
Wound-induced polyploidization is dependent on Integrin-Yki ...

2006;66:3715-3725. Cancer Res Christian Spangenberg, Ekkehart U. Lausch, Tatjana M. Trost, et al. Under Adverse ConditionsIntegrin Fibronectin Receptor Promotes Tumor Cell Survival
1β5αERBB2-Mediated Transcriptional Up-regulation of the
Updated version
http://cancerres.aacrjournals.org/content/66/7/3715
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/66/7/3715.full.html#ref-list-1
This article cites by 73 articles, 35 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/66/7/3715.full.html#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from Research.
on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

ERBB2-Mediated Transcriptional Up-regulation of the A5B1
Integrin Fibronectin Receptor Promotes Tumor Cell
Survival Under Adverse Conditions
Christian Spangenberg,1Ekkehart U. Lausch,
1Tatjana M. Trost,
1Dirk Prawitt,
1Andreas May,
1
Romy Keppler,1Stephan A. Fees,
1Dirk Reutzel,
1Carolin Bell,
2Steffen Schmitt,
3
Ilka B. Schiffer,4Achim Weber,
5Walburgis Brenner,
6Matthias Hermes,
7
Ugur Sahin,2Ozlem Tureci,
2Heinz Koelbl,
4Jan G. Hengstler,
7
and Bernhard U. Zabel1
1Children’s Hospital, 2Department of Internal Medicine, 3FACS Core Facility, 4Department of Obstetrics and Gynecology, 5Institute ofPathology, 6Department of Urology, University of Mainz, Mainz, Germany; 7Center for Toxicology, Institute of Legal Medicine andRudolf-Boehm Institute of Pharmacology and Toxicology, University of Leipzig, Leipzig, Germany
Abstract
Oncogenic activation of the receptor tyrosine kinase ERBB2 isa key event in the development of a number of epithelialmalignancies. In these tumors, high levels of ERBB2 arestrongly associated with metastatic disease and poor progno-sis. Paradoxically, an inherent cellular response to hyper-mitogenic signaling by ERBB2 and other oncogenes seems tobe growth arrest, rather than proliferation. Molecularcharacterization of this yet undefined antiproliferative statein independent cell lines overexpressing either wild-typeERBB2 or the mutationally activated receptor unveiled adramatic induction of the A5B1 integrin fibronectin receptor.A5 Integrin up-regulation is mainly a transcriptional responsemediated by the hypoxia-inducible transcription factors (HIF),leading to a massive increase in membrane-resident receptormolecules and enhanced fibronectin adhesiveness of therespective cells. Functionally, ERBB2-dependent ligation offibronectin results in improved survival of mammary adeno-carcinoma cells under adverse conditions, like serum with-drawal, hypoxia, and chemotherapy. HIF-1A is an independentpredictor of poor overall survival in patients with breastcancer. In particular, HIF-1A overexpression correlatessignificantly with early local relapse and distant metastasis,a phenotype also highly characteristic of ERBB2-positivetumors. As HIF-1A is known to be stabilized by ERBB2signaling under normoxic conditions, we propose that A5integrin is a major effector in this regulatory circuit and mayrepresent the molecular basis for the HIF-1A-dependentaggressiveness observed in ERBB2-overexpressing breastcarcinomas. Hypermitogenic ERBB2 signaling and tumorhypoxia may act synergistically to favor the establishment ofchemoresistant dormant micrometastatic cells frequentlyobserved in patients with breast cancer. This new insightcould be the basis for additional approaches complementingcurrent cancer therapy. (Cancer Res 2006; 66(7): 3715-25)
Introduction
The ERBB2 proto-oncogene is a member of the EGFR genefamily consisting of ERBB1 (EGFR or HER1), ERBB2 (HER2 orNEU), ERBB3 (HER3), and ERBB4 (HER4). The genes encode type Ireceptor tyrosine kinases (RTK, except for ERBB3, which codes fora receptor harboring an impaired kinase-domain; ref. 1) that areinvolved in the transmission of proliferative as well as differenti-ation signals (2, 3). ERBB2 plays an important role in thedevelopment of human epithelial neoplasias and high levels ofERBB2 expression correlate with poor prognosis in breast andovarian carcinoma (4, 5). Overexpression of the receptor was notonly described in human breast and ovarian carcinomas (6), butalso in other types of cancer, e.g., bladder carcinoma (7), non–smallcell lung cancer (8), in tumors of the gastrointestinal tract (9), aswell as in other human malignancies (10). At least for breastcarcinogenesis, it is believed that oncogenic activation of ERBB2 isan early event because it could be found in preinvasive lesions aswell as in advanced tumors (11, 12). Aberrant ERBB2 activation isimplicated in the tumorigenic process through its influence onproliferation, survival of tumor cells, angiogenesis, invasive growth,metastatic behavior, and therapy resistance (12–15). However, asfor some other proto-oncogenes, like RAS and RAF, inherentcellular anticarcinogenic mechanisms have been identified pre-venting aberrant growth of affected cells and leading to aproliferative halt on oncogenic signaling (ref. 16; recent reviews:refs. 17, 18).Exploiting the tetracycline expression system, we have previously
shown that inducible hypermitogenic ERBB2 signaling results ingrowth arrest of MCF-7 breast carcinoma cells, and that the cyclin-dependent kinase inhibitor, P21, is an important mediator ofthis cellular response (16). Expression screening focused on theregulation of integrin extracellular matrix (ECM) receptors byoncogenic ERBB2 signaling revealed a strong up-regulation ofseveral a-integrin subunits, including a5 integrin, in response toERBB2 induction. Integrin ligation influences signal transductioncascades that promote cell proliferation, cell migration, cellsurvival, as well as angiogenesis. Unlike growth factor receptors,they lack intrinsic enzymatic activity but participate in signaling bycoclustering with kinases and adaptor proteins. The a5h1 integrinheterodimer constitutes the major cellular receptor for the ECMcomponent fibronectin. Although its effect in tumorigenesis seemsto be cell type–specific, there are several studies providing evidencefor a pro-tumorigenic role of a5h1 integrin engagement (ref. 19 and
Note: C. Spangenberg and E.U. Lausch contributed equally to this work.Requests for reprints: Christian Spangenberg, Children’s Hospital, University of
Mainz, Obere Zahlbacher Strasse 63, 55131 Mainz, Germany. Phone: 49-6131-393-3339;Fax: 49-6131-393-0227; E-mail: [email protected].
I2006 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-05-2823
www.aacrjournals.org 3715 Cancer Res 2006; 66: (7). April 1, 2006
Research Article
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

references therein). It has recently been shown that adhesion tofibronectin mediated by the heterodimer is responsible forimproved survival of different cell lines, including breast carcinomacells, under adverse conditions (20–23). Furthermore, a5h1 integrinfibronectin receptor ligation is the key mediator of cell survival inan in vitro bone marrow metastasis model in which breastcarcinoma cells undergo a transition to a nonproliferative, dormantstate in response to fibroblast growth factor 2 (FGF2) treatment(24). For patients with breast cancer, the presence of singledormant cells in their bone marrow is indicative of fatal relapseincluding metastatic disease in bone, and ERBB2 has beenimplicated in the survival of such disseminated cells (25, 26).In this work, we explore the molecular characteristics and
consequences of a hypermitogenic state induced by stringentlycontrolled overexpression of ERBB2 in MCF-7 breast carcinomacells. In expression analyses, we observe a strong induction ofITGA5 transcripts and protein in response to oncogenic ERBB2signaling. We characterize this regulatory interaction and investi-gate the functional effect on tumor cell physiology. Our experi-ments identify the regulation of cellular adhesion to fibronectin viahypoxia-inducible factor (HIF)-dependent a5h1 integrin inductionas a novel mechanism by which oncogenic ERBB2 signalingpromotes cellular survival under adverse conditions (serumdeprivation, hypoxia, chemotherapeutic drugs) in vitro , thus addinginsight to the processes of ERBB2-driven mammary carcinogenesisat the molecular level. This regulatory circuit helps to explainhitherto uncharacterized consequences of oncogenic ERBB2activation, especially during the early stages of the carcinogenicprocess, in which strong ERBB2 signals may lead to nondividing,latent tumor cells exhibiting improved survival characteristics.
Materials and Methods
Antibodies and reagents. ERBB2 (sc-284) and actin (sc-1616)-specificantibodies were obtained from Santa Cruz (Heidelberg, Germany), the a5integrin-specific antibody (610633) used for immunoblotting was purchased
from BD PharMingen (Heidelberg, Germany), and the h1 integrin-specific
antibody was purchased from BioGenex (San Ramon, CA). The a5 integrin-specific antibody used for flow cytometric detection as well as for function
blocking experiments was purchased from Calbiochem (Schwalbach,
Germany; integrin a5 fibronectin receptor Ab-1, clone P1D6). Doxycyclinewas used as the inducer of ERBB2/NeuT in all experiments. It was obtained
as the hydrochloride salt from Sigma (Munich, Germany) and stored as a
200 Ag/mL aqueous stock solution at �20jC. Human plasma fibronectin
was purchased from Calbiochem. RGD (GRGDSPK, Sigma) and RGE(GRGESP, Takara Bio Inc., distributed by Cambrex, Potsdam, Germany)
peptides were reconstituted in PBS according to the manufacturer’s
instructions. Inhibitors PD98059, SB203580, and wortmannin were pur-
chased from Sigma, SP600125 and GO6976 were obtained from Calbiochem.They were dissolved as stock solutions in DMSO and added to a final
concentration of 10 Amol/L (Wortmannin), 50 Amol/L (PD98059),
20 Amol/L (SB203580), 10 Amol/L (SP600125), and 5 Amol/L (GO6976).
Solutions of chemotherapeutic drugs [cisplatin (CDDP), 5-fluorouracil(5-FU)] were prepared by the pharmacy of the Mainz University Hospital.
