
VCAM-1 on activated endothelium interacts with the leukocyte integrin VLA4 at a site distinct from...
8
Cell, Vol. 60, 577-564, February 23, 1990, Copyright 0 1990 by Cell Press VCAM-1 on Activated Endothelium Interacts with the Leukocyte lntegrin VLA-4 at a Site Distinct from the VLA=4/Fibronectin Binding Site Mariano J. Elites: Laurelee Osborn,t Yoshikazu Takada,’ Carol Grouse,’ Stefan Luhowskyj,t Martin E. Hemler: and Roy R. Lobbt l Division of Tumor Virology Dana-Farber Cancer Institute Boston, Massachusetts 02115 t Biogen, Inc. . 14 Cambridge Center Cambridge, Massachusetts 02142 Summary Cytokine-activated human endothelial cells express vascular cell adhesion molecule-l (VCAM-l), which binds lymphocytes. We now identify the in&grin VLA-4 as a receptor for VCAM-1 because VLA4 surface ex- pfession on K-562 cells (following transfection of the VLA a4 subunit cDNA) resulted in specific cell adhe- sion to VCAM-1, and antiYLA4 antibodies completely inhibited VCAM-l-dependent cell-cell attachment. In addition, VLA-4 expression allowed K-562 cells to at- tach to the heparin II binding region (FN-40) of fibro- nectin. However, VLA-4NCAM-1 and VLR4/FN-40 in- teractions are readily distinguishable: only the former was inhibited by the antiYLA4 monoclonal antibody HPl13, and only the latter was inhibited by soluble FN- 40. The VCAM-lNL&4 iigand-receptor pair may play a major role in the recruitment of mononuclear leuko- cytes to inflammatory sites in vivo. introduction Vascular endothelial cells constitute the lining of blood vessels and normally exhibit a low affinity for circulating leukocytes (Harlan, 1965). The release of cytokines at sites of inflammation, and in response to immune reac- tions, causes their activation and results in the increased expression of a host of surface antigens (Collins et al., 1986; Pober et al., 1986; Bevilacqua et al., 1987; Leeuwen- berg et al., 1989). These include the adhesion proteins ELAM-1, which binds neutrophils (Bevilacqua et al., 1987, 1989), and ICAM-1, which interacts with all leukocytes (Dustin et al., 1986; Pober et al., 1986; Boyd et al., 1988; Dustin and Springer, 1988). In addition, we have recently described a new adhesion protein, VCAM-1, which binds lymphocytes (Osborn et al., 1989). These cytokine-in- duced adhesion molecules appear to play an important role in leukocyte recruitment to extravascular tissues. The integrins are a group of cell-extracellular matrix and cell-cell adhesion receptors exhibiting an ap hetero- dimeric structure, with a widespread cell distribution and a high degree of conservation throughout evolution (Hynes, 1987; Marcantonio and Hynes, 1988). The integrins have been subdivided into three major subgroups: the p2 sub- family of integrins (LFA-1, Mac-l, and p150,95) is mostly in- volved in cell-cell interactions within the immune system (Kishimoto et al., 1989), whereas members of the p, and p3 integrin subfamilies predominantly mediate cell attach- ment to the extracellular matrix (Hynes, 1987; Ruoslahti, 1988). In particular, the PI integrin family, also termed VLA proteins, includes at least six receptors that specifi- cally interact with fibronectin, collagen, and/or laminin (Hemler, 1990). Within the VLA family, VLA-4 is atypical because it is mostly restricted to lymphoid and myeloid cells (Hemler et al., 1987b), and indirect evidence has sug- gested that it may be involved in various cell-cell interac- tions (Clayberger et al., 1987; Takada et al., 1989; Holz- mann et al., 1989; Bednarczyk and McIntyre, 1990). In addition, VLA-4 has recently been shown to mediate T and B lymphocyte attachment to the heparin II binding fragment of human plasma fibronectin (FN) (Wayner et al., 1989). VCAM-1, like ICAM-1, is a member of the immunoglobu- lin gene superfamily (Osborn et al., 1989). Since ICAM- is known to interact with the integrin LFA-1 (Marlin and Springer, 1987; Makgoba et al., 1988), we examined possi- ble VCAM-llintegrin receptor interactions. The present re- port demonstrates that VCAM-1 and VLA-4 represent a ligand-receptor pair that allows attachment of lympho- cytes to activated endothelium. Moreover, we show that VLA-4 binding to VCAM-1 occurs by a mechanism inde- pendent of the VLA-4 interaction with FN. Thus, VLA-4 represents a singular example of a PI integrin receptor participating in both cell-cell and cell-extracellular matrix adhesion functions by means of the defined ligands VCAM-1 and FN. Results Antibodies to VLA-4 Block Lymphocyte Attachment to VCAM-1 In a previous study, we showed conclusively that p2 inte- grins (which include LFA-1) were not involved in the selec- tive binding of the B cell line Ramos, or the T cell line Jur- kat, to VCAM-1 (Osborn et al., 1989). To evaluate the possible role of PI integrins in VCAM-l-dependent cell- ceil adhesion, the inhibitory effect of specific monoclonal antibodies (MAbs) on Ramos attachment to COS and hu- man umbilical vein endothelial cell (HUVEC) monolayers was assessed. Adhesion of Ramos to both TNF-a-treated HUVECs and VCAM-l-transfected COS cells was abro- gated by the anti-integrin p, MAb BlEll, but not by con- trol MAbs (Figures 1A and 1B). Likewise, complete inhi- bition was achieved using goat anti-& heteroantiserum (not shown). Since VLA-4 is the major receptor in the PI integrin family expressed on Ramos (see below), anti- VLA-4 antibodies were then tested. As shown in Figures 1A and lB, the anti-VLA-4 MAb HP2/1 blocked completely the attachment of Ramos to both TNF-a-activated HUVECs and VCAM-l-transfected COS cells. A second anti-VLA-4 MAb, HP1/3, displayed an inhibitory potency similar to HP2/1 (see Figure 6C), while B-5G10, a third MAb specific
Transcript of VCAM-1 on activated endothelium interacts with the leukocyte integrin VLA4 at a site distinct from...

