
Reproductive factors affecting survivorship in captive callitrichidae
15
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 84:291-305 (1991) Reproductive Factors Affecting Survivorship in Captive Callitrichidae CASHELL E. JAQUISH, TIMOTHY B. GAGE, AND SUZETTE D. TARDIF Department of Anthropology, Northwestern University, Evanston, Illinois 60208 (C.E.J.); Department of Anthropology, State University of New York, Albany, New York 12222 (T.B.G.); Oak Ridge Associated Universities, Oak Ridge, Tennessee 37831-011 7 (S.D.T.) KEY WORDS Optimal litter size, Maternal effects, Survivor- ship, Callitrichidae ABSTRACT Efforts to breed Callitrichidae in captivity have resulted in high fertility accompanied by high infant mortality. This paper investigates the relationship between reproductive characteristics and survivorship in the Oak Ridge and Associated Universities (ORAU)callitrichid colony. Records of 2,834 individuals were analyzed using Cox Proportional Hazards regression to investigate factors affecting infant survivorship. Species and birth cohort were found to have a significant (2 ' < 0.05) effect on survivorship. Litter size also was an important factor in determining survivorship. Individuals born into singlet and twin litters had significantly (P < 0.001) higher survivorship than triplets. Factors which affect survivorship perinatally (0-1 months of age), prior to breeding age and subsequent to the first month of life were then investigated. Litter size was a significant (P < 0.05) factor in determining survivorship perinatally, with twins having significantly higher survivorship than both singlets and triplets. After the first month oflife, litter size no longer showed a significant effect on survivorship. Records of 145 dams were ana- lyzed using the same procedures to investigate reproductive characteristics associated with survivorship. Age at first litter showed a significant (P < 0.Q1) positive relationship with dam survivorship. These results suggest various factors affect survivorship at specific temporal points during development, and also suggest selection for survivorship perinatally to be strongly associ- ated with litter size, possibly through sibling competition and maternal effects. Primates are ordinarily characterized as having one or two offspring per litter, one being the most common. There are three primate families in which species have mean litter sizes greater than or equal to 1.5. These are the Lemuridae (4 species), Lorisidae (2 species), and the Callitrichidae (all species) (Harvey and Clutton-Brock, 1985).The Callitrichidae clearly stand out in that all of the species in the family average 21.5 offspring per litter. This study ad- dresses variation in litter size in the Callitri- chidae as a potential factor affecting survi- vorship. A general understanding of callitrichid re- production is necessary to identify the mech- anisms by which litter size can respond to selection, and to then develop an accurate view of the interaction of litter size and offspring survivorship after birth. In captiv- ity, most s ecies are able to reproduce year months, Saguinus oedipus being the longest at almost 8 months (Tardif et al., 1984; Ziegler et al., 1987). Dizygotic twins are the norm; therefore, there is the possibility of a pair producing four offspring in 1 year. The gestation period varies by species, but is usually in the range of 135-184 days (Sussman and Kinzey, 1984; Ziegler et al., 1987). Captive common marmosets are able round, wit g an average birth interval of five Received June 28,1989; accepted June 27,1990. Address correspondence to Cashell E. Jaquish, Oak Ridge Associated Universities, Marmoset Research Center, P.O. Box 117, Oak Ridge, TN 37831-0117. @ 1991 WILEY-LISS, INC.
Transcript of Reproductive factors affecting survivorship in captive callitrichidae

AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 84:291-305 (1991)
Reproductive Factors Affecting Survivorship in Captive Callitrichidae
CASHELL E. JAQUISH, TIMOTHY B. GAGE, AND SUZETTE D. TARDIF Department of Anthropology, Northwestern University, Evanston, Illinois 60208 (C.E.J.); Department of Anthropology, State University of New York, Albany, New York 12222 (T.B.G.); Oak Ridge Associated Universities, Oak Ridge, Tennessee 37831-011 7 (S.D.T.)
KEY WORDS Optimal litter size, Maternal effects, Survivor- ship, Callitrichidae
ABSTRACT Efforts to breed Callitrichidae in captivity have resulted in high fertility accompanied by high infant mortality. This paper investigates the relationship between reproductive characteristics and survivorship in the Oak Ridge and Associated Universities (ORAU) callitrichid colony. Records of 2,834 individuals were analyzed using Cox Proportional Hazards regression to investigate factors affecting infant survivorship. Species and birth cohort were found to have a significant (2' < 0.05) effect on survivorship. Litter size also was an important factor in determining survivorship. Individuals born into singlet and twin litters had significantly (P < 0.001) higher survivorship than triplets. Factors which affect survivorship perinatally (0-1 months of age), prior to breeding age and subsequent to the first month of life were then investigated. Litter size was a significant (P < 0.05) factor in determining survivorship perinatally, with twins having significantly higher survivorship than both singlets and triplets. After the first month oflife, litter size no longer showed a significant effect on survivorship. Records of 145 dams were ana- lyzed using the same procedures to investigate reproductive characteristics associated with survivorship. Age at first litter showed a significant (P < 0.Q1) positive relationship with dam survivorship. These results suggest various factors affect survivorship at specific temporal points during development, and also suggest selection for survivorship perinatally to be strongly associ- ated with litter size, possibly through sibling competition and maternal effects.
Primates are ordinarily characterized as having one or two offspring per litter, one being the most common. There are three primate families in which species have mean litter sizes greater than or equal to 1.5. These are the Lemuridae (4 species), Lorisidae (2 species), and the Callitrichidae (all species) (Harvey and Clutton-Brock, 1985). The Callitrichidae clearly stand out in that all of the species in the family average 21.5 offspring per litter. This study ad- dresses variation in litter size in the Callitri- chidae as a potential factor affecting survi- vorship.
A general understanding of callitrichid re- production is necessary to identify the mech- anisms by which litter size can respond to selection, and to then develop an accurate
view of the interaction of litter size and offspring survivorship after birth. In captiv- ity, most s ecies are able to reproduce year
months, Saguinus oedipus being the longest at almost 8 months (Tardif et al., 1984; Ziegler et al., 1987). Dizygotic twins are the norm; therefore, there is the possibility of a pair producing four offspring in 1 year. The gestation period varies by species, but is usually in the range of 135-184 days (Sussman and Kinzey, 1984; Ziegler et al., 1987). Captive common marmosets are able
round, wit g an average birth interval of five
Received June 28,1989; accepted June 27,1990. Address correspondence to Cashell E. Jaquish, Oak Ridge
Associated Universities, Marmoset Research Center, P.O. Box 117, Oak Ridge, TN 37831-0117.
