
A two-step process for the reflooding of the Mediterranean after the Messinian Salinity Crisis
Upload
univ-bordeauxCategory
view
0download
0
REVIEW OF
PALAEOBOTANY AND
PALYNOLOGY ELSEVIER Review of Palaeobotany and Palynology 87 (1995) 51-79
Evolution of the Messinian Mediterranean environments: the Tripoli Formation at Capodarso (Sicily, Italy)
Jean-Pierre Suc ", Donata Violanti b Laurent Londeix c, Claude Poumot d
Christian Robert e , Georges Clauzon f, Frangois Gautier ", Jean-Louis Turon c ,
Jacqueline Ferrier ~, Hafida Chikhi ", Genevi+ve Cambon a
a Palkoenvironnements et Palynologie, Institut des Sciences de l'Evolution, URA 327 CNRS, UniversitO Montpellier II, case 061, F-34095 Montpellier COdex 5, France
b Dipartimento di Scienze della Terra, Universitgt di Torino, via Accademia delle Seienze, 5, 1-10123 Torino, Italy c DOpartement de GOologie et d'OcOanographie, URA 197 CNRS, Universitk Bordeaux I, avenue des FacultOs, F-33405
Talenee COdex, France a DOpartement GOologie SOdimentaire, Elf-Aquitaine Production, F-64018 Pau COdex, France
e Laboratoire de Gkologie du Quaternaire, UPR 1201 CNRS, CNRS-Luminy, case 907, F-13288 Marseille COdex 9, France
: Institut de GOographie, URA 903 CNRS, 29 avenue R. Schuman, F-13621 Aix-en-Provenee COdex, France
Received 24 May 1993; revised and accepted 29 April 1994
Abstract
New field data have been collected from the Capodarso section (Caltanissetta basin, Sicily); 48 samples from the Tripoli Formation were analysed for foraminifera, dinocysts, palynofacies, CaCO3, pollen grains and clay minerals. The uppermost clays of the Terravecchia Formation are clearly the result of deeper deposition than those corresponding to the lowermost analysed samples within the overlying Tripoli Formation. So, the general evolution of the basin was from normal marine conditions to confinement, during which the sedimentation of diatomite vs. clay was mainly controlled by relative sea-level changes. Three successive types of processes are considered to have generated the diatomites:
(1) in the lower part of the section, the sea level was relatively high and deposition of diatomites was aided by sea- level rises of unequal importance and variations of continental precipitations and run-off;
(2) then, relatively minor sea-level oscillations prevailed; however, the second part of the sequence is characterized by a generally strong shallowing trend leading to basin confinement;
(3) the explanation for diatomites during the last member does not rely as much on the occurrence of substantial sea-level rises in an increasingly desiccated basin; on the other hand, an increased number of rivers reached the central area of the basin, which helped to produce the youngest diatomites.
The warm (subtropical to tropical) and xeric climate predisposed to such an evolution, inducing continuous high salt-concentration of marine waters; so, the smallest influx of less saline (oceanic) waters and/or continental freshwater (run-off) could produce water-mass stratification and diatomite deposition. However, not all lithologic breaks are likely to correspond to recorded environmental transitions.
Bio- and magnetostratigraphy of the section allow consistent relationships between the sea-level curve at Capodarso and the 6 '80 curve at Site 588 (southwest Pacific), attesting global eustatic control on the sedimentation of the Caltanissetta basin up to the beginning of the Messinian salinity crisis.
0034-6667/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0034-6667(94)00144-8

52 J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
1. Introduction
The Tripoli Formation (Ogniben, 1957) is gen- erally considered in its entirety as an intermediate lithostratigraphic unit found between, on the one hand, the uppermost Tortonian and lowermost Messinian blue clays (Terravecchia Formation), regarded as corresponding to normal marine con- ditions, and, on the other, the basal limestone (Calcare di Base) and lowermost evaporites (Gessoso-Solfifera Formation) (Colalongo et al., 1979; McKenzie et al., 1979; Rouchy, 1980; Broquet et al., 1981b; Busson, 1990). Several authors have assumed that the Tripoli Formation was partly deposited in a normal marine environ- ment (Gersonde and Schrader, 1984; Suc et al., 1991) or even deep-water conditions (Arambourg, 1925; Selli, 1952; Leonardi, 1959; Schrader, 1975; Hst~ et al., 1978; McKenzie et al., 1979). Most studies have noted the regular diatomite-clay alternations in the Tripoli Formation and have interpreted the diatomite deposition as a conse- quence of alternations in water-mass stratification (McKenzie et al., 1979; Meulenkamp et al., 1979; Broquet et al., 1981b; Gersonde and Schrader, 1984; Thunell et al., 1984; van der Zwaan and Gudjonsson, 1986; Grasso et al., 1990; Suc et al., 1991).
Water-mass stratification led to repetitive diato- mites; however, their cause is still being discussed, and several hypotheses have been proposed:
~ cO o"
~t.# ~ ~ < . : ~ G a s p a
~!2~m~ oi ~ -.~-.-.---.--<;:::y~-.a?--~.-;-.:•:~ Falconara ~ /
--occurrence of periodic upwellings (Gersonde, 1978; Thunell et al., 1984);
--warming of the surface water (Broquet et al., 1981b);
--occurrence of Atlantic water inflow-mass (van der Zwaan and Gudjonsson, 1986; Benson et al., 1991; Suc et al., 1991);
~ ~ 4 8 • [ / I . - 4 7
I I - - - 4 6
I I I -45 ,--. ~
- - ~42 ~41 ~40 ~39 ~38
l m
in
1
J i h
g
f e
d c
b
Ia
-24 ~23
-ii D
-'18 "17 -16 -15
Limestone
F 2 =] Brown clay
Diatomite
Grey clay
Fig. 1. Location map of the Capodarso and other relevant Fig. 2. The Tripoli Formation at the Capodarso section. sections. 1 ~ the Caltanissetta basin (after Decima et al., 1988). Location of samples and identification of units a r.

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 53
After Bennet (1980) This paper After Gersonde & Schrader (1984)
1"
. . . . . . . . . . ii . . . . . . p
" " ' n .
i n
k J i h
Limestone
Brown clay [:2-:]
~LJ . . . . . . . . . . . . . . . . ~
i i -E
_ ° ° . . - ° ° °
Diatomite
Grey clay
~ ]
°
- _
D-----~ 1
° .~ ° o
°
a I lIn
Fig. 3. The section of the Tripoli Formation at Capodarso, studied in this paper, compared to those of Bennet (1980) and Gersonde and Schrader (1984).
- - increase of continental run-off (Meulenkamp et al., 1979).
In addition, Bennet (1980), Gersonde (1980) and McKenzie et al. (1979) have proposed the conjunction of upwelling occurrences and global sea-level rises, and Grasso et al. (1990) have emphasized local tectonic control. Increases of continental water run-off are generally considered to cause Pliocene laminated or diatomaceous deposits in the Central Mediterranean (van der
Zwaan and Gudjonsson, 1986; Hilgen, 1987; Howell et al., 1988).
In order to better understand this situation, we have carried out a detailed sedimentological and paleobiological study of the Tripoli Formation at Capodarso (Fig. 1 ). For the first time, the sampling (48 samples) has taken into account each diato- mite-clay alternation. This study enlarges a former investigation which mainly considered the diatomi- tic layers, and compared them to the underlying

54 J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51 79
clays and the two thin clay layers within the Calcare di Base (Suc et al., 1991). The originality of this work consists in comparing indicators of marine and continental origin (foraminifera, dinocysts, amorphous organic matter, CaCO 3 content, clay minerals, vegetal organic matter, pollen grains).
2. The Capodarso section
The Capodarso section (Caltanissetta basin) is the lower part of the Messinian stratotype pro- posed by Selli (1960). It consists of 170 m of claystones overlain by the Tripoli Formation (thickness 25 m) and the Calcare di Base (thickness 12 m). Only the upper 17 m of the Tripoli Formation are exposed because of a scree hiding the lowermost 8 m. Nevertheless, the study was performed on this section because of the richness of pollen grains due to its location close to the lands bordering the Caltanissetta basin to the north (Fig. 1), unlike other reference sections such as Falconara (the present stratotype; D'Onofrio et al., 1975) and Monte Giammoia (Fig. 1). Eighteen diatomite-clay alternations have been identified (from a to r) and sampled (21 diatomite and 23 clay samples) (Fig. 2). In addition, four samples were taken from the three lowermost clayey intercalations within the Calcare di Base (Fig. 2). This is therefore the most detailed sam- pling ever carried out in the Tripoli Formation. Consistent comparisons are possible with reports
on the Capodarso section by Bennet (1980) and Gersonde and Schrader (1984) (Fig. 3).
Our paleomagnetic investigation of the Tripoli Formation at Capodarso shows that the polarity of the lower part of the section (diatomite a - g ) is reversed and the upper part is normal, except for the latest diatomite (r) and the Calcare di Base which are reversed.
During our sampling in 1990, we observed diverging strong inclinations (50-60 ° ) in the clays of the lower part of the section, revealing an anticlinal axis (Fig. 4). This axis joins the Calcare di Base just where the Tripoli Formation is pinch- ing out (Fig. 4), whereas the Tripoli Formation is thickest in the adjacent syncline (Fig. 4). This observation supports the idea that the considerable variation in thickness of the Tripoli Formation in the Caltanissetta basin was controlled by substra- tum strains (Grasso et al., 1990) and very active synsedimentary tectonics. This refutes the conclu- sions derived from the former sampling in the Terravecchia Formation, especially samples "1" to "15" (Suc and Bessais, 1990), taken from vertical parallel gullies (Fig. 4). All samples of the previous study were taken in the same gully ( "1" -"6" ; "7" - "9" ; "10 . . . . 15") and, as a consequence, are almost contemporaneous. So the three clusters of samples represent three contracted successive time intervals (Fig. 4). As a consequence, the previous sampling on the Terravecchia Formation from the Capodarso section should be considered meaning- less. This observation could explain some aber-
Fig. 4. Photograph of the Capodarso section. Dashed white and black line: anticline axis. Arrows: dips. Continuous white lines and white numbers: previous sampling by Suc and Bessais (1990). Continuous black line and black numbers: present study sampling. Te = Terravecchia Formation; Tr = Tripoli Formation; CB = Calcare di Base Formation.

