 1378-1384.
doi:10.1212/01.wnl.0000340982.01727.6e
van den Heuvel, M. P., Mandl, R. C., Stam, C. J., Kahn, R. S., & Pol, H. E. H. (2010).
Aberrant frontal and temporal complex network structure in schizophrenia: a
graph theoretical analysis. J Neurosci, 30(47), 15915-15926.
van den Heuvel, M. P., Sporns, O., Collin, G., Scheewe, T., Mandl, R. C., Cahn, W., . . .
Kahn, R. S. (2013). Abnormal rich club organization and functional brain
dynamics in schizophrenia. JAMA Psychiatry, 70(8), 783-792.
doi:10.1001/jamapsychiatry.2013.1328
van den Heuvel, M. P., Stam, C. J., Kahn, R. S., & Pol, H. E. H. (2009). Efficiency of
functional brain networks and intellectual performance. J Neurosci, 29(23), 7619-
7624.
van Swieten, G. (1742). commentaria in Hermanni Boer|haavii aphorismos de
cognoscendis et curandis morbis (Vol. 5): Lugduni Batavorum.
Vandenbossche, J., Deroost, N., Soetens, E., Coomans, D., Spildooren, J., Vercruysse, S.,
. . . Kerckhofs, E. (2013). Freezing of gait in Parkinson's disease: disturbances in
automaticity and control. Front Hum Neurosci, 6. doi:ARTN 356
10.3389/fnhum.2012.00356
Vecchio, F., Miraglia, F., Curcio, G., Altavilla, R., Scrascia, F., Giambattistelli, F., . . .
Rossini, P. M. (2015). Cortical brain connectivity evaluated by graph theory in
dementia: a correlation study between functional and structural data. J Alzheimers
Dis, 45(3), 745-756. doi:10.3233/jad-142484
Walton, C. C., Shine, J. M., Hall, J. M., O'Callaghan, C., Mowszowski, L., Gilat, M., . . .
Lewis, S. J. (2015). The major impact of freezing of gait on quality of life in
Parkinson's disease. J Neurol, 262(1), 108-115. doi:10.1007/s00415-014-7524-3
Wang, H., Gao, H., Jiao, T., & Luo, Z. (2016). A meta-analysis of the pedunculopontine
nucleus deep-brain stimulation effects on Parkinson’s disease. Neurorep, 27(18),
1336-1344.

37
Weil, R. S., Schrag, A. E., Warren, J. D., Crutch, S. J., Lees, A. J., & Morris, H. R.
(2016). Visual dysfunction in Parkinson’s disease. Brain, 139(11), 2827-2843.
doi:10.1093/brain/aww175
Wen, M. C., Xu, Z., Lu, Z., Chan, L. L., Tan, E. K., & Tan, L. C. S. (2017).
Microstructural network alterations of olfactory dysfunction in newly diagnosed
Parkinson's disease. Sci Rep, 7(1), 12559. doi:10.1038/s41598-017-12947-7
Zalesky, A., Fornito, A., Seal, M. L., Cocchi, L., Westin, C.-F., Bullmore, E. T., . . .
Pantelis, C. (2011). Disrupted axonal fiber connectivity in schizophrenia. Biol
Psychiatry, 69(1), 80-89.
Zhang, Y., Wu, I. W., Tosun, D., Foster, E., Schuff, N., & the Parkinson’s Progression
Markers, I. (2016). Progression of Regional Microstructural Degeneration in
Parkinson’s Disease: A Multicenter Diffusion Tensor Imaging Study. PLoS One,
11(10), e0165540. doi:10.1371/journal.pone.0165540

CHAPTER2DIFFUSIONALTERATIONSASSOCIATEDWITH
PARKINSON’SDISEASESYMPTOMATOLOGY:AREVIEW
OFTHELITERATURE
This study is published in Parkinsonism and Related Disorders
Hall, J. M., Ehgoetz Martens, K. A., Walton, C. C., O’Callaghan, C., Keller, P. E., Lewis, S. J. G., Moustafa, A. A. Diffusion alterations associated with Parkinson's disease
symptomatology: A review of the literature. Parkinsonism and Related Disorders, 2016. DOI:10.1016/j.parkreldis.2016.09.026

39
Chapter 2: Diffusion alterations associated with Parkinson’s disease
symptomatology: A review of the literature
Abstract
Parkinson’s disease (PD) is a heterogeneous neurological disorder with a variety of motor
and non-motor symptoms. The underlying mechanisms of these symptoms are not fully
understood. An increased interest in structural connectivity analyses using diffusion
tensor imaging (DTI) in PD has led to an expansion of our understanding of the impact of
abnormalities in diffusivity on phenotype. This review outlines the contribution of these
abnormalities to symptoms of PD including bradykinesia, tremor and non-tremor
phenotypes, freezing of gait, cognitive impairment, mood, sleep disturbances, visual
hallucinations and olfactory dysfunction. Studies have shown that impairments in
cognitive functioning are related to diffusion abnormalities in frontal and parietal regions,
as well as in the corpus callosum and major fibres connecting midbrain and subcortical
structures with the neocortex. However, the impact of diffusion alterations on motor,
mood and other symptoms of PD are less well understood. The findings presented here
highlight the challenges faced and the potential areas of future research avenues where
DTI may be beneficial. Larger cohort studies and standardized imaging protocols are
required to investigate current promising preliminary findings.
Key words: Parkinson’s disease; white matter tracts; diffusion tensor imaging;
heterogeneity
Abbreviations: PD = Parkinson’s disease; DTI = diffusion tensor imaging; FA =
fractional anisotropy; MD = mean diffusivity; TBSS = tract-based spatial statistics; ROI
= region of interest; SN = substantia nigra; SLF = superior longitudinal fasciculus; IF-OF
= inferior fronto-occipital fasciculus; PPN = pedunculopontine nucleus; PIGD = postural
instability and gait disorder; FOG = freezing of gait; MCI = mild cognitive impairment;
VH = visual hallucinations; RBD = REM sleep behaviour disorder.

40
Introduction
Parkinson’s disease (PD) is classified as a movement disorder with its cardinal motor
symptoms of bradykinesia, muscle rigidity and resting tremor predominantly arising from
nigrostriatal dopaminergic denervation. However, it has become evident that PD is a
multisystem disorder extending beyond the loss of dopamine (Langston, 2006). Indeed,
PD can be better characterised as a heterogeneous disorder with a wide variety of both
motor and non-motor symptoms, including cognitive impairments, sleep-wake
disturbances, mood disorders, autonomic dysfunction and visual hallucinations
(Chaudhuri et al., 2006). The complexity of this disorder is emphasised by its clinical
diversity with symptoms differing greatly across patients (Lewis et al., 2005). This
degree of heterogeneity has hampered the understanding of the neural underpinnings of
each symptom, leaving many symptoms poorly managed or even unable to be treated. In
an attempt to mitigate these issues, magnetic resonance imaging (MRI) techniques have
been used to investigate tissue alteration and abnormalities that may be present in order to
provide a more thorough understanding of the neural basis underlying symptomology in
PD (Goveas et al., 2015; Weingarten et al., 2015).
One such structural imaging technique, diffusion tensor imaging (DTI) is a non-invasive
imaging approach that analyses the diffusivity in brain tissues. It is sensitive to the flow
of water molecules, and as water predominantly diffuses along axons, DTI can be used to
infer white matter tracts throughout the brain (Le Bihan, 1985; Merboldt et al., 1985).
The organisation of axons and their properties (i.e. axonal ordering, diameter, density,
myelination, microstructural complexity and white matter integrity changes) are the main
contributors that influence the flow of water molecules along neural pathways. Water
molecules move much faster along parallel white matter fibres compared to perpendicular
fibres. Fractional anisotropy (FA) is a measure that reflects the degree of diffusivity in
the different directions and can inform understanding of the microstructural organization
of the tensors; an organized multidimensional array of numerical values, expressing a
relationship between vectors (Basser et al., 1996). A decrease in FA values may arise
from demyelination, axonal loss or changes in the size of axons. Mean diffusivity (MD)
represents the overall movement of water molecules within the brain. Structurally intact
white matter has high FA and low MD, whereas structurally compromised white matter

41
has low FA and high MD. Additionally, DTI provides an estimate of diffusivity along
(axial diffusivity; AxD) and perpendicular (radial diffusivity; RxD) to the main fibre
direction. RxD increases in response to demyelination (Song et al., 2005), whereas a
decrease in AxD is indicative of axonal damage (Song et al., 2003). Grey matter
abnormalities can also be detected with DTI analyses. Anisotropy in grey matter is much
lower as the cellular membranes are not positioned along a preferential direction
(Pierpaoli et al., 1996). As such, MD, rather than FA, is an appropriate metric for cortical
and subcortical grey matter, and can inform about the possible breakdown of barriers to
diffusion, resulting in higher MD values. DTI can detect microscopic tissue abnormalities
and might be able to detect these changes earlier than conventional structural MRI
sequences. DTI data can be analysed using: i) tract based spatial statistics (TBSS)(Smith
et al., 2006) (white matter) and voxel-based analyses (VBA; white and grey matter),
which entail a whole-brain unbiased hypothesis-free approach, and/or ii) a probabilistic
tractography approach which estimates the most likely fibre orientation at each voxel
tracing a single tract (in contrast with streamline or deterministic tractography).
Probabilistic tractography is a valuable and informative technique for testing a priori
hypotheses.
Diffusion tensor MRI can detect alterations in white matter in the early stages of PD and
has therefore proven useful as a biomarker in our understanding of PD (Vaillancourt et
al., 2009). Lewy neurites, a pathological hallmark of PD, might affect axons and
dendrites in the brain and can be present from the early stages of PD (Kanazawa et al.,
2012). Lewy neurites might not only appear before Lewy bodies but they could extend
subsequently to develop Lewy bodies (Braak et al., 2003). White matter damage could
add to the PD pathophysiology by disrupting neuromodulator projection systems (Double
et al., 1996). Indeed, white matter atrophy has been shown to have a greater influence on
remote metabolism than grey matter alterations in Alzheimer’s disease (Villain et al.,
2010). Additionally, altered patterns of functional connectivity correlate with axonal
injury (Palacios et al., 2013; Sharp et al., 2011).
DTI analysis of white matter changes has been shown to successfully distinguish PD
patients from healthy controls (Vaillancourt et al., 2009) and from Parkinsonian

42
syndromes (Cochrane et al., 2013). Several studies using an a priori region of interest
(ROI) approach have confirmed the widespread nature of PD by reporting alterations in
diffusivity in subcortical structures as well as an association between reduced FA in the
substantia nigra (SN) pars compacta and increasing motor severity (Ofori et al., 2015;
Vaillancourt et al., 2009; Zhan et al., 2012).
Although these interesting results provide insight into PD pathology, it is important to
investigate the clinical diversity of this disorder in order to determine better treatment
options and predictions of progression. Therefore, the aim of this review is to provide an
overview of studies that have investigated the association between alterations in
diffusivity and PD motor symptoms, such as tremor, bradykinesia and freezing of gait
(FOG) as well as non-motor symptoms, including cognitive impairment, mood
disturbances, visual hallucinations (VH), REM sleep behaviour disorder (RBD) and
autonomic dysfunction. With the accumulated knowledge regarding the neural
underpinnings of such symptoms, a more complete understanding of the underlying
heterogeneity in PD will potentially result in improved symptom management and
highlight the challenges faced and the potential areas of future research avenues where
DTI may be beneficial.
Search Strategy
A literature search was carried out in September – December 2015 using the search
engines PubMed and Scopus using the criteria of English language and human studies
with no time scale restrictions. The following key words were used in the literature
searches: Parkinson's disease (MeSH); diffusion tensor imaging (MeSH), white matter;
fractional anisotropy, mean diffusivity, tractography. In total, 243 articles were found and
91 articles were selected based on titles and abstracts. Articles were excluded when they
were unrelated to the topic (e.g. focusing on Parkinsonism). The results were further
examined and articles were excluded if they used DTI to investigate anatomical
differences between PD and healthy controls (without relation to a specific PD symptom
(28)), anatomical studies of the progression of PD (6), to inform deep brain surgery (16),
or the influence of medication (1). Seven articles were added after scanning the reference

43
lists of the selected articles. A total of 47 articles met the criteria and were included in
this review. An overview of these results is provided in Table 1.
Figure 1. Flowchart of search strategy
Results
Motor correlates
Motor dysfunctions are typically the initial signs of PD that lead to diagnosis and the
administration of medication. The severity and progression of these motor symptoms can
be assessed by rating scales including the Unified Parkinson’s Disease Rating Scale
(UPDRS)(Goetz et al., 2008), the Hoehn and Yahr clinical motor scale (H&Y) (Hoehn &
Yahr, 1967) and scale Schwab and England (SE) scale (Schwab et al., 1968). Lenfeldt
and colleagues investigated the progression of alterations in diffusivity with motor

44
deterioration (Lenfeldt et al., 2013). They reported that worsening scores on the H&Y
and SE scale correlated with an increase in MD in the putamen, globus pallidus and
thalamus, while this correlation was not observed for the UPDRS (Lenfeldt et al., 2013).
However, these are measures of generalised motor functioning and more specific
symptoms have also been explored and will be discussed below. Nevertheless, these
results demonstrate that progression of diffusivity abnormalities are associated with
worsening of motor symptoms in PD.
To accommodate for the heterogeneity in the clinical symptoms, several motor subtypes
have been proposed that generally focus on features of tremor and non-tremor (Jankovic
et al., 1990; Lewis et al., 2005; Zetusky et al., 1985). A few studies have investigated
white matter pathology between the tremor dominant and postural instability and gait
difficulty (PIGD) subgroups. The tremor dominant phenotype is characterized by modest
motor symptoms other than tremor and relatively spared cognitive function (Lewis et al.,
2005). In contrast, patients in the PIGD or non-tremor dominant phenotype more often
experience executive dysfunction and mood disorders in addition to gait and balance
difficulties (Lewis et al., 2005). Differences in white matter integrity between these
subtypes are reported to be already present in the early stages of PD (Tessa et al., 2008).
For example, whole brain analyses have shown FA reductions in the bilateral superior
longitudinal fasciculus (SLF) in patients within the dominant PIGD phenotype compared
to tremor dominant patients (Gu et al., 2014). The SLF connects the parietal, temporal
and occipital lobes with ipsilateral frontal cortices and abnormalities within this tract
could impair communication between these circuits (Makris et al., 2005), which might
contribute to why patients with the PIGD phenotype have a greater severity and variety of
symptoms. This study identified the subgroups based on the PIGD/tremor ratio derived
from UPDRS scores. However, the PIGD score did not differ between the groups,
suggesting the categorisation into subgroups was primarily based on tremor severity (Gu
et al., 2014). Additionally, a region of interest analysis on diffusivity differences between
tremor dominant patients and patients who initially presented with predominant postural
instability, frequent falls, freezing and walking difficulties demonstrated reduced FA in
the corpus callosum tracts and increased MD values in the thalamus, putamen and corpus

45
callosum in PIGD (Chan et al., 2014). Furthermore, higher MD values in the thalamus,
corona radiata, centrum semiovale and the corpus callosum are shown in PIGD patients
compared to control participants. These findings are in keeping with the notion that PIGD
pathophysiology is widespread with greater involvement of the corticostriatal motor loop
and frontal areas compared to those patients with tremor dominant pathology (Lewis et
al., 2005). Future studies should also take non-motor symptoms into account when
classifying subtypes, as they form an important aspect of identifying phenotypes in PD.
Several studies have investigated the association between diffusivity changes and a
specific motor symptom, such as bradykinesia. Based on the notion that dopamine
depletion in the SNpc is implicated in slowness of movement, Ofori et al. (2015)
analysed ROI free-water imaging, a method for diffusion MRI which separately models
the contribution of extracellular free water and water that is in the vicinity of cellular
tissue. The authors found that an increase in free water values in the posterior SN could
predict the progression of bradykinesia over one year. Additionally, it has been shown
that frontal areas also contribute to bradykinesia (Lee et al., 2014). Lee and colleagues
(2014) investigated 12 de novo PD patients and found that bradykinesia negatively
correlated with FA values in the long association fibres connecting the frontal lobes to the
temporal, parietal, and occipital lobes (the SLF, the uncinate fasciculus and the inferior
fronto-occipital fasciculus (IF-OF)). The authors hypothesized that these regions are
unable to correct hypokinetic movements due to the dysfunction of the error detector
centre postulated to be within the anterior cingulate cortex (Carter et al., 1998; Lee et al.,
2014).
Diffusivity changes have further been associated with FOG, a sudden and transient gait
disturbance described as the inability to move forward despite the intention to walk
(Giladi et al., 1992). Studies investigating FOG focused predominantly on the brainstem
locomotor regions and found white matter abnormalities in the corticopontine and (right)
pontine-cerebellar tracts in freezers (Canu et al., 2015; Fling et al., 2013a; Schweder et
al., 2010; Vercruysse et al., 2015; Youn et al., 2015). The pedunculopontine nucleus
(PPN) is associated with gait regulation and posture (Fraix et al., 2013; Karachi et al.,

46
2010) and has frequently been identified as a key hub in this debilitating symptom (Nutt
et al., 2011). In addition to locomotor areas, the involvement of frontal regions in FOG
has become increasingly recognized (Shine et al., 2013a; Vercruysse et al., 2014). A
recent study found white matter abnormalities in the subcortico-cortical tracts
(Vercruysse et al., 2015). Whole brain analyses further demonstrated white matter
alterations within the major cortico-cortical, several frontostriatal tracts (see Table 1) as
well as in regions in the cerebellum in FOG patients compared to non-FOG patients
(Canu et al., 2015; Vercruysse et al., 2015). These results are in agreement with the
notion that complimentary yet competing frontostriatal pathways might induce FOG that
process motor, cognitive and limbic information (Lewis & Barker, 2009). However, the
authors note that the difference between freezers and non-freezers in the whole brain
analyses was partly driven by increased FA values in non-freezers compared to controls,
which may indicate a possible compensation mechanism.
These results emphasize the involvement of the long association fibres and frontal areas
in the manifestation of different motor symptoms, but also highlight the distinct neural
abnormalities underlying these symptoms in PD. However, more research is needed to
address the inconsistencies in the findings using objective measures to measure motor
performance.
Cognitive correlates
Cognitive impairment is one of the most common non-motor aspects of PD and manifests
in many domains such as attention, executive, visuospatial functioning and memory and
can progress to dementia (Svenningsson et al., 2012). White matter tracts are key to
efficient cognitive processing and changes in anisotropy have consistently been
associated with cognitive deficiencies.
The relationship between white matter damage and executive functions in PD has been
investigated in several studies where DTI combined with neuropsychological testing is
valuable for identifying correlations. Executive dysfunction has been associated with

47
widespread diffusion abnormalities in the left parietal and frontal regions (Auning et al.,
2014; Duncan et al., 2016; Gallagher et al., 2013; Koshimori et al., 2015; Matsui et al.,
2007b; Melzer et al., 2013; Rae et al., 2012; Theilmann et al., 2013; Zheng et al., 2014),
corpus callosum and major fibres connecting midbrain and subcortical structures with the
neocortex, such as the posterior thalamic radiation, anterior corona radiate and the
internal capsule, as well as regions within the striatum (Al Serafy et al., 2015; Koshimori
et al., 2015; Melzer et al., 2013; Zheng et al., 2014). Attentional capacity and language
have also been associated with changes in areas implicated in executive dysfunction, such
as in the major cortical tracts and thalamic radiation. However, unique patterns of
diffusivity have also be identified for these separate cognitive domains, such as changes
in the cingulate gyrus are associated with attention dysfunction (Zheng et al., 2014) and
Broca’s and Wernicke’s areas relating to speech impairment (Lee, 2011). Response
inhibition, as measured by NoGo errors, was not found to be associated with any FA
reductions (Ye et al., 2014). Short-term memory impairment correlates inversely with the
MD values within the fornix, whereas long-term memory performance inversely
correlates with MD in the right anterior corona radiate (Zheng et al., 2014). Evidently, the
white matter abnormalities as well as diffusivity changes in grey matter in cognitively
impaired PD patients are widespread in keeping with the heterogeneous nature of
cognitive impairments in PD.
Dementia in PD (PDD) is common in the advanced stages and estimated to affect 48-83%
of PD patients with disease duration over 15-20 years (Hely et al., 2005; 2008). Mild
cognitive impairment (MCI) is a transitional state between normal cognitive functioning
and dementia and several studies have investigated diffusion abnormalities in MCI and
PDD. Correlations between cognitive assessments (Montreal Cognitive Assessment) and
decreased FA values in major cortico-cortical tracts and hippocampus have been reported
in PD (Carlesimo et al., 2012; Chen et al., 2015). Furthermore, in patients with PD-MCI,
widespread FA reductions are also reported in the major cortical tracts (see Table 1), as
well as in the corpus callosum, anterior limb of internal capsule compared with healthy
controls. Compared to cognitively intact patients, one study reported diffusion alterations
in the corona radiata, corpus collosum, IF-OF and SLF in PD-MCI (Agosta et al., 2014).

48
However, Rossi and colleagues only found a tendency towards decreasing FA values in
MCI (Rossi et al., 2014) but also noted that younger age at disease onset was associated
with increased FA values in frontal and parietal lobes, as well as in the right temporal and
occipital lobes and thalamus. Compared to non-demented patients and healthy controls,
PDD patients show significantly lower FA values in bilateral posterior cingulate bundles
(Kamagata et al., 2012; Matsui et al., 2007b), bilateral anterior cingulate bundles
(Kamagata et al., 2012), corpus callosum splenium (Deng et al., 2013) and left
hippocampus (Chen et al., 2015). However, not all studies support these findings in
patients with mild PDD (Wiltshire et al., 2010). Nevertheless, cognitive impairments in
PD are generally shown to be associated with widespread white matter alterations,
predominantly in the frontal and parietal regions, corpus callosum and the subcortico-
cortical pathways, as well as with diffusivity changes within the hippocampus.
Mood disturbances
Mood disturbances are common in PD and are more frequently observed in patients
within the non-tremor dominant phenotype. Recently, it has even been suggested that
Parkinsonian phenotypes could be classified based on psychological well-being (Brown
et al., 2011; Landau et al., 2016). Though often experienced concurrently, it is now
understood that depression, anxiety and apathy are dissociable symptoms in PD (Brown
et al., 2011; Levy et al., 1998). The aetiology of these mood disturbances remains
unclear. In fact, to our knowledge, no studies have investigated anxiety using DTI and
only one study examined diffusivity changes in apathy, reporting no significant
abnormalities associated with this symptom (Carriere et al., 2014).
Additionally, only few reports with a limited number of participants have explored the
contribution of white matter abnormalities to depression in PD (Huang et al., 2014).
Damaged long association fibres and alterations in the left anterior thalamic radiation and
left forceps minor were reported in patients with depression. This indicates prominent
involvement of the neocortex, additionally to the locus coeruleus, which is thought to
play a key role in mood disorders (Chan-Palay et al., 1989). However, another study
reported no significant FA findings when looking at the uncinate fasciculus (Surdhar et

49
al., 2012), though this study only included six depressed patients. Furthermore, Li and
colleagues investigated the involvement of the thalamus in PD depression using whole
brain analyses and found decreased FA values in the bilateral medial dorsal thalamus
(van den Heuvel et al., 2010). Decreased FA in the thalamus may reflect a reduction in
either incoming or projecting white matter tracts. The medial dorsal thalamus is believed
to relay inputs from the amygdala and olfactory cortex to the frontal cortex and limbic
system (Tham et al., 2009). The medial dorsal thalamus receives numerous dopaminergic
projections (Garcia-Cabezas et al., 2009) and loss of dopamine is also found in the limbic
system in depressed patients (Remy et al., 2005). Finally, decreased FA values in the
anterior cingulate bundle have been reported (Matsui et al., 2007a). The anterior
cingulate is related to motivation, initiation of movement and apathy (Remy et al., 2005).
Moreover, emotion recognition, particularly identifying negative emotions might be
impaired in PD (Dujardin et al., 2004). FA levels in the portion of the right IF-OF
correlate positively to the ability to recognize sadness as a facial expression (Baggio et
al., 2012). Despite these interesting results, it is evident that more studies are needed to
elucidate the contribution of diffusion abnormalities to mood disturbances.

50
Table 1. An overview of the DTI literature in Parkinson’s disease
Study n Mean age ± SD
Field strength Tesla
No. of direc-tions
b-values s/mm2
Analyses methods
Symptom studied / contrast
Regions studied FA reductions MD (AD and RD)
increases
Statis-tical thres-hold, cor-rection
Motor correlates
Chan et al. 2014
46 PD (25 PIGD; 21 TD), 19 HC
PIGD: 72. ± 4.8; TD: 73.3 ± 6.2; HC: 71.5 ± 5.0
3 30 800 Tractography Subtype Whole brain analyses
PIGD < TD: CC body PIGD < controls: CC genu, body
PIGD > TD: Thal, PU, CC body, splenium and centrum semiovale; PIGD > controls: SN, Thal, corona radiata, centrum semiovale (CST), CC body, centrum semiovale; PD > controls: SN;
P < 0.05; logistic regression
Gu et al. 2014 24 PD (12 PIGD, 12 TD)
PIGD: 55.7 ± 8.1; TD: 56.0 ± 8.4
3 15 1000 VBA Subtype SLF, anterior corona radiata, genu of CC
Bilateral SLF
P < 0.05 FWE (cluster size >10 voxels)
Tessa et al. 2008
27 PD (de novo; 11 PIGD, 13 TD, 3 mixed), 16 HC
PIGD: 62.2 ± 8.3; TD: 57.9 ± 10.4; Mixed: 69.3 ± 7.7; HC: 59.4 ± 9.4
1.5 6 1000 Histogram analyses Subtype
Whole brain histograms FA and MD
PD and PIGD < controls: increase of the 25th percentile of FA histograms.
n.s.
P < 0.01

51
Lee et al. 2014
12 PD (de novo) 60.3 ± 9.1 3 45 600 TBSS Bradykinesi
a Whole brain analyses
Bilateral SLF, UF in the contralateral hemisphere, IF-OF in the ipsilateral hemisphere of the affected hand (no correlated area for less affected hand)
Cingulum, posterior / retrolenticular limb of internal capsule, posterior Thal radiation and cingulum (hippocampus) in the contralateral hemisphere; SLF, corona radiata, CST and the inferior CP in the ipsilateral hemisphere, stria terminalis, anterior limb of internal capsule, CP, CC body and CC splenium, middle CP, and pontine crossing tract in the bilateral hemisphere
P < 0.05 TFCE
Ofori et al. 2015
25 PD; 19 HC
Baseline: PD: 65.3 ± 9.1; HC: 65.3 ± 10.1
3 64 1000 ROI (free water mapping)
Bradykinesia, cognition (MoCA) (& progression)
Anterior and posterior SN
n.s. Free water: posterior SN correlated with progression of bradykinesia and MoCA
n.s.
P < 0.05 FDR
Canu et al. 2014
22 PD (+FOG); 35 HC (28 PD-FOG, 25 PD; 30 HC*)
PD: 66.9 ± 8.0; HC: 67.7 ± 7.6
3 (1.5*) 35 (65*) 900 TBSS and
tractography FOG Whole brain analyses and PPN tract
Primary motor, premotor, prefrontal. orbitofrontal and inferior parietal cortices, cingulum, bilateral SLF, bilateral CP, CC genu, anterior body and splenium, temporo-occipital tracts
Primary motor, premotor, prefrontal. orbitofrontal and inferior parietal cortices, cingulum, bilateral SLF, left CP, left CST, Thal radiation, bilateral external capsule, cerebellum in right lobule VIII, bilateral lobule IX (also increased AD and RD in these areas; increased AD in left PPN tract) PD-FOG vs. PD*: AD in CC genu and body
P < 0.05 FWE, TFCE

52
and WM under right parietal cortex and bilateral PPN tract)
Fling et al. 2013
26 PD (14 PD-FOG; 12 PD), 15 HC
PD-FOG: 67.1± 5.4; PD: 65.5 ± 7.0; HC: 66.7 ± 7.6
3 30 1000 Tractography FOG PPN
PD-FOG vs. PD and HC: PUT, internal GPi, cingulate, Thal, precentral and postcentral gyrus, superior and middle frontal gyrus, SMA, pre-SMA and bilateral cerebellar locomotor regions PPN fibre tract in the right hemisphere
P < 0.05 Bonferroni
Peterson et al. 2014
25 PD (13 PD-FOG; 12 PD)
PD-FOG: 66.2 ± 5.5; PD: 64.9 ± 6.9
3 30 1000 Tractography
FOG, dual tasks, stride length, stride length variability
PPN
Greater quantity in PPN tract in the right hemisphere; Correlation in FOG: lateralized PPN connectivity and larger dual task interference, stride length during normal and dual task walking
P < 0.05

53
Vercruysse et al. 2014
26 PD (11 FOG; 15 PD), 15 HC
PD-FOG: 68.6 ± 8.7; PD: 67.6 ± 5.6; HC: 68.1 ± 6.5
3 20/40/75*
700/1000/2800*
TBSS and tractography FOG
Whole brain analyses and subcortical-cortical connections
Left (lobule VI) and right (Crus I, lobule VIIIb) hemisphere of the cerebellum, left temporal part of the SLF; connections between the left caudate nucleus and ACC and orbitofrontal cortex, between the left middle CP and PPN
Right anterior part of the capsula interna and corona radiata, the superior frontal cortex, and a left-hemispheric cluster in cerebellum Crus II; connecting the caudate, PUT, GP, and STN to frontal (ACC, superior and orbitofrontal cortex), motor (pre-SMA, SMA) and sensory (S1 and S2) cortical regions predominantly in the left hemisphere
P < 0.001 (cluster forming threshold); P < 0.05 (cluster size threshold), FDR
Youn et al. 2015
42 PD (19 PD-FOG; 23 PD), 33 HC
PD-FOG: 71.1 ± 6.1; PD: 67.7 ± 6.4; HC: 69.6 ± 5.8
3 45 600 VBA and ROI FOG
Whole brain analyses and PPN and projection fibres (ROI)
Bilateral PPN Basal ganglia, Thal and cerebellum (in PPN tract); bilateral PPN
P < 0.05 FDR
Cognitive correlates
Gallagher et al. 2013
15 PD; 15 HC
PD: 62.7 ± 10; HC: 60.3 ± 6.5
3 25 Not reported TBSS Executive
function Whole brain analyses
Anterior limb of internal capsule, anterior corona radiata, CC body, sagittal stratum (ILF and IF-OF), UF, and deep cerebellar WM, right midbrain and in WM subjacent to several frontal. temporal. parietal. and occipital gyri
"More extensive than FA changes and present in portions of most tracks."
P < 0.05 FWE, TFCE
Matsui et al. 2007a
21 PD (6 PD with low EF scores; 15 high EF
PD-Low-EF: 71.5 ± 5.83; PD-High EF: 68.9 ± 10.3
1.5 6 1000 ROI Executive function (WCST)
Parietal WM (posterior to the central sulcus on the most caudalslice),
PD-Low EF < PD-High EF: Left parietal WM
P < 0.05 Bonferroni

54
scores) temporal WM (posterolateral to the lateral fissure on the most caudal slice where the lateral fissure was present), occipital WM (within the optic radiations on the most caudal slice where the occipital horn of the lateral ventricle was imaged).
Rae et al. 2012
14 PD; 15 HC
PD: 65 (range: 51-78); HC: 64 (range: 50-75)
3 12; 30**
250/500/750/950/1200; 600/1200**
TBSS and VBA
Executive function
Whole brain analyses
Forceps minor, CC, forceps major, anterior Thal radiation, UF, IF-OF, CST, SLF, ILF
Forceps minor, CC, forceps major, anterior Thal radiation, UF, IF-OF, CST, SLF, ILF, internal and external capsule
P < 0.01 FWE, TFCE
Theilmann et al. 2013
25 PD; 26 HC
PD: 68.0 ± 8.9; HC: 65.9 ± 8.4
1.5 51 1000 TBSS and ROI
Executive function
ROI generated from the voxels that showed significant group differences
CC, cingulum, anterior and superior corona radiata, anterior Thal radiation, forceps minor, forceps major, IF-OF, SLF, IFL
CC, cingulum, anterior corona radiata, anterior Thal radiation, forceps minor, forceps major, IF-OF, SLF, IFL
P < 0.05
Melzer et al. 2013
109 PD (18 PDD; 28 PD-MCI; 63 PD), 32 HC
PDD: 73.7 ± 6.5; PD-MCI: 71.0 ± 7.3; PD: 64.0 ± 9.2; HC: 70.1 ± 9.0
3 28 1000 TBSS Global Whole brain analyses n.s.
Correlations: Bilateral CC genu and body, internal capsule anterior and posterior limb, external capsule, anterior and superior corona radiata, anterior Thal radiation, left CST, left cingulum, bilateral IF-OF, UF, ILF, right SLF
P < 0.05 TFCE

55
Executive function n.s.
Correlations: bilateral CC genu and body, internal capsule anterior and posterior limb, external capsule, anterior, posterior and superior corona radiata, anterior Thal radiation, left cingulum, bilateral IF-OF and UF, right ILF and SLF
Attention
Correlations: CC body, external capsule, anterior and superior corona radiata, anterior Thal radiation, CST, IF-OF, ILF, SLF (all right)
Correlations: bilateral CC genu and body, internal capsule anterior and posterior limb, external capsule, anterior and superior corona radiata, right posterior corona radiata, bilateral anterior Thal radiation, CST, left cingulum, bilateral IF-OF, SLF and UF, right ILF
Memory n.s.
Correlations: bilateral CC genu and left body, bilateral internal capsule anterior and posterior limb, external capsule, anterior and superior corona radiata, bilateral anterior Thal radiation, bilateral IF-OF and UF, right ILF and right SLF
Visuospatial n.s. n.s.

56
MCI, Dementia
PD-MCI and PDD < HC: bilateral CC genu, body, splenium, medial lemniscus (PD-MCI: left; PDD: bilateral), inferior (PDD) and superior CP, internal capsule anterior and posterior limb, external capsule, anterior superior and posterior corona radiata, anterior Thal radiation, CST (all bilateral); cingulum (PD-MCI-bilateral. PDD: left), IF-OF, ILF, SLF, UF (all bilateral). PD-MCI < PD: bilateral CC genu, right CC body, bilateral internal capsule anterior limb, right external capsule. PD < PD: right superior corona radiata, right CST
PD > HC: bilateral CC body, left splenium, bilateral superior corona radiata, left cingulum. PD-MCI and PDD > HC: bilateral CC genu, body, splenium, right superior CP (PDD), left (MCI) and right (PDD) CP, internal capsule anterior and posterior limb, external capsule, anterior superior and posterior corona radiata, anterior Thal radiation, CST cingulum, IF-OF, ILF, SLF (all bilateral) UF (left PD-MCI, bilateral PDD). PD-MCI > PD: bilateral external capsule and internal capsule anterior limb, left anterior and superior corona radiata, left IF-OF and SLF, bilateral UF and SLF PDD > PD: bilateral CC genu and right body, bilateral internal capsule anterior limb, right internal capsule superior limb and external capsule, bilateral anterior and superior corona radiata, left Thal radiation, right CST, bilateral IF-OF, left UF.

57
Zheng et al. 2014 16 PD 62.2 ± 9.6 3 20** 1000 ROI
Executive function
ROI derived from significant correlations of FA and MD with cognitive domains
Correlation: right anterior corona radiata, left anterior limb of internal capsule, genu, right posterior Thal radiation
Correlations: bilateral anterior corona radiata, left anterior limb of internal capsule, genu, right posterior Thal radiation, left sagittal stratum, left SFO
P < 0.01
Attention
Correlations: bilateral cingulate gyrus, right external capsule, bilateral posterior Thal radiation, left retrolenticular part of internal capsule, left sagittal stratum, bilateral SFO
Correlations: left cingulate gyrus, left anterior corona radiata, bilateral superior corona radiata, left CST, left hippocampus, bilateral posterior corona radiata, bilateral posterior Thal radiation, left retrolenticular part of internal capsule, left sagittal stratum, splenium, bilateral SFO
Linguistic performance
Correlations: left anterior limb of internal capsule, right sagittal stratum
Correlations: bilateral corona radiata, genu, bilateral sagittal stratum, left SFO
Visuospatial n.s. n.s. Short term memory n.s. Fornix
Long term memory n.s. Anterior corona radiata
Al Serafy, 2015
40 PD (10 PD-MCI; 23 PD), 20 HC
PD (total): 59.9 ± 7.5; HC: 56.4 ± 5.1
1.5 25 800 ROI
Attention Caudate, PUT, GP, Thal, SN, PFC, hippocampus.
Correlations: Thal, GP, caudate, PFC
P < 0.05
Working memory
Correlations: Thal, GP, caudate, PFC
Immediate and delayed recall
Correlations: Thal, PUT, PFC, hippocampus

58
Alternating and action verbal fluency
Correlations: Thal, GP, caudate, PFC
Naming, visuoperceptual. visouconstructual
n.s.
MCI PD-MCI < PD: Thal, GP, PUT, caudate, PFC
Auning et al. 2014
18 PD; 19 HC
PD: 66.7 ± 5.1; HC: 64.6 ± 6.5
1.5 12 750 TBSS and ROI
Cognitive performance
Middle frontal gyrus (rostral division)
n.s.
RD: correlation executive function tests (composite score, trail making task part B and colour-word interference task) and rostral middle frontal cortex
P < 0.05 TFCE
Ye et al. 2014 21 PD; 20 HC
PD: 64.0 ± 8.1; HC: 65.3 ± 5.7
3 63 Not reported TBSS Response
inhibition Whole brain analyses n.s.
P < 0.05 TFCE
Chen et al. 2015
30 PD (19 PD; 11 PDD), 21 HC
PDD: 64.1 ± 11.4; PD: 59.5 ± 8.8; HC: 61.1 ± 8.3
3 Not reported
Not reported ROI
Cognitive performance (MoCA)
Body and splenium of CC, fornix, SFO, cingulate gyrus, hippocampus, IF-OF, ILF, SLF, UF
PD vs. HC: n.s.; PDD < PD: left hippocampus
HC vs. PD: n.s.; PDD > PD: right SLF, right cingulate gyrus, left ILF, left SLF, bilateral UF; PDD > HC: splenium and body of CC, right SFO, left SLF, bilateral UF
P < 0.05 Bonferroni
Duncan et al. 2016
125 PD (newly diagnosed), 50 HC
PD: 66.0 ± 10.5; HC: 65.8 ± 8.0
3 64 1000 TBSS Cognitive performance
Whole brain analyses n.s.
Forceps minor, cingulum, SLF, ILF, IF-OF, CST, CC and internal capsule
P < 0.05 FWE, TFCE
Koshimori et al. 2015
16 PD; 14 HC
PD: 70.5 ± 5.6; HC: 3 60 1000 TBSS Cognitive
performance Whole brain analyses n.s. Forceps minor and
major, cingulum, P < 0.05 TFCE

59
67.13 ± 5.1 anterior thalamic radiation, superior corona radiata, external capsule, body of CC, UF, IF-OF, SLF and ILF.
Carlesimo et al. 2012
25 PD; 25 HC
PD: 65.0 ± 8.4; HC: 65.0 ± 8.9
3 30 1000 ROI Declarative memory Hippocampus
Right hippocampus in PD compared to HC; hippocampus correlated to memory performance
P < 0.05 FWE
Agosto et al. 2014
43 PD (30 PD-MCI, 13 PD); 33 HC
PD-MCI: 63.9 ± 7.1; PD: 66.6 ± 8.2; HC: 64.0 ± 7.3
1.5 12 1000 TBSS MCI Whole brain analyses
PD-MCI vs. PD: bilateral anterior and superior corona radiata, CC genu and body anterior inferior IF-OF/uncinate, anterior SLF; PD-MCI vs. HC: bilateral anterior and superior corona radiata, CC body, anterior SLF
n.s.
P < 0.05 FWE, TFCE
Rossi et al. 2014
33 PD; 19 HC
PD: median: 71 (range 42 – 86; MCI and cognitively intact), HC: median: 73 (range: 44-87)
3 20 1000 TBSS MCI Whole brain analyses
n.s. (Correlation: older age at disease onset associated and bilateral frontal and parietal lobes, right temporal and occipital lobes and right thalamus)
P < 0.05
Deng et al. 2012
64 PD (10 PDD; 30 PD-MCI; 24 PD), 21 HC
PDD: 69.0 ± 9.7; PD-MCI: 65.1 ± 11.8; PD: 62.1 ± 8.6; HC: 60.1 ± 13.6
3 16 1000 ROI MCI; Dementia
Temporal WM (posterolateral to the lateral fissure on most caudal slice), frontal WM (anterior /posterior
PD < HC: left occipital WM and the left anterior cingulate bundle; PD-MCI and PDD < HC: left frontal and right temporal WM, bilateral anterior
P < 0.05 Fisher least significant difference t-test

60
cingulate bundle, CC genu and splenium, and SLF), parietal WM (in the region posterior to the central sulcus on the most caudal slice), occipital WM (within the optic radiations on the most caudal slice), CST at the levels of the red nucleus and internal capsule.
cingulate bundles; PDD < all groups: left anterior cingulate bundle and CC splenium; PDD vs. PD: left occipital WM. Correlations cognitive status: left frontal. right temporal. left occipital WM, bilateral anterior cingulate bundles and CC splenium
Hattori et al. 2012
85 PD (25 PDD; 28 PD-MCI; 32 PD), 40 HC
PDD: 80.0 ± 4.5; PD-MCI: 76.3 ± 6.4; PD: 75.9 ± 2.0; HC: 76.9 ± 4.9
1.5 12 1000 TBSS MCI; Dementia
Whole brain analyses
PD vs. HC: n.s. PD-MCI and PDD < HC: SLF, ILF, IF-OF, UF, cingulum, CC; PDD vs. PD: SLF, IFL, IF-OF, UF, cingulum, CC
P < 0.05 TFCE
Kamagata et al. 2012
30 PD (15 PDD; 15 PD), 15 HC
PDD: 71.3 ± 5.6; PD: 69.8 ± 5.9; HC: 69.5 ± 6.9
3 32 1000 Tractography Dementia
Anterior and posterior cingulate bundle, CST
PD < HC: anterior cingulate bundle; PDD < HC: anterior and posterior cingulate bundle; PDD vs. PD: n.s.
PD > HC: anterior cingulate bundle; PDD vs. HC: n.s.; PDD vs. PD: n.s.
P < 0.05 Scheffé correction, Bonferroni
Kamagata et al. 2013
40 PD patients (20 PDD, 20 PD), 20 HC
PDD: 71.7 ± 5.3; PD: 71.6 ± 4.3; HC: 72.2 ± 3.3
3 32 1000 TBSS and tractography Dementia
Whole brain analyses and significant clusters found in TBSS as ROIs (genu CC)
PDD < PD: anterior part of the IF-OF (adjacent to the prefrontal area) and in part of the genu of CC; PDD < HC: SLF, ILF, IF-OF, UF, cingulum, anterior limb of internal
PDD > PD: anterior part of the IF-OF (adjacent to the prefrontal area) and in part of the genu of CC; PDD > HC: SLF, ILF, IF-OF, UF, cingulum, anterior and posterior limb of internal capsule, SN (all
P < 0.05 Bonferroni

61
capsule, SN (all bilateral). Correlation all PD patients and MMSE in genu CC.
bilateral). Correlation all PD patients and MMSE in genu CC.
Matsui et al. 2007b
37 PD (11 PDD; 26 PD)
PDD: 75.0 ± 7.7; PD: 70.0 ± 8.6
1.5 6 1000 ROI Dementia
Bilateral parietal. temporal. frontal. occipital WM, bilateral anterior and posterior cingulate bundle
Bilateral posterior cingulate bundles
P < 0.05 Fisher’s PLSD
Wiltshire et al. 2010
35 PD (6 PDD; 29 PD), 15 HC
PDD: 71.4 ± 4.2; PD: 70.9 ± 4.6; HC: 70.7 ± 4.0
1.5 6 1000 Tractography Dementia, cognition CC, cingulum n.s.
PDD vs. PD vs. HC: n.s. Correlation: MMSE and CC anterior and posterior
P < 0.05 Bonferroni
Lee et al., 2011
11 PD; 11 HC
PD: 70.4 ± 4.7; HC: 67.0 ± 3.8
3 32 Not reported ROI Speech
impairment
Broca’s and Wernicke’s areas
Broca’s and Wernicke’s areas n.s.
P < 0.05
Mood disturbances
Carriere et al. 2014
20 PD (10 PD-Apathy; 10 PD), 10 HC
PD-Apathy: 67.2 ± 8.4; PD: 60.7 ± 11.1; HC: 66.8 ± 6.8
3 15 1000 TBSS Apathy Whole brain analyses n.s. n.s.
P < 0.05 TFCE
Huang et al. 2014
30 PD (15 PD-Depression; 15 PD)
PD-Depression: 54.5 ± 12.2; PD: 54.8 ± 10.1
3 31 1000 TBSS and ROI Depression
Whole brain analyses and significant clusters found in TBSS as ROIs
Left UF, left SLF, left anterior Thal radiation, left forceps minor, ILF; Correlation deep temporal cortex with depression rating scale
n.s.
P < 0.01 cluster-based thresholding method (cluster size > 100 voxels)

62
Li et al. 2010
32 PD (14 PD-Depression; 18 PD)
PD-Depression: 65.3 ± 8.9; PD: 61.1 ± 10.2
3 20 1000 ROI Depression Thal
PD-depressed < PD: Bilateral mediodorsal Thal; Correlation between bilateral mediodorsal Thal and depression rating scale
n.s.
P < 0.05
Matsui et al. 2007c
28 PD (14 PD-Depression; 14 PD)
PD-Depression: 72.1 ± 9.9; PD: 69.3 ± 8.1
1.5 6 1000 ROI Depression
Orbitofrontal. prefrontal. temporal. occipital. parietal WM, anterior and posterior cingulate bundles
PD-depressed < PD: Bilateral anterior cingulate bundles
P < 0.05
Surdhar et al. 2012
12 PD (6 PD-Depression; 6 PD), 6 HC
PD-Depression: 70.1 ± 2.9; PD: 68.6 ± 2.0; HC: 70.7 ± 2.4
1.5 6 1000 Tractography Depression (mild) CC, UF n.s. n.s.
P < 0.05 Bonferroni
Baggio et al. 2012
43 PD; 23 HC
PD: 63.5 ± 11.4; HC: 61.0 ± 9.7
3 30 1000 TBSS, ROI Facial emotion recognition
IF-OF, SLF, ILF, UF, cingulum, forceps minor
Sadness: IF-OF in right frontal lobe and left temporal lobe, forceps minor, CC body, left CC semiovale, IF-OF in left temporal lobe.
P < 0.05 FWE
Visual hallucinations
Yao et al. 2015
27 PD (12 PD-VH; 15 PD), HC: 14
PD-VH: 70 (percentile: 64, 72.75); PD: 66 (percentile: 62, 72); HC: 63 (percentile: 62,68.75)
3 16 1000 ROI VH Hippocampus Not reported
PD-VH > PD: Right posterior hippocampal regions and left hippocampal body
P < 0.05 FWE

63
Zhong et al. 2013
66 PD (18 PD-Psychosis; 48 PD)
PD-Psychosis: 56.3 ± 23.7; PD: 55.6 ± 22.3
3 15 1000 ROI Psychosis
SN, GP, frontal and occipital lobe, cingulate gyrus, hippocampus
Bilateral frontal and occipital lobe, cingulate gyrus and left hippocampus
P < 0.05
REM Sleep behaviour disorder
Ford et al. 2013
124 PD patients (46 PD-RBD; 78 PD)
PD-RBD: 66.4 ± 9.9; PD: 65.8 ± 10.9
3 64 1000 TBSS RBD Whole brain analyses
n.s. (before correcting for multiple comparisons: Right IF-OF, left ILF, left SLF, right CST)
n.s. (before correcting for multiple comparisons: bilateral ILF)
P < 0.05 TFCE
Lim et al. 2016
38 PD (24 PD-RBD; 14 PD), 25 HC
PD-RBD: 69.8 ± 6.4; PD: 69.7 ± 7.2; HC: 68.5 ± 6.6
3 15 800 TBSS, ROI and tractography
RBD
Whole brain analyses and PPN and projection fibres
PD and PD-RBD < HC: bilateral frontal areas
n.s.
P < 0.05 TFCE
Autonomic function and miscellaneous studies
Ibarretxe-Bilbao et al. 2010
24 PD (9 PD-anosmic; 9 PD-hyposmic; 6 PD), 24 HC
PD-anosmic: 57.9 ± 7.9; PD-hyposmic: 58.0 ± 5.8; PD: 50.7 ± l1.3; HC: 57.3 ± 8.9
3 30 1000 TBSS and ROI
Olfactory impairment
Whole brain analyses and WM adjacent to the orbitofrontal cortex (superficial WM structures), medial temporal lobe WM including WM adjacent to the amygdala, piriform cortex, and entorhinal cortex (primary olfactory system), UF
Hypnosmic and anosmic PD < PD and HC: WM adjacent to bilateral gyrus rectus; Anosmic vs. PD and HC: primary olfactory cortex (adjacent to entorhinal cortex). Correlation olfactory tests in PD: left primary olfactory cortex, right gyrus rectus
P < 0.05 FWE

64
PD = Parkinson’s disease, HC = healthy controls, FA = fractional anisotropy, MD = mean diffusivity, AD = axial diffusivity, RD = radial diffusivity, TBSS = tract-based spatial statistics, VBA = voxel-based analyses; ROI = region of interest, WM = WM, UPDRS = Unified Parkinson’s Disease Rating Scale, H&Y = Hoehn and Yahr, SE = Schwab and England, PIGD = postural instability and gait disorder, TD = tremor dominant; FOG = freezing of gait, PD-MCI = Parkinson’s disease with mild cognitive impairment, PDD = Parkinson’s disease with dementia, AD = Alzheimer’s disease; WCST = Wisconsin Card Sorting Test; VH = visual hallucinations, RBD = REM sleep behaviour disorder, MoCA = Montreal Cognitive Assessment CC = corpus callosum, SN = substantia nigra, Thal = thalamus, PUT = putamen, GP = globus pallidus (GPi/e = globus pallidus internus/externus), SLF = superior longitudinal fasciculus, ILF = inferior longitudinal fasciculus, UF = uncinate fasciculus, IF-OF = inferior fronto-occipital fasciculus; CST = corticospinal tract, PPN = pedunculopontine nucleus, CP = cerebral peduncle, STN = subthalamic nucleus, ACC = anterior cingulate cortex, SMA = supplementary motor area, PFC: prefrontal cortex, n.s. = not significant, FWE: family-wise error, TFCE: threshold-free cluster enhancement, FDR: false discovery rate, PLSD: partial least-squares difference
*Subsequent analyses **Different imaging protocols used ***One subject underwent a 60-direction acquisition
Rolheiser et al. 2011
14 PD; 14 HC
PD: 56 ± 4.8; HC: 55.2 ± 6.2
1.5 31 900 TBSS and ROI
Olfactory impairment and early diagnosis PD
Whole brain analyses and ROI: Anterior olfactory structures and SN
SN, anterior olfactory structures RD: SN
P < 0.05 Bonferroni
Skorpil et al. 2012
14 PD; 15 HC
PD: 64 (range: 47-73); HC: 60 (range: 44-72)
1.5 32 900 ROI
Olfactory impairment, Parkinsonian severity
Olfactory bulb, SN, CC (two DTI sequences)
Olfactory bulb (first series), SN
Olfactory bulb (first series)
P < 0.05
Zhang et al. 2009
25 PD; 25 HC
PD: 58.4 ± 9.8; HC: 58.4 ± 9.3
3 12 1000 VBA Olfactory impairment
Whole brain analyses
Bilateral cerebellar hemisphere and right rectus gyrus; Correlation with olfactory tests: left medial cerebellum white matter
Bilateral orbitofrontal cortices and bilateral inferior temporal gyri; bilateral parietal lobes and left precentral gyrus; Correlation with olfactory tests: right cerebellum, white matter in the left inferior parietal lobule
P < 0.05 Monte Carlo simulations cluster size for P < 0.001 Regression: P < 0.001 uncorrected
Bertrand et al. 2012
26 PD (13 PD-MCI; 13 PD), 7 HC
26 PD: 64.1 ± 8.6; HC: 70.4 ± 9.9
3 64 700 TBSS Colour discrimination
Whole brain analyses n.s.
Temporal and parietal portions of the right SLF. RD: right IF-OF, CC splenium to posterior region of the body, IFL
P < 0.05 Voxel-wise correction

65
Visual hallucinations and psychosis
VH in PD exist along a perceptual spectrum from simple images without a recognizable
form, to complex well-formed hallucinations, which comprise images of people, faces,
animals or scenery (Santhouse et al., 2000). VH are more often present in the advanced
stages of PD and previous studies have shown that patients in the non-tremor dominant
phenotype have a higher risk of developing visual hallucinations (VH) (Emre et al.,
2007). Patients with VH show higher MD values in the posterior regions of the
hippocampus compared to non-VH patients and healthy controls (Yao et al., 2014), a
finding related to impaired visuospatial memory. Moreover, the hippocampus has been
identified as a key hub in the default mode network (Buckner et al., 2008). This network
supports introspection, mental imagery and the capacity to retrieve and manipulate
episodic memories (Buckner et al., 2008) and has recently been proposed to play a key
role in the occurrence of VH (Shine et al., 2014; Shine et al., 2011a). Reductions in FA
values in the hippocampus are also found in PD patients with psychosis (Zhong et al.,
2013). Furthermore, FA reductions within the cingulate gyrus, another region of the
default mode network, have also been reported in this patient group, along with FA
abnormalities in the frontal and occipital lobe. The majority of these patients also
reported VH, however, it has yet to be investigated whether or not these changes were
driven by this symptom.
REM sleep behaviour disorder
RBD is a parasomnia associated with PD. Patients with RBD not only lose muscle atonia
during REM sleep but also show dream enactments, which are more vivid and often
comprise aggressive or frightening content (Boeve, 2010). Muscle atonia is regulated by
nuclei in the brainstem (Boeve, 2010), however, dream alteration in RBD indicates the
involvement of neocortical structures. Studies investigating idiopathic RBD identify
diffusivity changes in the brainstem, midbrain, pons and SN compared to age-matched
controls (Scherfler et al., 2011; Unger et al., 2010). Surprisingly, the two studies using
DTI to investigate the presence of white matter abnormalities in PD-RBD patients, both

66
report changes in the bilateral frontal areas but only before correcting for multiple
comparisons (Ford et al., 2013; Lim et al., 2016).
Autonomic dysfunction and miscellaneous studies
Our search yielded no studies using DTI analyses to investigate bladder dysfunction,
constipation, blood pressure, fatigue or other autonomic dysfunctions.
Loss of smell is common in PD and often predates the clinical motor symptoms. Studies
using DTI analyses have identified reduced FA values in the olfactory region (Ibarretxe-
Bilbao et al., 2010; Rolheiser et al., 2011; Skorpil et al., 2012; Zhang et al., 2011).
Additionally, a correlation between reduced FA values in bilateral cerebellar white matter
was reported (Zhang et al., 2011). The cerebellum modulates the diaphragm and
intercostal muscles and it has been suggested that smell could be improved by extra sniffs
created by these muscles.
Colour discrimination deficiency, another common symptom that might precede PD, is
associated with increased diffusivity in the long association fibres in the temporal and
parietal regions as well as in the corpus callosum (Bertrand et al., 2012). However, the
authors suggest that the measure they used to test this symptom might reflect cognitive
impairment instead of colour discrimination deficits, as the long fasciculi such as the SLF
and IF-OF project to the frontal lobe, which is associated with cognitive performance.
Conclusions and future directions
This review has outlined the contribution of diffusivity abnormalities to different
symptoms of PD using DTI analyses. The increased interest in axonal analyses using DTI
in PD in the last decade has led to an expansion of our understanding of the impact of
white matter damage and changes in membrane permeability in grey matter in
symptomology of PD. White matter alterations within the long association and inter-
hemispheric fibres have implications for many symptoms of PD due to their extensive
coverage of multiple regions. For example, the SLF is a large bundle of bidirectional

67
fibres that integrate motor and decision-making centres with visual, auditory and sensory
regions (Makris et al., 2005). Damage within the SFL contributes to bradykinesia, FOG,
executive dysfunction and depression (see summary of results in Table 1). Moreover,
disruptions within the corpus callosum disrupt the frontal inter-hemispheric connections
and have a huge impact on complex integrated behaviour such as motor programs and
gait (Bhadelia et al., 2009) as well as cognitive functioning (Wiltshire et al., 2010).
Abnormalities within other brain tissues are more symptom specific, such as changes in
diffusivity in the putamen and its projection fibres (motor symptoms) or forceps minor
(executive function and depression) and hippocampus (dementia and hallucinations).
Investigations into these specific alterations are relevant for the understanding the neural
underpinnings of these PD features and possibly provide new opportunities for disease
modifying therapies. It further provides insights into the heterogeneous character of PD
and could aid in identifying and predicting different PD phenotypes. However, whilst the
association between changes in structural connectivity and cognitive correlates in PD are
well studied, further research is needed to obtain a better understanding of several motor
symptoms. It has been suggested that an increase in axonal density in pathways of the
cortico-basal-ganglia-thalamo-cortical loop might reflect a potential compensation
mechanism (Mole et al., 2016; Zhan et al., 2012). Increased FA values have been found
in these motor pathways and should be taken into account when investigating white
matter in PD in future studies. Importantly, this review also highlights the limited
research investigating symptoms such as autonomic dysfunction, anxiety, VH and RBD.
Limitations and considerations for future research
Of the studies reported here, many are limited by a low number of subjects, resulting in
low statistical power, which might explain the conflicting results. Future studies should
also include a large control cohort, as it has been shown that differences between healthy
participants and PD patients might rely on high FA values in the control group (Schwarz
et al., 2013). Furthermore, studies correlating imaging variables and symptoms can be
confounded by disease severity. Whilst it is clinically difficult to distinguish the two,
general disease severity should be taken into account when assessing symptomology of

68
PD in the research setting. Furthermore, studies on grey matter are relatively limited and
replication of the current results is very important. All of the studies included in our
review included patients with a clinical diagnosis of PD; however this does not rule out
the potential for other underlying pathologies, for example progressive supranuclear
palsy or multiple system atrophy. An important future direction will be to corroborate
DTI investigations in PD with postmortem data. Unique insights into PD symptomology
can further be gained by conducting multimodal studies, combining advanced DTI
analyses of structural connectivity with functional MRI (task-related and resting state) as
well as grey matter volumetric MRI and cortical thinning analyses. Such studies are
required to unravel how white matter injuries are affected by grey matter atrophy as well
as their influence on functional connectivity. Finally, exciting approaches such as graph
theory could provide a more systematic analyses white matter topology.
Protocol considerations
Currently, DTI protocols vary across centres and these differences might impact the
results. Firstly, the number of diffusion encoding directions differs across studies. A
minimum of six non-collinear diffusion-encoding directions are required to measure the
full diffusion tensor. However, the use of more than six gradient directions (e.g. 30 or 60)
improves the accuracy of fibre-tract orientations. The longer scanning time required with
the use of a higher number of gradient directions influences the signal to noise ratio
(Lebel et al., 2012). Larger voxel sizes can improve this image signal to noise ratio,
although this will increase partial volume averaging of tissues, leading to an increased
risk of errors in the fits to the diffusion tensor model. Secondly, higher field strength
(e.g. 3 Tesla vs. 1.5 Tesla) may improve image quality and diagnostic accuracy. Less
scanning time is required for a similar image, which can improve signal to noise ration by
25%. Diffusion weighted gradients (or b-values) may also impact research outcomes. The
use of weighted gradients for the brain of less than 700 s/mm2 is discouraged, and b-
values of 1000 s/mm2 are most commonly used. Higher weighted gradients show
increased contrast and expose lesions more accurately (Alexander et al., 2007).
Evidently, these different protocol considerations are highly dependent on each other.

69
Undeniably, it should be highlighted that DTI only allows for indirect validation and the
measurements are error-prone and can be difficult to interpret. Furthermore, the motion
of water molecules does not inform about the directionality of axons. However, it is the
only non-invasive technique that allows for analysing in vivo structural connectivity of
the brain and is valuable for addressing new scientific questions. Given that DTI analyses
is able to identify microscopic changes in white and grey matter in the early stages of PD,
this technique could be valuable in predicting the manifestation of certain symptoms of
PD. Furthermore, the quality of DTI analyses has improved substantially over the last
decade. Advances such as the use of increased field strengths and diffusion-encoding
directions, as well as improved motion-correction techniques have led to better quality of
FA and MD images.
Overall, clinical manifestations of PD are related to diffusivity abnormalities within
divergent axons to extended brain regions. However, several conflicting results call for
further research using DTI analyses to clarify these abnormalities in more detail. DTI
could be a valuable technique to identify early alterations, which may lead to early
interventions to treat key features of PD.

70
References
Agosta, F., Canu, E., Stefanova, E., Sarro, L., Tomic, A., Spica, V., . . . Filippi, M.
(2014). Mild cognitive impairment in Parkinson's disease is associated with a
distributed pattern of brain white matter damage. Hum Brain Mapp, 35(5), 1921-
1929. doi:10.1002/hbm.22302
Al Serafy, O. A., Soliman, R. H., Asaad, R. E., Youssof, A. M. A., & Tawfik, M. H.
(2015). Cognitive functions in patients with parkinson’s disease: I. The effect of
cerebral microstructural changes. Egytp J Neurol Psychiatry Neurosurg, 52(1),
29-36.
Alexander, A. L., Lee, J. E., Lazar, M., & Field, A. S. (2007). Diffusion tensor imaging
of the brain. Neurotherapeutics, 4(3), 316-329. doi:10.1016/j.nurt.2007.05.011
Auning, E., Kjaervik, V. K., Selnes, P., Aarsland, D., Haram, A., Bjornerud, A., . . .
Fladby, T. (2014). White matter integrity and cognition in Parkinson's disease: a
cross-sectional study. BMJ Open, 4(1), e003976. doi:10.1136/bmjopen-2013-
003976
Baggio, H. C., Segura, B., Ibarretxe-Bilbao, N., Valldeoriola, F., Marti, M. J., Compta,
Y., . . . Junque, C. (2012). Structural correlates of facial emotion recognition
deficits in Parkinson's disease patients. Neuropsychologia, 50(8), 2121-2128.
doi:10.1016/j.neuropsychologia.2012.05.020
Basser, P. J., & Pierpaoli, C. (1996). Microstructural and physiological features of tissues
elucidated by quantitative-diffusion-tensor MRI. J Magn Res Series B, 111(3),
209-219. doi:Doi 10.1006/Jmrb.1996.0086
Bertrand, J. A., Bedetti, C., Postuma, R. B., Monchi, O., Genier Marchand, D., Jubault,
T., & Gagnon, J. F. (2012). Color discrimination deficits in Parkinson's disease
are related to cognitive impairment and white-matter alterations. Mov Disord,
27(14), 1781-1788. doi:10.1002/mds.25272
Bhadelia, R. A., Price, L. L., Tedesco, K. L., Scott, T., Qiu, W. Q., Patz, S., . . .
Bergethon, P. (2009). Diffusion tensor imaging, white matter lesions, the corpus
callosum, and gait in the elderly. Stroke, 40(12), 3816-3820.
doi:10.1161/STROKEAHA.109.564765

71
Boeve, B. F. (2010). REM sleep behavior disorder Updated review of the core features,
the REM sleep behavior disorder-neurodegenerative disease association, evolving
concepts, controversies, and future directions. Year Neurol 2, 1184, 15-54.
Braak, H., Del Tredici, K., Rub, U., de Vos, R. A., Jansen Steur, E. N., & Braak, E.
(2003). Staging of brain pathology related to sporadic Parkinson's disease.
Neurobiol Aging, 24(2), 197-211.
Brown, R. G., Landau, S., Hindle, J. V., Playfer, J., Samuel, M., Wilson, K. C., . . .
Group, P.-P. S. (2011). Depression and anxiety related subtypes in Parkinson's
disease. J Neurol Neurosurg Psychiatry, 82(7), 803-809.
doi:10.1136/jnnp.2010.213652
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L. (2008). The brain's default
network: anatomy, function, and relevance to disease. Ann N Y Acad Sci, 1124, 1-
38. doi:10.1196/annals.1440.011
Canu, E., Agosta, F., Sarasso, E., Volonte, M. A., Basaia, S., Stojkovic, T., . . . Filippi,
M. (2015). Brain structural and functional connectivity in Parkinson's disease
with freezing of gait. Hum Brain Mapp, 36(12), 5064-5078.
doi:10.1002/hbm.22994
Carlesimo, G. A., Piras, F., Assogna, F., Pontieri, F. E., Caltagirone, C., & Spalletta, G.
(2012). Hippocampal abnormalities and memory deficits in Parkinson disease: a
multimodal imaging study. Neurology, 78(24), 1939-1945.
doi:10.1212/WNL.0b013e318259e1c5
Carriere, N., Besson, P., Dujardin, K., Duhamel, A., Defebvre, L., Delmaire, C., &
Devos, D. (2014). Apathy in Parkinson's disease is associated with nucleus
accumbens atrophy: a magnetic resonance imaging shape analysis. Mov Disord,
29(7), 897-903. doi:10.1002/mds.25904
Carter, C. S., Braver, T. S., Barch, D. M., Botvinick, M. M., Noll, D., & Cohen, J. D.
(1998). Anterior cingulate cortex, error detection, and the online monitoring of
performance. Science, 280(5364), 747-749.
Chan, L. L., Ng, K. M., Rumpel, H., Fook-Chong, S., Li, H. H., & Tan, E. K. (2014).
Transcallosal diffusion tensor abnormalities in predominant gait disorder

72
parkinsonism. Parkinsonism Relat Disord, 20(1), 53-59.
doi:10.1016/j.parkreldis.2013.09.017
Chan-Palay, V., & Asan, E. (1989). Alterations in catecholamine neurons of the locus
coeruleus in senile dementia of the Alzheimer type and in Parkinson's disease
with and without dementia and depression. J Comp Neurol, 287(3), 373-392.
doi:10.1002/cne.902870308
Chaudhuri, K. R., Healy, D. G., Schapira, A. H., & National Institute for Clinical, E.
(2006). Non-motor symptoms of Parkinson's disease: diagnosis and management.
Lancet Neurol, 5(3), 235-245. doi:10.1016/S1474-4422(06)70373-8
Chen, B., Fan, G. G., Liu, H., & Wang, S. (2015). Changes in anatomical and functional
connectivity of Parkinson's disease patients according to cognitive status. Eur J
Radiol, 84(7), 1318-1324. doi:10.1016/j.ejrad.2015.04.014
Cochrane, C. J., & Ebmeier, K. P. (2013). Diffusion tensor imaging in parkinsonian
syndromes: a systematic review and meta-analysis. Neurology, 80(9), 857-864.
doi:10.1212/WNL.0b013e318284070c
Deng, B., Zhang, Y., Wang, L., Peng, K., Han, L., Nie, K., . . . Wang, J. (2013).
Diffusion tensor imaging reveals white matter changes associated with cognitive
status in patients with Parkinson's disease. Am J Alzheimers Dis Other Demen,
28(2), 154-164. doi:10.1177/1533317512470207
Double, K. L., Halliday, G. M., McRitchie, D. A., Reid, W. G., Hely, M. A., & Morris, J.
G. (1996). Regional brain atrophy in idiopathic parkinson's disease and diffuse
Lewy body disease. Dementia, 7(6), 304-313.
Dujardin, K., Blairy, S., Defebvre, L., Duhem, S., Noel, Y., Hess, U., & Destee, A.
(2004). Deficits in decoding emotional facial expressions in Parkinson's disease.
Neuropsychologia, 42(2), 239-250.
Duncan, G. W., Firbank, M. J., Yarnall, A. J., Khoo, T. K., Brooks, D. J., Barker, R. A., .
. . O'Brien, J. T. (2016). Gray and white matter imaging: A biomarker for
cognitive impairment in early Parkinson's disease? Mov Disord, 31(1), 103-110.
doi:10.1002/mds.26312

73
Emre, M., Aarsland, D., Brown, R., Burn, D. J., Duyckaerts, C., Mizuno, Y., . . . Dubois,
B. (2007). Clinical diagnostic criteria for dementia associated with Parkinson's
disease. Mov Disord, 22(12), 1689-1707; quiz 1837. doi:10.1002/mds.21507
Fling, B. W., Cohen, R. G., Mancini, M., Nutt, J. G., Fair, D. A., & Horak, F. B. (2013).
Asymmetric pedunculopontine network connectivity in parkinsonian patients with
freezing of gait. Brain, 136(Pt 8), 2405-2418. doi:10.1093/brain/awt172
Ford, A. H., Duncan, G. W., Firbank, M. J., Yarnall, A. J., Khoo, T. K., Burn, D. J., &
O'Brien, J. T. (2013). Rapid eye movement sleep behavior disorder in Parkinson's
disease: magnetic resonance imaging study. Mov Disord, 28(6), 832-836.
doi:10.1002/mds.25367
Fraix, V., Bastin, J., David, O., Goetz, L., Ferraye, M., Benabid, A. L., . . . Debu, B.
(2013). Pedunculopontine nucleus area oscillations during stance, stepping and
freezing in Parkinson's disease. PLoS One, 8(12), e83919.
doi:10.1371/journal.pone.0083919
Gallagher, C., Bell, B., Bendlin, B., Palotti, M., Okonkwo, O., Sodhi, A., . . . Alexander,
A. (2013). White matter microstructural integrity and executive function in
Parkinson's disease. J Int Neuropsychol Soc, 19(3), 349-354.
doi:10.1017/S1355617712001373
Garcia-Cabezas, M. A., Martinez-Sanchez, P., Sanchez-Gonzalez, M. A., Garzon, M., &
Cavada, C. (2009). Dopamine innervation in the thalamus: monkey versus rat.
Cereb Cortex, 19(2), 424-434. doi:10.1093/cercor/bhn093
Giladi, N., McMahon, D., Przedborski, S., Flaster, E., Guillory, S., Kostic, V., & Fahn, S.
(1992). Motor blocks in Parkinson's disease. Neurology, 42(2), 333-339.
Goetz, C. G., Tilley, B. C., Shaftman, S. R., Stebbins, G. T., Fahn, S., Martinez-Martin,
P., . . . Movement Disorder Society, U. R. T. F. (2008). Movement Disorder
Society-sponsored revision of the Unified Parkinson's Disease Rating Scale
(MDS-UPDRS): scale presentation and clinimetric testing results. Mov Disord,
23(15), 2129-2170. doi:10.1002/mds.22340
Goveas, J., O'Dwyer, L., Mascalchi, M., Cosottini, M., Diciotti, S., De Santis, S., . . .
Giannelli, M. (2015). Diffusion-MRI in neurodegenerative disorders. Magn Reson
Imaging, 33(7), 853-876. doi:10.1016/j.mri.2015.04.006

74
Gu, Q., Huang, P., Xuan, M., Xu, X., Li, D., Sun, J., . . . Zhang, M. (2014). Greater loss
of white matter integrity in postural instability and gait difficulty subtype of
Parkinson's disease. Can J Neurol Sci, 41(6), 763-768. doi:10.1017/cjn.2014.34
Hely, M. A., Morris, J. G., Reid, W. G., & Trafficante, R. (2005). Sydney Multicenter
Study of Parkinson's disease: non-L-dopa-responsive problems dominate at 15
years. Mov Disord, 20(2), 190-199. doi:10.1002/mds.20324
Hely, M. A., Reid, W. G., Adena, M. A., Halliday, G. M., & Morris, J. G. (2008). The
Sydney multicenter study of Parkinson's disease: the inevitability of dementia at
20 years. Mov Disord, 23(6), 837-844. doi:10.1002/mds.21956
Hoehn, M. M., & Yahr, M. D. (1967). Parkinsonism: onset, progression and mortality.
Neurology, 17(5), 427-442.
Huang, P., Xu, X., Gu, Q., Xuan, M., Yu, X., Luo, W., & Zhang, M. (2014). Disrupted
white matter integrity in depressed versus non-depressed Parkinson's disease
patients: a tract-based spatial statistics study. J Neurol Sci, 346(1-2), 145-148.
doi:10.1016/j.jns.2014.08.011
Ibarretxe-Bilbao, N., Junque, C., Marti, M. J., Valldeoriola, F., Vendrell, P., Bargallo, N.,
. . . Tolosa, E. (2010). Olfactory impairment in Parkinson's disease and white
matter abnormalities in central olfactory areas: A voxel-based diffusion tensor
imaging study. Mov Disord, 25(12), 1888-1894. doi:10.1002/mds.23208
Jankovic, J., McDermott, M., Carter, J., Gauthier, S., Goetz, C., Golbe, L., . . . et al.
(1990). Variable expression of Parkinson's disease: a base-line analysis of the
DATATOP cohort. The Parkinson Study Group. Neurology, 40(10), 1529-1534.
Kamagata, K., Motoi, Y., Abe, O., Shimoji, K., Hori, M., Nakanishi, A., . . . Hattori, N.
(2012). White matter alteration of the cingulum in Parkinson disease with and
without dementia: evaluation by diffusion tensor tract-specific analysis. AJNR Am
J Neuroradiol, 33(5), 890-895. doi:10.3174/ajnr.A2860
Kanazawa, T., Adachi, E., Orimo, S., Nakamura, A., Mizusawa, H., & Uchihara, T.
(2012). Pale neurites, premature alpha-synuclein aggregates with centripetal
extension from axon collaterals. Brain Pathol, 22(1), 67-78. doi:10.1111/j.1750-
3639.2011.00509.x

75
Karachi, C., Grabli, D., Bernard, F. A., Tande, D., Wattiez, N., Belaid, H., . . . Francois,
C. (2010). Cholinergic mesencephalic neurons are involved in gait and postural
disorders in Parkinson disease. J Clin Invest, 120(8), 2745-2754.
doi:10.1172/JCI42642
Koshimori, Y., Segura, B., Christopher, L., Lobaugh, N., Duff-Canning, S., Mizrahi, R., .
. . Strafella, A. P. (2015). Imaging changes associated with cognitive
abnormalities in Parkinson's disease. Brain Struct Funct, 220(4), 2249-2261.
doi:10.1007/s00429-014-0785-x
Landau, S., Harris, V., Burn, D. J., Hindle, J. V., Hurt, C. S., Samuel, M., . . . Brown, R.
G. (2016). Anxiety and anxious-depression in Parkinson's disease over a 4-year
period: a latent transition analysis. Psychol Med, 46(3), 657-667.
doi:10.1017/S0033291715002196
Langston, J. W. (2006). The Parkinson's complex: parkinsonism is just the tip of the
iceberg. Ann Neurol, 59(4), 591-596. doi:10.1002/ana.20834
Le Bihan, D. (1985). [Microcomputer simulation of nuclear magnetic resonance imaging
contrasts]. J Radiol, 66(4), 303-308.
Lebel, C., Benner, T., & Beaulieu, C. (2012). Six is enough? Comparison of diffusion
parameters measured using six or more diffusion-encoding gradient directions
with deterministic tractography. Magn Reson Med, 68(2), 474-483.
doi:10.1002/mrm.23254
Lee, E., Lee, J. E., Yoo, K., Hong, J. Y., Oh, J., Sunwoo, M. K., . . . Kang, S. Y. (2014).
Neural correlates of progressive reduction of bradykinesia in de novo Parkinson's
disease. Parkinsonism Relat Disord, 20(12), 1376-1381.
doi:10.1016/j.parkreldis.2014.09.027
Lee, J., Lee, KB., Kim, SY., Lee, DW., Choe, BY., Choi, YB. (2011). Evaluation of
Fractional Anisotropy and Apparent Diffusion Coefficient of Broca’s Area in
Patients with Parkinson’s Disease: Quantitative MR Diffusion Tensor Imaging
Study at 3 Tesla. J Korean Physical Soc, 58(2), 343-348. doi:10.3938/jkps.58.343
Lenfeldt, N., Hansson, W., Larsson, A., Nyberg, L., Birgander, R., & Forsgren, L.
(2013). Diffusion tensor imaging and correlations to Parkinson rating scales. J
Neurol, 260(11), 2823-2830. doi:10.1007/s00415-013-7080-2

76
Levy, M. L., Cummings, J. L., Fairbanks, L. A., Masterman, D., Miller, B. L., Craig, A.
H., . . . Litvan, I. (1998). Apathy is not depression. J Neuropsychiatry Clin
Neurosci, 10(3), 314-319. doi:10.1176/jnp.10.3.314
Lewis, S. J., & Barker, R. A. (2009). A pathophysiological model of freezing of gait in
Parkinson's disease. Parkinsonism Relat Disord, 15(5), 333-338.
doi:10.1016/j.parkreldis.2008.08.006
Lewis, S. J., Foltynie, T., Blackwell, A. D., Robbins, T. W., Owen, A. M., & Barker, R.
A. (2005). Heterogeneity of Parkinson's disease in the early clinical stages using a
data driven approach. J Neurol Neurosurg Psychiatry, 76(3), 343-348.
doi:10.1136/jnnp.2003.033530
Li, W., Liu, J., Skidmore, F., Liu, Y., Tian, J., & Li, K. (2010). White matter
microstructure changes in the thalamus in Parkinson disease with depression: A
diffusion tensor MR imaging study. AJNR Am J Neuroradiol, 31(10), 1861-1866.
doi:10.3174/ajnr.A2195
Lim, J. S., Shin, S. A., Lee, J. Y., Nam, H., Lee, J. Y., & Kim, Y. K. (2016). Neural
substrates of rapid eye movement sleep behavior disorder in Parkinson's disease.
Parkinsonism Relat Disord, 23, 31-36. doi:10.1016/j.parkreldis.2015.11.027
Makris, N., Kennedy, D. N., McInerney, S., Sorensen, A. G., Wang, R., Caviness, V. S.,
Jr., & Pandya, D. N. (2005). Segmentation of subcomponents within the superior
longitudinal fascicle in humans: a quantitative, in vivo, DT-MRI study. Cereb
Cortex, 15(6), 854-869. doi:10.1093/cercor/bhh186
Matsui, H., Nishinaka, K., Oda, M., Niikawa, H., Komatsu, K., Kubori, T., & Udaka, F.
(2007a). Depression in Parkinson's disease. Diffusion tensor imaging study. J
Neurol, 254(9), 1170-1173. doi:10.1007/s00415-006-0236-6
Matsui, H., Nishinaka, K., Oda, M., Niikawa, H., Komatsu, K., Kubori, T., & Udaka, F.
(2007b). Wisconsin Card Sorting Test in Parkinson's disease: diffusion tensor
imaging. Acta Neurol Scand, 116(2), 108-112. doi:10.1111/j.1600-
0404.2006.00795.x
Matsui, H., Nishinaka, K., Oda, M., Niikawa, H., Kubori, T., & Udaka, F. (2007).
Dementia in Parkinson's disease: diffusion tensor imaging. Acta Neurol Scand,
116(3), 177-181. doi:10.1111/j.1600-0404.2007.00838.x

77
Melzer, T. R., Watts, R., MacAskill, M. R., Pitcher, T. L., Livingston, L., Keenan, R. J., .
. . Anderson, T. J. (2013). White matter microstructure deteriorates across
cognitive stages in Parkinson disease. Neurology, 80(20), 1841-1849.
doi:10.1212/WNL.0b013e3182929f62
Merboldt, K. D., Hanicke, W., & Frahm, J. (1985). Self-Diffusion Nmr Imaging Using
Stimulated Echoes. J Magn Res, 64(3), 479-486. doi:Doi 10.1016/0022-
2364(85)90111-8
Mole, J. P., Subramanian, L., Bracht, T., Morris, H., Metzler-Baddeley, C., & Linden, D.
E. (2016). Increased fractional anisotropy in the motor tracts of Parkinson's
disease suggests compensatory neuroplasticity or selective neurodegeneration.
Eur Radiol. doi:10.1007/s00330-015-4178-1
Nutt, J. G., Bloem, B. R., Giladi, N., Hallett, M., Horak, F. B., & Nieuwboer, A. (2011).
Freezing of gait: moving forward on a mysterious clinical phenomenon. Lancet
Neurol, 10(8), 734-744. doi:10.1016/S1474-4422(11)70143-0
Ofori, E., Pasternak, O., Planetta, P. J., Li, H., Burciu, R. G., Snyder, A. F., . . .
Vaillancourt, D. E. (2015). Longitudinal changes in free-water within the
substantia nigra of Parkinson's disease. Brain, 138(Pt 8), 2322-2331.
doi:10.1093/brain/awv136
Palacios, E. M., Sala-Llonch, R., Junque, C., Roig, T., Tormos, J. M., Bargallo, N., &
Vendrell, P. (2013). Resting-state functional magnetic resonance imaging activity
and connectivity and cognitive outcome in traumatic brain injury. JAMA Neurol,
70(7), 845-851. doi:10.1001/jamaneurol.2013.38
Pierpaoli, C., Jezzard, P., Basser, P. J., Barnett, A., & Di Chiro, G. (1996). Diffusion
tensor MR imaging of the human brain. Radiol, 201(3), 637-648.
doi:10.1148/radiology.201.3.8939209
Rae, C. L., Correia, M. M., Altena, E., Hughes, L. E., Barker, R. A., & Rowe, J. B.
(2012). White matter pathology in Parkinson's disease: the effect of imaging
protocol differences and relevance to executive function. Neuroimage, 62(3),
1675-1684. doi:10.1016/j.neuroimage.2012.06.012

78
Remy, P., Doder, M., Lees, A., Turjanski, N., & Brooks, D. (2005). Depression in
Parkinson's disease: loss of dopamine and noradrenaline innervation in the limbic
system. Brain, 128(Pt 6), 1314-1322. doi:10.1093/brain/awh445
Rolheiser, T. M., Fulton, H. G., Good, K. P., Fisk, J. D., McKelvey, J. R., Scherfler, C., .
. . Robertson, H. A. (2011). Diffusion tensor imaging and olfactory identification
testing in early-stage Parkinson's disease. J Neurol, 258(7), 1254-1260.
doi:10.1007/s00415-011-5915-2
Rossi, M. E., Ruottinen, H., Saunamaki, T., Elovaara, I., & Dastidar, P. (2014). Imaging
brain iron and diffusion patterns: a follow-up study of Parkinson's disease in the
initial stages. Acad Radiol, 21(1), 64-71. doi:10.1016/j.acra.2013.09.018
Santhouse, A. M., Howard, R. J., & ffytche, D. H. (2000). Visual hallucinatory
syndromes and the anatomy of the visual brain. Brain, 123 ( Pt 10), 2055-2064.
Scherfler, C., Frauscher, B., Schocke, M., Iranzo, A., Gschliesser, V., Seppi, K., . . .
Group, S. (2011). White and gray matter abnormalities in idiopathic rapid eye
movement sleep behavior disorder: a diffusion-tensor imaging and voxel-based
morphometry study. Ann Neurol, 69(2), 400-407. doi:10.1002/ana.22245
Schwab, J. F., & England, A. C. (1968). Projection technique for evaluating surgery in
Parkinson’s disease. Paper presented at the Third symposium on Parkinson’s
disease., Livingstone, Edinburgh.
Schwarz, S. T., Abaei, M., Gontu, V., Morgan, P. S., Bajaj, N., & Auer, D. P. (2013).
Diffusion tensor imaging of nigral degeneration in Parkinson's disease: A region-
of-interest and voxel-based study at 3 T and systematic review with meta-
analysis. Neuroimage Clin, 3, 481-488. doi:10.1016/j.nicl.2013.10.006
Schweder, P. M., Hansen, P. C., Green, A. L., Quaghebeur, G., Stein, J., & Aziz, T. Z.
(2010). Connectivity of the pedunculopontine nucleus in parkinsonian freezing of
gait. Neuroreport, 21(14), 914-916. doi:10.1097/WNR.0b013e32833ce5f1
Sharp, D. J., Beckmann, C. F., Greenwood, R., Kinnunen, K. M., Bonnelle, V., De
Boissezon, X., . . . Leech, R. (2011). Default mode network functional and
structural connectivity after traumatic brain injury. Brain, 134(Pt 8), 2233-2247.
doi:10.1093/brain/awr175

79
Shine, J. M., Halliday, G. M., Gilat, M., Matar, E., Bolitho, S. J., Carlos, M., . . . Lewis,
S. J. (2014). The role of dysfunctional attentional control networks in visual
misperceptions in Parkinson's disease. Hum Brain Mapp, 35(5), 2206-2219.
doi:10.1002/hbm.22321
Shine, J. M., Halliday, G. M., Naismith, S. L., & Lewis, S. J. (2011). Visual
misperceptions and hallucinations in Parkinson's disease: dysfunction of
attentional control networks? Mov Disord, 26(12), 2154-2159.
doi:10.1002/mds.23896
Shine, J. M., Matar, E., Ward, P. B., Bolitho, S. J., Gilat, M., Pearson, M., . . . Lewis, S.
J. (2013). Exploring the cortical and subcortical functional magnetic resonance
imaging changes associated with freezing in Parkinson's disease. Brain, 136(Pt 4),
1204-1215. doi:10.1093/brain/awt049
Skorpil, M., Soderlund, V., Sundin, A., & Svenningsson, P. (2012). MRI diffusion in
Parkinson's disease: using the technique's inherent directional information to
study the olfactory bulb and substantia nigra. J Parkinsons Dis, 2(2), 171-180.
doi:10.3233/JPD-2012-12091
Smith, S. M., Jenkinson, M., Johansen-Berg, H., Rueckert, D., Nichols, T. E., Mackay, C.
E., . . . Behrens, T. E. (2006). Tract-based spatial statistics: voxelwise analysis of
multi-subject diffusion data. Neuroimage, 31(4), 1487-1505.
doi:10.1016/j.neuroimage.2006.02.024
Song, S. K., Sun, S. W., Ju, W. K., Lin, S. J., Cross, A. H., & Neufeld, A. H. (2003).
Diffusion tensor imaging detects and differentiates axon and myelin degeneration
in mouse optic nerve after retinal ischemia. Neuroimage, 20(3), 1714-1722.
Song, S. K., Yoshino, J., Le, T. Q., Lin, S. J., Sun, S. W., Cross, A. H., & Armstrong, R.
C. (2005). Demyelination increases radial diffusivity in corpus callosum of mouse
brain. Neuroimage, 26(1), 132-140. doi:10.1016/j.neuroimage.2005.01.028
Surdhar, I., Gee, M., Bouchard, T., Coupland, N., Malykhin, N., & Camicioli, R. (2012).
Intact limbic-prefrontal connections and reduced amygdala volumes in
Parkinson's disease with mild depressive symptoms. Parkinsonism Relat Disord,
18(7), 809-813. doi:10.1016/j.parkreldis.2012.03.008

80
Svenningsson, P., Westman, E., Ballard, C., & Aarsland, D. (2012). Cognitive
impairment in patients with Parkinson's disease: diagnosis, biomarkers, and
treatment. Lancet Neurol, 11(8), 697-707. doi:10.1016/S1474-4422(12)70152-7
Tessa, C., Giannelli, M., Della Nave, R., Lucetti, C., Berti, C., Ginestroni, A., . . .
Mascalchi, M. (2008). A whole-brain analysis in de novo Parkinson disease.
AJNR Am J Neuroradiol, 29(4), 674-680. doi:10.3174/ajnr.A0900
Tham, W. W., Stevenson, R. J., & Miller, L. A. (2009). The functional role of the medio
dorsal thalamic nucleus in olfaction. Brain Res Rev, 62(1), 109-126.
doi:10.1016/j.brainresrev.2009.09.007
Theilmann, R. J., Reed, J. D., Song, D. D., Huang, M. X., Lee, R. R., Litvan, I., &
Harrington, D. L. (2013). White-matter changes correlate with cognitive
functioning in Parkinson's disease. Front Neurol, 4, 37.
doi:10.3389/fneur.2013.00037
Unger, M. M., Belke, M., Menzler, K., Heverhagen, J. T., Keil, B., Stiasny-Kolster, K., . .
. Knake, S. (2010). Diffusion tensor imaging in idiopathic REM sleep behavior
disorder reveals microstructural changes in the brainstem, substantia nigra,
olfactory region, and other brain regions. Sleep, 33(6), 767-773.
Vaillancourt, D. E., Spraker, M. B., Prodoehl, J., Abraham, I., Corcos, D. M., Zhou, X. J.,
. . . Little, D. M. (2009). High-resolution diffusion tensor imaging in the
substantia nigra of de novo Parkinson disease. Neurology, 72(16), 1378-1384.
doi:10.1212/01.wnl.0000340982.01727.6e
Vercruysse, S., Gilat, M., Shine, J. M., Heremans, E., Lewis, S., & Nieuwboer, A.
(2014). Freezing beyond gait in Parkinson's disease: a review of current
neurobehavioral evidence. Neurosci Biobehav Rev, 43, 213-227.
doi:10.1016/j.neubiorev.2014.04.010
Vercruysse, S., Leunissen, I., Vervoort, G., Vandenberghe, W., Swinnen, S., &
Nieuwboer, A. (2015). Microstructural changes in white matter associated with
freezing of gait in Parkinson's disease. Mov Disord, 30(4), 567-576.
doi:10.1002/mds.26130
Villain, N., Fouquet, M., Baron, J. C., Mezenge, F., Landeau, B., de La Sayette, V., . . .
Chetelat, G. (2010). Sequential relationships between grey matter and white

81
matter atrophy and brain metabolic abnormalities in early Alzheimer's disease.
Brain, 133(11), 3301-3314. doi:10.1093/brain/awq203
Weingarten, C. P., Sundman, M. H., Hickey, P., & Chen, N. K. (2015). Neuroimaging of
Parkinson's disease: Expanding views. Neurosci Biobehav Rev, 59, 16-52.
doi:10.1016/j.neubiorev.2015.09.007
Wiltshire, K., Concha, L., Gee, M., Bouchard, T., Beaulieu, C., & Camicioli, R. (2010).
Corpus callosum and cingulum tractography in Parkinson's disease. Can J Neurol
Sci, 37(5), 595-600.
Yao, N., Cheung, C., Pang, S., Shek-Kwan Chang, R., Lau, K. K., Suckling, J., . . .
McAlonan, G. M. (2014). Multimodal MRI of the hippocampus in Parkinson's
disease with visual hallucinations. Brain Struct Funct. doi:10.1007/s00429-014-
0907-5
Ye, Z., Altena, E., Nombela, C., Housden, C. R., Maxwell, H., Rittman, T., . . . Rowe, J.
B. (2014). Selective serotonin reuptake inhibition modulates response inhibition
in Parkinson's disease. Brain, 137(Pt 4), 1145-1155. doi:10.1093/brain/awu032
Youn, J., Lee, J. M., Kwon, H., Kim, J. S., Son, T. O., & Cho, J. W. (2015). Alterations
of mean diffusivity of pedunculopontine nucleus pathway in Parkinson's disease
patients with freezing of gait. Parkinsonism Relat Disord, 21(1), 12-17.
doi:10.1016/j.parkreldis.2014.10.003
Zetusky, W. J., Jankovic, J., & Pirozzolo, F. J. (1985). The heterogeneity of Parkinson's
disease: clinical and prognostic implications. Neurology, 35(4), 522-526.
Zhan, W., Kang, G. A., Glass, G. A., Zhang, Y., Shirley, C., Millin, R., . . . Schuff, N.
(2012). Regional alterations of brain microstructure in Parkinson's disease using
diffusion tensor imaging. Mov Disord, 27(1), 90-97. doi:10.1002/mds.23917
Zhang, K., Yu, C., Zhang, Y., Wu, X., Zhu, C., Chan, P., & Li, K. (2011). Voxel-based
analysis of diffusion tensor indices in the brain in patients with Parkinson's
disease. Eur J Radiol, 77(2), 269-273. doi:10.1016/j.ejrad.2009.07.032
Zheng, Z., Shemmassian, S., Wijekoon, C., Kim, W., Bookheimer, S. Y., & Pouratian, N.
(2014). DTI correlates of distinct cognitive impairments in Parkinson's disease.
Hum Brain Mapp, 35(4), 1325-1333. doi:10.1002/hbm.22256

82
Zhong, J., Wu, S., Zhao, Y., Chen, H., Zhao, N., Zheng, K., . . . Wu, K. (2013). Why
psychosis is frequently associated with Parkinson's disease? Neural Regen Res,
8(27), 2548-2556. doi:10.3969/j.issn.1673-5374.2013.27.006

CHAPTER3FREEZINGOFGAITANDITSASSOCIATIONSINTHE
EARLYANDADVANCEDCLINICALMOTORSTAGESOF
PARKINSON'SDISEASE
This study is published in Journal of Parkinson’s disease
Hall, J. M., Shine, J. M., Walton, C. C., Gilat, M., O’Callaghan, C., Naismith, S. L., Lewis, S. J. G. Freezing of Gait and its associations in the early and advanced stages of
Parkinson’s disease. Journal of Parkinson’s Disease, 2015.

84
Chapter 3: Freezing of gait and its associations in the early and advanced clinical motor
stages of Parkinson’s disease: a cross-sectional study
Abstract
Background: Freezing of gait is a common disabling symptom of Parkinson’s disease (PD)
with limited treatment options. The pathophysiological mechanisms of freezing behaviour are
still contentious.
Objective: To investigate the prevalence of freezing of gait and its associations with
increasing disease severity to gain a better understanding of the underlying pathophysiology.
Method: This exploratory study included 389 idiopathic PD patients, divided into four
groups; early and advanced PD with freezing of gait, and early and advanced PD without
freezing of gait. Motor, cognitive and affective symptoms, REM sleep behaviour disorder and
autonomic function were assessed.
Results: Regardless of disease stage, patients with freezing of gait had more severe motor
symptoms and a predominant non-tremor phenotype. In the early stages, freezers had a
selective impairment in executive function and had more marked REM sleep behaviour
disorder. Autonomic disturbances were not associated with freezing of gait across early or
advanced disease stages.
Conclusions: These findings support the notion that impairments across the frontostriatal
pathways are intricately linked to the pathophysiology underlying freezing of gait across all
stages of PD. Features of REM sleep behaviour disorder suggest a contribution to freezing
from brainstem pathology but this does not extend to more general autonomic dysfunction.
Key words: Parkinson’s disease, freezing of gait, disease stage, pathophysiology, executive
function

85
Introduction
Freezing of gait (FOG) is a common symptom in Parkinson’s disease (PD) that is defined as
a sudden involuntary motor arrest during walking (Nutt et al., 2011). With limited treatment
options (Walton et al., 2014), FOG has a substantial impact on independence, quality of life
and nursing home placement (Gray et al., 2000; Walton et al., 2015). Previous work has
reported that FOG is present in around 80% of patients in the more advanced stages of PD
whilst only 10% of patients experience this symptom in the early stages (Giladi et al., 2001;
Macht et al., 2007).
A number of phenotypic features have been associated with the presence of FOG. In relation
to motor features, FOG has been commonly associated with a non-tremor dominant subgroup
rather than tremulous PD (Factor et al., 2011; Lewis et al., 2005). Patients with this
phenotype typically experience a more rapid disease progression and have more severe motor
impairments, such as increased muscle rigidity with a concomitant deterioration of gait and
balance (Lewis et al., 2005; Perez-Lloret et al., 2014). In addition, non-tremor dominant
patients typically manifest a selective pattern of executive dysfunction (Lewis et al., 2005)
and a higher risk of dementia (Kehagia et al., 2013).
One recent study has confirmed that these distinct phenotypic differences are present between
patients with and without FOG in the early clinical stages of PD (Hall et al., 2014).
Specifically, patients in this study had disease duration of less than five years and were
Hoehn and Yahr stage 2 or less. Those patients with FOG had a non-tremor motor phenotype
with selective executive impairments but did not differ from non-freezers on measures of
tremor and more general cognition. Moreover, in this sample of early clinical stage patients,
no differences were found on measures of autonomic function, REM sleep behaviour disorder
(RBD) or mood between freezers and non-freezers (Hall et al., 2014). This finding is in
distinction from earlier reports with less stringently selected patients who had more varied
disease stages and durations (Giladi et al., 2006; Lieberman, 2006; Maidan et al., 2010;
Romenets et al., 2012).

86
The finding that FOG was not related to autonomic dysfunction, RBD or mood disorder in
patients with early disease (Hall et al., 2014) suggests that the underlying pathophysiology of
freezing at this stage may be more closely related to disturbances in frontostriatal rather than
brainstem circuitry. However, it must be acknowledged that good evidence exists implicating
the mesencephalic locomotor region and pedunculopontine nuclei (PPN) of the brainstem in
gait and freezing (Fling et al., 2013; Lam et al., 2014; Snijders et al., 2011). Early
involvement of the frontostriatal pathways in FOG, however, contradicts the commonly
accepted Braak hypothesis of disease spread in PD emphasises early brainstem pathology
(Braak et al., 2003). Although Braak’s model elegantly integrates motor and non-motor
symptoms, it is unable to currently account for the appearance of non-motor functions that
precede the motor symptoms (Diederich et al., 2012), suggesting that the pathological
degradation of regions outside the brainstem might be responsible for the complex, non-
motor symptoms of PD. Little is known about whether patients who experience freezing in
the more advanced stages of PD have the same clinical phenotype that has been characterized
for those in the early clinical stages of disease. Furthermore, it is not clear whether non-
freezers in the more advanced stages of PD demonstrate a distinct clinical phenotype that
might also inform the mechanisms underlying FOG. The aim of this exploratory cross-
sectional study was to determine how key phenotypic features, including motor symptoms,
cognition, mood, RBD and dysautonomia might be related to the freezing phenomenon FOG
in the advanced stages (H&Y ≥ 2.5) and if this demonstrates a similar pattern as in the early
stages (H&Y 1-2). We hypothesized that patients with FOG show impairments on variables
related to frontostriatal dysfunctions. Conversely, we expected no differences on variables
related to brainstem pathology in both the early and advanced stages.
Methods
Subjects
Data was prospectively collected from 389 patients between 2009 and 2013 at the
Parkinson’s Disease Research Clinic at the Brain and Mind Research Institute, University of
Sydney. Idiopathic PD was diagnosed based on the UKPDSBB criteria (Hughes et al., 1992)
and confirmed by a trained Neurologist (SJGL). Exclusion criteria included the presence of
other neurological diseases or other conditions that are linked to gait impairment or dementia

87
as rated by the Movement Disorder Society (MDS) Task Force criteria (Emre et al., 2007).
Freezers were firstly identified as those who were observed to freeze during MDS-UPDRS
assessments (question 3.11). Additionally, patients were included if they a positive score on
FOG-Questionnaire item 3 (“Do you feel that your feet get glued to the floor while walking,
making a turn or when trying to initiate walking (freezing)?”) (Giladi et al., 2000). This item
is the most accessible tool and has previously been shown to be a reliable screening
instrument (Shine et al., 2012) and was included to avoid false negative classification that is
common when relying on clinically observed ON-state FOG only. Patients were divided into
four different groups, on the basis of the presence of FOG and the severity of PD according to
the modified Hoehn and Yahr (H&Y) motor clinical staging (Hoehn & Yahr, 1967)
distinguished by postural instability. Patients in H&Y stages 1 and 2 were classified as early
PD patients with FOG (E-FOG) and early PD patients without FOG (E-NF). Patients in H&Y
stages 2.5-5 with FOG were classified as advanced patients with (A-FOG) and without FOG
(A-NF). All patients were assessed on their regular medication and their dopamine dose
equivalence (LEDD; mg/day)(Tomlinson et al., 2010) was recorded. Written informed
consent for this study was obtained from each patient, and the University of Sydney Human
Research and Ethics Committee approved the study.
Data collection
Age and time since diagnosis were recorded for each patient. The Movement Disorder
Society-Unified Parkinson’s Disease Rating Scale (MDS-UPDRS) (Goetz et al., 2008) was
obtained as well as the motor sub-score, excluding question 3.11 (“Freezing of gait”). Using
a previously described method (Lewis et al., 2005), the tremor score was derived from the
average of questions 2.10 and 3.15–3.18 of the MDS-UPDRS and a global score for non-
tremor features was obtained by averaging questions 2.1, 2.9, 2.11, 2.12, 3.1–3.10, 3.12-3.14.
All patients were assessed on the Mini Mental State Examination (MMSE)(Folstein et al.,
1975) to measure general cognitive function for descriptive purposes. More detailed
neuropsychological testing included psychomotor speed (TMTA) and attentional set-shifting
(TMTB) (Army Battery, 1944). To control for psychomotor speed in part B of the TMT, the
time to complete part A was subtracted from the time to complete part B (TMTB-A)(Corrigan

88
et al., 1987). The Digit Span (DS) Forward and Backward sub-tests (total score) were used to
assess attention and working memory and Logical Memory (LM) Retention score (Wechsler
Memory Scale – III) assessed verbal memory (Wechsler, 1997). Verbal (letters F, A, S) and
semantic fluency (Animals) was also assessed (Benton, 1983).
The Hospital Anxiety and Depression Scale (HADS) was obtained to assess affective
function (Snaith et al., 1983). HADS depression and anxiety scores were compared between
the groups, as well as the percentage of patients who scored above the cut-off score of 8,
which indicates clinically significant symptomatology (Snaith et al., 1983).
The RBD Screening Questionnaire (RBDSQ) (Stiasny-Kolster et al., 2007) was used to
investigate symptoms of RBD. The autonomic function score was derived from averaging
questions 1.10, 1.11, 1.12, 1.13 and 2.2 of the MDS-UPDRS.
Statistical analyses
All data analyses were performed using IBM SPSS Version 22. Due to the differences in
sample size and inequalities of variances, Kruskal Wallis tests were used to determine
differences between the four groups. To investigate whether there are any phenotypical
differences between the groups, E-FOG was compared to E-NF, and A-FOG was compared
to A-NF. In addition, to investigate how phenotypical changes progress, E-FOG was
compared to A-FOG and E-NF was compared to A-NF. Significant results were followed up
with pairwise comparisons with a Mann Whitney test to determine which groups were
significantly different from each other. In view of the exploratory nature of the study, p-
values were reported before corrections for multiple comparisons were made due to the
increased risk of false negatives (Rothman, 1990). However, Dunn’s Bonferroni approach
was also implemented to control for multiple comparisons and significant p-values that
survived this correction are marked with a + in Table 2. Furthermore, Jonckheere-Terpstra
tests were used to calculate effect sizes. Gender differences and the presence of RBD,
depression and anxiety were analysed using the Pearson’s Chi-Square test. Z-scores for TMT
(Tombaugh et al., 1996) were calculated and scores for verbal fluency (Tombaugh et al.,
1999) and LM (Wechsler, 1997) were adjusted for age and years of education. Z-scores based

89
on normative data were not computed for DS Forward and Backward tests, as normalized
data was not available for these scores individually. Furthermore, the raw scores of the TMT
were used to measure TMTB-A. In keeping with previous studies, the cut off score for TMTA
was set at 180s and at 300s for TMTB (Thompson et al., 1999). All analyses used an alpha of
0.05 and were two-tailed.
Results
In total 148 patients were classified as non-freezers (38%) whereas 241 patients (62%) were
classified as freezers. The percentages of freezers and non-freezers across all H&Y stages are
presented in Figure 1. Sixty-nine patients were identified as E-FOG, the E-NF group
consisted of 113 patients. The A-FOG consisted of 172 patients and the A-NF of 35 patients.
Clinical characteristics
The medians and 25th and 75th percentiles of demographic data are presented in Table 1.
Table 2 demonstrates the statistical differences between the four groups. As expected,
patients in the advanced stages were older than patients in the early stages. No differences in
age or gender were observed between the FOG and NF patients. Both the freezer groups had
a significantly longer time since diagnosis than their respective non-freezer groups. E-FOG
patients had significantly higher LEDD scores compared to E-NF. Interestingly, LEDD
scores did not differ between the early and advanged stages. Only three E-FOG patients
(5%) had a LEDD score of zero and four of the A-FOG patients (2%) was untreated or
refused to take medication. Seven patients in the E-NF group (7%) and five patients in the
A-NF group (15%) had a LEDD score of zero. In the A-NF group, all unmedicated patients
had discontinued medications due to side-effect of confusion.
Motor function
As demonstrated in Table 1 and 2, both the A-FOG and A-NF groups displayed higher total
MDS-UPDRS, motor MDS-UPDRS-III and non-tremor scores comparing to the E-FOG and
E-NF group respectively. A significant increase in tremor score was observed in A-NF
compared to E-NF group. Compared to E-NF, E-FOG had significantly higher scores on the

90
total MDS-UPDRS, and both FOG groups performed more poorly on the non-tremor
measures compared to the NF groups. Despite higher scores for the NF groups, the tremor
score was not significantly different between FOG and NF patients after controlling for
multiple comparisons.
Figure 1. The percentage of freezers and non-freezers identified with the Freezing of Gait
Questionnaire Item 3 across different Hoehn and Yahr stages (measured in the ON-state) of
the disease.(Number of patient in each group presented in bars).
CognitionCompared to E-FOG patients, the A-FOG group had significantly worse scores on the
MMSE and LM Retention. Compared to E-NF patients, A-NF patients scored significantly
worse on the MMSE and TMTB-A. No other differences were observed between these groups.
E-FOG patients scored more poorly than E-NF patients on the MMSE, TMTA, TMTB, TMTB-
13 56
43 74
39 16
241
35 78
14 16
4 1
148
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
1 2 2.5 3 4 5 Total Modified Hoehn and Yahr clinical motor stages
Frequency of Freezing of Gait by clincal motor stages in the on state Non-
Freezers

91
A, DS-Backward, phonemic and semantic verbal fluency, however, after controlling for the
multivariate approach only the differences on TMTA, phonemic and semantic verbal fluency
performance remained significant between the groups. In the advanced stages, compared to
the A-NF group, the A-FOG group had significantly poorer scores on DS- Backward, but this
difference did not survive after controlling for multiple comparisons. However, due to the
small number of patients in the A-NF group, these results should be interpreted with caution.
Mood
Depression scores increased in both FOG and NF as the disease progressed. In contrast,
anxiety was not related to disease stage (see Table 1 and 2). Freezers in both the early and
advanced stages had higher anxiety and depression scores compared to non-freezers, however
the difference did not reach significance when using a multivariate approach. Clinically
significant anxiety was found in 25.0% in the E-FOG group and in 31.9% of the patients in
the A-FOG group, compared to 9.9% and 17.1% in the E-NF and A-NF groups. In 6.3% of
the E-NF patients and 22.9% of the A-NF patients, clinically significant depression
symptoms were reported, whilst the prevalence of depression in the E-FOG and A-FOG
groups were 17.9% and 39.2% respectively.
Autonomic function and REM Sleep Behaviour Disorder
Compared to early disease stage, both the A-FOG as well as the A-NF group had
significantly higher autonomic scores, but no differences were found between freezers and
non-freezers in both early and advanced disease. RBD was independent of disease stage in
both groups. Both FOG groups had higher RBDSQ scores than than the NF patients, reaching
significance in the early clinical disease stages.

92
Table 1. Characteristics of Parkinson’s disease patients with and without freezing of gait (FOG) in the early and advanced stages (median (25th percentile – 75th percentile) FOG Non-FOG
Early Advanced Early Advanced
N Median N Median N Median N Median
Clinical characteristics
Age (y) 66 65.03 (59.61 – 71.77) 160 75.15 (68.47 – 80.91) 104 66.47 (61.49 – 72.32) 33 78.50 (68.64 – 84.51)
Gender (% male) 69 65.2% 164 59.3% 113 66.4% 35 48.6%
Time since diagnosis (y) 69 6.21 (3.50 – 9.44) 166 9.95 (4.24 – 16.62) 111 3.43 (1.70 – 6.28) 34 4.34 (1.72 – 7.05)
LEDD (mg/day) 63 750.00 (450.00 – 1100.00) 166 675.00 (300.00 – 1045.00) 104 456.25 (300.00 – 774.25) 30 425.00 (300.00 – 785.00)
Minute since last dose 65 120.00 (60.00 – 205.00) 166 120.00 (60.00 – 225.00) 100 120.00 (60.00 – 240.00) 30 115.00 (13.75 – 212.50)
Motor functions
H&Y 69 2.00 (2.00 – 2.00) 172 3.00 (2.63 – 4.00) 113 2.00 (1.00 – 2.00) 35 3.00 (2.50 – 3.00)
MDS- UPDRS total 69 50.00 (37.00 – 67.00) 168 93.50 (62.25 – 117.00) 112 39.00 (27.25 – 50.75) 35 72.00 (56.00 – 89.00)
MSD-UPRDS III 69 29.00 (18.50 – 40.00) 168 51.00 (40.00 – 64.00) 112 21.50 (13.00 – 28.75) 35 43.00 (30.00 – 54.00)
Non-Tremor 69 1.15 (0.69 – 1.52) 169 2.04 (1.54 – 2.54) 112 0.85 (0.50 – 1.12) 35 1.42 (1.08 – 2.00)
Tremor 69 0.27 (0.18 – 0.55) 169 0.45 (0.18 – 0.91) 112 0.36 (0.11 – 0.63) 35 0.64 (0.36 – 1.00)
Ratio score 69 0.43 (0.21 – 0.74) 169 0.32 (0.18 – 0.69) 112 0.56 (0.29 – 1.01) 35 0.43 (0.27 – 0.89)
MDS-UPDRSQ46- FOG 69 1.00 (0.00 -1.00) 169 1.00 (0.00 – 2.00) 113 0.00 (0.00 – 0.00) 35 0.00 (0.00 - 0.00)
FOG-Q item 3 69 2.00 (1.00 – 3.00) 172 3.00 (2.00 – 4.00) 113 0.00 (0.00 – 0.00) 35 0.00 (0.00 – 0.00)
Sleep and autonomic functions

93
LEDD = Levodopa Equivalent Daily Dose; MDS-UPDRS = Movement Disorder Society – Unified Parkinson’s Disease Rating Scale; MDS UPDRS iii = Motor part of the MDS-UPDRS excluding question 46 about freezing of gait; FOG-Q = Freezing of Gait Questionnaire; RBDSQ sub-score = REM Sleep Behaviour Screening Questionnaire, question 3, 6i, 6ii, 6iii; MMSE = Mini Mental State Examination; TMT = Trail Making Test; VF = Verbal Fluency; LM = Logical Memory, ASS = Age Scaled Score; HADS = Hospital Anxiety and Depression Scale. Anxiety Frequency = Percentage of patients with sum HADS Anxiety score ≥ 8; Depression Frequency = Percentage of patients with sum HADS Depression score ≥ 8.
RBDSQ 68 6.00 (3.25 – 9.00) 167 6.00 (3.00 – 9.00) 112 4.00 (2.00 – 7.00) 33 4.00 (2.00 – 6.50)
Autonomic function 69 0.83 (0.58 – 1.17) 169 1.67 (1.00 – 2.33) 113 0.67 (0.33 - 1.17) 35 1.33 (1.00 – 1.83)
Cognitive functions
MMSE 62 28.00 (26.00 – 30.00) 142 26.00 (23.00 – 29.00) 99 29.00 (28.00 – 30.00) 31 28.00 (24.00 – 29.00)
TMTA z-score 55 -0.10 (-1.11 – 0.49) 86 -0.90 (-2.45 – 0.33) 85 0.50 (-0.34 – 0.99) 15 -0.38 (-0.60 – 0.91)
TMTB z-score 54 0.01 (1.15 – 0.43) 73 -0.64 (-2.61 – 0.27) 84 0.31 (-0.32 – 0.80) 15 -0.30 (-1.00 – 0.00)
TMTB-A time 55 55.00 (453.00 – 96.00) 74 85.50 (41.50 – 164.00) 84 41.50 (29.25 – 66.00) 15 64.00 (55.00 – 103.00)
Digit Span Forward 55 10.00 (9.00 – 12.00) 89 10 .00 (8.00 - 11.00) 89 11.00 (9.00 – 12.00) 17 10.00 (9.50 -12.50)
Digit Span Backward 55 6.00 (5.00 – 7.00) 89 5.00 (4.00 – 7.00) 89 7.00 (6.00 – 8.00) 17 7.00 (6.00 – 7.50)
VF Animals z-score 55 -0.50 (-1.14 – 0.67) 87 -0.56 (-1.72 – 0.37) 91 0.40 (-0.43 – 1.14) 16 -0.43 (-0.96 – 0.81)
VF Letters z-score 55 -0.41 (-1.30 – 0.41) 88 -0.71 (-1.56 – 0.37) 92 0.08 (-0.42 – 0.83) 16 -0.25 (-0.77 – 0.89)
LM Retention ASS 55 12.00 (9.00 – 15.00) 87 10.00 (7.00 – 12.00) 83 11.00 (9.00 – 14.00) 16 10.50 (7.50 – 13.00)
Mood
HADS Anxiety 68 4.00 (2.00 – 7.75) 160 5.00 (2.00 – 8.00) 111 3.00 (1.00 – 5.00) 35 3.00 (1.00 – 6.00)
Anxiety Frequency 68 25.0% 160 31.9% 111 9.9% 35 17.1%
HADS Depression 67 3.00 (2.00 – 6.00) 161 6.00 (3.00 – 9.00) 111 2.00 (1.00 – 4.00) 35 3.00 (2.00 – 7.00)
Depression Frequency 67 17.9% 161 39.2 % 111 6.3% 35 22.9%

94
Table 2. Evaluation of clinical characteristics of Parkinson’s disease patients with and without freezing of gait (FOG) in the early and
advanced stages, illustrating the statistical differences between the four groups (Standard Test Statistics, p-values and Jonckheere
effect sizes)
FOG Non-FOG Early stages Advanced stages
Early vs Advanced stages Early vs Advanced stages FOG vs Non-FOG FOG vs Non-FOG
Std. Test
Stat
p-value r Std. Test
Stat
p-value r Std. Test
Stat
p-value r Std. Test
Stat
p-value r
Clinical characteristics
Age (y) -5.820 <.001+ -0.39 -5.022 <.001+ -0.43 0.279 1.000 0.02 1.025 1.000 0.07
Gender (% male) Χ2(1) = 4.354; p = .226
Time since diagnosis (y) -2.926 .003+ -0.19 -1.007 .314 -0.08 -3.549 <.001+ -0.26 -4.068 <.001+ -0.29
LEDD (mg/day) 1.662 .097 0.11 .235 .814 0.02 -3.535 <.001+ -0.27 -1.885 .064 -0.13
Time since last dose (min) H(3) 1.542; p = .673
Motor functions
H&Y -12.346 <.001+ -0.86 -8.469 <.001+ -0.65 -0.665 .506 -0.05 -1.223 .211 -0.12
MDS-UPDRS total -7.749 <.001+ -0.54 -5.990 <.001+ -0.46 -3.117 .002+ -0.23 -2.288 .022 -0.22
MDS-UPRDS III -7.948 <.001+ -0.55 -5.733 <.001+ -0.44 -2.502 .012 -0.19 -2.199 .028 -0.22
Non-Tremor -7.966 <.001+ -0.56 -5.294 <.001+ -0.41 -2.745 .006+ -0.21 -2.870 .004+ -0.28
Tremor -2.197 .028 -0.16 -3.791 <.001+ -0.29 0.166 .868 0.01 2.400 .016 0.24
Ratio score 0.848 .396 0.06 0.849 .396 0.07 1.892 .059 0.14 0.185 1.000 0.13
MDS-UPDRSQ46 - FOG -2.483 .013 -0.17 0.000 1.000 0.00 -6.079 <.001+ -0.45 -6.911 <.001+ -0.68
FOG-Q item 3 -3.371 .001+ -0.24 0.000 1.000 0.00 -8.945 <.001+ -0.66 -9.839 <.001+ -0.96
Sleep and autonomic functions

95
LEDD = Levodopa Equivalent Daily Dose; MDS-UPDRS = Movement Disorder Society – Unified Parkinson’s Disease Rating Scale; MDS UPDRS iii = Motor part of the MDS-UPDRS excluding question 46 about freezing of gait; FOG-Q = Freezing of Gait Questionnaire; RBDSQ = REM Sleep Behaviour Screening Questionnaire; MMSE = Mini Mental State Examination; TMT = Trail Making Test; VF = Verbal Fluency; LM = Logical Memory, ASS = Age Scaled Score; HADS = Hospital Anxiety and Depression Scale. Anxiety Frequency = Percentage of patients with sum HADS Anxiety score ≥ 8; Depression Frequency = Percentage of patients with sum HADS Depression score ≥ 8. + p – values significant after adjustments for multiple comparisons
RBDSQ 0.915 .360 0.06 0.531 .595 0.04 -2.845 .004+ -0.21 -2.157 .031 -0.21
Autonomic functions -6.161 <.001+ -0.43 -4.658 <.001+ -0.36 -1.185 .236 -0.09 -0.863 .388 -0.08
Cognitive functions
MMSE 3.510 <.001+ 0.27 3.271 .001+ 0.26 2.474 .013 0.19 1.321 .187 0.14
TMTA, z-score 1.954 .051 0.19 1.274 .203 0.14 2.790 .005+ 0.21 1.657 .098 0.20
TMTB, z-score 1.747 .081 0.19 2.437 .015 0.29 2.467 .014 0.21 0.830 1.000 0.03
TMTB-A, time -2.187 .029 -0.23 -2.921 .003+ -0.34 -2.263 .024 -0.19 0.130 .896 0.02
DS Forward 1.431 .251 0.14 0.055 .955 0.01 1.906 .078 0.16 2.096 .067 0.25
DS Backward 1.167 .380 0.12 0.007 .994 0.00 2.403 .023 0.20 2.295 .042 0.27
VF Animals 1.660 .109 0.17 1.738 .083 0.19 2.789 .005+ 0.24 1.036 .288 0.12
VF Letters 1.118 .288 0.11 0.889 .378 0.10 2.986 .003+ 0.25 1.665 .091 0.20
LM Retention ASS 3.113 .007+ 0.31 0.951 .339 0.10 -0.105 .758 -0.01 0.784 .572 0.09
Mood
HADS Anxiety -1.173 .241 -0.08 -0.402 .688 -0.03 -2.012 .044 -0.15 -2.153 .031 -0.21
Anxiety Frequency Χ2(1) =
1.078
.299 Χ2(1) =
1.353
.245 Χ2(1)
=
7.277
.007 Χ2(1)
=
3.013
.083
HADS Depression -3.998 < 001+ -0.29 -2.968 .003+ -0.23 -2.492 .013 -0.19 -2.095 .036 -0.21
Depression Frequency Χ2(1) =
11.740
.001 Χ2(1) =
7.907
.005 Χ2(1)
=
5.901
.015 Χ2(1)
=
4.282
.039

96
Discussion
This large-scale cross-sectional study investigated the association between different
characteristics of PD and FOG relative to disease severity. Here, we provide an in-depth
comparison of the clinical phenotype of patients with and without FOG, at early versus
advanced clinical disease stages. In doing so, a number of critical novel findings have
emerged, which inform possible mechanistic explanations of the emergence of FOG
across disease stages. Regardless of disease stage, patients with FOG performed worse
on their motor assessment particularly in relation to non-tremor symptoms. However,
freezers and non-freezers demonstrated similar tremor scores in both clinical stages, but
an increase in tremor score between the clinical stages was only found in non-freezers.
Although tremor and FOG share some characteristics such as an increased prevalence
during mental stress and an episodic character, there are significant distinct mechanisms
underlying each symptom (Lewis et al., 2005). Specifically, the cerebello-thalamo-
cortical circuit is likely to be involved in the incidence of tremor (Helmich et al., 2012)
rather than arising from frontostriatal pathways.
Previous studies have demonstrated a selective cognitive impairment in freezers that
typically affects executive functions such as verbal fluency (Amboni et al., 2008),
conflict resolution (Vandenbossche et al., 2012; Walton et al., 2014) and set-shifting
(Shine et al., 2012). Indeed, the current study demonstrated that FOG patients scored
worse on measures for phonemic and semantic verbal fluency in the early stages.
Additionally, though non-significant after controlling for multiple comparisons, freezers
performed more poorly on set shifting in the early stages of the disease and working
memory in both the early and advanced stages. In contrast, memory retention
performance, which is known to reflect temporal lobe function, did not differ between
groups across disease stages, thus reinforcing the possible selectivity of frontostriatal
impairment in FOG. Although current results do not show a clear division between FOG
and NF patients, a difference between these patient groups is plausible and in accordance
with recent work using functional neuroimaging techniques. Namely, it has been shown
that there is a paroxysmal functional decoupling of activity between neural networks
including the cognitive control network and the basal ganglia in patients with FOG

97
(Shine, 2013a). Such impaired information processing in the frontostriatal system might
be influenced by a faulty selective gating mechanism across the basal ganglia, which
would normally be responsible for the rapid updating of information (van Schouwenburg
et al., 2010). However, it is important to note that not all freezers present with executive
dysfunction, pointing out the likelihood of different causes of FOG (Ferraye et al., 2013).
Indeed, FOG is not due to executive dysfunction per se, but rather to impairments in
conflict processing independent of the processing system of which the conflict occurred.
As such, motor, cognitive, affective or perceptual conflict may underlie freezing
behaviour, given that dysfunctional activity within each of these circuits is processed via
a final common neural pathway, i.e. the subthalamic nucleus (Lewis & Shine, 2016).
The current study demonstrated that in contrast with depression, anxiety is not related to
disease stage. Freezers showed a trend towards significantly higher average anxiety
scores. This is in accordance with an elegant study recently published, showing
differences on depression and anxiety scores between FOG and non-FOG patient.
However, this difference disappeared when using a multivariate approach (Perez-Lloret et
al., 2014). The exact etiology of mood disturbances remains unknown (Aarsland et al.,
2012). The interplay of emotion and FOG might have an underlying neurobiological basis
in shared frontostriatal limbic circuitry, an interpretation consistent with a SPECT study
that found that bilateral ventromedial prefrontal cortex showed decreased perfusion in
freezers compared to non-freezers (Rahman et al., 2008). However, previous work has
also highlighted involvement of the locus coeruleus (Gesi et al., 2000; Pontone et al.,
2009). Therefore, as the current results are inconclusive, the association between freezing
and anxiety and depression needs further exploration, as well as its influence on other
variables associated with FOG, such as executive functioning, methods of assessments
and cut-off scores, and their underlying pathophysiology. A relationship between these
phenomena has potentially significant implications for future treatment strategies, such as
targeted cognitive-behavioural therapy that would be designed to manage anxiety
(Proudfoot et al., 2004), a non-pharmacological approach that would be advantageous in
a patient population often requiring polypharmacy (Walton et al., 2014).

