
REACTIONS OF CAPTIVE HARBOR PORPOISES (PHOCOENA PHOCOENA) TO PINGER-LIKE SOUNDS
Upload
independentCategory
view
4download
0
Development of Infant Independence in a Captive Group of Lowland Gorillas
MICHAEL P. HOFF RONALD D. NADLER
TERRY L. MAPLE Departments of Psychology and Sociology
and the Yerkes Regional Primate Research Center Emory University Atlanta, Georgia
and School of Psychology Georgia Institute of Technology
Atlanta, Georgia
In March 1976,3 lowland gorillas (Gori//ugoril/ugori//u) were born to primiparous females living with an adult male in a large compound at the field station of the Yerkes Regional Primate Research Center of Emory University. Observations of parent and infant behavior began at the birth of the infants, using several methods of data collection. This report focuses on the development of independence in these infants over the 1st 1 % years of life. As expected, measures of mother-infant contact and proximity decreased with age. Several measures suggested that infant independence developed as an interactive process between mothers and infants, with primary responsibility changing over the months of study. Maternal behaviors that served to maintain mother-infant contact were found to decrease with age, with an eventual shift to infant responsibility for contact maintenance. Additionally, the adult male appeared to influence developing independence as reflected in the maternal protectiveness evoked by his behavior.
The development of independence by infant primates is a complex, multifacted phenomenon, characterized primarily by a decrease in mother-infant contact with increasing age of the infant. Among the nonhuman primates studied in this respect, the monkeys, especially the macaques, have received the most extensive attention. Hansen (1966), Harlow, Harlow, and Hansen (1963), Hinde and White (1974), Itoigawa (1973), Jensen, Bobbitt, and Gordon (1967), Kaufmann (1966), Kaufman and Rosenblum (1969), and Vessey (1968) reported similar sharp decreases in ventro- ventral contact over the first months of life in several macaque species. Also evident in these reports are an increasing time spent off the mother and an increasing distance an infant traveled from the mother with age. Included among the many variables identified by research on the macaques as associated with the development of independence are mother and infant departure (leaving) and approach (e.g., Hinde & Atkinson, 1970; Jensen et al., 1967), maternal protection (e.g., Hansen, 1966; Hinde & Spencer-Booth, 1967; Itoigawa, 1973), nipple contact (e.g., Harlow et al., 1963; Hinde
Reprint requests should be sent to Michael P. Hoff, Department of Psychology, Emory University, Atlanta, Georgia 30322, U.S.A.
Received for publication 28 March 1980 Revised for publication 21 August 1980 Deve/opmental Psychobiology, 14(3):251-265 (1981) 0 1981 by John Wiley & Sons, Inc. CCC 0012-1630/8 I /030251-15$0l.50

252 HOFF, NADLER, AND MAPLE
& Spencer-Booth, 1967; Jensen et al., 1967), maternal punishment (e.g., Hansen, 1966; Harlow et al., 1963; Hinde & White, 1974), weaning (e.g., Hinde & Spencer-Booth, 1967), other group members (e.g., Altmann, 1959; Hansen, 1966; Itoigawa, 1973; Kaufmann, 1966; White & Hinde, 1975), and environmental factors (e.g., Jensen et al., 1967, 1968a; Kaufmann, 1966). Several studies have recognized the importance of both the mother and infant in developing independence, although differential emphasis (e.g., Hinde, 1974, 1975; Kaufman & Rosenblum, 1969) has been given to the importance of each.
Reports concerning great ape infant development are generally less complete than those of monkeys, with only a few providing developmental data on mother-infant contact. Only fragmentary data are available regarding orang-utans (Pongopygmaeus). For example, Maple, Wilson, Zucker, and Wilson (1978) reported mother-infant contact first broken at 18 weeks in a captive pair living with an adult male. Kingsley (1977) observed this at 12 weeks. Additionally, she found that a great majority of mother-infant time was spent in ventral contact through 19 weeks. van Lawick- Goodall reported that mother-infant contact among feral chimpanzees (Pan Iroglodytes) was first broken at 14-22 weeks. Infants subsequently began moving away from their mothers to greater distances and for longer periods of time. Nicolson (1977), in a comparative study of captive and feral chimpanzees, found almost continuous ventro-ventral contact over the 1st 4 months of life in both feral and captive mother-infant dyads living in groups. Another captive pair in this study (Nicolson, 1977), living only with an adolescent male, exhibited a precipitous decrease in ventro-ventral contact over the same time period. A more extensive study of group-living, captive chimpanzees (Horvat, Coe, & Levine, in press) reported that mother-infant contact dropped from almost total contact in the 1st 6 months to approximately 30% of the observation time by 18 months. Concurrent with this decrease in contact was an increase in time that mothers and infants were out of contact but within 5 m of one another beginning at 6 months. Mother-infant distances greater than 5 m were occasionally observed during the 1st year, but not regularly observed until the beginning of the 2nd year.
Mother-infant relations in gorillas, as in the other great apes, are characterized by a long-term, intensive relationship early in life, gradually decreasing in intensity. This relationship lasts at least into the juvenile (4-6) years (see Fossey, 1979; Schaller, 1963), and probably beyond. However, even fewer data are available on infant development of gorillas than of chimpanzees. For wild mountain gorillas (Gorilla gorilla beringei), Schaller (1 963) reported that mother-infant contact was initially broken at about 3 months of age, maximal intradyad distance increased to approximately 3 m by 4 months, and increased further to about 6 m by 8 months. By 1 year, infants wandered away from their mothers and throughout the group. Fossey (1979) reported that mother-infant contact in mountain gorillas was not initially broken until approxi- mately the 5th month. By 1 year of age, infants and mothers were spending approximately 50% of observation time in contact. At this age, when off their mothers, infants were almost exclusively within 1 m. By 2 years, infants were in contact 30% of the time and within I m of their mothers 30% of the time. The same developmental trends of less contact and increasing distance from the mother, often beyond 5 m, were reported at 3 years.