~ita uvula
70 8,0 90 loo
% Globigerinoides % Globigerina obliquus obliquus % Turborotalia
spp. ÷ G. obliquus extremus quinqueloba
0 10 20 30 0 10 20 30 40 50 60 70 80 0 10 20 30
% Turborotali~ multilobt
o ~ ~ ~
~?~ :~i~i:ii: ~ : ~ i ~ i ~
i~ ~/~ii~ ~ ~ ii~i~i ~ ~ii~i!~i~i~
i iilili/i ili/~/iiiil ¸i~:

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
J 0
% Orbulina suturalis + O. universa
l,o 20 30 40 50 60 70 80 90 100
% Neogloboquadrina acostaens
0 10 20 30 40 50 60 70 80 i i t t i i t | I
S ~
1
, k
J i h
l m
m
f e
d
C
b
I.
q ~ ~42 - 4 ] . . . . . . . . . . ~40
° ' ' : i i ~ - . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ~ - - ~ 3 3
Fig. 5. Foraminifera records.

pp. 57-58 J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
/
B
-1-
- 4 8
~47
P/(P+B) Foramin i f e r s N u m b e r o f P lank ton ic
F o r a m i n i f e r spec ies Dinofl~
Dive r s i ty I
0 10 20 30 40 50 60 70 80 90 100% 0 10 20 30 0.5 1
. ~ - . - ~ - 4 5
r I w ~ ; 42 q 41
-40
o - - a . ~ ' - 3 8
- - :1~ n ~ : :
m
- - - t : ] ~
-28 ii
g .v :i: "15 .:
w "14 ~ " 1 3
f ~ - - :i~ e ~ , , : :
e ~ . 6
l m - - - - - 2 a ~ - I
: ! : : :i? ̧ : i i i : : i i : i : i? ii:?~!:il :i:i:i:?i : i!
H • : :
?ill ! iiiii ¸ i:i:i::il : !ii~il ii iil #iilil
iiii~iiiiiili~ili!i!!iiiiiii!iiiiiiiiiiiiiii:il
10 20 30 4
D o m i n ,
( . . . . . c h a n g e it
Fig. 6. Comparison of some data concerning the marine environment: foraminifers, dinocysts, amorphous organic mat1

30
Neritic (on left) and Oceanic (on right) dinocysts
concentrations (nb/g dry sed.) A.O.M. Halophytes
0 5 lO 15 20 25 x lO 3 0 lO 20 30 40 50 60 70% 0 lO 20 30 40 50 60%
%, ............... i ̧ i ̧¸̧ ~ ~
iiiiiiiiiiiiiii!iiiii~iiii~iii~i%~ ii(:111! i ~
f H
E::I } : ] i> : : : : i ? . . . . ~ ' ~ ' ~ ' t " i i i i i i l l ¸i!!¸¸:?¸i¸> i
!!i'ii!-!!ii-!!i-~i~i-!iii-i!iii-ii!
- i i iil i ' - ~ i ii?~ii?i ¸ i ~i~ii?~? iiiiiii E ~ ~ ......
~i~ ~ i ~:~: ~ : ~: ~ ~ ~ ~ ~ ~ ¢
~ : : ~ x ~ : ~ : iiiiiiii~i~i~ ~ ~ : ~ ~ ~ ~ ~ ~i~ ~ i~:

: i _ ::~:~ii~iii~i~iJ ~ i ~i~i ~
sp~ooa.* l s , ~ o u ! G "L "~!~I
• . / ,+
. . . . J L --
i f - [ ~ ~! ~iil i iiiii ̧ i i:;il iii
i~/ii~iii '~iil/i?i~ ~ 7 __---/' I L ,YlI)I?/~i~ii ̧ "----" I
I£-.,
/ / i / l t t / I t / t t l t t / I / / / I l i l / / / l / / I / / / I / I t % % % % b % % % % % % % % " . . . . % % % % % % % % % % % % % % % % % % % / / / / l / / t t t t l / . I S ~ n O M l l / I l l / l / t i t / t r i l l % % % % + % % % % % % % % " . ~ * ~ % / % % % % % % % % % % % % % % % % % t / l / l / t / / / / t / / / / / / / / / I t / t / l / t / I t / / / / / / / % % % % ~ % % % % % % % % % % % % % % % % % % % % % % % % % % % % % % % % % / / / t i l l / t / t t / t / 1 / t / / / t / t / / / / t / / / / t / /
I
I,[-4 - - _ _ - -
LI - - - -
" "81 - - - - - -
U I
6 ~ ~ 0 t , - 4 - . . . . '
~ t , ' - q - -
91'-4 L~-J- - J - L ~
O'I 0 0'[
.,,,,,+oe Off Or, 0£ O~ O[ 0 ~OlXOt, I O ~ l O O I 08 O' 0 9 Or , O g
+,o*
q :)
P
t
? f
~ i '
I
U
o
d
b
.1
~9-6~ (~66I ) /g Xg°lOUXlVd pun Xumoqoavlv d fo a4a!,la~t/7v ~a ans; "d- f

, @
o o @
30 40 50 60 70 0 10 20 0 10 20 30 40 0 10 20 30 40 50 0 10 20 0 10 I I I I I I I i a i J i i I
~ % % % % % % % % % % % % % % % % % % % % % % % % ~. % % % • % % % % % % % % % % % %' .%%
% % % % % % % % % "-. % % % ~ % % % % . . % % % % % % % % % . . . % % % % % % % % % • % • % % % • % t . t . t . t . t . l . # % % N l l l l l l l l l l l l l l l , t l l l l l l l l l i i l l l l l l I l l l t l l l l l l l l l l
::)i :

sp.~oooJ l s , ~ o u ! ( I "L "fi'!~I
. . . . . . . . ~ ~ . ~ - I ~ ' ~ - ~
iii i iii: i : i i :¸¸ > i i i i i ; ' l ii!:i:i iiii iii i i l ) L i L i L ; ; " ~ / '
q
~'~ ~i ~i~i~ ~ i!i !~'~i ! 'i~ ii~ii~i~ii~'~!i!~ !i~ ~ ii'~"iiiilt'~il ~'~''~'~ ~i '~ ~ ~'~ ~ ~ 6 ~ ~
~ i ~ i~ i~iiiii~ii !i~i!~i!iii~i~i~i!~i!!ii iiii!ii'~i!,il )i~{!ili~i:~: ~ ~
i~ i ¸ ~ ~i i i i :i i i
o£
:)
P
I
q !
f
I
I l l
0 8~ 6~ d 01, It, b
P ~ # ~ # # # # # # # f g / ~ # # ~ # #
x % % % * % % % % % % % % . r x x ~ ' t l t l l l t t l l l l t l l l t l t l % % % % ~ % % % % % % % % % % % % % % %
' t i t t l l t l I l t l t l t l t t
f i z ~ _1 - " I J .......... "t",'s't't'¢'~'¢'¢'t'¢'¢'t't's'~'~'/'t ~Ig~ I I I
, , , , , , , , , , , , , . . . . . , y v , - - , - - , - - " g g l g g l i l l l l g l l l g l l ~ % % % % % % % % % % % % % ~ % % % %
g l g l l l l l l g l g l l l l l l % % % % % % % % % % % % % % % % % % %
l ' g l l l l l g g l g g l l g l l ~
f / /
~9 6~ (~661) zg Y~°l°UlflVd puv ~fuvtoqoavlvd fo ataz.,ta~I/7V la ans "d-T


J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 63
rant early records of Globorotalia conomiozea (Zachariasse, 1975; J. Cravatte, unpubl, data). It is clear that this should not be considered the stratotype of the lower part of the Messinian, as already emphasized by Colalongo et al. (1979).
3. General results
The Tripoli section at Capodarso can be sub- divided into three parts as follows (from bottom to top): Part I: samples 1-17 (thickness 6.75 m); this
part is equally composed of diatomites and claystones;
Part 11: samples 18-31 (thickness 5 m); diatomites being dominant;
Part 111". samples 32-48 (thickness 7.25 m), con- sisting mainly of claystones interbedded in the uppermost 2 m with the Calcare di Base.
3.1. Part I
For foraminiferal studies, in each sample about 100-150 g of sediment were washed with water and hydrogen peroxide at 120 volumes on 250, 150 and 40 #m sieves. The specific distribution was studied on the three grain-size fractions. Countings of planktic and benthic species were carried out on 300-400 specimens from the >150 pm total residues (> 250 and 150-250 pm fractions). In the statistical analysis, some related taxa were grouped under one name: Globigerina bulloides percentages are the total of the sum of G. bulbosa, G. bulloides, G. falconensis, G. parabulloides, and G. praebul- loides; Orbulina bilobata, O. suturalis, and O. uni- versa are grouped under the name of O. universa.
Planktic foraminifera are very abundant in part (up to 21 species), with Orbulina spp. being pre- dominant (Figs. 5 and 6). Turborotalita multiloba, an indicator of the early Messinian in the Mediterranean (D'Onofrio et al., 1975), is present in sample 2. The number of planktic species decreases in some clayey samples (2, 9, 11, 13, 14) as does their frequency, in favour of the benthic species (mainly Bulirnina echinata) (Figs. 5 and 6). Bulirnina aculeata, a living species related to B. echinata and recorded in upper epibathyal associa-
tions, is also common in the lower epibathyal associations (from 500-700 to 1000-1300 m; Wright, 1978). In most of the samples, B. echinata, an index form of the Mediterranean early Messinian (Colalongo et al., 1979), is the dominant benthic species; it is very abundant in some upper clayey layers. Only into the lower samples, rare and small (40-150 #m) specimens of Bolivina arta, B. dentellata, B. pseudoplicata, Brizalina dilatata, B. spathulata, B. aculeata, Cassidulina crassa, C. laevigata, Cibicides lobatulus, C. refulgens, Cibicidoides cf. kullenbergi, Elphidiurn semistria- turn, E. complanaturn, E. rnacellum, Eponides tumidulus, Fissurina marginata, Gobobulimina ovata, Neoconorbina terquemi, Pleurostomella alternans, and Rectouvigerina gaudryinoides are present. Such paleobathymetry is in agreement with the high values of the "planktic/planktic+ benthic" ratio. An assessment of the paleobathy- metry is difficult, because of thermospheric condi- tions affecting the salinity and oxygenation in the basin. However, the lack of diversity of the benthic association, generally diversified or oligotypic in central areas and richer in marginal areas (van der Zwaan, 1982), seems to indicate a lower epibathyal environment.
The change in coiling (from sinistral to dextral) of Neogloboquadrina acostaensis (Iaccarino, 1985) has been recorded in sample 8 (Fig. 5).
Using Goodman's method (1979), the antago- nism between the dinocyst diversity index, H(s), and the dominance index led us to identify six dinocyst "communities" (Fig. 6). Such changes in dinocyst "communities" should be related to varia- tions of surface water ecological parameters. The ratio between the neritic taxa (mainly Lingulodinium rnachaerophorum and Oper- culodinium israelianum) and the oceanic taxa (mainly Impagidiniurn patulum and, tentatively, Operculodinium janduchenei), and the dinocyst concentrations (an average of 25,000 dinocysts/g dry sediment) (Figs. 6 and 7), corresponds to a hemipelagic environment. Samples 1 and 8 corre- spond to two water depth maxima. Samples 9-14 correlate with an early minimum; the correspond- ing environment could be at least the outer shelf, and this assumption is not contradicted by fora- miniferal data. Some reworked dinocysts (dated Paleogene [mainly] to Lower Miocene) in samples