98
Higher RBD scores found in this study are in contrast with our previous study that used
more stringent inclusion criteria, pointing out a role of disease progression. However,
only in the early stages the RBD scores remained significantly different between the FOG
and Non-FOG groups when using a multivariate approach. RBD suggest brainstem
pathology (Grinberg et al., 2010), yet associations with executive dysfunction and mild
cognitive impairment have been frequently reported (Gagnon et al., 2012; Gagnon et al.,
2009). Finally, autonomic functions did not differ between FOG and NF regardless of
disease stage. These findings suggest that the pathological processes within the brainstem
underlying autonomic functions are not intrinsically associated with freezing behaviour
(Brooks et al., 2010).
Braak and colleagues have identified distinct stages in the progression of PD that spreads
rostrally from several nuclei in the brainstem with little inter-individual variability (Braak
et al., 2003). Despite this pattern of progression, disease heterogeneity is well recognised
in PD. Specifically, in relation to FOG, the results of our study do not indicate that early
or advanced brainstem pathology uniquely contributes to FOG. Indeed, a recent positron
emission tomography study demonstrated that the effects of cholinergic degeneration
associated with FOG were driven by neocortical degeneration but not by PPN-thalamic
degeneration, which was more associated with postural reflex impairments (Bohnen et
al., 2014). This finding would be aligned with our observations and suggests that
pathology within the PPN by itself is unlikely to be the underlying cause of FOG in PD.
Whilst the partial amelioration of FOG by dopaminergic agents presumably operating
through frontostriatal networks is well documented (Nantel et al., 2014; Schaafsma et al.,
2003), there is evidence for a dopamine resistant form of freezing (Espay et al., 2012;
Perez-Lloret et al., 2014). Furthermore, cholinergic degeneration in PD has been
associated with a non-tremor dominant phenotype (Bohnen et al., 2011) along with
executive and attentional dysfunction (Voytko, 1996), depression and apathy (Bohnen et
al., 2007). These observations highlight the complex interplay required between
complementary networks and neurotransmitter systems that when compromised lead to

99
FOG via dysfunction across cortical and subcortical regions. Furthermore, the
paroxysmal nature of FOG suggests that FOG does not result from structural lesions, but
rather from bouts of decreased neuronal activity. Indeed, FOG is best conceptualized as a
functional disorder with multiple possible pathological underpinnings across a range of
different areas in the brain.
Limitations and further directions
Whilst the current study raises several important clinical issues in relation to FOG in PD,
it does suffer with a number of limitations. Group sizes were not equal and the data was
only attained at one time point. Therefore, it was not possible to determine when FOG
had initially presented in the different groups. As such, more accurate temporal
relationships between neuropathology and phenotypic features could not be explored. All
patients were assessed on their optimal ON state and no differences were observed for
time since last dose. However, it is hard to elicit ON-state FOG in the clinical setting;
Levodopa helps in reducing the frequency and duration of 'off'-related FOG (Schaafsma
et al., 2007), and thus many patients report significant FOG at home (Aarsland et al.,
2012) (Schaafsma et al., 2003). Therefore, we have included a subjective questionnaire to
identify freezers. Future studies will be necessary to assess the effect of dopaminergic
medication, including potential wearing-off phenomena along with other neurotransmitter
disturbances in PD patients with FOG. Furthermore, the inability to match subgroups on
all demographic features may also have influenced our findings, but it should be
acknowledged that it might not be possible to control for the effects of FOG, such as the
use of higher dopaminergic therapy in an attempt to reduce freezing episodes. Both FOG
groups had a significantly longer time since diagnosis. Therefore, future longitudinal
studies observing the evolution of FOG within patients may offer important further
insights to those offered here. In addition, the executive functioning scores should be
interpreted with caution. Only 15-17 patients in the A-NF group performed these tests
and not all differences remained significant after controlling for multiple comparisons.
Finally, whilst we acknowledge that using solely behavioural tests in this study limits our
speculation regarding the pathophysiology underlying FOG, our results are aligned with
previous functional imaging data (Shine et al., 2011; 2013b).

100
Conclusions
In conclusion, this study suggests that FOG is associated with non-tremor motor severity,
selective executive dysfunction, mood disturbance and RBD but not more general
autonomic dysfunction or tremor. In patients with more severe tremor, the frontostriatal
pattern of neuropathology might be less affected in these patients compared to patients in
the non-tremor phenotype. The current findings do not support the notion that distinctly
different pathophysiological processes may underlie the freezing phenomenon across
advancing disease stages, however the differences between the groups become less strict
with progressing pathology. Future prospective clinicopathological studies may help
better understand these associations and could provide insights into novel therapies.
Acknowledgements
We wish to thank our patient volunteers of the Parkinson’s Disease Research Clinic for their
time and efforts.
Financial disclosures: JM Hall is supported by a University of Western Sydney
Postgraduate Research Award. JM Shine is supported by a NHMRC CJ Martin Fellow
Award. C O’Callaghan is supported by a NHMRC Neil Hamilton Fairley Award. CC
Walton is supported by an Australian Postgraduate Award at the University of Sydney. M
Gilat is supported by an International Postgraduate Research Scholarship at the
University of Sydney. SL Naismith is supported by a National Health and Medical
Research Council Career Development Award No. 1008117. SJG Lewis is supported by a
National Health and Medical Research Council Practitioner Fellowship No. 1003007.
Conflict of interest: The authors report no conflicts of interest.

101
References
Aarsland, D., Pahlhagen, S., Ballard, C. G., Ehrt, U., & Svenningsson, P. (2012).
Depression in Parkinson disease-epidemiology, mechanisms and management.
Nat Rev Neurol, 8(1), 35-47. doi:DOI 10.1038/nrneurol.2011.189
Amboni, M., Cozzolino, A., Longo, K., Picillo, M., & Barone, P. (2008). Freezing of gait
and executive functions in patients with Parkinson's disease. Mov Disord, 23(3),
395-400. doi:10.1002/mds.21850
Army Battery (1944). Manual of Directions and Scoring. Washington, DC: War
Department, Adjutant General’s Office.
Benton, A. L., Hamsher, K., Sivan, A. B. . (1983). Multilingual aphasia examination (3rd
ed.). Iowa City, IA AJA Associates.
Bohnen, N. I., & Albin, R. L. (2011). The cholinergic system and Parkinson disease.
Behav Brain Res, 221(2), 564-573. doi:DOI 10.1016/j.bbr.2009.12.048
Bohnen, N. I., Frey, K. A., Studenski, S., Kotagal, V., Koeppe, R. A., Constantine, G. M.,
. . . Muller, M. L. (2014). Extra-nigral pathological conditions are common in
Parkinson's disease with freezing of gait: An in vivo positron emission
tomography study. Mov Disord, 29(9), 1118-1124. doi:10.1002/mds.25929
Bohnen, N. I., Kaufer, D. I., Hendrickson, R., Constantine, G. M., Mathis, C. A., &
Moore, R. Y. (2007). Cortical cholinergic denervation is associated with
depressive symptoms in Parkinson's disease and parkinsonian dementia. J Neurol
Neurosurg Psychiatry, 78(6), 641-643. doi:DOI 10.1136/jnnp.2006.100073
Braak, H., Del Tredici, K., Rub, U., de Vos, R. A., Jansen Steur, E. N., & Braak, E.
(2003). Staging of brain pathology related to sporadic Parkinson's disease.
Neurobiol Aging, 24(2), 197-211.
Brooks, D. J., & Pavese, N. (2010). Imaging non-motor aspects of Parkinson's disease.
Prog Brain Res, 184, 205-218. doi:10.1016/S0079-6123(10)84011-7
Corrigan, J. D., & Hinkeldey, N. S. (1987). Relationships between parts A and B of the
Trail Making Test. J Clin Psychol, 43(4), 402-409.
Diederich, N. J., & Parent, A. (2012). Parkinson's disease: acquired frailty of archaic
neural networks? J Neurol Sci, 314(1-2), 143-151. doi:10.1016/j.jns.2011.10.003

102
Emre, M., Aarsland, D., Brown, R., Burn, D. J., Duyckaerts, C., Mizuno, Y., . . . Dubois,
B. (2007). Clinical diagnostic criteria for dementia associated with Parkinson's
disease. Mov Disord, 22(12), 1689-1707; quiz 1837. doi:10.1002/mds.21507
Espay, A. J., Fasano, A., van Nuenen, B. F., Payne, M. M., Snijders, A. H., & Bloem, B.
R. (2012). "On" state freezing of gait in Parkinson disease: a paradoxical
levodopa-induced complication. Neurology, 78(7), 454-457.
doi:10.1212/WNL.0b013e3182477ec0
Factor, S. A., Steenland, N. K., Higgins, D. S., Molho, E. S., Kay, D. M., Montimurro, J.,
. . . Payami, H. (2011). Postural instability/gait disturbance in Parkinson's disease
has distinct subtypes: an exploratory analysis. J Neurol Neurosurg Psychiatry,
82(5), 564-568. doi:10.1136/jnnp.2010.222042
Ferraye, M. U., Ardouin, C., Lhommee, E., Fraix, V., Krack, P., Chabardes, S., . . . Debu,
B. (2013). Levodopa-resistant freezing of gait and executive dysfunction in
Parkinson's disease. Eur Neurol, 69(5), 281-288. doi:10.1159/000346432
Fling, B. W., Cohen, R. G., Mancini, M., Nutt, J. G., Fair, D. A., & Horak, F. B. (2013).
Asymmetric pedunculopontine network connectivity in parkinsonian patients with
freezing of gait. Brain, 136, 2405-2418. doi:Doi 10.1093/Brain/Awt172
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). "Mini-mental state". A practical
method for grading the cognitive state of patients for the clinician. J Psychiatr
Res, 12(3), 189-198.
Gagnon, J. F., Bertrand, J. A., & Genier Marchand, D. (2012). Cognition in rapid eye
movement sleep behavior disorder. Front Neurol, 3, 82.
doi:10.3389/fneur.2012.00082
Gagnon, J. F., Vendette, M., Postuma, R. B., Desjardins, C., Massicotte-Marquez, J.,
Panisset, M., & Montplaisir, J. (2009). Mild cognitive impairment in rapid eye
movement sleep behavior disorder and Parkinson's disease. Ann Neurol, 66(1),
39-47. doi:10.1002/ana.21680
Gesi, M., Soldani, P., Giorgi, F. S., Santinami, A., Bonaccorsi, I., & Fornai, F. (2000).
The role of the locus coeruleus in the development of Parkinson's disease.
Neurosci Biobehav Rev, 24(6), 655-668.

103
Giladi, N., & Hausdorff, J. M. (2006). The role of mental function in the pathogenesis of
freezing of gait in Parkinson's disease. J Neurol Sci, 248(1-2), 173-176.
Giladi, N., Shabtai, H., Simon, E. S., Biran, S., Tal, J., & Korczyn, A. D. (2000).
Construction of freezing of gait questionnaire for patients with Parkinsonism.
Parkinsonism Relat Disord, 6(3), 165-170.
Giladi, N., Treves, T. A., Simon, E. S., Shabtai, H., Orlov, Y., Kandinov, B., . . .
Korczyn, A. D. (2001). Freezing of gait in patients with advanced Parkinson's
disease. J Neural Transm, 108(1), 53-61.
Goetz, C. G., Tilley, B. C., Shaftman, S. R., Stebbins, G. T., Fahn, S., Martinez-Martin,
P., . . . Movement Disorder Society, U. R. T. F. (2008). Movement Disorder
Society-sponsored revision of the Unified Parkinson's Disease Rating Scale
(MDS-UPDRS): scale presentation and clinimetric testing results. Mov Disord,
23(15), 2129-2170. doi:10.1002/mds.22340
Gray, P., & Hildebrand, K. (2000). Fall risk factors in Parkinson's disease. J Neurosci
Nurs, 32(4), 222-228.
Grinberg, L. T., Rueb, U., Alho, A. T. D., & Heinsen, H. (2010). Brainstem pathology
and non-motor symptoms in PD. J Neurol Sci, 289(1-2), 81-88. doi:DOI
10.1016/j.jns.2009.08.021
Hall, J. M., Shine, J. M., Walton, C. C., Gilat, M., Kamsma, Y. P., Naismith, S. L., &
Lewis, S. J. (2014). Early phenotypic differences between Parkinson's disease
patients with and without freezing of gait. Parkinsonism Relat Disord, 20(6), 604-
607. doi:10.1016/j.parkreldis.2014.02.028
Helmich, R. C., Hallett, M., Deuschl, G., Toni, I., & Bloem, B. R. (2012). Cerebral
causes and consequences of parkinsonian resting tremor: a tale of two circuits?
Brain, 135, 3206-3226. doi:Doi 10.1093/Brain/Aws023
Hoehn, M. M., & Yahr, M. D. (1967). Parkinsonism: onset, progression and mortality.
Neurology, 17(5), 427-442.
Hughes, A. J., Daniel, S. E., Kilford, L., & Lees, A. J. (1992). Accuracy of clinical
diagnosis of idiopathic Parkinson's disease: a clinico-pathological study of 100
cases. J Neurol Neurosurg Psychiatry, 55(3), 181-184.

104
Kehagia, A. A., Barker, R. A., & Robbins, T. W. (2013). Cognitive impairment in
Parkinson's disease: the dual syndrome hypothesis. Neurodegener Dis, 11(2), 79-
92. doi:10.1159/000341998
Lam, S., Moro, E., Poon, Y. Y., Lozano, A. M., & Fasano, A. (2014). Does dominant
pedunculopontine nucleus exist? Brain. doi:10.1093/brain/awu225
Lewis, S. J. G., Foltynie, T., Blackwell, A. D., Robbins, T. W., Owen, A. M., & Barker,
R. A. (2005). Heterogeneity of Parkinson's disease in the early clinical stages
using a data driven approach. J Neurol Neurosurg Psychiatry, 76(3), 343-348.
doi:10.1136/jnnp.2003.033530
Lewis, S. J. G. & Shine, J. M. (2016). The Next Step: A Common Neural Mechanism for
Freezing of Gait. Neuroscientist. doi:10.1177/1073858414559101
Lieberman, A. (2006). Are freezing of gait (FOG) and panic related? J Neurol Sci, 248(1-
2), 219-222. doi:10.1016/j.jns.2006.05.023
Macht, M., Kaussner, Y., Moller, J. C., Stiasny-Kolster, K., Eggert, K. M., Kruger, H. P.,
& Ellgring, H. (2007). Predictors of freezing in Parkinson's disease: a survey of
6,620 patients. Mov Disord, 22(7), 953-956. doi:10.1002/mds.21458
Maidan, I., Plotnik, M., Mirelman, A., Weiss, A., Giladi, N., & Hausdorff, J. M. (2010).
Heart rate changes during freezing of gait in patients with Parkinson's disease.
Mov Disord, 25(14), 2346-2354. doi:10.1002/mds.23280
Nantel, J., & Bronte-Stewart, H. (2014). The effect of medication and the role of postural
instability in different components of freezing of gait (FOG). Parkinsonism Relat
Disord, 20(4), 447-451. doi:10.1016/j.parkreldis.2014.01.017
Nutt, J. G., Bloem, B. R., Giladi, N., Hallett, M., Horak, F. B., & Nieuwboer, A. (2011).
Freezing of gait: moving forward on a mysterious clinical phenomenon. Lancet
Neurol, 10(8), 734-744. doi:10.1016/S1474-4422(11)70143-0
Perez-Lloret, S., Negre-Pages, L., Damier, P., Delval, A., Derkinderen, P., Destee, A., . . .
Rascol, O. (2014). Prevalence, determinants, and effect on quality of life of
freezing of gait in Parkinson disease. JAMA Neurol, 71(7), 884-890.
doi:10.1001/jamaneurol.2014.753
Pontone, G. M., Williams, J. R., Anderson, K. E., Chase, G., Goldstein, S. A., Grill, S., . .
. Marsh, L. (2009). Prevalence of anxiety disorders and anxiety subtypes in

105
patients with Parkinson's disease. Mov Disord, 24(9), 1333-1338.
doi:10.1002/mds.22611
Proudfoot, J., Ryden, C., Everitt, B., Shapiro, D. A., Goldberg, D., Mann, A., . . . Gray, J.
A. (2004). Clinical efficacy of computerised cognitive-behavioural therapy for
anxiety and depression in primary care: randomised controlled trial. Br J
Psychiatry, 185, 46-54.
Rahman, S., Griffin, H. J., Quinn, N. P., & Jahanshahi, M. (2008). The factors that induce
or overcome freezing of gait in Parkinson's disease. Behav Neurol, 19(3), 127-
136.
Romenets, S. R., Gagnon, J. F., Latreille, V., Panniset, M., Chouinard, S., Montplaisir, J.,
& Postuma, R. B. (2012). Rapid eye movement sleep behavior disorder and
subtypes of Parkinson's disease. Mov Disord, 27(8), 996-1003.
doi:10.1002/mds.25086
Rothman, K. J. (1990). No adjustments are needed for multiple comparisons.
Epidemiology, 1(1), 43-46.
Schaafsma, J. D., Balash, Y., Gurevich, T., Bartels, A. L., Hausdorff, J. M., & Giladi, N.
(2003). Characterization of freezing of gait subtypes and the response of each to
levodopa in Parkinson's disease. Eur J Neurol, 10(4), 391-398.
Schaafsma, J. D., Balash, Y., Gurevich, T., Bartels, A. L., Hausdorff, J. M., & Giladi, N.
(2007). Characterization of freezing of gait subtypes and the response of each to
levodopa in Parkinson's disease. Eur J Neurol, 10(4), 391-398.
Shine, J. M., Matar, E., Ward, P. B., Bolitho, S. J., Gilat, M., Pearson, M., . . . Lewis, S.
J. G. (2013). Exploring the cortical and subcortical fMRI changes associated with
freezing in Parkinson's disease. Brain, 136(4), 1204-1215.
Shine, J. M., Matar, E., Ward, P. B., Frank, M. J., Moustafa, A. A., Pearson, M., . . .
Lewis, S. J. (2013). Freezing of gait in Parkinson's disease is associated with
functional decoupling between the cognitive control network and the basal
ganglia. Brain, 136(Pt 12), 3671-3681. doi:10.1093/brain/awt272
Shine, J. M., Moore, S. T., Bolitho, S. J., Morris, T. R., Dilda, V., Naismith, S. L., &
Lewis, S. J. G. (2012). Assessing the utility of Freezing of Gait Questionnaires in

106
Parkinson's Disease. Parkinsonism Relat Disord, 18(1), 25-29.
doi:10.1016/j.parkreldis.2011.08.002
Shine, J. M., Naismith, S. L., Palavra, N. C., Lewis, S. J., Moore, S. T., Dilda, V., &
Morris, T. R. (2012). Attentional set-shifting deficits correlate with the severity of
freezing of gait in Parkinson's disease. Parkinsonism Relat Disord, 19(3), 388-
390. doi:10.1016/j.parkreldis.2012.07.015
Shine, J. M., Ward, P. B., Naismith, S. L., Pearson, M., & Lewis, S. J. G. (2011).
Utilising functional MRI (fMRI) to explore the freezing phenomenon in
Parkinson's disease. J Clin Neurosci, 18(6), 807-810. doi:S0967-5868(11)00110-
X [pii] 10.1016/j.jocn.2011.02.003
Snaith, R. P., & Zigmond, A. S. (1983). The hospital anxiety and depression scale. Br
Med J (Clin Res Ed), 292(6516), 344.
Snijders, A. H., Leunissen, I., Bakker, M., Overeem, S., Helmich, R. C., Bloem, B. R., &
Toni, I. (2011). Gait-related cerebral alterations in patients with Parkinson's
disease with freezing of gait. Brain, 134(Pt 1), 59-72. doi:10.1093/brain/awq324
Stiasny-Kolster, K., Mayer, G., Schafer, S., Moller, J. C., Heinzel-Gutenbrunner, M., &
Oertel, W. H. (2007). The REM sleep behavior disorder screening questionnaire--
a new diagnostic instrument. Mov Disord, 22(16), 2386-2393.
doi:10.1002/mds.21740 [doi]
Thompson, M. D., Scott, J. G., Dickson, S. W., Schoenfeld, J. D., Ruwe, W. D., &
Adams, R. L. (1999). Clinical utility of the Trail Making Test practice time. Clin
Neuropsychol, 13(4), 450-455. doi:10.1076/1385-4046(199911)13:04;1-Y;FT450
Tombaugh, T. N., Kozak, J., & Rees, L. (1999). Normative data stratified by age and
education for two measures of verbal fluency: FAS and animal naming. Arch Clin
Neuropsychol, 14(2), 167-177.
Tombaugh, T. N., Rees, L., & McIntyre, N. (1996). Normative data for the trail making
test In A Compendium of Neuropsychological Tests: Administration, Norms and
Commentary, 2nd ed New York: Oxford University Press.
Tomlinson, C. L., Stowe, R., Patel, S., Rick, C., Gray, R., & Clarke, C. E. (2010).
Systematic review of levodopa dose equivalency reporting in Parkinson's disease.
Mov Disord, 25(15), 2649-2653. doi:10.1002/mds.23429

107
van Schouwenburg, M. R., den Ouden, H. E., & Cools, R. (2010). The human basal
ganglia modulate frontal-posterior connectivity during attention shifting. J
Neurosci, 30(29), 9910-9918. doi:10.1523/JNEUROSCI.1111-10.2010
Vandenbossche, J., Deroost, N., Soetens, E., Zeischka, P., Spildooren, J., Vercruysse, S., .
. . Kerckhofs, E. (2012). Conflict and freezing of gait in Parkinson's disease:
support for a response control deficit. Neurosci, 206, 144-154.
doi:10.1016/j.neuroscience.2011.12.048
Voytko, M. L. (1996). Cognitive functions of the basal forebrain cholinergic system in
monkeys: memory or attention? Behav Brain Res, 75(1-2), 13-25.
Walton, C. C., Shine, J. M., Hall, J. M., O'Callaghan, C., Mowszowski, L., Gilat, M., . . .
Lewis, S. J. (2015). The major impact of freezing of gait on quality of life in
Parkinson's disease. J Neurol, 262(1), 108-115. doi:10.1007/s00415-014-7524-3
Walton, C. C., Shine, J. M., Mowszowski, L., Gilat, M., Hall, J. M., O'Callaghan, C., . . .
Lewis, S. J. (2014). Impaired cognitive control in Parkinson's disease patients
with freezing of gait in response to cognitive load. J Neural Transm.
doi:10.1007/s00702-014-1271-6
Walton, C. C., Shine, J. M., Mowszowski, L., Naismith, S. L., & Lewis, S. J. (2014).
Freezing of gait in Parkinson's disease: current treatments and the potential role
for cognitive training. Restor Neurol Neurosci, 32(3), 411-422. doi:10.3233/RNN-
130370
Wechsler, D. (1997). Wechsler Memory Scale. San Antonio: The Psychological
Corporation.

108
Box 2a: Evidence for the association between freezing of gait and executive
dysfunction
Walton, C. C., Shine, J. M., Mowszowski, L., Gilat, M., Hall, J. M., O'Callaghan, C.,
Naismith, S. L., Lewis, S. J. G. Impaired cognitive control in Parkinson's disease
patients with freezing of gait in response to cognitive load. Journal of Neural
Transmission, 2015
In this study, we further explored the association between freezing of gait and executive
dysfunction and showed specific deficits in inhibition and cognitive control. Specifically,
we explored self-monitoring ability of performance using the colour word interference
test. During the inhibition task, patients showed more self corrected, but not uncorrected
errors compared to non-freezers. However, when the switching element was introduced,
freezers were unable to self-correct their errors. These results indicated that freezers fail
to effectively monitor their performance with increasing cognitive load. Therefore,
patients with freezing of gait may use additional cognitive resources to perform difficult
tasks, impeding mechanisms to monitor errors.
Box 2b: Evidence for the association between freezing of gait and executive
dysfunction
Walton, C. C., O'Callaghan, C., Hall, J. M., Gilat, M., Mowszowski, L., Naismith, S.
L., Burrell, J. R., Shine, J. M., Lewis, S. J. G. Antisaccade errors reveal cognitive
control deficits in Parkinson's disease with freezing of gait. Journal of Neurology,
2015
In this study, we assessed antisaccade performance between groups and found no
differences on the simple saccade task, whilst freezers performed more poorly on the anti-
saccade task compared to non-freezers, showing disruptions with inhibiting the reflexitve
saccade. Furthermore, a reduction in grey matter density in the fronto-parietal regions and
occipital cortex was related to the anti-saccade task performance, a finding that supports
the involvement of cortical disruptions in freezing of gait.

109
Box 3: Evidence for the association between freezing of gait and mood
disturbances
Ehgoetz Martens, K. A., Hall, J. M., Gilat, M., Georgiades, M. J., Walton, C. C., Lewis, S.
J. G. Anxiety is associated with freezing of gait and attentional set-shifting in Parkinson’s
disease: A new perspective for early intervention. Gait and Posture 2016
Using a similar cohort of patients as presented in Chapter 3, this study further explored the
association between anxiety and the freezing of gait. We showed that freezers reported
significantly higher anxiety scores compared to non-freezers, and this effect remained
significant even when controlled for age disease duration, severity, global cognitive
andexecutive functioning and depression. Furthermore, both anxiety and depression were
correlated to set-shifting and freezing of gait. Finally, anxiety and depression were
significant predictors of freezing. In the presence of clinical anxiety, a patient was 3.4 times
more likely to have freezing of gait. However, both anxiety and depression as well as
executive dysfunction were not present in all freezers, therefore, these symptoms do not
entirely explain freezing. It can be hypothesized that diffuse cortical and sub-cortical
changes contribute to ineffective information processing, leading to functional disturbances,
such as freezing of gait.

CHAPTER4ALTERATIONSINWHITEMATTERNETWORKTOPOLOGY
CONTRIBUTETOFREEZINGOFGAITINPARKINSON’S
DISEASE
This study is published in Journal of Neurology
Hall, J. M., Shine, J. M., Ehgoetz Martens, K. A., Gilat, M., Broadhouse, K. M., Szeto, J.
Y. Y., Walton, C. C., Moustafa, A. A., Lewis, S. J. G. Alterations in white matter network
topology contribute to freezing of gait in Parkinson’s disease. Journal of Neurology,
2018.

111
Chapter 4: Alterations in white matter network topology contribute to freezing of
gait in Parkinson’s disease
Abstract
Freezing of gait (FOG) is a common symptom in advanced Parkinson’s disease (PD).
Despite current advances, the neural mechanisms underpinning this disturbance remain
poorly understood. To this end, we investigated the structural organization of the white
matter connectome in PD freezers and non-freezers. We hypothesized that freezers would
show an altered network architecture, which could hinder the effective information
processing that characterizes the disorder. Twenty-six freezers and twenty-four well-
matched non-freezers were included in this study. Using diffusion tensor imaging, we
investigated the modularity and integration of the regional connectome by calculating the
module degree z-score and the participation coefficient, respectively. Compared to non-
freezers, freezers demonstrated lower participation coefficients in the right caudate,
thalamus and hippocampus, as well as within superior frontal and parietal cortical
regions. Importantly, several of these nodes were found within the brain’s ‘rich club’.
Furthermore, group differences in module degree z-scores within cortical frontal regions
and in sensory processing areas were found. Together, our results suggest that changes in
the structural network topology contribute to the manifestation of FOG in PD,
specifically due to a lack of structural integration between key information processing
hubs of the brain.
Key Words: Parkinson’s disease; Freezing of Gait; Connectome; Diffusion imaging

112
Introduction
Freezing of gait (FOG) is a sudden and involuntary gait cessation that is common in
idiopathic Parkinson’s disease (PD) (Nutt et al., 2011) and has a prominent impact on
quality of life (Walton et al., 2015). Freezing behaviour can be provoked by a range of
triggers, including turning, dual tasking and emotional stress (Plotnik et al., 2012).
Treatment options are limited, and although dopaminergic medication can partially
ameliorate freezing, patients are often forced to rely on behavioural strategies that
temporarily enhance their gait pattern using goal directed strategies (Nutt et al., 2011).
Furthermore, patients with freezing often experience executive dysfunction, mood
disorders and perceptual problems (Ehgoetz Martens et al., 2014a, 2014b; Naismith et al.,
2010). These observations have led to the suggestion that FOG is not simply the result of
dysfunctional locomotor output, but rather results from a diffuse, network-level
dysfunction that ultimately manifests as a loss of top-down control over the brainstem
structures that control gait (Lewis & Shine, 2016).
A range of studies has been conducted to determine whether individuals with FOG
demonstrate white matter abnormalities. For instance, widespread alterations in structural
connections between cortical, subcortical and brainstem regions have been reported in
studies using diffusion tensor imaging (DTI), a technique that infers the integrity of inter-
regional white matter. Compared to non-freezers, freezers have shown abnormalities in
the corticopontine, pontine-cerebellar (Fling et al., 2013; Schweder et al., 2010;
Vercruysse et al., 2015; Youn et al., 2015), major cortico-cortical and frontostriatal tracts
(Pietracupa et al., 2018; Vercruysse et al., 2015). Conversely, differences were not
observed in a study investigating the frontostriatal hyper-direct pathway (Fling et al.,
2014) or in a whole brain analysis between freezers and non-freezers (Canu et al., 2015).
These divergent results suggest that structural lesions in freezers might be subtle and
dispersed, and hence raise the question of whether FOG actually stems from a focal
anatomical lesion at all (Lewis & Shine, 2016). Indeed, a key caveat of the majority of
studies to date is that they have used diffusion imaging in an attempt to identify lesions
within specific regions or tracts. As such, they offer insights that are relatively limited to

113
the field of interest and therefore miss more complex, distributed patterns that might exist
in the data.
Given the multi-dimensional nature of FOG, the primary objective of this study was to
investigate the structural brain network architecture in patients with FOG and this
debilitating phenomenon. Specifically, we investigated whether white matter changes that
contribute to freezing might emerge as alterations in structural network topology. The
brain displays a modular structure, in which discrete communities (‘modules’) with
densely interconnected nodes are only sparsely connected with nodes of other
communities (Shine et al., 2017; Sporns et al., 2016). These specialist modules allow for
processing without unnecessary cross-talk (Bassett et al., 2015). Conversely, the
integration of communities is crucial for incorporating complex information streams of
different modalities, such as in cognitively challenging tasks (Figure 1b) (Bertolero et al.,
2015; Shine et al., 2017).
Graph theory offers a robust mathematical framework for interrogating neural network
topology, and can offer a detailed in vivo analysis of the brain at the level of circuits and
modules (Sporns et al., 2004). Importantly, whole brain graph theoretical embraces the
inherent complexity of interacting subsystems, rather than isolating a particular region
within a circuit. Here, we used a combination of DTI and graph theoretical analyses to
investigate the global and regional topology of the structural connectome in individuals
with PD and FOG. Given the known impairments in multi-domain behaviour in patients
with freezing, we predicted that freezers should exhibit impairments in inter-modular
connectivity, especially in frontoparietal and subcortical networks (Lewis & Shine,
2016).
Materials and methods
Fifty patients with mild to moderate idiopathic PD, recruited from the Parkinson’s
Disease Research Clinic at the Brain and Mind Centre, University of Sydney were
included in this study. Demographic and clinical details including age, gender, disease
duration, levodopa equivalent daily dose and motor severity were obtained and are listed

114
in Table 1. Motor severity was measured by the Movement Disorder Society Unified
Parkinson’s Disease Rating Scale (MDS-UPDRS) part III (Goetz et al., 2008) and Hoehn
and Yahr (H&Y) scale (Hoehn & Yahr, 1967). Patients with an H&Y score of > 3 were
excluded from the study. Cognitive screening was performed using the Mini Mental State
Examination (Folstein et al., 1975) and patients with a score of ≤ 25 on this test were also
excluded from the study. Finally, Logical Memory (Wechsler, 1997), the Trail Making
Test Part B – A (TMTB-A) (Army Battery, 1944) and Digit Span Backward (DSB)
(Wechsler, 1997) were used as measures of memory and executive functioning ability.
Patients were identified as ‘freezers’ if they scored positively on the Freezing of Gait
Questionnaire item 3 (Giladi et al., 2000; Shine et al., 2012) and as ‘non-freezers’ if they
scored a 0 on this item.
MRI Acquisition
Neuroimaging was acquired on a 3-Tesla General Electric Discovery MR750 scanner
(GE Medical Systems) using an eight-channel phased array head coil. 3D T1-weighted,
anatomical images (voxel size 1 × 1 × 1 mm) were obtained for parcellation of cortical
regions, using the following protocol: Repetition time 7.2 ms, echo time 2.7 ms, inversion
time 450 ms, 196 transverse slices, 256 × 256 matrix; field of view 256 x 256 mm, flip
angle 12°. Diffusion weighted images (DWI) were acquired in 61-gradient directions,
using the following echoplanar imaging protocol: repetition time 7025 ms, echo time 80
ms, 55 transverse slices, 2.5 mm slice thickness, 256 × 256 matrix; field of view 240 x
240 mm. The effective diffusion weighting was b = 1000 s/mm2. Four volumes with no
diffusion weighting (b = 0 s/mm2) were acquired at the beginning of each diffusion
sequence and used for robust individual registration between T1-weighted anatomical and
the diffusion weighted images.
Image pre-processing
Diffusion image pre-processing was performed using the FMRIB Software Library (FSL,
http://fsl.fmrib.ox.ac.uk). Raw DWI were corrected for susceptibility, head motion and
eddy current distortions using FSL’s tool eddy and skull stripped using BET. T1-weighted

115
images were segmented into white and grey matter (cortical and subcortical structures)
and cerebrospinal fluid using FreeSurfer (version 5.3; http://surfer.nmr.mgh.harvard.edu).
The cortical surface was further parcellated into 219 cortical regions using the Lausanne
anatomical atlas (Cammoun et al., 2012) distributed as part of the Connectome Mapping
Toolkit (Daducci et al., 2012) and validated for diffusion MRI analysis. Using
FreeSurfer, fourteen subcortical structures were segmented including the left and right
thalamus, caudate, putamen, pallidum, amygdala, hippocampus and nucleus accumbens
area by an atlas based segmentation (Fischl et al., 2002). This atlas was chosen because
this scale has been shown to provide local interpretability whilst containing homogeneous
clusters (Cammoun et al., 2012). T1-weighted images were registered in DWI space to
the b0 volume. Fibre reconstruction was performed using robust tensor fitting. The main
diffusion direction of each voxel was estimated after decomposition of the tensor into
eigenvectors and eigenvalues, and fractional anisotropy values, a measure of magnitude
of directionally of diffusion in white matter fibres, were computed. White matter
pathways were constructed using deterministic fibre tracking approach. The deterministic
approach was chosen to control for the number of false positives, which are unfavourable
when investigating network topology (Zalesky et al., 2016). Using the fibre assignment
by continuous tracking algorithm, streamlines were traced following the main diffusion
direction starting from eight seeds evenly distributed in each white matter voxel and
stopping when one of the following criteria was met: 1) fractional anisotropy < 0.1; 2)
turn > 45 degrees; 3) the end of the brain mask was reached. Connectome maps were
reconstructed using the Connectome Mapper (Daducci et al., 2012). By combining the
parcellation map and the reconstructed fibres a connectivity matrix was created for each
subject, using the average fractional anisotropy of voxels of the reconstructed
streamlines. The pipeline is illustrated in Figure 1a. Finally, to account for spurious
streamlines, values of tracts that were present in less than 50% of all participants were set
to zero by applying a threshold on the individual connectivity matrices (de Reus et al.,
2013). To verify the results were not skewed by the choice of threshold, the analysis was
also performed using thresholds of 40% and 60%.

116
Figure 1a) Imaging processing flow. b) Representation of a brain network with a
modular structure. Different graph metrics are highlighted.
Graph Theoretical Analyses
Graph metrics were calculated on the thresholded adjacency matrices using MATLAB
scripts from the Brain Connectivity Toolbox (Rubinov et al., 2010). First, we confirmed
that the group mean structural connectome demonstrated a ‘small-world’ architecture, by
comparing the ratio of the mean clustering coefficient (the average intensity of triangles

117
around a node) to the shortest path length (a metric of information transmission) of the
thresholded connectivity matrix. We next sought to identify significant differences in
graph metrics between the two groups. To this end, network modularity was calculated
using the Louvain modularity algorithm (Rubinov et al., 2010). This algorithm iteratively
assigns nodes to different communities, until the maximum possible modularity statistic
Q is reached. In this way, the network is subdivided into non-overlapping groups of
highly connected nodes that minimizes between module edges.
! = !
! !!" − !!" !!!!!!" [1] Equation 1 – Louvain modularity algorithm, where v is the total weight of the network (sum of all positive connections), wij is the weighted connection between regions i and j, eij is the strength of a connection divided by the total weight of the network, and δMiMj is set to 1 when regions are in the same community and 0 otherwise.
Due to the stochastic nature of the Louvain algorithm, the community assignment for
each region was assessed 100 times on the group average connectome and a consensus
partition was identified using a fine-tuning algorithm. To define an appropriate value for
the resolution parameter (γ), we iterated the Louvain algorithm across a range of values
(0.5 – 2.5 in steps of 0.1) for 100 iterations and then estimated the similarity of the
resultant partitions using mutual information. A γ parameter of 1.0 provided the most
robust estimates of topology across these iterations, and thus was used for the main
experiment.
To estimate the topology at the regional level, we calculated the module degree z-score
(Wi; a measure of intra-module connectivity; see equation 2) and participation coefficient
(Bi; a measure of inter-module connectivity; see equation 3) for each node. Positive Wi-
values reflect nodes with more connections to other members of the same community
(compared to the average number of connections), and vice versa. In contrast, the Bi
values ranges between 0 and 1, with a value of 0 indicating that all connections are within
its community, whereas a value of 1 indicates uniformly distributed connections across
modules (github.com/juliemaehall/topology).

118
!! =!!!!!!!!!!
[2]
Equation 2 – Module degree z-score Wi, where κi is the strength of the connections of region i to other regions in its module si, !!! is the average of κ over all the regions in si, and !!!! is the standard deviation of κ in si
!! = 1− !!"!!
!!!!!! [3]
Equation 3 - Participation coefficient Bi, where κis is the strength of the positive connections of region i to regions in module s, and κi is the sum of strengths of all positive connections of region i. The participation coefficient of a region is therefore close to 1 if its connections are uniformly distributed among all the modules and 0 if all of its links are within its own module.
Rich Club Analysis
We next identified the structural ‘rich club’ of the binarized group mean connectivity
matrix of the entire sample. The ‘rich club’ of a network represents a set of high degree
nodes (i.e. nodes with a large number of edges) that are more densely interconnected with
other high degree nodes than would be expected by chance (van den Heuvel et al., 2011).
To estimate these regions, the degree of each node i was determined by taking the sum of
the number of connections node i shared with k other nodes in the network. Then a sub-
network (the ‘k core’) was obtained by removing all nodes that had connections ≤ k. For
each value of k, the rich-club coefficient Φk was then computed as the ratio of the number
of connections between the nodes in the k core network and the sum of possible
connections that would be present if the network was completely connected (see equation
4).
Φ! = !!!!
!!!(!!!!!) [4]

119
Equation 4 – Rich club coefficient Φ! at a specific k level is the fraction of edges E that connect nodes of a specific level of k or higher out of the maximum number of edges that nodes may share.
Since random networks also show an increasing function of Φk, we next normalized the
rich club coefficient in reference to a null model. To this end, we created 5000 random
networks with a preserved degree distribution, and then the average rich-club coefficient
Φrand over the set of random networks was calculated for each level of k. We then divided
Φk by Φrand, and to define the normalized rich club coefficient (ΦZ). Values of k with a ΦZ
> 1 can thus be said to contain a stronger rich club architecture than expected under the
conditions of the null model (van den Heuvel et al., 2011). The identity of rich club nodes
for the highest value of k with ΦZ > 1 can be found by identifying nodes that were present
within the k core network for that value of k. The 85th percentile of nodes was selected as
part of the rich club.
Statistical analyses
Data analyses for demographic and clinical variables were performed using IBM SPSS
Version 19. Independent t-tests and Mann-Whitney U-tests were performed, depending
on the distribution of the data. Missing values for LEDD (n = 1; 2% of the sample),
Logical Memory (n=1; 2% of the sample), TMTB-A (n = 2; 4% of the sample) and DBS (n
= 1, 2% of the sample) were replaced using the Expectation Minimization algorithm. The
results are presented in Table 1. To assess statistical significance for the global and
regional level metrics, a non-parametric permutation testing approach was used (5000
iterations) that effectively controls for multiple comparisons (Nichols et al., 2002).
Additionally, permutation testing with 5000 iterations was applied following a
Spearman’s correlation analysis between the significant Bi and Wi-values and TMTB-A
and DSB scores.
To determine whether there was overlap between the rich club and regions with altered
topology in our two clinical groups, we compared the spatial overlap between the three
variables. To analyse whether this overlap was statistically significant, a permutation test

120
was performed in which we randomly permuted the rich club identity of each region
(5000 iterations) and used the overlap between the randomized vector and the
significantly different Wi and Bi scores to populate two separate null distributions. We
then compared the empirical overlap between rich club and topological outcome
measures with the null distributions and rejected the null hypothesis of no overlap if the
true overlap was less than the 97.5th percentile of null distribution (i.e. the top 2.5%).
The difference scores between the groups on the Bi and Wi values were compared across
the three different thresholds, and moderate to very strong correlations were found,
confirming the robustness of the findings (see Supplementary Materials).
Results
Demographics
Twenty-six patients were identified as freezers and twenty-four as non-freezers. There
were no differences between the groups for age, disease duration, dopaminergic
medication, disease severity as measured by the MDS-UPDRS part III and H&Y scale
and the MMSE, Logical Memory, TMTB-A and DSB (see Table 1).
Graph metrics
A small-world organisation of the structural brain networks was confirmed in both patient
groups (van den Heuvel et al., 2011). Table 2 and Figure 2a demonstrate the 23 nodes
that had significantly lower Bi-values in freezers compared to non-freezers (nodes
coloured in green in Figure 2a). Three of these more segregated nodes encompassed
subcortical nuclei, including the right caudate, thalamus and hippocampus. In the left
hemisphere, we found six nodes in the frontal cortex, six nodes in the parietal cortex and
one node in the posterior cingulate gyrus. In the right hemisphere, six nodes in the frontal
cortex and one node in the anterior cingulate gyrus showed significantly lower values in
freezers compared to non-freezers. Conversely, the caudal middle frontal gyrus showed a
significantly higher Bi-value in freezers. Nine nodes in distributed cortical nodes with a
lower Wi-score were found (nodes coloured in green in Figure 2b) and six cortical nodes

121
with a higher z-score (nodes coloured in orange in Figure 2b) in PD freezers were found
(see Table 3).
Table 1. Demographic and clinical data of patients with and without freezing of gait Freezers Non-freezers p-value
n 26 24 -
Age, years, mean ± SD 64.2 ± 9.4 66.9 ± 5.3 0.220
Disease duration, years, mean ± SD 5.2 ± 3.8 5.2 ± 3.0 0.970
LEDD, mg, mean (range)* 656.7 (0 - 1548) 526.6 (0 - 1200) 0.431
MDS-UPDRS III, mean ± SD 25.5 ± 13.0 23.3 ± 13.5 0.558
H&Y, mean (range)* 2.0 (1 - 2.5) 1.8 (1 - 2.5) 0.131
MMSE, mean (range) * 28.5 (26 - 30) 29.1 (26 - 30) 0.051
Logical Memory I, mean ± SD 36.1 ± 10.0 39.8 ± 10.3 0.201
Logical Memory II, mean ± SD 21.3 ± 7.8 22.5 ± 10.4 0.657
TMTB-A, sec, mean (range) * 61.6 (17 - 216) 41.0 (-1 - 92) 0.063
Digit Span Backward, mean ± SD 6.38 ± 1.8 7.57 ± 3.3 0.118
* Mann-Whitney U test; LEDD = levodopa equivalence daily dose; MDS-UPDRS III = motor part of the Movement Disorder Society Unified Parkinson’s Disease Rating Scale; H&Y = Hoehn and Yahr motor scale; MMSE = Mini-Mental State Examination; TMTB-A = Trail Making Test part B - A
Of the regions that showed significantly different Bi-values, two nodes in the superior
parietal cortex negatively correlated with the TMTB-A scores ('lh-superior-parietal_2': r =
-0.32, p < 0.05; 'lh-superior-parietal_4': r = -0.34, p < 0.05), whilst the right hippocampus
and node ‘rh-superiorfrontal_4’ showed a positive correlation with the DSB (r = 0.35, p <
0.01 and r = 0.27, p < 0.05 respectively). One node with a significantly different Wi-value
showed a positive correlation (left 'superior-frontal': r = 0.30, p < 0.05) with the TMTB-A.
Finally, node ‘rh-superiorfrontal_8’ showed a negative correlation with the DSB (r =
0.17, p < 0.05), whilst node ‘rh-rostralmiddlefrontal_4’ showed a negative correlation
with this test performance (r = -0.27 p < 0.05).
A rich-club network encompassing 35 nodes was identified. Of the 24 nodes that had a
significantly different Bi-value between groups, six nodes were identified as part of the

122
rich-club, which was deemed significant (p < 0.05; see Table 2, Figure 3 and
Supplementary materials). Of the 15 nodes with different Wi-values between groups,two
nodes were considered within the rich-club network but this overlap was not significant
(p > 0.05).
Figure 2a: Nodes with a significantly different Bi score in freezers compared to non-
freezers, b: nodes with a significantly different Wi score in freezers compared to non-
freezers.
a
b

123
Figure 3. Representation of the rich-club network.
Discussion
Here we used graph theoretical methods to assess the structural network topology of PD
patients with FOG. This approach allowed us to investigate both integrative as well as
specialized properties of individual nodes within the brain’s structural connectivity
network in a data-driven manner, thus obviating any a priori assumptions about selected
regions. In doing so, we demonstrated that freezers had a significantly lower participation
coefficient in 23 of the 233 nodes, compared to non-freezers. Specifically, these more
segregated nodes included the right subcortical caudate, thalamus and hippocampus, as
well as nodes within bilateral frontal and parietal regions. Moreover, the six of these
nodes were also part of the rich-club network that forms the backbone of the structural
connectome by reducing path length (as opposed to multiple short-path connections) and
hence permits faster and less noisy signal transmission (Bertolero et al., 2015; Bullmore
et al., 2012; van den Heuvel et al., 2011).
Therefore, in keeping with other biological models (Crossley et al., 2014; Robbins et al.,
2009), we hypothesize that the symptoms of FOG may in part arise due to a lack of
structural integration between key information processing hubs of the brain including
motor, cognitive and limbic structures.

124
Table 2. Nodes with significantly different participation coefficients between groups
lh = left hemisphere; rh = right hemisphere
Node Region x y z Freezers Non-freezers Rich club
Non-freezers > freezers
Subcortical
R caudate Basal ganglia 15 10 11 0.690 ± 0.05
0.731 ± 0.05
✓
R hippocampus Hippocampus 28 -22 -13 0.613 ± 0.09 0.668 ± 0.08 ✓ R thalamus Thalamus 13 -17 7 0.699 ± 0.07 0.752 ± 0.05 ✓ Frontal
lh-lateralorbitofrontal_4 Orbitofrontal cortex -12 52 -19 0.562 ± 0.12 0.628 ± 0.10 -
rh-lateralorbitofrontal_4 Orbitofrontal cortex 16 52 -18 0.525 ± 0.13 0.612 ± 0.10 ✓ lh-medialorbitofrontal_2 Orbitofrontal cortex -5 33 -20 0.596 ± 0.11 0.676 ± 0.07 -
lh-superiorfrontal_5 Prefrontal cortex -11 36 47 0.594 ± 0.10 0.653 ± 0.09 -
lh-superiorfrontal_7 Premotor cortex -8 14 52 0.579 ± 0.16 0.660 ± 0.08 -
lh-superiorfrontal_8 Premotor cortex -8 -2 63 0.524 ± 0.13 0.628 ± 0.10 -
lh-superiorfrontal_9 Premotor cortex -17 -1 67 0.524 ± 0.10 0.608 ± 0.09 -
rh-superiorfrontal_3 Prefrontal cortex 11 40 40 0.580 ± 0.16 0.670 ± 0.11 -
rh-superiorfrontal_4 Prefrontal cortex 18 34 47 0.571 ± 0.17 0.674 ± 0.09 -
rh-superiorfrontal_5 Premotor cortex 12 18 52 0.583 ± 0.14 0.679 ± 0.09 -
rh-superiorfrontal_6 Premotor cortex 11 2 60 0.521 ± 0.15 0.611 ± 0.09 -
rh-paracentral_2 Primary motor cortex 10 -16 59 0.199 ± 0.18 0.325 ± 0.18 -
Parietal
lh-inferiorparietal_4 Somatosensory association cortex
-36 -71 42 0.304 ± 0.16 0.423 ± 0.18 -
lh-postcentral_2 Primary somatosensory cortex
-26 -31 65 0.596 ± 0.11 0.676 ± 0.10 -
lh-postcentral_3 Primary somatosensory cortex
-40 -30 57 0.479 ± 0.15 0.565 ± 0.16 -
lh-superiorparietal_2 Somatosensory association cortex
-33 -43 55 0.239 ± 0.16 0.373 ± 0.17 -
lh-superiorparietal_4 Somatosensory association cortex
-20 -62 61 0.574 ± 0.14 0.681 ± 0.08 ✓
lh-superiorparietal_7 Visual association cortex -13 -87 34 0.548 ± 0.15 0.640 ± 0.10 ✓ Cingulate
lh-posteriorcingulate_1 Posterior cingulate gyrus -4 -11 35 0.409 ± 0.16 0.512 ± 0.18 -
rh-caudalanteriorcingulate_1 Anterior cingulate gyrus 9 36 5 0.497 ± 0.17 0.602 ± 0.18 -
Non-freezers < freezers
rh-caudalmiddlefrontal_1 Prefrontal cortex 36 2 47 0.333 ± 0.17 0.235 ± 0.16 -

125
Table 3. Nodes with significantly different module degree z-scores between groups
lh = left hemisphere; rh = right hemisphere
A decreased ability to recruit alternative routes for parallel information processing
(Achard et al., 2006) would lead to a lack of resilience when there are functional network
challenges (such as cognitive dual-task performance) and also offer an explanation for the
known characteristic set-shifting deficits in individuals with FOG (Naismith et al.,
2010b). The motor manifestation of FOG could thus arise due to a flow-on effect,
Node Region x y z Freezers Non-freezers
Non-freezers > freezers
Frontal
lh-medialorbitofrontal_1 Orbitofrontal cortex -7 52 -11
-0.482 ± 0.70 0.189 ± 0.95
lh-parstriangularis_1 Prefrontal cortex -43
33 2 0.371 ± 0.74 0.847 ± 0.88
rh-parsorbitalis_1 Prefrontal cortex 43 43 -10
0.324 ± 0.80 0.904 ± 0.66
rh-superiorfrontal_8 Prefrontal cortex 22 10 58 -0.574 ± 0.49 -0.181 ± 0.51
Parietal
lh-postcentral_5 Primary sensory cortex -55
-15
43 -0.686 ± 0.48 -0.367 ± 0.52
rh-postcentral_5 Primary sensory cortex 28 -32
69 0.249 ± 0.72 0.826 ± 0.91
Occipital
lh-lateraloccipital_5 Visual association cortex -34
-83
-10
-0.222 ± 0.80 0.534 ± 0.78
Cingulate
lh-rostralanteriorcingulate_1
Anterior cingulate -7 37 2 0.566 ± 1.00 1.234 ± 0.83
rh-rostralanteriorcingulate_1
Anterior cingulate 14 -33
52 0.713 ± 0.93 1.397 ± 0.50
Non-freezers < freezers
Frontal
lh-superiorfrontal_4 Prefrontal cortex -8 32 37 -0.556 ± 0.87 -1.082 ± 0.57
rh-rostralmiddlefrontal_4 Prefrontal cortex 34 52 10 -0.837 ± 0.54 -1.187 ± 0.55
Parietal
lh-postcentral_7 Retrosubicular area -52
-11
19 -0.354 ± 0.75 -0.724 ± 0.52
rh-superiorparietal_1 Somatosensory association cortex 33 -43
54 0.550 ± 0.85 0.095 ± 0.79
rh-precuneus_1 Visual association cortex 23 -65
21 0.282 ± 0.76 -0.183 ± 0.71
Temporal
lh-inferiortemporal_1 Inferior temporal gyrus -41
-3 -38
-0.100 ± 0.61 -0.518 ± 0.68

126
whereby inefficient network architecture would hamper the effective communication
between major cortical and subcortical integration centres (Lewis & Shine, 2016).
Nodes with high participating coefficients have previously been characterized as
‘connector hubs’ (Guimera et al., 2005), as they tend to interconnect otherwise segregated
communities. In the brain, nodes with high participation are thought to be crucial for
incorporating complex information streams of different modalities. Affected participation
scores, especially across these connector hubs, may cause disruptions extending across
cognitive, motor and limbic domains (Bertolero et al., 2015). Nodes with high
participation coefficients exhibit more activity when coordinated activity occurs across
communities, such as during cognitive task performance (Shine et al., 2017). These hubs
provide an integrative network function to an even greater extent than rich club nodes
(Bertolero et al., 2017) and importantly, ten regions affected in freezers were amongst the
nodes with the highest participation scores. Hence they likely play a crucial role in
disrupted information processing in these patients.
The caudate nucleus and thalamus have previously been implicated in the freezing
pathophysiology with studies showing reduced grey matter volume (Herman et al., 2014)
and hypo-activity within subcortical structures (Shine et al., 2013b). Importantly, the
thalamic nuclei receive input from and project to diffuse areas in the cortex, basal ganglia
and cerebellum play a role in modulating activity in the rest of the brain. Rather than
simply being a relay station, the thalamic nuclei are a major hub for controlling the
information flow across cortical and subcortical regions (Sherman, 2016). The caudate
nucleus also receives inputs from various prefrontal regions (Jarbo et al., 2015) and plays
a major part in task-switching behaviour, which can be understood in terms of activating
different modules for specialised function. Despite not reaching significance, the left
thalamus and caudate also showed a lower participation coefficient in freezers (p = 0.07
and p = 0.08, respectively), hence there was no strong lateralisation effect observed.
Thus, the lower participation scores of these nodes may contribute to the pathophysiology
of FOG through either ineffective integration of motor, sensory and limbic activity during

127
gait or inefficient shifting between functionally distinct but complementary neural
pathways (Lewis & Shine, 2016).
Previous findings of neuromodulatory systems have demonstrated the existence of an
inverted U-shaped curve on executive functioning (Robbins et al., 2009), and recent
conceptual work has suggested that this relationship may also be present in network
topology (Shine et al., 2017). Consistent with the well-established association between
FOG and impaired executive function (Hall et al., 2015; Vandenbossche et al., 2012;
Walton et al., 2015b), our findings showed less integrated nodes in the caudate, bilateral
dorsolateral prefrontal (DLPFC) and anterior cingulate cortex. Moreover, participation
coefficients of nodes within the DLPFC, superior parietal cortex and hippocampus were
associated with a performance on tasks investigating set-shifting and working memory.
Thus, is it possible that changes in integration scores in areas associated with cognitive
performance affect effective information transfer across the network, and might explain
these phenotypic changes.
Lower participation coefficients were also found across frontoparietal structures,
including the premotor, primary motor cortex and somatosensory association cortices,
whilst a node with a higher participation coefficient was also found in the premotor
cortex, perhaps reflecting a rerouting of pathways in this region. Automated movements
are performed by activity in segregated modules (Bassett et al., 2015; Shine et al., 2017),
and these movements are affected by PD pathology (Redgrave et al., 2010). As such, PD
patients rely more on effective communication in superior frontoparietal regions since
combining different sources of information between these regions is fundamental for
successful completion of an accurate and context-appropriate movement (Wu et al.,
2005). When a region becomes too specialized or segregated, it loses its flexibility that is
necessary to navigate through changing circumstances, which may underpin the
difficulties freezers experience navigating complex environments.
A number of fMRI studies using region of interest analyses comparing PD freezers to
non-freezers, have shown alterations in similar regions as reported in the current study.

128
Specifically, a functional decoupling between the bilateral cognitive control network and
the basal ganglia during freezing episodes has been reported during motor arrests
provoked by a virtual reality paradigm (Shine, 2013a). Furthermore, using this paradigm,
freezing episodes were associated with alterations in BOLD response within
frontoparietal sensorimotor regions, basal ganglia, thalamus and in the mesencephalic
locomotor region (Shine et al., 2013b). Altered resting state functional connectivity
within the cognitive control network, visual networks and locomotor regions has further
been shown to be associated with FOG severity (Fling et al., 2014; Tessitore et al., 2012).
However, future research avenues should explore alterations in functional network
topology associated with freezing of gait.
The current study also compared within-module connectivity across groups. Segregation
into isolated modules allows for specialized processing of an action and increases
efficiency, flexibility and robustness to perturbation (Bullmore et al., 2012). Abnormal
within-module scores can affect these characteristics by increasing effective metabolic
costs (Bullmore et al., 2012). Compared to non-freezers, freezers showed lower module
degree z-scores in nodes within the prefrontal cortex (pars orbitalis, pars triangularis,
orbitofrontal and rostral anterior cingulate), primary sensory, primary motor and visual
association cortex. Perhaps surprisingly, higher module degree z-scores were observed in
patients with FOG in prefrontal cortices (DLPFC and dorsal anterior cingulate), and
various sensory information-processing areas such as the somatosensory association,
visual association and inferior temporal regions. Interestingly, higher module degree z-
scores are considered favourable for behaviours that require specific, unimodal function,
such as certain sensory processing systems (Park et al., 2013). As such, freezers may
need to rely more heavily on exogenous stimuli to gather information to guide their
actions, perhaps due to a loss of automaticity (Vandenbossche et al., 2012). Indeed,
previous studies have shown a proprioceptive deficit in PD patients with (Ehgoetz
Martens et al., 2013) and without freezing (Wu et al., 2005) along with a greater
dependence on visual information (Davidsdottir et al., 2005). Therefore, it could be
speculated that a reweighting of sensory systems occurs in PD patients with freezing.
Specifically, there may be an increased emphasis placed on discrete modality specific

129
pathways that process afferent information, however if the system is unable to effectively
integrate this information, the dynamic evolution of the gait pattern may suffer.
Limitations and future directions
The current study has several limitations. Firstly, we grouped the participants based on a
subjective question of the FOGQ. Currently, there is no gold standard to define FOG and
whilst objective measures are preferable, FOG can be difficult to elicit in a clinical or
experimental environment and might remain unnoticed (Nieuwboer et al., 1998). Another
possible weakness of this study’s methodology is that the Louvain algorithm does not
allow for overlapping modules, as a node is only assigned to one community.
Furthermore, although DTI is an established method for measuring white matter integrity
and has led to great insights in PD (de Oliveira et al., 2017; Hall et al., 2016), it suffers
from inherent limitations. Specifically, diffusion imaging measures the motion of water
molecules along a tract and thus only allows for indirect validation of streamlines. The
deterministic diffusion tensor fitting approach used in this study requires sufficient
directional information at each voxel and is incapable of resolving crossing or ‘kissing’
fibres. However, the deterministic diffusion tensor fitting approach used in this study
requires sufficient directional information at each voxel and is incapable of resolving
crossing or ‘kissing’ fibres. At an ambiguous directional point in a voxel, fractional
anisotropy values might drop leading to a premature termination of the reconstruction of
a tract. This could be especially problematic in structures with complex fibre architecture
such as subcortical nuclei. Future research should use more sophisticated methods, such
as constrained spherical deconvolution, that are better equipped to deal with complex
fibre configurations. Finally, this study used single shell DTI data, without EPI distortion
corrections, which could have impacted the registration accuracy between the DTI and T1
images, creating suboptimal connectomes. Hence, these results should be interpreted with
caution and replication studies using multi-shell sequences are necessary to fully
understand the changes in network topology associated with FOG. Despite its discernible
limitations, diffusion imaging is the only non-invasive in-vivo structural imaging
technique and has successfully addressed important scientific matters.

130
Conclusion
This is the first study to use graph theory to demonstrate subtle alterations in network
topology in PD patients with FOG compared to those without this symptom. Specifically,
we showed that freezers have lower participation scores in subcortical regions as well as
higher order and integrative cortical cortices. We argue that this increase in segregation
hinders efficient communication between major cortical and subcortical centres, resulting
in ineffective integration of motor, sensory and limbic activity, as well as inefficient
shifting between specialised modules that could disrupt gait in PD. Decreased intra-
module connectivity might reflect a less cost effective processing of information, whereas
an increase in these scores in the sensory cortex of freezers, suggests a possible
compensatory strategy.
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical standards
All persons gave their informed consent prior to their inclusion in registries. The
registries were approved by the Human Research Ethics Committee of the University of
Sydney. Patients were included after informed written consent had been obtained, as set
forth by the Declaration of Helsinki (WMA, 1964–2014).
Acknowledgements
We thank the patients and their families who contribute to our research at the Parkinson’s
Disease Research Clinic. This research was supported by Sydney Informatics Hub,
funded by the University of Sydney. JMH is supported by a Western Sydney University
Postgraduate Research Award; JMS is supported by a National Health and Medical
Research Council CJ Martin Fellowship (1072403); KAEM is supported by a Parkinson
Canada Fellowship; MG is supported by a University of Sydney International
Scholarship; Professor Lewis is supported by National Health and Medical Research
Council-Australian Research Council Dementia Fellowship (#1110414) and this work

131
was supported by funding to Forefront, a collaborative research group dedicated to the
study of non-Alzheimer disease degenerative dementias, from the National Health and
Medical Research Council of Australia program grant (#1037746 and #1095127). KMB,
JYYS, CCW and AAM have no funding source to disclose.

132
References
Achard, S., Salvador, R., Whitcher, B., Suckling, J., & Bullmore, E. (2006). A resilient,
low-frequency, small-world human brain functional network with highly
connected association cortical hubs. J Neurosci, 26(1), 63-72.
doi:10.1523/JNEUROSCI.3874-05.2006
Army Battery (1944). Manual of Directions and Scoring. Washington, DC: War
Department, Adjutant General’s Office.
Bassett, D. S., Yang, M., Wymbs, N. F., & Grafton, S. T. (2015). Learning-induced
autonomy of sensorimotor systems. Nat Neurosci, 18(5), 744-751.
doi:10.1038/nn.3993
Bertolero, M. A., Yeo, B. T. T., & D'Esposito, M. (2017). The diverse club. Nat
Commun, 8(1), 1277. doi:10.1038/s41467-017-01189-w
Bertolero, M. A., Yeo, B. T. T., & D’Esposito, M. (2015). The modular and integrative
functional architecture of the human brain. Proc Natl Acad Sci USA, 112(49),
E6798-E6807. doi:10.1073/pnas.1510619112
Bullmore, E., & Sporns, O. (2012). The economy of brain network organization. Nat Rev
Neurosci, 13(5), 336-349. doi:10.1038/nrn3214
Cammoun, L., Gigandet, X., Meskaldji, D., Thiran, J. P., Sporns, O., Do, K. Q., . . .
Hagmann, P. (2012). Mapping the human connectome at multiple scales with
diffusion spectrum MRI. J Neurosci Methods, 203(2), 386-397.
doi:10.1016/j.jneumeth.2011.09.031
Canu, E., Agosta, F., Sarasso, E., Volonte, M. A., Basaia, S., Stojkovic, T., . . . Filippi,
M. (2015). Brain structural and functional connectivity in Parkinson's disease with
freezing of gait. Hum Brain Mapp, 36(12), 5064-5078. doi:10.1002/hbm.22994
Crossley, N. A., Mechelli, A., Scott, J., Carletti, F., Fox, P. T., McGuire, P., & Bullmore,
E. T. (2014). The hubs of the human connectome are generally implicated in the
anatomy of brain disorders. Brain, 137(Pt 8), 2382-2395.
doi:10.1093/brain/awu132
Daducci, A., Gerhard, S., Griffa, A., Lemkaddem, A., Cammoun, L., Gigandet, X., . . .
Thiran, J.-P. (2012). The Connectome Mapper: An Open-Source Processing

133
Pipeline to Map Connectomes with MRI. PLoS One, 7(12), e48121.
doi:10.1371/journal.pone.0048121
Davidsdottir, S., Cronin-Golomb, A., & Lee, A. (2005). Visual and spatial symptoms in
Parkinson’s disease. Vis Res, 45(10), 1285-1296.
doi:http://dx.doi.org/10.1016/j.visres.2004.11.006
de Oliveira, R. V., & Pereira, J. S. (2017). The role of diffusion magnetic resonance
imaging in Parkinson's disease and in the differential diagnosis with atypical
parkinsonism. Radiol Brasileira, 50(4), 250-257. doi:10.1590/0100-3984.2016-
0073
de Reus, M. A., & van den Heuvel, M. P. (2013). Estimating false positives and negatives
in brain networks. Neuroimage, 70, 402-409.
doi:http://dx.doi.org/10.1016/j.neuroimage.2012.12.066
Ehgoetz Martens, K. A., Ellard, C. G., & Almeida, Q. J. (2014a). A closer look at
mechanisms underlying perceptual differences in Parkinson's freezers and non-
freezers. Neuroscience, 274, 162-169. doi:10.1016/j.neuroscience.2014.05.022
Ehgoetz Martens, K. A., Ellard, C. G., & Almeida, Q. J. (2014b). Does anxiety cause
freezing of gait in Parkinson's disease? PLoS One, 9(9), e106561.
doi:10.1371/journal.pone.0106561
Ehgoetz Martens, K. A., Pieruccini-Faria, F., & Almeida, Q. J. (2013). Could sensory
mechanisms be a core factor that underlies freezing of gait in Parkinson's disease?
PLoS One, 8(5), e62602. doi:10.1371/journal.pone.0062602
Fischl, B., Salat, D. H., Busa, E., Albert, M., Dieterich, M., Haselgrove, C., . . . Dale, A.
M. (2002). Whole brain segmentation: automated labeling of neuroanatomical
structures in the human brain. Neuron, 33(3), 341-355.
Fling, B. W., Cohen, R. G., Mancini, M., Carpenter, S. D., Fair, D. A., Nutt, J. G., &
Horak, F. B. (2014). Functional Reorganization of the Locomotor Network in
Parkinson Patients with Freezing of Gait. PLoS One, 9(6). doi:ARTN e100291
10.1371/journal.pone.0100291
Fling, B. W., Cohen, R. G., Mancini, M., Nutt, J. G., Fair, D. A., & Horak, F. B. (2013).
Asymmetric pedunculopontine network connectivity in parkinsonian patients with
freezing of gait. Brain, 136(Pt 8), 2405-2418. doi:10.1093/brain/awt172

134
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). "Mini-mental state". A practical
method for grading the cognitive state of patients for the clinician. J Psychiatr
Res, 12(3), 189-198.
Giladi, N., Shabtai, H., Simon, E. S., Biran, S., Tal, J., & Korczyn, A. D. (2000).
Construction of freezing of gait questionnaire for patients with Parkinsonism.
Parkinsonism Relat Disord, 6(3), 165-170.
Goetz, C. G., Tilley, B. C., Shaftman, S. R., Stebbins, G. T., Fahn, S., Martinez-Martin,
P., . . . Movement Disorder Society, U. R. T. F. (2008). Movement Disorder
Society-sponsored revision of the Unified Parkinson's Disease Rating Scale
(MDS-UPDRS): scale presentation and clinimetric testing results. Mov Disord,
23(15), 2129-2170. doi:10.1002/mds.22340
Guimera, R., & Amaral, L. A. (2005). Cartography of complex networks: modules and
universal roles. J Stat Mech, 2005(P02001), nihpa35573. doi:10.1088/1742-
5468/2005/02/P02001
Hall, J. M., Ehgoetz Martens, K. A., Walton, C. C., O'Callaghan, C., Keller, P. E., Lewis,
S. J., & Moustafa, A. A. (2016). Diffusion alterations associated with Parkinson's
disease symptomatology: A review of the literature. Parkinsonism Relat Disord,
33, 12-26. doi:10.1016/j.parkreldis.2016.09.026
Hall, J. M., Shine, J. M., O'Callaghan, C., Walton, C. C., Gilat, M., Naismith, S. L., &
Lewis, S. J. G. (2015). Freezing of Gait and its Associations in the Early and
Advanced Clinical Motor Stages of Parkinson's Disease: A Cross-Sectional Study.
J Parkinsons Dis, 5(4), 881-891. doi:10.3233/Jpd-150581
Herman, T., Rosenberg-Katz, K., Jacob, Y., Giladi, N., & Hausdorff, J. M. (2014). Gray
matter atrophy and freezing of gait in Parkinson's disease: Is the evidence black-
on-white? Mov Disord, 29(1), 134-139. doi:10.1002/mds.25697
Hoehn, M. M., & Yahr, M. D. (1967). Parkinsonism: onset, progression and mortality.
Neurology, 17(5), 427-442.
Jarbo, K., & Verstynen, T. D. (2015). Converging Structural and Functional Connectivity
of Orbitofrontal, Dorsolateral Prefrontal, and Posterior Parietal Cortex in the
Human Striatum. J Neurosci, 35(9), 3865-3878. doi:10.1523/jneurosci.2636-
14.2015

135
Lewis, S. J., & Shine, J. M. (2016). The Next Step: A Common Neural Mechanism for
Freezing of Gait. Neuroscientist, 22(1), 72-82. doi:10.1177/1073858414559101
Naismith, S. L., Shine, J. M., & Lewis, S. J. G. (2010). The Specific Contributions of Set-
Shifting to Freezing of Gait in Parkinson's Disease. Mov Disord, 25(8), 1000-
1004. doi:10.1002/mds.23005
Nichols, T. E., & Holmes, A. P. (2002). Nonparametric permutation tests for functional
neuroimaging: A primer with examples. Hum Brain Mapp, 15(1), 1-25.
doi:10.1002/hbm.1058
Nieuwboer, A., De Weerdt, W., Dom, R., & Lesaffre, E. (1998). A frequency and
correlation analysis of motor deficits in Parkinson patients. Disabil Rehabil,
20(4), 142-150.
Nutt, J. G., Bloem, B. R., Giladi, N., Hallett, M., Horak, F. B., & Nieuwboer, A. (2011).
Freezing of gait: moving forward on a mysterious clinical phenomenon. Lancet
Neurol, 10(8), 734-744. doi:10.1016/S1474-4422(11)70143-0
Park, H. J., & Friston, K. (2013). Structural and functional brain networks: from
connections to cognition. Science, 342(6158), 1238411.
doi:10.1126/science.1238411
Pietracupa, S., Suppa, A., Upadhyay, N., Giannì, C., Grillea, G., Leodori, G., . . .
Pantano, P. (2018). Freezing of gait in Parkinson’s disease: gray and white matter
abnormalities. J Neurol, 265(1), 52-62. doi:10.1007/s00415-017-8654-1
Plotnik, M., Giladi, N., & Hausdorff, J. M. (2012). Is Freezing of Gait in Parkinson's
Disease a Result of Multiple Gait Impairments? Implications for Treatment.
Parkinsons Dis, 2012, 8. doi:10.1155/2012/459321
Redgrave, P., Rodriguez, M., Smith, Y., Rodriguez-Oroz, M. C., Lehericy, S., Bergman,
H., . . . Obeso, J. A. (2010). Goal-directed and habitual control in the basal
ganglia: implications for Parkinson's disease. Nat Rev Neurosci, 11(11), 760-772.
Robbins, T. W., & Arnsten, A. F. T. (2009). The Neuropsychopharmacology of Fronto-
Executive Function: Monoaminergic Modulation. Ann Rev Neurosci, 32, 267-287.
doi:10.1146/annurev.neuro.051508.135535

136
Rubinov, M., & Sporns, O. (2010). Complex network measures of brain connectivity:
uses and interpretations. Neuroimage, 52(3), 1059-1069.
doi:10.1016/j.neuroimage.2009.10.003
Schweder, P. M., Hansen, P. C., Green, A. L., Quaghebeur, G., Stein, J., & Aziz, T. Z.
(2010). Connectivity of the pedunculopontine nucleus in parkinsonian freezing of
gait. Neuroreport, 21(14), 914-916. doi:10.1097/WNR.0b013e32833ce5f1
Sherman, S. M. (2016). Thalamus plays a central role in ongoing cortical functioning.
Nat Neurosci, 16(4), 533-541. doi:10.1038/nn.4269
Shine, J. M., Matar, E., Ward, P. B., Bolitho, S. J., Gilat, M., Pearson, M., . . . Lewis, S.
J. G. (2013). Exploring the cortical and subcortical fMRI changes associated with
freezing in Parkinson's disease. Brain, 136(4), 1204-1215.
Shine, J. M., Matar, E., Ward, P. B., Frank, M. J., Moustafa, A. A., Pearson, M., . . .
Lewis, S. J. (2013). Freezing of gait in Parkinson's disease is associated with
functional decoupling between the cognitive control network and the basal
ganglia. Brain, 136(Pt 12), 3671-3681. doi:10.1093/brain/awt272
Shine, J. M., Moore, S. T., Bolitho, S. J., Morris, T. R., Dilda, V., Naismith, S. L., &
Lewis, S. J. G. (2012). Assessing the utility of Freezing of Gait Questionnaires in
Parkinson's Disease. Parkinsonism Relat Disord, 18(1), 25-29.
doi:10.1016/j.parkreldis.2011.08.002
Shine, J. M., & Poldrack, R. A. (2017). Principles of dynamic network reconfiguration
across diverse brain states. Neuroimage. doi:10.1016/j.neuroimage.2017.08.010
Sporns, O., & Betzel, R. F. (2016). Modular Brain Networks. Annu Rev Psychol, 67, 613-
640. doi:10.1146/annurev-psych-122414-033634
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C. (2004). Organization,
development and function of complex brain networks. Trends Cogn Sci, 8(9),
418-425. doi:10.1016/j.tics.2004.07.008
Tessitore, A., Amboni, M., Esposito, F., Russo, A., Picillo, M., Marcuccio, L., . . .
Barone, P. (2012). Resting-state brain connectivity in patients with Parkinson's
disease and freezing of gait. Parkinsonism Relat Disord, 18(6), 781-787.
doi:10.1016/j.parkreldis.2012.03.018

137
van den Heuvel, M. P., & Sporns, O. (2011). Rich-Club Organization of the Human
Connectome. J Neurosci, 31(44), 15775-15786. doi:10.1523/jneurosci.3539-
11.2011
Vandenbossche, J., Deroost, N., Soetens, E., Coomans, D., Spildooren, J., Vercruysse, S.,
. . . Kerckhofs, E. (2012). Freezing of gait in Parkinson's disease: disturbances in
automaticity and control. Front Hum Neurosci, 6, 356.
doi:10.3389/fnhum.2012.00356
Vandenbossche, J., Deroost, N., Soetens, E., Zeischka, P., Spildooren, J., Vercruysse, S., .
. . Kerckhofs, E. (2012). Conflict and freezing of gait in Parkinson's disease:
support for a response control deficit. Neurosci, 206, 144-154.
doi:10.1016/j.neuroscience.2011.12.048
Vercruysse, S., Leunissen, I., Vervoort, G., Vandenberghe, W., Swinnen, S., &
Nieuwboer, A. (2015). Microstructural Changes in White Matter Associated With
Freezing of Gait in Parkinson's Disease. Mov Disord, 30(4), 567-576.
doi:10.1002/mds.26130
Walton, C. C., O'Callaghan, C., Hall, J. M., Gilat, M., Mowszowski, L., Naismith, S. L., .
. . Lewis, S. J. G. (2015). Antisaccade errors reveal cognitive control deficits in
Parkinson's disease with freezing of gait. J Neurol, 262(12), 2745-2754.
doi:10.1007/s00415-015-7910-5
Walton, C. C., Shine, J. M., Hall, J. M., O'Callaghan, C., Mowszowski, L., Gilat, M., . . .
Lewis, S. J. (2015). The major impact of freezing of gait on quality of life in
Parkinson's disease. J Neurol, 262(1), 108-115. doi:10.1007/s00415-014-7524-3
Wechsler, D. S. (1997). Wechsler Memory Scale. San Antonio XT: The Psychological
Corporation.
Wu, T., & Hallett, M. (2005). A functional MRI study of automatic movements in
patients with Parkinson's disease. Brain, 128(Pt 10), 2250-2259.
doi:10.1093/brain/awh569
Youn, J., Lee, J. M., Kwon, H., Kim, J. S., Son, T. O., & Cho, J. W. (2015). Alterations
of mean diffusivity of pedunculopontine nucleus pathway in Parkinson's disease
patients with freezing of gait. Parkinsonism Relat Disord, 21(1), 12-17.
doi:10.1016/j.parkreldis.2014.10.003

138
Zalesky, A., Fornito, A., Cocchi, L., Gollo, L. L., van den Heuvel, M. P., & Breakspear,
M. (2016). Connectome sensitivity or specificity: which is more important?
Neuroimage, 142, 407-420. doi:10.1016/j.neuroimage.2016.06.035

139
Supplementary Materials
Figure 1. Average degree of the rich club nodes *Nodes with significantly lower participation coefficient in freezers compared to non-freezers lh = left hemisphere; rh = right hemisphere
Table 1. Pearson correlation coefficient (p-value) between different thresholds across difference scores between freezers and non-freezers 40% vs. 50% 50% vs. 60% 40% vs. 60%
Participation coefficient 0.75 (<0.0001) 0.56 (<0.0001) 0.45 (<0.0001)
Module degree z-score 0.90 (<0.0001) 0.61 (<0.0001) 0.64 (<0.0001)

CHAPTER5
DYSFUNCTIONINATTENTIONALPROCESSINGIN
PATIENTSWITHPARKINSON’SDISEASEANDVISUAL
HALLUCINATIONS
This study is published in Journal of Neural Transmission Hall, J. M., O’Callaghan, C., Shine, J. M., Muller, A. J., Phillips, J. R., Walton, C. C., Lewis, S. J. G., Moustafa, A. A. Dysfunction in attentional processing in patients with Parkinson's disease and visual hallucinations. Journal of Neural Transmission, 2016.

141
Chapter 5: Dysfunction in attentional processing in patients with Parkinson’s
disease and visual hallucinations
Abstract
Mechanistic insights into visual hallucinations (VH) in Parkinson’s disease (PD) have
suggested a role for impaired attentional processes. The current study tested 25 PD
patients with and 28 PD patients without VH on the Attentional Network Test.
Hallucinators had significantly lower accuracy rates compared to non-hallucinators, but
no differences were found in reaction times. This suggests that hallucinators show deficits
in attentional processes and conflict monitoring. Our findings provide novel behavioural
insights that dovetail with current neurobiological frameworks of VH.
Key words: Parkinson’s disease, visual hallucinations, attentional network test, dorsal
attentional network

142
Introduction
One of the most disabling non-motor symptoms of Parkinson’s disease (PD) is the
emergence of visual hallucinations (VH) (Hely et al., 2008). VH often appear recurrently
without external stimuli (Goetz et al., 2010; Sanchez-Ramos et al., 1996) and exist along
a perceptual spectrum; from simple images without a recognizable form, to complex
hallucinations, which consist of well-formed images of faces, animals or scenery
(Santhouse et al., 2000). Once established, VH not only become more frequent over time
but many patients also report their VH as becoming more threatening (Goetz et al., 2001).
At present, several competing hypotheses have been proposed regarding the development
and neurobiology of VH (Muller et al., 2014). One proposal supported by recent
empirical evidence is that VH arise from a breakdown between large-scale attentional
networks across the brain (Shine et al., 2014). These comprise the dorsal attentional
network (DAN), which is involved in goal-driven and sustained attention, the ventral
attention network (VAN), which is responsible for the re-orienting of attention towards a
salient stimulus, and the default mode network (DMN), a network that supports
introspection, mental imagery and the capacity to retrieve and manipulate episodic
memories (Buckner et al., 2008). In the Attentional Networks Hypothesis, VH can
emerge due to a failure in recruiting the DAN during periods of conflict resolution, such
that an ambiguous visual input is incorporated in the visual processing stream (Shine et
al., 2011a). Interpreting incoming perceptual information then relies on the VAN and
DMN. Importantly, increased activity in the DMN allows for augmented connectivity of
the visual cortices (Shine et al., 2015), influencing ongoing activity within the visual
stream. This increased coupling may predispose an individual to hallucinate. In the
context of over-activation of the DMN, autobiographical information can be incorporated
to resolve the ambiguous input, such that familiar people or objects might be
hallucinated, and aberrant activation of the VAN can lead to an increased salience
attached to the percept (Shine 2011a; Yao et al., 2014). Finally, the frontoparietal control
network mediates activity between the three attentional networks (Petersen et al., 2012).
Here, we further explore attentional processing deficits in PD patients with VH using the

143
Attentional Network Test (ANT) (Fan et al., 2002). This computerized task is a
combination of a cued reaction time task (Posner, 1980) and a flanker task (Eriksen et al.,
1974). Successful performance of this task requires sustained attention and visual search,
which are regulated by the DAN, and conflict resolution abilities, a task of the
frontoparietal control network (Fan et al., 2002). Therefore, we hypothesized that a
disruption within and between these attentional networks, as seen in PD VH, would lead
to a deficit in the overall performance of the ANT, with a profound impairment on the
conflict resolution task.
Methods
Participants
Fifty-three non-demented PD patients were recruited from the Parkinson’s Disease
Research Clinic at the Brain and Mind Centre, University of Sydney. All patients
satisfied the UKPDS Brain Bank Criteria and informed consent was obtained from all
individuals in the study. Permission was acquired from the local Ethics Committee. In
keeping with previous work (Goldman et al., 2014), PD patients were classified as
hallucinators (n=25) or non-hallucinators (n=28) according to their response on question
2 of the MDS-UPDRS (Goetz et al, 2010): “Over the past week have you seen, heard,
smelled or felt things that were not really there? [If yes, examiner asks patient or
caregiver to elaborate and probes for information]” (hallucinators ≥1 (slight to severe
VH); non-hallucinators = 0 (no hallucinations or psychotic behaviour).
Assessments
Groups were matched on age, disease duration, Levodopa equivalent dose (LED), motor
severity (Hoehn and Yahr clinical staging; H&Y (Hoehn & Yahr, 1967) and MDS-
UPDRS part III (Goetz et al., 2008), mood (Beck’s Depression Inventory (BDI-II)) (Beck
et al., 1961) and general cognition (Mini Mental State Examination (MMSE) (Folstein et
al., 1983). See Table 1 for demographics and clinical characteristics.

144
Table 1. Demographical data and clinical characteristics (Mean (SD)) PD Non-VH PD-VH p-value
N 28 25 n.a.
Age (y) 66.65 (8.20) 69.28 (5.74) .187
Duration (y)a,b 6.18 (3.37) 6.74 (5.22) .652
H&Y 2.11 (0.39) 2.24 (0.60) .406
MDS-UPDRS iiib,c 28.15 (11.95) 34.63 (15.26) .096
LEDa 590.50 (259.79) 688.50 (395.63) .553
MMSEa 28.58 (1.47) 28.72 (1.72) .596
BDI-IIc 7.30 (6.38) 12.64 (11.64) .054
PD Non-VH = Parkinson’s disease patients without visual hallucinations; PD-VH = Parkinson’s disease patients with visual hallucinations; H&Y = Hoehn and Yahr clinical motor staging; MDS-UPDRS iii = Part three of the Movement Disorder Society Unified Parkinson’s Disease Rating Scale; LED = Levodopa equivalent dose; MMSE = Mini Mental State Examination; BDI-II = Beck’s Depression Inventory (revised) a PD Non-VH: n= 26; b PD-VH: n = 24; c PD Non-VH: n = 27; Note: The removal of RTs of patients with a response percentage < 70% did not affect these results.
Attentional Network Test
A shortened version of the ANT (Fan et al., 2002) was used to explore attentional
processes. The task and its procedure are explained elsewhere in more detail (Fan et al.,
2002). Both accuracy and reaction time (RT) were recorded. Each participant completed
96 trials. Warning cues appeared before the presentation of the target. There were four
types of warning cues: double (two asterisk at the top and bottom of the screen) centre
(one asterisk in the centre of the screen), spatial (one asterisk in the position where the
target would appear) or no asterisk. Congruent or incongruent flanker arrows or neutral
lines surround the target arrow. In line with the original study of Fan and colleagues
(2002), three attentional processes can be calculated using the following methods:
Alerting = RT no cue - double cue condition; Orienting = RT centre cue – RT spatial cue
condition; Executive network = RT incongruent – RT congruent condition
Data processing and statistical analyses
All data analyses were performed using IBM SPSS version 22. All analyses were two-
tailed using a significance level of 0.05. Due to the non-parametric nature of the task
data, response percentage and accuracy were analysed using Mann Whitney U tests for
different flanker and cue conditions for the groups. Corrections for multiple comparisons

145
were not implemented to keep the risk of a Type 2 error minimal (Perneger, 1998).
However, effect sizes of the accuracy data were calculated to determine the strength of
the outcomes (Table 2). RT of the ANT was analysed using 2 (group) x 4 (cue condition)
x 3 (flanker type) mixed factorial ANOVAs. For the alerting, orienting and executive
aspects of the task we performed three separate 2x2 repeated measures ANOVAs with
group as between-subject factor, and cue type (alerting: no- and double-cue trials;
orienting: centre- and spatial-cue trials) or target type (executive: incongruent and
congruent flankers) as within-subject factors. The RT of erroneous responses were
excluded from the analyses. Furthermore, patients with a response percentage of < 70%
(five hallucinators and one non-hallucinator) were excluded from further analyses as their
RT were not considered reliable.
Results
ANT Accuracy
Missed and erroneous responses were combined to calculate accuracy percentages.
Overall, hallucinators had significantly lower accuracy compared to non-hallucinators (p
= 0.016). As shown in Table 2, hallucinators had more erroneous responses on all
incongruent flanker condition (p-values < 0.05) apart from the no cue condition (p =
0.115), and in selected trials of the neutral flanker condition (no cue and spatial cue
condition: p-values < 0.05) compared to non-hallucinators. No differences were found
between the groups on the congruent flanker condition (p-values > 0.05; see table 2).
ANT Reaction times (RT)
Reaction times are shown in Table 2. No interaction effect was found for group x cue x
flanker type (p = 0.566), cue x flanker type (p = 0.821), group x cue (p = 0.904) and
group x flanker type (p = 0.284). Main effects for cue condition (p = 0.002) and flanker
type (p < 0.001) were found. Overall, both groups were slower to respond to targets
surrounded by incongruent flankers compared to neutral and congruent flankers, and
when no, a centre or double cue was presented in comparison to a spatial cue. Finally, no
main group differences were found (p = 0.145).

146
Attentional Processes
As shown in Table 2, the 2x2 repeated measures showed no interaction effects (alerting: p
= 0.719; orienting: p = 0.600; executive control: p = 0.815). A significant main effect
was found for the orienting condition (p = 0.001), demonstrating that RT were lower for
the spatial cue condition compared to the centre condition across all patients.
Furthermore, a significant executive control effect was found (p < 0.001), demonstrating
that RT were longer for the incongruent condition compared to the congruent flanker
condition across all patients. No alerting effect was found (p = 0.649). Finally, no main
group differences were found for the alerting (p = 0.139), orienting (p = 0.115) or
executive control processes (p = 0.146).
Table 2. Accuracy percentages for each cue and flanker condition (Mean (SD)) and
reaction times of the attentional networks (Mean (SD)) Flanker Cue PD-VH PD-Non VH U z r p-value
Incongruent Total 73.00 (28.80) 88.84 (14.71) 218.50 -2.361 -0.32 .018
Spatial 71.50 (33.76) 89.29 (18.55) 241.00 -2.108 -0.29 .035
Double 72.50 (30.83) 89.73 (14.46) 228.50 -2.272 -0.31 .023
Centre 74.50 (30.93) 91.07 (12.66) 246.50 -1.976 -0.27 .048
None 73.50 (30.26) 87.95 (15.40) 265.50 -1.576 -0.22 .115
Congruent Total 84.13 (27.06) 94.64 (10.31) 262.00 -1.651 -0.23 .099
Spatial 83.50 (26.45) 93.75 (13.82) 279.50 -1.471 -0.20 .141
Double 86.00 (28.71) 93.75 (13.39) 329.00 -0.483 -0.07 .629
Centre 83.50 (28.80) 93.75 (13.82) 295.00 -1.167 -0.16 .243
None 83.50 (28.12) 93.75 (12.03) 268.00 -1.667 -0.23 .095
Neutral Total 86.25 (19.97) 94.31 (13.23) 199.50 -2.771 -0.38 .006
Spatial 88.00 (15.92) 95.54 (9.14) 247.00 -2.124 -0.29 .034
Double 89.00 (20.19) 95.09 (13.75) 273.00 -1.734 -0.24 .083
Centre 86.00 (25.60) 92.86 (14.20) 305.00 -0.954 -0.13 .340
None 82.00 (23.96) 95.54 (10.87) 205.00 -2.943 -0.40 .003
F (df, df error) η2 p-value
Alerting RT Group 0.86 (60.82) 7.30 (60.82) 2.265 (1, 45) 0.048 .139
Cue 0.210 (1, 45) 0.005 .649
Group x cue 0.131 (1, 45) 0.003 .719
Orienting RT Group 38.74 (61.80) 28.96 (63.29) 2.580 (1, 45) 0.054 .115
Cue 13.410 (1, 45) 0.230 .001
Group x cue 0.280 (1, 45) 0.006 .600
Executive RT Group 158.15 (115.04) 151.44 (80.77) 2.185 (1, 45) 0.046 .146

147
PD Non-VH = Parkinson’s disease patients without visual hallucinations; PD-VH = Parkinson’s disease patients with visual hallucinations; RT = reaction time; Executive = Executive control
Discussion
Here we explored attentional processing in PD patients with VH and identified a specific
impairment in their accuracy levels. Whilst no differences in RT were observed,
hallucinators had more missed and erroneous responses compared to non-hallucinators.
This difference was most apparent in the incongruent task conditions, where the accuracy
rate of hallucinators dropped to 73% reflecting greater impairments in attentional
processing with increasing task demands. Our findings are consistent with the
hypothesised attentional network dysfunction thought to play a role in the emergence of
hallucinations in PD.
Sustained attention and visual search, both crucial for successful performance on the
overall and incongruent task condition is regulated by the DAN. It is worth noting that
clinically more VH occur in the context of cluttered or distracting environments. Conflict
monitoring is regulated by the frontoparietal control regions (Petersen et al., 2012; Wang
et al., 2010), and disruptions within this network could add to the deficiency in the
incongruent task condition. Moreover, the frontoparietal control network mediates
activity between the DAN and DMN (Buckner et al., 2008). Failure to recruit the DAN is
in keeping with recently proposed hypothesis that VH patients increasingly rely on the
VAN and DMN (Buckner et al., 2008; Shine et al., 2014).
Similar RT scores for the alerting, orienting and executive control networks were found
between the groups, suggesting that the incorrect responses were not mediated by
increased response latencies in the hallucinators. This suggests that when coordination
within and between each of the major attentional networks is effective, hallucinators are
able to perform just as well on the attentional task as non-hallucinators. However, when a
disruption of these attentional processes arise, VH patients fail to resolve conflicting
Cue 117.689 (1, 45) 0.723 <.001
Group x cue 0.055 (1, 45) 0.001 .815

148
information resulting in an erroneous response rather than a delayed response, which is in
keeping with the episodic nature of VHs.
In conclusion, we have demonstrated that hallucinators showed marked impairments in
attentional processing, especially in more distracting conditions. Our results support the
crucial role for deficits within attentional networks in the manifestation of VH in PD,
which represents an important target for future pharmacological and behavioural
treatment strategies.
Acknowledgements
We wish to thank our patient volunteers of the Parkinson’s Disease Research Clinic for
their time and efforts.

149
References
Beck, A. T., Ward, C. H., Mendelson, M., Mock, J., Erbaugh, J. (1961) An inventory for
measuring depression. Arch Gen Psychiatry, 4:561-571
Buckner, R. L., Andrews-Hanna, J. R., Schacter, D. L. (2008) The brain's default
network: anatomy, function, and relevance to disease. Ann NY Acad Sci, 1124:1-
38 doi:10.1196/annals.1440.011
Eriksen, B. A., Eriksen, C. W. (1974) Effects of noise letters upon the identification of a
target letter in a nonsearch task. Percept Psychophysics, 16:143-149
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., Posner, M. I. (2002) Testing the
efficiency and independence of attentional networks. J Cogn Neurosci, 14:340-
347 doi:10.1162/089892902317361886
Folstein, M. F., Robins, L. N., Helzer, J. E. (1983) The Mini-Mental State Examination.
Arch Gen Psychiatry, 40:812
Goetz, C. G., Tilley, B. C., Shaftman, S. R., Stebbins, G. T., Fahn, S., Martinez-Martin,
P., … Movement Disorder Society UPDRS Revision Task Force. (2010)
Movement Disorder Society-sponsored revision of the Unified Parkinson's
Disease Rating Scale (MDS-UPDRS): scale presentation and clinimetric testing
results. Mov Disord, 23(15): 2129-70. doi: 10.1002/mds.22340.
Goetz, C. G., Leurgans, S., Pappert, E. J., Raman, R., Stemer, A. B. (2001) Prospective
longitudinal assessment of hallucinations in Parkinson's disease. Neurology,
57:2078-2082
Goetz, C. G., Ouyang, B., Negron, A., Stebbins, G. T. (2010) Hallucinations and sleep
disorders in PD: ten-year prospective longitudinal study. Neurology, 75:1773-
1779 doi:WNL.0b013e3181fd6158 [pii]10.1212/WNL.0b013e3181fd6158
Goldman, J. G., Stebbins, G. T., Dinh, V., Bernard, B., Merkitch, D., deToledo-Morrell,
L., Goetz, C. G. (2014) Visuoperceptive region atrophy independent of cognitive
status in patients with Parkinson's disease with hallucinations. Brain, 137:849-859
doi:10.1093/brain/awt360

150
Hely, M. A., Reid, W. G., Adena, M. A., Halliday, G. M., Morris, J. G. (2008) The
Sydney multicenter study of Parkinson's disease: the inevitability of dementia at
20 years. Mov Disord, 23:837-844 doi:10.1002/mds.21956 [doi]
Hoehn, M. M., Yahr, M. D. (1967) Parkinsonism: onset, progression and mortality.
Neurology, 17:427-442
Muller, A. J., Shine, J.M., Halliday, G. M., Lewis, S. J. G. (2014) Visual hallucinations in
Parkinson's disease: theoretical models. Mov Disord, 29:1591-1598
doi:10.1002/mds.26004
Perneger, T. V. (1998) What's wrong with Bonferroni adjustments. Bmj, 316:1236-1238
Petersen, S. E., Posner, M. I. (2012) The attention system of the human brain: 20 years
after. Ann Rev Neurosci 35:73-89 doi:10.1146/annurev-neuro-062111-150525
Posner, M. I. (1980) Orienting of attention. Quarterly J Exp Psychology, 32:3-25
Sanchez-Ramos, J.R., Ortoll, R., Paulson, G.W (1996) Visual hallucinations associated
with Parkinson disease. Arch Neurol, 53:1265-1268
Santhouse, A. M., Howard, R. J, ffytche, D. H. (2000) Visual hallucinatory syndromes
and the anatomy of the visual brain. Brain, 123 (Pt 10):2055-2064
Shine, J. M., Halliday, G. M., Naismith, S. L., Lewis, S. J. G. (2011a) Visual
misperceptions and hallucinations in Parkinson's disease: dysfunction of
attentional control networks? Mov Disord, 26:2154-2159 doi:10.1002/mds.23896
Shine, J. M., Muller, A. J., O'Callaghan, C., Hornberger, M., Halliday. G. M., Lewis, S. J.
G. (2015) Abnormal connectivity between the default mode and the visual system
underlies the manifestation of visual hallucinations in Parkinson's disease: a task-
based fMRI study. NPJ Parkinson's Dis, 4
Shine, J. M., O'Callaghan, C., Halliday, G. M., Lewis, S. J. G. (2014) Tricks of the mind:
Visual hallucinations as disorders of attention. Prog Neurobiol, 116:58-65
doi:10.1016/j.pneurobio.2014.01.004
Shine J. M., Ward, P. B., Naismith, S. L., Pearson, M., Lewis, S. J. G. (2011b) Utilising
functional MRI (fMRI) to explore the freezing phenomenon in Parkinson's
disease. J Clin Neurosci, 18:807-810 doi:10.1016/j.jocn.2011.02.003

151
Wang, L,, Liu, X., Guise, K. G., Knight, R. T., Ghajar, J., Fan, J. (2010) Effective
connectivity of the fronto-parietal network during attentional control. J Cogn
Neurosci, 22:543-553 doi:10.1162/jocn.2009.21210
Yao, N., Shek�Kwan Chang, R., Cheung, C., Pang, S., Lau, K. K., Suckling, J., . . .
Chua, SE. (2014). The default mode network is disrupted in Parkinson's disease
with visual hallucinations. Hum Brain Mapp, 35(11), 5658-5666.

CHAPTER6
CHANGESINSTRUCTURALNETWORKTOPOLOGY
CORRELATEWITHSEVERITYOFHALLUCINATORY
BEHAVIOURSINPARKINSON’SDISEASE
This study is published in Network Neuroscience
Hall, J. M., O’Callaghan, C., Muller, A. J., Ehgoetz Martens, K.A., Phillips, J. R., Moustafa, A. A., Lewis, S. J. G., Shine, J. M. Changes in structural network topology correlate with severity of hallucinatory behaviour in Parkinson's disease. Network Neurocience, 2019.
DOI:10.1101/383141

153
Chapter 6: Changes in structural network topology correlate with severity of
hallucinatory behaviour in Parkinson’s disease
Abstract
Inefficient integration between bottom-up visual input and higher-order visual processing
regions is implicated in visual hallucinations in Parkinson’s disease (PD). Here, we
investigated white matter contributions to this perceptual imbalance hypothesis. Twenty-
nine PD patients were assessed for hallucinatory behaviour. Hallucination severity was
correlated to connectivity strength of the network using the Network Based Statistic
approach. The results showed that hallucination severity was associated with reduced
connectivity within a sub-network that included the majority of the diverse club. This
network showed overall greater between-module scores compared to nodes not associated
with hallucination severity. Reduced between-module connectivity in the lateral occipital
cortex, insula and pars orbitalis, as well as decreased within-module connectivity in the
prefrontal, somatosensory and primary visual cortices were associated with hallucination
severity. Conversely, hallucination severity was associated with increased between- and
within-module connectivity in the orbitofrontal and temporal cortex, as well as regions
comprising the dorsal attentional and default mode network. These results suggest that
hallucination severity is associated with marked alterations in structural network topology
with changes in participation along the perceptual hierarchy. Therefore, impaired
integration across the perceptual hierarchy may result in the inefficient transfer of
information that gives rise to hallucinations in PD.
Key words: Parkinson’s disease, visual hallucinations, diffusion tensor imaging, network
topology, graph theory, connectomics

154
Introduction
Visual hallucinations (VHs) in Parkinson’s disease (PD) exist on a spectrum ranging
from simple misperceptions, to complex well-formed images (Barnes & David, 2001).
With disease progression and loss of insight, VHs constitute a major source of distress for
the patient (Goetz, 2009; Schrag, 2004) and comprise a high degree burden for caregivers
(Aarsland et al., 2007). Risk factors of VHs include older age and disease duration, sleep
and mood disturbances as well as cognitive decline (Barnes & David, 2001; Fénelon,
Mahieux, Huon, & Ziégler, 2000; Lenka, Hegde, Arumugham, & Pal, 2017).
Furthermore, previous work has shown that patients with VHs show disruptions in
attentional processing (Hall et al., 2016), reduced performance on visuoperceptive tasks
(Barnes, Boubert, Harris, Lee, & David, 2003; Gallagher et al., 2011; Ramirez-Ruiz,
Junque, Marti, Valldeoriola, & Tolosa, 2006), decreased visual contrast sensitivity,
colour discrimination (Diederich et al., 1998) and acuity (Matsui et al., 2006). Current
models of VHs have therefore focused on the interaction of perceptual and attentional
dysfunction (for review, see Muller, Shine, Halliday, & Lewis, 2014). Specifically, it has
been proposed that failure to effectively integrate information from different processing
sites across the perceptual hierarchy is likely to contribute to VHs and misperceptions in
PD (Collerton, Perry & McKeith, 2005; Diederich, Goetz, & Stebbins, 2005; Muller et
al., 2014; Shine, Halliday, Carlos, Naismith, & Lewis, 2012).
Attention, prior experience and expectations strongly influence perception. Perceptual
predictions, generated from a myriad of modalities across the brain, guide perceptual
processes to facilitate the interpretation of noisy and ambiguous input (Bar, 2009; Engel,
Fries, & Singer, 2001; Summerfield et al., 2006). The orbitofrontal cortex (OFC) process
coarse information projected from the visual cortex and provides an “initial guess” of an
object’s identity (Summerfield & Koechlin, 2008). Previous work in PD patients with
VHs has shown that the accumulation of sensory evidence is slow and inefficient, which
may result in an over-reliance on these top-down predictions (O’Callaghan et al., 2017).
Importantly, top-down visual processing regions can modulate neural activity in early
visual regions, with expected stimuli leading to reduced activity (Meyer & Olson, 2011).
Additionally, activity within the default mode network (DMN), a network involved in

155
mediating endogenous perception, has shown to be increased during a misperception in
this patient population (Shine et al., 2014). Therefore, VHs may arise when perceptual
input is not properly integrated and internally generated images interfere with the
perceptual process (Fletcher & Frith, 2008; Intaite, Noreika, Soliunas, & Falter, 2013;
O’Callaghan et al., 2017; Powers, Kelley, & Corlett, 2016).
While functional neuroimaging studies have made significant contributions to our
understanding (Hepp, Foncke, Dubbelink, et al., 2017; Ramírez�Ruiz et al., 2008;
Shine, Halliday, et al., 2014; Shine, Muller, et al., 2015; Yao et al., 2014), less is known
about the involvement of white matter changes in the manifestation of VHs in PD.
Experiments using diffusion tensor imaging (DTI) have reported altered white matter
integrity in the optic nerve and optic radiation (Lee et al., 2016) as well as ascending
tracts from the cholinergic nucleus basalis of Meynert to parietal and occipital cortical
regions (Hepp, Foncke, Berendse, et al., 2017). However, given the involvement of large-
scale brain networks in perception, unique insights into white matter changes associated
with VHs can be gained by investigating whole brain network topology. Topological
features of the human connectome allow us to describe the arrangement of connections
within and between segregated sub-modules (Bullmore & Sporns, 2009). Specifically,
nodes that integrate these specialist communities are crucial for incorporating information
streams of different modalities, which is essential for processes such as perception
(Bertolero, Yeo, & D’Esposito, 2015; Muller, O'Callaghan, Walton, Shine, & Lewis,
2017). Therefore, investigating network topology can provide novel insights in changes
across different perceptual hierarchies.
The current study aimed to examine whether VHs are associated with changes in
structural network topology. To identify hallucinatory behaviour in patients with PD, we
assessed performance on a computerized task capable of inducing misperceptions (Shine
et al., 2012), in combination with a validated questionnaire that evaluates hallucinatory
behaviour in PD patients (Shine et al., 2015). We aimed to circumvent the sole reliance
on self-reported symptom occurrence whilst controlling for the possibility of
misclassifying hallucinators as non-hallucinators, when patients did not experience visual

156
hallucinations during their clinic visit or when ‘passage’ hallucinations go unreported.
Additionally, by creating this composite score, we are able to assess visual hallucinatory
severity, rather than classifying patients into artificial dichotomous patient groups.
We hypothesized that the severity of hallucinatory behaviour would be associated with
ineffective information processing as shown by reduced between-module scores in visual
networks, reflecting reduced visual input to integration centres. Furthermore, increased
between-module scores across top-down perceptual prediction areas and the DMN could
indicate an over-reliance on regions involved in the generation of internal percepts
(Shine, O'Callaghan, Halliday, & Lewis, 2014).
Methods
Twenty-nine patients with idiopathic PD were included in this study. Demographic
information including age, disease duration and levodopa dose equivalent (LEDD) were
obtained for all participants. All patients were assessed on the Hoehn & Yahr clinical
stage (Hoehn & Yahr, 1998) and the motor aspect of the MDS-UPDRS (part III) (Goetz
et al., 2008). Global cognition was assessed using the Mini-Mental State Examination
(MMSE) (Folstein, Robins, & Helzer, 1983) and set-shifting performance was assessed
using the Trail Making Test part B minus part A (TMTB-A) (Tombaugh, 2004). The study
was approved by the ethics committee of the University of Sydney and was in accordance
with the principles of the Helsinki Declaration. Written informed consent was obtained
from all participants before participation.
Bistable Percept Paradigm
All patients performed the Bistable Percept Paradigm (BPP) (James M Shine et al.,
2012), a behavioural task capable of inducing misperceptions in susceptible patients. In
this task, patients were presented with either single or bistable percepts (i.e. “hidden”
images as shown in Figure 1) for a maximum of 30 sec in a randomised order. The
participant had to decide whether the stimulus was a single or hidden image by a button
press and describe to the examiner what they had seen. The recorded responses included
the following: 1) correct single or correct hidden, 2) “missed”, recorded when the subject

157
perceived a single image when a bistable percept was presented and 3) “misperceptions”,
recorded when a subject incorrectly identified a single image as a bistable image, i.e.
incorrectly reported an image that was not presented on the screen.
Figure 1. Example of single and hidden images of the BPP (Shine, Halliday, et al., 2012a)
Psychosis and Hallucinations Questionnaire
All patients completed the Psychosis and Hallucinations Questionnaire (PsycH-Q)
(Shine, Mills, et al., 2015). The PsycH-Q (part A) consists of three subscales including
1) visual misperceptions, which includes questions about the presence of VHs, passage
hallucinations and three frequently reported contents of VHs including people, animals
and objects; 2) sensory misperceptions, including audition, touch, olfaction, and
gustation, and; 3) disordered thought and psychotic behaviour. Participants rated the
frequency of their symptoms on a 5-point Likert scale, ranging from 0 (“never
experienced”) to 4 (“experienced daily”). The total score was calculated by summing the
responses (Shine et al., 2015) (see Supplementary Materials). Part B of the PsycH-Q
assesses symptoms related to VHs (i.e. attention and sleep) and was not included in this
study.
Composite score
The percentage of misperceptions on the BPP (‘indirect’ measure of VH) and the total
score on the PsycH-QA (‘direct’ measure of hallucinatory behaviour) were standardized

158
and then summed to create a composite score that reflected the severity of hallucinatory
behaviour (hereafter referred to as the hallucination severity score (HSS)). The HSS was
correlated with the demographic variables using parametric or non-parametric
correlations depending on the distribution of the variables and was used as a correlate in
the imaging analysis.
MRI Acquisition
All participants underwent magnetic resonance imaging (MRI) using a 3-Tesla General
Electric Discovery MR750 scanner (GE Medical Systems) with an 8-channel phased
array head coil. Diffusion weighted images (DWI) were obtained by using echo-planar
imaging sequences with 61 different motion probing gradient directions (TR/TE: 7025/80
ms, 55 transverse slices, slice thickness: 2.5 mm, matrix: 256 × 256, FOV: 240 x 240
mm). The effective diffusion weighting was b = 1000 s/mm2 and four volumes with no
diffusion weighting (b = 0 s/mm2) were obtained at the beginning of each diffusion
sequence. 3D T1-weighted, anatomical images were obtained (TR/TE/TI: 7.2/2.7/450 ms,
voxel size 1 × 1 × 1 mm, 196 transverse slices, 256 × 256 matrix, FOV: 256 x 256 mm,
flip angle 12°). The 3D-T1 images were used for individual registration between T1
weighted anatomical and the DWI images and cortical parcellation using FreeSurfer
(version 5.3; http://surfer.nmr.mgh.harvard.edu).
Diffusion tensor imaging pre-processing and deterministic fibre tracking
DTI pre-processing was performed using the FMRIB Software Library (FSL,
http://fsl.fmrib.ox.ac.uk). The pre-processing steps were as follows: i) DTI images were
corrected for susceptibility, head motion and eddy current induced geometrical distortions
using FSL’s tool eddy; ii) a binary brain mask was created using bet; iii) images were
realigned using a rigid body registration to the b = 0 image; then iv) a tensor was fitted in
each voxel (Chang, Jones, & Pierpaoli, 2005); followed by v) the computation of the
fractional anisotropy (FA) level based on the eigenvalues for each voxel, in order to
determine the preferred diffusion direction within a voxel. FA thus serves as a surrogate
measure of white matter integrity, with lower levels of FA reflecting reduced white

159
matter integrity (Sun et al., 2003; van den Heuvel & Sporns, 2011; Verstraete et al.,
2010). The preferred diffusion direction information was then used to reconstruct the
white matter tracts of the brain using a deterministic tracking approach based on the fibre
assignment by continuous tracking (FACT) algorithm (Mori, Crain, Chacko, & van Zijl,
1999). Deterministic tractography yields less false positive tracts compared to
probabilistic methods (Bastiani, Shah, Goebel, & Roebroeck, 2012). False positives are
detrimental in network modularity as they occur more prevalently between than within
modules (Zalesky et al., 2016). A streamline was started from eight seeds within each
voxel of the brain (grey and white matter) following the main diffusion direction of the
voxel and stopped when: i) the FA value < 0.1; ii) the traced fibre made a turn >45°; or
iii) when the tract left the brain mask. The images were acquired when reverse phase-
encoding direction approaches were not the standard procedure within acquisition
protocols, which could have influenced the registration of diffusion and anatomical
images. Therefore, anatomically constrained tractography was not applied (Smith,
Tournier, Calamante, & Connelly, 2012). The atlas presented by Cammoun et al. (2012)
was used, including 219 cortical regions and 14 subcortical regions. The weighted brain
network was calculated for each participant and consistency thresholding at 50% was
applied (i.e. including the tracts found in 50% of the patients) (de Reus & van den
Heuvel, 2013). The mean density of the thresholded group matrix was 8.7%. To verify
the results were not skewed by the choice of threshold, we also applied the thresholding
method that retained most consistent edges across subjects but controlling for their
distance (i.e. the consistency of edges within "bins" based on their length to avoid
preferential retention of short edges (Misic et al., 2015)). The mean density of the group
matrix using this threshold was 13.2%.
Network Based Statistic
A Network Based Statistic (NBS) analysis was applied to investigate whether the HSS
was associated with altered connectivity strength in an interconnected sub-network of the
brain (Zalesky, Fornito, & Bullmore, 2010). NBS is a non-parametric method for
connectome-wide analysis, which aims to detect specific pairs of brain regions showing a
significant effect of interest, while controlling for family-wise-error (FWE) rate.

160
Importantly, no inferences of individual connections are made; instead the null-
hypothesis can only be rejected at the sub-network level. As such, NBS is similar to the
cluster-based multiple-comparisons approaches used in standard functional MRI analysis.
To identify changes in sub-networks associated with the HSS, the t-statistic was set at 1.7
determined using the critical value of the t-distribution for our sample size (Field, 2009).
Additionally, we verified the robustness of the results by controlling for disease severity
using the MDS-UPDRS III (motor part) as covariate. Connections were deemed
significant at FWE-corrected p-value < 0.05 (one-sided) using 5000 permutations.
To investigate whether the sub-network involved particular functional networks, we
investigated whether nodes in the sub-network that correlated with the HSS overlapped
with previously defined resting state networks. To this end, seven canonical resting state
networks from the Yeo et al. atlas (2011) were overlaid with the structural parcellation
and the percentage of nodes from each network included within the structural sub-
network that inversely related to HSS was calculated for each resting state network. To
analyse whether this overlap occurred significantly above chance, we randomly permuted
the resting state network identity of each region (5000 iterations) and used the overlap
between the randomized vector and the original node assignment to populate a null
distribution. To test whether each individual resting state network overlapped with the
significant sub-network, their overlap was compared with the null distributions. A resting
state network was identified as targeted if the true overlap was more than the 97.5th
percentile of null distribution (i.e. the top 2.5%). A network was considered not to be
associated with the HSS if the overlap was less than the 2.5th percentile of the null
distribution.
Graph theoretical analysis
The graph organizational measures were computed using the Brain Connectivity Toolbox
(http://www.brain-connectivity-toolbox.net) (Rubinov & Sporns, 2010). The thresholded,
weighted brain networks were then partitioned into modules, which are non-overlapping
groups of highly connected nodes that are only sparsely connected with other modules,
using the Louvain algorithm (Rubinov & Sporns, 2010). To account for the stochastic

161
nature of the Louvain algorithm, a consensus partition was identified by calculating the
module assignment for each node 500 times. To define an appropriate value for the
resolution parameter (γ), the Louvain algorithm was iterated 100 times across a range of
values (0.5 – 2.0 in steps of 0.1) of the group mean connectivity matrix and then
estimated the similarity of the resultant partitions using mutual information. The γ
parameter of 1.9 provided the most robust estimates of topology across the iterations and
was used to determine the optimal resolution of the network modularity.
After the nodes were assigned to their modules, their intra- and inter-modular
connectivity were calculated. Intra-modular connectivity was calculated using the module
degree z-score Wi (see equation 1), in which a positive score reflects high within-module
connections (compared to the node’s average number of connections), and negative z-
scores denote the opposite. Inter-modular connectivity was calculated using the
participation coefficient Bi (see equation 2). Low Bi values indicate few between-module
connections, whereas high Bi values indicate uniformly distributed connections across
modules (github.com/juliemaehall/topology). High Wi and high Bi scores are not mutually
exclusive (Guimerà & Nunes Amaral, 2005).
!! =!!!!!!!!!!
[1]
Equation 1 – Module degree z-score Wi, where κi is the strength of the connections of region i to other regions in its
module si, κ!! is the average of κ over all the regions in si, and !!!! is the standard deviation of κ in si
!! = 1− !!"!!
!!!!!! [2]
Equation 2 - Participation coefficient Bi, where κis is the strength of the positive connections of region i to regions in
module s, and κi is the sum of strengths of all positive connections of region i. The participation coefficient of a region
is therefore close to 1 if its connections are uniformly distributed among all the modules and 0 if all of its links are
within its own module.
To test whether nodes within the sub-network identified using the NBS analysis differed
from nodes not included in the sub-network, the average Wi and Bi of the sub-network

162
were contrasted against the average Wi and Bi of the nodes not included in the sub-
network using non-parametric permutation testing.
To test whether the HSS correlated with the Wi and Bi nodes across the whole brain
connectome, a Spearman’s rho correlation was performed followed by a non-linear
permutation test using 5000 iterations to control for multiple comparisons (Nichols &
Holmes, 2002), using an alpha of 0.05. This approach was repeated using the different
threshold (Misic et al., 2015) and the outcome was correlated to the Wi and Bi using the
original threshold. Both the Wi and Bi scores calculated using the aforementioned
threshold highly correlated with the Wi and Bi scores calculated with the consensus
threshold (r = 0.92 and r = 0.94, respectively), indicating that the results were not biased
by the chosen thresholding method.
Diverse club analysis
We identified the ‘diverse club’ of the network, which comprised of the top 20% of Bi
nodes (Bertolero, Yeo, & D'Esposito, 2017). These nodes play an important role in
network integration and changes to these nodes could affect between module
communication (Bertolero et al., 2017). We normalized the diverse club coefficient in
reference to a null model: a random vector with a preserved modular structure was
created by randomizing the mean participation coefficient of each node for 5000
iterations. The diverse club was identified as those regions with a participation
coefficient greater than the 95th percentile of the permuted distribution. We investigated
whether the number of diverse club nodes was significantly higher within the sub-
network associated with the HSS, compared to nodes that were not included in the sub-
network identified using the NBS analysis.
Results
Demographics
Table 1 presents the descriptive variables of the 29 patients. The mean percentage of
misperceptions on the BPP was 18.48 (range: 0 – 49), and the mean score on the PsycH-
QA was 9.48 (range: 0 – 34 (max score = 52)), highlighting a diverse range of

163
hallucinatory behaviour in the patient cohort. The PsycH-QA and the BPP score showed a
positive, significant correlation (r = 0.52, p = 0.004). Finally, to verify the severity score
to the ‘gold standard’, we correlated the HSS in a large cohort of patients with PD and
Lewy Body Dementia (n = 75) to the MDS-UPDRS item 2 and found a correlation of r =
0.53 (p < 0.001). However, given higher construct validity (Shine et al., 2015), we opted
to include the PsycH-QA and the BPP scores in the composite score (HSS) for the
remainder of our analysis.
Table 1. Demographics and clinical variables
Variable Mean (range) Correlation with HSS
r (p-value) Age (y) 66.8.1 ± 8 (51 – 84) 0.18 (0.345)
Duration (y) 5.8 ± 4 (1.2 – 16) -0.05 (0.785)
LEDD 617.8 ± 392 (125 – 1548) -0.06 (0.767)
H&Y 1.9 ± 0.5 (1 – 3) 0.01 (0.949)
MDS-UPDRS III 30.0 ± 14 (7 – 55) 0.29 (0.125)
MMSE 28.5 ± 2 (25 – 30) -0.06 (0.762)
TMTB-A 78.9 ± 65 (-1 – 123) 0.33 (0.08)
BPP % misperceptions 18.48 ± 17 (0 – 49) -
PsycH-QA 9.48 ± 8 (0 – 34) -
LEDD = Levodopa equivalence daily dose; MDS-UPDRS III = motor part of the Movement Disorder Society Unified Parkinson’s
Disease Rating Scale; H & Y = Hoehn and Yahr; MMSE = Mini Mental State Examination; TMTB-A = Trail Making Test part B –
part A; BPP = Bistable Percept Paradigm; PsycH-QA = Psychosis and Hallucinations Questionnaire, part A
The HSS showed a positive correlation trending towards significance with the TMTB-A (r
= 0.33, p = 0.08). No significant correlations were observed between the HSS and other
demographic and clinical variables.
The HSS correlated with decreased connectivity in a large sub-network
As illustrated in Figure 2, the NBS analysis revealed a sub-network comprising 183 edges
(8% of the edges in the thresholded connectivity matrix) and 127 nodes with reduced FA-
based connectivity strength correlated to the HSS (p < 0.05). Using disease severity as a
covariate, the NBS analysis revealed a similar sub-network comprising 177 edges and
135 nodes that showed a correlation with the HSS that was trending towards significance

164
(p = 0.059). The effects presented with a fairly liberal threshold, suggesting the changes
related to the HSS are subtle yet topological extended (Zalesky et al., 2010). The size of
networks identified using a range of t-statistics are presented in the Supplementary
Materials. No significant sub-network was identified in the opposite direction (positive
correlation between the HSS and connectivity strength).
Furthermore, the group average Bi score within the sub-network was 0.506, which was
significantly higher (p < 0.05) than nodes outside this network, which show a group
average of 0.310. The group average Wi score of the nodes within the network was higher
than the group average Wi score of nodes outside the network (0.149 and -0.178,
respectively), yet this difference did not remain significant when controlling for disease
severity (p = 0.104).
Figure 2. NBS analysis reveals a sub-network, comprising 183 edges with reduced connectivity strength
correlated to increased HSS (p < 0.05). Figure visualized with BrainNet Viewer (Xia et al., 2013).

