
Exploitation du monde animal sur le versant nord des Pyrénées durant le Tardiglaciaire. In :...
27
INSTITUT D’ESTUDIS CERETANS ELS PIRINEUS I LES ÀREES CIRCUMDANTS DURANT EL TARDIGLACIAL. MUTACIONS I FILIACIONS TECNOCULTURALS, EVOLUCIÓ PALEOAMBIENTAL (16000-10000 BP) XIV COL·LOQUI INTERNACIONAL D’ARQUEOLOGIA DE PUIGCERDÀ HOMENATGE AL PROFESSOR GEORGES LAPLACE Puigcerdà - 10-12 de novembre de 2006 I N S T I T U T d ’ E S T U D I S C E R E T A N S
Transcript of Exploitation du monde animal sur le versant nord des Pyrénées durant le Tardiglaciaire. In :...

INSTITUT D’ESTUDIS CERETANS
ELS PIRINEUS I LES ÀREES CIRCUMDANTS DURANT
EL TARDIGLACIAL.MUTACIONS I FILIACIONS
TECNOCULTURALS, EVOLUCIÓ PALEOAMBIENTAL
(16000-10000 BP)
XIV COL·LOQUI I N T E R N AC I O N A LD’ARQUEOLOGIA DE PUIGCERDÀ
HOMENATGE AL PROFESSOR GEORGES LAPLACE
Puigcerdà - 10-12 de novembre de 2006
INST
ITU
Td’
ESTUDIS CERETA
NS
Nicolas
Zone de texte
Fullola, J.-M., Valdeyron, N., Langlais, M. (dir.). Les Pyrénées et leurs marges durant le Tardiglaciaire. Mutations et filiations techno-culturelles, évolutions paléo-environnementales, actes du XIVème colloque international d'archéologie de Puigcerda, novembre 2006, Hommages à Georges Laplace, Institut d'Estudis Ceretans

President del CIAP: Josep Padró
Comitè científic del CIAP: Sara AliagaPere Campmajó Denis CrabolJosep M. FullolaOriol Mercadal Nicolas Valdeyron
Coordinadors científics del CIAPi de les actes: Josep Maria Fullola Nicolas ValdeyronMathieu Langlais
Coordinador de la publicació: Oriol Mercadal i Fernàndez
Comitè organitzador: Joan Mateu i Lladó (President de l’IEC)Josep Brunet i ComasPere Font i AlartJoaquim Carbonell i Cornejo
Col·laboradors/es personals:Joan AntonNeus BallbéÀngels CasadesúsOlga GarcíaEsperanza Sánchez
INST
ITU
Td’
ESTUDIS CERETA
NS
Organitza:
Institut d’Estudis Ceretans
Patrocina:
Amb el suport de:
Amb la col·laboració de:
Ajuntament de Puigcerdà
Foto de la portada: jaciment magdalenià de Montlleó (Prats i Sansor). Autor: O. MercadalDibuix de la portada: a partir d’elements de l’art parietal paleolític. Autora: S. Aliaga
Edita:INSTITUT D’ESTUDIS CERETANS
Passeig 10 d’Abril, 2n, 2a - 17520 Puigcerdà
Primera edició: novembre de 2009Reservats tots els drets. No és permesa la reproducció total o parcial d’aquest llibre amb qualsevol tècnica o mitjà,inclosa la fotocòpia, sense permís conjunt, previ i per escrit, dels respectius titulars dels drets d’autor.© Pels textos i imatges: autors corresponents© Per l’edició: Institut d’Estudis Ceretans
Imprès a MAIDEU, SL - Ripoll 2009ISBN: 978-84-933111-8-9-7Dipòsit Legal GI 1321-2009
UNIVERSITAT DE BARCELONA
U
B
S E R PS E R P

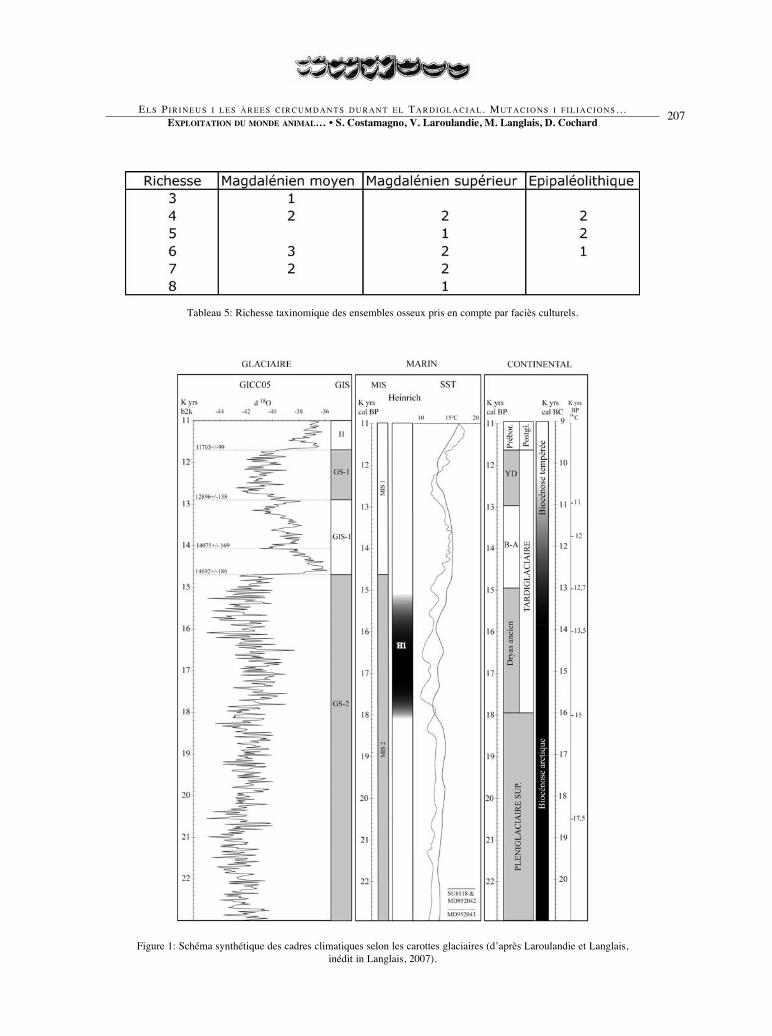
Le Tardiglaciaire est marqué pard’importants changements climatiquescomme en témoignent les archives gla-ciaires, continentales et marines (Cachoet al., 2001; Elliot et al., 2002; Pailler etBard, 2002; Andersen et al., 2006; Ras-mussen et al., 2006; Svensson et al.,2006) (fig. 1). Dans sa version longue(18000 – 11700 calBP), il débute par leDryas ancien qui s’étend de 15000 à12700 BP, soit 18000 – 15000 calBP(Magny, 1995). Cette chronozone coïnci-de avec l’événement de Heinrich 1(He1), daté entre 15000 et 13400 BP, soit18000 - 15200 calBP (Elliot et al., 2002).L’interstade Bølling/Allerød, qui luisuccède, correspond peu ou prou à l’in-terstade glaciaire GIS-11 (14700 – 12900b2k) enregistré dans les carottes glaciai-res (Rasmussen et al., 2006). Il existeainsi un décalage de 300 à 500 ans entre
la fin du Dryas ancien / He1 et les débutsdu GIS-1, décalage qui coïncide d’ai-lleurs avec un plateau du 14C (Reimer etal., 2004). Le Dryas récent, dont la limi-te supérieure est datée d’environ 11700calBP, représente la dernière phase duTardiglaciaire.
Les changements climatiques interve-nus durant cette période ont eu des réper-cussions importantes sur les biocénosesvégétales (e.g. Beaulieu et al., 1988;Jalut et al., 1992; Reille et Andrieu,1995; Jalut et al., 1998; Richard et al.,2000; Naughton et al., 2007; Jalut etTuru i Michels, ce volume). Ainsi, l’évè-nement de He1 se marque par une aug-mentation des Armoises et Poacées,suivies par les Genévriers (colonisationfin He1) et les Bouleaux dont le dévelop-pement coïncide avec le réchauffementdu GIS-1 (Jalut et Turu i Michels, cevolume). La répartition des populationsanimales s’en trouve affectée en particu-lier celle des Ongulés qui, se situant à labase de la pyramide écologique, dépen-dent étroitement de la végétation (Del-pech et al., 1983; Delpech, 1989, 1999;Bridault et Fontana, 2003; Costamagnoet al., 2008). Sur le versant nord des
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…185
EXPLOITATION DU MONDE ANIMAL SUR LE VERSANT
NORD DES PYRÉNÉES AU TARDIGLACIAIRE
Sandrine COSTAMAGNO*, Véronique LAROULANDIE**, Mathieu LANGLAIS***, David COCHARD**
* TRACES – UMR 5608 du CNRS – Université ToulouseLe Mirail, Maison de la Recherche, 5 allées AntonioMachado, 31058 Toulouse cedex 9, France.
** PACEA – UMR 5199 du CNRS - Université Bordeaux I,Avenue des Facultés, 33405 Talence Cedex, France.
***TRACES – UMR 5608 du CNRS – Université ToulouseLe Mirail, Maison de la Recherche, 5 allées AntonioMachado, 31058 Toulouse cedex 9, France & SERP-Uni-versité de Barcelone.
Nicolas
Zone de texte
Fullola, J.-M., Valdeyron, N., Langlais, M. (dir.). Les Pyrénées et leurs marges durant le Tardiglaciaire. Mutations et filiations techno-culturelles, évolutions paléo-environnementales, actes du XIVème colloque international d'archéologie de Puigcerda, novembre 2006, Hommages à Georges Laplace, Institut d'Estudis Ceretans

Pyrénées, comment les communautésanimales ont-elles réagi à cette forte ins-tabilité climatique et environnementale?Quel est l’impact de ces changements surles stratégies de subsistance développéespar les chasseurs-cueilleurs du Tardigla-ciaire? Par une revue synthétique desdonnées disponibles, nous essayonsd’apporter des éléments de réponse à cesquestions.
Choix du corpus
Ces dernières années, les différentessynthèses réalisées sur la faune du ver-sant nord pyrénéen au cours du Paléoli-thique ont pris en compte l’ensemble desunités stratigraphiques dont les assem-blages osseux avaient fait l’objet d’étudeplus ou moins détaillée (Costamagno etLaroulandie, 2004; Costamagno et Mate-os Cachorro, 2007; Costamagno et al.,2008). Dans le cadre de cette contribution,nous n’avons retenu que les ensemblesfauniques datés radiochronologiquement(dates AMS préférentiellement aux datesconventionnelles et sigma inférieur à300) afin de caler le plus précisémentpossible nos données aux courbes paléo-climatiques et reconstitutions paléoenvi-ronnementales. De ce fait, notre corpusest beaucoup plus limité que celui géné-ralement utilisé2. Seuls quinze sites, quicorrespondent à 24 unités stratigraphi-ques, répartis sur l’ensemble de la chaîneont ainsi pu être mobilisés dans le cadrede cette analyse (tabl. 1 et fig. 2). Ponc-tuellement, des ensembles osseux nondatés ont pu être utilisés dans les discus-sions.
Cortèges fauniques documentés
Le Dryas ancien / He1 voit l’avène-ment du Magdalénien moyen puis satransformation en Magdalénien supérieurautour de 13500 BP, soit 16000 calBP.Cette évolution se marque, en particulier,par des changements dans la gestion deséquipements et une diversification desarmatures (Lacombe, 1998; Dachary,2002; Langlais, 2007). Parmi les ensem-bles osseux de cette période, celui d'Au-rensan (Hautes-Pyrénées) (Clot, 1983)présente un spectre faunique particulier,dominé par les carnivores (en particulierle Renard) qui représentent plus de 44%des restes osseux (tabl. 2). La couche 7de Gazel (Aude) se singularise aussi desautres gisements de cette période en rai-son de l’abondance des restes de Lièvrevariable (Fontana, 2003). Dans les autressites, les Ongulés sont dominants (tabl.2): Renne, Cheval et grands Bovidés3
sont signalés dans tous les ensemblesosseux. En terme d’occurrences, vien-nent ensuite le Cerf et les Ongulés demontagne, respectivement onze et neufoccurrences. Les Ongulés inféodés auxmilieux boisés (Sanglier et Chevreuil)sont, en revanche, rarement signalés dansles assemblages du Dryas ancien. L’Anti-lope saïga, qui évite les zones accidenté-es (Costamagno, 2001), n’a été identifiéeque dans la couche 3 des Conques(Moigne, 2003) et le niveau 5/6 de l’abriDufaure (Altuna et Mariezkurrena,1995).
Parmi les Ongulés, le Renne dominedans huit ensembles osseux (Dufaurec.6/5, Dufaure c.4, Duruthy c.3, Espèche(Clot et Omnès, 1979), Enlène c.3 (Del-
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard186