@ 1991 WILEY-LISS, INC.

292 C.E. JAQUISH ET AL
to breed at approximately 13 months and conceive at approximately 18 months, and female tamarins reach sexual maturity at approximately 18 months (Sussman and Knzey, 1984). After birth, other group mem- bers, especially the male parent, care for the offspring. This cooperative rearing includes carrying and grooming infants (Cleveland and Snowdon, 1984).
Attempts to breed Callitrichidae in captiv- ity have resulted in high fertilit accompa- nied by high infant mortality (&bourn et al., 1983; Kirkwood et al., 1983; Tardif et al., 1986). This suggests that along with selec- tion for increased litter size, there is strong selection perinatall on offspring phenotype.
chids and the observed high infant mortality have prompted interest in the relationship between litter size and overall reproductive success. Two previous approaches to the evolution of litter size will be contrasted to provide a theoretical background for the study.
Litter size has often been investigated as a component of reproductive success (the num- ber of offspring a parent is capable of suc- cessfully raising to weaning). Thus it has been examined primarily as a trait in the parental generation. This has provided the basis for many models of the evolution of litter and clutch size that evoke selection acting to increase litter size within certain constraints. Lack (1947) developed an opti- mization model of the evolution of clutch size; in this model, the average clutch size in the population reflected the most young which a parent could raise to independence. This model evoked selection to increase clutch size and aspects of parental care to constrain clutch size resulting in an opti- mum. Cody (1966) also integrated a tradeoff between parental survival and fecundity (within the same optimization framework), assuming that those individuals which pro- duce larger litters have reduced survivor- ship. Clutch size manipulation studies have shown mixed results and the negative corre- lation between fecundity and survival is not always apparent (Rockwell et al., 1987).
Selection may also act on litter size prior to birth. There can be a progressive loss of potential embryos as ovulation rate in- creases. In Booroola merino sheep when ovu- lation rate exceeds three, there is little or no increase in litter size at birth (Bindon, 1984). The loss of potential embryos is most likely due to maternal uterine environment; i.e.,
The unusual repro B uctive biology of callitri-
crowding, inadequate nutrition, etc., but could be due to the viability of the embryo. High rates of in-utero loss are found in most prolific species (Bindon, 1984; Hanrahan, 1980). Gengozian and Merritt (1970) found that in a sample of births from unilateral ovariectomized female marmosets, all single births had either two placentae or were chi- meric by analysis of sex chromosomes, sug- gesting loss of a sibling in utero.
A second approach to the evolution of litter size considers enetic and environmental
ation and their interactions. Maternal genes can affect litter size and subsequent survi- vorship through number of eggs ovulated, uterine capacity, and aspects of maternal performance, such as milk production (Will- ham, 1972; Robison, 1981). The offspring genes can affect litter size by directly affect- ing offspring survivorship in utero and after birth. Litter size also affects offspring survi- vorship through sibling competition and ma- ternal ability to care for increased numbers of offspring. Thus, litter size is an extremely complex character, which is determined by various genetic and environmental factors prior to birth, and subsequent to birth litter size affects offspring survivorship. Studies of maternal effects have shown that maternal performance contributes a large proportion to the variance in offspring phenotypes early in life and decreases during ontogeny, whereas non-maternal environmental ef- fects increase in relative importance during development (Cheverud et al., 1983; Hanra- han and Eisen, 1974; Legates, 1972; Rut- ledge et al., 1972).
Many species showing variation in litter size also show an inverse relationship be- tween litter size and postnatal survivorship (Cameron, 1973, rodents; Bindon, 1984, sheep). Decreases in survivorship correlated with increased litter size are most likely due to a combination of lowered birthweight and nutritional stress. It is apparent that survi- vorship subsequent to birth is dependent on parental ability to provide adequate care and nutrition, as well as the direct genetic com- ponent of offspring phenotype and the envi- ronment provided by the number of siblings, i.e., litter size.
Evolution of litter size in the Callitri- chidae is of particular interest due to high parental investment and the practice of co- operative rearing, since both of these factors would be important to the predictions from optimality models. The extremely high in-
factors in both t a e parental and filial gener-

REPRODUCTION AND SURVIVORSHIP IN CALLITRICHIDAE 293
fant mortality found in captive colonies sug- gests strong selection on offs ring phenotype
the question as to whether the Callitrichidae are increasing their reproductive success by having larger litters.
The extensive records for captive callitri- chid colonies provide an opportunity to in- vestigate the relationship between charac- ters in the parental and filial generation and survivorship. In particular, those covariates associated with reproductive success which influence mortality will be identified and two predictions from optimal litter size mod- els tested; 1) that the most common litter size will have the highest survivorship until reproductive age, and 2) that dams with higher average litter sizes show lower survi- vorship.
during the first few weeks o ! life. This raises
MATERIALS AND METHODS The Oak Ridge Associated Universities
(ORAU) Marmoset Research Center has been in operation for over 20 years. It was begun by Dr. Nazareth Gengozian (Gengo- zian, 19691, as part of the Medical and Health Sciences Division of the ORAU group. The majority of the colony is com- posed of three species of Callitrichidae, Saguinus oedipus, Saguinus fuscicollis (sub- species: nigrifrons, lagonotus, leucogenys and illigeri) and Callithrix jacchus. As of 1985 approximately 415 tamarins and mar- mosets composed the colony; 267 of these were S. oedipus (Clapp and Tardif, 1985).
As of 1979 a breeding facility, which is physically separate from the main research building, houses the majority of animals. Individuals are chosen for breeding based on two criteria: 1) genetic representation in the population. Animals are chosen for breeding such that there is a relatively even genetic representation of the population’s founders. 2) Rearing experience. Only individuals who have had prior experience with infant sib- lings are bred (Clapp and Tardif, 1985). These criteria have been used from 1979 to
present. Prior to 1979, virtually all adult animals were bred. Individuals are closely observed for compatibility and incompatible pairs are remated. Family units are left in- tact through the second and sometimes third consecutive litter. Thereafter, older off- spring are housed in individual cages, at the main facility, until bred.