Cell, Vol. 60, 577-564, February 23, 1990, Copyright 0 1990 by Cell Press
VCAM-1 on Activated Endothelium Interacts with the Leukocyte lntegrin VLA-4 at a Site Distinct from the VLA=4/Fibronectin Binding Site
Mariano J. Elites: Laurelee Osborn,t Yoshikazu Takada,’ Carol Grouse,’ Stefan Luhowskyj,t Martin E. Hemler: and Roy R. Lobbt l Division of Tumor Virology Dana-Farber Cancer Institute Boston, Massachusetts 02115 t Biogen, Inc. . 14 Cambridge Center Cambridge, Massachusetts 02142
Summary
Cytokine-activated human endothelial cells express vascular cell adhesion molecule-l (VCAM-l), which binds lymphocytes. We now identify the in&grin VLA-4 as a receptor for VCAM-1 because VLA4 surface ex- pfession on K-562 cells (following transfection of the VLA a4 subunit cDNA) resulted in specific cell adhe- sion to VCAM-1, and antiYLA4 antibodies completely inhibited VCAM-l-dependent cell-cell attachment. In addition, VLA-4 expression allowed K-562 cells to at- tach to the heparin II binding region (FN-40) of fibro- nectin. However, VLA-4NCAM-1 and VLR4/FN-40 in- teractions are readily distinguishable: only the former was inhibited by the antiYLA4 monoclonal antibody HPl13, and only the latter was inhibited by soluble FN- 40. The VCAM-lNL&4 iigand-receptor pair may play a major role in the recruitment of mononuclear leuko- cytes to inflammatory sites in vivo.
introduction
Vascular endothelial cells constitute the lining of blood vessels and normally exhibit a low affinity for circulating leukocytes (Harlan, 1965). The release of cytokines at sites of inflammation, and in response to immune reac- tions, causes their activation and results in the increased expression of a host of surface antigens (Collins et al., 1986; Pober et al., 1986; Bevilacqua et al., 1987; Leeuwen- berg et al., 1989). These include the adhesion proteins ELAM-1, which binds neutrophils (Bevilacqua et al., 1987, 1989), and ICAM-1, which interacts with all leukocytes (Dustin et al., 1986; Pober et al., 1986; Boyd et al., 1988; Dustin and Springer, 1988). In addition, we have recently described a new adhesion protein, VCAM-1, which binds lymphocytes (Osborn et al., 1989). These cytokine-in- duced adhesion molecules appear to play an important role in leukocyte recruitment to extravascular tissues.
The integrins are a group of cell-extracellular matrix and cell-cell adhesion receptors exhibiting an ap hetero- dimeric structure, with a widespread cell distribution and a high degree of conservation throughout evolution (Hynes, 1987; Marcantonio and Hynes, 1988). The integrins have been subdivided into three major subgroups: the p2 sub- family of integrins (LFA-1, Mac-l, and p150,95) is mostly in-
volved in cell-cell interactions within the immune system (Kishimoto et al., 1989), whereas members of the p, and p3 integrin subfamilies predominantly mediate cell attach- ment to the extracellular matrix (Hynes, 1987; Ruoslahti, 1988). In particular, the PI integrin family, also termed VLA proteins, includes at least six receptors that specifi- cally interact with fibronectin, collagen, and/or laminin (Hemler, 1990). Within the VLA family, VLA-4 is atypical because it is mostly restricted to lymphoid and myeloid cells (Hemler et al., 1987b), and indirect evidence has sug- gested that it may be involved in various cell-cell interac- tions (Clayberger et al., 1987; Takada et al., 1989; Holz- mann et al., 1989; Bednarczyk and McIntyre, 1990). In addition, VLA-4 has recently been shown to mediate T and B lymphocyte attachment to the heparin II binding fragment of human plasma fibronectin (FN) (Wayner et al., 1989).
VCAM-1, like ICAM-1, is a member of the immunoglobu- lin gene superfamily (Osborn et al., 1989). Since ICAM- is known to interact with the integrin LFA-1 (Marlin and Springer, 1987; Makgoba et al., 1988), we examined possi- ble VCAM-llintegrin receptor interactions. The present re-
port demonstrates that VCAM-1 and VLA-4 represent a ligand-receptor pair that allows attachment of lympho- cytes to activated endothelium. Moreover, we show that VLA-4 binding to VCAM-1 occurs by a mechanism inde- pendent of the VLA-4 interaction with FN. Thus, VLA-4 represents a singular example of a PI integrin receptor participating in both cell-cell and cell-extracellular matrix adhesion functions by means of the defined ligands VCAM-1 and FN.
Results
Antibodies to VLA-4 Block Lymphocyte Attachment to VCAM-1 In a previous study, we showed conclusively that p2 inte-
grins (which include LFA-1) were not involved in the selec- tive binding of the B cell line Ramos, or the T cell line Jur- kat, to VCAM-1 (Osborn et al., 1989). To evaluate the possible role of PI integrins in VCAM-l-dependent cell- ceil adhesion, the inhibitory effect of specific monoclonal antibodies (MAbs) on Ramos attachment to COS and hu- man umbilical vein endothelial cell (HUVEC) monolayers was assessed. Adhesion of Ramos to both TNF-a-treated HUVECs and VCAM-l-transfected COS cells was abro- gated by the anti-integrin p, MAb BlEll, but not by con- trol MAbs (Figures 1A and 1B). Likewise, complete inhi- bition was achieved using goat anti-& heteroantiserum (not shown). Since VLA-4 is the major receptor in the PI integrin family expressed on Ramos (see below), anti- VLA-4 antibodies were then tested. As shown in Figures 1A and lB, the anti-VLA-4 MAb HP2/1 blocked completely the attachment of Ramos to both TNF-a-activated HUVECs and VCAM-l-transfected COS cells. A second anti-VLA-4 MAb, HP1/3, displayed an inhibitory potency similar to HP2/1 (see Figure 6C), while B-5G10, a third MAb specific

Cell 578
Adhesion to HUVEC A
Untreated
F
Treated
with
TNFo
I 1 I I -J 0 200 400 600 800 1,000 1,200
Ramos Cells Bound/mm*
Adhesion B to cos I I
VCAM-I Anti-VIA-4
d
Anti-VLAO
I/ 0 100 200 300 400
Ramos Cells Bound/mm2
Figure 1. Inhibition of Ramos Cell Attachment to VCAM-l-Expressing Cell Monolayers
Ramos cells wre labeled with P?S]Met overnight, washed with serum- free RPM containing 1% BSA. and plated in triplicate on monolayers of (A) HUVECs with or without prior treatment with TNFa (10 @ml), or (B) COS cells transfected with either VCAM-1 or expression vector alone (see Experimental Procedures). Inhibition of Ramos binding to cell monolayers was carried out using MAbs P3 (control), HP2/l (anti- VLA-4) and BlEll (anti-VLA 3,). Specific adhesion of Ramos cells was analyzed by measuring the radioactivity of 0.1% SDS cell lysates in a beta counter (experimental variation was less than 10%).
for VLA-4, did not inhibit (not shown). Results similar to those presented in Figures 1A and 1B were also obtained using the VLA4-expressing T lymphoblastoid cell line HPB-ALL (not shown). These MAb blocking experiments strongly suggested an interaction between VCAM-1 and VLA-4.