pech, 1981), Gazel c.7, Canecaude c.II(Fontana, 1998a), Les Conques c.3). Àl’Ouest de la chaîne pyrénéenne maiségalement à la grotte de Labastide, lesOngulés de grande taille (Cheval, Bovi-nés) occupent une place prépondérante(Clot et Omnès, 1979). Dans la couche 5de Duruthy, le Cheval représente plus de60% des restes déterminés suivi par lesgrands Bovidés (30,7%); dans la couche4, ce sont les Bovinés qui dominent(44,3%), le Cheval et le Renne présen-tant des fréquences comparables (envi-ron 25%) (Delpech, 1978). Certainsgisements se caractérisent par une prédo-minance des Ongulés de montagne. ÀAurensan, si l’on exclut les restes de Vul-pinés, ils représentent plus de 40% desrestes. À la Vache (Pailhaugue, 1995,1998), aux Eglises (Delpech et Le Gall,1983) et dans une moindre mesure dansla couche 3 de Belvis (Fontana, 1998b),le spectre des Mammifères est largementdominé par le Bouquetin. En revanche,l’Isard n’est présent que dans de faiblesproportions sur les sites du Dryas ancien.Il en est de même du Cerf dont la fré-quence n’excède que rarement 15% àl’exception de la couche 4 de Dufaure,des couches 3 de Duruthy et des Conques(Moigne et al., 1998; Moigne, 2003) etde la couche B3 d’Arancou (Fosse, 2000)dans laquelle c’est l’espèce dominante.La présence du Chevreuil et du Sanglierest anecdotique puisqu’elle n’est jamaissupérieure à 2%.
Certains sites présentent des dates éta-lées à la fois sur le Dryas ancien et leBølling/Allerød sans montrer de vérita-ble changement dans la diète. C’est le casde Belvis c.3 (Magdalénien supérieur à
triangles) (Sacchi, 1986; Langlais, 2007)dominé par le Bouquetin et le Renne(Sacchi, 1994; Fontana, 1998a) ou LaVache. Ce site livre un abondant matérielattribuable au Magdalénien supérieurclassique (Clottes et Delporte, 2003).Toutefois, des pointes à dos et certainsharpons plats suggèrent l’existence d’oc-cupations aziliennes non distinguées à lafouille (Schmider, 2003). De plus, la pré-sence de pointes à base fourchues pose laquestion d’un phasage interne de l’en-semble, ces pièces témoignant plutôtd’une phase ancienne du Magdaléniensupérieur relativement aux harpons(Pétillon, 2006). Cette dilation chrono-culturelle se marque d’ailleurs à traversl’ensemble des dates 14C s’étalant de11650 à 13770 BP et recouvrant ainsi unegrande partie du Tardiglaciaire.
Durant l’interstade Bølling/Allerød,on assiste à la fin du Magdalénien quiévolue progressivement vers un Epipa-léolithique ancien polymorphe (Langlais,2007; Martzluff, ce volume). Au cours dece réchauffement climatique, les restesde Léporidés sont, comme au Dryasancien, peu nombreux (tabl. 3). Au con-traire, le Cerf, présent sur tous les gise-ments, prend de l’importance dans lescortèges de faune dès le début de cetinterstade. C’est le premier gibier chassédans l’ensemble osseux B2 d’Arancou(Fosse, 2000), la couche 7a de Troubat(Costamagno, 2005) et le foyer 6 de Rho-des II (Delpech, 1983). Dans les foyers 5et 7 de ce gisement, il arrive en deuxièmeposition après le Sanglier dans F7 et leBouquetin dans F5. Parallèlement, lesespèces inféodées aux milieux boisésprennent leur essor aux dépens du Renne
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
187

dont la présence devient sporadique:Sanglier et Chevreuil représentent entre23 et 62% des restes fauniques. Dans lacouche B2 d’Arancou, les Ongulés step-piques (Cheval, Bovinés) sont relative-ment bien représentés ce qui contrasteavec les autres gisements de cette pério-de.
Pour le Dryas récent, au cours duquelévolue l’Azilien classique (Martzluff, cevolume; Valdeyron et Detrain, ce volu-me), seuls deux assemblages osseuxayant fait l’objet d’une étude quantifiéebénéficient de datations radiocarbones:la couche 3 de Dufaure et le niveau 6 deTroubat. À l’abri Dufaure, le Cerf estultra-majoritaire (83,7%), suivi par leChevreuil (8,9%) (tabl. 3) (Altuna etMariezkurrena, 1995). La présence dequelques restes de rennes est à noter. Desrestes de rennes sont également signalésdans la couche 2 de la grotte de Duruthytoute proche (Delpech, 1978). À Troubatc.6, le Cerf et les Ongulés de montagnesont les gibiers dominants et le Renne estabsent (Martin, 1994).
Recomposition végétale et animale
sur le versant nord des Pyrénées
La distribution géographique desMammifères, en particulier celle desHerbivores, est largement dépendantedes conditions environnementales: topo-graphie, ressources alimentaires, climatsont autant de facteurs qui entrent enligne de compte. Dans ce chapitre, nousessayons de documenter dans quellemesure les cortèges fauniques reflètentles changements climatiques et environ-nementaux enregistrés durant le Tardi-
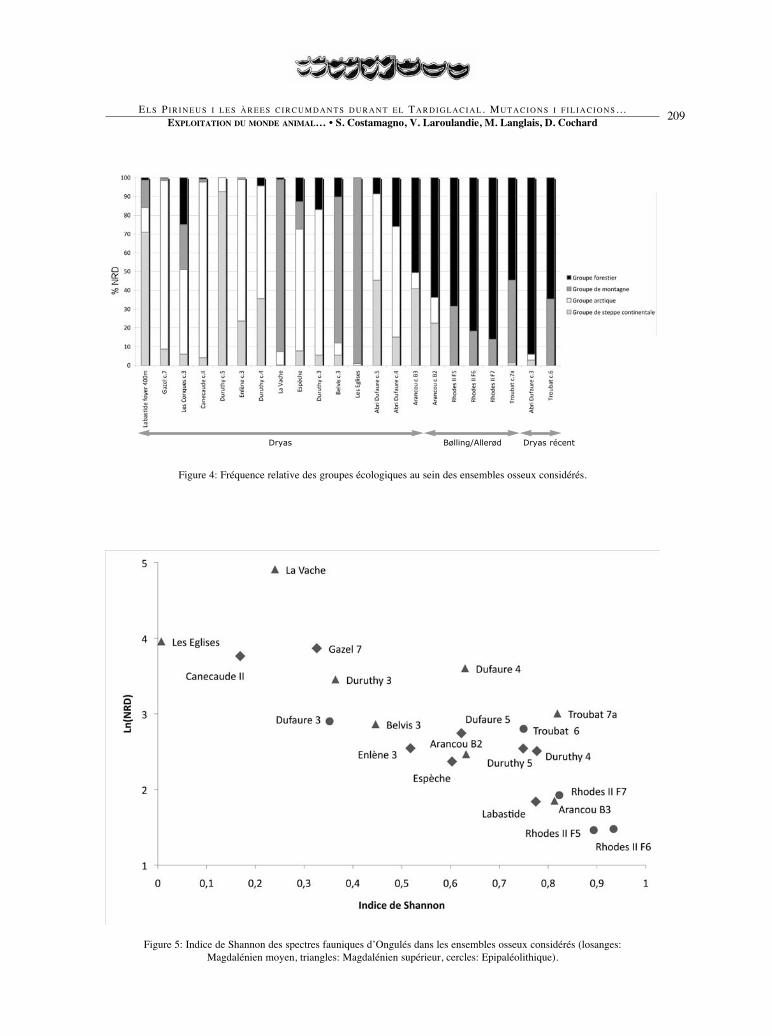
glaciaire. Dans cet objectif, les différentsensembles osseux ont été classés parordre chronologique croissant, sur labase de la datation calBP plancher la plusancienne (fig. 3). Les quatre groupesécologiques définis par Ch. Griggo(1995) ont ensuite été utilisés pour cha-que ensemble osseux (fig. 4). Dans lescortèges fauniques documentés à cettepériode sur le versant nord des Pyrénées,le groupe arctique se compose du Lièvrevariable, du Renard polaire, du Gloutonet du Renne4; le groupe de montagne duBouquetin et de l’Isard, le groupe fores-tier du Lièvre d’Europe, du Lapin, duChat sauvage, du Lynx, du Blaireau, dela Martre, de la Fouine, de l’Hermine, duSanglier, du Cerf et du Chevreuil et legroupe de steppe continentale du Cheval,du Bison et de l’Antilope saïga.
D’après l’analyse pollinique desdépôts lacustres pyrénéens, le couvertforestier au Dryas ancien reste très limitéjusque vers 15000 - 14700 calBP (Jalut etTuru i Michels, ce volume). Si l’onexclut les assemblages osseux dans les-quels les Ongulés de montagne tiennentune place prépondérante, les cortègesfauniques de cette période se caractéri-sent par une large prédominance, soit dugroupe arctique (Renne essentiellement,parfois Lièvre variable), soit du groupede steppe continentale (Cheval et/ouBison). L’abondance des Ongulés steppi-ques est surtout perceptible dans la partieoccidentale de la chaîne, ce qui pourraittémoigner de conditions climatiques plussèches dans cette zone relativement à lapartie orientale. Durant ce stade glaciai-re, le groupe forestier est représentéquasi-exclusivement par le Cerf, qui,
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard188