64 J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
7 and 8 were found to correspond to a relative rise in sea level. This could indicate coastal erosion due to a transgression or to the presence of olistol- ithes (Broquet et al., 1981a).
Dinocysts and organic matter show positive correlations (Figs. 6 and 8). The components of the organic matter (van der Zwan, 1990), as observed in the palynological residue, are esti- mated in percentages:
- -amorphous elements (A.O.M.) directly inher- ited from phytoplankton (Raynaud et al., 1989);
--vegetal organic matter (V.O.M.) consisting of well preserved particles of wood, parenchyma, leaf cuticules, etc.;
- -black organic debris which is opaque and non-identifiable; this can be transported over long distances and is often associated with the rework- ing of ancient sediment during a sea-level fall.
I m
m
f
e
d
e ¸
b
I.
q - i S
- - m
E
E
~32
j A.O.M.
0 10 20 30 40 50 60 70% ' ' ' I I I I I
-22
- - l
- 16 ?
4
~ 7 •
~ f
Black debris V.O.M.
10 20 30 40 50 60 70 80% 0 10 20 30 40% i , , I I I I I I ~ I i
_ _ |
"-. I i
B .
Fig. 8. Organic matter records. A. O.M. = amorphous organic matter; E O. M. = vegetal organic matter.

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 65
Amorphous organic matter, of marine origin, commonly prevails in the lower part of the section, whereas vegetal organic matter, associated with land proximity, mostly dominates in the upper part (Fig. 8).
The pollen flora contains herbaceous halophytic plants (Amaranthaceae-Chenopodiaceae, Caryo- phyllaceae, Plumbaginaceae) which is commonly found in modern environments adjacent to salt- water marshes. These elements are mainly present in samples 2-4 and 11-17 (Fig. 6). Their increas- ing percentages denote coastal influence.
X-ray analyses of clay particles were conducted following the method described by Holtzapffel (1985). Clay minerals in Messinian sediments of Sicily are generally considered to be terrigenous particles eroded from the continental soil cover (Chamley, 1989; de Visser, 1991). On the other hand, authigenesis and erosion of ancient sedi- ments may have occurred locally (de Visser, 1991 ). Paleogeographic and structural reconstructions (Dercourt et al., 1986; Ricou et al., 1986) show that during the Late Miocene the Caltanissetta basin was located on the African margin, southeast of the subduction zone of the African plate below the Calabrian arc: it is likely that most clay miner- als were supplied from the emerged low-relief areas in Tunisia and Lybia. This is especially the case for smectite and palygorskite, which develop in poorly drained soils (with alternating wet and dry seasons) and evaporitic areas as well as continental calcareous crusts (Chamley, 1989). Chlorite, illite and randomly mixed layers are derived from parent rock and poorly developed soils, most probably located in steep relief areas of the European plate where tectonic activity increased the unstability of the drainage basins. Formation of kaolinite is favoured in warm climates where precipitation and drainage ensure an intensive leaching of the parent rock: these conditions are generally fulfilled in upstream parts of the drainage basins and moun- tain areas. The clay mineral association of Part I of the section is dominated by smectite (Figs. 9 and 10), indicating the importance of the African detrital supply in the Caltanissetta basin. This also suggests the occurrence of warm climates, with alternating wet and dry seasons, in the source areas (mostly on the African plate).
Gersonde and Schrader (1984) studied diatoms from the same part of the section, and were of the opinion that continuous open marine conditions had prevailed.
The assignment of the lowermost layers of Part I to a relatively high sea level contrasts with the interpretation of the uppermost layers of the underlying Terravecchia Formation, that were probably deposited during low sea level (Suc et al., 1991), as indicated by the very small concen- trations of dinocysts, the low percentages of amor- phous organic matter, an abundance of reworked organisms, the relatively high percentages of fern spores (increased fluvial transport), a large amount of immature foraminifera, the presence of algal colonies of Botryococcus and the appearance of red fluorescent organic matter (usually observed in modern salt-marshes; Poumot, 1984), and the continuous record of pyrite.
3.2. Part H
Foraminiferal associations are largely domi- nated by planktic species but the diversity decreases, especially in the clayey levels (3-4 species) (Figs. 5 and 6). The number of small specimens progressively increases upwards.
The opposition between dinocyst diversity and dominance indices disappears (Fig. 6). Nevertheless, the correlations between dinocyst concentrations (Fig. 7) and amorphous organic matter (Fig. 6) still support the autochtonous origin of the dinocysts. The increased abundance of neritic taxa (e.g. samples 20, 21, 23, 25) (Fig. 6) could correspond to a relatively slight lowering of the sea level. The presence of reworked dinocysts in samples 20-24 (Paleogene [mainly] to Lower Miocene) supports this interpretation (increasing coastal influence).
Halophytes are relatively poorly represented in the pollen flora of the upper Part II, indicating a weak coastal influence (Fig. 6).
Relatively low values of the smectite/illite and kaolinite/illite indices are associated with a some- what increased kaolinite/smectite index (Figs. 9 and 10). Intensified precipitation may have favoured the development of kaolinite soils in well- drained areas and concomitant erosion of sub-

66 J. -P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51 79
m
/ 0
47
46 45
- - ~ - 4 2 ~ q -41
- 4 0
- - m i - -
o34 o33 b32
10 20 30 40 50 60 70 80 90 100
' ' ' ' ' ' ' ' ' / t
;::i:!:i:~:i:
~ %
l m I a
g
f
e ~
C
b ~
14 ~13 - 1 2
9 8 7 6 5 4 3 2
- 1
Chlorite [----] Smectite ~ ] Illite ~ Kaolinite
Random ~=~ Palygorskite mixed-layers
Fig. 9. Clayey minera l records.
strates by run-off. Also, a lowering of the longitudi- nal profile of the drainage basins associated with a sea-level drop could have resulted in increased river discharge of terrigenous materials from upstream areas and substratums. A comparison with marine indicators (dinocysts) suggests that the mineralogical evolution in this part of the
section principally results from a lowering of sea level. Indeed, it is noteworthy that samples 29-33 indicate an interval, with the occurrence of reworked dinocysts (mainly Paleogene taxa).
The upper Part II is furthermore characterized by the decrease and then disappearance of oceanic diatoms (Gersonde and Schrader, 1984).

mi f e d c b
lmI
a
/ o
-48
- 47
i
-46
-45
-42
-33
.....
-32
[
-24
-18_
_ -1
7
-14
P -1
3 ,
-9
b
!,iiii:i
it -
2 l,
- I
.H
.
% C
aCO
3
10
20
30
40
50
60
-...,
Com
posi
tae
+ Ly
seum
G
ram
inae
7,0
o !
lO
Kao
lini
te
/ Il
lite
0 1
I ,,
I I
I ,2
o
Fig
. 10
. C
om
par
iso
n o
f so
me
reco
rds
conc
erni
ng t
he c
onti
nent
al e
nvir
onm
ent.
Kao
lim
te/S
mec
fite
S
m~t
ite/
Illi
te
1 2
01
23
4
L
7
i ~ ~
:~
:~
~ii
q~
,.q
L~