Cell lines, cell culture, and transfections. The MCF-7/pTet-NeuT and
enhanced green fluorescent protein (EGFP) cell lines harboring doxycycline-inducible expression constructs respond to doxycycline administration with
expression of EGFP alone (MCF-7/pTet-EGFP) or in combination with a
mutationally activated ERBB2-isoform isolated from rat glioblastoma (MCF-
7/pTet-NeuT). They have been described in detail in ref. (16). MCF-7/pTet-ERBB2 cell lines expressing the human ERBB2 protein in response to
doxycycline treatment were generated similarly. All MCF-7-derived cell lines
were kept in DMEM/10% fetal bovine serum in humidified 5% CO2
atmosphere at 37jC during routine culture. 184B5 transformed mammary
epithelial cells derived from normal mammary tissue obtained from areduction mammoplasty were obtained from ATCC (CRL-8799) and
cultured as described for MCF-7 cells.
The wild-type ERBB2 cDNA was amplified using the following
oligonucleotides as primers (ERBB2/Nhe-For, 5V-AAAAAGCTAGCAGCCG-CAGTGAGCACCAT-3V; ERBB2/Nhe-Rev, 5V-AAAAAGCTAGCGGTTCACACT-GGCACGTC-3V; restriction sites introduced for cloning are underlined). The
resulting cDNA was digested and inserted into the NheI site of pINSpBI-
NeuT/EGFP following removal of the NeuT cDNA.Retroviral gene transfer was done using high-titer retroviral stocks
generated by transient calcium phosphate transfection of BING ampho-
tropic packaging cells with a pBabe-puro vector containing the NeuT cDNA
(pBabe-NeuT) or vector without insert (pBabe) as previously described(16). Early-passage 184B5 cells were incubated with titered retroviral super-
natants in the presence of 8 Ag/mL polybrene for 12 hours. Twenty-four
hours after infection, cells were selected for 5 days in the presence of2 Ag/mL puromycin. Individual cell lines emerging after 2 weeks were
analyzed for ERBB2 expression by immunoblotting.
Luciferase reporter gene constructs harboring relevant parts of the
5V-flanking region of the ITGA5 gene (as described in ref. 27) were generatedinserting a proofreading PCR-amplified DNA fragment of the respective
region (the primers used for amplification were ITGA5prom-F, 5V-GTTAC-TCGAGGTCTTGAACTCCTGGCCTCA-3V and ITGA5prom-R, 5V-GTTACTC-GAGCCGCTCTTCCCTGTCCTG-3V; restriction sites introduced for cloningare underlined) into the XhoI site of pGL2-basic (Promega, Mannheim,
Germany). All constructs were verified by sequence analysis.
Transfections for reporter gene analysis were carried out in 24-well tissue
culture vessels. One day before transfection, 3 � 104 cells per well were
plated for duplicate analyses. The next day, 900 AL of fresh DMEM/10% fetal
bovine serum (with or without doxycycline) per well were added, followed
by the addition of 1.5 Ag pGL2-DNA (pGL2-basic or pGL2-ITGA5), and
0.5 Ag of the secreted alkaline phosphatase (SEAP2) control plasmid
(BD Clontech, Heidelberg, Germany), which had been complexed with 5 ALFUGENE 6 transfection reagent (Roche, Mannheim, Germany) according to
the manufacturer’s instructions. To transfect human embryonic kidney
(HEK) cells, 1 Ag of pGL2-DNA (pGL2-basic or pGL2-ITGA5), 1 Ag of pBabe-DNA (pBabe or pBabe-NeuT), and 0.5 Ag of SEAP2 control plasmid were
treated with FUGENE 6 accordingly. Cotransfection experiments using von
Hippel-Lindau (VHL) tumor suppressor protein expression constructs were
carried out in analogy: 0.5 Ag pGL2-DNA (pGL2-basic or pGL2-ITGA5),
0.5 Ag pcDNA3 or pcDNA3-VHL, and 0.1 Ag SEAP2 control plasmid
complexed with 3 AL of FUGENE 6 transfection reagent were added to
NeuT48 cells 16 hours prior to doxycycline induction. Reporter gene assays
were done 48 hours after transfection (in the case of VHL cotransfection
experiments, reporter gene assays were done 48 hours after the addition of
doxycycline, i.e., 64 hours following transfection).
Luciferase assays were done using the firefly luciferase assay kit
(Promega). SEAP assays used the Great EscAPe SEAP chemiluminescence
assay kit (BD Clontech). Relative light units (RLU) were calculated as thequotient of luciferase and SEAP activities. All transfection experiments were
done at least thrice with similar results.
Quantitative real-time reverse transcription-PCR. RNA isolation,reverse transcription, and subsequent quantitative real-time reverse
transcription-PCR experiments were done essentially as described (16).
The primers used for amplification of ITGA5 were ITGA5-F (5V-GGGCCAA-GACTTTCTTGCAG-3V) and ITGA5-R (5V-TGAAGAATCCAAGCTTGTA-GAGG-3V). Expression of ITGB1 was quantified using the following
primers and cycling conditions: ITGB1-F (5V-TACTTGTGAAGCCAG-CAACG-3V), ITGB1-R (5V-GGGGTAATTTGTCCCGACTT-3V). Cycling condi-
tions were: for ITGA5 amplification 95jC for 2 seconds, 61jC for 5 seconds,72jC for 10 seconds, 40 cycles; for ITGB1 amplification 95jC for 2 seconds,
61jC for 5 seconds, 72jC for 13 seconds, 40 cycles; for pyruvate dehydro-
genase h (PDH) amplification 95jC for 2 seconds, 61jC for 5 seconds, 72jCfor 8 seconds, 40 cycles. Both programs were preceded by an initial
incubation for 10 minutes at 95jC. Relative quantification of cDNA
concentrations was done using the Data analysis function of the Roche
Molecular Biochemicals Light Cycler Software (V. 3.5). Integrin expression
Cancer Research
Cancer Res 2006; 66: (7). April 1, 2006 3716 www.aacrjournals.org
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

levels were calculated in relation to the expression level of PDH as thereference gene [primers for PDH amplification were PDH-F (5V-GGAGTT-GAATGTGAGGTGATAAA-3V) and PDH-R (5V-ACGCAGGACCTTCCATGAT-3V)]. For quantification using real-time reverse transcription-PCR, at least
two independent experiments with duplicate amplifications were done.Immunoblotting. Protein extract preparation, immunoblotting, and
antibody detection was done as described in ref. (16). a5- and h1 integrin-
specific antibodies were used as a 1:1,000 dilution.
Flow cytometry. For flow cytometric detection of a5 integrin,subconfluent cells that were grown for 48 hours in the presence or absence
of doxycycline in 75 cm2 tissue culture flasks were harvested and washed
twice with PBS/EDTA. After incubation in PBS/1% bovine serum albumin
(BSA) with the primary antibody specific for the a5 integrin protein (1:100dilution) for 30 minutes at room temperature, the cells were washed thrice
with PBS/1% BSA at 4jC. The secondary antibody (phycoerythrin-
conjugated goat anti-mouse IgG; obtained from Santa Cruz) was used ina 1:200 dilution in PBS/1% BSA and the resulting cell suspension was kept
at room temperature for 20 minutes. Fluorescence-activated cell sorting
(FACS) analysis (FACSCalibur, Becton Dickinson, Heidelberg, Germany) was
done after three additional washes in PBS/1% BSA at 4jC. The primaryantibody was omitted in controls to check for background fluorescence in
order to allow assessment of basal cell surface a5 integrin levels.
Adhesion assay and quantification of living cells. Fibronectin-coatingof tissue culture vessels was done by adding 50 AL of a fibronectin solution(50 Ag/mL) per well of a 96-well tissue culture plate and a 16-hour
incubation at 4jC. After removal of excess fibronectin solution, a 10 mg/mL
BSA in PBS solution was added, and the plates were kept at 25jC for anadditional hour. Subsequently, the wells were washed once with PBS and
directly used for adhesion or survival assays. For profiling and adhesion
assays, subconfluent cells were cultured in the presence or absence of
doxycycline for 48 hours, harvested by incubation in PBS/EDTA for f10minutes and washed once more in PBS/EDTA. Following an additional wash
in serum-free DMEM, the cells were resuspended at a final concentration of
2 � 105 cells per mL in DMEM.
Initial profiling experiments determining the expression of a-integrinsubunits in NeuT48 and EGFP control cells were done using the a-integrinmediated cell adhesion array (Chemicon, Hofheim, Germany) according to
the manufacturer’s instructions. Cells (2 � 104; 100 AL) were added to thereconstituted wells and allowed to bind to the wells coated with different
antibodies for 2 hours at 37jC. Following aspiration of the medium, wells
were washed twice with assay buffer, 100 AL crystal violet containing Cell
Stain Solution were added to each well and bound cells were incubated for5 minutes at room temperature. Subsequently, the staining solution was
removed, the cells were washed four times with deionized water, air-dried,
and cell-bound dye was extracted by the addition of 100 AL extraction
buffer to each well. The amount of solubilized dye was quantified using aMRX II Elisa reader (Dynex Technologies, Inc., Chantilly, VA).
For adhesion assays, 100 AL of the cell suspensions were plated as
quadruplicates in a 96-well tissue culture plate previously coated with
fibronectin, BSA, or uncoated, and left at 37jC for 1 hour. Subsequently,
nonadherent cells were removed by three washings with PBS, and 100 ALDMEM per well were added. To quantify adherent cells, 10 AL of WST-1
reagent (Roche) were added to the wells, and the plate was kept at 37jC for
1 hour, followed by photometric quantification in a MRX II Elisa reader
(Dynex). Quantification of plated cells by omitting the washings controlled
for differences in the number of respective cell populations.
To quantify the influence of ERBB2-dependent a5h1 integrin inductionon cell growth (as the combined effect of proliferation and survival) under
adverse conditions, cells grown in the presence or absence of doxycycline
for 30 hours were harvested and washed as described for adhesion assays.5 � 103 cells (100 AL) per well were plated as triplicates in pretreated
96-well tissue culture vessels (day 0) and kept in DMEM (with or without
doxycycline) in a 5% CO2-humidified atmosphere or hypoxia (5% CO2, 0.1%
O2, 94.5% N2). At the indicated time periods, 10 AL of WST-1 reagent(Roche) were added to quantify living cells. Wells of the plating control were
quantified directly following plating on day 0. The specificity of a5h1integrin ligation on cell growth kinetics was shown by adhesion blockade,
preincubating the cells either with a function-blocking monoclonalantibody against a5 integrin (clone P1D6), or nonspecific isotype control
antibody for 30 minutes at 37jC before plating. Fibronectin specificity was
documented by adhesion blockade, preincubating the cells either with an
RGD- or with an RGE-containing peptide.For the analysis of chemoresistance, experimental media containing
either specific chemotherapeutic agents dissolved in serum-reduced
OptiMEM I (Invitrogen, Karlsruhe, Germany) or control media without
drugs were added after cell adherence. Serial dilutions were done todetermine and span the effective dose range for each drug. Drugs in use for
clinical breast cancer were selected, including antimetabolites (5-FU) and
platinum compounds (CDDP). Following incubation for 72 hours at 37jCin a humidified atmosphere containing 5% CO2, plates were analyzed byadding the WST-1 reagent as described above. The fraction of surviving cells
relative to control were plotted against the log of drug concentration, and
the IC50 was interpolated from the sigmoidal curve resulting from nonlinearregression applying a modified Hill model (28). All assays described were
done at least thrice with similar results.