d’après les études menées sur les isoto-pes du Carbone et du Nitrogène, peut s’a-dapter à des milieux très ouverts(Drucker et Célérier, 2001; Drucker etal., 2003; Drucker et Bridault, 2004). Endehors de la couche 4 de Dufaure et de lacouche B3 d’Arancou datées de l’extrê-me fin du Dryas ancien, la fréquence desespèces dites boisées ne dépasse jamaisles 15%: seule exception, la couche 3 dugisement méditerranéen des Conques(17410 – 16910 calBP), dans laquelle unquart des restes identifiés correspond àdu Cerf. Sans d’autres gisements datés decette période à proximité5, il est difficilede savoir si cette abondance est liée à unenvironnement local plus favorable audéveloppement de cette espèce. La pré-sence de lapins pourrait cependant indi-quer de possibles contaminations avecdes niveaux plus récents. Par ailleurs, unreste de Renne issu de la couche 3 deDuruthy a récemment été daté par leLaboratoire de Poznan. La nouvelle dateobtenue (Poz 15983 12 930 ± 60 BP, tabl.1) remet en cause l’attribution de ceniveau à la fin de l’Allerød. L’outillagelithique de Duruthy c.3, parfaitementcohérent avec du Magdalénien supérieurclassique (Arambourou, 1978; Dachary,2002), va d’ailleurs plutôt dans le sens dela nouvelle date AMS, au détriment del’ancienne (11150 BP). La couche 3 deDuruthy, qui se serait déposée durant leDryas ancien, se caractérise donc par desassociations fauniques tout à fait compa-rable à celles retrouvées dans des ensem-bles pénécontemporains (fig. 4).
Durant l’interstade Bølling/Allerød, legroupe forestier prend une place prépon-dérante au sein des cortèges de faune.
Dans la couche B2 d’Arancou, le Cerf estle seul représentant de ce groupe, ce quicorroborerait les données polliniques. Eneffet, au cours du GI-1e (équivalent duBølling) (Rasmussen et al., 2006), bienque la température et les précipitationsaugmentent, le milieu reste encore trèsouvert (Jalut et Turu i Michels, ce volu-me) donc peu propice au développementdu Chevreuil, qui se nourrit de jeunespousses d’arbres et d’arbustes et defruits, et du Sanglier, qui affectionne lesforêts de feuillus. L’abondance de cesdeux espèces dans les foyers 5 et 6 deRhodes II, datés tous deux du Bølling,soulèvent des interrogations quant à lapertinence des dates par rapport aux don-nées environnementales. D’après lestaxons en présence, ces ensembles trou-veraient mieux leur place au sein de l’A-llerød, interstade au cours duquel lecouvert forestier devient plus dense(Jalut et Turu i Michels, ce volume). Tou-tefois, à partir de certains éléments ensilex, le foyer 5 de Rhodes II est attribuépar certains auteurs à un Epipaléolithique/ Azilien ancien pour lequel l’apparte-nance à la seconde moitié du Bølling esttout à fait envisageable (Barbaza, 1996;Langlais, 2007; Martzluff, ce volume).Les deux ensembles documentant l’Al-lerød sont largement dominés par les ani-maux de milieu boisé; le Chevreuil et leSanglier représentent 62 % des restesdans le foyer 7 de Rhodes II et 25% dansla couche 7a de Troubat. Dans ces deuxensembles, le Renne est absent, ce quipourrait indiquer sa migration vers deszones septentrionales plus propices à sesexigences écologiques.
Au sein des enregistrements lacustres
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
189

pyrénéens, le refroidissement du Dryasrécent est bien perceptible. Il se traduitpar «une interruption du développementdes arbres et une augmentation généra-lement limitée des herbacées et des cha-méphytes héliophiles de caractèresteppique» (Jalut et Turu i Michels, cevolume). Peu d’ensembles osseux docu-mentent ce stade. Il est donc difficile d’é-valuer l’impact de ce refroidissement surles biocénoses animales. Dans la couche6 de Troubat, le cortège faunique est trèssimilaire à celui de la couche 7a sousjacente. On observe cependant une nettediminution du Chevreuil (conséquencedu refroidissement ou choix des chas-seurs?). Dans la couche 3 de Dufaure, leRenne fait sa réapparition. Sa présenceest documentée dans la couche 2 deDuruthy, également azilienne. Pourexpliquer la persistance du Renne dansdes couches aussi récentes, F. Delpech(1989, 2003) a émis l’hypothèse de laprésence de zones refuges dans desrégions montagneuses au cours de cettepériode. À la faveur du refroidissementdu Dryas récent, le Renne pourrait égale-ment avoir recolonisé le Sud-Ouest de laFrance, d’autres ensembles contempo-rains de ce refroidissement livrent eneffet quelques restes de rennes: Sainte-Eulalie c.1 (Gif 2193 10 400 ± 300 BP(12 746 – 11 823 calBP); Gif 1697 10830± 200 BP (13051 – 12 644 calBP)) (Del-pech, 1989), Gazel c.6 (Fontana, 1998a).Cependant, comme nous l’avons vu, lesrestes de rennes de la couche 3 deDuruthy, jusqu’ici attribuée à l’Allerød,semblent contemporains du Dryasancien. De la même façon, les ossementsde Renne récoltés dans la couche 3 de
Dufaure (Azilien) pourraient provenir decontaminations avec des couches sous-jacentes. En effet, un os de Renne envo-yé au laboratoire de Poznan a fourni unedate de 12 260 ± 60 BP Poz 15984, soit14 200 - 14 030 calBP, donc contempo-raine du Bølling. Ces premières datesobtenues sur des restes de rennes identi-fiés dans des niveaux de la fin de l’in-terstade Bølling/Allerød iraient dans lesens d’une disparition du Renne dans lesPyrénées au début de l’Allerød commecela est documenté dans d’autres régionsfrançaises (Bridault et al., 2000; Bridaultet Fontana, 2003). La forte taille duRenne observée à Duruthy (Delpech,1983) et à Dufaure (Weinstock, 2000)pour cette période constituerait un argu-ment supplémentaire. Un datage systé-matique des restes de rennes retrouvés encontexte tardif dans le Sud-Ouest de laFrance est en cours de réalisation afin dedocumenter le rythme de sa disparition.
En résumé, du point de vue des cortè-ges fauniques, des changements sem-blent s’amorcer dès le début del’interstade Bølling/Allerød avec, notam-ment, une augmentation des effectifs despopulations de Cerf. Ces dernièresvenant probablement de la Péninsule ibé-rique, qui a servi de zone refuge durantles périodes glaciaires (Delpech, 1989;Costamagno et Mateos Cachorro, 2007),recolonisent rapidement le versant norddes Pyrénées. Les Ongulés communs auxbiocénoses glaciaires sont toujours pré-sents mais leurs effectifs semblent dimi-nuer (Delpech, 1988). D’après F.Delpech (1999), à partir de 12500 BP(15000 – 14500 calBP), l’aire de réparti-tion du Renne se morcelle, annonçant le
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard190

début d’une période contraignante sousnos latitudes pour cette espèce. Dansd’autres régions et notamment en Aqui-taine, l’Antilope saïga se raréfie (Del-pech, 1999). Mais la véritable ruptured’équilibre se situe au début de l’Allerød(12000 BP, soit 13800 calBP): les popu-lations de rennes, de bisons puis de che-vaux se raréfient puis disparaissent duversant nord pyrénéen (Costamagno etal., 2008) et sont remplacées par desespèces forestières (Chevreuil, Sanglier).Ce changement s’inscrit à l’échelle plusvaste du Sud de l’Europe comme entémoignent les archéofaunes d’autresrégions (Delpech, 1989, 1999; Bridaultet Fontana, 2003).
Changements environnementaux et
subsistance
Du point de vue des ressources ali-mentaires et, selon l’approche «naturalis-te», les bouleversements climatiquesintervenus à la fin du Tardiglaciaireseraient responsables de l’intégration denouvelles ressources alimentaires, abou-tissant à un élargissement de la diète deschasseurs-cueilleurs (Rowley-Conwy,1986). Ce postulat repose sur le modèlede l’amplitude de la diète (diet breadthmodel) qui est fondé sur le principe d’op-timisation (Winterhalder, 1981). Selon cemodèle, la sélection des ressources ali-mentaires est guidée par la contraintecoût/bénéfice et ce, afin d’obtenir lemeilleur rendement énergétique possible.Observe-t-on au cours du Tardiglaciaireune diversification des spectres de chas-se dans le massif pyrénéen? L’intégrationde la petite faune au régime alimentaire
de ces derniers chasseurs-cueilleurs est-elle attestée? Dans ce chapitre, outre lesgrandes subdivisions paléoclimatiques,sont également prises en compte les attri-butions culturelles (Magdalénien moyen,Magdalénien supérieur, Epipa-léolithique) afin de mettre en évidenced’éventuelles spécificités tant du point devue des stratégies de chasse mises enœuvre que des gibiers recherchés ou del’exploitation des ressources alimentai-res.
Degré de spécialisation des spectresfauniques
La recomposition des cortèges fauni-ques observée à la fin du Tardiglaciaireest marquée par une diminution puis unedisparition des grands troupeaux d’ongu-lés (chevaux, rennes) au profit d’espècesplus solitaires (sangliers, chevreuils). Lespremiers, pouvant être abattus en masselors de chasses collectives, offrent par-fois un rang tellement élevé par rapportaux autres ongulés disponibles dans l’en-vironnement que, dans certaines régions,ils ont été les seuls gibiers exploités, aumoins à certaines saisons de l’année, parles groupes paléolithiques (Enloe, 1999;Costamagno, 2003). Au contraire, lesOngulés de milieu fermé en raison deleurs mœurs solitaires possèdent desrangs équivalents ce qui doit aboutir,selon le principe d’optimisation, à desspectres de chasse plus diversifiés. Unchangement dans les tactiques de chasseest d’ailleurs évoqué comme un des fac-teurs de l’évolution de l’armement deschasseurs magdaléniens et aziliens,notamment du Bassin parisien (Pelegrin,
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
191

2000; Valentin, 2005), passant ainsi pro-gressivement d’une activité collective àdes actions individuelles.
La richesse taxinomique c’est-à-direle nombre de taxons présents au sein duspectre faunique identifie le nombred’espèces6 introduites dans l’alimenta-tion. Sur le versant nord des Pyrénées, ledegré de richesse des spectres fauniquessemble relativement comparable duranttout le Tardiglaciaire (tabl. 5) Au coursde l’Epipaléolithique, le nombre detaxons chassés semble cependant un peuplus restreint, en raison probablement dela disparition progressive des Ongulésglaciaires.
La diversité des spectres des onguléschassés a été documentée grâce à l’indi-ce de Shannon7 (Grayson, 1984): plusl’indice est fort, plus le spectre est équi-libré; des valeurs proches de zéro indi-quent des ensembles osseuxmonospécifiques, c’est-à-dire spéciali-sés. La taille des échantillons ne joue pasde rôle dans le degré de diversité desensembles osseux considérés puisque lesensembles osseux les plus importantsprésentent les indices de diversité lesplus faibles (fig. 5). La diversité descortèges apparaît comme une constantetout au long de la période considérée. Demanière générale, les spectres fauniquespyrénéens sont diversifiés à très diversi-fiés, les indices de Shannon étant majori-tairement compris entre 0,4 et 0,9. Seulsquelques ensembles contemporains duDryas ancien mais aussi du début de l’in-terstade glaciaire se caractérisent par desspectres fauniques spécialisés sur unongulé particulier. Ces ensembles repré-sentent environ 20% des accumulations
considérées. Ils sont présents dans tousles faciès du Magdalénien: pour le Mag-dalénien moyen, la couche II de Cane-caude et la couche 7 de Gazel8 secaractérisent par des spectres fauniquesspécialisés sur le Renne tandis qu’auMagdalénien supérieur, les sites desEglises et de la Vache montrent des spec-tres spécialisés sur le Bouquetin. Demanière générale, les niveaux épipaléo-lithiques présentent les spectres les plusdiversifiés, à l’exception de la couche 3de Dufaure largement dominée par leCerf. Sur la bordure nord du massif pyré-néen, la diversité des économies de sub-sistance mise en évidence contraste aveccelle des chasseurs-cueilleurs du versantsud et des Cantabres, basée principale-ment sur le Cerf, et dans une moindremesure le Bouquetin (Costamagno etMateos Cachorro, 2007).
Stratégies d’acquisition et d’exploita-tion des Ongulés
Les données relatives aux stratégiesd’acquisition et d’exploitation des res-sources carnées sont très limitées. En cequi concerne les saisons d’abattage, lesite des Eglises se caractérise par unechasse très saisonnière du Bouquetin, dela fin de l’automne au début de l’hiver(Delpech et Le Gall, 1983). Dans la cou-che 4 de Dufaure, le Renne comme lesautres Ongulés semblent avoir été abat-tus de la fin de l’automne à la fin de l’hi-ver (Straus et Spiess, 1985; Spiess,1995). À Duruthy, les chasseurs étaientprésents de septembre à mars (Delpech,1978). À Canecaude c.II et Gazel c.7, lesrennes ont été abattus tout au long de
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard192

