
Estimates of Natural Selection in a Salmon Population in Captive and Natural Environments
12
Contributed Paper Estimates of Natural Selection in a Salmon Population in Captive and Natural Environments MICHAEL J. FORD, ∗ § JEFFREY J. HARD, ∗ BRANT BOELTS,† ERIC LAHOOD, ∗ AND JASON MILLER ∗ ∗ Conservation Biology Division, Northwest Fisheries Science Center, 2725 Montlake Boulevard E. Seattle, WA 98112, U.S.A. †Washington Department of Fish and Wildlife, 600 Capital Way N, Olympia, WA 98501-1091, U.S.A. Abstract: Captive breeding is a commonly used strategy for species conservation. One risk of captive breeding is domestication selection—selection for traits that are advantageous in captivity but deleterious in the wild. Domestication selection is of particular concern for species that are bred in captivity for many generations and that have a high potential to interbreed with wild populations. Domestication is understood conceptually at a broad level, but relatively little is known about how natural selection differs empirically between wild and captive environments. We used genetic parentage analysis to measure natural selection on time of migration, weight, and morphology for a coho salmon (Oncorhynchus kisutch) population that was subdivided into captive and natural components. Our goal was to determine whether natural selection acting on the traits we measured differed significantly between the captive and natural environments. For males, larger individuals were favored in both the captive and natural environments in all years of the study, indicating that selection on these traits in captivity was similar to that in the wild. For females, selection on weight was significantly stronger in the natural environment than in the captive environment in 1 year and similar in the 2 environments in 2 other years. In both environments, there was evidence of selection for later time of return for both males and females. Selection on measured traits other than weight and run timing was relatively weak. Our results are a concrete example of how estimates of natural selection during captivity can be used to evaluate this common risk of captive breeding programs. Keywords: artificial propagation, captive breeding, domestication selection, hatchery, Oncorhynchus kisutch, parentage analysis Estimaciones de Selecci´ on Natural en una Poblaci´ on de Salm´ on en Ambientes Cautivos y Naturales Resumen: La reproducci´ on en cautiverio es una estrategia usada com´ unmente para la conservaci´ on de especies. Un riesgo de la reproducci´ on en cautiverio es la selecci´ on por domesticaci´ on—la selecci´ on de atrib- utos que son ventajosos en cautiverio pero delet´ ereos en el medio silvestre. La selecci´ on por domesticaci´ on es de inter´ es particular para especies que han sido reproducidas en cautiverio por muchas generaciones y que tienen un alto potencial de entrecruzarse con poblaciones silvestres. La domesticaci´ on es entendida conceptualmente a nivel general, pero se conoce poco sobre como difiere emp´ ıricamente la selecci´ on natural entre ambientes silvestres y cautivos. Utilizamos an´ alisis parental gen´etico para medir la selecci´ on natural sobre el tiempo de migraci´ on, peso y morfolog´ ıa de una poblaci´ on de salm´ on Oncorhynchus kisutch que fue subdividida en componentes cautivos y naturales. Nuestra meta fue determinar si la selecci´ on natu- ral que act´ ua sobre los atributos que medimos difiere significativamente entre los ambientes cautivos y naturales. Para machos, los individuos m´ as grandes fueron favorecidos tanto en los ambientes cautivos como en los naturales, en todos los a˜ nos del estudio, lo que indica que la selecci´ on de estos atributos en cautiverio era similar a la del medio silvestre. Para hembras, la selecci´ on de peso fue significativamente mayor en el ambiente natural que en el cautivo en 1 a˜ no y similar en los 2 ambientes en los otros dos a˜ nos. En ambos ambientes hubo evidencia de selecci´ on de un mayor tiempo de retorno tanto para machos §email [email protected] Paper submitted April 10, 2007; revised manuscript accepted December 17, 2007. 783 Conservation Biology, Volume 22, No. 3, 783–794 Journal compilation C 2008 Society for Conservation Biology. No claim to original US government works. DOI: 10.1111/j.1523-1739.2008.00965.x
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Estimates of Natural Selection in a Salmon Population in Captive and Natural Environments

Contributed Paper
Estimates of Natural Selection in a SalmonPopulation in Captive and Natural EnvironmentsMICHAEL J. FORD,∗§ JEFFREY J. HARD,∗ BRANT BOELTS,† ERIC LAHOOD,∗
AND JASON MILLER∗∗Conservation Biology Division, Northwest Fisheries Science Center, 2725 Montlake Boulevard E. Seattle,WA 98112, U.S.A.†Washington Department of Fish and Wildlife, 600 Capital Way N, Olympia, WA 98501-1091, U.S.A.
Abstract: Captive breeding is a commonly used strategy for species conservation. One risk of captive breeding
is domestication selection—selection for traits that are advantageous in captivity but deleterious in the wild.
Domestication selection is of particular concern for species that are bred in captivity for many generations
and that have a high potential to interbreed with wild populations. Domestication is understood conceptually
at a broad level, but relatively little is known about how natural selection differs empirically between wild
and captive environments. We used genetic parentage analysis to measure natural selection on time of
migration, weight, and morphology for a coho salmon (Oncorhynchus kisutch) population that was subdivided
into captive and natural components. Our goal was to determine whether natural selection acting on the
traits we measured differed significantly between the captive and natural environments. For males, larger
individuals were favored in both the captive and natural environments in all years of the study, indicating
that selection on these traits in captivity was similar to that in the wild. For females, selection on weight was
significantly stronger in the natural environment than in the captive environment in 1 year and similar in
the 2 environments in 2 other years. In both environments, there was evidence of selection for later time
of return for both males and females. Selection on measured traits other than weight and run timing was
relatively weak. Our results are a concrete example of how estimates of natural selection during captivity can
be used to evaluate this common risk of captive breeding programs.
Keywords: artificial propagation, captive breeding, domestication selection, hatchery, Oncorhynchus kisutch,parentage analysis
Estimaciones de Seleccion Natural en una Poblacion de Salmon en Ambientes Cautivos y Naturales
Resumen: La reproduccion en cautiverio es una estrategia usada comunmente para la conservacion de
especies. Un riesgo de la reproduccion en cautiverio es la seleccion por domesticacion—la seleccion de atrib-
utos que son ventajosos en cautiverio pero deletereos en el medio silvestre. La seleccion por domesticacion
es de interes particular para especies que han sido reproducidas en cautiverio por muchas generaciones
y que tienen un alto potencial de entrecruzarse con poblaciones silvestres. La domesticacion es entendida
conceptualmente a nivel general, pero se conoce poco sobre como difiere empıricamente la seleccion natural
entre ambientes silvestres y cautivos. Utilizamos analisis parental genetico para medir la seleccion natural
sobre el tiempo de migracion, peso y morfologıa de una poblacion de salmon Oncorhynchus kisutch que
fue subdividida en componentes cautivos y naturales. Nuestra meta fue determinar si la seleccion natu-
ral que actua sobre los atributos que medimos difiere significativamente entre los ambientes cautivos y
naturales. Para machos, los individuos mas grandes fueron favorecidos tanto en los ambientes cautivos
como en los naturales, en todos los anos del estudio, lo que indica que la seleccion de estos atributos en
cautiverio era similar a la del medio silvestre. Para hembras, la seleccion de peso fue significativamente
mayor en el ambiente natural que en el cautivo en 1 ano y similar en los 2 ambientes en los otros dos
anos. En ambos ambientes hubo evidencia de seleccion de un mayor tiempo de retorno tanto para machos
§email [email protected] submitted April 10, 2007; revised manuscript accepted December 17, 2007.
783Conservation Biology, Volume 22, No. 3, 783–794Journal compilation C©2008 Society for Conservation Biology. No claim to original US government works.DOI: 10.1111/j.1523-1739.2008.00965.x

784 Selection during Captivity
como hembras. La seleccion de los atributos medidos distintos al peso y el tiempo de corrida fue relativa-
mente debil. Nuestros resultados son un ejemplo concreto de como las estimaciones de seleccion natural
durante el cautiverio pueden ser utilizadas para evaluar este riesgo comun en los programas de reproduccion
en cautiverio.