Direct Evidence that VLA-4 Can Interact with VCAM-1 To confirm directly that the integrin heterodimer VLA-4 (a4f3r) acts as a receptor for VCAM-1, a cell line express- ing VLA4 after transfection with an a4 cDNA clone was tested for VLA-Cdependent binding to VCAM-1. K-562
erythroleukemia cells were chosen as appropriate host cells because they are devoid of VLA4 surface expres- sion (Hemier et al., 1967b) and have no endogenous VCAM-1 binding capability (see below). The recently cloned cDNA for the a4 subunit of VLA4 (Takada et al., 1969) was transfected into K-562 cells, and stable, long-term subiines were selected that express VLA4 as a result of association of transfected a4 with endogenous 6, (see below). Figure 2 illustrates the ability of K-562 a4 trans- fectants to adhere to COS cells transfected with VCAM-1 (Figure 2b), or to TNF-a-activated HUVECs (Figure 2d). In contrast, a control transfectant expressing VLA-2, pre- pared using the a* cDNA, failed to bind to VCAM-1 ex- pressed either on COS cells or on HUVECs (Figures 2a and 2~).
Quantitation of the results shown in Figure 2 was then carried out. Adhesion of K-562 a4 transfectant cells to a monolayer of COS cells transfected with VCAM-1 (Figure 3A) or to TNF-a-treated HUVECs (Figure 38) was approxi- mately 20 times higher than K-562 a4 transfectant cell binding to negative controls. Both TNF-a-activated HUV- ECs and VCAM-1-transfected COS cells failed to support attachment of control K-562 a2 transfectants (Figures 3A and 38) or of the parent cell line K-562 (not shown), indi- cating that a4 expression is essential for attachment. In addition, as in Figures 1A and lB, heterotypic interactions between the K-562 a4 transfectant and either VCAM-1 on COS cells, or VCAM-1 on TNF-a-treated HUVECs were abolished by MAbs to both VLA-4 and VLA 9, (Figures 3A and 38). These results demonstrate directly that VLA4 is a specific cell surface receptor for VCAM-1.
Characterixation of VLA Proteins Expressed on Ramos and Transfected Cell Lines To verify integrin cell surface phenotypes, flow cytometry analyses of Ramos cells and K-562 a2 and K-562 a4 transfectant cells were performed using a panel of MAbs to a and fi subunits (Figure 4). On the B lymphoblastoid cell line Ramos, VLA4 showed a high level of expression, with a mean Ruorescence intensity value almost equal to that for VLA 6, (Figure 4A). All other known VLA proteins (i.e., VLA-1, -2, -3, -5, and -6) were either undetectable or present in trace amounts. The K-562 a4 transfectant ex- pressed VLA 9, and VLA4 on the surface, as expected (Figure 48). in addition, VLA-5 levels were found to be comparable to those found in the parent ceil line K-562 (not shown), indicating that background integrin expres- sion had not changed upon transfection. Ail other known VLA proteins (i.e., VLA-1, -2, -3, and -6) were absent from the K-562 transfectant (not shown). The control K-562 a2 transfectant exhibited substantial surface expression of VLA-2 but was negative for VLA4 (Figure 4C). This trans- fectant also retained surface levels of VLA-5 similar to those present on the parent cell line K-562 (not shown). In addition, both K-562 a4 and a2 transfectants displayed comparable mean fluorescence intensities for the VLA 6, subunit (Figures 48 and 4C), implying that total expres- sion of 6, integrins was similar in both ceils.
lmmunoprecipitation analyses of Ramos and transfec- tant ceils yielded results consistent with the flow cytome-

Activated Endothelial VCAM-1 Binds VLA4 lntegrin 579
Adhesion to cos
A I I
Mock-transfected
r Control Transf.
Control mAb
VCAM-1 i
Anti-VIA4
I An&VlAR
0 too 200 300 400 Transfectant Cells Bound/mm2
Adhesion B I I
Treated Control rnAk
with
TNFa Ant&VU 4
0 100 200 300 400 500 600 700 Transfectant Cells Bound/mmZ
Figure 3. inhibition of K-562 a4 Transfectant Cell Adhesion to COS and HUVEC Monolayers
Attachment of K-562 a4 transfectants to either (A) COS cells trans- fected with VCAM-1 or (B) TNF-a-treated HUVECs, and inhibition by MAbs P3 (control), HP2/1 (anti-VLA-4) and BlEll (anti-VLA 6,) were performed as indicated in the legend to Figure 1.
Figure 2. Binding of K-562 a4 Transfectants to VCAM-1 Expressed on Either COS or HUVEC Monolayers
K-562 a4 transfectants (b and d) or control K- 562 as transfectants (a and c) were plated on monolayers of COS ceils transfected with VCAM-1 (a and b) or HUVECs incubated with TNF-a (c and d) as described in Experimental Procedures. After washing to remove unbound cells, plates were photographed (magnification 54x).
try results. On both Flamos and K-562 a4 transfectant ceils, anti-VLA-4 and anti-& MAbs immunoprecipitated the expected pattern of a4 and 8, subunits, including vari- able amounts of 80,000 and 70,000 M, fragments derived from the 150,000 M, intact a4 (Hemler et al., 1987a, 1987b). There was no evidence of any integrin 6 subunit, other than PI, associated with the a4 subunit on either Ramos cells or K-562 a4 transfectants (not shown).
VLA-4-Dependent Adhesion to Fibmnectin Recent studies have shown that VLA-4 mediates cell at- tachment to the CS-1 site within the heparin II binding fragment of human plasma FN (Wayner et al., 1989). Bind- ing of K-562 transfectants and Ramos cells to FN frag- ments confirmed this observation. Dose-dependent at- tachment to the C-terminal heparin II binding region of FN (FN-40) was observed for the VLA4-expressing cell lines Ramos and the K-562 a4 transfectant, but not for the VLA-Cnegative K-562 a2 transfectant (Figure 5A). In con- trast, both the K-562 a2 and a4 transfectants (Figure 58) as well as the parent cell line K-562 (not shown) showed similar degrees of adherence to the cell binding region of FN (FN-120) which contains the Arg-GlyAsp (RGD) se- quence recognized by VLAQ (Ruoslahti, 1988). As ex- pected, Ramos, which is negative for VLA-5 (see above), did not attach to FN-120 (Figure 5B).