Results
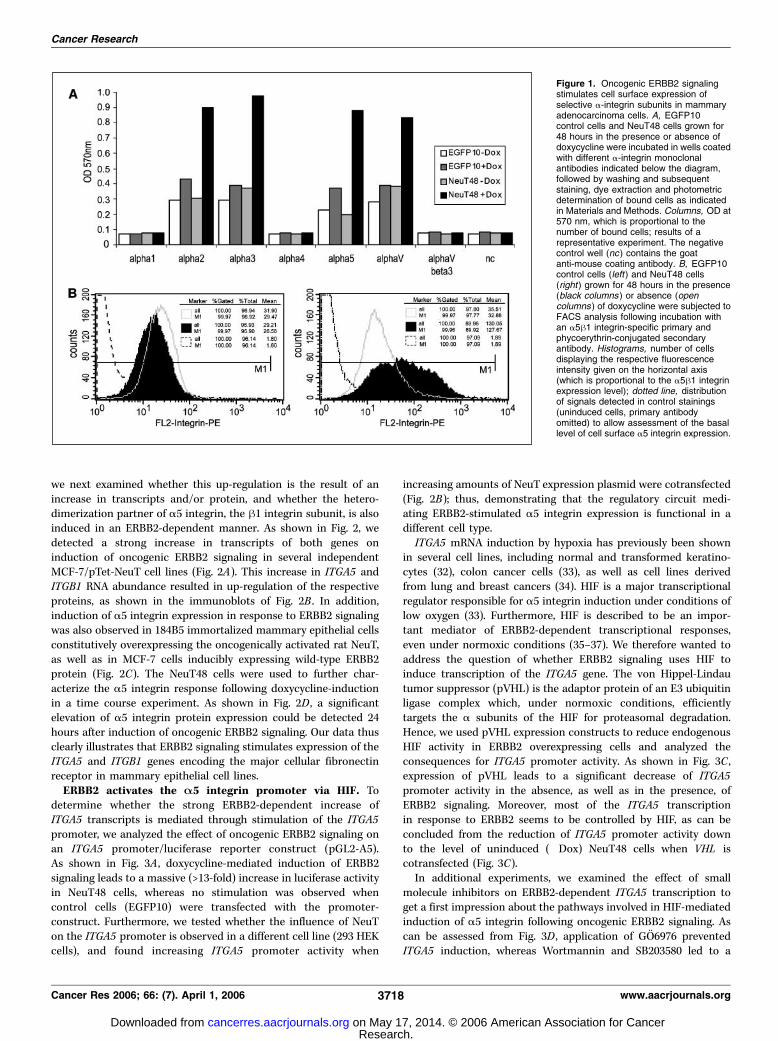
Identification of A-integrins as targets of oncogenic ERBB2signaling. Specific interactions of tumor cells and surroundingECM via cellular integrin receptors have been shown to haveprofound effects on carcinogenesis due to the modulation ofcellular variables influencing tumor cell physiology, like prolifera-tion, survival, as well as invasion and metastatic spread. Recentevidence suggests that interactions between integrin ECMreceptors and growth factor receptors play a substantial role incarcinogenesis (29–31). Because overexpression of the RTK ERBB2is causally involved in the development of f30% of mammarycarcinomas as well as several other epithelial malignancies, weexamined whether aberrant ERBB2 signaling results in thederegulation of integrin receptor expression in tumor cells. Wetherefore analyzed MCF-7 cells conditionally expressing ERBB2 (16)for an ERBB2-dependent response of a subset of relevant a-integrinsubunits. As shown in Fig. 1, doxycycline administration led toselective up-regulation of a2, a3, a5, and aV integrin proteins onlyin NeuT48 cells, suggesting a role for oncogenic ERBB2 signalingin regulating the tumor cell integrin receptor repertoire. Specificadhesion to fibronectin via the a5h1 integrin heterodimer hasrecently been shown to promote survival of nonproliferative,dormant breast carcinoma cells under conditions of bone marrowmetastasis (24). Given this apparent analogy to our observationsregarding the effect of oncogenic ERBB2 signal transduction inmammary adenocarcinoma cells (i.e., RTK-signaling provoking aproliferative arrest and a concomitant regulation of integrins)we looked more carefully for expression of the a5h1 integrinfibronectin receptor. We therefore analyzed induced and unin-duced NeuT48 as well as EGFP10 control cells by flow cytometryfor extracellular a5 integrin. As depicted in the histogram plot ofFig. 1B , a strong increase in membrane-resident a5 integrin proteincould be detected in NeuT48 cells in response to hypermitogenicERBB2 signaling: The mean relative fluorescence of doxycycline-induced NeuT48 cells was 127.67, nearly 4-fold higher comparedwith uninduced NeuT48 or EGFP10 control cells. Taken together,these results clearly show that oncogenic ERBB2 signaling leads toa selective up-regulation of specific a-integrin subunits in theplasma membrane of breast carcinoma cells, including the a5h1integrin fibronectin receptor.Identification of ITGA5 and ITGB1 as target genes of
oncogenic ERBB2 signaling in mammary adenocarcinomacells. Given the increase of membrane-resident a5 integrin protein,
ERBB2 Induces the a5b1 Integrin Fibronectin Receptor
www.aacrjournals.org 3717 Cancer Res 2006; 66: (7). April 1, 2006
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
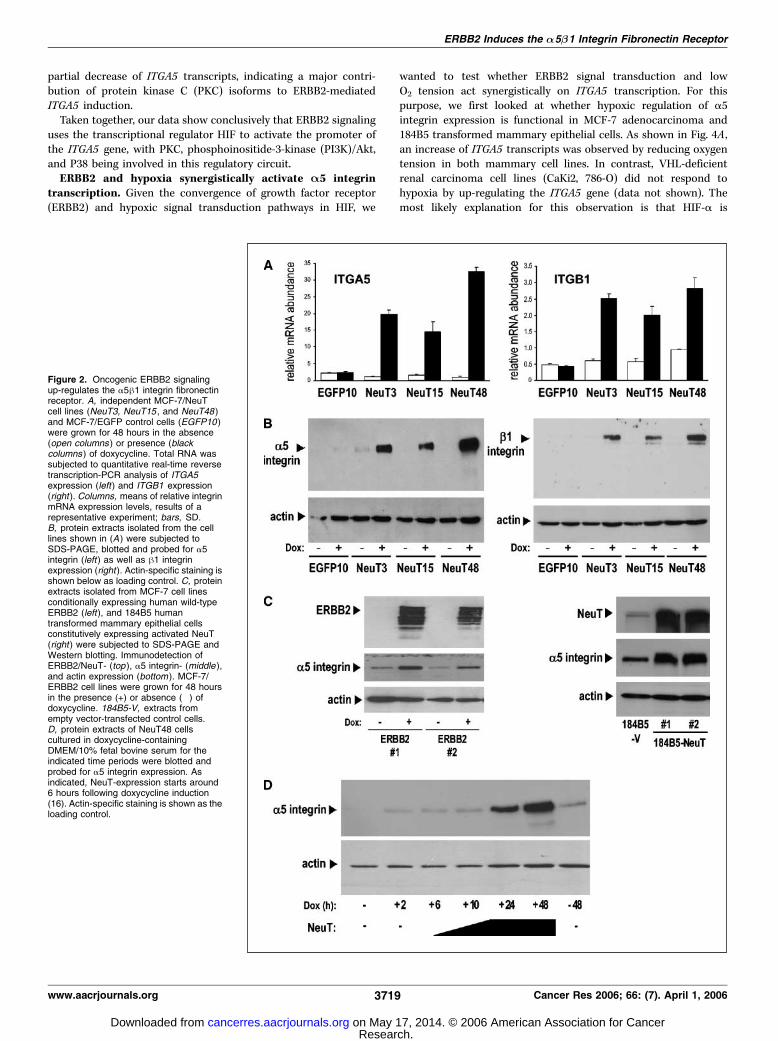
we next examined whether this up-regulation is the result of anincrease in transcripts and/or protein, and whether the hetero-dimerization partner of a5 integrin, the h1 integrin subunit, is alsoinduced in an ERBB2-dependent manner. As shown in Fig. 2, wedetected a strong increase in transcripts of both genes oninduction of oncogenic ERBB2 signaling in several independentMCF-7/pTet-NeuT cell lines (Fig. 2A). This increase in ITGA5 andITGB1 RNA abundance resulted in up-regulation of the respectiveproteins, as shown in the immunoblots of Fig. 2B . In addition,induction of a5 integrin expression in response to ERBB2 signalingwas also observed in 184B5 immortalized mammary epithelial cellsconstitutively overexpressing the oncogenically activated rat NeuT,as well as in MCF-7 cells inducibly expressing wild-type ERBB2protein (Fig. 2C). The NeuT48 cells were used to further char-acterize the a5 integrin response following doxycycline-inductionin a time course experiment. As shown in Fig. 2D , a significantelevation of a5 integrin protein expression could be detected 24hours after induction of oncogenic ERBB2 signaling. Our data thusclearly illustrates that ERBB2 signaling stimulates expression of theITGA5 and ITGB1 genes encoding the major cellular fibronectinreceptor in mammary epithelial cell lines.ERBB2 activates the A5 integrin promoter via HIF. To
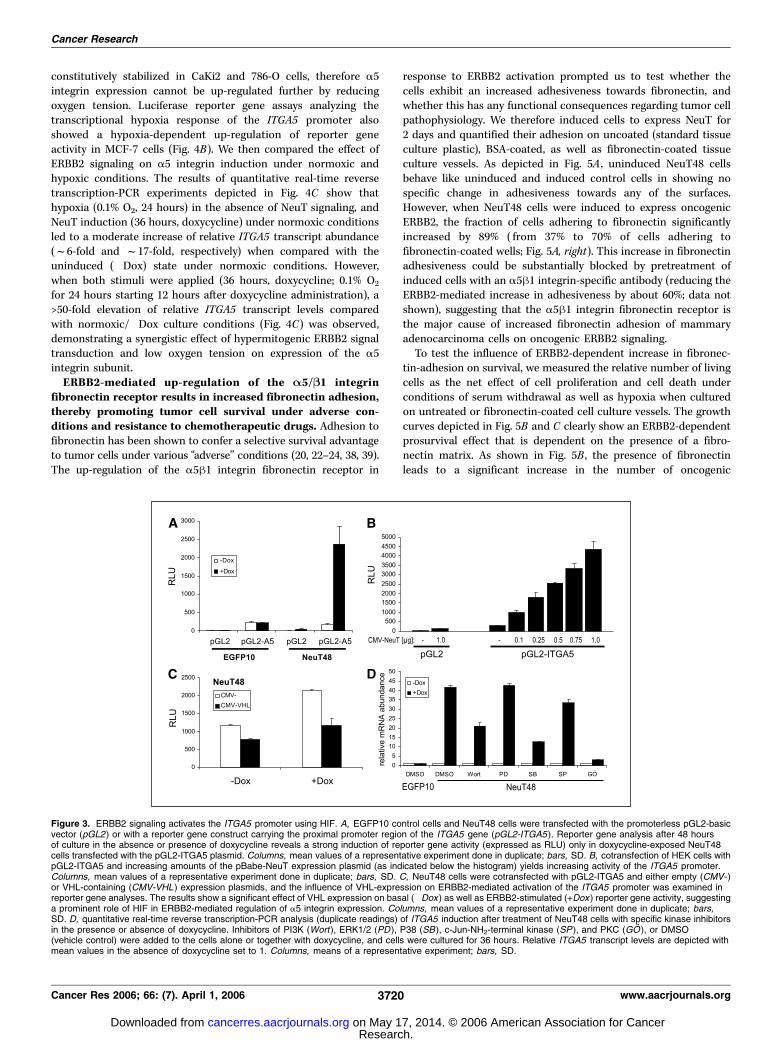
determine whether the strong ERBB2-dependent increase ofITGA5 transcripts is mediated through stimulation of the ITGA5promoter, we analyzed the effect of oncogenic ERBB2 signaling onan ITGA5 promoter/luciferase reporter construct (pGL2-A5).As shown in Fig. 3A , doxycycline-mediated induction of ERBB2signaling leads to a massive (>13-fold) increase in luciferase activityin NeuT48 cells, whereas no stimulation was observed whencontrol cells (EGFP10) were transfected with the promoter-construct. Furthermore, we tested whether the influence of NeuTon the ITGA5 promoter is observed in a different cell line (293 HEKcells), and found increasing ITGA5 promoter activity when
increasing amounts of NeuT expression plasmid were cotransfected(Fig. 2B); thus, demonstrating that the regulatory circuit medi-ating ERBB2-stimulated a5 integrin expression is functional in adifferent cell type.