A brief report (Lang, 1959) of the development of the 1st captive-born, mother- reared lowland gorilla (Gorilla gorilla gorilla) suggests a pattern similar to that reported by Schaller (1963) and Fossey (1979) for group-living, wild mountain gorillas. The male infant apparently began leaving its mother when it was 4-5 months

1NDEPENDENCE IN INFANT GORILLAS 253
of age. Weaning was described as gradual; the infant rarely nursed at 18 months and not at all by 2 years of age. A more detailed study of another captive lowland gorilla mother-infant dyad was reported recently by Kingsley (1977). Ventro-ventral contact was the most prevalent form of contact for the 1st 6 weeks of life, but had decreased to less than 30% of the observed time by 5 months of age, by which time the infant was off the mother but within arm’s reach of her about half the time.
These limited data on gorillas provide, at best, a rather tentative basis for generalizations about infant development in this species. The data available on group-living, wild infants, most appropriate for defining species-typical patterns, are limited overall and especially lacking in details. The data on captive individuals are also limited in terms of the number of subjects and the period of study, as well as in terms of the restricted nature of the physical and social environment. The present study was an attempt to supplement the data available on infant development of gorillas with data on a captive group of 3 mothers and infants living with an adult male in an outdoor compound. This approach, similar to that recently reported for chimpanzees by Horvat et al. (in press), has the advantage of permitting detailed observations characteristic of laboratory research while maintaining the animals under social conditions that more closely resemble the natural groupings of the species in the wild.
Methods
Subjects
The subjects were 3 wild-born, primiparous female lowland gorillas (Gorilla gorilla gorilla), their 1 female and 2 male captive-born infants, and 1 adult wild-born male, who probably sired all of the infants (see Table 1). All the adult gorillas were obtained from the wild as infants and had spent approximately 12 years in captivity as the study began.
Maintenance
Approximately 9 months before this study was initiated, the adult gorillas were transferred from cages at the Yerkes Regional Primate Research Center to a 75 X 100 ft (22.8 X 30.5 m) outdoor compound at the Yerkes Field Station. The history and housing ofthe group are described in detail elsewhere(Nadler, 1976; Tilford & Nadler, i 978).
TABLE 1. Subject Information.
Name Sex Born Parentage
Rann M 1963 Feral born Choornba F I963 Feral born Shamba F 1959 Feral born Segou F 1963 Feral born Machi F 1 March 1976 Rann or Calabar X Choomba Akbar M 9 March 1976 Rann or Calabar X Shamba Born-Born M 30 March 1976 Rann X Segou

254 HOFF, NADLER, AND MAPLE
Procedure
Observations were made in either early morning or afternoon, both ti,mes of maximal activity among gorillas in the field, according to Jones and Sabater Pi ( 1 97 1) and Schaller (1963). The period of study extended through the infants’ 1st 18 months of life, excluding a 6-week period ( 1 1 th lunar month) in which no observations were made due to extreme cold weather in late 1976 and early 1977. Two methods of data collection were utilized in the study. One-zero sampling using a 1-min interval was utilized in the 1st 3 and last 5 months. For the remainder of the study, a focal animal, continuous clock, sequential activity record was employed. Data were acquired by focusing on each mother-infant dyad for 30-min periods on each observation day. Daily order of observation was randomly determined.
During each test session, a variety of mother-infant and infant solitary behaviors exhibited by the focal dyad were recorded. In addition, focal infant-other mother, adult male-infant, and infant-infant behaviors were recorded. The infant member of each mother-infant dyad was the primary target. Thus, if the focal infant left its mother, it was followed. Although a multitude of behaviors was scored, only those related to developing infant independence are included herein. (See Table 2.)
Analysis
Data were tabulated on summary sheets and averaged over 4-week time periods, hereafter referred to as “lunar months” or “months.” Data collected utilizing the focal animal technique were tabulated as one-zero frequencies to maintain consistency across both methods of data collection. 1 The average number of 30-min observation sessions in each month was 27.3. Developmental profiles were plotted for the various
TABLE 2. Behavioral Definitions.
Venrro-ventral: Animals’ ventral surfaces in contact. Dorso-ventral: One animal’s ventrum pressed against another animal’s dorsum. Herein used exclusively in
Ventro-dorsal: As above. Herein used exclusively as infant’s dorsum against its mother’s ventrum. Orher contact: Any form of contact excluding ventro-ventral, dorso-ventral and ventro-dorsal. Proximate: Animals not in contact, but within 3 ft ( 1 m) (arm’s length) of one another. Disfant: Animals between 3 and 15 ft ( 1 and 5 m) apart. Far distant: Animals beyond 15 ft ( 5 m) apart. Leave: Moving from a position of contact to proximate, distant or far distant. Refuvn: Reestablishing contact with another animal. Scored for the animal reestablishing contact,
Social approach: Animal moving from distant or far distant to proximate (contact not established). FoNow: Moving behind and in the same direction as another animal. Restrain: Prevent another animal from moving away. Struxgle: Animal attempting to leave another while being restrained. Retrieval: Move another animal into closer spatial proximity as defined by the above contact variables. Nipple contact: Animal’s mouth placed on another’s nipple with or without suckling. Wean: Remove an infant from the nipple. Remove: Move another animal into a position of less close spatial proximity as defined by the above contact
Social examination: Inspect another animal with fingers, feet, and/ or mouth (includes stare). Object examination: Inspection and/ or manipulation of articles in the environment (does not include
PLotect: Saving an animal from possible harm (by retrieval, crouching over the animal, holding arm up,
“back ride” position (infant lying on mother’s back).
regardless of which animal left.
variables.
solitary play using objects).
etc.).