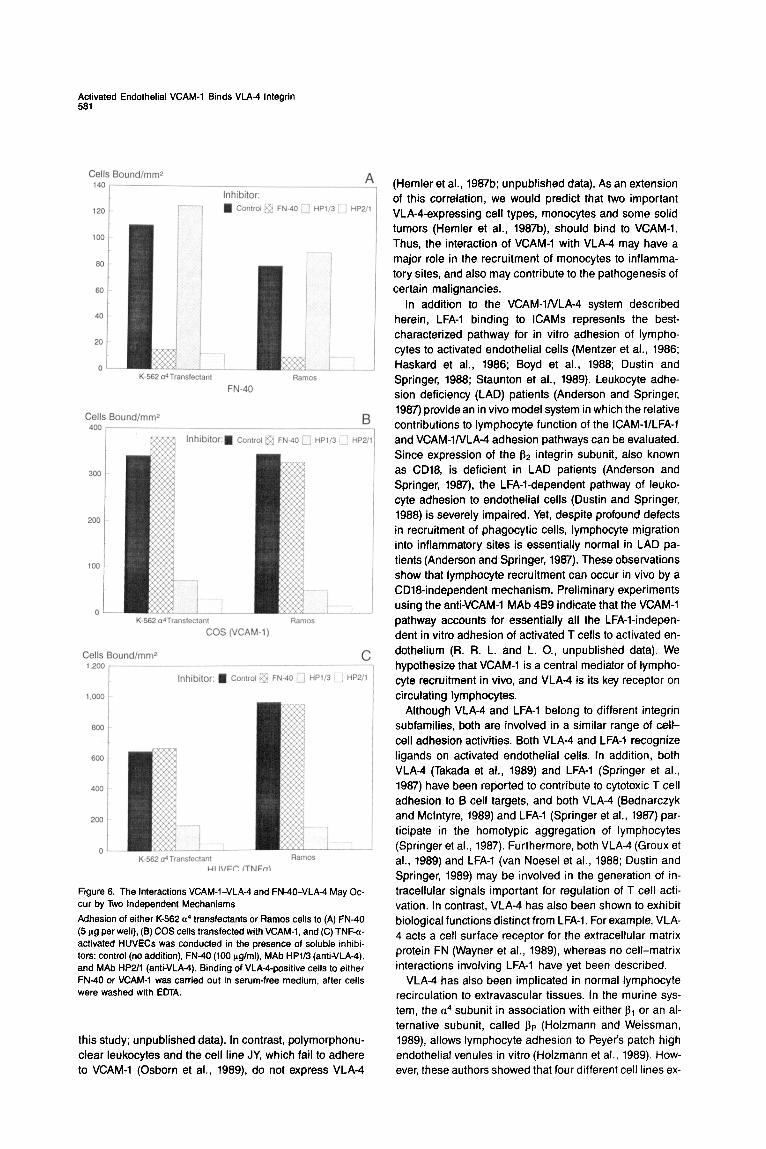
Cell-Cell and Cell-Matrix Adhesion Functions of VLA-4 Are independent Since VLA-4 mediates cell attachment to both VCAM-1 and FN-40, we addressed the question of whether the same or different sites on VLA-4 were responsible for both functions. Attachment of either K-562 a4 transfectant or Ramos cells to FN-40 could be completely inhibited by preincubation of cells with soluble FN-40 fragment (Figure 6A). In contrast, heterotypic adhesion of either K-562 a4 transfectant cells or Ramos to either VCAM-1-transfected COS cells (Figure 6B) or TNF-a-activated HUVECs (Fig-

Cell 580
A. RAMOS 6. K-562 C. (a41 )
K-562 , (Control)
-- ..- &LA-4
FLUORESCENCE INTENSITY
Figure 4. Cell Surface Expression of VLA Heterodimers on Ramos Cells and K-562 a4 and K-562 a2 (Control) Transfectants
Flow cytometry of cell lines was carried out using the MAbs P3 (con- trol), AlA (anti-VLA 6,) B5GlO (anti-WA a4), and Gil4 (anti-VLA a?, followed by incubation with fluorescein-labeled goat anti-mouse IgG as described in Experimental Procedures. Surface expression of B2 inte- grins was undetectable on both K-562 transfectants (not shown).
ure 6C) was unaffected by their prior treatment with solu- ble FN-40. This result shows that the VLA4/VCAM-1 inter- action is independent of VLA4/FN40 binding.
Experiments performed in parallel also showed that the anti-VLA-4 MAb HP1/3 did not inhibit attachment of either K-562 a4 transfectants or Ramos cells to FN40 (Figure 6A), yet completely inhibited adhesion of either K-562 a4 transfectants or Ramos cells to VCAM-1 expressed on COS (Figure 66) or TNF-a-treated HUVECs (Figure 6C). Taken together, the reciprocal inhibition patterns observed for soluble FN40 and HP1/3 indicate unequivocally that distinct sites on VLA-4 mediate attachment to FN40 and VCAM-1.
Even though VLA~IVCAM-1 and VLA-4IFN-40 interac- tions were readily distinguishable using soluble FN40 and HP113 as inhibitors, the anti-VLA4 MAb HP2/1 in- hibited VLA4 attachment to both FN40 (Figure 6A) and VCAM-1 (Figures 66 and 6C). This suggests that the two VLA-4 ligand binding sites, although functionally distinct, may be adjacent.
Discussion
Our data strongly suggest that the molecules VCAM-1 and VLA4 constitute a ligand-receptor pair. First, the VCAM- lNLA4 interaction was reconstituted using a heterotypic adhesion assay in which K-562 cells transfected with the a4 subunit of VLA4 bound to monolayers of COS cells transfected with VCAM-1. And second, adhesion of K-562 a4 transfectant cells to COS cells bearing VCAM-1 was abrogated by MAbs to both the a4 and the 8, subunit of the VLA4 protein.
K-562 a4 transfectant cells described herein were also shown to attach specifically to the heparin II binding (FN- 40) fragment of FN (Pierschbacher et al., 1961) thus directly confirming previous results obtained in antibody blocking studies (Wayner et al., 1969). Although there is no obvious sequence homology between VCAM-1 (Os- born et al., 1989) and the 25 amino acid CS-1 region in-
Adhesion to FN-40 fragment .* ,.a’
__-’ _,.-
,_,’ . ,_*’
.:’ )/
_,’ K-562 ol Transf.
,,I’ ,,,’
,’ . . Ramos ,-
,:’
,,‘* K-562 o? Transf.
,u”
,‘..u l
i 1 I
0 2 4
(LlGAND~@glwell) s 10 12
aw B
Adhesion to FN-120 fragment
0 . . . . . 7 . ,, .
1 0 2 4
(LlGAND~@g/well) s 10
Figure 5. Attachment of Ramos and K-562 Transfectants to Proteolytic Fragments of Human Plasma Fibronectin
Ramos cells, K-562 a4 transfectants, and K-562 control transfectants were tested for adhesion to microtiter plates coated with various amounts of either (A) the heparin II binding domain (FN-40) or(B) the cell binding domain (FN-120) of human plasma fibronectin (see Ex- perimental Procedures).
volved in VLA4 binding to FN (Wayner et al., 1989) it was possible that VCAM-1 interacted with the FN binding site on VLA-4. In addition, even though the transfection experi- ments show a direct VCAM-iNLA-4 interaction, it was con- ceivable that this adhesive function was mediated by FN, which can be made by T cells (Godfrey et al., 1988). How- ever, we found that VLA4-dependent binding to FN40 was inhibited by soluble FN40 but not by the anti-VLA4 MAb HP3/t, and, conversely, VLA4-dependent binding to VCAM-1 was inhibited by MAb HP3/l but not by soluble FN40. Thus, VCAM-1 interacts with a site on VLA4 func- tionally distinct from that which interacts with FN, and FN itself is not involved in the VCAM-lNLA4 interaction.