68 J.-P. Suc et al./Review of Palaeobotany and Palvnology 87 (1995) 51-79
3.3. PartlII Most of the samples have shown a very poor
diversity in foraminiferal fauna (mainly composed of benthic species, such as Bulimina echinata, asso- ciated with small planktic specimens) (Figs. 5 and 6). Samples 41-48 are sterile. Samples 38 and 39 are once again rich in planktic taxa, frequently including Orbulina suturalis, O. universa and Neogloboquadrina acostaensis. The association differs from that of Part II, in the fact that Globigerinoides obliquus obIiquus, Turborotalita multiloba, T. quinqueloba and Globigerinita uvula are missing. Such an impoverished assemblage denotes a confined environment with intermittent inflows of oceanic water (samples 35, and 38 39). Maximum salinity tolerated by the modern Orbulina universa, Globigerinoides saccuIifer and G. ruber (equivalent of the fossil Globigerinoides obliquus; Thunell, 1979) range from 46%o (O. uni- versa) to 49%o (G. ruber) (Hemleben et al., 1989). Such values should have been reached 5 m below the Calcare di Base. The proposed trend to con- finement is also supported by the brown colour of clays overlying the diatomite p and by the occur- rence of gypsum from sample 42 upwards.
The concentrations of neritic and oceanic dino- cyst taxa fluctuate in the same way. In addition, there is no conflict between the diversity and dominance indices; total dinocyst concentrations do not correlate with the abundance of amorphous organic matter (Figs. 6 and 7). This probably indicates the allochtonous origin of dinoflagellate cysts, probably associated with the influx of oceanic water masses, indicated by increasing con- centration of oceanic taxa. Samples 45-48 are devoid of dinocysts; their absence is not due to poor preservation, since pollen grains are present, but suggests that the oceanic influx might have been greatly reduced during the deposition of the Calcare di Base. One must bear in mind the monospecific flora with I. patulum, found in the first clayey level within the Calcare di Base, which indicates an ultimate oceanic water inflow (Suc et al., 1991).
This part of the section, therefore, expresses an irregular but continuous evolution towards a con- fined environment. This progression is very consis- tent with the evolution of halophyte frequency
(Fig. 6), which shows several peaks (sample 32, 29%; sample 34, 33%; sample 38, 38%) before culminating in sample 48 to 52%, which is higher than percentages observed in some modern Mediterranean environments (J.-P. Suc, unpubl. data):
halophytes reach 30% in a brackish water lagoon and 38% in a salt marsh of the Rh6ne delta;
--halophytes range from 4 to 12% in six recent coastal sediments (VICOMED cruise) from the south- west Mediterranean sea (Alboran sea, Algeria to Sicily).
Smectite noticeably increases in this part of the section, and is locally associated with significant occurrences of palygorskite (Figs. 9 and 10). This mineralogical evolution could indicate an augmen- tation in seasonality, marked by an increasing importance of the dry season. From this moment, clay mineral associations are principally controlled by continental climate. The trend towards more arid conditions increases in the upper part of the section, after a brief return towards a more humid environment (samples 43 and 45). This evolution ultimately led to clay associations containing smec- tire (mostly largely dominant) and associated paly- gorskite in the upper part of the Messinian (Chamley et al., 1978; Chamley and Robert, 1980).
Confinement is also the obvious solution of Gersonde and Schrader (1984), who observed the presence of only one, broadly tolerant to salinity, diatom species (Asterolampra acutiloba) in the second clay level within the Calcare di Base.
The progression towards confinement, therefore, was interrupted by several irregular sea-level rises. The lower part of the Tripoli Formation was deposited during a slight rise in sea level, being higher than during the deposition of the uppermost Terravecchia Formation (Suc et al., 1991); these results contradict the hypotheses of Rouchy (1981), McKenzie and Oberhfinsli (1985) and Thunell et al. (1987), who generally considered the Tripoli Formation as deposited during a low stage of sea level.
4. The diatomite-clay alternations
In order to explain the diatomite deposition, Gersonde (1978) evoked the occurrence of upwell-
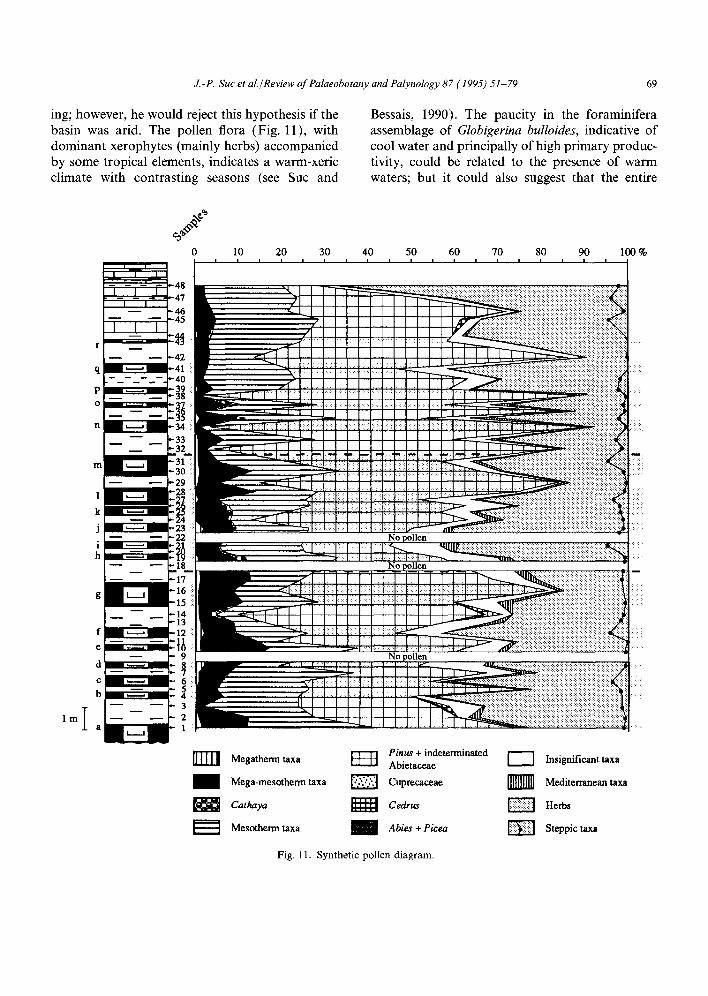
J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 69
ing; however, he would reject this hypothesis if the basin was arid. The pollen flora (Fig. 11), with dominant xerophytes (mainly herbs) accompanied by some tropical elements, indicates a warm-xeric climate with contrasting seasons (see Suc and
Bessais, 1990). The paucity in the foraminifera assemblage of Globigerina bulloides, indicative of cool water and principally of high primary produc- tivity, could be related to the presence of warm waters; but it could also suggest that the entire
J 0 10 20 30 40 50 60 70 80 90 100%
m
l m
f e
d
C
b
I, Megatherm taxa
Mega-mesotherm taxa
Cathaya
[ ~ Mesotherm taxa
~ ] Pinus + indeterminated Abietaceae
Cuprecaceae
C edr us
Abies + Picea
Fig. l 1. Synthetic pollen diagram.
- - ' - ] Insignificant taxa
Mediterranean taxa
Herbs
Steppic taxa

70 J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
productivity, even elevated in diatomites, did not reach the values of upwelling areas. The plank- tic foraminifera assemblage confirms the high temperature (tropical to subtropical) of the Mediterranean Sea surface water during the early Messinian (Catalano and Sprovieri, 1971; Hst~ et al., 1973; Iaccarino, 1981; Glagon et al., 1990). Only the interval between samples 14 and 18, from which Orbulina universa and Neogloboquadrina acostaensis (warm water forms) are missing and Globigerinita glutinata, Globigerina bulloides (opportunist forms presently living in cool waters; Hemleben et al., 1989) and G. praebulloides (Keller, 1985) dominate, could indicate a slight decrease in temperature or a change in circulation or availability of nutrients. The pollen diagram (Fig. 11) shows slightly increased percentages of Cathaya and Cedrus, respectively mid- and mid- high altitude trees, in samples 13-19. Taxodiaceae (Sequoia-type: a coastal to mid-altitude tree) also increased during this interval. This suggests moder- ate cooling, because of the continuous presence of megathermic plants. We conclude, therefore, that there is no suitable evidence for a dominant influ- ence of upwelling phenomena.
There are no data to be found in the pollen distribution (shown in Fig. 11) that support the theory of recurrent warmings in order to explain diatomite genesis, as proposed by Broquet et al. (1981b).
Since two of the four hypotheses (upwelling; warming of surface water; Atlantic water inflow; increase of run-off) generally considered to explain deposition of diatomites, have now been rejected, we will review the lithologic changes with special regard to information relevant to variations in sea level and/or continental run-off. Benson et al. (1991) explained the presence of diatomites and their alternation by periodic injections of high nutrient waters by a strong, eastward flowing, bottom current which started just before the depos- ition of the Tripoli Formation.
The lithologic variations generated by eustacy will be deduced from a comparison of data con- cerning foraminifera (Figs. 5 and 6), dinocyst concentrations (Fig. 7), organic matter (i.e. paly- nofacies; Fig. 6) and, additionally, halophyte pollen grains (Fig. 6).
Planktic foraminifera associations can be regu- larly subdivided in two groups:
(1) Orbulina universa, O. suturalis, Neo- globoquadrina acostaensis, Globigerinella obesa, Globigerina bulloides, G. praebulloides, Globigeri- noides obliquus obliquus and G. obliquus extremus are, especially in the middle-high part of the section, more abundant in the diatomites, indicating major primary productivity and more active circulation (McKenzie et al., 1979; Cita and McKenzie, 1986; Suc et al., 1991); Orbulina sutur- alis and O. universa seem to behave rather differently from the other species in relation with primary productivity or partly transport and gra- nulometric selection;
(2) Turborotalita multiloba and, secondarily, Globigerinita glutinata, G. uvula and T. quinqueloba reach their highest percentages in the claystone levels, also characterized by low organic produc- tivity and poorly diversified foraminifera associations.
The periodic changes in the distribution of dino- cysts cannot be linked to diatomite-clay alternations.
The influence of continental run-off on the litho- logic cyclothems will be estimated using the CaCO3 content and the Compositae+Lygeum/Graminae pollen ratio (Fig. 10).
In their study of the Lower Pliocene rhythmites of the Trubi Formation at Capo Rossello (Southern Sicily), de Visser et al. (1989) concluded that the occurrence of less calcareous layers is due to periodic increases of run-off (i.e. intensified precipitations) which may also increase the supply of continental organic matter.
The synthetic pollen diagram (Fig. 11) does not show any noticeable climatic change during the deposition of the diatomite layers. Nevertheless, the opposite pattern of the Compositae and Graminae curves is remarkable. Today, the Compositae/Graminae ratio is highly correlative with latitudinal change in Europe (P. Cour, pers. commun., 1993). Compositae predominate in Northern Africa, due to a dryer climate, while Graminae predominate in more humid regions. In the Capodarso section we tested the Compositae + Lygeum/Graminae ratio (Lygeum is a Mediterranean steppe Graminae living in warm