INDEPENDENCE IN INFANT GORILLAS 255
behaviors, using mean number of I-min intervals in which a given behavior was observed per 30 min of observation as the ordinate and infant age as the abscissa. Mean individual scores (rounded to the nearest unit) per month are presented in Table 3 to allow an examination of individual differences. Behaviors presented are those in which there was a difference between any 2 animals of 4 or more I-min intervals per 30-min test over 2 or more consecutive time blocks.
Results
Ventro-Ventral, Dorso-Ventral, Ventro-Dorsal, Other Contact
Through the 1st 6 months of life, mothers and infants quickly assumed less intimate positions as ventro-ventral contact decreased and was replaced by other
TABLE 3. Selected Behaviors (see text).
Mean Individual Scores (rounded to the nearest unit) Per Month for
Month
Behavior 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Ventro-ventral A292624 4 4 3 4 2 9 2 X 4 4 5 1 5 2 I 0 0 B30302511 710 7 4 1 X X 4 2 4 0 I I 2 2 X M302928302513 7 4 4 8 X 0 8 211 3 3 2 4 3
Dorso-ventral
Ventro-dorsal
A 0 0 0 0 5 5 4 7 712 X 4 5 4 3 2 2 2 2 3 B 0 0 0 0 0 0 0 1 1 X X 0 1 1 1 1 1 3 1 X M 0 0 0 0 0 0 1 1 3 4 X 5 0 3 0 3 2 4 9 3
A - - - 4 4 O I I I O X I O O O O O O O O B - - 3 9 9 5 2 4 9 X X 2 O O O O I O O X M - - - O O l l O O O X O O l O l l O O O
Other contact A 1 4 6151216181710 7 XI010 9 9 7 4 6 3 3 B 0 0 6141614141712 X X111112 7 8 6 8 5 X M 0 I 2 I 61817191513 XI3 912 7 9 9 9 1 1 1 0
Proximate
Distant
Far distant
Social Exam (Infant)
Object Exam
A 0 0 019131618151012 XI3 7 5 6 5 4 6 3 0 B 0 0 2 0 8101210 9 X X10 5 5 7 7 5 1 1 7 X
M 0 0 0 0 2 7161314 6 XI9 6141111 61013 9
A 0 0 0 0 5 8 4 12 8 12 X 19 9 14 10 15 15 13 14 12 B 0 0 0 0 4 6 8 914 X XI416 911 16221418 X M 0 0 0 0 0 1 2 5 5 8 X 31117121415151316
A - - - 0 0 0 0 0 0 0 X 0 0 7161020182023 B - - 0 0 0 0 0 0 0 X X 0 4131712 71218 X
M - - - 0 0 0 0 0 0 O X 0 0 5 2 8 1 0 9 3 5
A 0 0 1 1 4 5 7 5 3 3 X 4 3 5 4 4 5 5 5 3 B 0 0 2 6 6 6 2 4 2 X X 2 5 4 3 3 4 4 1 X
M 0 0 0 0 310 8 7 5 3 X 1 2 5 2 5 5 5 7 4
A - - - 4 I I 17 20 23 16 21 X 26 15 18 21 17 20 23 19 25 B - - 4 13 18 18 18 20 25 X X 18 20 18 22 20 21 22 20 X
M - - - 1 5 15 21 20 22 16 X 20 13 26 14 25 23 21 21 21
_ - - behavior not scored: X = no data taken: A = Akbar (male); B = Bom-Born (male); M = Machi (female).

256 HOFF, NADLER, AND MAPLE
22 \ v) _I
2 I8
2 8 f
W + 14
r 10
2 6
2
2 4 6 8 10 12 14 16 18 20 AGE (Lunar months)
Fig. I . Incidence of ventro-ventral ( O - O ) , ventro-dorsal (m-m), dorso-ventral(0- a), and other (e--e) contact as a function of infant age.
contact positions. Over the next several months, other forms of contact than ventro- ventral became increasingly common. Shortly before I year of age, all forms of contact began dropping off to rather low terminal levels. (See Fig. 1 .)
The incidence of ventro-ventral contact decreased from a high of 29.7 intervals per test in the 1st lunar month to a low of 1.5 intervals in the final month. An especially sharp decrease was evident during the 4th month, with a gradually slowing trend thereafter. By the 8th month, ventro-ventral contact occurred in 3.4 intervals and remained at or near this level throughout the remainder of the study.
The dorso-ventral “back-ride’’ position first occurred in the 5th month with a frequency of 1.6 intervals. Through the remainder of the study, the frequency was generally between 1.5 and 3.0 intervals, with a single peak of 8.0 in the 10th month.
The ventro-dorsal position was first observed and scored at a low level in the 3rd lunar month. It increased to the highest level-4.5 intervals-in the 5th month, decreased thereafter, and was rarely observed beyond the 9th month.
Other contact occurred in the 1st month with a very low incidence. There was a rapid rise in other contact concurrent with the drop in ventro-ventral contact in the4th month. A peak of 17.8 intervals per test was reached in the 8th lunar month; the occurrence thereafter gradually declined to 6.3 intervals at the end of the study.
Individual variation did exist in several of the contact behaviors, especially during the 1st year (see Table 3). The female infant, Machi, spent more of the 1st 6 months in ventro-ventral contact, and less in other contact positions than did the male infants. Also duringthefinal months, Machi was in the other contact position more than were the males. There were also differences between the 2 male infants at a number of points, but these were generally less striking than the male-female differences. The early predominance of dorso-ventral contact by Akbar and ventro-dorsal contact by Born-Born were largely related to mother-specific modes of transporting (back-ride and tripedal carry, respectively) infants (see Hoff, Nadler, & Maple, 1980).