ITGA5 mRNA induction by hypoxia has previously been shownin several cell lines, including normal and transformed keratino-cytes (32), colon cancer cells (33), as well as cell lines derivedfrom lung and breast cancers (34). HIF is a major transcriptionalregulator responsible for a5 integrin induction under conditions oflow oxygen (33). Furthermore, HIF is described to be an impor-tant mediator of ERBB2-dependent transcriptional responses,even under normoxic conditions (35–37). We therefore wanted toaddress the question of whether ERBB2 signaling uses HIF toinduce transcription of the ITGA5 gene. The von Hippel-Lindautumor suppressor (pVHL) is the adaptor protein of an E3 ubiquitinligase complex which, under normoxic conditions, efficientlytargets the a subunits of the HIF for proteasomal degradation.Hence, we used pVHL expression constructs to reduce endogenousHIF activity in ERBB2 overexpressing cells and analyzed theconsequences for ITGA5 promoter activity. As shown in Fig. 3C ,expression of pVHL leads to a significant decrease of ITGA5promoter activity in the absence, as well as in the presence, ofERBB2 signaling. Moreover, most of the ITGA5 transcriptionin response to ERBB2 seems to be controlled by HIF, as can beconcluded from the reduction of ITGA5 promoter activity downto the level of uninduced (�Dox) NeuT48 cells when VHL iscotransfected (Fig. 3C).In additional experiments, we examined the effect of small
molecule inhibitors on ERBB2-dependent ITGA5 transcription toget a first impression about the pathways involved in HIF-mediatedinduction of a5 integrin following oncogenic ERBB2 signaling. Ascan be assessed from Fig. 3D , application of GO6976 preventedITGA5 induction, whereas Wortmannin and SB203580 led to a
Figure 1. Oncogenic ERBB2 signalingstimulates cell surface expression ofselective a-integrin subunits in mammaryadenocarcinoma cells. A, EGFP10control cells and NeuT48 cells grown for48 hours in the presence or absence ofdoxycycline were incubated in wells coatedwith different a-integrin monoclonalantibodies indicated below the diagram,followed by washing and subsequentstaining, dye extraction and photometricdetermination of bound cells as indicatedin Materials and Methods. Columns, OD at570 nm, which is proportional to thenumber of bound cells; results of arepresentative experiment. The negativecontrol well (nc ) contains the goatanti-mouse coating antibody. B, EGFP10control cells (left ) and NeuT48 cells(right ) grown for 48 hours in the presence(black columns ) or absence (opencolumns ) of doxycycline were subjected toFACS analysis following incubation withan a5h1 integrin-specific primary andphycoerythrin-conjugated secondaryantibody. Histograms, number of cellsdisplaying the respective fluorescenceintensity given on the horizontal axis(which is proportional to the a5h1 integrinexpression level); dotted line, distributionof signals detected in control stainings(uninduced cells, primary antibodyomitted) to allow assessment of the basallevel of cell surface a5 integrin expression.
Cancer Research
Cancer Res 2006; 66: (7). April 1, 2006 3718 www.aacrjournals.org
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
partial decrease of ITGA5 transcripts, indicating a major contri-bution of protein kinase C (PKC) isoforms to ERBB2-mediatedITGA5 induction.Taken together, our data show conclusively that ERBB2 signaling
uses the transcriptional regulator HIF to activate the promoter ofthe ITGA5 gene, with PKC, phosphoinositide-3-kinase (PI3K)/Akt,and P38 being involved in this regulatory circuit.ERBB2 and hypoxia synergistically activate A5 integrin
transcription. Given the convergence of growth factor receptor(ERBB2) and hypoxic signal transduction pathways in HIF, we
wanted to test whether ERBB2 signal transduction and lowO2 tension act synergistically on ITGA5 transcription. For thispurpose, we first looked at whether hypoxic regulation of a5integrin expression is functional in MCF-7 adenocarcinoma and184B5 transformed mammary epithelial cells. As shown in Fig. 4A ,an increase of ITGA5 transcripts was observed by reducing oxygentension in both mammary cell lines. In contrast, VHL-deficientrenal carcinoma cell lines (CaKi2, 786-O) did not respond tohypoxia by up-regulating the ITGA5 gene (data not shown). Themost likely explanation for this observation is that HIF-a is
Figure 2. Oncogenic ERBB2 signalingup-regulates the a5h1 integrin fibronectinreceptor. A, independent MCF-7/NeuTcell lines (NeuT3, NeuT15 , and NeuT48 )and MCF-7/EGFP control cells (EGFP10 )were grown for 48 hours in the absence(open columns ) or presence (blackcolumns ) of doxycycline. Total RNA wassubjected to quantitative real-time reversetranscription-PCR analysis of ITGA5expression (left) and ITGB1 expression(right ). Columns, means of relative integrinmRNA expression levels, results of arepresentative experiment; bars, SD.B, protein extracts isolated from the celllines shown in (A ) were subjected toSDS-PAGE, blotted and probed for a5integrin (left ) as well as h1 integrinexpression (right ). Actin-specific staining isshown below as loading control. C, proteinextracts isolated from MCF-7 cell linesconditionally expressing human wild-typeERBB2 (left), and 184B5 humantransformed mammary epithelial cellsconstitutively expressing activated NeuT(right ) were subjected to SDS-PAGE andWestern blotting. Immunodetection ofERBB2/NeuT- (top ), a5 integrin- (middle ),and actin expression (bottom ). MCF-7/ERBB2 cell lines were grown for 48 hoursin the presence (+) or absence (�) ofdoxycycline. 184B5-V, extracts fromempty vector-transfected control cells.D, protein extracts of NeuT48 cellscultured in doxycycline-containingDMEM/10% fetal bovine serum for theindicated time periods were blotted andprobed for a5 integrin expression. Asindicated, NeuT-expression starts around6 hours following doxycycline induction(16). Actin-specific staining is shown as theloading control.