VLA4 is probably the major, if not the only, receptor for VCAM-1. Anti-VLA4 antibodies completely inhibited bind- ing of the lymphoid cell lines Ramos and HPBALL to VCAM-1 expressed in COS cells. Furthermore, we have found an absolute positive correlation between VLA4 ex- pression and VCAM-1 binding. Nine different cell types (HL60, U937, Ramos, Raji, Jurkat, HPBALL, THPl, pe- ripheral blood lymphocytes, and activated T cells) that bind to VCAM-1 (Osborn et al., 1989; and unpublished data) all express cell surface VLA4 (Hemler et al., 1987b;
&ivated Endothelial VCAM-1 Binds WA-4 lntegrin
A Inhibitor: 4 Control # FN-40 :-A HPlI3 0 HP20
Cells Bound/mm2 140
120
100
- K-562 +Transfectant Fiamos
FN-40
Cells Bound/mm2 4oc
300
200
100
0
B m Inhibitor:m control q ~~-40 r] HP1/3 r’i HP2/ 1
K-562 a4Transfectant Ramos COS (VCAM-1)
Cells Bound/mm2 C 1,200 r
1,000
800
600
Inhibitor: m Control @ FN-40 g HPlI3 I HPZIt
K-562 dTransfectant Ramos HUVEC (TNFa)
Figure 6. The Interactions VCAM-I-VLA-4 and FN-40-VLA-4 May Oc- cur by Two Independent Mechanisms
Adhesion of either K-562 a4 transfectants or Ramos cells to (A) FN-40 (5 fig per well), (6) COS cells transfected with VCAM-1, and (C) TNF-a- activated HUVECs was conducted in the presence of soluble inhibi- tors: control (no addition), FN40 (100 &ml), MAb HP1/3 (anti-VLA4), and MAb HP2/1 (anti-VLM). Binding of VLA4-positive cells to either FN-40 or VCAM-1 was carried out in serum-free medium, after cells were washed with EDTA.
this study; unpublished data). In contrast, polymorphonu- clear leukocytes and the cell line JY, which fail to adhere to VCAM-1 (Osborn et al., 1989), do not express VLA-4
(Hemler et al., 1987b; unpublished data). As an extension of this correlation, we would predict that two important VLA-Cexpressing cell types, monocytes and some solid tumors (Hemler et al., 1987b), should bind to VCAM-1. Thus, the interaction of VCAM-1 with VLA-4 may have a major role in the recruitment of monocytes to inflamma- tory sites, and also may contribute to the pathogenesis of certain malignancies.
In addition to the VCAM-l/VLA-4 system described herein, LFA-1 binding to ICAMs represents the best- characterized pathway for in vitro adhesion of lympho- cytes to activated endothelial cells (Mentzer et al., 1988; Haskard et al., 1986; Boyd et al., 1988; Dustin and Springer, 1988; Staunton et al., 1989). Leukocyte adhe- sion deficiency (LAD) patients (Anderson and Springer, 1967) provide an in vivo model system in which the relative contributions to lymphocyte function of the ICAM-l/LFA-1 and VCAM-lNLA-4 adhesion pathways can be evaluated. Since expression of the p2 integrin subunit, also known as CD18, is deficient in LAD patients (Anderson and Springer, 1987), the LFA-l-dependent pathway of leuko- cyte adhesion to endothelial cells (Dustin and Springer, 1988) is severely impaired. Yet, despite profound defects in recruitment of phagocytic cells, lymphocyte migration into inflammatory sites is essentially normal in LAD pa- tients (Anderson and Springer, 1987). These observations show that lymphocyte recruitment can occur in vivo by a CDlE-independent mechanism. Preliminary experiments using the anti-VCAM-1 MAb 489 indicate that the VCAM-1 pathway accounts for essentially all the LFA-l-indepen- dent in vitro adhesion of activated T cells to activated en- dothelium (Ft. R. L. and L. O., unpublished data). We hypothesize that VCAM-1 is a central mediator of lympho- cyte recruitment in vivo, and VLA-4 is its key receptor on circulating lymphocytes.
Although VLA4 and LFA-1 belong to different integrin subfamilies, both are involved in a similar range of ceil- cell adhesion activities. Both VLA-4 and LFA-1 recognize ligands on activated endothelial cells. In addition, both VLA-4 (Takada et al., 1989) and LFA-1 (Springer et al., 1987) have been reported to contribute to cytotoxic T cell adhesion to B cell targets, and both VLA4 (Bednarczyk and McIntyre, 1989) and LFA-1 (Springer et al., 1987) par- ticipate in the homotypic aggregation of lymphocytes (Springer et al., 1987). Furthermore, both VLA-4 (Groux et al., 1989) and LFA-1 (van Noesel et al., 1988; Dustin and Springer, 1989) may be involved in the generation of in- tracellular signals important for regulation of T cell acti- vation. In contrast, VLA4 has also been shown to exhibit biological functions distinct from LFA-1. For example, VLA- 4 acts a cell surface receptor for the extracellular matrix protein FN (Wayner et al., 1989), whereas no cell-matrix interactions involving LFA-1 have yet been described.
VLA4 has also been implicated in normal lymphocyte recirculation to extravascular tissues. In the murine sys- tem, the a4 subunit in association with either PI or an al- ternative subunit, called fip (Holzmann and Weissman, 1989), allows lymphocyte adhesion to Peyer’s patch high endothelial venules in vitro (Holzmann et al., 1989). How- ever, these authors showed that four different cell lines ex-

Cell 582
pressing a481 (VLA-4) but not a4f3p, did not bind to Peyer’s patch frozen sections (Holzmann and Weissman, 1989) implying that VLA4 is not itself sufficient for adhe- sion. An intriguing hypothesis is that a4fip provides selec- tivity for Peyer’s patch endothelium via an undefined ligand, perhaps one resembling a vascular addressin (Streeter et al., 1988; Berg et al., 1989), while a48r pro- vides selectivity for inflamed endothelium via VCAM-1. If VLA4 (and also a48p) is not itself a primary homing receptor, it instead may provide accessory functions for lymphocyte homing events, in a manner analogous to LFA-1 (Pals et al., 1988; Hamann et al., 1988). By exten- sion, a potential role for VLA-4 in abnormal lymphocyte trafficking should also be considered. In this regard, the related integrin LFA-1 has been suggested to be involved in lymphoma invasion and metastasis (Roossien et al., 1989).