Proximate, Distant, Far Distant
The mother-infant dyads maintained constant contact through the 1st 1 I weeks of life. At this time, contact was broken but very close separation distances were maintained. Through the remainder of the 1st year, the infants gradually ranged

INDEPENDENCE IN INFANT GORILLAS 257
Fig. 2. Incidence of proximate ("-0). distant (0-*), and far distant (0-0) as a function of infant age.
farther from their mothers, and were away longer, but still remained within 5 m. At about 1 year of age, infants and mothers began moving more than 5 m apart, and toward the end of the study were often seen across the compound from one another, that is, more than 5 m apart. Consistent with the previous contact behaviors, these also show the female, Machi, generally remaining closer to her mother (Table 3), whereas the male-male differences were less consistent.
The proximate position (Fig. 2) 1st occurred with a very low incidence in the 3rd month, with a sharp rise in the 4th, and a continuing increase to a high of 15.3 intervals per test in the 7th month. Thereafter, its occurrence generally declined to a final incidence of 4.5 intervals.
A mother and her infant were first observed to be distant from one another in the 16th week (Fig. 2). The increase in occurrence of this measure followed the pattern of the proximate position. The increase, however, was not as rapid as that of other contact or the proximate position, but peaked higher at 17.1 intervals per test during the 17th lunar month. The final frequency was 13.8 intervals during the 20th month.
No instance of mother and infant being beyond 5 m apart (far distant) was observed until the 13th month (Fig. 2). From a very low rate of 1.4 intervals, the frequency rapidly increased in the next month to 8.6, and continued increasing through the remainder of the study to a frequency of 14.1 intervals in the final month.
Leave, Return
Both the mothers and infants began leaving each other in the 4th month (Fig. 3). However, a great difference existed between them in the frequency of occurrence of this behavior. The infants started and maintained a much greater rate of leaving than did their mothers. The mean number of intervals in which infants left their mothers increased from 1.1 per test initially to a high of 5.9 in the 16th month, and dropped to a final incidence of 3.5 intervals. The mothers,however, initially left their infants in only . 1 interval per test, this continually increasing to a high of 2.9 in the 20th lunar month.
Both mothers and infants exhibited a pattern of returning to the other similar to that of leaving(Fig. 3). Mothers first began returning to their infants in the 5th month and reached a high of 1.3 intervals per test in the 9th month. The incidence dropped to .4 interval in the 12th month, remaining near that level for the remainder of the study. Infants returned to mothers initially in the 4th month with an incidence of .6 interval.

258 HOFF, NADLER, AND MAPLE
7 rn W t-
2 6 z 0
P 5
g 4 (0
a
W t-
w + 3 2 3
$ 2
H I Return to M W M Relurn to I
2 4 6 8 I0 12 14 16 18 20 AGE (Lunar months)
Fig. 3. Incidence of leave and return as a function of infant age.
Infant returns rapidly increased between the 9th and 10th months, to a high of 6.8 intervals in the 10th month, thereafter decreasing to 3.2 intervals in the final month.
Social Approach, Follow
Throughout the study, social approach, moving from the distant or far distant to the proximate position, was typically less frequent for the mothers than for the infants. In fact, this behavior was relatively uncommon in the mothers and was maintained at about the same level from its inception through the entire 18 months.
Maternal social approach toward infants began in the 5th month with an incidence of .3 interval per test and fluctuated at low levels throughout the remainder of the study. Infant social approach also began in the 5th month, with an incidence of .2 interval. It increased slowly to a high of 2.1 in the 18th month, dropping thereafter to a terminal frequency of .8 interval.
Following was observed in both mothers and infants beginning in the 5th month. Mothers did not regularly follow infants; this behavior was observed infrequently only in the 5th-8th, and 16th-19th lunar months. It began at .3 interval per test and never rose to greater than .6 interval throughout the study. Infants followed mothers more regularly and the development of this behavior paralleled that of maternal leave (see Figure 3), but at slightly lower levels. Infant following began in the 5th month at . l interval, and gradually increased to a high of 1.4 intervals in the final lunar month.
Restrain, Struggle, Retrieve
Both maternal restraint and the infant response of struggling (Fig. 4) were short-lived behaviors, peaking at relatively high levels early in life. They were rarely observed thereafter. These behaviors appeared at about the same time that infants were moving from ventro-ventral to less intimate contact positions (Fig. 1). This is also the same time that mothers and infants were first observed proximate (Fig. 2), and when infants first began examining the physical environment (Fig. 6).
Restrain and struggle both began in the 3rd month. Both behaviors peaked rapidly in the 4th month at 3.4 intervals per test for restraint and 1.5 intervals per test for struggle. Maternal restraint decreased to .2 interval by the 9th month, subsequently occurring only infrequently. Infant struggling dropped to .2 interval in the 5th month and was completely absent after the 6th month.

INDEPENDENCE IN INFANT GORILLAS 259
w I Struggle o-a M Restrah I
8 H M Retrieve I \ 4
Y 3 F 3
3 2
8 1
z w I-
z
3 = 0 w
2 4 6 8 10 12 14 16 I8 20 AGE (Lunar mths)
Fig. 4. Incidence of struggle, restrain, and retrieve as a function of infant age
Mothers began retrieving infants to closer positions (Fig. 4) at about the same time that they were restraining the infants. Mothers continued to retrieve infants, however, after restraint dropped out. This occurred most frequently in the 1st 12 months, but mothers did retrieve infants throughout the entire 18 months of study. Retrieval began in the 3rd month with an incidence of .6 interval. It increased to a high of 4.3 in the 8th month, then decreased, with a terminal frequency of .5 interval. We cannot account for the large decline in maternal retrieve following the 6-week break in observations. However, the decline does not seem to reflect the absence of the observer, as it is the only behavior which showed a precipitous change over the break.
Nipple Contact, Wean, Remove
Infant nipple contact (Fig. 5), with or without suckling, was observed at low frequencies throughout the study. The highest frequency of occurrence was in the 1st month at 3.5 intervals per test. Thereafter, this behavior generally decreased with a slight resurgence in the final months to a terminal frequency of .9 interval.