ERBB2 Induces the a5b1 Integrin Fibronectin Receptor
www.aacrjournals.org 3719 Cancer Res 2006; 66: (7). April 1, 2006
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
constitutively stabilized in CaKi2 and 786-O cells, therefore a5integrin expression cannot be up-regulated further by reducingoxygen tension. Luciferase reporter gene assays analyzing thetranscriptional hypoxia response of the ITGA5 promoter alsoshowed a hypoxia-dependent up-regulation of reporter geneactivity in MCF-7 cells (Fig. 4B). We then compared the effect ofERBB2 signaling on a5 integrin induction under normoxic andhypoxic conditions. The results of quantitative real-time reversetranscription-PCR experiments depicted in Fig. 4C show thathypoxia (0.1% O2, 24 hours) in the absence of NeuT signaling, andNeuT induction (36 hours, doxycycline) under normoxic conditionsled to a moderate increase of relative ITGA5 transcript abundance(f6-fold and f17-fold, respectively) when compared with theuninduced (�Dox) state under normoxic conditions. However,when both stimuli were applied (36 hours, doxycycline; 0.1% O2
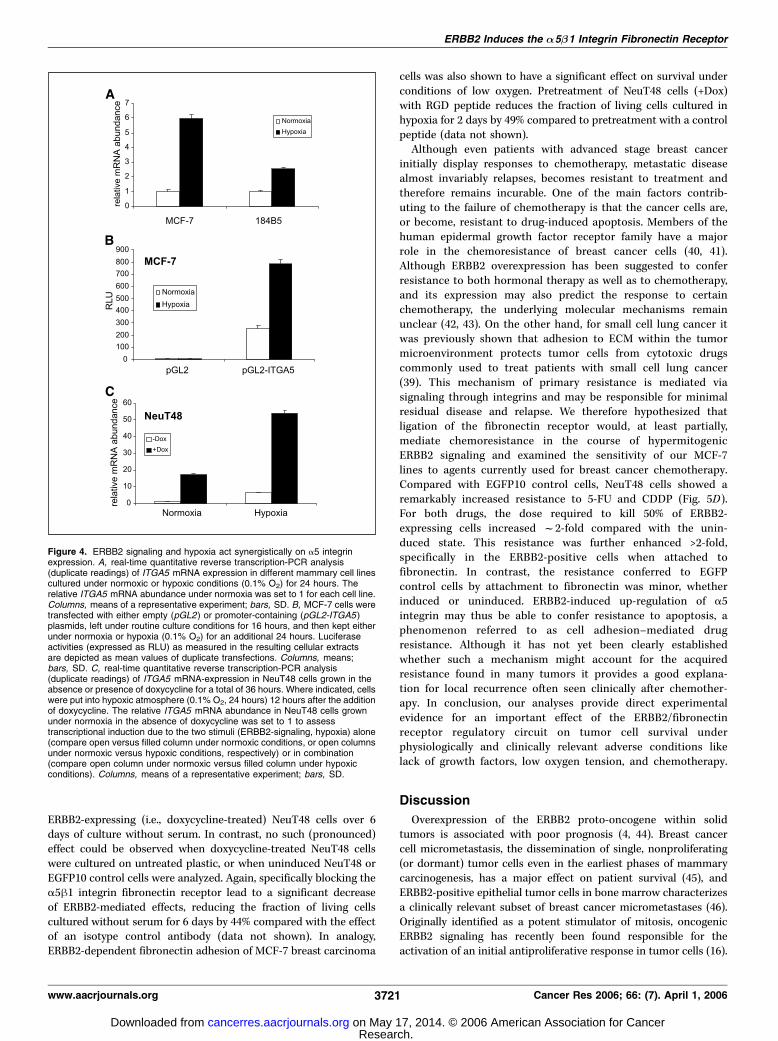
for 24 hours starting 12 hours after doxycycline administration), a>50-fold elevation of relative ITGA5 transcript levels comparedwith normoxic/�Dox culture conditions (Fig. 4C) was observed,demonstrating a synergistic effect of hypermitogenic ERBB2 signaltransduction and low oxygen tension on expression of the a5integrin subunit.ERBB2-mediated up-regulation of the A5/B1 integrin
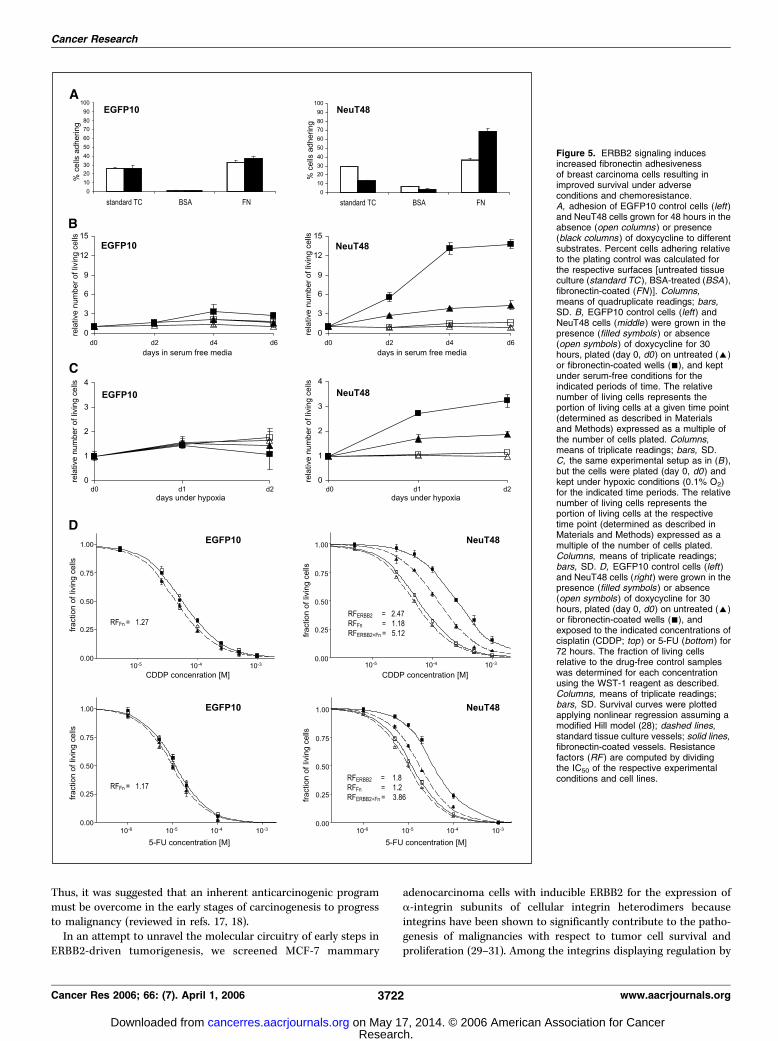
fibronectin receptor results in increased fibronectin adhesion,thereby promoting tumor cell survival under adverse con-ditions and resistance to chemotherapeutic drugs. Adhesion tofibronectin has been shown to confer a selective survival advantageto tumor cells under various ‘‘adverse’’ conditions (20, 22–24, 38, 39).The up-regulation of the a5h1 integrin fibronectin receptor in
response to ERBB2 activation prompted us to test whether thecells exhibit an increased adhesiveness towards fibronectin, andwhether this has any functional consequences regarding tumor cellpathophysiology. We therefore induced cells to express NeuT for2 days and quantified their adhesion on uncoated (standard tissueculture plastic), BSA-coated, as well as fibronectin-coated tissueculture vessels. As depicted in Fig. 5A , uninduced NeuT48 cellsbehave like uninduced and induced control cells in showing nospecific change in adhesiveness towards any of the surfaces.However, when NeuT48 cells were induced to express oncogenicERBB2, the fraction of cells adhering to fibronectin significantlyincreased by 89% ( from 37% to 70% of cells adhering tofibronectin-coated wells; Fig. 5A, right). This increase in fibronectinadhesiveness could be substantially blocked by pretreatment ofinduced cells with an a5h1 integrin-specific antibody (reducing theERBB2-mediated increase in adhesiveness by about 60%; data notshown), suggesting that the a5h1 integrin fibronectin receptor isthe major cause of increased fibronectin adhesion of mammaryadenocarcinoma cells on oncogenic ERBB2 signaling.To test the influence of ERBB2-dependent increase in fibronec-
tin-adhesion on survival, we measured the relative number of livingcells as the net effect of cell proliferation and cell death underconditions of serum withdrawal as well as hypoxia when culturedon untreated or fibronectin-coated cell culture vessels. The growthcurves depicted in Fig. 5B and C clearly show an ERBB2-dependentprosurvival effect that is dependent on the presence of a fibro-nectin matrix. As shown in Fig. 5B , the presence of fibronectinleads to a significant increase in the number of oncogenic
Figure 3. ERBB2 signaling activates the ITGA5 promoter using HIF. A, EGFP10 control cells and NeuT48 cells were transfected with the promoterless pGL2-basicvector (pGL2) or with a reporter gene construct carrying the proximal promoter region of the ITGA5 gene (pGL2-ITGA5 ). Reporter gene analysis after 48 hoursof culture in the absence or presence of doxycycline reveals a strong induction of reporter gene activity (expressed as RLU) only in doxycycline-exposed NeuT48cells transfected with the pGL2-ITGA5 plasmid. Columns, mean values of a representative experiment done in duplicate; bars, SD. B, cotransfection of HEK cells withpGL2-ITGA5 and increasing amounts of the pBabe-NeuT expression plasmid (as indicated below the histogram) yields increasing activity of the ITGA5 promoter.Columns, mean values of a representative experiment done in duplicate; bars, SD. C, NeuT48 cells were cotransfected with pGL2-ITGA5 and either empty (CMV- )or VHL-containing (CMV-VHL ) expression plasmids, and the influence of VHL-expression on ERBB2-mediated activation of the ITGA5 promoter was examined inreporter gene analyses. The results show a significant effect of VHL expression on basal (�Dox ) as well as ERBB2-stimulated (+Dox ) reporter gene activity, suggestinga prominent role of HIF in ERBB2-mediated regulation of a5 integrin expression. Columns, mean values of a representative experiment done in duplicate; bars,SD. D, quantitative real-time reverse transcription-PCR analysis (duplicate readings) of ITGA5 induction after treatment of NeuT48 cells with specific kinase inhibitorsin the presence or absence of doxycycline. Inhibitors of PI3K (Wort ), ERK1/2 (PD ), P38 (SB), c-Jun-NH2-terminal kinase (SP ), and PKC (GO ), or DMSO(vehicle control) were added to the cells alone or together with doxycycline, and cells were cultured for 36 hours. Relative ITGA5 transcript levels are depicted withmean values in the absence of doxycycline set to 1. Columns, means of a representative experiment; bars, SD.