In summary, we have unequivocally demonstrated that VLA-4 is a receptor for VCAM-1. We also have confirmed that VLA4 mediates attachment to the heparin II binding region of FN, and have shown that the VCAM-lNLA4 and FN40NLA-4 interactions are distinct. This opens the in- triguing possibility that VLA-Cpositive mononuclear leu- kocytes independently regulate these two adhesive func- tions during an inflammatory response in vivo. Thus as the need may arise, they may bind either to activated en- dothelium through the cellular ligand (VCAM-1) or, alterna- tively, to FN deposited at inflammatory sites.
Exoerimentai Procedures
Antibodies, Ceils, and Matrix Proteins Monoclonal antibodies utilized were anti-VLA-1, TS2I7 (Hemler et al., 1984); anti-VLM, Gil4 (Kiefel et al., 1989); anti-VLA-3, J143 (Fradet et al., 1984); anti-VLA-4. B-5610 (Hemler et al., 1987b), HP113, and HP2/1 (Sanchez-Madrid et al., 1966); anti-VLA-5, PlD6 (Wayner et al., 1968); anti-VLA-6, 0836 (Hsi et al.. 1987); anti-VLA 8,, A-lA5 (Hemler et al., 1983) and BlEll (from Dr. C. Damsky, University of California, San Francisco); and anti& integrin, TSI/t8 (Sanchez-Madrid et al., 1982). A goat heteroantiserum to purified VLA 8, was prepared by Dr. M. Ginsberg (Scripps institute, San Diego).
Ramos, HP&ALL, and K-562 cells were maintained in RPM1 1640 supplemented with 10% fetal bovine serum (FBS). COS7 cells were grown in HEPES-buffered Duibeccos modified Eagles medium con- taining 10% FBS. HUVECs were isolated and subcultured as previ- ously described (Osborn et al., 1969). K-562 a2 and a4 transfectants were maintained in 1 mg/mi geneticin in RPMI, 10% FBS.
FN from human plasma and the chymotryptic fragments FN-120 (cell binding domain) and FN-40 (heparin II binding domain) were ob- tained from Telios Pharmaceuticals. San Diego.
Construction of Plasmids Containing VLA a Subunit cDNA and Selection of Stabie lkansfectants The VLA a4 cDNA clones 4.39 (base pairs 1-1625) and 4.43 base pairs (1041-3805) subcloned into the Bluescript piasmid have been de- scribed previously (Takada et al., 1989). Assembly of the entire VLA a4 coding region into the pFneo expression vector (Saito et al., 1987) was accomplished in stepwise fashion. First, clone 439 was digested with both Sal1 (which cuts in the Bluescript multicloning site) and Xmnl (which cuts clone 4.39 at base pair 1300). while clone 4.43 was digested with BamHl (which also cuts in the Bluescript multicloning site) and Xmnl, and pFneo DNA was treated with both Sall and BarnHI. Then, the corresponding 1.3 kb Sall-Xmnl, 2.5 kb Xmnl-BamHI, and 5.9 kb pFneo digestion fragments were gel purified and ligated to- gether. The resulting construct was used to transform DH5 bacteria, and clones of the correct size and restriction pattern were selected.
To prepare a pFneo construct containing inserted control DNA, an
integrin VLA a* subunit cDNA clone was employed. Clone 2.72 (Takada and Hemler, 1989) in the Bluescript piasmid was digested completely with Sall and partially with BamHl to yield a 5.4 kb insert containing the entire VLA a2 coding sequence. After gel purification, this fragment was ligated to pFneo that had been previously cut with Sall and BamHl as above. VLA a*-pfneo and VLA a4-pFneo con- structs were transfected into the human erythroteukemic cell line K-552 by electroporation (Potter et al., 1984). Transfectants were selected by growth in RPM1 1640 supplemented with 10% FBS and 1 mg/ml geneti- tin (G-416 sulfate, Gibco Co.). After 2 weeks, transfectants were sub- cloned by limiting dilution and maintained thereafter in culture medium containing geneticin.
Adhesion of Suspended Ceils to COS and HUVEC Monolayers Confluent monolayers of HUVECs, and COS cells expressing surface &AM-l. were prepared in 48-weil cluster plates as described previ- ously (Osborn et al., 1989). When indicated, HUVECs were incubated for 24 hr with TNF-a (10 @ml, Biogen). Suspended cell lines (Ramos, HPBALL. and K-562 a2 and a4 transfectants) were labeled by over- night incubation with 2 &i/ml IeeS)methionine (Met) in Met-free RPM1 1640 supplemented with 10% FBS. After washing first with 1 mM EDNA in phosphate-buffered saline (PBS) and then with serum-free RPM1 containing 1% bovine serum albumin (BSA, Calbiochem). labeled cells were resuspended in RPMI, 1% BSA and plated on top of either HUVEC or COS monolayers in triplicate aliquots (3 x 105 ceils per well). FSjMet-labeled ceils were incubated for 10 min at room tem- perature, and unbound cells were removed by washing the plates three times with RPMI, 1% BSA, followed by centrifugation of the plates in an inverted position (560 rpm for 1 min) and finally three additional washes with RPMI, 1% BSA. Bound cells were lysed with 0.1% SDS and 0.1 N NaOH, and radioactivity in cell lysates was analyzed in a beta counter (Beckman). For inhibition experiments, labeled cells (i.e., ei- ther Ramos or K-562 a* and a4 transfer&ants) were preincubated with MAbs (5-20 ng/mi) for 20-30 min at room temperature and added directly to the previously washed cell monolayers.
Flow Cytometry Ramos cells, as well as K-562 a* and K-562 a4 transfectants, were washed twice with PBS containing 1% BSA and 4% human serum (HS, Gibco Co.), followed by preincubation of the ceils in PBS, 1% BSA, 4% HS for 30 min at mom temperature. Subsequently, 2-3 x 105 cell aliquots were incubated individually with MAbs to VLA al-as, VLA 51, and integrins 52 subunit (see above) at saturating concentra- tions in PBS, 1% BSA, 4% HS for 45 min at 4OC. Cells were washed three times with PBS, 1% BSA, 4% HS, then treated with goat anti- mouse IgG coupled to fluorescein (Cappel) for 45 min at 4OC, washed with PBS, 1% BSA, 4% HS three times, and finally resuspended in PBS, 1% BSA, 4% HS. Fluorescein-labeled cells were analyzed using a FACScan (Becton Dickinson).
Matrix Adhesion Assay Ninety-six-well microtiter plates were incubated with matrix protein (FN-40 or FN-120. as indicated in the legends to Figures 5 and 6) in 0.1 M NaHCOe for 2 hr at 3pc, and then with RPMI, 1% BSA for 1 hr at 5pc. Cells were labeted overnight either with 51Cr (50 @/ml) in RPM1 containing 10% FBS. or with pS]Met as above. After sequen- tial washes with 1 mM EmA in PBS followed by RPMI, 1% BSA, radio- actively labeled cells were resuspended in RPMI, 1% BSA and plated in triplicate on ligandcoated plates (5 x lti cells per well) for 20-30 min at 3pc. Unbound cells were removed by washing three times with RPMI, 1% BSA. Remaining bound cells were lysed with 0.1% SDS and 0.1 N NaOH, and radioactivity in cell lysates was analyzed in a gamma counter (5tCr) or a beta counter (“S).