1.0
0.5
0.0 2 4 6 8 10 12 14 16 18 20
AGE (Lunar months) Fig. 5 . Incidence of nipple contact ( 0 - O ) , and wean (B-D) as a function of infant age.
260 HOFF, NADLER, A N D MAPLE
y2*1 M I Object exam H I Social exam M
1 r I 1
2 4 6 8 10 I2 14 16 18 20 AGE (Lunar months)
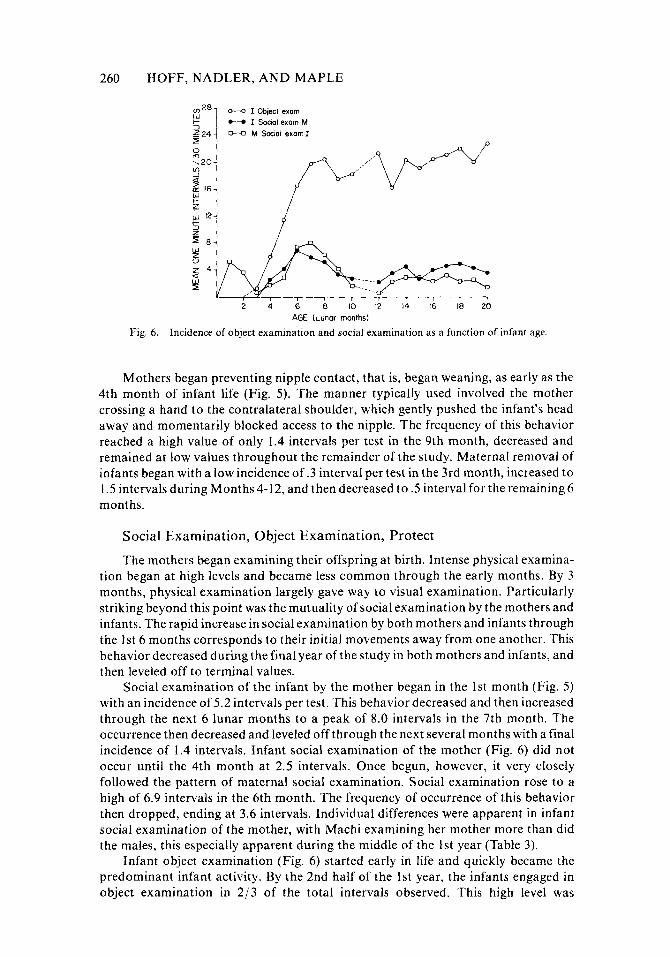
Fig. 6. Incidence of object examination and social examination as a function of infant age.
Mothers began preventing nipple contact, that is, began weaning, as early as the 4th month of infant life (Fig. 5). The manner typically used involved the mother crossing a hand to the contralateral shoulder, which gently pushed the infant's head away and momentarily blocked access to the nipple. The frequency of this behavior reached a high value of only 1.4 intervals per test in the 9th month, decreased and remained at low values throughout the remainder of the study. Maternal removal of infants began with a low incidence of . 3 interval per test in the 3rd month, increased to 1.5 intervals during Months 4-12, and then decreased to .5 interval for the remaining6 months.
Social Examination, Object Examination, Protect
The mothers began examining their offspring at birth. Intense physical examina- tion began at high levels and became less common through the early months. By 3 months, physical examination largely gave way to visual examination. Particularly striking beyond this point was the mutuality of social examination by the mothers and infants. The rapid increase in social examination by both mothers and infants through the 1st 6 months corresponds to their initial movements away from one another. This behavior decreased during the final year of the study in both mothers and infants, and then leveled off to terminal values.
Social examination of the infant by the mother began in the 1st month (Fig. 5 ) with an incidence of 5.2 intervals per test. This behavior decreased and then increased through the next 6 lunar months to a peak of 8.0 intervals in the 7th month. The occurrence then decreased and leveled off through the next several months with a final incidence of 1.4 intervals. Infant social examination of the mother (Fig. 6) did not occur until the 4th month at 2.5 intervals. Once begun, however, it very closely followed the pattern of maternal social examination. Social examination rose to a high of 6.9 intervals in the 6th month. The frequency of occurrence of this behavior then dropped, ending at 3.6 intervals. Individual differences were apparent in infant social examination of the mother, with Machi examining her mother more than did the males, this especially apparent during the middle of the 1st year (Table 3) .
Infant object examination (Fig. 6) started early in life and quickly became the predominant infant activity. By the 2nd half of the 1st year, the infants engaged in object examination in 2 /3 of the total intervals observed. This high level was

INDEPENDENCE IN INFANT GORILLAS 261
maintained through the remainder of the study. Scoring of this behavior began in the 3rd month, at which time it occurred in 1.2 intervals per test. Object examination increased rapidly to an 8-month value of 20.4 intervals. This behavior remained at approximately this level throughout the remaining months, ending at 22.8 intervals per test at the conclusion of the study. The female infant, Machi, exhibited this behavior (Table 3) at a lower frequency than did the males early and at later scattered points through the study.
Protective behaviors by a mother were almost exclusively related to attempts by the adult male to examine an infant. Under these circumstances, a mother typically retrieved the infant and turned her back toward the male, sometimes pushing his hand away. A mother also protected her infant when any adult lunged toward the mother- infant dyad. She typically retrieved the infant, crouched over it, and raised her arm, thereby blocking any blows. Scoring of this behavior began in the 3rd month at . l interval and continued at low frequencies until the 13th month, after which there were no further observed instances of maternal protection.