Palabras Clave: analisis parental, criadero de peces, propagacion artificial, Oncorhynchus kisutch, repro-duccion en cautiverio, seleccion por domesticacion
Introduction
Understanding how populations evolve is becomingan increasingly important part of conservation biology.There are several reasons for this. Global warming andassociated climate change have brought attention to thefact that many species live in environments that arechanging rapidly. The ability of a species to adapt to thesechanges is an important factor in the extinction risk ofthat species (e.g., Lynch & Lande 1993; Lande & Shannon1996; Etterson & Shaw 2001). In addition to global warm-ing, most species face a myriad of environmental changesthat alter the selective landscape they experience, suchas land-use changes and invasions by non-native species.Another important form of environmental change occursduring captive breeding. Many species of conservationconcern are propagated artificially in an attempt to in-crease their abundance. Most captive breeding programsare not intended to be permanent substitutions for viablenatural populations, however, and seek to minimize theamount of evolution that occurs in captivity. Whether in-tentional or not, selection for traits that are advantageousor neutral in captivity but disadvantageous in the wild(domestication selection) is expected to be a particularproblem if a species is propagated for many generationsin captivity (e.g., Frankham et al. 1986; Snyder et al. 1996;Bryant & Reed 1999).
Seven species of Pacific salmon (Oncorhynchus spp.)spawn in rivers on the west coast of North America,and all contain populations that have declined to thepoint of conservation concern (Nehlsen et al. 1991).Twenty-seven evolutionarily significant units (ESUs)of Pacific salmon have been listed as threatened orendangered under the U.S. Endangered Species Act(http://www.nwr.noaa.gov/). Salmon populations havedeclined for several reasons common to other threat-ened species, including loss and degradation of habitatand overharvest (National Research Council 1996). An-other potential factor for the decline of wild salmon pop-ulations is the genetic and ecological effects of artificialpropagation. In much of their range, salmonids are ar-tificially propagated on a very large scale. For example,over 4 billion anadromous juvenile salmon are releasedannually into the North Pacific Ocean from hatcheriesin North America and Asia (Beamish et al. 1997). Similarhatchery programs and large-scale closed-pen fish farm-
ing operations exist for Atlantic salmon (Salmo salar) inthe North Atlantic Ocean and Baltic Sea.
Artificial propagation poses both ecological and ge-netic risks to salmonids (reviewed by National ResearchCouncil 1996; Waples & Drake 2004). For example, thepotential effects of supportive breeding on effective pop-ulation size have been modeled extensively (Ryman &Laikre 1991; Ryman et al. 1995; Wang & Ryman 2001).Domestication selection may be another particularly im-portant genetic risk because salmonids are often prop-agated artificially for many generations. Several investi-gators have found evidence for domestication in salmon(e.g., Reisenbichler & McIntyre 1977; Fleming & Gross1993; Araki et al. 2007a, 2007b), and evolutionary mod-els suggest that domestication has the potential to causedeclines in the fitness of natural populations that arepartially supported by artificial propagation (e.g., Lynch& O’Hely 2001; Ford 2002). The predictions of thesemodels are quite sensitive to their underlying parame-ters, however, including the strength and form of naturalselection that occurs in the captive and wild environ-ments. Obtaining an empirical understanding of how se-lection differs between captive and natural environmentsis therefore important for predicting the outcomes oflong-term supplementation programs (Hershberger et al.1990; Ford 2002; Araki et al. 2007a).
We quantified differences in how selection operates inthe wild and in captivity for a salmon population withcomponents that experienced either a wild or a cap-tive environment. Using genetic parentage assignmentmethods, we estimated the number of progeny producedby individual fish in either of 2 distinct environments, asalmon hatchery or a natural stream. Between the 2 envi-ronments, we then compared selection gradients (regres-sions of relative progeny number upon standardized traitvalue) for several traits, including time of return to thestream, weight, and body shape. We were particularly in-terested in determining whether optimum trait values dif-fered in wild and captive environments or whether selec-tion in captivity was “relaxed” (weaker) compared withwhat we observed in the wild. The natural populationwe studied had already experienced many generationsof artificial propagation, so the situation we analyzed isprobably more typical of a captive population readaptingto the wild environment than a wild population adaptingto a captive environment.
Conservation Biology
Volume 22, No. 3, 2008

Ford et al. 785
Methods
Study Site
Minter Creek is a small stream draining into HendersonBay in Puget Sound, Washington (U.S.A.; Fig. 1). Cohosalmon in Minter Creek have a life-history pattern typi-cal for the species at this stream’s latitude (Sandercock1991), with juveniles spending about 18 months in thestream prior to smoltification and migration to salt water.The fish then typically spend another 18 months at sea,returning to freshwater as 3-year-olds. In some years a siz-able fraction of the males return as 2-year-olds (“jacks”)after approximately 6 months at sea (Salo & Bayliff 1958;Sandercock 1991), and other male and female life-historypatterns also occasionally occur.
A hatchery operated by the Washington Departmentof Fish and Wildlife (WDFW) is located near the mouthof the creek. Coho salmon have been bred and releasedfrom the hatchery since 1938, when WDFW selected thestream as a study site representative of the hundreds ofsmall Pacific Northwest streams inhabited by coho (On-
corhynchus kisutch) and chum (O. keta) salmon (Salo &
Figure 1. Map of the Minter Creek, Washington, study
area showing the location of the Minter Creek
Hatchery (including the adult sampling weir) and the
juvenile (smolt) collections weirs.
Bayliff 1958). Production-scale (>1,000,000 smolts/year)coho salmon releases were initiated in 1960 (WDFW,Olympia, Washington). There have been no sustainedattempts to keep the hatchery and natural populationsisolated from each other, and in the years leading up toour study an unquantified (probably >90%) fraction ofthe natural spawning population consisted of hatchery-produced fish. The natural population of coho salmonin Minter Creek is therefore clearly not a “pristine” wildpopulation; rather, it is representative of many of thenatural salmon populations in the Pacific Northwest thathave experienced high levels of hatchery stocking formany decades. In a previous study (Ford et al. 2006), wefound no significant differences in fitness between hatch-ery and naturally produced fish in Minter Creek. There-fore, hatchery and natural-origin fish were combined inthe analyses reported here.
In 2000 through 2004 all coho salmon were inter-cepted at a weir located at the head end of tidewater(river kilometer 0.8) as they returned from the ocean tospawn, and a fraction were detained for sampling. Eachsampled fish was sexed, weighed, and photographed,and each fish had a small piece (<1 cm2) of caudal fintissue removed for subsequent DNA analysis. Fish wereallowed to recover in fresh flowing water before eitherbeing released upstream (all years) or into the hatcheryholding pond (2001 through 2003).
In addition to our study fish, each year several thou-sand hatchery-origin fish were passed directly into thehatchery holding pond without being sampled, but nounsampled fish were allowed to pass above the weir tospawn naturally. Wild fish were identified by the pres-ence of an adipose fin (hatchery coho salmon releasedin the Puget Sound area typically have their adipose finremoved prior to release) and by scale analysis (see Fordet al. 2006 for details). In 2001 between 26 Septemberand 31 October, every third sampled fish (whether ofwild or hatchery origin) was released into the hatcheryholding pond. Prior to 26 September and after 31 Oc-tober, all but 10 fish were passed upstream so that fishmanagement spawning goals for wild fish would be met.The mean run timing (day of return to the weir measuredas days after September 1) of males put into the pond wasa few days earlier than the mean run timing of the malesput into the stream (day 45.8 [SD 15.03] vs. day 48.3[18.8], p = 0.08, t test). The mean run timing of femalesput into the pond was approximately 5 days earlier thanthe mean of the females put into the stream (day 45.1 [SD13.6] vs. 51.6 [17.7], p < 0.001, t test). All sampled fishreleased into the holding pond were tagged with num-bered metal jaw tags so that they could be identified atthe time of spawning. We followed a similar protocol fordirecting fish into the stream and hatchery pond in 2002and 2003, but these fish were not included as part ofthe parentage analyses because their progeny were notgenotyped.
Conservation Biology
Volume 22, No. 3, 2008

786 Selection during Captivity
For the 2001 cohort spawning in the hatchery occurredon 6 days in November (5, 7, 14, 19, 20, and 27). Spawn-ing dates were determined by the hatchery staff and werethe same as for the general (nonexperimental) hatcherypopulation. We spawned the fish by randomly select-ing sexually mature, tagged males and females from thehatchery pond. Each female was spawned with one male,and >95% of the males were spawned with a single fe-male. We kept progeny from the sampled fish spawnedin the hatchery separate from the general hatchery pop-ulations until they were large enough to be sampled andtagged in summer of 2002. At that time all progeny fromsampled fish received a coded wire tag (CWT) implantedinto their snout so that they could be identified as ourstudy fish when they returned to the stream as adults in2004.