Cages are kept visually isolated from one another, and olfactory and auditory expo- sure is kept to a minimum. The BAM diet has been fed to the animals at ORAU since 1978; prior to this a diet based on Purina Primate chow was fed. The BAM diet consists of a morning feeding of bananas, applesauce, and high protein monkey chow, with the addition of vitamins DS, C, multiple B, folic acid, and copper (Clapp and Tardif, 1985). The afternoon feeding consists of canned marmoset diet with a variety of dietary sup- plements.
A number of changes in husbandry and management deserve mention. With the completion of the breeding facility (1979), ca3ge volume increased from 7.76 ft3 to 150 ft , and cage complexity was increased. Ani- mals were also exposed to temperate lighting and ambient summertime temperatures in the breeding facility. Gengozian (1969) and Clapp and Tardif (1985) give detailed de- scriptions of management before and after 1979.
Records of 2,834 individuals (spanning 23 years) were used for analysis of survivorship with respect to traits commonly associated with reproductive success using Cox Propor- tional Hazards regression (Lee, 1980). Sam- ple sizes, listed by species and litter size, ap ear in Table 1.
8ox Proportional Hazards model uses log- likelihood regression; thus, the effects of fac- tors not continuously distributed, such as sex, species, etc., can be investigated. The resulting regression coefficients are inter- preted in the same manner as standard re- gression coefficients, but they differ in sign, i.e., the coefficient of a covariate that has a positive effect on survivorship will have a
TABLE 1 . Sample size by cohort litter size and species
Litter size S. fuscicollis S. oedipus C. jacchus Total
One 29 1 143 24 458
Three 162 159 57 378 Total 1,833 734 267 2,834
Two 1,380 432 186 1,998

294 C.E. JAQUISH ET AL.
negative sign. An advantage of the Cox re- gression is that it is semi-parametric, in that it allows for an arbitrary baseline hazard function (the probability of dying during a specified time interval is not fitted to a par- ticular shape, as in a Weibull model), but also assumes that covariates induce propor- tional shifts of this arbitrary hazard function throughout the lifetime. The proportional hazards assumption implies that groups with different values of the covariates expe- rience the same relative hazard functions over time. Thus, if sex were run as a covari- ate, the ratio of male to female hazard func- tions should remain constant over time.
The survival function estimated by the Cox model is as follows:
h(t,z) = h,ft)expfz’p) where h,(t) is the baseline hazard function and exp(z,’p) is the proportional shift of the covariate vector z and the coefficient vector p. Thus, h,(t) gives a measure of the shape of the hazard function and expiz,’p) gives a measure of the direction and magnitude of the change in the hazard function attribut- able to a particular covariate.
To determine if the hazard differentials identified by this study were truly propor- tional with respect to age, survivorship was analyzed through the whole life and during specific age intervals. If survivorship curves for each covariate being compared were roughly parallel within the time interval, then the assumption of proportional hazards was not violated. All covariates were visually examined in this manner during specific age intervals to verify that the assumptions of the model were not violated. This also pro- vided a method of identifying covariates which affected survivorship only over a por- tion of the life span. Cox beta coefficients were converted to relative risk values for ease of interpretation using the following equation:
RR = expfbeta) Covariates were divided into two groups,
those which affect offspring survivorship through maternal characteristics, i.e., dam’s age and dam’s average litter size and those affecting offspring survivorship in the filial generation, i.e., litter size born into (cohort litter size), species, and sex. Due to manage- ment changes in the colony, outlined above, time interval born into (birth cohort) was
also considered as a covariate. The time period covered was divided into four inter- vals for comparison: 1962-1970,1971-1975, 1976-1980, and 1981-1985, approximately corresponding to management changes in housing and nutrition.
Indicator variables were used for covari- ates with more than two cate ories. Birth cohort was coded so that all su % sequent in- tervals were compared to the first (1962-70). The species variable was coded so that all species were compared to Saguinus fuscicol- lis. S. fuscicollis was chosen for corn arison
the longest period of time. Litter sizes of one and three were compared to litter size of two. Individuals with unknown sex were coded separately and both females and unknowns were compared to males.
Survivorship analysis was performed 1) through the whole life, 2) during the first month of life, 3) subsequent to the first month of life, and 4) during the first 2 years of life (2 years corresponds to the approxi- mate age at which the animals in this colony are first bred). From the data it is not known if those not surviving the first day of life are still born or live born; it can only be assumed that they were near full term. Therefore, analysis of survivorship subsequent to the first day of life was performed to ensure that no stillbirths were included in the sample.
Survival analysis of 145 dams (those with known birth dates) was performed in the same manner using the covariates of aver- age litter size, age at first litter, species, and birth cohort to investigate the relationship between factors which affected reproductive success and survivorship.
Variation between and within species in litter size was analyzed using a standard ANOVA. Repeatability of litter size was cal- culated, and the relationship between litter size and age of dam was analyzed by regres- sion of litter size on dam’s age. The average litter size in this sample was compared to the expected value of two using a Student’s t- test; the same comparison was made within each species. The average number of off- spring per litter surviving the first month was calculated for each litter size.
Results are presented by age interval ana- lyzed. Only a brief discussion of those covari- ates not associated with reproductive char- acters, Le., species, birth cohort, and sex, will be given, since they do not directly relate to the hypotheses investigated in this paper.
because it has been in the ORAU co P ony for

REPRODUCTION AND SURVIVORSHIP IN CALLITRICHIDAE 295
RESULTS 1. Whole life suruiuorship
Whole life survivorship analysis of all in- dividuals born into the colony (see Table 2; Fig. 1) showed that individuals born into litters of two had significantly higher survi- vorship (P < 0.001) than those born into triplet litters. Singlets also showed lower survivorship than twins, but it was not sig- nificant at the 0.05 level. Species showed a significant (P < 0.001) affect on lifelong sur- vivorship, with Callithrix jacchus having the best survivorship, followed by Saguinus oe- dipus and then Saguinus fuscicollis (see Fig. 2). The most recent time interval (1981- 1985) showed significantly (P < 0.05) higher survivorship than previous time intervals. Male and female survivorship was not signif- icantly different.