Discussion
The infant gorillas in this study exhibited a trend toward increasing independence throughout the 1st 1 % years of life. Based on the studies of monkeys (e.g., Hansen, 1966; Harlow et al., 1963; Hinde & Spencer-Booth, 1967; Hinde &White, 1974) and apes (e.g., Fossey, 1979; Horvat et al., in press; Kingsley, 1977), the most salient measure of increasing independence is decreasing spatial proximity of a mother-infant dyad. These studies typically reported that ventro-ventral contact, providing the closest measure of contact within a dyad, decreases rapidly in early infancy with a concurrent increase in other forms of contact. This is similar to what was found in the present study. Ventro-ventral contact dropped rapidly, the dramatic decrease between the 3rd and 4th months being similar to that reported by Kingsley (1977) for an infant lowland gorilla. Because the other reports on great ape development (i.e., Fossey, 1979; Horvat et al., in press) did not separate ventro-ventral contact from total contact, no direct comparisons between them and the present study can be made.
The development of the variables of other contact and proximate positions were similar, although other contact was initiated earlier and peaked 1 month later at a slightly higher frequency than did the proximate position. These were both supplanted by the developing distant variable at about I year of age. The infants were also first observed beyond 5 m from their mothers at about this age. At the termination of this study, at 1 $$ years of age, the little contact that occurred between mothers and infants was largely other contact. Additionally, both proximate and distant measures had peaked and were on the decline, while the frequency of far distant slightly surpassed distant.
The findings of the present study regarding mother-infant spatial proximity amplify and extend those of other studies of gorillas. In our study, a break in contact was first observed at I 1 weeks, as compared with Kingsley’s ( 1 977) finding of 6 weeks. Kingsley’s captive infant, however, lived with only its mother and 1 adult female (S. Kingsley, personal communication). The difference between these studies, therefore, may be due to the less complex social environment employed by Kingsley (1977), resulting in less maternal restraint of infant movements. Nicolson (1977), in fact, reported such a relationship among chimpanzees living in various social situations. Alternatively, due to the small sample sizes in all 3 of these studies, the differences may simply reflect individual differences among mother-infant dyads. With regard to reports of feral mountain gorillas, the present results are similar to those of Schaller

262 HOFF, NADLER, AND MAPLE
(1963), but somewhat different from those of Fossey (1979). Schaller (1963) reported contact broken at about 3 months; Fossey (1979) at 5 months. Additionally, when compared to those studied by Fossey (1979) and Schaller (1963), the Yerkes infants were intermediate in moving to greater distances from their mothers. Schaller observed infants beyond 5 m from their mothers at 8 months and Fossey at approximately 18 months. In our study, infants reached this distance from their mothers at approximately 1 year.
The increasing incidence of object examination is also indicative of increasing infant independence in that it reflects a shift in the infant’s attention away from its mother and toward the environment. Jensen, Bobbitt, and Gordon (1967) proposed that in pigtailed macaques, increasing environmental interaction by infants cor- responds to detachment from the mother and attachment to other objects. The present data provide support for this hypothesis. From the 3rd to the 8th months, a sharp increase in object examination largely coincided with a sharp decrease in ventro- ventral contact. Additionally, object examination attained its highest levels as the infant moved farther away from its mother.
In many primate species, weaning is associated with the development of infant independence. Weaning was initiated at low frequency in this captive group of gorillas during the 4th month of infant life. This is much earlier than weaning times reported for chimpanzees (Clark, 1977) and orang-utans (Horr, 1977), for which weaning occurs only after several years. No other longitudinal studies of the gorilla exist that contain quantitative data on weaning. The present findings, however, are not inconsistent with observations by both Schaller (1963) and Fossey (1979), who reported some evidence of weaning within the 1st year of life. Whether a more active weaning process takes place in later infancy in the feral gorilla than among captive groups such as the one currently under study is yet to be determined. As in feral chimpanzees (Clark, 1977) and gorillas (Schaller, 1963), weaning in this captive group was initiated without maternal aggression and was usually accomplished by the mother simply covering her breasts with her arms.
The role of the adult male in developing independence, although not the subject of direct study in the present project, also appears to be of some importance. This was the male’s first adult experience with infants, and he was persistent in his attempts to examine and play with the infants (see Tilford & Nadler, 1978). The mothers’ response to this is shown in the protection data. Virtually all protection occurred in response to the presence of the adult male, and resulted in increased maternal restrictiveness. This type and this amount of maternal protection were not reported for wild populations by either Schaller (1963) or Fossey (1979). Stewart (1977), however, did observe protective behavior by feral gorilla mothers in response to inquisitiveness by juveniles, especially by juvenile females.
A major question addressed in our study is the relative contribution of mother and infant to developing infant independence. Considerable controversy exists in the literature regarding this question. Major infant responsibility is implied in many field reports (e.g., DeVore, 1963; Goodall, 1965; Jay, 1963; van Lawick-Goodall, 1967; Schaller, 1963) and explicitly stated in several laboratory studies (e.g., Kaufman, 1974; Kaufman & Rosenblum, 1967; Rosenblum, 1971). In the above studies, monkey and ape infants moved away from their mothers and toward other objects in the environment. Object examintion and peer play were reported to be primary in attracting the infants away from their mothers. In all these cases, the mothers played a role in developing independence through the weaning process, although independence occurred much sooner and was more quickly absolute in monkeys than in chimpanzees

INDEPENDENCE IN INFANT GORILLAS 263
or gorillas. Contrary to these reports are those of Hansen (1966), Hinde (1974, 1975), Hinde and White (1974), and Jensen et al. (1967, 1973), among others. These studies found predominant maternal responsibility for infant independence as indicated by a variety of measures. Maternal rejection was seen as a major instigating force behind developing independence (see Suomi, 1976, for a review of this controversy). In her sample of feral mountain gorillas, Fossey (1979) also concluded that infant independence was maternally determined. She did not provide supporting quantitative data, however, regarding variables such as leave, return, and restrain. In order to investigate the question of the relative contributions of mother and infant to developing independence in the present study, a variety of behavioral categories were used.