Wild juvenile progeny were sampled at the fry (firstsummer after emergence) and smolt (second spring/summer after emergence) stages between March 2001and June 2003 as described in Ford et al. (2006). Fry weresampled by electrofishing extensively throughout the wa-tershed, and smolts were sampled at weir traps through-out their outmigration period. These juvenile sampleswere a small fraction (1–5%) of the total juvenile pop-ulation. Adult progeny from the 2000 and 2001 broodswere sampled at the weir in the fall of 2003 and 2004,respectively. Progeny were assigned to parents with themaximum likelihood method implemented in the pro-gram FAMOZ (Gerber et al. 2003). See Ford et al. (2006)for a detailed description of the parentage analysis.
Morphological Analyses
We measured 6 morphological traits (Fig. 2) from the dig-ital images taken at the time of sampling. Each trait wasdigitized and measured to the nearest millimeter withthe computer program TPSDIG (Rohlf 2001). Weightswere converted to the linear scale of the other traits bycube-root transformation. To reduce allometric correla-tions among traits, we regressed each morphological traitother than weight on transformed weight with model I
Figure 2. Morphological traits that were studied for
their effects on relative fitness of coho salmon
spawning in Minter Creek. Fish image source:
National Oceanic and Atmospheric Administration
(NOAA) Photo Library.
regression and then used residuals from these regressionsas the trait values in further analyses. Some fish had miss-ing data because of occasional failures of the scale orcamera, especially for the 2000 sampling season. Timeof return was essentially independent of weight (correla-tion coefficients ranged from −0.11 to 0.14, dependingon the sex and cohort), so time of return was analyzedindependently.
Selection and Fitness Analyses
All statistical analyses were conducted with the generallinear model (GLM) function in the SYSTAT computerpackage (version 11, Systat Software, San Jose, Califor-nia). Traits were standardized within each sex and cohortby subtracting the mean and dividing by the standarddeviation. We made separate fitness estimates by count-ing progeny at fry, smolt, and adult life stages. Absolutefitness (progeny counts) within sexes, cohorts, and lo-cations (stream vs. hatchery) was converted to relativefitness by dividing by the mean fitness. In addition, weestimated the minimum number of unique mates an in-dividual had by summing the other unique parents ofthe progeny assigned to that individual regardless of lifestage. We estimated standardized linear and quadratic se-lection gradients (Lande & Arnold 1983) individually foreach trait with the following model: progeny = constant+ trait + (trait)2. We tested differences in linear selectionterms between the hatchery and natural environmentswith analysis of covariance with the model: progeny =constant + trait + location ∗ trait. No attempt was madeto correct p values for multiple tests, but results wereinterpreted with an appreciation that a large number oftests were conducted. We focused primarily on resultsthat were both highly significant at the level of individ-ual tests and that followed a consistent pattern for thedifferent life stages sampled.
As an additional measure of selection we comparedthe mean trait values of the group of parents inferred tohave produced offspring versus those inferred to haveproduced no sampled offspring. We calculated selectiondifferentials as the difference in trait means of the sam-pled population and the subset of individuals that wereinferred to produce offspring. For fish in the hatcherypond, we also estimated selection differentials indepen-dently from the parentage analysis by comparing the traitsof spawned fish as recorded in the hatchery records ver-sus all tagged fish put into the hatchery pond.
Results
All progeny assignments were the same as described inFord et al. (2006), with the exception of the adult off-spring from the parents spawned in the hatchery in 2001,which were conducted with the same methods but were
Conservation Biology
Volume 22, No. 3, 2008

Ford et al. 787
Table 1. Summary of parent and progeny sample sizes for genetic parentage analysis of Minter Creek coho salmon in stream and hatcheryenvironments.
Parents Offspring
Brood year, location females males (jacks)a fry smolts adults
2000, stream 422 473 (28) 534 801 924b
2001, stream 480 566 (8) 699 677 1683c
2001, hatchery pond 139 154 (1) 370 — 128
aMales includes all males (age 2 and 3 years). Numbers in parentheses are the total number of males that were 2 years old (jacks).bIncludes 901 3-year-old fish sampled in 2003 and 23 2-year-old fish sampled in 2002.cIncludes 1657 3-year-old fish sampled in 2004 and 26 2-year-old fish sampled in 2003.
not reported in the previous paper. Sample sizes for bothparents and progeny are in Table 1. For the 2000 brood95.4% of the potential parents sampled at the weir weregenotyped for parentage analyses. For the 2001 brood92.6% of the potential parents in the stream and 98.1% ofthe potential parents in the hatchery were genotyped forparentage analysis. The majority of the potential spawn-ers in each of the years were 3-year-old fish, with 2-year-olds making up only 6% of the males in 2000 and 1.4%of the males in 2001 (Table 1). Similarly, in 2002, 2003,and 2004, when offspring from the 2 parental years in thestudy returned to spawn, >97% of the returning adultswere 3-year-old fish. Depending on the life stage and yearsampled, 50–70% of the progeny were unambiguouslyassigned to a single pair of spawning parents.
In the stream environment significant selection onweight (size) was detected for both sexes and in bothyears (Table 2). Linear selection gradients for weightwere positive, indicating that for both males and fe-males larger individuals were more successful at produc-ing offspring (Fig. 3). For both sexes selection gradientswere generally similar among the different life stages ofprogeny that we assessed, although the smolt-stage as-sessments produced the largest selection gradients (Ta-ble 2). Selection on weight was similar between years.After taking into account the effects of size, there was nosignificant effect of male age on the number of offspringproduced (analysis of covariance, results not shown). Sig-nificant selection was also detected on run timing, aswas reported previously by Ford et al. (2006). The lin-ear terms were generally positive (indicating selectionfor later than average return time), and quadratic termswere generally negative, indicating stabilizing selectionagainst returning at the extremes of the run (Table 2).The absolute values of the selection gradients on run tim-ing differed slightly from the earlier study because in thisstudy fitness was standardized by dividing by the meanfitness (Lande & Arnold 1983), whereas the earlier es-timates were standardized by subtracting the mean anddividing by the standard deviation. For males significantlinear selection on caudal peduncle size was also detectedin the 2000 cohort but not the 2001 cohort. In 2001 sig-nificant quadratic selection was detected for males on
fin size, body depth, snout length, and caudal pedunclesize, but these results depended on a single outlying indi-vidual with high relative fitness that was unusually lightfor its size (data not shown). In general there were noclear patterns of selection on traits other than weight andrun timing after correcting for correlations with weight(Table 2).
For males selection in the hatchery pond was similar tothat observed in the stream. In particular, linear selectiongradients for weight were positive and highly significant(Table 2 & Fig. 3). Selection on weight in the hatcherypond was significantly stronger than selection on weightin the stream when progeny were counted as fry but indis-tinguishable from selection in the stream when progenywere counted at the smolt or adult life stages (Table 2).Selection on run timing was also positive and significantand not statistically distinguishable from selection on runtiming in the stream, indicating that, as in the stream envi-ronment, later-returning males produced more progenythan earlier-returning males (Table 2). In the pond, un-like the stream, significant selection was also detected ondorsal fin size, when selection was measured as numberof mates or progeny at the fry stage (Table 2).
For females in the hatchery pond, selection on weightin 2001 was weaker than was observed in the stream.In particular, linear selection gradients on weight werenot significantly greater than zero and were significantlyless than the selection gradients estimated for femalesin the stream environment for 2 of the 3 measures offitness (Table 2). Selection on run timing was similar tothat observed in the stream, with later-returning femalesproducing more progeny than earlier-returning ones, al-though the p values were generally less significant thanthey were for the stream coefficients (Table 2).
The selection differentials, which were based onlyon whether or not an individual was inferred to havespawned, were consistent with but smaller than the selec-tion gradients, which incorporated both spawning suc-cess and progeny number. Selection differentials for maleweight were significant in both environments (Table 3).For females the selection differential for weight in 2001was significant in the stream environment but not inthe pond environment, which was consistent with the
Conservation Biology
Volume 22, No. 3, 2008

788 Selection during Captivity
Tabl
e2.