165
Figure 3: Overlap between the identified structural and functional resting-state networks. (A) The
percentage of nodes included in the sub-network for each resting state network. DMN = default mode
network; VAN = ventral attentional network; DAN = dorsal attentional network. (B) The functional resting
state networks of the Yeo atlas (Yeo et al., 2011).
The sub-network includes all sub-cortical nodes but did not target a specific cortical
resting state network
The sub-network that showed decreased connectivity strength correlated with the HSS
included all 14 sub-cortical nodes (p < 0.05). As illustrated in Figure 3, the sub-network
further included nodes across the cortex. However, none of the other the resting state
networks were significantly correlated with the HSS (p > 0.05), yet the somatomotor
network was relatively spared (p < 0.05).
Nodes in the sub-network show high participation scores
Eighteen nodes were included in the diverse club (see Supplementary Materials).
Seventeen of the eighteen nodes (94%) of the diverse club were included in the
aforementioned sub-network, which was deemed significantly above chance (p < 0.001).
As illustrated in Figure 4, nodes with high participation coefficients were more often part
of the sub-network.
0%
20%
40%
60%
80%
100%
Percentage of nodes include in the sub-network per functional network
A B

166
The HSS correlated with Wi and Bi scores
When investigating the whole structural connectome, the HSS positively correlated to
regional Bi (i.e. higher participation scores were associated with higher severity values)
for nodes in the left medial OFC, a node in the right anterior and left posterior cingulate,
precuneus, and the caudal middle frontal gyrus. Furthermore, nodes in the right occipital,
pars orbitalis and insula showed negative correlations between the HSS and participation
coefficient (i.e. lower participation scores were associated with higher scores on the HSS;
see Table 2 and Figure 5). However, when controlling for disease severity, the insula and
medial OFC were only trending towards significance, whilst the lateral occipital cortex
and anterior posterior cingulate did not remain significant.
Increased HSS scores were further associated with increased module degree z-scores in
the right thalamus, the bilateral lingual, left medial OFC, pars opercularis and
supramarginal gyrus and superior temporal cortices. The right lateral occipital cortex also
showed a positive correlation but this did not remain significant after controlling for
disease severity. Decreased HSS were associated with increased module degree z-scores
in the bilateral precuneus, the left parts triangularis, rostral middle frontal and superior
parietal cortex. The right pericalcarine and precentral gyrus also a showed negative
correlation with the HSS, yet this did not remain significant after controlling for disease
severity (see Table 3 and Figure 5).

167
Figure 4. Nodes ranked according to the Bi scores. Blue: nodes included in the sub-network; grey: nodes
not included in the sub-network correlated to the HSS.
Table 2. Spearman’s rho correlation between the participation coefficient and the HSS (p
< 0.05; permutation test) Node X Y Z Rho Rho* Sub-network
Positively correlated
Frontal
ctx-lh-medialorbitofrontal_2 -5 33 -20 0.37 0.35# �
ctx-rh-caudalmiddlefrontal_2 40 15 39 0.43 0.53 -
Parietal
ctx-lh-precuneus_2 -10 -44 46 0.40 0.37 �
Cingulate
ctx-lh-posteriorcingulate_2 -8 -43 21 0.57 0.53 �
ctx-rh-superiorfrontal_3 11 40 40 0.36 0.31^ �
Negatively correlated
Frontal
ctx-rh-parsorbitalis_1 43 43 -10 -0.46 -0.46 �
Occipital
ctx-rh-lateraloccipital_5 47 -73 2 -0.39 -0.21^ �
Insular
ctx-rh-insula_1 36 -18 13 -0.40 -0.32# �
* Rho after controlling for disease severity; # p < 0.1; ^ p > 0.1. ctx-lh = left hemisphere; ctx-rh = right hemisphere

168
Table 3. Spearman’s rho correlation between the module degree z-score and the HSS (p
< 0.05; permutation test) Node X Y Z Rho Rho* Sub-network
Positively correlated
Subcortical
Right-Thalamus-Proper 13 -17 7 0.49 0.45 �
Frontal
ctx-lh-medialorbitofrontal_1 -7 52 -11 0.40 0.43 -
ctx-lh-parsopercularis_2 -47 14 15 0.41 0.44 -
Parietal
ctx-lh-supramarginal_3 -55 -34 35 0.45 0.53 -
Temporal
ctx-rh-superiortemporal_4 46 -6 -11 0.57 0.47 -
Occipital
ctx-rh-lateraloccipital_3 27 -94 -6 0.39 0.34# -
ctx-lh-lingual_2 -10 -70 0 0.42 0.50 �
ctx-rh-lingual_1 14 -83 -7 0.41 0.37 -
Negatively correlated
Frontal
ctx-lh-parstriangularis_1 -43 33 2 -0.50 -0.43 �
ctx-lh-rostralmiddlefrontal_5 -31 56 2 -0.57 -0.50 �
ctx-rh-precentral_6 22 -17 67 -0.39 -0.35# �
Parietal
ctx-lh-precuneus_2 -10 -44 46 -0.46 -0.43 �
ctx-lh-superiorparietal_6 -22 -76 29 -0.41 -0.47 �
ctx-rh-precuneus_2 12 -54 26 -0.39 -0.43 �
Occipital
ctx-rh-pericalcarine_1 15 -75 12 -0.375 -0.24^ �
* Rho after controlling for disease severity; # p < 0.1; ^ p > 0.1. ctx-lh = left hemisphere; ctx-rh = right hemisphere

169
Figure 5) Significant changes in between- and within-module scores associated with the HSS. (A) Nodes
with a significant correlation between Bi scores and the HSS; (B) Nodes with a significant correlation
between the Wi scores and HSS. Green indicates a positive correlation, red indicates a negative
correlation. Figure visualized with BrainNet Viewer (Xia et al., 2013).
Discussion
The aim of this study was to determine whether changes in structural network topology
were associated with hallucinatory behaviour in PD. We showed that severity of
hallucinatory behaviour was negatively correlated with connectivity across a bilateral
sub-network. Regions within this sub-network showed higher participation compared to
regions outside this network. The loss of connectivity strength may force the system to
adapt and reroute information across less efficient pathways, impeding the standard
sensory integration process. Importantly, 94% of the nodes in the diverse club were
included in this sub-network. This community of high participation nodes is thought to
control the integration of relatively segregated regions (Bertolero et al., 2017). Indeed,
the diverse connectivity pattern of these nodes makes them crucial for the functional
coordination of brain regions during tasks, and activity in these nodes predicts changes in
A
B

170
the coupling of other regions (Bertolero et al., 2017). Severity of hallucinatory behaviour
may thus be the result of impaired integration and segregation of brain networks or
‘modules’, affecting effective information transfer. Finally, we showed regional changes
in participation associated with hallucination severity (the HSS score), with a positive
correlation between participation scores in the medial OFC, cingulate, precuneus and
middle frontal gyrus and the HSS and negative correlation with participation scores in the
lateral occipital cortex, pars orbitalis and insula. These findings suggest a reweighting of
the regions along the perceptual hierarchy, which may give rise to hallucinations.
Lower participation of the lateral occipital cortex may reflect reduced early visual
processing, resulting in ineffective accumulation of visual information from the
environment. Previous work using a Bayesian drift diffusion model has demonstrated that
accumulation speed and quality of perceptual information are reduced in PD patients with
VH (O’Callaghan et al., 2017). Furthermore, reduced quality or integration of visual
stimuli may increase perceptual uncertainty, a suggestion that aligns with increased
participation in the dorsal anterior cingulate cortex (Keri, Decety, Roland, & Gulyas,
2004). Perceptual uncertainty may place excessive emphasis on top-down prediction
centres, which subsequently could lead to a reduced activity in early visual regions
(Meyer & Olson, 2011). However, the reduced participation score in the occipital and
anterior cingulate cortex may not be specific to hallucinatory behaviour, as the negative
correlations did not remain significant after controlling for disease severity.
This emphasis on top-down visual processing centres is supported by the increased
participation coefficient and module degree z-score in the medial OFC. The OFC has an
integrative function across brain networks as evidenced by its high participation
coefficient. Additionally, this region is thought to facilitates recognition during visual
perception by integrating incoming sensory information with previous experiences and
expectations (Panichello, Cheung, & Bar, 2012). During typical visual perception, the
OFC is activated early in response to visual stimuli, receiving low spatial frequency
signals from the visual cortex (Bar et al., 2006). Notably, only stimuli resembling known
objects are shown to activate the OFC, which in turn generates a semantic association and

171
provides a predictive signal to the visual system (Chaumon, Kveraga, Barrett, & Bar,
2014). Conversely, visual stimuli that carry no meaningful association do not activate the
OFC in healthy individuals. Hence, it could be speculated that due to decreased quality of
visual input, inappropriate recruitment of the OFC occurs, which may result in ascribing
false associative information to visual stimuli.
The manifestation of VHs has previously been recognized as a dysfunction between the
attentional networks (Shine, Muller, et al., 2015). Specifically, patients with VHs are
shown to be less able to recruit the dorsal attentional network (DAN), which enables the
selection of appropriate sensory stimuli (Maurizio Corbetta, Patel, & Shulman, 2008).
With reduced control of this network, ambiguous stimuli might instead be interpreted by
the ventral attentional network (VAN) and the DMN, which are less well equipped for
this task. Our results showed increased participation in the posterior cingulate cortex
(PCC), a key hub of the DMN. PCC activity has been implicated in regulating the focus
of attention, specifically, the shift from the external world into internal mentation
(Weissman, Roberts, Visscher, & Woldorff, 2006). Furthermore, the PCC is involved in
mind wandering and supports internally directed cognition (Leech & Sharp, 2013). A
failure to suppress PCC activity may lead to the intrusion of internal thoughts into task
performance (Sonuga-Barke & Castellanos, 2007). Moreover, a positive correlation was
found between the HSS and the module degree z-score of the left pars opercularis, a node
in the VAN, a network that is activated when expectations in perception are violated
(Corbetta & Shulman, 2002; Shine, Halliday, et al., 2014). Conversely, a negative
correlation between the HSS scores and module degree score and participation
coefficients was found in other nodes of the VAN, namely the left pars triangularis, the
right pars orbitalis and insula. The left pars triangularis supports resolving competition
between simultaneously active representations (Badre & Wagner, 2007), whilst the insula
plays an important role in dynamically shifting attention between the attentional control
networks (Menon & Uddin, 2010). The anterior insula has previously been shown to be
reduced in volume in PD patients with VH (Ibarretxe-Bilbao et al., 2010; Shine, Halliday,
et al., 2014). Together, these results suggest that ineffective communication between
attentional networks in the brain may predispose an individual to hallucinate.

172
Surprisingly, the participation coefficient of a node within the DAN (‘ctx-rh-
caudalmiddlefrontal_2’) showed a positive correlation with the HSS. This node was not
part of the sub-network and it may be possible that this is a compensatory response to the
loss of connectivity strength in the other DAN regions. Notably, the connectivity matrix
shows between module connections of this region with nodes in the somatomotor and the
frontoparietal network, but not with the DMN or VAN.
Finally, all nodes that showed negative correlations with the HSS were included in the
sub-network. Decreased within-module scores were found across the prefrontal and the
somatosensory association cortex, as well as in the primary visual cortex, whilst there
was a positive correlation between the HSS and the bilateral secondary visual cortex,
perhaps as a result of the decreased visual input from V1. Additionally, the supramarginal
gyrus, a node that has been shown to be functionally active during spatial perception but
also during visual imagery (Ganis, Thompson, & Kosslyn, 2004) showed an increased
module degree z-score with increasing severity of VHs. Furthermore, a positive
correlation with the HSS and the module degree score in the superior temporal cortex, a
region involved in auditory processing was found. It could be speculated that increased
visual uncertainty may stimulate other sensory processing areas. Conversely, previous
work in schizophrenia has shown increased activation in the superior temporal cortex
during a mis-match between expected and perceived auditory feedback (Fu et al., 2008).
The HSS also included hallucinations of other sensory modalities and it could be
speculated that a greater emphasize is being placed on predictions of auditory stimuli, as
reflected as increased within-module connectivity in the superior temporal cortex.
This study has several limitations worth noting. Firstly, the DWI data was acquired
without EPI distortion correction. This may have affected the accuracy of registration
between DWI and T1 images in the frontal and temporal cortices. Due to relatively low
diffusion weighting used in the current MRI protocol, it was chosen to employ DTI rather
than more sophisticated methods such as constrained spherical deconvolution, an
algorithm that more adequately deals with multiple fibre directions within one voxel than
DTI. Furthermore, after controlling for disease severity, the identified sub-network was

173
only trending towards significance. The correlation between the HSS and the
participation coefficients of the lateral occipital and anterior cingulate did not remain
significant, whilst the OFC and insula were only trending towards significance, and
similar patterns were found for the module degree z-score: correlation with the HSS and
nodes in the occipital and precentral regions did not remain significant. However, this
may be expected given the close relationship between overall symptom severity and
phenotypic symptom severity. Importantly, the correlation coefficients showed notable
consistency across the analyses, suggesting the HSS was able to distinguish inter-
individual variability. Whilst the current study included PD patients with no self-reported
hallucinations (n=6), no data from a healthy control group was acquired, which could
have aided in disassociating the significant network measures and disease burden. The
LEDD score was not associated with the HSS in the current study, and the relation
between hallucinations and a LEDD-related side effect has been dismissed in previous
work (Fénelon et al., 2000; Graham et al., 1997; Sanchez-Ramos et al., 1996; Shergill et
al., 1998). Furthermore, hallucinations may occur in the absence of dopaminergic
treatment in PD or Dementia with Lewy Bodies (Ala et al., 1997). Therefore, the current
study chose not to include LEDD score as a covariate. Furthermore, the investigation was
conducted in a relatively small group of PD patients and future studies should replicate
our findings in a larger sample size. Finally, this study chose to include the PsycH-Q to
assess hallucinations. This is an in-depth questionnaire with high construct validity,
however, patients have to report their hallucinations themselves. Ideally, a trained
neurologist or qualified researcher, who can probe the patients when in doubt, should
assist with this process.
Conclusions
We cautiously conclude that hallucinatory behaviour in PD patients is associated with
marked alterations in structural network topology. Severity of hallucinatory behaviour
was associated with decreased connectivity in a large sub-network that included the
majority of the diverse club. These changes may result in an inefficient rerouting of
information across less efficient pathways, which may lead to impaired visual integration
processes. Furthermore, nodes within the orbitofrontal cortex and temporal lobes showed

174
increased participation scores, whilst the visual association cortex, insula and middle
frontal gyrus showed decreased scores associated with the HSS score. These findings
suggest that impaired integration across different regions along the perceptual hierarchy
may result in inefficient transfer of information. A failure to effectively switch between
attentional networks and the intrusion of internal percepts could give rise to perceptual
glitches, such as misperceptions and hallucinations.
Acknowledgements
We thank the patients and their families who contribute to our research at the Parkinson’s
Disease Research Clinic. We thank Dr Váša for sharing his thresholding code
(github.com/frantisekvasa/matlab_general). The DWI data was processed during the
10kin1day initiative at the Dutch Connectome Lab. This research was supported by
Sydney Informatics Hub, funded by the University of Sydney. JMH is supported by a
Western Sydney University Postgraduate Research Award; CO is supported by a National
Health and Medical Research Council Neil Hamilton Fairley Fellowship (1091310) and
by the Wellcome Trust (200181/Z/15/Z). AJM is supported by an Australian
Postgraduate Award at the University of Sydney; KAEM is supported by a University of
Sydney Postdoctoral Fellowship; JRP is supported by an Australian Postgraduate Award
at the Western Sydney University. SJGL is supported by National Health and Medical
Research Council-Australian Research Council Dementia Fellowship (#1110414) and
this work was supported by funding to Forefront, a collaborative research group
dedicated to the study of non-Alzheimer disease degenerative dementias, from the
National Health and Medical Research Council of Australia program grant (#1037746
and #1095127). JMS is supported by a National Health and Medical Research Council CJ
Martin Fellowship (1072403). AAM has no funding source to disclose.

175
References
Aarsland, D., Bronnick, K., Ehrt, U., De Deyn, P. P., Tekin, S., Emre, M., & Cummings,
J. L. (2007). Neuropsychiatric symptoms in patients with Parkinson's disease and
dementia: frequency, profile and associated care giver stress. J Neurol Neurosurg
Psychiatry, 78(1), 36-42.
Ala, T. A., Yang, K. H., Sung, J. H., & Frey, W. H. (1997). Hallucinations and signs of
parkinsonism help distinguish patients with dementia and cortical Lewy bodies
from patients with Alzheimer's disease at presentation: a clinicopathological
study. Journal of Neurology, Neurosurgery & Psychiatry, 62(1), 16-21.
doi:10.1136/jnnp.2005.083113
Badre, D., & Wagner, A. D. (2007). Left ventrolateral prefrontal cortex and the cognitive
control of memory. Neuropsychologia, 45(13), 2883-2901.
doi:https://doi.org/10.1016/j.neuropsychologia.2007.06.015
Bar, M. (2009). The proactive braerin: memory for predictions. Philosophical
Transactions of the Royal Society B: Biological Sciences, 364(1521), 1235-1243.
doi:10.1098/rstb.2008.0310
Bar, M., Kassam, K. S., Ghuman, A. S., Boshyan, J., Schmid, A. M., Dale, A. M., . . .
Halgren, E. (2006). Top-down facilitation of visual recognition. Proc Natl Acad
Sci U S A, 103(2), 449-454. doi:10.1073/pnas.0507062103
Barnes, J., Boubert, L., Harris, J., Lee, A., & David, A. S. (2003). Reality monitoring and
visual hallucinations in Parkinson's disease. Neuropsychologia, 41(5), 565-574.
Barnes, J., & David, A. (2001). Visual hallucinations in Parkinson's disease: a review and
phenomenological survey. Journal of Neurology, Neurosurgery & Psychiatry,
70(6), 727-733.
Bastiani, M., Shah, N. J., Goebel, R., & Roebroeck, A. (2012). Human cortical
connectome reconstruction from diffusion weighted MRI: The effect of
tractography algorithm. Neuroimage, 62(3), 1732-1749.
doi:https://doi.org/10.1016/j.neuroimage.2012.06.002
Bertolero, M. A., Yeo, B. T. T., & D'Esposito, M. (2017). The diverse club. Nat
Commun, 8(1), 1277. doi:10.1038/s41467-017-01189-w

176
Bertolero, M. A., Yeo, B. T. T., & D’Esposito, M. (2015). The modular and integrative
functional architecture of the human brain. Proceedings of the National Academy
of Sciences, 112(49), E6798-E6807. doi:10.1073/pnas.1510619112
Bullmore, E., & Sporns, O. (2009). Complex brain networks: graph theoretical analysis
of structural and functional systems. Nature Reviews Neuroscience, 10(3), 186.
Chang, L. C., Jones, D. K., & Pierpaoli, C. (2005). RESTORE: robust estimation of
tensors by outlier rejection. Magn Reson Med, 53(5), 1088-1095.
doi:10.1002/mrm.20426
Chaumon, M., Kveraga, K., Barrett, L. F., & Bar, M. (2014). Visual predictions in the
orbitofrontal cortex rely on associative content. Cereb Cortex, 24(11), 2899-2907.
doi:10.1093/cercor/bht146
Collerton, D., Perry, E., & McKeith, I. (2005). Why people see things that are not there: a
novel Perception and Attention Deficit model for recurrent complex visual
hallucinations. Behav Brain Sci, 28(6), 737-757; discussion 757-794.
doi:10.1017/S0140525X05000130
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The Reorienting System of the Human
Brain: From Environment to Theory of Mind. Neuron, 58(3), 306-324.
doi:10.1016/j.neuron.2008.04.017
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven
attention in the brain. Nat Rev Neurosci, 3(3), 201-215. doi:10.1038/nrn755
de Reus, M. A., & van den Heuvel, M. P. (2013). Estimating false positives and negatives
in brain networks. Neuroimage, 70, 402-409.
doi:10.1016/j.neuroimage.2012.12.066
Diederich, N. J., Goetz, C. G., Raman, R., Pappert, E. J., Leurgans, S., & Piery, V.
(1998). Poor visual discrimination and visual hallucinations in Parkinson's
disease. Clin Neuropharmacol, 21(5), 289-295.
Diederich, N. J., Goetz, C. G., & Stebbins, G. T. (2005). Repeated visual hallucinations
in Parkinson's disease as disturbed external/internal perceptions: focused review
and a new integrative model. Mov Disord, 20(2), 130-140.
doi:10.1002/mds.20308

177
Engel, A. K., Fries, P., & Singer, W. (2001). Dynamic predictions: Oscillations and
synchrony in top–down processing. Nature Reviews Neuroscience, 2, 704.
doi:10.1038/35094565
Fénelon, G., Mahieux, F., Huon, R., & Ziégler, M. (2000). Hallucinations in Parkinson's
disease: prevalence, phenomenology and risk factors. Brain, 123(4), 733-745.
Field, A. (2009). Discovering Statistics Using SPSS. London, UK: SAGE Publications
Ltd.
Fletcher, P. C., & Frith, C. D. (2008). Perceiving is believing: a Bayesian approach to
explaining the positive symptoms of schizophrenia. Nature Reviews
Neuroscience, 10, 48. doi:10.1038/nrn2536
Folstein, M. F., Robins, L. N., & Helzer, J. E. (1983). The mini-mental state examination.
Archives of general psychiatry, 40(7), 812-812.
Fu, C. H. Y., Brammer, M. J., Yágüez, L., Allen, P., Matsumoto, K., Johns, L., . . . van
Haren, N. (2008). Increased superior temporal activation associated with external
misattributions of self-generated speech in schizophrenia. Schizophrenia
Research, 100(1), 361-363. doi:https://doi.org/10.1016/j.schres.2007.10.023
Gallagher, D. A., Parkkinen, L., O'sullivan, S. S., Spratt, A., Shah, A., Davey, C. C., . . .
Lees, A. J. (2011). Testing an aetiological model of visual hallucinations in
Parkinson's disease. Brain, 134(11), 3299-3309.
Ganis, G., Thompson, W. L., & Kosslyn, S. M. (2004). Brain areas underlying visual
mental imagery and visual perception: an fMRI study. Cognitive Brain Research,
20(2), 226-241. doi:https://doi.org/10.1016/j.cogbrainres.2004.02.012
Goetz, C. G. (2009). Scales to evaluate psychosis in Parkinson's disease. Parkinsonism
Relat Disord, 15 Suppl 3, S38-41. doi:10.1016/S1353-8020(09)70777-1
Goetz, C. G., Tilley, B. C., Shaftman, S. R., Stebbins, G. T., Fahn, S., Martinez-Martin,
P., . . . Dodel, R. (2008). Movement Disorder Society-sponsored revision of the
Unified Parkinson's Disease Rating Scale (MDS-UPDRS): scale presentation and
clinimetric testing results. Movement disorders: official journal of the Movement
Disorder Society, 23(15), 2129-2170.

178
Graham, J. M., Grünewald, R. A., & Sagar, H. J. (1997). Hallucinosis in idiopathic
Parkinson’s disease. Journal of Neurology, Neurosurgery & Psychiatry, 63(4),
434-440.
Guimerà, R., & Nunes Amaral, L. A. (2005). Functional cartography of complex
metabolic networks. Nature, 433, 895. doi:10.1038/nature03288
Hall, J. M., O'Callaghan, C., Shine, J. M., Muller, A. J., Phillips, J. R., Walton, C. C., . . .
Moustafa, A. A. (2016). Dysfunction in attentional processing in patients with
Parkinson's disease and visual hallucinations. J Neural Transm (Vienna), 123(5),
503-507. doi:10.1007/s00702-016-1528-3
Hepp, D. H., Foncke, E. M. J., Berendse, H. W., Wassenaar, T. M., Dubbelink, K. T. E.
O., Groenewegen, H. J., . . . Schoonheim, M. M. (2017). Damaged fiber tracts of
the nucleus basalis of Meynert in Parkinson’s disease patients with visual
hallucinations. Scientific Reports, 7(1), 10112.
Hepp, D. H., Foncke, E. M. J., Dubbelink, K. T. E. O., Berg, W. D. J. v. d., Berendse, H.
W., & Schoonheim, M. M. (2017). Loss of Functional Connectivity in Patients
with Parkinson Disease and Visual Hallucinations. Radiology, 285(3), 896-903.
doi:10.1148/radiol.2017170438
Hoehn, M. M., & Yahr, M. D. (1998). Parkinsonism: onset, progression, and mortality.
Neurology, 50(2), 318-318.
Ibarretxe-Bilbao, N., Ramirez-Ruiz, B., Junque, C., Marti, M. J., Valldeoriola, F.,
Bargallo, N., . . . Tolosa, E. (2010). Differential progression of brain atrophy in
Parkinson's disease with and without visual hallucinations. J Neurol Neurosurg
Psychiatry, 81(6), 650-657. doi:10.1136/jnnp.2009.179655
Intaite, M., Noreika, V., Soliunas, A., & Falter, C. M. (2013). Interaction of bottom-up
and top-down processes in the perception of ambiguous figures. Vision Res, 89,
24-31. doi:10.1016/j.visres.2013.06.011
Keri, S., Decety, J., Roland, P. E., & Gulyas, B. (2004). Feature uncertainty activates
anterior cingulate cortex. Hum Brain Mapp, 21(1), 26-33. doi:10.1002/hbm.10150
Lee, J.-Y., Yoon, E. J., Lee, W. W., Kim, Y. K., Lee, J.-Y., & Jeon, B. (2016). Lateral
geniculate atrophy in Parkinson's with visual hallucination: A trans-synaptic
degeneration? Movement Disorders, 31(4), 547-554. doi:10.1002/mds.26533

179
Leech, R., & Sharp, D. J. (2013). The role of the posterior cingulate cortex in cognition
and disease. Brain, 137(1), 12-32.
Lenka, A., Hegde, S., Arumugham, S. S., & Pal, P. K. (2017). Pattern of cognitive
impairment in patients with Parkinson's disease and psychosis: A critical review.
Parkinsonism & Related Disorders, 37, 11-18.
doi:https://doi.org/10.1016/j.parkreldis.2016.12.025
Matsui, H., Udaka, F., Tamura, A., Oda, M., Kubori, T., Nishinaka, K., & Kameyama, M.
(2006). Impaired visual acuity as a risk factor for visual hallucinations in
Parkinson's disease. J Geriatr Psychiatry Neurol, 19(1), 36-40.
doi:10.1177/0891988705284739
Menon, V., & Uddin, L. Q. (2010). Saliency, switching, attention and control: a network
model of insula function. Brain Struct Funct, 214(5-6), 655-667.
doi:10.1007/s00429-010-0262-0
Meyer, T., & Olson, C. R. (2011). Statistical learning of visual transitions in monkey
inferotemporal cortex. Proceedings of the National Academy of Sciences, 108(48),
19401-19406. doi:10.1073/pnas.1112895108
Misic, B., Betzel, R. F., Nematzadeh, A., Goni, J., Griffa, A., Hagmann, P., . . . Sporns,
O. (2015). Cooperative and Competitive Spreading Dynamics on the Human
Connectome. Neuron, 86(6), 1518-1529. doi:10.1016/j.neuron.2015.05.035
Mori, S., Crain, B. J., Chacko, V. P., & van Zijl, P. C. (1999). Three-dimensional
tracking of axonal projections in the brain by magnetic resonance imaging. Ann
Neurol, 45(2), 265-269.
Muller, A. J., O'Callaghan, C., Walton, C. C., Shine, J. M., & Lewis, S. J. (2017).
Retrospective Neuropsychological Profile of Patients With Parkinson Disease
Prior to Developing Visual Hallucinations. J Geriatr Psychiatry Neurol, 30(2),
90-95. doi:10.1177/0891988716686830
Muller, A. J., Shine, J. M., Halliday, G. M., & Lewis, S. J. (2014). Visual hallucinations
in Parkinson's disease: theoretical models. Movement Disorders, 29(13), 1591-
1598.
Nichols, T. E., & Holmes, A. P. (2002). Nonparametric permutation tests for functional
neuroimaging: a primer with examples. Human brain mapping, 15(1), 1-25.

180
O’Callaghan, C., Hall, J. M., Tomassini, A., Muller, A. J., Walpola, I. C., Moustafa, A.
A., . . . Lewis, S. J. G. (2017). Visual Hallucinations Are Characterized by
Impaired Sensory Evidence Accumulation: Insights From Hierarchical Drift
Diffusion Modeling in Parkinson’s Disease. Biological Psychiatry: Cognitive
Neuroscience and Neuroimaging, 2(8), 680-688.
doi:https://doi.org/10.1016/j.bpsc.2017.04.007
Panichello, M. F., Cheung, O. S., & Bar, M. (2012). Predictive Feedback and Conscious
Visual Experience. Frontiers in Psychology, 3, 620.
doi:10.3389/fpsyg.2012.00620
Powers, A. R., Kelley, M., & Corlett, P. R. (2016). Hallucinations as Top-Down Effects
on Perception. Biological Psychiatry: Cognitive Neuroscience and Neuroimaging,
1(5), 393-400. doi:https://doi.org/10.1016/j.bpsc.2016.04.003
Ramirez-Ruiz, B., Junque, C., Marti, M. J., Valldeoriola, F., & Tolosa, E. (2006).
Neuropsychological deficits in Parkinson's disease patients with visual
hallucinations. Mov Disord, 21(9), 1483-1487. doi:10.1002/mds.20965
Ramírez-Ruiz, B., Martí, M. J., Tolosa, E., Falcón, C., Bargalló, N., Valldeoriola, F., &
Junqué, C. (2008). Brain response to complex visual stimuli in Parkinson's
patients with hallucinations: a functional magnetic resonance imaging study.
Movement Disorders, 23(16), 2335-2343.
Rubinov, M., & Sporns, O. (2010). Complex network measures of brain connectivity:
uses and interpretations. Neuroimage, 52(3), 1059-1069.
doi:10.1016/j.neuroimage.2009.10.003
Sanchez-Ramos, J. R., Ortollm, R., & Paulson, G., W. (1996). Visual hallucinations
associated with Parkinson disease. Archives of Neurology, 53(23), 1265-1268.
Schrag, A. (2004). Psychiatric aspects of Parkinson's disease--an update. J Neurol,
251(7), 795-804. doi:10.1007/s00415-004-0483-3
Shergill, S., S., Walker, Z., Le Katona, C. (1998). A preliminary investigation of
laterality in Parkinson’s disease and susceptibility to psychosis. Journal of
Neurology, Neurosurgery & Psychiatry, 65(4), 610-611.

181
Shine, J. M., Halliday, G. H., Carlos, M., Naismith, S. L., & Lewis, S. J. (2012).
Investigating visual misperceptions in Parkinson's disease: a novel behavioral
paradigm. Movement Disorders, 27(4), 500-505.
Shine, J. M., Halliday, G. M., Gilat, M., Matar, E., Bolitho, S. J., Carlos, M., . . . Lewis,
S. J. (2014). The role of dysfunctional attentional control networks in visual
misperceptions in Parkinson's disease. Hum Brain Mapp, 35(5), 2206-2219.
doi:10.1002/hbm.22321
Shine, J. M., Mills, J. M. Z., Qiu, J., O'Callaghan, C., Terpening, Z., Halliday, G. M., . . .
Lewis, S. J. G. (2015). Validation of the Psychosis and Hallucinations
Questionnaire in Non-demented Patients with Parkinson's Disease. Movement
Disorders Clinical Practice, 2(2), 175-181. doi:10.1002/mdc3.12139
Shine, J. M., Muller, A. J., O'Callaghan, C., Hornberger, M., Halliday, G. M., & Lewis,
S. J. (2015). Abnormal connectivity between the default mode and the visual
system underlies the manifestation of visual hallucinations in Parkinson's disease:
a task-based fMRI study. NPJ Parkinsons Dis, 1, 15003.
doi:10.1038/npjparkd.2015.3
Shine, J. M., O'Callaghan, C., Halliday, G. M., & Lewis, S. J. (2014). Tricks of the mind:
Visual hallucinations as disorders of attention. Prog Neurobiol, 116, 58-65.
doi:10.1016/j.pneurobio.2014.01.004
Smith, R. E., Tournier, J. D., Calamante, F., & Connelly, A. (2012). Anatomically-
constrained tractography: improved diffusion MRI streamlines tractography
through effective use of anatomical information. Neuroimage, 62(3), 1924-1938.
doi:10.1016/j.neuroimage.2012.06.005
Sonuga-Barke, E. J. S., & Castellanos, F. X. (2007). Spontaneous attentional fluctuations
in impaired states and pathological conditions: A neurobiological hypothesis.
Neuroscience & Biobehavioral Reviews, 31(7), 977-986.
doi:https://doi.org/10.1016/j.neubiorev.2007.02.005
Summerfield, C., Egner, T., Greene, M., Koechlin, E., Mangels, J., & Hirsch, J. (2006).
Predictive Codes for Forthcoming Perception in the Frontal Cortex. Science,
314(5803), 1311-1314. doi:10.1126/science.1132028

182
Summerfield, C., & Koechlin, E. (2008). A Neural Representation of Prior Information
during Perceptual Inference. Neuron, 59(2), 336-347.
doi:https://doi.org/10.1016/j.neuron.2008.05.021
Sun, Z., Wang, F., Cui, L., Breeze, J., Du, X., Wang, X., . . . Zhang, D. (2003). Abnormal
anterior cingulum in patients with schizophrenia: a diffusion tensor imaging
study. Neuroreport, 14(14), 1833-1836.
doi:10.1097/01.wnr.0000094529.75712.48
Tombaugh, T. N. (2004). Trail Making Test A and B: normative data stratified by age
and education. Archives of clinical neuropsychology, 19(2), 203-214.
Turner, T. H., Cookson, J. C., Wass, J. A., Drury, P. L., Price, P. A., Besser, G. M.
(1984). Psychotic reactions during treatment of pituitary tumours with dopamine
agonists. British Medical Journal (Clin Res Ed), 289(6452), 1101-1103.
van den Heuvel, M. P., & Sporns, O. (2011). Rich-club organization of the human
connectome. J Neurosci, 31(44), 15775-15786. doi:10.1523/JNEUROSCI.3539-
11.2011
Verstraete, E., van den Heuvel, M. P., Veldink, J. H., Blanken, N., Mandl, R. C.,
Hulshoff Pol, H. E., & van den Berg, L. H. (2010). Motor network degeneration
in amyotrophic lateral sclerosis: a structural and functional connectivity study.
PLoS One, 5(10), e13664. doi:10.1371/journal.pone.0013664
Weissman, D. H., Roberts, K., Visscher, K., & Woldorff, M. (2006). The neural bases of
momentary lapses in attention. Nature neuroscience, 9(7), 971.
Yao, N., Shek-Kwan Chang, R., Cheung, C., Pang, S., Lau, K. K., Suckling, J., . . . Chua,
S. E. (2014). The default mode network is disrupted in Parkinson's disease with
visual hallucinations. Human brain mapping, 35(11), 5658-5666.
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D., Hollinshead, M., .
. . Buckner, R. L. (2011). The organization of the human cerebral cortex estimated
by intrinsic functional connectivity. J Neurophysiol, 106(3), 1125-1165.
doi:10.1152/jn.00338.2011
Zalesky, A., Fornito, A., & Bullmore, E. T. (2010). Network-based statistic: identifying
differences in brain networks. Neuroimage, 53(4), 1197-1207.
doi:10.1016/j.neuroimage.2010.06.041

183
Zalesky, A., Fornito, A., Cocchi, L., Gollo, L. L., van den Heuvel, M. P., & Breakspear,
M. (2016). Connectome sensitivity or specificity: which is more important?
Neuroimage, 142, 407-420. doi:10.1016/j.neuroimage.2016.06.035

184
Supplementary Materials Supplementary Table 1. Nodes of the diverse club Node Participation coefficient Inclusion sub-network
'Left-Putamen' 0.814 ✓
'Right-Pallidum' 0.811 ✓
'Right-Putamen' 0.808 ✓
'Left-Pallidum' 0.807 ✓
'Right-Thalamus-Proper' 0.805 ✓
'Right-Caudate' 0.768 ✓
'ctx-rh-precuneus_4' 0.734 ✓
'ctx-lh-superiorparietal_5' 0.731 ✓
'ctx-lh-superiorparietal_4' 0.724 ✓
'ctx-rh-superiorfrontal_5' 0.721 ✓
'ctx-rh-isthmuscingulate_1' 0.716 ✓
'Left-Hippocampus' 0.713 ✓
'ctx-lh-insula_3' 0.705 ✓
'ctx-rh-rostralmiddlefrontal_5' 0.695 ✓
'ctx-lh-superiorfrontal_3' 0.664 ✓
'ctx-rh-superiorparietal_3' 0.658 ✓
'ctx-lh-lateraloccipital_1' 0.653 ✓
'ctx-rh-postcentral_5’ 0.612 -
ctx-lh = left hemisphere; ctx-rh = right hemisphere

185
Psychosis and Hallucinations Questionnaire, part A

186

187
Supplementary figure 1. Mean connectivity matrix (thresholded) Connectivity matrix is deposited at github.com/juliemaehall
Supplementary figure 2. Size of sub-networks correlated with the HSS for different t-statistics. Blue bars represent the number of edges; white bars represent the number of nodes. Asterisks (*) represent a significant correlation (p < 0.05).
*
* *
* *
0
50
100
150
200
250
300
1.5 1.6 1.7 1.8 1.9 2
Num
ber o
f nod
es /
edge
s
t-statistic
Number of edges in sub-network correlated with the HSS for different t-statistic thresholds

CHAPTER7
GENERALDISCUSSION

188
7. General discussion
The preceding Chapters in this thesis aimed to outline how new insights into the
heterogeneous character could be gained by considering Parkinson’s disease as a
‘disconnection disorder’. Specifically, this thesis has shown that two heretofore
poorly understood symptoms of Parkinson’s disease, namely freezing of gait and
visual hallucinations, could be recognized as a failure of effective communication
within and between large-scale neuronal networks. The specific aims of this thesis
were as follows: 1) to provide an overview of the current literature on white matter
changes associated with different symptoms of Parkinson’s disease using DTI
(Chapter 2), 2) To investigate whether freezing of gait could be result from
ineffective communication between widespread brain networks (Chapter 3 and
Chapter 4) and 3) to investigate whether visual hallucinations in Parkinson’s disease
arise due to a reweighting of attentional and perceptual brain networks (Chapter 5
and Chapter 6). The key findings of each chapter are summarized chiefly in their
respective discussion sections. However, these points are briefly outlined again
below in relation to how they meet the initial aims of the thesis.
7.1 Summary of key findings
7.1.1 To provide an overview of the current literature on white matter changes associated
with different symptoms of Parkinson’s disease using DTI (Chapter 2)
The first aim of this thesis was to provide an in depth overview of the current
literature investigating diffusion alterations associated with specific symptoms of
Parkinson’s disease. Chapter 2 demonstrated that distinct symptoms have unique
neurobiological patterns and thus emphasised the importance of investigating
homogeneous patient groups with Parkinson’s disease to avoid confounding results.
This review further highlighted that changes in white matter associated with several
symptoms, including visual hallucinations and freezing of gait, but also REM sleep
behaviour disorders and mood disturbances, remain poorly understood, either due to
a lack of research into these phenomena or because of conflicting results. One
explanation for this variability can be found in the different MRI protocols used

189
across studies. However, it is also possible that symptoms of Parkinson’s disease are
not the result of a focalised lesion, but rather due to changes in network structure.
These changes might go unexposed when using traditional methods such as TBSS or
tractography. Instead, assessing whole brain connectivity using a data driven
approach may reveal complex, distributed patterns in white matter tracts. Rather than
trumpeting one region of a circuit as responsible for a specific function, this
approach allows for the inherent complexity of interacting subsystems and their
connections to be studied and therefore may provide unique insights into white
matter pathology in Parkinson’s disease.
The findings of the two symptoms central to this dissertation, freezing of gait and
visual hallucinations, will be briefly discussed in the next section.
Freezing of gait
The review presented in Chapter 2 showed that studies investigating freezing of gait
in Parkinson’s disease have focused predominantly on the brainstem locomotor
regions and reported white matter abnormalities in the corticopontine and pontine-
cerebellar tracts. Cholinergic pedunculopontine nucleus projections regulate gait
initiation, continuation and modulation in healthy adults (Grabli et al., 2012;
Rochester et al., 2012). Furthermore, increasing evidence suggests that the
pedunculopontine nucleus underlies an integrative function of distinct functional
neuronal networks; hence disruptions in its ascending pathway could have
implications beyond motor control (Snijders et al., 2016). Diffusivity changes were
also found in major cortico-cortical and several frontostriatal tracts, highlighting the
role of extensive cortical and subcortical changes. Finally, a recent study not
included in the review chapter showed that diffusivity changes in long-range
associative fibres that are involved in integration between left and right frontal lobes
as well as intra-hemispheric connections between associative cortical areas (Sara
Pietracupa et al., 2018). These findings are consistent with behavioural studies
showing frontal impairment in patients with freezing (de Souza Fortaleza et al.,
2017; Hall et al., 2014; Naismith et al., 2010; Vastik et al., 2017; Walton et al.,

190
2015), as well as functional connectivity abnormalities reported in the basal ganglia
and frontal and parietal cortices (Mi et al., 2017; Shine et al., 2011b; Tessitore et al.,
2012). Deficits in complimentary yet competing pathways between motor, cognitive
and limbic structures may contribute to freezing behaviour.
Visual hallucinations
At the time the search for this literature review was performed, only one study had
been performed on diffusivity alterations underlying visual hallucinations. This study
observed diffusivity changes in the hippocampus (Yao et al., 2014), a node that
strongly projects to the default mode network (Lee & Xue, 2018). Since this review
was published, two other studies have been performed that investigated white matter
changes in Parkinson’s disease patients with visual hallucinations. Firstly, a study
reported diffusivity changes in the optic nerve and optic radiation, which suggest that
both pre- and postsynaptic fibres of the visual afferent neurons undergo degeneration
in hallucinators (Lee et al., 2016). Secondly, changes in diffusivity were observed in
tracts connecting the nucleus basalis of Meynert with parietal and occipital cortical
regions and the authors suggested that this might be due to a loss of cholinergic
innervation (Hepp et al., 2017). This hypothesis is supported by the tendency of
anticholinergic drugs to induce visual hallucinations (Bullock et al., 2002; Burn et al.,
2006; Perry et al., 1995). Taken together, these studies demonstrate the likelihood of
deficits in primary visual regions, as well as in regions associated with cognitive
performance (Gratwicke et al., 2018; Squire et al., 1991) in patients with visual
hallucinations.
Studies investigating the white matter integrity associated with freezing and
hallucinations have aided our understanding of these phenomena. Evidently, there are
some discrepancies in the presented data. Studies have often targeted different
regions of interest and used a differing array of methods (i.e. TBSS or tractography).
Whilst tractography focuses on a specific tract rather than on the whole connectome,
TBSS may not be sensitive enough to reveal subtle changes in connectivity strength.
Graph theory allows for the measurement of global and regional changes in the

191
whole brain connectome without a predefined region of interest. By studying the
structural connectome, subtle changes in the organisation of brain networks may
become evident.
7.1.2. To investigate whether freezing of gait could be a result to ineffective
communication between brain networks (Chapter 3 and Chapter 4)
This thesis has made an important contribution to the understanding of the
phenotypical associations across disease stages (Chapter 3), as well as to the
neurobiological mechanisms (Chapter 4) of freezing of gait.
Consistent with previous work (Naismith et al., 2010; Walton et al., 2015a; 2015b),
results presented in Chapter 3 showed a significant association between executive
dysfunction and freezing of gait, suggesting the involvement of frontostriatal
networks (Shine et al., 2011b). This study extended previous work by mapping out a
number of different phenotypical features in a large patient cohort across different
disease stages. The results showed that freezing of gait was associated with the non-
tremor dominant motor phenotype. Furthermore, differences in mood and sleep
disturbances were observed, with a higher prevalence of depression, anxiety and
REM sleep behaviour disorder in freezers than in non-freezers. Depression scores
increased with advancing disease stages in all patients, whilst anxiety and REM sleep
behaviour disorder were not related to disease stage. These results support the
hypothesis that a deficit in conflict processing, which can occur within and between
motor, cognitive, affective or perceptual processing systems, may underlie freezing
of gait results (Lewis & Barker, 2009; Lewis & Shine, 2016; Vandenbossche et al.,
2012). Finally, autonomic disturbances were not associated with freezing of gait
across early or advanced disease stages, suggesting freezing behaviour does not
simply arise as a result of a brainstem lesion, nor is it likely that one pathological
lesion in a certain brain area will result in this transient and episodic symptom.
Rather, we hypothesize that ineffective information processing across subcortical and
cortical sites may underlie freezing of gait.

192
The results from Chapter 3 provided the theoretical foundation for the study
presented in Chapter 4. Namely, I considered the organisation of the white matter
connectome, allowing for assessing changes on the network level. Indeed,
investigating changes in network topology can elucidate changes in nodes that are
important in the integration and segregation of different networks (Sporns et al.,
2004). Changes in these scores could inform how effective the processing of
information is within and across different brain networks (Bertolero et al., 2015). A
group comparison between patients with and without freezing of gait showed low
participation scores in several nodes across subcortical, superior frontal and superior
parietal cortices. Moreover, a significant number of affected nodes were part of the
‘rich-club’, a group of high degree nodes that are densely connected with each other
and form the skeleton of the connectome. The rich-club plays a crucial role in global
information integration between different modules in the brain (van den Heuvel et
al., 2011). Moreover, the findings from Chapter 4 demonstrated that investigating
network topology could be advantageous for understanding a symptom that is
associated with pathological impairments at multiple levels across cortical and
subcortical regions.
The findings presented above are consistent with extensive evidence from functional
MRI studies showing reduced activity within, and decoupling between cortico-
cortical and cortico-striatal networks (Shine, 2013a; 2013b; Tessitore et al., 2012).
Whilst these results provide additional evidence for the hypothesis that freezing may
arise from inefficient shifting between functionally distinct but complementary
neural pathways (Lewis & Shine, 2016), the findings can also be interpreted as the
interplay between decreased gait automaticity and impairments in cognitive control
(Vandenbossche et al., 2012). The decreased participation scores in the basal ganglia
could indicate impairment in automatic processing. In such circumstances, patients
might need to rely on cortical control to operate an effective gait pattern. The study
presented in Chapter 4 showed decreased participation scores in both frontal and
parietal regions, which could indicate ineffective regulation of cognitive control
necessary to compensate for the loss of automaticity. Alternative freezing of gait

193
models, such as the decoupling model (Jacobs et al., 2009) and the abnormal gait
pattern generation hypothesis (Plotnik et al., 2005) were not directly investigated in
Chapter 4. Indeed, the brain atlas used in Chapter 4 did not include the
pedunculopontine nucleus as a region of interest. However, the findings in Chapter 3
did not allude to specific brainstem lesions (as indicated by similar autonomic
functioning across groups), and the current results argue that deficiencies across
multiple cortical and subcortical domains contribute to the freezing phenomenon.
7.1.3. To investigate whether visual hallucinations in Parkinson’s disease arise due to a
reweighting of attentional and perceptual brain networks (Chapter 5 and Chapter 6)
Visual hallucinations in Parkinson’s disease are common but the incomplete
knowledge of its pathophysiology limits treatment strategies. Previous work has
associated attentional dysfunction to hallucination proneness (Hepp et al., 2013;
Meppelink et al., 2008). Attention encompasses several processes such as alerting
and maintaining awareness, orienting, noticing and selecting stimuli for processing
or executive control. One test that can dissociate different attentional processes is the
Attentional Network Task (Fan et al., 2002). As such, to refine our understanding of
attentional deficits, Chapter 5 explored attentional processes in patients with visual
hallucinations using the Attentional Network Task. This study showed that
hallucinators had more missed and erroneous responses, especially in incongruent
trial conditions, compared to non-hallucinators. Hence, with increasing task
demands, impairments in sustained attention and conflict processing arise. The
reaction times between the groups on correct trials did not differ, suggesting a
selective communication breakdown between attentional networks.
However, not all patients with impaired attentional processes experience visual
hallucinations. Indeed, several models that have tried to explain the underlying
processes have suggested that hallucinations and misperceptions arise due to failure
of effective coordinated activities within and between attentional networks (Shine et
al., 2014; 2015). As such, when investigating the structural changes associated with
hallucinatory behaviour, it may be beneficial to investigate the communication

194
between different perceptual processing sites. A system-level perspective can
provide unique insights into between- and within-network communication, which
was employed in Chapter 6 of this thesis. This study investigated changes in
structural network topology associated with severity of hallucinatory behaviour,
which was assessed using a composite score including both a behavioural test and a
subscale of a questionnaire. This study found that the severity of hallucinatory
behaviour was associated with an overall reduction in connectivity strength in a sub-
network, spanning both hemispheres. Importantly, nodes within this network on
average showed higher between- and within-module scores compared to the
unaffected network and the majority of diverse club nodes were included in this
network. As such, this reduced connectivity associated with severity of
hallucinations may have extensive implications for processing speed and efficiency
across the connectome. Furthermore, a number of connections between nodes of the
diverse club (a network of nodes with high participation coefficients) also showed
reduced connectivity strength. The diverse club is thought to organise network
integration and segregation (Bertolero et al., 2015), thus changes between these
nodes could have detrimental effects for network communication. On a regional
level, the findings showed reduced participation in the right lateral occipital cortex,
pars orbitalis and insula, whilst increased participation was observed the left medial
orbitofrontal cortex, left posterior and right anterior cingulate, regions typically
involved in generating perceptual predictions (Bar et al., 2006; Chaumon et al.,
2014), endogenous attention (Andrews-Hanna et al., 2010; Vincent et al., 2006) and
error monitoring (Botvinick et al., 2004; 1999; Carter et al., 1998) respectively.
These findings are in accordance with the hypothesis presented in Chapter 5 and the
findings presented in Box 5 (O’Callaghan et al., 2017). This structural pattern supports
the assertion that visual misperceptions may arise due to increased reliance on areas
involved in predictive processing, in combination with reduced visual input and less
effective top-down control mechanisms that fail to recruit the appropriate networks
(Shine et al., 2014). Interestingly, one node within the right dorsal attentional network
(DAN) (‘ctx-rh-caudalmiddlefrontal_2’) was positively correlated with a higher

195
hallucination severity score. This finding was unexpected and contradictory to our
hypothesis mentioned in Chapter 5 of this thesis, namely that a loss of connectivity
between the DAN and other attentional networks. However, there were no connections
between this node and nodes from the DAN or default mode network. As such, the higher
between-module score may reflect a compensatory response to the loss of connectivity
strength in other regions of the DAN.
Finally, the current results also align with the Perception and Attention Deficit
Model (Collerton et al., 2005), which proposes that a hallucination, or the intrusion
of an incorrect proto-object, interfere with perception due to a combination of
impaired attention and visual perception. This model does not specify where or how
these unfitting proto-objects may arise. However, the results of Chapter 6 within this
thesis, as well as previous findings (Shine et al., 2014) would suggest that the
processes occurring in the default mode network represent a candidate mechanism
for the generation of these internal perceptual intrusions.
Finally, the Perception and Action Deficit Model as well as other previous work (Hepp et
al., 2017; Bullock et al., 2002; Burn et al., 2006; Perry et al., 1995) have shown
involvement of the cholinergic system in the occurrence of hallucinations in Parkinson’s
disease. Acetylcholine modulates the orienting network (Posner, et al., 2008), but also
influences the communication between attentional networks (Demeter & Sarter, 2012).
Deficits in sustained attention may also indicate impairments in the cholinergic system
(Turchi & Sarter, 2001). A follow-up unpublished analysis on the same dataset used in
Chapter 5 investigated fluctuations in reaction times, but showed no differences
between hallucinators and non-hallucinators on the coefficient of variation when
investigating variability across all trials, nor when analysing congruent, incongruent
and neutral trials separately. However, deficits in sustained attention may not be
specific to patients with hallucincations. More research is needed to investigate this
relationship.

196
7.1.4 Converging themes: similarities and differences between freezing of gait and visual
hallucinations
Albeit with differing specific manifestations, the current thesis has illustrated the
pathophysiological overlap between seemingly disparate symptoms and suggests that
freezing and hallucinations may occur due to a failure of effective information
processing between large-scale neuronal networks.
Consistent with the presented results, previous work has shown that both freezing and
hallucinations in Parkinson’s disease can be considered as an outcome of a
communication breakdown between functional attentional networks (Shine, 2013a) and
this may instigate the occurrence of transient, paroxysmal unwanted behaviours. Conflict
in information processing can further explain the increasing occurrence of both of these
symptoms under conditions where there is additional cognitive load. For example,
freezing behaviour is exacerbated during dual-tasking or stress, whilst hallucinations are
more likely to occur in cluttered environments with multiple competing stimuli that
require processing.
However, both hallucinations and freezing of gait also showed unique anatomical
signatures. Of the 29 patients included in the study presented in Chapter 6, sixteen
patients scored positively on the Freezing of Gait Questionnaire item 3, indicating
they have experienced freezing of gait. Specifically, thirteen patients self-reported
both freezing of gait and hallucinations (a positive score on PsycH-QA), three
patients reported freezing of gait but no hallucinations and thirteen patients reported
hallucinations but no freezing of gait. This indicates that these symptoms are likely
part of the same non-tremor dominant phenotype, yet occur often independently. In
Chapter 4 of this thesis, patients with freezing showed predominantly lower
participation scores, which is interpreted at an overall loss of effective information
processing across subcortical, frontal and parietal sites, whilst hallucinatory
behaviour (in Chapter 6) showed both decreased and increased scores across top-
down and bottom-up perceptual processing regions, reflecting a reweighting of the
perceptual system. The most notable difference between the two symptoms was

197
found in the cingulate gyrus. Freezing was associated with lower participation in the
anterior and posterior cingulate, whilst hallucinations were associated with higher
participation in these regions. Diffusivity changes as well as cortical thinning in
freezing of gait in the cingulate gyrus have previously been reported (Pietracupa et
al., 2018; Vastik et al., 2017). Conversely, previous work has not observed
diffusivity changes in the cingulate tracts in PD hallucinators (Hepp et al., 2017),
whilst functional imaging studies have shown increased activity within posterior
cingulate (Yao et al., 2014). This region further showed increased functional activity
in Dementia with Lewy Bodies, a Parkinsonian condition in which visual
hallucinations are highly prevalent, compared to Alzheimer’s disease patients
(Franciotti et al., 2013).
These differences between freezing and hallucinatory behaviour highlight the
importance of assessing well-matched groups when investigating different symptoms
of Parkinson’s disease individually. Indeed, changes in network architecture
associated with a specific symptom might skew the results when these issues are not
taken into account. However, it needs to be acknowledged that patients present with
co-morbid symptoms (Lewis et al., 2005). For example, both freezing of gait and
visual hallucinations are both associated with the non-tremor dominant phenotype
hence they commonly co-occur (Lewis et al., 2005) (see also Chapter 3). The
refinement of our understanding of the different subtypes within Parkinson’s disease
is therefore important in our ability to recognise the behavioural and
pathophysiological interaction of symptoms.
7.2 Limitations of the current work
The work in this thesis does have some limitations. In the next section, the
limitations and their implications will be discussed.

198
7.2.1 Self-reported symptoms
An important limitation of the empirical work presented in this thesis is where the
group allocation of patients is based on self-report questionnaire data. In Chapter 2
and 3, freezing of gait was identified using the Freezing of Gait Questionnaire (FOG-
Q) (Giladi et al., 2000). Whilst previous work has shown that this is a reliable tool to
identify freezers (Shine et al., 2012), ideally freezing of gait should be observed in
the clinical setting, using objective gait assessments, such as a gait mat,
accelerometry or Timed Up and Go tests (Galna et al., 2013). Objective assessments
however, lend themselves to false-negative classification, since an affected
individual might not freeze during their brief visit to the clinic, especially when
taking their optimal dose of dopaminergic medication. As such, most research
investigating freezing has used self-reported questionnaires (Iansek et al., 2016;
Nieuwboer et al., 2013). Furthermore, Chapter 4 of this thesis identified patients
with visual hallucinations using question 2 of the MDS-UPDRS. Hallucinations
occur on a continuum and it is possible that patients with benign misperceptions
might not yet recognize these more subtle events as being abnormal. On the other
end of the spectrum, patients can also lose insight into their visual hallucinations.
Both of these scenarios may cause discrepancies in responding to this item on the
MDS-UPDRS. At the University of Sydney, Parkinson’s Disease Research Clinic,
the MDS-UPDRS is obtained by a trained neurologist, who can probe the patient
(and informant) if there is doubt. This approach may thus overcome some of the
difficulties in capturing hallucinatory symptoms.
Whilst objective assessment of any symptom is desirable, this bears particular difficulties
when assessing visual hallucinations, as they are not observable by anyone other than the
affected individual. To circumvent this issue, the Bistable Percept Paradigm, an objective
test that can elicit misperceptions in patients with Parkinson’s disease, was designed
(Shine et al., 2012a). In Chapter 6, visual hallucinations were assessed using a detailed
validated questionnaire about hallucinatory behaviour (Shine et al., 2015) and the
Bistable Percept Paradigm (BPP). The BPP is a computerized task that aims to elucidate
visual misperceptions, and can be compared to the Pareidolia test (Uchiyama et al.,

199
2012), which is developed for patients with dementia with Lewy bodies. Whilst the
Pareidoloa test has been shown to have a good inter-rater and test-retest reliability and
exhibits high sensitivity to discriminate dementia with Lewy bodies from Alzheimer’s
disease, it does not correlate with clinical visual hallucinations (Mamiya et al., 2016). A
new simplified version of the Pareidolia test has shown to be a valid and reliable
assessment of visual hallucinations in patients with dementia with Lewy bodies (Mamiya
et al., 2016). Future work should compare the BPP and the adjusted Pareidolia test
(Mamiya et al., 2016), which was not available at time of testing of the current work.
The Psychosis and Hallucinations Questionnaire (PsycH-Q), includes items to probe
a broad spectrum of hallucinatory symptoms and their frequency (Shine et al., 2015).
This questionnaire is written in ‘non-scientific’ language and aimed to improve the
sensitivity to changes in the frequency and severity of hallucinatory and psychotic
symptoms in Parkinson’s disease. Using a combined score of objective and
subjective assessments, we aimed to circumvent the sole reliance on self-reported
symptom occurrence whilst controlling for the possibility of misclassifying
hallucinators as non-hallucinators, when patients did not experience visual
hallucinations during their clinic visit. Additionally, by creating this composite
score, we are able to assess visual hallucinatory severity, rather than classifying
patients into more artificial dichotomous patient groups.
Finally, whilst hallucinations in other modalities, including auditory, olfactory, tactile and
gustatory systems, also exist in Parkinson’s disease, this thesis has mainly discussed
hallucinations in the visual system. Importantly, the measurements used in Chapter 5
(MDS-UPDRS item 2) and Chapter 6 (PsycH-Q) assess hallucinations in all sensory
modalities. Hence, a firm conclusion about hallucinations in the visual domain cannot be
made. However, sensory hallucinations other than visual hallucinations are rare in
Parkinson’s disease, especially when unaccompanied by visual hallucinations or
misperceptions (Inzelberg et al., 1998). Therefore, the majority of the hallucinations of
our participants are likely of visual nature. Lastly, hallucinations in other sensory
domains are poorly understood due to a dearth of studies in this area. However, the
hypothesis proposed in Chapter 6 is not limited to visual hallucinations. In fact, previous

200
work has proposed that hallucinations in different sensory domains in mental illness are
due to a bias towards top-down information (for review, see Powers et al., 2016).
7.2.2 Cross-sectional studies
The studies presented in this thesis used a cross-sectional study design. A cross-
sectional design limits the interpretation of a temporal relationship between two
measures. For example, in the study presented in Chapter 3, I presented the
prevalence of freezing across different disease stages. However, future work should
carry out a longitudinal design to track disease progression in freezers compared to
non-freezers to further elucidate temporal changes and their dependence on specific
characteristics associated with this phenomenon.
7.2.3 Limitations of DTI
Chapter 4 and 6 of this thesis used DTI analyses to investigate white matter changes
in Parkinson’s disease patients. However, DTI suffers from a number of inherent
limitations. Firstly, diffusion imaging measures the motion of water molecules along
Box 4: Identifying risk factors for freezing of gait
Ehgoetz Martens, K. A., Lukasik, E., Georgiades, M, J., Gilat, M., Hall, J.M., Walton,
C. C., Lewis, S. J. G. Predicting the onset of freezing of gait: A Longitudinal study.
Movement Disorders, 2017.
This longitudinal study used a logistic regression model including demographic
factors, motor, executive and mood disturbances, to identify risk factors for the
development of freezing of gait. The model found that the Freezing of Gait
Questionnaire (item 1 and 2) together with the Hospital Anxiety and Depression Scale
could predict with > 70% accuracy if patients would transition into freezers at follow
up (1 year). These results suggest that patients at risk for freezing could be identified,
and allow for targeted interventions to delay or prevent the development of freezing
behaviour in patients with Parkinson’s disease.

201
a tract from which no directionality can be differentiated. Additionally, this method
only allows for an indirect validation of white matter tracts and therefore it is not
possible to infer whether demyelination or axonal injury is the source of changes in
anisotropy values. Secondly, DTI tractography approaches are incapable of resolving
crossing or ‘kissing’ fibres (Lanyon, 2012). At an ambiguous directional point in a
voxel, fractional anisotropy values fall below the cut-off threshold leading to a
premature termination of the reconstructed tract. Therefore, connectomes constructed
in these Chapters may potentially miss important axons. More sophisticated methods
such as constrained spherical deconvolution (CSD) are capable of producing
biologically expected ‘fan-shaped’ configuration of tracts (Tournier et al., 2004). So
far, several studies investigating Parkinson’s disease have employed CSD.
Specifically, a recent study compared a multi-shell, multi-tissue (MSMT) to single-
shell, single tissue (SSST) CSD approach to investigate structural network topology
in Parkinson’s disease patients compared to healthy controls (Kamagata et al., 2018).
Whilst the SSST technique was able to show differences in global strength and
small-worldness, the MSMT CSD also revealed differences in efficiency and
clustering between the groups, though it should be noted that the specificity was not
investigated in this study. Nevertheless, these developments are promising to unravel
white matter changes in Parkinson’s disease and other neurological disorders.
Finally, our data was acquired when field maps or phase encodings in different
directions were not readily available and as such, DTI scans were not corrected for
inhomogeneities in field distortions. This can have implications for registration of
images among individual diffusion weighted images that bias microstructural
assessment. To address this issue, the b0 images were registered to the segmented T1
using boundary-based registration (bbregister in FreeSurfer (Greve et al., 2009).
Despite these approaches, it should be appreciated that the connectomes might have
been suboptimal and the results should be validated, ideally implementing the
aforementioned techniques such as MSMT CSD in future work.

202
7.3 Future directions
To ensure progress in understanding the nature of freezing and hallucinatory
behaviour and how they relate to neural markers, candidate mechanisms need to be
identified of how these symptoms might arise based on a growing understanding of
the relevant systems. Whilst the work presented in this thesis has made a significant
contribution to this understanding, more work is required to fully understand the
pathophysiology of these symptoms. Recommendations for future research into these
symptoms are discussed below. Furthermore, the use of graph theory can open up
exciting avenues for further research in Parkinson’s disease, which are discussed in
the next section.
7.3.1 Assessing freezing of gait and visual hallucinations
Objective testing
To further our understanding of freezing behaviour, future research should explore
novel, minimally invasive and time-consuming ways to identify this symptom, for
example using body-worn technologies or a gait carpet, which is able to measure
specific gait characteristics such as step length, and velocity, variability, and single
and double stance time objectively (Mazilu et al., 2016). Unfortunately, these
systems currently lack the ability to accurately detect freezing. Techniques for
recording objective hallucinations (such as from ambulatory electroencephalographic
recordings) are at a much less developed stage. The results presented in Chapter 6
suggested that visual hallucinations might arise due to an over-reliance of top-down
prediction centres. This assumption should further be tested in a clinical setting. An
objective test that can identify shifts towards prior expectations has successfully been
used to test patients with psychosis (Teufel et al., 2015). In this test, participants
were shown two-toned images that were meaningless without prior knowledge of
their content, which was only provided in a number of cases.

203
Participants that rely more heavily on prior knowledge showed an advantage in
discriminating the ambiguous images when prior knowledge about the image was
available. Therefore, objective tasks capitalizing on subtle variation in perception can be
applied in Parkinson’s disease patients with visual hallucinators to test the assumption of
a shift towards prediction centres. Furthermore, computational explanations of changes
along the perceptual processing sites can be tested in terms of Bayesian models of
perception within a predictive coding framework, for example, using a drift diffusion
model (O’Callaghan et al., 2017), in order to test the understanding of the reweighting
occurring within the perceptual system.
Box 5: Evidence for impaired sensory evidence accumulation in Parkinson’s disease
patients with visual hallucinations
O’Callaghan, C., Hall, J. M., Tomassini, A., Muller, A. J., Walpola, I. C., Moustafa, A.
A., Shine, J. M., Lewis, S. J. G. Visual Hallucinations Are Characterized by Impaired
Sensory Evidence Accumulation: Insights From Hierarchical Drift Diffusion Modeling in
Parkinson's Disease. Biological Psychiatry: Cognitive Neuroscience and Neuroimaging,
2017
Computational explanations of changes along the perceptual hierarchy can be tested in
terms of Bayesian models of perception within a predictive coding framework. Recent
work has made an important contribution to perception research in Parkinson’s disease
(O’Callaghan et al., 2017). This study fitted a hierarchical drift diffusion model to the
ANT to identify the drift rates of the different task components. This study observed
slower drift rates and wider decision boundaries in patients with visual hallucinations,
suggesting reduced perceptual precision. The relative amount of evidence that bottom-up
and top-down processes provide mediates the integration of information of each
component. Perceptual information in this patient group might be less informative, and
therefore down-weighted, which could result in an over-reliance on prior expectations.

204
Confirming lack of integration using transmagnetic stimulation
Whilst increased severity of hallucinations (i.e. increased % misperceptions and/or
increased frequency) is often accompanied by a lack of insight (Amar et al., 2014),
this was not specifically tested in this thesis. However, Chapter 6 of this thesis
showed that the severity of hallucinations was associated with reduced connectivity
between a large-scale network and between several nodes of the diverse club. As
such, it is possible that changes in the environmental awareness are associated with
decreased network integration. Previous work has shown that transmagnetic
stimulation (TMS) can induce an interaction between cortical areas, resulting in a
more integrated state of these regions (Massimini et al., 2005). Thus, TMS may be
capable of identifying a node’s integrative ability. As such, this quality of nodes may
in future be testable using this an approach in patients with and without visual
hallucinations or freezing of gait.
7.3.4 Temporal course of pathophysiology
Both freezing of gait and visual hallucinations are likely to be functional disorders,
as evidenced by their transient and episodic nature. If these symptoms arose merely
due to a structural lesion, then these phenomena would be more predictable. Hence,
to fully understand the impact of the structural changes associated with freezing of
gait and visual hallucination in Parkinson’s disease, future research should combine
structural with functional with functional analysis, such as fMRI or
electroencephalogram (EEG), to elucidate their interaction.
Furthermore, longitudinal studies should investigate whether changes in white matter
precede functional abnormalities and/or grey matter lesions. Recent work in early
Parkinson’s disease patients has shown that white matter alterations are present
before grey matter degeneration and hereby challenged the traditional view that grey
matter is initially affected (Rektor et al., 2018). Therefore, treatment strategies
should focus on targeting any white matter abnormalities early in the course of
disease. Previous research has shown that white matter health can be improved by a

205
combination of physical exercise and social engagement in healthy older adults
(Burzynska et al., 2017). Importantly, this indicates that age-related changes in white
matter integrity are at least partly reversible or modifiable. Such behavioural
treatments could be beneficial for patients at risk of developing freezing or
hallucinations (Burzynska et al., 2017), especially since these phenomena are not
consistently responsive to dopaminergic treatment (Combs et al., 2017; Giladi,
2008).
Functional changes could occur before any cell death or demyelination has emerged.
Functional activity is more intimately tied to synaptic firing, which is regulated by
neurotransmitters and cellular events (Lodish, 2000). Furthermore, Lewy body
pathology intrusion does not immediately result in atrophy of cortical brain structure
(Sterling et al., 2016). Therefore, another important research avenue is to elucidate
the effect of changes in the balance of different neurotransmitters on structural
integrity and whether future disease modifying therapies that impact on disease
progression could also reduce white matter disturbance.
Box 6: Cognitive training in patients with freezing of gait
Walton, C. C., Mowszowski, L. Gilat, M., Hall, J. M., O’Callaghan, C., Muller, A. J.,
Georgiades, M. J., Szeto, J. Y. Y., Ehgoetz Martens, K. A., Shine, J. M., Naismith, S.
L., Lewis, S. J. G. Cognitive training for freezing of gait in Parkinson’s disease: a
randomized controlled trial. NPJ Parkinson's Disease, 2018; 4(15).
Our research group has recently completed a randomised controlled trial in which
patients with freezing of gait underwent either a cognitive training program or an active
sham program. The results were cautiously promising: patients in the training group
reduced their time percent frozen (in the OFF state) significantly compared to the
control group. Changes in functional MRI are currently being analysed, but
unfortunately, diffusion weighted imaging was not included in this study, and future
studies are necessary to assess if such a program could instigate changes in white matter
in patients with a neurodegenerative disorder.

206
7.3.5 Network topology in Parkinson’s disease heterogeneity
By reframing Parkinson’s disease as a ‘disconnection disorder’, this thesis has shown
that unique insights in the pathophysiology of individual symptoms can be gained
using graph theoretical methods. Graph theory can also be applied to elucidate the
progression of neurodegenerative disorders and therefore will be able to verify or
rearticulate current models.
Parkinson’s disease pathology affects dissociated but dispersed brain networks,
suggesting that the pathology transmits changes from the initial injury site to diffuse
networks towards the later stages of the disease. Vulnerability of these secondary
sites may be governed by the connectional structure of the brain, in which nodes with
high participation coefficients are particularly vulnerable due to their greater
susceptibility to oxidative stress, as a result of their myriad of connections and high
metabolic demands (Buckner et al., 2009; Saxena et al., 2011). Indeed, previous
work in other neurodegenerative disorders including Alzheimer’s disease, cortical
basal syndrome, semantic and frontotemporal dementia, as well as schizophrenia, has
shown that vulnerability of highly connected nodes could be predicted by the total
functional connectional flow through the node in combination with the path length to
the affected injury site (Crossley et al., 2014). Importantly, the anatomical location
of the affected hub regions (“epicentres”) may be specific to an individual disorder.
For example, in Alzheimer’s disease, the temporal lobe and the parietal regions were
predominantly affected (Crossley et al., 2014; Park et al., 2017), whereas in
schizophrenia, medial frontal and anterior cingulate regions were most affected
(Brugger et al., 2017; Crossley et al., 2014; Rubinov et al., 2013). Parkinson’s
disease could also be explained by this “transneuronal spread hypothesis” (Crossley
et al., 2014), in which the striatal nuclei would serve as the epicentre and fulfil the
criteria of high vulnerability: a short path length to a dominant injury site, the
substantia nigra and high participation across different modules.
Furthermore, the type of connections might dictate the vulnerability of a node to
Lewy body pathology. Previous work has shown that long-range connections or

207
widely distributed axonal projections appear particularly vulnerable to amyloid
(Wirths et al., 2006) whilst short-range or clustered dendritic and inter-neuronal
connections are particularly vulnerable to certain tauopathies (McMillan et al.,
2008). Thus, specific pathogenic disfigurements direct how pathology disseminates
across a network (Warren et al., 2013). Previous work has shown that Lewy body
pathology appears to be transmitted retrogradely via axon terminals (Surmeier et al.,
2017). Whether or not Lewy body pathology prefers a certain configuration of
connections remains to be investigated. Using graph theoretical models, the
connectivity profile of each node could be determined and in combination with
pathological studies, future studies could test whether nodes with high degree or high
participation are more vulnerable to Lewy body intrusion, compared to nodes with
lower connectivity strength. If so, investigations into network topology may not only
predict the progression of Parkinson’s disease but could also allow insights to further
explain the heterogeneity observed in this condition. Specifically, the site of
pathology intrusion may inform how the pathology will spread. It could be
hypothesized that patients in the non-tremor dominant phenotype, including
symptoms such as freezing of gait and visual hallucinations, show a greater hub
vulnerability in frontoparietal regions including connector hubs compared to patients
in the tremor dominant phenotype (see Figure 1). Lesions of these rich club nodes
lead to a more profound deterioration of network efficiency compared to changes
peripheral nodes. In future, such a method combined with computational modelling
may be implemented to test this hypothesis.
Furthermore, longitudinal investigations into network topology changes are
necessary to further understand the progression of Parkinson’s disease. Importantly,
after neuronal injury to a node, the module or modules may reconfigure and therefore
hyperconnectivity between other parts of the network can be observed (Hillary et al.,
2017). Longitudinal investigations into network architecture can thus provide
information about insult as well as compensation strategies of the network over time.

208
Figure 1: A hypothetical overview of how pathology may spread if nodes with high
connectivity strength display increased vulnerability, depicting two possible scenarios,
representing different clinical phenotypes of Parkinson’s disease. Red: affected nodes;
grey: non-affected nodes; dashed lines: affected edges; blue arrows: pathology intrusion
Zooming in
If changes in network integration and segregation underlie behavioural symptoms of
Parkinson’s disease, an important next step is to identify which nodes play a crucial
role in network configuration. As shown in this thesis, nodes belonging to the rich or
diverse club are crucial for effective communication across networks. Specifically,
cross-modular connections through rich clubs enhance network complexity (Zamora-
Lopez et al., 2016). Importantly, few candidate mechanisms have been proposed for
the optimisation of integration and segregation. For example, disruptions in the
coordinated projection of dopamine to the striatum likely result in a failure to timely
activate and inhibit behaviours (Chakravarthy et al., 2018; Howard et al., 2017) and
may be an obvious mechanism for changes in network topology in Parkinson’s
disease. However, this dopamine-centric model cannot solely account for the
multitude of non-motor symptoms observed in PD and other neuromodulatory
systems of the brainstem are likely to be involved in the tuning of functional

209
connection strength and therefore the optimal global synchronisation. Indeed, Shine
and colleagues (Shine et al., 2018) showed that nodes in the ascending arousal
system, including the noradrenergic locus coeruleus, modulate neural gain. Neural
gain controls the extent to which a particular region is excitable and thus susceptible
to interacting with other regions within or between networks. Additionally, recent
work in rodents has further highlighted the notion that integration does not depend
on the largest network hubs, but can be coordinated by a set of low-degree nodes but
highly influential nodes (Del Ferraro et al., 2018).
It is possible that changes in nodes with a high collective influence underlie
symptoms such as visual hallucinations and freezing of gait. As such, future studies
should investigate whether local activity in nodes that previously have been
implicated in the pathophysiology of these symptoms (e.g. the pedunculopontine
nucleus and nucleus basalis of Meynert) influence global computation by calculating
their intrinsic ignition (Deco et al., 2017). Identifying nodes with high collective
influence could provide a new avenue for treatment strategies.
7.4 Concluding remarks
This thesis has made a notable contribution to the understanding of freezing of gait
and visual hallucinations in Parkinson’s disease. The work has added to the known
phenotypical associations of freezing of gait by comparing patients with and without
this symptom across different disease stages. In addition, the relationship between
visual hallucinations and attention were assessed through previously unexplored
tasks. Importantly, to date most symptoms have been assessed using traditional DWI
measures, which in some cases has led to conflicting results (as outlined in Chapter
2). Paroxysmal symptoms of Parkinson’s disease, such as freezing of gait and visual
hallucinations are not likely to be the result of a focalized disruption but rather stem
from a network reconfiguration. Investigating the structural network topology
associated with freezing and hallucinatory behaviour was able to elucidate this
hypothesis. Disruptions in effective information processing that may underlie
different symptoms of Parkinson’s disease, might be amplified by these affected hub

210
regions. Indeed, these connector hubs play a crucial role in selecting and inhibiting
behaviours, and disruption in the integration within and between neurological
networks can result in a multitude of symptoms across cognitive, motor and affective
domains. Finally, the future use of the approaches implemented in this thesis may
provide critical information to our understanding of disease progression,
heterogeneity and the targeting of future therapies to improve the lives of the
millions of people affected by Parkinson’s disease.

211
7.5 References
Amar, B. R., Yadav, Ravi, Janardhan Reddy, Y. C., & Pal, Pramod Kumar. (2014). A
clinical profile of patients with Parkinson's disease and psychosis. Ann Indian
Acad Neurol, 17(2), 187-192. doi:10.4103/0972-2327.132625
Andrews-Hanna, J. R., Reidler, J. S., Sepulcre, J., Poulin, R. & Buckner, R. L. (2010).
Functional-Anatomic Fractionation of the Brain's Default Network. Neuron,
65(4), 550-562. doi:https://doi.org/10.1016/j.neuron.2010.02.005
Bar, M., Kassam, K. S., Ghuman, A. S., Boshyan, J., Schmid, A. M., Dale, A. M., . . .
Halgren, E. (2006). Top-down facilitation of visual recognition. Proc Natl Acad
Sci U S A, 103(2), 449-454. doi:10.1073/pnas.0507062103
Bertolero, M. A., Yeo, B. T., & D’Esposito, M. (2015). The modular and integrative
functional architecture of the human brain. Proc Natl Acad Sci U S A, 112(49),
E6798-E6807. doi:10.1073/pnas.1510619112
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004). Conflict monitoring and anterior
cingulate cortex: an update. Trends Cogni Sci, 8(12), 539-546.
Botvinick, M., Nystrom, L. E., Fissell, K., Carter, C. S., & Cohen, J. D. (1999). Conflict
monitoring versus selection-for-action in anterior cingulate cortex. Nature, 402,
179. doi:10.1038/46035
Brugger, S. P., & Howes, O. D. (2017). Heterogeneity and homogeneity of regional brain
structure in schizophrenia: A meta-analysis. JAMA Psychiatry, 74(11), 1104-
1111. doi:10.1001/jamapsychiatry.2017.2663
Buckner, R. L., Sepulcre, J., Talukdar, T., Krienen, F. M., Liu, H., Hedden, T., . . .
Johnson, K. A. (2009). Cortical hubs revealed by intrinsic functional connectivity:
mapping, assessment of stability, and relation to Alzheimer's disease. J Neurosci,
29(6), 1860-1873. doi:10.1523/jneurosci.5062-08.2009
Bullock, R., & Cameron, A. (2002). Rivastigmine for the treatment of dementia and
visual hallucinations associated with Parkinson's disease: a case series. Curr Med
Res Opin, 18(5), 258-264. doi:10.1185/030079902125000813
Burn, D., Emre, M., McKeith, I., De Deyn, P. P., Aarsland, D., Hsu, C., & Lane, R.
(2006). Effects of rivastigmine in patients with and without visual hallucinations

212
in dementia associated with Parkinson's disease. Mov Disord, 21(11), 1899-1907.
doi:10.1002/mds.21077
Burzynska, A. Z., Jiao, Y., Knecht, A. M., Fanning, J., Awick, E. A., Chen, T., . . .
Kramer, A. F. (2017). White Matter Integrity Declined Over 6-Months, but Dance
Intervention Improved Integrity of the Fornix of Older Adults. Front Aging
Neurosci, 9(59). doi:10.3389/fnagi.2017.00059
Carter, C. S, Braver, T. S, Barch, D. M, Botvinick, M. M., Noll, D., & Cohen, ¥ D.
(1998). Anterior cingulate cortex, error detection, and the online monitoring of
performance. Science, 280(5364), 747-749.
Chakravarthy, V. S., & Moustafa, A. A. (2018). Computational Neuroscience Models of
the Basal Ganglia. Singapore: Springer Nature.
Chaumon, M., Kveraga, K., Barrett, L. F., & Bar, M. (2014). Visual Predictions in the
Orbitofrontal Cortex Rely on Associative Content. Cereb Cortex, 24(11), 2899-
2907. doi:10.1093/cercor/bht146
Collerton, D., Perry, E., & McKeith, I. (2005). Why people see things that are not there: a
novel Perception and Attention Deficit model for recurrent complex visual
hallucinations. Behav Brain Sci, 28(6), 737-757; discussion 757-794.
doi:10.1017/S0140525X05000130
Combs, B. L., & Cox, A. G. (2017). Update on the treatment of Parkinson’s disease
psychosis: role of pimavanserin. Neuropsychiatric Dis Treatm, 13, 737-744.
doi:10.2147/NDT.S108948
Crossley, N. A., Mechelli, A., Scott, J., Carletti, F., Fox, P. T., McGuire, P., & Bullmore,
E. T. (2014). The hubs of the human connectome are generally implicated in the
anatomy of brain disorders. Brain, 137(Pt 8), 2382-2395.
doi:10.1093/brain/awu132
de Souza Fortaleza, A. C., Mancini, M., Carlson-Kuhta, P., King, L. A., Nutt, J. G.,
Chagas, E. F., . . . Horak, F. B. (2017). Dual task interference on postural sway,
postural transitions and gait in people with Parkinson's disease and freezing of
gait. Gait Posture, 56, 76-81. doi:10.1016/j.gaitpost.2017.05.006

213
Deco, G., & Kringelbach, M. L. (2017). Hierarchy of Information Processing in the
Brain: A Novel ‘Intrinsic Ignition’ Framework. Neuron, 94(5),
961-968. doi:10.1016/j.neuron.2017.03.028
Del Ferraro, G., Moreno, A., Min, B., Morone, F., Pérez-Ramírez, U., Pérez-Cervera, L.,
. . . Makse, H. A. (2018). Finding influential nodes for integration in brain
networks using optimal percolation theory. Nature Comm, 9(1), 2274.
doi:10.1038/s41467-018-04718-3
Demeter, E., & Sarter, M. (2012). Leveraging the cortical cholinergic system to enhance
attention. Neuropharmacology, 64(1), 294-304.
Fan, J., McCandliss, B. D., Sommer, T., Raz, A., & Posner, M. I. (2002). Testing the
efficiency and independence of attentional networks. J Cogn Neurosci, 14(3),
340-347. doi:10.1162/089892902317361886
Franciotti, R., Falasca, N. W., Bonanni, L., Anzellotti, F., Maruotti, V., Comani, S., . . .
Onofrj, ¥. (2013). Default network is not hypoactive in dementia with fluctuating
cognition: an Alzheimer disease/dementia with Lewy bodies comparison.
Neurobiol Aging, 34(4), 1148-1158.
doi:https://doi.org/10.1016/j.neurobiolaging.2012.09.015
Galna, B., Lord, S., Rochester, L. (2013). Is gait variability reliable in older adults and
Parkinson's disease? Towards an optimal testing protocol. Gait & Posture, 37(4);
580-585.
Giladi, N. (2008). Medical treatment of freezing of gait. Mov Disord, 23 Suppl 2, S482-
488. doi:10.1002/mds.21914
Giladi, N., Shabtai, H., Simon, E. S., Biran, S., Tal, J., & Korczyn, A. D. (2000).
Construction of freezing of gait questionnaire for patients with Parkinsonism.
Parkinsonism Relat Disord, 6(3), 165-170.
Grabli, D., Karachi, C., Welter, M. L., Lau, B., Hirsch, E. C., Vidailhet, M., & Francois,
C. (2012). Normal and pathological gait: what we learn from Parkinson's disease.
J Neurol Neurosurg Psychiatry, 83(10), 979-985. doi:10.1136/jnnp-2012-302263
Gratwicke, J. P., & Foltynie, T. (2018). Early nucleus basalis of Meynert degeneration
predicts cognitive decline in Parkinson’s disease. Brain, 141(1), 7-10.
doi:10.1093/brain/awx333

214
Greve, D. N., & Fischl, B. (2009). Accurate and Robust Brain Image Alignment using
Boundary-based Registration. Neuroimage, 48(1), 63-72.
doi:10.1016/j.neuroimage.2009.06.060
Hall, J. M., Shine, J. M., Walton, C. C., Gilat, M., Kamsma, Y. P., Naismith, S. L., &
Lewis, S. J. (2014). Early phenotypic differences between Parkinson's disease
patients with and without freezing of gait. Parkinsonism Relat Disord, 20(6), 604-
607. doi:10.1016/j.parkreldis.2014.02.028
Hepp, D. H., da Hora, C. C., Koene, T., Uitdehaag, B. M., van den Heuvel, O. A., Klein,
M., . . . Foncke, E. M. (2013). Cognitive correlates of visual hallucinations in non-
demented Parkinson's disease patients. Parkinsonism Relat Disord, 19(9), 795-
799. doi:10.1016/j.parkreldis.2013.04.026
Hepp, D. H., Foncke, E. M. J., Berendse, H. W., Wassenaar, T. M., Dubbelink, K. T. E.
O., Groenewegen, H J., . . . Schoonheim, M. M. (2017). Damaged fiber tracts of
the nucleus basalis of Meynert in Parkinson’s disease patients with visual
hallucinations. Sci Rep, 7(1), 10112.
Hillary, F. G., & Grafman, J. H. (2017). Injured Brains and Adaptive Networks: The
Benefits and Costs of Hyperconnectivity. Trends Cogn Sci, 21(5), 385-401.
doi:10.1016/j.tics.2017.03.003
Howard, C. D., Li, H., Geddes, C. E., & Jin, X. (2017). Dynamic Nigrostriatal Dopamine
Biases Action Selection. Neuron, 93(6), 1436-1450.e1438.
doi:10.1016/j.neuron.2017.02.029
Iansek, R. & Danoudis, M. (2016). Freezing of Gait in Parkinson's Disease: Its
Pathophysiology and Pragmatic Approaches to Management. Mov Disord Clin
Prac, n/a-n/a. doi:10.1002/mdc3.12463
Inzelberg, R., Kipervasser, S., Korczyn, A. D. (1998). Auditory hallucinations in
Parkinson’s disease. J Neurol Neurosurg Psychiatry, 64:533-535.
Jacobs, J. V., Nutt, J. G., Carlson-Kuhta, P., Stephens, M., & Horak, F. B. (2009). Knee
trembling during freezing of gait represents multiple anticipatory postural
adjustments. Exp Neurol, 215(2), 334-341. doi:10.1016/j.expneurol.2008.10.019
Kamagata, K., Zalesky, A., Hatano, T., Di Biase, M. A., El Samad, O., Saiki, S., . . .
Pantelis, C. (2018). Connectome analysis with diffusion MRI in idiopathic

215
Parkinson's disease: Evaluation using multi-shell, multi-tissue, constrained
spherical deconvolution. Neuroimage Clin, 17, 518-529.
doi:10.1016/j.nicl.2017.11.007
Lanyon, L. J. (2012). Diffusion tensor imaging: Structural connectivity insights,
limitations and future directions. Neuroimaging Methods. InTech.
Latora, V., & Marchiori, M. (2003). Economic small-world behavior in weighted
networks. Eur Phys J B Condens Matter, 32(2), 249-263. doi:10.1140/epjb/e2003-
00095-5
Lee, T. W. & Xue, S. W. (2018). Functional connectivity maps based on hippocampal
and thalamic dynamics may account for the default-mode network. Eur J Neuosci
47(5):388-398. doi: 10.1111/ejn.13828.
Lee, J. Y., Yoon, E. J., Lee, W. W., Kim, Y. K., Lee, JY., & Jeon, B. (2016). Lateral
geniculate atrophy in Parkinson's with visual hallucination: A trans-synaptic
degeneration? Mov Disord, 31(4), 547-554. doi:10.1002/mds.26533
Lewis, S. J. G., & Barker, R. A. (2009). A pathophysiological model of freezing of gait in
Parkinson's disease. Parkinsonism Relat Disord, 15(5), 333-338.
doi:10.1016/j.parkreldis.2008.08.006
Lewis, S. J. G., & Shine, J. M. (2016). The Next Step: A Common Neural Mechanism for
Freezing of Gait. Neuroscientist, 22(1), 72-82. doi:10.1177/1073858414559101
Lewis, S. J. G., Foltynie, T., Blackwell, A. D., Robbins, T. W., Owen, A. M., & Barker,
R. A. (2005). Heterogeneity of Parkinson’s disease in the early clinical stages
using a data driven approach. J Neurol Neursurg Psychiatry, 76(3), 343-348.
Lodish, H., Berk, A,. Zipursky, S. L. (2000). Molecular Cell Biolology (4th edition ed.).
New York: W. H. Freeman.
Massimini, M., Ferrarelli, F., Huber, R., Esser, S. K., Singh, H., & Tononi, G. (2005).
Breakdown of cortical effective connectivity during sleep. Science, 309(5744),
2228-2232. doi:10.1126/science.1117256
Mazilu, S., Blanke, U., Calatroni, A., Gazit, E., Hausdorff, J. M., & Tröster, G. (2016).
The role of wrist-mounted inertial sensors in detecting gait freeze episodes in
Parkinson’s disease. Pervasive Mob Comput, 33, 1-16.
doi:http://dx.doi.org/10.1016/j.pmcj.2015.12.007

216
McMillan, P., Korvatska, E., Poorkaj, P., Evstafjeva, Z., Robinson, L., Greenup, L., . . .
D'Souza, I. (2008). Tau isoform regulation is region- and cell-specific in mouse
brain. J Comp Neurol, 511(6), 788-803. doi:10.1002/cne.21867
Meppelink, A. M., Koerts, J., Borg, M., Leenders, K. L., & van Laar, T. (2008). Visual
object recognition and attention in Parkinson's disease patients with visual
hallucinations. Mov Disord, 23(13), 1906-1912. doi:10.1002/mds.22270
Mi, T. M., Mei, S. S., Liang, P. P., Gao, L. L., Li, K. C., Wu, T., & Chan, P. (2017).
Altered resting-state brain activity in Parkinson's disease patients with freezing of
gait. Sci Rep, 7(1), 16711. doi:10.1038/s41598-017-16922-0
Naismith, S. L., Shine, J. M., & Lewis, S. J. G. (2010). The specific contributions of set-
shifting to freezing of gait in Parkinson's disease. Mov Disord, 25(8), 1000-1004.
doi:10.1002/mds.23005
Nieuwboer, A., & Giladi, N. (2013). Characterizing freezing of gait in Parkinson's
disease: models of an episodic phenomenon. Mov Disord, 28(11), 1509-1519.
doi:10.1002/mds.25683
O’Callaghan, C., Hall, J. M., Tomassini, A., Muller, A. J., Walpola, I. C., Moustafa, A.
A., . . . Lewis, S. J. G. (2017). Visual Hallucinations Are Characterized by
Impaired Sensory Evidence Accumulation: Insights From Hierarchical Drift
Diffusion Modeling in Parkinson’s Disease. Biol Psychiatry Cogn Neurosci
Neuroimaging, 2(8), 680-688. doi:/10.1016/j.bpsc.2017.04.007
Park, J. Y., Na, H. K., Kim, S., Kim, H., Kim, H. J., Seo, S. W., . . . Alzheimer’s Disease
Neuroimaging, Initiative. (2017). Robust Identification of Alzheimer’s Disease
subtypes based on cortical atrophy patterns. Sci Rep, 7, 43270.
doi:10.1038/srep43270
Perry, E. K., & Perry, R. H. (1995). Acetylcholine and hallucinations: disease-related
compared to drug-induced alterations in human consciousness. Brain Cogn, 28(3),
240-258. doi:10.1006/brcg.1995.1255
Pietracupa, S., Suppa, A., Upadhyay, N., Gianni, C., Grillea, G., Leodori, G., . . .
Pantano, P. (2018). Freezing of gait in Parkinson's disease: gray and white matter
abnormalities. J Neurol, 265(1), 52-62. doi:10.1007/s00415-017-8654-1

217
Pietracupa, S., Suppa, A., Upadhyay, N., Giannì, C., Grillea, G., Leodori, G., . . .
Pantano, P. (2018). Freezing of gait in Parkinson’s disease: gray and white matter
abnormalities. J Neurol, 265(1), 52-62. doi:10.1007/s00415-017-8654-1
Plotnik, M., Giladi, N., Balash, Y., Peretz, C., & Hausdorff, J. M. (2005). Is freezing of
gait in Parkinson's disease related to asymmetric motor function? Ann Neurol,
57(5), 656-663. doi:10.1002/ana.20452
Posner, M. I., Fan, J., & Pomerantz, J. R. (2008). Topics in integrative neuroscience.
Powers, A. R., Kelley, M., & Corlett, P. R. (2016). Hallucinations as top-down effects on
perception. Biol Psychiatry. Cogn Neurosci Neuroimaging, 1(5), 393-400.
Rektor, I., Svátková, A., Vojtíšek, L., Zikmundová, I., Vaníček, J., Király, A., & Szabó,
N. (2018). White matter alterations in Parkinson’s disease with normal cognition
precede grey matter atrophy. PLoS One, 13(1), e0187939.
doi:10.1371/journal.pone.0187939
Rochester, L., Yarnall, A. J., Baker, M. R., David, R. V., Lord, S., Galna, B., Burn, D. J.
(2012). Cholinergic dysfunction contributes to gait disturbance in early
Parkinson’s disease, Brain, 135(9); 779–2788. doi:10.1093/brain/aws207
Rubinov, M., & Bullmore, E. (2013). Schizophrenia and abnormal brain network hubs.
Dial Clin Neurosci, 15(3), 339-349.
Saxena, S., & Caroni, P. (2011). Selective neuronal vulnerability in neurodegenerative
diseases: from stressor thresholds to degeneration. Neuron, 71(1), 35-48.
doi:10.1016/j.neuron.2011.06.031
Shine, J. M. (2013). Attentional Network Dysfunction in Parkinson’s disease (PhD),
University of Sydney, Sydney.
Shine, J. M., Aburn, M. J., Breakspear, M., & Poldrack, R. A. (2018). The modulation of
neural gain facilitates a transition between functional segregation and integration
in the brain. Elife, 7. doi:10.7554/eLife.31130
Shine, J. M., Halliday, G. H., Carlos, M., Naismith, S. L., & Lewis, S. J. (2012).
Investigating visual misperceptions in Parkinson's disease: a novel behavioral
paradigm. Mov Disord, 27(4), 500-505. doi:10.1002/mds.24900
Shine, J. M., Halliday, G. M., Gilat, M., Matar, E., Bolitho, S. J., Carlos, M., . . . Lewis,
S. J. G. (2014). The role of dysfunctional attentional control networks in visual

218
misperceptions in Parkinson's disease. Hum Brain Mapp, 35(5), 2206-2219.
doi:10.1002/hbm.22321
Shine, J. M., Matar, E., Ward, P. B., Bolitho, S. J., Gilat, M., Pearson, M., . . . Lewis, S.
J. G. (2013). Exploring the cortical and subcortical functional magnetic resonance
imaging changes associated with freezing in Parkinson's disease. Brain, 136(Pt 4),
1204-1215. doi:10.1093/brain/awt049
Shine, J. M., Moore, S. T., Bolitho, S. J., Morris, T. R., Dilda, V., Naismith, S. L., &
Lewis, S. J. G. (2012). Assessing the utility of Freezing of Gait Questionnaires in
Parkinson's Disease. Parkinsonism Relat Disord, 18(1), 25-29.
doi:10.1016/j.parkreldis.2011.08.002
Shine, J. M., Muller, A. J., O'Callaghan, C., Hornberger, M., Halliday, G. M., & Lewis,
S. J. G. (2015). Abnormal connectivity between the default mode and the visual
system underlies the manifestation of visual hallucinations in Parkinson's disease:
a task-based fMRI study. NPJ Parkinsons Dis, 1, 15003.
doi:10.1038/npjparkd.2015.3
Shine, J. M., Ward, P. B., Naismith, S. L., Pearson, M., & Lewis, S. J. G. (2011).
Utilising functional MRI (fMRI) to explore the freezing phenomenon in
Parkinson's disease. J Clin Neurosci, 18(6), 807-810. doi:S0967-5868(11)00110-
X [pii] doi:10.1016/j.jocn.2011.02.003
Shine, J. M., Mills, J. M. Z., Qiu, J., O'Callaghan, C., Terpening, Z., Halliday, G. M, . . .
Lewis, S. J. G. (2015). Validation of the Psychosis and Hallucinations
Questionnaire in Non�demented Patients with Parkinson's Disease. Mov Disord
Clin Pract, 2(2), 175-181.
Snijders, A. H., Takakusaki, K., Debu, B., Lozano, A. M., Krishna, V., Fasano, A., . . .
Hallett, M. (2016). Physiology of freezing of gait. Ann Neurol, 80(5), 644-659.
doi:10.1002/ana.24778
Sporns, O., Chialvo, D., Kaiser, M., Hilgetag, C. C. (2004) Organization, development
and function of complex brain networks. Trends Cogn Sci., 8:418–425
Squire, L. R., & Zola-Morgan, S. (1991). The medial temporal lobe memory system.
Science, 253(5026), 1380-1386.

219
Sterling, N. W., Lewis, M. M., Du, G., & Huang, X. (2016). Structural imaging and
Parkinson’s disease: Moving toward quantitative markers of disease progression.
J Parkinsons Dis, 6(3), 557-567.
Surmeier, D. J., Obeso, J. A., & Halliday, G. M. (2017). Selective neuronal vulnerability
in Parkinson disease. Nat Rev Neurosci, 18(2), 101-113.
doi:10.1038/nrn.2016.178
Tessitore, A., Amboni, M., Esposito, F., Russo, A., Picillo, M., Marcuccio, L., . . .
Barone, P. (2012). Resting-state brain connectivity in patients with Parkinson's
disease and freezing of gait. Parkinsonism Relat Disord, 18(6), 781-787.
doi:10.1016/j.parkreldis.2012.03.018
Tessitore, A., Esposito, F., Vitale, C., Santangelo, G., Amboni, M., Russo, A., . . .
Tedeschi, G. (2012). Default-mode network connectivity in cognitively
unimpaired patients with Parkinson disease. Neurology, 79(23), 2226-2232.
doi:10.1212/WNL.0b013e31827689d6
Teufel, C., Subramaniam, N., Dobler, V., Perez, J., Finnemann, J., Mehta, P. R., . . .
Fletcher, P. C. (2015). Shift toward prior knowledge confers a perceptual
advantage in early psychosis and psychosis-prone healthy individuals. Proc Natl
Acad Sci U S A, 112(43), 13401-13406. doi:10.1073/pnas.1503916112
Tournier, J. D., Calamante, F., Gadian, D. G., Connelly, A. Direct estimation of the fiber
orientation density function from diffusion-weighted MRI data using spherical
deconvolution. NeuroImage, 23(3), 1176-1185.
doi:10.1016/j.neuroimage.2004.07.0.37
Turchi, J., & Sarter, M. (2001). Bidirectional modulation of basal forebrain N-methyl-D-
aspartate receptor function differentially affects visual attention but not visual
discrimination performance. Neuroscience, 104(2), 407-417.
van den Heuvel, M. P., & Sporns, O. (2011). Rich-club organization of the human
connectome. J Neurosci, 31(44), 15775-15786. doi:10.1523/JNEUROSCI.3539-
11.2011
Vandenbossche, J., Deroost, N., Soetens, E., Coomans, D., Spildooren, J., Vercruysse, S.,
. . . Kerckhofs, E. (2012). Freezing of gait in Parkinson's disease: disturbances in

220
automaticity and control. Front Hum Neurosci, 6, 356.
doi:10.3389/fnhum.2012.00356
Vandenbossche, J., Deroost, N., Soetens, E., Zeischka, P., Spildooren, J., Vercruysse, S., .
. . Kerckhofs, E. (2012). Conflict and freezing of gait in Parkinson's disease:
support for a response control deficit. Neuroscience, 206, 144-154.
doi:10.1016/j.neuroscience.2011.12.048
Vastik, M., Hok, P., Valosek, J., Hlustik, P., Mensikova, K., & Kanovsky, P. (2017).
Freezing of gait is associated with cortical thinning in mesial frontal cortex.
Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub, 161(4), 389-396.
doi:10.5507/bp.2017.035
Vincent, J. L., Snyder, A. Z., Fox, M. D., Shannon, B. J., Andrews, J. R., Raichle, M. E.,
& Buckner, R. L. (2006). Coherent spontaneous activity identifies a hippocampal-
parietal memory network. J Neurophysiol, 96(6), 3517-3531.
Vossel, S., Geng, J. J., & Fink, G. R. (2014). Dorsal and Ventral Attention Systems:
Distinct Neural Circuits but Collaborative Roles. Neuroscientist, 20(2), 150-159.
doi:10.1177/1073858413494269
Walton, C. C., Mowszowski, L., Gilat, M., Hall, J. M., O'Callaghan, C., Muller, A. J., . . .
Lewis, S. J. G. (2018). Cognitive training for freezing of gait in Parkinson's
disease: a randomized controlled trial. NPJ Parkinsons Dis, 4, 15.
doi:10.1038/s41531-018-0052-6
Walton, C. C., O'Callaghan, C., Hall, J. M., Gilat, M., Mowszowski, L., Naismith, S. L., .
. . Lewis, S. J. G. (2015). Antisaccade errors reveal cognitive control deficits in
Parkinson's disease with freezing of gait. J Neurol, 262(12), 2745-2754.
doi:10.1007/s00415-015-7910-5
Walton, C. C., Shine, J. M., Mowszowski, L., Gilat, M., Hall, J. M., O'Callaghan, C., . . .
Lewis, S. J. G. (2015). Impaired cognitive control in Parkinson's disease patients
with freezing of gait in response to cognitive load. J Neural Transm (Vienna),
122(5), 653-660. doi:10.1007/s00702-014-1271-6
Warren, J. D., Rohrer, J. D., Schott, J. M., Fox, N. C., Hardy, J., & Rossor, M. N. (2013).
Molecular nexopathies: a new paradigm of neurodegenerative disease. Trends
Neurosci, 36(10), 561-569. doi:10.1016/j.tins.2013.06.007

221
Wirths, O., Weis, J., Szczygielski, J., Multhaup, G., & Bayer, T. A. (2006). Axonopathy
in an APP/PS1 transgenic mouse model of Alzheimer's disease. Acta
Neuropathol, 111(4), 312-319. doi:10.1007/s00401-006-0041-4
Yao, N., Cheung, C. Pang, S., Shek-Kwan, C. R., Lau, K. K., Suckling, J. … McAlonan,
G. M. (2014). Multimodal MRI of the hippocampus in Parkinson’s disease with
visual hallucinations. Brain Struct Func. doi:10.1007/s00429-014-0907-5.
Yao, N., Shek�Kwan C., Richard, C., Charlton, P., Shirley, L., Kui K., Suckling, J., . . .
Chua, S. E. (2014). The default mode network is disrupted in Parkinson's disease
with visual hallucinations. Hum Brain Mapp, 35(11), 5658-5666.
Zamora-Lopez, G., Chen, Y., Deco, G., Kringelbach, M. L., & Zhou, C. (2016).
Functional complexity emerging from anatomical constraints in the brain: the
significance of network modularity and rich-clubs. Sci Rep, 6, 38424.
doi:10.1038/srep38424

APPENDICES

ORIGINAL COMMUNICATION
The major impact of freezing of gait on quality of lifein Parkinson’s disease
Courtney C. Walton • James M. Shine • Julie M. Hall • Claire O’Callaghan •
Loren Mowszowski • Moran Gilat • Jennifer Y. Y. Szeto • Sharon L. Naismith •
Simon J. G. Lewis
Received: 6 August 2014 / Revised: 8 September 2014 / Accepted: 30 September 2014 / Published online: 16 October 2014! Springer-Verlag Berlin Heidelberg 2014
Abstract Freezing of gait (FOG) is a disabling motorsymptom experienced by a large proportion of patients
with Parkinson’s disease (PD). While it is known that FOG
contributes to lower health-related quality of life (HRQoL),previous studies have not accounted for other important
factors when measuring the specific impact of this symp-
tom. The aim of this study was to examine FOG andHRQoL while controlling for other factors that are known
to impact patient well-being, including cognition, motor
severity, sleep disturbance and mood. Two hundred andthree patients with idiopathic PD (86 with FOG) were
included in the study. All patients were between Hoehn and
Yahr stages I–III. A forced entry multiple regression modelevaluating the relative contribution of all symptoms was
conducted, controlling for time since diagnosis and current
dopaminergic treatment. Entering all significantly corre-lated variables into the regression model accounted for the
majority of variance exploring HRQoL. Self-reported
sleep–wake disturbances, depressive and anxious symp-toms and FOG were individually significant predictors.
FOG accounted for the highest amount of unique variance.
While sleep–wake disturbance and mood have a significantnegative impact on HRQoL in PD, the emergence of FOG
represents the most substantial predictor among patients in
the earlier clinical stages of disease. This finding presum-ably reflects the disabling loss of independence and fear of
injury associated with FOG and underlines the importance
of efforts to reduce this common symptom.
Keywords Parkinson’s disease ! Freezing of gait !Quality of life ! Depression ! Anxiety ! Sleep
Introduction
Parkinson’s disease (PD) is a common neurodegenerative
disorder [1] in which the combination of both motor and
non-motor symptoms has a significant and negative impacton patients’ quality of life. Health-related quality of life
(HRQoL) is the most typically used outcome measure
representing an assessment made by patients about thedetrimental impact of disease and its consequences on their
life. This is a crucial aspect of modern medicine given theimportance of applying biopsychosocial models to treat-
ment [2].
Many studies have now assessed HRQoL in PD, mostcommonly using the Parkinson’s Disease Questionnaire
(PDQ-39) [3]. These studies have revealed that disease
severity, motor complications and dopaminergic treatmentall appear to contribute to HRQoL [4–6]. However, a
recent large-scale multicenter study showed that the com-
bined effect of non-motor symptoms was the most signif-icant predictor of HRQoL, when compared to the motor
aspects of the disease [7]. More specifically, the study
identified mood disturbance and sleep problems/fatigue ashaving the strongest negative impact on HRQoL. Indeed,
C. C. Walton ! J. M. Shine ! J. M. Hall ! L. Mowszowski !M. Gilat ! J. Y. Y. Szeto ! S. L. Naismith ! S. J. G. Lewis (&)Parkinson’s Disease Research Clinic, Brain and Mind ResearchInstitute, University of Sydney, Sydney, NSW, Australiae-mail: [email protected]
C. C. Walton ! L. Mowszowski ! S. L. NaismithHealthy Brain Ageing Program, Brain and Mind ResearchInstitute, University of Sydney, Sydney, NSW, Australia
C. O’CallaghanNeuroscience Research Australia and School of MedicalSciences, University of New South Wales, Sydney, NSW,Australia
123
J Neurol (2015) 262:108–115
DOI 10.1007/s00415-014-7524-3

this is a finding that replicates those shown previously in
the literature [8, 9]. Given the frequency with which non-motor symptoms occur [10], understanding their significant
impact on individuals with PD is critical for effective
patient management.While appreciating the significant impact of non-motor
aspects of PD is important, the same is true of freezing of
gait (FOG), which is a symptom also experienced by manypatients as their disease progresses. The phenomenon of
FOG presents as the sudden inability to generate effectivestepping and forward progression despite the intention to
do so [11]. The occurrence of FOG significantly increases a
patient’s likelihood of falling and subsequent nursing homeplacement [12]. While research into this symptom is
growing in interest, treatment options for FOG are pres-
ently limited [13] representing the currently incompleteunderstanding of the underlying pathophysiology [14].
Given the functional impact of FOG it is not surprising
that it has previously been linked to reduced HRQoL in PDpatients [15–17]. Moore and colleagues [17] have demon-
strated that FOG has a strong and significant impact on
HRQoL, irrespective of more general disease severitymeasures. Similarly, Rahman and colleagues [16] have also
reported that FOG negatively impacts on HRQoL, although
this study used a non-validated questionnaire to identifyfreezing and did not include this measure in their main
regression analyses. In a very recent study, Perez-Lloret
et al. [15] have also been able to correlate FOG withHRQoL. They used logistic regression to show that this
relationship occurred irrespective of disease severity and
duration, in addition to motor complications.Thus, previous studies exploring the impact of FOG on
HRQoL have not controlled well for variables outside of
disease and motor severity when assessing the impact ofthis symptom, which represents a significant limitation in
interpreting the relative strength of these findings. Prob-
lems with depression [18], anxiety [19] and sleep [20] havebeen related to FOG but are also large contributors to
HRQoL in PD more generally [7].
The aim of the current study was to assess whetherseverity of self-reported FOG still independently predicted
HRQoL, after controlling for the contribution of other
factors within PD. The current study introduced new fac-tors which have not previously been considered concur-
rently with FOG to determine the specific effects on
HRQoL. Based on previous findings in the field, we choseto analyze key variables of generalized cognition, mood,
sleep disturbance and motor function [6–9]. It was
hypothesized that the combination of these measures wouldaccount for a high proportion of variance in HRQoL scores.
However, our primary goal was to explore the unique
contribution of FOG to HRQoL while controlling for thecombined impact of these other key variables.
Methods
Sample
Data was collected from the baseline assessment of patientsat the ‘Parkinson’s Disease Research Clinic’ at the Brain
and Mind Research Institute, University of Sydney. Par-
ticipants included were assessed between December 2008and November 2013. The diagnosis of idiopathic PD was
based on the UK Brain Bank clinical criteria [21] and was
confirmed by a neurologist (SJGL). Patients with anMMSE of less than 24 were excluded due to possible
dementia and thus questionable validity of their self-report
questionnaires. Patients did not meet MDS criteria for PDwith dementia [22]. To explore the impact of FOG on
patients in the earlier clinical stages of disease where
paroxysmal limitation on mobility was more likely to be ofdaily significance and impact, the sample was restricted to
those patients rated as Hoehn and Yahr (H and Y) stages I–
III [23]. This research was approved by the HumanResearch Ethics Committee of The University of Sydney,
and written informed consent was obtained from all
participants.A sample of 285 patients met criteria for inclusion in the
study. However, 78 were removed due to missing data for
questionnaire or clinical variables. During initial datascreening, four participants were further removed from the
analysis due to being strongly significant outliers. There-fore, a sample of 203 patients was included in the study. Of
these, 86 reported a score of 1 or higher on ‘Question 3’ of
the Freezing of Gait Questionnaire (FOG-Q) [‘‘Do you feelthat your feet get glued to the floor while walking, making
a turn, or trying to initiate walking (freezing)?’’], which has
been shown to be a reliable screening tool to identifyfreezers [24]. As such, 43 % of the sample was deemed to
experience FOG by subjective report. Patient characteris-
tics are shown in Table 1.
Measures
We assessed HRQoL using the scaled index of the PDQ-39
[3]. This questionnaire consists of 39 multiple choice items
covering eight dimensions of ‘mobility’, ‘activities of dailyliving’, ‘emotional well-being’, ‘stigma’, ‘social support’,
‘cognitions’ ‘communication’ and ‘bodily discomfort’.
Scores are scaled between 0 and 100 with higher scoressuggesting poorer HRQoL. To assess FOG severity, we
used the FOG-Q [24]. The FOG-Q consists of six multiple
choice questions which assess a patient’s severity andfrequency of freezing. Scores range from 0 to 24, with
higher scores suggesting more severe FOG. Section III of
the Movement Disorder Society—Unified Parkinson’sDisease Rating Scale (MDS-UPDRS) [25] was used to
J Neurol (2015) 262:108–115 109
123

assess for motor severity. This assessment is conducted by
a clinician and contains 33 scores over 18 items. Scores
range from 0 to 132 with higher scores indicating worsemotor symptoms. Global cognitive functioning was asses-
sed using the Mini-Mental State Examination (MMSE)
[26]. The second edition of the Beck Depression Inventory(BDI-II) [27] was used to measure depressive symptom
severity. This self-report questionnaire comprises 21 itemsand yields a total score ranging from 0 to 63. Higher scores
suggest more severe depressive symptoms. Anxiety was
measured using the anxiety subscale of the Hospital Anx-iety and Depression Scale (HADS-anxiety) [28]. The scale
consists of 7 questions, with scores ranging from 0 to 21.
Higher scores suggest worse symptoms of anxiety. TheSCOPA-Sleep (SCOPA-S) [29] was used to measure
nighttime and daytime disturbance. Scores range from 0 to
15 for nighttime sleep and 0–18 for daytime sleepiness,with higher scores indicating poorer sleep quality. For
medically treated patients, dopamine dose equivalence
scores (DDE) were calculated (mg/day) [30]. Time since
diagnosis was calculated in months from date of diagnosis
to date of assessment.
Statistical analyses
Data analyses were conducted using the Statistical Package
for the Social Sciences (SPSS) version 21. All analyses
used an alpha of 0.05 and were two tailed. Pearson bivar-iate correlations were employed to assess for relationships
between the predictor and the outcome variables forregression. Significantly correlated predictor variables
were entered at once into a two-block multiple linear
regression model to assess their contribution to PDQ-39scores. Statistical assumptions of normality and hetero-
scedasticity were not met in some variables included in the
model. For this reason, we performed more conservativebootstrapping [31] with bias corrected and accelerated
confidence intervals for regression analyses. One thousand
bootstrap samples were implemented. The multicollinearityassumption was met, assessed by correlations and collin-
earity statistics.
Results
Patients defined as freezers scored higher on the PDQ-39
(M = 29.57, SD = 15.29) than those defined as non-
freezers (M = 15.22, SD = 9.36). More specifically,average PDQ-39 scores across responses to FOG-Q
Question 3 are illustrated in Fig. 1. This figure shows that
higher scores on this question show a linear relationshipwith worsened PDQ-39 scores.
Table 1 Clinical characteristics of sample
Mean Standard deviation
Age (years) 66.77 8.9
Gender (n)
Male 139
Female 64
Time since diagnosis (months) 61.30 61.3
Hoehn and Yahr stage (n)
1 39
1.5 7
2 98
2.5 39
3 20
DDE (mg/day) 615.81 487.8
UPDRS III 27.60 15.1
MMSE 28.35 1.6
PDQ-39 21.21 14.0
SCOPA-S day 4.29 3.5
SCOPA-S night 4.67 4.2
BDI-II 9.77 7.0
HADS-anxiety 3.95 3.4
FOG-Q total 5.34 5.7
n = 203, DDE dopamine dose equivalence, MDS-UPDRS IIIMovement Disorder Society Unified Parkinson’s Disease RatingScale (Motor subsection), MMSE Mini-Mental State Examination,PDQ-39 39-item Parkinson’s Disease Questionnaire, SCOPA-S Scales for Outcomes in Parkinson’s Disease (Sleep), BDI-II BeckDepression Inventory 2nd Edition, HADS Hospital Anxiety andDepression Scale (Anxiety scale), FOG-Q Freezing of GaitQuestionnaire
Fig. 1 Mean PDQ-39 scores for patients responding at differingseverity levels to Question 3 of the FOG-Q. Error bars represent 1standard error
110 J Neurol (2015) 262:108–115
123