l’hiver et du printemps (Fontana, 1998a).Les bouquetins ont été chassés de l’au-tomne au printemps à la Vache (Pailhau-gue, 1995), du début de printemps audébut de l’automne à Belvis (Fontana,1998b). Dans la couche 6 de Troubat, leschasses aux cerfs et bouquetins se sontdéroulées durant l’été (Martin, 1994). AuMagdalénien, cette grotte pourrait avoirété occupée à la même saison commesemble l’indiquer la saison de capturedes poissons (Le Gall, inédit). À Gazel etCanecaude, l’étude de l’âge et du sexemontre, par ailleurs, une chasse nonsélective des rennes. La chasse au Bou-quetin s’inscrit dans le même registrecomme en témoigne la présence de jeu-nes et d’adultes mais également de mâleset de femelles, à la Vache, aux Eglises età Belvis. Dans la couche 6 de Troubat, cesont, en revanche, essentiellement descerfs et des bouquetins adultes qui ontété abattus. Enfin sur la plupart des gise-ments documentés, les animaux sem-blent avoir été introduits entiers.
Les quelques données disponibles surle traitement des carcasses en relationavec leur exploitation alimentaire mon-trent de nombreuses similitudes d’un siteà l’autre quelle que soit la période consi-dérée. Les carcasses transportées au cam-pement font l’objet d’une exploitationgénéralement intensive. Dans la plupartdes gisements documentés, de nombreu-ses traces de décarnisation sont présentessur les ossements. Elles peuvent êtreliées soit au décharnement des ossementspour leur fracturation, soit à la confectionde filets de viande en vue d’un séchagepour une consommation différée. Cettepratique n’a été mise en évidence que sur
le site des Eglises (Delpech et Villa,1993). Sur les autres gisements, l’abon-dance des stries de décharnement pou-rrait être liée à la fracturation desossements en vue d’une exploitation dela moelle. Les os longs complets sontextrêmement rares sur la plupart desgisements documentés. La présence detraces de percussion ainsi que la formedes bords de fracture qui est majoritaire-ment caractéristique d’une fracturationsur os frais indique que cette fragmenta-tion est d’origine anthropique. Cetteexploitation de la moelle est intensivepuisque les mandibules tout comme lespremières phalanges font l’objet d’unefracturation quasi-systématique de la partdes Magdaléniens mais également desAziliens (Costamagno, 1999, 2003;Ferrié, inédit). De telles pratiques ontégalement été mises en évidence sur leversant sud des Pyrénées (MateosCachorro, 2000/2001, 2003; Costamagnoet Mateos Cachorro, 2007). La consom-mation de la graisse contenue dans lesportions spongieuses est particulièrementdifficile à mettre en évidence. Cette pra-tique n'est signalée que pour la couche 7de Troubat (Costamagno, 2005). Enfin,l’emploi de l’os comme combustible estattesté dans les niveaux inférieurs deTroubat attribués au Magdalénien moyen(Costamagno, inédit) ainsi qu’à Enlène(Fosse, com. orale).
De ces données éparses, il est trèsdélicat de tirer des généralités. Quoiqu’ilen soit, il n’apparaît pas au cours du Tar-diglaciaire de véritables ruptures tant dupoint de vue des stratégies d’acquisitiondes proies que de l’exploitation des res-sources alimentaires. Malgré une aug-
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
193

mentation de la saison végétative desplantes durant l’interstade Bølling /Allerød et donc potentiellement d’uneconsommation plus importante de végé-taux, la graisse semble avoir été unesubstance systématiquement recherchéequelles que soient les saisons et forte-ment prisée par tous les groupes de chas-seurs-cueilleurs.
Exploitation des petits gibiers dansles Pyrénées
Bien que présents dans la plupart desgisements répertoriés, la fréquence del’ensemble de ces petits gibiers excèderarement 10% des restes osseux détermi-nés (Costamagno et Laroulandie, 2004).Sur les gisements ayant fait l’objet d’a-nalyses archéozoologiques, la gammedes petits Mammifères exploités apparaîtvariée tout au long de la période considé-rée (Lièvre, Renard, Blaireau, Chatforestier…). Seuls les niveaux corres-pondant au Dryas ancien de la grotteGazel témoignent d’une exploitationimportante du Lièvre variable9 (Fontana,1998a, 2003). Parmi les Oiseaux, si lesossements de plusieurs espèces de gran-de taille ont été utilisés comme matièrepremière (Laroulandie, sous presse), lamajorité de la documentation concernantla chasse du gibier à plumes durant leTardiglaciaire intéresse trois taxons: leslagopèdes Lagopus lagopus et L. mutus,la Chouette harfang Bubo scandiacus, etle Chocard Pyrrhocorax graculus(Laroulandie, 2003, sous presse). Dans ledétail, les séries du Magdalénien moyende Gazel et du Mas d’Azil montrent uneexploitation des Lagopèdes et, dans une
moindre mesure, du Chocard (Vilette,1983, 1999; Laroulandie et Vilette, sou-mis-a). Dans les séries attribuées auMagdalénien supérieur (post He1 etBølling), les Lagopèdes ont été chassés àLa Vache10 (Laroulandie, 2000, 2005),aux Eglises (Laroulandie, 1998), à Istu-ritz (Pétillon et al., sous presse), à Rho-des II (Bouchard, 2007) ainsi qu’àBerroberria (Diez Fernandez-Lomana etal., 1995). L’exploitation du Chocard estquant à elle documentée à Isturitz (Pétil-lon et al., sous presse) et à La Vache(Laroulandie, 2000). Les sites ayant livrédes preuves de la chasse à la ChouetteHarfang se situent à l’Ouest de la chaînepyrénéenne: il s’agit d’Arancou (East-ham, 1998), Dufaure (Eastham, 1995),Duruthy (Laroulandie, 2006) et Isturitz(Pétillon et al., sous presse). Les donnéesconcernant les séries épipaléolithiquess.l. sont encore peu nombreuses. La cou-che 6 azilienne de Troubat livre une peti-te série aviaire indiquant une utilisationde Lagopède, Anatidés, Perdrix grise,Tétras Lyre et Aigle Royal (Laroulandie,2007). Dans les niveaux épipaléolithi-ques de Gazel, le Pigeon prend peu à peude la place au détriment des Lagopè-des(Laroulandie et Vilette, soumis-b).Ces changements dans le spectre indi-quent vraisemblablement une variationde la biogéographie des espèces arcti-ques exploitées par les chasseurs Magda-léniens.
Les quelques études archéozoologi-ques disponibles sur les petits gibiersindiquent une diversité dans les techni-ques bouchères mises en œuvre et dansles sous-produits recherchés (viande,moelle, peau, plume, ossements…). La
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard194

consommation de la viande est attestéepour l’ensemble des petits animaux chas-sés pour toute la période étudiée. Sur lesite magdalénien de la Vache, l’abondan-ce des stries de boucherie correspond àun prélèvement quasi-systématique de laviande des lagopèdes et des chocards(Laroulandie, 2000, 2005). En revanche,dans le site voisin des Eglises, la faiblefréquence de traces de découpe sur lescarcasses de lagopèdes témoigne de pra-tiques de boucherie distinctes (Laroulan-die, 1998, 2005). Pour les Léporidés,parallèlement à l’exploitation de la vian-de, l’extraction de la moelle apparaîtcomme un phénomène récurrent(Cochard, 2004). Au delà de l’aspect ali-mentaire, les petits gibiers participent àl’enrichissement de la gamme des matiè-res premières exploitables. Ainsi, lesMagdaléniens auraient utilisé les fourru-res de lièvres variables sur le site deGazel c.7 (Fontana, 2003) et les griffesde chouettes harfang sur les gisements deDufaure et d’Arancou (Eastham, 1998).Des baguettes osseuses ont par exempleété extraites d’un tibia de lièvre au Masd’Azil11 et d’os longs d’oiseaux à RhodesII (Bouchard, 2007). Enfin, de très nom-breux exemplaires d’os longs d’oiseauxdécorés sont décrits dans des niveaux duMagdalénien moyen et supérieur. (e.g.bilan dans Laroulandie, 2000, sous pres-se). L’utilisation des plumes pour lesempennages et comme éléments de décorest probable mais reste difficilementdémontrable par le seul registre archéo-zoologique en raison du caractère labilede cette matière. Néanmoins, à RhodesII, l’abondance des traces de découpe surl’ulna pourrait indiquer le prélèvement
des plumes (Bouchard, 2007). L’exploitation des Oiseaux au Magda-
lénien supérieur sur le versant nord desPyrénées s’intègre dans un espace géo-graphique plus large qui couvre le Sud del’Europe. En effet, des témoignages pro-viennent de Suisse (Chaix, 2003), du Sudde la France (Mourer-Chauviré, 1983;Vilette, 1999, 2003; Laroulandie, souspresse) et de l’Espagne (Diez Fernandez-Lomana et al., 1995; Garcia Petit, 1997).Durant la fin du Tardiglaciaire, la chasseaux Oiseaux est également attestée enItalie dans des niveaux de l’Épigravettienfinal (Cassoli et Tagliacozzo, 1997;Tagliacozzo et Gala, 2002). La chasse sepratique essentiellement aux dépens desLagopèdes et du Chocard dans les zonesmontagneuses, de l’Outarde canepetière,de la Grande Outarde et des Oies au Sudde cette aire géographique. La ChouetteHarfang est quant-elle chassée dans leBassin aquitain. Par ailleurs, dans le Sudde la France, la chasse aux Oiseaux s’ac-compagne de la pêche, surtout aux Sal-monidés (Le Gall, 1999, 2003) et del’exploitation du Lièvre variable au coursdu Bølling, puis du Lapin (Cochard,2004).
L’incorporation au régime alimentairede ces petites proies semble s’initier aucours de l’He1 et s’intensifier à la fin decet épisode climatique, lors du Bølling,alors même que les grands troupeauxd’Ongulés de milieu ouvert sont toujoursabondants (Costamagno et al., 2008).Cependant, d’après F. Delpech (1999,2003), la biomasse des Ongulés com-mencerait à diminuer avant le début duBølling, aux environs de 15500 - 15 000calBP. L’intensification de la capture de
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
195