Line
ar(B
)an
dqu
adra
tic(G
)re
gres
sion
coef
ficie
nts
desc
ribi
ngth
ere
latio
nshi
pbe
twee
nre
lativ
efit
ness
(num
ber
ofin
ferr
edm
ates
and
num
ber
ofsa
mpl
edfr
y,sm
olt,
and
adul
tpro
geny
per
indi
vidu
alst
anda
rdiz
edby
the
mea
nnu
mbe
rof
prog
eny
per
indi
vidu
al)
and
the
trai
tsth
atw
ere
mea
sure
don
coho
salm
onsa
mpl
edat
the
Min
ter
Cree
kw
eir.
Loca
tion
a
stre
am
pon
d
yea
r
20
00
20
01
20
01
Inte
ract
ion
sc
Lif
e
Tra
itSex
sta
ge
bB
GB
GB
Gp
yea
rp
loca
tion
Wei
ght
mal
esm
ates
0.50
(0.0
0)0.
09(0
.09)
0.35
(0.0
0)0.
07(0
.15)
0.27
(0.0
0)−0
.03
(0.5
2)0.
490.
32fr
y0.
67(0
.00)
0.16
(0.0
6)0.
26(0
.01)
0.11
(0.1
0)0.
89(0
.00)
−0.1
0(0
.61)
0.09
0.01
smo
lts
0.75
(0.0
0)0.
18(0
.04)
0.64
(0.0
0)0.
04(0
.69)
––
0.96
–ad
ult
s0.
67(0
.00)
0.18
(0.0
6)0.
36(0
.00)
0.06
(0.4
1)0.
26(0
.03)
0.00
(0.9
9)0.
090.
43fe
mal
esm
ates
0.34
(0.0
0)0.
08(0
.17)
0.50
(0.0
0)−0
.01
(0.9
1)0.
04(0
.48)
−0.0
2(0
.58)
0.16
0.00
fry
0.27
(0.1
2)0.
27(0
.02)
0.56
(0.0
0)0.
14(0
.11)
0.30
(0.1
4)−0
.32
(0.0
4)0.
080.
06sm
olt
s0.
50(0
.00)
0.03
(0.8
0)1.
19(0
.00)
0.47
(0.0
0)–
–0.
00ad
ult
s0.
35(0
.01)
0.08
(0.3
4)0.
62(0
.00)
−0.0
1(0
.93)
0.09
(0.3
4)−0
.03
(0.6
9)0.
100.
01A
rriv
alm
ales
mat
es0.
07(0
.35)
−0.2
0(0
.00)
0.10
(0.1
3)−0
.12
(0.0
4)0.
25(0
.00)
−0.0
3(0
.64)
0.47
0.13
tim
efr
y0.
26(0
.06)
−0.1
9(0
.12)
0.20
(0.0
3)0.
03(0
.72)
0.59
(0.0
1)−0
.17
(0.4
0)0.
980.
23sm
olt
s0.
14(0
.27)
−0.3
9(0
.00)
0.29
(0.0
4)−0
.02
(0.8
8)–
–0.
13–
adu
lts
0.15
(0.2
5)−0
.15
(0.1
9)0.
15(0
.09)
−0.1
3(0
.09)
0.16
(0.1
6)−0
.16
(0.1
2)0.
860.
62fe
mal
esm
ates
0.15
(0.0
3)−0
.28
(0.0
0)0.
27(0
.00)
−0.0
8(0
.22)
0.20
(0.0
0)−0
.05
(0.3
2)0.
230.
69fr
y0.
26(0
.05)
−0.1
3(0
.27)
0.27
(0.0
1)−0
.08
(0.3
2)0.
39(0
.08)
−0.1
3(0
.54)
0.98
0.57
smo
lts
0.06
(0.6
7)−0
.40
(0.0
0)0.
18(0
.28)
−0.1
7(0
.22)
––
0.56
–ad
ult
s0.
15(0
.17)
−0.2
5(0
.01)
0.21
(0.0
5)−0
.10
(0.2
5)0.
04(0
.73)
0.07
(0.4
6)0.
700.
86Le
ngt
hm
ales
mat
es−0
.15
(0.1
1)−0
.03
(0.5
2)0.
01(0
.86)
0.00
(0.9
9)0.
02(0
.79)
0.11
(0.0
3)0.
110.
80fr
y−0
.17
(0.2
7)−0
.03
(0.6
8)0.
01(0
.91)
0.02
(0.5
5)0.
02(0
.92)
0.09
(0.6
5)0.
230.
97sm
olt
s−0
.36
(0.0
3)0.
03(0
.70)
0.05
(0.7
3)0.
01(0
.84)
––
0.08
–ad
ult
s−0
.30
(0.0
9)−0
.03
(0.7
7)0.
05(0
.61)
−0.0
2(0
.62)
−0.0
7(0
.59)
0.05
(0.6
3)0.
050.
69fe
mal
esm
ates
−0.0
8(0
.39)
−0.0
1(0
.71)
−0.0
6(0
.50)
0.01
(0.8
8)0.
03(0
.53)
−0.0
1(0
.88)
0.84
0.63
fry
− 0.1
9(0
.30)
0.03
(0.7
1)−0
.08
(0.4
4)−0
.03
(0.6
1)0.
27(0
.16)
−0.0
7(0
.53)
0.65
0.13
smo
lts
−0.1
1(0
.50)
−0.0
3(0
.62)
−0.1
2(0
.51)
−0.0
3(0
.80)
––
0.99
–ad
ult
s0.
03(0
.79)
−0.0
2(0
.64)
0.01
(0.9
0)0.
00(0
.97)
0.02
(0.8
1)0.
09(0
.13)
0.95
0.84
Bo
dy
mal
esm
ates
0.19
(0.1
8)0.
10(0
.14)
0.09
(0.1
7)0.
06(0
.03)
−0.0
6(0
.38)
0.10
(0.0
6)0.
720.
14d
epth
fry
0.15
(0.5
4)0.
17(0
.16)
0.06
(0.5
4)0.
13(0
.00)
−0.1
1(0
.63)
0.30
(0.1
2)0.
380.
10sm
olt
s0.
36(0
.13)
0.21
(0.0
7)−0
.04
(0.8
1)0.
04(0
.44)
––
0.58
–ad
ult
s0.
30(0
.24)
0.18
(0.1
6)0.
04(0
.71)
0.02
(0.5
7)0.
07(0
.53)
0.12
(0.2
3)0.
680.
95fe
mal
esm
ates
−0.0
9(0
.44)
−0.0
2(0
.66)
−0.1
1(0
.19)
0.02
(0.5
5)−0
.13
(0.0
2)0.
04(0
.25)
0.71
0.77
fry
0.26
(0.3
4)0.
04(0
.65)
−0.0
9(0
.43)
0.02
(0.6
5)−0
.37
(0.0
7)0.
16(0
.18)
0.18
0.76
smo
lts
0.02
(0.9
3)−0
.01
(0.9
2)0.
17(0
.36)
0.09
(0.2
9)–
–0.
79–
adu
lts
−0.2
0(0
.25)
−0.0
3(0
.62)
−0.1
0(0
.38)
0.03
(0.5
2)−0
.04
(0.7
3)0.
08(0
.22)
0.80
0.80
con
tin
ued
Conservation Biology
Volume 22, No. 3, 2008

Ford et al. 789
Tabl
e2.
cont
inue
d.
Loca
tion
a
stre
am
pon
d
yea
r
20
00
20
01
20
01
Inte
ract
ion
sc
Lif
e
Tra
itSex
sta
ge
bB
GB
GB
Gp
yea
rp
loca
tion
Sno
ut
mal
esm
ates
0.21
(0.1
0)−0
.05
(0.5
3)0.
00(1
.00)
0.09
(0.0
0)0.
08(0
.24)
0.09
(0.0
3)0.
240.
68le
ngt
hfr
y0.
25(0
.25)
−0.0
4(0
.75)
0.21
(0.0
2)0.
23(0
.00)
0.31
(0.2
1)0.
35(0
.15)
0.77
0.04
smo
lts
0.35
(0.1
0)−0
.09
(0.4
8)0.
06(0
.67)
0.05
(0.5
1)–
–0.
38–
adu
lts
0.39
(0.0
9)−0
.02
(0.8
7)−0
.04
(0.6
7)0.
12(0
.01)
−0.0
8(0
.55)
0.09
(0.2
2)0.
070.
84fe
mal
esm
ates
0.01
(0.9
4)−0
.01
(0.8
8)−0
.02
(0.8
1)0.
07(0
.11)
−0.0
6(0
.29)
0.01
(0.8
6)0.
770.
89fr
y−0
.25
(0.3
3)0.