2. Survivorship during the first month of life
When survivorship analysis was performed only during the first month of life, singlets and triplets both show significantly (sin- glets: P < 0.05; triplets: P < 0.01) lower sur- vivorship than twins (Fig. 3). C. jacchus showed significantly (P < 0.001) higher sur- vivorship than Saguinus fuscicollis; survi- vorship of S. oedipus was not significantly different from that of S. fuscicollis (see Table 3).
Maternal characteristics, i.e., dam’s age and dam’s average litter size, did not show a significant effect on offspring survivorship during the first month of life (see Table 4).
Sample size for analysis of maternal effects on survivorship was restricted to only those individuals whose dam was born in the col- ony(N = 968).
Calculation of the number of offspring per litter surviving the first month revealed that on average, one out of two twins survived and one out of three triplets survived the first month. Approximately one-third (0.377) of all singlets survived the first month.
3. Survivorship during the first 2 years of life
Analysis of survivorship up to breeding age (Table 5; Fig. 4) showed the same rela- tionship between cohort litter size as in the whole life analysis, i.e., triplets having sig- nificantly lower survivorship when com- pared to twins. The same species effect on survivorship which was found in the whole life analysis was found up to breeding age; C. jacchus had significantly higher survivor- ship, followed by s. oedipus and s. fuscicol- lis, respectively.
4. Survivorship subsequent to the first month of life
Survivorship analysis of those individuals who survived their first month (Table 6 ) showed litter size not to be a significant factor in subsequent survivorship. Although not significant, singlets had the best survi- vorship when compared to twins and triplets had the worst (see Fig. 5). S. oedipus had the highest survivorship subsequent to the first month, followed by C. jacchus and S. fus-
TABLE 2. Cox Proportional Hazard regression coefficients for whole life survivorship analysis’
95% confidence
Covariate Sample interval
size Estimate2 STD Lower Upper Relative risk Time interval
1971-75 851 0.025 0.052 -0.078 0.127 0.975 1976-80 840 -0.030 0.063 -0.153 0.092 1.035 1981 -85 457 0.193* 0.085 0.027 0.359 0.825
C. jacchus 271 0.602** 0.092 0.422 0.783 0.548 S. oedipus 738 0.298** 0.060 0.181 0.416 0.742
One 458 -0.075 0.058 -0.189 0.039 1.078 Three 378 -0.473** 0.062 -0.594 -0.351 1.605
Female 1,170 0.037 0.044 -0.049 0.124 0.964 Unknown 414 -0.982** 0.065 -1.109 -0.855 2.670
Species
Litter size
Sex
‘N = 2,834. Wote: the sign of the covariate estimate was changed to reflect the interpreted effect of the covariate *Significant at 0.05 level. **Significant at 0.001 level.

296 C.E. JAQUISH ET AL
S U
V
V
r
i
a I
P r 0 b
1
0.8
0.6
0.4
0.2
c
SINGLETS A
+ TWINS
* TRIPLETS
0 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 age (years)
Fig. 1. Effect of litter size born into on whole life survivorship (N = 2,834). Triplets show significantly (P < 0.001) lower survivorship than singlets and twins,
especially during early age intervals. Note that age (x-axis) is given in years.
S U
V
V
r
i
a I
P r 0 b
0 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5
age (years) Fig. 2. Effect of species on whole life survivorship
(N = 2,834). S. oeclzppus and C.jacchus have significantly (P < 0.001) higher survivorship than S. fusczcolZis. Note that age (x-axis) is given in years.

REPRODUCTION AND SURVIVORSHIP IN CALLITRICHIDAE 297
S U
V
V
r
i
a I
P r 0 b
1
0.8
0.6
0.4
0.2
C 0 2 4 5 8 10 12 14 18 18 20 22 24 25 28 30
age (days) Fig, 3. Effect of litter size born into on survivorship
during the first month of life (N = 2,834). During the first month oflife, both singlets and triplets show signif- icantly (P < 0.01) lower survivorship than twins. Note
TABLE 3. Cox Proportional Hazard regression coefficients for survivorship analysis of all individuals during the first month of life’
95% confidence interval Sample
Covariate size Estimate2 STD Lower Upper Relative risk
Time interval 1971-75 851 1976-80 840
0.018 0.069 -0.116 0.153 -0.090 0.078 -0.243 0.063
0.982 1.094
1981-85 457 0.140 0.100 -0.057 0.336 0.869
C. jacchus 271 0.799** 0.127 0.549 1.049 0.450 S. oedipus 738 0.030 0.072 -0.112 0.171 0.970
Species
Litter size One 458 -0.159* 0.070 -0.297 -0.022 1.172 Three
SPX 378 -0.536** 0.071 -0.675 -0.397 1.709
- -.. Female 1,170 0.070 0.059 -0.045 0.185 0.932 Unknown 414 -0.954** 0.068 -1.088 -0.820 2.545
‘N = 2,834. *Note: the sign of the covariate estimate was changed to reflect the interpreted effect of the covariate *Significant at 0.05 level. **Significant at 0.001 level.

298 C.E. JAQUISH ET AL.
TABLE 4. Cox Proportional Hazard regression coefficients for survivorship during the first month of life when maternal characteristics are included as covariates'
95% confidence interval Sample
Covariate size Estimate2 STD Lower Upper Relative risk
Time interval 1971-75 326 0.175 0.159 -0.136 0.486 0.839 1976-80 306 0.253 0.165 -0.071 0.577 0.776 1981-85 289 0.639*** 0.187 -0.272 1.006 0.528
Species C. jacchus 133 0.473** 0.153 0.172 0.773 0.623 s. oediDus 158 -0.184 0.117 -0.412 0.045 1.202
Litter size One 125 -0.263* 0.107 -0.053 -0.474 1.301 Three 138 -0.366*** 0.104 -0.569 -0.162 1.442
Female 406 0.012 0.078 -0.141 0.165 0.988 Unknown 158 -0.753*** 0.106 -0.961 -0.544 2.123
Dam age 968 0.020 0.018 -0.015 0.056 0.980 Dam 968 -0.034 0.087 -0.205 0.137 1.035
Sex
Maternal characteristics
average litter size
'N = 968. "Note: the sign of the covariate estimate was changed to reflect the interpreted effect of the covariate. *Significant at 0.05 level. **Significant at 0.01 level. ***Significant at 0.001 level.