Cancer Research
Cancer Res 2006; 66: (7). April 1, 2006 3720 www.aacrjournals.org
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
ERBB2-expressing (i.e., doxycycline-treated) NeuT48 cells over 6days of culture without serum. In contrast, no such (pronounced)effect could be observed when doxycycline-treated NeuT48 cellswere cultured on untreated plastic, or when uninduced NeuT48 orEGFP10 control cells were analyzed. Again, specifically blocking thea5h1 integrin fibronectin receptor lead to a significant decreaseof ERBB2-mediated effects, reducing the fraction of living cellscultured without serum for 6 days by 44% compared with the effectof an isotype control antibody (data not shown). In analogy,ERBB2-dependent fibronectin adhesion of MCF-7 breast carcinoma
cells was also shown to have a significant effect on survival underconditions of low oxygen. Pretreatment of NeuT48 cells (+Dox)with RGD peptide reduces the fraction of living cells cultured inhypoxia for 2 days by 49% compared to pretreatment with a controlpeptide (data not shown).Although even patients with advanced stage breast cancer
initially display responses to chemotherapy, metastatic diseasealmost invariably relapses, becomes resistant to treatment andtherefore remains incurable. One of the main factors contrib-uting to the failure of chemotherapy is that the cancer cells are,or become, resistant to drug-induced apoptosis. Members of thehuman epidermal growth factor receptor family have a majorrole in the chemoresistance of breast cancer cells (40, 41).Although ERBB2 overexpression has been suggested to conferresistance to both hormonal therapy as well as to chemotherapy,and its expression may also predict the response to certainchemotherapy, the underlying molecular mechanisms remainunclear (42, 43). On the other hand, for small cell lung cancer itwas previously shown that adhesion to ECM within the tumormicroenvironment protects tumor cells from cytotoxic drugscommonly used to treat patients with small cell lung cancer(39). This mechanism of primary resistance is mediated viasignaling through integrins and may be responsible for minimalresidual disease and relapse. We therefore hypothesized thatligation of the fibronectin receptor would, at least partially,mediate chemoresistance in the course of hypermitogenicERBB2 signaling and examined the sensitivity of our MCF-7lines to agents currently used for breast cancer chemotherapy.Compared with EGFP10 control cells, NeuT48 cells showed aremarkably increased resistance to 5-FU and CDDP (Fig. 5D).For both drugs, the dose required to kill 50% of ERBB2-expressing cells increased f2-fold compared with the unin-duced state. This resistance was further enhanced >2-fold,specifically in the ERBB2-positive cells when attached tofibronectin. In contrast, the resistance conferred to EGFPcontrol cells by attachment to fibronectin was minor, whetherinduced or uninduced. ERBB2-induced up-regulation of a5integrin may thus be able to confer resistance to apoptosis, aphenomenon referred to as cell adhesion–mediated drugresistance. Although it has not yet been clearly establishedwhether such a mechanism might account for the acquiredresistance found in many tumors it provides a good explana-tion for local recurrence often seen clinically after chemother-apy. In conclusion, our analyses provide direct experimentalevidence for an important effect of the ERBB2/fibronectinreceptor regulatory circuit on tumor cell survival underphysiologically and clinically relevant adverse conditions likelack of growth factors, low oxygen tension, and chemotherapy.
Discussion
Overexpression of the ERBB2 proto-oncogene within solidtumors is associated with poor prognosis (4, 44). Breast cancercell micrometastasis, the dissemination of single, nonproliferating(or dormant) tumor cells even in the earliest phases of mammarycarcinogenesis, has a major effect on patient survival (45), andERBB2-positive epithelial tumor cells in bone marrow characterizesa clinically relevant subset of breast cancer micrometastases (46).Originally identified as a potent stimulator of mitosis, oncogenicERBB2 signaling has recently been found responsible for theactivation of an initial antiproliferative response in tumor cells (16).
Figure 4. ERBB2 signaling and hypoxia act synergistically on a5 integrinexpression. A, real-time quantitative reverse transcription-PCR analysis(duplicate readings) of ITGA5 mRNA expression in different mammary cell linescultured under normoxic or hypoxic conditions (0.1% O2) for 24 hours. Therelative ITGA5 mRNA abundance under normoxia was set to 1 for each cell line.Columns, means of a representative experiment; bars, SD. B, MCF-7 cells weretransfected with either empty (pGL2) or promoter-containing (pGL2-ITGA5 )plasmids, left under routine culture conditions for 16 hours, and then kept eitherunder normoxia or hypoxia (0.1% O2) for an additional 24 hours. Luciferaseactivities (expressed as RLU) as measured in the resulting cellular extractsare depicted as mean values of duplicate transfections. Columns, means;bars, SD. C, real-time quantitative reverse transcription-PCR analysis(duplicate readings) of ITGA5 mRNA-expression in NeuT48 cells grown in theabsence or presence of doxycycline for a total of 36 hours. Where indicated, cellswere put into hypoxic atmosphere (0.1% O2, 24 hours) 12 hours after the additionof doxycycline. The relative ITGA5 mRNA abundance in NeuT48 cells grownunder normoxia in the absence of doxycycline was set to 1 to assesstranscriptional induction due to the two stimuli (ERBB2-signaling, hypoxia) alone(compare open versus filled column under normoxic conditions, or open columnsunder normoxic versus hypoxic conditions, respectively) or in combination(compare open column under normoxic versus filled column under hypoxicconditions). Columns, means of a representative experiment; bars, SD.
ERBB2 Induces the a5b1 Integrin Fibronectin Receptor
www.aacrjournals.org 3721 Cancer Res 2006; 66: (7). April 1, 2006
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
Thus, it was suggested that an inherent anticarcinogenic programmust be overcome in the early stages of carcinogenesis to progressto malignancy (reviewed in refs. 17, 18).In an attempt to unravel the molecular circuitry of early steps in
ERBB2-driven tumorigenesis, we screened MCF-7 mammary
adenocarcinoma cells with inducible ERBB2 for the expression ofa-integrin subunits of cellular integrin heterodimers becauseintegrins have been shown to significantly contribute to the patho-genesis of malignancies with respect to tumor cell survival andproliferation (29–31). Among the integrins displaying regulation by
Figure 5. ERBB2 signaling inducesincreased fibronectin adhesivenessof breast carcinoma cells resulting inimproved survival under adverseconditions and chemoresistance.A, adhesion of EGFP10 control cells (left)and NeuT48 cells grown for 48 hours in theabsence (open columns ) or presence(black columns ) of doxycycline to differentsubstrates. Percent cells adhering relativeto the plating control was calculated forthe respective surfaces [untreated tissueculture (standard TC ), BSA-treated (BSA ),fibronectin-coated (FN)]. Columns,means of quadruplicate readings; bars,SD. B, EGFP10 control cells (left) andNeuT48 cells (middle ) were grown in thepresence (filled symbols ) or absence(open symbols ) of doxycycline for 30hours, plated (day 0, d0) on untreated (E)or fibronectin-coated wells (n), and keptunder serum-free conditions for theindicated periods of time. The relativenumber of living cells represents theportion of living cells at a given time point(determined as described in Materialsand Methods) expressed as a multiple ofthe number of cells plated. Columns,means of triplicate readings; bars, SD.C, the same experimental setup as in (B ),but the cells were plated (day 0, d0) andkept under hypoxic conditions (0.1% O2)for the indicated time periods. The relativenumber of living cells represents theportion of living cells at the respectivetime point (determined as described inMaterials and Methods) expressed as amultiple of the number of cells plated.Columns, means of triplicate readings;bars, SD. D, EGFP10 control cells (left)and NeuT48 cells (right ) were grown in thepresence (filled symbols ) or absence(open symbols ) of doxycycline for 30hours, plated (day 0, d0) on untreated (E)or fibronectin-coated wells (n), andexposed to the indicated concentrations ofcisplatin (CDDP; top ) or 5-FU (bottom ) for72 hours. The fraction of living cellsrelative to the drug-free control sampleswas determined for each concentrationusing the WST-1 reagent as described.Columns, means of triplicate readings;bars, SD. Survival curves were plottedapplying nonlinear regression assuming amodified Hill model (28); dashed lines,standard tissue culture vessels; solid lines,fibronectin-coated vessels. Resistancefactors (RF ) are computed by dividingthe IC50 of the respective experimentalconditions and cell lines.