41(0
.01)
−0.0
7(0
.49)
0.20
(0.0
0)−0
.33
(0.1
2)−0
.09
(0.5
9)0.
880.
31sm
olt
s0.
03(0
.89)
0.08
(0.5
7)−0
.08
(0.6
5)0.
16(0
.13)
––
0.60
–ad
ult
s−0
.05
(0.7
7)−0
.02
(0.8
3)−0
.06
(0.5
6)0.
09(0
.16)
−0.0
6(0
.59)
0.08
(0.3
3)0.
880.
74A
nal
mal
esm
ates
−0.0
2(0
.91)
0.05
(0.5
0)0.
04(0
.51)
0.03
(0.2
9)0.
02(0
.77)
0.02
(0.7
2)0.
640.
81fi
nfr
y0.
07(0
.75)
0.13
(0.3
5)0.
17(0
.06)
0.14
(0.0
0)0.
00(1
.00)
−0.0
1(0
.97)
0.65
0.68
smo
lts
−0.0
6(0
.78)
−0.0
5(0
.74)
0.02
(0.8
7)0.
04(0
.59)
––
0.83
–ad
ult
s0.
10(0
.67)
0.06
(0.7
0)0.
07(0
.45)
0.07
(0.1
0)−0
.08
(0.5
0)0.
10(0
.19)
0.86
0.36
fem
ales
mat
es0.
01(0
.94)
0.01
(0.8
9)0.
10(0
.26)
−0.0
2(0
.65)
0.04
(0.5
5)0.
02(0
.59)
0.50
0.71
fry
−0.0
2(0
.94)
0.05
(0.7
5)0.
04(0
.69)
−0.0
2(0
.80)
0.14
(0.5
3)0.
02(0
.86)
0.71
0.87
smo
lts
−0.1
9(0
.42)
0.04
(0.7
8)−0
.05
(0.7
8)−0
.03
(0.8
0)–
–0.
56–
adu
lts
−0.2
7(0
.11)
−0.0
1(0
.92)
0.15
(0.1
8)−0
.07
(0.3
4)0.
05(0
.66)
0.11
(0.0
6)0.
030.
34D
ors
alm
ales
mat
es−0
.02
(0.8
7)0.
16(0
.09)
−0.0
1(0
.89)
0.15
(0.0
0)0.
16(0
.00)
0.04
(0.2
2)0.
990.
29fi
nfr
y0.
11(0
.60)
0.56
(0.0
0)0.
14(0
.14)
0.20
(0.0
0)0.
54(0
.00)
0.00
(0.9
8)0.
740.
05sm
olt
s−0
.09
(0.6
9)0.
07(0
.65)
−0.2
0(0
.18)
0.14
(0.1
3)–
–0.
70–
adu
lts
0.00
(1.0
0)0.
00(0
.98)
0.03
(0.7
5)0.
16(0
.01)
0.13
(0.1
5)−0
.02
(0.6
9)0.
770.
76fe
mal
esm
ates
0.06
(0.6
0)−0
.14
(0.0
2)0.
09(0
.26)
−0.0
3(0
.62)
−0.0
2(0
.74)
−0.0
2(0
.53)
0.36
0.74
fry
−0.2
1(0
.46)
−0.0
6(0
.65)
0.15
(0.1
7)0.
01(0
.85)
0.01
(0.9
5)−0
.04
(0.7
5)0.
090.
40sm
olt
s−0
.02
(0.9
3)−0
.15
(0.2
1)0.
08(0
.67)
−0.0
2(0
.88)
––
0.49
–ad
ult
s0.
12(0
.51)
−0.1
6(0
.07)
0.03
(0.8
0)−0
.01
(0.8
8)0.
03(0
.79)
−0.0
1(0
.89)
0.86
0.76
Cau
dal
mal
esm
ates
0.33
(0.0
1)−0
.03
(0.6
6)0.
02(0
.73)
0.09
(0.0
0)−0
.02
(0.7
5)0.
04(0
.25)
0.06
0.58
ped
.fr
y0.
42(0
.05)
−0.0
8(0
.50)
0.05
(0.6
4)0.
23(0
.00)
−0.0
6(0
.77)
0.08
(0.5
7)0.
260.
09sm
olt
s0.
57(0
.01)
0.09
(0.4
5)0.
13(0
.42)
−0.0
1(0
.90)
––
0.13
–ad
ult
s0.
75(0
.00)
0.12
(0.3
1)0.
03(0
.78)
0.12
(0.0
0)−0
.07
(0.4
8)0.
05(0
.44)
0.00
0.44
fem
ales
mat
es0.
05(0
.69)
0.02
(0.6
7)0.
00(0
.98)
−0.0
4(0
.35)
0.08
(0.1
3)0.
03(0
.43)
0.77
0.45
fry
−0.1
7(0
.54)
0.05
(0.5
9)0.
16(0
.14)
0.01
(0.8
5)0.
23(0
.24)
−0.0
3(0
.80)
0.13
0.91
smo
lts
−0.4
4(0
.06)
−0.0
3(0
.71)
0.06
(0.7
4)−0
.06
(0.5
6)–
–0.
13–
adu
lts
−0.0
1(0
.95)
0.03
(0.6
3)−0
.10
(0.3
5)−0
.09
(0.1
5)0.
18(0
.07)
0.10
(0.1
1)0.
680.
15
aLoca
tion
refe
rsto
wh
eth
er
the
sam
ple
dfi
shw
ere
allow
ed
tosp
aw
nn
atu
rally
inth
est
rea
m(s
tea
m)
or
were
pu
tin
toth
eh
atc
hery
pon
d(p
on
d).
Th
eB
an
dG
term
sre
fer
toth
eli
nea
ra
nd
qu
adra
tic
coeff
icie
nts
of
the
regre
ssio
n,re
spect
ively
:re
lati
ve
pro
gen
y=
con
sta
nt+
B(t
rait
)+
G(t
rait
)2.Sig
nif
ica
nce
test
sfo
rth
ere
gre
ssio
nco
eff
icie
nts
(pva
lues,
un
corr
ect
ed
for
mu
ltip
le
test
s)a
rein
pa
ren
these
s.bLif
est
age
refe
rsto
the
age
at
wh
ich
pro
gen
yw
ere
sam
ple
d.P
rogen
yw
ere
sam
ple
da
nd
ass
ign
ed
topa
ren
tsa
tth
efr
y(<
1yea
rold
),sm
olt
(appro
xim
ate
ly1
.5yea
rsold
),a
nd
adu
lt
(pri
ma
rily
3yea
rsold
)li
fest
ages.
Ma
tes
refe
rsto
the
min
imu
mest
ima
teof
the
nu
mber
of
ma
tes
on
the
ba
sis
of
all
the
sam
ple
dpro
gen
y.
cSig
nif
ica
nce
of
inte
ract
ion
term
sfo
ryea
ra
nd
loca
tion
,re
spect
ively
,by
ea
chm
ea
sure
dtr
ait
,fo
rli
nea
rte
rms
on
ly.