We thank Drs. T Albino, C. Damsky, M. Ginsberg, J. Harlan, F. Sanchez-Madrid, S. Santoso, T. Springer, E. Wayner. and G. Yeh for providing antibodies used in this study. M. J. E. is a fellow of the Can- cer Research Institute (New York). This work was supported in part by National Institutes of Health grant GM33903 (M. E. H.).
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby

;;vated Endothelial VCAM-1 Binds WA-4 lntegrin
marked hdveftisemenf” in accordance with 16 U.S.C. Section 1734 solely to indicate this fact.
Received.November 17, 1969.
Anderson, D. C., and Springer, T A. (1967). Leukocyte adhesion defi- ciency: an inherited defect in the Mac-l, LFA-1, and p150,95 glycopro- teins. Annu. Rev. Med. 38, 175-194.
Bednarczyk, J., and McIntyre, B. W. (1990). A monoclonal antibody to VtA-4 a chain (CDw49).induces homotypic lymphocyte aggregation. J. Immunol., in press.
Berg, E. L., Goldstein, L. A., Jutila, M. A., Nakache, M., Picker, L. J., Streeter, f? Ft., Wu, N. W., Zhou, D., and Butcher, E. C. (1989). Homing receptors and vascular addressins: cell adhesion molecules that direct lymphocyte traffic. Immunol. Rev. 708, 1-18.
Bevilacqua, M. P, Pober, J. S., Mendrick, D. L., Cotran, R. S., and Gimbrone, M. A. (1987). Identification of an inducible endothelial- leukocyte adhesion molecule. Proc. Natl. Acad. Sci. USA 84, 9238- 9242.
Bevilacqua, M. P, Stengelin, S., Gimbrome, M. A., and Seed, B. (1989). Endothelial leukocyte adhesion molecule 1: an inducible recep- tor for neutrophils related to complement regulatory proteins and lec- tins. Science 243, 1160-1185.
Boyd, A. W., Wawryk, S. O., Burns, G. F., and Fecondo, J. V (1966). In- tercellular adhesion molecule 1 (ICAM-1) has a central role in cell- cell contact-mediated immune mechanisms. Proc. Natl. Acad. Sci. USA 85, 30953099.
Clayberger, C., Krensky, A. M., McIntyre, B. W., Keller, T D., Parham, f?, Brodsky, F. Linn, D. J., and Evans, E. L. (1967). Identification and characterization of two novel lymphocyte function-associated anti- gens, L24 and L25. J. Immunol. 738, 1510-1514.
Collins, T, Lapierre, L. A., Fiers, W., Strominger, J. L., and Pober, J. S. (1986). Recombinant human tumour necrosis factor increases mRNA levels and surface expression of HLAA,B antigens in vascular en- dothelial cells and dermal fibroblasts in vitro. Proc. Natl. Acad. Sci. USA 83, 446-450.
Dustin, M. L., and Springer, T A. (1968). Lymphocyte function- associated antigen-l (LFA-1) interaction with intercellular adhesion molecule-l (ICAM-1) is one of at least three mechanisms for lympho- cyte adhesion to cultured endothelial cells. J. Cell Biol. 107 321-331.
Dustin, M. L., and Springer, T A. (1969). Linkage between the T cell antigen receptor and LFA-1 regulates adhesion and deadhesion. Na- ture 347, 619-624.
Dustin, M. L., Rothlein, R., Bhan, A. K., Dinarello, C. A., and Springer, T A. (1966). Induction by P-1 and interferon-y: tissue distribution, bio- chemistry, and function of a natural adherence molecule (ICAM-1). J. Immunol. 737, 245-254.
Fradet, Y., Cordon-Cardo, C., Thomson, T., Daly, M. E., Whitmore, W. F, Jr., Lloyd, K. 0.. Melamed, M. R., and Old, L. J. (1964). Cell sur- face antigens of human bladder cancer defined by mouse monoclonal antibodies. Proc. Natl. Acad. Sci. USA 87, 224-228.
Godfrey, H. P, Canfield, L. S., Kindler, H. L., Angadi, C. V., Tomasek, J. J., and Goodman, J. W. (1966). Production of a fibronectin-associ- ated lymphokine by cloned mouse T cells. J. Immunol. 747, 1506-1515.
Groux. H., Huet, S., Valentin, H., Pham, D.. and Bernard, A. (1969). Suppressor effects and cyclic AMP accumulation by the CD29 mole- cule of CD4+ lymphocytes. Nature 339, 152-164.
Hamann, A., Jablonski-Westrich, D., Duijvestijn, A., Butcher, E. C., Baisch, H., Harder, R.. and Theile. H.-G. (1966). Evidence for an acces- sory role of LFA-1 in lymphocyte-high endothelium interaction during homing. J. Immunol. 740, 693699.
Harlan, J. M. (1965). Leukocyte-endothelial interactions. Blood 65, 513-525.
Haskard, D., Cavender, D., Beatty, l?, Springer, T., and Ziff, M. (1986). T lymphocyte adhesion to endothelial cells: mechanisms demon- strated by anti-LFA-1 monoclonal antibodies. J. Immunol. 137, 2901- 2906.
Hemler, M. E. (1990). VLA proteins in the integrin family: structures, functions and their role on leukocytes. Annu. Rev. Immunol. 8, 365 400.
Hemler, M. E., Ware, C. F, and Strominger, J. L. (1963). Characteriza- tion of a novel differentiation antigen complex recognized by a mono- clonal antibody (A-lA5): unique activation-specific molecular forms on stimulated T cells. J. Immunol. 131, 334-340.
Hemler, M. E., Sanchez-Madrid, F, Flotte, T. J., Krensky, A. M., Burakoff. S. J., Bhan, A. K., Springer, T. A., and Strominger, J. L. (1984). Glycoproteins of 210,000 and 130,000 m.w. on activated T cells: cell distribution and antigenic relation to components on resting cells and T cell lines. J. Immunol. 732, 3011-3018.
Hemler, M. E., Huang, C., and Schwarz, L. (1967a). The VLA protein family. Characterization of five distinct cell surface heterodimers each with a common 130,000 molecular weight 6 subunit. J. Biol. Chem. 282,3300-3309.
Hemler, M. E., Huang, C., Takada, Y., Schwarz, L., Strominger, J. L., and Clabby, M. L. (1967b). Characterization of the cell surface hetero- dimer VLA4 and related peptides. J. Biol. Chem. 262, 11476-11485.
Holzmann, B., and Weissman, I. L. (1969). Peyer’s patch-specific lym- phocyte homing receptors consist of a VLM-like c chain associated with either of two integrin 6 chains, one of which is novel. EMBO J. 8. 1735-1741.