TABLE 5. Cox Proportional Hazard regression coefficients for survivorship analysis of all individuals during first two Years o f life'
95% confidence Sample interval
Covariate size estimate2 STD Lower Uuuer Relative risk
Time interval 1971-75 851 -0.049 0.059 -0.164 0.066 1.050 1976-80 840 -0.059 0.069 -0.194 0.076 1.061 1981-85 457 -0.156 0.082 -0.317 0.006 1.169
C. jacchus 271 0.454* 0.091 0.276 0.631 0.635 S. oedipus 738 0.206** 0.064 0.080 0.332 0.814
Species
Litter size One Three
458 -0.099 0.062 -0.220 0.022 1.104 378 -0.472*** 0.064 -0.597 -0.348 1.603
Sex Female 1,170 0.043 0.048 -0.052 0.137 0.958 Unknown 414 -1.001*** 0.065 -1.128 -0.874 2.721
'N = 2,834. 'Note: the sign of the covariate estimate was changed to reflect the interpreted effect of the covariate. *Significant at 0.05 level. **Significant at 0.01 level. ***Significant at 0.001 level.
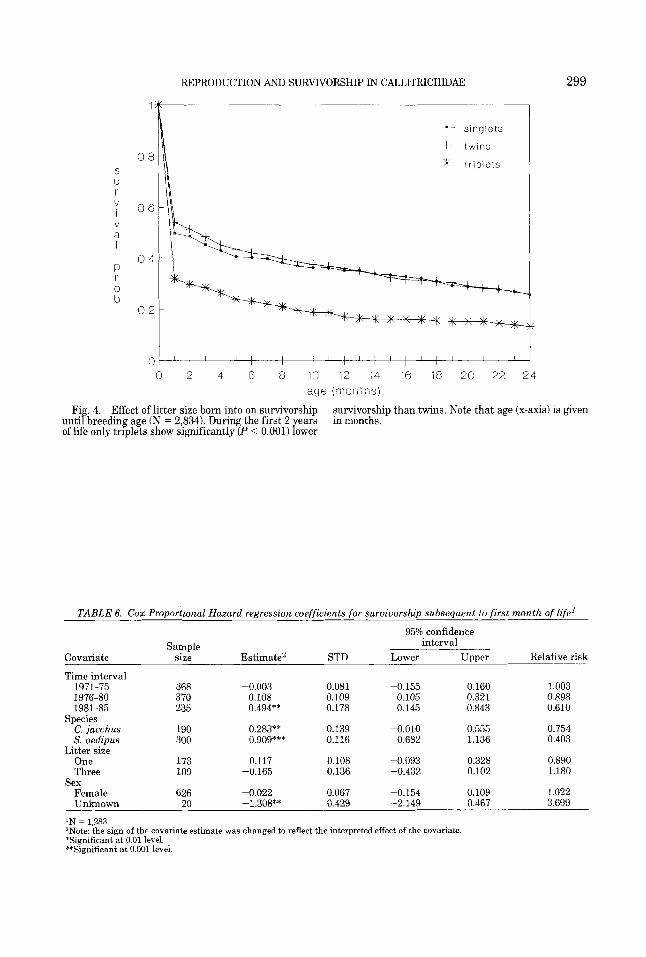
REPRODUCTION AND SURVIVORSHIP IN CALLITRICHIDAE 299
S U
V
V
r
i
a I
P r 0 b
1-
08
06
04
0 2
0
I singlets t
+ twins
+ triplets
I 1 I 1 1 l l i i l 1 I l l I I 1 1 I I I I I I I I I I I I
0 2 4 6 8 10 12 14 16 18 20 22 2 4 age (months)
survivorship than twins. Note that age (x-axis) is given in months.
Fi? 4. Effect of litter size born into on survivorship unti breeding age (N = 2,834). During the first 2 ears of life only triplets show significantly (P < 0.001) Lwer
TABLE 6. Cox Proportional Hazard regression coefficients for surviuorslup subsequent to first mon th o f life'
95% confidence interval Sample
Covariate size Estimate2 STD Lower Upper Relative risk
Time interval 1971-75 368 -0.003 0.081 -0.155 0.160 1.003 1976-80 370 0.108 0.109 -0.105 0.321 0.898 1981 -85 285 0.494** 0.178 0.145 0.843 0.610
Species C. jacchus 190 0.283** 0.139 -0.010 0.555 0.754 S. oedinus 300 0.909*+* 0.116 0.682 1.136 0.403
Litter size One Three
S P X
173 0.117 0.108 -0.093 0.328 0.890 109 -0.165 0.136 -0.432 0.102 1.180
I___
Female 626 -0.022 0.067 -0.154 0.109 1.022 Unknown 20 -1.308** 0.429 -2.149 -0.467 3.699
'N = 1,283. 2Note: the sign of the covariate estimate was changed to reflect the interpreted effect of the covariate *Significant at 0.01 level. **Significant at 0.001 level.

300
1 .o
08 S U r
06 V a
0 4 P r 0 b
0;:
0. c 0
C.E. JAQUISH ET AL.
- singlets
+ twins
++ tr ip le ts
,Z 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
age (years)
Fig. 5 . Effect of litter size born into on survivorship subsequent to the first month of life (N = 1,283). Subse- quent to the first month of life litter size born into no
cicollis, respectively. The most recent birth cohort showed significantly higher survivor- ship than earlier time intervals. 5. Survivorship subsequent to the first day
of life Survivorship analysis of all those individ-
uals which were definitely live births (those who survived their first day of life) revealed that singlets and twins did not show signifi- cantly different survivorship. Triplet survi- vorship remained significantly lower than twins (see Table 7; Fig. 6). S. oedipus showed significantly lower survivorship than the other species.