petits gibiers pourrait-elle être la consé-quence de cette diminution de la disponi-bilité des ressources de haut rang? Cettehypothèse est envisageable mais il fautnoter que, dans les Pyrénées, l’importan-ce de ces proies reste relative puisque,lorsqu’elles sont exploitées, elles four-nissent au maximum 5% du poids desressources carnées (Costamagno etLaroulandie, com. orale). Par ailleurs, laminutie apportée à la préparation culinai-re des oiseaux, notamment sur le site dela Vache, dénote un fort investissement,réduisant d’autant la rentabilité de cesressources. Enfin, élément essentiel, leurplace dans l’économie n’est pas unique-ment alimentaire mais également techni-que et esthétique. Parallèlement, lespremiers développements d’essencesvégétales tels que le Genévrier, suivi duBouleau et du Saule qui sont enregistrésdans plusieurs carottes continentales desPyrénées (Jalut et al., 1992, 1998) autourde 15 500 calBP constituent un environ-nement favorable aux Lagopèdes notam-ment qui se nourrissent de ces plantes.Selon cette hypothèse, il est envisageableque la disponibilité de cette ressourceaviaire potentielle ait augmenté durantcette période. Si d’autres facteurs ontconjointement pu motiver l’acquisitiondes ces proies, ces conditions l’ont, aminima, rendu possible. L’intégration deces nouvelles ressources (alimentaire ettechnique) dans l’économie des sociétésmagdaléniennes implique de nouvellesactivités que ce soit dans leur acquisition,leur traitement ou leur utilisation. Lesmodes de représentation que les chas-seurs-cueilleurs ont de leur environne-ment et les codes sociaux qui y sont
associés s’en trouvent également modi-fiés; ces changements économiques par-ticipent de modifications plus vastesperceptibles dans d’autres sphères d’acti-vités liées à l’exploitation des ressourcesminérales, à l’armement et à l’universsymbolique. Leurs causes doivent êtreappréhendées dans un système complexed’actions et de rétroactions.
Conclusion
Au cours du Tardiglaciaire sur le ver-sant nord des Pyrénées, de profondschangements sont perceptibles du pointde vue de la grande faune. Dès le débutde l’interstade Bølling/Allerød, on obser-ve une diminution des effectifs de renneset d’ongulés steppiques, en particulier duBison, et une recrudescence des popula-tions de Cerf. Cette baisse progressive dela biomasse en ongulés steppiquescorrespond également à un changementculturel à partir de 12500 BP, soit vers15000-14500 calBP. On observe alors lacoexistence de sites attribués au Magda-lénien supérieur et à l’Azilien ancien.Cette contemporanéité ne pourrait êtrequ’une illusion du plateau 14C et l’hy-pothèse d’une azilianisation précoce duMagdalénien (au cours du Bølling) peutêtre envisagée au regard également deplusieurs dates de l’Epipaléolithiqueancien dans l’ensemble de l’Europe del’Ouest. Cette évolution se marque par leremplacement des pointes lithiques audétriment des pointes osseuses armées debarbelures lithiques, transformation ducarquois qui semble définitive à partir del’Allerød. C’est alors que le développe-ment du couvert forestier permet la mise
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard196

en place d’une biocénose animale fores-tière. Les premières dates obtenues surdes os de rennes issus des couches 3 deDuruthy et de Dufaure semblent d’ai-lleurs remettre en cause l’hypothèsed’une perduration tardive du Renne dansle massif pyrénéen. Si la disparition défi-nitive de cet animal dans cette régionsemble ainsi postérieure aux débuts del’azilianisation du Magdalénien, engagéedès la fin du Dryas ancien à travers ledéveloppement de pointes lithiques(Langlais, 2007), il faut sans doutedésormais envisager ce processus nonplus comme une rupture abrupte maiscomme un lent processus de désagréga-tion des troupeaux qui a participé très tôtà certains changements économiques.L’intensification de la chasse aux petitsgibiers, qui, sur le versant nord des Pyré-nées, concerne principalement les Oise-aux, s’amorce vers 16000 calBP alorsque la recomposition de la grande faunen’a pas encore débuté. La diminution dela biomasse des Ongulés vers 15500calBP (Delpech, 1999, 2003) elle-mêmeen relation avec les changements du cou-vert végétal pourrait être un des facteursdéclenchants, les mutations socio-écono-miques marquant le développement duMagdalénien supérieur ne devant pourautant pas être sous-estimées (Langlais,2007; Costamagno et al., 2008).
Bibliographie
ALTUNA J., MARIEZKURRENA K. (1995) -Les restes osseux de macromammifères,Straus L.G. (Ed.), Les derniers chasseurs derennes du monde pyrénéen. L'abri Dufaure: ungisement tardiglaciaire en Gascogne, Paris,Mémoire de la Société Préhistorique françaiseXXII, 181-211.
ANDERSEN K.K., SVENSSON A.M., JOHN-SEN S.J., RASMUSSEN S.O., BIGLER M.,RÖTHLISBERGER R., RUTH U., SIGGA-ARD-ANDERSEN M.-L., STEFFENSENJ.P., DAHL-JENSEN D., VINTHER B.M.,CLAUSEN H.B. (2006) - The Greenland IceCore chronology 2005, 15-42 ka. Part 1: cons-tructing the time scale, Quaternary ScienceReviews, 25, 3246-3257.
ARAMBOUROU R. (Ed.) (1978) - Le gisementpréhistorique de Duruthy à Sorde l'Abbaye(Landes). Bilan des recherches de 1958 à1975, Paris, Mémoire de la Société préhistori-que française XIII, 153 p.
BARBAZA M. (1996) - Le Magdalénien termi-nal des Pyrénées françaises, L'art préhistori-que des Pyrénées, Paris, Musée des Antiquitésnationales, Réunion des Musées nationaux,124-131.
BEAULIEU J.-L. (de), PONS A., REILLE M.(1988) - Histoire de la flore et de la végétationdu Massif central (France) depuis la fin de ladernière glaciation, Cahier de Micropaléonto-logie, 3, 5-36.
BOUCHARD L. (2007) - L’avifaune du site tar-diglaciaire de Rhodes II (Ariège): Approchetaphonomique, archéozoologique et paléoen-vironnementale, Mémoire de Master 2, Men-tion Anthropologie Biologie et Préhistoire,Université de Bordeaux I, 71 p.
BRIDAULT A., CHAIX L., PION G., OBER-LIN C., THIEBAULT S., ARGANT J.(2000)- Position chronologique du Renne(Rangifer tarandus L.) à la fin du Tardiglaciai-re dans les Alpes du Nord françaises et le Juraméridional, Le Paléolithique supérieur récent:nouvelles données sur le peuplement et l'envi-ronnement, Paris, Société préhistoriquefrançaise, Actes de la table ronde de Cham-bery, 12-13 mars 1999, 47-57.
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
197

BRIDAULT A., FONTANA L. (2003) - Enregis-trement des variations environnementales parles faunes chassées, dans les zones de moyen-ne montagne d'Europe occidentale, au Tardi-glaciaire et au début de l'Holocène,Patou-Mathis M., Bocherens H. (Eds.), Le rôlede l'environnement dans les comportementsdes chasseurs-cueilleurs préhistoriques,Oxford, BAR International Series 1105, 55-65.
BRONK RAMSEY C. (1995) - Radiocarboncalibration and analysis of stratigraphy: TheOxCal program, Radiocarbon, 37, 425-430.
BRONK RAMSEY C. (2001) - Development ofthe radiocarbon calibration program OxCal,Radiocarbon, 43, 355-363.
CACHO I., GRIMALT J.O., CANALS M.,SBAFFI L., SHACKLETON N.J., SCHÖN-FELD J., ZAHN R. (2001) - Variability of thewestern Mediterranean Sea surface temperatu-re during the last 25,000 years and its connec-tion with the Northern Hemisphere climaticchanges, Paleoceanography, 16, 40-52.
CASSOLI P.F., TAGLIACOZZO A. (1997) -Butchering and Cooking of Birds in the Palae-olithic Site of Grotta Romanelli (Italy), Inter-national Journal of Osteoarchaeology, 7,303-320.
CHAIX L. (2003) - L’exploitation de la faune auMagdalénien en Suisse et dans les régionslimitrophes, Costamagno S., Laroulandie V.(Eds.), Mode de vie au Magdalénien: lesapports de l'archéozoologie, Oxford, BARInternational Series 1144, Liège, XIV°Congrès UISPP, 2-8 septembre 2001, 65-71.
CHOLLOT M. (1964) - Musée des AntiquitésNationales - Collection Piette - Art mobilierpréhistorique, Paris, Musées Nationaux, 479 p.
CLOT A. (1983) - Déterminations de paléontolo-gie quaternaire dans le bassin de l'Adour (Pre-mière série, Pyrénées Atlantiques),Archéologie des Pyrénées occidentales, 3,121-139.
CLOT A., OMNÈS J. (1979) - Premiers datagesradiocarbone du Magdalénien des Hautes-Pyrénées, Bulletin de la Société PréhistoriqueFrançaise, 76, 324-339.
CLOTTES J., DELPORTE H. (Eds.) (2003) - LaGrotte de La Vache (Ariège). Fouilles Romain
Robert, Paris, R.M.N./C.T.H.S., 2 vol.,407/463 p.
COCHARD D. (2004) - Les Léporidés dans lasubsistance des Paléolithiques du Sud de laFrance, Bordeaux, Thèse de Doctorat, Univer-sité de Bordeaux I, 346 p.
COSTAMAGNO S. (1999) - Stratégies de chas-se et fonction des sites au Magdalénien dans lesud de la France, Bordeaux, Thèse de Docto-rat, Université de Bordeaux I, 2 t., 495 p., 329tabl., 132 fig., 128 pl.
COSTAMAGNO S. (2001) - Exploitation del’Antilope saïga au Magdalénien en Aquitaine:méthodes d’étude et applications archéologi-ques, Paleo, 13, 111-127.
COSTAMAGNO S. (2003) - Exploitation de lagrande faune au Magdalénien dans le sud de laFrance, Costamagno S., Laroulandie V. (Eds.),Mode de vie au Magdalénien: les apports del'archéozoologie, Oxford, BAR InternationalSeries 1144, Liège, XIV° Congrès UISPP, 2-8septembre 2001, 73-88.
COSTAMAGNO S. (2005) - Mobilité, territoiresde chasse et ressources animales au Magdalé-nien final en contexte pyrénéen: le niveau 7ade la grotte-abri du Moulin (Troubat, Hautes-Pyrénées), Jaubert J., Barbaza M. (Eds.),Territoires, déplacements, mobilité, échanges,Paris, C.T.H.S., Actes du 126eme congrés natio-nal des sociétés historiques et scientifiques,Toulouse, du 9 au 14 avril 2001, 371-383.
COSTAMAGNO S. (2006) - Archéozoologie desgrands mammifères des gisements de la falai-se du Pastou, Dachary M. (Ed.), Les Magdalé-niens à Duruthy, La Rochelle, ImprimerieRochelaise, 20-29.
COSTAMAGNO S., COCHARD D., FERRIÉ J.-G., LAROULANDIE V., CAZALS N., LAN-GLAIS M., VALDEYRON N., DACHARYM., BARBAZA M., GALOP D., MARTIN H.,PHILIBERT S. (2008) - Nouveaux milieux,nouveaux gibiers, nouveaux chasseurs? Evo-lution des pratiques cynégétiques dans lesPyrénées du Tardiglaciaire au début du Post-glaciaire, Bulletin de la Société PréhistoriqueFrançaise, 105, 17-27.
COSTAMAGNO S., LAROULANDIE V.(2004) - L'exploitation des petits vertébrésdans les Pyrénées françaises du Paléolithique
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard198