Cancer Research
Cancer Res 2006; 66: (7). April 1, 2006 3722 www.aacrjournals.org
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

oncogenic ERBB2 signal transduction in our cells, we selected theconstituents of the major cellular fibronectin receptor (a5 and h1integrin proteins) for further in-depth investigation, mainlybecause of its known effect on tumor cell survival andproliferation. By specifically adhering to the ECM componentfibronectin, the receptor improves the survival of different celltypes under adverse conditions (see refs. 20–24; see refs. 29, 47–50for reviews). Thus, adhesion on fibronectin prevents the apoptosisof Chinese hamster ovary cells upon serum withdrawal, and thisprosurvival/antiapoptotic effect of fibronectin-adhesion has beenattributed to the selective up-regulation of the antiapoptotic BCL-2protein, involving SHC, FAK, and RAS subsequently activating thePI3K/AKT pathway in these cells (20, 51). In the recent work ofKorah et al., the interaction of a5h1 integrin and fibronectinmaintains survival of different growth-arrested mammary adeno-carcinoma cells in an in vitro model system of bone marrowmetastasis. Comparable to results obtained in our inducible cellsystem, the group elegantly shows that selective up-regulation ofthe a5h1 integrin fibronectin receptor by FGF2 treatment—thereby simulating the conditions of bone marrow metastasis—leads to long-term survival of dormant clones (24).Tumor cell dormancy is a concept used to characterize
nonproliferating single tumor cells that have been shed by theprimary tumor and extravasated in a distant target organ (refs.52, 53; see refs. 25, 26, 54–56 for reviews). Metastatic growth oftumor cells at distant sites is generally considered a late stepduring tumorigenesis, but current metastasis models hypothe-size that dissemination of single tumor cells begins even in thevery early phases of malignancy (25, 26, 57). Such disseminatedmicrometastases can be found in a number of organs, like thebone marrow, as solitary cells, where they may survive for a longperiod of time, eventually leading to metastatic disease evendecades later. On the molecular level, dormancy is not wellcharacterized. One prominent feature of dormant cells is thealtered spectrum of integrin cell surface receptors involved insurvival signaling. Engagement of specific signaling pathways,like the activity ratio of ERKMAPK to P38MAPK, seems to be animportant determinant of tumor cell fate—i.e., proliferativegrowth versus dormancy (58, 59). Interestingly, the a5h1 integrinfibronectin receptor, in concert with uPAR, is known tomodulate the respective mitogen-activated protein kinase(MAPK) activities and therefore represents a critical relay inthe tumor cell with respect to proliferation/dormancy (58).Given that a5 and h1 integrins, as shown in the present study,are targets of oncogenic ERBB2 signaling, it is intriguing tospeculate that this regulatory circuit represents an importantmechanism of breast cancer cell dormancy in vivo , that mighteven be augmented by additional growth factor signals [e.g.,FGF2 as documented by Korah et al. (24)]. The observation thatthe ERBB2 conditional cells display a profound activation ofboth relevant MAPK signal transduction cascades in response tooncogenic ERBB2 signaling, with p38 MAPK being the pathwayresponsible for transducing the growth arrest signal (16), furthersupports this concept.Expression profiling experiments have identified a molecular
signature associated with bone marrow micrometastasis in humanbreast cancer (60). By using cDNA arrays to comparatively monitorprimary tumors of patients with bone marrow–positive and bonemarrow-negative breast cancer, the authors showed that differen-tially expressed genes include those involved in ECM remodeling,adhesion, and signal transduction. In this study, HIF-1a was
among those genes most profoundly up-regulated in bonemarrow–positive primary tumors. The consistent regulation ofseveral known HIF-1 target genes in the samples analyzed suggeststhat pathways connected with HIF impinge on bone marrowmetastasis (60). Whether HIF-dependent regulation of ECMreceptors is responsible for the association of HIF-1a over-expression and unfavorable prognosis (shorter overall survival,shorter disease-free survival; refs. 61–63) in patients with advancedstage breast cancer remains to be determined. Other integrin ECMreceptors, like a6h4 and a6h1 integrin heterodimers which areassociated with breast cancer progression have been shown to playimportant roles in cellular responses to hypoxia (64, 65). As HIF-1ais known to be stabilized by ERBB2 signaling under normoxicconditions (35–37), we propose that the a5h1 integrin fibronectinreceptor is a major effector in this regulatory circuit and mayrepresent the molecular basis for the HIF-1a-dependent aggres-siveness observed in ERBB2-overexpressing breast carcinomas.According to this hypothesis, HIF-1a-dependent up-regulationof the a5h1 integrin fibronectin receptor in response to hypoxia(as shown in refs. 33, 34; and in the present study) and/or ERBB2(this study) might be one decisive factor selecting for living butnondividing, i.e., dormant chemoresistant cells frequently observedin the bone marrow of patients with breast cancer. The functionalrelevance of the cellular a5h1 integrin fibronectin receptor forbreast cancer cell dormancy has recently been described (24).Hence, we suggest that the ERBB2-HIF-fibronectin receptorregulatory interaction is involved in increasing the portion ofsurviving hypoxic cells overexpressing ERBB2 described recently byDragowska and coworkers (66). This notion is supported by ourfunctional studies demonstrating a prosurvival effect of ERBB2signaling under hypoxia that is dependent on fibronectin adhesion(Fig. 5C).Resistance to chemotherapeutic or radiotherapeutic interven-
tion has been associated with ERBB2 overexpression in tumors(41, 44, 67–70). Given the stimulation of fibronectin adhesivenessfollowing oncogenic ERBB2 signaling, we also examined theinfluence of this interaction on tumor cell chemoresistance andfound that fibronectin ligation lead to a 5-fold increase inresistance against drugs used in breast cancer chemotherapyprotocols (5-FU and CDDP), further supporting the clinicalrelevance of the ERBB2/a5 integrin regulatory interaction. Onehallmark of tumor cell dormancy is the inherent resistance ofnondividing cells against chemotherapeutic agents (26, 55, 71). Assuggested by the work of Korah et al. (24), as well as by ourfunctional in vitro studies, the a5h1 integrin/fibronectin interac-tion may be a (perhaps essential) marker for dormant cells, andtherefore a very interesting target for breast cancer therapy. Givenits well accessible cellular location, peptides targeting fibronectinadhesion via the a5h1 integrin heterodimer, or small-moleculeinhibitors targeting downstream survival signaling could be used astherapeutic tools—even in combination with strategies directedagainst the ERBB2 RTK—to prevent signal transduction essentialfor long-term survival of dormant tumor cells (29, 72). Further-more, the identification of HIF as a central mediator of ERBB2-dependent regulation of tumor cells’ fibronectin adhesiveness addsan attractive drug target (73) that might be of special importance,given the synergistic activation of fibronectin receptor expressionby ERBB2-signaling and low oxygen tension observed in the workpresented.In summary, we identify a5 integrin, as well as its heterodi-
merization partner, h1 integrin, as targets of oncogenic ERBB2
ERBB2 Induces the a5b1 Integrin Fibronectin Receptor
www.aacrjournals.org 3723 Cancer Res 2006; 66: (7). April 1, 2006
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

signaling. Using Tet-inducible epithelial breast adenocarcinomacells, we show that ERBB2 signaling leads to the accumulationof ITGA5 and ITGB1 transcripts and protein. Functionally,oncogene-induced expression of the a5h1 integrin fibronectinreceptor on the surface of MCF-7 cells enhances their adhesive-ness to fibronectin, and adhesion on fibronectin leads to chemo-resistance and enhanced survival of ERBB2-overexpressing cellsunder adverse conditions (specifically nutrient deprivation andhypoxia). We discuss that increased fibronectin adhesivenessinduced by oncogenic ERBB2 signaling plays an importantrole in the course of ERBB2-driven carcinogenesis, and thatthis may be particularly relevant in the subsistence of long-surviving, dormant micrometastatic cells in the early phase
following activation of the ERBB2 proto-oncogene. Hence, thenovel regulatory interactions described here could lead toadditional (complementary) therapeutic options for ERBB2-dependent malignancies.
Acknowledgments
Received 8/9/2005; revised 1/20/2006; accepted 1/26/2006.Grant support: Stiftung Rheinland-Pfalz fur Innovation (386261/618), Deutsche
Forschungsgemeinschaft (SP 682-1/1), and Maifor (C. Spangenberg); DeutscheForschungsgemeinschaft (LA 1381-3/2, E.U. Lausch).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
The authors wish to thank Elke Deiss and Marcel Engel for excellent technicalassistance.
References1. Guy PM, Platko JV, Cantley LC, Cerione RA, CarrawayKL III. Insect cell-expressed p180erbB3 possesses animpaired tyrosine kinase activity. Proc Natl Acad SciU S A 1994;91:8132–6.2. Olayioye MA, Neve RM, Lane HA, Hynes NE. TheErbB signaling network: receptor heterodimerization indevelopment and cancer. EMBO J 2000;19:3159–67.3. Yarden Y, Sliwkowski MX. Untangling the ErbBsignalling network. Nat Rev Mol Cell Biol 2001;2:127–37.4. Slamon DJ, Clark GM, Wong SG, et al. Human breastcancer: correlation of relapse and survival with ampli-fication of the HER-2/neu oncogene. Science 1987;235:177–82.5. Tanner B, Kreutz E, Weikel W, et al. Prognosticsignificance of c-erB-2 mRNA in ovarian carcinoma.Gynecol Oncol 1996;62:268–77.6. Slamon DJ, Godolphin W, Jones LA, et al. Studies of theHER-2/neu proto-oncogene in human breast andovarian cancer. Science 1989;244:707–12.7. Sauter G, Moch H, Moore D, et al. Heterogeneity oferbB-2 gene amplification in bladder cancer. Cancer Res1993;53:2199–203.8. Weiner DB, Nordberg J, Robinson R, et al. Expressionof the neu gene-encoded protein (P185neu) in humannon-small cell carcinomas of the lung. Cancer Res 1990;50:421–5.9. Ross JS, McKenna BJ. The HER-2/neu oncogene intumors of the gastrointestinal tract. Cancer Invest 2001;19:554–68.10. Menard S, Casalini P, Campiglio M, et al. HER2overexpression in various tumor types, focussing on itsrelationship to the development of invasive breastcancer. Ann Oncol 2001;12:S15–19.11. DiGiovanna MP, Chu P, Davison TL, et al. Activesignaling by HER-2/neu in a subpopulation of HER-2/neu-overexpressing ductal carcinoma in situ : clinico-pathological correlates. Cancer Res 2002;62:6667–73.12. Menard S, Casalini P, Campiglio M, Pupa SM,Tagliabue E. Role of HER2/neu in tumor progressionand therapy. Cell Mol Life Sci 2004;61:2965–78.13. Yu D, Hung MC. Role of erbB2 in breast cancerchemosensitivity. Bioessays 2000;22:673–80.14. Eccles SA. The role of c-erbB-2/HER2/neu in breastcancer progression and metastasis. J Mammary GlandBiol Neoplasia 2001;6:393–406.15. Holbro T, Civenni G, Hynes NE. The ErbB receptorsand their role in cancer progression. Exp Cell Res 2003;284:99–110.16. Trost TM, Lausch EU, Fees SA, et al. Prematuresenescence is a primary fail-safe mechanism of ERBB2-driven tumorigenesis in breast carcinoma cells. CancerRes 2005;65:840–9.17. Lowe SW, Cepero E, Evan G. Intrinsic tumoursuppression. Nature 2004;432:307–15.18. Dimri GP. What has senescence got to do withcancer? Cancer Cell 2005;7:505–12.19. Adachi M, Taki T, Higashiyama M, et al. Significanceof integrin a5 gene expression as a prognostic factor in
node-negative non-small cell lung cancer. Clin CancerRes 2000;6:96–101.20. Zhang Z, Vuori K, Reed JC, Ruoslahti E. The a5h1integrin supports survival of cells on fibronectin andup-regulates Bcl-2 expression. Proc Natl Acad Sci U S A1995;92:6161–5.21. Varner JA, Emerson DA, Juliano RL. Integrin a5h1expression negatively regulates cell growth: reversal byattachment to fibronectin. Mol Biol Cell 1995;6:725–40.22. Nista A, Leonetti C, Bernardini G, Mattioni M,Santoni A. Functional role of a4h1 and a5h1 integrinfibronectin receptors expressed on Adriamycin-resistantMCF-7 human mammary carcinoma cells. Int J Cancer1997;72:133–41.23. Lee JW, Juliano RL. a5h1 Integrin protects intestinalepithelial cells from apoptosis through a phosphatidy-linositol 3-kinase and protein kinase B-dependentpathway. Mol Biol Cell 2000;11:1973–87.24. Korah R, Boots M, Wieder R. Integrin a5h1 promotessurvival of growth-arrested breast cancer cells: anin vitro paradigm for breast cancer dormancy in bonemarrow. Cancer Res 2004;64:4514–22.25. Pantel K, Brakenhoff RH. Dissecting the metastaticcascade. Nat Rev Cancer 2004;4:448–56.26. Wieder R. Insurgent micrometastases: sleeper cellsand harboring the enemy. J Surg Oncol 2005;89:207–10.27. Birkenmeier TM, McQuillan JJ, Boedeker ED, et al.The a5h1 fibronectin receptor. Characterization of thea5 gene promoter. J Biol Chem 1991;266:20544–9.28. Gardner SN. A mechanistic, predictive model ofdose-response curves for cell cycle phase-specific and-nonspecific drugs. Cancer Res 2000;60:1417–25.29. Jin H, Varner J. Integrins: roles in cancer developmentand as treatment targets. Br J Cancer 2004;90:561–5.30. Danen EH. Integrins: regulators of tissue functionand cancer progression. Curr Pharm Des 2005;11:881–91.31. Guo W, Giancotti FG. Integrin signalling duringtumour progression. Nat Rev Mol Cell Biol 2004;5:816–26.32. Denko NC, Fontana LA, Hudson KM, et al. Investi-gating hypoxic tumor physiology through gene expres-sion patterns. Oncogene 2003;22:5907–14.33. Koike T, Kimura N, Miyazaki K, et al. Hypoxiainduces adhesion molecules on cancer cells: a missinglink between Warburg effect and induction of selectin-ligand carbohydrates. Proc Natl Acad Sci U S A 2004;101:8132–7.34. Wykoff CC, Sotiriou C, Cockman ME, et al. Genearray of VHL mutation and hypoxia shows novelhypoxia-induced genes and that cyclin D1 is a VHLtarget gene. Br J Cancer 2004;90:1235–43.35. Laughner E, Taghavi P, Chiles K, Mahon PC, SemenzaGL. HER2 (neu) signaling increases the rate of hypoxia-inducible factor 1a (HIF-1a) synthesis: novel mecha-nism for HIF-1-mediated vascular endothelial growthfactor expression. Mol Cell Biol 2001;21:3995–4004.36. Li YM, Zhou BP, Deng J, et al. A hypoxia-independenthypoxia-inducible factor-1 activation pathway inducedby phosphatidylinositol-3 kinase/Akt in HER2 over-expressing cells. Cancer Res 2005;65:3257–63.