The measures of infant behavior considered to reflect its own contribution to its developing independence include struggle, leave, and object examination. Infant behavior indicative of the infant’s efforts to maintain the mother-infant bond include return, social approach, and follow. The infants were observed struggling primarily during the 3rd and 4th months of life, during which time they were in close contact with the mother. Struggling decreased after the 4th month as infants increasingly left their mothers and spent increased time proximate to and distant from their mothers. Object examination was the main infant activity reported in the present study; by the 8th month of life it was observed in approximately 67% of the total intervals through the end of the study. Relevant to the field reports suggesting that peer play groups are important in developing infant independence, in the present study peer play away from the mothers began in the 5th month, becoming more frequent and complex through 18 months. (See Hoff, Nadler, & Maple, in press, for a report on infant play.)
Although the infants left their mothers with increasing frequency from the 4th- 10th months of life, they also returned to and socially approached the mothers at relatively high frequencies and, especially during the latter months, increasingly followed their mothers. These data suggest that the infants initially struggle for independence from maternal contact, but subsequently act to maintain a relationship by regular, periodic returns to the mother and increased following of the mother in response to her leaving.
The measures of maternal behavior that reflect the mother’s efforts to maintain the integrity of the dyad are restrain, retrieve, and return. Measures that reflect the mother’s behavior directed toward increasing the independence of the infants are wean and leave. During Months 3-5, the mothers responded to infant struggling with restraint, and during Months 4-10, they responded to infant leaving by retrieval. Maternal returning contributed little to reestablishment of contact. The mother’s role during much of the 1st year of life, therefore, was directed toward reestablishing and maintaining the mother-infant bond. Only during the last 3-4 months of this study did the mothers begin leaving the infants at frequencies approaching those at which the infants left the mothers. As noted above, the increase in maternal leaving correlated with increases in infant following. These data suggest, therefore, that the mother’s behavior changed during the 1st 1 ‘/2 years of infant development, restricting and restraining independence (activity away from mother) during an early stage of development and, later, actively dissociating herself from the infant.
Individual differences were apparent in a number of the 25 behaviors which we report. The most consistent differences appeared in the various contact and separation measures. These show the female infant, Machi, in closer contact with her mother during early life, and later, when not in contact with her mother, being more often within I m and less often beyond 5 m relative to the males. With our small sample size,

264 HOFF, NADLER, AND MAPLE
we cannot, of course, differentiate between individual and gender-related differences, but this indication of relatively more contact with the mother by the female infant is consistent with reports of macaques (e.g. Jensen, Bobbitt, & Gordon, 1968b).
Notes
‘In light of the recent controversy regarding sampling methodology (e.g., Altmann, 1974; Rhine & Flanigon, 1978) and following Rhine and Flanigon (1978). correlations were performed between the one-zero frequencies and actual frequencies for a variety of behaviors. Data for these correlations were taken from the 10 months of focal animal sampling that were originally tabulated as one-zero frequencies. The behaviors included both maternal and infant leave, return and social approach, and additionally. maternal protect. remove, and retrieve. The correlations ranged from .92 to .99. All of these are significant (p < . O l ) and indicate that for these behaviors in our sample, the one-zero methodology provides an accurate representation of behavior as it would have been measured by the focal animal technique. Furthermore. it is noted by the authors that the term “frequency” is used in this paper to refer to the frequency of intervals in which a given behavior occurred, rather than to the actual frequency of behavior.
This work was supported by N S F Grant BNS75-06287 and P H S Grant RR-00165 from NIH. The authors express their gratitude t o Dr. K . Wallen for reading an early version of the manuscript. Dr. R. B. Fischer was responsible for the first 3 months of data collection.
References
Altmann, J . ( 1974). Observational study of behavior: Sampling methods. Behaviour, 49:227-267. Altmann, S. A. (1959). Field obscrvations on a howling monkey society. J. Mammal.. 40:317-330. Clark. C. B. (1977). A preliminary report on weaning among chimpanzees of the Gombe National Park,
Tanzania. In S. Chevalier-Skolnikoff and F. E. Poirier (eds.), Primare Bio-social Development. New York: Garland. Pp. 235-260.
DeVore. I. (1963). Mother-infant relations in free-ranging baboons. In H. I>. Rheingold (ed.), Mafernal Behavior in Manima1.s. New York: Wiley. Pp. 305-335.
Fossey. D. (1979). Development of the mountain gorilla (Gorilla gorilla beringei): The first thirty-six months. I n D. A. Hamburg and E. R. McCown (eds.), 7he Great Apes. Menlo Park, Calif.: Benjamin/Cummings. Pp. 139-184.
Goodall, J . (1965). Chimpanzees of the Gombe Stream Reserve. In 1. DeVore(ed.), Primare Behavior. New York: Holt, Rinehart, & Winston. Pp. 425-473.
Hansen, E. W. (1966). The development of maternal and infant behavior in the rhesus monkey. Behaviour,
Harlow. H. F.. Harlow, M. K., and Hansen. E. W. (1963). The maternal affectional system of rhesus monkeys. In H. L. Rheingold (ed.), Maternal Behavior in Mammals. New York: Wiley. Pp. 254-281.
Hinde. R. A. (1974). Mother/infant relations in rhesus monkeys. In N. F. White (ed.), Et:rhology and P.svchia/r,v. Toronto: University of Toronto Press. Pp. 29-46.
Hinde, R. A. (1975). Mothers’ and infants’ roles: Distinguishing the questions to be asked. Parent-lnfant lnreraction, Ciba Foundation Symposium 33. Pp. 5-13.
Hinde. R. A., and Atkinson, S. (1970). Assessing the roles of social partners in maintaining mutual proximity, as exemplified by mother-infant relations in rhesus monkeys. Anim. Behav., I R : 169- 176.
Hinde. R. A., and Spence-Booth, Y. (1967). The behavior ofsocially living rhesus monkeys in their first two and a half years. Anim. Behav., 15:169-196.