Pearson bivariate correlations are displayed in Table 2
and show that PDQ-39 scores were significantly correlatedwith our control variables of time since diagnosis and
DDE. Age and gender did not show significant correlations
with the PDQ-39 and were not included in the regressionmodel. All remaining predictor variables (i.e., MMSE,
MDS-UPDRS III, FOG-Q, BDI-II, HADS-anxiety, SCO-
PA-S day and SCOPA-S night) correlated significantlywith the PDQ-39. The strongest relationship was between
FOG-Q and PDQ-39 scores, and this correlation is illus-trated in Fig. 2. Expanding on this, Table 3 also displays
correlations for measures of FOG with each of the eight
domains of the PDQ-39, showing that FOG significantlycorrelated with all.
To control for time since diagnosis and dopaminergic
medication use, these variables were first force entered intothe model as a block. These variables alone significantly
predicted 17.8 % of the variance in PDQ-39 scores
(F2,202 = 21.651, p \ 0.0001). However, once the addi-tional predictors were entered at the second stage, a DR2 of
54.4 % was found, suggesting a considerably strongermodel (72.2 % of total variance in HRQoL explained). At
this stage, however, time since diagnosis, DDE and MDS-
UPDRS III all became non-significant predictors, while the
Table 2 Pearson correlations between PDQ-39 and initial predictor variables
PDQ-39
Timesincediagnosis
DDE Age Gender MDS-UPDRSIII
MMSE SCOPA-S day
SCOPA-S night
BDI-II HADS-anxiety
FOG-Q
PDQ-39 – 0.374** 0.371** 0.093 0.092 0.313** -0.178* 0.522** 0.299** 0.615** 0.554** 0.628**
Time sincediagnosis
– 0.467** 0.111 -0.043 0.319** -0.221* 0.242** -0.001 0.147* 0.054 0.493*
DDE – 0.028 -0.066 0.160* -0.194** 0.351** 0.149* 0.144* 0.091 0.344**
Age – -0.054 0.342* -0.271** 0.141* 0.001 0.046 -0.155* 0.200**
Gender – -0.100 0.164* -0.010 0.131 0.146* 0.136 0.022
MDS-UPDRSIII
– -0.103 0.185** 0.013 0.195 0.037 0.419**
MMSE – -0.124 -0.028 -0.078 -0.041 -0.218**
SCOPA-S day – 0.356** 0.309** 0.226** 0.303**
SCOPA-Snight
– 0.209** 0.208** 0.050
BDI-II – 0.561** 0.293**
HADS-anxiety – 0.202**
FOG-Q –
* p B 0.05
** p B 0.01
Fig. 2 Scatterplot displaying individual FOG-Q and PDQ-39 scores
Table 3 Correlations between FOG measures and PDQ-39 subscales
FOG-Q total FOG-Q question 3
Mobility 0.801** 0.665**
Activities of daily living 0.560** 0.526**
Emotional well-being 0.352** 0.278**
Stigma 0.254** 0.210**
Social support 0.352** 0.303**
Cognitions 0.279** 0.250**
Communication 0.424** 0.441**
Bodily discomfort 0.369** 0.309**
* p B 0.05
** p B 0.01
J Neurol (2015) 262:108–115 111
123

predictor variables of FOG-Q, BDI-II, HADS-anxiety and
both SCOPA-S scores were significant contributors to themodel. MMSE was not a significant predictor. This mod-
el’s overall predictive power was statistically significant
(F9,202 = 57.696, p \ 0.0001).We next analyzed semi-partial correlations of the sig-
nificant contributors to account for the unique variance in
PDQ-scores that could be attributed to each of the signif-icant predictors in the full model. While accounting for the
other variables in the model, FOG-Q scores uniquely
contributed 9.4 %. The contribution of BDI-II to the modelwas 4.4 % and HADS-anxiety was 4.2 %. SCOPA-S day
scores contributed 3.2 % while night scores contributed
less than 1 %. Table 4 displays beta values along withother relevant statistics from the regression models.
Motor phenotype analysis
As an additional analysis, we calculated motor phenotypescores for ‘tremor’, ‘non-tremor’ and ‘rigidity’, as descri-
bed elsewhere [32, 33]. Tremor scores showed a weak
correlation with PDQ-39 (r = 0.145, p \ 0.05), non-tre-mor showed a medium to strong correlation (r = 0.447,
p \ 0.01), while rigidity did not significantly correlate
(r = 0.120, p [ 0.05). Importantly, however, non-tremorscores were strongly correlated with FOG-Q (r = 0.597,
p \ 0.01) and given this likely confound were not con-
sidered appropriate for inclusion in the regression model.
Discussion
The results of the current study have shown for the first
time that the independent contribution of FOG-Q scoresrepresents the strongest predictor of HRQoL in the earlier
clinical stages of PD, after accounting for the contributionof other key variables (general cognition, sleep distur-
bance, mood problems and motor severity). The variance
that could be independently attributed to FOG was morethan double that of depression, anxiety and sleep distur-
bance scores. The contribution of general cognition, motor
severity, dopaminergic medication use and time sincediagnosis was minimal when considering these variables.
These findings consolidate the work of Moore et al. [17],
Rahman et al. [16] and Perez-Lloret et al. [15] who foundFOG to be related to HRQoL. However, the current results
significantly extend these findings by introducing addi-
tional clinical factors known to strongly contribute toHRQoL, and illustrate that FOG still remains the most
significant predictor.
These results also confirm the hypothesis that measuresof self-reported FOG, mood and sleep disturbance all
contribute significantly to HRQoL in PD. This finding is
directly in agreement with previous studies highlighting theimportance of these factors in patient well-being [6–9, 15–
17]. The study also confirmed previous findings that non-
motor aspects of PD appear to have a more significantimpact on HRQoL than broad measures of motor severity
[7], with MDS-UPDRS III scores being a non-significant
contributor when included in the second stage of theregression model.
While it might seem initially surprising that FOG can be
more disabling to HRQoL than sleep and mood distur-bance, or motor severity, previously strong arguments have
been made for why this may be the case [17]. For example,
FOG is an episodic, paroxysmal phenomenon that canoccur in public or dangerous situations, which are stress
provoking (e.g., crossing roads or entering crowded spaces)
[34]. Thus, the disabling impact of a freezing episode canoccur at times which significantly impact patients’ sense of
safety, fear of falling and feeling of independence. Such
events can be embarrassing and frustrating and may lead topatients feeling isolated and of burden to their carers and/or
family [35]. Thus, negative responses could be anticipated
in a HRQoL assessment and indeed the correlations inTable 3 of domains such as ‘activities of daily living’,
‘social support’ and ‘emotional well-being’ support these
notions. In addition, our sample consisted of patients in the
Table 4 Linear model predictors of PDQ-39 scores showing boot-strapped beta scores with the corresponding standard error, signifi-cance values, and semi-partial correlations
Beta Std. errorof beta
Significancevalue
Semi-partialcorrelation
Model 1
Time sincediagnosis
0.052 0.017 0.003 0.203
DDE 0.008 0.002 0.002 0.234
Model 2
Time sincediagnosis
0.005 0.011 0.652 0.017
DDE 0.002 0.001 0.078 0.068
MDS-UPDRSIII
0.020 0.040 0.622 0.019
MMSE -0.051 0.383 0.874 -0.006
SCOPA-Snight
0.283 0.148 0.044 0.077
SCOPA-Sday
0.842 0.179 0.001 0.178
BDI-II 0.527 0.098 0.001 0.210
HADS-anxiety
1.041 0.211 0.001 0.206
FOG-Q 0.955 0.142 0.001 0.306
112 J Neurol (2015) 262:108–115
123

earlier clinical stages of the disease. The loss of indepen-
dence may be one of the most significant challenges forthese patients to cope with, possibly contributing to chan-
ges for example in social engagement or early retirement.
Depression and anxiety were the next most importantpredictors of HRQoL. Initial correlational analyses showed
that FOG was significantly correlated with these variables.
These relationships provide further support for the argu-ment above that FOG may have its negative impact on
patient well-being by making patients feel isolated and ofburden to their nearest and dearest [17]. The impact of
sleep disturbance on HRQoL was less strong. Interestingly,
it was daytime sleep disturbance that showed the strongercontribution, with night problems having a minimal impact
in the model. Our results suggest that duration of disease
and dopaminergic medication use do not strongly impacton HRQoL relative to these other clinical measures.
Currently, treatment options for FOG are poor. Classi-
cally, pharmacological intervention and cueing therapieshave been employed; however, deep brain stimulation has
increasingly grown in appeal [36]. Unfortunately, the
effects of cueing and pharmacological options are oftenshort lived, while deep brain stimulation is invasive and
unfortunately often suitable for only a small number of
patients [13]. It has previously been suggested that com-puterized cognitive training and virtual reality training may
be a beneficial option for the treatment of FOG and gait
disorders [13, 37]. However, the current results also pro-vide strong support for the role of more rounded remedi-
ation techniques employing concurrent group-based
psychoeducation, for example [38]. Such interventionshave been proposed as important factors in possibly
improving quality of life and well-being [39]. Patients with
FOG could be presumed to benefit from theseinterventions.
Limitations
It is important to consider the limitations to this study.
Primarily, we assessed freezing using self-report ques-tionnaire, the most accessible and widely used research
tool, and the results are uncompromisingly strong in favor
of the contribution of the FOG-Q to HRQoL, suggesting avery significant relationship. However, where resources
allow, correlating actual measures of freezing episodes
[e.g., 40] would be beneficial in future work. Secondly,MMSE showed little impact on HRQoL, however, given
the role of cognitive impairment in FOG [e.g., 41, 42], a
more detailed cognitive assessment may have influencedthe model. Future studies could explore this impact more
precisely. Finally, we had a much higher proportion of
male participants, but this is typically reflective of thedisease [43].
Conclusions
The current results have shown that self-reported FOG
severity is a strong independent contributor to HRQoL in
individuals with PD. This is likely to stem from the dailyimpact of FOG on mobility and independence, causing
stress and anxiety in addition to individuals perceiving
caregiver burden [35]. Follow-up studies to examine thespecific effect of FOG relative to these predictors in care-
giver QoL would be beneficial. These results highlight the
increasing need for a more solid understanding of patho-physiological mechanisms underlying FOG leading to
better treatment options. Clinicians in the field of neurol-
ogy must show increased awareness to the impact of FOGfor optimal management of PD patients.
Acknowledgments We would like to thank the patients of theParkinson’s Disease Research Clinic for being so generous with theirtime and efforts. CC Walton is supported by an Australian Post-graduate Award at the University of Sydney. C O’Callaghan is sup-ported by an Alzheimer’s Australia PhD Scholarship at the Universityof New South Wales. M Gilat is supported by an InternationalPostgraduate Research Scholarship at the University of Sydney. SLNaismith is supported by an NHMRC Career Development AwardNo. 1008117. SJG Lewis is supported by an NHMRC PractitionerFellowship No. 1003007. JM Shine, JM Hall, L Mowszowski andJYY Szeto have no financial disclosures.
Conflicts of interest The authors declare no conflict of interest.
Ethical standard This study has been approved by the appropriateethics committee and has therefore been performed in accordancewith the ethical standards laid down in the 1964 Declaration ofHelsinki.
References
1. Jankovic J (2008) Parkinson’s disease: clinical features anddiagnosis. J Neurol Neurosurg Psychiatry 79(4):368–376. doi:10.1136/jnnp.2007.131045
2. Martinez-Martin P (1998) An introduction to the concept of‘‘quality of life in Parkinson’s disease’’. J Neurol 245(Suppl1):S2–S6
3. Peto V, Jenkinson C, Fitzpatrick R, Greenhall R (1995) Thedevelopment and validation of a short measure of functioning andwell being for individuals with Parkinson’s disease. Qual LifeRes Int J Qual Life Asp Treat Care Rehabil 4(3):241–248
4. Muslimovic D, Post B, Speelman JD, Schmand B, de Haan RJ,Group CS (2008) Determinants of disability and quality of life inmild to moderate Parkinson disease. Neurology 70(23):2241–2247. doi:10.1212/01.wnl.0000313835.33830.80
5. Chapuis S, Ouchchane L, Metz O, Gerbaud L, Durif F (2005)Impact of the motor complications of Parkinson’s disease on thequality of life. Mov Disord Off J Mov Disord Soc 20(2):224–230.doi:10.1002/mds.20279
6. Slawek J, Derejko M, Lass P (2005) Factors affecting the qualityof life of patients with idiopathic Parkinson’s disease—a cross-sectional study in an outpatient clinic attendees. ParkinsonismRelat Disord 11(7):465–468. doi:10.1016/j.parkreldis.2005.04.006
J Neurol (2015) 262:108–115 113
123

7. Martinez-Martin P, Rodriguez-Blazquez C, Kurtis MM, Chau-dhuri KR, Group NV (2011) The impact of non-motor symptomson health-related quality of life of patients with Parkinson’sdisease. Mov Disord Off J Mov Disord Soc 26(3):399–406.doi:10.1002/mds.23462
8. Santos-Garcia D, de la Fuente-Fernandez R (2013) Impact ofnon-motor symptoms on health-related and perceived quality oflife in Parkinson’s disease. J Neurol Sci 332(1–2):136–140.doi:10.1016/j.jns.2013.07.005
9. Naismith SL, Hickie IB, Lewis SJ (2010) The role of milddepression in sleep disturbance and quality of life in Parkinson’sdisease. J Neuropsychiatry Clin Neurosci 22(4):384–389. doi:10.1176/appi.neuropsych.22.4.384
10. Chaudhuri KR, Healy DG, Schapira AH, National Institute forClinical E (2006) Non-motor symptoms of Parkinson’s disease:diagnosis and management. Lancet Neurol 5(3):235–245. doi:10.1016/S1474-4422(06)70373-8
11. Nutt JG, Bloem BR, Giladi N, Hallett M, Horak FB, NieuwboerA (2011) Freezing of gait: moving forward on a mysteriousclinical phenomenon. Lancet Neurol 10(8):734–744. doi:10.1016/S1474-4422(11)70143-0
12. Kerr GK, Worringham CJ, Cole MH, Lacherez PF, Wood JM,Silburn PA (2010) Predictors of future falls in Parkinson disease.Neurology 75(2):116–124. doi:10.1212/WNL.0b013e3181e7b688
13. Walton CC, Shine JM, Mowszowski L, Naismith SL, Lewis SJG(2014) Freezing of gait in Parkinson’s disease: current treatmentsand the potential role for cognitive training. Restor NeurolNeurosci 32(3):411–422. doi:10.3233/RNN-130370
14. Heremans E, Nieuwboer A, Vercruysse S (2013) Freezing of gaitin Parkinson’s disease: where are we now? Curr Neurol NeurosciRep 13(6):350. doi:10.1007/s11910-013-0350-7
15. Perez-Lloret S, Negre-Pages L, Damier P, Delval A, DerkinderenP, Destee A, Meissner WG, Schelosky L, Tison F, Rascol O(2014) Prevalence, determinants, and effect on quality of life offreezing of gait in Parkinson disease. JAMA Neurol. doi:10.1001/jamaneurol.2014.753
16. Rahman S, Griffin HJ, Quinn NP, Jahanshahi M (2008) Quality oflife in Parkinson’s disease: the relative importance of the symp-toms. Mov Disord Off J Mov Disord Soc 23(10):1428–1434.doi:10.1002/mds.21667
17. Moore O, Peretz C, Giladi N (2007) Freezing of gait affectsquality of life of peoples with Parkinson’s disease beyond itsrelationships with mobility and gait. Mov Disord Off J MovDisord Soc 22(15):2192–2195. doi:10.1002/mds.21659
18. Shine JM, Naismith SL, Lewis SJG (2013) The differential yetconcurrent contributions of motor, cognitive and affective dis-turbance to freezing of gait in Parkinson’s disease. Clin NeurolNeurosurg 115(5):542–545. doi:10.1016/j.clineuro.2012.06.027
19. Lieberman A (2006) Are freezing of gait (FOG) and panic related?J Neurol Sci 248(1–2):219–222. doi:10.1016/j.jns.2006.05.023
20. Videnovic A, Marlin C, Alibiglou L, Planetta PJ, VaillancourtDE, Mackinnon CD (2013) Increased REM sleep without atoniain Parkinson disease with freezing of gait. Neurology81(12):1030–1035. doi:10.1212/WNL.0b013e3182a4a408
21. Hughes AJ, Daniel SE, Kilford L, Lees AJ (1992) Accuracy ofclinical diagnosis of idiopathic Parkinson’s disease: a clinico-pathological study of 100 cases. J Neurol Neurosurg Psychiatry55(3):181–184
22. Emre M, Aarsland D, Brown R, Burn DJ, Duyckaerts C, MizunoY, Broe GA, Cummings J, Dickson DW, Gauthier S, Goldman J,Goetz C, Korczyn A, Lees A, Levy R, Litvan I, McKeith I,Olanow W, Poewe W, Quinn N, Sampaio C, Tolosa E, Dubois B(2007) Clinical diagnostic criteria for dementia associated withParkinson’s disease. Mov Disord Off J Mov Disord Soc22(12):1689–1707. doi:10.1002/mds.21507 (quiz 1837)
23. Hoehn MM, Yahr MD (1967) Parkinsonism: onset, progressionand mortality. Neurology 17(5):427–442
24. Giladi N, Shabtai H, Simon ES, Biran S, Tal J, Korczyn AD(2000) Construction of freezing of gait questionnaire for patientswith Parkinsonism. Parkinsonism Relat Disord 6(3):165–170
25. Goetz CG, Tilley BC, Shaftman SR, Stebbins GT, Fahn S,Martinez-Martin P, Poewe W, Sampaio C, Stern MB, Dodel R,Dubois B, Holloway R, Jankovic J, Kulisevsky J, Lang AE, LeesA, Leurgans S, LeWitt PA, Nyenhuis D, Olanow CW, Rascol O,Schrag A, Teresi JA, van Hilten JJ, LaPelle N, Movement Dis-order Society URTF (2008) Movement Disorder Society-spon-sored revision of the Unified Parkinson’s Disease Rating Scale(MDS-UPDRS): scale presentation and clinimetric testing results.Mov Disord Off J Mov Disord Soc 23(15):2129–2170. doi:10.1002/mds.22340
26. Folstein MF, Folstein SE, McHugh PR (1975) ‘‘Mini-mentalstate’’. A practical method for grading the cognitive state ofpatients for the clinician. J Psychiatr Res 12(3):189–198
27. Beck A, Steer R, Brown G (1996) Manual for the BDI-II. PsycholCorp, San Antonio
28. Zigmond AS, Snaith RP (1983) The hospital anxiety anddepression scale. Acta Psychiatr Scand 67(6):361–370
29. Marinus J, Visser M, van Hilten JJ, Lammers GJ, Stiggelbout AM(2003) Assessment of sleep and sleepiness in Parkinson disease.Sleep 26(8):1049–1054
30. Tomlinson CL, Stowe R, Patel S, Rick C, Gray R, Clarke CE(2010) Systematic review of levodopa dose equivalency reportingin Parkinson’s disease. Mov Disord Off J Mov Disord Soc25(15):2649–2653. doi:10.1002/mds.23429
31. Efron B, Tibshirani RJ (1993) An introduction to the bootstrap.Chapman & Hall, New York
32. Hall JM, Shine JM, Walton CC, Gilat M, Kamsma YP, NaismithSL, Lewis SJG (2014) Early phenotypic differences betweenParkinson’s disease patients with and without freezing of gait.Parkinsonism Relat Disord 20(6):604–607. doi:10.1016/j.parkreldis.2014.02.028
33. Lewis SJG, Foltynie T, Blackwell AD, Robbins TW, Owen AM,Barker RA (2005) Heterogeneity of Parkinson’s disease in theearly clinical stages using a data driven approach. J NeurolNeurosurg Psychiatry 76(3):343–348. doi:10.1136/jnnp.2003.033530
34. Rahman S, Griffin HJ, Quinn NP, Jahanshahi M (2008) Thefactors that induce or overcome freezing of gait in Parkinson’sdisease. Behav Neurol 19(3):127–136
35. Schrag A, Hovris A, Morley D, Quinn N, Jahanshahi M (2006)Caregiver-burden in parkinson’s disease is closely associatedwith psychiatric symptoms, falls, and disability. ParkinsonismRelat Disord 12(1):35–41. doi:10.1016/j.parkreldis.2005.06.011
36. Vercruysse S, Vandenberghe W, Munks L, Nuttin B, Devos H,Nieuwboer A (2014) Effects of deep brain stimulation of thesubthalamic nucleus on freezing of gait in Parkinson’s disease: aprospective controlled study. J Neurol Neurosurg Psychiatry85(8):871–877. doi:10.1136/jnnp-2013-306336
37. Mirelman A, Rochester L, Reelick M, Nieuwhof F, Pelosin E,Abbruzzese G, Dockx K, Nieuwboer A, Hausdorff JM (2013)V-TIME: a treadmill training program augmented by virtual realityto decrease fall risk in older adults: study design of a randomizedcontrolled trial. BMC Neurol 13:15. doi:10.1186/1471-2377-13-15
38. Naismith SL, Mowszowski L, Diamond K, Lewis SJG (2013)Improving memory in Parkinson’s disease: a healthy brain ageingcognitive training program. Mov Disord Off J Mov Disord Soc28(8):1097–1103. doi:10.1002/mds.25457
39. Walton CC, Mowszowski L, Lewis SJG, Naismith SL (2014)Stuck in the mud: time for change in the implementation ofcognitive training research in ageing? Front Aging Neurosci 6:43.doi:10.3389/fnagi.2014.00043
114 J Neurol (2015) 262:108–115
123

40. Nonnekes J, Janssen A, Mensink SG, Oude Nijhuis L, Bloem B,Snijders A (2014) Short rapid steps to provoke freezing of gait inParkinson’s disease. J Neurol. doi:10.1007/s00415-014-7422-8
41. Walton C, Shine J, Mowszowski L, Gilat M, Hall J, O’CallaghanC, Naismith SL, Lewis SJG (2014) Impaired cognitive control inParkinson’s disease patients with freezing of gait in response tocognitive load. J Neural Transm. doi:10.1007/s00702-014-1271-6
42. Naismith SL, Shine JM, Lewis SJG (2010) The specific contri-butions of set-shifting to freezing of gait in Parkinson’s disease.
Mov Disord Off J Mov Disord Soc 25(8):1000–1004. doi:10.1002/mds.23005
43. Wooten GF, Currie LJ, Bovbjerg VE, Lee JK, Patrie J (2004) Are menat greater risk for Parkinson’s disease than women? J Neurol Neuro-surg Psychiatry 75(4):637–639. doi:10.1136/jnnp.2003.020982
J Neurol (2015) 262:108–115 115
123

APPENDIX(2((
(Walton, C. C., Shine, J. M., Mowszowski, L., Gilat, M., Hall, J. M., O’Callaghan, C.,
Naismith, S. L., Lewis, S. J. G. Impaired cognitive control in Parkinson’s disease
patients with freezing of gait in response to cognitive load. Journal of
Neurotransmission, 2015

NEUROLOGY AND PRECLINICAL NEUROLOGICAL STUDIES - ORIGINAL ARTICLE
Impaired cognitive control in Parkinson’s disease patientswith freezing of gait in response to cognitive load
Courtney C. Walton • James M. Shine • Loren Mowszowski • Moran Gilat •
Julie M. Hall • Claire O’Callaghan • Sharon L. Naismith • Simon J. G. Lewis
Received: 31 May 2014 / Accepted: 3 July 2014 / Published online: 22 July 2014! Springer-Verlag Wien 2014
Abstract Freezing of gait is a frequent and disablingsymptom experienced by many patients with Parkinson’s
disease. A number of executive deficits have been shown to
be associated with the phenomenon suggesting a commonunderlying pathophysiology, which as of yet remains
unclear. Neuroimaging studies have also implicated the role
of the cognitive control network in patients with freezing. Toexplore this concept, the current study examined error-
monitoring as a measure of cognitive control. Thirty-four
patients with and 38 without freezing of gait, who wereotherwise well matched on disease severity, completed a
colour-word interference task that allowed the specific
assessment of error monitoring during conflict. Whilst bothgroups performed colour-naming and word-reading tasks
equally well, those patients with freezing showed a pattern
between conditionswhereby theywere better able tomonitorperformance and self-correct errors in the pure inhibition
task but not after a switching rule was introduced. The novel
results shown here provide insight into possible pathophys-iological mechanisms involved in cognitive load and error
monitoring in patients with freezing of gait. These resultsprovide further evidence for the role of functional fronto-
striatal circuitry impairments in patients with freezing of gait
and have implications for future studies and possible thera-peutic interventions.
Keywords Freezing of gait ! Parkinson’s disease !Executive function ! Error monitoring ! Stroop task !Cognitive control
Introduction
Freezing of gait (FOG) is a debilitating symptom experi-enced by many patients with Parkinson’s disease (PD)
(Giladi et al. 2001). The complex phenomenon is clinically
defined as the sudden inability to generate effective step-ping and forward progression despite the intention to do so
and is often described as feeling as though one’s feet are
glued to the ground (Nutt et al. 2011). The current treat-ment options for FOG are limited (Walton et al. 2014), and
its occurrence leads to a significantly increased risk of fallsand reduced quality of life (Gray and Hildebrand 2000;
Moore et al. 2007) highlighting the importance of gaining
an increased understanding of its manifestation.At present the pathophysiology underlying FOG is
poorly understood (Heremans et al. 2013b), although dys-
function of frontostriatal pathways is thought to be criti-cally involved (Hall et al. 2014; Lewis and Barker 2009;
Shine et al. 2013d). One area of increasing interest in the
literature is the distinct pattern of neuropsychologicaldeficits demonstrated by PD patients with FOG, which may
inform our understanding of the underlying neural mech-
anisms of freezing. A range of specific executive impair-ments are greater in patients with FOG than in those
C. C. Walton ! J. M. Shine ! L. Mowszowski ! M. Gilat !J. M. Hall ! S. L. Naismith ! S. J. G. Lewis (&)Parkinson’s Disease Research Clinic, Brain and Mind ResearchInstitute, University of Sydney, Sydney, NSW, Australiae-mail: [email protected]
C. C. Walton ! L. Mowszowski ! S. L. NaismithHealthy Brain Ageing Program, Brain and Mind ResearchInstitute, University of Sydney, Sydney, NSW, Australia
C. O’CallaghanNeuroscience Research Australia and School of MedicalSciences, University of New South Wales, Sydney, NSW,Australia
123
J Neural Transm (2015) 122:653–660
DOI 10.1007/s00702-014-1271-6

without. This has led many authors to suggest that partic-
ular cognitive functions may be intimately related tofreezing behaviour (For reviews see: Heremans et al.
2013a; Walton et al. 2014). General executive deficits have
been reported in patients with FOG (Amboni et al. 2010,2008), more specifically in the domains of set-shifting
(Naismith et al. 2010; Shine et al. 2013e), conflict resolu-
tion and inhibitory control (Cohen et al. 2014; Matar et al.2013; Vandenbossche et al. 2011, 2012) and implicit
learning (Vandenbossche et al. 2013b). These findings havebeen further validated by recent functional neuroimaging
studies, which have implicated the dysfunctional role of a
putative cognitive control network (CCN) operatingthrough both fronto-parietal cortices and subcortical
structures including the subthalamic nucleus (STN) and
striatum (Shine et al. 2013a, b, c).These neuroanatomical insights raise the possibility that
the executive deficits documented in FOG may be under-
pinned to an extent by impairments in cognitive control,which refers to the ability to control goal directed behav-
iour by organizing and optimising appropriate information
processing to flexibly respond to changes in predictedbehavioural outcomes (Narayanan et al. 2013; Ridderink-
hof et al. 2004). Taking these findings together leads to an
intriguing proposition: given FOG has been hypothesisedto occur as a result of a decreased neural reserve for coping
with demands on the frontostriatal system (Lewis and
Barker 2009), it could be suggested that under highlydemanding cognitive situations, patients with FOG display
an inefficient recruitment of cognitive control.
Therefore, the aim of the current study was to furtherexplore impairments in cognitive control using increasingly
demanding conditions of a verbal Stroop task. By using a
verbal rather than computerized format, the stimulus used inthe task matches the form of response needed (i.e., word–
word rather than word-button) leading to more automatic
responding (Kornblum et al. 1990). This is important inadequately differentiating between automatic versus effort-
ful processing (Vandenbossche et al. 2013a). Further, the
current Stroop task records both corrected and uncorrectederrors. Thus, different interpretations regarding cognitive
control can be recorded than in previous measures of Stroop
performance in FOG (Cohen et al. 2014; Vandenbosscheet al. 2012). An error that is corrected would imply cognitive
control has been enforced, albeit late. Conversely, uncor-
rected errors are suggestive of poor cognitive control.In the current study, we sought to investigate novel
measures of error making and monitoring in PD patients
with and without FOG. We propose that if the CCNunderpins the pathophysiology of FOG, patients with FOG
should produce more uncorrected errors than patients
without FOG during the most demanding condition of thetask which requires set-shifting. Such a finding would
demonstrate inefficient recruitment of cognitive control
regions under high cognitive load, specifically occurringunder switching demands.
Experimental procedures
Participants
Participants were recruited from a larger cohort of patientsprospectively evaluated at the Parkinson’s Disease
Research Clinic at the Brain and Mind Research Institute,
University of Sydney. The diagnosis of idiopathic PD wasbased on the UK brain bank clinical criteria (Hughes et al.
1992) and was confirmed by a neurologist (SJGL). The 109
patients originally included in this study were divided intotwo groups based on their score on the FOG-Questionnaire
item 3 [FOGQ-3: ‘‘Do you feel that your feet get glued to
the floor while walking, making a turn, or trying to initiatewalking (freezing)?’’], which has been shown to be a
reliable screening tool to identify freezers (Giladi et al.
2000). Higher scores on FOGQ-3 indicate increasedfreezing severity and patients who scored 2 or higher were
included in the FOG group, while those scoring 0 were
placed in the no-FOG group. Patients scoring 1 were notincluded in the study to minimise the possibility of mis-
classification. Exclusion criteria included the presence of
other neurological diseases, other conditions that wouldimpair gait and a score of B24 on the mini mental state
examination (MMSE) (Folstein et al. 1975). All patients
were assessed on their regular medication. This researchwas approved by the Human Research Ethics Committee of
The University of Sydney, and written informed consent
was obtained from all participants.
Clinical assessments
A series of demographic details were recorded, including
age, gender, and predicted premorbid intellectual func-
tioning (revised national adult reading test; NART-R)(Nelson and Willison 1991). Clinical features including
disease duration, Hoehn & Yahr stage (H&Y) (Hoehn and
Yahr 1967), dopamine dose equivalence (DDE; mg/day)(Tomlinson et al. 2010) and disease severity using the
Movement Disorder Society-Unified Parkinson’s Disease
Rating Scale (MDS-UPDRS) (Goetz et al. 2008), of whichthe total score and section III (motor sub-score) were
assessed. Section ‘‘Demographic and clinical data’’ of the
MDS-UPDRS was also individually recorded to ensureverbal responses to the task did not simply reflect problems
with speech output. Additionally, self-reported mood dis-
turbance using the beck depression inventory (BDI-II)(Beck et al. 1996) and global cognitive functioning using
654 C. C. Walton et al.
123

the MMSE (Folstein et al. 1975) were used to assess for
depression and dementia, respectively.
Inhibition and switching tasks
The task used in the current study was the Delis–Kaplan
executive function system (D-KEFS) colour-word interfer-
ence test (Delis et al. 2001). This task contains four condi-tions: colour-naming (condition 1), word-reading
(condition 2), inhibition (condition 3) and inhibition/switching (condition 4). In all conditions, the patient is
asked to read the colours or words aloud depending on the
rule given, as quickly as possible while minimising errors.There are 50 words presented on the page in each condition.
Participants practice on ten word trials prior to the recorded
version to ensure they understand the rules. During the task,participants are prompted if they make three consecutive
errors and reminded of the rules. This occurs only once
during each condition, however. Participants are told theycan correct any errors as they progress through the task.
In condition 1, participants must name aloud the colour
of red, green or blue squares on the page. In condition 2,participants must read through a page containing the words
‘‘red’’, ‘‘green’’ and ‘‘blue’’ printed in black ink. In con-
dition 3, the same words in differing order are presented inincongruent ink colour (e.g., the word ‘‘red’’ appears in
blue ink). The participant is asked to read aloud the colour
of the ink as quickly as possible without making mistakes.This task, therefore, requires the participant to inhibit a
more automatic response (word reading). In condition 4,
the participant is presented with incongruent colour wordsas in the previous condition but a selection of the colour
words are surrounded by a black rectangular outline. For
these selected words, the patient is asked to read the wordwritten and not to name the colour of the ink, thus requiring
them to set-shift in addition to maintaining inhibitory
control (condition 4 requires 37 switches).Outcome measures include the total time taken to
complete each condition (raw and age scaled scores) (Delis
et al. 2001) as well as the number of uncorrected (UC),self-corrected (SC) and total errors. In this study, contrast
scores between conditions were not calculated due to their
known unreliability for this test (Crawford et al. 2008).
Statistical analysis
This study employed a matched-groups sample to address
common critiques of other neuropsychological assessments
of FOG, which do not often account for disease severity(Heremans et al. 2013a). Therefore, cases were removed
prior to task analysis where outliers for descriptive and
clinical variables led to significantly differing groups. Thiswas done in a stepwise fashion, removing patients with
particularly high or low MDS-UPDRS scores until there
was no significant group difference on this variable. Sub-sequently, 34 patients with FOG and 38 without were
retained in the sample, matched on most key demographic
variables (see Table 1). Data analyses were conductedusing the Statistical Package for the Social Sciences (SPSS)
version 21. Due to the non-parametric nature of the task
data, Mann–Whitney U tests were conducted to assess fordifferences between groups. Due to unequal variances for
error scores, the data were additionally analysed by cate-gorising the data and using Pearson’s Chi Square test to
ensure the robustness and reliability of the results. For these
analyses, error scores were split into those who scored 0 or 1error, and those who scored 2 or more errors (multiple
errors). Where the assumption of expected counts was
violated, Fisher’s exact test statistic was used (FET). Wheredata met parametric assumptions for descriptive statistics,
t-tests were used. Wilcoxon signed ranks test was used to
assess within-groups differences across conditions. Allanalyses used an alpha of 0.05 and were two-tailed. The
effect size for differences between groups using Mann–
Whitney U and Wilcoxon signed ranks tests was producedusing the following formula (r = Z/HN) while Cramer’s V
was used for the Chi Square statistic. For both values, 0.10
represents a small, 0.30 a medium, and 0.50 a large effect.One patient with FOG was unable to complete condition 4
despite sufficiently completing the prior three conditions,
and, therefore, a maximum time score was applied inaccordance with standard D-KEFS procedure and their error
scores were not included in the analysis of this condition.
Results
Demographic and clinical data
Table 1 displays the median, range, mean and standarddeviations for demographic and clinical data.
Colour-word interference test
For condition 1 and 2, there were no significant differences
between groups for any measure, suggesting no differencebetween groups in simple reading or naming speed.
Table 2 displays the median, range, mean, standard devi-
ation and effect sizes for condition 3 and 4 representing thecritical trials in the study. For condition 3, there was no
significant difference between groups in completion time
for raw (U = 513.5, Z = -1.50, p = 0.135) or age-scaledtime scores (U = 502, Z = -1.63, p = 0.102). There was
also no significant difference between groups in UC errors
(U = 576, Z = -1.06, p = 0.288). However, there weresignificant differences in SC errors (U = 473.5, Z = -2.04,
Impaired cognitive control in Parkinson’s disease patients 655
123

p = 0.042) and total errors (U = 457, Z = -2.20,
p = 0.028), with patients with FOG producing more. Incondition 4, patients with FOG were significantly slower in
both raw (U = 474, Z = -1.94, p = 0.052) and age-
scaled time scores (U = 470, Z = -2.01, p = 0.045).Patients with FOG also committed significantly more UC
errors than those without FOG (U = 434.5, Z = -2.39,
p = 0.017). However, groups did not differ significantly onSC errors (U = 585.0, Z = -0.50, p = 0.614) or total
errors (U = 507, Z = -1.41, p = 0.158).
In order to confirm these results, a Chi Square test wasused to compare error rates between groups. Again, no
significant differences were found for any measures on
condition 1 and 2. In condition 3, the number of UC errorswas not significantly different between groups (FET (1),
p = 1.00), while patients with FOG produced significantly
more SC [v2 (1) = 5.46, p = 0.019] and total errors [v2
(1) = 6.55, p = 0.010]. In condition 4, participants with
FOG recorded significantly more UC errors [v2 (1) = 3.77,
p = 0.052], while SC [v2 (1) = 0.09, p = 0.761] and totalerrors did not differ [v2 (1) = 0.96, p = 0.327] between
groups. Thus, all previous results were confirmed.
To further explore the pattern of error-making acrossconditions, a within-group analysis using Wilcoxon signed
Table 1 Descriptive and clinical data for patients with and without FOG
FOG No FOG Significance
N 34 38 –
Gender (% male) 68 % 63 % 0.81a
Age (years) 66.44 ± 8.8 66.76 ± 8.1 0.87b
FOGQ-3 3 (2)/2.82 ± 0.8 – –
NART-R 106 (35)/107.88 ± 9.4 115 (55)/112.00 ± 10.5 0.06
MMSE 28 (5)/28.21 ± 1.5 29 (3)/29.05 ± 1.1 0.01
Disease duration (years) 6.54 ± 3.9 5.7 ± 3.6 0.37b
Hoehn & Yahr stage 2 (3)/2.18 ± 0.6 2 (1)/2.08 ± 0.2 0.40
DDE 650 (1820)/723.44 ± 452.9 575 (1508)/605.53 ± 310.4 0.42
MDS-UPDRS total 51.39 ± 20.4 44.79 ± 18.9 0.16b
MDS-UPDRS-III 22 (58)/28.06 ± 15.3 23.5 (68)/26.47 ± 13.0 0.95
MDS-UPDRS-speech 1 (2)/0.75 ± 0.7 1 (4)/0.86 ± 0.9 0.71
BDI-II 9 (30)/11.81 ± 7.4 7 (25)/8.63 ± 6.1 0.04
Scores represent median (range)/mean ± standard deviation
FOGQ-3 Freezing of gait questionnaire-question 3, NART-R revised national adult reading test, MMSE minimental state examination, DDEdopamine dose equivalence, MDS-UPDRS movement disorder society-unified Parkinson’s disease rating scale: total, motor score (III), and part3.1 (speech), BDI-II beck depression inventorya Pearson’s Chi Square test usedb Independent samples t test used
Table 2 Descriptive data andeffect sizes for conditions 3 and4
Scores represent median(range)/mean ± standarddeviationa Independent samples t testusedns p [ 0.05
* p B 0.05
** p B 0.01
Effect size
FOG No FOG Cramer’s V r
Condition 3 (Inhibition)
Time 69 (130)/75.50 ± 27.9 61 (126)/68.11 ± 27.1 – 0.18
Age scaled score 9.53 ± 3.5 10.66 ± 3.6 – 0.07ns,a
Uncorrected errors 0 (12)/0.85 ± 2.3 0 (3)/0.37 ± 0.9 0.02ns 0.12ns
Self-corrected Errors 1 (13)/2.15 ± 2.9 1 (4)/0.92 ± 1.2 0.28* 0.24*
Total errors 2 (14)/3.00 ± 4.0 1 (6)/1.29 ± 1.6 0.30** 0.26*
Condition 4 (Switching)
Time 74.50 (142)/86.21 ± 33.3 64.5 (96)/74.66 ± 25.9 – 0.23*
Age scaled score 8.97 ± 4.0 10.42 ± 3.5 – 0.24*,a
Uncorrected errors 1 (11)/2.18 ± 3.0 0 (7)/0.87 ± 1.6 0.23* 0.28*
Self-corrected errors 1 (5)/1.42 ± 1.4 1 (8)/1.29 ± 1.5 0.04ns 0.06ns
Total errors 2 (14)/3.61 ± 3.9 2 (9)/2.16 ± 2.4 0.12ns 0.17ns
656 C. C. Walton et al.
123

ranks test was conducted. As shown in Fig. 1, there was a
medium to strong sized significant difference in UC errorsfor patients with FOG between conditions 3 and 4 (Z =
-2.89, p = 0.004, r = 0.38). Conversely, UC errors for
patients without FOG (Z = -1.61, p = 0.108), and the SCerrors for those with (Z = -1.47, p = 0.143) and without
FOG (Z = -1.20, p = 0.230) between conditions were
not significant. Both patients with FOG (Z = -2.20,p = 0.028) and those without FOG (Z = -2.15,
p = 0.031) showed significantly more total errors incondition 4.
Discussion
This study presents the first investigation into self-moni-toring of conflict-induced errors in FOG under differing
levels of cognitive load. The key finding was that the
introduction of switching demands to the task impairedperformance in patients with FOG, particularly in their UC
error making. More precisely, in condition 3, freezers
produced significantly more errors than non-freezers(automatic responding), but also demonstrated the cogni-
tive control necessary to self-correct immediately after
responding. However, when the additional switching ele-ment was introduced in condition 4, patients with FOG
displayed a large and significant increase in UC errors
while also being less likely to self-correct. These novelresults suggest that in the presence of intensified cognitive
load induced by switching, patients with FOG may no
longer be able to recruit appropriate monitoring mecha-nisms to adjust response selection (Brown 2013; Lewis and
Barker 2009). This finding of a threshold-crossing is sim-
ilar to that of PD in general when compared to healthycontrols (Brown and Marsden 1988).
This finding is consistent with the understanding that
patients with FOG have deficits in both conflict interfer-ence (Cohen et al. 2014; Matar et al. 2013; Vandenbossche
et al. 2011, 2012) and switching ability (Naismith et al.2010; Shine et al. 2013e). However, the current results
extend this understanding by further exploring the precise
inability of patients with FOG to track and adapt theirresponses under increased cognitive demand (combination
of conflict interference and switching). This raises impor-
tant questions regarding the underlying anatomical regionsassociated with performance monitoring and how these
may be related to pathophysiological mechanisms under-
lying FOG.Understanding the processes involved in error monitor-
ing is complex. For example, separating differing aspects
such as monitoring when aware and/or unaware of errors,which may be self-made and/or external (incongruent
stimuli) in nature, in addition to the subsequent detection
and correction of these errors is extremely complex, andnot well understood (Orr and Hester 2012; Wessel 2012).
Generally, the process of detection and correction of errors
has been shown to involve the anterior cingulate, medialfrontal, posterior parietal, pre-supplementary motor area
(pSMA) and insular cortices (Garavan et al. 2003; Klein
et al. 2007; Narayanan et al. 2013; Orr and Hester 2012;Ridderinkhof et al. 2004; Sharp et al. 2010). However, the
precise mechanism for how these areas interact in error
detection and correction remains unclear (Narayanan et al.2013). These cortical regions are known to commonly co-
activate as part of a large-scale network (Niendam et al.
2012), and this network is thought to be involved in theprocessing of response conflict and the effective switching
of attention during cognitively demanding tasks (Wager
et al. 2004). Additionally, sub-cortical circuitry has alsobeen associated with cognitive demands of inhibitory and
switching control, with the STN known to play key roles in
such processes (Aron and Poldrack 2006; Frank 2006),including in PD patients (Alegre et al. 2013; Brittain et al.
2012; Obeso et al. 2014).
Interestingly, dysfunction within these regions is con-sistent with findings from recent functional neuroimaging
experiments demonstrating impaired processing through
the pSMA, anterior insula, ventral striatum and STN duringcognitive processing in patients with FOG (Shine et al.
2013b). Furthermore, an additional study showed that
episodes of freezing were associated with increased activ-ity within the fronto-parietal regions which are required to
Fig. 1 Mean error scores across conditions 3 and 4 for patients withand without FOG. Error bars represent 1 SE. * p B 0.05, ** p B 0.01
Impaired cognitive control in Parkinson’s disease patients 657
123

perform goal-directed behaviours (Shine et al. 2013a).
Expanding on these results, it has since been shown that inFOG, there is a paroxysmal functional decoupling of
activity between multiple neural networks, including the
basal ganglia and CCN, and cross-talk between left andright portions of the CCN (Shine et al. 2013c). Together,
these studies suggest that patients with FOG may require
activation of more cognitive resources to effectively per-form challenging tasks, leaving less reserve available for
the ongoing monitoring of errors. This suggestion is similarto that proposed of Pieruccini-Faria et al. (2014) who found
cognitive overload to impede on complex motor planning
in patients with FOG and that these deficits were associatedwith executive dysfunction.
Alternatively, the inability to properly recruit appropri-
ate cortical regions during high cognitive load may be dueto the role of the STN. One hypothesis is that as cognitive
load increases in response to these executive tasks, it drives
an overwhelming increase in STN firing through the hyper-direct pathway (Aron and Poldrack 2006), leading to
increased inhibition of basal ganglia output nuclei to the
thalamus. Although the hyper-direct pathway of the basalganglia was originally hypothesised as important for purely
motor cessation (Nambu et al. 2002), recent work has
implicated this network in stopping behaviour in executivedomains (Aron, Robbins and Poldrack 2014; Haynes and
Haber 2013). As such, overactivity in the executive regions
of this hyper-direct pathway may result in thalamo-corticalinhibition, which would then mean that error-monitoring
hubs, including areas of medial and prefrontal cortex, are
unable to process information efficiently during periods ofhigh cognitive load (DeLong and Wichmann 2007). This
would be reflected in increased uncorrected errors, as
shown in the current study.
Conclusions and future directions
Further research employing behavioural and neuroimaging
techniques should be employed to help disentangle these
preliminary interpretations. In future studies, the role ofswitching and inhibitory control along with the influence of
cognitive load could be manipulated using novel paradigms
to provide additional insight into these processes and howthey may relate to such functional networks. The current
findings may have implications for understanding the role
of performance monitoring during gait (Amboni et al.2013; Cohen et al. 2014). For example, the current results
may well relate to the impaired ability of patients to
appropriately monitor and plan gait while dealing withcognitive overload in overwhelming situations (Pieruccini-
Faria et al. 2014). Virtual reality studies may be employed
for this goal (Mirelman et al. 2013). Finally, the current
findings help advise on specific targets for cognitive
training in patients with FOG (Walton et al. 2014).Overall, these novel findings demonstrate that in
patients with FOG, there is a specific deficit in monitoring
for self-made errors under high cognitive load. They raiseimportant questions regarding the underlying anatomical
regions associated with performance monitoring and the
pathophysiological mechanisms underlying FOG. Futurestudies are needed to untangle this complex area of cog-
nitive control in patients with FOG.
Acknowledgments The authors wish to thank Dr. Dafydd Llewelynfor his contribution to data collection that made up part of this study.Additionally, we thank the patients of the Parkinson’s DiseaseResearch Clinic for being so generous with their time and efforts.
Financial disclosures CC Walton is supported by an AustralianPostgraduate Award at the University of Sydney. M Gilat is supportedby an International Postgraduate Scholarship at the University ofSydney. C O’Callaghan is supported by an Alzheimer’s Australia PhDScholarship at the University of New South Wales. SL Naismith issupported by a National Health and Medical Research Council CareerDevelopment Award No. 1008117. SJG Lewis is supported by aNational Health and Medical Research Council Practitioner Fellow-ship No. 1003007. JM Shine, L Mowszowski, JM Hall have nofinancial disclosures.
Conflict of interest The authors report no Conflicts of interest.
References
Alegre M et al (2013) The subthalamic nucleus is involved insuccessful inhibition in the stop-signal task: a local fieldpotential study in Parkinson’s disease. Exp Neurol 239:1–12.doi:10.1016/j.expneurol.2012.08.027
Amboni M, Cozzolino A, Longo K, Picillo M, Barone P (2008)Freezing of gait and executive functions in patients withParkinson’s disease. Mov Disord 23:395–400. doi:10.1002/mds.21850
Amboni M, Barone P, Picillo M, Cozzolino A, Longo K, Erro R,Iavarone A (2010) A two-year follow-up study of executivedysfunctions in parkinsonian patients with freezing of gait at on-state. Mov Disord 25:800–802. doi:10.1002/mds.23033
Amboni M, Barone P, Hausdorff JM (2013) Cognitive contributionsto gait and falls: evidence and implications. Mov Disord28:1520–1533. doi:10.1002/mds.25674
Aron AR, Poldrack RA (2006) Cortical and subcortical contributionsto Stop signal response inhibition: role of the subthalamicnucleus. J Neurosci 26:2424–2433. doi:10.1523/JNEUROSCI.4682-05.2006
Aron AR, Robbins TW, Poldrack RA (2014) Inhibition and the rightinferior frontal cortex: one decade on. Trends Cogn Neurosci18(4):177–185. doi:10.1016/j.tics.2013.12.003
Beck A, Steer R, Brown G (1996) Manual for the BDI-II.Psychological Corporation, San Antonio
Brittain JS et al (2012) A role for the subthalamic nucleus in responseinhibition during conflict. J Neurosci 32:13396–13401. doi:10.1523/JNEUROSCI.2259-12.2012
Brown JW (2013) Beyond conflict monitoring: cognitive control andthe neural basis of thinking before you act. Curr Dir Psychol Sci22:179–185. doi:10.1177/0963721412470685
658 C. C. Walton et al.
123

Brown RG, Marsden CD (1988) Internal versus external cues and thecontrol of attention in Parkinson’s disease brain. J Neurol 111(Pt2):323–345
Cohen RG et al (2014) Inhibition, executive function, and freezing ofgait. J Parkinson’s Dis 4:111–122. doi:10.3233/JPD-130221
Crawford JR, Sutherland D, Garthwaite PH (2008) On the reliabilityand standard errors of measurement of contrast measures fromthe D-KEFS. J Int Neuropsychol 14:1069–1073. doi:10.1017/S1355617708081228
Delis DC, Kaplan E, Kramer JH (2001) Delis-Kaplan executivefunction system (D-KEFS). Psychological Corporation, SanAntonio, TX
DeLong MR, Wichmann T (2007) Circuits and circuit disorders of thebasal ganglia. Arch Neurol 64:20–24. doi:10.1001/archneur.64.1.20
Folstein MF, Folstein SE, McHugh PR (1975) Mini-mental state Apractical method for grading the cognitive state of patients forthe clinician. J Psychiatr Res 12:189–198
Frank MJ (2006) Hold your horses: a dynamic computational role forthe subthalamic nucleus in decision making. Neural Netw19:1120–1136. doi:10.1016/j.neunet.2006.03.006
Garavan H, Ross TJ, Kaufman J, Stein EA (2003) A midlinedissociation between error-processing and response-conflictmonitoring. Neuroimage 20:1132–1139. doi:10.1016/S1053-8119(03)00334-3
Giladi N, Shabtai H, Simon ES, Biran S, Tal J, Korczyn AD (2000)Construction of freezing of gait questionnaire for patients withParkinsonism. Parkinsonism Relat Disord 6:165–170
Giladi N et al (2001) Freezing of gait in patients with advancedParkinson’s disease. J Neural Transm 108:53–61
Goetz CG et al (2008) Movement Disorder Society-sponsoredrevision of the unified Parkinson’s disease rating scale (MDS-UPDRS): scale presentation and clinimetric testing results. MovDisord 23:2129–2170. doi:10.1002/mds.22340
Gray P, Hildebrand K (2000) Fall risk factors in Parkinson’s disease.J Neurosci Nurs 32:222–228
Hall JM, Shine JM, Walton CC, Gilat M, Kamsma YP, Naismith SL,Lewis SJ (2014) Early phenotypic differences between Parkin-son’s disease patients with and without freezing of gait. Parkin-sonism Relat Disord. doi:10.1016/j.parkreldis.2014.02.028
Haynes WI, Haber SN (2013) The organization of prefrontal-subthalamic inputs in primates provides an anatomical substratefor both functional specificity and integration: implications forBasal Ganglia models and deep brain stimulation. J Neurosci33:4804–4814. doi:10.1523/JNEUROSCI.4674.12.2013
Heremans E et al (2013a) Cognitive aspects of freezing of gait inParkinson’s disease: a challenge for rehabilitation. J NeurTransm 120:543–557. doi:10.1007/s00702-012-0964-y
Heremans E, Nieuwboer A, Vercruysse S (2013b) Freezing of gait inParkinson’s disease: where are we now? Curr Neurol NeurosciRep 13:350. doi:10.1007/s11910-013-0350-7
Hoehn MM, Yahr MD (1967) Parkinsonism: onset, progression andmortality. Neurology 17:427–442
Hughes AJ, Daniel SE, Kilford L, Lees AJ (1992) Accuracy ofclinical diagnosis of idiopathic Parkinson’s disease: a clinico-pathological study of 100 cases. J Neurol Neurosurg Psychiatry55:181–184
Klein TA, Endrass T, Kathmann N, Neumann J, von Cramon DY,Ullsperger M (2007) Neural correlates of error awareness. Neuro-image 34:1774–1781. doi:10.1016/j.neuroimage.2006.11.014
Kornblum S, Hasbroucq T, Osman A (1990) Dimensional overlap:cognitive basis for stimulus-response compatibility—a modeland taxonomy. Psychol Rev 97:253–270
Lewis SJ, Barker RA (2009) A pathophysiological model of freezingof gait in Parkinson’s disease. Parkinsonism Relat Disord15:333–338. doi:10.1016/j.parkreldis.2008.08.006
Matar E, Shine JM, Naismith SL, Lewis SJ (2013) Using virtual realityto explore the role of conflict resolution and environmentalsalience in Freezing of Gait in Parkinson’s disease. ParkinsonismRelat Disord. doi:10.1016/j.parkreldis.2013.06.002
Mirelman A, Maidan I, Deutsch JE (2013) Virtual reality and motorimagery: promising tools for assessment and therapy in Parkin-son’s disease. Mov Disord 28:1597–1608. doi:10.1002/mds.25670
Moore O, Peretz C, Giladi N (2007) Freezing of gait affects quality oflife of peoples with Parkinson’s disease beyond its relationshipswith mobility and gait. Mov Disord 22:2192–2195. doi:10.1002/mds.21659
Naismith SL, Shine JM, Lewis SJ (2010) The specific contributions ofset-shifting to freezing of gait in Parkinson’s disease. MovDisord 25:1000–1004. doi:10.1002/mds.23005
Nambu A, Tokuno H, Takada M (2002) Functional significance of thecortico-subthalamo-pallidal ‘hyperdirect’ pathway. Neurosci Res43:111–117
Narayanan NS, Cavanagh JF, Frank MJ, Laubach M (2013) Commonmedial frontal mechanisms of adaptive control in humans androdents. Nat Neurosci 16:1888–1895. doi:10.1038/nn.3549
Nelson H, Willison J (1991) The revised national adult reading test–test manual Windsor: NFER-Nelson
Niendam TA, Laird AR, Ray KL, Dean YM, Glahn DC, Carter CS(2012) Meta-analytic evidence for a superordinate cognitivecontrol network subserving diverse executive functions. CognAffect Behav Neurosci 12:241–268. doi:10.3758/s13415-011-0083-5
Nutt JG, Bloem BR, Giladi N, Hallett M, Horak FB, Nieuwboer A(2011) Freezing of gait: moving forward on a mysterious clinicalphenomenon. Lancet Neurol 10:734–744. doi:10.1016/S1474-4422(11)70143-0
Obeso I et al (2014) The subthalamic nucleus and inhibitory control:impact of subthalamotomy in Parkinson’s disease. Brain137:1470–1480. doi:10.1093/brain/awu058
Orr C, Hester R (2012) Error-related anterior cingulate cortex activityand the prediction of conscious error awareness. Front HumNeurosci 6:177. doi:10.3389/fnhum.2012.00177
Pieruccini-Faria F, Jones JA, Almeida QJ (2014) Motor planning inParkinson’s disease patients experiencing freezing of gait: theinfluence of cognitive load when approaching obstacles. BrainCogn 87:76–85. doi:10.1016/j.bandc.2014.03.005
Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S (2004)The role of the medial frontal cortex in cognitive control.Science 306:443–447. doi:10.1126/science.1100301
SharpDJ, Bonnelle V,DeBoissezonX, BeckmannCF, James SG, PatelMC, Mehta MA (2010) Distinct frontal systems for responseinhibition, attentional capture, and error processing. Proc NatlAcad Sci USA 107:6106–6111. doi:10.1073/pnas.1000175107
Shine JM et al (2013a) Exploring the cortical and subcorticalfunctional magnetic resonance imaging changes associated withfreezing in Parkinson’s disease. Brain 136:1204–1215. doi:10.1093/brain/awt049
Shine JM, Matar E, Ward PB, Bolitho SJ, Pearson M, Naismith SL,Lewis SJ (2013b) Differential neural activation patterns inpatients with Parkinson’s disease and freezing of gait in responseto concurrent cognitive and motor load. PloS One 8:e52602.doi:10.1371/journal.pone.0052602
Shine JM et al (2013c) Freezing of gait in Parkinson’s disease isassociated with functional decoupling between the cognitivecontrol network and the basal ganglia. Brain 136:3671–3681.doi:10.1093/brain/awt272
Shine JM, Moustafa AA, Matar E, Frank MJ, Lewis SJ (2013d) Therole of frontostriatal impairment in freezing of gait in Parkin-son’s disease Frontiers in Systems Neuroscience 7 doi:10.3389/fnsys.2013.00061
Impaired cognitive control in Parkinson’s disease patients 659
123

Shine JM, Naismith SL, Palavra NC, Lewis SJ, Moore ST, Dilda V,Morris TR (2013e) Attentional set-shifting deficits correlate withthe severity of freezing of gait in Parkinson’s disease. Parkin-sonism Relat Disord 19:388–390. doi:10.1016/j.parkreldis.2012.07.015
Tomlinson CL, Stowe R, Patel S, Rick C, Gray R, Clarke CE (2010)Systematic review of levodopa dose equivalency reporting inParkinson’s disease. Mov Disord 25:2649–2653. doi:10.1002/mds.23429
Vandenbossche J, Deroost N, Soetens E, Spildooren J, Vercruysse S,Nieuwboer A, Kerckhofs E (2011) Freezing of gait in Parkinsondisease is associated with impaired conflict resolution. Neuro-rehabil Neural Repair 25:765–773. doi:10.1177/1545968311403493
Vandenbossche J et al (2012) Conflict and freezing of gait inParkinson’s disease: support for a response control deficit.Neuroscience 206:144–154. doi:10.1016/j.neuroscience.2011.12.048
Vandenbossche J et al (2013a) Freezing of gait in Parkinson’s disease:disturbances in automaticity and control. Front Hum Neurosci6:356. doi:10.3389/fnhum.2012.00356
Vandenbossche J et al (2013b) Impaired implicit sequence learning inParkinson’s disease patients with freezing of gait. Neuropsy-chology 27:28–36. doi:10.1037/a0031278
Wager TD, Jonides J, Reading S (2004) Neuroimaging studies ofshifting attention: a meta-analysis. NeuroImage 22:1679–1693.doi:10.1016/j.neuroimage.2004.03.052
Walton CC, Shine JM, Mowszowski L, Naismith SL, Lewis SJ (2014)Freezing of gait in Parkinson’s disease: current treatments andthe potential role for cognitive training. Restor NeurologyNeurosci 32:411–422. doi:10.3233/rnn-130370
Wessel JR (2012) Error awareness and the error-related negativity:evaluating the first decade of evidence. Front Hum Neurosci6:88. doi:10.3389/fnhum.2012.00088
660 C. C. Walton et al.
123

APPENDIX(3((
Gilat, M., Shine, J. M., Walton, C. C., O’Callaghan, C., Hall, J. M., Lewis, S. J. G.
Brain activation underlying turning in Parkinson’s disease patients with and without
Freezing of Gait. NPJ Parkinson’s Disease, 2015
!

ARTICLE OPEN
Brain activation underlying turning in Parkinson’s diseasepatients with and without freezing of gait: a virtual realityfMRI studyMoran Gilat1, James M Shine1,2, Courtney C Walton1, Claire O’Callaghan1,3, Julie M Hall1,4 and Simon JG Lewis1
BACKGROUND: Freezing of gait is a debilitating symptom affecting many patients with Parkinson’s disease (PD), causing severeimmobility and decreased quality of life. Turning is known to be the most common trigger for freezing and also causes the highestrates of falls. However, the pathophysiological basis for these effects is not well understood.METHODS: This study used a virtual reality paradigm in combination with functional magnetic resonance imaging to explore theneural correlates underlying turning in 17 PD patients with freezing of gait (FOG) and 10 PD patients without FOG while off theirdopaminergic medication. Participants used foot pedals to navigate a virtual environment, which allowed for blood oxygen level-dependent (BOLD) responses and footstep latencies to be compared between periods of straight “walking” and periods of turningthrough 90°. BOLD data were then analyzed using a mixed effects analysis.RESULTS: Within group similarities revealed that overall, PD patients with freezing relied heavily on cortical control to enableeffective stepping with increased visual cortex activation during turning. Between groups differences showed that when turning,patients with freezing preferentially activated inferior frontal regions that have been implicated in the recruitment of a putativestopping network. In addition, freezers failed to activate premotor and superior parietal cortices. Finally, increased task-basedfunctional connectivity was found in subcortical regions associated with gait and stopping within the freezers group during turning.CONCLUSIONS: These findings suggest that an increased propensity towards stopping in combination with reduced sensorimotorintegration may underlie the neurobiology of freezing of gait during turning.
npj Parkinson's Disease (2015) 1, 15020; doi:10.1038/npjparkd.2015.20; published online 22 October 2015
INTRODUCTIONTurning is an integral yet complex task of daily mobility thatcommonly precipitates falls in the elderly population.1 This effectis greater in Parkinson’s disease (PD),2 significantly increasing therisk of falls and related injuries, such as hip fractures1 leading tonursing home placement.One reason for the increased incidence of falls in PD is known to
be freezing of gait (FOG), which is described as a brief, episodicabsence or marked reduction of forward progression of the feetdespite the intention to walk.3 Importantly, turning is recognizedto be the most frequent trigger of this phenomenon.4 Thisdebilitating symptom impacts around half of PD patients, causingregular falls and a decreased quality of life (for review see Nuttet al.).3
Behavioral measures of turning difficulties and their associationwith FOG have been widely studied. For instance, patients withFOG turn more slowly, take more steps, are more variable in theirstep times and implement a different turning strategy whencompared with PD patients without FOG and healthy controls.2,5
Turning difficulties in PD patients with FOG are only partlyimproved by dopaminergic medication2 and any ameliorationachieved through cueing has only a short lasting carry over effectafter cue removal.6
Due to difficulties inherent in the neuroimaging of gait, thepathophysiological mechanisms linking turning and freezing are
currently poorly understood, limiting our ability to developadequate therapeutic interventions. Recent insights have beengained from the effects of deep brain stimulation in thesubthalamic nucleus (STN-DBS)7 and saccadic functioning8 inParkinson’s disease. The STN, with its striatal, cerebellar and hyper-direct supplementary motor and other frontal cortexconnections,9–11 is thought to be involved in a common neuralpathway underlying FOG causing inhibition of the gait relatedsubcortical structures.12,13 As turning is a provocative trigger forFOG, one might predict that abnormal STN activation by itself, orindirectly through abnormal activation in its cerebellar, striatal andfrontal cortex connections is at least in part responsible for thedeficits in turning kinematics seen in PD patients with FOG.Indeed, Lohnes and Earhart7 showed that STN-DBS in PD patientsshortened their turn duration, whereas it also improved saccadicfunctions that are important for turning.7 STN-DBS has also beenshown to improve visuospatial attention14 and decrease interseg-mental latencies (e.g., eye–head, eye–foot, and head–trunk), whichare both affected in PD patients, especially in those thatexperience freezing.7,15 However, though these studies providevaluable information regarding the STN’s role, little informationexists about other regions of the brain that are likely to beinvolved in the functional impairments that cause turningdifficulties and FOG in PD patients.12
1Parkinson’s Disease Research Clinic, Brain and Mind Research Institute, The University of Sydney, Sydney, NSW, Australia; 2Department of Psychology, Stanford University,Stanford, CA, USA; 3Department of Psychology, Behavioural and Clinical Neuroscience Institute, University of Cambridge, Cambridge, UK and 4School of Social Sciences andPsychology, University of Western Sydney, Sydney, NSW, Australia.Correspondence: SJG Lewis ([email protected])Received 21 April 2015; revised 21 July 2015; accepted 24 August 2015
www.nature.com/npjparkdAll rights reserved 2373-8057/15
© 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited

It is therefore clear that novel paradigms are needed to improveour understanding of the pathophysiology underlying turningdeficits in Parkinson’s disease, especially when exploring freezingof gait. As such, the current study set out to investigate the effectof turning during the performance of an interactive virtual reality(VR) paradigm that has previously been used in combination withfunctional magnetic resonance imaging (fMRI) to investigate thepathophysiology underlying behavioral freezing episodes13 andthe effects of other known triggers of FOG, such as high cognitiveload.16,17 However, the current study will be the first to combinean adjusted version of the VR with fMRI to explore the widespreadbrain regions that underpin turning behavior in Parkinson’sdisease patients with and without freezing of gait. The currentVR paradigm allows for the investigation of complex sensorimotorintegration during turning, as it requires subjects to generateeffective lower limb motor output while updating changes in theirvisual environment. We hypothesized that PD patients with FOGwould be slower and more variable in their step times during thenavigation of a turn in the VR18,19 and that turning would elicitaltered activation patterns in the STN7 and its hyperdirect frontalcortex connections.9–11 In addition, we expected to find differ-ences across the cortical, striatal and other subcortical regions thathave previously been identified in PD patients with FOG duringstructural MRI,11,20 fMRI resting state,21 PET gait imagery tasks22
and during fMRI with a gait imagery tasks,23,24 upper limb motortask,25 and VR performance.13,16,17
MATERIALS AND METHODSPatient details and study protocolA total of 27 levodopa-responsive patients with idiopathic PD wererecruited from the Parkinson’s Disease Research Clinic, Brain and MindResearch Institute, the University of Sydney having satisfied UKPDS BrainBank clinical diagnostic criteria. Seventeen Parkinson’s disease patientswith freezing of gait (PD+FOG) were selected based on a previouslyobtained positive score on the Freezing of Gait Questionnaire question 3(FOG-Q3)26 (“do you feel that your feet get glued to the floor whilewalking, making a turn or when trying to initiate walking (freezing)?”) and10 non-freezing Parkinson’s disease− patients (PD−NF) were selectedbased on a null score on this question. All patients underwent neurologicaland neuropsychological assessment, completed a gait protocol andperformed an fMRI scanning session in their practically defined ‘off’ state,having been withdrawn from dopaminergic medication overnight formore than 12 h before testing. The patients in the current study were
measured “off” their dopaminergic medications to increase the likelihoodof eliciting freezing like behaviors in a standardized environment, to allowus to better investigate dopamine dependent basal ganglia dysfunctionsand finally, to enable fair comparisons with previous fMRI studies.13,23,24
Ethical approval for this study was obtained from the University of SydneyHuman Research Ethics Committee and written informed consent wasobtained from each patient.
Neurological and cognitive assessmentAll patients were assessed on the motor section of the Unified Parkinson’sDisease Rating Scale (UPDRS-III) and Hoehn and Yahr Stage in their “off”state. In addition, the Mini Mental State Examination, Montreal CognitiveAssessment, and Hospital Anxiety and Depression Scale (HADS) wereobtained and their daily Dopamine Dose Equivalency was calculated.
Gait protocolEach subject completed eight video recorded 5-m long timed up and gotasks, during which each turn was performed inside a 50-cm2 taped box onthe ground. The timed up and go tasks incorporated 180 and 540° turns inboth directions, taking short steps around the outline of the box in bothdirections and two vocal dual tasks (i.e., naming multiples of nine andmonths of the year backwards) with 180° turns in both directions. Thevideos were scored offline for periods of freezing as defined by a brief,episodic absence or marked reduction of forward progression of the feetdespite the intention to walk.3 The timed up and go tasks clinicallyconfirmed FOG in all but one subjects in the PD+FOG group, whereas noneof the PD−NF patients experienced any freezing episode. The one subjectin the PD+FOG group was still included into the study based on a positivescore on the FOG-Q3 and UPDRS question 3.11 and because he was seenby an experienced physician to have experienced freezing when arrivinginto the clinic.
Virtual reality paradigmPatients performed the VR while lying inside the fMRI scanner. The tasktook ~ 6min to complete and was presented on a screen that could beclearly viewed via a mirror mounted onto the head coil. The virtualenvironment was a three-dimensional corridor presented in thefirst-person. Forward progression through this corridor was accomplishedby alternately depressing left and right foot pedals at least 30° belowparallel in a “physiological” sequence (e.g., left–right). Out of sequencesteps (left–left or right–right) did not result in forward progression andwere disregarded from the analyses. Patients were instructed to tap thepedals in a comfortable rhythm. The VR only contained turns and simple“STOP” cues presented in the color red, followed by a simple “WALK” cuespresented in the color green (Figure 1), which were added to ensure thatthe patients were still paying attention to the task. No other environmentaltriggers (e.g., doorways) or complex cognitive cues were presented duringthis experiment in distinction to our previous reports.13 The turns in the VRwere 90° and randomly presented in both directions (Figure 1). An averageof 23 turns were presented during the trial and patients had to takebetween 3 and 6 steps to complete a turn, based on their steppinglatencies. The VR automatically presented the navigation of a turn as areaction to the physiological sequence of foot pedal depressions. Wechose to leave out an additional motor task to prevent dual tasking frominducing any freezing.27 As such, no difference in behavioral motoractivation was required between periods of straight walking and periods ofturning, except for potential eye movements induced by the updating ofvisual information as the turn was presented on the screen. Similar toprevious studies,2,24 no distinction was made between left and right turnsin order to increase the power of the analyses.
Behavioral measuresFootstep latencies were calculated by measuring the time between twoconsecutive foot pedal depressions. The mean and standard deviationwere then used to calculate the coefficient of variation for the three stepsthat were taken during a turn and for three randomly selected steps thatwere unrelated to any turn or “STOP” and “WALK” cues (hereafter definedas “walking”). In addition, we calculated the longest footstep latency inthose three steps (defined as the maximum footstep latency). Anymaximum footstep latency that was greater than twice the modal footsteplatency was considered to be a behavioral freezing episode, as described inmore detail elsewhere.13,19 All behavioral freezing episodes were removed
Figure 1. Representation of the virtual reality task showing a turn, aperiod of virtual reality walking and a WALK and STOP cue. WALK,walking.
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
2
npj Parkinson's Disease (2015) 15020 © 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited

from the current analyses to ensure that any results in this study wereowing to the effect of turning and were not being driven by theoccurrence of any freezing episodes.17
NeuroimagingEvent related analysis. The image acquisition and image preprocessingsteps are described elsewhere.17 Individual first-level spatial maps werecreated in Statistical parametric mapping software (SPM8, Wellcome TrustCentre for Neuroimaging, London, UK, http://www.fil.ion.ucl.ac.uk/spm/software/) using a general linear model analysis within an epoch-relateddesign in a fixed-effects analysis. A design matrix was created for eachpatient by entering two regressors for each trial, namely, a regressor thatmodeled the specific onset times and associated temporal derivatives foreach turn and a regressor that similarly modeled periods of walking. Thewalking epochs were randomly selected and scaled to the number ofturning epochs, both covering the total duration of the task to control forpossible effects of fatigue. All patients were instructed to minimize headmotion by only moving the ankles, while not raising the legs andpreventing hip rotation. In addition, a brief trial run was performed insidethe scanner before the start of the task. This allowed a researcher to adjustthe position of the patient’s feet and give additional instructions ifextensive head motion was detected. This, together with the placement ofcushions inside the head coil ensured optimal performance with the leastamount of head motion. After data collection, any trial with 43-mm headmotion was excluded from the analyses and six motion and nuisanceregressors were added into the first level analysis per subject, controllingfor movement artifacts in the three directions of translation and axes ofrotation. Contrast images from the first-level analyses were then enteredinto a second-level random-effects independent samples t-test designanalysis to determine the group differences on the contrast of interest(turning4walking). This contrast was chosen as it minimizes thedifferences between the two conditions, both requiring bi-pedaling motoroutput while watching a screen, with the only difference being goingthrough a turn. It therefore controls for the variance associated withbi-pedaling motor output and watching a screen, while allowing theresultant brain activation pattern to be interpreted as the effectsassociated with turning. HADS anxiety, HADS depression and MontrealCognitive Assessment scores were entered as covariates at thesecond level. Whole brain voxel maps were displayed using XjView(www.alivelearn.net/xjview) software (Po0.005). To decrease the risk oftype-II error, we used a large cluster size threshold (k420 voxels).28
Region of interest analysis. Spherical 8-mm regions of interest (ROI) weredrawn around the peak voxels from the second level T-map by using theMarsBar toolbox in SPM8 (ref. 29). The peak voxel values and theircoordinates are presented in Table 2. Importantly, these values were notused for any further statistical inference, but instead allowed us to furtherexplore the linear direction of blood oxygen level-dependent (BOLD)response patterns found in the whole brain analysis, as describedelsewhere.17
In addition, previous studies have implicated key striatal and subcorticalregions in turning and behavioral freezing in patients with Parkinson’sdisease.7,11,23,24 As such, we subsequently explored the images from thefirst-level analysis using predefined regions of interest, which wereanalyzed independently from the whole brain analyses. Spherical ROI’swere drawn around the following left and right striatal regions: caudatenucleus, putamen and ventral striatum, and subcortical regions: mesence-phalic locomotor regions (MLR), globus pallidus internus (GPi), STN and thebilateral cerebellar locomotor region (CLR; see Supplementary Table 1 forcoordinates in Montreal Neurological Institute (MNI) space). MarsBar29 wasused to extract percent signal change values for each region and adifference score was calculated between periods of turning and periods ofwalking. Two-sided independent sampled t-tests were performed on thegroup level and paired sampled t-tests were used within groups. Alphalevels were set to 0.05.
Task-based functional connectivity. On the basis of current perceptionsthat FOG is likely due to functional network deficits,12,16 we aimed toexplore the task based functional connectivity patterns30 associated withturning in the VR. As such, the Marsbar toolbox was also used to extractraw ROI data (beta weights; or β) of each ROI for each patient.MATLAB (The MathWorks Inc., Natick, MA, USA) was used to calculatethe temporal derivative (TD= β (Tn)—β (Tn-1)) of the raw β weights for eachROI. After scaling each time course by its variance, the temporal derivatives
of each ROI were multiplied by the temporal derivatives of the other ROI’sfor each time point, such that a positive score reflected ‘functionalcoupling’ between a pair of ROIs. For each subject, we multiplied thefunctional coupling score for each temporal derivative by the convolvedtime points associated with either turning or walking periods in the virtualreality task. We then calculated the non-zero average for each ROI pair forboth contrasts for each patient. Paired sampled t-tests were employed toanalyze the differences in these non-zero average scores between periodsof turning and periods of walking. Finally, the non-zero average scoreswere organized into two 13-by-13 matrices for each subject, one for theperiods of turning and one for the periods of walking for each of the 13ROI’s. These matrices were then compared statistically using the NetworkBased Statistics Toolbox31 with a threshold value of 3.0, Po0.05 as well asthe False Detection Rate option (Po0.05) in the network-based statisticssoftware to control for multiple comparisons.
RESULTSPatient demographicsThe demographic statistics, results of the gait assessments and thebehavioral measures from the VR are presented in Table 1. Thegroups were matched for key demographics such as age, diseaseduration, dopamine dose equivalency, mini mental state examina-tion scores, disease severity and motor severity (UPDRS-III) afterremoving the gait and freezing items. Moreover, both groupsincluded more males (χ2(1) = 0.764, P= 0.382). As is commonlyreported, the PD+FOG group did have significantly lower MontrealCognitive Assessment scores and higher HADS anxiety and HADSdepression scores (Table 1).17 The current study thereforecontrolled for the significant group differences in MontrealCognitive Assessment and HADS scores through the use ofcovariate analyses. Additional non-parametric analyses revealedthat the groups were also matched for Hoehn and Yahr stages andthat only the PD+FOG group scored positively on the FOG-Q3(Table 1). Finally, 53% of the PD+FOG group and 60% of thePD−NF group (χ2(1) = 0.127, P= 0.722) had worse Parkinson’sdisease symptoms on the left side of the body as calculated by aratio of the sum of UPDRS-III items related to symptom severity onthe right side and left side of the body. Finally, no significantdifferences were found between the groups on handedness(χ2(1) = 1.27, P=0.260), foot tapping abilities and leg agility asobtained by the UPDRS questions 3.7 and 3.8, respectively (Table 1).In addition, toe tapping and leg agility scores did not correlate withthe behavioral measures of the VR task (results not shown).
Gait assessmentTurning indeed proved to be a provocative trigger for FOG, as 76%of the patients in the PD+FOG group froze during the 540° turnsand 41% froze during the 180° turns. In addition, only 35% of thepatients froze when having to perform a cognitive dual taskduring straight walking while 71% froze when dual tasking wasperformed during the performance of a 180° turn (Table 1).PD−NF patients did not experience any freezing. There was nosignificant difference in the amount of freezing experienced by PD+FOG patients between left and right turns (540°: t= 1.541,P= 0.143 and 180°: t= 0.490, P= 0.631).
Virtual reality taskTurning during the VR provoked behavioral freezing episodes in10 PD+FOG patients with an average of 13% of turns eliciting afreeze in those patients, whereas none of the PD−NF patientsexperienced a behavioral freeze during turning (U= 35,Z=− 2.793, Po0.01). In addition, even when removing allbehavioral freezing episodes, PD+FOG still had significantly higherscaled maximum footstep latencies when turning compared withwalking (t(16) = 2.17, P= 0.045), whereas PD−NF had similarscaled maximum footstep latencies (t(9) = 0.693, P= 0.506). Aspredicted PD+FOG also had higher step time variability compared
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
3
© 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited npj Parkinson's Disease (2015) 15020

with the non-freezer group as shown by an increased coefficientof variation during both turning and walking (Table 1).19 Withingroups, the coefficient of variation was slightly higher but notsignificantly different during turning compared with walking(PD+FOG: t=0.815, P=0.427, PD−NF: t=0.437, P=0.672). It isimportant to note that although PD+FOG patients had a slightlyhigher cadence, no significant differences in modal footstep latencieswere found between the groups, which together with similarUPDRS-III questions 3.7 (toe tapping) and 3.8 (leg agility) scores,indicates that any group differences found during turning wereunlikely to be due to an overall difference in motor performance.
Neuroimaging resultsWithin-group similarities. Figure 2 shows the whole brain BOLDresponse patterns for periods of walking and turning for eachgroup separately. The results of the within group turning4walk-ing contrasts are presented in Figure 3. The PD+FOG grouprequired widespread activation across the motor and visualcortices, cerebellum, and MLR region to achieve stepping in thevirtual reality task when compared with the selective corticalrecruitment in the PD−NF group. The PD+FOG group alsoactivated the cerebellum during turning, but this time with amore caudal region of the medulla. PD−NF required lessactivation across the motor, visual and cerebellar cortices, whilerecruiting more medial frontal regions.
Between group differences. Comparing the groups for thecontrast of turning4walking revealed four brain areas withsignificantly different BOLD responses (Figure 4). PD+FOG showed
decreased BOLD responses across the left supplementary motorarea (SMA) extending to the left premotor area and the leftsuperior parietal lobule with increased BOLD responses in boththe left and right inferior frontal gyrus when compared withPD−NF. Peak voxel statistics and coordinates are presented inTable 2. Although the left superior parietal lobule was significantlydifferent with the current statistical settings, the right superiorparietal lobule also appeared as to have decreased BOLDactivation during turning in PD+FOG compared with PD−NFwhen lowering the cluster size threshold to 13 voxels (t=− 3.08,P= 0.003).
Predefined ROI analysis. No significant group differences werefound for percent signal changes of striatal and subcortical ROI’swhen contrasting turning with walking in the VR. However, withinthe PD+FOG group a significant increased percent signal changein the left caudate nucleus (t16 = 2.75, P= 0.014) was found in thedifference score between turning and walking, whereas nodifferences were found for PD−NF. The right caudate alsoshowed increased percent signal change for the PD+FOG group,but this did not reach statistical significance (t16 = 1.72, P= 0.106).Finally, a supplementary analysis using a predefined spherical ROIof the pre-SMA (MNI: − 3; 6; 53)32 revealed reduced activation,although not significantly different, in the PD+FOG group duringturning compared with walking in the VR (Average beta: − 0.664,t= 1.55, P= 0.138).
Task-based functional connectivity. Four highly significantfunctional connectivity scores survived network-based statisticscorrection31 with stringent threshold settings (threshold = 3.0,
Table 1. Demographic statistics and behavioral results
PD+FOG (n= 17) PD−NF (n= 10)
Demographics Mean s.d. Mean s.d. T-value P-value
Age 67.4 6.2 64.8 4.1 1.15 0.262Disease duration 116 63 92.4 28 1.09 0.285DDE 776 321 785 296 0.07 0.945UPDRS-III 37.2 12 30.1 11 1.53 0.139MMSE 28.0 2.2 29.4 0.7 1.96 0.061MOCA 25.6 3.2 28.3 2.3 2.36 0.027HADS (Anxiety) 6.12 3.3 3.40 2.4 2.29 0.031HADS (Depression) 5.59 2.6 1.40 1.7 4.53 o0.01
Median Range Median Range Z-value P-value
H&Ya 2.5 2.0–3.0 2.0 2.0–2.5 − 1.55 0.122FOG-Q3a 3.10 2.0–4.0 0 0 − 4.14 o0.01UPDRS 3.7a (Toe tap R)a 2.0 0–3.0 1.0 0–2.0 − 1.70 0.127UPDRS 3.7b (Toe tap L)a 2.0 0–3.0 1.0 0–2.0 − 1.13 0.286UPDRS 3.8a (Leg agility R)a 1.0 0–3.0 1.0 0–2.0 − 1.67 0.127UPDRS 3.8b (Leg agility L)a 1.0 0–3.0 1.0 0–2.0 − 1.33 0.223
Virtual Reality task Mean s.d. Mean s.d. T-value P-value
Max SFSL turning 1.14 0.09 1.07 0.10 2.33 0.028Max SFSL walking 1.12 0.09 1.06 0.02 2.34 0.027CV turning 16.5 5.84 9.60 3.9 3.31 o0.01CV walking 15.7 5.44 9.26 3.9 3.27 o0.01Modal FSL 0.56 0.11 0.62 0.10 1.39 0.172
Abbreviations: PD+FOG, Parkinson’s disease patients with freezing of gait; PD−NF, Parkinson’s disease patients without freezing of gait.Independent sample t-test results presented unless otherwise indicated.Demographics: Disease duration given in months, DDE, daily dopamine dose equivalence; FOG-Q3, question 3 of the freezing of gait questionnaire;HADS, hospital anxiety and depression scale; H&Y, Hoehn and Yahr; MMSE, Mini Mental State Examination; MOCA, Montreal Cognitive Assessment; UPDRS-III,motor section of the unified Parkinson’s disease rating scale without Q10 (gait) and Q11 (freezing); UPDRS 3.7 and 3.8, Average scores on Q3.7a and Q3.7b and3.8a and 3.8b of the UPDRS (Left and right toe tapping and leg agility, respectively). Virtual Reality task: CV, coefficient of variation; Max SFSL, maximum scaledfootstep latency as scaled to the modal FSL; Modal FSL, modal footstep latency. All analyses were two-tailed with an alpha of 0.05.aMann–Whitney U-test used.
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
4
npj Parkinson's Disease (2015) 15020 © 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited

P= 0.007) when comparing periods of turning with walking withinthe PD+FOG group (Figure 5). Specifically, we observed increasedconnectivity between the bilateral MLR (t= 5.16, Po0.001),between the left GPi and both the right (t= 4.14, Po0.001) andthe left (t= 3.53, Po0.01) MLR and finally, between the right GPiand the left STN (t= 3.32, Po0.01). At a lower statistical thresholdof 2.5 (P= 0.023), additional significant functional connectivityscores were revealed between the right GPi and the right STN(t= 2.83, P= 0.01) and between the bilateral CLR with both the left(t= 2.86, Po0.01) and right MLR (t= 2.84, P= 0.01). The increasedfunctional connectivity score between the bilateral MLR remainedsignificant (Po0.05) when using the false detection rate option inthe network-based statistics software. No changes in functionalconnectivity were found within the PD−NF group betweenperiods of turning and walking. In addition, no significant groupdifferences in functional connectivity were found that survivednetwork based statistic or false detection rate correction.
DISCUSSIONThis is the first study to investigate brain activation patternsunderlying turning during a virtual reality task in Parkinson’sdisease patients with and without freezing of gait. Within groupresults revealed that PD+FOG relied heavily on widespread corticalcontrol of their movements, whereas PD−NF achieved moresuccessful stepping with a selective motor network. Betweengroup results showed that during turning, patients with freezingof gait displayed increased BOLD responses in bilateral inferiorfrontal regions and decreased BOLD across the left premotorcortex and left superior parietal cortex when compared withPD−NF. In addition, PD+FOG showed increased percent signalchanges in the left caudate nucleus and displayed strongfunctional connectivity between the GPi, STN, cerebellar andmesencephalic locomotor regions during turning relative to
walking. Importantly, an additional analysis without the subjectin the PD+FOG group that did not freeze during the gaitassessment (n= 16) revealed the same brain activation patterns,aiding towards the robustness of our findings.Turning during walking is a difficult motor task to investigate
using current neuroimaging techniques. Most evidence regardingthe effects of turning in PD patients with FOG therefore comesfrom indirect and spatially limited techniques, such as the effect ofSTN-DBS, transcranial magnetic stimulation and the effects ofdopaminergic medication.2,7,33 Thus far only one study has usedfMRI to investigate the effects of turning in PD patients with FOG.Peterson et al.24 used gait imagery of simple (forward) andcomplex (backward or turning) movements in combination withfMRI to report BOLD responses of five locomotor regions ofinterest in PD patients with and without FOG.24 They found thatfor PD patients without FOG, gait imagery of simple forwardwalking compared with rest was associated with increased BOLDresponses in several locomotor regions, including the SMA.However, patients with FOG actually showed decreased activationin the globus pallidus and mesencephalic locomotor region withtrends towards decreased activation in the right SMA.24 However,despite the large clinical impact turning has on freezing, nosignificant differences were found in that study between imaginedforward walking and turning in PD patients with and without FOG.In addition, none of the patients in that study reported freezingduring imagined gait.24 This indicates that the complexitybetween the gait imagery tasks might be too subtle to inducedetectable changes in brain activation within a Parkinson’s diseasecohort.24 These methods may also have lacked key challenges insensorimotor integration associated with turning and freezing ofgait.34–36 As such, the current study set out to investigate turning-related brain activation by using an interactive virtual reality taskthat required patients to perform a lower limb motor task while
Figure 2. Within-group whole brain results for periods of walking (WALK) and turning (TURN) in the virtual reality task. A, anterior, L, left; P,posterior, R, right; k420 and Po0.005.
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
5
© 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited npj Parkinson's Disease (2015) 15020

having to update a changing visual environment similar toturning.
Within groups: walkingThe within-group similarities during periods of walking revealedthat when off medication, PD+FOG patients required widespread
activation across their motor cortices, cerebellum and MLR region,whereas PD−NF used focused activation patterns across themotor cortex, frontal cortex, and cerebellum to perform the task.The widespread cortical involvement seen in the PD+FOG groupmight indicate that even during simple forward walking in the VR,these patients are unable to rely on lower-order automaticbehaviors and therefore must employ a more cortically controlledlocomotor network.3,11,12,37 Indeed, we found that PD+FOG wereunable to keep a consistent rhythm between their steps, asindicated by high step time variability scores, when comparedwith PD−NF. This result is consistent with previous studieslooking at virtual reality task performance19 and over groundwalking18 and is at least in part indicative to a loss of automaticityin locomotion, as stride time variability has been shown toimprove with dopaminergic medication while worsening duringdual tasking,18,38 indicating an ineffective basal gangliainvolvement.3,39 Our findings also support the notion thatalthough FOG is a paroxysmal phenomenon, patients withfreezing already have altered brain activation patterns duringeffective walking.18 This could increase their susceptibility tofreeze when additional information from competing pathways(e.g., sensorimotor, cognitive, and limbic) require concurrentprocessing by the basal ganglia causing response conflict thateventually presents itself as FOG.12
Within groups: turningWithin group similarities during periods of turning in the VRrevealed that PD+FOG relied more heavily on visual informationwhile decreasing the recruitment of medial motor and left parietalcortices. This might indicate that the PD+FOG were over-reliant onvisual information, possibly as a learned response to poorkinesthetic feedback.35,40–42 Turning in the VR might worsen thiseffect by preventing the subjects from actually rotating theirbodies according to the visually presented turn, altering theirexpected kinesthetic feedback as associated with turning overground.Alternatively, the extra visuo-parietal activation seen in the PD
+FOG group could reflect saccadic abnormalities that could leadto unsuccessful sensorimotor integration.8,43,44 Indeed, we also
Figure 4. Results of the whole brain second level group effect for turning4walking in the virtual reality task. (a) Screenshots from the wholebrain second level independent t-test for the turning4walking contrast showing PD+FOG4PD−NF results (P= Posterior, A=Anterior, k420,Po0.005, uncorrected, see Table 2), while using MOCA and HADS scores as covariates and. (b) The average beta intensities of the peak voxel8 mm spherical ROI’s for the turning 4walking contrast in the virtual reality task for both Parkinson’s disease patients with freezing ofgait (FOG) and without freezing of gait (NF). HADS, Hospital Anxiety and Depression Scale; MOCA, Montreal Cognitive Assessment;PD+FOG, Parkinson’s disease patients with freezing of gait; PD−NF, Parkinson’s disease patients without freezing of gait; ROI, regions ofinterest.
Figure 3. Within-group whole brain results for the contrast turn-ing4walking in the virtual reality task. A, anterior, L, left; P, posterior,R, right; k420 and Po0.005.
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
6
npj Parkinson's Disease (2015) 15020 © 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited

observed an increased percent signal change during turning inthe caudate nucleus for the PD+FOG group, an area implicated insaccadic functioning.45 Because of strong functional interactionsbetween the oculomotor system and basal ganglia circuitry,46 anyinappropriate visuomotor integration could cause responseconflict between predicted and actual motor outcomes, thuspotentially eliciting FOG.12 Indeed, the failure to recruit medialmotor and medial frontal regions in the PD+FOG group duringturning could indicate a difficulty with facilitating internally drivenmotor actions when visual support falls away.47 This notion issupported by the fact that externally driven motor actions, such asachieved through visual cueing techniques can alleviate freezingof gait,48 whereas turning is the most provocative trigger forfreezing of gait.27 Future studies are now needed to confirmsaccadic dysfunctions during turning in the VR and to determinewhether dopaminergic medication improves the basal gangliacircuitry during turning and thus visuomotor integration.49
The neural correlates underlying behavioral freezing episodes ina similar VR task are described in detail elsewhere.13,16,17 Such ananalysis was not performed in the current study owing to thelimited amount of behavioral freezing episodes recorded. Thesmall amount of freezing episodes can partly be explained bythe turns being 90°, with previous studies showing that sharperturns are more likely to cause FOG.5,27 Future studies are thereforeencouraged to implement sharper turns in virtual reality tasks toincrease the likelihood of eliciting FOG, allowing for thoseepisodes to be modeled with sufficient power when using fMRI.In addition, studies are encouraged to use ambulatory
electroencephalography systems to provide information aboutcortical activation underlying other critical sensorimotorchallenges associated with turning (e.g., balance, changing steplengths, and posture) that could not be modeled in the currentstudy. Finally, no objective measures were obtained duringthe current gait tasks, which were solely implemented to ensureaccurate group allocations. Future studies using objectivekinematical measures during turning are therefore needed to testwhether the VR and neuroimaging findings presented here areindeed related to the deficits seen during over ground turning.
Between groups: turning4walkingBetween group differences showed that navigating a virtual turncaused PD+FOG to recruit similar regions as those associated withthe putative “stopping network”10 that has also been implicated indetecting salient stimuli and attentional processes.50 In addition,strong functional connectivity patterns were found in the PD+FOGgroup during turning between subcortical regions and the STN,which has neural projections to the regions of this stoppingnetwork.10 The stopping network is most commonly implicatedthrough stop-signal tasks, showing activation of the right inferiorfrontal gyrus together with the pre-SMA, which is both func-tionally and structurally connected with the inferior frontalgyrus.10 This network implements inhibition through the STN,which regulates its effects via the Substantia Nigra pars reticulata,leading to thalamic inhibition when stopping is successful.10
Although little is known regarding the involvement of this stoppingnetwork across a broader range of motor tasks, a generalizabilityof the brain’s network for simple stop signal tasks has been shownto exist in more ecologically valid scenarios.51 In addition, theregions of this network have already been implicated in severalpathophysiological hypotheses of FOG.11,12,39,52 The recruitmentof this braking network could be an adaptive brain function thatarose during the gradual development of FOG as a result of fear offalling53 and difficulties navigating challenging environments.35 Ifso, this braking network might be over recruited during suchlocomotor challenges, precipitating more FOG.A recent pathophysiological model suggests that the main
output structures of the basal ganglia (i.e., GPi and SubstantiaNigra pars reticulata) provide tonic GABAergic inhibitory tone overthe brainstem structures that control gait (such as the MLR anddorsal pendunculopontine nucleus) and the motor thalamus,preventing any unwanted movements at rest.12 Importantly, thistonic inhibition can be deactivated when an appropriate motorplan from the motor cortex is processed by the basal ganglia,relieving this inhibitory output.12 However, one can argue thatwhen patients with PD and FOG proactively recruit a corticallycontrolled braking network as described above, additionalactivation of the STN might overrule any inhibitory reliefaccomplished through the basal ganglia system. Furthermore, inpatients with PD and FOG the motor plans presented mightalready be inappropriate due to abnormal sensory integration,34
Figure 5. Representation of the task based functional connectivityresults. Figure shows the comparison between periods of turningand periods of walking in the virtual reality task within theParkinson’s disease patients with freezing of gait group.CLR, cerebellar locomotor region; GPi, globus pallidusinternus; MLR, mesencephalic locomotor region; STN, subthalamicnucleus.
Table 2. Brain areas with significantly different BOLD responses in the second level using an independent t-test design for the contrastturning4walking in the virtual reality task, showing the PD+FOG4PD−NF peak voxel statistics
Brain area x y z # voxels T-value Z-value P-value
L—SMA—Premotor − 21 − 7 64 59 − 4.17 3.54 o0.001L—Parietal superior − 30 − 46 58 57 − 3.91 3.37 o0.001L—Inferior frontal − 30 35 13 35 3.58 3.14 o0.002R—Inferior frontal 36 20 25 20 4.04 3.46 o0.001
Abbreviations: BOLD, blood oxygen level-dependent; L—SMA, left supplementary motor area; PD+FOG, Parkinson’s disease patients with freezing of gait;PD−NF, Parkinson’s disease patients without freezing of gait; R, right.k420, Po0.005, uncorrected with coordinates in MNI space.
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
7
© 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited npj Parkinson's Disease (2015) 15020

especially during challenging situations,42 which together with adopamine depleted basal ganglia system decreases the likelihoodof successful cortico-basal relief of gait inhibition.The current study also found increased BOLD responses in the
PD+FOG group across the inferior frontal gyrus bilaterally alongwith increased functional connectivity among subcortical regionsassociated with stopping (e.g., STN and GPi) and movement(e.g., MLR and CLR). Importantly however, another key region ofthe stopping network, namely the pre-SMA, was absent while theleft premotor area actually showed significant decreased activa-tion patterns in PD+FOG. Indeed, a predefined ROI analysis of thepre-SMA revealed reduced activation, although not significantlydifferent, in the PD+FOG group during turning compared walkingin the VR. The lack of significant group differences across the basalganglia, thalamic and subcortical regions associated with stoppingcan potentially be explained by the removal of all behavioralfreezing episodes from the analyses. As such, our results arealigned with previous studies that also showed reduced premotoractivation in PD patient with FOG during gait imagery23,24 andbehavioral freezing in the virtual reality task,13 along withprofound structural20,54 and similarly increased functionalconnectivity impairments11 that have been reported in PD+FOGbetween the (pre-) SMA, STN and the CLR and MLR.Based on the current results it can be speculated that PD
patients with FOG have an inherent over activity across a stoppingnetwork when turning and that during this time the pre-SMA failsto provide necessary contextual input, rendering the striatumunable to correctly update the ongoing motor plan.11,52 Thisforces the GPi and STN to “shut down” activity in its’ efferenttargets through the synchronization of the MLR and CLR,12 whichin turn prevents the movement centers of the brain (such as themotor cortex, thalamus, and cerebellum) from receiving sensoryinformation to adjust the gait cycle. To make matters worse, muchof the sensory information gathered will already be inadequate, asPD+FOG have known proprioceptive, postural, saccadic andsensorimotor integration deficits,34,41–43 which are all essentialelements for the successful execution of a turn. Indeed, reducedBOLD responses were found in the current study in both the leftpremotor cortex along with the left and right superior parietallobule, which are thought to be involved in the prediction ofsomatosensory consequences of a motor plan and the integrationof sensorimotor information.23 The movements during a turn willtherefore no longer match their predictions, leading to increasedresponse conflict, which together with the prospective recruit-ment of a braking network, explains why turning is so likely totrigger FOG.
ConclusionDuring the navigation of a turn in the virtual reality task,Parkinson’s disease patients with freezing of gait show alteredBOLD responses across regions that implicate the prospectiverecruitment of a stopping network, which may be manifestedpathologically as a freeze when sensorimotor processing becomesmore complex.
ACKNOWLEDGMENTSWe would like to thank Dr Paul Clouston, Matthew Georgiades, Deborah Hammond,Alana Muller and Jennifer Szeto, for their help with data collection. We would finallylike to thank our patient volunteers for their participation in this study.
CONTRIBUTIONSConceived and designed the experiments: MG JMS CCW SJGL. Performed theexperiments: MG CCW JMH. Analyzed the data: MG JMS CO. Wrote the firstmanuscript: MG. Review and critique of the manuscript: JMS CCW CO JMH SJGL.Guarantor: SJGL.
COMPETING INTERESTSNone of the authors have any competing financial interests to declare in relation tothe work described.
FUNDINGMoran Gilat is supported by a University of Sydney International PostgraduateResearch Scholarship. James M. Shine is supported by an NHMRC CJ MartinFellowship. Courtney C. Walton is supported by the Australian Postgraduate Award.Claire O’Callaghan is supported by an NHMRC Neil Hamilton Fairley Fellowship. JulieM. Hall is supported by a University of Western Sydney International Scholarship.Simon J.G. Lewis received funding for this project from a NHMRC Project Grant(1062319).
REFERENCES1 Cumming RG, Klineberg RJ. Fall frequency and characteristics and the risk of hip
fractures. J Am Geriatr Soc 1994; 42: 774–778.2 McNeely ME, Earhart GM. The effects of medication on turning in people with
Parkinson disease with and without freezing of gait. J Parkinsons Dis 2011; 1:259–270.
3 Nutt JG, Bloem BR, Giladi N, Hallett M, Horak FB, Nieuwboer A. Freezing of gait:moving forward on a mysterious clinical phenomenon. Lancet Neurol 2011; 10:734–744.
4 Schaafsma JD, Balash Y, Gurevich T, Bartels AL, Hausdorff JM, Giladi N.Characterization of freezing of gait subtypes and the response of each to levo-dopa in Parkinson's disease. Eur J Neurol 2003; 10: 391–398.
5 Bhatt H, Pieruccini-Faria F, Almeida QJ. Dynamics of turning sharpness influencesfreezing of gait in Parkinson's disease. Parkinsonism Relat Disord 2013; 19:181–185.
6 Spildooren J, Vercruysse S, Meyns P, Vandenbossche J, Heremans E, Desloovere Ket al. Turning and unilateral cueing in Parkinson's disease patients with andwithout freezing of gait. Neuroscience 2012; 207: 298–306.
7 Lohnes CA, Earhart GM. Effect of subthalamic deep brain stimulation on turningkinematics and related saccadic eye movements in Parkinson disease. Exp Neurol2012; 236: 389–394.
8 Rieger JW, Kim A, Argyelan M, Farber M, Glazman S, Liebeskind M et al. Corticalfunctional anatomy of voluntary saccades in Parkinson disease. Clin EEG Neurosci2008; 39: 169–174.
9 Haynes WIA, Haber SN. The organization of prefrontal-subthalamic inputs inprimates provides an anatomical substrate for both functional specificity andintegration: implications for Basal Ganglia models and deep brain stimulation.J Neurosci 2013; 33: 4804–4814.
10 Aron AR, Robbins TW, Poldrack RA. Inhibition and the right inferior frontal cortex:one decade on. Trends Cogn Sci 2014; 18: 177–185.
11 Fling BW, Cohen RG, Mancini M, Carpenter SD, Fair DA, Nutt JG et al. Functionalreorganization of the locomotor network in Parkinson patients with freezingof gait. PLoS One 2014; 9: e100291.
12 Lewis SJG, Shine JM. The next step: a common neural mechanism for freezingof gait. Neuroscientist 20141073858414559101.
13 Shine JM, Matar E, Ward PB, Bolitho SJ, Gilat M, Pearson M et al. Exploring thecortical and subcortical functional magnetic resonance imaging changes asso-ciated with freezing in Parkinson's disease. Brain 2013; 136: 1204–1215.
14 Schmalbach B, Günther V, Raethjen J, Wailke S, Falk D, Deuschl G et al. Thesubthalamic nucleus influences visuospatial attention in humans. J Cogn Neurosci2014; 26: 543–550.
15 Spildooren J, Vercruysse S, Heremans E, Galna B, Vandenbossche J, Desloovere Ket al. Head-pelvis coupling is increased during turning in patients with Parkinson'sdisease and freezing of gait. Mov Disord 2013; 28: 619–625.
16 Shine JM, Matar E, Ward PB, Frank MJ, Moustafa AA, Pearson M et al. Freezing ofgait in Parkinson's disease is associated with functional decoupling between thecognitive control network and the basal ganglia. Brain 2013; 136: 3671–3681.
17 Shine JM, Matar E, Ward PB, Bolitho SJ, Pearson M, Naismith SL et al. Differentialneural activation patterns in patients with Parkinson's disease and freezing of gaitin response to concurrent cognitive and motor load. PLoS One 2013; 8: e52602.
18 Hausdorff JM, Schaafsma JD, Balash Y, Bartels AL, Gurevich T, Giladi N. Impairedregulation of stride variability in Parkinson's disease subjects with freezing of gait.Exp Brain Res 2003; 149: 187–194.
19 Gilat M, Shine JM, Bolitho SJ, Matar E, Kamsma YPT, Naismith SL et al. Variability ofStepping during a virtual reality paradigm in Parkinson's disease patients withand without freezing of gait. PLoS One 2013; 8: e66718.
20 Vercruysse S, Leunissen I, Vervoort G, Vandenberghe W, Swinnen S, Nieuwboer A.Microstructural changes in white matter associated with freezing of gait inParkinson's disease. Mov Disord 2015; 30: 567–576.
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
8
npj Parkinson's Disease (2015) 15020 © 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited

21 Tessitore A, Amboni M, Esposito F, Russo A, Picillo M, Marcuccio L et al. Resting-state brain connectivity in patients with Parkinson's disease and freezing of gait.Parkinsonism Relat Disord 2012; 18: 781–787.
22 Maillet A, Thobois S, Fraix V, Redouté J, Le Bars D, Lavenne F et al. Neural substratesof levodopa-responsive gait disorders and freezing in advanced Parkinson's dis-ease: a kinesthetic imagery approach. Hum Brain Mapp 2015; 36: 959–980.
23 Snijders AH, Leunissen I, Bakker M, Overeem S, Helmich RC, Bloem BR et al. Gait-related cerebral alterations in patients with Parkinson's disease with freezingof gait. Brain 2010; 134: 59–72.
24 Peterson DS, Pickett KA, Duncan R, Perlmutter J, Earhart GM. Gait-related brainactivity in people with Parkinson disease with freezing of gait. PLoS One 2014; 9:e90634.
25 Vercruysse S, Spildooren J, Heremans E, Wenderoth N, Swinnen SP, Vanden-berghe W et al. The neural correlates of upper limb motor blocks in Parkinson'sdisease and their relation to freezing of gait. Cereb Cortex 2014; 24: 3154–3166.
26 Giladi N, Shabtai H, Simon E, Biran S, Tal J, Korczyn A. Construction of freezing ofgait questionnaire for patients with Parkinsonism. Parkinsonism Relat Disord 2000;6: 165–170.
27 Spildooren J, Vercruysse S, Desloovere K, Vandenberghe W, Kerckhofs E, Nieuw-boer A. Freezing of gait in Parkinson's disease: the impact of dual-tasking andturning. Mov Disord 2010; 25: 2563–2570.
28 Lieberman MD, Cunningham WA. Type I and Type II error concerns in fMRIresearch: re-balancing the scale. Soc Cogn Affect Neurosci. 2009; 4: 423–428.
29 Brett M, Anton J-L, Valabregue R, Poline J-B. Region of interest analysis using anSPM toolbox. Presented at the 8th International Conference on FunctionalMapping of the Human Brain, June 2-6 2002, Sendai, Japan. Available on CD-ROMin NeuroImage, Vol 16, No. 2, abstract 497.
30 Friston KJ. Functional and effective connectivity: a review. Brain Connect 2011; 1:13–36.
31 Zalesky A, Fornito A, Bullmore ET. Network-based statistic: identifying differencesin brain networks. Neuroimage 2010; 53: 1197–1207.
32 Mayka MA, Corcos DM, Leurgans SE, Vaillancourt DE. Three-dimensional locationsand boundaries of motor and premotor cortices as defined by functional brainimaging: a meta-analysis. Neuroimage 2006; 31: 1453–1474.
33 Lee SY, Kim M-S, Chang WH, Cho J-W, Youn J-Y, Kim Y-H. Effects of repetitivetranscranial magnetic stimulation on freezing of gait in patients withParkinsonism. Restor Neurol Neurosci 2014; 32: 743–753.
34 Earhart GM, Stevens ES, Perlmutter JS, Hong M. Perception of active and passiveturning in Parkinson disease. Neurorehabil Neural Repair 2007; 21: 116–122.
35 Cowie D, Limousin P, Peters A, Day BL. Insights into the neural control of loco-motion from walking through doorways in Parkinson's disease. Neuropsychologia2010; 48: 2750–2757.
36 Herman T, Giladi N, Hausdorff JM. Neuroimaging as a window into gait dis-turbances and freezing of gait in patients with Parkinson's disease. Curr NeurolNeurosci Rep 2013; 13: 411–413.
37 Vandenbossche J, Deroost N, Soetens E, Coomans D, Spildooren J, Vercruysse Set al. Freezing of gait in Parkinson's disease: disturbances in automaticity andcontrol. Front Hum Neurosci 2012; 6: 356.
38 Pieruccini-Faria F, Jones JA, Almeida QJ. Motor planning in Parkinson's diseasepatients experiencing freezing of gait: the influence of cognitive load whenapproaching obstacles. Brain Cogn 2014; 87: 76–85.
39 Lewis SJG, Barker RA. A pathophysiological model of freezing of gait inParkinson's disease. Parkinsonism Relat Disord 2009; 15: 333–338.
40 Azulay J-P, Mesure S, Blin O. Influence of visual cues on gait in Parkinson's disease:contribution to attention or sensory dependence? J Neurol Sci 2006; 248:192–195.
41 Tan T, Almeida QJ, Rahimi F. Proprioceptive deficits in Parkinson's disease patientswith freezing of gait. Neuroscience 2011; 192: 746–752.
42 Ehgoetz Martens KA, Pieruccini-Faria F, Almeida QJ. Could sensory mechanismsbe a core factor that underlies freezing of gait in Parkinson's disease? PLoS One2013; 8: e62602.
43 Lohnes CA, Earhart GM. Saccadic eye movements are related to turning perfor-mance in Parkinson disease. J Parkinsons Dis 2011; 1: 109–118.
44 Galna B, Lord S, Daud D, Archibald N, Burn D, Rochester L. Visual sampling duringwalking in people with Parkinson’s disease and the influence of environment anddual-task. Brain Res 2012; 1473: 35–43.
45 Kitama T, Ohno T, Tanaka M, Tsubokawa H, Yoshida K. Stimulation of the caudatenucleus induces contraversive saccadic eye movements as well as head turning inthe cat. Neurosci Res 1991; 12: 287–292.
46 Sieger T, Bonnet C, Serranová T, Wild J, Novák D, Růžička F et al. Basal ganglianeuronal activity during scanning eye movements in Parkinson's disease. PLoSOne 2013; 8: e78581.
47 van der Hoorn A, Renken RJ, Leenders KL, de Jong BM. Parkinson-related changesof activation in visuomotor brain regions during perceived forward self-motion.PLoS One 2014; 9: e95861.
48 Velik R, Hoffmann U, Zabaleta H, Marti Masso JF, Keller T. The effect of visual cueson the number and duration of freezing episodes in Parkinson's patients. ConfProc IEEE Eng Med Biol Soc 2012; 2012: 4656–4659.
49 Nantel J, Bronte-Stewart H. The effect of medication and the role of posturalinstability in different components of freezing of gait (FOG). Parkinsonism RelatDisord 2014; 20: 447–451.
50 Sebastian A, Jung P, Neuhoff J, Wibral M, Fox PT, Lieb K et al. Dissociableattentional and inhibitory networks of dorsal and ventral areas of the right inferiorfrontal cortex: a combined task-specific and coordinate-based meta-analyticfMRI study. Brain Struct Funct 2015; e-pub ahead of print 1 February 2015; doi:10.1007/s00429-015-0994-y.
51 Wessel JR, Aron AR. Inhibitory motor control based on complex stopping goalsrelies on the same brain network as simple stopping. Neuroimage 2014; 103:225–234.
52 Shine JM, Moustafa AA, Matar E, Frank MJ, Lewis SJG. The role of frontostriatalimpairment in freezing of gait in Parkinson's disease. Front Syst Neurosci 2013;7: 61.
53 Ehgoetz Martens KA, Ellard CG, Almeida QJ. Does anxiety cause freezing of gait inParkinson's disease? PLoS One 2014; 9: e106561.
54 Schweder PM, Hansen PC, Green AL, Quaghebeur G, Stein J, Aziz TZ. Connectivityof the pedunculopontine nucleus in parkinsonian freezing of gait. Neuroreport2010; 21: 914–916.
This work is licensed under a Creative Commons Attribution 4.0International License. The images or other third party material in this
article are included in the article’s Creative Commons license, unless indicatedotherwise in the credit line; if the material is not included under the Creative Commonslicense, users will need to obtain permission from the license holder to reproduce thematerial. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
Supplementary Information accompanies the paper on the npj Parkinson’s Disease website (http://www.nature.com/npjparkd)
Neuroimaging turning in Parkinson’s diseaseM Gilat et al
9
© 2015 Parkinson's Disease Foundation/Macmillan Publishers Limited npj Parkinson's Disease (2015) 15020

APPENDIX(4((
(Walton C.C., O’Callaghan, C., Hall J. M., Gilat, M., Mowszowski, L., Naismith,
S.L., Burrell, J. R., Shine, J.M., Lewis, S. J. G. Antisaccade errors reveal cognitive
control deficits in Parkinson’s disease with freezing of gait. Journal of Neurology,
2015
!