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51 79 71
and xeric conditions) in order to discern minor variations in climate (Fig. 10). High values of this ratio should indicate dryer conditions; low values should imply less xeric conditions and, therefore, increasing run-off.
The samples taken from the diatomite levels generally show higher values of both smectite/illite and kaolinite/illite indices than the samples from the marls (Fig. 10). This indicates a development of deeply weathered soils on the emerged continen- tal margins during deposition of the diatomites, probably as a consequence of improved climatic conditions (including precipitation on continental relief).
The diatomite-clay couplets will be described following the stratigraphic succession within each part of the section (part I: rhythmites a-g; part II: rhythmites h-m; part III: rhythmites n to r and limestone-clay alternations within the Calcare di Base Formation). A synthetic presentation has been choosen, which emphasizes the most relevant events and clarifies the interpretations. The latest are mainly referred to the above-mentioned argu- ments, that avoids some continuous repetitions.
on the whole increasing amorphous organic matter; Fig. 6), temporarily disturbed by the diato- mite layer c (reduced oceanic dinocysts and amor- phous organic matter; Fig. 6). The data indicate a moderate continuous continental run-off (low quantity of CaCO3 and domination of Graminae in samples 4 and 5; Fig. 10).
d The diatomite layer d is thought to be deposted
during a relatively high sea level (supremacy of planktic foraminifera and oceanic dinocysts, an increasing dinocyst concentration, importance of amorphous organic matter, scarcity of halophyte pollen grains; Fig. 6) during which the sedimenta- tion was also influenced by a moderate continental run-off (decreasing CaCO3, dominance of Graminae; Fig. I0). The overlying claystone (sample 9) seems to correspond to an obvious relative sea-level fall, based on decreasing planktic foraminifera, oceanic dinocysts and amorphous organic matter (Fig. 6). A continuously important continental run-off is supposed because of the lower quantity of CaCO3 (Fig. 10).
4.1. Couplets a-g
a
Diatomite a is considered to be deposited during a relative sea-level fall based on the low dinocyst concentrations (Figs. 6 and 7), and the poor amount of amorphous organic matter (Fig. 6). Nevertheless, the sea level was, as explained before, clearly higher than during the deposition of the uppermost clays of the Terravecchia Formation. The sea-level fall is mainly expressed in the clays of the couplet through the decrease in planktic foraminifera and the increase in halophyte pollen grains (Fig. 6). There is some evidence of efficient continental run-off in the lower part of the claystone (sample 2), based on the low percentages in CaCO3, and the dominance of Graminae (Fig. 10).
b and c These cycles are more problematic. For the two
sequences, the data suggest a relative sea-level rise (continuous dominance of planktic foraminifera,
e, f g These couplets are similarly interpreted as corre-
sponding to successive positive (diatomite layers) and negative (claystones) limited sea-level fluctua- tions. Such an interpretation is supported by the following, sometimes a little opposed, data. The planktic foraminifera are prevailing in the diato- mite layers and generally infrequent to scarce (except for sample 17) in the claystones (Fig. 6). The dinocyst concentration increases in diatomite layers e (neritic dinocysts), f (neritic and oceanic dinoxysts) and g (neritic dinocysts) (Fig. 6); a minor increasing concentration (both neritic and oceanic taxa) occurs in the claystone of cycle g (sample 17) (Fig. 6). The amorphous organic matter is on the whole abundant in the diatomite layers ( f a n d g: sample 15) but not in sample 10 (diatomite e) (Fig. 6). The halophyte pollen grains are regularly more abundant in the claystones (samples 11, 13, 14, 17) (Fig. 6). During this period, a maintained effect of a more or less important continental run-off is deduced from the low values of CaCO 3 and Compositae+

72 J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
Lygeum/Graminae ratio (samples 10, 12, 15) (Fig. 10).
Therefore, a detailed analysis of part I of the section reveals the prevalent control of sea-level fluctuations on the diatomite-clay alternations. Diatomite layers seem to be generally consistent with relative sea-level rises but some lithologic breaks probably do not correspond to environmen- tal transitions. In addition, a regular and more or less important continental run-off appears to have also influenced the lithologic changes.
4.2. Couplets h - m
k and l These couplets can be regarded as caused by
almost comparable limited sea-level fluctuations: the maxima occurred in diatomite k and upper diatomite l (importance of planktic foraminifera, higher concentrations in dinocysts, development of amorphous organic matter; Fig. 6) and the minima in samples 26 (claystone), 27 (lower diato- mite l) and 29 (claystone) (especially high quantity of halophyte pollen grains; Fig. 6). The low content in CaCO3 in diatomite l, associated to a moderate prevalence of Graminae (sample 27; Fig. 10), evokes some influence of continental run-off.
h and i These two diatomite layers and the intercalated
claystone seem to correspond to a relatively high sea level, taking into account the continuous high frequencies of planktic foraminifera and amor- phous organic matter (Fig. 6). Dinocyst records support this interpretation but the dominance of neritic taxa point to a lower sea level than in the early part I of the section (Figs. 6 and 7). The relatively low values of halophyte pollen grains (Fig. 6) are consistent as well. The uppermost claystone of these couplets (sample 22) is interpre- ted to be relatively low sea level because of the decrease in planktic foraminifera, dinocyst concen- trations and amorphous organic matter (Fig. 6). Continental run-off was not clear in diatomite layers h and i (relatively low frequency of CaCO3, predominant Graminae; Fig. 10).
J Diatomite j is interpreted as due to a slight sea-
level rise (large frequency of planktic foraminifera and increasing amorphous organic matter; Fig. 6). In contrast, the overlying claystone is considered to be corresponding to a slight sea-level fall (decrease in dinocyst concentration without change in the planktic foraminifer frequency, decreasing amorphous organic matter and higher percentage of halophyte pollen grains; Fig. 6). Continental run-off probably played an important role in diato- mite layer j, as suggested by very low values in CaCO3 associated to the prevalence of Graminae (Fig. 10).
m
Only the predominance of planktic foraminifera and the moderate concentrations of oceanic dino- cysts (Fig. 6) are in favour of a slight rise in sea level to explain this diatomite deposition. In con- trast, the lower part of the overlying claystone (sample 32) seems to correspond to a relevant relative sea-level fall (disappearance of planktic foraminifera, low value of amorphous organic matter, high frequency of halophyte pollen grains; Fig. 6) whereas sample 33 (upper part of the claystone) may announce a possible new rise in sea level (increase in amorphous organic matter, low frequency of halophyte pollen grains; Fig. 6). Continental run-off may have been effective during the beginning of diatomite deposition (sample 30: low values of CaCO3 and prevalence of Graminae; Fig. 10).
Except for the influence of continental run-off, the lithologic differences recorded in part II of the section seem to be mainly due primarily to sea- level changes. Nevertheless, this evolution must be understood in the context of a strong seaqevel fall. As shown for the previous period (part I), the lithologic subdivisions do not correlate constantly with the environmental changes.
4.3. Couplets n r
Data often appear contradictory when explain- ing diatomite origin in a recently confined basin.
We think that diatomites n and o originated during relatively low sea-level falls (disappearance of planktic foraminifera, disappearance and/or