Hinde, R. A, , and White, L. E. (1974). Dynamics of a relationship: Rhesus mother-infant ventro-ventral contact. J. Comp. Phxsiol. Psycho/.. 863-23.
Hoff. M. P., Nadler, R. D., and Maple, T. (1980). Infant motor development in a captive group of lowland gorillas. Paper presented a t the American Society of Primatologists, Winston-Salem, North Carolina. June, 1980.
Hoff. M.P.. Nadler. R .D. , and Maple, T. (in press). The development of infant play in a captive group of lowland gorillas (Gorilla gorilla gorilla). A m . J. Primaro.
Horr, D. A. (1977). Orang-utan maturation: Growing up in a female world. In S. Chevalier-Skolnikoffand F. E. Poirier (eds.), Primate Bio-social Developmenf. New York: Garland. Pp. 289-321.
Horvat, J. R., Coe, C. L . , and Levine, S. (in press). Infant development and maternal behavior in captive chimpanzees. In W. Smotherman and R. Bell (eds.), Ear/r Experience and Behavior. Jamaica, N. Y.: Spectrum.
27: 107- 149.

INDEPENDENCE 1N INFANT GORlLLAS 265
Itoigawa, N. (1973). Group organization of a natural troop of Japanese monkeys and mother-infant interactions. In C. R. Carpenter (ed.,), Behavioral Regulators of Behavior in Primates. Cranbury, N.J.: Associated University Presses. Pp. 229-250.
Jay, P. (1963). Mother-infant relations in langurs. In H. L. Rheingold (ed.), Maternal Behavior in Mammals. New York: Wiley. Pp. 282-304.
Jensen, G. D., Bobbitt, R. A., and Gordon, B. N. (1967). The development of mutual independence in mother-infant pigtail monkeys, Maraca nemestrina. In S. A. Altmann (ed.), Social Communication Among Primates. Chicago: University of Chicago Press. Pp. 43-53.
Jensen, G. D., Bobbitt, R. A., and Gordon, B. N. (1968a). Effects of environment on the relationship between mother and infant pigtailed monkeys (Macaca nemestrina). J . Comp. Ph,vsiol. Psjdml.,
Jensen, G. D., Bobbitt. R. A,, and Gordon, 8. N. (1968b). Sex differences in the development of independence in infant monkeys. Behaviour, 30: 1-14.
Jensen, G. D., Bobbitt, R. A., and Gordon, B. N. (1973). Mothers’and infants’ roles in the development of independence of Macaca nemestrina. Primates. 14:79-88.
Jones, C . , and Sabater Pi, J . (1971). Comparative ecology of Gorillagorilla (Savage & Wyman) and Pan rroglodvtes (Blumenbach) in Rio Muni, West Africa. Bib/. Primatol., 13: 1-96.
Kaufman, 1. C. (1974). Mother-infant relations in monkeys and humans: A reply to Professor Hinde. In N. F. White (ed.), Ethology and Psychiatr.y. Toronto: University of Toronto Press. Pp. 47-68.
Kaufman, 1. C., and Rosenblum, L. A. (1969). The waning of the mother-infant bond in two species of macaque. In B. M. Foss (ed.), Determinants oflnfani Behaviour. IY London: Methuen. Pp. 41-59.
Kaufmann, J. (1966). Behavior of infant rhesus monkeys and their mothers in a free-ranging band. Zoologica, 51: 17-28.
Kingsley, S. (1977). Early mother-infant behaviour in two species of great apes. Dodo (J. Jersqr Wild. Preserv. TTust), 1455-65.
Lang. E. M. (1959). The birth of a gorilla at Bade Zoo. Int. Zoo Yrbk., 1:3-7. van Lawick-Goodall, J . (1967). Mother-offspring relationships in free-ranging chimpanzees. In D. Morris
Maple, T., Wilson, M. E., Zucker, E. L., and Wilson, S. F. (1978). Notes on the development of a
Nadler, R. D. (1976). Rann versus Calibar: A study in gorilla behavior. Yerkes Newsltr, 13:l 1-14. Nicolson, N. A. (1977). A comparison ofearly behavioral development in wild and captive chimpanzees. In
S. Chevalier-Skolnikoff and F. E. Poirier (eds.), Primate Bio-socia/ Development. New York: Garland. Pp. 529-560.
Rhine, R. J., and Flanigon, M. (1978). An empirical comparison of one-zero, focal animal, and instantaneous methods of sampling spontaneous primate social behavior. Primates, 19:353-361.
Rosenblum, L. A. (1971). The ontogeny of mother-infant relations in macaques. In H. Moltz (ed.), The Onto,qen,p of Vertebrate Behavior. New York: Academic Press. Pp. 3 15-367.
Schaller, G. (1963). The Mountain Gorilla. Chicago: University of Chicago Press. Stewart, K. J . (1977). The birth of a wild mountain gorilla (Gorilla gorilla heringei). Primates. 18:965-976. Suomi, S. J. (1976). Mechanisms underlying social development: A re-examination of mother-infant
interactions in monkeys. In A. Pick (ed.), Minnesota Synzposium on Child Ps.p~cho1og.v. Volume 10. Minneapolis: University of Minnesota Press. Pp. 201-228.
Tilford, B. L., and Nadler, R. D. (1978). Male parental behavior in a captive group of lowland gorillas (Gorilla gorilla gorilla). Folia Primatol., 29:218-228.
Vessey, S. H. ( I 968). Behavior of free-ranging rhesus monkeys in the first year of life. Amer. Zool., 8:740. White, L. E., and Hinde, R. A. (1975). Some factors affecting mother-infant relations in rhesus monkeys.
66:259-263.
(ed.), Primate EthologL,. London: Weidenfeld. Pp. 287-346.
mother-reared orang-utan: The first six months. Primares, l9:593-602.
Anim. Behav., 23:527-542.
Copyright © 2022 FDOKUMEN




