au Mésolithique: un inventaire taphonomiqueet archéozoologique, Brugal J.-P., Desse J.(Eds.), Petits animaux et sociétés humaines,Antibes, APDCA, XXIV° Rencontres Interna-tionales d'Archéologie et d'Histoire d'Antibes,23-25 octobre 2003, 403-416.
COSTAMAGNO S., MATEOS CACHORRO A.(2007) - Milieu animal de part et d'autre de lachaîne pyrénéenne: implications sur les modesde subsistance au Magdalénien, Cazals N.,González Urquijo J., Terradas X. (Eds.), Fron-tières naturelles et frontières culturelles dansles Pyrénées préhistoriques, Santander, PUbli-Can-Ediciones de la Universidad de Canta-bria, 53-74.
DACHARY M. (2002) - Les Magdaléniens desPyrénées occidentales, Paris, Thèse de Docto-rat, Université de Paris X, 2 t., 299p., 247 fig.,XLVII pl.
DELPECH F. (1978) - Les faunes magdalénien-nes et aziliennes du gisement de Duruthy, àSorde l'Abbaye (Landes), Arambourou R.(Ed.), Le gisement préhistorique de Duruthy àSorde l'Abbaye (Landes). Bilan des recherchesde 1958 à 1975, Paris, Mémoire de la Sociétépréhistorique française XIII, 110-116.
DELPECH F. (1981) - La faune magdaléniennede la Salle des Morts à Enlène, Montesquieu-Avantès (Ariège), Clottes J. (Ed.), La Préhis-toire du Quercy dans le contexte deMidi-Pyrénées. Compte Rendu de la XXIe ses-sion, Congrès Préhistorique de France, Paris,S.P.F., C.N.R.S., 65-69.
DELPECH F. (1983) - Les faunes du Paléolithi-que supérieur dans le Sud-Ouest de la France,Bordeaux, Cahier du Quaternaire, C.N.R.S.,453 p.
DELPECH F. (1988) - Les réponses des Ongulésdu Pléistocène Supérieur aux changements cli-matiques en Aquitaine (Sud-Ouest de la Fran-ce): quelques exemples, Geobios, 20, 495-503.
DELPECH F. (1989) - L'environnement animaldes magdaléniens, Otte M. (Ed.), Le Magdalé-nien en Europe, Liège, ERAUL, Actes duColloque de Mayence, XI° congrès U.I.S.P.P.,5-30.
DELPECH F. (1999) - Biomasse d'Ongulés auPaléolithique et inférences sur la démograp-hie, Paleo, 11, 19-42.
DELPECH F. (2003) - L'environnement animaldes européens au Paléolithique supérieur, Des-brosse R., Thévenin A. (Eds.), Préhistoire del'Europe. Des origines à l'Âge du Bronze,Paris, C.T.H.S., Actes du 125è Congrès natio-nal des Sociétés historiques et scientifiques,Lille 2000, 271-289.
DELPECH F., DONARD E., GILBERT A.,GUADELLI J.-L., GALL O.L., JACQUINA.M., PAQUEREAU M.-M., PRAT F.,TOURNEPICHE J.-F. (1983) - Contribution àla lecture des paléoclimats quaternaires d'aprèsles données de la paléontologie en milieu con-tinental. Quelques exemples de flore et defaune d'Ongulés pris dans le Pléistocène supé-rieur, Paléoclimats, Bulletin de l'Institut géo-logique du Bassin d'Aquitaine, Bordeaux,C.N.R.S., Cahiers du Quaternaire, 165-177.
DELPECH F., LE GALL O. (1983) - La faunemagdalénienne de la grotte des Eglises (Ussat,Ariège), Bulletin de la Société Préhistoriquede l'Ariège, 36, 91-118.
DELPECH F., VILLA P. (1993) - Activités dechasse et de boucherie dans la grotte des l'E-glises, Desse J., Audoin-Rouzeau F. (Eds.),Exploitation des animaux sauvages à travers letemps. XIII° Rencontre Internationales d'Ar-chéologie et d'Histoire d'Antibes. IV° Collo-que international de l'Homme et de l'Animal,Antibes, APDCA, 79-102.
DIEZ FERNANDEZ-LOMANA C., SANCHEZMARCO A., MORENO LARA V. (1995) -Grupos avicaptores del Tardiglaciar: Las avesde Berroberria, Munibe, 47, 3-22.
DRUCKER D., BOCHERENS H., BRIDAULTA., BILLIOU D. (2003) - Carbon and nitrogenisotopic composition of red deer (Cervus elap-hus) collagen as a tool for tracking palaeoen-vironmental change during the Late-Glacialand Early Holocene in the northen Jura (Fran-ce), Palaeogeography, Palaeoclimatology,Palaeoecology, 195, 375-388.
DRUCKER D., BRIDAULT A. (2004) - Déve-loppement du couvert forestier dans le Juradepuis le dernier interglaciaire: traçage par lesteneurs en carbone-13 du collagène des cerfs,Lettre pigb-pmrc, 16, 59-60.
DRUCKER D., CÉLÉRIER G. (2001) - Teneursen carbone-13 du collagène de grands mam-
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
199

mifères du site de Pont-d'Ambon (Dordogne,France): implications pour l'environnement etson exploitation au Tardiglaciaire dans le sud-ouest de la France, Paleo, 13, 145-158.
EASTHAM A. (1998) - Magdalenians andSnowy-Owls: bones recovered at the grotte deBourouilla (Arancou, Pyrénées atlantiques),Paleo, 10, 95-107.
EASTHAM A.S. (1995) - L'écologie Avienne,Straus L.G. (Ed.), Les derniers chasseurs derennes du monde pyrénéen. L'abri Dufaure: ungisement tardiglaciaire en Gascogne, PAris,Mémoire de la Société Préhistorique françaiseXXII, 219-245.
ELLIOT M., LABEYRIE L., DUPLESSY J.-C.(2002) - Changes in North Atlantic deep-waterformation associated with the Dansgaard-Oeschger temperature oscillations (60-10 ka),Quaternary Science Reviews, 21, 1153-1165.
ENLOE J.G. (1999) - Hunting specialization:single-species focus and human adaptation,Brugal J.-P., David F., Enloe J.G., Jaubert J.(Eds.), Le Bison: gibier et moyen de subsis-tance des hommes du Paléolithique aux Palé-oindiens des grandes plaines, Antibes,APCDA, Actes du Colloque international,Toulouse 1995, 501-509.
FONTANA L. (1998a) - Mobilité et subsistanceau Magdalénien dans le Languedoc occidentalet le Roussillon, Paris, Thèse de Doctorat,Université de Paris I, 2 t., 285p., 297fig.,296tabl. p.
FONTANA L. (1998b) - Subsistance et territoireau Magdalénien supérieur dans les Pyrénées:l'apport des données archéozoologiques de lagrotte de Belvis, Bulletin Préhistoire du sud-Ouest, Nouvelles Etudes, 5, 131-146.
FONTANA L. (2003) - Characterization andExploitation of the Arctic Hare (Lepus timi-dus) during the Magdalenian: Surprising Datafrom Gazel Cave (Aude, France), CostamagnoS., Laroulandie V. (Eds.), Mode de vie auMagdalénien: les apports de l'archéozoologie,Oxford, BAR International Series 1144,Liège, XIV° Congrès UISPP, 2-8 septembre2001.
FOSSE P. (2000) - La grande faune mammalien-ne: remarques préliminaires, Chauchat C.(Ed.), L'habitat magdalénien de la grotte du
Bourouilla à Arancou (Pyrénées-Atlantiques) -Gallia Préhistoire, 41, 98-113.
GARCIA PETIT L. (1997) - Les restes d'oiseauxdes sites de Seriyá (Pays Catalan), FullolaJ.M., Soler N. (Eds.), El Món Mediterrani des-prés del Pleniglacial (18,000-12,000 BP),Museu d'Arqueologia de Catalunya-Girona,Girona, 329-344.
GRAYSON D.K. (1984) - Quantitative Zooar-chaeology Topics in the Analysis of Archaeo-logical Faunas, New York, Academic Press,202 p.
GRIGGO C. (1995) - Significations paléoenvi-ronnementales des communautés animalespléistocènes reconnues dans l'abri Suard (Cha-rente) et la Grotte de Bois-Ragot (Vienne):Essai de quantification de variables climati-ques, Bordeaux, Thèse de Doctorat, Universi-té de Bordeaux I, 206 p.
HÉLÉNA P. (1928) - La stratigraphie de la grot-te de la Crouzade, Bull. Comm. archéol. Nar-bonne, XVII, 50 p.
JALUT G., GALOP D., BELET J.M., AUBERTS., ESTEBAN AMAT A., BOUCHETTE A.,DEDOURAT J.-J., FONTUGNE M. (1998) -Histoire des forêts du versant nord des Pyré-nées au cours des 30 000 dernières années, J.Bot. Soc. Fr., 5, 73-84.
JALUT G., MONSERRAT MARTI J., FON-TUGNE M., DELIBRIAS G., VILAPANAJ.M., JULIA R. (1992) - Glacial to Interglacialvegetation changes in the northern and sout-hern Pyrénées: Deglaciation, vegetation coverand chronology, Quaternary Science Reviews,11, 449-480.
JALUT G., TURU I MICHELS V. (ce volume) -La végétation des Pyrénées françaises lors dudernier épisode glaciaire et durant la transitionglaciaire-interglaciaire.
LACOMBE S. (1998) - Préhistoire des groupesculturels au Tardiglaciaire dans les Pyrénéescentrales, Toulouse, Thèse de Doctorat, Uni-versité de Toulouse II, 2 t., 385 p.
LANGLAIS M. (2007) - Dynamiques culturellesdes sociétés magdaléniennes dans leurs cadresenvironnementaux. Enquête sur 7000 ans d'é-volution de leurs industries lithiques entreRhône et Ebre, Toulouse/Barcelone, Thèse deDoctorat, Université de Toulouse II, Universi-
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard200