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 73
paucity of dinocysts, strong increases in halophyte pollen grains, but also a large amount of amor- phous organic matter in sample 34; Fig. 6) whereas the immediately overlying clays suggest oceanic water inflows (increasing quantities of planktic foraminifera and dinocysts, large amount of amor- phous organic matter in sample 38, but also a high frequency of halophyte pollen grains in sample 38; Fig. 6). To explain these "atypic" diatomites, one can imagine the following process: the confined basin had such a reduced surface that the increas- ing river streams were able to reach its central area more easily. Diatomites n and o contain the largest percentages of palygorskite (Fig. 9) indicating the development of calcareous crusts and/or evapora- tive peri-marine environments in relation to an encroaching aridity on the emerged continental margins.
Diatomites p, q and r seem to be derived from inflows of oceanic waters related to minor sea- level rises. These inflows are shown by the abun- dance of poorly diversified foraminifera (Orbulina spp., Neogloboquadrina acostaensis, Globigerinella obesa and Globigerina spp. in diatomite p; Fig. 5) and by the presence of allochtonous dinoflagellate cysts (i.e. Batiacasphaera spp. in diatomite p [the ecology of this taxon is unknown]; Impagidinium patulum in diatomite r; Fig. 7). More humid climatic phases inducing continental run-off are indicated by the diatomite of sequence o and by the clay of sequence q (decreasing CaCO3 and prevalent Graminae; Fig. 10).
Another argument supports this point of view: the high percentages of Orbulina universa and Neogloboquadrina acostaensis (Fig. 5) suggest a short period of water inflow or a synsedimentary reworking (samples 38, 39). Thus, the diatomite deposition became less influenced by sea-level rises and/or run-off of climatic origin, but was almost exclusively controlled by the confinement of the basin. Indeed, large amounts of halophytes from sequence m (Fig. 6) suggest an increased influence of emerged continental areas.
4.4. The clay intercalations within the Caleare di Base
The clays appear to represent increasingly reduced oceanic inflows, as illustrated by our previ-
ous discovery (Suc et al., 1991) of large amounts of Impagidinium patulum in the lowest intercal- ation. The short incursions of marine waters in a more and more coastal environment (no foramini- fera, disappearance of dinocysts, high percentages of halophytes: Figs. 6 and 7; large quantities of black debris and vegetal organic matter: Fig. 8) are also attested by decreasing amorphous organic matter (samples 46-48; Fig. 6). The hypersaline marine waters in the Caltanissetta basin were so shallow that the oceanic influxes were unable to provoke anoxic conditions (Suc et al., 1991). Decima et al. (1988) concluded that the Calcare di Base deposition resulted from an intense sea- level lowering; temporary alterations of hyper- saline environments by influxes of marine and/or meteoric water were responsible for the precipita- tion of aragonite which later recrystallized into calcite.
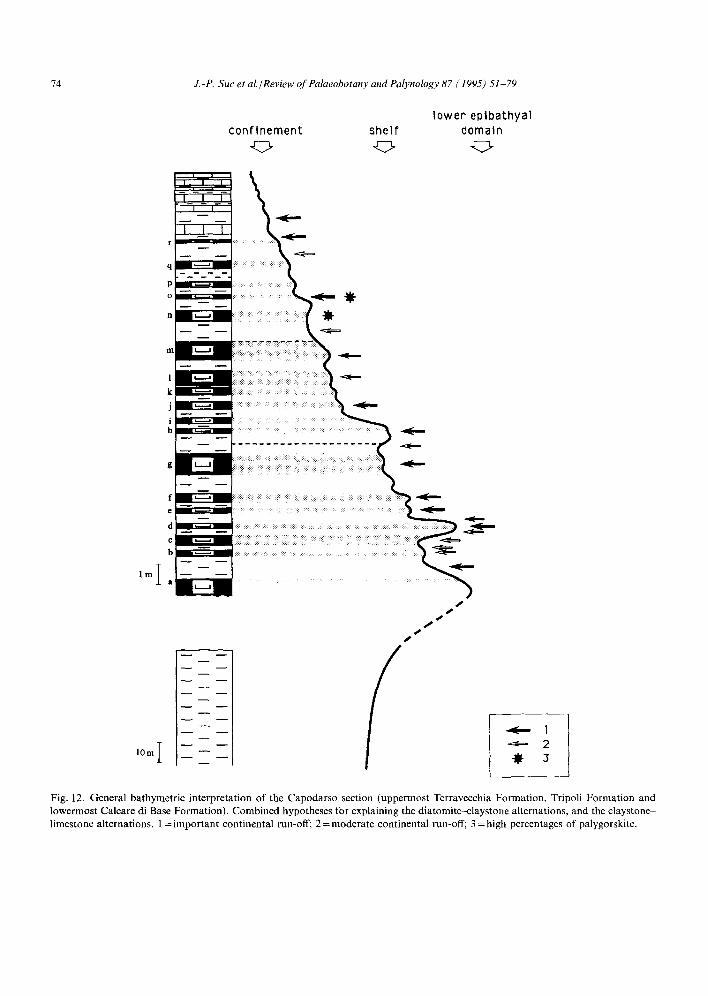
To summarize, three more or less combined types of processes could have generated the diato- mites (Fig. 12). In the lower part of the section, the sea level was relatively high and deposition of diatomites was aided by sea-level rises of unequal importance and variations of continental precipit- ations and run-off. Relatively minor sea-level oscil- lations prevailed then and also during the second part of the sequence, characterized by a generally strong decreasing bathymetry leading to basin confinement. The explanation for the diatomites during the last member now relies less on the occurrence of substantial sea-level rises in an increasingly desiccated basin; an increased number of rivers reached the central area of the basin, helping to produce the youngest diatomites. Fig. 12 shows the evolution of marine and continental environments during the deposition of the Tripoli Formation.
Van der Zwaan (1982) and Grasso et al. (1990) thought of increasing salinity as the main factor controlling foraminiferal distribution in diato- mites, whereas Sturani and Samp6 (1973) gave a greater importance to the weak ventilation and oxygenation of this moderatly eutrophic basin (Cita, 1982). The depth where the foraminifera species lived, their trophic requirements, and their strategy of life influenced the distribution of species more than any chemical-physical factors
74 J.-P. Suc et aL/Review of Palaeobotany and Palynology 87 (1995) 51 79
lower eplbathyal conf inement shel f domain
lm I
[ . . . .
10m I
, t s
s
S s
-a== 2 3
Fig. 12. General bathymetric interpretation of the Capodarso section (uppermost Terravecchia Formation, Tripoli Formation and lowermost Calcare di Base Formation). Combined hypotheses for explaining the diatomite-claystone alternations, and the claystone- limestone alternations. 1 = important continental run-off; 2 = moderate continental run-off; 3 = high percentages of palygorskite.

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 75
(Coulbourn et al., 1980; Hallock et al., 1991 ). The Capodarso microfauna is characterized by a low diversity and the presence of more or less oppor- tunist species living in surface waters (Globigerinita glutinata, G. uvula, Globigerinoides obliquus obliquus, and Turborotalita quinqueloba) and in intermediate waters (Globigerina bulloides, Orbulina universa, Neogloboquadrina acostaensis and probably Globigerinella obesa). Mesopelagic, oligotrophic and oligohaline, herbivorous (Globorotalia spp.) and carnivorous (Sphaeroidi- nellopsis spp.) forms are not present or very scarce. All these properties are typical of eutrophic unstable environments, subject to broad seasonal variations (also shown by continental records: pollen flora, clay minerals), but unable to promote a large variety of primary and secondary consumers (Hallock et al., 1991 ). The often contra- dictory behaviour of Orbulina spp. and Globigerinoides obliquus obliquus could be caused by competition, probably between two predatory taxa. The foraminifera distribution in claystones is chiefly influenced by a reduced availability of nutrients (for the planktic species) and bottom oxygenation (for the benthic species), probably accompanied by an increase in salinity. The micro- fauna was dominated by opportunist species able to proliferate in drastically confined environments. Turborotalita multiloba and Bulimina echinata appear to have been not only tolerant but also specific forms for such an environment: indeed, they have not been reported outside the Mediterranean.
5. Discussion
Suc et al. (1991) have shown differences between the underlying Terravecchia Formation claystones and the Tripoli Formation. These differences are larger than those existing between the clays and diatomite layers within the Tripoli Formation (at least in parts I and II of the Capodarso section). The sea-level rise, which occurred between depos- ition of the Terravecchia Formation and the Tripoli Formation, was of great amplitude. The deposition of the uppermost Terravecchia Formation clay- stones was already controlled by some confinement
(Suc et al., 1991) as represented by the presence of Botryococcus colonies, red fluorescence typical of modern salt-marshes (Poumot, 1984), numerous immature foraminifera and the occurrence of pyrite.
The diatomite-claystone couplets within the Tripoli Formation are generally interpreted as the result of sea-level fluctuations. The following pro- cess is proposed: warm (subtropical to tropical) and xeric climatic conditions (Suc and Bessais, 1990) induced a high salt-concentration of marine waters; the smallest influx of less saline (oceanic) waters and/or continental freshwaters (run-off) could produce water-mass stratification and diato- mite deposition. The process probably stopped when the influx of surface waters diminished, aiding the homogenisation of the water-mass due to intense evaporitic conditions. However, in con- trast to previous studies (unfortunately working with inadequate sampling; McKenzie et al., 1979; Suc et al., 1991), we think that not all diatomites can be considered as marine influxes and, con- versely, not all clays can be considered a result of sea-level drops. The exceptions are diatomites c, n and o and, partly, the clays of sequences b, c, f, g, h and n. It is important to note that the lithological difference between diatomite and clay is, in the field, sometimes very small.
Reworked materials (foraminifera, dinocysts, pollen grains) are frequent in the uppermost part of the Terravecchia Formation, but they are rare in the Tripoli Formation (Suc et al., 1991); varia- tions in their content is probably related to sea- level changes.
In addition, the physiography of the basin and tectonic activity may have played a significant role. For example, the Contrada Gaspa section, 12 km north of Capodarso (Fig. 1), where the apparent thickness of the Tripoli Formation reaches 108 m, is almost totally constituted by more or less dia- tomitic to clayey laminites (Grasso et al., 1990). The environmental information provided by fora- minifera is similar to that presented in this paper, but their relationship with sedimentological cycles is not clear. We envisage a small basin intermit- tently isolated from the Caltanissetta basin during the Tripoli sedimentation. This example empha- sizes the difficulty of correlating the Tripoli sections

76 J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
(especially those nearest to the edge of the Caltanissetta basin) without a detailed knowledge of the regional paleogeography and its evolution. In the same way, lithological relationships are hard to establish between the Capodarso and the Falconara sections, located in a more open marine context (Fig. 1). Only similarly detailed studies will provide suitable stratigraphic relationships between the sections. Such a program would deter- mine whether the secondary sea-level changes recorded at Capodarso have a local or more wide- spread significance.
Indeed, numerous climatic fluctuations (gen- erally associated to eustatic oscillations) have been recorded for the same period in the world ocean (McKenzie and Oberh~insli, 1985; Hodell et al., 1986; Hodell and Kennett, 1986; Keigwin et al., 1987; Aharon et al., 1993). The question is, are all the sea-level cycles registered at Capodarso related to global eustacy? Every sea-level rise at Capodarso is shown by the occurrence of oceanic taxa among foraminifera and/or dinocysts which attest, even for the latest incursions, water flows of Atlantic origin (Hodell et al., 1989; Benson and Rakic-E1 Bied, 1991; Benson et al., 1991). The correlation of each positive event at Capodarso to global eustacy is suggested. Thus, another question arises: was each sea-level drop at Capodarso caused by global eustacy, or only by strong but regional evaporitic conditions, maybe amplified by the paleogeographic isolation? It has been established that each outstanding sea-level rise in the Caltanissetta basin corresponds to an ocean high- stand, during which a maximum of oceanic water flowed through the Betic and Rifian corridors. The Central Mediterranean xeric climate rapidly con- centrated the Mediterranean waters, resulting in a fall in sea level. The progressive but irregular evolution of the Caltanissetta basin towards con- finement and desiccation can be thus explained. Accordingly, the curve of large amplitude sea-level changes deduced from the study of the Capodarso section correlates with the global eustatic curve.
Unattenuating the probable effects of continen- tal run-off on diatomite deposition, one could propose tentative relationships between the Capodarso sea-level curve and the very detailed climatic curve from the southwest Pacific DSDP
Site 588 (Hodell et al., 1986). The reversal deposits of the Tripoli Formation at Capodarso represent a fraction of the reverse event 3An.lr (= middle part of the Chron 5; Cande and Kent, 1992), due to the location of the change in coiling of Neogloboquadrina acostaensis that is in agreement with the data from the Bou Regreg section in Morocco (Benson and Rakic-E1 Bied, 1991). The overlying normal samples belong to the following normal event 3An.ln (=upper part of the Chron 5; Cande and Kent, 1992). At last, the couplet r and the Calcare di Base belong to the earliest Chron 3r (= the earliest Chron Gilbert; Cande and Kent, 1992). So, it is possible to correlate the relative highstand of sea level evidenced at Capodarso (diatomites a-d; Fig. 12) to the warm episode (i.e. a global sea-level rise) recorded at Site 588 within the reversed event of Chron 5 (Hodell et al., 1986). As a consequence, the following drop in sea level shown at Capodarso (Fig. 12) must be related to the Antarctic cooling (i.e. a global sea-level fall) recorded at Site 588 in the upper part of Chron 5 and the earliest Chron Gilbert (Hodell et al., 1986).
6. Conclusion
A detailed and multidisciplinary study of the Tripoli Formation at Capodarso has resulted in new information regarding the processes that led to the isolation and desiccation of the Caltanissetta basin. In the context of almost stable tropical- subtropical climatic conditions, a model is pro- posed where sea-level changes are the governing factor. These fluctuations of sea level correlate with global eustacy before the interruption of communications with the Atlantic Ocean.
The influence of other environmental factors (paleogeographic feature of the basin, continental run-off) has been of secondary importance. As confinement progressed, the physiography of the basin and the influence of the increasing fluvial effect, due to the closer proximity of continent, played a more important role.
The Tripoli Formation cannot yet be simply considered as an intermediate term between normal marine conditions and confinement. The