Conservation Biology
Volume 22, No. 3, 2008
790 Selection during Captivity
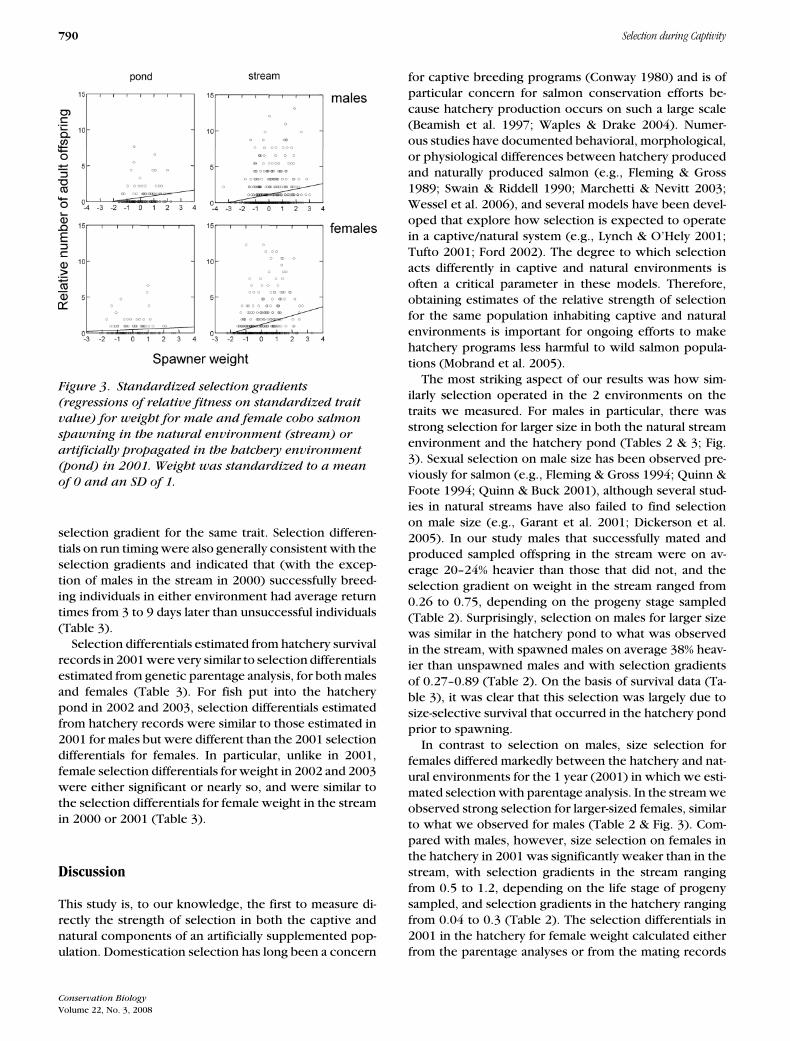
Figure 3. Standardized selection gradients
(regressions of relative fitness on standardized trait
value) for weight for male and female coho salmon
spawning in the natural environment (stream) or
artificially propagated in the hatchery environment
(pond) in 2001. Weight was standardized to a mean
of 0 and an SD of 1.
selection gradient for the same trait. Selection differen-tials on run timing were also generally consistent with theselection gradients and indicated that (with the excep-tion of males in the stream in 2000) successfully breed-ing individuals in either environment had average returntimes from 3 to 9 days later than unsuccessful individuals(Table 3).
Selection differentials estimated from hatchery survivalrecords in 2001 were very similar to selection differentialsestimated from genetic parentage analysis, for both malesand females (Table 3). For fish put into the hatcherypond in 2002 and 2003, selection differentials estimatedfrom hatchery records were similar to those estimated in2001 for males but were different than the 2001 selectiondifferentials for females. In particular, unlike in 2001,female selection differentials for weight in 2002 and 2003were either significant or nearly so, and were similar tothe selection differentials for female weight in the streamin 2000 or 2001 (Table 3).
Discussion
This study is, to our knowledge, the first to measure di-rectly the strength of selection in both the captive andnatural components of an artificially supplemented pop-ulation. Domestication selection has long been a concern
for captive breeding programs (Conway 1980) and is ofparticular concern for salmon conservation efforts be-cause hatchery production occurs on such a large scale(Beamish et al. 1997; Waples & Drake 2004). Numer-ous studies have documented behavioral, morphological,or physiological differences between hatchery producedand naturally produced salmon (e.g., Fleming & Gross1989; Swain & Riddell 1990; Marchetti & Nevitt 2003;Wessel et al. 2006), and several models have been devel-oped that explore how selection is expected to operatein a captive/natural system (e.g., Lynch & O’Hely 2001;Tufto 2001; Ford 2002). The degree to which selectionacts differently in captive and natural environments isoften a critical parameter in these models. Therefore,obtaining estimates of the relative strength of selectionfor the same population inhabiting captive and naturalenvironments is important for ongoing efforts to makehatchery programs less harmful to wild salmon popula-tions (Mobrand et al. 2005).
The most striking aspect of our results was how sim-ilarly selection operated in the 2 environments on thetraits we measured. For males in particular, there wasstrong selection for larger size in both the natural streamenvironment and the hatchery pond (Tables 2 & 3; Fig.3). Sexual selection on male size has been observed pre-viously for salmon (e.g., Fleming & Gross 1994; Quinn &Foote 1994; Quinn & Buck 2001), although several stud-ies in natural streams have also failed to find selectionon male size (e.g., Garant et al. 2001; Dickerson et al.2005). In our study males that successfully mated andproduced sampled offspring in the stream were on av-erage 20–24% heavier than those that did not, and theselection gradient on weight in the stream ranged from0.26 to 0.75, depending on the progeny stage sampled(Table 2). Surprisingly, selection on males for larger sizewas similar in the hatchery pond to what was observedin the stream, with spawned males on average 38% heav-ier than unspawned males and with selection gradientsof 0.27–0.89 (Table 2). On the basis of survival data (Ta-ble 3), it was clear that this selection was largely due tosize-selective survival that occurred in the hatchery pondprior to spawning.
In contrast to selection on males, size selection forfemales differed markedly between the hatchery and nat-ural environments for the 1 year (2001) in which we esti-mated selection with parentage analysis. In the stream weobserved strong selection for larger-sized females, similarto what we observed for males (Table 2 & Fig. 3). Com-pared with males, however, size selection on females inthe hatchery in 2001 was significantly weaker than in thestream, with selection gradients in the stream rangingfrom 0.5 to 1.2, depending on the life stage of progenysampled, and selection gradients in the hatchery rangingfrom 0.04 to 0.3 (Table 2). The selection differentials in2001 in the hatchery for female weight calculated eitherfrom the parentage analyses or from the mating records
Conservation Biology
Volume 22, No. 3, 2008

Ford et al. 791
Tabl
e3.
Infe
rred
and
estim
ated
mor
talit
yra
tes
and
sele
ctio
ndi
ffere
ntia
lsof
coho
salm
onfo
rw
eigh
tand
run
timin
gin
the
stre
aman
dha
tche
rypo
nden
viro
nmen
ts.
Mea
nru
nti
min
g,
Mea
nw
eig
ht,
g(S
D)
da
ys
aft
er
1Sep.(S
D)
Sa
mple
Mort
ali
ty
Sex
Yea
rLoca
tion
aM
eth
od
bsi
ze
(%)c
spa
wn
ed
not
spa
wn
ed
Dd
(p)
spa
wn
ed
not
spa
wn
ed
Dd
(p)
Mal
e20
00st
ream
gen
etic
449
6123
75(9
72)
1983
(929
)0.
26(0
.001
)47
.6(2
0.5)
49.2
(23.
5)−0
.04
(0.4
48)
2001
stre
amge
net
ic53
153
2593
(125
1)20
97(9
99)
0.23
(<0.
001)
50.1
(19.
4)46
.7(1
9.4)
0.10
(0.0
4)20
01p
on
dge
net
ic15
452
2778
(108
0)20
11(9
01)
0.36
(<0.
001)
50.3
(14.
6)41
.1(1
4.1)
0.25
(<0.
001)
2001
po
nd
reco
rd15
451
2817
(109
2)20
11(9
01)
0.39
(<0.
001)
46.1
(10.
6)40
.2(1
3.0)
0.20
(0.0
02)
2002
po
nd
reco
rd10
558
2543
(891
)22
23(9
57)
0.23
(0.0
9)50
.2(1
6.5)
42.7
(18.
5)0.
24(0
.033
)20
03p
on
dre
cord
244
6424
07(9
87)
1888
(965
)0.
36(<
0.00
1)39
.3(1
2.1)
32.2
(10.
4)0.
39(<
0.00
1)
Fem
ale
2000
stre
amge
net
ic40
756
2393
(692
)21
38(6
07)
0.20
(0.0
01)
55.5
(18.
0)50
.6(2
1.9)
0.13
(0.0
14)
2001
stre
amge
net
ic43
956
2518
(862
)22
15(8
53)
0.21
(<0.
001)
55.7
(16.
9)48
.5(1
7.8)
0.24
(<0.
001)
2001
po
nd
gen
etic
139
5023
18(8
09)
2244
(887
)0.
05(0
.61)
48.4
(14.
0)41
.3(1
2.1)
0.10
(0.0
01)
2001
po
nd
reco
rd13
950
2319
(810
)22
62(8
81)
0.03
(0.6
9)43
.7(8
.0)
40.4
(10.
4)0.
12(0
.036
)20
02p
on
dre
cord
8354
2587
(637
)23
13(6
92)
0.24
(0.0
7)55
.3(1
2.1)
42.6
(17.
1)0.
42(<
0.00
1)20
03p
on
dre
cord
208
6025
59(7
86)
1979
(932
)0.
45(<
0.00
1)43
.2(1
3.8)
35.1
(12.
0)0.