6. Dam survivorship Survivorship analysis of dams (Table 8)
revealed species and age at first litter to be significant (P < 0.05) factors affecting dam survivorship. Average litter size of the dam showed no relationship with dam survivor- ship. S. oedipus showed higher survivorship than other species. Age at first litter showed a positive relationship with survivorship, therefore, dams which started reproducing later in life showed higher survivorship. Birth cohort showed dams born between 1971 and 1975 to have significantly ( P <
longer has a significant effect on survivorship. Note that age (x-axis) is given in years.
0.05) better survivorship than those born in the preceding time interval. 7. Variation of litter size within the sample
Repeatability of litter size was 0.128 based on a minimum of two recorded litters per dam; when three recorded litters were used the repeatability rose to 0.304. Regression of litter size on dam’s age showed no significant (P > 0.05) relationship. Analysis of variance of litter size with species as the factor showed a significant (P < 0.05) amount of variance between species in litter size. The average litter size in the population was 1.845, a Student’s t-test showed this value to be significantly (P < 0.05, N = 1648) lower than the expected value of two. Analysis of average litter size by species using paired samples t-tests showed S. fuscicollis (av- erage litter size of 1.827) and S. oedipus (average litter size of 1.816) to have average litter sizes significantly (P < 0.05) less than two, while average litter size of C. jacchus (1.985) was not significantly less than two.
DISCUSSION Overall survivorship of individuals born
into the ORAU colony shows litter size, spe- cies, and birth cohort to be significant factors

REPRODUCTION AND SURVIVORSHIP IN CALLITRICHIDAE 301
TABLE 7. Cox Proportional Hazard regression coefficients for survivorship subsequent to first day of life'
95% confidence Sample interval
Covariate size Estimate2 STD Lower Umer Relative risk
Time interval 1971-75 466 -0.253 0.166 -0.578 0.073 1.288 1976-80 523 -0.486** 0.181 -0.841 -0.130 1.626 198 1-85 300 -0.008 0.219 -0.437 0.422 1.008
C. jacchus 236 0.277 0.198 -0.110 0.664 0.758 S. oedipus 441 -0.375* 0.145 -0.660 -0.091 1.455
One 216 -0.203 0.165 -0.527 0.121 1.259 Three 179 -0.791** 0.141 -1.067 -0.515 2.206
Species
Litter size
Sex Female 805 0.002 0.106 -0.206 0.209 0.998 Unknown 33 -0.956 0.303 -1.549 -0.363 2.601
'N = 1,658. *Note: the sign of the covariate estimate was changed to reflect the interpreted effect of the covariate. *Significant a t 0.01 level. '*Significant a t 0.001 level.
1
08 S U r
06 I V a I
P r 0 b
04
0 2 singleis
-+ twins
+ trip le ts
0 1 3 5 7 9 11 13 15 17 13 21 23 25 27 29
age (days) Fig. 6. Effect of litter size born into on survivorship
subsequent to the first da of life (N = 1,658). Subse- quent to the first day of life only triplets show signifi-
affecting survivorship. The lower survivor- ship of individuals of unknown sex was not biologically significant. These individuals were necessarily neonates, because they were first observed dead and in unidentifi- able condition; these individuals were in- cluded because they were an important source of neonatal mortality. The effects of litter size on survivorship will be discussed
cantly (P < 0.001) lower survivorship than twins. Note that age (x-axis) is given in years.
by age interval analyzed; the effects of the other covariates will be summarized at the end of the discussion.
Litter size was found to have a significant effect on survivorship perinatally (singlets and triplets with lower survivorship), but not subsequent to the first month of life. Subsequent to the first month singlet, survi- vorship is not statistically different from

302 C.E. JAQUISH ET AL.
TABLE 8. Cox Proportional Hazard regression coefficients for survivorship analysis of darns'
95% confidence Sample interval
Covariate size Estimate2 STD Lower Upper Relative risk
Dam age at 145 0.266** 0.091 0.087 0.444 0.766
Average litter 145 0.290 0.292 -0.283 0.863 0.748 1st litter
size - ~ ~ .
Time interval 1971-75 36 1976-80 46
0.575* 0.259 -0.068 1.082 -0.102 0.356 -0.799 0.595
0.562 1.107
1981-85 9 -1.874 1.120 -4.069 0.321 6.514 SDecies
C. jacchus 24 0.070 0.500 -0.910 1.050 0.932 5'. oedipus 36 0.772* 0.320 0.144 1.400 0.462
'N = 145. *Note: the sign of the covariate estimate was changed to reflect the interpreted effect of the covariate *Significant at 0.05 level. **Significant at 0.01 level.
twins which have significantly better survi- vorship than triplets. Thus, it does not ap- pear that being born into a singlet litter confers an advantage on an individual dur- ing the time spent within the family group (before breeding age).
One possible explanation for the lower than expected survivorship of singlets is that they are actually the remaining fetus of a twin pregnancy. Cases of singlet births would then occur when one embryo does not survive and is resorbed or expelled at partu- rition. There have not been many longitudi- nal studies of prenatal survivorship, but many studies have found evidence (blood chimerism, presence of two placentae and/or two corpora lutea) of loss of a co-twin in utero (Wislocki, 1939; Gengozian and Merritt, 1970; Chambers and Hearn, 1985; Tardif, personal communication). If ovulation and fertilization of two ova were standard, the singleton may not receive the advantage of increased birth weight and prenatal nutri- tion. But this situation would not confer a disadvantage on singlets when compared to twins, and therefore their survivorship should be approximately the same as twins. This survivorship pattern is what is seen if potential stillbirths are discounted from the population, suggesting a higher number of stillbirths in singlet litters as opposed to twins and triplets. The inclusion of near- term stillbirths could explain the apparent lower survivorship of singlets, and is in agreement with previous results showing higher rates of singlet stillbirths than to
twin or triplet stillbirths (Kilbourn et al., 1983; Tardif et al., 1984).
A possible explanation for the apparent high incidence of stillbirths in singlets is that due to increased body size, some singlets may be difficult births. Unpublished data from the ORAU colony (Tardif, personal communication) show twins to have signifi- cantly higher birth weight than triplets, but there are insufficient data on singlet birth weight from the colony. Chambers and Hearn (1985) have found an inverse relation- ship between birth weight and litter size. Twinning in the Callitrichidae has been ex- plained as an adaptation allowing small pri- mates to give birth to large-brained offspring (Leutenegger, 1973; Harvey and Clutton- Brock, 1985). Thus, if some singlets are larger at birth, passage of the head through the birth canal could be difficult, resulting in a large number of stillbirths. This would suggest that those singlets which have higher birth weights may not have an advantage because they are more likely to be stillbirths.