tat de Barcelona, 549 p.LAROULANDIE V. (1998) - Etudes archéozoo-
logique et taphonomique des lagopèdes dessaules de la grotte magdalénienne des Eglises(Ariège), Anthropozoologica, 28, 45-54.
LAROULANDIE V. (2000) - Taphonomie etarchéozoologie des oiseaux en grotte: Appli-cations aux sites paléolithiques du Bois-Ragot(Vienne), de Combe Saunière (Dordogne) etde la Vache (Ariège), Bordeaux, Thèse d'Uni-versité, Université de Bordeaux I, 396 p.
LAROULANDIE V. (2003) - Exploitation desOiseaux au Magdalénien en France: état deslieux, Costamagno S., Laroulandie V. (Eds.),Mode de vie au Magdalénien: les apports del'archéozoologie, Oxford, BAR InternationalSeries 1144, Liège, XIV° Congrès UISPP, 2-8septembre 2001, 129-138.
LAROULANDIE V. (2005) - Bird exploitation:the cas of Ptarmigan Lagopus sp. in the UpperMagdalenian site of La Vache (Ariège, Fran-ce), Grupe G., Peters J. (Eds.), Feathers, gritand symbolism. Birds and humans in theancient Old and New Worlds, Rahden/Westf,Verlag Marie Leidorf, Proceedings of the 5thMeeting of the ICAZ Bird Working Group,Munich, 26-28 July 2004, 165-178.
LAROULANDIE V. (2006) - Les restes d’oise-aux des gisements de la falaise du Pastou,Dachary M. (Ed.), Les Magdaléniens àDuruthy, Imprimerie Rochelaise, La Rochelle,20-29.
LAROULANDIE V. (2007) - Les restes aviairesdes niveaux aziliens de la grotte-abri du Mou-lin (Troubat, Hautes-Pyrénées): paléoenviron-nement et modalités d’exploitation,Préhistoire du sud-Ouest, 14, 19-29.
LAROULANDIE V. (sous presse) - De la plumeà l’œuf: exploitation des ressources aviaires auMagdalénien dans le Sud de la France, Fonta-na L., Chauvière F.-X., Bridault A. (Eds.),Exploitation du monde animal par les sociétésde chasseurs-cueilleurs préhistoriques. Envi-ronnements, subsistance et comportementstechniques, Oxford, BAR International Series,Lisbonne, XV° Congrès UISPP, 4-9 septem-bre 2006.
LAROULANDIE V., VILETTE P. (soumis-a) -Les ressources aviaires dans le Magdalénien
moyen de Gazel, Sacchi D. (Ed.), Monograp-hie de la grotte Gazel.
LAROULANDIE V., VILETTE P. (soumis-b) -Notes sur les ressources aviaires dans l’Épi-magdalénien et l’Aziloïde de Gazel, Sacchi D.(Ed.), Monographie de la grotte Gazel.
LE GALL O. (1999) - Ichtyophagie et pêchespréhistoriques. Quelques données de l'Europeoccidentale, Bordeaux, Thèse d'Etat, Universi-té Bordeaux I, 476 p.
LE GALL O. (2003) - Des Magdaléniens et... despoissons, Costamagno S., Laroulandie V.(Eds.), Mode de vie au Magdalénien: lesapports de l'archéozoologie, Oxford, BARInternational Series 1144, Liège, XIV°Congrès UISPP, 2-8 septembre 2001, 119-128.
MAGNY M. (1995) - Une histoire du climat -Des derniers mammouths au siècle de l'auto-mobile, Paris, Errance, 176 p.
MARTIN H. (1994) - Nouveaux milieux, nouve-aux chasseurs, une approche des comporte-ments au Post-glaciaire à travers l'étude dessaisons de capture du gibier, Toulouse, Thèsede Doctorat, Université de Toulouse 2 - LeMirail, 383 p.
MARTZLUFF M. (ce volume) - L'Azilien pyré-néen entre Garonne et Ebre: un état de la ques-tion.
MATEOS CACHORRO A. (2000/2001) - Frac-turation anthropique intentionnelle sur mandi-bules et phalanges dans le niveau VIII de lagrotte de Las Caldas (Asturies, Espagne),Préhistoire Européenne, 16/17, 255-270.
MATEOS CACHORRO A. (2003) - Estudio dela fragmentación de falanges y mandíbulas enloa secuencia temporal del 19000-13000 BPde la cueva de las Caldas (Priorio, Oveido).Implicaciones paleoeconómicas: nutrición ysubsistencia, Gallaecia, 22, 9-20.
MOIGNE A.-M. (2003) - Les faunes de grandsmammifères de la grotte des Conques à Vin-grau, Baills H. (Ed.), Les Conques: des chas-seurs magdaléniens et leur territoire, ERAUL,Liège, 75-103.
MOIGNE A.-M., BAILLS H., GRÉGOIRE S.(1998) - Les magdaléniens de la grotte desConques (Pyrénées-Orientales), caractérisa-tion du site d'après les restes osseux et l'outi-
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
201

llage, Brugal J.-P., Meignen L., Patou-MathisM. (Eds.), Economie préhistorique: les com-portements de subsistance au Paléolithique,Antibes, APCDA-CNRS, XVIII° RencontresInternationales d'Archéologie et d'Histoired'Antibes, 397-411.
MOURER-CHAUVIRÉ C. (1983) - Les oiseauxdans les habitats paléolithiques: gibier deshommes ou proies des rapaces?, Grigson C.,Clutton-Brock J. (Eds.), Animals in Archaeo-logy: Shell middens, fishes, and birds, Oxford,BAR international series 163, 111-124.
NAUGHTON F., SANCHEZ GOÑIA M.F.,DESPRATA S., TURONA J.-L., DUPRATAJ., MALAIZÉA B., JOLIA C., CORTIJOC E.,DRAGOD T., FREITASE M.C. (2007) - Pre-sent-day and past (last 25 000 years) marinepollen signal off western Iberia, MarineMicropaleontology, 62, 91-114.
PAILHAUGUE N. (1995) - La faune de la SalleMonique, Grotte de la Vache (Alliat, Ariège),Bulletin de la Société Préhistorique de l'Ariè-ge, L, 225-289.
PAILHAUGUE N. (1998) - Faune et saisonsd'occupation de la salle Monique au Magdalé-nien pyrénéen, Grotte de la Vache (Alliat,Ariège, France), Quaternaire, 9, 385-400.
PAILLER D., BARD E. (2002) - High frequencypalaeoceanographic changes during the past140000 yr recorded by the organic matter insediments of the Iberian Margin, Palaeogeo-graphy, Palaeoclimatology, Palaeoecology,181, 431-452.
PELEGRIN J. (2000) - Les techniques de débita-ge laminaire au Tardiglaciaire: critères dediagnose, Valentin B., Bodu P., ChristensenM. (Eds.), L'Europe centrale et septentrionaleau Tardiglaciaire, Nemours, APRAIF, Actesde la Table-ronde internationale de Nemours,14-16 mai 1997, 73-86.
PÉTILLON J.-M. (2006) - La chasse au Magda-lénien, Pour la Science, 341, 47-53.
PÉTILLON J.-M., LETOURNEUX C., LAROU-LANDIE V. (sous presse) - Archéozoologiedes collections anciennes: le cas de la faune duMagdalénien supérieur de la grotte d'Isturitz,Normand C. (Ed.), Les recherches archéologi-ques dans les grottes d’Isturitz et d’Oxocelha-ya de 1912 à nos jours: une synthèse des
résultats, Paleo hors-série, Actes de la tableronde du cinquantenaire du classement commeMonument Historique des grottes d’Isturitz etd’Oxocelhaya, Hasparren, 14-15 novembre2003.
RASMUSSEN S.O., ANDERSEN K.K.,SVENSSON A.M., STEFFENSEN J.P.,VINTHER B.M., CLAUSEN H.B., DAHL-JENSEN D., BIGLER M., RÖTHLISBER-GER R., FISCHER H., GOTO-AZUMA K.,HANSSON M.E., RUTH U. (2006) - A newgreenland ice core chronology for the last gla-cial termination, Journal of Geophysical Rese-arch, 111, 1-14.
REILLE M., ANDRIEU V. (1995) - The latePleistocene and Holocene in the LourdesBasin, Westren Pyrénées, France: new pollenand chronological data, Vegetation Historyand Archaeobotany, 4, 1-21.
REIMER P.J., BAILLIE M.G.L., BARD E.,BAYLISS A., BECK J.W., BERTRAND C.,BLACKWELL P.G., BUCK G., BURR C.E.,CUTLER K.B., DAMON P.E., EDWARDSR.L., FAIRBANKS R.G., FRIEDRICH M.,GUILDERSON T.P., HUGHEN K.A., KRO-MER B., MCCORMAC F.G., MANNING S.,BRONK R.C., REIMER R.W., REMMELES., SOUTHON J.R., STUIVER M., TALA-MO S., TAYLOR F.W., VAN DER PLICHTJ., WEYHENMEYER C. (2004) - IntCal04Atmospheric radiocarbon age calibration, 26-0ka BP, Radiocarbon, 46, 1029-1058.
RICHARD H., BÉGEOT C., GAUTHIER E.,RUFFALDI P. (2000) - Evolution du couvertvégétal du Tardiglaciaire et du début de l'Ho-locène sur la chaîne jurassienne: nouveauxrésultats, Les derniers chasseurs-cueilleursd'Europe occidentale (13000 - 5500 av. J.-C.),Besançon, Presses Universitaires Franc-com-toises, Actes du Colloque internationale deBesançon, 23-25 octobre 1998, 29-36.
ROWLEY-CONWY P. (1986) - Between cavepainters and crop planters: aspects of the tem-perate European Mesolithic, Zvelebil M.(Ed.), Hunters in Transition. Mesolithic Socie-ties of Temperate Eurasia and their Transitionto Farming, Cambridge, Cambridge Univer-sity Press, 13-32.
SACCHI D. (1986) - Le Paléolithique supérieur
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard202