ORIGINAL COMMUNICATION
Antisaccade errors reveal cognitive control deficits in Parkinson’sdisease with freezing of gait
Courtney C. Walton1 • Claire O’Callaghan1,2 • Julie M. Hall1,3 • Moran Gilat1 •
Loren Mowszowski1,4 • Sharon L. Naismith1,4 • James R. Burrell5 •
James M. Shine5,6 • Simon J. G. Lewis1
Received: 23 July 2015 / Revised: 2 September 2015 / Accepted: 14 September 2015! Springer-Verlag Berlin Heidelberg 2015
Abstract Freezing of gait is a poorly understood symptomof Parkinson’s disease (PD) that is commonly accompanied
by executive dysfunction. This study employed an antisac-
cade task to measure deficits in inhibitory control in patientswith freezing, and to determine if these are associated with a
specific pattern of grey matter loss using voxel-based mor-
phometry. PD patients with (n = 15) and without (n = 11)freezing along with 10 age-matched controls were included.
A simple prosaccade task was administered, followed by a
second antisaccade task that required subjects to either looktowards or away from a peripheral target. Behavioral results
from the antisaccade task were entered as covariates in the
voxel-based morphometry analysis. Patient and controlgroups performed equally well on the first task. However,
patients with freezing were significantly worse on the sec-
ond, which was driven by a specific impairment in sup-pressing their responses toward the target on the antisaccade
trials. Impaired antisaccade performance was associatedwith grey matter loss across bilateral visual and fronto-
parietal regions. These results suggest that patients with
freezing have a significant deficit of inhibitory control that isassociated with volume reductions in regions crucial for
orchestrating both complex motor behaviors and cognitive
control. These findings highlight the inter-relationshipbetween freezing of gait and cognition and confirm that
dysfunction along common neural pathways is likely to
mediate the widespread cognitive dysfunction that emergeswith this symptom.
Keywords Antisaccades ! Cognitive control !Freezing of gait ! Parkinson’s disease
Introduction
Freezing of gait (FOG) is a debilitating motor symptom
experienced by a significant number of people withParkinson’s disease (PD), which manifests when a patient
becomes unable to progress forward while walking [1]. The
symptom is highly related to poorer quality of life [2],highlighting the importance in understanding its complex
nature. A number of mechanisms for FOG have beenhypothesized; however, our understanding of the phe-
nomenon is still evolving. Therefore, further investigations
into underlying neural processes employing new and novelexperimental designs are warranted, in order to advance the
potential for clinical treatments and biomarkers.
There now stands a significant body of evidence tosuggest that FOG is fundamentally related to dysfunction
across fronto-parietal and striatal regions [3, 4], which has
been identified via convergent evidence from neuropsy-chological testing and both structural and functional
& Simon J. G. [email protected]
1 Parkinson’s Disease Research Clinic, Brain and Mind Centre,University of Sydney, 94 Mallett Street, Camperdown, NSW,Australia
2 Behavioural and Clinical Neuroscience Institute, Universityof Cambridge, Cambridge, UK
3 School of Social Sciences and Psychology, Western SydneyUniversity, Parramatta, NSW, Australia
4 Healthy Brain Ageing Program, School of Psychology; Brainand Mind Centre & The Charles Perkins Centre, Universityof Sydney, Sydney, NSW, Australia
5 Neuroscience Research Australia, University of New SouthWales, Sydney, NSW, Australia
6 School of Psychology, Stanford University, Stanford, CA,USA
123
J Neurol
DOI 10.1007/s00415-015-7910-5

neuroimaging [5, 6]. Yet, the classical neuropsychological
tasks that have been used to establish the cognitive profileassociated with FOG tend to be highly multidimensional,
relying on diverse cognitive and behavioral domains (in-
cluding verbal and motor responses), which can complicatetheir interpretation. In contrast, saccadometry is a novel,
objective tool that can be used to probe discrete aspects of
neural decision-making behavior, even in patients withsignificant motor and speech impairments [7]. Thus, it can
provide highly informative data relating to the neuralunderpinnings of cognitive processing.
This tool measures saccades, the eye movements made
when rapidly shifting focus across the external environ-ment. A typical experiment involves presenting a target
stimulus to a participant, who must then either direct their
attention towards (prosaccades) or away (antisaccades)from a second peripheral stimulus, depending on the
paradigm rule. As such, during the antisaccade task the
patient is required to: (1) suppress the automatic responseto look to the target; and (2) to produce an internal vol-
untary command to look away from the target (see Fig. 1).
Additionally, working memory is required to keep theparadigm rules online.
Though clearly involving visuospatial processing, the
cognitive demands of the antisaccade task have beendescribed in terms of inhibitory control and are linked with
functioning in the basal ganglia and frontal regions such as
the supplementary and frontal eye fields and dorsolateralprefrontal cortex (DLPFC). These regions are involved in
an oculomotor network in which the superior colliculus is
the final region responsible for saccade responses. Appro-priate processing therefore requires frontal inhibition onto
this structure [8]. Thus, antisaccade impairments are typi-
cally inferred as representative of frontostriatal dysfunction[9]. Accordingly, performance in this task has been shown
to be impaired in various conditions with known frontos-
triatal dysfunction, including fronto-temporal dementia [7],schizophrenia [10], attention deficit hyperactivity disorder
[11], frontal lobe lesions [12] and, of relevance here, PD.
There have been significant variations to the way sac-cadic behavior has been investigated in PD, leading to
contradictory findings in the literature [13, 14]. Typically, a
higher proportion of research has focused on reflexive,visually guided or memory-guided saccades, rather than
antisaccades. Nonetheless, Rivaud-Pehoux and colleagues[15] have shown that in tasks where prosaccades and
antisaccades are combined together, patients with PD show
significant errors in antisaccade production. This is similarto recent work [16] which has shown that even in drug-
naıve PD, patients show significantly more antisaccade
errors compared to controls. In another particularly inter-esting study [17], the antisaccade task was broken down
into three variations to assess the components necessary for
successful completion. This study suggested that deficits inworking memory for paradigm rules could not explain
differences in antisaccade performance in PD compared to
controls. Rather, they discovered that patients with PDshowed a generalized slowing of voluntary saccade pro-
duction. Additionally, they found that patients showed
significant problems in inhibiting their responses in adelayed version of the task, concluding this to be core to
the deficits observed. This is in line with a Go/NoGo ver-
sion of the task which highlighted impaired inhibition of apartially prepared saccadic response in patients with PD
[18].
However, antisaccades are not consistently impaired inall patients with PD [19], and therefore it is plausible that
the heterogeneity present in the disease may impact on
antisaccade performance across study samples. Given theobserved frontostriatal deficits in patients with FOG, and in
Fig. 1 Representation of the antisaccade task used in this study.a The participant keeps their eyes focused on the central cue (showinggreen and red lights overlapping). A change in the color of the centralcue will then require one of two responses, as follows: b (prosaccade),
the patient must direct their eyes towards the peripheral light (as thecentral cue is green) while in c (antisaccade), the patient must lookaway from the peripheral light when it appears (as the central cue isred)
J Neurol
123

particular their difficulty with inhibitory control [20, 21], it
could be anticipated these patients would have an impairedability to perform antisaccades accurately. However, the
specific investigation of inhibitory eye movements is yet to
be investigated in PD patients with FOG.In the current study, we aimed to explore whether
antisaccade performance was impaired in patients with
FOG, compared to those without, in relation to simpleprosaccades. Given that numerous structural changes have
been related to FOG [22–26], we aimed to investigatewhether structural imaging could help define some of the
core processes underlying poor performance of antisac-
cades. Thus, we used voxel-based morphometry (VBM) todetermine the neuroanatomical basis of any saccadic
abnormalities discovered in the antisaccade task. Specifi-
cally, we hypothesized that patients with and without FOGwould show similar levels of performance on the first,
simple prosaccade task. However, we expected those with
FOG to show an impaired ability to perform quickly andaccurately in the antisaccade task, reflecting a diminished
ability to activate effective cognitive control. Finally, we
anticipated poor antisaccade performance to correlate withatrophy in visual and fronto-parietal areas involved in
cognitive control.
Methods
Recruitment of sample
Patient data were collected from the Parkinson’s DiseaseResearch Clinic at the Brain and Mind Centre, University
of Sydney. All patients met UK Brain Bank clinical criteria
[27] and did not meet MDS criteria for PD with dementia[28]. We defined patients with FOG as those who were
observed to freeze during Movement Disorders Society-
Unified Parkinson’s Disease Rating Scale (MDS-UPDRS)[29] assessments or alternatively, if they reported FOG on
Question 3 of the Freezing of Gait Questionnaire (FOG-Q)
[30] and to a clinician during part II of the MDS-UPDRS.Ten healthy age- and education-matched controls were
recruited from a volunteer panel, having been screened for
cognitive impairment or history of psychiatric or neuro-logical disorders. This research was approved by the
Human Research Ethics Committee of The University of
Sydney, and written informed consent was obtained fromall participants.
Sample selection
We actively matched our groups prior to experimental
analysis to account for the possible impact of diseaseseverity and dopaminergic treatment on saccade
performance [31, 32]. Owing to the typical nature of
comparison between patients with (FOG) and without FOG(no-FOG), significant group differences were shown in key
clinical characteristics for this initial sample. We chose to
remove two patients from the no-FOG group who were notundergoing medical treatment and one patient who had
only recently been diagnosed, thus significantly altering the
profile of the no-FOG group. The groups still stronglydiffered on disease duration and Hoehn and Yahr (H&Y)
staging [33] and thus we removed patients in a stepwisemanner starting with those in the FOG group with the
highest disease duration until no statistical difference
remained (four removed). This method has been usedpreviously [20] and led to groups being statistically
equivalent on key clinical variables prior to carrying out
any experimental testing (i.e., p[ .05; see Table 1).
Procedure
A standardized saccadometry protocol was administered
using a saccadometer Advanced (Ober Consulting,
Poland), which is a head-mounted device containing lasersthat project the target stimuli 2 m in front of the participant
on to a blank white wall [34]. Two experimental proce-
dures were carried out, each consisting of 100 trials thatwere preceded by 20 calibration trials.
The first experiment (‘‘STEP task’’) was a 10-degree
STEP task designed to evoke saccades directly toward thevisual stimulus (prosaccades). Each experimental trial
began with a central target (presented during a random
fore-period of 1–2 s), followed by a peripheral targetlocated 10" randomly presented to the left or right. In this
task, the peripheral target was presented at the time of the
central stimulus removal, thus typically generating fewerearly saccades in normal subjects [7]. The participant was
asked to direct their eyes to this peripheral target as soon as
it appeared. The data recorded from this experiment wereused to calculate median saccadic latency and a number of
parameters (l, r, and rE) as specified in the Linear
Approach to Threshold with Ergodic Rate (LATER) modelof saccadometry analysis developed by Carpenter and
colleagues [35]. In this model, the value of l represents the
mean rate of decision processes within the main distribu-tion of responses (with larger values indicating faster
decision-making). Additionally, r is the standard deviation
of l and thus represents variability of this rate. Finally, rErepresents the incidence of early saccades. These measures
are typically used to report data in prosaccade trials.
The second experiment (Peripheral Conflict withOverlap; ‘‘PCO Task’’) was a cued antisaccade task (see
Fig. 1). Each trial began with overlapping central cues (1
red and 1 green) presented vertically for 1.5–2.5 s. Asingle red or green central cue was then presented with a
J Neurol
123

simultaneous and randomly sequenced left or right
peripheral target (red). If the single central cue was green,participants were instructed to look at the peripheral tar-
get, whereas if the single central cue was red, they were
instructed to look away from the peripheral target, i.e., inthe opposite direction. Each individual trial was followed
by a gap of 750 ms, following which both central cues
would reappear. In this study, the primary outcomemeasure was the percentage of correct responses for
overall (all saccades) and specific (pro- vs antisaccades)scores in the PCO task. We did not anticipate any influ-
ence of fatigue on performance, with each trial taking
approximately 5 min to complete (including calibration).Additionally, there is a small gap between trials, making
any oculomotor fatigue unlikely.
Raw saccadometry data were analyzed using Laten-cyMeter (Ober Consulting, Version 4.9). Invalid traces
were automatically eliminated by LatencyMeter based on
a statistical analysis of the position and velocity profile ofeach individual trace. Data were then exported to Sac-
cadic Programming and Instrumentation Computer soft-
ware (http://www.cudos.ac.uk/spic.html), for estimationof best-fit LATER parameters of l, r, and rE. For the
STEP task, percentages and latency scores were derived
from LatencyMeter. The saccadometry tool and software
used in the current study have been used extensively insaccade research, including in neurodegenerative diseases
[7, 36].
Clinical assessments
A series of demographic details were recorded, includingage, gender, and education. Clinical features including
disease duration, Hoehn and Yahr stage (H&Y) [33],dopamine dose equivalence (DDE; mg/day) [37] and dis-
ease severity using the Movement Disorder Society-Uni-
fied Parkinson’s Disease Rating Scale (MDS-UPDRS) [29]of which section III (motor sub-score) was assessed.
Additionally, self-reported mood disturbance using the
Beck Depression Inventory (BDI-II) [38] and global cog-nitive functioning using the Minimental State Examination
(MMSE) [39] and the Montreal Cognitive Assessment
(MOCA) [40] were employed. A score comprising part Bminus part A of the trail-making test (TMT B - A) was
calculated as a typical measure of executive functioning
[41]. Color discrimination was assessed using the Farns-worth–Munsell 100-Hue Test [42] to ensure there were no
significant visual differences between groups.
Table 1 Clinical anddemographic characteristics ofthe sample including those with(FOG), without (no-FOG)freezing, and control subjects
FOG No-FOG Control p value
N 15 11 10
Age (years) 68.00 (7.7) 65.73 (5.2) 63.90 (7.5) .36a
Gender (male/female) 11/4 9/2 5/5 .26d
MDS-UPDRS III 33.27 (16.3) 22.45 (9.6) N/A .06b
H&Y stage (N) N/A .05d
1 1 1
2 6 9
2.5 3 0
3 4 1
5 1 0
DDE 632.00 (1300.0) 550.00 (992.0) N/A .53c
Years since diagnosis 8.13 (4.2) 5.27 (2.9) N/A .06b
Education (years) 13.86 (2.6) 13.30 (3.6) 14.25 (2.8) .78a
MMSE 30.00 (3.0) 30.00 (6.0) Not collected .84c
MOCA 28.00 (1.6) 27.60 (2.6) Not collected .64b
BDI-II 9.00 (27) 7.00 (21) Not collected .32c
Color contrast 167.00 (73.0) 120.00 (70.6) Not collected .13b
TMT B-A 54.20 (27.7) 56.36 (47.3) 43.44 (26.0) .68a
FOG-Q total 10.20 (6.0) 3.00 (2.5) N/A .00b
Means and standard deviations displayed unless data is non-parametric (c), in which case median (range)givena ANOVA usedb Independent samples t test usedc Mann–Whitney U test usedd Chi-square test used
J Neurol
123

Statistical analyses
Analyses of variance (ANOVA) or Kruskal–Wallis H tests
were conducted across the three groups for parametric and
non-parametric data, respectively. Independent samplest tests or Mann–Whitney U tests were conducted to assess
for differences between the two PD groups for parametric
and non-parametric data, respectively. Chi-square test wasused for categorical variables. The majority of saccadom-
etry data was non-parametric, and as such was entered at
once for a Kruskal–Wallis H test. Significant differenceswere followed by post hoc pairwise comparisons using a
Dunn’s Bonferroni approach to control for multiple
comparisons.
Imaging acquisition
A subset of 20 PD patients (12 freezers, 8 non-freezers)
underwent structural magnetic resonance imaging (MRI).This subset did not differ from the remaining population in
antisaccade performance or any other clinical variables (all
p[ .05). Whole-brain T1 images were acquired using aGeneral Electric 3 T MRI (General Electric, Milwaukee,
USA) with standard quadrature head coil (8 channels). The
3D T1-weighted sequences were acquired as follows:coronal orientation, matrix 256 9 256, 200 slices,
1 9 1 mm2 in-plane resolution, slice thickness 1 mm, TE/
TR = 2.6/5.8 ms.
Voxel-based morphometry (VBM) analysis
Voxel-based morphometry (VBM) was performed on the
three-dimensional T1-weighted scans, using the FSL-VBM
toolbox in the FMRIB software library package. Pre-pro-
cessing of the scans firstly involved extracting the brain
from all scans using the BET algorithm in FSL, using afractional intensity threshold of 0.22 [43]. A grey matter
template, specific to this study, was then built from can-
vassing the maximum equal amounts from both groups(i.e., FOG and no-FOG). Template scans were then regis-
tered to the Montreal Neurological Institute Standard space
(MNI 152) using non-linear b-spline representation of theregistration warp field, resulting in a study-specific grey
matter template at 2 mm3 resolution in standard space.
Simultaneously, the brain-extracted scans were also pro-cessed with the FMRIB’s automatic segmentation tool
(FAST v4.0) [44] to achieve tissue segmentation into
cerebrospinal fluid, grey matter and white matter. TheFAST algorithm also corrected for spatial intensity varia-
tions such as bias field or radio-frequency inhomogeneities
in the scans, resulting in partial volume maps of the scans.In the next step, grey matter partial volume maps were non-
linearly registered to the study-specific template via non-
linear b-spline representation of the registration warp.These maps were then modulated by dividing by the
Jacobian of the warp field, to correct for any contraction/
enlargement caused by the non-linear component of thetransformation [45]. After normalization and modulation,
smoothing the grey matter maps occurred using an iso-
tropic Gaussian kernel (standard deviation = 3 mm; fullwidth half maximum = 8 mm). Finally, a voxel-wise
general linear model (GLM) was applied and permutation-based non-parametric testing was used to form clusters
with the Threshold-Free Cluster Enhancement method.
Percent correct scores from the antisaccade task wereentered as covariates in a GLM matrix and a positive
Fig. 2 Boxplots representing error rates between groups on the PCO task; **p\ .01, *p\ .05, ns not significant; a percentage of correctprosaccades; b percentage of correct antisaccades
J Neurol
123

t-contrast was tested with 5000 permutations and reported
at a significance level of p\ .001 uncorrected and at aconservative cluster threshold of greater than 100 con-
tiguous voxels.
Results
Demographic statistics and descriptives are presented in
Table 1. Table 2 shows detailed results for saccadometry
performance. Of note, there were no significant differencesacross the groups for any of the STEP task outcomes (all
p values [.05), suggesting intact performance on basicsaccadometry metrics for both patient groups relative to
controls. However, the PCO task revealed specific deficits
for patients with FOG, evidenced by significantly lowerpercentage of correct saccades relative to controls
(p = .00) and the same trend relative to the no-FOG group
(p = .07). Importantly, when separating the performancebetween prosaccade and antisaccade trials (Fig. 2), it was
clear that the FOG groups’ poor performance on the anti-
saccade trials was driving this result, with their accuracysignificantly below the control and no-FOG groups
(p = .01). In contrast, the three groups were equivalent in
their accuracy for prosaccade trials (p = .23). The FOG
group had slower latencies compared to controls for anti-
saccade trials they performed correctly (p = .01), but notcompared to those without FOG. Both patient groups had
slower latencies than controls in the incorrect trials but this
was not statistically significant.
Relationship between antisaccade performanceand voxel-based morphometry
Across the PD patients, poorer performance (i.e., percent-age of incorrect antisaccades) on the PCO task was related
to atrophy in occipital regions, in addition to a fronto-
parietal network. Specifically, clusters where found in theoccipital cortex, bilateral posterior parietal lobe, supple-
mentary motor cortex, left posterior lateral temporal cortex,
right inferior frontal gyrus, anterior/paracingulate cortexand left orbitofrontal cortex (see Fig. 3; Table 3).
Discussion
In the current study, we have shown that the ability tocorrectly execute antisaccades is significantly compro-
mised in patients with FOG. This was despite PD patients
with and without FOG showing relatively equal
Table 2 Descriptives and non-parametric test statistics across groups for all saccadometry variables
Saccadometryoutcomes
Group Test statistics
FOG No-FOG Control Overall groupdifference
FOG vs no-FOG FOG vs control
STEP parameters
l 4.77 (4.6) 3.96 (3.8) 5.15 (1.2) H = 3.97,p = .14
N/A N/A
r 1.02 (1.6) 1.20 (1.3) 0.88 (1.3) H = 2.70,p = .26
N/A N/A
rE 5.06 (11.7) 0.00 (6.4) 0.00 (5.8) H = 5.37,p = .07
N/A N/A
STEP latency 209.64 (529.4) 252.50 (297.3) 194.40 (47.0) H = 11.42,p = .13
N/A N/A
STEP % correct 98.00 (36.0) 99.00 (31.0) 100.00 (9.0) H = 3.64,p = .16
N/A N/A
PCO % correct 62.00 (27.0) 66.00 (33.0) 73.00 (40.0) H = 11.42,p = .00
H = -9.51,z = -2.28, p = .07
H = -13.78,z = -3.21, p = .00
PCO % prosaccadescorrect
92.00 (43.2) 85.70 (37.9) 90.65 (20.0) H = 2.92,p = .23
N/A N/A
PCO % antisaccadescorrect
32.00 (62.3) 58.00 (55.9) 56.65 (69.3) H = 12.86,p = .00
H = -13.25,z = -3.17, p = .01
H = -12.18,z = -2.83, p = .01
PCO correctantisaccades latency
534.00 (757.0) 476.00 (541.0) 387.00 (241.0) H = 9.88,p = .01
H = -5.28,z = 1.26, p = .62
H = -13.52,z = 3.14, p = .01
PCO incorrectantisaccades latency
297.00 (323.0) 334.00 (336.0) 219.00 (101.0) H = 11.28,p = .00
H = -5.13,z = -1.23, p = .66
H = -10.08,z = 2.35, p = .06
J Neurol
123

performance in a simple STEP task along with neuropsy-
chological measures of cognitive functioning. Moreover,reduced performance of antisaccades related to atrophy in
fronto-parietal regions and occipital cortex, which have
previously been implicated in the pathophysiology of FOG.These results suggest that saccadometry may be a useful
behavioral marker for FOG, and provide further evidence
of widespread visual and cognitive control regions being
associated with its pathology.Results on the first task showed no significant differ-
ences across patient and control groups in terms of early
saccades and decision-making processes. Additionally,measures of saccade latency and accuracy were similar for
this task across the groups. This is an important consider-
ation relative to the PCO task and suggests that differencesbetween these groups are not simply due to basic decision-
making or visual dysfunction. Rather, the more complexhigher cognitive demands induced by the PCO task led to
the significant group differences. However, it should be
noted that though not statistically significant, the rE vari-able was trending towards a difference. Median rates across
groups clearly suggest a higher incidence of early saccades
in patients with FOG compared to patients without, andcontrols. Table 2 shows that the median rE was 0 for both
no-FOG and control groups, while patients with FOG
showed much higher responses (5.06). This may be rele-vant, as an increased incidence of early saccades could
indicate reduced inhibitory control of reflexive saccades
[7], consistent with impaired performance on the PCO task.The trending difference found here may have become more
apparent in a larger sample. Even so, this is an area that is
not well understood in terms of the neural mechanics [46].It is thought that early saccades may be a result of ineffi-
cient inhibition of the frontal cortex onto the superior
colliculus, though further exploration is needed to ade-quately define the neural and neuropsychological correlates
of this measure.
Patients with FOG performed significantly worse on thePCO task in terms of the overall percentages of trials that
were correctly responded to. Moreover, the errors in this
Fig. 3 VBM results showing regions of grey matter loss thatcorrelated with lower performance (i.e., higher error percentage) onthe antisaccade task. a Right orbitofrontal cortex; b right inferior/middle frontal gyri; c left posterior middle/superior temporal gyri;d supplementary motor cortex/primary motor cortex/frontal eyefields, anterior cingulate, occipital lobe. Results uncorrected atp\ .001; t[ 3.8
Table 3 VBM results showing areas of significant grey matter intensity decrease across all PD patients that correlated with reduced performanceon the antisaccade task
Region Hemisphere(L/R/B)
MNI coordinates for voxel of maximal intensity Number ofvoxels
T value
X Y Z
Occipital pole B 10 -92 28 696 4.90
Middle/superior temporal gyri L -50 -40 -6 567 4.90
Occipital pole R 20 -90 18 304 3.89
Inferior/middle frontal gyri R 44 8 34 203 3.89
Supramarginal gyrus/superior parietal lobule L -50 -42 4 201 4.90
Supplementary motor cortex/primary motorcortex/frontal eye fields
B -10 -8 62 165 4.90
Anterior/paracingulate cortex B 14 24 34 141 3.89
Lateral occipital cortex/superior parietal lobule R 36 -60 46 139 3.89
Orbitofrontal cortex L -20 26 -12 125 3.58
Lateral occipital cortex/superior parietal lobule L -22 -64 64 117 4.90
Superior parietal lobule/postcentral gyrus R 12 -46 68 102 3.89
Results are uncorrected at p\ .001 and at a cluster threshold of greater than 100 contiguous voxels
J Neurol
123

task could be specifically attributed to the antisaccade tri-
als, rather than the prosaccades. Thus, the primary deficitobserved in patients with FOG was in inhibiting the
reflexive saccade, and the production of a voluntary anti-
saccade without error. These results are aligned with recentbehavioral findings, which show that patients with FOG
make significantly more errors during incongruent tasks
which require control of inhibitory processes compared topatients without FOG [20, 21]. Overall, the behavioral
results presented here dovetail with prior literature toconfirm that patients with FOG have deficient cognitive
control.
Cognitive control has previously been implicated asinvolving a fronto-parietal network which is needed for the
orchestration of complex executive functions. In particular,
inhibitory control—a crucial function needed to performantisaccades—has been shown to recruit these regions. A
large-scale meta-analysis incorporating neuroimaging
results from a wide array of inhibitory tasks (Antisaccade,Flanker, Go/No go, Simon and Stroop tasks) in 1112 par-
ticipants showed that the middle, inferior and medial
regions of frontal cortex along with the superior and infe-rior parietal cortex are most consistently implicated in this
process [47]. In particular, the right inferior frontal cortex
is thought to behave as a brake to suppress or pauseinappropriate responses, along with involvement of sub-
cortical regions [48].
To complement our behavioral findings, and to assess ifantisaccade performance in PD was associated with atro-
phy in the aforementioned regions, antisaccade perfor-
mance was correlated with VBM analysis. Antisaccadedeficits were related to grey matter loss along key regions
in the visual processing stream, including occipital, tem-
poral and parietal cortex. Broadly, these areas are critical invisual perception and saccade production. We also found
evidence of frontal involvement in relation to poorer per-
formance. The critical contribution of these regions,including but not limited to the frontal eye fields, superior
parietal cortex and supramarginal gyrus, is aligned with
previous explorations of antisaccade performance [9, 49,50]. We also identify regions in the right inferior prefrontal
cortex that correlate with antisaccade performance, regions
previously implicated in various inhibitory tasks, includingantisaccades [47]. Although we did not identify correlates
in the DLPFC specifically, which has been previously
shown to be involved in antisaccades [50], it is likely thatsuccessful performance of antisaccade tasks relies on
integration between these many regions.
Taken together, the results of the current study suggestthat antisaccade measures are sensitive to FOG highlight-
ing that the condition is specifically associated with deficits
in cognitive control which may interact with other net-works in the brain, presumably visual and motor, to cause
freezing episodes [4]. Our imaging results suggest that grey
matter atrophy in key visual and cognitive processingregions mediates performance on the task, highlighting that
these regions are likely to be critically involved in the
pathophysiology of FOG. As we have previously proposed,it is plausible that a more paroxysmal functional deficit in
communication between cortical regions described above,
and subcortical nuclei involved in locomotion are respon-sible for the phenomenon [51]. More specifically, such a
decoupling of these regions may lead to an overwhelminginhibition of locomotor nuclei via the subthalamic nucleus
[3]. How structural changes such as those found in this
study may influence this functional instability [51, 52] is aninteresting avenue for further research.
It is important to recognize, however, that the impaired
performance of antisaccades is not likely to be unique topatients with FOG, but rather appears to be more exten-
sively impaired in those patients that experience FOG.
Antisaccades have indeed been shown to be impaired in PDcompared to healthy controls, similarly to executive func-
tioning [13]. We also highlight that the current results were
found in a relatively small sample size and thus needreplication. An important future direction of study would
be to assess whether antisaccade performance can act as an
early neurophysiological marker for subsequent FOGevolution. Interestingly, antisaccade performance is
already in use as a potential predictor of motor and non-
motor progression in PD [16]. This is particularly relevantgiven the current sample did not show differences in
classical neuropsychological measures of global cognition
and executive function (e.g., MoCA and trail-making testscores). Therefore, saccadometry may provide a novel and
objective technique for tapping into early deficits, which
cannot be detected in more general neuropsychologicaltasks. Only more extensive longitudinal studies could
answer these questions.
These findings also point to rehabilitation strategieswhich target the fronto-parietal executive networks of the
brain, that are likely involved in the pathogenesis of
freezing episodes. As previously argued [53], we suggesttechniques such as cognitive training which is known to be
effective on executive functions in PD [54], or non-inva-
sive magnetic/electrical stimulation [55] are worth furtherexploration to this end. Finally, given the functional nature
of FOG [3, 4], antisaccade tasks could be employed while
patients undergo functional imaging [50], offering a novelwindow into the way in which executive and visual net-
works are dysfunctional in PD patients with FOG.
The current results have shown that patients with FOGshow a significant inability to inhibit reflexive saccades
prior to initiating goal-directed voluntary saccades. This
finding is aligned with the literature, and supportshypotheses of a cognitive control deficit in FOG. The VBM
J Neurol
123

analysis was able to uncover anatomical abnormalities
which may be responsible for these deficits, and infer thatcognitive control and visual-motor networks are central to
the behavioral deficits observed. Further research to
explore these processes in real-time are needed, and mayhelp to uncover important and vitally needed treatments for
this disabling condition.
Acknowledgments We thank the patients and their families whocontribute to our research at the Parkinson’s Disease Research Clinic.We also thank Mr. David Foxe for his help collecting some of the dataused in the study.
Compliance with ethical standards
Conflicts of interest CC Walton is supported by an AustralianPostgraduate Award at the University of Sydney. C. O’Callaghan issupported by a National Health and Medical Research Council ofAustralia (NHMRC) Neil Hamilton Fairley Postdoctoral Fellowship(GNT1091310) at the University of Cambridge. J.M. Hall is sup-ported by an International Postgraduate Research Scholarship at theUniversity of Western Sydney. M. Gilat is supported by an Interna-tional Postgraduate Research Scholarship at the University of Sydney.L. Mowszowski is supported by an Alzheimer’s Australia DementiaResearch Foundation Postdoctoral Research Fellowship at theUniversity of Sydney. S.L. Naismith is supported by an NHMRCCareer Development Award No. 1008117. J.R. Burrell was supportedin part by funding to Forefront, a collaborative research group dedi-cated to the study of frontotemporal dementia and motor neurondisease, from the NHMRC program Grant (APP1037746) and theAustralian Research Council Centre of Excellence in Cognition andits Disorders Memory Node (CE110001021). J.R. Burrell isalso supported by an NHMRC Early Career Fellowship(APP1072451). J.M. Shine is supported by a NHMRC CJ MartinPostdoctoral Fellowship (GNT1072403) at Stanford University.S.J.G. Lewis is supported by an NHMRC Practitioner Fellowship No.1003007.
References
1. Nutt JG, Bloem BR, Giladi N, Hallett M, Horak FB, NieuwboerA (2011) Freezing of gait: moving forward on a mysteriousclinical phenomenon. Lancet Neurol 10(8):734–744. doi:10.1016/S1474-4422(11)70143-0
2. Walton CC, Shine JM, Hall JM, O’Callaghan C, Mowszowski L,Gilat M, Szeto JY, Naismith SL, Lewis SJ (2015) The majorimpact of freezing of gait on quality of life in Parkinson’s disease.J Neurol 262(1):108–115. doi:10.1007/s00415-014-7524-3
3. Lewis SJG, Shine JM (2014) The next step: a common neuralmechanism for freezing of gait. Neuroscientist. doi:10.1177/1073858414559101
4. Lewis SJ, Barker RA (2009) A pathophysiological model offreezing of gait in Parkinson’s disease. Parkinsonism Relat Dis-ord 15(5):333–338. doi:10.1016/j.parkreldis.2008.08.006
5. Fasano A, Herman T, Tessitore A, Strafella AP, Bohnen NI(2015) Neuroimaging of freezing of gait. J Parkinson’s Dis.doi:10.3233/JPD-150536
6. Heremans E, Nieuwboer A, Spildooren J, Vandenbossche J,Deroost N, Soetens E, Kerckhofs E, Vercruysse S (2013) Cog-nitive aspects of freezing of gait in Parkinson’s disease: a chal-lenge for rehabilitation. J Neural Transm 120(4):543–557. doi:10.1007/s00702-012-0964-y
7. Burrell JR, Hornberger M, Carpenter RH, Kiernan MC, HodgesJR (2012) Saccadic abnormalities in frontotemporal dementia.Neurology 78(23):1816–1823. doi:10.1212/WNL.0b013e318258f75c
8. Terao Y, Fukuda H, Yugeta A, Hikosaka O, Nomura Y, SegawaM, Hanajima R, Tsuji S, Ugawa Y (2011) Initiation and inhibi-tory control of saccades with the progression of Parkinson’sdisease: changes in three major drives converging on the superiorcolliculus. Neuropsychologia 49(7):1794–1806. doi:10.1016/j.neuropsychologia.2011.03.002
9. Munoz DP, Everling S (2004) Look away: the anti-saccade taskand the voluntary control of eye movement. Nat Rev Neurosci5(3):218–228
10. Nieman DH, Bour LJ, Linszen DH, Goede J, Koelman JH,Gersons BP, Ongerboer de Visser BW (2000) Neuropsychologi-cal and clinical correlates of antisaccade task performance inschizophrenia. Neurology 54(4):866–871
11. Munoz DP, Armstrong IT, Hampton KA, Moore KD (2003)Altered control of visual fixation and saccadic eye movements inattention-deficit hyperactivity disorder. J Neurophysiol90(1):503–514. doi:10.1152/jn.00192.2003
12. Guitton D, Buchtel HA, Douglas RM (1985) Frontal lobe lesionsin man cause difficulties in suppressing reflexive glances and ingenerating goal-directed saccades. Exp Brain Res 58(3):455–472
13. Anderson TJ, MacAskill MR (2013) Eye movements in patientswith neurodegenerative disorders. Nat Rev Neurol 9(2):74–85.doi:10.1038/nrneurol.2012.273
14. Yerram S, Glazman S, Bodis-Wollner I (2013) Cortical control ofsaccades in Parkinson disease and essential tremor. J NeuralTransm 120(1):145–156. doi:10.1007/s00702-012-0870-3
15. Rivaud-Pechoux S, Vidailhet M, Brandel JP, Gaymard B (2007)Mixing pro- and antisaccades in patients with parkinsonian syn-dromes. Brain 130(Pt 1):256–264. doi:10.1093/brain/awl315
16. Antoniades CA, Demeyere N, Kennard C, Humphreys GW, HuMT (2015) Antisaccades and executive dysfunction in early drug-naive Parkinson’s disease: the discovery study. Mov Disord30(6):843–847. doi:10.1002/mds.26134
17. Amador SC, Hood AJ, Schiess MC, Izor R, Sereno AB (2006)Dissociating cognitive deficits involved in voluntary eye move-ment dysfunctions in Parkinson’s disease patients. Neuropsy-chologia 44(8):1475–1482. doi:10.1016/j.neuropsychologia.2005.11.015
18. Joti P, Kulashekhar S, Behari M, Murthy A (2007) Impairedinhibitory oculomotor control in patients with Parkinson’s dis-ease. Exp Brain Res 177(4):447–457. doi:10.1007/s00221-006-0687-0
19. Mosimann UP, Muri RM, Burn DJ, Felblinger J, O’Brien JT,McKeith IG (2005) Saccadic eye movement changes in Parkin-son’s disease dementia and dementia with Lewy bodies. Brain128(Pt 6):1267–1276. doi:10.1093/brain/awh484
20. Walton CC, Shine JM, Mowszowski L, Gilat M, Hall JM,O’Callaghan C, Naismith SL, Lewis SJ (2015) Impaired cogni-tive control in Parkinson’s disease patients with freezing of gaitin response to cognitive load. J Neural Transm 122(5):653–660.doi:10.1007/s00702-014-1271-6
21. Vandenbossche J, Deroost N, Soetens E, Zeischka P, SpildoorenJ, Vercruysse S, Nieuwboer A, Kerckhofs E (2012) Conflict andfreezing of gait in Parkinson’s disease: support for a responsecontrol deficit. Neuroscience 206:144–154. doi:10.1016/j.neuroscience.2011.12.048
22. Kostic VS, Agosta F, Pievani M, Stefanova E, Jecmenica-LukicM, Scarale A, Spica V, Filippi M (2012) Pattern of brain tissueloss associated with freezing of gait in Parkinson disease. Neu-rology 78(6):409–416. doi:10.1212/WNL.0b013e318245d23c
23. Tessitore A, Amboni M, Cirillo G, Corbo D, Picillo M, Russo A,Vitale C, Santangelo G, Erro R, Cirillo M, Esposito F, Barone P,
J Neurol
123

Tedeschi G (2012) Regional gray matter atrophy in patients withParkinson disease and freezing of gait. AJNR Am J Neuroradiol33(9):1804–1809. doi:10.3174/ajnr.A3066
24. Snijders AH, Leunissen I, Bakker M, Overeem S, Helmich RC,Bloem BR, Toni I (2011) Gait-related cerebral alterations inpatients with Parkinson’s disease with freezing of gait. Brain134(Pt 1):59–72. doi:10.1093/brain/awq324
25. Herman T, Rosenberg-Katz K, Jacob Y, Giladi N, Hausdorff JM(2014) Gray matter atrophy and freezing of gait in Parkinson’sdisease: is the evidence black-on-white? Mov Disord29(1):134–139. doi:10.1002/mds.25697
26. Sunwoo MK, Cho KH, Hong JY, Lee JE, Sohn YH, Lee PH(2013) Thalamic volume and related visual recognition areassociated with freezing of gait in non-demented patients withParkinson’s disease. Parkinsonism Relat Disord19(12):1106–1109. doi:10.1016/j.parkreldis.2013.07.023
27. Hughes AJ, Daniel SE, Kilford L, Lees AJ (1992) Accuracy ofclinical diagnosis of idiopathic Parkinson’s disease: a clinico-pathological study of 100 cases. J Neurol Neurosurg Psychiatry55(3):181–184
28. Emre M, Aarsland D, Brown R, Burn DJ, Duyckaerts C, MizunoY, Broe GA, Cummings J, Dickson DW, Gauthier S, Goldman J,Goetz C, Korczyn A, Lees A, Levy R, Litvan I, McKeith I,Olanow W, Poewe W, Quinn N, Sampaio C, Tolosa E, Dubois B(2007) Clinical diagnostic criteria for dementia associated withParkinson’s disease. Mov Disord 22(12):1689–1707. doi:10.1002/mds.21507 (quiz 1837)
29. Goetz CG, Tilley BC, Shaftman SR, Stebbins GT, Fahn S,Martinez-Martin P, Poewe W, Sampaio C, Stern MB, Dodel R,Dubois B, Holloway R, Jankovic J, Kulisevsky J, Lang AE, LeesA, Leurgans S, LeWitt PA, Nyenhuis D, Olanow CW, Rascol O,Schrag A, Teresi JA, van Hilten JJ, LaPelle N, MovementDisorder Society URTF (2008) Movement Disorder Society-sponsored revision of the Unified Parkinson’s Disease RatingScale (MDS-UPDRS): scale presentation and clinimetric testingresults. Mov Disord 23(15):2129–2170. doi:10.1002/mds.22340
30. Giladi N, Shabtai H, Simon ES, Biran S, Tal J, Korczyn AD(2000) Construction of freezing of gait questionnaire forpatients with Parkinsonism. Parkinsonism Relat Disord 6(3):165–170
31. Kitagawa M, Fukushima J, Tashiro K (1994) Relationshipbetween antisaccades and the clinical symptoms in Parkinson’sdisease. Neurology 44(12):2285–2289
32. Michell AW, Xu Z, Fritz D, Lewis SJ, Foltynie T, Williams-GrayCH, Robbins TW, Carpenter RH, Barker RA (2006) Saccadic latencydistributions in Parkinson’s disease and the effects of L-dopa. ExpBrain Res 174(1):7–18. doi:10.1007/s00221-006-0412-z
33. Hoehn MM, Yahr MD (1967) Parkinsonism: onset, progressionand mortality. Neurology 17(5):427–442
34. Ober JK, Przedpelska-Ober E, Gryncewicz W, Dylak J, CarpenterRHS, Ober JJ (2003) Hand-held system for ambulatory mea-surement of saccadic durations of neurological patients. In: GajdaJ (ed) Modelling and Measurement in Medicine Komitet Biocy-bernityki i Inzyneierii Biomedycznej PAN, Warsaw, pp 187–198
35. Carpenter RH, Williams ML (1995) Neural computation of loglikelihood in control of saccadic eye movements. Nature377(6544):59–62. doi:10.1038/377059a0
36. Burrell JR, Carpenter RH, Hodges JR, Kiernan MC (2013) Earlysaccades in amyotrophic lateral sclerosis. Amyotroph LateralScler Frontotemporal Degener 14(4):294–301. doi:10.3109/21678421.2013.783077
37. Tomlinson CL, Stowe R, Patel S, Rick C, Gray R, Clarke CE(2010) Systematic review of levodopa dose equivalency reportingin Parkinson’s disease. Mov Disord 25(15):2649–2653. doi:10.1002/mds.23429
38. Beck A, Steer R, Brown G (1996) Manual for the BDI-II. Psy-chological Corporation, San Antonio
39. Folstein MF, Folstein SE, McHugh PR (1975) ‘‘Mini-mentalstate’’. A practical method for grading the cognitive state ofpatients for the clinician. J Psychiatr Res 12(3):189–198
40. Nasreddine ZS, Phillips NA, Bedirian V, Charbonneau S,Whitehead V, Collin I, Cummings JL, Chertkow H (2005) TheMontreal cognitive assessment, MoCA: a brief screening tool formild cognitive impairment. J Am Geriatr Soc 53(4):695–699.doi:10.1111/j.1532-5415.2005.53221.x
41. Reitan RM (1979) Manual for administration of neuropsycho-logical test batteries for adults and children. NeuropsychologyLaboratory, Indiana University medical Center
42. Viliunas V, Lukauskien _e R, Svegzda A, Zukauskas A (2006)End-box scoring artefact evaluation of the Farnsworth–Munsell100-Hue colour vision test. Ophthalmic Physiol Opt26(6):580–586. doi:10.1111/j.1475-1313.2006.00411.x
43. Smith SM (2002) Fast robust automated brain extraction. HumBrain Mapp 17(3):143–155. doi:10.1002/hbm.10062
44. Zhang Y, Brady M, Smith S (2001) Segmentation of brain MRimages through a hidden Markov random field model and theexpectation-maximization algorithm. IEEE Trans Med Imaging20(1):45–57. doi:10.1109/42.906424
45. Andersson JL, Jenkinson M, Smith S (2007) Non-linear optimi-sation. FMRIB technical report TR07JA1. Practice 2007a Jun
46. Carpenter RHS (2012) Analysing the detail of saccadic reactiontime distributions. Biocybern Biomed Eng 32(2):49–63
47. Niendam TA, Laird AR, Ray KL, Dean YM, Glahn DC, CarterCS (2012) Meta-analytic evidence for a superordinate cognitivecontrol network subserving diverse executive functions. CognAffect Behav Neurosci 12(2):241–268. doi:10.3758/s13415-011-0083-5
48. Aron AR, Robbins TW, Poldrack RA (2014) Inhibition and theright inferior frontal cortex: one decade on. Trends Cogn Sci18(4):177–185. doi:10.1016/j.tics.2013.12.003
49. Matsuda T, Matsuura M, Ohkubo T, Ohkubo H, Matsushima E,Inoue K, Taira M, Kojima T (2004) Functional MRI mapping ofbrain activation during visually guided saccades and antisac-cades: cortical and subcortical networks. Psychiatry Res131(2):147–155. doi:10.1016/j.pscychresns.2003.12.007
50. Ettinger U, Ffytche DH, Kumari V, Kathmann N, Reuter B,Zelaya F, Williams SC (2008) Decomposing the neural correlatesof antisaccade eye movements using event-related FMRI. CerebCortex 18(5):1148–1159. doi:10.1093/cercor/bhm147
51. Shine JM, Matar E, Ward PB, Frank MJ, Moustafa AA, PearsonM, Naismith SL, Lewis SJ (2013) Freezing of gait in Parkinson’sdisease is associated with functional decoupling between thecognitive control network and the basal ganglia. Brain 136(Pt12):3671–3681. doi:10.1093/brain/awt272
52. Shen K, Hutchison RM, Bezgin G, Everling S, McIntosh AR(2015) Network structure shapes spontaneous functional con-nectivity dynamics. J Neurosci 35(14):5579–5588. doi:10.1523/JNEUROSCI.4903-14.2015
53. Walton CC, Shine JM, Mowszowski L, Naismith SL, Lewis SJ(2014) Freezing of gait in Parkinson’s disease: current treatmentsand the potential role for cognitive training. Restor NeurolNeurosci 32(3):411–422. doi:10.3233/RNN-130370
54. Leung IHK, Walton CC, Hallock H, Lewis SJG, Valenzuela M,Lampit A (2015) Cognitive training in Parkinson’s disease: asystematic review and meta-analysis. Neurology. doi:10.1212/WNL.0000000000002145
55. Lee SY, Kim MS, Chang WH, Cho JW, Youn JY, Kim YH(2014) Effects of repetitive transcranial magnetic stimulation onfreezing of gait in patients with Parkinsonism. Restor NeurolNeurosci 32(6):743–753. doi:10.3233/RNN-140397
J Neurol
123

APPENDIX(5((
Ehgoetz Martens, K. A., Hall, J. M., Gilat, M., Georgiades, M. J., Walton, C. C.,
Lewis, S. J. G. Anxiety is associated with freezing of gait and attentional set-shifting
in Parkinson's disease: A new perspective for early intervention. Gait & Posture,
2016
!

Full length article
Anxiety is associated with freezing of gait and attentional set-shiftingin Parkinson’s disease: A new perspective for early intervention
K.A.Ehgoetz Martensa,*, J.M. Halla,b, M. Gilata, M.J. Georgiadesa, C.C. Waltona,S.J.G. Lewisaa Parkinson’s Disease Research Clinic, Brain and Mind Centre, University of Sydney, 100 Mallet Street, Camperdown, New South Wales, 2050, Australiab School of Social Sciences and Psychology, Western Sydney University, Horsley Rd & Bullecourt Road, Milperra New South Wales, 2214, Australia
A R T I C L E I N F O
Article history:Received 19 February 2016Received in revised form 25 July 2016Accepted 26 July 2016
Keywords:AnxietyFreezing of gaitParkinson’s diseaseAttentionSet-shifting
A B S T R A C T
Previous research has shown that anxiety in Parkinson’s disease (PD) is associated with freezing of gait(FOG), and may even contribute to the underlying mechanism. However, limited research hasinvestigated whether PD patients with FOG (PD + FOG) have higher anxiety levels when compareddirectly to non-freezing PD patients (PD-NF) and moreover, how anxiety might contribute to FOG. Thecurrent study evaluated whether: (i) PD + FOG have greater anxiety compared to PD-NF, and (ii) anxiety inPD is related to attentional set-shifting, in order to better understand how anxiety might be contributingto FOG. In addition, we explored whether anxiety levels differed between those PD patients with mildFOG (PD + MildFOG) compared to PD-NF. Four hundred and sixty-one patients with PD (231 PD-NF, 180PD + FOG, 50 PD + MildFOG) were assessed using the Freezing of Gait Questionnaire item 3 (FOG-Q3),Hospital Anxiety and Depression Scale (HADS), Digit Span Test, Logical Memory Retention Test and TrailMaking Tests. Compared to PD-NF, PD + FOG had significantly greater anxiety (p < 0.001). PD + MildFOG,however, demonstrated similar levels of anxiety as the PD + FOG. In all patients, the severity of anxietysymptoms was significantly correlated to their degree of self-reported FOG on FOG-Q3 (p < 0.001) andTMT B-A (p = 0.039). Similar results were found for depression. In conclusion, these results confirm thekey role played by anxiety in FOG and also suggest that anxiety might be a promising biomarker for FOG.Future research should consider whether treating anxiety with pharmacological and/or cognitivebehavioural therapies at early stages of gait impairment in PD may alleviate troublesome FOG.
ã 2016 Elsevier B.V. All rights reserved.
1. Introduction
Anxiety disorders such as generalized anxiety disorder, panicdisorder and phobias have been argued to be one of the earliestmanifestations of Parkinson’s disease (PD) [1–3]. Patients with PDand their caregivers rank anxiety and falls as the top twosymptoms that require better management in order to improvequality of life [4,5]. One of the most common causes of falls in PD isfreezing of gait (FOG), which is characteristically described as atransient cessation of gait, where patients report feeling as thoughtheir feet become glued to the floor [6]. A relationship betweengeneralized symptoms of anxiety and severe gait impairments hasbeen noted [7–9], and mood disorders (such as generalized anxietyor depression) have also been suggested to be more common inthose with PD who experience FOG [7,10]. Panic attacks are also
often associated with freezing episodes [11,12], and recent findingshave shown that walking in threatening situations which provokeanxiety, also provoke more FOG episodes [9].
Although, the underlying pathophysiological mechanisms ofFOG are still unclear, recent research has proposed that anxietymight contribute to the underlying mechanism of FOG [9,11,13], byoverloading the capacity of the basal ganglia to process competingyet concurrent inputs (i.e. cognitive, sensorimotor, emotional)[13,14]. Interconnections between the limbic and motor circuitswithin the basal ganglia (nucleus accumbens) permit emotionalinput to interfere with motor outputs (resulting in FOG) [9].
Attention and executive function deficits are also features ofbasal ganglia pathology even in early PD. Attentional set-shiftingdeficits, which reflect disturbance in the fronto-striatal pathway[14–17], are often worse in PD + FOG compared to PD-NF [18–21].These cognitive impairments are especially detrimental to thosewho have severe gait impairments, since they rely on attention tocompensate for their movement impairment [22]. Given thatindividuals with PD, especially PD + FOG, rely on attention to
* Corresponding author.E-mail address: [email protected] (K.A.E. Martens).
http://dx.doi.org/10.1016/j.gaitpost.2016.07.1820966-6362/ã 2016 Elsevier B.V. All rights reserved.
Gait & Posture 49 (2016) 431–436
Contents lists available at ScienceDirect
Gait & Posture
journal homepage: www.else vie r .com/locate /gai t post

compensate for their gait impairments (i.e. loss of automaticity),anxiety might also contribute to FOG by consuming attentionalresources [23–25]. As a result, anxiety may interfere with executiveprocesses (e.g. attentional set-shifting) that are needed in order tocompensate for severe gait impairments [9,26,27]. Therefore,anxiety might play a key role in the development of FOG, and assuch may be important to consider for early interventions andfuture therapies to delay or prevent the onset of FOG.
The aim of the current study was to evaluate (i) whetherPD + FOG have greater levels of anxiety compared to PD-NF, (ii)whether anxiety in PD is associated with attentional set-shiftingdeficits, and (iii) whether anxiety might be a useful predictor ofthose who develop FOG. In addition, we explored whether anxietylevels differed between those patients with PD in the early stagesof gait impairment (PD + MildFOG) compared to PD-NF. Wehypothesized that both the PD + FOG and PD + MildFOG groupswill demonstrate higher clinical levels of anxiety compared to thePD-NF group. We also expected that attentional set-shiftingdeficits will be related to the severity of anxiety in PD. Finally, itwas expected that the severity of anxiety might predict those whoexperience FOG. By addressing these questions, we hope toextend previous research by examining the relationship betweenFOG, anxiety and attentional set-shifting, in order to provideinsight as to the feasibility of using anxiety as a FOG biomarkerand as a potential target for early interventions and future FOGtherapies.
2. Methods
2.1. Subjects
The subjects included in this study were recruited from alarger cohort of cases prospectively evaluated between 2008 and2015 at the PD Research Clinic at the Brain and Mind Centre,University of Sydney. All participants have a confirmed diagnosisof idiopathic PD (based on the United Kingdom Brain Bank clinicalcriteria) by a trained neurologist (SGJL), and all patients gavewritten informed consent to the study, which was approved bythe University of Sydney Human Research and Ethics Committee.Four hundred and sixty-one PD patients were included in thisstudy. Participants were divided into three groups (PD-NF,PD + FOG, PD + MildFOG) based on their score on the FOG-Questionnaire item 3 (FOG-Q3) (“Do you feel that your feet getglued to the floor while walking, making a turn or when trying toinitiate walking (freezing)?”) [28]. This measure is a reliablescreening tool to identify ‘freezers’[29]. Two hundred and thirty-one patients were classified as PD-NF (scored 0-“Not at all” onFOG-Q3), 180 patients were classified as PD + FOG (scoredbetween 2 and 4: 2-“rarely, about once a week”; 3-“often, aboutonce a day”; 4-“always, whenever walking” on FOG-Q3), and 50patients were classified as PD + MildFOG (scored of 1-“Very rarely,about once a month” on FOG-Q3). Exclusion criteria included: thepresence of neurological diseases other than PD, psychiatricdisorders other than an affective disorder, and/or mini-mentalstate examination (MMSE) score below 24 indicating dementia.One hundred and twenty-nine patients (62 PD-NF; 54 PD + FOG;13 PD + MildFOG) in this study were taking anti-depressants and/or anxiolytic medications.
2.2. Data collection and analyses
To evaluate anxiety and depression separately, the HospitalAnxiety and Depression Scale was administered. The ‘Anxiety’subscale (HADS-A) and ‘Depression’ subscale (HADS-D) wassubtotalled and compared between groups. FOG-Q3 was selectedto reflect FOG severity rather than the total FOG-Q score, based on
findings by Shine and colleagues which demonstrated that item 3from the FOG-Q was most strongly associated with actualbehavioural freezing events compared to the total score of theFOG-Q [29]. To evaluate attentional and executive functionsparticipants also completed the Trail Making Test Parts A (TMT-A) and B (TMT-B), as well as the Digit Span Test (backwards andforwards). The primary measure of set-shifting ability was thedifference in time to complete Parts A and B of the Trail Making test(TMT B-A). Whereas, attention and working memory was assessedby the forward and backward Digit Span respectively (refer toTable 1). In order to demonstrate a selective relationship betweenanxiety and executive functions, a Logical Memory Test (LM%retention) was also carried out. Since this test is not an executivefunction, it was not expected to be related to anxiety or FOG andthus acted as a control measure to assess whether the relationshipbetween anxiety, FOG and cognition is specific to executivefunctioning. Finally, global cognitive functioning was examinedwith MMSE. Participants were assessed on their regular dopami-nergic medication.
An ANOVA was conducted across the three groups andsignificant findings were further analysed using Tukey’s posthoc tests. Due to the group differences in MMSE, UPDRS-III andDisease duration at baseline, a covariate analysis was alsoconducted.
Pearson’s correlations were performed to test associationsbetween neuropsychiatric, neurocognitive, and clinical variables(reported in Table 2). Stepwise multiple regression analyses wereperformed to examine the relative contribution of neuropsychiat-ric, neurocognitive and disease related variables to FOG in PD.
In order to determine whether HADS anxiety score might be auseful measure to predict FOG, we collapsed the PD + MildFOG andPD + FOG into one PD + FOG group (N = 230), compared to the PD-NF group (N = 231). Then patients were categorized as havinganxiety and/or depression if they scored 8 or above on the HADS-Aand/or HADS-D section. Using this criterion, the PD-NF group had24 patients with anxiety and 207 without, as well as 23 patientswith depression, and 208 without (10 of the patients in the PD-NFgroup had both anxiety and depression). In the PD + FOG groupthere were 65 patients with anxiety and 165 without, and 66patients with depression and 164 without 39 of the patients in thePD + FOG group had both anxiety and depression. Odds ratio,positive predictive values and negative predictive values werereported.
3. Results
3.1. Group differences in anxiety
A significant main effect of group was found for anxiety (F(2,458) = 17.97, p < 0.001). PD + FOG had significantly greater levelsof anxiety compared to PD-NF (p < 0.001). However, PD + MildFOGwas not significantly different from PD + FOG (p = 0.18) (see Fig. 1).Notably, even when UPDRS-III, disease duration and MMSE wereentered as covariates, the group effect remained significant (F(2,371) = 11.92, p < 0.001). This group effect also remained signifi-cant when depression (HADS-D) was added as a covariate (F(2,368) = 4.23, p = 0.015), and likewise when TMT B-A andbackwards Digit Span were added as covariates (F(2,257) = 3.03,p = 0.05).
3.2. Group differences in executive functioning
A significant main effect of group was found for the time tocomplete TMT-A (F(2,289) = 8.27, p < 0.001), TMT-B (F(2,281) = 8.62, p < 0.001) and TMT B-A (F(2,281) = 7.22, p = 0.001).PD + FOG (p = 0.001) and PD + MildFOG (p = 0.041) took
432 K.A.E. Martens et al. / Gait & Posture 49 (2016) 431–436

significantly longer to complete TMT-A compared to PD-NF.PD + FOG (p = 0.001) and PD + MildFOG (p = 0.021) also tooksignificantly longer to complete TMT-B compared to PD-NF.However, only the PD + FOG group scored significantly higher onthe TMT B-A (p = 0.002) compared to PD-NF. The PD + MildFOGgroup scored marginally higher on the TMT B-A compared to thePD-NF (p = 0.058). It should be noted that only the group effect seen
in the TMT-A remained significant when UPDRS-III, Diseaseduration and MMSE were entered as covariates (TMT-A: F(2,273) = 3.08), p = 0.048, TMT-B: F(2,266) = 2.65, p = 0.073; TMTB-A: F(2,266) = 1.49, p = 0.228). The group effect for TMT-A alsoremained significant if HADS-A and HADS-D were added ascovariates (F(2,269) = 3.43, p = 0.037).
A significant main effect of group was also found for thebackwards Digit Span (F(2,288) = 6.07, p = 0.003). PD + FOG per-formed significantly worse on the backwards digit span comparedto PD-NF (p = 0.011). PD + MildFOG also performed significantlyworse on the backwards digit span compared to PD-NF (p = 0.027).The main effect of group did not remain significant when UPDRS-III, Disease duration and MMSE (F(2,270) = 2.55, p = 0.08) wereentered as covariates.
3.3. Predictors of freezing of gait
Disease duration, UPDRS-III, MMSE, HADS-A, HADS-D, TMT-A,TMT-B, TMT B-A, backwards Digit Span were entered into astepwise regression analysis to predict FOG-Q3 (Table 2). The bestpredictors of FOG were disease duration, depression, motorsymptom severity, and anxiety, which accounted for approximate-ly 28% of the variance (F(4,261) = 25.17, p < 0.001). The resultantregression equation was:
Predicted FOG = !0.304 + 0.057(disease duration) + 0.061(HADS-D) + 0.018(UPDRS-III)+ 0.057(HADS-A).
Table 2Correlations between FOG, neuropsychological, neuropsychiatric and demographicvariables.
FOG-Q3 HADS-A HADS-D
FOG-Q3 0.276*** 0.316***
HADS-A 0.276*** 0.619***
HADS-D 0.316*** 0.619***
Age 0.092 !0.120* 0.084UPDRS-III 0.379*** 0.123* 0.332***
Disease Duration 0.292*** 0.101* 0.187***
MMSE !0.154** !0.169** !0.229***
TMT-A 0.231*** 0.026 0.174**
TMT-B 0215*** 0.104 0.179**
TMT B-A 0.199** 0.117* 0.153*
Forward Digit Span !0.049 !0.006 !0.025Backwards Digit Span !0.165** !0.024 !0.029Logical Memory (% retention) !0.088 !0.012 !0.072
R values reported in the table.* Denotes a significant relationship at the p < 0.05 level.** Indicates a significant relationship at the p < 0.01 level.*** Denotes a significant relationship at the p < 0.001 level.
Fig. 1. highlights the group differences between PD + FOG and PD-NF on their HADSanxiety score. * Indicates significant difference (p < 0.05).
Table 1Demographic data and outcome measures of patient groups.
PD-NF PD + Mild FOG PD + FOG P-value
AgeM 67.8 69.3 69.2 P = 0.303SE 0.64 1.09 0.77N 228 49 173
Gender150M 34M 109M81F 16F 71F
Disease DurationM 3.8 6.3 8.7 P < 0.001SE 0.56 0.87 0.6N 218 47 164
Age of OnsetM 63.5 63.3 59.3 P = 0.008SE 0.7 1.4 1.4N 222 46 168
UPDRS-IIIM 28.3 33.5 41.5 *P < 0.001SE 1.1 2.5 1.3N 217 46 168
MMSEM 28.2 27.6 27.4 *P < 0.005SE 0.16 0.24 0.30N 203 41 148
FOG-Q3M 0 1 3 *P < 0.001SE 0 0 0.05N 231 50 180
HADS-AM 3.5 4.7 5.8 *P < 0.001SE 0.22 0.55 0.31N 231 50 180
HADS-DM 3.6 4.2 6.2 *P < 0.001SE 0.23 0.48 0.31N 230 50 179
TMT-A (sec)M 41.0 52.1 52.1 P = 0.072SE 1.47 5.17 2.74N 161 31 100
TMT-B (sec)M 101.8 141.6 138.6 P = 0.093SE 5.12 17.77 8.28N 159 31 94
TMT B-A (sec)M 61.2 89.5 89.8 P = 0.241SE 4.3 13.9 7.0N 159 31 94
Forward Digit SpanM 10.3 9.8 10.1 P = 0.801SE 0.19 0.43 0.26N 160 31 101
Backward Digit SpanM 6.8 5.7 6.0 P = 0.087SE 0.18 0.35 0.18N 160 31 100
LM% RetentionM 75.0 77.3 70.6 P = 0.353SE 1.57 3.33 2.06N 158 31 97
Note: Bolded p-values denote p-values from covariate analysis where Age, UPDRS-III and MMSE were factored out of outcome measure analysis. M = mean,SE = standard error, N = number of participants, UPDRS-III = Unified Parkinson’sDisease Rating Score, motor subsection.
* Reflects significant findings.
K.A.E. Martens et al. / Gait & Posture 49 (2016) 431–436 433

3.4. Using anxiety and depression to determine the odds of developingFOG
Finally, a significant odds ratio of 3.40 [95% CI: 2.04–5.66] wasfound (z = 4.69, p < 0.0001) when anxiety (HADS-A score 8 + ) wasused as the exposure variable and FOG as the outcome.Additionally, the HADS anxiety score yielded a Positive PredictiveValue of 28.26% [95% CI: 22.26%–34.55%] for identifying patientswith FOG and a Negative Predictive Value of 89.61% [95% CI:84.94%–93.23%] for identifying patients without FOG.
A similar odds ratio of 3.63 [95% CI: 2.17–6.1] was found (z = 4.9,p < 0.0001) when depression (HADS-D score 8 + ) was used as theexposure variable and FOG as the outcome. The HADS depressionscore yielded a similar Positive Predictive Value of 28.7% [95% CI:22.94%–35.01%] for identifying patients with FOG and a NegativePredictive Value of 90.04% [95% CI: 85.44%–93.58%] for identifyingpatients without FOG.
4. Discussion
We sought to determine whether anxiety might be a usefulpredictor of FOG by evaluating whether PD + FOG had more severegeneralized symptoms of anxiety compared to PD-NF and its’relation to executive function and attentional set-shifting deficits.First, PD + FOG reported significantly worse anxiety symptomscompared to the PD-NF group as we had predicted. It is noteworthythat this group effect remained significant after accounting formotor symptom severity, disease duration, global cognitive status,depression, set-shifting ability and working memory performance.Individuals in the Mild + FOG group demonstrated more anxietysymptoms than PD-NF but less than PD + FOG. Both freezer groupsalso demonstrated attention and executive function deficits.Second, anxiety and depression was significantly correlated withset-shifting ability and FOG. Third, anxiety and depression was asignificant predictor of FOG and provided a significant odds ratio aswell as promising positive and negative predictive values. To ourknowledge, this is the first study to provide evidence that anxietymay be a useful predictor of FOG, and also the first to demonstrate arelationship between anxiety and set-attentional shifting deficitsin PD.
Consistent with previous literature, the current study providesadditional evidence that anxiety and FOG are associated [7,10,11],and supports the notion that anxiety may be an importantindependent contributor to the underlying pathophysiology ofFOG [9]. Anxiety has been postulated to be a prodromal symptomof PD [30], resulting from damage to the brainstem nuclei (e.g.locus coeruleus), striatum and amygdala [31–36]. Furthermore,FOG has been suggested to manifest in instances where thedopamine-depleted striatum is unable to process the competingyet concurrent inputs from the cognitive-, motor- and limbic-loops [13,14]. Researchers have suggested that anxiety mightcontribute to FOG by overloading the limbic input into the striatumand thus interfere with competing information processing withinthe BG [9]. If anxiety does indeed present prior to motorsymptoms, then one explanation for why anxiety might predictFOG is that as the basal ganglia degenerates, attention becomesnecessary to compensate for movement impairments. However,anxiety or emotional dysregulation may also progressivelygenerate more interference by demanding greater attentionalresources and disrupting one’s ability to employ compensatorystrategies as the disease advances. Research has demonstrated thestrength of the limbic response, such that threat-related process-ing receives privileged access and priority to attentional processingresources, since the amygdala can allocate these resources throughthe ventral attentional network to prioritize threat-relatedprocessing and disrupt other cognitive processes [23–25]. In
support of this notion, greater anxiety was associated with longertime to switch attention between tasks in the current study.Previous research has also shown that anxiety in PD can disruptsensory feedback processing during walking, and provoke FOGepisodes [9,37]. Considering that anxiety might play a key role notonly provoking FOG but developing the phenomenon, this presentsa new avenue for FOG prevention and therapy. For example,treating anxiety with pharmacological and cognitive behaviouraltherapies when PD patients first develop freezing might alter theprogression and impact of this symptom. Unfortunately, there havebeen no studies to date that have reported treating FOG byattempting to reduce anxiety either pharmacologically or non-pharmacologically so further research is needed.
Based on this sample, the chance of someone having FOG was3.4 times more likely if PD patients had clinical anxiety (HADS-Ascore 8 + ) compared to those who did not. The HADS-A alsoprovided an excellent negative predictive value of 89.61% (i.e.correctly identifying those patients who do not have FOG),whereas it had a much more modest positive predictive value of28.26% (i.e. correctly identifying just over a quarter of thosepatients who do have FOG). Depression revealed a similar oddsratio, positive and negative predictive values as anxiety scores. Thiswas not a surprising result considering the comorbidity of theseaffective symptoms. Furthermore, both anxiety and depressionwere significant predictors of FOG in addition to disease durationand motor symptom severity. Taken together, these findingssuggest that both anxiety and depression are important factors toconsider when trying to predict individuals with PD who willdevelop FOG in order to implement early interventions.
It is important to point out that the HADS-A (or HADS-D) onlypredicted a quarter of individuals that had FOG. Therefore, theseaffective symptoms do not fully explain FOG but rather mightcontribute to FOG in some individuals. Indeed, given that FOGremains a difficult phenomenon to study, due to the heterogeneityand unpredictable episodes, it is possible that different subtypes ofPD + FOG exist and that anxiety/depression may represent onesignificant modifiable factor.
As predicted, we observed deficits in attentional and executivefunctions in both FOG groups. More specifically, set-shifting abilityand working memory was worse in PD + FOG and PD + MildFOGcompared to PD-NF. These findings are similar to previous reports[18–21] which have been argued to reflect disturbance infrontostriatal pathways [14–17]. Although both of these measureswere associated with FOG, only attentional set-shifting was relatedto anxiety and depression. This result in addition to the null resultsfound for logical memory provide support for our hypothesissuggesting that anxiety in PD is selectively related to attentionalset-shifting.
We would like to acknowledge some limitations within thisstudy. Although the reliability and convergent validity for HADS isgood, it has poor discriminant validity, and recently has not beenrecommended to diagnose anxiety in PD [38,39]. Others haveargued that HADS-A has clinical and research utility for evaluatinggeneralized anxiety symptom severity[40]. One of the majorconcerns is with the sensitivity rather than the specificity of HADSin PD. Taking this into consideration, our findings are likely aconservative representation of the relationship between anxiety/depression and FOG. We would recommend that future researchshould investigate this relationship between anxiety/depressionand FOG with more sensitive measures of anxiety and depression,and evaluate whether these measures can identify at riskindividuals who might respond to targeted early interventionaltherapy to reduce troublesome FOG.
It is also important to recognize that the relationship betweenanxiety and FOG does not dissociate whether anxiety precedesthe onset of FOG. PD + MildFOG reported more severe anxiety
434 K.A.E. Martens et al. / Gait & Posture 49 (2016) 431–436

than PD-NF but less than PD + FOG, but the PD + MildFOG did notstatistically differ from either group. This could be interpreted asindirectly supporting the notion that anxiety may precede FOG,since those with less FOG had similar levels of anxiety as thosewho experience FOG much more frequently. Alternatively, onemight suggest that the onset of FOG leads to heighted anxiety dueto the unpredictable nature and debilitating consequences ofthese episodes (such as falling). Unfortunately, a fall history orfear of falling report was not assessed in the current study. Werecommend that future research should consider implementingthese measures in order to determine whether anxiety might be apromising biomarker of FOG. Furthermore, limited research hasinvestigated the influence of anxiety on cognition in PD. In orderto better understand the relationship between anxiety, FOG andexecutive functions, more detailed neuropsychological investiga-tion needs to be done in order to understand how anxiety mightbe affecting cognition in individuals with PD prior to developingFOG.
In conclusion, this study provides direct evidence that anxiety isgreater in PD patients who experience troublesome FOG comparedto those who do not, and may be a useful predictor of those whomight develop FOG. This study also suggests that anxiety isassociated with set-shifting deficits common to PD + FOG. Futureresearch should consider whether treating anxiety with pharma-cological and/or cognitive behavioural therapies at early stages ofgait impairment in PD may alter the onset and progression of FOG.
Conflicts of interest
None.
Funding
KA Ehgoetz Martens is supported by an Australian EndeavourPostdoctoral Fellowship Award from the Australian Government.JM Hall is supported by an International Postgraduate ResearchScholarship at the University of Western Sydney. M Gilat issupported by an International Postgraduate Research Scholarshipat the University of Sydney. CC Walton is supported by anAustralian Postgraduate Award at the University of Sydney. SJGLewis is supported by an NHMRC-ARC Dementia Fellowship.
Disclosure of conflicts of interest
The funding sources of the study had not role in the design, datacollection, data management, data analysis, and data interpreta-tion of the study. The authors declare no financial or other conflictsof interest.
References
[1] R.D. Prediger, F.C. Matheus, M.L. Schwarzbold, M.M. Lima, M.A. Vital, Anxiety inParkinson’s disease: a critical review of experimental and clinical studies,Neuropharmacology 62 (2012) 115–124, doi:http://dx.doi.org/10.1016/j.neuropharm.2011.08.039.
[2] G.M. Pontone, J.R. Williams, K.E. Anderson, G. Chase, S.A. Goldstein, S. Grill,et al., Prevalence of anxiety disorders and anxiety subtypes in patients withParkinson’s disease, Mov. Disord. 24 (2009) 1333–1338, doi:http://dx.doi.org/10.1002/mds.22611.
[3] S. Coakeley, K.A. Ehgoetz Martens, Q.J. Almeida, Management of anxiety andmotor symptoms in Parkinson’s disease, Expert Rev. Neurother. 14 (2014) 937–946, doi:http://dx.doi.org/10.1586/14737175.2014.936388.
[4] K.H.O. Deane, H. Flaherty, D.J. Daley, R. Pascoe, B. Penhale, C.E. Clarke, et al.,Priority setting partnership to identify the top 10 research priorities for themanagement of Parkinson’s disease, BMJ Open 4 (2014), doi:http://dx.doi.org/10.1136/bmjopen-2014-006434 e006434–e006434.
[5] K.K. Hanna, A. Cronin-Golomb, Impact of anxiety on quality of life inParkinson’s disease, Parkinsons Dis. 2012 (2012) 640707, doi:http://dx.doi.org/10.1155/2012/640707.
[6] J.G. Nutt, B.R. Bloem, N. Giladi, M. Hallett, F.B. Horak, A. Nieuwboer, Freezing ofgait: moving forward on a mysterious clinical phenomenon, Lancet Neurol. 10(2011) 734–744, doi:http://dx.doi.org/10.1016/S1474-4422(11)70143-0.
[7] D.J. Burn, S. Landau, J.V. Hindle, M. Samuel, K.C. Wilson, C.S. Hurt, et al.,Parkinson’s disease motor subtypes and mood, Mov. Disord. 27 (2012) 379–386, doi:http://dx.doi.org/10.1002/mds.24041.
[8] K.A. Ehgoetz Martens, C.G. Ellard, Q.J. Almeida, Evaluating the link betweendopaminergic treatment, gait impairment, and anxiety in Parkinson’s disease,Mov. Disord. Clin. Pract. (2016), doi:http://dx.doi.org/10.1002/mdc3.12298.
[9] K.A. Ehgoetz Martens, C.G. Ellard, Q.J. Almeida, Does anxiety cause freezing ofgait in Parkinson’s disease? PLoS One 9 (2014) e106561, doi:http://dx.doi.org/10.1371/journal.pone.0106561.
[10] J.M. Hall, J.M. Shine, C. O’Callaghan, C.C. Walton, M. Gilat, S.L. Naismith, et al.,Freezing of gait and its associations in the early and advanced clinical motorstages of Parkinson’s disease: a cross-sectional study, J. Parkinsons Dis. 5 (4)(2015) 881–891, doi:http://dx.doi.org/10.3233/JPD-150581.
[11] A. Lieberman, Are freezing of gait (FOG) and panic related? J. Neurol. Sci. 248(2006) 219–222, doi:http://dx.doi.org/10.1016/j.jns.2006.05.023.
[12] I. Maidan, M. Plotnik, A. Mirelman, A. Weiss, N. Giladi, J.M. Hausdorff, Heartrate changes during freezing of gait in patients with Parkinson’s disease, Mov.Disord. 25 (2011) 2346–2354, doi:http://dx.doi.org/10.1002/mds.23280.
[13] S.J.G. Lewis, R.A. Barker, A pathophysiological model of freezing of gait inParkinson’s disease, Parkinsonism Relat. Disord. 15 (2009) 333–338, doi:http://dx.doi.org/10.1016/j.parkreldis.2008.08.006.
[14] S.J.G. Lewis, J.M. Shine, The next step: a common neural mechanism forfreezing of gait, Neuroscience (2014), doi:http://dx.doi.org/10.1177/1073858414559101 1073858414559101.
[15] S.L. Naismith, S.J.G. Lewis, The specific contributions of set-shifting to freezingof gait in Parkinson’s disease, Mov. Disord. 25 (2010) 1000–1004.
[16] N. Giladi, V. Huber-Mahlin, T. Herman, J.M. Hausdorff, Freezing of gait in olderadults with high level gait disorders: association with impaired executivefunction, J. Neural Transm. 114 (2007) 1349–1353.
[17] M. Amboni, A. Cozzolino, K. Longo, M. Picillo, P. Barone, Freezing of gait andexecutive functions in patients with Parkinson’s disease, Mov. Disord. 23(2008) 395–400.
[18] J.M. Shine, S.L. Naismith, N.C. Palavra, S.J.G. Lewis, S.T. Moore, V. Dilda, et al.,Attentional set-shifting deficits correlate with the severity of freezing of gait inParkinson’s disease, Parkinsonism Relat. Disord.19 (2013) 388–390, doi:http://dx.doi.org/10.1016/j.parkreldis.2012.07.015.
[19] J.M. Hall, J.M. Shine, C.C. Walton, M. Gilat, Y.P.T. Kamsma, S.L. Naismith, et al.,Early phenotypic differences between Parkinson’s disease patients with andwithout freezing of gait, Parkinsonism Relat. Disord. 20 (2014) 604–607, doi:http://dx.doi.org/10.1016/j.parkreldis.2014.02.028.
[20] C.C. Walton, J.M. Shine, L. Mowszowski, M. Gilat, J.M. Hall, C. O’Callaghan, et al.,Impaired cognitive control in Parkinson’s disease patients with freezing of gaitin response to cognitive load, J. Neural Transm. 122 (2015) 653–660.
[21] C.C. Walton, C. O’Callaghan, J.M. Hall, M. Gilat, L. Mowszowski, S.L. Naismith,et al., Antisaccade errors reveal cognitive control deficits in Parkinson’s diseasewith freezing of gait, J. Neurol. 262 (2015) 2745–2754, doi:http://dx.doi.org/10.1007/s00415-015-7910-5.
[22] L. Rochester, B. Galna, S. Lord, D. Burn, The nature of dual-task interferenceduring gait in incident Parkinson’s disease, Neuroscience 265 (2014) 83–94,doi:http://dx.doi.org/10.1016/j.neuroscience.2014.01.041.
[23] E. Fox, R. Russo, R. Bowles, K. Dutton, Do threatening stimuli draw or holdvisual attention in subclinical anxiety? J. Exp. Psychol. Gen. 130 (2001) 681–700.
[24] L. Pessoa, How do emotion and motivation direct executive control? TrendsCogn. Sci. 13 (2010) 160–166, doi:http://dx.doi.org/10.1016/j.tics.2009.01.006.
[25] P.C. Holland, M. Gallagher, Amygdala circuitry in attentional andrepresentational processes, Trends Cogn. Sci. 3 (1999) 65–73, doi:http://dx.doi.org/10.1016/S1364-6613(98)01271-6.
[26] K.A. Ehgoetz Martens, C.G. Ellard, Q.J. Almeida, Anxiety-provoked gait changesare selectively dopa-responsive in Parkinson’s disease, Eur. J. Neurosci. (2015)1–8, doi:http://dx.doi.org/10.1111/ejn.12928.
[27] K.A. Ehgoetz Martens, C.G. Ellard, Q.J. Almeida, Virtually-induced threat inParkinson’s: dopaminergic interactions between anxiety and sensory-perceptual processing while walking, Neuropsychologia (2015) 1–10, doi:http://dx.doi.org/10.1016/j.neuropsychologia.2015.05.015.
[28] N. Giladi, H. Shabtai, E.S. Simon, S. Biran, J. Tal, A.D. Korczyn, Construction offreezing of gait questionnaire for patients with Parkinsonism, ParkinsonismRelat. Disord. 6 (2000) 165–170, doi:http://dx.doi.org/10.1016/S1353-8020(99)00062-0.
[29] J.M. Shine, S.T. Moore, S.J. Bolitho, T.R. Morris, V. Dilda, S.L. Naismith, et al.,Assessing the utility of freezing of gait questionnaires in Parkinson’s disease,Parkinsonism Relat. Disord. 18 (2012) 25–29, doi:http://dx.doi.org/10.1016/j.parkreldis.2011.08.002.
[30] E. Jacob, N. Gatto, A. Thompson, Y. Bordelon, B. Ritz, Occurrence of depressionand anxiety prior to Parkinson’s disease, Parkinsonism Relat. Disord. 16 (2010)576–581, doi:http://dx.doi.org/10.1016/j.parkreldis.2010.06.014.
[31] P. Remy, M. Doder, A. Lees, N. Turjanski, D. Brooks, Depression in Parkinson’sdisease: loss of dopamine and noradrenaline innervation in the limbic system,Brain 128 (2005) 1314–1322, doi:http://dx.doi.org/10.1093/brain/awh445.
[32] H. Braak, E. Ghebremedhin, U. Rüb, H. Bratzke, K. Del Tredici, Stages in thedevelopment of Parkinson’s disease-related pathology, Cell Tissue Res. 318(2004) 121–134, doi:http://dx.doi.org/10.1007/s00441-004-0956-9.
K.A.E. Martens et al. / Gait & Posture 49 (2016) 431–436 435

[33] R.Y. Moore, Organization of midbrain dopamine systems and thepathophysiology of Parkinson’s disease, Parkinsonism Relat. Disord. 9(2003) 65–71, doi:http://dx.doi.org/10.1016/S1353-8020(03)00063-4.
[34] A. Tessitore, A.R. Hariri, F. Fera, W.G. Smith, T.N. Chase, T.M. Hyde, et al.,Dopamine modulates the response of the human amygdala: a study inParkinson’s disease, J. Neurosci. 22 (2002) 9099–9103.
[35] A.J. Harding, E. Stimson, J.M. Henderson, G.M. Halliday, Clinical correlates ofselective pathology in the amygdala of patients with Parkinson’s disease, Brain125 (2002) 2431–2445, doi:http://dx.doi.org/10.1093/brain/awf251.
[36] Y. Ouchi, E. Yoshikawa, H. Okada, M. Futatsubashi, Y. Sekine, M. Iyo, et al.,Alterations in binding site density of dopamine transporter in the striatum,orbitofrontal cortex, and amygdala in early Parkinson’s disease: compartmentanalysis for beta-CFT binding with positron emission tomography, Ann.Neurol. 45 (1999) 601–610.
[37] K.A. Ehgoetz Martens, C.G. Ellard, Q.J. Almeida, Virtually-induced threat inParkinson’s: dopaminergic interactions between anxiety and sensory-
perceptual processing while walking, Neuropsychologia 2 (2015), doi:http://dx.doi.org/10.1016/j.neuropsychologia.2015.05.015.
[38] A.F.G. Leentjens, K. Dujardin, L. Marsh, P. Martinez-Martin, I.H. Richard, S.E.Starkstein, et al., Anxiety rating scales in Parkinson’s disease: critique andrecommendations, Mov. Disord. 23 (2008) 2015–2025, doi:http://dx.doi.org/10.1002/mds.22233.
[39] E. Torbey, N.A. Pachana, N.N.W. Dissanayaka, Anxiety rating scales inParkinson’s disease: a critical review updating recent literature, J. Affect.Disord. 184 (2015) 216–224, doi:http://dx.doi.org/10.1016/j.jad.2015.05.059.
[40] A. Leentjens, K. Dujardin, L. Marsh, I.H. Richard, S.E. Starkstein, P. Martinez-Martin, Anxiety rating scales in Parkinson’s disease: a validation study of theHamilton anxiety rating scale, the Beck anxiety inventory, and the hospitalanxiety and depression scale, Mov. Disord. 26 (2011) 407–415.
436 K.A.E. Martens et al. / Gait & Posture 49 (2016) 431–436

APPENDIX(6((
(Ehgoetz Martens, K. A., Szeto, J. Y. Y., Muller, A. J., Hall, J. M., Gilat, M., Walton,
C. C., Lewis, S. J. G. Cognitive function in Parkinson's disease patients with and
without anxiety. Neurology Research International, 2016
!

Research ArticleCognitive Function in Parkinson’s Disease Patients with andwithout Anxiety
K. A. Ehgoetz Martens, J. Y. Y. Szeto, A. J. Muller, J. M. Hall, M. Gilat,C. C. Walton, and S. J. G. Lewis
Brain and Mind Centre, Faculty of Medicine, University of Sydney, 100Mallet Street, Camperdown, Sydney, NSW 2050, Australia
Correspondence should be addressed to K. A. Ehgoetz Martens; [email protected]
Received 1 August 2016; Revised 6 September 2016; Accepted 18 September 2016
Academic Editor: Jeff Bronstein
Copyright © 2016 K. A. Ehgoetz Martens et al.This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
Research on the implications of anxiety in Parkinson’s disease (PD) has been neglected despite its prevalence in nearly 50% ofpatients and its negative impact on quality of life. Previous reports have noted that neuropsychiatric symptoms impair cognitiveperformance in PD patients; however, to date, no study has directly compared PD patients with and without anxiety to examinethe impact of anxiety on cognitive impairments in PD.This study compared cognitive performance across 50 PD participants withand without anxiety (17 PDA+; 33 PDA−), who underwent neurological and neuropsychological assessment. Group performancewas compared across the following cognitive domains: simple attention/visuomotor processing speed, executive function (e.g., set-shifting), working memory, language, and memory/new verbal learning. Results showed that PDA+ performed significantly worseon the Digit Span forward and backward test and Part B of the Trail Making Task (TMT-B) compared to the PDA− group. Therewere no group differences in verbal fluency, logical memory, or TMT-A performance. In conclusion, anxiety in PD has ameasurableimpact on working memory and attentional set-shifting.
1. Introduction
Anxiety affects quality of life in those living with Parkinson’sdisease (PD) more so than overall cognitive status, motordeficits, apathy, and depression [1–3]. Although anxiety anddepression are often related and coexist in PD patients [4],recent research suggests that anxiety rather than depressionis themost prominent and prevalent mood disorder in PD [5,6]. Yet, our current understanding of anxiety and its impacton cognition in PD, as well as its neural basis and best treat-ment practices, remains meager and lags far behind that ofdepression [7].
Overall, neuropsychiatric symptoms in PD have beenshown to be negatively associated with cognitive perfor-mance. For example, higher depression scores have beencorrelated with lower scores on the Mini-Mental State Exam(MMSE) [8, 9] as well as tests of memory and executive func-tions (e.g., attention) [10–14]. Likewise, apathy and anhedoniain PD patients have been associated with executive dysfunc-tion [10, 15–23]. However, few studies have specifically inves-tigated the relationship between anxiety and cognition in PD.
One study showed a strong negative relationship betweenanxiety (both state and trait) and overall cognitive perfor-mance (measured by the total of the repeatable battery forthe assessment of neuropsychological status index) withina sample of 27 PD patients [24]. Furthermore, trait anxietywas negatively associated with each of the cognitive domainsassessed by the RBANS (i.e., immediatememory, visuospatialconstruction, language, attention, and delayedmemory). Twofurther studies have examined whether anxiety differentiallyaffects cognition in patients with left-sided dominant PD(LPD) versus right-sided dominant PD (RPD); however,their findings were inconsistent. The first study found thatworking memory performance was worse in LPD patientswith anxiety compared to RPD patients with anxiety [25],whereas the second study reported that, in LPD, apathy butnot anxiety was associated with performance on nonverballymediated executive functions and visuospatial tasks (e.g.,TMT-B,WMS-III spatial span), while in RPD, anxiety but notapathy significantly correlated with performance on verballymediated tasks (e.g., clock reading test and Boston naming
Hindawi Publishing Corporation
Neurology Research International
Volume 2016, Article ID 6254092, 7 pages
http://dx.doi.org/10.1155/2016/6254092

2 Neurology Research International
Table 1: Demographic and clinical variables in PD participants.
Variable PDA−(" = 33) PDA+
(" = 17) # valuesDemographicAge (years) 68.3 (8.9) 64.9 (8.1) # = 0.20Gender (% male) 19/14 (57.6%) 9/8 (52.9%) # = 0.75ClinicalDisease duration(years) 6.6 (4.2) 6.6 (5.6) # = 0.66UPDRS-III 31.9 (13.0) 31.2 (15.3) # = 0.65LPD/RPD ratio(% LPD) 15/17 (45.5%) 6/11 (35.3%) # = 0.38H&Y stage,median (range) 2.0 (1–4) 2.0 (1–5) # = 0.43LEDD, mg/day 674.0 (550.5) 710.2 (478.7) # = 0.80HADS-A 2.03 (1.6) 10.5 (1.9) ∗# < 0.001HADS-D 3.2 (1.7) 4.1 (1.6) # = 0.06HADS-total 5.2 (2.7) 14.7 (2.9) ∗# < 0.001Note: values reflect mean (SD). LEDD: levodopa equivalent daily dose. ∗indicates significant group differences.
test) [15]. Furthermore, anxiety was significantly correlatedwith neuropsychological measures of attention and executiveand visuospatial functions [15]. Taken together, it is evidentthat there are limited and inconsistent findings describing therelationship between anxiety and cognition in PD and morespecifically how anxiety might influence particular domainsof cognition such as attention and memory and executivefunctioning. It is also striking that, to date, no study hasexamined the influence of anxiety on cognition in PD bydirectly comparing groups of PD patients with and withoutanxiety while excluding depression. This was the primaryobjective of the current study.
Given that research on healthy young adults suggests thatanxiety reduces processing capacity and impairs processingefficiency, especially in the central executive and attentionalsystems of working memory [26, 27], we hypothesizedthat PD patients with anxiety would show impairments inattentional set-shifting and working memory compared toPD patients without anxiety. Furthermore, since previouswork, albeit limited, has focused on the influence of symptomlaterality on anxiety and cognition, we also explored thisrelationship.
2. Methods
Seventeen PD patients with anxiety and thirty-three PDpatients without anxiety were included in this study (seeTable 1).The cross-sectional data from these participants wastaken from a patient database that has been compiled over thepast 8 years (since 2008) at the Parkinson’s Disease ResearchClinic at the Brain and Mind Centre, University of Sydney.Inclusion criteria involved a diagnosis of idiopathic PDaccording to theUnitedKingdomParkinson’sDisease SocietyBrain Bank criteria [28] and were confirmed by a neurologist(SJGL). Patients also had to have an adequate proficiency
in English and have completed a full neuropsychologicalassessment. Ten patients in this study (5 PD with anxiety;5 PD without anxiety) were taking psychotropic drugs (i.e.,benzodiazepine or selective serotonin reuptake inhibitor).Patients with an MMSE of less than 24 were excluded.Patients were also excluded if they had other neurologicaldisorders, psychiatric disorders other than affective disorders(such as anxiety), or if they reported a score greater thansix on the depression subscale of the Hospital Anxiety andDepression Scale (HADS). Thus, all participants who scoredwithin a “depressed” (HADS-D > 6) range were excludedfrom this study, in attempt to examine a refined sample ofPD patients with and without anxiety in order to determinethe independent effect of anxiety on cognition.This researchwas approved by the Human Research Ethics Committee ofthe University of Sydney, and written informed consent wasobtained from all participants.
Self-reported HADS was used to assess anxiety in PDand has been previously shown to be a useful measure ofclinical anxiety in PD [29]. A cut-off score of>8on the anxietysubscale of the HADS (HADS-A) was used to identify PDcases with anxiety (PDA+), while a cut-off score of <6 onthe HADS-A was used to identify PD cases without anxiety(PDA−).This criterionwasmore stringent thanusual (>7 cut-off score) [30], in effort to create distinct patient groups.
All participants underwent a neurological assessmentconducted while on their medications. The neurologicalevaluation rated participants according to Hoehn and Yahr(H&Y) stages [31] and assessed their motor symptoms usingpart III of the revised MDS Task Force Unified Parkinson’sDisease Rating Scale (UPDRS) [32]. In a similar way, symp-tom laterality was determined by taking a left/right ratio.Thiswas determined by calculating a total left and right scorefrom rigidity items 30–35, voluntary movement items 36–43,and tremor items 50–57 from the MDS-UPDRS part III (seeTable 1).
Neuropsychological functioning was assessed using stan-dardized tests and appropriate normative data. A detaileddescription of the neuropsychological battery was reportedelsewhere [33]. Processing speed was assessed using the TrailMaking Test, Part A (TMT-A, z-score) [34]. Attentional set-shifting was measured using the Trail Making Test, PartB (TMT-B, z-score) [34]. Working memory was assessedusing the Digit Span forward and backward subtest of theWechsler Memory Scale-III (raw scores) [35]. Language wasassessed with semantic and phonemic verbal fluency via theControlled Oral Word Associated test (COWAT animals andletters, z-score) [36]. The ability to retain learned verbalmemory was assessed using the Logical Memory subtestfrom the Wechsler Memory Scale-III (LM-I z-score, LM-IIz-score, % LM retention z-score) [35].TheMini-Mental StateExamination (MMSE) was used to assess global cognition.
2.1. Statistical Analyses. The Shapiro-Wilk test was performedto determine normal distribution.Demographic, clinical, andneuropsychological variables were compared between thetwo groups with the independent t-test or Mann–Whitney$ test, depending on whether the variable met parametricassumptions. Chi-square tests were used to examine gender

Neurology Research International 3
Table 2: Neuropsychological performance.
PDA−(" = 33) PDA+
(" = 17) # valuesGlobal cognitionMMSE 28.2 (1.5) 27.5 (1.8) # = 0.23Attention/visuomotorprocessing speedTMT-A, %-score −0.15 (1.2) 0.66 (2.23) # = 0.62Executive functionTMT-B, %-score −0.02 (1.1) −0.85 (1.8) ∗# = 0.048Working memoryDigit Span forward total 10.6 (2.3) 8.8 (3.2) ∗# = 0.031Digit Span backward total 7.2 (2.8) 5.7 (1.5) ∗# = 0.012LanguageSemantic verbal fluency,%-score −0.04 (1.0) −0.26 (1.1) # = 0.49Phonemic verbal fluency,%-score −0.10 (1.1) −0.23 (1.2) # = 0.70Memory/new verballearningLM-I immediate recall,%-score −0.21 (1.1) −0.83 (1.1) # = 0.059LM-II, %-score 0.10 (1.1) −0.29 (1.1) # = 0.20% LM retention, %-score 0.40 (1.2) 0.14 (1.2) # = 0.43Note: values represent mean (SD). ∗ indicates significant differences at the" < 0.05 level.and symptom laterality differences between groups. All anal-yses employed an alpha level of # < 0.05 and were two-tailed. Spearman correlations were performed separately ineach group to examine associations between anxiety and/ordepression ratings and cognitive functions.
3. Results
3.1. Demographic and Clinical Characteristics. As expected,the PDA+ group reported significant greater levels of anxietyon the HADS-A ($ = 0, # < 0.001) and higher total scoreon the HADS ($ = 1, # < 0.001) compared to the PDA−group (Table 1). Groups were matched in age (&(48) = 1.31,# = 0.20), disease duration ($ = 259, # = 0.66), UPDRS-III score ($ = 250.5, # = 0.65), H&Y ($ = 245, # = 0.43),LEDD ($ = 159.5, # = 0.80), and depression (HADS-D)($ = 190.5, # = 0.06). Additionally, all groups were matchedin the distribution of gender ('2 = 0.098, # = 0.75) and side-affected ('2 = 0.765, # = 0.38).3.2. Cognitive Functioning. There were no group differencesfor TMT-A performance ($ = 256, # = 0.62) (Table 2); how-ever, the PDA+ group had worse performance on the TrailMaking Test Part B (&(46) = 2.03, # = 0.048) compared to thePDA− group (Figure 1).The PDA+ group also demonstratedsignificantly worse performance on the Digit Span forwardsubtest (&(48) = 2.22, # = 0.031) and backward subtest ($ =190.5, # = 0.016) compared to the PDA− group (Figures 2(a)
∗0.40.20
−0.2−0.4−0.6−0.8−1
−1.2−1.4 PDA− PDA+
TMT-
B (z
-sco
re)
Figure 1: PD patients with anxiety performed significantly worse onPart B of the Trail Making Task compared to PD patients withoutanxiety. ∗ denotes # values < 0.05. Error bars represent standarderror.
and 2(b)). Neither semantic verbal fluency (&(47) = 0.70, # =0.49) nor phonemic verbal fluency (&(47) = 0.39, # = 0.70)differed between groups. Logical Memory I immediate recalltest ($ = 176, # = 0.059) showed a trend that the PDA+group had worse new verbal learning and immediate recallabilities than the PDA− group. However, Logical Memory IItest performance ($ = 219, # = 0.204) and Logical Memory% retention ($ = 242.5, # = 0.434) did not differ betweengroups. There were also no differences between groups inglobal cognition (MMSE) ($ = 222.5, # = 0.23).3.3. Comparison within LPD and RPD Patients. Partici-pants were split into LPD and RPD, and then furthergroup differences were examined between PDA+ and PDA−.Importantly, the groups remained matched in age, diseaseduration, UPDRS-III, DDE, H&Y stage, and depressionbut remained significantly different on self-reported anxiety.LPDA+ demonstrated worse performance on the Digit Spanforward test (&(19) = 2.29, # = 0.033) compared to LPDA−,whereas RPDA+ demonstrated worse performance on theDigit Span backward test ($ = 36.5, # = 0.006), LM-Iimmediate recall ($ = 37.5,# = 0.008), and LM-II ($ = 45.0,# = 0.021) but not LM % retention ($ = 75.5, # = 0.39)compared to RPDA−.4. Discussion
This study is the first to directly compare cognition betweenPDpatients with andwithout anxiety.The findings confirmedour hypothesis that anxiety negatively influences attentionalset-shifting andworkingmemory in PD.More specifically, wefound that PD patients with anxiety were more impaired onthe Trail Making Test Part B which assessed attentional set-shifting, on both Digit Span Tests which assessed workingmemory and attention, and to a lesser extent on the Log-ical Memory test which assessed memory and new verballearning compared to PD patients without anxiety. Takentogether, these findings suggest that anxiety in PD mayreduce processing capacity and impair processing efficiency,especially in the central executive and attentional systems ofworking memory in a similar way as seen in young healthyadults [26, 27].

4 Neurology Research International
∗
PDA− PDA+
12
10
8
6
4
2
0
Dig
it Sp
an fo
rwar
ds
(a)
∗
PDA− PDA+
12
10
8
6
4
2
0
Dig
it Sp
an b
ackw
ards
(b)
Figure 2: PD patients with anxiety performed significantly worse on the Digit Span forward (a) and backward (b) test compared to PDpatients without anxiety. ∗ denotes # values < 0.05. Error bars represent standard error of the means.
Although the neurobiology of anxiety in PD remainsunknown, many researchers have postulated that anxietydisorders are related to neurochemical changes that occurduring the early, premotor stages of PD-related degeneration[37, 38] such as nigrostriatal dopamine depletion, as wellas cell loss within serotonergic and noradrenergic brainstemnuclei (i.e., raphe nuclei and locus coeruleus, resp., whichprovide massive inputs to corticolimbic regions). Over time,chronic dysregulation of adrenocortical and catecholaminefunctions can lead to hippocampal damage as well as dys-functional prefrontal neural circuitries [39, 40], which playa key role in memory and attention [24]. Recent functionalneuroimaging work has suggested that enhanced hippocam-pal activation during executive functioning and workingmemory tasks may represent compensatory processes forimpaired frontostriatal functions in PD patients comparedto controls [41]. Therefore, chronic stress from anxiety, forexample, may disrupt compensatory processes in PD patientsand explain the cognitive impairments specifically inworkingmemory and attention seen in PD patients with anxiety. It hasalso been suggested that hyperactivation within the putamenmay reflect a compensatory striatal mechanism to maintainnormal working memory performance in PD patients; how-ever, losing this compensatory activation has been shownto contribute to poor working memory performance [42].Anxiety in mild PD has been linked to reduced putamendopamine uptake which becomes more extensive as the
disease progresses [43].This further supports the notion thatanxiety may disrupt compensatory striatal mechanisms aswell, providing another possible explanation for the cognitiveimpairments observed in PD patients with anxiety in thisstudy.
Noradrenergic and serotonergic systems should also beconsidered when trying to explain the mechanisms by whichanxiety may influence cognition in PD. Although these neu-rotransmitter systems are relatively understudied in PD cog-nition, treating the noradrenergic and serotonergic systemshas shown beneficial effects on cognition in PD. Selectiveserotonin reuptake inhibitor, Citalopram, was shown toimprove response inhibition deficits in PD [44], while nora-drenaline reuptake blocker, atomoxetine, has been recentlyreported to have promising effects on cognition in PD [45,46].
Overall, very few neuroimaging studies have been con-ducted in PD in order to understand the neural correlatesof PD anxiety and its underlying neural pathology. Futureresearch should focus on relating anatomical changes andneurochemical changes to neural activation in order to gaina clearer understanding on how these pathologies affectanxiety in PD. To further understand how anxiety andcognitive dysfunction are related, future research shouldfocus on using advanced structural and function imagingtechniques to explain both cognitive and neural breakdownsthat are associated with anxiety in PD patients. Research has

Neurology Research International 5
indicated that those with amnestic mild cognitive impair-ment who have more neuropsychiatric symptoms have agreater risk of developing dementia compared to those withfewer neuropsychiatric symptoms [10]. Future studies shouldalso examine whether treating neuropsychiatric symptomsmight impact the progression of cognitive decline andimprove cognitive impairments in PD patients.
4.1. Does Symptom Laterality Influence the Effect of Anxietyon Cognition? Previous studies have used PD symptomlaterality as a window to infer asymmetrical dysfunctionof neural circuits. For example, LPD patients have greaterinferred right hemisphere pathology, whereas RPD patientshave greater inferred left hemisphere pathology [15]. Thus,cognitive domains predominantly subserved by the left hemi-sphere (e.g., verballymediated tasks of executive function andverbal memory) might be hypothesized to be more affectedin RPD than LPD; however, this remains controversial [47].It has also been suggested that since anxiety is a commonfeature of left hemisphere involvement [48, 49], cognitivedomains subserved by the left hemisphere may also be morestrongly related to anxiety [15]. Results from this studyshowed selective verbal memory deficits in RPD patientswith anxiety compared to RPD without anxiety, whereasLPD patients with anxiety had greater attentional/workingmemory deficits compared to LPDwithout anxiety. Althoughthese results align with previous research [15], interpretationsof thesefindings should bemadewith caution due to the smallsample size in the LPD comparison specifically.
4.2. Limitations, Considerations, and Future Directions. Wewould like to acknowledge a few shortcomings of this study.Recent work has suggested that the HADS questionnaire mayunderestimate the burden of anxiety related symptomologyand therefore be a less sensitive measure of anxiety in PD[30, 50]. In addition, our small sample size also limited thestatistical power for detecting significant findings. Based onthese limitations, our findings are likely conservative andunderrepresent the true impact anxiety has on cognition inPD. Additionally, the current study employed a very briefneuropsychological assessment including one or two testsfor each cognitive domain. Future studies are encouraged tocollect a more complex and comprehensive battery from alarger sample of PDparticipants in order to better understandthe role anxiety plays on cognition in PD.
Another limitation of this study was the absence ofdiagnostic interviews to characterize participants’ psychiatricsymptoms and specify the type of anxiety disorders includedin this study. Future studies should perform diagnosticinterviews with participants (e.g., using DSM-V criteria)rather than relying on self-reported measures to group par-ticipants, in order to better understand whether the type ofanxiety disorder (e.g., social anxiety, phobias, panic disorders,and generalized anxiety) influences cognitive performancedifferently in PD.
One advantage the HADS questionnaire provided overother anxiety scales was that it assessed both anxiety anddepression simultaneously and allowed us to control forcoexisting depression. Although there was a trend that the
PDA+ group self-reported higher levels of depression thanthe PDA− group, all participants included in the study scored<6 on the depression subscale of the HADS. Controlling fordepressionwhile assessing anxiety has been identified as a keyshortcoming in the majority of recent work [10]. Consideringmany previous studies have investigated the influence ofdepression on cognition in PD without accounting for thepresence of anxiety and the inconsistent findings reportedto date, we recommend that future research should try todisentangle the influence of anxiety versus depression oncognitive impairments in PD.
Considering the growing number of clinical trials fortreating depression, there are few if any for the treatment ofanxiety in PD. Anxiety is a key contributor to decreased qual-ity of life in PD and greatly requires better treatment options.Moreover, anxiety has been suggested to play a key role infreezing of gait (FOG) [51], which is also related to atten-tional set-shifting [52, 53]. Future research should examinethe link between anxiety, set-shifting, and FOG, in orderto determine whether treating anxiety might be a potentialtherapy for improving FOG.
Competing Interests
The authors have no competing interests to report.
Acknowledgments
This work was supported by a Postdoctoral Fellowship fromParkinson’s Canada (K. A. Ehgoetz Martens); an Interna-tional Postgraduate Research Scholarship at the University ofWestern Sydney (J. M. Hall); an International PostgraduateResearch Scholarship at the University of Sydney (M. Gilatand C. C. Walton); a NHMRC-ARC Dementia Fellowship (S.J. G. Lewis). The authors would like to thank the patientsand their families who contributed to the research at theParkinson’s Disease Research Clinic.
References
[1] K. K. Hanna and A. Cronin-Golomb, “Impact of anxiety onquality of life in Parkinson’s disease,” Parkinson’s Disease, vol.2012, Article ID 640707, 8 pages, 2012.
[2] J. D. Jones, L. C. Butterfield,W. Song et al., “Anxiety and depres-sion are better correlates of Parkinson’s disease quality of lifethan apathy,” Journal of Neuropsychiatry and Clinical Neurosci-ences, vol. 27, no. 3, pp. 213–218, 2015.
[3] K. H. O. Deane, H. Flaherty, D. J. Daley et al., “Priority settingpartnership to identify the top 10 research priorities for themanagement of Parkinson’s disease,” BMJ Open, vol. 4, no. 12,Article ID e006434, 2014.
[4] L. Marsh, “Neuropsychiatric aspects of Parkinson’s disease,”Psychosomatics, vol. 41, no. 1, pp. 15–23, 2000.
[5] S. Landau, V. Harris, D. J. Burn et al., “Anxiety and anxious-depression in Parkinson’s disease over a 4-year period: a latenttransition analysis,” Psychological Medicine, vol. 46, no. 3, pp.657–667, 2016.
[6] R. G. Brown, S. Landau, J. V. Hindle et al., “Depression and anx-iety related subtypes in Parkinson’s disease,” Journal of Neuro-logy, Neurosurgery and Psychiatry, vol. 82, no. 7, pp. 803–809,2011.

6 Neurology Research International
[7] K. Seppi, D.Weintraub,M. Coelho et al., “Themovement disor-der society evidence-based medicine review update: treatmentsfor the non-motor symptoms of Parkinson’s disease,”MovementDisorders, vol. 26, supplement 3, pp. S42–S80, 2011.
[8] H. H. Fernandez, R. H. See, M. F. Gary et al., “Depressive symp-toms in Parkinson disease correlate with impaired global andspecific cognitive performance,” Journal of Geriatric Psychiatryand Neurology, vol. 22, no. 4, pp. 223–227, 2009.
[9] E. Cubo, B. Bernard, S. Leurgans, and R. Raman, “Cognitive andmotor function in patients with Parkinson’s disease with andwithout depression,” Clinical Neuropharmacology, vol. 23, no. 6,pp. 331–334, 2000.
[10] H. Alzahrani and A. Venneri, “Cognitive and neuroanatomicalcorrelates of neuropsychiatric symptoms in Parkinson’s disease:a systematic review,” Journal of the Neurological Sciences, vol.356, no. 1-2, pp. 32–44, 2015.
[11] A. Costa, A. Peppe, G. A. Carlesimo, P. Pasqualetti, and C. Cal-tagirone, “Major andminor depression in Parkinson’s disease: aneuropsychological investigation,” European Journal of Neuro-logy, vol. 13, no. 9, pp. 972–980, 2006.
[12] E. Stefanova, A. Potrebic, L. Ziropadja, J. Maric, I. Ribaric,and V. S. Kostic, “Depression predicts the pattern of cognitiveimpairment in early Parkinson’s disease,” Journal of the Neuro-logical Sciences, vol. 248, no. 1-2, pp. 131–137, 2006.
[13] J. Uekermann, I. Daum, S. Peters, B. Wiebel, H. Przuntek, andT. Muller, “Depressed mood and executive dysfunction in earlyParkinson’s disease,”ActaNeurologica Scandinavica, vol. 107, no.5, pp. 341–348, 2003.
[14] G. Spalletta, S. Fagioli, G. Meco et al., “Hedonic tone and itsmood and cognitive correlates in Parkinson’s disease,” Depres-sion and Anxiety, vol. 30, no. 1, pp. 85–91, 2013.
[15] Y. Bogdanova and A. Cronin-Golomb, “Neurocognitive corre-lates of apathy and anxiety in parkinson’s disease,” Parkinson’sDisease, vol. 2012, Article ID 793076, 9 pages, 2012.
[16] L. C. Butterfield, C. R. Cimino, L. E. Oelke, R. A. Hauser, andJ. Sanchez-Ramos, “The independent influence of apathy anddepression on cognitive functioning in Parkinson’s disease,”Neuropsychology, vol. 24, no. 6, pp. 721–730, 2010.
[17] K. Dujardin, P. Sockeel,M.Delliaux, A. Destee, and L. Defebvre,“Apathy may herald cognitive decline and dementia in Parkin-son’s disease,”MovementDisorders, vol. 24, no. 16, pp. 2391–2397,2009.
[18] V. Isella, P. Melzi, M. Grimaldi et al., “Clinical, neuropsycho-logical, and morphometric correlates of apathy in Parkinson’sdisease,”Movement Disorders, vol. 17, no. 2, pp. 366–371, 2002.
[19] G. C. Pluck and R. G. Brown, “Apathy in Parkinson’s disease,”Journal of Neurology Neurosurgery and Psychiatry, vol. 73, no. 6,pp. 636–642, 2002.
[20] D. J. Zgaljardic, J. C. Borod, N. S. Foldi et al., “Relationshipbetween self-reported apathy and executive dysfunction innondemented patients with Parkinson disease,” Cognitive andBehavioral Neurology, vol. 20, no. 3, pp. 184–192, 2007.
[21] G. Santangelo, C. Vitale, L. Trojano et al., “Relationship betweendepression and cognitive dysfunctions in Parkinson’s diseasewithout dementia,” Journal of Neurology, vol. 256, no. 4, pp. 632–638, 2009.
[22] G. Santangelo, L. Morgante, R. Savica et al., “Anhedonia andcognitive impairment in Parkinson’s disease: Italian validationof the Snaith-Hamilton Pleasure Scale and its application in theclinical routine practice during the PRIAMO study,” Parkin-sonism and Related Disorders, vol. 15, no. 8, pp. 576–581, 2009.
[23] F. Assogna, L. Cravello, C. Caltagirone, and G. Spalletta, “Anhe-donia in Parkinson’s disease: a systematic review of the liter-ature,”Movement Disorders, vol. 26, no. 10, pp. 1825–1834, 2011.
[24] K. A. Ryder, S. T. Gontkovsky, K. L. Mcswan et al., “Cognitivefunction in Parkinson’s disease: association with anxiety but notdepression,”Aging, Neuropsychology, andCognition, vol. 9, no. 2,pp. 77–84, 2002.
[25] P. S. Foster, V. Drago, G. P. Crucian et al., “Anxiety and depres-sion severity are related to right but not left onset Parkinson’sdisease duration,” Journal of the Neurological Sciences, vol. 305,no. 1-2, pp. 131–135, 2011.
[26] M. W. Eysenck, N. Derakshan, R. Santos, and M. G. Calvo,“Anxiety and cognitive performance: attentional control the-ory,” Emotion, vol. 7, no. 2, pp. 336–353, 2007.
[27] M.W. Eysenck, C. MacLeod, and A. Mathews, “Cognitive func-tioning and anxiety,” Psychological Research, vol. 49, no. 2-3, pp.189–195, 1987.
[28] A. J. Hughes, S. E. Daniel, L. Kilford, and A. J. Lees, “Accuracyof clinical diagnosis of idiopathic Parkinson’s disease: a clinico-pathological study of 100 cases,” Journal of Neurology, Neuro-surgery & Psychiatry, vol. 55, no. 3, pp. 181–184, 1992.
[29] J. Marinus, A. F. G. Leentjens, M. Visser, A. M. Stiggelbout, andJ. J. Van Hilten, “Evaluation of the hospital anxiety and depres-sion scale in patients with Parkinson’s disease,” Clinical Neu-ropharmacology, vol. 25, no. 6, pp. 318–324, 2002.
[30] I. Bjelland, A. A. Dahl, T. T. Haug, and D. Neckelmann, “Thevalidity of the hospital anxiety and depression scale: an updatedliterature review,” Journal of Psychosomatic Research, vol. 52, no.2, pp. 69–77, 2002.
[31] M. M. Hoehn and M. D. Yahr, “Parkinsonism: onset, progres-sion, and mortality,” Neurology, vol. 17, no. 5, pp. 427–442, 1967.
[32] C. G. Goetz, S. Fahn, P. Martinez-Martin et al., “Movementdisorder society-sponsored revision of the unified Parkinson’sdisease rating scale (MDS-UPDRS): process, format, and clini-metric testing plan,”Movement Disorders, vol. 22, no. 1, pp. 41–47, 2007.
[33] J. Y. Y. Szeto, L. Mowszowski, M. Gilat, C. C. Walton, S. L.Naismith, and S. J. G. Lewis, “Assessing the utility of the Move-ment Disorder Society Task Force Level 1 diagnostic criteria formild cognitive impairment in Parkinson’s disease,” Parkinson-ism and Related Disorders, vol. 21, no. 1, pp. 31–35, 2015.
[34] R. M. Reitan,Manual for Administration for NeuropsychologicalTest Batteries for Adults and Children, Reitan Neuropsychologi-cal Laboratory, Tucson, Ariz, USA, 1979.
[35] D. Wechsler,WAIS-III WMS-III Technical Manual, Psychologi-cal Corporation, San Antonio, Tex, USA, 1997.
[36] O. Spreen and E. A. Strauss, A Compendium of Neuropsycho-logical Tests: Administration, Norms, and Commentary, OxfordUniversity Press, New York, NY, USA, 1998.
[37] H. Braak, E. Ghebremedhin, U. Rub, H. Bratzke, and K. DelTredici, “Stages in the development of Parkinson’s disease-related pathology,” Cell and Tissue Research, vol. 318, no. 1, pp.121–134, 2004.
[38] R. D. S. Prediger, F. C. Matheus, M. L. Schwarzbold, M. M. S.Lima, and M. A. B. F. Vital, “Anxiety in Parkinson’s disease:a critical review of experimental and clinical studies,” Neuro-pharmacology, vol. 62, no. 1, pp. 115–124, 2012.
[39] G. M. Sullivan, J. D. Coplan, J. M. Kent, and J. M. Gorman, “Thenoradrenergic system in pathological anxiety: a focus on panicwith relevance to generalized anxiety and phobias,” BiologicalPsychiatry, vol. 46, no. 9, pp. 1205–1218, 1999.

Neurology Research International 7
[40] E. F. Espejo, “Selective dopamine depletion within the medialprefrontal cortex induces anxiogenic-like effects in rats placedon the elevated plus maze,” Brain Research, vol. 762, no. 1-2, pp.281–284, 1997.
[41] A. Dagher, A. M. Owen, H. Boecker, and D. J. Brooks, “The roleof the striatum and hippocampus in planning: a PET activationstudy in Parkinson’s disease,” Brain, vol. 124, no. 5, pp. 1020–1032, 2001.
[42] K. L. Poston, S. YorkWilliams, K. Zhang et al., “Compensatoryneural mechanisms in cognitively unimpaired Parkinson dis-ease,” Annals of Neurology, vol. 79, no. 3, pp. 448–463, 2016.
[43] D. Weintraub, A. B. Newberg, M. S. Cary et al., “Striatal do-pamine transporter imaging correlates with anxiety and depres-sion symptoms in Parkinson’s disease,” Journal of Nuclear Medi-cine, vol. 46, no. 2, pp. 227–232, 2005.
[44] Z. Ye, E. Altena, C. Nombela et al., “Selective serotonin reuptakeinhibition modulates response inhibition in Parkinson’s dis-ease,” Brain, vol. 137, no. 4, pp. 1145–1155, 2014.
[45] D. Weintraub, S. Mavandadi, E. Mamikonyan et al., “Atomox-etine for depression and other neuropsychiatric symptoms inParkinson disease,” Neurology, vol. 75, no. 5, pp. 448–455, 2010.
[46] L. Marsh, K. Biglan, M. Gerstenhaber, and J. R. Williams, “Ato-moxetine for the treatment of executive dysfunction in Parkin-son’s disease: a pilot open-label study,”Movement Disorders, vol.24, no. 2, pp. 277–282, 2009.
[47] C. Pellicano, F. Assogna, L. Cravello et al., “Neuropsychiatricand cognitive symptoms andbody side of onset of parkinsonismin unmedicated Parkinson’s disease patients,” Parkinsonism andRelated Disorders, vol. 21, no. 9, pp. 1096–1100, 2015.
[48] G. Gainotti, “Emotional behavior and hemispheric side of thelesion,” Cortex, vol. 8, no. 1, pp. 41–55, 1972.
[49] D. Galin, “Implications for psychiatry of left and right cerebralspecialization. A neurophysiological context for unconsciousprocesses,”Archives of General Psychiatry, vol. 31, no. 4, pp. 572–583, 1974.
[50] E. Torbey, N. A. Pachana, and N. N. W. Dissanayaka, “Anxietyrating scales in Parkinson’s disease: a critical review updatingrecent literature,” Journal of AffectiveDisorders, vol. 184, pp. 216–224, 2015.
[51] K. A. Ehgoetz Martens, C. G. Ellard, and Q. J. Almeida, “Doesanxiety cause freezing of gait in Parkinson’s disease?” PLoSONE, vol. 9, no. 9, Article ID e106561, 2014.
[52] J. M. Shine, S. L. Naismith, N. C. Palavra et al., “Attentional set-shifting deficits correlate with the severity of freezing of gait inParkinson’s disease,”ParkinsonismandRelatedDisorders, vol. 19,no. 3, pp. 388–390, 2013.
[53] K. A. EhgoetzMartens, J. M. Hall, M. Gilat, M. J. Georgiades, C.C.Walton, and S. J. G. Lewis, “Anxiety is associatedwith freezingof gait and attentional set-shifting in Parkinson’s disease: a newperspective for early intervention,” Gait & Posture, vol. 49, pp.431–436, 2016.

APPENDIX(7(
Gilat, M., Hall, J. M., Ehgoetz Martens, K. A., Shine, J. M., Walton, C. C.,
MacDougall,H. G., Moore, S. T., Lewis, S. J. G. Staircase climbing is not solely a
visual compensation strategy to alleviate freezing of gait in Parkinson's disease.
Journal of Neurology, 2017
!

LETTER TO THE EDITORS
Staircase climbing is not solely a visual compensation strategyto alleviate freezing of gait in Parkinson’s disease
M. Gilat1 • J. M. Hall1,2 • K. A. Ehgoetz Martens1 • J. M. Shine3 •
C. C. Walton1 • H. G. MacDougall4 • S. T. Moore5,6 • Simon J. G. Lewis1
Received: 14 August 2016 / Revised: 16 October 2016 / Accepted: 26 October 2016 / Published online: 29 November 2016! Springer-Verlag Berlin Heidelberg 2016
Dear Editors,
In a recent case study by Janssen et al. [1], the authors
reported that Freezing of Gait (FOG) was alleviated in an
individual with Parkinson’s disease (PD) when climbingstairs, an effect that was preserved when walking on a
painted 3D staircase illusion. This is an important obser-
vation, as the clinical management of FOG remains chal-lenging [2] due in part to a lack of consensus regarding the
underlying mechanisms driving effective compensation
[1, 3, 4]. The authors provided three hypotheses as to why3D visual cues helped to alleviate FOG: enhanced visual
feedback from 3D cues were thought to either: (1) force
individuals to lift their feet higher; (2) perform larger lat-eral weight shifts; and/or (3) compensate for visuomotor
deficits [1]. Here, we refine these hypotheses by providing
evidence that the compensatory mechanisms involvedduring staircase climbing in PD patients with FOG are not
solely visual [1, 3, 4].
In this study, we asked seven male PD patients withclinically confirmed FOG and ‘off’ their regular
dopaminergic medication to perform two Timed up and Go
(TUG) tasks and to climb up and down a staircase twice,first with their eyes open and then with their eyes closed
(Fig. 1). All trials were videotaped for post hoc analysis of
freezing episodes, which were defined as brief, episodicabsence or marked reduction of forward progression of the
feet despite the intention to walk [5]. We hypothesized that
if visual information is essential for achieving compensa-tion, then patients would experience more FOG with the
eyes closed and any improvements observed during stair-
case climbing should disappear in the absence of visualinput. As expected [6], the patients experienced more FOG
when performing the TUG tasks with their eyes closed as
compared to eyes open (Z = -2.36, p\ 0.05) during over-ground TUG trials, confirming the importance of visual
feedback during over-ground walking in PD patients with
FOG [6]. Interestingly however, a strong beneficial effectwas observed during staircase climbing, with no patient
experiencing FOG during either the eyes open or eyesclosed conditions. Our data thus indicates that the com-
pensatory effect of staircase climbing can persist in the
absence of visual input, suggesting that enhanced visualfeedback is not the sole mechanism involved in the
compensation.
Indeed, there are other fundamental differences betweenstaircase climbing and over-ground walking that could
explain this finding. For instance, staircase climbing could
be a more discrete motor task that is carried out in an open-loop fashion (e.g. lifting feet to a predictable target of set
height) [7, 8]. Discrete movements only require the initi-
ation and execution of a motor plan (open loop motorcontrol) and are likely controlled through higher cortical
planning areas and preserved cortico-cerebellar circuitry
& Simon J. G. [email protected]
1 Parkinson’s Disease Research Clinic, Brain and Mind Centre,The University of Sydney, 100 Mallett Street, Camperdown,Sydney, NSW 2050, Australia
2 School of Social Sciences and Psychology, Western SydneyUniversity, Sydney, Australia
3 Department of Psychology, Stanford University, Stanford,CA, USA
4 School of Psychology, The University of Sydney, Sydney,Australia
5 Icahn School of Medicine, Mount Sinai, New York, NY,USA
6 School of Engineering and Technology, Central QueenslandUniversity, Rockhampton, QLD, Australia
123
J Neurol (2017) 264:174–176
DOI 10.1007/s00415-016-8329-3

[7, 9], without the need for online sensorimotor feedback in
the impaired basal ganglia [6, 10]. In contrast, rhythmic
and continuous over-ground walking requires attention inParkinson’s disease and is, therefore, under closed-loop
motor control and heavily dependent on vision, which PD
patients with FOG use to compensate for sensorimotorimpairments [6, 11, 12]. Removing vision, therefore,
increases the load on the impaired basal ganglia system
during overground walking, resulting in an increased riskfor FOG [13], which is bypassed during staircase climbing
since this can be carried out in a feed-forward manner. Our
results support this hypothesis, although future studies areneeded to confirm that staircase climbing is a less rhythmic
task for PD patients with FOG as compared to overground
walking. Furthermore, similar to cycling, the tactile cuesprovided by each stair may also regulate the timing and
amplitude of each movement, further mitigating sensori-motor processing in the basal ganglia [14]. Finally, the
patients in the current study first performed the tasks with
the eyes open and were thus able to obtain visuospatialmemory of the staircase, which could have allowed them to
plan their movements in advance in a feed-forward manner
during the eyes-closed condition. Visuospatial memory of
an upcoming task could, therefore, contribute to the com-
pensatory effect of staircase climbing and 3D visual cues
without the need for online visual feedback [1].We conclude that although vision is undoubtedly
important for PD patients with FOG [1, 6], online visual
feedback merely reflects one aspect of what makesstaircase climbing (and its painted illusion) an effective
compensatory strategy for FOG in PD [1, 3, 4]. Future
studies investigating the precise mechanisms underlyingcompensatory strategies for FOG in PD (including feed-
forward motor control and visuospatial memory) in a
standardized manner will, therefore, aid the developmentof more effective treatments for this devastating symp-
tom [2].
Acknowledgements M Gilat is supported by a University of SydneyInternational Scholarship. JM Hall is supported by a Western SydneyUniversity Postgraduate Research Award. KA Ehgoetz Martens issupported by Parkinson Canada. JM Shine is supported by a NationalHealth and Medical Research Council CJ Martin Fellowship. CCWalton is supported by an Australian Postgraduate Award at theUniversity of Sydney. HG MacDougall is supported by a grant fromthe Garnett Passe and Rodney Williams Memorial Foundation. STMoore is supported by NASA grant NNX12AM25G. SJG Lewis issupported by an NHMRC-ARC Dementia Fellowship.
Fig. 1 Experimental set-up showing the overground walking trial (Timed up and Go) in the top panels and staircase climbing in the bottompanels with eye tracker confirmation for eyes-open (left panels) and eyes-closed (right panels) conditions
J Neurol (2017) 264:174–176 175
123

Compliance with ethical standards
Conflicts of interest All authors have nothing to disclose and declareto have no conflicts of interest. The funding sources of the study hadno influence on the preparation, review or approval of thismanuscript.
Ethical standard Ethical approval for this study was obtained fromthe University of Sydney Human Research Ethics Committee andwritten informed consent was obtained from each patient prior to theirinclusion in the study as per the ethical standards laid down in the1964 Declaration of Helsinki and its later amendments.
References
1. Janssen S, Soneji M, Nonnekes J, Bloem BR (2016) A paintedstaircase illusion to alleviate freezing of gait in Parkinson’s dis-ease. J Neurol 263:1661–1662. doi:10.1007/s00415-016-8195-z
2. Nonnekes J, Snijders AH, Nutt JG et al (2015) Freezing of gait: apractical approach to management. Lancet Neurol 14:768–778.doi:10.1016/S1474-4422(15)00041-1
3. Snijders AH, Jeene P, Nijkrake MJ et al (2012) Cueing forfreezing of gait: a need for 3-dimensional cues? Neurologist18:404–405. doi:10.1097/NRL.0b013e31826a99d1
4. Rahman S, Griffin HJ, Quinn NP, Jahanshahi M (2008) Thefactors that induce or overcome freezing of gait in Parkinson’sdisease. Behav Neurol 19:127–136
5. Nutt JG, Bloem BR, Giladi N et al (2011) Freezing of gait:moving forward on a mysterious clinical phenomenon. LancetNeurol 10:734–744. doi:10.1016/S1474-4422(11)70143-0
6. Ehgoetz Martens KA, Pieruccini-Faria F, Almeida QJ (2013)Could sensory mechanisms be a core factor that underliesfreezing of gait in Parkinson’s disease? PLoS One 8:e62602.doi:10.1371/journal.pone.0062602
7. Howard IS, Ingram JN, Wolpert DM (2011) Separate represen-tations of dynamics in rhythmic and discrete movements: evi-dence from motor learning. J Neurophysiol 105:1722–1731.doi:10.1152/jn.00780.2010
8. Pisotta I, Molinari M (2014) Cerebellar contribution to feedfor-ward control of locomotion. Front Hum Neurosci 8:475. doi:10.3389/fnhum.2014.00475
9. Schaal S, Sternad D, Osu R, Kawato M (2004) Rhythmic armmovement is not discrete. Nat Neurosci 7:1136–1143. doi:10.1038/nn1322
10. Wolpert DM, Landy MS (2012) Motor control is decision-mak-ing. Curr Opin Neurobiol 22:996–1003. doi:10.1016/j.conb.2012.05.003
11. Almeida QJ, Lebold CA (2010) Freezing of gait in Parkinson’sdisease: a perceptual cause for a motor impairment? J NeurolNeurosurg Psychiatr 81:513–518. doi:10.1136/jnnp.2008.160580
12. Azulay J-P, Mesure S, Blin O (2006) Influence of visual cues ongait in Parkinson’s disease: contribution to attention or sensorydependence? J Neurol Sci 248:192–195. doi:10.1016/j.jns.2006.05.008
13. Lewis SJG, Shine JM (2014) The next step: a common neuralmechanism for freezing of gait. Neuroscientist. doi:10.1177/1073858414559101
14. Snijders AH, Toni I, Ruzicka E, Bloem BR (2011) Bicyclingbreaks the ice for freezers of gait. Mov Disord 26:367–371.doi:10.1002/mds.23530
176 J Neurol (2017) 264:174–176
123

APPENDIX(8(
Gilat, M., Bell, P. T., Ehgoetz Martens, K. A., Georgiades, M. J., Hall, J. M., Walton,
C. C., Lewis, S. J. G., Shine, J. M. Dopamine depletion impairs gait automaticity by
altering cortico-striatal and cerebellar processing in Parkinson's disease. NeuroImage,
2017
!