Individuals born into triplet litters should show lower birth weight coupled with inabil- ity of parents to care for three offspring, and therefore have lower survivorship than twins. Subsequent to the first month, defi- cits such as lower birth weight do not signif- icantly affect survivorship. Therefore, it ap- pears that selection related to maternal performance and sibling competition acts perinatally, rather than through breeding age. In general, the effects of maternal per- formance are very strong perinatally and

REPRODUCTION AND SURVIVORSHIP IN CALLITRICHIDAE 303
decline throughout development (Cheverud et al., 1983; Hanrahan and Eisen, 1974; Legates, 1972; Rutledge et al., 1972); if sur- vivorship is not analyzed at different points in development, it incorrectly appears that maternal and sib effects act until breeding age, as predicted by optimal litter size mod- els. This is due to the persistent effects of extremely high perinatal mortality and con- tributes to the problem of assuming propor- tional hazard differentials in this data set.
Analysis of average survivorship er litter
cally only one offspring survived the first month regardless of whether twins or trip- lets were born. One-third of all singletons survived the first month. Therefore, from the parental perspective, there was no difference in number of offspring surviving the perina- tal period when twins and triplets were com- pared; singlets would be the least productive since the entire litter is lost on average 63.3% of the time. Optimal litter size models would predict that twins are the most com- mon in the population because twins place less stress on the dam than triplets and thereby enable her to have higher survivor- ship. This was not found to be the case. Average litter size of dam showed no rela- tionship with dam survivorship (Table 7).
Analysis of average litter size in the popu- lation showed it to be significantly lower than the value of two predicted by optimal litter size theory. This could be explained by opposing responses to selection in the paren- tal and filial generation. Selection may act directly on the parental generation to in- crease litter size, thereby increasing the po- tential number of offspring which may sur- vive to weaning. A problem arises in that large litter sizes generally result in lower perinatal survivorship of offspring. The “litter size genes” carried by the mother indi- rectly affect the offspring through maternal performance and directly effect the offspring because they are carrying one-half of the maternal litter size genes. The indirect and direct genetic components are negatively correlated. Response to direct selection in the parental generation for increased litter size would potentially result in an increase; the correlated response in the filial genera- tion would result in a decrease through in- creased survivorship of offspring born into smaller litters. If selection for increased lit- ter size and increased offspring survivorship occur simultaneously (as in optimality mod- els), the negative correlated response of lit- ter size to direct selection on survivorship is
during the first month revealed t f: at typi-
potentially much stronger than the positive response of litter size to direct selection. This suggests that litter size would typically show a smaller value than predicted by optimal litter size models. It is possible that negative direct-maternal covariances are a cause of the mean litter size being lower than pre- dicted by optimality models. At this time, there are not sufficient data to directly test this hypothesis.
Repeatability of litter size was not differ- ent from previous results of other species (Hanrahan and Eisen, 1974; Falconer, 1981; Turner and Young, 1969); a value of 0.128 shows the character to be highly variable. Since repeatability is the proportion of vari- ance in litter size between dams to the total variance, it sets an upper limit for heritabil- ity (Falconer, 1981). Therefore, heritability of litter size cannot exceed 0.128; most esti- mates have been approximately 0.20 (Eisen, 1970; Falconer, 1981). This upper bound of heritability suggests a strong environmental component affecting litter size and very little genetic variation in dams for litter size. Se- lection on offspring phenotype appears to be a more plausible route for the evolution of litter size in callitrichids. Analysis of aver- age litter size by species was consistent with results of a study earlier in the colony’s history (Tardif et al., 1984).
Analysis of dam survivorship showed age at first litter to have a significant (P < 0.05) positive effect on survivorship. This suggests early reproduction to be costly in terms of optimality models; therefore there should be a trade-off between the cost of early repro- duction and the benefit of increased repro- ductive output. For management practices, this suggests that delaying breeding may increase survivorship and possibly reproduc- tive output of dams. Average litter size and dam survivorship did not show the expected inverse relationship. The lack of relationship between average litter size and dam survi- vorship could be due to the high plane of nutrition received by these animals when compared to those in the wild.
Species relationship with survivorship agrees with that found in previous studies (Tardif et al., 1984; Wolfe et al., 1975; Hamp- ton et al., 1978) and suggests that C.jacchus can withstand the colony environment bet- ter than Suguinus species and that more care must be taken in designing a colony environment for Saginus species. This is especially apparent earlier in life, since in analyses of survivorship later in life (subse- quent to the first month of life and the

304 C.E. JAQUISH ET AL
analysis of dams) S. oedipus had lower rela- tive risk than C. jacchus. Alternatively, Saguinus species may have lower natural survivorship, but there are currently insuffi- cient field data to test this hypothesis. Anal- ysis of variance showed C. jacchus to have higher average litter size than either Sagui- nus species. This is in agreement with a previous study of species differences in re- productive performance in the ORAU colony (Tardif et al., 19841, which found C.jacchus to have a higher percentage of twins and lower percentage of singletons than Sagui- nus species. This further suggests that C. jacchus is better suited to colony breeding.
Time interval showed a significant effect on whole life survivorship and survivorship subsequent to first month of life. The in- creased survivorship in the most recent time interval in these two analyses suggests that improvements in management are signifi- cantly increasing survivorship at later ages, rather than perinatally. This is consistent with changes in diet (to the BAM diet) in the late seventies, which have eliminated mar- moset wasting syndrome as a major cause of death in the colony (Clapp and Tardif, 1985).
Survivorship to breeding age has often been attributed to parental performance and sibling competition. It can be seen that par- titioning survivorship prior to breeding age reveals factors such as litter size affecting survivorship perinatally, rather than until reproductive age, as assumed in optimal lit- ter size models. Survivorship to breeding age is affected by several factors which act at different temporal points in development. Identification of the factors affecting survi- vorship during specific age intervals is the first step in understanding the mechanisms by which these survival patterns are pro- duced. This study has identified some of the factors affecting captive callitrichid survi- vorship, now the mechanisms must be inves- tigated.