du Languedoc occidental et du Roussillon,C.N.R.S., Paris, 284 p.
SACCHI D. (1994) - Un site paléolithique supé-rieur de moyenne altitude dans les Pyrénées:La Cauna de Belvis (France), Human adapta-tions to the moutain environnement in theUpper Palaeolithic and Mesolithic, PreistoriaAlpina, Trente, 1992, 59-90.
SCHMIDER B. (2003) - L'Outillage lithique dela salle Monique, Clottes J., Delporte H.(Eds.), La grotte de La Vache (Ariège). Foui-lles Romain Robert. Vol. 1, Les occupationsdu Magdalénien, Paris, R.M.N./C.T.H.S., 169-196.
SPIESS A. (1995) - Etude de la saison d'habita-tion au moyen du cément dentaire, Straus L.G.(Ed.), Les derniers chasseurs de rennes dumonde pyrénéen. L'abri Dufaure: un gisementtardiglaciaire en Gascogne, Mémoire de laSociété Préhistorique française XXII, Paris,213-217.
STRAUS L.G., SPIESS A.E. (1985) - Le Magda-lénien final de l'Abri Dufaure (Sordes de l'Ab-baye, Landes): un aperçu de la chronologie etde la saison d'occupation humaine, Bulletin dela Société Préhistorique de l'Ariège, 40, 169-184.
SVENSSON A.M., ANDERSEN K.K., BIGLERM., CLAUSEN H.B., DAHL-JENSEN D.,DAVIES S.M., JOHNSEN S.J., MUSCHE-LER R., RASMUSSEN S.O., STEFFENSENJ.P., VINTHER B.M. (2006) - The GreenlandIce Core chronology 2005, 15-42 ka. Part 2:comparison to other records, QuaternaryScience Reviews, 25, 3258-3267.
TAGLIACOZZO A., GALA M. (2002) - Exploi-tation of Anseriformes at two Upper Palaeolit-hic sites in Southern Italy: Grotta Romanelli(Lecce, Apulia) and Grotta del Santuario dellaMadonna a Praia a Mare (Cosenza, Calabria),Bochenski Z.M., Bochenski Z., Stewart J.R.(Eds.), Proceedings of the 4th Meeting of theICAZ Bird Working Group, Krakow, Poland,11-15 september 2001, Acta Zoologica Craco-viensia 45 (special issue), 117-131.
VALDEYRON N., DETRAIN L. (ce volume) -La fin du Tardiglaciaire en Agenais, Périgordet Quercy: état de la question, perspectives.
VALENTIN B. (2005) - Transformations de l'in-
dustrie lithique pendant l'Azilien. Etude desniveaux 3 et 4 du Bois-Ragot, Chollet A.,Dujardin V. (Eds.), La grotte du Bois-Ragot àGouex (Vienne). Magdalénien et Azilien,Paris, Mémoire de la Société préhistoriquefrançaise XXXVIII, 89-182.
VILETTE P. (1983) - Avifaunes du Pléistocènefinal et de l'Holocène dans le Sud de la Franceet en Catalogne, Atacina, 11.
VILETTE P. (1999) - Bilan provisoire sur lachasse aux oiseaux pendant le Leptolithiquedans le sud de la France, Sacchi D. (Ed.), Lesfaciès leptolithiques du nord-ouest méditerra-néen: milieux naturels et culturels, Paris,Société préhistorique française, XXIV°Congrès Préhistorique de France, Carcassonne26-30 septembre 1994, 267-276.
WEINSTOCK J. (2000) - Late Pleistocene rein-deer populations in Middle and Western Euro-pe: an osteometrical study of Rangifertarandus, BioArchaeologica, Tübingen, 307 p.
WINTERHALDER B.P. (1981) - Optimal fora-ging strategies and hunter-gatherer research inanthropology: theory and models, Winterhal-der B.P., Smith E.A. (Eds.), Hunter-GathererForaging Strategies: Ethnographic and Archa-eological Analyses, Chicago, University ofChicago Press, 13-36.
Notes
1 GIS-1: Greenland Interstadial 12 15 sites au lieu, par exemple, des 53 utilisés pour la
synthèse sur l’exploitation des petits Vertébrés dans lesPyrénées (Costamagno et Laroulandie, 2004).
3 Bison priscus a été identifié à Dufaure; pour les autresgisements, il n’y a pas d’identification au niveau spécifi-que des restes de Bovinés, en dehors des Conques danslequel est mentionnée la présence de Bos primigenius(Moigne, 2003). Les arguments présentés sur ce site noussemblant insuffisants, nous avons préféré classer ces res-tes dans la sous-famille des Bovinés
4 Les Oiseaux, qui ne sont pas toujours dénombrés, ne sontpas pris en compte
5 Les ensembles osseux du gisement de la Crouzade n’ontpas fait l’objet de quantification. La présence de restes deCerf dans les dépôts attribués au Magdalénien moyen estsignalé (Héléna 1928).
6 Seuls sont pris en compte les Ongulés7 Indice de Shannon: -∑pi/ln(pi))/ln(S) avec pi =
NRDi/NRD, S correspond au nombre de taxons présentsdans l’assemblage, NRDi est le nombre de restes détermi-
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
203
nés pour une espèce donnée, NRD, le nombre des restesdéterminés total
8 Exclusion faite des Lagomorphes9 Ce gisement a livré plus de 8000 restes de Lièvre qui
représentent près de 50% des restes osseux identifiés.
10 Ce gisement a livré près de 50 000 restes de Lagopèdes,soit 4 566 individus.
11 Cette pièce est décrite comme un os d’oiseau dans M.Chollot (1964).
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard204
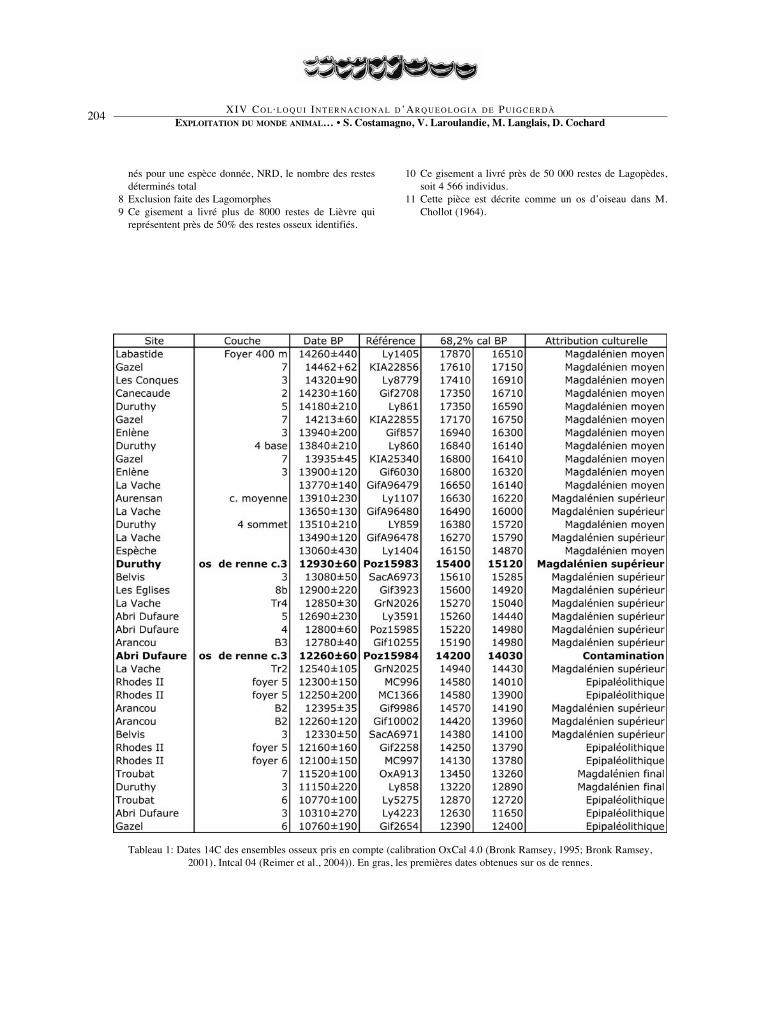
Tableau 1: Dates 14C des ensembles osseux pris en compte (calibration OxCal 4.0 (Bronk Ramsey, 1995; Bronk Ramsey,2001), Intcal 04 (Reimer et al., 2004)). En gras, les premières dates obtenues sur os de rennes.

EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
205
Tab
leau
2: F
réqu
ence
s re
lativ
es d
es g
rand
s M
amm
ifèr
es a
u D
ryas
anc
ien
(NR
D: n
ombr
e de
res
tes
déte
rmin
és)
(d’a
près
Clo
t et O
mnè
s, 1
979;
Del
pech
, 198
1; C
lot,
1983
;D
elpe
ch e
t Le
Gal
l, 19
83 ;
Altu
na e
t Mar
iezk
urre
na, 1
995;
Pai
lhau
gue,
199
5; F
onta
na, 1
998a
, 199
8b; F
osse
, 200
0 ; M
oign
e, 2
003;
Cos
tam
agno
, 200
6 su
r la
bas
e de
sdé
com
ptes
de
F. D
elpe
ch (
1978
) à
Dur
uthy
).
XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
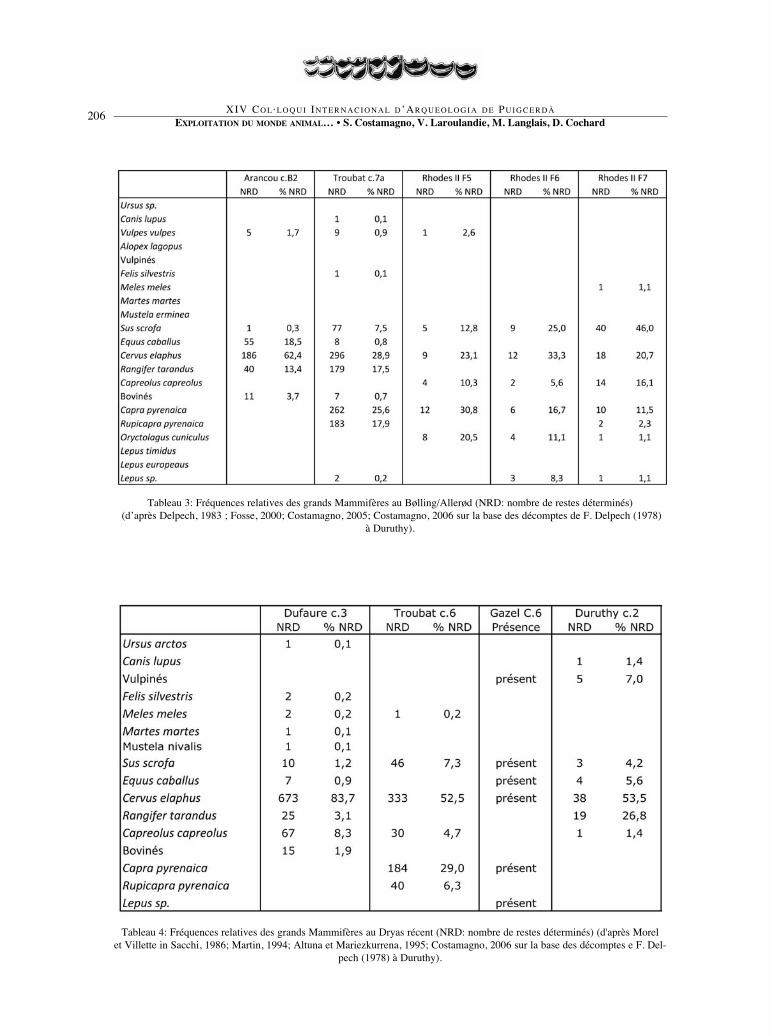
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard206
Tableau 3: Fréquences relatives des grands Mammifères au Bølling/Allerød (NRD: nombre de restes déterminés)(d’après Delpech, 1983 ; Fosse, 2000; Costamagno, 2005; Costamagno, 2006 sur la base des décomptes de F. Delpech (1978)
à Duruthy).
Tableau 4: Fréquences relatives des grands Mammifères au Dryas récent (NRD: nombre de restes déterminés) (d'après Morel et Villette in Sacchi, 1986; Martin, 1994; Altuna et Mariezkurrena, 1995; Costamagno, 2006 sur la base des décomptes e F. Del-
pech (1978) à Duruthy).
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
207
Tableau 5: Richesse taxinomique des ensembles osseux pris en compte par faciès culturels.
Figure 1: Schéma synthétique des cadres climatiques selon les carottes glaciaires (d’après Laroulandie et Langlais,inédit in Langlais, 2007).

XIV CO L·L O Q U I IN T E R N A C I O N A L D’AR Q U E O L O G I A D E PU I G C E R D À
EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard208
Figure 2: Carte de répartition des sites pris en compte.
Figure 3: Représentation des dates 14C des ensembles osseux considérés.
EL S PI R I N E U S I L E S À R E E S C I R C U M D A N T S D U R A N T E L TA R D I G L A C I A L. MU T A C I O N S I F I L I A C I O N S…EXPLOITATION DU MONDE ANIMAL… • S. Costamagno, V. Laroulandie, M. Langlais, D. Cochard
209
Figure 5: Indice de Shannon des spectres fauniques d’Ongulés dans les ensembles osseux considérés (losanges:Magdalénien moyen, triangles: Magdalénien supérieur, cercles: Epipaléolithique).
Figure 4: Fréquence relative des groupes écologiques au sein des ensembles osseux considérés.
Dryas Bølling/Allerød Dryas récent

















![“Le Monde à son image: le cinéma et le mythe d’Icare [Guest Lecture].”](https://static.fdokumen.com/doc/165x107/633d5f5ad0a2f101870ab50b/le-monde-a-son-image-le-cinema-et-le-mythe-dicare-guest-lecture.jpg)