36(<
0.00
1)
aLoca
tion
refe
rsto
wh
eth
er
the
sam
ple
dfi
shw
ere
allow
ed
tosp
aw
nn
atu
rally
inM
inte
rC
reek
(str
ea
m)
or
were
pu
tin
toth
eh
atc
hery
pon
d(p
on
d).
bM
eth
od
refe
rsto
wh
eth
er
mem
bers
hip
of
the
spa
wn
ed
vers
us
un
spa
wn
ed
gro
ups
wa
sin
ferr
ed
on
the
ba
sis
of
resu
lts
of
the
gen
eti
cpa
ren
tage
an
aly
ses
(gen
eti
c)or
from
the
ha
tch
ery
ma
tin
g
reco
rds
(reco
rd).
cM
ort
ali
tyre
fers
toth
eest
ima
ted
pre
spa
wn
ing
mort
ali
tyra
te,ca
lcu
late
da
sth
en
um
ber
of
indiv
idu
als
kn
ow
nor
infe
rred
toh
ave
spa
wn
ed
div
ided
by
the
tota
ln
um
ber
of
indiv
idu
als
inth
e
sam
ple
gro
up
(row
).Th
epre
spa
wn
ing
mort
ali
tyra
tes
est
ima
ted
from
the
gen
eti
cpa
ren
tage
resu
lts
are
ma
xim
um
est
ima
tes
beca
use
itis
likely
tha
tso
me
indiv
idu
als
infa
ctsp
aw
ned
bu
tby
cha
nce
did
not
pro
du
cesa
mple
dpro
gen
y.
dSta
nda
rdiz
ed
sele
ctio
ndif
fere
nti
al(s
tan
da
rdiz
ed
dif
fere
nce
betw
een
the
tota
lpopu
lati
on
tra
itm
ea
na
nd
the
tra
itm
ea
nof
the
indiv
idu
als
kn
ow
nor
infe
rred
toh
ave
spa
wn
ed).
Th
ep
va
lues
were
calc
ula
ted
from
a2
-sa
mple
tte
stth
at
com
pa
red
the
dif
fere
nce
betw
een
the
spa
wn
ed
an
du
nsp
aw
ned
gro
ups.
Conservation Biology
Volume 22, No. 3, 2008

792 Selection during Captivity
were also much smaller than the selection differentialsestimated for females in the stream (Table 3). These re-sults suggest that selection for female size was relaxedin the hatchery in 2001 compared with selection in thenatural stream setting. This result was not consistent overtime, however, and in 2002 and 2003 selection on femaleweight in the hatchery was similar to that in the streamin 2001 (Table 3).
Conceptually there could be several mechanisms lead-ing to selection for larger size in the hatchery. Large sizemight be related to good general health, for example.Interactions among fish in the hatchery pond could alsolead to size selection. For example, the fish, particularlymales, in the hatchery pond may interact and fight witheach other in a manner similar to what they do in thestream, resulting in selection for larger size in the hatch-ery even though the hatchery staff does not actively selectfor large size when choosing fish to spawn.
The similar strength of selection in the 2 environmentsin these years does not imply that the selective mecha-nisms were necessarily the same in the 2 environments,however. For example, selection for larger female size inthe wild has been related to a variety of behavioral fac-tors that would have no opportunity to be expressed inthe Minter Creek Hatchery, such as ability to dig deepernests in the gravel, defend superior spawning locations,or attract more males (e.g., van den Berghe & Gross 1989;Fleming & Gross 1994; Quinn et al. 2001). Although wehave clearly documented that selection on size occursin both environments, we have no direct data on theselective mechanism in either environment.
Natural selection for run timing was qualitatively simi-lar for both sexes in both environments, although the sta-tistical significance varied considerably among compar-isons. In both environments there was some evidence forselection for later than average time of return, with suc-cessfully breeding individuals returning anywhere fromseveral days to about a week later than unsuccessful in-dividuals (Tables 2 & 3). These results should be inter-preted in light of the long-term changes that have beenobserved in the run timing of Minter Creek coho salmon(Ford et al. 2006). Both hatchery and natural-origin fishare returning to Minter Creek more than a month ear-lier on average than they did at the time when recordswere first kept in the 1940s (Salo & Bayliff 1958). Thelikely cause of this trend toward earlier time of return isprevious selection for earlier-spawning fish in the MinterCreek Hatchery.
We previously estimated selection gradients on runtiming for those salmon that spawned in the stream (Fordet al. 2006) and hypothesized that the selection for laterrun time indicated that the current, relatively early, runtime was evidence of hatchery-induced selection. It was,therefore, initially surprising to find that selection forlater return time was just as strong in the hatchery asit was in the wild. Nevertheless, for the last decade,
hatcheries throughout Washington have been subject toreview and reform efforts designed to make them morebenign to natural salmon populations (e.g., Mobrand et al.2005). As a result, at the time we initiated our study, theMinter Creek Hatchery began to deliberately reverse itsprevious spawning protocols in favor of protocols thatwould avoid selection for early-returning spawners (D.Popochock, personal communication). The finding of se-lection for later return time therefore appears consistentwith the current spawning protocols employed by thehatchery. In addition, the high level of prespawning mor-tality that occurred in all years of the study (Table 3) likelycontributed to selection for later return time simply be-cause few early-returning fish survived long enough to bespawned.
Consistent with an earlier study on coho salmon (Flem-ing & Gross 1994), we found little evidence for strongselection on the other morphological traits we measuredafter correcting for overall size (Table 2). An importantlimitation of our study, however, was that selection couldonly be estimated on the relatively small number of traitswe measured. Other traits, including behavior and phys-iology, were likely to have been under selection in bothcaptive and natural environments, but would have goneundetected in our study. In addition, in our study all traitswere measured at the weir, often on fish that had not yetbecome fully sexually mature. Many of the morpholog-ical traits we measured vary depending on the state ofsexual maturation (Sandercock 1991), and the measure-ments made at the weir likely differ somewhat from whatthey were at the time that selection actually occurred ineither environment.
The Minter Creek coho salmon population has somecharacteristics that should be taken into account whenconsidering the patterns of selection we observed. Forexample, the low proportion of jacks during the years ofour study may explain why we found no evidence of dis-ruptive selection on male size (Fig. 3), in contrast to someprevious studies (e.g., Gross 1985). The approximately50% mortality rate of our study fish in the hatchery wasalso much higher than the 1–10% that is more typical fora coho salmon hatchery (D. Popochock, personal com-munication), and the patterns of selection we observedin the hatchery may therefore not be typical of hatcherypopulations generally. In particular, relaxation of selec-tion, at least on traits associated with breeding success,may be more typical of hatcheries with very high pre-spawning survival rates. The selection we measured inthe stream setting may also be different from what mighthave been observed had the population not already beenaffected by several decades of hatchery stocking.
Despite these caveats, our results provide an impor-tant glimpse into how natural selection operates in cap-tive and natural environments. Contrary to our initialexpectations, we found that patterns of selection weresimilar between the hatchery and stream environments,
Conservation Biology
Volume 22, No. 3, 2008

Ford et al. 793
especially for males. The observation of similar outcomesof selection in the 2 environments suggests that selectionon these traits in the hatchery would not be expected tocreate a large selective load that contributes to poor pop-ulation fitness in the natural environment. The case forfemales is somewhat different, with evidence for relaxedselection on weight in the hatchery compared with thenatural stream, at least for 1 of the 3 years studied. Differ-ences in selection regimes between captive and naturalenvironments can lead to reduced fitness in the naturalcomponent of supplemented populations (Bryant & Reed1999; Lynch & O’Hely 2001; Ford 2002). It is not clear,however, whether the degree of relaxation we observedfor selection on female weight in captivity will producea significant selective load on the natural population inMinter Creek. Addressing the fitness consequences ofthe selection we observed will require additional anal-yses that take into account factors such as the geneticbasis of the trait and the rates of gene flow between thecaptive and natural components of the population.
Acknowledgments
We particularly acknowledge the Minter Creek hatcherycrew, D. Popochock, R. Henderson, E. Kinney, S. Davis,D. Watkins, and W. Campbell, whose kind assistance, pa-tience, and unfailing good humor were essential to thesuccess of this project. R. Waples, H. Araki, I. Fleming,M. Chilcote, P. Hulett, and L. Laikre provided useful com-ments on an earlier version of this paper. This projectwas funded in large part by a grant from the HatcheryScientific Review Group, whose assistance we gratefullyacknowledge.