Contents lists available at ScienceDirect
NeuroImage
journal homepage: www.elsevier.com/locate/neuroimage
Dopamine depletion impairs gait automaticity by altering cortico-striataland cerebellar processing in Parkinson's disease
Moran Gilata,⁎, Peter T. Bellb, Kaylena A. Ehgoetz Martensa, Matthew J. Georgiadesa, JulieM. Halla, Courtney C. Waltona, Simon J.G. Lewisa,1, James M. Shinec,d,1
a Parkinson's Disease Research Clinic, Brain and Mind Centre, The University of Sydney, Sydney, NSW, Australiab University of Queensland Centre for Clinical Research, University of Queensland, Brisbane, QLD, Australiac Department of Psychology, Stanford University, Stanford, CA, United States of Americad Neuroscience Research Australia, Neuroscience Research Australia, University of New South Wales, Sydney, NSW, Australia
A R T I C L E I N F O
Keywords:Parkinson's diseaseAutomaticityDopamineBasal GangliafMRI
A B S T R A C T
Impairments in motor automaticity cause patients with Parkinson's disease to rely on attentional resourcesduring gait, resulting in greater motor variability and a higher risk of falls. Although dopaminergic circuitry isknown to play an important role in motor automaticity, little evidence exists on the neural mechanismsunderlying the breakdown of locomotor automaticity in Parkinson's disease. This impedes clinical managementand is in great part due to mobility restrictions that accompany the neuroimaging of gait. This study thereforeutilized a virtual reality gait paradigm in conjunction with functional MRI to investigate the role ofdopaminergic medication on lower limb motor automaticity in 23 patients with Parkinson's disease that weremeasured both on and off dopaminergic medication. Participants either operated foot pedals to navigate acorridor (‘walk’ condition) or watched the screen while a researcher operated the paradigm from outside thescanner (‘watch’ condition), a setting that controlled for the non-motor aspects of the task. Step time variabilityduring walk was used as a surrogate measure for motor automaticity (where higher variability equates toreduced automaticity), and patients demonstrated a predicted increase in step time variability during thedopaminergic “off” state. During the “off” state, subjects showed an increased blood oxygen level-dependentresponse in the bilateral orbitofrontal cortices (walk >watch). To estimate step time variability, a parametricmodulator was designed that allowed for the examination of brain regions associated with periods of decreasedautomaticity. This analysis showed that patients on dopaminergic medication recruited the cerebellum duringperiods of increasing variability, whereas patients off medication instead relied upon cortical regions implicatedin cognitive control. Finally, a task-based functional connectivity analysis was conducted to examine the mannerin which dopamine modulates large-scale network interactions during gait. A main effect of medication wasfound for functional connectivity within an attentional motor network and a significant condition by medicationinteraction for functional connectivity was found within the striatum. Furthermore, functional connectivitywithin the striatum correlated strongly with increasing step time variability during walk in the off state(r=0.616, p=0.002), but not in the on state (r=−0.233, p=0.284). Post-hoc analyses revealed that functionalconnectivity in the dopamine depleted state within an orbitofrontal-striatal limbic circuit was correlated withworse step time variability (r=0.653, p < 0.001). Overall, this study demonstrates that dopamine amelioratesgait automaticity in Parkinson's disease by altering striatal, limbic and cerebellar processing, thereby informingfuture therapeutic avenues for gait and falls prevention.
http://dx.doi.org/10.1016/j.neuroimage.2017.02.073Received 27 November 2016; Accepted 24 February 2017
⁎ Correspondence to: 100 Mallett Street, Camperdown 2050, NSW, Australia.
1 These authors contributed equally to this work.E-mail address: [email protected] (M. Gilat).
Abbreviations: BOLD, Blood Oxygenation Level Dependent; CV, Coefficient of Variation; DC, Dorsal Caudate; DCP, Dorsal Caudal Putamen; DLPFC, Dorsolateral Prefrontal Cortex;DRP, Dorsal Rostral Putamen; FDR, False Discovery Rate; FSL, Footstep Latency; MNI, Montreal Neurological Institute and Hospital; PCP, Postcommissural putamen; PMd, DorsalPremotor Cortex; PPC, Posterior Parietal Cortex; Pre-SMA, Pre-Supplementary Motor Area; ROIs, Regions of Interest; SD, Standard Deviation; SPM12, Statistical Parametric Mappingversion 12; TFCE, Threshold-Free Cluster Enhancement; UPDRS-III, Motor section of the Unified Parkinson's Disease Rating Scale; VRP, Ventral Rostral Putamen; VSi, Inferior VentralStriatum; VSs, Superior Ventral Striatum
NeuroImage 152 (2017) 207–220
Available online 03 March 20171053-8119/ © 2017 Elsevier Inc. All rights reserved.
MARK

Introduction
Impairments in motor automaticity are a hallmark feature ofParkinson's disease that cause patients to increasingly demand corticalresources in order to execute basic motor operations via attentionalprocesses (Bohnen and Jahn, 2013; Wu et al., 2015). A reduction inmotor automaticity is of particular concern during gait in Parkinson'sdisease, as the cortical resources that would be used for compensationare not optimized for the fast and parallel processing required duringlocomotion (Clark, 2015). Furthermore, the excessive attentionaldemand of walking in Parkinson's disease demands a high computa-tional cost and interferes with gait control in conditions of highworkload (Schneider and Chein, 2003; Lewis and Barker, 2009;Lewis and Shine, 2014; Clark, 2015; Wu et al., 2015). As a result,patients with Parkinson's disease have a greater risk of adversemobility outcomes and falls, especially during more complex everydaysituations where a secondary task is performed in parallel with gait(Schaafsma et al., 2003; Clark, 2015; Hausdorff et al., 2003a;Vandenbossche et al., 2012; Lewis and Shine, 2014; Strouwen et al.,2015; Wu et al., 2015). Despite the clinical importance of gait inParkinson's disease, the precise neural mechanisms underlying im-pairments in locomotor automaticity remain poorly understood, thusimpeding targeted management (Wu et al., 2015).
Increased step time variability is a robust predictor for falls inParkinson's disease (Hausdorff et al., 1998) and has previously beensuggested as a surrogate measure for reduced locomotor automaticity(Frenkel-Toledo et al., 2005; Yogev et al., 2005; Peterson and Horak,2016). A recently proposed framework by Wu and colleagues (2015)outlines several features that indicate whether a motor deficit is directlylinked to an underlying impairment in motor automaticity inParkinson's disease, namely: i) the motor skill is performed automa-tically (and without behavioural interference) in healthy subjects; ii)dual task performance results in significant deterioration in the motorskill in patients with Parkinson's disease as compared to healthysubjects; and iii) external cueing (or attention) significantly improvesthe performance of this motor skill (Wu et al., 2015). If this theoreticalmodel is applied to the existing literature, it indeed becomes evidentthat step time variability fits these criteria as an index of locomotorautomaticity: i) stride time variability is resistant to interference duringdual-task walking in healthy subjects (Yogev et al., 2005), suggestingthat regulation of stride time variability is an automated motor skill inhealthy subjects (Friedman et al., 1982; Yogev et al., 2005; Wu et al.,2015); ii) in Parkinson's disease, walking while performing a cognitivedual task exacerbates stride time variability (Hausdorff et al., 2003b;Yogev et al., 2005; Plotnik et al., 2011) and patients experience higherdual task cost during walking as compared to healthy subjects (Yogev
et al., 2005); iii) stride time variability in Parkinson's disease has beenshown to reduce in the presence of external auditory cues (Willemset al., 2006; Hausdorff et al., 2007; Rochester et al., 2011). Therefore,this evidence suggests that step time variability, the regulation of whichis an automated process in the healthy population, is impaired inParkinson's disease and reflective of reduced motor automaticity(Yogev et al., 2005; Bohnen and Jahn, 2013; Lewis and Shine, 2014;Wu et al., 2015; Peterson and Horak, 2016).
Motor automaticity is typically achieved through motor learning,followed by the timely initiation and maintenance of automated motorsequences, even during interference (Wu et al., 2015). During motorlearning, the posterior striatum (putamen) is thought to ‘chunk’ motoraction sequences under the influence of dopamine (Graybiel, 1998).This process allows performance of well-learned motor patterns to beexecuted as a single unit of activity rather than multiple serialcomputations, enhancing neural efficiency and reducing motor varia-bility (Schneider and Chein, 2003; Poldrack et al., 2005; Wymbs et al.,2012; Wu et al., 2015). The initiation of an automatic sequence isthought to involve a shift from the anterior associative fronto-striatalcircuit to the posterior sensorimotor striatum (Miyachi et al., 1997;Lehéricy et al., 2005; Wymbs et al., 2012; Wu et al., 2015), which freesup frontal attentional resources that can then be used to processconcurrent secondary demands (Carbon and Marié, 2003; Monchiet al., 2007; Lewis and Shine, 2014; Wu et al., 2015). The most welldescribed circuits involved with maintaining an automated motorsequence include the spinal cord, brainstem locomotor regions, poster-ior striatum, primary cortical motor regions and cerebellum (Fig. 1)(Poldrack et al., 2005; Bohnen and Jahn, 2013; Clark, 2015; Wu et al.,2015). Doyon et al. (2002) further proposed that during motorlearning, a transfer of experience-dependent changes from the cere-bellar cortex to the deep cerebellar dentate nucleus takes place, andthen with extended practice, from a cerebellar-cortical to a striatal-cortical network (Doyon et al., 2002). Evidently, a well functioningstriatum is integral for effective locomotor automaticity (Wu et al.,2015).
Ascending dopaminergic neurons in the midbrain provide richinnervation to the entire striatum, thereby exerting neuromodulatorycontrol over information processing across the striatum and parallelcortico-striatal loops that underpin the execution of coordinatedbehaviours (Alexander et al., 1986; Kelly et al., 2009; Helmich et al.,2010; Surmeier et al., 2010; Hacker et al., 2012; Sharman et al., 2013;Bell et al., 2014). In Parkinson's disease, the pathological degenerationof nigrostriatal dopaminergic neurons impacts on the communicationacross the striatum and parallel cortico-striatal circuits (Alexanderet al., 1986; Kelly et al., 2009; Helmich et al., 2010; Surmeier et al.,2010; Hacker et al., 2012; Sharman et al., 2013; Bell et al., 2014),
Fig. 1. Schematic representation of the locomotor automaticity processes in health and hypothesized neural mechanisms underlying automaticity impairments in Parkinson's diseasewith and without dopaminergic replacement therapy. Left image – hypothesized posterior motor network underlying locomotor automaticity in health; Middle image – dopaminergicpathology in posterior striatum in Parkinson's disease is thought to cause patients to utilize attention demanding cortical resources to operate gait; Right image – dopaminergicreplacement therapy is thought to normalize locomotor automaticity impairments in Parkinson's disease. NOTE: “Off”=Dopamine depleted state; “On”=On dopaminergic replacementtherapy.
M. Gilat et al. NeuroImage 152 (2017) 207–220
208

which likely impairs motor learning and automaticity (Fig. 1)(Vandenbossche et al., 2012; Bohnen and Jahn, 2013; Lewis andShine, 2014; Hamacher et al., 2015; Wu et al., 2015; Peterson andHorak, 2016). Indeed, the dopaminergic insult is most severe in theposterior striatum (i.e. sensorimotor putamen) that is involved withmotor learning and automaticity (Brooks et al., 1990; Poldrack et al.,2005; Wu et al., 2015). Furthermore, step time variability improveswith dopaminergic replacement therapies in Parkinson's disease (Fig. 1)(Hausdorff et al., 2003a; Schaafsma et al., 2003; Bryant et al., 2016). Thedopamine depletion in the posterior putamen is also likely to impact onthe anterior-to-posterior striatal shift (Ashby et al., 2010; Everitt andRobbins, 2016), thus placing additional load on the frontal attentionalresources that further prevents patients from effectively utilizing com-pensatory locomotor control strategies when automaticity is reduced(Lewis and Barker, 2009; Helmich et al., 2010; Vandenbossche et al.,2012; Lewis and Shine, 2014; Wu et al., 2015). Finally, it has often beensuggested that Parkinson's patients utilize the cortico-cerebellar path-ways involved during early motor learning in order to maintain motorfunctions following the cortico-striatal impairments (Hanakawa et al.,1999; Wu and Hallett, 2013; Peterson and Horak, 2016). However, todate little evidence exists describing the neural mechanisms underlyingdeficits in locomotor automaticity and the role of compensatorystrategies in Parkinson's disease due in great part to the mobilityrestrictions that accompany the neuroimaging of gait per se (Shineet al., 2013a; Wu et al., 2015). As such, we currently lack a preciseunderstanding of how dopaminergic pathology impairs motor automa-ticity during gait (Vandenbossche et al., 2012; Bohnen and Jahn, 2013;Lewis and Shine, 2014; Hamacher et al., 2015; Wu et al., 2015; Petersonand Horak, 2016). Furthermore, the role of dopaminergic medication inregulating locomotor automaticity in Parkinson's disease remains poorlyunderstood. Finally, understanding compensatory mechanisms for over-coming locomotor automaticity impairments in Parkinson's diseaseremains an important unresolved clinical question (Nonnekes et al.,2015; Wu et al., 2015; Peterson and Horak, 2016).
In this study we set out to investigate the neural mechanisms ofdopamine on repetitive lower limb movements in Parkinson's disease.Twenty-three patients with Parkinson's disease performed a virtualreality gait paradigm in conjunction with functional MRI both on andoff their dopaminergic medication. The virtual reality task requiredpatients to either operate foot pedals to navigate a virtual corridor(walk condition) or to watch the screen while a researcher operated theparadigm from outside the scanner (watch condition). We hypothe-sized that in the off medication state, patients with Parkinson's diseasewould show increased variability in their step times as compared to themedicated state, indicative of reduced motor automaticity (Hausdorffet al., 2003b; Yogev et al., 2005; Gilat et al., 2013). We furtherhypothesized that this increase in variability would be associated withincreased Blood Oxygenation Level Dependent (BOLD) in frontal andparietal cortical regions that are associated with the attentional controlof movements (Ouchi et al., 2001; Bohnen and Jahn, 2013; Wu et al.,2011; 2015; Peterson and Horak, 2016). In addition, a recent restingstate functional MRI study showed that the strength of connectivityacross the striatum was significantly reduced in Parkinson patients offtheir dopaminergic medication, which even at rest impacted on large-scale sensorimotor network dynamics (Bell et al., 2014). As such, wehypothesized that dopamine denervation would also have the strongesteffect on the connectivity across the striatum during the virtual gaittask (Bell et al., 2014), as shown by a significant condition (watch,walk) by medication (off, on) interaction effect. Furthermore, wehypothesized that dopamine depleted connectivity changes across thestriatum would correlate with increasing step time variability(Hausdorff et al., 2003a; Schaafsma et al., 2003 Bryant et al., 2016)and impact on the connectivity across large-scale motor automaticityand attentional motor control networks involved with gait inParkinson's disease (Kelly et al., 2009; Helmich et al., 2010; Hackeret al., 2012; Bohnen and Jahn, 2013; Bell et al., 2014; Wu et al., 2015).
Materials and methods
Study protocol
Twenty-three patients with Parkinson's disease that satisfied theUnited Kingdom Parkinson's Disease Society Brain Bank clinicaldiagnostic criteria were recruited for this study from the Parkinson'sDisease Research Clinic, Brain and Mind Centre, The University ofSydney. None of the patients were diagnosed with dementia accordingto the Movement Disorders Society guidelines (Goetz et al., 2008) ormajor depression according to Diagnostic and Statistical Manual ofMental Disorders-IV guidelines of the American PsychiatricAssociation (Diagnostic and Statistical Manual of Mental Disorders(4th ed., text rev.)). None of the participants had any additionalneurological comorbidities including no history of stroke or significanthead injury. In addition, high-resolution T1-weighted images of eachsubject passed visual inspection by an experienced radiologist forabsence of any pathological white or grey matter lesions. All patientsreceived dopamine-replacement therapy as part of their daily clinicalmanagement. Specifically, twelve patients were on levodopa mono-therapy; two patients were on dopamine agonist monotherapy; fourpatients were on levodopa plus a dopamine agonist; two patients wereon levodopa plus a monoamine oxidase inhibitor; three patients wereon levodopa plus a dopamine agonist plus a monoamine oxidaseinhibitor. Patients were tested on two separate occasions, once whilston their usual medications and once in the practically defined off state,having been withdrawn from their dopaminergic medication overnightfor more than 12 h before testing. The order of testing (ON/OFF) wasrandomized and counterbalanced between subjects with a minimuminterval of three weeks between trials (mean: 7.4±5 weeks).
In addition to PD participants, we also acquired imaging data in anindependent group of twelve healthy age matched control participants(see Materials and methods: Regions of interest) that were scanned ona single occasion. The acquisition of healthy control data enabled data-driven functional localization of regions involved in lower limb motorcontrol during the virtual gait paradigm in an independent andmatched cohort. Ethical approval for this study was obtained fromthe University of Sydney Human Research Ethics Committee andwritten informed consent was obtained from each subject in accor-dance with the Declaration of Helsinki.
Cognitive and neurological assessment
All subjects were assessed on the Mini Mental State Examination.In addition, the motor section of the Movement Disorders SocietyUnified Parkinson's Disease Rating Scale (UPDRS-III) and the Hoehnand Yahr Stages (HY), both on and off medications were obtained frompatients with Parkinson disease. Motor symptom severity was assessedper body side by calculating a sum score of the UPDRS-III items for theleft and right body side separately (items 3.3–3.8 and 3.15–3.17). Thebody side with the highest sum score on these UPDRS-III items wasdefined as being most affected. Furthermore, item 3 of the Freezing ofGait Questionnaire (FOG-Q3) “Do you feel that your feet get glued tothe floor while walking, making a turn or when trying to initiatewalking (freezing)?” was obtained from each subject. Finally, dopa-mine dose equivalency scores were recorded. Cognitive and neurologi-cal measures were compared between medication states using a paired-samples t-test or non-parametric Wilcoxon Signed Rank Sum test(alpha=0.05).
Virtual reality task
The virtual reality task was performed while subjects lay supineinside the MRI scanner. The duration of the task was 10 min. The taskstimuli were presented on a screen that could be clearly viewed via amirror mounted onto the head coil. The virtual environment consisted
M. Gilat et al. NeuroImage 152 (2017) 207–220
209

of a straight corridor presented in the first person that contained noadditional environmentally salient or cognitive triggers, which havepreviously been utilised to trigger freezing behaviour (motor arrest)(Fig. 2A) (Shine et al., 2013a; Gilat et al., 2013; 2015). Indeed verylittle behavioural freezing (less than 0.05% of the total paradigm) wasobserved in the current experiment. Progression through this corridorwas accomplished by alternately depressing left and right foot pedals atleast 30° below parallel in a physiological sequence (e.g. left-right-left-…). Out of sequence steps (left-left or right-right) did not result inforward progression and were disregarded from behavioural analyses.Subjects were instructed to tap the pedals in a comfortable rhythm asper previous work (Shine et al., 2013a; Gilat et al., 2013; 2015). Theparadigm was made up of alternating blocks that instructed theparticipants to perform either of two rules. Walk blocks were initiated
with the word “WALK” being presented on screen instructing theparticipants to start operating the pedals. In addition, participantsperformed a control watch block. Watch blocks were initiated with theword “WATCH” being presented on screen instructing participants torefrain from operating the pedals and watch the screen while aresearcher operated the task from outside the MRI scanner in a similarrhythm as the participants. Subjects were therefore presented with thesame visual input and baseline cognitive and limbic processes as duringwalk, without the need for any motor activation of the legs. Each blocklasted for approximately one minute depending on the subject's latencyand was ended when the word “STOP” was presented on screen in thecolour red. Six seconds after participants had stopped accordingly, thealternative of the previous instruction (either “WALK” or “WATCH”)was presented. Each participant completed 4–5 alternating blocks ofboth conditions with the first condition always being a walk block toallow the researcher to ensure that the patient had understood theinstructions.
Behavioural measures
Footstep latency was calculated during walk by measuring the timebetween two consecutive foot pedal depressions. The first five stepsfollowing a “WALK” cue and any step following a “STOP” cue wereexcluded from the analyses to remove the effects of motor initiationand cessation (Georgiades et al., 2016). The mean and standarddeviation were then used to calculate the coefficient of variation, whichis a measure of step time variability (Hausdorff et al., 2003a; Gilatet al., 2013). A coefficient of variation in footstep latencies was alsocalculated for each foot separately. A repeated measures ANOVA wasthen used to investigate the interaction between medication (off, on)and symptom side (most affected, least affected) on the coefficient ofvariation for each leg separately. In addition, the coefficient of variationwas compared between the most- and least affected side using a paired-sampled t-test for both medication states. These analyses wereperformed to assess whether any increase in variability may beconfounded by asymmetry in symptom severity. No behaviouralmeasures were obtained during watch blocks, however all participantsresponded adequately to the “WALK” cue that followed a watch block (< 2 s initiation times), indicating that they were likely to be payingclose attention to the task.
Neuroimaging
Image acquisitionA General Electric 3T MRI was used to obtain T2*-weighted echo
planar functional images in sequential order with repetition time (TR)=3 s, echo time=32ms, flip angle=90°, 32 axial slices covering thewhole brain, field of view=220 mm, interslice gap=0.4 mm and rawvoxel size=3.9 mm×3.9 mm×4 mm thick. High-resolution 3D T1-weighted anatomical images with voxel size=0.4×0.4×0.9 mm wereobtained for co-registration with functional scans.
Image pre-processingImage processing and analyses were performed using Statistical
Parametric Mapping Software (SPM12, Wellcome Trust Centre forNeuroimaging, London, UK). Functional images were pre-processedusing the standard pre-processing pipeline provided with SPM12.Functional scans were: (i) manually realigned along the anterior-posterior commissure; (ii) slice-time corrected to the median (17th)slice in each scan; (iii) realigned to create a mean realigned image andmeasures of 6 degrees of rigid head movements were created for lateruse in the correction of minor head movements; (iv) unwarped to dealwith residual movement related variance induced by the susceptibility-by-movement interaction effects; (v) spatially normalized using the T1-weighted image to improve segmentation accuracy; (vi) co-registeredand estimated; and (vii) smoothed using an 8-mm full-width at half
Fig. 2. Whole Brain analysis for walk >watch. A=visual representation of the virtualreality task; B=BOLD responses within the Parkinson's disease group (n=23) betweenmedication states (off > on) on the condition of interest (walk>watch); p < 0.005, k > 20,TFCE corrected; C=Average beta values associated with the significant clusters as perfigure B. NOTE: OFC=Orbitofrontal cortex; PD=Parkinson's disease; OFF=Dopaminedepleted state; ON=On dopaminergic replacement therapy.
M. Gilat et al. NeuroImage 152 (2017) 207–220
210

maximum isotropic Gaussian kernel. Spatial normalization was thenmanually checked for quality assurance.
Head motion correctionMultiple precautions were taken to ensure head motion was fully
accounted for: (i) all subjects were instructed to minimize head motionby only moving the ankles, while not raising the legs and restrict hiprotation; (ii) cushions were placed inside the head coil to ensureoptimal performance with the least amount of head motion; (iii)following data collection, trials with > 3 mm or 3° of scan-to-scanmovement were considered a-priori exclusion criterion; (iv) six motionand nuisance regressors were added into the first level analysis persubject, controlling for minor movement artefacts in the three direc-tions of translation and axes of rotation; (v) each trial was analysedusing ArtRepair (Mazaika et al., 2007) and trials with a large amount ofglobal drift or scan-to-scan head movement > 1.5 mm were correctedusing the interpolation method; and finally, (vi) mean frame-wisedisplacement was calculated per trial and compared across sessions,showing no significant differences between the on and off states(Parkinson patients off versus on: Z=−0.304, p=0.761) or betweengroups (Parkinson patients off versus controls: U=124, Z=−0.487,p=0.644; Parkinson patients on versus controls: U=126, Z=−0.417,p=0.694).
Event related whole brain analysisIndividual first-level spatial maps were created in SPM12 using a
general linear model analysis within an epoch-related design in a fixed-effects analysis. A design matrix was created for each subject byentering two regressors for each trial: a regressor that modelled thespecific onset times and associated temporal derivatives of walk blocksand a regressor that similarly modelled the watch blocks. Contrastimages from the first-level analyses were then entered into a second-level random-effects dependent samples t-test design to test the effectsof dopaminergic medication within the Parkinson's disease group onthe condition of interest (walk >watch), whereas a one-sample t-testdesign was used for the healthy control group. To further investigateimaging-behaviour associations, 8mm ROIs were created around thepeak voxel for each significant second-level cluster found whencontrasting walk >watch between medication states in Parkinson'sdisease.
In addition, a whole brain parametric modulation analysis was usedto investigate which brain regions were involved in the modulation ofstep time variability. First, a normalized footstep variability value perTR was calculated for the walk blocks for each subject. The normalizedfootstep variability was calculated as an absolute Z-score of footsteplatencies, which were then de-meaned and averaged per TR. Thisparametric modulator vector was then entered into a general linearmodel together with the six head motion regressors and time deriva-tives of the hemodynamic response function in the first level analysis ofSPM12. Contrast images from this analysis were then entered into asecond-level random-effects analysis: dependent samples t-test toinvestigate differences between dopaminergic states in theParkinson's disease group; and one-sample t-test analysis for thehealthy control group. Regions with a negative relationship to theparametric modulator were associated with maintaining low step timevariability, whereas an increased BOLD response indicates that thoseregions were involved during period of increasing step time variability,and hence, worsened automaticity.
The whole brain voxel maps for both analyses were displayed usingxjView (www.alivelearn.net/xjview) software (p < 0.005, k > 20)(Lieberman and Cunningham, 2009) and a threshold-free clusterenhancement (TFCE) for multiple corrections was performed on eachsecond level contrast (Smith and Nichols, 2009). To explore thedirection of pattern in the BOLD responses found in this study,significant clusters (See Supplementary Table 3 for MNI coordinates)from the within-Parkinson group second level T-maps were saved as
images and raw Beta scores were then extracted from these ROIs usingthe MarsBar toolbox (Brett et al., 2002) in SPM12, the values of whichwere averaged for reporting purposes only.
Task-based functional connectivityIn this analysis, we examined the effect of dopaminergic medication
on large-scale network interactions within and between motor auto-maticity, attentional motor control and striatal networks. Task basedfunctional connectivity in these networks was calculated using pre-defined regions of interest (ROI) as further described below. TheResponse Exploration for Neuroimaging datasets toolbox (Duff et al.,2007) was used to extract time series data of each predefined ROI foreach patient. Task-based functional connectivity was then calculatedusing the Multiplication of Temporal Derivatives statistical method(Shine et al., 2015). This method allows greater temporal resolution oftime-resolved connectivity in BOLD time series data when compared tothe conventional sliding-window Pearson's correlation coefficient(Shine et al., 2015, 2016). The code is freely available at http://github.com/macshine/coupling/ (Shine et al., 2015, 2016). First, meanfunctional connectivity was calculated within and between eachnetwork of interest and entered into condition (watch, walk) bymedication (on, off) repeated measures ANOVAs (alpha=0.05, FalseDiscovery Rate (FDR) correction 0.05). For significant interactionspost-hoc correlations were performed to further examine whether thetask-based functional connectivity was associated with worse step timevariability. Furthermore, to examine whether striatal dopaminemodulated the communication across large-scale networks, wefurther examined the effects of dopamine on internal networkconnectivity. A correlation analysis was performed on the amount offunctional connectivity during walk in the on state compared to the offstate (on-off) for the connectivity across each network.
Regions of interestRegions of interest were defined using a combination of data-driven
regions identified using the age-matched healthy control group duringperformance of the virtual gait task and pre-defined a priori regionsshown to be involved in attentional motor control in Parkinson'sdisease and the striatum (see Supplementary Materials: Regions ofinterest for full description). Defined regions of interest were thenutilized for subsequent functional connectivity analyses (seeSupplementary Table 2 for MNI coordinates). These regions composedthree different networks: motor automaticity, attention motor andstriatal networks.
Motor automaticity networkAs it is currently unclear how lower limb motor automaticity is
achieved, data from the twelve matched healthy control subjects wasused to explore which brain regions would be involved with theperformance of the virtual reality task (walk >watch), and specificallyto identify which brain regions would be associated with maintaininglow step time variability. Based on this data-driven approach weincluded the bilateral primary motor cortex area of the legs, thalamus,putamen, superior orbitofrontal gyrus, lateral cerebellum and anteriorcingulate into the motor automaticity network. In addition, thecerebellar locomotor and bilateral mesencephalic locomotor regionswere included as predefined a-priori regions of interest (SeeSupplementary Materials: Regions of interest and SupplementaryTables 1 and 2 for MNI coordinates).
Attention motor networkThe low variability in step times seen in the healthy control
participants did not allow for the investigation of the attentiondemanding cortical regions that are hypothesized to be associated withhigh step time variability in the Parkinson's disease group followingdopamine depletion (Wu and Hallett, 2005; Yogev et al., 2005; Bohnenand Jahn, 2013; Lewis and Shine, 2014; Clark, 2015; Wu et al., 2015;
M. Gilat et al. NeuroImage 152 (2017) 207–220
211

Peterson and Horak, 2016). As such, predefined ROIs were created forregions that have been shown to be involved with the attentionalcontrol of movements in Parkinson's disease, including the dorsallateral prefrontal cortex (DLPFC), pre-supplementary motor area (pre-SMA), dorsal premotor cortex (PMd) and posterior parietal cortex(PPC) (Cole and Schneider, 2007; Kurz et al., 2012; Shine, et al.,2013b; Wu et al., 2011; 2015).
Striatal networkThe striatal network consisted of seven predefined regions of
interest in the striatum of each hemisphere in the Parkinson's diseasegroup, as described by Bell et al. (2014). These striatal regionscorrespond to dissociable functional systems (Di Martino et al., 2008;Kelly et al., 2009) and enable broad coverage of the striatum (Bell et al.,2014). The ROIs included the bilateral: inferior ventral striatum (VSi),superior ventral striatum (VSs), dorsal caudate (DC), dorsal caudalputamen (DCP), dorsal rostral putamen (DRP), ventral rostral puta-men (VRP) and postcommissural putamen (PCP) (Di Martino et al.,2008; Kelly et al., 2009; Bell et al., 2014). Based on the study by DiMartino and colleagues (2008) we further defined the caudal putamenseeds (DCP and PCP) as being most involved with sensorimotor tasks,the dorsal caudate (DC) with cognitive control and the inferior ventralstriatum (VSi) seeds, which approximate the nucleus accumbens, withlimbic processing (Di Martino et al., 2008). These seeds were used todefine striatal cognitive-motor, limbic-motor and limbic-cognitiveconnections during a post-hoc analysis, as further described below.
Results
Participant demographics and behavioural outcomes
Patients with Parkinson disease and healthy control subjects werematched across multiple demographics (see Supplementary Table 4).The Parkinson's disease patients had a mean levodopa equivalent dailydose of 861 (±525) and disease duration (in years) of 6 (±2.9). Patientsdemonstrated clinical improvement following dopaminergic treatment,as indicated by a significantly lower UPDRS-III score and HY stages inthe on state (Table 1). The current cohort consisted of patients with arange of freezing of gait severity, with eight subjects scoring a 0(‘Never’), six subjects scoring a 1 (‘Very rarely’) and nine subjectsscoring > 1 (‘Rarely-Often-Always’) on the FOG-Q3. Importantly,Parkinson's disease patients off medication had similar modal footsteplatencies as compared to when medicated, which together with similarscores on UPDRS-III items 3.7 (toe tapping) indicate that any groupdifferences found were unlikely to be driven by an overall difference inlower limb motor performance (e.g. rigidity) (Table 1). However aspredicted, step time variability was significantly higher in Parkinson
patients off their medication as compared to when on medication(Table 1), suggesting between group differences in motor automaticity.Indeed, no significant interaction effect was found between medication(off, on) and symptom side severity (most affected, least affected) onthe coefficient of variation of footstep latencies (F=0.992, p=0.330).This analysis also revealed no main effect of medication (F=2.698,p=0.115) or main effect of symptom side severity (F=0.598, p=0.448).Furthermore, a difference score in toe tapping ability between themost- and least affected side did not significantly correlate with thecoefficient of variation in footstep latencies in either medication state(Spearman correlation OFF: ρ=0.318, p=0.139; ON: ρ=−0.311,p=0.149). Together, these results indicate that the change in step timevariability seen between medication states did not reflect asymmetry insymptom severity.
Neuroimaging results
Whole brain activation (walk >watch)The dopamine-depleted state of Parkinson's disease was associated
with a significantly positive BOLD response in the bilateral orbito-frontal cortex during walk compared to negative BOLD during watch(TFCE corrected, see Fig. 2B and Supplementary Table 3 for peak voxelcoordinates). In addition, within-group analysis revealed that in bothmedication states patients demonstrated activation of the primarymotor cortex. However, when on medication patients also utilized thepre-supplementary motor area, visual cortex and cerebellum, whereaspatients offmedication utilized the right mid frontal gyrus and bilateralorbitofrontal cortex (see Supplementary Fig. 2). For exploration (i.e.non-statistical) purposes only, post-hoc one-sampled and paired-sampled t-tests were performed on the mean beta values in theorbitofrontal clusters that were found to be significantly differentbetween medication states on the condition of interest (walk > watch)(See Fig. 2C). These results confirmed that in the off state, the betavalues in the orbitofrontal clusters were significantly different fromzero during both walk and watch blocks (all p < 0.01, Bonferonnicorrected) and significantly different between walk and watch blocks(Left orbitofrontal: t=4.36, p < 0.001; Right orbitofrontal: t=5.42, p <0.001, Bonferonni corrected). No differences in beta values were foundfor both orbitofrontal clusters in the dopaminergic on state betweenwalk and watch blocks (Left orbitofrontal: t=0.560, p=0.592; Rightorbitofrontal: t=0.881, p=0.151), which were also not significantlydifferent from zero (all p > 0.1).
Whole brain activation (parametric modulator)The goal of this analysis was to identify regions that were
consistently associated with greater step time variability. The BOLDresponses associated with the parametric modulator in the Parkinson'sdisease group are presented in Fig. 3. In the off state Parkinson'sdisease patients had significant negatively signed beta values across theright dorsal premotor cortex and left posterior parietal cortex incontrast to the slight positively signed beta values in these regions inthe on state (Fig. 3C and E). This suggests that a negative associationexists between BOLD response in these regions and step timevariability in the off state. Furthermore, patients on medication hadsignificant positively signed beta values in the right lateral cerebellumcompared to the off state (Fig. 3C and E), indicating that the cerebellarregions were recruited during periods of increasing step time variabilitywhen medicated, although these findings did not survive TFCE correc-tion for multiple comparisons. As tremor amplitude in Parkinson'sdisease has previously been associated with increased beta values in thecerebellum (Helmich et al., 2012), a post-hoc Spearman correlationanalysis was performed between the sum score of UPDRS-III tremoritems 3.15–3.18 and the beta weights of the right cerebellum clusterfound to be significantly associated with the parametric modulator. Theresults showed no significant correlations between tremor scores andcerebellar Beta weights for both medication states (OFF: ρ=0.199,
Table 1Demographic and Behavioural Statistics within the Parkinson's disease group (n=23)during different medication states (off/on).
Task OFF ON Test-value P-value
UPDRS-III 27.8 (14) 22.9 (14) 2.62 0.015a
HY 2 (1–3) 2 (1–3) −2.12 0.034b
Toe-Tap Right 1 (0–3) 1 (0–4) −0.428 0.669b
Toe-Tap Left 1 (0–3) 1 (0–3) −0.775 0.439b
Modal FSL 0.48 (0.15) 0.48 (0.12) −0.011 0.991a
CV 29.6 (20) 16.4 (8.3) 3.50 0.002a
NOTE: UPDRS-III=Movement Disorder Society Unified Parkinson's Disease RatingScale motor section III, Toe-Tap=Scores on item 3.7 of the Movement Disorder SocietyUnified Parkinson's Disease Rating Scale, Modal FSL=Modal Footstep Latency duringperformance of walk in the virtual reality task, CV=Coefficient of Variation in footsteplatencies during the performance of walk in the virtual reality task.
a Paired-samples t-test used and Mean (SD) and t-value reported.b Non-parametric Wilcoxon Signed Ranks Test used and Median (Range) and Z-value
reported for ordinal variables.
M. Gilat et al. NeuroImage 152 (2017) 207–220
212

p=0.362; ON: ρ=0.142, p=0.517), indicating that tremor was unlikelyto explain the results found.
To explore whether Parkinson patients utilized a cortico-cerebellarnetwork in order to operate movements as a compensation for cortico-striatal impairments (Rascol et al., 1997; Doyon et al., 2002), a post-hoc correlation analysis was performed between the loading of theparametric modulator regressor onto the right lateral cerebellum thatwas found to be significant during the parametric modulator analysiscontrast (OFF >ON, see Fig. 3C) and the functional connectivity withinthe attention motor control network. A positive correlation was foundbetween the loading of the variability regressor onto the right lateralcerebellum and the total functional connectivity within the regions ofthe cognitive network for the Parkinson's disease group when offmedication (r=0.452, p=0.030, uncorrected), whereas the negative
correlation found when patients were medicated did not reach statis-tical significance (r=−0.246, p=0.259) (Fig. 3F).
Task-based functional connectivityAs predicted a significant main effect of condition was found for all
network connections showing an increased functional connectivityduring walk as compared to watch (see Supplementary Fig. 3, FDRcorrected). Furthermore, a significant main effect of medication wasfound within the attention motor network (FDR corrected, see Fig. 4).A significant condition (watch, walk) by medication (off, on) interac-tion effect was found within the striatum (F(1,22)=5.022, p=0.035,uncorrected, see Fig. 4). Post hoc simple effect analysis showed thatpatients on medication were able to increase internal functionalconnectivity within the striatal network during walk compared to
Fig. 3. BOLD responses (p < 0.005, k > 20, uncorrected) associated with step time variability as per the parametric modulator analysis within the Parkinson's disease group (n=23),between medication states (off/on). A=Whole brain BOLD responses within the Parkinson's disease group off medication (PD OFF); B=Whole brain BOLD responses within theParkinson's disease group on medication (PD ON); C=Whole brain BOLD responses within the Parkinson's disease group between medication states (PD OFF > PD ON); D=Parametricmodulator data from one subjects in each group over the time course of the virtual reality task; E=Mean Beta values for the regions associated with the parametric modulator contrastOFF >ON, as per figure C; F=Scatterplot and linear correlation between the loading of the parametric modulator (Beta values) onto the right lateral cerebellum cluster that was found tobe significantly different on the contrast (OFF >ON) as per figure C and the total functional connectivity within regions of the attention motor control network for Parkinson patients offmedication (OFF) and on medication (ON). NOTE: ΔFSL=Absolute normalized and demeaned footstep latency per Repetition Time (3 s) over the course of the virtual reality task walkcondition; HC=Healthy Controls; CBM=Cerebellum, PMd=dorsal premotor cortex, PPC=Posterior Parietal Cortex; *indicates significant correlation.
M. Gilat et al. NeuroImage 152 (2017) 207–220
213

watch (Mean difference=0.367, p=0.003, FDR corrected), whereaspatients off medication were not (Mean difference=0.106, p=0.285).
To examine how striatal dopamine modulates the interactionsacross large-scale networks, we further examined the effects ofdopamine on internal network connectivity. Results showed thatdopaminergic modulation of the striatal network significantly corre-lated with the degree of dopaminergic modulation of internal motorautomaticity network (r=0.536, p=0.008, Bonferonni corrected) andinternal attention-motor network connectivity (r=0.464, p=0.026,uncorrected). As expected, there was no relationship between dopami-nergic modulation of the attention-motor network and motor auto-maticity networks (r=0.219, p=0.315), indicating that the dopaminer-gic innervation of the striatum may be a key driving factor inmodulating dopamine mediated change in functional connectivity inother large-scale networks.
Imaging-behaviour associationsWe further examined the relationship between the major imaging
findings in this study and behaviour, in order to explain how break-down in network communication perturbs locomotor automaticity inthe dopamine depleted state. Following the significant condition bymedication interaction effect within the striatum, a post-hoc correla-
tion analysis was performed between the amount of functionalconnectivity within the striatum during walk and step time variability(Bonferonni corrected for multiple comparisons). Internal striatalfunctional connectivity during walk correlated strongly with increasingstep time variability in the off state (r=0.616, p=0.002), whereas nosuch correlation was found in the on state (r=−0.233, p=0.284). Tofurther explore these findings, a separate post-hoc analysis wasperformed to see which striatal circuit (e.g. cognitive-motor, limbic-motor or limbic-cognitive) was driving this correlation with step timevariability during walk (Fig. 5A-B). Interestingly, a significant correla-tion was found between step time variability and the functionalconnectivity in the striatal limbic-motor (r=0.688, p < 0.001) andlimbic-cognitive (r=0.597, p=0.003) circuits, whereas the cognitive-motor (r=0.380, p=0.068) functional connectivity did not reachstatistical significance (Fig. 5A-B). No significant correlations werefound in the on state (limbic-motor: r=−0.153, p=0.487; limbic-cognitive: r=−0.227, p=0.297; cognitive-motor: r=−0.142, p=0.519).
As aforementioned, the whole brain analysis comparing walk >watch between medication states revealed increased bilateral BOLDresponses in the orbitofrontal cortex in the off state (see Fig. 2). Theorbitofrontal cortex is known to have connections with multiple areasof the striatum allowing it to adapt behaviour (Haber et al., 1995;
Fig. 4. Results for the Condition (watch, walk) by Medication (off, on) Repeated Measures ANOVA on functional connectivity values within and between the three networks inParkinson's disease patients (n=23). Top left - Main effect of Medication (off, on); Bottom left - Condition×Medication Interaction effect; Each matrix provides summary statistics fromthe Repeated measures ANOVA analysis for within- and between network functional connectivity. P-values are embedded within each cell of the matrix. The colour scale represents F-values. Top right - Mean MTD values within the attention motor control network showing the main effect of medication; Bottom right - Mean MTD values within the striatum networkshowing the significant condition×medication interaction effect. NOTE: Auto=Motor Automaticity Network; Attn=Attention Motor Network; Stri=Striatum Network; PD off=Parkinson'sdisease patients off dopaminergic replacement medication; PD on=Parkinson's disease patients on dopaminergic replacement medication; *p < 0.05, **p < 0.05 FDR corrected.
M. Gilat et al. NeuroImage 152 (2017) 207–220
214

Schoenbaum et al., 2009; Rolls, 2015). As such, to explore itsinfluences in the current study a second post-hoc analysis wasperformed to investigate whether the functional connectivity betweenthe orbitofrontal cortex (see supplementary table 3 for MNI coordi-nates) and intra-striatal circuits was correlated with step time varia-bility (Fig. 5C). The results showed that the functional connectivitybetween the bilateral orbitofrontal cortex and inferior ventral striatum(limbic, r=0.653, p < 0.001) and dorsal putamen seeds (motor,r=0.643, p < 0.001) were significantly correlated to step time variabilityin the off state (Bonferonni corrected for multiple comparisons),whereas no correlation was found between step time variability andthe functional connectivity between the orbitofrontal cortex and dorsalcaudate nucleus (cognitive, r=0.293, p=0.174) (Fig. 5C). Again nosignificant correlations were found in the on state (motor: r=−0.323,p=0.132; cognitive: r=−0.167, p=0.447; limbic: r=−0.304, p=0.159).
Finally, it has been proposed that local depletion of dopamine levelswithin the striatum in Parkinson's disease can lead to a reduced abilityto consecutively process information through complementary yetcompeting cortico-striatal neural pathways (e.g. motor, cognitive andlimbic pathways), which may eventually result in the inhibition ofbrainstem locomotor centres (Lewis and Barker, 2009; Lewis andShine, 2014). We therefore wanted to explore whether the increasedstriatal limbic functional connectivity in Parkinson's disease patientsoff medication could have been influenced by a reduced ability tointegrate this information into the consecutive motor or cognitivecortico-striatal pathways. As such, a post-hoc analysis was performedby correlating the degree of functional connectivity within the striatallimbic circuits (e.g. limbic-motor and limbic-cognitive) with theamount of functional connectivity between the primary motor cortex
and dorsal putamen (motor cortico-striatal loop) and between thedorsolateral prefrontal cortex and dorsal caudate (cognitive cortico-striatal loop) (Fig. 6A). Results showed that during the on state, thefunctional connectivity in intra-striatal limbic circuits correlatedstrongly with the functional connectivity between regions of thecognitive cortico-striatal loop (striatal limbic-motor: r=0.615,p=0.002; striatal limbic-cognitive: r=0.711, p < 0.001), but not themotor cortico-striatal loop (striatal limbic-motor: r=−0.125, p=0.569,limbic-cognitive: r=0.204, p=0.351) (Fig. 6B-C). Interestingly, nosignificant correlations were found in the off medication state foreither the cognitive cortico-striatal loop (striatal limbic-motor:r=−0.189, p=0.387; limbic-cognitive: r=0.077, p=0.728) or the motorcortico-striatal loop (striatal limbic-motor: r=0.080, p=0.718; limbic-cognitive: r=0.074, p=0.738).
Discussion
To our knowledge, this is the first study to investigate the role ofdopamine on the neural mechanisms underlying lower limb motorautomaticity impairments in Parkinson's disease. Twenty-three pa-tients with Parkinson's disease performed an interactive virtual realityparadigm consisting of two conditions (walk, watch) in conjunctionwith functional MRI both on and off dopaminergic medication. Themain results were: (i) Parkinson's disease patients had greater steptime variability off dopaminergic medication and recruited the bilateralorbitofrontal cortex when performing lower limb movements in thevirtual reality task, as compared to when appropriately medicated; (ii)in the dopamine-depleted “off” state, patients with Parkinson's diseasedemonstrated an over-reliance on regions associated with cognitive
Fig. 5. Post-hoc correlation analysis between functional connectivity in limbic circuits and step time variability during the walk condition of the virtual reality task in Parkinson'sdisease patients off their dopaminergic medication. A=Representation of the limbic intra-striatal and orbitofrontal-striatal functional connections; B=Scatter plots for the correlationanalyses between functional connectivity in the intra-striatal limbic circuits and step time variability; C=Scatter plots for the correlation analyses between functional connectivity in theorbitofrontal-striatal limbic circuits and step time variability. NOTE: PD OFF=Parkinson's disease patients off their dopaminergic replacement medication (n=23); Step time variabilitymeasured as the coefficient of variation in footstep latencies; DC=Dorsal Caudate; DCP=Dorsal Caudal Putamen; VSi=Inferior Ventral Striatum; OFC=Orbitofrontal Cortex;CV=Coefficient of Variation; *Indicates a significant p-value (p < 0.05) that survived Bonferonni correction for multiple comparisons.
M. Gilat et al. NeuroImage 152 (2017) 207–220
215

control, which is in contrast to the recruitment of the cerebellum tomaintain low variability in the “on” state; (iii) dopamine had ademonstrable influence on intra-striatal functional connectivity duringlower limb movements; (iv) in the dopamine depleted-state, functionalconnectivity in orbitofrontal-striatal limbic circuits was correlated withstep time variability; (v) with the administration of dopamine, theaforementioned striatal limbic circuits became coupled with cognitivecortico-striatal pathways that are putatively used to integrate the limbicinformation in order to maintain effective motor performance.
As predicted, patients with Parkinson's disease demonstrated anincreased step time variability during the virtual reality gait task in thedopamine depleted state compared to when medicated, indicative of aloss of motor automaticity (Hausdorff 2003b; Yogev et al., 2005;Plotnik et al., 2011; Kelly et al., 2012; Gilat et al., 2013). This findingis in accordance with a broad literature showing that dopaminergicmedication improves step time variability during over ground walkingin Parkinson's disease (Hausdorff et al., 2003b; Schaafsma et al., 2003;Bryant et al., 2016) and is thought to indicate a shift from anautomated towards a more attention demanding cognitive strategy ofmotor control (Bohnen and Jahn, 2013; Clark, 2015; Peterson andHorak, 2016).
During the walk condition, patients with Parkinson's disease in thedopamine depleted state demonstrated significantly greater BOLDactivation across the bilateral orbitofrontal cortex compared to the onstate (TFCE corrected). The orbitofrontal cortex is involved in manyfunctions including modulation of attention and goal-directed beha-viour (Rolls and Grabenhorst, 2008; Lewis and Barker, 2009;Takahashi et al., 2011; Hartikainen et al., 2012; Marinelli et al.,2015). These findings are in accordance with Ouchi et al. (2001) whoused dopamine transporter PET imaging (DAT and [11C]CFT) to showthat Parkinson's disease patients off their medication have significantlyincreased activation in dopaminergic neurons of the bilateral orbito-frontal cortices during continuous straight walking, whereas gait inhealthy controls activated the dopaminergic neurons in the putamen(Ouchi et al., 2001). Importantly, orbitofrontal [11C]CFT uptake inParkinson's disease was inversely correlated with cadence during gait(Ouchi et al., 2001).
The novel parametric modulator analysis used in the current studyshowed that during periods of increasing variability Parkinson's diseasepatients on medication engaged the bilateral cerebellum, a key hubknown to be important for automated feed-forward control of motortiming and adaptation (Rand et al., 1998; Lang and Bastian, 1999;Doya, 2000; Morton and Bastian, 2006; Wu and Hallett, 2013).Recruiting this region may have allowed patients on medication toappropriately adapt to sudden changes in step timing variabilitywithout the need for attentional control (Horak and Diener, 1994;Rand et al., 1998; Doya, 2000; Morton and Bastian, 2006)Withoutdopaminergic medication however, the same patients became unable torecruit the cerebellum and instead relied on cortical regions associatedwith cognitive control. The slow and serial processing of these cognitiveresources (Schneider and Chein, 2003) could have required a longertime for peripheral information to be integrated with the steppingpattern resulting in a higher step time variability (Lucas et al., 2013;Shine, et al., 2013a; Clark, 2015; Hamacher et al., 2015). These resultshowever did not survive TFCE correction for multiple comparisons,warranting cautious interpretation.
A post-hoc analysis further showed that the loading of thevariability regressor onto the cerebellum correlated significantly withthe amount of functional connectivity within the attentional motornetwork in the off state. This novel finding could reflect a compensatoryincrease in network level organization where the attentional motornetwork might be attempting to engage the cerebellum following a lossof motor automaticity in the striatum (Wu et al., 2009; 2011), althoughseemingly failing to do so. As the cerebellum receives relatively minordopaminergic innervation (localized mostly in the vermis) (Melchitzkyand Lewis, 2000) but shares strong reciprocal connections to the basalganglia (Morton and Bastian, 2004; Bostan et al., 2013), it could bethrough functional coupling with the basal ganglia, that dopaminemodulates cerebellar circuits. The failure to recruit the cerebellum inthe off state of Parkinson's disease could therefore be the result ofimpaired basal ganglia – cerebellar coupling (Morton and Bastian,2004; Bostan et al., 2013). This is in accordance with previous restingstate functional MRI studies showing reduced functional couplingbetween the striatum and the cerebellum in the dopamine depleted
Fig. 6. Post-hoc correlation analysis between the amount of functional connectivity in the cognitive and motor cortico-striatal loops and intra-striatal limbic pathways in Parkinson'sdisease patients on dopaminergic replacement medication (n=23). A=Representation of the cortico-striatal and intra-striatal limbic pathways used for the analysis; B=Scatter plots forthe correlations between the amount of functional connectivity between the cognitive cortico-striatal loop and intra-striatal limbic pathways; C=Scatter plots for the correlations betweenthe amount of functional connectivity between the motor cortico-striatal loop and intra-striatal limbic pathways. NOTE: PD ON=Parkinson's disease patients on dopaminergicreplacement medication; DC=Dorsal Caudate; DCP=Dorsal Caudal Putamen; VSi=Inferior Ventral Striatum; DLPFC=Dorsolateral Prefrontal Cortex; M1=Primary motor cortex of theleg area; *Indicates a significant p-value (p < 0.05) that survived Bonferonni correction for multiple comparisons.
M. Gilat et al. NeuroImage 152 (2017) 207–220
216

state of Parkinson's disease (Hacker et al., 2012; Jech et al., 2013; Bellet al., 2014).
Striatal dysfunction can lead to an over-activation of the outputnuclei of the basal ganglia (e.g. subthalamic nucleus and globuspallidus internus) that send inhibitory GABAergic projections to thecerebellum (via the pontine nuclei), thus hampering cerebellar com-pensation abilities in Parkinson patients off medication (Lewis andBarker, 2009; Bostan et al., 2013; Lewis and Shine, 2014). This isfurther evidenced by an increase in neuronal activation of deepcerebellar nuclei following high-frequency stimulation of the subtha-lamic nucleus in rats (Moers-Hornikx et al., 2011) and normalizedcerebellar activation in patients with Parkinson's disease that receivedsubthalamic deep brain stimulation (Asanuma et al., 2006; Graftonet al., 2006; Hill et al., 2013; Wu and Hallett, 2013). Future studies areencouraged to further investigate these important dopamine relatedcompensatory and pathological alterations in striatal-cerebellar andcortico-cerebellar networks during gait in Parkinson's disease (Wu andHallett, 2013). In addition, these results lend weight to investigatingthe potential therapeutic implications of non-invasive cerebellar sti-mulation techniques (e.g. transcranial magnetic stimulation) to im-prove gait in patients with Parkinson's disease (Koch et al., 2008). Thecerebellum also shares connections with the frontal and parietalcortices via the thalamus, which is also intimately involved in corti-co-basal ganglia circuitry (Lewis and Barker, 2009; Bohnen and Jahn,2013; Verlinden et al., 2016). It is therefore somewhat surprising thatthe current study did not find any significant thalamic influence on steptime variability. For instance, higher white matter microstructureradiations between the thalamus and cortical and cerebellar regionshave recently been shown to be associated with improved gaitmeasures, including step width variability in healthy elderly(Verlinden et al., 2016). Future studies specifically examining thala-mic-cortical connectivity as a function of motor automaticity may bemore sensitive to the effects of dopamine on thalamic circuitry. Inaddition, it remains to be determined whether white matter changesare associated with increased gait variability in Parkinson's disease.The negative association found in the off state between step timevariability and BOLD responses of the right dorsal premotor cortex andleft posterior parietal cortex indicate that Parkinson's disease patientsare unable to bring these brain regions online, leading to increasingstep time variability. Alternatively, patients may have utilized thesecortical regions as a compensatory strategy to improve their steppingperformance in the off state. Future studies are now needed to testthese hypotheses, as causality could not be inferred from these results.
The functional connectivity analysis revealed a significant main effectof condition where each within and between network connectionsignificantly increased its functional connectivity during walk comparedto watch, highlighting the involvement of these predefined networks inlower limbmotor performance in Parkinson's disease (Lewis and Barker,2009; Bohnen and Jahn, 2013; Hamacher et al., 2015; Wu et al., 2015;Peterson and Horak, 2016). Furthermore, a significant main effect ofmedication was found within the attentional motor network, showingoverall increased functional connectivity in the off state. This dopami-nergic effect likely reflects the attentional compensatory strategy em-ployed by Parkinson patients off medication as a result of impairedstriatal functioning (Bohnen and Jahn, 2013; Clark, 2015; Wu et al.,2015). Indeed, the dopaminergic innervation of the striatum signifi-cantly correlated with the effects of dopamine within the other networks,reflecting the importance of striatal dopamine in large-scale networkfunction during lower limb movements (Kelly et al., 2009; Jech et al.,2013; Bell et al., 2014). Furthermore, a significant condition bymedication interaction effect was found within the striatum. This findingadvances previous resting state functional MRI studies by showing thatimpaired integration across striatal subdivisions in the dopaminedepleted state of Parkinson's disease found during rest also affects lowerlimb motor performance (Helmich et al., 2010; Surmeier et al., 2010;Hacker et al., 2012; Sharman et al., 2013; Bell et al., 2014).
To our knowledge, this study is the first to show that impairedcrosstalk across the dopaminergically-depleted striatum of Parkinson'sdisease correlates with increased step time variability. Furthermore, ourpost-hoc analyses revealed that intra-striatal limbic circuits were drivingthis correlation with worse step time variability. In fact, the functionalconnectivity within the orbitofrontal-ventral striatum limbic circuit wasstrongly correlated with increased step time variability in the dopaminedepleted state. The current study therefore provides novel pathophysio-logical evidence to suggest that activation in the orbitofrontal cortexduring gait in Parkinson's disease is related to activity within a limbiccortico-striatal circuit that, in the context of reduced dopamine, inter-feres with the striatal control of lower limb motor function.
Our findings are supported by primate work showing that theterminals of the orbitofrontal cortex are extensive throughout thedopaminergic neurons, which influence a wide area of the striatum,particularly the ventral striatum and core of the nucleus accumbens(Haber et al., 1995). Furthermore, a resting state functional MRI studyby Di Martino et al. (2008) showed that in healthy adults thespontaneous fluctuations in BOLD response in the inferior ventralstriatum primarily correlated with the orbitofrontal cortex, indicatingthat strong functional connections exist between these regions inhumans (Di Martino et al., 2008). Yang et al. (2016) also recentlyshowed that functional connectivity during rest was increased betweenthe ventral striatum and orbitofrontal cortex in Parkinson's diseasepatients off dopaminergic medication as compared to when medica-tion, indicating that dopamine has a profound influence on thisorbitofrontal-striatal limbic circuitry (Yang et al., 2016).
The striatal projections allow the orbitofrontal cortex to regulatemotor actions under the influence of dopamine, for instance to adaptbehaviour in the face of unexpected outcomes (Schoenbaum et al.,2009; Takahashi et al., 2009; Rolls, 2015). It has previously beenproposed that error signals processed by the midbrain dopaminergicneurons that project to the striatum originate within regions of theorbitofrontal cortex that encode expected value and performanceoutcome, and which later connect to the ventral striatum (Takahashiet al., 2011; Rolls, 2015). The orbitofrontal cortex indeed receivesnegative prediction error feedback (i.e. sensory evidence that did notmatch the predicted performance outcome) from every sensory system,making it an important hub for multisensory integration that enablesplanning and learning performance outcomes (Kringelbach, 2005;Schoenbaum et al., 2009; Takahashi et al., 2009; Goble et al., 2011;Rolls, 2015; Chanes and Barrett, 2016). As Parkinson's disease patientshave known sensorimotor impairments (Nolano et al., 2008; Conteet al., 2013; Ehgoetz Martens et al., 2013; Lucas et al., 2013), theincreased BOLD responses found in the orbitofrontal cortex in theParkinson group off medication could reflect an attempt to adaptbehaviour following increased negative prediction error feedback(Kringelbach, 2005; Schoenbaum et al., 2009; Takahashi et al.,2009). Indeed, our results further showed that increased intra-striatallimbic functional connectivity was correlated with increased functionalconnectivity within regions of the cognitive cortico-striatal pathwayselectively in the “on” state. This might indicate that with sufficientdopaminergic resources, the striatum is able to functionally integratelimbic information into the parallel cortico-striatal pathways, whichcan then provide top-down control over motor performance to resolvethe prediction error and prevent motor deterioration (i.e. compensa-tion) (Postuma and Dagher, 2006; Lewis and Barker, 2009; Rolls,2015). However, dopamine depletion likely impairs such parallelcortico-striatal integration within the striatum, as evidenced by thelack of such correlations in “off” state (Lewis and Barker, 2009; Bellet al., 2014; Wu et al., 2015). The prediction error would thereforeremain unresolved and hence, continue to induce an increased limbicdrive within the striatal network. This could impair motor performancedirectly by innervating the inhibitory basal ganglia output structuresthat project to the brainstem locomotor centres (Lewis and Barker,2009; Lewis and Shine, 2014). In addition, the increased limbic drive
M. Gilat et al. NeuroImage 152 (2017) 207–220
217

likely demands a proportion of the depleted dopaminergic resourceswithin the striatum, further depriving the dorsal striatum from itsability to process automated motor sequences (Wu et al., 2015).Therefore, based on the results of the current study, it can bepostulated that although the ventral striatal-orbitofrontal circuit mayusually be employed as a compensatory strategy to overcome negativeerror feedback, without sufficient dopaminergic resources utilizing thisnetwork can actually deteriorate motor performance in Parkinson'sdisease patients.
This study now forms the basis for future work. The orbitofrontalcortex has previously been associated with the integration of emotionalinformation into decision processes (Bechara et al., 2000). In addition,anxiety and depression are associated with impaired gait performancein Parkinson's disease patients (Rochester et al., 2004, 2005; Lordet al., 2011, 2013; Ehgoetz Martens et al., 2015), and these deficits areamenable to dopaminergic medication (Ehgoetz Martens et al., 2015).Previous authors have postulated that depression and anxiety impairgoal-directed attentional processing (Rochester et al., 2004, 2005),possibly by increasing computational load of the ventral striatum-orbitofrontal pathway resulting in response conflict in the dopaminedepleted cortico-basal ganglia motor circuitry (Lewis and Barker, 2009;Ehgoetz Martens et al., 2013, 2015). Future studies should thereforeinvestigate the complex interaction between dopamine, mood distur-bance and gait impairments in Parkinson's disease.
In addition, the current patient cohort consisted mostly of patientswith moderate bilateral disease (1 subject scored HY=1; 18 subjectsscored HY=2–3; 3 subjects scored HY=3 and no subjects scored HY >3). As such, future work is needed to investigate whether the neuralbasis underlying gait variability changes with disease severity.Furthermore, it is recommended that future studies account forlaterality in brain pathology when analysing neuroimaging data in acohort of Parkinson's disease patients with obvious unilateral symptomseverities. It is also important to note that although dopamine evidentlyplays a key role, gait disturbances in Parkinson's disease likely reflect amultisystem neurodegenerative disorder beyond the loss of dopami-nergic neurons, especially as the disease progresses (Bohnen and Albin,2011; Bohnen and Jahn, 2013; Bohnen et al., 2013). For instance,prefrontal cholinergic neurons are key players in attentional controlfunctions that may be put under increasing pressure following striataland prefrontal deterioration in Parkinson's disease (Bohnen et al.,2013; Gonzales and Smith, 2015). Gait speed has indeed been shown tobe most affected in patients with both nigrostriatal dopaminergic andcortical cholinergic denervation (Bohnen et al., 2013). Future studiesusing larger cohorts are now needed to investigate the role ofdopaminergic, cholinergic and other neurotransmitter systems on gaitautomaticity impairments in Parkinson's disease and Parkinson'sdisease subgroups, for example Parkinson's disease patients with andwithout freezing of gait (Bohnen and Albin, 2011; Bohnen et al., 2014).
Furthermore, dual task interference provides an alternative avenueto study motor automaticity impairments (Yogev et al., 2005; Kellyet al., 2012; Wu et al., 2015). Future studies are therefore encouragedto further evaluate dual task interference to confirm that the increasedstep time variability seen in the Parkinson's disease patients offmedication reflects reduced motor automaticity (Poldrack et al.,2005; Wu et al., 2015). However, similar to walking and most dailybehaviours, simple motor tasks such as foot tapping are usuallyperformed automatically in healthy subjects (Wu et al., 2015).Furthermore, once a motor task is automatic it becomes difficult torevert back to controlled behaviour (Schneider and Chein, 2003; Wuet al., 2015). It can therefore be assumed that the performance inhealthy control participants and low variability seen in Parkinson’sdisease patients on medication indicated a more automatic motorperformance. In addition, a supplementary correlation analysis wasperformed between the functional connectivity values and anotherbehavioural motor outcome of the virtual reality paradigm, namelymodal footstep latency, showing no significant correlations (See
Supplementary Table 4). This further indicates that the findings ofthe current study are indeed specific to step time variability and thusmotor automaticity. In accordance with the findings in this study,previous behavioural research has shown that the administration ofdopaminergic replacement therapy often improves step time variabilityin Parkinson's disease patients (Hausdorff et al., 2003a; Lord et al.,2011; Rochester et al., 2011; Bryant et al., 2016). Furthermore,impairments in dual-task walking are also often improved by optimaldopaminergic replacement therapy (Camicioli et al., 1998; Lord et al.,2011; Elshehabi et al., 2016). Together, this evidence suggests thatdopaminergic therapy may influence gait-related neural computationsthrough modulation of motor automaticity.
While multiple motor skills rely upon motor automaticity, in thisstudy, step time variability was employed as a surrogate for motorautomaticity in an attempt to model simple gait. Equally, however, dualtask paradigms can also provide insights into the breakdown ofautomaticity in Parkinson's disease by loading additional attentionalresources with overt task demands. Future studies are now needed tofurther investigate the interaction between dopamine and dual taskingability during gait in patients with Parkinson's disease. Finally, alimitation inherent in the study of motor automaticity is that indirectsurrogate measures are required. Therefore, the component processes bywhich dopaminergic replacement therapy improves motor skills cannotbe precisely isolated. While it is hypothesized that dopamine replace-ment therapy exerts its major mechanism of action by improving motorautomaticity in subcortical structures leading to reduced step timevariability, dopamine may also modulate other sub-processes including,coordination, balance, proprioception, affective processing and cognitionthat may affect step time variability and motor automaticity.
In conclusion, this study showed that dopamine depletion inParkinson's disease impairs motor automaticity by reducing striatalfunctioning and cerebellar compensation strategies, which lead toincreased attentional motor control and excessive orbitofrontal-striatallimbic interference.
Funding
MG is supported by a University of Sydney InternationalScholarship; PTB has no funding to declare; KAEM is supported by aPostdoctoral Fellowship from Parkinson Canada; MJG is supported bythe University of Sydney RA Money Postgraduate Research Scholarshipin Neuroscience; JMH is supported by a Western Sydney UniversityPostgraduate Research Scholarship; CCW has no funding to declare;SJGL is supported by a NHMRC-ARC Dementia Fellowship 1110414;JMS is supported by a CJ Martin Fellowship from the National Healthand Medical Research Council (GNT1072403).
Acknowledgements
We would like to thank the patient volunteers and their families fortheir participation in this study. The authors declare that they have noconflicts of interest, financial or otherwise.
Appendix A. Supporting information
Supplementary data associated with this article can be found in theonline version at doi:10.1016/j.neuroimage.2017.02.073.
References
Alexander, G.E., DeLong, M.R., Strick, P.L., 1986. Parallel organization of functionallysegregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 9,357–381.
Asanuma, K., Tang, C., Ma, Y., Dhawan, V., Mattis, P., Edwards, C., et al., 2006. Networkmodulation in the treatment of Parkinson's disease. Brain 129, 2667–2678.
Ashby, F.G., Turner, B.O., Horvitz, J.C., 2010. Cortical and basal ganglia contributions tohabit learning and automaticity. Trends Cogn. Sci. 14, 208–215.
M. Gilat et al. NeuroImage 152 (2017) 207–220
218

Bechara, A., Damasio, H., Damasio, A.R., 2000. Emotion, decision making and theorbitofrontal cortex. Cereb. Cortex 10, 295–307.
Bell, P.T., Gilat, M., O'Callaghan, C., Copland, D.A., Frank, M.J., Lewis, S.J.G., et al.,2014. Dopaminergic basis for impairments in functional connectivity acrosssubdivisions of the striatum in Parkinson's disease. Hum. Brain Mapp. 36,1278–1291.
Bohnen, N.I., Albin, R.L., 2011. The cholinergic system and Parkinson disease. Behav.Brain Res. 221, 564–573.
Bohnen, N.I., Jahn, K., 2013. Imaging: what can it tell us about parkinsonian gait? Mov.Disord. 28, 1492–1500.
Bohnen, N.I., Frey, K.A., Studenski, S., Kotagal, V., Koeppe, R.A., Scott, P.J.H., et al.,2013. Gait speed in Parkinson disease correlates with cholinergic degeneration.Neurology 81, 1611–1616.
Bohnen, N.I., Frey, K.A., Studenski, S., Kotagal, V., Koeppe, R.A., Constantine, G.M.,et al., 2014. Extra-nigral pathological conditions are common in Parkinson's diseasewith freezing of gait: an in vivo positron emission tomography study. Mov. Disord.29, 1118–1124.
Bostan, A.C., Dum, R.P., Strick, P.L., 2013. Cerebellar networks with the cerebral cortexand basal ganglia. Trends Cogn. Sci. 17, 241–254.
Brett, M., Anton, J.-L., Valabregue, R., Poline, J.-B., 2002. Region of interest analysisusing an SPM toolbox. In: Proceedings of the Eight International Conference onFuncthional Mapping of the Human Brain.
Brooks, D.J., Ibanez, V., Sawle, G.V., Quinn, N., Lees, A.J., Mathias, C.J., et al., 1990.Differing patterns of striatal 18F-dopa uptake in Parkinson's disease, multiplesystem atrophy, and progressive supranuclear palsy. Ann. Neurol. 28, 547–555.
Bryant, M.S., Rintala, D.H., Hou, J.G., Collins, R.L., Protas, E.J., 2016. Gait variability inParkinson's disease: levodopa and walking direction. Acta Neurol. Scand. 134,83–86.
Camicioli, R., Oken, B.S., Sexton, G., Kaye, J.A., Nutt, J.G., 1998. Verbal fluency taskaffects gait in Parkinson's disease with motor freezing. J. Geriatr. Psychiatry Neurol.11, 181–185.
Carbon, M., Marié, R.-M., 2003. Functional imaging of cognition in Parkinson's disease.Curr. Opin. Neurol. 16, 475–480.
Chanes, L., Barrett, L.F., 2016. Redefining the role of limbic areas in cortical processing.Trends Cogn. Sci. 20, 96–106.
Clark, D.J., 2015. Automaticity of walking: functional significance, mechanisms,measurement and rehabilitation strategies. Front. Hum. Neurosci. 9, 246.
Cole, M.W., Schneider, W., 2007. The cognitive control network: integrated corticalregions with dissociable functions. Neuroimage 37, 343–360.
Conte, A., Khan, N., Defazio, G., Rothwell, J.C., Berardelli, A., 2013. Pathophysiologyof somatosensory abnormalities in Parkinson disease. Nat. Rev. Neurol. 9,687–697.
Di Martino, A., Scheres, A., Margulies, D.S., Kelly, A.M.C., Uddin, L.Q., Shehzad, Z.,et al., 2008. Functional connectivity of human striatum: a resting state FMRI study.Cereb. Cortex 18, 2735–2747.
Diagnostic and Statistical Manual of Mental Disorders (4th ed., text rev.).Doya, K., 2000. Complementary roles of basal ganglia and cerebellum in learning and
motor control. Curr. Opin. Neurobiol. 10, 732–739.Doyon, J., Song, A.W., Karni, A., Lalonde, F., Adams, M.M., Ungerleider, L.G., 2002.
Experience-dependent changes in cerebellar contributions to motor sequencelearning. Proc. Natl. Acad. Sci. USA 99, 1017–1022.
Duff, E.P., Cunnington, R., Egan, G.F., 2007. REX: response exploration forneuroimaging datasets. Neuroinformatics 5, 223–234.
Ehgoetz Martens, K.A., Pieruccini-Faria, F., Almeida, Q.J., 2013. Could sensorymechanisms be a core factor that underlies freezing of gait in Parkinson's disease?PLoS One 8, e62602.
Ehgoetz Martens, K.A., Ellard, C.G., Almeida, Q.J., 2015. Anxiety-provoked gait changesare selectively dopa-responsive in Parkinson's disease. Eur. J. Neurosci. 42,2028–2035.
Elshehabi, M., Maier, K.S., Hasmann, S.E., Nussbaum, S., Herbst, H., Heger, T., et al.,2016. Limited effect of dopaminergic medication on straight walking and turning inearly-to-moderate Parkinson's disease during single and dual tasking. Front. AgingNeurosci. 8, 4.
Everitt, B.J., Robbins, T.W., 2016. Drug addiction: updating actions to habits tocompulsions ten years on. Annu. Rev. Psychol. 67, 23–50.
Frenkel-Toledo, S., Giladi, N., Peretz, C., Herman, T., Gruendlinger, L., Hausdorff, J.M.,2005. Effect of gait speed on gait rhythmicity in Parkinson's disease: variability ofstride time and swing time respond differently. J. Neuroeng. Rehabil. 2, 23.
Friedman, A., Polson, M.C., Dafoe, C.G., Gaskill, S.J., 1982. Dividing attention withinand between hemispheres: testing a multiple resources approach to limited-capacityinformation processing. J. Exp. Psychol. Hum. Percept. Perform. 8, 625–650.
Georgiades, M.J., Gilat, M., Ehgoetz Martens, K.A., Walton, C.C., Bissett, P.G., Shine,J.M., et al., 2016. Investigating motor initiation and inhibition deficits in patientswith Parkinson's disease and freezing of gait using a virtual reality paradigm.Neuroscience 337, 153–162.
Gilat, M., Shine, J.M., Bolitho, S.J., Matar, E., Kamsma, Y.P.T., Naismith, S.L., et al.,2013. Variability of stepping during a virtual reality paradigm in Parkinson's diseasepatients with and without freezing of gait. PLoS One 8, e66718.
Gilat, M., Shine, J.M., Walton, C.C., O'Callaghan, C., Hall, J.M., Lewis, S.J.G., 2015.Brain activation underlying turning in Parkinson's disease patients with and withoutfreezing of gait: a virtual reality fMRI study. Nat. Partn. J. Park. Dis..
Goble, D.J., Coxon, J.P., Van Impe, A., Geurts, M., Doumas, M., Wenderoth, N., et al.,2011. Brain activity during ankle proprioceptive stimulation predicts balanceperformance in young and older adults. J. Neurosci. 31, 16344–16352.
Goetz, C.G., Emre, M., Dubois, B., 2008. Parkinson's disease dementia: definitions,guidelines, and research perspectives in diagnosis. Ann. Neurol. 64 (2), S81–S92.
Gonzales, K.K., Smith, Y., 2015. Cholinergic interneurons in the dorsal and ventralstriatum: anatomical and functional considerations in normal and diseasedconditions. Ann. N.Y. Acad. Sci. 1349, 1–45.
Grafton, S.T., Turner, R.S., Desmurget, M., Bakay, R., Delong, M., Vitek, J., et al., 2006.Normalizing motor-related brain activity: subthalamic nucleus stimulation inParkinson disease. Neurology 66, 1192–1199.
Graybiel, A.M., 1998. The basal ganglia and chunking of action repertoires. Neurobiol.Learn Mem. 70, 119–136.
Haber, S.N., Kunishio, K., Mizobuchi, M., Lynd-Balta, E., 1995. The orbital and medialprefrontal circuit through the primate basal ganglia. J. Neurosci. 15, 4851–4867.
Hacker, C.D., Perlmutter, J.S., Criswell, S.R., Ances, B.M., Snyder, A.Z., 2012. Resting statefunctional connectivity of the striatum in Parkinson's disease. Brain 135, 3699–3711.
Hamacher, D., Herold, F., Wiegel, P., Hamacher, D., Schega, L., 2015. Brain activityduring walking: a systematic review. Neurosci. Biobehav Rev. 15, 310–327.
Hanakawa, T., Fukuyama, H., Katsumi, Y., Honda, M., Shibasaki, H., 1999. Enhancedlateral premotor activity during paradoxical gait in Parkinson's disease. Ann. Neurol.45, 329–336.
Hartikainen, K.M., Ogawa, K.H., Knight, R.T., 2012. Orbitofrontal cortex biases attentionto emotional events. J. Clin. Exp. Neuropsychol. 34, 588–597.
Hausdorff, J.M., Cudkowicz, M.E., Firtion, R., Wei, J.Y., Goldberger, A.L., 1998. Gaitvariability and basal ganglia disorders: stride-to-stride variations of gait cycle timingin Parkinson's disease and Huntington's disease. Mov. Disord. 13, 428–437.
Hausdorff, J.M., Schaafsma, J.D., Balash, Y., Bartels, A.L., Gurevich, T., Giladi, N.,2003a. Impaired regulation of stride variability in Parkinson's disease subjects withfreezing of gait. Exp. Brain Res. 149, 187–194.
Hausdorff, J.M., Balash, J., Giladi, N., 2003b. Effects of cognitive challenge on gaitvariability in patients with Parkinson's disease. J. Geriatr. Psychiatry Neurol. 16,53–58.
Hausdorff, J.M., Lowenthal, J., Herman, T., Gruendlinger, L., Peretz, C., Giladi, N., 2007.Rhythmic auditory stimulation modulates gait variability in Parkinson's disease. Eur.J. Neurosci. 26, 2369–2375.
Helmich, R.C., Derikx, L.C., Bakker, M., Scheeringa, R., Bloem, B.R., Toni, I., 2010.Spatial remapping of cortico-striatal connectivity in Parkinson's disease. Cereb.Cortex 20, 1175–1186.
Helmich, R.C., Bloem, B.R., Toni, I., 2012. Motor imagery evokes increasedsomatosensory activity in Parkinson's disease patients with tremor. Hum. BrainMapp. 33, 1763–1779.
Hill, K.K., Campbell, M.C., McNeely, M.E., Karimi, M., Ushe, M., Tabbal, S.D., et al.,2013. Cerebral blood flow responses to dorsal and ventral STN DBS correlate withgait and balance responses in Parkinson's disease. Exp. Neurol. 241, 105–112.
Horak, F.B., Diener, H.C., 1994. Cerebellar control of postural scaling and central set instance. J. Neurophysiol. 72, 479–493.
Jech, R., Mueller, K., Schroeter, M.L., Růžička, E., 2013. Levodopa increases functionalconnectivity in the cerebellum and brainstem in Parkinson's disease. Brain 136,(e234–e234).
Kelly, C., de Zubicaray, G., Di Martino, A., Copland, D.A., Reiss, P.T., Klein, D.F., et al.,2009. L-dopa modulates functional connectivity in striatal cognitive and motornetworks: a double-blind placebo-controlled study. J. Neurosci. 29, 7364–7378.
Kelly, V.E., Eusterbrock, A.J., Shumway-Cook, A., 2012. A review of dual-task walkingdeficits in people with Parkinson's disease: motor and cognitive contributions,mechanisms, and clinical implications. Park. Dis. 2012, 918719.
Koch, G., Mori, F., Marconi, B., Codecà, C., Pecchioli, C., Salerno, S., et al., 2008. Changesin intracortical circuits of the human motor cortex following theta burst stimulationof the lateral cerebellum. Clin. Neurophysiol. 119, 2559–2569.
Kringelbach, M.L., 2005. The human orbitofrontal cortex: linking reward to hedonicexperience. Nat. Rev. Neurosci. 6, 691–702.
Kurz, M.J., Wilson, T.W., Arpin, D.J., 2012. Stride-time variability and sensorimotorcortical activation during walking. Neuroimage 59, 1602–1607.
Lang, C.E., Bastian, A.J., 1999. Cerebellar subjects show impaired adaptation ofanticipatory EMG during catching. J. Neurophysiol. 82, 2108–2119.
Lehéricy, S., Benali, H., Van de Moortele, P.-F., Pélégrini-Issac, M., Waechter, T.,Ugurbil, K., et al., 2005. Distinct basal ganglia territories are engaged in early andadvanced motor sequence learning. Proc. Natl. Acad. Sci. USA 102, 12566–12571.
Lewis, S.J.G., Barker, R.A., 2009. A pathophysiological model of freezing of gait inParkinson's disease. Park. Relat. Disord. 15, 333–338.
Lewis, S.J.G., Shine, J.M., 2014. The next step: a common neural mechanism for freezingof gait. Neuroscientist, (Epub ahead of print: 1073858414559101).
Lieberman, M.D., Cunningham, W.A., 2009. Type I and Type II error concerns in fMRIresearch: re-balancing the scale. Soc. Cogn. Affect. Neurosci. 4, 423–428.
Lord, S., Baker, K., Nieuwboer, A., Burn, D., Rochester, L., 2011. Gait variability inParkinson's disease: an indicator of non-dopaminergic contributors to gaitdysfunction? J. Neurol. 258, 566–572.
Lord, S., Galna, B., Coleman, S., Burn, D., Rochester, L., 2013. Mild depressive symptomsare associated with gait impairment in early Parkinson's disease. Mov. Disord. 28,634–639.
Lucas, M., Chaves, F., Teixeira, S., Carvalho, D., Peressutti, C., Bittencourt, J., et al.,2013. Time perception impairs sensory-motor integration in Parkinson's disease. Int.Arch. Med. 6, 39.
Marinelli, L., Piccardo, A., Mori, L., Morbelli, S., Girtler, N., Castaldi, A., et al., 2015.Orbitofrontal (18) F-DOPA uptake and movement preparation in Parkinson'sdisease. Park. Dis., 180940.
Mazaika, P., Whitfield-Gabrieli, S., Reiss, A.L., 2007. Artifact repair for fMRI data fromhigh motion clinical subjects. In: Presentation at Human Brain Mapping Conference.
Melchitzky, D.S., Lewis, D.A., 2000. Tyrosine hydroxylase- and dopamine transporter-immunoreactive axons in the primate cerebellum. Evidence for a lobular- andlaminar-specific dopamine innervation. Neuropsychopharmacology 22, 466–472.
M. Gilat et al. NeuroImage 152 (2017) 207–220
219

Miyachi, S., Hikosaka, O., Miyashita, K., Kárádi, Z., Rand, M.K., 1997. Differential rolesof monkey striatum in learning of sequential hand movement. Exp. Brain Res. 115,1–5.
Moers-Hornikx, V.M.P., Vles, J.S.H., Tan, S.K.H., Cox, K., Hoogland, G., Steinbusch,W.M.H., et al., 2011. Cerebellar nuclei are activated by high-frequency stimulation ofthe subthalamic nucleus. Neurosci. Lett. 496, 111–115.
Monchi, O., Petrides, M., Mejia-Constain, B., Strafella, A.P., 2007. Cortical activity inParkinson's disease during executive processing depends on striatal involvement.Brain 130, 233–244.
Morton, S.M., Bastian, A.J., 2004. Cerebellar control of balance and locomotion.Neuroscientist 10, 247–259.
Morton, S.M., Bastian, A.J., 2006. Cerebellar contributions to locomotor adaptationsduring splitbelt treadmill walking. J. Neurosci. 26, 9107–9116.
Nolano, M., Provitera, V., Estraneo, A., Selim, M.M., Caporaso, G., Stancanelli, A., et al.,2008. Sensory deficit in Parkinson's disease: evidence of a cutaneous denervation.Brain 131, 1903–1911.
Nonnekes, J., Snijders, A.H., Nutt, J.G., Deuschl, G., Giladi, N., Bloem, B.R., 2015.Freezing of gait: a practical approach to management. Lancet Neurol. 14, 768–778.
Ouchi, Y., Kanno, T., Okada, H., Yoshikawa, E., Futatsubashi, M., Nobezawa, S., et al.,2001. Changes in dopamine availability in the nigrostriatal and mesocorticaldopaminergic systems by gait in Parkinson's disease. Brain 124, 784–792.
Peterson, D.S., Horak, F.B., 2016. Neural control of walking in people withParkinsonism. Physiology 31, 95–107.
Plotnik, M., Dagan, Y., Gurevich, T., Giladi, N., Hausdorff, J.M., 2011. Effects of cognitivefunction on gait and dual tasking abilities in patients with Parkinson's diseasesuffering from motor response fluctuations. Exp. Brain Res. 208, 169–179.
Poldrack, R.A., Sabb, F.W., Foerde, K., Tom, S.M., Asarnow, R.F., Bookheimer, S.Y.,et al., 2005. The neural correlates of motor skill automaticity. J. Neurosci. 25,5356–5364.
Postuma, R.B., Dagher, A., 2006. Basal ganglia functional connectivity based on a meta-analysis of 126 positron emission tomography and functional magnetic resonanceimaging publications. Cereb. Cortex 16, 1508–1521.
Rand, M.K., Wunderlich, D.A., Martin, P.E., Stelmach, G.E., Bloedel, J.R., 1998.Adaptive changes in responses to repeated locomotor perturbations in cerebellarpatients. Exp. Brain Res. 122, 31–43.
Rascol, O., Sabatini, U., Fabre, N., Brefel, C., Loubinoux, I., Celsis, P., et al., 1997. Theipsilateral cerebellar hemisphere is overactive during hand movements in akineticparkinsonian patients. Brain 120 (Pt 1), 103–110.
Rochester, L., Hetherington, V., Jones, D., Nieuwboer, A., Willems, A.-M., Kwakkel, G.,et al., 2004. Attending to the task: interference effects of functional tasks on walkingin Parkinson's disease and the roles of cognition, depression, fatigue, and balance.Arch. Phys. Med. Rehabil. 85, 1578–1585.
Rochester, L., Hetherington, V., Jones, D., Nieuwboer, A., Willems, A.-M., Kwakkel, G.,et al., 2005. The effect of external rhythmic cues (auditory and visual) on walkingduring a functional task in homes of people with Parkinson's disease. Arch. Phys.Med. Rehabil. 86, 999–1006.
Rochester, L., Baker, K., Nieuwboer, A., Burn, D., 2011. Targeting dopa-sensitive anddopa-resistant gait dysfunction in Parkinson's disease: selective responses to internaland external cues. Mov. Disord. 26, 430–435.
Rolls, E.T., 2015. Limbic systems for emotion and for memory, but no single limbicsystem. Cortex 62, 119–157.
Rolls, E.T., Grabenhorst, F., 2008. The orbitofrontal cortex and beyond: from affect todecision-making. Prog. Neurobiol. 86, 216–244.
Schaafsma, J.D., Balash, Y., Gurevich, T., Bartels, A.L., Hausdorff, J.M., Giladi, N., 2003.Characterization of freezing of gait subtypes and the response of each to levodopa inParkinson's disease. Eur. J. Neurol. 10, 391–398.
Schneider, W., Chein, J.M., 2003. Controlled & automatic processing: behavior, theory,and biological mechanisms. Cogn. Sci. 27, 525–559.
Schoenbaum, G., Roesch, M.R., Stalnaker, T.A., Takahashi, Y.K., 2009. A new perspectiveon the role of the orbitofrontal cortex in adaptive behaviour. Nat. Rev. Neurosci. 10,885–892.
Sharman, M., Valabregue, R., Perlbarg, V., Marrakchi-Kacem, L., Vidailhet, M., Benali,H., et al., 2013. Parkinson's disease patients show reduced cortical-subcorticalsensorimotor connectivity. Mov. Disord. 28, 447–454.
Shine, J.M., Matar, E., Ward, P.B., Bolitho, S.J., Gilat, M., Pearson, M., et al., 2013a.Exploring the cortical and subcortical functional magnetic resonance imagingchanges associated with freezing in Parkinson's disease. Brain 136, 1204–1215.
Shine, J.M., Matar, E., Ward, P.B., Frank, M.J., Moustafa, A.A., Pearson, M., et al.,2013b. Freezing of gait in Parkinson's disease is associated with functionaldecoupling between the cognitive control network and the basal ganglia. Brain 136,3671–3681.
Shine, J.M., Koyejo, O., Bell, P.T., Gorgolewski, K.J., Gilat, M., Poldrack, R.A., 2015.Estimation of dynamic functional connectivity using multiplication of temporalderivatives. Neuroimage 122, 399–407.
Shine, J.M., Bissett, P.G., Bell, P.T., Koyejo, O., Balsters, J.H., Gorgolewski, K.J., et al.,2016. The dynamics of functional brain networks: integrated network states duringcognitive task performance. Neuron 92, 544–554.
Smith, S.M., Nichols, T.E., 2009. Threshold-free cluster enhancement: addressingproblems of smoothing, threshold dependence and localisation in cluster inference.Neuroimage 44, 83–98.
Strouwen, C., Molenaar, E.A.L.M., Münks, L., Keus, S.H.J., Bloem, B.R., Rochester, L.,et al., 2015. Dual tasking in Parkinson's disease: should we train hazardousbehavior? Expert Rev. Neurother. 15, 1031–1039.
Surmeier, D.J., Shen, W., Day, M., Gertler, T., Chan, S., Tian, X., et al., 2010. The role ofdopamine in modulating the structure and function of striatal circuits. Prog. BrainRes. 183, 149–167.
Takahashi, Y.K., Roesch, M.R., Stalnaker, T.A., Haney, R.Z., Calu, D.J., Taylor, A.R.,et al., 2009. The orbitofrontal cortex and ventral tegmental area are necessary forlearning from unexpected outcomes. Neuron 62, 269–280.
Takahashi, Y.K., Roesch, M.R., Wilson, R.C., Toreson, K., O'Donnell, P., Niv, Y., et al.,2011. Expectancy-related changes in firing of dopamine neurons depend onorbitofrontal cortex. Nat. Neurosci. 14, 1590–1597.
Vandenbossche, J., Deroost, N., Soetens, E., Coomans, D., Spildooren, J., Vercruysse, S.,et al., 2012. Freezing of gait in Parkinson's disease: disturbances in automaticity andcontrol. Front. Hum. Neurosci. 6, 356.
Verlinden, V.J.A., de Groot, M., Cremers, L.G.M., van der Geest, J.N., Hofman, A.,Niessen, W.J., et al., 2016. Tract-specific white matter microstructure and gait inhumans. Neurobiol. Aging 43, 164–173.
Willems, A.M., Nieuwboer, A., Chavret, F., Desloovere, K., Dom, R., Rochester, L., et al.,2006. The use of rhythmic auditory cues to influence gait in patients with Parkinson'sdisease, the differential effect for freezers and non-freezers, an explorative study.Disabil. Rehabil. 28, 721–728.
Wu, T., Hallett, M., 2005. A functional MRI study of automatic movements in patientswith Parkinson's disease. Brain 128, 2250–2259.
Wu, T., Hallett, M., 2013. The cerebellum in Parkinson's disease. Brain 136, 696–709.Wu, T., Hallett, M., Chan, P., 2015. Motor automaticity in Parkinson's disease.
Neurobiol. Dis. 82, 226–234.Wu, T., Wang, L., Chen, Y., Zhao, C., Li, K., Chan, P., 2009. Changes of functional
connectivity of the motor network in the resting state in Parkinson's disease.Neurosci. Lett. 460, 6–10.
Wu, T., Wang, L., Hallett, M., Chen, Y., Li, K., Chan, P., 2011. Effective connectivity ofbrain networks during self-initiated movement in Parkinson's disease. Neuroimage55, 204–215.
Wymbs, N.F., Bassett, D.S., Mucha, P.J., Porter, M.A., Grafton, S.T., 2012. Differentialrecruitment of the sensorimotor putamen and frontoparietal cortex during motorchunking in humans. Neuron 74, 936–946.
Yang, W., Liu, B., Huang, B., Huang, R., Wang, L., Zhang, Y., et al., 2016. Altered resting-state functional connectivity of the striatum in Parkinson's disease after Levodopaadministration. PLoS One 11, e0161935.
Yogev, G., Giladi, N., Peretz, C., Springer, S., Simon, E.S., Hausdorff, J.M., 2005. Dualtasking, gait rhythmicity, and Parkinson's disease: which aspects of gait are attentiondemanding? Eur. J. Neurosci. 22, 1248–1256.
M. Gilat et al. NeuroImage 152 (2017) 207–220
220

APPENDIX(9(
Ehgoetz Martens, K. A., Lukasik, E., Georgiades, M, J., Gilat, M., Hall, J.M.,
Walton, C. C., Lewis, S. J. G. Predicting the onset of freezing of gait: A longitudinal
study. Movement Disorders, 2017
!

Predicting the Onset of Freezing of Gait: A Longitudinal Study
Kaylena A. Ehgoetz Martens, PhD ,1* Emily L. Lukasik, MBMSc,1 Matthew J. Georgiades, BSc,1 Moran Gilat, MSc,1
Julie M. Hall, MSc,1,2 Courtney C. Walton, PhD,1 and Simon J.G. Lewis, MBBCh, MD1
1Parkinson’s Disease Research Clinic, Brain and Mind Centre, University of Sydney, Sydney, Australia2School of Social Sciences and Psychology, Western Sydney University, Sydney, Australia
ABSTRACT: Background: Freezing of gait is a dis-abling symptom of Parkinson’s disease that ultimatelyaffects approximately 80% of patients, yet very littleresearch has focused on predicting the onset of freezingof gait and tracking the longitudinal progression of symp-toms prior to its onset. The objective of the current studywas to examine longitudinal data spanning the transitionperiod when patients with PD developed freezing of gaitto identify symptoms that may precede freezing and cre-ate a prediction model that identifies those “at risk” fordeveloping freezing of gait in the year to follow.Methods: Two hundred and twenty-one patients withPD were divided into 3 groups (88 nonfreezers, 41 tran-sitional freezers, and 92 continuing freezers) based ontheir responses to the validated Freezing of Gait-Questionnaire item 3 at baseline and follow-up. Criticalmeasures across motor, cognitive, mood, and sleepdomains were assessed at 2 times approximately 1year apart.
Results: A logistic regression model that included age,disease duration, gait symptoms, motor phenotype,attentional set-shifting, and mood measures could predictwith 70% and 90% accuracy those patients who wouldand would not develop, respectively, freezing of gait overthe next year. Notably, the Freezing of Gait-Questionnairetotal and the anxiety section of the Hospital Anxiety andDepression Scale were the strongest predictors and alonecould significantly predict if one might develop freezing ofgait in the next 15 months with 82% accuracy.Conclusions: Our results suggest that it is possible toidentify the majority of patients who will develop freez-ing of gait in the following year, potentially allowing tar-geted interventions to delay or possibly even preventthe onset of freezing. VC 2017 International Parkinsonand Movement Disorder Society
Key Words: Parkinson’s disease; freezing of gait;cognition; anxiety; motor; sleep
Freezing of gait (FOG) is a common and complex symp-tom of Parkinson’s disease (PD) with a poorly understoodonset1 and limited options for successful treatment.2
Although approximately 80% of patients ultimatelydevelop FOG,3 predicting who and when is currently notpossible, thus thwarting targeted interventions.
To date, only 2 studies have followed early-stage PDpatients over time to examine the risk factors associ-ated with the development of FOG.4,5 Findings fromthe DATATOP cohort suggested that the risk of devel-oping FOG was significantly higher in more advancedPD patients with predominant gait, balance, speech,and bradykinetic motor symptoms rather than tremor-dominant symptoms.5 A more recent study also identi-fied an akinetic-rigid phenotype as a primary riskfactor for FOG along with lower education, not usinga dopamine agonist, and higher scores on the mood,cognitive disturbance, and sleep disorder domains ofthe Hamilton Depression Rating Scale in a Chinesepopulation when followed over 3 years.4
------------------------------------------------------------*Correspondence to: Kaylena A. Ehgoetz Martens, Parkinson’s DiseaseResearch Clinic, Brain and Mind Centre, University of Sydney, 100 MalletStreet, Sydney, New South Wales 2050; [email protected]
Relevant conflicts of interest/financial disclosures: Kaylena A.Ehgoetz Martens, Emily L. Lukasik, Matthew J. Georgiades, Moran Gilat,Julie M. Hall, Courtney C. Walton, and Simon J.G. Lewis report nodisclosures.
Funding agencies: This work was supported by funding to Forefront, a col-laborative research group dedicated to the study of non-Alzheimer’s diseasedegenerative dementias from the National Health and Medical ResearchCouncil of Australia program granst 1037746 and #1095127. Kaylena A.Ehgoetz Martens is supported by a Parkinson Canada Postdoctoral fellow-ship. Matthew J. Georgiades is supported by the Sydney Medical School RAMoney Postgraduate Research Scholarship in Neuroscience. Moran Gilat issupported by a University of Sydney International Scholarship. Julie M. Hallis supported by a Western Sydney University Postgraduate Research Award.These funding organizations did not play any role in the design and conduc-tor of the study; collection, management, analysis or interpretation of thedata; nor the preparation, review, approval of the manuscript, or decision tosubmit the manuscript for publication.
Received: 27 July 2017; Revised: 29 September 2017; Accepted: 6October 2017
Published online 18 November 2017 in Wiley Online Library(wileyonlinelibrary.com). DOI: 10.1002/mds.27208
R E S E A R C H A R T I C L E
128 Movement Disorders, Vol. 33, No. 1, 2018

Previous work has also reported that freezing isassociated with cognitive dysfunction such as atten-tional set-shifting, conflict resolution, and visuospatialprocessing.6-9 Furthermore, research has shown thatcognition declines at a faster rate in freezers comparedwith nonfreezers over a 2-year period.10 However, lit-tle research has disentangled whether these cognitivedeficits precede the onset of FOG or whether cognitivedecline accelerates after FOG develops. Affective dis-orders have also been linked to FOG,11 with bothanxiety and depression more common in freezers thanin nonfreezers.12 It is still debated whether these affec-tive disturbances precede the onset and possibly con-tribute to the development of FOG or whether anxietyand depression arise as a result of developing FOG.13
Finally, heterogeneity studies have highlighted an over-lap between the nontremor phenotype and rapid eyemovement (REM) sleep behavior disorder (RBD), inwhich patients experience dream enactment.14 It hasbeen postulated that the neuropathology underlyingRBD may lie within the brain stem,15-17 an anatomicalregion that contains the pedunculopontine nucleus, astructure that is believed to play a critical role in gaitfunction.18,19 Work investigating patients with FOGhas demonstrated that although RBD is associatedwith freezing in the early stages of disease, this rela-tionship is not observed in advanced PD.15 Thus, itremains unclear whether RBD might be useful to con-sider when predicting who might develop FOG in thefuture. Together, this body of evidence suggests thatspecific clinical symptoms may represent useful bio-markers for predicting the onset of FOG.
The objective of the current study was to create amore substantive predictive profile for transitionalfreezers by combining potential indicators of FOGfrom previously highlighted motor, cognitive, affec-tive, and sleep domains. The current study assessed 3groups of PD participants: (1) those who remainednonfreezers over time, (2) those who developed FOG,and (3) those who continued to freeze. First, wesought to identify symptoms that preceded FOG, thencreated a logistic regression model to predict thosewho would or would not develop FOG in the nextyear. Based on previous research, we hypothesizedthat in addition to obvious risk factors such as ageand disease duration, a non-tremor-dominant pheno-type with executive dysfunction, greater affective dis-turbance, and RBD might significantly predict theonset of FOG.
Methods
Participants
Longitudinal data were analyzed from 221 Parkin-son’s disease patients who attended the Parkinson’sDisease Research Clinic at the Brain and Mind Centre,
University of Sydney, between 2008 and 2016. Ethicalapproval was obtained from the University of SydneyHuman Research and Ethics Committee and writteninformed consent obtained from each patient. Partici-pant inclusion criteria included: a confirmed diagnosisof PD based on the UK Brain Bank clinical criteria20
by a trained neurologist (S.J.G.L.) and longitudinaldata from at least 2 points approximately 6-24months apart. Patients were identified as freezers ifthey had responded positively on a validated screeninginstrument for identifying freezers, namely, item 3 ofthe Freezing of Gait-Questionnaire (FOG-Q3): “Doyou feel that your feet get glued to the floor whilewalking, making a turn or when trying to initiatewalking (freezing)?”1,21 Based on their response toFOG-Q3 at baseline and follow-up, participants wereclassified into 3 groups: nonfreezers (n 5 88) — thosewho reported 0 at both baseline and follow-up; transi-tional freezers (n 5 41) — those who reported 0 atbaseline and 1 or more at follow-up; and continuingfreezers (n 5 92) — those who reported 1 or more atboth baseline and follow-up. All participants wereassessed on their regular dose of medication.
Motor, Cognitive, Limbic, and Sleep Domains
The Unified Parkinson’s Disease Rating Scale motorsubsection (UPDRS-III) assessed motor symptoms22
with the exclusion of question 3.11 (“Freezing ofgait”).23 A tremor-dominant (TD)/non-TD phenotyperatio was also calculated.24,25 The FOG-Questionnairetotal (FOG-Q total) was used to assess global gait dis-turbance; however, item 3 was removed from thetotal, given that it was used to classify participants.Finally, dopamine dose equivalence (DDE) was alsorecorded when available.
Cognition was evaluated using the following mea-sures: Digit Span (DS) Forwards and Backwards toexamine attention and working memory, the Mini-Mental State Examination (MMSE) to measure globalcognitive functioning, age-scaled Logical Memory(LM-I and LM-II) retention scores to assess verbalmemory, and the Trail-Making Test parts A (TMT-A)and B (TMT-B) to assess psychomotor speed andattentional set-shifting ability, respectively.26,27 Anxi-ety and depression were evaluated using the HospitalAnxiety and Depression Scale subscores (HADS-A andHADS-D).28 Finally, RBD was evaluated using a sub-score of 4 questions (RBDSQ4) pertaining specificallyto movements during REM sleep on the RBD Screen-ing Questionnaire.29
Statistical Analyses
Linear mixed-effects models were employed toexamine all variables longitudinally across all 3groups. Age and disease duration were entered ascovariates. Bonferroni post hoc analyses were carried
A N X I E T Y I S A P R E D I C T O R O F F O G O N S E T
Movement Disorders, Vol. 33, No. 1, 2018 129

out to examine significant group effects and interac-tions (see Table 1). To further unpack the relationshipbetween freezing of gait (frequency and severity) andcognitive and affective factors, correlational analyseswere also carried out on baseline data (see Table 2).
A backward stepwise (likelihood ratio) binary logis-tic regression was used to identify potential baselinepredictors of FOG approximately 1 year before freez-ing onset (continuing freezers were excluded from thisanalysis). The dependent variable for the regressionanalysis was the presence or absence of FOG atfollow-up. The independent variables included: theTD/non-TD ratio, FOG-Q total, UPDRS-III, TMT-Aand TMT-B, DS-Forwards, DS-Backwards, LM-II,HADS-A and HADS-D, RBDSQ4, MMSE, and ageand disease duration at baseline. The stepwise methodwas chosen based on the exploratory nature of thestudy, and a backward model was employed to reducethe type II error rate. The significance level at whichvariables were entered and removed from the modelwas P 5 0.05 and P 5 0.1 respectively. The Homerand Lemeshow test was used to examine the goodnessof fit for the model. In addition, in an effort to maxi-mize clinical relevance, a post hoc reduced-factormodel was constructed on the basis of the most signifi-cant predictive factors from the backward logisticregression described above. The aim of this secondaryanalysis was to understand which tools could beimplemented in clinical settings to detect those at riskfor developing FOG and the success rate of thesepredictors.
Results
Longitudinal Analyses
Table 1 reports mean 6 SD scores between groupsand P values for main effects and interactions fromthe linear mixed-effects models. Table 2 illustrates therelationship between freezing severity and cognitiveand affective symptoms across all participants.
Motor
Overall group effects were found for DDE(F2,05.18 5 7.23, P 5 0.001), UPDRS-III (F2,213.97
5 44.30, P<0.001), and TD/non-TD ratio (F2,216.84
5 10.02, P<0.001). Post hoc analyses revealed thatnonfreezers were receiving a lower dose of dopaminer-gic medication and had lower symptom severity com-pared with the transitional and continuing freezers.Continuing freezers also had significantly worse motorsymptom severity compared with transitional freezers.In addition, nonfreezers had a significantly higher TD/non-TD ratio compared with both freezer groups(indicating that freezers had a greater non-TD score).A main effect of time was also found for UPDRS-III(F1,175.26 5 4.11, P 5 0.044), such that all PD patients
had significantly worse symptom severity at follow-upcompared with baseline.
A significant interaction between group and time wasfound for FOG-Q total (F2,218 5 24.76, P<0.001).Post hoc analyses revealed that at baseline, continuingfreezers had significantly worse gait severity than transi-tional freezers and nonfreezers, but there was no differ-ence between transitional freezers and nonfreezers.However, at follow-up, transitional freezers had signifi-cantly more impaired gait than nonfreezers. Continuingfreezers continued to have the most severe gait impair-ment, which was significantly greater than transitionalfreezers and nonfreezers.
Cognition
Overall group effects were found for MMSE(F2,160.75 5 16.72, P< 0.001), DS-Backwards (F2,136.59
5 6.63, P 5 0.002), and TMT-B-A (F2,128.13 5 6.17,P 5 0.003). Post hoc analyses showed that there were nosignificant differences in performance on the MMSE, DS-Backwards, or TMT-B-A between transitional freezers andnonfreezers. Transitional freezers performed significantlybetter on the MMSE compared with continuing freezersbut were not different from continuing freezers on the DS-Backwards or TMT-B-A. Continuing freezers performedsignificantly worse on the MMSE, DS-Backwards, andTMT-B-A compared with nonfreezers. A main effect oftime was found for MMSE (F1,123.17 5 4.88, P<0.001),LM-I and LMT-II (F1,120.36 5 4.39, P 5 0.038; F1,118.69
5 14.54, P<0.001), and TMT-B-A (F1,116.75 5 4.05,P 5 0.046), which showed that although MMSE andTMT-B-A scores got worse over time, scores for LM-I andLMT-II improved.
A significant interaction between group and timewas found for TMT-B (F2,119.21 5 3.08, P 5 0.050).Post hoc analysis revealed that at baseline transitionalfreezers were not different from nonfreezers or con-tinuing freezers. However, by follow-up, transitionalfreezers performed significantly worse on the TMT-Bcompared with nonfreezers, and continuing freezerswere significantly worse compared with transitionalfreezers and nonfreezers. Thus, both freezing groupsshowed a significant increase in TMT-B scores atfollow-up compared with baseline, whereas the non-freezers did not change (Figure 1).
Mood
A group effect was found for HADS-A(F2.209.99 5 8.78, P< 0.001; Figure 2). Post hoc analysesrevealed that both transitional freezers and continuingfreezers had significantly greater anxiety compared withnonfreezers. A main effect of time was found forHADS-A (F1,198.42 5 56.27, P<0.001), which revealedthat anxiety worsened over time across all groups.
There was a significant interaction between groupand time for HADS-D (F2,200.30 5 3.61, P 5 0.029).
E H G O E T Z M A R T E N S E T A L
130 Movement Disorders, Vol. 33, No. 1, 2018

TA
BL
E1.
Dem
og
rap
hic
,clin
icaland
neuro
psy
cho
log
icalchara
cte
rist
ics
at
base
line
and
follo
w-u
p
Gro
up
No
nfr
eeze
rsTr
ansi
tionalfr
eeze
rsC
ontin
uin
gfr
eeze
rs
Tim
e1
21
21
2G
roup
eff
ect
Tim
eeff
ect
Gro
up
3tim
ein
tera
ctio
n
n88
4192
Timebetweenbaselineand
follow-up(months)
14.8
64.6
156
4.6
12.8
64.6
Male/female
52/36
26/15
56/36
Age(years)
65.37
69.84
70.75
69.39
70.24
610.85
P5
0.002a
Diseaseduration(years)
3.16
64.32
5.97
64.14
9.66
67.50
P<0.001a
DDE
504.77
6355.21
584.49
6293.34
832.15
6452.28
992.33
6422.81
909.92
6549.31
804.57
6478.61
P<0.001a
p50.49
P50.271
MDS
-UPD
RSIII
24.51
613.62
25.51
614.40
35.14
617.07
38.42
615.33
48.64
620.68
49.81
621.06
P<0.001a
p50.044a
P50.539
TD/non-TDratio
0.80
60.82
0.65
60.54
0.51
60.36
0.48
60.39
0.45
60.43
0.37
60.32
P<0.001a
p50.055
P50.624
FOG-Qtotal
1.41
61.37
1.68
61.60
2.68
61.74
6.17
62.39
10.23
64.19
10.79
64.02
P<0.001a
P<0.001a
P<0.001a
MMSE
28.60
61.78
28.65
62.17
28.45
61.93
27.39
62.92
26.58
63.50
24.89
65.53
P<0.001a
P5
0.029a
P5
0.053
DigitS
panForwards
10.18
62.51
10.59
62.49
10.42
61.90
9.96
61.81
9.41
62.33
9.70
62.21
P5
0.106
P5
0.69
P5
0.282
DigitS
panBackwards
6.98
61.76
7.19
61.94
6.30
61.69
6.40
62.18
5.92
62.03
5.97
61.72
P5
0.002a
P5
0.493
P5
0.861
TMTA(seconds)
39.75
619.36
38.40
633.75
43.03
620.73
46.18
623.65
47.90
620.74
50.43
629.15
P5
0.084
P5
0.424
P5
0.591
TMTB(seconds)
96.99
677.25
87.70
649.39
117.93
665.42
139.54
6102.67
131.03
681.06
149.50
693.69
P5
0.005a
P5
0.014a
P5
0.050
a
TMTB-A(seconds)
56.94
662.38
49.67
634.18
74.90
653.98
93.57
685.08
83.64
668.77
99.08
671.77
P5
0.003a
P5
0.046a
P5
0.102
Logicalm
emory1
9.79
62.74
10.62
63.27
10.54
63.83
10.04
63.51
8.13
63.25
9.81
63.40
P5
0.007a
P5
0.038a
P5
0.07
Logicalm
emory2
10.32
63.33
10.92
63.18
10.81
63.45
11.60
63.20
9.03
63.08
10.81
63.31
P5
0.159
P<0.001a
P5
0.426
HADS
-A1.18
62.31
3.62
62.93
3.65
63.08
5.18
64.68
2.94
63.89
5.38
64.19
P<0.001a
P<0.001a
P5
0.526
HADS
-D4.10
63.80
3.54
63.45
6.14
64.80
5.31
64.14
8.96
65.48
6.61
64.20
P<0.001a
P<0.001a
P5
0.029a
RBDS
Q41.64
62.02
1.84
62.02
2.23
62.35
2.73
62.29
2.69
62.30
2.63
62.12
P5
0.006a
P5
0.077
P5
0.279
Mean
sco
re6
stand
ard
devi
atio
nare
pre
sente
d.
aS
ignifi
cant
eff
ects
.N
F,no
nfr
eeze
r;T
F,tr
ansi
tional
freeze
r;C
F,co
ntin
uin
gfr
eeze
r;D
DE
,d
op
am
ine
do
seeq
uiv
ale
nce;
MD
S-U
PD
RS
,M
ove
ment
Dis
ord
er
So
cie
ty-U
nifi
ed
Park
inso
n’s
Dis
ease
Ratin
gS
cale
;T
D,
trem
or
do
min
ant;
FO
G-Q
,F
reezi
ng
of
Gait
Quest
ionnaire;
MM
SE
,M
ini-
Menta
lS
tate
Exa
min
atio
n;
TM
T(A
and
B,
B-A
),Tr
ail-
Maki
ng
Test
part
sA
and
B,
part
B-p
art
A;
HA
DS
(A,
D),
Ho
spita
lA
nxi
ety
and
Dep
ress
ion
Scale
(Anxi
ety
,D
ep
ress
ion
sub
sco
res;
RB
DS
QR
EM
Sle
ep
Behavi
or
Dis
ord
er
Quest
ionnaire.
A N X I E T Y I S A P R E D I C T O R O F F O G O N S E T
Movement Disorders, Vol. 33, No. 1, 2018 131

Post hoc analyses showed that continuing freezers hadsignificantly greater depression than transitional freez-ers and nonfreezers at baseline, but were not differentfrom transitional freezers at follow-up. Continuingfreezers also reported significantly lower depressionscores at follow-up compared with baseline.
Sleep
Overall group effects were found for RBDSQ4(F2,215.50 5 5.21, P 5 0.006). Post hoc analysesrevealed that transitional freezers were not differentfrom nonfreezers or continuing freezers; however, con-tinuing freezers had significantly higher RBDSQ4scores than nonfreezers.
Predictors of Freezing
A model (see Table 3) was found to be a good fitfor the data (v2
ð7Þ5 5.91, P 5 0.55), which included 75participants (52 nonfreezers and 23 transitional freez-ers). The following predictors were included in thefinal model: age (B 5 0.11, P 5 0.054), disease dura-tion (B 5 0.15, P 5 0.091), FOG-Q total (B 5 0.85,P 5 0.01), TD/non-TD ratio (B 5 -1.60, P 5 0.085),TMT-B (B 5 -0.02, P 5 0.055), HADS-A (B 5 0.92,P 5 0.001), and HADS-D (B 5 -0.32, P 5 0.067). Theoverall predictive success of the model was 84%, andit correctly classified 69.6% of transitional freezersand 90.4% of nonfreezers.
Given that FOG-Q total and HADS-A were thestrongest predictors in the above model, we examinedthe predictive power of using only these factors in aneffort to maximize clinical relevance. A model wasfound to be a good fit for the data (v2
ð8Þ5 6.23,P 5 0.62), which included 117 participants (80 non-freezers and 37 transitional freezers). The model hadan overall predictive success of 82.1%, which cor-rectly classified 64.9% of patients who developedFOG and 90% of patients who remained nonfreezers.The following equation represents the probability (p)of developing FOG in the following year based on thisrestricted model:
Logðp=1 2 pÞ 5 22:383 1 0:457 ðFOG2Q totalÞ 1 0:305 ðHADS2AÞ
Discussion
The overall aim of this study was to identify symp-toms that preceded FOG and to create a predictivemodel that could be used to identify those at risk fordeveloping freezing in the following year. Althoughthere were many expected differences at baselinebetween freezers and nonfreezers,15,24 there were afew measures that differentiated between transitionalfreezers and nonfreezers at baseline (ie, 1 year prior tothe onset of FOG).
FIG. 1. Transitional freezers showed greater deficits in set-shiftingcompared with nonfreezers at follow-up but not baseline.
FIG. 2. Transitional freezers and continuing freezers had greater anxi-ety compared with nonfreezers at baseline and follow-up.
E H G O E T Z M A R T E N S E T A L
132 Movement Disorders, Vol. 33, No. 1, 2018

Symptoms That Precede FOG
Motor aspects such as the DDE, UPDRS-III, and anon-TD phenotype were found to be greater in transi-tional freezers (and continuing freezers) than in non-freezers at baseline and follow-up. These findings areconsistent with previous research showing that freezershave worse motor symptoms, especially nontremorsymptoms.4,5,15,24,30 Such patients require greaterdopaminergic therapy to manage these more severesymptoms.31,32 Anxiety was also greater in transi-tional freezers (and continuing freezers) comparedwith nonfreezers regardless of time, which providesstrong evidence that anxiety precedes the onset ofFOG. These results are in support of previous worksuggesting that anxiety is greater in freezers than innonfreezers12 and might be an important contributorto the underlying mechanism of FOG.33 However,depression was not significantly different in transi-tional freezers than nonfreezers at baseline or follow-up and therefore may not predict FOG onset. Never-theless, continuing freezers had worse depression thantransitional and nonfreezers at baseline, which is simi-lar to previous accounts.5,12 Interestingly, depression
in continuing freezers was reduced at follow-up com-pared with baseline and no longer differed from tran-sitional freezers, suggesting that this symptom wasbetter managed by follow-up.
Surprisingly, there were no differences at baselinebetween transitional freezers and nonfreezers acrossthe array of cognitive or sleep outcomes. Rather, oncetransitional freezers had developed FOG by follow-up,attentional set-shifting impairments (TMT-B) werefound compared with nonfreezers. Furthermore, onlytransitional freezers and continuing freezers (but notthe nonfreezer group) showed a significant decline inset-shifting capabilities over time. Furthermore, theseverity of FOG measured by the total score on theFOG-Q (including item 3) was associated with wors-ened cognition (regardless of neuropsychologicalassessment), greater affective symptom severity, andgreater RBD score. Taken together, these findingsmight suggest that cognitive dysfunction and sleep dis-turbances either become gradually more impaired asFOG onset approaches as a function of disease pro-gression or that FOG onset and impaired cognitionand sleep occur within a similar time frame because ofshared neurobiological underpinnings such as choliner-gic denervation within the pedunculopontine nucleus,for example.
Predicting Freezing of Gait
This study demonstrates that by examining specificsymptoms (ie, TD/non-TD ratio, FOG-Q total, TMT-B, anxiety, depression) along with age and diseaseduration, we could successfully predict with 84%accuracy those PD patients who transitioned tobecome freezers and those who did not in the follow-ing year. The current findings are aligned with previ-ous work,4,5 which also showed that more severe gaitdisability, having a non-tremor-dominant phenotype,and anxiety and depression were significant FOG riskfactors. Notably, the only domain that was not pre-sent in this model was sleep. Because RBD is generallyconsidered a prodromal marker of PD, it may not bea useful indicator of later-stage symptoms such asfreezing despite biological overlap between RBD andFOG.
The ability to predict FOG has tremendous implica-tions for patients, caregivers, and clinicians, because itprovides a unique opportunity to implement earlytherapeutic interventions to delay or even prevent theonset of FOG. This is likely to prolong patients’mobility and quality of life. For this reason, we alsoevaluated a more restricted model to predict FOGincluding simply 2 questionnaires that could beadministered by a nonspecialist. Although the predic-tive power was not as strong as the full modeldescribed above, by using only the FOG-Q total andthe HADS-A score, we could predict whether one
TABLE 2. Relationships between freezing severity andcognitive, affective, and sleep symptoms
FOGQ-Total (severity)
MMSE 20.409b
TMT-A 20.288b
TMT-B 20.218a
Digit Span Forward 20.232b
Digit Span Backwards 20.245b
LM-I -0.305b
LM-II 20.272b
HADS-A 0.206b
HADS-D 0.499b
RBDSQ4 0.185b
aP< 0.05.bP< 0.01.
TABLE 3. Motor and nonmotor predictors of freezingof gait
B (SE) Odds ratio 95% CI
Age 0.11 (0.06) 1.11 0.10-1.24Disease duration 0.15 (0.09) 1.16 0.98-1.39TD/non-TD ratio 21.60 (0.93) 0.20 0.03-1.24FOG-Q total 0.85a (0.33) 2.34 1.23-4.45TMT B raw score 20.02 (0.01) 0.99 0.97-1.00HADS-A 0.92a (0.28) 2.50 1.46-4.30HADS-D 20.32 (0.17) 0.73 0.52-1.02Constant 27.94a (3.58)
R2 5 0.45 (Cox & Snell), R2 5 0.64 (Nagelkerke). Homer and LemeshowGoodness of fit v2
ð7Þ5 5.83, P 5 0.56.aWald statistic P< 0.05.TD, tremor dominant; FOG-Q, Freezing of Gait Questionnaire; TMT, Trail-Making Test; HADS-A and -D, Hospital Anxiety and Depression Scale anxi-ety and depression subscores.
A N X I E T Y I S A P R E D I C T O R O F F O G O N S E T
Movement Disorders, Vol. 33, No. 1, 2018 133

might develop FOG in the next year with 82.1% accu-racy. It should be noted that this model is much betterat classifying those who will not develop FOG (90%accuracy) than those who will (65% accuracy). None-theless, it may be a useful screening tool to identifyindividuals at risk who may need further assessment,such as comprehensive testing to improve predictivepower, and could also identify candidates at risk oreligible for early intervention studies. Because 80% ofpatients will develop PD eventually, it is likely that a1-year assessment is not enough, and additional fac-tors that were not measured here may also contributeto these prediction models. Thus, future researchshould consider flagging and following at-risk individ-uals for a longer period, beyond the time frame cap-tured in this study, to determine whether thepredictive accuracy improves over time.
Limitations and Clinical Implications
A limitation of this study was the lack of DDE datafor numerous participants, which led us to excludeDDE from the logistic regression. Because DDE is typ-ically associated with disease duration and symptomseverity, it might be expected to contribute in a similarway to these other variables that were included in themodel. Alternatively, given that dopaminergic dysfunc-tion may impact the degree to which network loopscross over, dopamine agonists may play a crucial rolein the development of FOG and may be a predictivefactor worth exploring in future research. A secondlimitation that should be noted is that a subjectivequestionnaire was used to classify freezers, rather thanan objective measure of freezing, which may be supe-rior at detecting freezers and minimizing misclassifica-tion. In the current study, a video of FOG was notpresented to participants prior to answering the FOG-Q, although many of the participants had clinicallyconfirmed freezing and continued to report FOG insubsequent visits. Nonetheless, future work shouldaim to use a video of FOG to minimize classificationerrors and if possible clinically confirm all participantswho report FOG.
Despite these limitations, the findings from thisstudy provide clinicians with means to better predictthe likelihood that a patient will develop FOG. Futurestudies should focus on developing appropriate inter-ventions, with the goal of delaying or even halting theonset of FOG for those most at risk of transitioning.Current treatment options for FOG are very limitedand hindered by inconsistent results2; thus, preventivemeasures may prove more effective. Cognitive training(CT) has been speculated to be a promising treatmentfor FOG.34 Given that CT is targeted at improvingexecutive functions in PD,35 it aims to promote theactivation of compensatory networks and alleviate thecognitive interference that precedes FOG.34
Alternatively, cholinesterase inhibitors might be usedto target cognitive dysfunction preceding FOG onset.Research has linked FOG and neocortical cholinergicdenervation36 and has suggested cholinergic denerva-tion as a potential mechanism underlying depression-modulated cognitive impairment, which can exacer-bate the cognitive symptoms found in PD.37 Impor-tantly, a recent review concluded that cholinesteraseinhibitors can enhance cognitive function in PDpatients with cognitive impairment38 and have alsobeen shown to be promising for reducing falls inPD.39 Thus, further trials examining cholinesteraseinhibitors are a much-needed undertaking.40 Finally,given that anxiety is a strong predictor of FOG onset,anxiety should be targeted as part of preventive effortsfor potential freezers, especially because to date, norandomized, controlled trials have been undertaken toexplore how anxiety management could delay thedevelopment or progression of FOG.
Conclusion
A careful assessment of selected symptoms can accu-rately predict the onset of FOG over a 15-month inter-val. These findings will hopefully allow future targetedtreatments for FOG in PD, which would greatlyenhance the quality of life for a substantial proportionof PD patients.
Acknowledgments: We are grateful to the research participantsinvolved in this work.
References1. Giladi N, Treves TA, Simon ES, et al. Freezing of gait in patients
with advanced Parkinson’s disease. J Neural Transm 2001;108:53-61.
2. Nonnekes J, Snijders AH, Nutt JG, et al. Freezing of gait: a practi-cal approach to management. Lancet Neurol 2015;14:768-778.
3. Macht M, Kaussner Y, M€oller JC, et al. Predictors of freezing inParkinson’s disease: a survey of 6,620 patients. Mov Disord 2007;22:953-956.
4. Zhang H, Yin X, Ouyang Z, et al. A prospective study of freezingof gait with early Parkinson disease in Chinese patients. Medicine(Baltimore) 2016;95:e4056.
5. Giladi N, McDermott MP, Fahn S, et al. Freezing of gait in PD:prospective assessment in the DATATOP cohort. Neurology 2001;56:1712-1721.
6. Naismith SL, Shine JM, Lewis SJG. The specific contributions ofset-shifting to freezing of gait in Parkinson’s disease. Mov Disord2010;25:1000-1004.
7. Amboni M, Cozzolino A, Longo K, et al. Freezing of gait andexecutive functions in patients with Parkinson’s disease. Mov Dis-ord 2008;23:395-400.
8. Walton CC, Shine JM, Mowszowski L, et al. Impaired cognitivecontrol in Parkinson’s disease patients with freezing of gait inresponse to cognitive load. J Neural Transm 2015;122:653-660.
9. Walton CC, O’Callaghan C, Hall JM, et al. Antisaccade errorsreveal cognitive control deficits in Parkinson’s disease with freezingof gait. J Neurol 2015;262:2745-2754.
10. Amboni M, Barone P, Picillo M, et al. A two-year follow-up studyof executive dysfunctions in parkinsonian patients with freezing ofgait at on-state. Mov Disord 2010;25:800-802.
E H G O E T Z M A R T E N S E T A L
134 Movement Disorders, Vol. 33, No. 1, 2018

11. Lieberman A. Are freezing of gait (FOG) and panic related?J Neurol Sci 2006;248:219-222.
12. Ehgoetz Martens KA, Hall JM, Gilat M, et al. Anxiety is associ-ated with freezing of gait and attentional set-shifting in Parkinson’sdisease: a new perspective for early intervention. Gait Posture2016;49:431-436.
13. Ehgoetz Martens KA, Ellard CG, Almeida QJ. Evaluating the linkbetween dopaminergic treatment, gait impairment, and anxiety inParkinson’s disease. Mov Disord Clin Pract 2016;3(4):389-394.
14. Kumru H, Santamaria J, Tolosa E, et al. Relation between subtypeof Parkinson’s disease and REM sleep behavior disorder. SleepMed 2007;8:779-783.
15. Hall JM, Shine JM, O’Callaghan C, et al. Freezing of gait and itsassociations in the early and advanced clinical motor stages of Par-kinson’s disease: a cross-sectional study. J Parkinsons Dis 2015;5(4):881-891.
16. Postuma RB, Gagnon J-F, Vendette M, et al. Manifestations ofParkinson disease differ in association with REM sleep behaviordisorder. Mov Disord 2008;23:1665-1672.
17. Romenets SR, Gagnon J-F, Latreille V, et al. Rapid eye movementsleep behavior disorder and subtypes of Parkinson’s disease. MovDisord 2012;27:996-1003.
18. Lau B, Welter ML, Belaid H, et al. The integrative role of thepedunculopontine nucleus in human gait. Brain 2015;138:1284-1296.
19. Windels F, Thevathasan W, Silburn P, et al. Where and what isthe PPN and what is its role in locomotion? Brain 2015;128:1133-1134.
20. Hughes AJ, Daniel SE, Kilford L, et al. Accuracy of clinical diag-nosis of idiopathic Parkinson’s disease: a clinico-pathologicalstudy of 100 cases. J Neurol Neurosurg Psychiatry 1992;55:181-184.
21. Shine JM, Moore ST, Bolitho SJ, et al. Assessing the utility ofFreezing of Gait Questionnaires in Parkinson’s Disease. Parkinson-ism Relat Disord 2012;18:25-29.
22. Goetz CG, Tilley BC, Shaftman SR, et al. Movement DisorderSociety-sponsored revision of the Unified Parkinson’s Disease Rat-ing Scale (MDS-UPDRS): Scale presentation and clinimetric testingresults. Mov Disord 2008;23:2129-2170.
23. Georgiades MJ, Gilat M, Ehgoetz Martens KA, et al. Investigatingmotor initiation and inhibition deficits in patients with Parkinson’sdisease and freezing of gait using a virtual reality paradigm. Neu-roscience 2016;337:153-162.
24. Hall JM, Shine JM, Walton CC, et al. Early phenotypic differencesbetween Parkinson’s disease patients with and without freezing ofgait. Park Relat Disord 2014;20:604-607.
25. Lewis SJG, Foltynie T, Blackwell AD, et al. Heterogeneity of Par-kinson’s disease in the early clinical stages using a data drivenapproach. J Neurol Neurosurg Psychiatry 2005;76:343-348.
26. Reitan R, Wolfson D. The Halstead-Reitan neuropsychological testbattery: theory and clinical interpretation. 1985.
27. Wechsler D. Wechsler Memory Scale (WMS-III). San Antonio, TX:Psychological Corporation: 1997.
28. Zigmond AS, Snaith RP. The Hospital Anxiety and DepressionScale. Acta Psychiatr Scand 1983;67:361-370.
29. Bolitho SJ, Naismith SL, Terpening Z, et al. Investigating rapid eyemovement sleep without atonia in Parkinson’s disease using therapid eye movement sleep behavior disorder screening question-naire. Mov Disord 2014;29:736-742.
30. Factor SA, Steenland NK, Higgins DS, et al. Postural instability/gaitdisturbance in Parkinson’s disease has distinct subtypes: an explor-atory analysis. J Neurol Neurosurg Psychiatry 2011;82:564-568.
31. Vervoort G, Bengevoord A, Strouwen C, et al. Progression of posturalcontrol and gait deficits in Parkinson’s disease and freezing of gait: Alongitudinal study. Parkinsonism Relat Disord 2016;28:73-79.
32. Gray P, Hildebrand K. Fall risk factors in Parkinson’s disease.J Neurosci Nurs 2000;32:222-228.
33. Ehgoetz Martens KA, Ellard CG, Almeida QJ, et al. Does anxiety causefreezing of gait in Parkinson’s disease? PLoS One 2014;9:e106561.
34. Walton CC, Shine JM, Mowszowski L, et al. Freezing of gait inParkinson’s disease: current treatments and the potential role forcognitive training. Restor Neurol Neurosci 2014;32:411-422.
35. Leung IHK, Walton CC, Hallock H, et al. Cognitive training inParkinson disease. A systematic review and meta-analysis. Neurol-ogy 2015;85:1843-1851.
36. Bohnen NI, Frey KA, Studenski S, et al. Extra-nigral pathological condi-tions are common in Parkinson’s disease with freezing of gait: An in vivopositron emission tomography study. Mov Disord 2014;29:1118–24.
37. Bohnen NI, Albin RL. The cholinergic system and Parkinson dis-ease. Behav Brain Res 2011;221:564-573.
38. Wang H-F, Yu J-T, Tang S-W, et al. Efficacy and safety of cholines-terase inhibitors and memantine in cognitive impairment in Parkin-son’s disease, Parkinson’s disease dementia, and dementia withLewy bodies: systematic review with meta-analysis and trial sequen-tial analysis. J Neurol Neurosurg Psychiatry 2015;86:135-143.
39. Chung KA, Lobb BM, Nutt JG, et al. Effects of a central cholines-terase inhibitor on reducing falls in Parkinson disease. Neurology2010;75:1263-1269.
40. Fling BW, Cohen RG, Mancini M, et al. Functional reorganizationof the locomotor network in Parkinson patients with freezing ofgait. PLoS One 2014;9:e100291.
A N X I E T Y I S A P R E D I C T O R O F F O G O N S E T
Movement Disorders, Vol. 33, No. 1, 2018 135

APPENDIX(10(
Ehgoetz Martens, K. A., Hall, J. M., Georgiades, M, J., Gilat, M., Walton, C. C.,
Matar, E., Lewis, S. J. G., Shine, J. M. The functional network signature of
heterogeneity in freezing of gait. Brain, 2018
!