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 77
t r end to conf inement is a l r eady d o c u m e n t e d in the u p p e r m o s t Ter ravecchia F o r m a t i o n (Suc et al. , 1991) wi thou t p r o d u c i n g cyclic l i thologica l changes c o m p a r a b l e to those in the over ly ing Tr ipo l i F o r m a t i o n . But rhy thmi tes exist for exam- ple in the M o n t e G i a m m o i a sect ion in the mar l s under ly ing the Tr ipol i F o r m a t i o n ( M e u l e n k a m p et al. , 1979). The lack o f s imi lar a l t e rna t ions at C a p o d a r s o could be expla ined by active tectonics dur ing depos i t i on o f the Ter ravecchia clays, which d i s tu rbed the s ed imen ta t ion process. A n o t h e r i m p o r t a n t ques t ion remains unsolved: wha t deter- mined the passage f rom the rhy thmi tes o f the Ter ravecchia F o r m a t i o n to the cycles o f the Tr ipo l i F o r m a t i o n ?
In o rde r to e l abo ra t e a sui table s t r a t ig raphy and evo lu t ion o f the M e d i t e r r a n e a n env i ronments at the beg inn ing o f the Mess in ian sal in i ty crisis, sim- i lar mul t id i sc ip l inary studies should be conduc t ed on o ther sect ions f rom Sicily, as well as in the Betic and Rif ian cor r idors .
Acknowledgements
This p a p e r is con t r i bu t ion No. 648 o f the C N R S - I N S U " D y n a m i q u e et Bi lan de la Terre P r o g r a m " ( topic "Message s6dimenta i re" ) and pub l i ca t ion No. 94-092 o f the Ins t i tu t des Sciences de l 'Evo lu t ion ( U R A 327 C N R S ) . This s tudy was also s u p p o r t e d by D G I C Y T (agreement No. PB 90-0489) (J.-P. S.). Pa l aeomagne t i c measu remen t s have been car r ied ou t a t the L a b o r a t o i r e de Pa l6omagn6t i sme o f the Cent re G6olog ique et G6ophys ique o f Montpe l l i e r . The au tho r s are indeb ted to Dr. R i c h a r d H. Benson for Engl ish cor rec t ion o f the manusc r i p t and helpful scientific comments ; they also t hank the referees who pro- posed i m p o r t a n t improvemen t s o f the paper .
References
Aharon, P., Goldstein, S.L., Wheeler, C.W. and Jacobson, G., 1993. Sea-level events in the South Pacific linked with the Messinian salinity crisis. Geology, 21: 771-775.
Arambourg, C., 1925. Rdvision des poissons fossiles de Licata (Sicile). Ann. Pal6ontol., 14: 39-96.
Bennet, G.G., 1980. The Sedimentology, Diagenesis and Palaeoceanography of Diatomites from the Miocene of Sicily. Thesis. Univ. Durham, Durham, 223 pp.
Benson, R.H. and Rakic-E1 Bied, K., 1991. The Messinian parastratotype at Cuevas del Almanzora, Vera basin, SE Spain: refutation of the deep-basin shallow-water hypothesis? Micropaleontology, 37(3): 289-302.
Benson, R.H., Rakic-E1 Bied, K. and Bonaduce, G., 1991. An important current reversal (influx) in the Rifian corridor (Morocco) at the Tortonian-Messinian boundary: the end of the Tethys ocean. Paleoceanography, 6(1): 164-192.
Broquet, P., Du6e, G., Mascle, G. and Truillet, R., 1981a. Evolution structurale alpine r6cente de la Sicile et sa signification g6odynamique. Rev. G6ol. Dyn. G6ogr. Phys., 23(2): 75-85.
Broquet, P., Mascle, G. and Monnier, M., 1981b. La formation tripolis du bassin de Caltanissetta (Sicile). Rev. G6ol. Dyn.
G6ogr. Phys., 23(2): 87-98. Busson, G., 1990. Le Messinien de la M6diterran6e ... vingt
ans apr~s. G6ol. Fr., 3/4: 3-58. Cande, S.C. and Kent, D.V., 1992. A new geomagnetic polarity
time scale for the Late Cretaceous and Cenozoic. J. Geophys. Res., 97(B10): 13,917-13,951.
Catalano, R. and Sprovieri, R., 1971. Biostratigrafia di alcune serie saheliane (Messiniano inferiore) in Sicilia. Plankt. Conf., Rome, pp. 211-249.
Chamley, H. and Robert, C., 1980. S6dimentation argileuse au tertiaire sup~rieur dans le domaine m~diterran6en. G~ol. M6diterr., 7: 25-34.
Chamley, H., Dunoyer de Segonzac, G. and Meli~res, F., 1978. Clay minerals in Messinian sediments of the Mediterranean area. Init. Rep. Deep-Sea Drill. Proj., 42(1): 389-394.
Chamley, R., 1989. Clay Sedimentology. Springer, Heidelberg, 620 pp.
Cita, M.B., 1982. Impact of paleoceanographic events on the marine biotas of the Mediterranean in the least ten million years. Proc. 1st Symp. Paleontology, Essential of Historical Geology, pp. 123-149.
Cita, M.B. and McKenzie, J.A., 1986. The terminal Miocene event. Mesozoic and Cenozoic Oceans. Am. Geophys. Union Geodyn. Ser., 15: 123-140.
Colalongo, M.L., Di Grande, A., D'Onofrio, S., Gianelli, L., Iaccarino, S., Mazzei, R., Romeo, M. and Salvatorini, G., 1979. Stratigraphy of Late Miocene Italian sections straddling the Tortonian Messinian boundary. Boll. Soc. Paleontol. Ital., 18(2): 258-302.
Coulbourn, W.T., Parker, F.L. and Berger, W.H., 1980. Faunal and solution patterns of planktonic foraminifera in surface sediments of the North Pacific. Mar. Micropaleontol., 5: 329-399.
De Visser, J.P., 1991. Clay mineral stratigraphy of Miocene to recent marine sediments in the Central Mediterrranean. Geol. Ultraiectina, 75, 243 pp,
De Visser, J.P., Ebbing, J.H.J., Gudjonsson, L., Hilgen, F.J., Jorissen, F.J., Verhallen, P.J.J.M. and Zevenboom, D., 1989. The origin of rhythmic bedding in the Pliocene Trubi