37. Xu Q, Briggs J, Park S, et al. Targeting Stat3 blocksboth HIF-1 and VEGF expression induced by multipleoncogenic growth signaling pathways. Oncogene 2005;24:5552–60.38. Aoudjit F, Vuori K. Integrin signaling inhibitspaclitaxel-induced apoptosis in breast cancer cells.Oncogene 2001;20:4995–5004.39. Sethi T, Rintoul RC, Moore SM, et al. Extracellularmatrix proteins protect small cell lung cancer cellsagainst apoptosis: a mechanism for small cell lungcancer growth and drug resistance in vivo . Nat Med1999;5:662–8.40. Yu D, Hung MC. Overexpression of ErbB2 incancer and ErbB2-targeting strategies. Oncogene 2000;19:6115–21.41. Knuefermann C, Lu Y, Liu B, et al. HER2/PI-3K/Akt activation leads to a multidrug resistance inhuman breast adenocarcinoma cells. Oncogene 2003;22:3205–12.42. Perez-Soler R, Kemp B, Wu QP, et al. Response anddeterminants of sensitivity to paclitaxel in human non-small cell lung cancer tumors heterotransplanted innude mice. Clin Cancer Res 2000;6:4932–8.43. Leitzel K, Teramoto Y, Konrad K, et al. Elevatedserum c-erbB-2 antigen levels and decreased responseto hormone therapy of breast cancer. J Clin Oncol 1995;13:1129–35.44. Hengstler JG, Lange J, Kett A, et al. Contribution ofc-erbB-2 and topoisomerase IIa to chemoresistance inovarian cancer. Cancer Res 1999;59:3206–14.45. Braun S, Pantel K, Muller P, et al. Cytokeratin-positivecells in the bone marrow and survival of patients withstage I, II, or III breast cancer. N Engl J Med 2000;342:525–33.46. Braun S, Schlimok G, Heumos I, et al. ErbB2overexpression on occult metastatic cells in bonemarrow predicts poor clinical outcome of stage I-IIIbreast cancer patients. Cancer Res 2001;61:1890–5.47. Kumar CC. Signaling by integrin receptors. Oncogene1998;17:1365–73.48. Ruoslahti E. Fibronectin and its integrin receptors incancer. Adv Cancer Res 1999;76:1–20.49. Jones JL, Walker RA. Integrins: a role as cell signallingmolecules. Mol Pathol 1999;52:208–13.50. Buttery RC, Rintoul RC, Sethi T. Small cell lungcancer: the importance of the extracellular matrix. Int JBiochem Cell Biol 2004;36:1154–60.51. Matter ML, Ruoslahti E. A signaling pathway fromthe a5h1 and a(v)h3 integrins that elevates bcl-2transcription. J Biol Chem 2001;276:27757–63.52. Luzzi KJ, MacDonald IC, Schmidt EE, et al. Multistepnature of metastatic inefficiency: dormancy of solitarycells after successful extravasation and limited survivalof early micrometastases. Am J Pathol 1998;153:865–73.53. Naumov GN, MacDonald IC, Weinmeister PM, et al.Persistence of solitary mammary carcinoma cells in asecondary site: a possible contributor to dormancy.Cancer Res 2002;62:2162–8.54. Naumov GN, MacDonald IC, Chambers AF, GroomAC. Solitary cancer cells as a possible source of tumourdormancy? Semin Cancer Biol 2001;11:271–6.
Cancer Research
Cancer Res 2006; 66: (7). April 1, 2006 3724 www.aacrjournals.org
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

55. Chambers AF, Groom AC, MacDonald IC. Dissemi-nation and growth of cancer cells in metastatic sites.Nat Rev Cancer 2002;2:563–72.56. Cairns RA, Khokha R, Hill RP. Molecular mechanismsof tumor invasion and metastasis: an integrated view.Curr Mol Med 2003;3:659–71.57. Riethmuller G, Klein CA. Early cancer cell dissemi-nation and late metastatic relapse: clinical reflectionsand biological approaches to the dormancy problem inpatients. Semin Cancer Biol 2001;11:307–11.58. Aguirre-Ghiso JA, Liu D, Mignatti A, Kovalski K,Ossowski L. Urokinase receptor and fibronectin regulatethe ERK(MAPK) to p38(MAPK) activity ratios thatdetermine carcinoma cell proliferation or dormancyin vivo . Mol Biol Cell 2001;12:863–79.59. Aguirre-Ghiso JA, Ossowski L, Rosenbaum SK. Greenfluorescent protein tagging of extracellular signal-regulated kinase and p38 pathways reveals noveldynamics of pathway activation during primary andmetastatic growth. Cancer Res 2004;64:7336–45.60. Woelfle U, Cloos J, Sauter G, et al. Molecularsignature associated with bone marrow micrometastasisin human breast cancer. Cancer Res 2003;63:5679–84.61. Schindl M, Schoppmann SF, Samonigg H, et al.
Overexpression of hypoxia-inducible factor 1a is asso-ciated with an unfavorable prognosis in lymph node-positive breast cancer. Clin Cancer Res 2002;8:1831–7.62. Bos R, van der GP, Greijer AE, et al. Levels of hypoxia-inducible factor-1a independently predict prognosis inpatients with lymph node negative breast carcinoma.Cancer 2003;97:1573–81.63. Giatromanolaki A, Koukourakis MI, Simopoulos C, etal. c-erbB-2 related aggressiveness in breast cancer ishypoxia inducible factor-1a dependent. Clin Cancer Res2004;10:7972–7.64. Chung J, Yoon S, Datta K, Bachelder RE, MercurioAM. Hypoxia-induced vascular endothelial growthfactor transcription and protection from apoptosis aredependent on a6h1 integrin in breast carcinoma cells.Cancer Res 2004;64:4711–6.65. Yoon SO, Shin S, Mercurio AM. Hypoxia stimulatescarcinoma invasion by stabilizing microtubules andpromoting the Rab11 trafficking of the a6h4 integrin.Cancer Res 2005;65:2761–9.66. Dragowska WH, Warburton C, Yapp DT, et al. HER-2/neu overexpression increases the viable hypoxic cellpopulation within solid tumors without causing changesin tumor vascularization. Mol Cancer Res 2004;2:606–19.
67. Yu D, Jing T, Liu B, et al. Overexpression of ErbB2 blocksTaxol-induced apoptosis by up-regulation of p21Cip1,which inhibits p34Cdc2 kinase. Mol Cell 1998;2:581–91.68. Tan M, Jing T, Lan KH, et al. Phosphorylation ontyrosine-15 of p34(Cdc2) by ErbB2 inhibits p34(Cdc2)activation and is involved in resistance to taxol-inducedapoptosis. Mol Cell 2002;9:993–1004.69. Liang K, Lu Y, Jin W, et al. Sensitization of breastcancer cells to radiation by trastuzumab. Mol CancerTher 2003;2:1113–20.70. Soderlund K, Perez-Tenorio G, Stal O. Activation ofthe phosphatidylinositol 3-kinase/Akt pathway preventsradiation-induced apoptosis in breast cancer cells. Int JOncol 2005;26:25–32.71. Naumov GN, Townson JL, MacDonald IC, et al.Ineffectiveness of doxorubicin treatment on solitarydormant mammary carcinoma cells or late-developingmetastases. Breast Cancer Res Treat 2003;82:199–206.72. Paulhe F, Manenti S, Ysebaert L, et al. Integrinfunction and signaling as pharmacological targets incardiovascular diseases and in cancer. Curr Pharm Des2005;11:2119–34.73. Semenza GL. Targeting HIF-1 for cancer therapy. NatRev Cancer 2003;3:721–32.
ERBB2 Induces the a5b1 Integrin Fibronectin Receptor
www.aacrjournals.org 3725 Cancer Res 2006; 66: (7). April 1, 2006
Research. on May 17, 2014. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN




