The functional network signature ofheterogeneity in freezing of gaitKaylena A. Ehgoetz Martens,1,2 Julie M. Hall,1,2,3 Matthew J. Georgiades,1,2 Moran Gilat,1,2
Courtney C. Walton,1,2 Elie Matar,1,2 Simon J. G. Lewis1,2 and James M. Shine1,2
Freezing of gait is a complex, heterogeneous, and highly variable phenomenon whose pathophysiology and neural signature
remains enigmatic. Evidence suggests that freezing is associated with impairments across cognitive, motor and affective domains;
however, most research to date has focused on investigating one axis of freezing of gait in isolation. This has led to inconsistent
findings and a range of different pathophysiological models of freezing of gait, due in large part to the tendency for studies to
investigate freezing of gait as a homogeneous entity. To investigate the neural mechanisms of this heterogeneity, we used an
established virtual reality paradigm to elicit freezing behaviour in 41 Parkinson’s disease patients with freezing of gait and
examined individual differences in the component processes (i.e. cognitive, motor and affective function) that underlie freezing
of gait in conjunction with task-based functional MRI. First, we combined three unique components of the freezing phenotype:
impaired set-shifting ability, step time variability, and self-reported anxiety and depression in a principal components analysis to
estimate the severity of freezing behaviour with a multivariate approach. By combining these measures, we were then able to
interrogate the pattern of task-based functional connectivity associated with freezing (compared to normal foot tapping) in a sub-
cohort of 20 participants who experienced sufficient amounts of freezing during task functional MRI. Specifically, we used the first
principal component from our behavioural analysis to classify patterns of functional connectivity into those that were associated
with: (i) increased severity; (ii) increased compensation; or (iii) those that were independent of freezing severity. Coupling between
the cognitive and limbic networks was associated with ‘worse freezing severity’, whereas anti-coupling between the putamen and
the cognitive and limbic networks was related to ‘increased compensation’. Additionally, anti-coupling between cognitive cortical
regions and the caudate nucleus were ‘independent of freezing severity’ and thus may represent common neural underpinnings of
freezing that are unaffected by heterogenous factors. Finally, we related these connectivity patterns to each of the individual
components (cognitive, motor, affective) in turn, thus exposing latent heterogeneity in the freezing phenotype, while also identifying
critical functional network signatures that may represent potential targets for novel therapeutic intervention. In conclusion, our
findings provide confirmatory evidence for systems-level impairments in the pathophysiology of freezing of gait and further
advance our understanding of the whole-brain deficits that mediate symptom expression in Parkinson’s disease.
1 Parkinson’s Disease Research Clinic, Brain and Mind Centre, University of Sydney, Australia2 ForeFront, Brain and Mind Centre, University of Sydney, Australia3 School of Social Sciences and Psychology, Western Sydney University, Australia
Correspondence to: Kaylena A. Ehgoetz MartensBrain and Mind Centre100 Mallet Street Camperdown NSW 2050 AustraliaE-mail: [email protected]
Keywords: freezing of gait; Parkinson’s disease; functional connectivity; functional magnetic resonance imaging; virtual reality
Abbreviation: FCI = Freezing Component Index; HADS = Hospital Anxiety and Depression Scale; TMT = Trail Making Test
doi:10.1093/brain/awy019 BRAIN 2018: Page 1 of 16 | 1
Received October 14, 2017. Revised December 6, 2017. Accepted December 12, 2017.! The Author(s) (2018). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved.For Permissions, please email: [email protected]
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

IntroductionFreezing of gait is a complex and poorly understood symp-tom of Parkinson’s disease that typically presents as a par-oxysmal and transient episode in which patients reportfeeling as though their feet are ‘glued’ to the floor(Schaafsma et al., 2003). Management of the condition re-mains difficult, partly because of the heterogeneous natureof the disorder. For instance, freezing of gait can be pro-voked in a wide variety of situations (e.g. when distracted,turning or when feeling anxious) (Nutt et al., 2011). Thesecommon ‘triggers’ can be broadly classified into impair-ments within three behavioural categories: cognitive,motor and limbic.
Impairments in cognitive flexibility, namely attentional set-shifting deficits are a widely recognized ‘cognitive signature’associated with freezing of gait (Amboni et al., 2008; Hallett,2008; Shine et al., 2013d; Szeto et al., 2015). For instance,patients with freezing of gait typically perform significantlyworse on part B of the Trail Making Test (TMT)(Hall et al., 2014, 2015; Beck et al., 2015), and intra-extradimensional set shifting test (Stefanova et al., 2014).Additionally, patients with freezing of gait show greaterdual-task interference (Peterson et al., 2015) and commitmore errors (Pieruccini-Faria et al., 2014) when asked toperform a secondary cognitive task while walking. Thesebehavioural deficits align with the observation that dividingattention, or switching motor programmes often provokesfreezing of gait (Schaafsma et al., 2003; Nutt et al., 2011;Beck et al., 2015), whereas dual motor-cognitive interventionscan improve clinical freezing (Killane et al., 2015).
The motor signature of freezing is generally characterizedby impairments in gait automaticity, which is normallyoperationalized by an increase in step-to-step variability(Hausdorff et al., 2003), step length variability (Thevathasanet al., 2012; Barbe et al., 2014) and/or step timevariability (Pieruccini-Faria et al., 2015; Weiss et al., 2015)during regular walking, even in the absence of freezing.Additionally, step time variability in patients with freezing ofgait is also exacerbated during split belt treadmill walking(Nanhoe-Mahabier et al., 2013), while approaching narrowdoorways (Almeida and Lebold, 2010; Ehgoetz Martenset al., 2013b; Silveira et al., 2015), walking in threateningsituations (Ehgoetz Martens et al., 2013a, 2014b), duringsharp turns (Bhatt et al., 2013), and while dual-tasking(Pieruccini-Faria et al., 2014; Beck et al., 2015). In fact, vari-ability in stepping has even been shown to be worse in pa-tients with freezing of gait while performing a stationaryvirtual reality tapping task (Gilat et al., 2013) and repetitivestepping in place task (Nantel et al., 2011).
Finally, emotional disturbances, such as anxiety, havebeen suggested to represent a key limbic signature of freez-ing (Giladi and Hausdorff, 2006). Panic attacks (Vazquezet al., 1993; Lieberman, 2006) and physiological heart ratechanges (Maidan et al., 2011) have been temporally linkedto the onset of freezing of gait episodes, and threatening or
stressful situations (e.g. walking over an elevated virtualplank) can also reliably trigger freezing of gait (Bloemet al., 2004; Ehgoetz Martens et al., 2014b). Recent re-search has also shown that patients with freezing of gaithave worse symptoms of anxiety and depression comparedto those without freezing of gait (Burn et al., 2012; EhgoetzMartens et al., 2016c), and greater anxiety is also asso-ciated with worse freezing severity (Ehgoetz Martenset al., 2016b).
Several models have been proposed to explain the patho-physiology of freezing (Nieuwboer and Giladi, 2013).However, most mechanistic explanations have focused onimpairments in one component domain (e.g. cognitive ormotor), and thus essentially underestimate the role ofsystem-level impairments in freezing. In contrast, the‘cross-talk’ model (Lewis and Barker, 2009) explicitly em-braces the interactions between the component dimensionsof freezing. Specifically, the model suggests that freezing ofgait is triggered by episodic discordant ‘cross-talk’ betweencompeting yet complimentary frontostriatal circuits thatspan motor, cognitive and limbic cortical areas. This‘cross-talk’ is proposed to overload the information process-ing capacity within the dopamine-depleted striatum, thusproducing momentary synchronous firing in the outputnuclei of the basal ganglia, which in turn would lead toincreased inhibition in brainstem locomotor areas, and con-sequently freezing of gait (Lewis and Shine, 2016). In thismodel, the corticostriatal architecture is critical for balan-cing integration and segregation across disparate motor,cognitive, and limbic circuits, which is required for optimalinformation processing and hence guided the regions ofinterest selected for this study. It can be proposed that thecorticostriatal dysfunction seen in Parkinson’s disease maylead to interference and over-integration between thesenormally segregated networks.
Functional imaging studies have provided broad empiricalsupport for the cross-talk model. Specifically, patients withParkinson’s disease and freezing have been shown to demon-strate reduced frontal and parietal cortical activation butincreased subcortical activation (mesencephalic locomotorregion) during imagined walking (Snijders et al., 2011),while the opposite pattern (i.e. increased frontal and parietalactivation but reduced subcortical activation) is observedduring freezing episodes (Shine et al., 2013b). Others havelooked beyond the role of individual regions, and examinedthe interplay between regions, as well as large-scale networks.For example, reduced functional coupling in fronto-parietalnetworks at rest was identified in patients with freezing ofgait compared to those patients without (Tessitore et al.,2012b), and reduced coupling between the cognitive controlnetwork and the basal ganglia network was shown to bespecifically associated with episodes of freezing behaviourelicited during functional MRI scanning (Shine et al.,2013c). These studies suggest that freezing of gait canbe thought of as occurring due to dynamic dysfunctionalinteractions across normally coordinated neural networks(Fasano et al., 2015). However, recent reviews have
2 | BRAIN 2018: Page 2 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

promoted the need to characterize the heterogeneity andcomplexity of freezing of gait, rather than to study contribu-tors to freezing in isolation (Fasano et al., 2015; Snijderset al., 2016). Therefore, a crucial next step towards under-standing the freezing phenomenon is to examine the neuralsignature of freezing across multiple levels of neural networkorganization.
Here, we used a task-based functional MRI approachin combination with a validated virtual reality task(Gilat et al., 2013) to investigate the neural signature ofthe heterogeneity of freezing of gait in Parkinson’s disease.We aimed to clarify: (i) how specific regions are coordi-nated during freezing compared to normal foot tapping(i.e. a proxy for normal walking); (ii) how the freezingsignature varies as a function of the first principal compo-nent (which relates to ‘freezing severity’); and (iii) whetherdifferent behavioural signatures of freezing (i.e. cognitive,motor, limbic) are associated with unique neural underpin-nings. Based on previous research (Shine et al., 2013c), wehypothesized that freezing epsiodes would be characterizedby abnormal patterns of functional connectivity within andbetween anatomical cortico-striatal circuitry. This wasthought to reflect a loss of specificity between the cortexand the striatum, and hence to increase the vulnerability ofthe system to ‘overload’. Given the heterogeneity in freez-ing, we further hypothesized that the freezing networksignature would be uniquely associated with individualdifferences in the cognitive, motor, limbic componentprocesses of freezing.
Materials and methods
ParticipantsForty-one patients with Parkinson’s disease and freezing of gaitwere studied at the Brain and Mind Centre at the University ofSydney. Inclusion criteria included: (i) a diagnosis of Parkinson’sdisease in accordance with the United Kingdom ParkinsonDisease Society Brain Bank Diagnostic Criteria; (ii) a self-re-ported score of 51 on question 3 of the Freezing of GaitQuestionnaire (FOGQ3): ‘do you feel that your feet get gluedto the floor while walking, making a turn or when trying toinitiate walking (freezing)?’; (iii) clinically-evident freezing ofgait, confirmed visually by a neurologist (S.J.G.L.); and (iv) thecompletion of the virtual gait paradigm in the MRI scanner inthe ‘OFF’ state (i.e. after a minimum of 12 h withdrawal fromtheir regular dopaminergic medication). Exclusion criteriaincluded: (i) any additional neurological comorbidities (e.g. his-tory or stroke or head injury); and (ii) any pathological lesionsor abnormalities that were identified by an experienced radiolo-gist from the participants’ structural high-resolution T1-weightedimage. Because of the nature of our study (i.e. to investigate aclinical phenomenon that only occurs in a particular group ofindividuals), we did not include a traditional ‘control’ group. Thecurrent study received ethical approval from the University ofSydney Human Research Ethics Committee and was conductedin accordance with the Declaration of Helsinki. All participantsprovided written informed consent.
Data collection and analysis
Clinical and neuropsychological assessment
Participants’ motor symptom severity was assessed with PartIII of the Movement Disorders Society Unified Parkinson’sDisease Rating Scale. Global cognition was assessed with theMini-Mental State Examination, and attentional set-shiftingwas assessed with the TMT, parts A and B. Finally, affectivedisturbance was assessed using the Hospital Anxiety andDepression Scale (HADS).
Virtual gait task
Participants lay supine inside the MRI scanner with a mirrorthat was mounted to the head coil, which enabled participantsto have a clear view of the screen where the virtual environ-ment was projected. As detailed in previous work (Shine et al.,2013a), foot pedals were positioned under the participants’feet (Fig. 1A), and participants were instructed to flex andextend their ankle in order to ‘tap’ the pedals, which in turnallowed the participants to navigate forward through the vir-tual environment (Shine et al., 2013a). Forward progressionwas only achieved when participants alternately depressedthe pedals (i.e. left-right-left).
The virtual environment consisted of a straight corridor(presented in first-person view) that contained environmentallysalient triggers, such as wide and narrow gaps and doorways.As in previous experiments, cognitive cues were also includedto exacerbate set-shifting deficits. Both narrow doorways andcognitive cues have previously been shown to elicit freezingbehaviour in susceptible individuals (Shine et al., 2013a).Walking and stopping in the virtual environment was initiatedby simple (e.g. WALK and STOP) and complex cue words(e.g. ‘WALK’ if the colour-word pair match and ‘STOP’ ifthe colour-word pair do not match) that were displayedon the screen (for further details see Shine et al., 2013b).Performance on the task was titrated to acceptable levelsprior to scanning. Patients were familiarized with the taskprior to scanning to ensure acceptable performance. Sixteenparticipants completed the full 10-min protocol, and 25participants completed a shorter 5-min protocol.
The timing of each participants’ footsteps during the taskwas collected by recording the onset of each sequential pedaldepression. From this output, the modal footstep latency(i.e. step time) for the duration of the protocol was calculatedas the weighted average after removing all cognitive cues andfreezing of gait episodes (Shine et al., 2013b). Freezing of gaitwas defined by any footstep latency that was longer than twicethe modal footstep latency. This definition is in line with pastwork and has been previously shown to correlate with theseverity freezing of gait episodes experienced in the ‘real’world (Gilat et al., 2013; Matar et al., 2013; Shine et al.,2013a). The coefficient of variation of step time (i.e. steptime variability) was calculated for the duration of the proto-col for which freezing of gait episodes were also removed.
Functional MRI acquisition and preprocessing
A General Electric 3 T MRI was used to obtain T2*-weightedecho planar functional images were acquired in sequentialorder with repetition time = 3000 ms, echo time = 40 ms,flip angle = 90!, 40 axial slices covering the wholebrain, field of view = 220 mm, interslice gap = 0.4 mm and
The network signature of freezing of gait BRAIN 2018: Page 3 of 16 | 3
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

raw voxel size = 3.9 mm " 3.9 mm " 4 mm thick. High-reso-lution 3D T1-weighted anatomical images with voxelsize = 0.4 " 0.4 " 0.9 mm were obtained for co-registrationwith functional scans.
Statistical Parametric Mapping Software (SPM12, WellcomeTrust Centre for Neuroimaging, London, UK, http://www.fil.ion.ucl.ac.ak/spm/software) was used for image preprocessing.Functional scans were: (i) manually realigned along theanterior-posterior commissure; (ii) slice time corrected to the
median (21st) slice in each repetition time; (iii) realigned tocreate a mean realigned image and measures of 6! of rigidhead movements were calculated for later use in the correctionof minor head movements; (iv) unwarped to deal with residualmovement-related variance induced by the susceptibility-by-movement interaction effects; (v) spatially normalized usingthe T1-weighted image to improve segmentation accuracy;(vi) co-registered; and (vii) smoothed using an 8-mm full-width at half-maximum isotropic Gaussian kernel.
Figure 1 Experimental design and protocol. (A) The left panel illustrates a participant lying in the MRI scanner with the foot pedals at their
feet. The right panel shows an example of the virtual environment that the participant views while lying in the MRI scanner. As the participant
performs the task, depressing the pedals (e.g. left-right-left-right), step time is recorded and periods of normal foot tapping and freezing are
calculated. The relationship between the percentage of time spent frozen and the FCI (B: r = 0.40, P = 0.01); inverse of TMT- B (C: r = 0.35,
P = 0.02); step time variability (D: r = 0.56, P5 0.001); and HADS (E: r = 0.14, P = 0.37) are also illustrated. PCA = principle component analysis.
4 | BRAIN 2018: Page 4 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

Multiple precautions were taken to ensure strict control ofthe effects of head motion: (i) all participants were instructedto minimize head motion by keeping the legs and hips station-ary and only moving the ankles to depress the foot pedals;(ii) cushions were placed inside the head coil to limit the phys-ical possibility of head motion; (iii) ArtRepair was used toanalyse each trail and applied the interpolation method tocorrect for large amounts of global drift or scan-to-scanhead movement 41.5 mm; and (iv) six motion and nuisanceregressors were regressed out of each participants’ extractedtime series during statistical modeling.
The freezing component index
To estimate a low-dimensional signature of the factors thatrelate to freezing across subjects, we ran a principal compo-nents analysis on a set of three behavioural variables that werehypothesized a priori to be associated with the freezing pheno-type: cognitive, TMT-B Z-score (inverted so that worse scoreswere positive); motor, step time variability; and limbic, HADStotal. We labelled the first principal component (which ex-plained 79.24% of the variance) the ‘Freezing ComponentIndex’ (FCI). To confirm that the FCI related to freezing ofgait severity, we correlated the principal component tothe percentage of time spent frozen during the virtualreality task (any footstep latency that was longer than twicethe modal footstep latency) using a Spearman’s correlation(Fig. 1B).
Region of interest selection
To improve the specificity of our imaging analysis, we prede-fined a set of regions of interest. Based on previous work(Lewis and Barker, 2009), we chose cortical and subcorticalregions that covered the motor, cognitive and limbic networks,along with striatal regions that are functionally related toeach network. The cortical regions were chosen using theterm-based meta-analyses tool NeuroSynth (http://www.neuro-synth.org/). Specifically, we identified the top regions asso-ciated with the terms: ‘cognitive control network’, ‘motornetwork’ and ‘anxiety’. It should be noted that anxiety anddepression identified similar regions during the NeuroSynthsearch; however, we selected anxiety as the search term sinceit has been more robustly linked to freezing. The peakMNI coordinates for each region (Table 1) were then usedto construct 8 mm region of interest spheres (Fig. 2). Basedon previous research (Di Martino et al., 2008; Bell et al.,2015), we also defined a set of 2 mm region of interest spheresto parcellate the striatum into seven distinct regions.
Functional connectivity analysis
To conduct a task-based functional connectivity analysis, timeseries data were extracted from the first level general linearmodel for each of the 31 regions of interest (17 cortical, 14striatal) using the MarsBaR toolbox (http://marsbar.source-forge.net/). Further preprocessing steps included the normaliza-tion of each region to its own mean and standard deviation(SD), low pass filtering (f40.125 Hz), and removal of themean signal across all regions at each time point. SPM12software was then used to model freezing and normal foottapping (plus their temporal and spatial derivatives) using anepoch design, after which the regressors were convolved with acanonical haemodynamic response function. Normal foot
tapping epochs were defined as five consecutive footstepsthat were not interrupted by an environmentally salient cue(such as a doorway) or by a cognitive cue. The criterion ofa minimum of five freezing events was selected to permit ef-fective modelling of freezing events (Poldrack et al., 2011).Thus, participants who experienced less than five freezingevents during functional MRI scanning were removed fromall of the subsequent imaging analyses. Our cohort of freezerswas made up of participants from two separate studies, whichvaried the task length during functional MRI. Thus, our finalsample included 20 participants: 13 participants who com-pleted a 10-min protocol and seven who completed a 5-minprotocol.
To estimate the time-resolved functional connectivity be-tween each of the 31 regions of interest over the course ofthe task, we used the multiplication of temporal derivativesapproach (Shine et al., 2015) with a window size of 10 repe-tition times (i.e. 30 s). Briefly, this technique affords a win-dowed estimate of functional connectivity as it evolves overtime (code is freely available at http://github.com/macshine/coupling/). To determine whether functional connectivitybetween regions changed with respect to freezing behaviour,the pairwise connectivity estimates were fitted to a generallinear model comparing epochs of freezing with those ofnormal foot tapping. To control for multiple comparisons,we performed a non-linear permutation testing by creating anull distribution of 5000 randomly permuted connectivitymatrices (Nichols and Holmes, 2002). Edges were deemed sig-nificant if the connectivity strength was either stronger orweaker than the 97.5th (or 2.5th) percentile of the null distri-bution, respectively. To further minimize type I errors, we alsothresholded results using a large effect size (Cohen’s d5 0.8;Supplementary Table 1).
To determine the relationship between abnormal functionalconnectivity and freezing severity and heterogeneity, we sepe-rately correlated the freezing4 normal foot tapping functionalconnectivity matrix with the FCI and each individual component,respectively (Supplementary Table 2). A similar non-parametricpermutation approach as detailed above was used to correct formultiple comparisons. Specifically, we randomly re-sorted theFCI vector across participants and then estimated the strengthof correlation between the reordered FCI and each pairwise func-tional connection 5000 times per edge—edges 52.5th percentile(P5 0.05) were retained for further interpretation. Only the sig-nificant edges that both survived permutation testing and werealso significantly correlated with the FCI were reported.
To aid in the interpretation of our results, edges werequalitatively sorted according to their relationship with theFCI. We reasoned that edges that were similarly correlatedwith the FCI (e.g. an edge with increased coupling duringfreezing was positively correlated the FCI or an edge withdecreased coupling during freezing was inversely correlatedwith the FCI) may represent an index of freezing ‘severity’(i.e. greater coupling/anti-coupling was associated with agreater ‘freezing severity’) (Fig. 3). In contrast, if the FCIwas correlated in the opposite direction to the mean patternobserved in an edge (e.g. an edge with increased coupling wasassociated with a lower ‘severity’ score on the FCI), then theedge may be ‘compensatory’. Finally, if an edge was found tobe independent of the severity of freezing of gait correlates, itwas labelled as ‘independent’, suggesting that there was a sig-nificant pattern of abnormal connectivity that was consistently
The network signature of freezing of gait BRAIN 2018: Page 5 of 16 | 5
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018
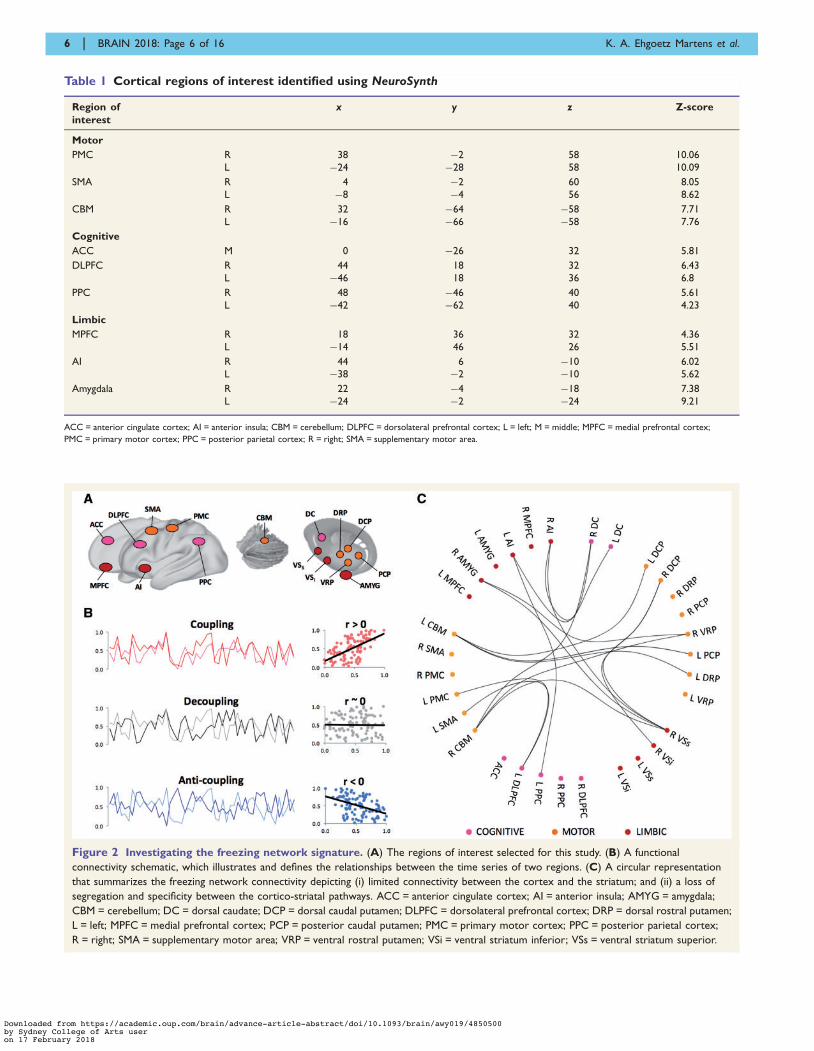
Table 1 Cortical regions of interest identified using NeuroSynth
Region ofinterest
x y z Z-score
Motor
PMC R 38 #2 58 10.06L #24 #28 58 10.09
SMA R 4 #2 60 8.05L #8 #4 56 8.62
CBM R 32 #64 #58 7.71L #16 #66 #58 7.76
Cognitive
ACC M 0 #26 32 5.81
DLPFC R 44 18 32 6.43L #46 18 36 6.8
PPC R 48 #46 40 5.61L #42 #62 40 4.23
Limbic
MPFC R 18 36 32 4.36L #14 46 26 5.51
AI R 44 6 #10 6.02L #38 #2 #10 5.62
Amygdala R 22 #4 #18 7.38L #24 #2 #24 9.21
ACC = anterior cingulate cortex; AI = anterior insula; CBM = cerebellum; DLPFC = dorsolateral prefrontal cortex; L = left; M = middle; MPFC = medial prefrontal cortex;
PMC = primary motor cortex; PPC = posterior parietal cortex; R = right; SMA = supplementary motor area.
Figure 2 Investigating the freezing network signature. (A) The regions of interest selected for this study. (B) A functional
connectivity schematic, which illustrates and defines the relationships between the time series of two regions. (C) A circular representation
that summarizes the freezing network connectivity depicting (i) limited connectivity between the cortex and the striatum; and (ii) a loss of
segregation and specificity between the cortico-striatal pathways. ACC = anterior cingulate cortex; AI = anterior insula; AMYG = amygdala;
CBM = cerebellum; DC = dorsal caudate; DCP = dorsal caudal putamen; DLPFC = dorsolateral prefrontal cortex; DRP = dorsal rostral putamen;
L = left; MPFC = medial prefrontal cortex; PCP = posterior caudal putamen; PMC = primary motor cortex; PPC = posterior parietal cortex;
R = right; SMA = supplementary motor area; VRP = ventral rostral putamen; VSi = ventral striatum inferior; VSs = ventral striatum superior.
6 | BRAIN 2018: Page 6 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

present in the cohort of patients with freezing of gait irrespect-ive of freezing severity. A similar analysis was then conductedusing the individual component scores (i.e. cognitive, motor,limbic); however, ‘independent’ edges were ignored due tooverlap across the three component domains.
Results
Executive summary
In this study, we made an explicit choice to expose thedetailed heterogeneity of the freezing of gait phenotype.To clarify our results, we would like to first emphasizethe overarching framework that we applied to investigatethe multivariate nature of freezing. First, by contrastingconnectivity during freezing episodes to normal foottapping we constructed a ‘Freezing Network Signature’(Figs 2C and 3). Second, we related the network signatureof freezing to the FCI (which is significantly correlated withper cent time spent frozen during the task) in order toassess which aspects might be associated with ‘severity’,‘compensation’, or ‘independent’ altogether (Fig. 4).Third, by decomposing the FCI into its cognitive, motorand limbic components, we examined the relationship be-tween individual behavioural components and the freezingnetwork signature further (Fig. 5). Characteristics of par-ticipants both included and excluded from the functionalMRI analysis are presented in Table 2. Notably the twogroups (MRI + and MRI#) were matched in age, symptomseverity, cognitive status, FOGQ3, anxiety, total HADS,processing speed, set-shifting ability and step time variabil-ity. There were a few group differences such that the cohortincluded in the functional MRI analysis (MRI + ) had asignificantly higher dopamine dose equivalence (P = 0.04),greater depression score (although not clinical significant)
(P = 0.03), faster foot step latency (P5 0.01), and greateramount of freezing (P5 0.01).
Freezing network signature
Figure 3 illustrates a summary of the overall network as afunction of freezing of gait (i.e. freezing4normal foot tap-ping). During freezing episodes, the functional connectivitywas disrupted between the striatum and the cognitivecontrol network and the motor network compared tonormal foot tapping. However, increased functional con-nectivity between the limbic network and striatum wasevident when freezing episodes were contrasted tobouts of normal foot tapping. Further details are providedbelow.
Cognitive network
During episodes of freezing, anti-coupling was observedbetween the cognitive control network and striatum,whilst increased coupling was observed within the cognitivecontrol network when compared to normal foot tapping.The cerebellum was also significantly more coupled to thecaudate and cognitive control network.
Motor network
There was substantial disruption to the motor networkduring freezing compared to normal foot tapping. Greateranti-coupling was found both within the cortical motornetwork, and within the putamen. Furthermore, themotor network was also decoupled from the putamenand and limbic structures and instead coupled to thedorsal caudate nucleus.
Limbic network
Increased coupling between the limbic network (corticaland subcortical) and the ventral striatum was found
Figure 3 Decomposing the freezing signature. During freezing episodes, functional connectivity was disrupted between the striatum and
the cognitive control network and the motor network compared to normal foot tapping. However, increased functional connectivity between the
limbic network and striatum was evident when freezing episodes were contrasted to bouts of normal foot tapping. C = caudate;
CBM = cerebellum; P = putamen; scLimbic = subcortical limbic; V = ventral striatum.
The network signature of freezing of gait BRAIN 2018: Page 7 of 16 | 7
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

during freezing compared to normal foot tapping.Furthermore, increased coupling between the limbic net-work (cortical and subcortical) and the cognitive controlnetwork was found. In contrast, anti-coupling betweenthe cortical and subcortical limbic network and within
the subcortical network was marked during freezing com-pared to normal foot tapping. Notably, the subcorticallimbic regions were tightly coupled to the dorsal caudate,despite that the cognitive control network was anti-coupledfrom the dorsal caudate nucleus.
Figure 4 Associations between the freezing network signature and the FCI. Functional connections identified in the Freezing
Network Signature, which were associated with a worse FCI are depicted in (A), whereas the connections that were associated with a better FCI
are depicted in (B). The remaining functional connections that were not significantly correlated to the FCI are depicted in C, and (D) dem-
onstrates a summary key that describes how the classification was determined. C = caudate; CBM = cerebellum; P = putamen; PC = principal
component; scLimbic = subcortical limbic; V = ventral striatum.
Figure 5 Unique network signatures for individual components of freezing. Functional connections identified in the freezing network
signature that were associated with more severe components are illustrated in the top panel (A, cognitive; B, motor; C, limbic). Functional connections
identified in the freezing network signature that were associated with reduced component scores (i.e. compensatory) are illustrated in the bottom panel.
C = caudate; CBM = cerebellum; COG = cognitive; P = putamen; scLimbic = subcortical limbic; STV = step time variability; V = ventral striatum.
8 | BRAIN 2018: Page 8 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

Relating the freezing network signa-ture to the freezing component index
To assess which aspects of the freezing network signaturewere associated with severity, compensation, or were inde-pendent of severity, we first verified there was a significantrelationship between the percentage of time spent frozenduring the task and the FCI (r = 0.40, P = 0.01; Fig. 1B).Furthermore, the FCI was also positively correlated withFOGQ3 (r = 0.32, P = 0.045). Figure 4 summarizes therelationship between functional connectivity and the FCI,wherein the FCI was correlated with each significantconnection from the freezing signature (freezing4 normalfoot tapping; Fig. 3).
Severity versus compensation
Overall, a worse FCI (a proxy for freezing severity) wasassociated with: (i) greater coupling within the cognitivecontrol network and also between the cognitive controlnetwork and the limbic cortex; (ii) anti-coupling withinthe putamen, and between the putamen and caudate nu-cleus, as well as a loss of anti-coupling between the puta-men and ventral striatum; and (iii) anti-coupling betweenthe motor cortex and the limbic subcortex, and cognitivecontrol network.
In contrast, reduced freezing severity (i.e. ‘Compensation’)was associated with: (i) anti-coupling between the putamenand cortical networks (cognitive control, limbic and motornetwork); (ii) anti-coupling between the ventral striatumand motor network; and (iii) increased coupling betweenthe cognitive control network and the cerebellum andsubcortical limbic network.
Freezing component signatures
To understand the heterogeneous network signature asso-ciated with freezing, we decomposed the FCI into its cog-nitive, motor and limbic parts, and further examined therelationship between individual behavioural componentsand the freezing network signature. Figure 5 depicts theedges associated with better (green) or worse (purple)scores for each of the three component domains (cognitive,motor and limbic).
Cognitive signature (TMT-B)
Worse set-shifting ability was associated with decoupling be-tween cognitive control network and the motor network, aswell as decoupling between the putamen and the ventral stri-atum. Increased coupling within the ventral striatum wasobserved, as well as between the ventral striatum and thecortical limbic network, and between the cerebellum andcaudate nucleus was also associated with worse set-shiftingability. Anti-coupling between the motor network and thesubcortical limbic network was related to worse set-shiftingability. In contrast, better set-shifting ability was associatedwith coupling between the cognitive control network and thelimbic network (cortex and subcortex), and anti-couplingbetween the cortical and subcortical limbic network.Decoupling between the cortex and striatum was alsoassociated with greater cognitive flexibility.
Motor signature (step time variability)
Worse step time variability was primarily associated withlimbic edges. Greater coupling between cortical limbic andcognitive control networks, and greater anti-coupling be-tween cortical and subcortical limbic networks as well as
Table 2 Demographic, clinical and neuropsychological and virtual reality characteristics
Total participants (n = 41) MRI + (n = 20) MRI# (n = 21)
Mean SD Mean SD Mean SD
Gender, n 9 F (22%) 3 F (15%) 6 F (29%)
Age 67.54 6.40 66.45 5.34 68.57 7.26
DDE 877.9 579.9 1069.5* 687.6 686.3* 374.4
MDS UPDRS-III 33.89 14.0 34.15 12.19 33.62 15.84
MMSE 27.90 2.07 27.65 2.16 28.14 2.01
FOGQ3 2.5 1.01 2.53 1.02 2.48 1.03
HADS-Anxiety 5.78 3.63 6.3 3.25 5.29 3.98
HADS-Depression 5.37 2.77 6.35* 2.83 4.43* 2.42
HADS Total 11.15 5.9 12.65 5.79 9.71 5.77
TMT-A (Z-score) #0.34 1.14 #0.26 0.97 #0.42 1.31
TMT-B (Z-score) #0.84 1.68 #0.80 1.40 #0.88 1.95
Step time variability 23.08 7.86 24.99 7.04 21.27 8.32
Modal foot step latency, s 0.52 0.27 0.38* 0.17 0.64* 0.29
Time spent frozen, % 0.10 0.09 0.14* 0.08 0.07* 0.14
Protocol completed (5-min:10-min) 25:16 7:13 18:3
*Significant group difference, P5 0.05.
DDE = dopamine dose equivalence; FOGQ3 = question 3 of the Freezing of Gait Questionnaire; MDS UPDRS-III = Movement Disorders Society Unified Parkinson’s Disease Rating Scale
motor subsection; MMSE = Mini-Mental State Examination; MRI + = participants included in the functional MRI analysis; MRI# = participants removed from the functional MRI analysis.
The network signature of freezing of gait BRAIN 2018: Page 9 of 16 | 9
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

within the putamen. Decoupling between the putamen andsubcortical limbic regions was also associated with greaterstep time variability.
In contrast, reduced step time variability was primarilyassociated with reduced coupling between the cortex andstriatum and across the striatum. More specifically, anti-coupling between the putamen and the cognitive controlnetwork and cortical limbic network, as well as anti-cou-pling between the dorsal caudate and the putamen wererelated to less variable step timing. Additionally, decouplingbetween ventral striatum and both the motor network andputamen, as well as decoupling between the motor networkand cognitive control network was also associated withreduced step time variability.
Limbic signature (HADS)
Worse anxiety and depression was associated withincreased coupling between the cortical limbic and cogni-tive control network, as well as between the motor networkand the dorsal caudate nucleus. Furthermore, anti-couplingbetween the subcortical limbic network and the motor net-work, within the putamen, and between the putamen andthe dorsal caudate was also related to worse affectivedisturbance.
Similar to the step time variability, less affective disturb-ance was associated primarily with anti-coupling betweenthe cortex and the striatum. Specifically, anti-coupling wasfound between the putamen and the cognitive control net-work and cortical limbic network, as well as between thedorsal caudate and the cognitive control network.Additionally, anti-coupling between the cortical and sub-cortical limbic network was found. Decoupling betweenthe putamen and the subcortical limbic network andmotor network as well as decoupling between the motornetwork and the various other cortical and striatal regions(i.e. the cognitive control network, subcortical limbic net-work, putamen, and the ventral striatum) was also relatedto less severe anxiety and depression symptoms.
DiscussionIn this study, we used a task-based connectivity analysis toidentify latent heterogeneity within the neural signatureunderlying freezing of gait in Parkinson’s disease. Whencompared to periods of normal foot tapping (i.e. a proxyfor normal walking), freezing episodes were characterizedby an overall loss of synchrony between the cortex and thestriatum, as well as a loss of segregation and specificitybetween the cortico-striatal pathways (Figs 2B and 3).Subsequent group-level behavioural interrogation allowedus to dissociate the freezing signature into three distinctcategories (Fig. 4): a set of intracortical and intrastriatalconnections that were related to freezing severity; a groupof cortico-striatal connections that were putatively relatedto compensation; and a network of cortico-striatal andstriato-cerebellar connections that were independent of
freezing severity. Finally, we related these deficits to indi-vidual differences in the behavioural factors (cognitive,motor, limbic) that predispose individuals towards freezingepisodes (Fig. 5). Together, these results provide confirma-tory evidence for systems-level impairments in the patho-physiology of freezing of gait in Parkinson’s disease.
Severity versus compensation
In contrast to previous studies, the approach used in thisstudy was able to decipher the patterns within the freezingof gait phenotype that were related to individual differencesin freezing severity. Specifically, worsened scores on thefirst principal component (i.e. inferring worse behaviouralfreezing) was associated with increased connectivity withinthe cognitive control network and between the cognitivecontrol network and other cortical networks (i.e. motorand limbic; Fig. 4). That is, greater cortical ‘cross-talk’was associated with worse freezing, and less cross-talk be-tween the cortex and striatum was associated with lessfreezing of gait. It is plausible that cortical areas maybecome too integrated due to striatal dysfunction or peri-ods of increased limbic drive. Alternatively, actively redu-cing the ‘cross-talk’ between the striatum and cortex (e.g.via maximizing temporal separation of regional activity)may reduce the severity of freezing of gait and reflect acompensatory strategy to overcome freezing episodes.There is growing evidence that dopaminergic replacementtherapy may restore the appropriate corticostriatal connect-ivity (Esposito et al., 2013; Tahmasian et al., 2015; Gilatet al., 2017), and normalize the disrupted networktopology (Berman et al., 2016; Kim et al., 2017), thus po-tentially minimizing ‘crosstalk’. Further research is neededto determine whether dopaminergic replacement therapyrestores network segregation (Nieuwhof and Helmich,2017), and whether network segregation versus integrationmight be protective against freezing of gait. The currentfindings also converge with previous suggestions that freez-ing represents dysfunction within a distributed network ofwidespread frontal and parietal cortical regions (Shineet al., 2013c), and supplements studies that have shown areduction in frontoparietal blood oxygen level-dependentactivation (Snijders et al., 2011; Shine et al., 2013b), greymatter atrophy in frontal and parietal cortices (Kostic et al.,2012; Tessitore et al., 2012a; Herman et al., 2014), andreduced perfusion rate in frontal networks (Imamura et al.,2012) in association with freezing severity. Interestingly,freezing severity has been associated with reduced func-tional connectivity within the ‘executive-attention’ neuralnetwork in the ‘resting’ state (Tessitore et al., 2012b),whereas here we demonstrate the opposite pattern and re-lationship during a task-based protocol. These differing re-sults could emphasize that freezing is likely related toaltered cortical control of gait, or perhaps via failed com-pensatory strategies that facilitate too much cortical inte-gration. These findings also indirectly support predictionsof the cross-talk model, which hypothesizes that an
10 | BRAIN 2018: Page 10 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

overflow of communication between the cortex and thestriatum, paired with a loss of segregation (demonstratedhere by pathological cortical coupling) among competingyet complimentary brain networks may overwhelm thestriatum, thus generating excessive inhibitory output fromthe globus pallidus to the brainstem structures controllinggait, ultimately manifesting freezing behaviour (Lewis andBarker, 2009).
Similar to recent work, the current study also identifiedpatterns of abnormal cerebellar connectivity that were asso-ciated with freezing of gait. In previous work, Fasano et al.(2017) used a novel lesion network mapping technique toshow that lesions in the dorsal midline cerebellum wereimplicated in generating freezing symptoms; however, itwas inconclusive whether the cerebellum’s role in lesion-induced freezing was relevant for parkinsonian freezing ofgait. Several studies have shown abnormalities in cerebellarlocomotor regions’ structural and functional connectivity inParkinson’s disease patients with freezing (Schweder et al.,2010; Fling et al., 2014; Vercruysse et al., 2015; Myerset al., 2017), yet a recent study investigating cerebellartheta burst stimulation found that it did not improvefreezing of gait in patients with Parkinson’s disease(Janssen et al., 2017). In the current study, the connectivitybetween the cerebellum and the striatum was a markedfeature of the network signature of freezing independentof severity, whereas coupling between the cerebellum andcognitive control network was found to be compensatory(i.e. associated with less freezing). Therefore, we providefurther evidence that the cerebellum plays an importantrole in the underlying pathophysiology of freezing of gait;however, future research is needed to understand itscompensatory relationship in order to help guide thedevelopment of effective new therapies.
Recent work by Nieuwhof et al. (2017) has also shown aloss of segregation within cortico-striatal loops during dual-task performance while foot-tapping in Parkinson’s disease(Nieuwhof et al., 2017). Here, we advanced these findingsby mapping patterns of abnormal connectivity during freez-ing, which were further related to individual differences inpredispositional components (Fig. 3). More specifically, weused a low dimensional behavioural signature to infer pat-terns of functional connectivity associated with increasedseverity; with increased compensation; or those that wereindependent of severity. Based on Nieuwhof’s predispos-ition hypothesis, it may have been expected that a loss ofcortico-striatal segregation (i.e. greater striatal overlap)might be independent of freezing severity, and thus repre-sent trait-like factors common in all patients with freezingof gait. Indeed, rather than the typical cortico-striatalcircuitries being functionally coupled (e.g. cognitive controlnetwork – dorsal caudate; motor network – dorsal puta-men; limbic network – ventral striatum), we observed in thecurrent study little specificity (i.e. greater overlap) in thefunctional connectivity between the cortical networks andthe striatum during freezing compared to normal foottapping. Moreover, all positive edges between the cortex
and striatum (e.g. motor network – caudate, subcorticallimbic – caudate) were independent of freezing severity.Furthermore, increased limbic connectivity with thestriatum, as well as disrupted communication within themotor network and between the cognitive control networkand the striatum were also independent of freezing severity.Therefore, one interpretation could be that these abnorm-alities in communication were consistently present acrossall patients with freezing of gait during episodes of freezing,and thus may represent the true neural underpinnings offreezing of gait that are unchanged by heterogeneoustrait-like factors.
In contrast, the first principal component (a proxy forfreezing severity) was associated with altered connectivityacross the striatal nuclei (Fig. 4). More specifically, worsefreezing behaviour was associated with a loss of couplingwithin the putamen and between the dorsal caudate and pu-tamen, and accompanied by greater coupling between theventral striatum and putamen. These findings are in keepingwith previous work which has demonstrated decreased bloodoxygen level-dependent signal in subcortical areas (such asthe caudate, globus pallidus internus, thalamus and mesen-cephalic locomotor region) during freezing (Shine et al.,2013b). Furthermore, these results may reflect the dopamin-ergic aetiology of freezing of gait, since dopamine degener-ation progresses posterior-dorsal to anterior-ventral withinthe striatum (Poldrack, 2005; Wu and Hallett, 2005), thusaffecting the putamen and dorsal caudate more so than theventral striatum. This degenerative process could lead torecruitment of the dopamine-preserved ventral striatum as acompensatory strategy to aid with cortico-striatal processing,which may also contribute to greater overlap and loss ofsegregation within cortico-striatal pathways. This notion isconsistent with research suggesting that a bottleneck of pro-cessing might occur when motor and cognitive inputs arefunnelled into the relatively spared ventro-anterior putamendue to the gradient dopamine depletion in the dorsoposteriorputamen (Lewis and Barker, 2009). Hypothetically, in in-stances where the ventral striatum may be needed for infor-mation processing within its own segregated cortico-striatallimbic loop, this over-recruitment may lead to vulnerabilityacross other systems that rely on compensatory ventralstriatum processing. In this way, limbic input may have agreater capacity to interfere with motor output such as walk-ing, which might offer a further explanation for the emergingrelationship between anxiety and freezing of gait.
Component signatures of freezing
Several of the regions that have been associated with freez-ing of gait are also known to be intimately involved withmotor, cognitive and limbic functions, which in turn mayexplain the well-known clinical relationship between freez-ing and impairment across multiple behavioural domains(Schaafsma et al., 2003; Giladi and Hausdorff, 2006;Naismith et al., 2010; Nutt et al., 2011; Ehgoetz Martenset al., 2014b; Hall et al., 2015). Indeed, this very feature of
The network signature of freezing of gait BRAIN 2018: Page 11 of 16 | 11
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

freezing forms an important basis of the ‘cross-talk’ model,which describes freezing as a transient overload of infor-mation processing capacity of within the broader networkof the brain (i.e. independent of the domain that catalysesoverload), ultimately leading to a breakdown in motorfunction and freezing (Lewis and Barker, 2009; Lewisand Shine, 2016). Given that the precise circuitry thatprovokes freezing likely differs depending on the specifictrigger that will cause an episode (cognitive, motor,limbic), this study aimed to explore how functionalconnectivity correlated to particular cognitive, motor, andlimbic components of freezing (i.e. set-shifting ability, steptime variability, anxiety and depression severity), in aneffort to better understand their unique contributions andneural underpinnings.
Cognitive signature
Set-shifting impairments are well known in Parkinson’s dis-ease, and have been linked with dopaminergic depletion inthe caudate nucleus as well as reduced structural and func-tional connectivity to frontal areas (Bartels et al., 2006;Shine et al., 2013d). In the current study, worse set-shiftingability was associated with greater coupling between thecerebellum and the caudate as well as between the ventralstriatum and cortical limbic regions. In contrast, greatercognitive flexibility was associated with coupling betweenthe cognitive control network and the limbic network andless connectivity between the cortex and the striatum. Inpatients with Parkinson’s disease, including patients withfreezing of gait, a relationship between affective disturbanceand performance on the TMT has been established, suchthat anxiety and depression can interfere with set-shiftingabilities (Ehgoetz Martens et al., 2016d). Our findings mayfit with this notion, since greater synchronization within thelimbic cortico-striatal pathway relates to worsened TMT-Bperformance, whereas greater ‘top-down’ control of thelimbic network is associated with more cognitive flexibility,which confirms the dependent nature of cognitive andaffective disturbance in patients with freezing of gait.
Motor signature
Increased step time variability has also been robustly linkedwith freezing of gait (Pieruccini-Faria et al., 2015; Weisset al., 2015) and is thought to reflect a loss in motor auto-maticity due to dopaminergic denervation predominately inthe posterior dorsal putamen (Wu and Hallett, 2005). Thisarea of the putamen is thought be responsible for chunkingmotor sequences to improve efficiency and free attentionalresources (Graybiel, 1998). Given that dopaminergicdepletion affects the posterior-dorsal putamen early on,compensatory attentional and cognitive control networkshave been postulated to compensate by being broughtonline to control and coordinate movements. Recent neu-roimaging work has indeed shown that greater activationin frontal areas (Wu and Hallett, 2005) and increased
intrastriatal functional connectivity was associated withgreater movement variability in patients with Parkinson’sdisease (Gilat et al., 2017). In patients with freezing ofgait, it might be expected that this loss of automaticity(represented by increased step time variability) places aneven greater demand on limited attentional resources andwill be similarly related to dysfunctional striatal connectiv-ity. Here, we found that worsened step time variability wasindeed related to disrupted connectivity within the striatum,predominately between the putaminal nuclei. Furthermore,less variability was associated with anti-coupling betweenthe putamen and the cortex, as well as reduced connectivitybetween intra-striatal nuclei and between the cognitive con-trol network and motor network. Overall these findings arein support of previous work suggesting a loss of dopaminein the dorsal striatum is related to greater step timevariability. Likewise, the pattern of results in this studycould also reflect a compensatory shift in processing, suchthat the ventral-anterior striatum may be recruited more toaid with cortico-striatal processing to control movementsince it may have more preserved dopaminergic neurons.Furthermore, individuals with less step time variability alsohad reduced connectivity between the cognitive control net-work and motor network to employ top-down control ofmovement, which is sensible given that cognitive control ofgait typically enhances variability and reduces rhythmicity.
Limbic signature
Although affective disturbance and freezing of gait are clo-sely related, little research has examined the neural correl-ates of anxiety or depression in Parkinson’s disease, letalone the integrity and functional connectivity of thelimbic cortico-striatal pathway in patients with freezing ofgait. Here, we demonstrate for the first time that limbic-striatal connectivity is a key component of the freezingneural signature. Moreover, increased input from thelimbic network may contribute to interference in processingat the level of the basal ganglia via ‘limbic load’, which hasbeen previously suggested based on behavioural results as apotential mechanism underpinning freezing of gait (EhgoetzMartens et al., 2014b). Considering the loss of segregationacross the striatum, the ventral striatum may become in-creasingly utilized for motor-related processing and move-ment control, which could be especially problematic inanxious patients, who have an ‘overactive’ limbic network(and threat detection system) to begin with. Surprisingly,however, we found that greater affective disturbance inpatients with freezing of gait was associated withpredominately motor network connectivity rather thanlimbic per se.
Similar to variability and set-shifting, less affective dis-turbance was associated with less functional connectivitybetween the cortex and the striatum. This might suggestthat anxiety in Parkinson’s disease, or perhaps that whichis associated with freezing of gait is intimately related tomovement control deficits. A common discussion point is
12 | BRAIN 2018: Page 12 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

whether anxiety is a response to movement impairments ora driving cause. It is well known that uncertainty in sensoryinformation provides the basis for anxious processes. Thus,perhaps sensory impairments that underlie movementsymptoms could also provoke anxiety over time inParkinson’s disease. Previous work has demonstrated thatsensory deficits are linked to freezing as well (EhgoetzMartens et al., 2013b, 2014a, 2016a), such that freezingwas exacerbated when asked to walk toward a doorway inthe dark compared to a lit frame, and also suggest that thethreat in an environment compounds freezing behaviourssince freezing was also greater when walking toward adoorway in the dark compared to a dark open space(Ehgoetz Martens et al., 2013a). Further research isneeded to test whether anxiety is related to sensory impair-ments in Parkinson’s disease early on in the disease course(Ehgoetz Martens et al., 2016b), and also to further disen-tangle whether anxiety is the chicken or egg to freezing.
Clinical implications
Overall, these findings suggest that unique neural signaturesexist and relate to different behavioural correlates of freez-ing. The suggestion that freezing of gait subtypes may existhas been previously put forward (Fasano et al., 2015);however, to date, limited evidence has been presented.This is the first study to examine multiple components offreezing, and demonstrate that unique neural signatures arepresent during freezing of gait episodes. Based on our re-sults, one could speculate that different subtypes of freezingof gait arise due to individual vulnerabilities within the di-verse systems required for dynamic control of adaptive be-haviour and locomotion. For instance, if one is highlyanxious, input from the limbic system to the striatumcould ‘tip the scale’, causing freezing episodes to occurwhen the processing capacity of the system exceeds itslimit. Similarly, if someone has significant impairments tomotor automaticity and hence, relies more heavily on cog-nitive control resources for normally automatic gait, theninstances that perturb or divide these necessary resourcesmay in turn provoke freezing of gait. However, these rela-tionships are not always straightforward and remainopaque, for example in situations where kinesia paradoxaenable freezers to execute movement in highly stressful situ-ations, cross-talk between circuitries might channel ad-equate compensation rather than overload the system(Nieuwhof and Helmich, 2017). Thus, further research iscertainly needed to illuminate whether segregation versusintegration is detrimental or compensatory across varioussituations specific to freezing.
It should be noted that these findings do not necessarilydispute the involvement of a common downstream path-way of freezing (Lewis and Shine, 2016), but rather ex-poses the fact that there are potentially distinct upstreamcauses that may represent dissociable targets for treatment.For example, treating anxiety may have beneficial effectsfor specific individuals with freezing of gait (Ehgoetz
Martens et al., 2016c), whereas cognitive training mightbe more effective for others (Walton et al., 2014; Leunget al., 2015). Alternatively, an individual with either amotor or anxious phenotype may benefit from sensory-based training programmes that work on improving theuse of sensory information and restoring gait automaticity(Sage and Almeida, 2009; Lefaivre and Almeida, 2015).However, much research is still needed to clarify thispoint and further research is also needed to characterizethese subtypes, and determine whether in a data-drivenapproach there is any evidence of these different types,and whether they lend support to different cortical mech-anisms and thus different ‘cortically’ geared therapies.
Limitations and considerations
It is important to point out the shortcomings and limita-tions of this work. First, we did not include any ‘ON’ statepatients with freezing of gait in this cohort. Although this isa rare subtype of freezing of gait, it is currently unclearwhether the findings presented here would generalize tothis subgroup. Second, although the virtual gait task isable to capture much of the coordination involved ingait, it also has many limitations. For example, the con-straints imposed by the functional MRI scanner are incom-patible with analyses that interrogate the delicate control ofupright stance and dynamic balance required whilewalking. Additionally, the freezing events detected couldbe confounded by fatigue or a loss in concentration.Another caveat in this study was that the participantsincluded in the imaging cohort had a faster modal footsteplatency on average than the cohort who were excluded.Although a slower foot tapping speed may set the thresholdto detect freezing higher than those with a faster foot steplatency, we did not observe an association between modalfoot step latency and percentage of time spent frozen(P4 0.1). Thus, it is unlikely that foot step latencyconfounds the neuroimaging results reported in this study.Finally, given that our focus was on investigating theparticular episodic characteristics of freezing, we did notinclude a control group in this study. However, futureresearch should consider examining differences in continu-ous gait patterns between Parkinson’s disease patients withand without freezing to further understand the role of themotor, cognitive and limbic networks and their contribu-tion to gait variability and gait disturbances.
ConclusionOverall, these findings provide confirmatory evidence forsystems-level impairments in the pathophysiology of freez-ing of gait. Importantly, the findings of this study showhow functional connectivity during freezing is correlatedto particular cognitive, motor and limbic features in effortto better understand their unique contributions andneural underpinnings. Thereby we further advance our
The network signature of freezing of gait BRAIN 2018: Page 13 of 16 | 13
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

understanding of the whole-brain deficits that mediatesymptom expression in Parkinson’s disease.
AcknowledgementsWe would like to thank the participants and their familymembers who gave us their time to take part in this study.
FundingK.A.E.M. is supported by a Parkinson Canada PostdoctoralFellowship; J.M.H. is supported by a Western SydneyUniversity Postgraduate Research Award; M.G. is supportedby a University of Sydney International Scholarship; C.C.W.is supported by an Australian Postgraduate Award at theUniversity of Sydney; S.J.G.L. is supported by a NationalHealth and Medical Research Council PractitionerFellowship (1003007); J.M.S. is supported by a NationalHealth and Medical Research Council CJ MartinFellowship (1072403).
Supplementary materialSupplementary material is available at Brain online.
ReferencesAlmeida QJ, Lebold CA. Freezing of gait in Parkinson’s disease: a
perceptual cause for a motor impairment? J Neurol NeurosurgPsychiatry 2010; 81: 513–18.
Amboni M, Cozzolino A, Longo K, Picillo M, Barone P. Freezing ofgait and executive functions in patients with Parkinson’s disease.Mov Disord 2008; 23: 395–400.
Barbe MT, Amarell M, Snijders AH, Florin E, Quatuor EL, SchonauE, et al. Gait and upper limb variability in Parkinson’s diseasepatients with and without freezing of gait. J Neurol 2014; 261:330–42.
Bartels AL, de Jong BM, Giladi N, Schaafsma JD, Maguire RP,Veenma L, et al. Striatal dopa and glucose metabolism in PDpatients with freezing of gait. Mov Disord 2006; 21: 1326–32.
Beck EN, Ehgoetz Martens KA, Almeida QJ. Freezing of gait inParkinson’s disease: an overload problem? [Internet]. PLoS One2015; 10: e0144986.
Bell PT, Gilat M, O’Callaghan C, Copland DA, Frank MJ, Lewis SJG,et al. Dopaminergic basis for impairments in functional connectivityacross subdivisions of the striatum in Parkinson’s disease. HumBrain Mapp 2015; 36: 1278–91.
Berman BD, Smucny J, Wylie KP, Shelton E, Kronberg E, Leehey M,et al. Levodopa modulates small-world architecture of functionalbrain networks in Parkinson’s disease. Mov Disord 2016; 31:1676–84.
Bhatt H, Pieruccini-Faria F, Almeida QJ. Dynamics of turning sharp-ness influences freezing of gait in Parkinson’s disease. ParkinsonismRelat Disord 2013; 19: 181–5.
Bloem BR, Hausdorff JM, Visser JE, Giladi N. Falls and freezing ofgait in Parkinson’s disease: a review of two interconnected, episodicphenomena. Mov Disord 2004; 19: 871–84.
Burn DJ, Landau S, Hindle JV, Samuel M, Wilson KC, Hurt CS, et al.Parkinson’s disease motor subtypes and mood. Mov Disord 2012;27: 379–86.
Di Martino A, Scheres A, Margulies DS, Kelly AMC, Uddin LQ,Shehzad Z, et al. Functional connectivity of human striatum: a rest-ing state fMRI study. Cereb Cortex 2008; 18: 2735–47.
Ehgoetz Martens KA, Ellard CG, Almeida QJ. A closer look at mech-anisms underlying perceptual differences in Parkinson’s freezers andnon-freezers. Neuroscience 2014a; 274: 162–9.
Ehgoetz Martens KA, Ellard CG, Almeida QJ. Does anxiety causefreezing of gait in Parkinson’s disease? PLoS One 2014b; 9:e106561.
Ehgoetz Martens KA, Ellard CG, Almeida QJ. Dopaminergic medica-tion amplifies sensory integration deficits during static balance inParkinson’s patients with freezing of gait. BAOJ Neurol 2016a; 2:1–6.
Ehgoetz Martens KA, Ellard CG, Almeida QJ. Evaluating the linkbetween dopaminergic treatment, gait impairment, and anxiety inParkinson’s disease. Mov Disord Clin Pract 2016b; 3: 389–94.Available from: http://doi.wiley.com/10.1002/mdc3.12298.
Ehgoetz Martens KA, Hall JM, Gilat M, Georgiades MJ, Walton CC,Lewis SJG. Anxiety is associated with freezing of gait and atten-tional set-shifting in Parkinson’s disease: a new perspective forearly intervention. Gait Posture 2016c; 49: 431–6.
Ehgoetz Martens KA, Pieruccini-Faria F, Almeida QJ. Could sensorymechanisms be a core factor that underlies freezing of gait inParkinson’s disease? PLoS One 2013a; 8: e62602.
Ehgoetz Martens KA, Pieruccini-Faria F, Silveira CRA, Almeida QJ.The contribution of optic flow to freezing of gait in left- and right-PD: different mechanisms for a common phenomenon?Parkinsonism Relat Disord 2013b; 19: 1046–8.
Ehgoetz Martens KA, Szeto JYY, Muller AJ, Hall JM, Gilat M,Walton CC, et al. Cognitive function in Parkinson’s disease patientswith and without anxiety. Neurol Res Int 2016d; 2016: 6254092.
Esposito F, Tessitore A, Giordano A, De Micco R, Paccone A,Conforti R, et al. Rhythm-specific modulation of the sensorimotornetwork in drug-naive patients with Parkinson’s disease by levo-dopa. Brain 2013; 136: 710–25.
Fasano A, Herman T, Tessitore A, Strafella AP, Bohnen NI.Neuroimaging of freezing of gait. J Parkinsons Dis 2015; 5: 241–54.
Fasano A, Laganiere SE, Lam S, Fox MD. Lesions causing freezing ofgait localize to a cerebellar functional network. Ann Neurol 2017;81: 129–41.
Fling BW, Cohen RG, Mancini M, Carpenter SD, Fair DA, Nutt JG,et al. Functional reorganization of the locomotor network inParkinson patients with freezing of gait. PLoS One 2014; 9:e100291.
Giladi N, Hausdorff JM. The role of mental function in the pathogen-esis of freezing of gait in Parkinson’s disease. J Neurol Sci 2006;248: 173–6.
Gilat M, Bell PT, Ehgoetz Martens KA, Georgiades MJ, Hall JM,Walton CC, et al. Dopamine depletion impairs gait automaticityby altering cortico-striatal and cerebellar processing in Parkinson’sdisease. Neuroimage 2017; 152: 207–20.
Gilat M, Shine JM, Bolitho SJ, Matar E, Kamsma YPT, Naismith SL,et al. Variability of stepping during a virtual reality paradigm inParkinson’s disease patients with and without freezing of gait.PLoS One 2013; 8: e66718.
Graybiel AM. The basal ganglia and chunking of action repertoires.Neurobiol Learn Mem 1998; 70: 119–36.
Hall JM, Shine JM, O’Callaghan C, Walton CC, Gilat M, NaismithSL, et al. Freezing of gait and its associations in the early andadvanced clinical motor stages of Parkinson’s disease: a cross-sec-tional study. J Parkinsons Dis 2015; 5: 881–91.
Hall JM, Shine JM, Walton CC, Gilat M, Kamsma YPT, Naismith SL,et al. Early phenotypic differences between Parkinson’s disease pa-tients with and without freezing of gait. Parkinsonism Relat Disord2014; 20: 604–7.
14 | BRAIN 2018: Page 14 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

Hallett M. The intrinsic and extrinsic aspects of freezing of gait. MovDisord 2008; 23: S439–43.
Hausdorff JM, Schaafsma JD, Balash Y, Bartels AL, Gurevich T,Giladi N. Impaired regulation of stride variability in Parkinson’sdisease subjects with freezing of gait. Exp Brain Res 2003; 149:187–94.
Herman T, Rosenberg-Katz K, Jacob Y, Giladi N, Hausdorff JM.Gray matter atrophy and freezing of gait in Parkinson’s disease: isthe evidence black-on-white? Mov Disord 2014; 29: 134–9.
Imamura K, Okayasu N, Nagatsu T. Cerebral blood flow and freezingof gait in Parkinson’s disease. Acta Neurol Scand 2012; 126:210–18.
Janssen AM, Munneke MAM, Nonnekes J, van der Kraan T,Nieuwboer A, Toni I, et al. Cerebellar theta burst stimulationdoes not improve freezing of gait in patients with Parkinson’sdisease. J Neurol 2017; 264: 963–72.
Killane I, Fearon C, Newman L, McDonnell C, Waechter SM, Sons K,et al. Dual motor-cognitive virtual reality training impacts dual-taskperformance in freezing of gait. IEEE J Biomed Heal Inf 2015; 19:1855–61.
Kim J, Criaud M, Cho SS, Dıez-Cirarda M, Mihaescu A, Coakeley S,et al. Abnormal intrinsic brain functional network dynamics inParkinson’s disease. Brain 2017; 140: 2955–67.
Kostic VS, Agosta F, Pievani M, Stefanova E, Jecmenica-Lukic M,Scarale A, et al. Pattern of brain tissue loss associated with freezingof gait in Parkinson disease. Neurology 2012; 78: 409–16.
Lefaivre SC, Almeida QJ. Can sensory attention focused exercise fa-cilitate the utilization of proprioception for improved balance con-trol in PD? Gait Posture 2015; 41: 630–3.
Leung IHK, Walton CC, Hallock H, Lewis SJG. Cognitive training inParkinson disease A systematic review and meta-analysis. Neurology2015; 85: 1843–51.
Lewis SJ, Barker RA. A pathophysiological model of freezing of gait inParkinson’s disease. Parkinsonism Relat Disord 2009; 15: 333–8.
Lewis SJ, Shine JM. The next step: a common neural mechanism forfreezing of gait. Neuroscientist 2016; 22: 72–82.
Lieberman A. Are freezing of gait (FOG) and panic related? J NeurolSci 2006; 248: 219–22.
Maidan I, Plotnik M, Mirelman A, Weiss A, Giladi N, Hausdorff JM.Heart rate changes during freezing of gait in patients withParkinson’s diseae. Mov Disord 2011; 25: 2346–54.
Matar E, Shine JM, Naismith SL, Lewis SJG. Using virtual reality toexplore the role of conflict resolution and environmental salience infreezing of gait in Parkinson’s disease. Parkinsonism Relat Disord2013; 19: 937–42.
Myers PS, McNeely ME, Koller JM, Earhart GM, Campbell MC.Cerebellar volume and executive function in Parkinson diseasewith and without freezing of gait. J Parkinsons Dis 2017; 7: 149–57.
Naismith SL, Shine JM, Lewis SJG. The specific contributions of set-shifting to freezing of gait in Parkinson’s disease. Mov Disord 2010;25: 1000–4.
Nanhoe-Mahabier W, Snijders AH, Delval A, Weerdesteyn V, DuysensJ, Overeem S, et al. Split-belt locomotion in Parkinson’s disease withand without freezing of gait. Neuroscience 2013; 236: 110–16.
Nantel J, de Solages C, Bronte-Stewart H. Repetitive stepping in placeidentifies and measures freezing episodes in subjects withParkinson’s disease. Gait Posture 2011; 34: 329–33.
Nichols TE, Holmes AP. Nonparametric permutation tests for func-tional neuroimaging experiments: a primer with examples. HumBrain Mapp 2002; 15: 1–25.
Nieuwboer A, Giladi N. Characterizing freezing of gait in Parkinson’sdisease: models of an episodic phenomenon. Mov Disord 2013; 28:1509–19.
Nieuwhof F, Bloem BR, Reelick MF, Aarts E, Maidan I, Mirelman A,et al. Impaired dual tasking in Parkinson’s disease is associated withreduced focusing of cortico-striatal activity. Brain 2017; 140: 1384–98.
Nieuwhof F, Helmich R. Entangled cerebral networks in Parkinson’sdisease. Brain 2017; 140: 2767–9.
Nutt JG, Bloem BR, Giladi N, Hallett M, Horak FB, Nieuwboer A.Freezing of gait: moving forward on a mysterious clinical phenom-enon. Lancet Neurol 2011; 10: 734–44.
Peterson D, Fling BW, Mancini M, Cohen RG, Nutt JG, Horak FB.Dual-task interference and brain structural connectivity in peoplewith Parkinson’s disease who freeze. J Neurol NeurosurgPsychiatry 2015; 86: 786–92.
Pieruccini-Faria F, Ehgoetz Martens KAE, Silveira CRA, Jones JA,Almeida QJ. Side of basal ganglia degeneration influences freezingof gait in Parkinson’s disease. Behav Neurosci 2015; 129: 214–18.
Pieruccini-Faria F, Jones JA, Almeida QJ. Motor planning inParkinson’s disease patients experiencing freezing of gait: the influ-ence of cognitive load when approaching obstacles. Brain Cogn2014; 87: 76–85.
Poldrack RA. The neural correlates of motor skill automaticity.J Neurosci 2005; 25: 5356–64.
Poldrack RA, Mumford JA, Nichols TE. Handbook of functional MRIanalysis. New York, NY: Cambridge University Press; 2011.
Sage MD, Almeida QJ. Symptom and gait changes after sensory at-tention focused exercise vs aerobic training in Parkinson’s disease.Mov Disord 2009; 24: 1132–8.
Schaafsma JD, Balash Y, Gurevich T, Bartels AL, Hausdorff JM,Giladi N. Characterization of freezing of gait subtypes and the re-sponse of each to levodopa in Parkinson’s disease. Eur J Neurol2003; 10: 391–8.
Schweder PM, Hansen PC, Green AL, Quaghebeur G, Stein J, AzizTZ. Connectivity of the pedunculopontine nucleus in parkinsonianfreezing of gait. Neuroreport 2010; 21: 914–16.
Shine JM, Koyejo O, Bell PT, Gorgolewski KJ, Poldrack RA.Estimation of dynamic functional connectivity using multiplicationof temporal derivatives. Neuroimage 2015; 122: 399–407.
Shine JM, Matar E, Bolitho SJ, Dilda V, Morris TR, Naismith SL,et al. Modeling freezing of gait in Parkinson’s disease with a virtualreality paradigm. Gait Posture 2013a; 38: 104–8.
Shine JM, Matar E, Ward PB, Bolitho SJ, Gilat M, Pearson M, et al.Exploring the cortical and subcortical functional magnetic resonanceimaging changes associated with freezing in Parkinson’s disease.Brain 2013b; 136: 1204–15.
Shine JM, Matar E, Ward PB, Frank MJ, Moustafa AA, Pearson M,et al. Freezing of gait in Parkinson’s disease is associated with func-tional decoupling between the cognitive control network and thebasal ganglia. Brain 2013c; 136: 3671–81.
Shine JM, Moustafa AA, Matar E, Frank MJ, Lewis SJG. The role offrontostriatal impairment in freezing of gait in Parkinson’s disease.Front Syst Neurosci 2013d; 7: 61.
Shine JM, Naismith SL, Palavra NC, Lewis SJG, Moore ST, Dilda V,et al. Attentional set-shifting deficits correlate with the severity offreezing of gait in Parkinson’s disease. Parkinsonism Relat Disord2013e; 19: 388–90.
Silveira CRA, Ehgoetz Martens KA, Pieruccini-Faria F, Bell-BoucherD, Roy EA, Almeida QJ. Disentangling perceptual judgment andonline feedback deficits in Parkinson’s freezing of gait. J Neurol2015; 262: 1629–36.
Snijders AH, Leunissen I, Bakker M, Overeem S, Helmich RC, BloemBR, et al. Gait-related cerebral alterations in patients withParkinson’s disease with freezing of gait. Brain 2011; 134: 59–72.
Snijders AH, Takakusaki K, Debu B, Lozano AM, Krishna V, FasanoA, et al. Physiology of freezing of gait. Ann Neurol 2016; 80:644–59.
Stefanova E, Jecmenica Lukic M, Ziropadja L, Markovic V, StojkovicT, Tomic A, et al. Attentional set-shifting in Parkinson’s diseasepatients with freezing of gait-acquisition and discrimination setlearning deficits at the background? J Int Neuropsychol Soc 2014;20: 929–36.
Szeto JYY, O’Callaghan C, Shine JM, Walton CC, Naismith SL,Halliday GM, et al. The relationships between mild cognitive im-pairment and phenotype in Parkinson’s disease. NPJ Parkinsons Dis2015; 1: 15015.
The network signature of freezing of gait BRAIN 2018: Page 15 of 16 | 15
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

Tahmasian M, Bettray LM, van Eimeren T, Drzezga A, TimmermannL, Eickhoff CR, et al. A systematic review on the applications ofresting-state fMRI in Parkinson’s disease: does dopamine replace-ment therapy play a role? Cortex 2015; 73: 80–105.
Tessitore A, Amboni M, Cirillo G, Corbo D, Picillo M, Russo A, et al.Regional gray matter atrophy in patients with Parkinson dis-ease and freezing of gait. AJNR Am J Neuroradiol 2012a; 33:1804–9.
Tessitore A, Amboni M, Esposito F, Russo A, Picillo M, Marcuccio L,et al. Resting-state brain connectivity in patients with Parkinson’sdisease and freezing of gait. Parkinsonism Relat Disord 2012b; 18:781–7.
Thevathasan W, Cole MH, Graepel CL, Hyam JA, Jenkinson N,Brittain JS, et al. A spatiotemporal analysis of gait freezing andthe impact of pedunculopontine nucleus stimulation. Brain 2012;135: 1446–54.
Vazquez A, Jimenez-Jimenez FJ, Garcia-Ruiz P, Garcia-Urra D. ‘Panicattacks’ in Parkinson’s disease. Acta Neurol Scand 1993; 87: 14–18.
Vercruysse S, Leunissen I, Vervoort G, Vandenberghe W, Swinnen S,Nieuwboer A. Microstructural changes in white matter associatedwith freezing of gait in Parkinson’s disease. Mov Disord 2015; 30:567–76.
Walton CC, Shine JM, Mowszowski L, Naismith SL, Lewis SJG.Freezing of gait in Parkinson’s disease: current treatments and thepotential role for cognitive training. Restor Neurol Neurosci 2014;32: 411–22.
Weiss A, Herman T, Giladi N, Hausdorff JM. New evidence for gaitabnormalities among Parkinson’s disease patients who suffer fromfreezing of gait: insights using a body-fixed sensor worn for 3 days.J Neural Transm 2015; 122: 403–10.
Wu T, Hallett M. A functional MRI study of automatic movements inpatients with Parkinson’s disease. Brain 2005; 128: 2250–9.
16 | BRAIN 2018: Page 16 of 16 K. A. Ehgoetz Martens et al.
Downloaded from https://academic.oup.com/brain/advance-article-abstract/doi/10.1093/brain/awy019/4850500by Sydney College of Arts useron 17 February 2018

APPENDIX(11(
Walton, C. C., Mowszowski, L. Gilat, M., Hall, J. M., O’Callaghan, C., Muller, A. J.,
Georgiades, M. J., Szeto, J. Y. Y., Ehgoetz Martens, K. A., Shine, J. M., Naismith, S.
L., Lewis, S. J. G. Cognitive training for freezing of gait in Parkinson’s disease: a
randomized controlled trial. NPJ Parkinson's Disease, 2018
!

ARTICLE OPEN
Cognitive training for freezing of gait in Parkinson’s disease: arandomized controlled trialCourtney C. Walton 1, Loren Mowszowski1,2, Moran Gilat1, Julie M. Hall1,3, Claire O’Callaghan1,4, Alana J. Muller1,Matthew Georgiades1, Jennifer Y. Y. Szeto1, Kaylena A. Ehgoetz Martens1, James M. Shine1,5, Sharon L. Naismith1,2 andSimon J. G. Lewis 1
The pathophysiological mechanism of freezing of gait (FoG) has been linked to executive dysfunction. Cognitive training (CT) is anon-pharmacological intervention which has been shown to improve executive functioning in Parkinson’s disease (PD). This studyaimed to explore whether targeted CT can reduce the severity of FoG in PD. Patients with PD who self-reported FoG and were freefrom dementia were randomly allocated to receive either a CT intervention or an active control. Both groups were clinician-facilitated and conducted twice-weekly for seven weeks. The primary outcome was percentage of time spent frozen during a TimedUp and Go task, assessed both on and off dopaminergic medications. Secondary outcomes included multiple neuropsychologicaland psychosocial measures. A full analysis was first conducted on all participants randomized, followed by a sample of interestincluding only those who had objective FoG at baseline, and completed the intervention. Sixty-five patients were randomized intothe study. The sample of interest included 20 in the CT group and 18 in the active control group. The primary outcome ofpercentage time spent frozen during a gait task was significantly improved in the CT group compared to active controls in the on-state. There were no differences in the off-state. Patients who received CT also demonstrated improved processing speed andreduced daytime sleepiness compared to those in the active control. The findings suggest that CT can reduce the severity of FoG inthe on-state, however replication in a larger sample is required.
npj Parkinson's Disease �(2018)�4:15� ; doi:10.1038/s41531-018-0052-6
INTRODUCTIONFreezing of gait (FoG) is a disabling symptom of Parkinson’sDisease (PD), which presents as a “brief, episodic absence ormarked reduction of forward progression of the feet, despite theintention to walk”.1 FoG is well-known to lead to falls2 and lowerquality of life, making it an important target for treatment.3 Thepathophysiological mechanism of FoG has been linked toexecutive dysfunction, particularly in aspects of cognitivecontrol,4 which aligns with neuroimaging evidence showingfronto-parietal and fronto-striatal impairments.5 Recent meta-analytic data suggests that cognitive training (CT) is aneffective6 and important7 behavioral intervention for improvingcognition, and in particular executive functions, in patientswith PD.Given that these executive deficits have been hypothesized to
underlie the pathophysiological mechanisms of FoG, it is plausiblethat reducing executive dysfunction via CT may lessen the severityof FoG, by mediating more effective fronto-striatal function.8 Anumber of studies have now shown that CT in PD can lead toneuroplastic changes by way of increased activity and functionalconnectivity in frontal-striatal regions.9–11 Given that FoG relatesto dysfunction in these areas, it is reasonable to hypothesize thatCT may facilitate more efficient processing between frontal andstriatal regions, leading to a reduction of FoG severity. Interestedreaders are directed to a previous review from our group, which
has provided more extensive evidence and rationale for thisproposal.12
In this study, a double-blind randomized controlled trial wasconducted to explore the efficacy of CT targeting executivefunctions in PD patients with FoG. We hypothesized thatparticipants receiving CT would show improvements as illustratedby the reduced severity of FoG after completion of theintervention. Additionally, we anticipated that secondary out-comes including cognitive and psychosocial measures wouldshow improvement following the CT program.
RESULTSParticipantsFigure 1 illustrates the flow of participants moving through thestudy. The first participant was randomized in April 2013 and thelast in June 2015. There were nine dropouts in the activecontrol condition (AC) (five prior to beginning the program) andone participant was lost to follow up. There were no dropouts inthe CT group. In addition, in the AC group one participant wasremoved entirely from the analysis as their diagnosis waschanged from PD to Progressive Supranuclear Palsy, and asecond was removed as they could not complete the TUGassessments due to severe motor disability. Two participantswere removed from the CT group due to inadvertent
Received: 17 January 2018 Revised: 11 April 2018 Accepted: 17 April 2018
1Parkinson’s Disease Research Clinic, Brain and Mind Centre, University of Sydney, Camperdown, NSW, Australia; 2Healthy Brain Ageing Program, Brain and Mind Centre & CharlesPerkins Centre, University of Sydney, Sydney, NSW, Australia; 3School of Social Sciences and Psychology, Western Sydney University, Sydney, NSW, Australia; 4Department ofPsychiatry and Behavioural and Clinical Neuroscience Institute, University of Cambridge, Cambridge, UK and 5School of Psychology, Stanford University, Palo Alto, CA, USACorrespondence: Simon J. G. Lewis ([email protected])
www.nature.com/npjparkd
Published in partnership with the Parkinson’s Foundation

incorrect randomization. These participants were retrospec-tively identified as not meeting the original inclusion criteria andthus it was determined their data would not be analysed atany point.
Upon intervention completion, TUG scoring indicated thatdespite self-reporting FoG, nine participants from the CT and fourfrom the AC group did not objectively exhibit FoG on baselineassessment. Therefore, we designed two analysis populations
Fig. 1 CONSORT Flow diagram
Cognitive training for freezing of gaitCC Walton et al.
2
npj Parkinson’s Disease (2018) �15� Published in partnership with the Parkinson’s Foundation
1234
56789
0():,;

post-hoc. The full analysis set (FAS) included all participantsrandomized into the study, whether or not they dropped out, orshowed objective FoG at BL. The sample of interest set (SIS) wasdecided upon as a post-hoc analysis, to account for the fact that anumber of participants did not display FoG at baseline. The SIStherefore included only those participants who showed objectiveFoG on BL testing, as identified in the baseline TUG, andcompleted the study in full. The SIS population is consideredthe analysis of interest and is therefore what is reported in theresults, however all analyses were initially run on the FAS sampleto confirm no sampling bias. Demographic data for both the FASand SIS samples are provided in Table 1.
Primary outcomeResults of the SIS analysis for the primary outcome are displayedin Table 2. This analysis showed that patients in the CT groupshowed a large and statistically significant reduction in FoGseverity in the on-state compared to participants in the AC. Therewas no difference in the off-state. The FAS analysis was consistentwith these results, suggesting that the SIS sample was not biased.We did not compare performance across each of the fourconditions separately, on and off as this was not part of thepredetermined outcome plan, and secondly, we felt it was aninappropriate exploratory analysis given the smaller than antici-pated sample.
Secondary outcomesResults of the SIS analysis for the secondary outcomes aredisplayed in Table 3. In the SIS analysis, there were no statisticallysignificant differences for any of the secondary outcomes overtime between groups.
Covariate analysisThe covariate analysis showed that the results for the primaryoutcome remained unchanged by the introduction of covariates(i.e., still statistically significant). However, in terms of secondaryoutcomes, the inclusion of DDE as a covariate led to TMT-A anddaytime sleep disturbance scores becoming significant, with thosein the CT group improving compared to the AC.
DISCUSSIONThis pilot study represents one of the largest RCTs of CT to date inPD. Though interpretation of the results must remain cautiousowing to the limitations outlined below, the results allude to thepotential for CT to reduce the severity of FoG in people with PD.We showed that CT led to a large and significant reduction of FoGseverity compared to AC while in the on-state, but this was notreplicated in the off-state. These results were consistent, whetherwe included participants who did not display FoG at baseline ornot, and when accounting for covariates. We suggest these resultswarrant larger scale replication, employing the suggestedmethodological adjustments we provide below.The result of FoG only improving during the on-state is
noteworthy. Firstly, we preface this discussion by stating this isthe clinically relevant behavioral state, as patients in their day-to-day life would take dopaminergic medications as prescribed tominimize time in the off-state. Our provisional supposition toexplain this result is that participants in the off-state were tooimpaired to benefit from any of the potential changes initiatedthrough CT. Training was expected to impact frontal processing
Table 1. Demographic data of participants in both analysis samples
FAS population SIS population
AC group (N= 32)
CT group(N= 29)
AC group(N= 18)
CT group(N= 20)
Gender (M/F) 22/10 21/8 11/7 14/6
Age, years 68.50 (7.5) 68.48 (8.0) 69.61 (7.8) 69.70 (7.6)
Years sincediagnosis
11.06 (6.6) 8.82 (4.9) 11.89 (6.6) 9.95 (4.4)
LEDD 934.71 (555.1) 769.37(340.7)
975.43(570.8)
828.8(315.3)
Education, years 13.97 (3.2) 13.59 (3.2) 14.44 (3.5) 13.55 (3.4)
MMSE 28.16 (1.8) 27.72 (2.0) 28.56 (1.6) 27.35 (2.0)
Sessionsattended
11.58 (3.7) 13.31 (0.9) 13.06 (1.2) 13.4 (0.9)
Days until followup
8.91 (6.9) 6.48 (5.6) 9.11 (7.4) 7.20 (5.7)
Has DBS 6 4 3 4
MDS-UPDRSMotor on
36.16 (13.7) 36.83 (13.3) 33.44 (12.6) 38.70 (13.7)
MDS-UPDRSMotor off
43.76(11.7) 42.12 (12.4) 43.27 (11.8) 46.13 (10.9)
Hoehn and Yahr stage
1 1 0 1 0
2 12 11 5 5
2.5 6 5 5 5
3 11 9 6 7
4 2 4 1 3
LEDD levadopa equivalence daily dose, MMSE mini-mental state examina-tion, DBS deep brain stimulation, MDS-UPDRS Movement Disorder SocietyUnified Parkinson’s Disease Rating Scale
Table 2. Primary outcome data between groups before and after intervention
AC group CT group Comparison of change
%TF ona
BL 6.61 (3.92, 11.15) 9.16 (5.52, 15.19) 0.3 (0.14, 0.62); T=−3.36; p= 0.002; d= 1.02
FU 11.99 (7.11, 20.23) 4.95 (2.99, 8.21)
Change from BL 1.81 (1.08, 3.05) 0.54 (0.32, 0.90)
%TF offa
BL 16.61 (9.22, 29.93) 8.02 (4.59, 13.98) 1.07 (0.64, 1.76); T= 0.26; p= 0.800; d= 0.08
FU 15.44 (8.57, 27.81) 7.94 (4.55, 13.85)
Change from BL 0.93 (0.64, 1.34) 0.99 (0.70, 1.40)
aAnalysis conducted on Log10 transformed data and represented by geometric means. The change from baseline is a ratio of the geometric means at follow-up compared with baseline. The comparison of the changes from baseline is the ratio of the change from baseline for the CT group compared the AC group.95% CIs are presented in brackets. Direction of change and statistical significance was matched in the FAS analysis
Cognitive training for freezing of gaitCC Walton et al.
3
Published in partnership with the Parkinson’s Foundation npj Parkinson’s Disease (2018) �15�

Table 3. Secondary outcome data between groups before and after intervention
AC group CT group Comparison of change
MOCA
BL 25.33 (23.57, 27.09) 24.15 (22.48, 25.82) 0.84 (−1.38, 3.06); T= 0.77;
FU 25.39 (23.63, 27.15) 25.05 (23.38, 26.72) p= 0.45;
Change from BL 0.06 (−1.55, 1.67) 0.9 (−0.63, 2.43) d= 0.24
HVLT-R total
BL 19.39 (16.83, 21.95) 18.50 (16.07, 20.93) −1.93 (−4.75, 0.88);
FU 21.72 (19.16, 24.28) 18.90 (16.47, 21.33) T=−1.39; p= 0.17;
Change from BL 2.33 (0.29, 4.38) 0.40 (−1.54, 2.34) d= 0.44
HVLT-R delayed
BL 5.67 (4.23, 7.10) 5.00 (3.64, 6.36) −0.53 (−2.21, 1.15);
FU 6.35 (4.89, 7.80) 5.15 (4.49, 6.751) T=−0.64; p= 0.53;
Change from BL 0.68 (−0.55, 1.92) 0.15 (−0.99, 1.29) d= 0.19
Digit span total
BL 16.00 (14.41, 17.59) 14.85 (13.34, 16.36) −0.93 (−2.18, 0.32);
FU 16.78 (15.19, 18.37) 14.70 (13.19, 16.21) T=−1.50; p= 0.14;
Change from BL 0.78 (−0.13, 1.69) −0.15 (−1.01, 0.71) d= 0.47
TMT-A timea,b
BL 44.74 (35.40, 56.48) 50.31 (40.10, 63.05) 0.78 (0.64, 0.95);
FU 45.67 (36.14, 57.64) 39.77 (31.65, 49.90) T=−2.57; p= 0.01;
Change from BL 1.02 (0.89, 1.18) 0.79 (0.69, 0.91) d= 0.62
TMT-B timea
BL 119.11 (95.63, 148.31) 136.08 (110.52, 167.51) 0.88 (0.75, 1.04);
FU 120.96 (97.11, 150.61) 122 (99.12, 150.29) T=−1.50; p= 0.14;
Change from BL 1.02 (0.90, 1.15) 0.90 (0.80, 1.01) d= 0.48
VF-phonemic
BL 37.22 (32.26, 42.19) 31.55 (26.84, 36.26) −0.47 (−3.96, 3.03);
FU 37.89 (32.93, 42.85) 31.75 (27.04, 36.46) T=−0.27; p= 0.79;
Change from BL 0.67 (−1.87, 3.2) 0.20 (−2.21, 2.61) d= 0.09
VF-semantic
BL 33.22 (28.46, 37.99) 30.45 (25.93, 34.72) −1.03 (−5.28, 3.23);
FU 34.00 (29.23, 38.77) 30.20 (25.68, 34.72) T=−0.49; p= 0.63;
Change from BL 0.78 (−2.31, 3.87) −0.25 (−3.18, 2.68) d= 0.15
VF-switching total
BL 12.11 (10.54, 13.68) 9.70 (8.21, 11.19) 1.18 (−0.51, 2.87);
FU 11.78 (10.20, 13.35) 10.55 (9.06, 12.04) T= 1.42; p= 0.16;
Change from BL −0.33 (−1.56, 0.89) 0.85 (−0.31, 2.01) d= 0.45
VF-switching accuracy
BL 10.83 (9.04, 12.63) 8.90 (7.20, 10.60) −0.09 (−2.54, 2.35);
FU 10.28 (8.48, 12.07) 8.25 (6.55, 9.95) T= 0.08; p= 0.94;
Change from BL −0.56 (−2.33, 1.22) −0.65 (−2.33, 1.03) d= 0.02
SDMT total
BL 35.44 (30.00, 40.89) 33.95 (28.78, 39.12) 0.47 (−3.74, 4.68);
FU 37.22 (31.77, 42.67) 36.20 (31.03, 41.37) T= 0.23; p= 0.82;
Change from BL 1.78 (−1.28, 4.83) 2.25 (−0.65, 5.15) d= 0.07
AGN latency
BL 595.75 (547.92, 643.58) 657.65 (608.42, 706.89) −29.89 (−66.37, 6.58);
FU 586.42 (538.89, 633.95) 618.43 (569.19, 667.67) T=−1.67; p= 0.10;
Change from BL −9.33 (−34.34, 15.68) −39.22 (−65.78, −12.67) d= 0.62
AGN omissionsa
BL 2.85 (1.49, 4.94) 4.35 (2.42, 7.37) 0.70 (0.46, 1.06);
FU 4.26 (2.42, 7.08) 4.15 (2.29, 7.05) T=−1.73; p= 0.09;
Change from BL 1.37 (1.03, 1.82) 0.96 (0.71, 1.3) d= 0.61
AGN commissionsa
Cognitive training for freezing of gaitCC Walton et al.
4
npj Parkinson’s Disease (2018) �15� Published in partnership with the Parkinson’s Foundation

and also occurred in the on-state. In the dopamine depleted state,it is conceivable that the striatal dysfunction overshadowed anybenefit of CT,8,13 and FoG could not be improved. Our futureanalysis of functional neuroimaging outcomes in this study maybe able to unravel this further.A number of trials in older adults have now shown that CT can
have a beneficial impact on multiple gait parameters.14–16 In PDspecifically, a pilot study by Milman and colleagues17 showed that12 weeks of CT could improve TUG performance. Unfortunately,this pilot study did not employ a control group. Therefore, thecurrent results are an important extension showing improvementson TUG performance via the reduction of FoG, compared to anactive control group.Given that FoG was reduced in the CT group, we expected there
to be additional improvements in tests of EF, which werepresumed to underlie any improvement of FoG. However, changeson these outcomes did not reach statistical significance. We weretherefore unable to confirm the hypothesis that improving EFwould be the driver of reduced severity of FoG. It is possible thatour smaller sample size was a factor however (though it wasdeemed inappropriate to conduct post-hoc power analysis18).Indeed, there were near medium-sized effect sizes (d ≥ 0.45) formany of the executive tests we anticipated improvements onincluding TMT-B, and shift-measures of the affective go-no-go test(AGN) and verbal fluency (VF) (see Table 2). We do note howeverthat when adjusting for the effect of dopaminergic medication,the CT group did show medium-sized significant improvementscompared to AC in processing speed and daytime sleepiness.
LimitationsThis study has limitations which warrant consideration. The first isthat we did not meet the projected sample size target due tofeasibility issues with recruitment. In addition, there were anumber of dropouts in the AC group, although we note that overhalf of these dropouts occurred prior to commencement of theintervention and only one was due a lack of interest. The thirdissue was that participants were randomized prior to TUG scoring.This was necessary to avoid delay to participants being enrolled asTUG scoring is time consuming, requiring skilled and trainedraters. Nonetheless, we attempted to address these limitations byrunning the FAS analysis, which confirmed our primary result.Additionally, despite random allocation, the groups were
unbalanced in their baseline FoG severity. The CT group hadmore FoG than the AC in the on and less in the off-state. Westratified the randomization by cognitive functioning (MOCAscores), however it may be more appropriate in future to stratifyby objective FoG scores at baseline. We highlight however thatthe results remained when accounting for the impact of LEDD,and that the on-state is the clinically meaningful state. Related tothis unbalanced severity, it is important to highlight that the ACgroup actually had worse FoG at follow-up compared to the CTgroup in the on-state and this pattern of results was replicated inthe FAS analysis. Replication with a larger sample is needed todemonstrate if this is a reliable finding, or represents the variabilityfound in small samples such as this one.
Table 3 continued
AC group CT group Comparison of change
BL 5.05 (3.35, 7.41) 4.28 (2.76, 6.42) 0.83 (0.54, 1.3);
FU 5.15 (3.45, 7.51) 3.48 (2.19, 5.30) T=−0.83; p= 0.41;
Change from BL 1.02 (0.75, 1.38) 0.85 (0.62, 1.17) d= 0.26
HADS total
BL 10.78 (7.85, 13.71) 12.70 (9.92, 15.48) −1.98 (−5.38, 1.42);
FU 11.56 (8.63, 14.48) 11.50 (8.72, 14.28) T=−1.18; p= 0.25;
Change from BL 0.78 (−1.69, 3.24) −1.2 (−3.54, 1.14) d= 0.37
PDQ-39 total
BL 26.69 (20.81, 32.58) 28.21 (22.63, 33.79) −3.33 (−8.07, 1.41);
FU 27.32 (21.43, 33.20) 25.50 (19.92, 31.09) T=−1.42; p= 0.16;
Change from BL 0.62 (−2.82, 4.06) −2.71 (−5.97, 0.55) d= 0.45
CBI total
BL 18.74 (10.62, 26.86) 33.51 (26.21, 40.81) −4.51 (−10.66, 1.64);
FU 20.85 (12.66, 29.04) 31.11 (23.86, 38.36) T=−1.49; p= 0.15;
Change from BL 2.11 (−2.52, 6.73) −2.4 (−6.45, 1.65) d= 0.53
SCOPA-S dayb
BL 4.77 (3.16, 6.38) 6.31 (4.72, 7.89) −1.66 (−3.27, −0.05);
FU 5.00 (3.39, 6.62) 4.88 (3.32, 6.44) T=−2.10; p= 0.04;
Change from BL 0.24 (−0.91, 1.38) −1.43 (−2.56, −0.29) d= 0.56
SCOPA-S night
BL 5.33 (3.18, 7.48) 6.00 (3.94, 8.06) −0.24 (−2.17, 1.68);
FU 5.61 (3.46, 7.76) 6.03 (3.97, 8.09) T=−0.26; p= 0.80;
Change from BL 0.28 (−1.09, 1.64) 0.03 (−1.32, 1.39) d= 0.06
MOCA Montreal cognitive assessment, HVLT-R Hopkins verbal learning test-revised, WMS-III Wechsler Memory Scale, VF verbal fluency, DKEFS Delis–Kaplanexecutive function system, SDMT symbol digit modalities test, TMT trail making test, AGN affective go-no-go test, HADS Hospital Anxiety & Depression Scale,SCOPA Scales for Outcomes in Parkinson’s disease, PDQ-39 the Parkinson’s disease questionnaire, CBI-R Cambridge behavioral inventory-revisedaAnalysis conducted on Log10 transformed data and represented by geometric means. The change frombaseline is a ratio of the geometric means at follow-up compared with baseline. The comparison of the changes from baseline is the ratio of the change frombaseline for the CT group compared the AC groupbResults are adjusted for DDE as a covariate. 95% CIs are presented in brackets
Cognitive training for freezing of gaitCC Walton et al.
5
Published in partnership with the Parkinson’s Foundation npj Parkinson’s Disease (2018) �15�

Future directionsWe believe there is reason to be hopeful for the use of these trialsin the future. Feedback from participants and family membersinvolved in the groups was overwhelmingly positive, our pilotresults highlight positive trends, and the importance of non-pharmacological trials including CT has become increasinglyclear.7,19,20 We suggest that replication of this trial is warranted.However, with the hope of improving any future work learningfrom some of the issues that were raised during this study, wesuggest authors consider some of the following suggestions.Future studies where possible should aim to score FoG severity
prior to enrollment. A certain threshold for severity (e.g., >5%)should be specified for eligibility, and stratification across groupscould also be based on this. Where possible, additional methodsof FoG measurement could increase the reliability of %FoG scores.This could be done through measures such as gait mats andaccelerometry data, and also repeat TuG assessments to addressmeasurement variability. Multisite recruitment would increase thepotential for sample size without relying on home-based CT,which we do not believe would be a viable option,21 particularly inthis sample. The inclusion of additional data to aid analysis such asmeasurement of expectancy effects and CT training data can beuseful. Finally, we did not include a long-term follow upassessment. This has often been used as a criticism against CT,though we rebut that very few interventions elicit sustainedimprovements after the cessation of treatment. Thus, it is likelythat clinically, CT needs to be continuously delivered in order tocontinue any found benefits, just like most other interventions(e.g., exercise, medications etc). Nevertheless, obtaining a betterunderstanding of how long such results are maintained22 is usefulfor future trial design and clinical applications, and thus futurestudies could try to obtain this information if feasible.
CONCLUSIONSThe current study provides preliminary evidence that CT canreduce the severity of FoG in PD during the on-state. Thisimprovement was seen without concurrent, significant changes toexecutive functioning (despite near-medium sized effects on thesemeasures), but in the context of improved processing speed anddaytime sleep disturbance. Despite the limitations of this study,these results add to the growing body of evidence showing thatCT is a useful therapeutic technique worthy of continuedexploration in PD.
METHODSStudy registrationThis study was registered in 2013 on the 5 April through the Australian andNew Zealand Clinical Trials Registry (ACTRN12613000359730) and wasapproved by the Human Research Ethics Committee of the University ofSydney. Written informed consent was obtained from all participants.
EligibilityEligible participants were those diagnosed with idiopathic Parkinson’sdisease based on the UK Brain Bank clinical criteria,23 with self-reportedFoG at the time of assessment, and who were free from dementia asdetermined by a score of ≥24 on the mini-mental state examination(MMSE).24
RecruitmentThe study was advertised in a local PD community magazine as well aslocal PD community support groups. Potential participants were alsorecruited from the Parkinson’s Disease Research Clinic at the Brain andMind Centre, University of Sydney. Interested participants were invited toparticipate if they had previously reported a positive score on Question 3of the Freezing of Gait Questionnaire (FOG-Q): “do you feel that your feet
get glued to the floor while walking, making a turn, or trying to initiatewalking (freezing)?”.25
Prior to recruitment, we used baseline data from a previously publishedtrial26 to conduct a power analysis using a conservative effect size estimateof at least 0.2 in the study’s primary outcome. This suggested the minimumsample size required for each group was 39 (based on power= 0.80 and α= 0.05).
Study designThe study was a double-blind randomized active controlled trial. Interestedpatients were enrolled by CCW and LM after they met eligibility criteriaduring a baseline assessment and were then randomized into either the CTor an AC group. Conditions were masked as either “morning” or“afternoon” sessions, and the order of these was randomized betweenrecruitment waves prior to trial commencement. In order to facilitateblinding, participants were told that each session involved differentcomputerized activities, but were not explicitly told of a treatment orcontrol group. Randomization of participants and morning/afternoonsessions was carried out using a randomly generated number sequenceallocated by a blinded researcher not involved in trial recruitment, datagathering, assessments or training. Randomization was undertaken usingpermuted blocks and stratified by cognitive functioning, with stratadefined by Montreal cognitive assessment (MOCA) scores of <26 or ≥26.Participants were advised of their allocation into the morning or afternoonsession by way of sealed opaque envelopes delivered by CCW uponcompletion of the baseline assessments. Post-intervention assessmentswere conducted by clinicians who were blinded to treatment allocation. Allparticipants allocated to the AC group were offered the opportunity tocomplete CT after their involvement in the trial was complete. Tenparticipants elected to complete this, with those who declined citing timecommitments as the primary reason.
AssessmentsBaseline and post-intervention assessments were each completed in twoparts: on and off-medications. The on-state assessment included aneuropsychological test battery, psychosocial measures, part III of theMovement Disorders Society’s revision of the Unified Parkinson’s DiseaseResearch Scale (MDS-UPDRS),27 and a modified timed up-and-go (TUG)task. These assessments were completed in a random counterbalancedorder and took approximately 2.5 h to complete. Baseline assessmentswere conducted within 3 weeks prior to training commencement, andfollow-up assessment was within 3 weeks of the intervention finishing.The practically defined off-state assessment was completed in themorning on a different day when participants were asked not to taketheir usual Parkinson’s medication until after the assessment, andcomprised a repeated MDS-UPDRS part III and TUG, taking approximately1 h. Those with deep brain stimulation did not complete the off-stateassessment. A random subset of participants also underwent neuroima-ging, however the investigation of any training-induced changes are atertiary outcome and are therefore not included in the currentmanuscript.
Primary outcomeThe primary outcome was percentage of time spent frozen (%TF) across allfour trials of a TUG assessment. Percentage was chosen as the primaryoutcome as it was anticipated to be more sensitive than a reduction on theFOG-Q, whilst accounting for inter-individual variability in gait speed andthe variable duration of freezing episodes across TUG tasks.26 In each task,the participant was requested to get up from a chair, walk to a square boxshape taped to the floor five meters ahead and complete both a left and aright turn (see Fig. 2). TUGs were video recorded and then scoredindependently post-assessment. Six scorers (MG, JMH, AJM, MG, JYYS, andKAEM) were randomly distributed videos of the TUGs. Scorers were givenbaseline and follow-up TUG videos in a random order for the sameparticipant, to minimize pre-post scoring variability. FoG was tagged in thevideo at any point when a participant made a paroxysmal and involuntarycessation of normal progression of the feet through the task. This includeda typical trembling of the feet, short shuffling steps of a few centimetres inlength or a complete motor block.28
The %TF outcome was calculated by summing all FoG episodes acrossthe four conditions, and dividing by the total time to complete across allconditions. Inter-rater variability amongst blinded scorers was strong, andcalculated by all scorers being given a random selection of the same six
Cognitive training for freezing of gaitCC Walton et al.
6
npj Parkinson’s Disease (2018) �15� Published in partnership with the Parkinson’s Foundation

videos to independently score. The intraclass correlation coefficient was.902 across all FoG episodes (average %TF: 9.56%). We note that two of thesix videos by chance did not contain FoG, however they were included toconfirm no false-positive scoring had occurred. As this inflated reliabilityacross scorers however, we re-calculated the coefficient with the twovideos removed to be sure. Scoring was still accurate across raters (.865)(average %TF: 14.31%).
Secondary outcomesCognitive assessment. To assess global functioning for descriptivepurposes, the MMSE24 and MOCA29 were used. Total and delayed Hopkinsverbal learning test-revised scores were used to assess verbal memory.30
To assess attention and working memory, the total score from the DigitSpan subtest of the Wechsler Memory Scale was used.31 To assess verbalfluency, the total words generated on each condition of the VF subtest ofthe Delis–Kaplan executive function system was used.32 In this test, part 1measures phonemic fluency across three letters, part 2 measures semanticfluency across two categories while parts 3 and 4 assess switchingbetween two differing categories. Part 3 represents the total number ofcorrect items, while part 4 represents the total number of correct switches.Processing speed was assessed by the number of correct responses on theoral symbol digit modalities test.33 For these measures, higher scores areindicative of better performance.To assess processing speed and cognitive flexibility, times for parts A
and B respectively of the trail making test (TMT) were used.34 Finally, theAGN of the Cambridge neuropsychological test automated battery35 wasused as a computerized measure of inhibitory control/switching. Meanlatency post-switch was used to determine performance in addition to thenumber of commissions or omissions. For these tests, lower scores wereindicative of better performance. Alternate versions of each test with theexception of Digit Span (not available) were used at baseline and follow-upto minimise potential practice effects.
Psychosocial measures. Participants completed several questionnairestargeting mood and wellbeing. The Hospital Anxiety & Depression Scalewas used to assess anxious and depressive symptoms.36 The Scales ofOutcomes in PD (SCOPA)-Sleep was used to assess sleep quality in terms ofboth daytime sleepiness and night time sleep disturbance.37 TheParkinson’s Disease Questionnaire (PDQ-39)38 was used as a measure ofquality of life. Finally, if a participant lived with a carer, the Cambridgebehavioral inventory-revised39 was used as an informant report of
cognitive and behavioral changes. For all of these scores, a higher scorewas indicative of more substantial impairment.
Intervention. Both the CT and AC groups attended sessions at the Brainand Mind Centre at the University of Sydney in our designated CTlaboratory, and both conditions were matched in terms of time, cliniciancontact, computer use and social interaction. In accordance with ourprevious CT programs for older adults (see,40,41) the intervention wascompleted in a group format (n ≤ 10), and comprised of 2-h sessions, twiceweekly over 7 weeks (14 sessions in total). Both groups were supervisedand facilitated by CCW & LM. The first hour of the session was identicalacross CT and AC groups: (i) 30–45min was designated to psycho-education on a number of topics relevant to PD including cognition, sleepand mood, and was delivered by multidisciplinary specialists andresearchers from the Brain and Mind Centre; (ii) participants then tookan enforced break of 10–15min. This first hour, whilst not CT per-se, wasincluded for both conditions as a means of increasing participantengagement and has previously been shown to support our excellentprogram adherence rates.40,41 The second hour of the session differedacross CT and AC groups:(A) CT: Participants in this group completed a program of computerized
CT tasks, selected for their focus on executive functions and on the basis ofour previous experience employing the “Neuropsychological EducationalApproach to Remediation” approach41,42 in providing computerized CT to>400 older adults,41,43,44 including those with PD.40 Tasks includeddesignated “brain training” programs (e.g., Attention Process Training-III45) as well as commercially-available software (e.g., computerisedSudoku), which were determined by clinical neuropsychologists (LM,SLN) to target the cognitive processes of most interest to FoG (inhibitorycontrol, attentional set-shifting, working memory, processing speed andvisuospatial skills).4,12 Performance was monitored by the facilitators, withthe focus of progressively making the tasks more difficult where possiblefor the participant. These changes were done in an individualized mannerbased on performance and in consultation with the participant. Thereforewhile the tasks delegated for each session where standardized across allparticipants, there were differences in how far each progressed in terms ofdifficulty. The majority of exercises provided the participant with feedbackin the form of scores, and this was further discussed between facilitatorsand participants to help them better understand their performance.(B) AC: Participants in this group completed a series of non-specific
computer-based tasks including predominantly watching informativenature videos and answering content-related questions as previously
Fig. 2 Each condition involved two trials with a left and right turn version. In the 180° condition, the participant walked to the box, turnedaround and returned to their chair; In the 540° condition, they completed a 540° turn in the box before returning to the chair; In the boxcondition, participants shuffled around the box, keeping their inside foot to the outside of the box; in the dual task condition, participants didthe same as in the 180°, however completed a cognitive task as they walked. This was either naming the months backwards or multiples of 9or 7 aloud. The %TF outcome was calculated by summing all FoG episodes across the four conditions, and dividing by the total time tocomplete across all conditions
Cognitive training for freezing of gaitCC Walton et al.
7
Published in partnership with the Parkinson’s Foundation npj Parkinson’s Disease (2018) �15�

used,46 as well as online “treasure hunts” devised by our team. These taskswere designed to provide broader, generalized cognitive engagementcompared to the targeted focus on specific cognitive functions in the CTcondition. Therefore, those in this group were not expected to havereduced FoG severity but were rather intended to match for clinician andpeer contact, along with computer use.
Statistical analysis. To minimize any potential bias, statistical analyseswere conducted by a consultant statistician experienced in RCTs who wasbased at the University of Sydney (see acknowledgements) and who wasnot involved in any other aspect of the trial. Data was analysed using SASsoftware version 9.4.The analysis took the form of a mixed effects model using fixed effects
fitted to all endpoints, to test the null hypothesis of no difference inchange over time across groups against the alternative hypothesis of adifference between the two arms. An additional term in the model wasfitted to account for the repeated measures pre- and post-intervention.“Participant” was declared a random effect. An unstructured covariancepattern between baseline and post-intervention was used. The Kenwardand Roger’s method for correcting for the fixed effects, standard error biasby inflation of the variance and Satterthwaite’s adjustment to the degreesof freedom has also been applied to cater for the small sample size.Analyses of endpoints with non-normal variance for analysis have beentransformed to the log10 scale for analysis. Results were back transformedfor interpretation and represent geometric means. Cohens d wascalculated as a measure of effect size with 0.2, 0.5, and 0.8 consideredsmall, medium and large effects in the CT compared to the AC group.47
Covariate analysis. An additional analysis was undertaken to investigatethe effects of the following covariates on all outcome measures: age,education, levodopa equivalency daily dose LEDD;48 years since diagnosis,and the amount of days between CT completion and FU assessment (daysuntil FU) on each outcome. Only significant results are reported.
Data availability statementThe data that support the findings of this study are available from thecorresponding author upon reasonable request.
ACKNOWLEDGEMENTSWe would like to thank the participants and their family members who gave us theirtime to take part in this study. We thank Caro-Anne Badcock AStat, an independentstatistical consultant at The University of Sydney who conducted the statisticalanalysis for this study. We would like to thank Dr Zoe Terpening and Dr Paul Cloustonfor their assistance with assessments in addition to Ms Amelia English and Ms NicoleCockayne for their assistance with randomization during the study. This study wassupported by Michael J Fox Foundation Grant (#4669.01). C.C.W. was supported by anAustralian Postgraduate Award at the University of Sydney; L.M. is supported by aAustralian Research Council Dementia Research Development Fellowship (1109618);M.G. is supported by a University of Sydney International Scholarship; J.M.H. issupported by a Western Sydney University Postgraduate Research Award; C.O. issupported by a National Health and Medical Research Council Neil Hamilton FairleyFellowship (1091310); M.G. is supported by the RA Money Postgraduate ResearchScholarship in Neuroscience at the University of Sydney; K.A.E.M. is supported by aPostdoctoral Fellowship from Parkinson’s Canada; J.M.S. is supported by a NationalHealth and Medical Research Council CJ Martin Fellowship (1072403), S.L.N. issupported by a National Health and Medical Research Career Development Award(1008117); S.J.G.L. is supported by a National Health and Medical Research CouncilPractitioner Fellowship (1003007); A.J.M. and J.Y.Y.S. report no financial disclosures.
AUTHOR CONTRIBUTIONSC.C.W., L.M., J.M.S., S.L.N., S.J.G.L. were involved in conceptualization and design of thestudy. C.C.W. and L.M. were responsible for delivery of the intervention. C.C.W., L.M.,M.G., J.M.H., C.O.C., A.J.M., M.G., J.Y.Y.S., K.A.M., and J.M.S. were involved in datacollection and/or blinded scoring. C.C.W. was responsible for data collation, andpreparation for analysis. All authors contributed to and approved the finalmanuscript.
ADDITIONAL INFORMATIONCompeting interests: The authors declare no competing interests.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claimsin published maps and institutional affiliations.
REFERENCES1. Nutt, J. G. et al. Freezing of gait: moving forward on a mysterious clinical phe-
nomenon. Lancet Neurol. 10, 734–744 (2011).2. Gray, P. & Hildebrand, K. Fall risk factors in Parkinson’s disease. J. Neurosci. Nurs.
32, 222–228 (2000).3. Walton, C. C. et al. The major impact of freezing of gait on quality of life in
Parkinson’s disease. J. Neurol. 262, 108–115 (2015).4. Heremans, E. et al. Cognitive aspects of freezing of gait in Parkinson’s disease: a
challenge for rehabilitation. J. Neural Transm. 120, 543–557 (2013).5. Fasano, A., Herman, T., Tessitore, A., Strafella, A. P. & Bohnen, N. I. Neuroimaging
of freezing of gait. J. Parkinson’s Dis. https://doi.org/10.3233/JPD-150536 (2015).6. Leung, I. H. et al. Cognitive training in Parkinson disease: a systematic review and
meta-analysis. Neurology 85, 1843–1851 (2015).7. Ventura, M. I., Edwards, J. D. & Barnes, D. E. More than just a movement disorder:
Why cognitive training is needed in Parkinson disease. Neurology. https://doi.org/10.1212/wnl.0000000000002158 (2015).
8. Shine, J. M., Moustafa, A. A., Matar, E., Frank, M. J. & Lewis, S. J. The role offrontostriatal impairment in freezing of gait in Parkinson’s disease. Front. Syst.Neurosci. 7, 61 (2013).
9. Diez-Cirarda, M. et al. Increased brain connectivity and activation after cognitiverehabilitation in Parkinson’s disease: a randomized controlled trial. Brain ImagingBehav. https://doi.org/10.1007/s11682-016-9639-x (2016).
10. Cerasa, A. et al. Neurofunctional correlates of attention rehabilitation in Parkin-son’s disease: an explorative study. Neurol. Sci. https://doi.org/10.1007/s10072-014-1666-z (2014).
11. Nombela, C. et al. Cognitive rehabilitation in Parkinson’s disease: evidence fromneuroimaging. Front. Neurol. 2, 82 (2011).
12. Walton, C. C., Shine, J. M., Mowszowski, L., Naismith, S. L. & Lewis, S. J. Freezing ofgait in Parkinson’s disease: current treatments and the potential role for cognitivetraining. Restor. Neurol. Neurosci. 32, 411–422 (2014).
13. Lewis, S. J. & Barker, R. A. A pathophysiological model of freezing of gait inParkinson’s disease. Park. Relat. Disord. 15, 333–338 (2009).
14. Li, K. Z. et al. Benefits of cognitive dual-task training on balance performance inhealthy older adults. J. Gerontol. A. Biol. Sci. Med. Sci. 65, 1344–1352 (2010).
15. Verghese, J., Mahoney, J., Ambrose, A. F., Wang, C. & Holtzer, R. Effect of cognitiveremediation on gait in sedentary seniors. J. Gerontol. A. Biol. Sci. Med. Sci. 65,1338–1343 (2010).
16. Steinmetz, J. P. & Federspiel, C. The effects of cognitive training on gait speed andstride variability in old adults: findings from a pilot study. Aging Clin. Exp. Res. 26,635–643 (2014).
17. Milman, U., Atias, H., Weiss, A., Mirelman, A. & Hausdorff, J. M. Can cognitiveremediation improve mobility in patients with Parkinson’s disease? Findings froma 12 week pilot study. J. Park. Dis. 4, 37–44 (2014).
18. Levine, M. & Ensom, M. H. H. Post hoc power analysis: an idea whose time haspassed? Pharmacotherapy 21, 405–409 (2001).
19. Bloem, B. R., de Vries, N. M. & Ebersbach, G. Nonpharmacological treatments forpatients with Parkinson’s disease. Mov. Disord. 30, 1504–1520 (2015).
20. Walton, C. C., Naismith, S. L., Lampit, A., Mowszowski, L. & Lewis, S. J. Cognitivetraining in Parkinson’s disease: a theoretical perspective. Neurorehabil. NeuralRepair. 31, 207–216 (2017).
21. Lampit, A., Hallock, H. & Valenzuela, M. Computerized cognitive training in cog-nitively healthy older adults: a systematic review and meta-analysis of effectmodifiers. PLoS Med. 11, e1001756 (2014).
22. Lampit, A. et al. The timecourse of global cognitive gains from supervisedcomputer-assisted cognitive training: a randomised, active-controlled trial inelderly with multiple dementia risk factors. J. Prev. Alzheimer’s Dis. 1, 33–39 (2014).
23. Hughes, A. J., Daniel, S. E., Kilford, L. & Lees, A. J. Accuracy of clinical diagnosis ofidiopathic Parkinson’s disease: a clinico-pathological study of 100 cases. J. Neurol.Neurosurg. Psychiatry 55, 181–184 (1992).
24. Folstein, M. F., Folstein, S. E. & McHugh, P. R. Mini-mental state. A practicalmethod for grading the cognitive state of patients for the clinician. J. Psychiatr.Res. 12, 189–198 (1975).
25. Giladi, N. et al. Construction of freezing of gait questionnaire for patients withParkinsonism. Park. Relat. Disord. 6, 165–170 (2000).
26. Shine, J. M. et al. Assessing the utility of Freezing of Gait Questionnaires inParkinson’s Disease. Park. Relat. Disord. 18, 25–29 (2012).
27. Goetz, C. G. et al. Movement Disorder Society-sponsored revision of the UnifiedParkinson’s Disease Rating Scale (MDS-UPDRS): scale presentation and clinimetrictesting results. Mov. Disord. 23, 2129–2170 (2008).
Cognitive training for freezing of gaitCC Walton et al.
8
npj Parkinson’s Disease (2018) �15� Published in partnership with the Parkinson’s Foundation

28. Schaafsma, J. D. et al. Characterization of freezing of gait subtypes and theresponse of each to levodopa in Parkinson’s disease. Eur. J. Neurol. 10, 391–398(2003).
29. Nasreddine, Z. S. et al. The Montreal cognitive assessment, MoCA: a briefscreening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 53, 695–699(2005).
30. Benedict, R. H. B., Schretlen, D., Groninger, L. & Brandt, J. Hopkins verbal learningtest—revised: normative data and analysis of inter-form and test-retest reliability.Clin. Neuropsychol. 12, 43–55 (1998).
31. Wechsler, D. WMS-III: Wechsler Memory Scale Administration And Scoring Manual(Psychological Corporation, 1997).
32. Delis, D. C., Kaplan, E. & Kramer, J. H. Delis–Kaplan Executive Function System (D-KEFS) (Psychological Corporation, 2001).
33. Smith, A. Symbol Digit Modalities Test (Western Psychological Services, 1991).34. Tombaugh, T. N. Trail making test A and B: normative data stratified by age and
education. Arch. Clin. Neuropsychol. 19, 203–214 (2004).35. Robbins, T. W. et al. Cambridge neuropsychological test automated battery
(CANTAB): a factor analytic study of a large sample of normal elderly volunteers.Dementia 5, 266–281 (1994).
36. Zigmond, A. S. & Snaith, R. P. The hospital anxiety and depression scale. ActaPsychiatr. Scand. 67, 361–370 (1983).
37. Marinus, J., Visser, M., van Hilten, J. J., Lammers, G. J. & Stiggelbout, A. M.Assessment of sleep and sleepiness in Parkinson disease. Sleep 26, 1049–1054(2003).
38. Jenkinson, C., Fitzpatrick, R., Peto, V., Greenhall, R. & Hyman, N. The Parkinson’sdisease Questionnaire (PDQ-39): development and validation of a Parkinson’sdisease summary index score. Age. Ageing 26, 353–357 (1997).
39. Wear, H. J. et al. The Cambridge behavioural inventory revised. Dement. Neu-ropsychol. 2, 102–107 (2008).
40. Naismith, S. L., Mowszowski, L., Diamond, K. & Lewis, S. J. Improving memory inParkinson’s disease: a healthy brain ageing cognitive training program. Mov.Disord. 28, 1097–1103 (2013).
41. Diamond, K. et al. Randomized controlled trial of a healthy brain ageing cognitivetraining program: effects on memory, mood, and sleep. J. Alzheimer’s Dis. 44,1181–1191 (2015).
42. Medalia, A. & Freilich, B. The neuropsychological educational approach to cog-nitive remediation (NEAR) model: practice principles and outcome studies. Am. J.Psychiatr. Rehabil. 11, 123–143 (2008).
43. Naismith, S. L. et al. Enhancing memory in late-life depression: the effects of acombined psychoeducation and cognitive training program. Am. J. Geriatr. Psy-chiatry. 19, 240–248 (2011).
44. Mowszowski, L. et al. Cognitive training enhances pre-attentive neurophysiolo-gical responses in older adults “at risk” of dementia. J. Alzeimer’s Dis. 41,1095–1108 (2014).
45. Sohlberg, M. M. & Mateer, C. A. Attention Process Training APT-3: A Direct AttentionTraining Program For Persons With Acquired Brain Injury (Lash & Associates Pub-lishing/Training Inc, 2011).
46. Borness, C., Proudfoot, J., Crawford, J. & Valenzuela, M. Putting brain training tothe test in the workplace: a randomized, blinded, multisite, active-controlled trial.PLoS ONE 8, e59982 (2013).
47. Cohen, J. Statistical Power Analysis For The Behavioral Sciences 20–26 (LawrenceEarlbaum Associates, Hillsdale, NJ, 1988).
48. Tomlinson, C. L. et al. Systematic review of levodopa dose equivalency reportingin Parkinson’s disease. Mov. Disord. 25, 2649–2653 (2010).
Open Access This article is licensed under a Creative CommonsAttribution 4.0 International License, which permits use, sharing,
adaptation, distribution and reproduction in anymedium or format, as long as you giveappropriate credit to the original author(s) and the source, provide a link to the CreativeCommons license, and indicate if changes were made. The images or other third partymaterial in this article are included in the article’s Creative Commons license, unlessindicated otherwise in a credit line to the material. If material is not included in thearticle’s Creative Commons license and your intended use is not permitted by statutoryregulation or exceeds the permitted use, you will need to obtain permission directlyfrom the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
© The Author(s) 2018
Cognitive training for freezing of gaitCC Walton et al.
9
Published in partnership with the Parkinson’s Foundation npj Parkinson’s Disease (2018) �15�

APPENDIX(12((
O’Callaghan, C., Hall, J. M., Tomassini, A., Muller, A. J., Walpola, I. C., Moustafa,
A. A., Shine, J. M., Lewis, S. J. G. Visual Hallucinations are characterized by
impaired sensory evidence accumulation: insights from hierarchical drift diffusion
modeling in Parkinson's Disease. Biological Psychiatry: Cognitive Neuroscience and
Neuroimaging, 2017
!