78 .I.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79
Formation of Sicily, Southern Italy. Palaeogeogr. Palaeoclimatol. Palaeoecol., 69:45 66.
Decima, A., McKenzie, J.A. and Schreiber, B.C., 1988. The origin of "evaporative'limestones: an example from the Messinian of Sicily (Italy). J. Sediment. Petrol., 58(2): 256 272.
Dercourt, J., Zonenshain, L.P., Ricou, L.E., Kazmin, V.G., Le Pichon, X., Knipper, A.L., Grandjacquet, C., Sbortshikov, I.M., Geyssant, J., Lepvrier, C., Perchersky, D.H., Boulin, J., Sibuet, J.C., Savostin, L.A., Sorokhtin, O., Westphal, M., Bazhenov, M.L., Lauer, J.P. and Bij-Duval, B., 1986. Geological evolution of the Tethys belt from the Atlantic to the Pamirs since the Lias. Tectonophysics, 123:241 315.
D'Onofrio, S., Giannelli, L., Iaccarino, S. Morlotti, E., Romeo, M., Salvatorini, G., Sampo, M. and Sprovieri, R., 1975. Planktonic foraminifera of the Upper Miocene from some Italian sections and the problem of the lower boundary of the Messinian. Boll. Soc. Paleontol. Ital., 14(2): 177-196.
Gersonde, R., 1978. Diatoms paleoecology in the Mediterranean Messinian. 4th Messinian Seminar, Rome, Abstr., pp. 29 30.
Gersonde, R., 1980. Study of paleoecology and biostratigraphy of diatom assemblages from the lower Messinian "Tripoli" Formation in the Caltanissetta basin (Sicily). 6th Int. Syrup. Living and Fossil Diatoms, Budapest, Abstr., 2 pp.
Gersonde, R. and Schrader, H., 1984. Marine planktic diatom correlation of Lower Messinian deposits in the Western Mediterranean. Mar. Micropaleontol., 9: 93-110.
Gla~on, G., Vergnaud Grazzini, C., Iaccarino, S., Rehault, J.-P., Randrianasolo, A., Sierro, J.F., Weaver, P., Channell, J., Torii, M. and Hawthorne, T., 1990. Planktonic foramini- feral events and stable isotope records in the Upper Miocene. Proc. Ocean Drill. Prog. Sci. Results, 107: 415-427.
Goodman, D.K., 1979. Dinoflagellate "communities" from the Lower Eocene Nanjermoy Formation of Maryland, U.S.A. Palynology, 3: 169-190.
Grasso, M., Pedley, H.M. and Romeo, M., 1990. The Messinian Tripoli Formation of North-Central Sicily: paleo- environmental interpretation based on sedimentological, micropaleontological and regional tectonic studies. Paldobiol. Cont., 17:189 204.
Hallock, P., Premoli Silva, I. and Boersma, A., 1991. Similarities between planktonic and larger foraminiferal evolutionary trends through Paleogene paleoceanographic changes. Palaeogeogr. Palaeoclimatol. Palaeoecol., 83: 49-64.
Hemleben, C., Spindler, M. and Anderson, O.R., 1989. Modern planktonic Foraminifera. Springer, Berlin, 335 pp.
Hilgen, F.J., 1987. Sedimentary rhythms and high-resolution chronostratigraphic correlations in the Mediterranean Pliocene. Newslett. Stratigr., 17(2): 109-127.
Hodell, D.A. and Kennett, J.P., 1986. Late Miocene~arly Pliocene stratigraphy and paleoceanography of the south Atlantic and southwest Pacific Oceans: a synthesis. Paleoceanography, 1: 285-311.
Hodell, D.A., Elmstron, K.M. and Kennett, J.P., 1986. Latest
Miocene benthic 3180 changes, global ice volume, sea level and the "Messinian salinity crisis". Nature, 320:411 414.
Hodell, D.A., Benson, R.H., Kennett, J.P and Rakic-E1 Bied, K., 1989. Stable isotope stratigraphy of Late Miocene~Early Pliocene sequences in Northwest Morocco: the Bou Regreg section. Paleoceanography, 4(4): 467 482.
Holtzapffel, T., 1985. Les min6raux argileux. Pr6paration, analyse diffractom6trique et d6termination. Publ. Soc. G~ol. Nord, 12, 136pp.
Howell, M.W., Thunell, R.C., Tappa, E., Rio, D. and Sprovieri, R., 1988. Late Neogene laminated and opal-rich facies from the Mediterranean region: geochemical evidence for mechanisms of formation. Palaeogeogr. Palaeoclimatol. Palaeoecol., 64:265 286.
Hsia, K.J., Cita, M.B. and Ryan, W.B.F., 1973. The origin of the Mediterranean evaporites. Init. Rep. Deep-Sea Drill. Proj., 13: 1203-1231.
HsO, K.J., Montadert, L., Bernoulli, D,, Cita, M.B., Erickson, A., Garrison, R.E., Kidd, R.B., Meli6res, F., Muller, C. and Wright, R., 1978. History of the Mediterranean salinity crisis. Init. Rep. Deep-Sea Drill. Proj., 42: 1053-1078.
Iaccarino, S., 1981. Late Tortonian Messinian paleoenviron- ments. Ann. G6ol. Pays Hell6n., 4: 171-183.
Iaccarino, S., 1985. Mediterranean Miocene and Pliocene planktic foraminifera. In: H.M. Bolli, J.B. Saunders and K. Perch-Nielsen (Editors), Plankton Stratigraphy. Cambridge Univ. Press, Cambridge, pp. 283 314.
Keigwin, L.D., Aubry, M.-P. and Kent, D.V., 1987. North Atlantic Late Miocene stable isotope stratigraphy, biostratig- raphy, and magnetostratigraphy. Init. Rep. Deep-Sea Drill. Proj., 94:935 963.
Keller, G., 1985. Depth stratification of planktonic foraminifers in the Miocene ocean. Geol. Soc. Am. Mem., 163: 177-195.
Leonardi, A., 1959. L'ittiofauna del "Tripoli" del Miocene superiore di Bessima (Enna). Palaeontogr. Ital., 54:115 175.
McKenzie, J.A. and Oberhhnsli, H., 1985. Paleoceanographic expressions of the Messinian salinity crisis. In: K.J. HsO and H.J. Weissert (Editors), South Atlantic Paleoceanography. Cambridge Univ. Press, Cambridge, pp. 99-123.
McKenzie, J.A., Jenkyns, H.C. and Bennet, G.G., 1979. Stable isotope study of the cyclic diatomite-claystones from the Tripoli Formation, Sicily: a prelude to the Messinian salinity crisis. Palaeogeogr. Palaeoclimatol. Palaeoecol., 29:125 141.
Meulenkamp, J.E., Driever, B.W.M., Jonkers, H.A., Spaak, P., Zachariasse, W.J. and van der Zwaan, G.J., 1979. Late Miocene Pliocene climatic fluctuations and marine "cyclic" sedimentation patterns. Ann. G~ol. Pays. Helldn., 2:831-842.
Ogniben, L., 1957. Petrografia della Serie Solfifera siciliana e considerazioni geologiche relative. Mem. Descr. Carta Geol. Ital., 33, 275 pp.
Poumot, C., 1984. Sedimentation of the palynological organic matter in the solar salt works of Santa Pola (Spain). Relation with Salins de Giraud (France). Rev. Int. Geol., 38/39: 221 228.
Raynaud, J.-F., Lugardon, B. and Lacrampe-Couloume, G,, 1989. Structures lamellaires et bactdries, composants essen-

J.-P. Suc et al./Review of Palaeobotany and Palynology 87 (1995) 51-79 79
tiels de la mati6re organique amorphe des roches m+res. Bull. Cent. Rech. Explor.-Prod. Elf-Aquitaine, 13(1): 1-21.
Ricou, L.E., Dercourt, J., Geyssant, J., Grandjacquet, C., Lepvrier, C. and Bij-Duval, B., 1986. Geological constraints on the Alpine evolution of the Mediterranean Tethys. Tectonophysics, 123: 83-122.
Rouchy, J.-M., 1980. La g6n6se des 6vaporites messiniennes de M6diterran6e: un bilan. Bull. Cent. Rech. Explor.-Prod. Elf- Aquitaine, 4( 1 ): 511-545.
Rouchy, J.-M., 1981. La gen6se des 6vaporites messiniennes de M6diterran6e. Thesis. Univ. Paris 6, Paris, 293 pp.
Schrader, H.J., 1975. Correlation of the neostratotype of the Messinian with Pacific and Indian Ocean deep-sea drilling section based on marine planktonic diatoms. 6th RCMNS Congr., Bratislava, Abstr., pp. 403-405.
Selli, R., 1952. I1 bacino del Metauro. G. Geol., 24: 1-268. Selli, R., 1960. I1 Messiniano Mayer-Eymar 1867. Proposta di
un neostratotipo. G. Geol. (2), 28(1): 1-34. Sturani, C. and Samp6, M., 1973. I1 Messiniano inferiore in
facies diatomitica nel Bacino Terziario Piemontese. Mem. Soc. Geol. It., 12: 335-358.
Suc, J.-P. and Bessais, E., 1990. P6rennit6 d'un climat thermo- x6rique en Sicile avant, pendant, apr~s la crise de salinit6 messinienne. C.R. Acad. Sci. Paris (2), 310:1701 1707.
Suc, J.-P., Combourieu-Nebout, N., Robert, C., Poumot, C., Turon, J.-L. and Irr, F., 1991. Changements dans la s6dimentation argileuse au N6og~ne sup~rieur en M6diterran6e centrale: les Tripoli messiniens de Capodarso
(Sicile) et les laminites plio-pl6istoc~nes de Crotone (Calabre). Palynosciences, 1: 89-111.
Thunell, R.C., 1979. Pliocene Pleistocene paleotemperature and paleosalinity history of the Mediterranean Sea: results from DSDP Sites 125 and 132. Mar. Micropaleontol., 4: 173-188.
Thunell, R.C., Williams, D.F. and Belyea, P., 1984. Anoxic events in the Mediterranean Sea in relation to the evolution of late Neogene climates. Mar. Geol., 59: 105-134.
Thunell, R.C., Williams, D.F. and Howell, M., 1987. Atlantic- Mediterranean water exchange during the Late Neogene. Paleoceanography, 2(6): 661-678.
Van der Zwaan, G.J., 1982. Paleoecology of Late Miocene Mediterranean Foraminifera. Utrecht Micropaleontol. Bull., 25, 202 pp.
Van der Zwaan, G.J. and Gudjonsson, L., 1986. Middle Miocene-Pliocene stable isotope stratigraphy and paleocean- ography of the Mediterranean. Mar. Micropaleontol., 10: 71-90.
Van der Zwan, C.J., 1990. Palynostratigraphy and palynofacies reconstruction of the Upper Jurassic to lowermost Cretaceous of the Draugen Field, offshore mid Norway. Rev. Palaeobot. Palynol., 62:157 186.
Wright R., 1978. Neogene paleobathymetry of the Mediterranean based on benthic foraminifers from DSDP Leg 42A. Init. Rep. Deep-Sea Drill. Proj., 42A: 837-844.
Zachariasse W.J., 1975. Planktonic foraminiferal biostratigra- phy of the Late Neogene of Crete (Greece). Utrecht Micropaleontol. Bull., 11, 171 p.
Copyright © 2022 FDOKUMEN















![Travels of Ali Bey [pseud.] in Morocco, Tripoli, Cyprus, Egypt ...](https://static.fdokumen.com/doc/165x107/6333e93b3108fad7760f46d2/travels-of-ali-bey-pseud-in-morocco-tripoli-cyprus-egypt-.jpg)