ACKNOWLEDGMENTS
Part of this research has been supported by NIH grant R24 RR02022-05, RR02229, and HD25346. We gratefully acknowledge Bennett Dyke and James Bridges for prepar- ing the data sets of analysis, as well as Oak Ridge Associated Universities for access to the data. We also thank James Cheverud and Malcolm Dow for their assistance and suggestions regarding the analysis. A pre- liminary version of this paper was presented
at the 1989 AAPA meetings in San Diego (Am. J . Phys. Anthropol., 81:224).
LITERATURE CITED Bindon BM (1984) Reproductive biology of the Booroola
Merino sheep. Aust. J. Biol. Sci. 37:163-189. Cameron GN (1973) Effect of litter size on postnatal
growth and survival in the desert woodrat. J. Mammol. 5 4 m : 489-493.
Chambers PL, and Hearn J P (1985) Embryonic, foetal and placental development in the common marmoset monkey (Catlzthrix jacchus). J. Reprod. Fertil. 56:23- 32.
Cheverud JM, Leamy JL, Atchley WR, and Rutledge JJ (1983) Quantitative genetics and the evolution of on- togeny: I. Ontogenetic changes in quantitative genetic variance components in inbred mice. Genet. Res. Camb. 42:65-75.
Clapp N, and Tardif SD (1985) Marmoset husbandry and nutrition. Dig. Dis. Sci. 30:17-23.
Cleveland J, and Snowdon CT (1984) Social development during the first twenty weeks in the cotton-top tama- rin (Saguinus o.oedzpus). Anim. Behav. 32432444.
Cody M (1966) A general theory of clutch size. Evolution 20:174-184.
Eisen EJ (1970) Maternal effects on litter size in mice. Can. J . Genet. Cytol. 12:209-216.
Falconer DS (1981) Introduction to Quantitative Genet- ics, 2nd ed. New York: Longman Pub.
Gengozian N (1969) Marmosets: Their potential in ex- perimental medicine. Ann. N.Y. Acad. Sci. 162:336, 367.
Gengozian N, and Merritt CB (1970) Effect of unilateral ovariectomy on twinning frequency in the marmoset. J. Reprod. Fertil. 23509-512.
Hampton SH, Gross MJ, Hampton JK (1978) A compar- ison of captive breeding erformance and offspring survival in the Callitrichicfae. In Gengozian N., Dein- hardt F. (eds.): Primates in Medicine; vol. 10. Proc. of symposium sponsored by ORAU, Oak Ridge, Tenn., Karger, Basel pp. 88-95.
Hanrahan J P (1980) Ovulation rate as the selection criterion for litter size in sheep. Anim. Prod. Aust. 13:405408.
Hanrahan JP, and Eisen EJ (1974) Genetic variation in litter size and 12-day weight in mice and their rela- tionship with post-weaning growth. h i m . Prod.
Harvey PH, and Clutton-Brock TH (1985) Life history variation in primates. Evolution 39(3):559-581.
Elbourn JA, Sehgal P, Johnson LD, Beland M, and Bronson RT (1983) A retrospective study of infant mortality of cotton-top tamarins (Saguinus oedipus) in captive breeding. Lab. Anim. Sci. 33:168-171.
Krkwood JJ , Epstein MA, and Terlecki AJ (1983) Fac- tors influencing population growth of a colony of cot- ton-tor, tamarins. Ibis 17:3541.
19: 13-23.
Lack D (1947) The significance of clutch size. Lab. Anim. 89:302-352.
Lee ET (1980) Statistical Methods for Survival Data Analysis. Belmont, CA: Life Time Learning Pub.,
Legates JE (1972) The role of maternal effects in animal breeding: IV. maternal effect in laboratory species. J. Anim. Sci. 35(6):1294-1302.
Leutenegger W (1973) Maternal-fetal relationships in primates. Folia Primatol. (Basel) 20:283-290.
pp. 136-139.

REPRODUCTION AND SURVIVORSHIP IN CALLITRICHIDAE 305
Robison OW (1981) The influence of maternal effects on the efficiency of selection; a review. Livestock Prod. Sci. 8~121-137.
Rockwell RF, Findlay CS, and Cooke F (1987) Is there an optimal clutch size in snow geese? Am. Nat. 130(61:
Rutledge JJ, Robison OW, Eisen EJ, and Legates J E (1972) Dynamics of genetic and maternal effects on mice. J . Anim. Sci. 35(5):911-918.
Sussman RW, and Kinzey WG (1984) The ecological role of the callitrichidae: a review. Am. J. Phys. Anthropol. 64~419449.
Tardif SD, Richter CB, and Carson RL (1984) Reproduc- tive performance of three species of callitrichidae. Lab. h i m . Sci. 34:272-275.
Tardif SD, Carson RL, and Clapp NK (1986) Breeding performance of captive-born cotton-top tamarin (Saguirzus oedipus) females: proposed explanations for colony differences. Am. J. Primatol. 11:271-275.
839-863.
Turner HN, and Young SSY (1969) Quantitative Genet- ics in Sheep Breeding. Ithaca, NY: Cornell University Press, p. 240.
Willham RL (1972) The role ofmaternal effects in animal breeding. I11 Biometrical aspects of maternal effects in animals. J. Anim. Sci. 35~1288-1293.
Wislocki GB (1939) Observations on twinning in marmo- sets. Am. J. h a t . 64:445-483.
Wolfe LG, Deinhardt F, Ogden JD, Adams MR, Fisher LE (1975) Reproduction of wild-caught and labora- tory-born marmoset s p e s used in biomedical re- search (Saguinus sp., allithrix jacchus). Lab. Anim. Sci. 25:802-812.
Ziegler TE, Bridson WE, Snowdon CT, and Eman S (1987) Urinary gonadotropin and estrogen excretion during the postpartum estrus, conception and preg- nancy of the cotton-top tamarin (Saguinus oedipus oedipus). Am. J. Primatol. 12:127-140.




