Archival Report
Visual Hallucinations Are Characterized byImpaired Sensory Evidence Accumulation:Insights From Hierarchical Drift DiffusionModeling in Parkinson’s DiseaseClaire O’Callaghan, Julie M. Hall, Alessandro Tomassini, Alana J. Muller, Ishan C. Walpola,Ahmed A. Moustafa, James M. Shine, and Simon J.G. Lewis
ABSTRACTBACKGROUND: Models of hallucinations emphasize imbalance between sensory input and top-down influencesover perception, as false perceptual inference can arise when top-down predictions are afforded too muchprecision (certainty) relative to sensory evidence. Visual hallucinations in Parkinson’s disease (PD) are associatedwith lower-level visual and attentional impairments, accompanied by overactivity in higher-order association brainnetworks. PD therefore provides an attractive framework to explore contributions of bottom-up versus top-downdisturbances in hallucinations.METHODS:We characterized sensory processing during perceptual decision making in patients with PD with (n = 20)and without (n = 25) visual hallucinations and control subjects (n = 12), by fitting a hierarchical drift diffusion model toan attentional task. The hierarchical drift diffusion model uses Bayesian estimates to decompose task performanceinto parameters reflecting drift rates of evidence accumulation, decision thresholds, and nondecision time.RESULTS: We observed slower drift rates in patients with hallucinations, which were less sensitive to changes in taskdemand. In contrast, wider decision boundaries and shorter nondecision times relative to control subjects were foundin patients with PD regardless of hallucinator status. Inefficient and less flexible sensory evidence accumulationemerges as a unique feature of PD hallucinators.CONCLUSIONS: We integrate these results with evidence accumulation and predictive coding models of halluci-nations, suggesting that in PD sensory evidence is less informative and may therefore be down-weighted, resulting inoverreliance on top-down influences. Considering impaired drift rates as an approximation of reduced sensoryprecision, our findings provide a novel computational framework to specify impairments in sensory processing thatcontribute to development of visual hallucinations.
Keywords: Bayesian, Bottom up, Hierarchical drift diffusion model, Parkinson’s disease, Perception, Precision, Topdown, Visual hallucinations
http://dx.doi.org/10.1016/j.bpsc.2017.04.007
Visual hallucinations (VHs) are common in Parkinson’s disease(PD), occurring in over 30% of newly diagnosed and early-stage patients and increasing to upwards of 70% by the latestages of the disease (1–3). However, despite their prevalence,VHs remain poorly understood, and treatment options arelimited (4). Continued characterization of the psychological andmechanistic correlates of VHs in PD will be crucial to informtherapeutic advances.
Proposed explanatory models for VHs in PD emphasize astate of reduced sensory input, where the ongoing perceptualprocess is vulnerable to influence from internally generatedimagery (5–8). This is in keeping with a transdiagnosticframework, where hallucinations arise when the balancebetween sensory input and top-town influence over perceptionis disrupted, such that sensory information is reduced or not
properly integrated, and there is a predominance of top-downinfluence (9–13). In this context, there is both impairment insensory evidence accumulation and a failure of Bayesianinference processes regulating top-down versus bottom-upinfluences on perception.
In formal computational terms, VHs can be considered falseperceptual inference (13). Perceptual inference can becomputationally implemented by predictive coding or Kalmanfiltering, in which incoming sensory input is predicted basedon past information, and a prediction error specifies thedifference between the expected and incoming evidence.Precision is a measure of certainty determining the weightthose errors will have (termed their Kalman gain) in influencingthe updating of subsequent estimates (14). This is importantin theories of hallucinations, where the top-down versus
SEE COMMENTARY ON PAGE 640
680 ª 2017 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI ISSN: 2451-9022
BiologicalPsychiatry:CNNI

bottom-up imbalance is driven by a possibly compensatoryimbalance of precision, in which top-down predictions areafforded too much precision in relation to sensory evidence. Aprediction that follows is that patients with VHs should showreduced sensory precision.
In PD, sensory input is affected by dopaminergic retinalchanges and impairments in lower-level visual processes andattention—all of which can be more pronounced in patientswith hallucinations (15). However, hallucinations can alsooccur in patients in whom ophthalmologic measures andperformances on lower-level perceptual tasks are equivalent tothose in nonhallucinating patients (16). A possibility is thatlower-level sensory impairment and reduced attention conferrisk factors for VHs in PD, but failures in the dynamic inte-gration of visual input and attention trigger their occurrence(6,8). We aimed to investigate the dynamic processes under-lying visual perception in PD hallucinators by applying a driftdiffusion model (DDM) to an attentional task. Crucially, atten-tional deficits (and the use of an attentional task) follow fromthe above aberrant precision account. In predictive coding,attention serves as a gain control mechanism to enhance theprecision of sensory prediction errors (17–21). Increased pre-cision implies more in-depth processing of a stimulus (22).Aberrant precision control is therefore consistent with anattentional deficit, where sensory evidence is not efficientlyselected and enhanced, leading to failures in belief updatingand perceptual inference.
DDMs are based on the premise that reaction time andresponse output can be decomposed into parametersreflecting the latent cognitive processes driving task perfor-mance (23). The DDM quantifies information extracted from astimulus (drift rate), the evidence needed to make a decision(boundary separation), and components related to stimulusencoding and response output (nondecision time) (24,25). Inthe context of predictive coding, the drift rate can be used toapproximate precision or Kalman gain, as the rate of sensoryevidence accumulation would correspond to the precision (i.e.,confidence or certainty) ascribed to the evidence being accu-mulated. In this study, we applied a Bayesian hierarchicalversion of the DDM, which is robust in the context of low trialnumbers (26,27). This makes the task suitable for clinicalcontexts where task duration is necessarily limited, and it hasbeen successfully fitted to data from patients with PD in pre-vious studies (28–30). We assessed participants on theattention network task (ANT) (31), which allowed us to measureperceptual decision making under conditions with differentlevels of difficulty as determined by perceptual conflict in thestimuli.
Under the DDM, we predicted that patients with VHswould show reduced sensory precision relative to patientswithout hallucinations, as evidenced by impairments in pa-rameters reflecting the integration or accumulation of sensoryevidence in the decision-making process, i.e., the drift rate orboundary separation. However, their nondecision compo-nents should be similar. We also predicted that control sub-jects and patients without hallucinations would modulate theirdrift rate and boundary separation in response to the differentlevels of perpetual conflict, but that patients with VHs wouldnot show the same level of flexibility in response to taskdemands.
METHODS AND MATERIALS
Case Selection
We recruited 50 patients from the Parkinson’s DiseaseResearch Clinic at the Brain and Mind Centre, University ofSydney. Patients were identified as hallucinators if they self-reported visual hallucinatory phenomena and scored $1 onquestion 2 of the Movement Disorder Society–sponsoredrevision of the Unified Parkinson’s Disease Rating Scale (i.e.,over the past week have you seen, heard, smelled or felt thingsthat were not really there? If yes, the examiner asks the patientor caregiver to elaborate and probes for information) (32). Thisresulted in 24 patients in the VH group and 26 patients in thenonVH group. Four patients from the VH group and 1 from thenonVH group were excluded from analysis owing to excessivemissed responses on the experimental task, leaving a finalcohort of 20 patients in the VH group and 25 patients in thenonVH group. A proportion of these patients were included in aprevious behavioral study of the ANT (33). We recruited 12age-matched control subjects from a volunteer panel.
All patients satisfied the United Kingdom Parkinson’s Dis-ease Society Brain Bank criteria and were not demented,scoring above the recommended Mini-Mental State Examina-tion cutoff of $26 (34). Patients were assessed on the Hoehnand Yahr Scale and the Motor Examination section of theUnified Parkinson’s Disease Rating Scale. The Mini-MentalState Examination and Montreal Cognitive Assessment wereadministered as measures of general cognition. Clinicalassessments and the experimental task were performed withpatients in the “on” state, having taken their regular dopami-nergic medication, and dopaminergic dose equivalence scoreswere calculated. No patients in the cohort were taking anti-psychotic medication or cholinesterase inhibitors. Controlsubjects were screened for a history of neurological or psy-chiatric disorders. The study was approved by the local ethicscommittees, and participants provided informed consent.
Attention Network Task
We administered a shortened version of the ANT (31), whichrequires participants to determine if a central arrow points leftor right. Central arrows are flanked by flat lines (neutral con-dition), arrows facing the same direction (congruent condition),or arrows facing a mixture of directions (incongruent condition)(Figure 1A). The perceptual conflict in the incongruent condi-tion is designed to place a greater demand on attentionalprocesses relative to the congruent and neutral conditions.See Supplement for task details.
Hierarchical DDM of the ANT
DDMs are widely applied to rapid, two-choice decision-makingtasks such as the ANT (25,35,36). DDMs are typicallydescribed by four main parameters: drift rate (v), boundary (a),decision bias (z), and nondecision time (T). The decision pro-cess is modeled as the gradual accumulation of information,reflected by the drift rate, which continues until a decisionboundary is reached (37). The decision bias parameter cap-tures a priori bias toward one of the two responses. Nonde-cision time incorporates components that are not part of theevidence accumulation process, including stimulus encoding,
Visual Hallucinations in PD
Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI 681
BiologicalPsychiatry:CNNI

extracting stimulus dimensions, and executing a response (36).Figure 1B is a schematic diagram of the drift diffusion process.We applied a hierarchical DDM (hDDM) using the hDDMtoolbox (http://ski.clps.brown.edu/hddm_docs/) (26). ThehDDM uses Bayesian estimation to generate posterior distri-butions of parameters. This approach optimizes the tradeoffbetween within-subject and between-subject random effects,accounting for both within-subject variability and group-levelsimilarities, as individual parameters are constrained by agroup-level distribution.
We tested three models that all assumed an unbiasedstarting point (z), given that left/right responses were coun-terbalanced, and assumed that nondecision time (T) would notbe expected to vary as a function of condition, as the stimulusencoding and motor responses required across conditionswere comparable. Model specifications were as follows: in thefirst model, only drift rate (v) was permitted to vary by condi-tion, and decision boundary (a) was held constant; in a secondmodel, decision boundary (a) could vary across conditions, butdrift rate (v) was held constant; in a third model, both v and awere free to vary across conditions. For all models, Markovchain Monte Carlo simulations were used to generate 120,000samples from the joint posterior parameter distribution. Thefirst 20,000 samples were discarded as burn-in, and we used athinning factor of 10, with outliers specified at 5%. Conver-gence was assessed by visually inspecting the Markov chainsand computing the R-hat Gelman-Rubin statistic where suc-cessful coverage is indicated by values ,1.1 (37). The best
model was determined by comparing the deviance informationcriterion (DIC) of each model, which evaluates a model’sgoodness-of-fit while accounting for model complexity (i.e.,number of free parameters), with lower DIC values indicatingbetter model fit (38). To further evaluate the best fitting model,we ran posterior predictive checks by averaging 500 simula-tions generated from the model’s posterior to confirm it couldreliably reproduce patterns in the observed data (26). The fittedANT data and hDDM source code can be found at https://github.com/claireocallaghan/hDDM_ANT_PD.
Statistical Analysis
Independent samples t tests and analyses of variance withTukey post hoc tests compared demographics and behavioralresults from the ANT. Parameters from the hDDM wereanalyzed using Bayesian hypothesis testing to determine theextent of overlap between the percentage of samples drawnfrom two posterior density distributions. Posterior probabilitiesare considered significantly different if ,5% of the distribu-tions overlap (26–28). The proportion of overlap in the posteriorprobabilities is denoted by P to distinguish it from the classicalfrequentist p values.
RESULTS
Participant Characteristics
The groups were matched for age (F2,54 = 1.31, p = .28).Performance on the Mini-Mental State Examination was similaracross groups (F2,54 = 1.07, p = .35), but the MontrealCognitive Assessment revealed significant differences, withthe nonVH group performing below the control group ([F2,54 =5.91, p , .01]; nonVH group vs. control group, p , .01; VHgroup vs. control and nonVH groups, p = .17 and p = .19). Thepatient groups did not differ in disease duration (t = 21.42,p = .16), Hoehn and Yahr stage (t = 21.33, p = .19), UnifiedParkinson’s Disease Rating Scale Motor Examination(t = 21.80, p = .08), or dopamine dose equivalence (t = 21.63,p = .11) (Table 1).
Left decision boundary
Right decision boundary
Left RT distribution
Right RT distribution
{
Non-decision time (T)
Boundary separation (a)
A
B
Neutral Congruent Incongruent
Time (msec)
Figure 1. Attention network task conditions and drift diffusion model. (A)The three conditions in the attention network task. Examples of left and rightstimulus cues for each condition are shown here; however, in the task, onlyone cue was presented per trial for a maximum of 1700 ms. (B) Schematicexample of drift diffusion trajectories. Evidence is noisily accumulated to-ward a left or right response (blue and red panels), which are separated bythe boundary threshold (a). The average evidence accumulation is denotedby drift rate (v). The evidence accumulation begins after a period ofnondecision time (T). Density plots show the distribution of observable re-action times (RT). [Adapted from Wiecki et al. (26) and Zhang and Rowe (73).]
Table 1. Demographics and Clinical Characteristics ofPatients With Parkinson’s Disease and Healthy ControlSubjects
Control GroupPD nonVHGroup
PD VHGroup
Number 12 25 20
Gender (M:F) 4:8 19:6 15:5
Age, Years 64.75 (5.97) 67.08 (8.22) 68.89 (6.05)
MMSE 29.08 (0.79) 28.39 (1.47) 28.79 (1.72)
MoCA 28.50 (1.45) 26.56 (2.11) 27.41 (1.50)
Disease Duration, Years — 5.52 (2.86) 7.11 (4.99)
Hoehn and Yahr Stage — 2.16 (0.47) 2.37 (0.60)
UPDRS III — 27.67 (11.54) 34.28 (13.97)
Values are mean (SD) or n.F, female; M, male; MMSE, Mini-Mental State Examination; MoCA,
Montreal Cognitive Assessment; nonVH, non–visual hallucination; PD,Parkinson’s disease; UPDRS III, Unified Parkinson’s Disease RatingScale Motor Examination; VH, visual hallucination.
Visual Hallucinations in PD
682 Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI
BiologicalPsychiatry:CNNI

ANT Behavioral Results
Participants who made no response on more than one-third oftrials were excluded from the study. Trials where no responsewas made were omitted from the behavioral and modelinganalyses, rather than using the upper limit reaction time, whichwould bias the model. More trials were removed for the VHgroup compared with the nonVH group, although patients andcontrol subjects did not differ significantly ([F2,54 = 4.06,p , .05]; control group vs. nonVH and VH groups, p = .10 andp = .09; VH group vs. nonVH group, p , .05).
After removal of no response trials, accuracy was 100%across the three groups. Reaction times are plotted in Figure 2.Global reaction times, regardless of condition, were fastest forthe control group followed by the nonVH group and then theVH group, with significant differences evidenced by a maineffect for group in the analysis of variance ([F2,162 = 8.15,p , .001]; VH group vs. control and nonVH groups, bothp , .01; nonVH group vs. control group, p = .59). A main effectof condition revealed that reaction times were significantlyslower for the incongruent condition compared with both thecongruent and the neutral conditions, whereas the congruentand neutral conductions were equivalent ([F2,162 = 17.02,p , .000001]; incongruent vs. congruent and neutral, bothp , .00001; congruent vs. neutral, p = .98). There was no sig-nificant interaction between group and condition, suggestingthat the relatively slowed reaction times for the incongruentcondition were consistent across groups (F4,162 = 0.04, p = .99).
hDDM Fit
All three models showed good convergence (see Supplementfor R-hat values). The best-fitting model was model three,which allowed v and a to vary across conditions (DIC model3 = 2636.496, compared with DIC model 1 = 2436.160, and
DIC model 2 = 2626.593). Posterior predictive checks showedgood agreement between the simulated and observed data asshown in Supplemental Figure S1 plotting the observed dataagainst the model prediction. Comparisons showed that thedifference between the summary statistics of the simulatedand the observed data fell within the 95% credible interval.
Analysis of Model Parameters
Comparisons Between Groups. Figure 3 shows theposterior probability density plots for the drift rate (top panel)and decision boundaries (bottom panel) for the three groupsacross each condition. The VH group had uniformly lowerdrift rates compared with the nonVH group; these differedsignificantly in all conditions (neutral, P = 0.05%; incongruent,P = 0.04%; congruent, P = 3.41%). Drift rates of patients inthe VH group were also significantly lower than for controlsubjects for the incongruent and neutral conditions (P =0.04% and P = 3.43%), although not for the congruentcondition (P = 12.72%). Posterior probabilities did not differsignificantly between nonVH and control groups for anycondition (neutral, P = 84.21%; congruent, P = 66.45%;incongruent, P = 41.19%).
For the decision boundary, there were no significant differ-ences between the VH and nonVH groups for any of theconditions (neutral, P = 31.19%; congruent, P = 50.96%;incongruent, P = 9.45%). Both the VH and nonVH groups hadsignificantly larger decision boundaries than the control groupin the neutral and congruent conditions (VH group, P = 0.12%and P = 0.03%; nonVH group, P = 0.03% and P = 0.03%), andthe nonVH group was also significantly larger in the incon-gruent condition (nonVH group, P = 0.68%; VH group, P =14.19%). As shown in Figure 4, the VH and nonVH groups hadsimilar nondecision times (P = 34.04%), which were
050
010
0015
00
Con
trol
050
010
0015
00
Non
VH
050
010
0015
00
VH
neutral congruent incongruent
Figure 2. Attention network task reaction times.Reaction time distributions for the attention networktask across the three levels of task difficulty. Milli-seconds are shown on the y axis. Bold black linesdesignate the mean reaction times for each condi-tion, and the dotted line shows the overall meanacross conditions. The filled colored plots show thedensity distributions of the reaction times; these aredensity plots of the data rotated and mirroredsymmetrically to show the shape of the data distri-bution. nonVH, non–visual hallucination; VH, visualhallucination.
Visual Hallucinations in PD
Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI 683
BiologicalPsychiatry:CNNI

significantly shorter than the control group (VH group, P =0.05%; nonVH group, P = 0.00%).
Comparisons Across Conditions. Figure 5 shows pos-terior probabilities within each group for the three conditions(control group, left panel; nonVH group, middle panel; VHgroup, right panel). For drift rates, the control group showed anexpected pattern with significantly longer drift rates in the
incongruent condition compared with the neutral andcongruent conditions (P = 0.28% and P = 0.32%) and similarrates in the neutral and congruent conditions (P = 50.92%).The nonVH group showed a similar pattern with significantlylonger drift rates in the incongruent relative to the neutral andcongruent conditions (P = 1.55% and P = 0.15%), with similarrates in the neutral and congruent conditions (P = 18.31%). Incontrast, compared with the control and nonVH groups, the VHgroup showed less of a difference between the incongruentand neutral conditions (P = 3.28%) and no significant differ-ence between the incongruent and congruent conditions (P =17.47%), with similar rates also between neutral and congruentconditions (P = 17.80%). All groups showed a similar patternfor decision boundaries across the conditions, with signifi-cantly larger decision boundaries in the incongruent conditionrelative to the neutral and congruent conditions (control group,P = 0.00% and P = 0.00%; nonVH group, P = 0.00% andP = 0.00%; VH group, P = 0.00% and P = 0.00%), with similarthresholds in the neutral and congruent conditions (controlgroup, P = 38.80%; nonVH group, P = 37.32%; VH group,P = 20.00%).
DISCUSSION
Our results demonstrate two features that characterize theperceptual decision-making process of hallucinating patients
0
1
2
3
2.5 3.0 3.5Neutral drift rate
Post
erio
r pro
babi
lity
dens
ity
0
1
2
3
2.4 2.8 3.2 3.6Congruent drift rate
0
1
2
3
2.5 3.0 3.5 4.0Incongruent drift rate
0.0
0.5
1.0
1.5
2.0
2.5
2.0 2.5 3.0 3.5 4.0 4.5Neutral decision boundary
Post
erio
r pro
babi
lity
dens
ity
0.0
0.5
1.0
1.5
2.0
2.0 2.5 3.0 3.5 4.0 4.5Congruent decision boundary
0.0
0.5
1.0
1.5
4 5Incongruent decision boundary
Control VH NonVH
Figure 3. Between-group comparisons of drift rates and decision boundaries. Posterior probability density plots for drift rates (top panel) and decisionboundaries (bottom panel). nonVH, non–visual hallucination; VH, visual hallucination.
0
10
20
30
0.15 0.20 0.25 0.30 0.35Non−decision time
Post
erio
r pro
babi
lity
dens
ity
Control
VH
NonVH
Figure 4. Nondecision time. Posterior probability density plots fornondecision time. nonVH, non–visual hallucination; VH, visual hallucination.
Visual Hallucinations in PD
684 Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI
BiologicalPsychiatry:CNNI

relative to nonhallucinating patients: lower drift rates andreduced ability to adjust drift rates to accommodate changesin perceptual conflict. Behaviorally, all groups had 100% ac-curacy on the ANT. Employing a task that patients with PDcould easily understand and execute was particularly impor-tant, given the possibility of cognitive impairment in this patientcohort. This ensured that we could accurately accessperceptual decision making without higher-order cognitivedeficits confounding performance. Hallucinating patients hadthe slowest reaction times on the ANT, although each groupshowed the expected pattern of shorter reaction times for theeasier (neutral and congruent) conditions and longer reactiontimes for the more difficult (incongruent) condition. Our find-ings from the hDDM reveal the added benefits of modelingthese data to uncover the cognitive processes underlying thebehavioral results.
The drift rate reflects how quickly information is accumu-lated and reflects the quality or precision of evidence thatenters the decision-making process (25). In all three condi-tions, the control and nonVH groups displayed similar driftrates. The VH group had consistently lower drift rates. Inbetween-condition comparisons, drift rates are modulated bytask difficulty (24). This effect was apparent in our results forboth the control and the nonVH groups. These groups hadlonger drift rates in the incongruent relative to the congruent
conditions, suggesting that their integration of information intothe decision-making process was flexibly modulated inresponse to imprecise or ambiguous sensory evidence. Incontrast, the VH group had similar drift rates across thecongruent and incongruent conditions, indicating less flexiblecontext-dependent modulation of sensory accumulation.
Previous work has identified neural correlates of evidenceaccumulation during perceptual decision making. In monkeys,neuronal populations in sensory areas (e.g., middle temporalvisual area) fire in response to properties of a stimulus, anddownstream regions (e.g., lateral intraparietal, frontal eyefields, dorsal lateral prefrontal cortex) integrate this informationover time until sufficient evidence is accumulated for a deci-sion (39–42). In humans, evidence accumulation has beenrelated to frontoparietal regions [i.e., dorsal and ventrolateralprefrontal cortex and frontal eye fields (43–46)] and the anteriorinsula (47) as well as integration across large-scale networks(48). In PD, abnormal local and network-level engagement offrontoparietal and insula regions has been found in patientswith hallucinations (49–52) and is typically equated withattentional dysfunction. In perceptual decision making, accu-mulation of sensory evidence and attention are highlycollinear—and possibly inseparable—processes (23), consis-tent with the relationship between attention and sensory pre-cision specified by predictive coding accounts. It follows that
0.0
0.5
1.0
1.5
2.0
2.5
2.5 3.0 3.5 4.0Control drift rate
Post
erio
r pro
babi
lity
dens
ity
0
1
2
3
3.0 3.5 4.0nonVH drift rate
0
1
2
3
2.25 2.50 2.75 3.00 3.25VH drift rate
0.0
0.5
1.0
1.5
2.0
2.5
2 3 4 5Control decision boundary
Post
erio
r pro
babi
lity
dens
ity
0.0
0.5
1.0
1.5
2.0
3 4 5nonVH decision boundary
0.0
0.5
1.0
1.5
3 4 5VH decision boundary
Congruent Incongruent Neutral
Figure 5. Within-group comparisons of drift rates and decision boundaries. Posterior probability density plots for drift rates (top panel) and decisionboundaries (bottom panel). nonVH, non–visual hallucination; VH, visual hallucination.
Visual Hallucinations in PD
Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI 685
BiologicalPsychiatry:CNNI

our finding of impaired drift rates may offer a more tangible,computational framework to better specify attentional impair-ments in patients with PD with VHs.
All groups modulated their decision boundaries in responseto the more difficult condition, displaying larger boundaries forthe incongruent relative to congruent and neutral conditions.Flexible adaptation of decision thresholds in response to taskdemands has been shown previously in patients with PD(28,29). Despite the flexible adaptation of thresholds, regard-less of hallucination status, patients with PD had larger deci-sion boundaries than control subjects. The lack of differencebetween the VH and nonVH groups suggests that a wideningof decision boundaries during perceptual decision making is afeature of PD more generally. This corresponds to findings inhealthy aging, where older adults display wider decisionboundaries relative to younger adults (53–55). In older adults,adopting conservative decision criteria is presumed to be acompensatory strategy to prevent errors in speed and/or ac-curacy tradeoff tasks, and this effect may be amplified in PD.
Hallucinating and nonhallucinating patients also had com-parable nondecision times, and these were shorter than timesfor control subjects. This might have seemed counterintuitivein light of the longer nondecision times shown in aging (56,57),but reduced nondecision times in PD have also been foundwith the application of an hDDM to a saccadic go/no-go task(30). Nondecision time encompasses diverse processes:encoding evidence from a stimulus and extracting its di-mensions to guide the decision-making process and executionof a motor response (36). Decreased latencies in stimulusencoding and extraction of details may drive this finding, orshorter nondecision times may reflect motor impulsivity, whichhas previously been documented in PD despite general motorslowing (58,59). Future work may make this distinction in PD,although currently nondecision time is a relatively under-specified term compared with the other parameters in theDDM, and few studies have successfully separated its sub-components (23).
Inefficient and less flexible evidence accumulation duringperceptual decision making emerges as a unique feature ofhallucinating patients with PD, whereas changes in cautious-ness and visuomotor processes are apparent regardless ofhallucinator status. Longer drift rate latencies lead to slowerdecisions, and because perceptual decision making is a noisyprocess, this increases the chance of errors (37,60). In hallu-cinating patients with PD, low drift rates that are invariant tochanging environmental demands would be a source of low-quality or imprecise information entering the perceptualprocess.
In terms of aberrant precision control in hallucinating pa-tients with PD, our results are consistent with the simulationsreported by Adams et al. (12), namely, an imbalance betweensensory precision and the precision of top-down prior beliefs.In PD, there is degeneration of ascending gain-setting neuro-modulatory systems (i.e., dopaminergic, cholinergic, andnoradrenergic) (61,62) that are associated with implementingprecision. Aberrant precision control may be an importantunifying principal across PD symptoms. For example, motorsymptoms (in particular bradykinesia) may speak to failures inthe estimation and attenuation of proprioceptive and somato-sensory uncertainty. In active inference accounts, estimating
sensory uncertainty is crucial for initiating movement, as thegoal of movement is to reduce the discrepancy between pre-dicted and actual sensory inputs (63,64). Failures of sensoryattenuation and compensatory increases in the precision and/or gain at higher levels of processing may underlie both thefalse inference in VHs and the failure to realize predictedmovements in the proprioceptive domain.
Aberrant precision control is ultimately reflected as a failureof Bayesian inference (11,13,65). In a Bayesian framework forperception, sensory input (likelihood) is integrated with knownstatistics about the environment (priors), forming an estimateof the external stimulus (the posterior). Contributions of thelikelihood and prior in generating the posterior estimate areweighted in accordance with their certainty. Noisy or imprecisesensory input is uncertain and carries less weight, shifting thebalance in favor of priors. In this context, perception isvulnerable to excessive influence from internally generatedbeliefs and expectations. In PD, if visual information is accu-mulated slowly and inefficiently and is therefore less informa-tive, this may contribute to the down weighting of bottom-upinformation in favor of top-down information.
While the Bayesian framework is a compelling description ofhallucinations, it does not necessarily favor a singlemechanisticexplanation (66). For example, both aberrant predictive codingacross hierarchal brain circuitry (67) and large-scale disruptionsin the excitatory-to-inhibitory tone of the brain (68) accommo-date a Bayesian formulation of hallucinations. Reconcilingcomputational descriptions of hallucinations with amechanisticframework will open important therapeutic avenues, as pre-dictive coding and excitatory-to-inhibitory accounts rely ondistinct neuromodulatory and neurotransmitter profiles. In PD,we are beginning to uncover the psychological and neural sig-natures associated with a bottom-up versus top-down imbal-ance in perception. Along with the results of the current studyand evidence of lower-level deficits in attention and visualprocessing, previous work has identified overactivity in thedefault network of patients with PD with hallucinations (69) andincreased coupling between the default network and visualcortex (50). Given the role of the default network in constructionof mental imagery (70) and its positioning as a transmodalsystem distinct from unimodal sensory regions (71), over-engagement of the default network during visual perceptionmay be a source of excessive top-down influence.
Hallucinations develop progressively in PD, beginning withminor illusions before complex hallucinations emerge (4). Pa-tients initially retain full insight into these symptoms, appreci-ating their occurrence as part of the disease process. Withdisease progression and cognitive decline, patients can loseinsight and develop delusional ideas around the hallucinations.Delusions themselves can be viewed as the instantiation ofnew priors, to accommodate increasingly noisy or imprecisesensory input (11,72). We employed a limited assessment ofhallucinations. Future studies may want to stratify patients inmore detail based on the severity and phenomenology of theirhallucinations to determine how the imbalance between top-down versus bottom-up influence evolves over the diseasecourse. Additionally, we chose a task manageable for patientswith PD; nevertheless, lower-level visual deficits could rendersuch a task more challenging. Future work should employmore detailed visual testing to determine the influence of
Visual Hallucinations in PD
686 Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI
BiologicalPsychiatry:CNNI

low-level vision on sensory evidence accumulation, and repli-cating the current findings in other perceptual decision-makingtasks will be important.
In conclusion, our results suggest that impaired drift ratecan be used to approximate precision, providing a novelcomputational framework encompassing the sensory pro-cessing and lower-level attentional deficits previouslydescribed in individuals with VHs. Alterations in the dynamicprocess of sensory evidence accumulation may therefore be avaluable marker to exploit in future explanatory and therapeuticstudies of PD VHs.
ACKNOWLEDGMENTS AND DISCLOSURESThis work was supported by the National Health and Medical ResearchCouncil Neil Hamilton Fairley Fellowship (Grant No. 1091310 to CO), MedicalResearch Council (Grant No. MC-A060-5PQ30 to AT), University of SydneyAustralian Postgraduate Award (to AJM), National Health and MedicalResearch Council CJ Martin Fellowship (Grant No. 1072403 to JMS), andNational Health and Medical Research Council Practitioner Fellowship(Grant No. 1003007 to SJGL).
The authors report no biomedical financial interests or potential conflictsof interest.
ARTICLE INFORMATIONFrom the Department of Psychiatry and Behavioural and Clinical Neuro-science Institute (CO) and Department of Clinical Neurosciences andCognition and Brain Sciences Unit (AT), University of Cambridge, Cam-bridge, United Kingdom; Brain and Mind Centre (CO, JMH, AJM, ICW, JMS,SJGL), University of Sydney; School of Social Sciences and Psychology(JMH, AAM), Western Sydney University, Sydney, Australia; and School ofPsychology (JMS), Stanford University, Palo Alto, California.
Address correspondence to Claire O’Callaghan, Ph.D., Herchel SmithBuilding for Brain & Mind Sciences, Cambridge Biomedical Campus,Cambridge CB2 0SZ, UK; E-mail: [email protected].
Received Feb 25, 2017; revised Mar 30, 2017; accepted Apr 23, 2017.Supplementary material cited in this article is available online at http://
dx.doi.org/10.1016/j.bpsc.2017.04.007.
REFERENCES1. Hely MA, Reid WG, Adena MA, Halliday GM, Morris JG (2008): The
Sydney multicenter study of Parkinson’s disease: The inevitability ofdementia at 20 years. Mov Disord 23:837–844.
2. Pagonabarraga J, Martinez-Horta S, Fernández de Bobadilla R,Pérez J, Ribosa-Nogué R, Marín J, et al. (2016): Minor hallucinationsoccur in drug-naive Parkinson’s disease patients, even from the pre-motor phase. Mov Disord 31:45–52.
3. Holroyd S, Currie L, Wooten G (2001): Prospective study of halluci-nations and delusions in Parkinson’s disease. J Neurol NeurosurgPsychiatry 70:734–738.
4. Ffytche DH, Creese B, Politis M, Chaudhuri KR, Weintraub D,Ballard C, et al. (2017): The psychosis spectrum in Parkinson disease.Nat Rev Neurol 13:81–95.
5. Shine JM, O’Callaghan C, Halliday GM, Lewis SJG (2014): Tricks of themind: Visual hallucinations as disorders of attention. Prog Neurobiol116:58–65.
6. Collerton D, Perry E, McKeith I (2005): Why people see things that arenot there: A novel perception and attention deficit model for recurrentcomplex visual hallucinations. Behav Brain Sci 28:737–757.
7. Hobson JA, Pace-Schott EF, Stickgold R (2000): Dreaming and thebrain: Toward a cognitive neuroscience of conscious states. BehavBrain Sci 23:793–842.
8. Diederich NJ, Goetz CG, Stebbins GT (2005): Repeated visual hallu-cinations in Parkinson’s disease as disturbed external/internal per-ceptions: Focused review and a new integrative model. Mov Disord20:130–140.
9. O’Callaghan C, Kveraga K, Shine JM, Adams RB Jr, Bar M (2017):Predictions penetrate perception: Converging insights from brain,behaviour and disorder. Conscious Cogn 47:63–74.
10. Powers AR, Kelley M, Corlett PR (2016): Hallucinations as top-downeffects on perception. Biol Psychiatry Cogn Neurosci Neuroimaging1:393–400.
11. Fletcher PC, Frith CD (2009): Perceiving is believing: A Bayesianapproach to explaining the positive symptoms of schizophrenia. NatRev Neurosci 10:48–58.
12. Adams RA, Stephan KE, Brown HR, Frith CD, Friston KJ (2013): Thecomputational anatomy of psychosis. Front Psychiatry 4:47.
13. Friston KJ (2005): Hallucinations and perceptual inference. BehavBrain Sci 28:764–766.
14. Rao RP (1999): An optimal estimation approach to visual perceptionand learning. Vision Res 39:1963–1989.
15. Weil RS, Schrag AE, Warren JD, Crutch SJ, Lees AJ, Morris HR (2016):Visual dysfunction in Parkinson’s disease. Brain 139:2827–2843.
16. Gallagher DA, Parkkinen L, O’Sullivan SS, Spratt A, Shah A, Davey CC,et al. (2011): Testing an aetiological model of visual hallucinations inParkinson’s disease. Brain 134:3299–3309.
17. Rao RP (2005): Bayesian inference and attentional modulation in thevisual cortex. Neuroreport 16:1843–1848.
18. Spratling MW (2008): Predictive coding as a model of biasedcompetition in visual attention. Vision Res 48:1391–1408.
19. Feldman H, Friston K (2010): Attention, uncertainty, and free-energy.Front Humn Neurosci 4:215.
20. Jiang J, Summerfield C, Egner T (2013): Attention sharpens thedistinction between expected and unexpected percepts in the visualbrain. J Neurosci 33:18438–18447.
21. Friston KJ, Stephan KE (2007): Free-energy and the brain. Synthese159:417–458.
22. Wiese W, Metzinger TK (2017): Vanilla PP for philosophers: A primeron predictive processing. In: Metzinger TK, Wiese W, editors. Phi-losophy and Predictive Processing. Frankfurt am Main: MIND Group.
23. Mulder M, Van Maanen L, Forstmann B (2014): Perceptual decisionneurosciences—a model-based review. Neuroscience 277:872–884.
24. Voss A, Rothermund K, Voss J (2004): Interpreting the parameters ofthe diffusion model: An empirical validation. Mem Cognit 32:1206–1220.
25. Ratcliff R, McKoon G (2008): The diffusion decision model: Theory anddata for two-choice decision tasks. Neural Comput 20:873–922.
26. Wiecki TV, Sofer I, Frank MJ (2013): HDDM: Hierarchical bayesianestimation of the drift-diffusion model in python. Front Neuroinform7:14.
27. Cavanagh JF, Wiecki TV, Kochar A, Frank MJ (2014): Eye tracking andpupillometry are indicators of dissociable latent decision processes.J Exp Psychol Gen 143:1476.
28. Herz DM, Zavala BA, Bogacz R, Brown P (2016): Neural correlates ofdecision thresholds in the human subthalamic nucleus. Curr Biol26:916–920.
29. Cavanagh JF, Wiecki TV, Cohen MX, Figueroa CM, Samanta J,Sherman SJ, et al. (2011): Subthalamic nucleus stimulation reversesmediofrontal influence over decision threshold. Nat Neurosci 14:1462–1467.
30. Zhang J, Rittman T, Nombela C, Fois A, Coyle-Gilchrist I, Barker RA,et al. (2016): Different decision deficits impair response inhibition inprogressive supranuclear palsy and Parkinson’s disease. Brain139:161–173.
31. Fan J, McCandliss BD, Sommer T, Raz A, Posner MI (2002): Testingthe efficiency and independence of attentional networks. J CognNeurosci 14:340–347.
32. Goetz CG, Tilley BC, Shaftman SR, Stebbins GT, Fahn S, Martinez-Martin P, et al. (2008): Movement Disorder Society-sponsored revisionof the Unified Parkinson’s Disease Rating Scale (MDS-UPDRS): Scalepresentation and clinimetric testing results. Mov Disord 23:2129–2170.
33. Hall JM, O’Callaghan C, Shine JM, Muller AJ, Phillips JR, Walton CC,et al. (2016): Dysfunction in attentional processing in patients withParkinson’s disease and visual hallucinations. J Neural Transm(Vienna) 123:503–507.
Visual Hallucinations in PD
Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI 687
BiologicalPsychiatry:CNNI

34. Martinez-Martin P, Falup-Pecurariu C, Rodriguez-Blazquez C,Serrano-Dueñas M, Carod Artal F, Rojo Abuin J, et al. (2011): Dementiaassociated with Parkinson’s disease: Applying the Movement DisorderSociety Task Force criteria. Parkinsonism Relat Disord 17:621–624.
35. Wagenmakers E-J (2009): Methodological and empirical de-velopments for the Ratcliff diffusion model of response times andaccuracy. Eur J Cogn Psychol 21:641–671.
36. Ratcliff R, Smith PL, Brown SD, McKoon G (2016): Diffusion decisionmodel: Current issues and history. Trends Cogn Sci 20:260–281.
37. Krypotos A-M, Beckers T, Kindt M, Wagenmakers E-J (2015):A Bayesian hierarchical diffusion model decomposition of perfor-mance in Approach-Avoidance Tasks. Cogn Emot 29:1424–1444.
38. Spiegelhalter DJ, Best NG, Carlin BP, Van Der Linde A (2002):Bayesian measures of model complexity and fit. J R Stat Soc Series BStat Methodol 64:583–639.
39. Newsome WT, Britten KH, Movshon JA (1989): Neuronal correlates ofa perceptual decision. Nature 341:52–54.
40. Gold JI, Shadlen MN (2001): Neural computations that underlie de-cisions about sensory stimuli. Trends Cogn Sci 5:10–16.
41. Shadlen MN, Newsome WT (2001): Neural basis of a perceptual de-cision in the parietal cortex (area LIP) of the rhesus monkey.J Neurophysiol 86:1916–1936.
42. Heekeren HR, Marrett S, Ungerleider LG (2008): The neural systemsthat mediate human perceptual decision making. Nat Rev Neurosci9:467–479.
43. Heekeren HR, Marrett S, Bandettini PA, Ungerleider LG (2004):A general mechanism for perceptual decision-making in the humanbrain. Nature 431:859–862.
44. Heekeren HR, Marrett S, Ruff DA, Bandettini P, Ungerleider LG (2006):Involvement of human left dorsolateral prefrontal cortex in perceptualdecision making is independent of response modality. Proc Natl AcadSci 103:10023–10028.
45. Liu T, Pleskac TJ (2011): Neural correlates of evidence accumulation ina perceptual decision task. J Neurophysiol 106:2383–2398.
46. Philiastides MG, Sajda P (2007): EEG-informed fMRI reveals spatio-temporal characteristics of perceptual decision making. J Neurosci27:13082–13091.
47. Ho TC, Brown S, Serences JT (2009): Domain general mechanisms ofperceptual decision making in human cortex. J Neurosci 29:8675–8687.
48. Shine JM, Bissett PG, Bell PT, Koyejo O, Balsters JH, Gorgolewski KJ,et al. (2016): The dynamics of functional brain networks: Integratednetwork states during cognitive task performance. Neuron 92:544–554.
49. Goetz CG, Vaughan CL, Goldman JG, Stebbins GT (2014): I finally seewhat you see: Parkinson’s disease visual hallucinations captured withfunctional neuroimaging. Mov Disord 29:115–117.
50. Shine JM, Muller AJ, O’Callaghan C, Hornberger M, Halliday GM,Lewis SJG (2015): Abnormal connectivity between the default modeand the visual system underlies the manifestation of visual hallucina-tions in Parkinson’s disease: A task-based fMRI study. NPJ Parkin-sons Dis 1:15003.
51. Stebbins G, Goetz C, Carrillo M, Bangen K, Turner D, Glover G, et al.(2004): Altered cortical visual processing in PD with hallucinations: AnfMRI study. Neurology 63:1409–1416.
52. Ramírez-Ruiz B, Martí MJ, Tolosa E, Falcón C, Bargalló N,Valldeoriola F, et al. (2008): Brain response to complex visual stimuli inParkinson’s patients with hallucinations: A functional magnetic reso-nance imaging study. Mov Disord 23:2335–2343.
53. Ratcliff R, Thapar A, McKoon G (2007): Application of the diffusionmodel to two-choice tasks for adults 75-90 years old. Psychol Aging22:56.
54. Spaniol J, Madden DJ, Voss A (2006): A diffusion model analysis ofadult age differences in episodic and semantic long-term memoryretrieval. J Exp Psychol Learn Mem Cogn 32:101.
55. Ratcliff R, Thapar A, McKoon G (2001): The effects of aging on reac-tion time in a signal detection task. Psychol Aging 16:323.
56. Starns JJ, Ratcliff R (2010): The effects of aging on the speed-accuracy compromise: Boundary optimality in the diffusion model.Psychol Aging 25:377.
57. Ratcliff R (2008): Modeling aging effects on two-choice tasks:Response signal and response time data. Psychol Aging 23:900.
58. O’Callaghan C, Naismith SL, Hodges JR, Lewis SJG, Hornberger M(2013): Fronto-striatal atrophy correlates of inhibitory dysfunction inParkinson’s disease versus behavioural variant frontotemporal de-mentia. Cortex 49:1833–1843.
59. Nombela C, Rittman T, Robbins TW, Rowe JB (2014): Multiple modesof impulsivity in Parkinson’s disease. PLoS One 9:e85747.
60. Herz DM, Bogacz R, Brown P (2016): Neuroscience: Impaireddecision-making in Parkinson’s disease. Curr Biol 26:R671–R673.
61. Del Tredici K, Rüb U, De Vos RA, Bohl JR, Braak H (2002): Where doesParkinson disease pathology begin in the brain? J Neuropathol ExpNeurol 61:413–426.
62. Surmeier DJ, Obeso JA, Halliday GM (2017): Selective neuronalvulnerability in Parkinson disease. Nat Rev Neurosci 18:101–113.
63. Friston K, Mattout J, Kilner J (2011): Action understanding and activeinference. Biol Cybern 104:137–160.
64. Palmer C, Zapparoli L, Kilner JM (2016): A new framework to explainsensorimotor beta oscillations. Trends Cogn Sci 20:321–323.
65. Corlett P, Frith CD, Fletcher P (2009): From drugs to deprivation: ABayesian framework for understanding models of psychosis. Psy-chopharmacology 206:515–530.
66. Teufel C, Fletcher PC (2016): The promises and pitfalls of applyingcomputational models to neurological and psychiatric disorders. Brain139:2600–2608.
67. Friston KJ, Stephan KE, Montague R, Dolan RJ (2014): Computationalpsychiatry: The brain as a phantastic organ. Lancet Psychiatry 1:148–158.
68. Jardri R, Hugdahl K, Hughes M, Brunelin J, Waters F, Alderson-Day B,et al. (2016): Are hallucinations due to an imbalance between excit-atory and inhibitory influences on the brain? Schizophr Bull 42:1124–1134.
69. Franciotti R, Delli Pizzi S, Perfetti B, Tartaro A, Bonanni L, Thomas A,et al. (2015): Default mode network links to visual hallucinations: Acomparison between Parkinson’s disease and multiple system atro-phy. Mov Disord 30:1237–1247.
70. Andrews-Hanna JR (2012): The brain’s default network and its adap-tive role in internal mentation. Neuroscientist 18:251–270.
71. Margulies DS, Ghosh SS, Goulas A, Falkiewicz M, Huntenburg JM,Langs G, et al. (2016): Situating the default-mode network along aprincipal gradient of macroscale cortical organization. Proc Natl AcadSci U S A 113:12574–12579.
72. Corlett PR, Fletcher PC (2015): Delusions and prediction error: Clari-fying the roles of behavioural and brain responses. Cogn Neuropsy-chiatry 20:95–105.
73. Zhang J, Rowe JB (2014): Dissociable mechanisms of speed-accuracytradeoff during visual perceptual learning are revealed by a hierarchicaldrift-diffusion model. Front Neurosci 8:69.
Visual Hallucinations in PD
688 Biological Psychiatry: Cognitive Neuroscience and Neuroimaging November 2017; 2:680–688 www.sobp.org/BPCNNI
BiologicalPsychiatry:CNNI

APPENDIX(13(
((Moustafa, A.A., Chakravarthy, S., Phillips, J. R., Crouse, J. J., Gupta, A., Frank, M.
J., Hall, J. M., Jahanshahi, M. Interrelations between cognitive dysfunction and
motor symptoms of Parkinson's disease: behavioural and neural studies. Reviews in
the Neurosciences, 2016
!

Rev. Neurosci. 2016; 27(5): 535–548
*Corresponding author: Ahmed A. Moustafa, School of Social Sciences and Psychology and Marcs Institute for Brain and Behaviour, Western Sydney University, Sydney, New South Wales 2214, Australia, e-mail: [email protected]; and Department of Veterans Affairs, New Jersey Health Care System, East Orange, NJ 07018, USA Srinivasa Chakravarthy and Ankur Gupta: Department of Biotechnology, Indian Institute of Technology, Madras, Chennai 600 036, IndiaJoseph R. Phillips, Jacob J. Crouse and Julie M. Hall: School of Social Sciences and Psychology, Western Sydney University, Sydney, New South Wales, AustraliaMichael J. Frank: Department of Cognitive, Linguistic Sciences and Psychological Sciences, Brown Institute for Brain Science, Brown University, Providence, RI 02912, USAMarjan Jahanshahi: Cognitive Motor Neuroscience Group and Unit of Functional Neurosurgery, Sobell Department of Motor Neuroscience and Movement Disorders, UCL Institute of Neurology, The National Hospital for Neurology and Neurosurgery, London, WC1N 3BG, UK
Ahmed A. Moustafa*, Srinivasa Chakravarthy, Joseph R. Phillips, Jacob J. Crouse, Ankur Gupta, Michael J. Frank, Julie M. Hall and Marjan Jahanshahi
Interrelations between cognitive dysfunction and motor symptoms of Parkinson’s disease: behavioral and neural studiesDOI 10.1515/revneuro-2015-0070Received December 15, 2015; accepted February 21, 2016; previously published online March 16, 2016
Abstract: Parkinson’s disease (PD) is characterized by a range of motor symptoms. Besides the cardinal symptoms (tremor, bradykinesia/akinesia, and rigidity), PD patients also show other motor deficits, including gait disturbance, speech deficits, and impaired handwriting. However, along with these key motor symptoms, PD patients also experience cognitive deficits in attention, executive func-tion, working memory, and learning. Recent evidence sug-gests that these motor and cognitive deficits of PD are not completely dissociable, as aspects of cognitive dysfunc-tion can impact motor performance in PD. In this article, we provide a review of behavioral and neural studies on the associations between motor symptoms and cognitive deficits in PD, specifically akinesia/bradykinesia, tremor, gait, handwriting, precision grip, and speech production. This review paves the way for providing a framework for understanding how treatment of cognitive dysfunction, for example cognitive rehabilitation programs, may in turn influence the motor symptoms of PD.
Keywords: akinesia; bradykinesia; cognitive dysfunction; cognitive training; freezing of gait; grip force; Parkinson’s disease; postural instability; tremor.
IntroductionParkinson’s disease (PD) is a degenerative neurological condition that affects 1–2% of the population (Kish et al., 1988). PD manifests when 60–80% of the dopaminergic neurons within the substantia nigra pars compacta (SNc) are degenerated, initially disrupting midbrain regions such as the thalamus and subthalamic nucleus, basal ganglia, globus pallidus, nucleus basalis of Meynert, and substantia nigra. This disruption is then echoed through-out various cortical areas, such as the prefrontal cortex, supplementary and primary motor cortices, as well as the cerebellum. These cortical regions are viewed as being responsible for the cardinal motor symptoms of PD, including bradykinesia, resting tremor, rigidity, gait dis-turbance, and postural instability (Jankovic and Tolosa, 2007; Lees et al., 2009). While these symptoms are salient characteristics of PD, there are many underlying cognitive deficits that are not so visible. These include disturbances to memory, executive function, visuospatial perception, and language (Weintraub and Burn, 2011). Each of these functions is influenced by the basal ganglia (Monchi et al., 2000; Cameron et al., 2010; Macoir et al., 2013; Caproni et al., 2014). The basal ganglia are connected with cortical regions via a complex network of parallel circuits. Five basal ganglia-thalamocortical loops have been recognized, comprising a motor, oculomotor, dorso-lateral prefrontal, lateral orbitofrontal, and anterior cin-gulate circuit. Each basal ganglia-thalamocorticol circuit receives inputs from these cortical regions and traverses specific regions of the basal ganglia and thalamus nuclei and projecting back to the cortex (Alexander et al., 1986). The operation of these circuits as fully segregated parallel loops has been challenged (Joel and Weiner, 1997), with more recent evidence supporting a degree of integration
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

536 A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism
of information across the basal ganglia-thalamocortical circuits (Haynes and Haber, 2013), allowing for interaction between the motor and cognitive circuits. The integration of these two circuits suggests that damage to one would manifest in both cognitive and motor deficits. It also sug-gests that treatment of cognitive deficits alongside physi-cal therapy would be highly beneficial for the patient. Additionally, anatomical studies show that there are additional open loops (i.e. loops in which the afferent and efferent cortical areas of the basal ganglia are different) (Joel and Weiner, 1997; Graybiel, 1998). Computational models have been incorporating such open loops (Amos, 2000; Djurfeldt et al., 2001). Further, besides the frontal cortex, the occipital cortex (Seger, 2013) and medial tem-poral cortex (Middleton and Strick, 1996) also receive pro-jections from the basal ganglia. It has been suggested that basal ganglia projections to these areas support visual and memory processes (Middleton and Strick, 1996; Seger, 2013). Basal ganglia projections to the prefrontal cortex has been suggested to play a role in working memory (Moustafa and Maida, 2007; Moustafa et al., 2008).
Indeed, a strong relationship between motor and cog-nitive processes is evident. For example, different corti-cal areas play a role in the cognitive processes that guide movement, including motor planning and goal-based motor processes (Matsumoto et al., 2003; Mushiake et al., 2006; Riggeal et al., 2007; Colman et al., 2009; Smulders et al., 2012; Wylie et al., 2012). Various studies in healthy individuals suggest a strong relationship between motor and cognitive processes (Matsumoto et al., 2003; Mushiake et al., 2006; Riggeal et al., 2007; Colman et al., 2009; Smulders et al., 2012; Wylie et al., 2012). However, while some studies have found clear links between motor and cognitive measures (Shine et al., 2013d; Schneider et al., 2015), these data suggest that the primary motor symp-toms and cognitive deficits of PD are associated with the different clinical course of the illness. However, the inter-relationships among motor and cognitive processes in PD are not well characterized. As an example, the prefrontal cortex is important for the maintenance of information (e.g. goals) in working memory, which aids production of motor responses, including hand and eye movements (Ohbayashi et al., 2003; Moustafa et al., 2013a). Indeed, locomotive dysfunction in the elderly is associated with brain volume changes (Kostic et al., 2012; Tessitore et al., 2012) and aberrant neural activity (Matsui et al., 2005) within the prefrontal cortex, suggesting a potential rela-tionship between working memory and locomotion in PD. The dorsolateral prefrontal loop also plays a key role in motivation (Lawrence et al., 1998). So it is not surprising that some PD patients experience motivational disorders
(Aarsland et al., 2009). Motivation places a value on per-ceived goals, so low motivation levels would affect goal-directed behaviors, which may exacerbate several PD symptoms, such as bradykinesia and gait impairments, which are discussed in the following sections. To our knowledge, the effect of motivation on these motor pro-cesses has not been studied in PD.
Currently, while the presence of the core motor symp-toms remains essential for diagnosis, many PD patients demonstrate marked cognitive impairment even at the time of first presentation (Foltynie et al., 2004). Deficits across various cognitive domains including attention, working memory, and executive function have been described and tend to worsen with disease progression in tremor-dominant and akinesia-dominant (also known as akineto-rigid or non-tremor dominant) phenotypes and young-onset and rapid disease progression subgroups (Zetusky and Jankovic, 1985; Jankovic et al., 1990; Rajput, 1993; Schiess et al., 2000; Lewis et al., 2005; Zaidel et al., 2009; Mure et al., 2011; Poletti et al., 2011b; Schillaci et al., 2011; Lee et al., 2012; Moustafa and Poletti, 2013). Patients in the akinesia-dominant subgroup experience a more rapid disease progression compared to tremor-dominant patients (Louis et al., 1999; Eggers et al., 2012). Impor-tantly, compared to tremor-dominant patients, patients with severe akinesia are more likely to experience severe cognitive impairments or even dementia (Aarsland et al., 2003; Williams-Gray et al., 2007b; Poletti et al., 2011a, 2012).
The anatomy of the neural circuitry gone awry in PD is summarized here, to serve as a reference point to be consulted throughout the following review. To begin with, the cortex, including primary motor cortex; ventral and dorsal premotor cortex; supplementary motor areas; and rostral, dorsal, and ventral cingulate areas, have one-way excitatory glutamatergic effects with the putamen, brain-stem, and spinal cord, as well as a bidirectional excitatory relationship with the thalamus. The direct pathway of the putamen, which has an inhibitory GABAergic connection with the internal segment of the globus pallidus and sub-stantia nigra pars reticulata, receives glutamatergic exci-tation from the SNc and thalamus. The indirect pathway of the putamen has an inhibitory GABAergic effect on the external segment of the globus pallidus, which itself goes on to inhibit the internal segment of the globus pal-lidus and substantia nigra pars reticulata. The subtha-lamic nucleus is inhibited by this indirect pathway of the putamen but in turn sends excitatory signals back to the external segment of the globus pallidus, as well as to the cortex, internal segment of the globus pallidus, substan-tia nigra pars reticulata, and pedunculopontine nucleus
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism 537
(PPN), which itself finally excites the brainstem and spinal cord and bidirectionally inhibits the thalamus and its subcomponents (DeLong and Wichmann, 2007).
This review addresses cognitive-motor relationships in PD, describing how specific changes in cognitive func-tion are associated with changes in motor performance. We aim to achieve this by highlighting the specific aspects of the motor symptoms in PD that are associated with cog-nitive deficits and suggest that they may have a common pathophysiology.
Akinesia/bradykinesiaAkinesia and bradykinesia are considered among the primary motor features in PD. These symptoms are char-acterized by difficulty initiating movements, poverty of action, slowness of movements, a lack of spontaneous movement (such as swinging of the arms during walking, automatic blinking, changes of facial expression, or ges-turing during speech) as well as sequential movements, which are affected due to difficulties in transitions between subcomponents of movement (Berardelli et al., 2001). Studies have found that PD patients with akinesia have more dopamine loss in the basal ganglia than do PD patients with predominant tremor symptoms (Schil-laci et al., 2011), with basal ganglia dysfunction consid-ered to be the neural mechanism underlying bradykinesia symptoms (Berardelli et al., 2001). Dopamine is excitatory on the direct (“Go”) pathway, which facilitates respond-ing, whereas it is inhibitory on the indirect (“No Go”) pathway, suppressing a response (Frank and O’Reilly, 2006). A reduction in dopamine levels in PD therefore results in difficulty initiating movements and slowness of movements. Notably, computational and mechanistic models suggest that the source of akinesia involves hyper-excitability of striatal neurons originating in the indirect pathway leading to suppression of movement (Wiecki and Frank, 2010; Collins and Frank, 2014). In addition, cholin-ergic neurotransmission plays a critical role in controlling voluntary movement. As dopamine levels in the striatum decrease, cholinergic levels increase, suggesting that an imbalance of these neurotransmitters may underlie the motor symptoms of PD (Zhao-Shea et al., 2010).
Associations with cognitive dysfunction
Dopamine plays not only a crucial role in motor function; it also substantially influences a diversity of cognitive
processes. Therefore, it is not surprising that akinesia-dominant PD patients often show a more severe pattern of cognitive or executive dysfunction (Jankovic et al., 1990; Vakil and Herishanu-Naaman, 1998; Lewis et al., 2005; Burn et al., 2006; Lyros et al., 2008; Oh et al., 2009; Domellof et al., 2011). For example, studies have shown an association between akinesia and poor performance on tests of executive function that are dopamine depend-ent, such as probabilistic learning tasks (Moustafa et al., 2013b). This pattern was also present in other studies in which PD patients with predominant tremor were less impaired than akinesia-dominant PD s at performing procedural learning tasks, including the Tower of Hanoi task (Vakil and Herishanu-Naaman, 1998), as well as on working memory (Domellof et al., 2011; Poletti et al., 2012) and set shifting (Domellof et al., 2011, 2013; Poletti et al., 2012; Moustafa et al., 2013a). Akinesia-dominant patients were also more likely to experience bradyphrenia (slow-ness of thinking and information processing) (Rafal et al., 1984; Berardelli et al., 2001), suggesting more widespread cognitive disturbance than in the tremor-dominant type of PD.
Specifically, in non-medicated PD patients, reduced levels of dopamine impair learning from positive feed-back, while facilitating learning from negative feed-back (Wiecki and Frank, 2010; Collins and Frank, 2014). In patients on dopaminergic medication, the opposite occurs: dopamine replacement therapy facilitates learn-ing from positive feedback, but patients are impaired at learning from negative feedback because the medication blocks the effect of normal dopamine dips (Wiecki and Frank, 2010; Collins and Frank, 2014). In addition, D2 receptor blockade results in progressively worse motor performance due to a learning process dissociable from the direct performance effects of DA blockade (Wiecki et al., 2009; Beeler et al., 2012).
Working memory and executive functions required for set shifting, for example those involved in the Wis-consin Card Sorting and the Trail Making Test, are often impaired in PD patients with the akinesia dominant subtype (Domellof et al., 2011, 2013; Poletti et al., 2012; Moustafa et al., 2013a). Working memory impairment in PD may be due to a lack of appropriate regulation of the gating mechanism of the basal ganglia, impair-ing information processing in the frontostriatal system (Frank et al., 2001). Similar to reinforcement learning, “Go” and “No Go” signals can facilitate and suppress the updating of information into prefrontal working memory representations (O’Reilly and Frank, 2006). Thus, the basal ganglia is assumed to operate as a gating mechanism to modulate information updates to
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

538 A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism
the prefrontal cortex (Frank et al., 2001; O’Reilly and Frank, 2006).
Furthermore, set-shifting deficits in non-medicated PD patients are also consistent with an inability to update a new attentional set, while medicated patients show atten-tional shifting deficits when having to ignore distractors that had previously been task relevant (Moustafa et al., 2008). It has been suggested that set-shifting abnormali-ties may result from a paroxysmal over-excitation within the output nuclei of the basal ganglia circuitry (Lewis and Barker, 2009). Thus, akinesia/bradykinesia, as well as certain cognitive deficits in PD, is likely due to dopamine depletion within the basal ganglia, possibly impairing an optimal basal ganglia output selection.
Moreover, tests of working memory and attention have robust correlations with cortical cholinergic activ-ity. Cholinergic system degeneration and dysfunction are significant contributors to cognitive impairment in PD (Pagano et al., 2015). Cognitive deficits caused by pre-frontal dopaminergic changes in PD are likely to be aug-mented by a cholinergic component (Bohnen and Albin, 2011). Furthermore, the akinesia phenotype is also associ-ated with the development of dementia. Again, basal fore-brain cholinergic system degeneration appears early in PD and worsens coincident with the appearance of dementia (Bohnen and Albin, 2011; Gratwicke et al., 2015a).
Non-tremor-dominant PD patients display more severe cognitive and executive dysfunction than do patients with tremor-dominant PD. This is likely due to the more severe degradation of dopamine and choline systems in these patients as cognitive and executive function has a strong reliance on these two systems (Bohnen and Albin, 2011; Moustafa et al., 2013b; Pagano et al., 2015).
TremorThe neural substrates underlying the development of tremor in PD are still unclear. However, tremor has been associated with cerebellar, thalamic, and subthalamic nucleus abnormalities (Kassubek et al., 2002; Probst-Cousin et al., 2003; Weinberger et al., 2009; Zaidel et al., 2009; Mure et al., 2011; Helmich et al., 2012). It has also been suggested that cerebellar activation in PD patients during movement is a compensation mechanism for corticostriatal motor circuit under-activation (Yu et al., 2007; Wu and Hallett, 2013). In addition, Rosenberg-Katz et al. (2013) found that PD patients with tremor show a lower mean loss of grey matter in the pre-supplementary and primary motor areas than in non-tremor patients. A
few studies argue that tremor is dopamine independent (Benamer et al., 2000, 2003; Pirker et al., 2002; Spiegel et al., 2007).
Associations with cognitive dysfunction
Patients with the tremor-dominant subtype of PD are less often cognitively affected in comparison to akinesia-dom-inant patients. Several studies have shown that tremor is not associated with impaired cognitive function (Lewis et al., 2005; Elgh et al., 2009; Hall et al., 2014). Indeed, the frontostriatal pathways, which play a key role in cognitive deficits in PD, are relatively spared in patients with predominantly tremor symptoms (Lewis et al., 2005; Williams-Gray et al., 2007a,b).
Gait impairmentGait impairment and postural instability are hallmarks of PD. Patients with PD have reduced stride length and walking speed while double support duration and cadence rate are increased (Morris et al., 1998). During later stages of the disease, patients may also experience freezing of gait (FOG) (Lewis and Barker, 2009), wherein their walking motion ceases, often leading to falls (Rudzinska et al., 2013). Postural instability is also known to cause falls in many patients (Koller et al., 1989). PD patients may display clinical features of postural instability that are evident during quiet, undisturbed stance, such as poor anticipatory postural responses, abnormal postural sway, and inadequately organized automatic postural reactions. Performing voluntary actions, such as leaning, may reveal reduced limits of stability. Additional deficits may be evident upon disequilibrium, including slower compensatory stepping reactions, direction-specific instability, and incorrectly directed protective arm move-ments (Horak et al., 2005; Mancini et al., 2008; Grimber-gen et al., 2009; Rocchi et al., 2012). Gait impairment and postural instability are more common among PD patients with rapid disease progression (Jankovic et al., 1990), as well as in advanced-stage rather than early-stage PD (Kim et al., 2013).
Gait disorders and postural instability respond poorly to dopaminergic drugs, which suggest that they are caused by non-dopaminergic lesions (Karachi et al., 2010). It has been shown that gait and posture are under the control of cholinergic neurons of the PPN (Lee et al., 2000), which is part of a pathway involved in the initiation, acceleration,
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism 539
deceleration, and termination of locomotion. This pathway is under the control of the deep cerebellar and basal ganglia nuclei, particularly via inputs from the medial globus pallidus, substantia nigra pars reticulata, and subthalamic nucleus (Lee et al., 2000). In addition, there is accumulating evidence that implicates the basal ganglia nuclei in the performance of “automatic” behav-iors, which walking normally is, and the dysfunction of the basal ganglia in PD is proposed to interfere with auto-matic execution of walking, which is then performed in a goal-directed fashion (Redgrave et al., 2010).
Thus, tasks such as walking, which are automated in healthy individuals, require focused attention in PD patients. Aberrant neural activity (Matsui et al., 2005) and brain volume changes (Kostic et al., 2012; Tessitore et al., 2012) within the prefrontal cortex, an area that is also important for the maintenance of information (e.g. goals) in working memory (Ohbayashi et al., 2003; Moustafa et al., 2013a), also play a key role in locomo-tor dysfunction. Further, functional decoupling of goal-directed frontostriatal systems might be responsible for triggering episodes of FOG. This impaired coupling would lead to the loss of inhibitory influence over the output structures of the basal ganglia (Pahapill and Lozano, 2000; Lewis and Barker, 2009; Shine et al., 2013b), poten-tially leading to decreased activity within in areas of the mesencephalic locomotor region that coordinate gait (Pahapill and Lozano, 2000). This phenomenon might be triggered through dopaminergic depletion in the stri-atum and over-activity within the subthalamic nucleus. In addition, the inconsistent effects of dopaminergic therapy on stability in PD have led some researchers to suggest that the postural and balance deficits in PD may result from lesions to both dopaminergic and non-dopa-minergic nuclei (Bloem et al., 1996; Rocchi et al., 2002; Grimbergen et al., 2009).
Associations with cognitive dysfunction
For both the pervasive and paroxysmal gait impairments in PD, there are robust and consistent links with impair-ments in cognition. Since gait dysfunction, postural instability, and akinesia are characteristics of PD often associated with the non-tremor-dominant phenotype (Lewis et al., 2005), associations with the aforementioned cholinergic cognitive dysfunction and development of dementia also apply to gait dysfunction in PD. In addition, a clear example of shared mechanisms is the difficulty that PD patients experience with walking while perform-ing a concurrent cognitive task. For example, in healthy
participants, studies found that texting while walking can impact gait (Schabrun et al., 2014) and overall walking performance (Demura and Uchiyama, 2009), possibly due to affecting attentional performance (Woollacott and Shumway-Cook, 2002). This effect is more prominent in patients with moderate PD, as their walking becomes impaired during simultaneous performance of a simple goal-directed secondary motor task (e.g. “walk and talk”) (Bond and Morris, 2000). With the addition of a more cog-nitively demanding secondary task, larger variations in the gait parameters of PD patients are observed ( Hausdorff et al., 2003), perhaps uncovering an underlying impair-ment in the allocation of or limitation of attentional resources or in neuronal efficiency. In healthy adults, dual tasking is attention demanding too, but healthy individ-uals tend to give priority to postural tasks. PD patients, however, adopt a “posture second” approach, increasing the risk of falls (Bloem et al., 2001).
There is also growing evidence of a striking association between FOG and executive dysfunction (Amboni et al., 2008; Naismith et al., 2010; Shine et al., 2013e; Hall et al., 2014; Walton et al., 2014). During clinical assessment, the addition of a cognitive task, such as counting out loud in multiples of seven while walking along a standardized track, can severely exacerbate the frequency and sever-ity of freezing (Yogev et al., 2005; Ricciardi et al., 2014). Other studies have found that walking dysfunction in PD is related to difficulty in resolving response interference produced by distractors (Plotnik et al., 2011; Vandenbos-sche et al., 2011). Interestingly, individuals with freezing also suffer from similar deficits in cognitive tasks that require multiple shifts between cognitive sets (Naismith et al., 2010; Shine et al., 2012; Hall et al., 2014), suggesting that the pathophysiological mechanisms underlying FOG may reflect impairments in cognitive processing under temporal pressure (Shine et al., 2013c). An explanation for the concurrence of cognitive and ambulatory deficits in PD may lie in impaired compensatory processes within the brain. The loss of automatic function would necessar-ily force an individual to use more cognitive architecture to accomplish what would otherwise be routine tasks (such as walking), thereby decreasing the individuals’ capacity to flexibly deal with changes in their surround-ing environment.
Locomotive dysfunction is due to aberrant neural activity (Matsui et al., 2005) and brain volume changes (Kostic et al., 2012; Tessitore et al., 2012) within the pre-frontal cortex, an area that is also important for the main-tenance of information (e.g. goals) in working memory (Ohbayashi et al., 2003; Moustafa et al., 2013a). Addi-tionally, the nucleus basalis of Meynert is located in the
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

540 A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism
forebrain, and its cholinergic neurons are known loci of degeneration in PD (for reviews, see Gratwicke et al., 2015a,b), with more severe cholinergic loss being associ-ated with cognitive impairment. In PD, neuropathology in the nucleus basalis of Meynert leads to impairment of both arousal and selective attention, thus mediating some components of cognitive dysfunction and dementia.
In sum, there are at least two main cognitive deficits involved in gait impairments. The first is damage to the basal ganglia, which results in impairment to “automatic process,” such as movement. This results in PD patients requiring more attentional resources to initiate and maintain the walking motion and postural stability. Gait impairment is much more common in akinesia/bradykin-esia-dominant PD patients, implying that they also suffer the same severe cognitive and executive dysfunction. This limitation on their attentional resources, combined with the increased load required for walking, results in the gait impairments and freezing observed in this group of PD patients.
HandwritingHandwriting involves the coordination of finger move-ments as well as integration of sensory input to guide suc-cessful motor output. Evidence suggests that handwriting can be used as an early detection tool for PD (Rosenblum et al., 2013), as PD patients typically exhibit a diminutive form of handwriting known as micrographia. In an early study on PD micrographia, McLennan et al. (1972) found that it is present in a large proportion of PD patients and is dissociable from other PD motor symptoms includ-ing tremor and rigidity. In PD, micrographia may be an example of a motor decrement where amplitude is lost with repetitive movements. In addition to micrographia, handwriting in PD is characterized by a jagged contour and sharp fluctuations in velocity and acceleration pro-files (Van Gemmert et al., 1999; Teulings et al., 2002). Aspects of handwriting including stroke size, peak accel-eration, stroke duration, ratio between mean and stand-ard deviation of stroke length, or duration have been used for PD diagnostics (Teulings and Stelmach, 1991; Phillips et al., 1993). Compared to healthy controls, PD patients were found to exhibit increased movement time, reduced maximum and minimum values of magnitude of pen velocity, and more velocity inversions.
Studies found that inhibition of the supplementary cortex has been shown to improve handwriting in PD (Randhawa et al., 2013). Dopaminergic treatment in PD
patients resulted in marked improvements of the kin-ematics of handwriting movements (Tucha et al., 2006). Indeed, through computational modeling, reducing DA levels to the basal ganglia, substantia nigra, and globus pallidus external pathways also leads to a reduction in letter size, as seen in PD (Gangadhar et al., 2008; for a review, see Helie et al., 2013).
Associations with cognitive dysfunction
In daily life, during handwriting, mental load is often present, such as simultaneous calculations or memory recall. Similar to walking, it was found that handwriting relies on cognitive processing (Thomassen and Teulings, 1985) and is sensitive to the availability of cognitive resources. For example, it was found that PD patients took significantly longer to write while dual tasking (Van Gemmert et al., 1998). However, with the provision of external cues (e.g. lines for writing), the symptoms of micrographia in PD were lessened (Oliveira et al., 1997). It is suggested that in PD patients who experience difficul-ties with performing automatic movements such as hand-writing, adding a secondary task may increase mental load, causing a reduction in writing size, because the writing task must then be performed in a more automated manner to free resources to fulfill the requirements of the concurrently performed secondary task (Van Gemmert et al., 1998).
In PD, small handwriting was found to correlate with disease severity, motor abnormalities, as well as cognitive impairment as measured by the Mini-Mental State Exami-nation (Wagle Shukla et al., 2012). While writing relies on sequential motor processing, that is the coordination of different fingers (Catalan et al., 1999), to our knowledge, no study has tested the relationship between these two domains in PD. However, it is not known whether this deficit is an example of a more general deficit in dual tasking and task switching and how it is affected by dopa-minergic medication.
A possible mechanism explaining this reduction in the size of handwriting in PD patients may be deficits in action-perception loops or changes in sensory feedback (e.g. visual or proprioceptive). Indeed, PD patients show a discrepancy between visual and kinesthetic feedback (Teulings et al., 2002). This may be due to a degenera-tion of DA neurons in the nigro-striatal pathway, which may lead to impairment in scaling the size (amplitude) of movements (Contreras-Vidal and Stelmach, 1995; Contreras-Vidal et al., 1995). Despite the linguistic com-ponent of writing, the association between cognition and
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism 541
motor performance is unclear for micrographia. However, importantly, the impact of optimal response selection of the basal ganglia that influences many cognitive and motor performances is also apparent in handwriting.
A number of studies have examined the role of feed-back in handwriting. Since complex movement emerges out of perception-action interactions, changes in sensory feedback (e.g. visual or proprioceptive) are likely to intro-duce corresponding changes in motor output, such as in handwriting. Teulings et al. (2002) controlled visual feedback for healthy controls and early-stage PD patients such that only vertical gain of handwriting was altered, while leaving horizontal gain intact. Both healthy con-trols and PD patients showed gradual adaptation that compensated for distorted visual feedback. Another study on the effect of delayed visual feedback on handwriting found that most PD patients learned to adjust to feedback delay analogous to healthy controls (Smith and Fucetola, 1995). Of the five patients who participated in the study, only one who suffered from micrographia exhibited dif-ficulty in adapting to delayed feedback, suggesting the further studies employing a large sample of PD patients with micrographia are needed to investigate the nature of sensory feedback and micrographia.
In sum, similar to posture and gait, impairments in handwriting in PD patients may be partly due to poor dual task control. However, there may also be an impairment of sequential motor processing that can contribute to handwriting deficits in PD patients, but more research is needed to confirm or refute this link.
Precision gripPrecision grip (PG) is defined as a grip formed by one finger (typically the index finger) and thumb to hold a small object (Napier, 1956). This form of grip enables us to make (Marzke, 1997; Susman, 1998) and dexterously use tools (Moyà-Solà et al., 1999; Ambrose, 2001; Young, 2003; Jones and Lederman, 2006).
Although sensorimotor control may seem to func-tion independently of the cognitive system (Flanagan and Beltzner, 2000), various studies reveal an overlap between the motor areas for skilled movements, percep-tual, and cognitive functions (Olivier et al., 2007; Grafton, 2010). For example, significant influence of a word’s physical representation like apple or grape (Glover et al., 2004) and numerical magnitude like numbers 1 or 9 (Badets et al., 2007; Andres et al., 2008) affect grip motor control.
Associations with cognitive dysfunction
In concurrent motor and perceptual task performance, individuals show longer reaction times, delayed and less accurate grip size adjustment, and impaired perpetual task performance. This indicates that motor and percep-tual systems share overlapping attentional resources (Hesse and Deubel, 2011; Hesse et al., 2012). The addition of a delay to an occluded target dual-task increases the grip aperture, showing that motor, visual processing, and memory utilize similar resources (Singhal et al., 2007). While attention, visual processing, memory, and motor processing employ overlapping resources, only limited aspects of PG are affected in a PG-lift task with increased cognitive load. Using a task involving PG-lift task and a complex visual search combined with counting, it was found that a longer preload phase, a higher peak and static grip force, and a higher safety margin was employed in the combined motor-cognitive task compared to motor task alone (Guillery et al., 2013). Therefore, the PG and cognitive tasks utilize similar resources for processing information.
Impairment in PG in PD has been attributed to two pos-sible cognitive deficits, the first being poor sensory-motor coordination. There are multiple studies supporting a sen-sorimotor impairment in PD patients (Shine et al., 2013a; Diaz-Hung et al., 2014). Fellows et al. (1998) have dem-onstrated that this impairment is evident in PD patients due to excessive grip levels and delayed lifting actions. However, Gupta et al. (2013) suggest that the impairment may lie in risk-taking behaviors as part of impulse control disorders (Voon et al., 2011; Catalan et al., 2013) that are prevalent in approximately 36% of PD patients (Callesen et al., 2014). According to Gupta et al. (2013), an excessive grip is an indication of an increased “safety margin” to ensure the object does not fall. This effect is more salient in PD patients while off, than on, medication, indicat-ing higher risk behavior, which is in line with research on the interrelationship between impulsivity and dopa-mine medications (Catalan et al., 2013; Piray et al., 2014). Further research aimed at investigating sensory-motor coordination and impulsive behaviors would provide greater insight into the interaction of cognition and motor control and help identify which cognitive impairments affect PG.
Speech problemsMost PD patients develop speech and voice disorders at some point during their illness (Ho et al., 1998), including
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

542 A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism
stuttering and difficulty initiating speech. Speech in PD is often hypophonic (i.e. hypokinetic, soft speech), monotonic (i.e. speech quality tends to be soft, hoarse, and monotonous), and/or festinating (i.e. excessively rapid, soft, poorly intelligible speech) (Jankovic, 2008). Dysarthria, which refers to slurred, slow, and difficult-to-understand speech, is also common in PD. There are few studies investigating neural substrates of speech dysfunction in PD. In one study in PD, it was found that the basal ganglia plays a role in speech production and syntactic ability (Lieberman et al., 1992) as well as phone-mic and semantic fluency test performance (Ellfolk et al., 2014). One recent study found that bradykinesia affects the prosody of the patients’ speech (Azevedo et al., 2013), perhaps suggesting a link between speech production and dopamine in PD.
Associations with cognitive dysfunction
Studies have found that speech and cognitive processes are not necessarily dissociable (Altmann et al., 2001; Wiseheart et al., 2009). For example, prior studies have found a correlation between verbal and cognitive meas-ures (Lieberman et al., 1992). A recent review has also highlighted the relationship between speech production and cognitive measures in PD (Altmann and Troche, 2011). In PD, evidence suggests that both working memory and executive function processes affect speech production (Troche and Lori, 2012). Verb production also correlates with cognitive measures, such as working memory in PD (Colman et al., 2009).
DiscussionThis review investigated the relationship between cog-nitive processes and motor symptoms in PD, including akinesia/bradykinesia, tremor, rigidity, handwriting, and speech problems. Not surprisingly, cognitive impair-ments in PD are strongly related to some motor symptoms more than others are. This might be due to some cognitive processes that rely on the integrity of the basal ganglia-thalamocortical loops along with some motor operations, while others do not. In addition, as discussed above, there is evidence of an interaction between the cogni-tive and motor loops in the basal ganglia, as opposed to each being fully segregated. For example, PD patients with the akinesia-dominant phenotype are more likely to suffer executive dysfunction, bradyphrenia (i.e. slowness
of thinking and information processing), and dementia than those with the tremor-dominant phenotype. Tremors have been suggested to not be associated with cognitive dysfunction, with the frontostriatal pathways important for healthy cognition appearing relatively spared in those patients suffering predominantly from tremors. Gait dys-function, which is often associated with akinesia in non-tremor-dominant patients, also appears to be linked with cognitive dysfunction. Not only are attentional deficits during locomotion evident, but there is also mounting evidence of a strong association between the episodic symptom of FOG and executive dysfunction. The relation-ship between cognitive and motor processes in PD is less clear for handwriting and speech. Given that both hand-writing and speech have a linguistic component, the lack of a clear association with cognitive dysfunction in PD is somewhat surprising.
Focusing future studies on these weaker relationships may shed some light on the interaction of cognition and motor symptoms in PD. Such research might additionally attempt to investigate these cognitive-motor symptom relationships beginning at early, preclinical stages, at which symptoms are just evident, and follow the trajec-tory of these unfolding impairments and the temporal and pathophysiological relationships to other symptoms. Another direction for research might be to attempt to break down motor processes into small component parts, to elu-cidate what types of cognitive processes are involved at each part and how cognitive dysfunction might contribute to motor impairment. The unraveling of such cognitive-motor associations is of importance as it may lead to the possibility of active interventions such as cognitive train-ing programs for the treatment of motor impairments, including akinesia/bradykinesia, gait dysfunction, and FOG in PD patients and also older adults.
While the focus of this paper has been on the interac-tion of cognition and motor function in PD, there is also evidence for motivational modulation of movement speed in PD from behavioral (Ballanger et al., 2006; Shiner et al., 2012; Kojovic et al., 2014; MacDonald and Byblow, 2015) and imaging studies (Ballanger et al., 2008). Regions of the cerebral cortex that are engaged in motivation, cogni-tion, and sensorimotor control, as well as brainstem struc-tures, feed into the striatum and the subthalamic nucleus, the principal input nuclei of the basal ganglia. This input is important for action selection in different motor and nonmotor domains by inhibiting irrelevant internal and external stimuli. Given the structural and functional simi-larity in information processing across the partially seg-regated motor, cognitive, and limbic circuits (Alexander et al., 1986), some degree of interaction and integration
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism 543
of information across circuits may occur (Joel and Weiner, 1994; Haynes and Haber, 2013). Thus, at least for some of the motor symptoms of PD, an imbalance between neu-rotransmitters such as dopamine and acetylcholine may underlie deficits with inhibiting irrelevant and selecting optimal responses by the basal ganglia (Aosaki et al., 2010), causing both motor and cognitive impairments.
ReferencesAarsland, D., Andersen, K., Larsen, J.P., Lolk, A., and
Kragh-Sorensen, P. (2003). Prevalence and characteristics of dementia in Parkinson disease: an 8-year prospective study. Arch. Neurol. 60, 387–392.
Aarsland, D., Bronnick, K., Alves, G., Tysnes, O.B., Pedersen, K.F., Ehrt, U., and Larsen J.P. (2009). The spectrum of neuropsychiatric symptoms in patients with early untreated Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 80, 928–930.
Alexander, G.E., DeLong, M.R., and Strick, P.L. (1986). Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 9, 357–381.
Altmann, L.J. and Troche, M.S. (2011). High-level language production in Parkinson’s disease: a review. Parkinsons Dis. 2011, Article ID 238956, 12.
Altmann, L.J., Kempler, D., and Andersen, E.S. (2001). Speech errors in Alzheimer’s disease: reevaluating morphosyntactic preservation. J. Speech Lang. Hear. Res. 44, 1069–1082.
Amboni, M., Cozzolino, A., Longo, K., Picillo, M., and Barone, P. (2008). Freezing of gait and executive functions in patients with Parkinson’s disease. Mov. Disord. 23, 395–400.
Ambrose, S.H. (2001). Paleolithic technology and human evolution. Science 291, 1748–1753.
Amos, A. (2000). A computational model of information processing in the frontal cortex and basal ganglia. J. Cogn. Neurosci. 12, 505–519.
Andres, M., Ostry, D.J., Nicol, F., and Paus, T. (2008). Time course of number magnitude interference during grasping. Cortex 44, 414–419.
Aosaki, T., Miura, M., Suzuki, T., Nishimura, K., and Masuda, M. (2010). Acetylcholine-dopamine balance hypothesis in the striatum: an update. Geriatr. Gerontol. Int. 10, S148–S157.
Azevedo, L.L.d., Reis, C.A.d.C., Souza, I.S.d., and Cardoso, F.E.C. (2013). Prosody and levodopa in Parkinson’s disease. Arq. Neuropsiquiatr. 71, 835–840.
Badets, A., Andres, M., Di Luca, S., and Pesenti, M. (2007). Number magnitude potentiates action judgements. Exp. Brain Res. 180, 525–534.
Ballanger, B., Thobois, S., Baraduc, P., Turner, R.S., Broussolle, E., and Desmurget, M. (2006). “Paradoxical kinesis” is not a hallmark of Parkinson’s disease but a general property of the motor system. Mov. Disord. 21 1490–1495.
Ballanger, B., Baraduc, P., Broussolle, E., Bars, D.L., Desmurget, M., and Thobois, S. (2008). Motor urgency is mediated by the contralateral cerebellum in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 79, 1110–1116.
Beeler, J.A., Frank, M.J., McDaid, J., Alexander, E., Turkson, S., Bernardez Sarria, M.S., McGehee, D.S., and Zhuang, X. (2012). A role for dopamine-mediated learning in the pathophysiology and treatment of Parkinson’s disease. Cell Rep. 2, 1747–1761.
Benamer, H.T., Patterson, J., Wyper, D.J., Hadley, D.M., Macphee, G.J., and Grosset, D.G. (2000). Correlation of Parkinson’s disease severity and duration with 123I-FP-CIT SPECT striatal uptake. Mov. Disord. 15, 692–698.
Benamer, H.T., Oertel, W.H., Patterson, J., Hadley, D.M., Pogarell, O., Hoffken, H., Gerstner, A., and Grosset, D.G. (2003). Prospective study of presynaptic dopaminergic imaging in patients with mild parkinsonism and tremor disorders: part 1. Baseline and 3-month observations. Mov. Disord. 18, 977–984.
Berardelli, A., Rothwell, J.C., Thompson, P.D., and Hallett, M. (2001). Pathophysiology of bradykinesia in Parkinson’s disease. Brain 124, 2131–2146.
Bloem, B.R., Beckley, D.J., van Dijk, J.G., Zwinderman, A.H., Remler, M.P., and Roos, R.A. (1996). Influence of dopaminergic medication on automatic postural responses and balance impairment in Parkinson’s disease. Mov. Disord. 11, 509–521.
Bloem, B.R., Valkenburg, V.V., Slabbekoorn, M., and van Dijk, J.G. (2001). The multiple tasks test. Strategies in Parkinson’s dis-ease. Exp. Brain Res. 137, 478–486.
Bohnen, N.I. and Albin, R.L. (2011). The cholinergic system and Parkinson disease. Behav. Brain Res. 221, 564–573.
Bond, J.M. and Morris, M. (2000). Goal-directed secondary motor tasks: their effects on gait in subjects with Parkinson disease. Arch. Phys. Med. Rehabil. 81, 110–116.
Burn, D.J., Rowan, E.N., Allan, L.M., Molloy, S., O’Brien, J.T., and McKeith, I.G. (2006). Motor subtype and cognitive decline in Parkinson’s disease, Parkinson’s disease with dementia, and dementia with Lewy bodies. J. Neurol. Neurosurg. Psychiatry 77, 585–589.
Callesen, M.B., Weintraub, D., Damholdt, M.F., and Moller, A. (2014). Impulsive and compulsive behaviors among Danish patients with Parkinson’s disease: prevalence, depression, and personality. Parkinsonism Relat. Disord. 20, 22–26.
Cameron, I.G., Watanabe, M., Pari, G., and Munoz, D.P. (2010). Executive impairment in Parkinson’s disease: response automaticity and task switching. Neuropsychologia 48, 1948–1957.
Caproni, S., Muti, M., Di Renzo, A., Principi, M., Caputo, N., Calabresi, P., and Tambasco, N. (2014). Subclinical visuospatial impairment in Parkinson’s disease: the role of basal ganglia and limbic system. Front. Neurol. 5, 7.
Catalan, M.J., Ishii, K., Honda, M., Samii, A., and Hallett, M. (1999). A PET study of sequential finger movements of varying length in patients with Parkinson’s disease. Brain 122, 483–495.
Catalan, M.J., de Pablo-Fernandez, E., Villanueva, C., Fernandez-Diez, S., Lapena-Montero, T., Garcia-Ramos, R., and López-Valdés, E. (2013). Levodopa infusion improves impulsivity and dopamine dysregulation syndrome in Parkinson’s disease. Mov. Disord. 28, 2007–2010.
Collins, A.G. and Frank, M.J. (2014). Opponent actor learning (OpAL): modeling interactive effects of striatal dopamine on reinforce-ment learning and choice incentive. Psychol. Rev. 121, 337–366.
Colman, K.S., Koerts, J., van Beilen, M., Leenders, K.L., Post, W.J., and Bastiaanse, R. (2009). The impact of executive functions on verb production in patients with Parkinson’s disease. Cortex 45, 930–942.
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

544 A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism
Contreras-Vidal, J.L. and Stelmach, G.E. (1995). A neural model of basal ganglia-thalamocortical relations in normal and parkinsonian movement. Biol. Cybern. 73, 467–476.
Contreras-Vidal, J.L., Teulings, H.L., and Stelmach, G.E. (1995). Micro-graphia in Parkinson’s disease. Neuroreport 6, 2089–2092.
DeLong, M.R. and Wichmann, T. (2007). Circuits and circuit disorders of the basal ganglia. Arch. Neurol. 64, 20–24.
Demura, S. and Uchiyama, M. (2009). Influence of cell phone email use on characteristics of gait. Eur. J. Sport Sci. 9, 303–309.
Diaz-Hung, M.L., Blanco, L., Pavon, N., Leon, R., Estupinan, B., Orta, E., MartíDnez, K., and Fernández, I. (2014). Sensory-motor performance after acute glutathione depletion by l-buthionine sulfoximine injection into substantia nigra pars compacta. Behav. Brain Res. 271, 286–293.
Djurfeldt, M., Ekeberg, Ö., and Graybiel, A.M. (2001). Cortex-basal ganglia interaction and attractor states. Neurocomputing 38–40, 573–579.
Domellof, M.E., Elgh, E., and Forsgren, L. (2011). The relation between cognition and motor dysfunction in drug-naive newly diagnosed patients with Parkinson’s disease. Mov. Disord. 26, 2183–2189.
Domellof, M.E., Forsgren, L., and Elgh, E. (2013). Persistence of associations between cognitive impairment and motor dysfunction in the early phase of Parkinson’s disease. J. Neurol. 260, 2228–2236.
Eggers, C., Pedrosa, D.J., Kahraman, D., Maier, F., Lewis, C.J., Fink, G.R., Schmidt, M., and Timmermann, L. (2012). Parkinson subtypes progress differently in clinical course and imaging pattern. PLoS One 7, e46813, 8.
Elgh, E., Domellof, M., Linder, J., Edstrom, M., Stenlund, H., and Forsgren, L. (2009). Cognitive function in early Parkinson’s disease: a population-based study. Eur. J. Neurol. 16, 1278–1284.
Ellfolk, U., Joutsa, J., Rinne, J.O., Parkkola, R., Jokinen, P., and Karrasch, M. (2014). Striatal volume is related to phonemic verbal fluency but not to semantic or alternating verbal fluency in early Parkinson’s disease. J. Neural. Transm. 121, 33–40.
Fellows, S.J., Noth, J., and Schwarz, M. (1998). Precision grip and Parkinson’s disease. Brain 121, 1771–1784.
Flanagan, J.R. and Beltzner, M.A. (2000). Independence of perceptual and sensorimotor predictions in the size-weight illusion. Nat. Neurosci. 3, 737–741.
Foltynie, T., Brayne, C.E., Robbins, T.W., and Barker, R.A. (2004). The cognitive ability of an incident cohort of Parkinson’s patients in the UK. The CamPaIGN study. Brain 127, 550–560.
Frank, M.J. and O’Reilly, R.C. (2006). A mechanistic account of striatal dopamine function in human cognition: psychopharmacological studies with cabergoline and haloperidol. Behav. Neurosci. 120, 497–517.
Frank, M.J., Loughry, B., and O’Reilly, R.C. (2001). Interactions between frontal cortex and basal ganglia in working memory: a computational model. Cogn. Affect. Behav. Neurosci. 1, 137–160.
Gangadhar, G., Joseph, D., and Chakravarthy, V.S. (2008). Understanding Parkinsonian handwriting through a computational model of basal ganglia. Neural Comput. 20, 2491–2525.
Glover, S., Rosenbaum, D.A., Graham, J., and Dixon, P. (2004). Grasping the meaning of words. Exp. Brain Res. 154, 103–108.
Grafton, S.T. (2010). The cognitive neuroscience of prehension: recent developments. Exp. Brain Res. 204, 475–491.
Gratwicke, J., Jahanshahi, M., and Foltynie, T. (2015a). Parkinson’s disease dementia: a neural networks perspective. Brain 138, 1454–1476.
Gratwicke, J., Kahan, J., Zrinzo, L., Hariz, M., Limousin, P., Foltynie, T., and Jahanshahi, M. (2015b). The nucleus basalis of Meynert: a new target for deep brain stimulation in dementia? Neurosci. Rev. 37, 2676–2688.
Graybiel, A.M. (1998). The basal ganglia and chunking of action repertoires. Neurobiol. Learn. Mem. 70, 119–136.
Grimbergen, Y.A., Langston, J.W., Roos, R.A., and Bloem, B.R. (2009). Postural instability in Parkinson’s disease: the adrenergic hypothesis and the locus coeruleus. Exp. Rev. Neurother. 9, 279–290.
Guillery, E., Mouraux, A., and Thonnard, J.-L. (2013). Cognitive-motor interference while grasping, lifting and holding objects. PLoS One 8, e80125, 8.
Gupta, A., Balasubramani, P.P., and Chakravarthy, S. (2013). Computa tional model of precision grip in Parkinson’s disease: a utility based approach [original research]. Front. Computat. Neurosci. 7, 16.
Hall, J.M., Shine, J.M., Walton, C.C., Gilat, M., Kamsma, Y.P., Naismith, S.L., and Lewis, S.J. (2014). Early phenotypic differences between Parkinson’s disease patients with and without freezing of gait. Parkinsonism Relat. Disord. 20, 604–607.
Hausdorff, J.M., Schaafsma, J.D., Balash, Y., Bartels, A.L., Gurevich, T., and Giladi, N. (2003). Impaired regulation of stride variability in Parkinson’s disease subjects with freezing of gait. Exp. Brain Res. 149, 187–194.
Haynes, W.I. and Haber, S.N. (2013). The organization of prefrontal-subthalamic inputs in primates provides an anatomical substrate for both functional specificity and integration: implications for basal ganglia models and deep brain stimulation. J. Neurosci. 33, 10.
Helie, S., Chakravarthy, S., and Moustafa, A.A. (2013). Exploring the cognitive and motor functions of the basal ganglia: an integrative review of computational cognitive neuroscience models. Front. Comput. Neurosci. 7, 174, 16.
Helmich, R.C., Hallett, M., Deuschl, G., Toni, I., and Bloem, B.R. (2012). Cerebral causes and consequences of parkinso-nian resting tremor: a tale of two circuits? Brain 135, 3206–3226.
Hesse, C. and Deubel, H. (2011). Efficient grasping requires attentional resources. Vision Res. 51, 1223–1231.
Hesse, C., Schenk, T., and Deubel, H. (2012). Attention is needed for action control: further evidence from grasping. Vision Res. 71, 37–43.
Ho, A.K., Iansek, R., Marigliani, C., Bradshaw, J.L., and Gates, S. (1998). Speech impairment in a large sample of patients with Parkinson’s disease. Behav. Neurol. 11, 131–137.
Horak, F.B., Dimitrova, D., and Nutt, J.G. (2005). Direction-specific postural instability in subjects with Parkinson’s disease. Exp. Neurol. 193, 504–521.
Jankovic, J. (2008). Parkinson’s disease: clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 79, 368–376.
Jankovic, J. and Tolosa, E. (2007). Parkinson’s Disease and Movement Disorders (Philadelphia: Lippincott Williams and Wilkins).
Jankovic, J., McDermott, M., Carter, J., Gauthier, S., Goetz, C., Golbe, L., Huber, S., Koller, W., Olanow, C., Shoulson, I., et al. (1990). Variable expression of Parkinson’s disease:
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism 545
a base-line analysis of the DATATOP cohort. The Parkinson Study Group. Neurology 40, 1529–1534.
Joel, D. and Weiner, I. (1994). The organization of the basal ganglia-thalamocortical circuits: open interconnected rather than closed segregated. Neuroscience 63, 363–379.
Joel, D. and Weiner, I. (1997). The connections of the primate subthalamic nucleus: indirect pathways and the open-interconnected scheme of basal ganglia-thalamocortical circuitry. Brain Res. Brain Res. Rev. 23, 62–78.
Jones, L.A. and Lederman, S.J. (2006). Human Hand Function (New York: Oxford University Press).
Karachi, C., Grabli, D., Bernard, F.A., Tande, D., Wattiez, N., Belaid, H., Bardinet, E., Prigent, A., Nothacker, H.P., Hunot, S., et al. (2010). Cholinergic mesencephalic neurons are involved in gait and postural disorders in Parkinson disease. J. Clin. Invest. 120, 2745–2754.
Kassubek, J., Juengling, F.D., Hellwig, B., Spreer, J., and Lucking, C.H. (2002). Thalamic gray matter changes in unilateral Parkinsonian resting tremor: a voxel-based morphometric analysis of 3-dimensional magnetic resonance imaging. Neurosci. Lett. 323, 29–32.
Kim, S.D., Allen, N.E., Canning, C.G., and Fung, V.S. (2013). Postural instability in patients with Parkinson’s disease. Epidemiology, pathophysiology and management. CNS Drugs 27, 97–112.
Kish, S.J., Shannak, K., and Hornykiewicz, O. (1988). Uneven pattern of dopamine loss in the striatum of patients with idiopathic Parkinson’s disease. Pathophysiologic and clinical implications. N. Engl. J. Med. 318, 876–880.
Kojovic, M., Mir, P., Trender-Gerhard, I., Schneider, S.A., Pareés, I., Edwards, M.J., Bhatia, K.P., and Jahanshahi, M. (2014). Motivational modulation of bradykinesia in Parkinson’s disease off and on dopaminergic medication. J. Neurol. 261, 1080–1089.
Koller, W.C., Glatt, S., Vetere-Overfield, B., and Hassanein, R. (1989). Falls and Parkinson’s disease. Clin. Neuropharmacol. 12, 98–105.
Kostic, V.S., Agosta, F., Pievani, M., Stefanova, E., Jecmenica-Lukic, M., Scarale, A., Spica, V., and Filippi, M. (2012). Pattern of brain tissue loss associated with freezing of gait in Parkinson disease. Neurology 78, 409–416.
Lawrence, A.D., Sahakian, B.J., and Robbins, T.W. (1998). Cognitive functions and corticostriatal circuits: insights from Hunting-ton’s disease. Trends Cogn. Sci. 2, 379–388.
Lee, M.S., Rinne, J.O., and Marsden, C.D. (2000). The pedunculopontine nucleus: its role in the genesis of movement disorders. Yonsei Med. J. 41, 167–184.
Lee, J.M., Koh, S.B., Chae, S.W., Seo, W.K., Kwon do, Y., Kim, J.H., Oh, K., Baik, J.S., and Park, K.W. (2012). Postural instability and cognitive dysfunction in early Parkinson’s disease. Can. J. Neurol. Sci. 39, 473–482.
Lees, A.J., Hardy, J., and Revesz, T. (2009). Parkinson’s disease. Lancet 373, 2055–2066.
Lewis, S.J. and Barker, R.A. (2009). A pathophysiological model of freezing of gait in Parkinson’s disease. Parkinsonism Relat. Disord. 15, 333–338.
Lewis, S.J.G., Foltynie, T., Blackwell, A.D., Robbins, T.W., Owen, A.M., and Barker, R.A. (2005). Heterogeneity of Parkinson’s disease in the early clinical stages using a data driven approach. J. Neurol. Neurosurg. Psychiatry 76, 343–348.
Lieberman, P., Kako, E., Friedman, J., Tajchman, G., Feldman, L.S., and Jiminez, E.B. (1992). Speech production, syntax comprehension, and cognitive deficits in Parkinson’s disease. Brain Lang. 43, 169–189.
Louis, E.D., Tang, M.X., Cote, L., Alfaro, B., Mejia, H., and Marder, K. (1999). Progression of parkinsonian signs in Parkinson disease. Arch. Neurol. 56, 334–337.
Lyros, E., Messinis, L., and Papathanasopoulos, P. (2008). Does motor subtype influence neurocognitive performance in Parkinson’s disease without dementia? Eur. J. Neurol. 15, 262–267.
MacDonald, H.J. and Byblow, W.D. (2015). Does response inhibition have pre- and postdiagnostic utility in Parkinson’s disease? J. Motivated Behav. 47, 29–45.
Macoir, J., Fossard, M., Mérette, C., Langlois, M., Chantal, S., and Auclair-Ouellet, N. (2013). The role of basal ganglia in language production: evidence from Parkinson’s disease. J. Parkinsons Dis. 3, 393–397.
Mancini, M., Rocchi, L., Horak, F.B., and Chiari, L. (2008). Effects of Parkinson’s disease and levodopa on functional limits of stability. Clin. Biomech. (Bristol, Avon) 23, 450–458.
Marzke, M.W. (1997). Precision grips, hand morphology, and tools. Am. J. Phys. Anthropol. 102, 91–110.
Matsui, H., Udaka, F., Miyoshi, T., Hara, N., Tamaura, A., Oda, M., Oda, M., Kubori, T., Nishinaka, K., and Kameyama, M. (2005). Three-dimensional stereotactic surface projection study of freezing of gait and brain perfusion image in Parkinson’s disease. Mov. Disord. 20, 1272–1277.
Matsumoto, K., Suzuki, W., and Tanaka, K. (2003). Neuronal correlates of goal-based motor selection in the prefrontal cortex. Science 301, 229–232.
McLennan, J.E., Nakano, K., Tyler, H.R., and Schwab, R.S. (1972). Micrographia in Parkinson’s disease. J. Neurol. Sci. 15, 141–152.
Middleton, F.A. and Strick, P.L. (1996). The temporal lobe is a target of output from the basal ganglia. Proc. Natl. Acad. Sci. U S A 93, 8683–8687.
Monchi, O., Taylor, J.G., and Dagher, A. (2000). A neural model of working memory processes in normal subjects, Parkinson’s disease and schizophrenia for fMRI design and predictions. Neural Netw. 13, 953–973.
Morris, M., Iansek, R., Matyas, T., and Summers, J. (1998). Abnormalities in the stride length-cadence relation in parkinsonian gait. Mov. Disord. 13, 61–69.
Moustafa, A.A. and Maida, A.S. (2007). Using TD learning to simulate working memory performance in a model of the prefrontal cortex and basal ganglia. Cogn. Syst. Res. 8, 262–281.
Moustafa, A.A. and Poletti, M. (2013). Neural and behavioral substrates of subtypes of Parkinson’s disease. Front. Syst. Neurosci. 7, 117.
Moustafa, A.A., Sherman, S.J., and Frank, M.J. (2008). A dopaminergic basis for working memory, learning and attentional shifting in Parkinsonism. Neuropsychologia 46, 3144–3156.
Moustafa, A.A., Bell, P., Eissa, A.M., and Hewedi, D.H. (2013a). The effects of clinical motor variables and medication dosage on working memory in Parkinson’s disease. Brain Cogn. 82, 137–145.
Moustafa, A.A., Krishna, R., Eissa, A.M., and Hewedi, D.H. (2013b). Factors underlying probabilistic and deterministic stimulus-response learning performance in medicated and unmedicated
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

546 A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism
patients with Parkinson’s disease. Neuropsychology 27, 498–510.
Moyà-Solà, S., Köhler, M., and Rook, L. (1999). Evidence of hominid-like precision grip capability in the hand of the Miocene ape Oreopithecus. Proc. Natl. Acad. Sci. 96, 313–317.
Mure, H., Hirano, S., Tang, C.C., Isaias, I.U., Antonini, A., Ma, Y., Dhawan, V., and Eidelberg, D. (2011). Parkinson’s disease tremor-related metabolic network: characterization, progression, and treatment effects. Neuroimage 54, 1244–1253.
Mushiake, H., Saito, N., Sakamoto, K., Itoyama, Y., and Tanji, J. (2006). Activity in the lateral prefrontal cortex reflects multiple steps of future events in action plans. Neuron 50, 631–641.
Naismith, S.L., Shine, J.M., and Lewis, S.J.G. (2010). The specific contributions of set-shifting to freezing of gait in Parkinson’s disease. Mov. Disord. 25, 1000–1004.
Napier, J.R. (1956). The prehensile movements of the human hand. J. Bone Joint Surg. Br. 38-B, 902–913.
O’Reilly, R.C. and Frank, M.J. (2006). Making working memory work: a computational model of learning in the prefrontal cortex and basal ganglia. Neural Comput. 18, 283–328.
Oh, J.Y., Kim, Y.S., Choi, B.H., Sohn, E.H., and Lee, A.Y. (2009). Relationship between clinical phenotypes and cognitive impairment in Parkinson’s disease (PD). Arch. Gerontol. Geriatr. 49, 351–354.
Ohbayashi, M., Ohki, K., and Miyashita, Y. (2003). Conversion of working memory to motor sequence in the monkey premotor cortex. Science 301, 233–236.
Olivier, E., Davare, M., Andres, M., and Fadiga, L. (2007). Precision grasping in humans: from motor control to cognition. Curr. Opin. Neurobiol. 17, 644–648.
Oliveira, R.M., Gurd, J.M., Nixon, P., Marshall, J.C., and Passingham, R.E. (1997). Micrographia in Parkinson’s disease: the effect of providing external cues. J. Neurol. Neurosurg. Psychiatry 63, 429–433.
Pagano, G., Rengo, G., Pasqualetti, G., Femminella, G.D., Monzani, F., Ferrara, N., and Tagliati, M. (2015). Cholinesterase inhibitors for Parkinson’s disease: a systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry 86, 767–773.
Pahapill, P.A. and Lozano, A.M. (2000). The pedunculopontine nucleus and Parkinson’s disease. Brain 123, 1767–1783.
Phillips, J.G., Bradshaw, J.L., Iansek, R., and Chiu, E. (1993). Motor functions of the basal ganglia. Psychol. Res. 55, 175–181.
Piray, P., Zeighami, Y., Bahrami, F., Eissa, A.M., Hewedi, D.H., and Moustafa, A.A. (2014). Impulse control disorders in Parkinson’s disease are associated with dysfunction in stimulus valuation but not action valuation. J. Neurosci. 34, 7814–7824.
Pirker, W., Djamshidian, S., Asenbaum, S., Gerschlager, W., Tribl, G., Hoffmann, M., Brücke, T. (2002). Progression of dopaminergic degeneration in Parkinson’s disease and atypical parkinsonism: a longitudinal beta-CIT SPECT study. Mov. Disord. 17, 45–53.
Plotnik, M., Giladi, N., Dagan, Y., and Hausdorff, J.M. (2011). Postural instability and fall risk in Parkinson’s disease: impaired dual tasking, pacing, and bilateral coordination of gait during the “ON” medication state. Exp. Brain Res. 210, 529–538.
Poletti, M., Emre, M., and Bonuccelli, U. (2011a). Mild cognitive impairment and cognitive reserve in Parkinson’s disease. Parkinsonism Relat. Disord. 17, 579–586.
Poletti, M., Frosini, D., Pagni, C., Lucetti, C., Del Dotto, P., Tognoni, G., Ceravolo, R., and Bonuccelli, U. (2011b). The
association between motor subtypes and alexithymia in de novo Parkinson’s disease. J. Neurol. 258, 1042–1045.
Poletti, M., Frosini, D., Pagni, C., Baldacci, F., Nicoletti, V., Tognoni, G., Lucetti, C., Del Dotto, P., Ceravolo, R., and Bonuccelli, U. (2012). Mild cognitive impairment and cognitive-motor relationships in newly diagnosed drug-naive patients with Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 83, 601–606.
Probst-Cousin, S., Druschky, A., and Neundorfer, B. (2003). Disappearance of resting tremor after “stereotaxic” thalamic stroke. Neurology 61, 1013–1014.
Rafal, R.D., Posner, M.I., Walker, J.A., and Friedrich, F.J. (1984). Cognition and the basal ganglia. Separating mental and motor components of performance in Parkinson’s disease. Brain 107, 1083–1094.
Rajput, A.H. (1993). Diagnosis of PD. Neurology 43, 1629–1630.Randhawa, B.K., Farley, B.G., and Boyd, L.A. (2013). Repetitive
transcranial magnetic stimulation improves handwriting in Parkinson’s disease. Parkinsons Dis. 2013, 751925.
Redgrave, P., Rodriguez, M., Smith, Y., Rodriguez-Oroz, M.C., Lehericy, S., Bergman, H., Bergman, H., Agid, Y., DeLong, M.R., and Obeso, J.A. (2010). Goal-directed and habitual control in the basal ganglia: implications for Parkinson’s disease. Nat. Rev. Neurosci. 11, 760–772.
Ricciardi, L., Bloem, B.R., Snijders, A.H., Daniele, A., Quaranta, D., Bentivoglio, A.R., and Fasano, A. (2014). Freezing of gait in Parkinson’s disease: The paradoxical interplay between gait and cognition. Parkinsonism Relat. Disord. 20, 824–829.
Riggeal, B.D., Crucian, G.P., Seignourel, P., Jacobson, C.E., Okun, M.S., Rodriguez, R., and Fernandez, H.H. (2007). Cognitive decline tracks motor progression and not disease duration in Parkinson patients. Neuropsychiatr. Dis. Treat. 3, 955–958.
Rocchi, L., Chiari, L., and Horak, F.B. (2002). Effects of deep brain stimulation and levodopa on postural sway in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 73, 267–274.
Rocchi, L., Carlson-Kuhta, P., Chiari, L., Burchiel, K.J., Hogarth, P., and Horak, F.B. (2012). Effects of deep brain stimulation in the subthalamic nucleus or globus pallidus internus on step initiation in Parkinson disease. J. Neurosurg. 117, 1141–1149.
Rosenberg-Katz, K., Herman, T., Jacob, Y., Giladi, N., Hendler, T., and Hausdorff, J.M. (2013). Gray matter atrophy distinguishes between Parkinson disease motor subtypes. Neurology 80, 1476–1484.
Rosenblum, S., Samuel, M., Zlotnik, S., Erikh, I., and Schlesinger, I. (2013). Handwriting as an objective tool for Parkinson’s disease diagnosis. J. Neurol. 260, 2357–2361.
Rudzinska, M., Bukowczan, S., Stozek, J., Zajdel, K., Mirek, E., Chwala, W., Wójcik-Pędziwiatr, M., Banaszkiewicz, K., and Szczudlik, A. (2013). Causes and consequences of falls in Parkinson disease patients in a prospective study. Neurol. Neurochir. Pol. 47, 423–430.
Schabrun, S.M., van den Hoorn, W., Moorcroft, A., Greenland, C., and Hodges, P.W. (2014). Texting and walking: strategies for postural control and implications for safety. PLoS One 9, e84312, 8.
Schiess, M.C., Zheng, H., Soukup, V.M., Bonnen, J.G., and Nauta, H.J. (2000). Parkinson’s disease subtypes: clinical classification and ventricular cerebrospinal fluid analysis. Parkinsonism Relat. Disord. 6, 69–76.
Schillaci, O., Chiaravalloti, A., Pierantozzi, M., Di Pietro, B., Koch, G., Bruni, C., and Stefani, A. (2011). Different patterns of
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism 547
nigrostriatal degeneration in tremor type versus the akinetic-rigid and mixed types of Parkinson’s disease at the early stages: molecular imaging with 123I-FP-CIT SPECT. Int. J. Mol. Med. 28, 881–886.
Schneider, J.S., Sendek, S., and Yang, C. (2015). Relationship between motor symptoms, cognition, and demographic characteristics in treated mild/moderate Parkinson’s disease. PLoS One 10, 11.
Seger, C.A. (2013). The visual corticostriatal loop through the tail of the caudate: circuitry and function. Front. Syst. Neurosci. 7, 104.
Shine, J.M., Naismith, S.L., Palavra, N.C., Lewis, S.J., Moore, S.T., Dilda, V., and Morris, T.R. (2012). Attentional set-shifting deficits correlate with the severity of freezing of gait in Parkinson’s disease. Parkinsonism Relat. Disord. 19, 388–390.
Shine, J.M., Matar, E., Ward, P.B., Bolitho, S.J., Gilat, M., Pearson, M., Naismith, S.L., and Lewis, S.J. (2013a). Exploring the cortical and subcortical functional magnetic resonance imaging changes associated with freezing in Parkinson’s disease [randomized controlled trial research support, non-U.S. Gov’t]. Brain 136, 1204–1215.
Shine, J.M., Matar, E., Ward, P.B., Bolitho, S.J., Pearson, M., Naismith, S.L., and Lewis, S.J. (2013b). Differential neural activation patterns in patients with Parkinson’s disease and freezing of gait in response to concurrent cognitive and motor load [research support, non-U.S. Gov’t]. PLoS One 8, e52602, 7.
Shine, J.M., Moustafa, A.A., Matar, E., Frank, M.J., and Lewis, S.J. (2013c). The role of frontostriatal impairment in freezing of gait in Parkinson’s disease. Front. Syst. Neurosci. 7, 61.
Shine, J.M., Naismith, S.L., and Lewis, S.J.G. (2013d). The differential yet concurrent contributions of motor, cognitive and affective disturbance to freezing of gait in Parkinson’s disease. Clin. Neurol. Neurosurg. 115, 542–545.
Shine, J.M., Naismith, S.L., Palavra, N.C., Lewis, S.J., Moore, S.T., Dilda, V., and Morris, T.R. (2013e). Attentional set-shifting deficits correlate with the severity of freezing of gait in Parkinson’s disease. Parkinsonism Relat. Disord. 19, 388–390.
Shiner, T., Seymour, B., Symmonds, M., Dayan, P., Bhatia, K.P., and Dolan, R.J. (2012). The effect of motivation on movement: a study of bradykinesia in Parkinson’s disease. PLoS One 7, e47138, 7.
Singhal, A., Culham, J.C., Chinellato, E., and Goodale, M.A. (2007). Dual-task interference is greater in delayed grasping than in visually guided grasping. J. Vision 7, 5.
Smith, M.C. and Fucetola, R. (1995). Effects of delayed visual feedback on handwriting in Parkinson’s disease. Hum. Mov. Sci. 14, 109–123.
Smulders, K., van Nimwegen, M., Munneke, M., Bloem, B.R., Kessels, R.P., and Esselink, R.A. (2012). Involvement of specific executive functions in mobility in Parkinson’s disease. Parkinsonism Relat. Disord. 114, 331–335
Spiegel, J., Hellwig, D., Samnick, S., Jost, W., Mollers, M.O., Fassbender, K., Kirsch, C.M., and Dillmann, U. (2007). Striatal FP-CIT uptake differs in the subtypes of early Parkinson’s disease. J. Neural. Transm. 114, 331–335.
Susman, R.L. (1998). Hand function and tool behavior in early hominids. J. Hum. Evol. 35, 23–46.
Tessitore, A., Amboni, M., Cirillo, G., Corbo, D., Picillo, M., Russo, A., Vitale, C., Santangelo, G., Erro, R., Cirillo, M., et al. (2012).
Regional gray matter atrophy in patients with Parkinson disease and freezing of gait. AJNR Am. J. Neuroradiol. 33, 1804–1809.
Teulings, H.L. and Stelmach, G.E. (1991). Control of stroke size, peak acceleration, and stroke duration in Parkinsonian handwriting. Hum. Mov. Sci. 10, 315–334.
Teulings, H.L., Contreras-Vidal, J.L., Stelmach, G.E., and Adler, C.H. (2002). Adaptation of handwriting size under distorted visual feedback in patients with Parkinson’s disease and elderly and young controls. J. Neurol. Neurosurg. Psychiatry 72, 315–324.
Thomassen, A.J.W.M. and Teulings, H.L. (1985). Time, size, and shape in handwriting: exploring spatio-temporal relationships at different levels. Time, Mind, and Behavior. J.B.J.J.A. Michon, ed. (Heidelberg: Springer-Verlag), pp. 253–263.
Troche, M.S. and Lori, J.P.A. (2012). Sentence production in Parkinson disease: effects of conceptual and task complexity. Appl. Psycholinguistics 33, 225–251.
Tucha, O., Mecklinger, L., Thome, J., Reiter, A., Alders, G.L., Sartor, H., Naumann, M., and Lange, K.W. (2006). Kinematic analysis of dopaminergic effects on skilled handwriting movements in Parkinson’s disease. J. Neural. Transm. 113, 609–623.
Vakil, E. and Herishanu-Naaman, S. (1998). Declarative and procedural learning in Parkinson’s disease patients having tremor or bradykinesia as the predominant symptom. Cortex 34, 611–620.
Van Gemmert, A.W.A., Teulings, H.L., and Stelmach, G.E. (1998). The influence of mental and motor load on handwriting movements in Parkinsonian patients. Acta Psychol. 100, 161–175.
Van Gemmert, A.W., Teulings, H.L., Contreras-Vidal, J.L., and Stelmach, G.E. (1999). Parkinson’s disease and the control of size and speed in handwriting. Neuropsychologia 37, 685–694.
Vandenbossche, J., Deroost, N., Soetens, E., Spildooren, J., Vercruysse, S., Nieuwboer, A., and Kerckhofs, E. (2011). Freezing of gait in Parkinson disease is associated with impaired conflict resolution. Neurorehabil. Neural Rep. 25, 765–773.
Voon, V., Gao, J., Brezing, C., Symmonds, M., Ekanayake, V., Fernandez, H., Dolan, R.J., and Hallett, M. (2011). Dopamine agonists and risk: impulse control disorders in Parkinson’s disease. Brain 134, 1438–1446.
Wagle Shukla, A., Ounpraseuth, S., Okun, M.S., Gray, V., Schwankhaus, J., and Metzer, W.S. (2012). Micrographia and related deficits in Parkinson’s disease: a cross-sectional study. BMJ Open 2, 6.
Walton, C.C., Shine, J.M., Mowszowski, L., Gilat, M., Hall, J.M., O’Callaghan, C., Naismith, S.L., and Lewis, S.J. (2014). Impaired cognitive control in Parkinson’s disease patients with freezing of gait in response to cognitive load. J. Neural Transm. 122, 653–660.
Weinberger, M., Hutchison, W.D., Lozano, A.M., Hodaie, M., and Dostrovsky, J.O. (2009). Increased gamma oscillatory activity in the subthalamic nucleus during tremor in Parkinson’s disease patients. J. Neurophysiol. 101, 789–802.
Weintraub, D. and Burn, D.J. (2011). Parkinson’s disease: the quintessential neuropsychiatric disorder. Mov. Disord. 26, 1022–1031.
Wiecki, T.V. and Frank, M.J. (2010). Neurocomputational models of motor and cognitive deficits in Parkinson’s disease. Prog. Brain Res. 183, 275–297.
Wiecki, T.V., Riedinger, K., von Ameln-Mayerhofer, A., Schmidt, W.J., and Frank, M.J. (2009). A neurocomputational account of
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM

548 A.A. Moustafa et al.: Cognitive and motor symptoms in Parkinsonism
catalepsy sensitization induced by D2 receptor blockade in rats: context dependency, extinction, and renewal. Psychopharmacology (Berl) 204, 265–277.
Williams-Gray, C.H., Foltynie, T., Brayne, C.E., Robbins, T.W., and Barker, R.A. (2007a). Evolution of cognitive dysfunction in an incident Parkinson’s disease cohort. Brain 130, 1787–1798.
Williams-Gray, C.H., Hampshire, A., Robbins, T.W., Owen, A.M., and Barker, R.A. (2007b). Catechol O-methyltransferase Val158Met genotype influences frontoparietal activity during planning in patients with Parkinson’s disease. J. Neurosci. 27, 4832–4838.
Wiseheart, R., Altmann, L.J., Park, H., and Lombardino, L.J. (2009). Sentence comprehension in young adults with developmental dyslexia. Ann. Dyslexia 59, 151–167.
Woollacott, M. and Shumway-Cook, A. (2002). Attention and the control of posture and gait: a review of an emerging area of research. Gait Posture 16, 1–14.
Wu, T. and Hallett, M. (2013). The cerebellum in Parkinson’s disease. Brain 136, 696–709.
Wylie, S.A., van den Wildenberg, W., Ridderinkhof, K.R., Claassen, D.O., Wooten, G.F., and Manning, C.A. (2012). Differential susceptibility to motor impulsivity among
functional subtypes of Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 83, 1149–1154.
Yogev, G., Giladi, N., Peretz, C., Springer, S., Simon, E.S., and Hausdorff, J.M. (2005). Dual tasking, gait rhythmicity, and Parkinso’s disease: which aspects of gait are attention demanding? Eur. J. Neurosci. 22, 1248–1256.
Young, R.W. (2003). Evolution of the human hand: the role of throwing and clubbing. J. Anat. 202, 165–174.
Yu, H., Sternad, D., Corcos, D.M., and Vaillancourt, D.E. (2007). Role of hyperactive cerebellum and motor cortex in Parkinson’s disease. Neuroimage 35, 222–233.
Zaidel, A., Arkadir, D., Israel, Z., and Bergman, H. (2009). Akineto-rigid vs. tremor syndromes in Parkinsonism. Curr. Opin. Neurol. 22, 387–393.
Zetusky, W.J. and Jankovic, J. (1985). Laterality and symptom association in Parkinson’s disease. Arch. Neurol. 42, 1132–1133.
Zhao-Shea, R., Cohen, B.N., Just, H., McClure-Begley, T., Whiteaker, P., Grady, S.R., Salminen, O., Gardner, P.D., Lester, H.A., and Tapper, A.R. (2010). Dopamine D2-receptor activation elicits akinesia, rigidity, catalepsy, and tremor in mice expressing hypersensitive {alpha}4 nicotinic receptors via a cholinergic-dependent mechanism. FASEB J. 24, 49–57.
Brought to you by | University of SydneyAuthenticated
Download Date | 1/10/18 4:36 AM
Copyright © 2022 FDOKUMEN