Literature Cited
Araki, H., W. Ardren, E. Olsen, B. Cooper, and M. Blouin. 2007a. Re-productive success of captive-bred steelhead trout in the wild: eval-uation of three hatchery programs in the Hood River. ConservationBiology 21:181–190.
Araki, H., B. Cooper, and M. S. Blouin. 2007b. Genetic effects of cap-tive breeding cause a rapid, cumulative fitness decline in the wild.Science 318:100–103.
Beamish, R. J., C. Mahnken, and C. M. Neville. 1997. Hatchery andwild production of Pacific salmon in relation to large-scale, naturalshifts in the productivity of the marine environment. ICES Journalof Marine Science 54:963–964.
Bryant, E., and D. Reed. 1999. Fitness decline under relaxed selectionin captive populations. Conservation Biology 13:665–669.
Conway, W. G. 1980. An overview of captive propagation. Pages 199–208 in M. E. Soule and B. A. Wilcox, editors. Conservation biology:an evolutionary-ecological perspective. Sinauer Associates, Sunder-land, Massachusetts.
Dickerson, B., K. Brinck, M. Willson, P. Bentzen, and T. Quinn. 2005.Relative importance of salmon body size and arrival time at breedinggrounds to reproductive success. Ecology 86:347–353.
Etterson, J., and R. Shaw. 2001. Constraint to adaptive evolution inresponse to global warming. Science 294:151–154.
Fleming, I. A., and M. R. Gross. 1989. Evolution of adult female lifehistory and morphology in a Pacific salmon (coho: Oncorhynchus
kisutch). Evolution 43:141–157.Fleming, I. A., and M. R. Gross. 1993. Breeding success of hatchery and
wild coho salmon (Oncorhynchus kisutch) in competition. Ecolog-ical Applications 3:230–245.
Fleming, I. A., and M. R. Gross. 1994. Breeding competition in a PacificSalmon (coho: Oncorhynchus kisutch): measures of natural andsexual selection. Evolution 48:637–657.
Ford, M., H. Fuss, B. Boelts, E. LaHood, J. Hard, and J. Miller. 2006.Changes in run timing and natural smolt production in a naturallyspawning coho salmon (Oncorhynchus kisutch) population after60 years of intensive hatchery supplementation. Canadian Journalof Fisheries and Aquatic Sciences 63:2343–2355.
Ford, M. J. 2002. Selection in captivity during supportive breeding mayreduce fitness in the wild. Conservation Biology 16:815–825.
Frankham, R. H., H O. Ryder, E. Cothran, M. Soule, N. Murray, and M.Snyder. 1986. Selection in captive populations. Zoo Biology 5:127–138.
Garant, D., J. J. Dodson, and L. Bernatchez. 2001. A genetic evaluation ofmating system and determinants of individual reproductive successin Atlantic salmon (Salmo salar L.). The Journal of Heredity 92:137–145.
Gerber, S., P. Chabrier, and A. Kremer. 2003. FaMoz: a software forparentage analysis using dominant, codominant and uniparentallyinherited markers. Molecular Ecology Notes 3:479–481.
Gross, M. R. 1985. Disruptive selection for alternative life histories insalmon. Nature 313:47–48.
Hershberger, W., J. Myers, R. Iwamoto, W. Mcauley, and A. Sax-ton. 1990. Genetic changes in the growth of coho salmon (On-
corhynchus kisutch) in marine net-pens, produced by 10 years ofselection. Aquaculture 85:187–197.
Lande, R., and S. J. Arnold. 1983. The measurement of selection oncorrelated characters. Evolution 37:1210–1226.
Lande, R., and S. Shannon. 1996. The role of genetic variation in adap-tation and population persistence in a changing environment. Evo-lution 50:434–437.
Lynch, M., and R. Lande. 1993. Evolution and extinction in responseto environmental change. Sinauer Associates, Sunderland, Mas-sachusetts.
Lynch, M., and H. O’Hely. 2001. Captive breeding and the genetic fitnessof natural populations. Conservation Genetics 2:363–378.
Marchetti, M., and G. Nevitt. 2003. Effects of hatchery rearing on brainstructures of rainbow trout, Oncorhynchus mykiss. EnvironmentalBiology of Fishes 66:9–14.
Mobrand, L. E., J. Barr, L. Blankenship, D. E. Campton, T. T. P. Evelyn,T. A. Flagg, C. V. W. Mahnken, L. W. Seeb, P. R. Seidel, and W. W.Smoker. 2005. Hatchery reform in Washington State: principles andemerging issues. Fisheries 30:11–33.
National Research Council. 1996. Upstream: salmon and society in thePacific Northwest. National Academy Press, Washington, D.C.
Nehlsen, W., J. E. Williams, and J. A. Lichatowich. 1991. Pacific salmonat the crossroads: stocks at risk from California, Oregon, Idaho, andWashington. Fisheries 16:4–21.
Quinn, T. P., and G. B. Buck. 2001. Size- and sex-selective mortality ofadult sockeye salmon: bears, gulls, and fish out of water. Transac-tions of the American Fisheries Society 130:995–1005.
Quinn, T. P., and C. J. Foote. 1994. The effects of body sizeand sexual dimorphism on the reproductive behavior of sock-eye salmon, Oncorhynchus nerka. Animal Behavior 48:751–761.
Quinn, T. P., A. P. Hendry, and G. B. Buck. 2001. Balancing natural andsexual selection in sockeye salmon: interactions between body size,reproductive opportunity and vulnerability to predation by bears.Evolutionary Ecology Research 3:917–937.
Reisenbichler, R. R., and J. D. McIntyre. 1977. Genetic differences ingrowth and survival of juvenile hatchery and wild steelhead trout,
Conservation Biology
Volume 22, No. 3, 2008

794 Selection during Captivity
Salmo gairdneri. Journal of the Fisheries Research Board of Canada34:123–128.
Rohlf, F. J. 2001. TPSDIG Program for digitizing images for analysis bythin plate splines. State University of New York, Stony Brook, NewYork. Available from http://life.bio.sunysb.edu/morph/ (accessedMay, 2002).
Ryman, N., and L. Laikre. 1991. Effects of supportive breeding on thegenetically effective population size. Conservation Biology 5:325–329.
Ryman, N., P. E. Jorde, and L. Laikre. 1995. Supportive breeding andvariance effective population size. Conservation Biology 9:1619–1628.
Salo, E. O., and W. H. Bayliff. 1958. Artificial and natural production ofsilver salmon (Oncorhynchus kisutch) at Minter Creek, Washington.Washington Department of Fisheries, Olympia.
Sandercock, F. K. 1991. Life history of coho salmon (Oncorhynchus
kisutch). Pages 395–446 in C. Groot and L. Margolis, editors. Pa-cific salmon life histories. University of British Columbia Press,Vancouver.
Snyder, N., S. Derrickson, S. Beissinger, J. Wiley, T. Smith, W. Toone,and B. Miller. 1996. Limitations of captive breeding in endangeredspecies recovery. Conservation Biology 10:338–348.
Swain, D. P., and B. E. Riddell. 1990. Variation in agonistic behaviorbetween newly emerged juveniles from hatchery and wild popula-tions of coho salmon, Oncorhynchus kisutch. Canadian Journal ofFisheries and Aquatic Sciences 47:566–571.
Tufto, J. 2001. Effects of releasing maladapted individuals: ademographic-evolutionary model. The American Naturalist 158:331–340.
Van Den Berghe, E. P., and M. R. Gross. 1989. Natural selection result-ing from female breeding competition in a Pacific salmon (coho:Oncorhynchus kisutch). Evolution 43:125–140.
Wang, J., and N. Ryman. 2001. Genetic effects of multiple gen-erations of supportive breeding. Conservation Biology 16:1619–1631.
Waples, R. S., and J. Drake. 2004. Risk/benefit considerations for marinestock enhancement: a Pacific salmon perspective. Pages 260–306in L. M. Leber, S. Kitada, T. Svasand, and H. L. Blankenship, editors.Proceedings of the second international symposium on marine stockenhancement. Blackwell Publishing, Oxford, United Kingdom.
Wessel, M., W. Smoker, R. Fagen, and J. Joyce. 2006. Variation of ag-onistic behavior among juvenile Chinook salmon (Oncorhynchus
tshawytscha) of hatchery, hybrid, and wild origin. Canadian Jour-nal of Fisheries and Aquatic Sciences 63:438–447.
Conservation Biology
Volume 22, No. 3, 2008




