Holzmann, B., McIntyre, B. W.. and Weissman, I. L. (1989). Identifica- tion of a murine Peyer’s patch-specific lymphocyte homing receptor as an integrin molecule with an a chain homologous to human VLA-4~. Cell 56, 37-46.
Hsi, B.-L., Yeh, C-J. G., Samson, M., and Fehlmann, M. (1987). Mono- clonal antibody raised against human trophoblast recognizes a novel epithelial antigen. Placenta 8, 209-217.
Hynes, R. 0. (1987). Integrins: a family of cell surface receptors. Cell 48, 549-554.
Kiefel, V, Santoso, S., Katzmann, B., and Mueller-Eckhardt, C. (1989). The BralBrb alloantigen system on human platelets. Blood 73, 2219- 2223.
Kishimoto, T. K., Larson, R. S., Corbi, A. L., Dustin, M. L., Staunton, D. E., and Springer, T A. (1969). The leucocyte integrins. Adv. Im- munol. 46, 149-182.
Leeuwenberg, J. F. M., Jeunhomme, T M. A. A., and Buurman, W. A. (1969). Induction of an activation antigen on human endothelial cells in vitro. Eur. J. Immunol. 79, 715-720.
Makgoba, M. W., Sanders, M. E.. Lute, G. E. G., Dustin, M. L., Springer, T A., Clark, E. A., Mannoni, P., and Shaw, S. (1988). ICAM- a ligand for LFA-l-dependent adhesion of B, T and myeloid cells. Na- ture 337, 66-66.
Marcantonio, E. E.. and Hynes, R. 0. (1988). Antibodies to the con- served cytoplasmic domain of the integrin 5, subunit react with pro- teins in vertebrates, invertebrates and fungi. J. Cell Biol. 708, 1765- 1772.
Marlin, S. D., and Springer, T. A. (1967). Purified intercellular adhesion molecule-l (ICAM-1) is a ligand for lymphocyte function-associated an- tigen 1 (LFA-1). Cell 51, 813-619.
Mentzer, S. J., Burakoff, S. J., and Failer, D. V. (1986). Adhesion of T lymphocytes to human endothelial cells is regulated by the LFA-1 mem- brane molecule. J. Cell Physiol. 126, 265-290.
Osborn, L., Hession, C., Tizard, R., Vassallo, C., Luhowskyj, S., Chi- Rosso, G., and Lobb, R. (1989). Direct cloning of vascular cell adhesion molecule 1, a cytokine-induced endothelial protein that binds to lym- phocytes. Cell 59, 1203-1211.
Pals, S. T, den Otter, A.. Miedema. F., Kabel, f?, Keizer, G. D., Scheper, R. J., and Meijer, C. J. L. M. (1968). Evidence that leukocyte function- associated antigen-l is involved in recirculation and homing of human lymphocytes via high endothelial venules. J. Immunol. 148, 1651-1653.
Pierschbacher, M. D., Hayman, E. G., and Ruoslahti, E. (1981). Loca- tion of the cell-attachment site in fibronectin with monoclonal anti- bodies and proteolytic fragments of the molecule. Cell 26, 259-267
F’ober, J. S., Gimbrone, M. A., Lapierre, L. A., Mendrick, D. L., Fiers, W., Rothlein, R., and Springer, T A. (1968). Overlapping patterns of ac-

Cell 504
tivation of human endothelial cells by interleukinl, tumor necrosis fac- tor, and immune interferon. J. Immunol. 137, 1893-1896. Potter, H., Weir, L., and Leder, P (1984). Enhancer-dependent expres- sion of human K immunoglobulin genes introduced into mouse pre-B lymphocytes by electroporation. Proc. Natl. Acad. Sci. USA 81, 7l81- 7i65.
Roossien, F F., DeRijk. D., Bikker, A., and Roes, E. (1989). Involvement of LFA-I in lymphoma invasion and metastasis demonstrated with LFA l-deficient mutants. J. Cell Biol. 768, 1979-1986. Ruoslahti, E. (1986). Fibronectin and its receptors. Annu. Rev. Bio-
them. 57, 376413. Saito, T, Weiss, A., Miller, J., Norcross, M. A., and Germain, R. N. (1967). Specific antigen-la activation of transfected human T cells ex- pressing Ti a8-human T3 receptor complexes. Nature 325, 126-136. Sanchez-Madrid, F., Krensky, A. M., Ware, C. F, Robbins, E., Strominger, J. L.. Burakoff, S. J., and Springer, T. A. (1982). Three dis- tinct antigens associated with human T lymphocyte-mediated cytoly- sis: LFA-1, LFA2 and LFA3. Proc. Natl. Acad. Sci. USA 79.7469-7493.
Sanchez-Madrid, F, De Landaxuri, M. 0.. Morago, G., Cebrian, hf., Acevedo, A., and Bemabeu. C. (1968). VLAS: a novel polypeptide as- sociation within the VLA molecular complex: cell distribution and bio- chemical characterization. Eur. J. Immunol. 16, 1348-1349. Springer, T A., Dustin, M. L., Kishimoto, T. K., and Marlin, S. D. (1987). The lymphocyte function-associated LFAI, CD2, and LFA-3 mole- cules: cell adhesion receptors of the immune system. Annu. Rev. Im- munol. 6223-262. Staunton, D. E., Dustin, M. L.. and Springer, T. A. (1989). Functional cloning of ICAM-2, a cell adhesion ligand for LFA-1 homologous to ICAM-1. Nature 339, 61-64. Sheeter, f? R.. Berg, E. L., Rouse, B. T. N.. Bargatze, R. F., and Butcher, E. C. (1968). A tissue-specific endothelial cell molecule in- volved in lymphocyte homing. Nature 337. 41-46. Takada, Y, and Hemler, M. E. (1989). The primary structure of the VLA- 2Icollagen receptor a* subunit (platelet GP la): homology to other inte- grins and the presence of a possible collagen-binding domain. J. Cell Biol. 100, 397-407.
Takada. Y., Elites, M. J., Grouse, C., and Hemler, M. E. (1989). The pri- mary structure of the a4 subunit of VLA-4: homology to other integrins and a possible cell-cell adhesion function. EMBO J. 8, 1361-1368. van Noesel, C., Miedema, F.. Brouwer, M., de Rie, M. A., Aarden, L. A., and van Lier, R. A. W. (1986). Regulatory properties of LFA-1 a and 6 chains in human T-lymphocyte activation. Nature 333, 850-852. Wayner. E. A., Carter, W. G., Piotrowicz, R. S., and Kunicki. T J. (1968). The function of multiple extracellular matrix receptors in medi- ating cell adhesion to extraceflular matrix: preparation of monoclonai antibodies to the fibronectin receptor that specifically inhibit cell adhe sion to fibronectin and react with platelet glycoproteins Ic-lla. J. Cell Biol. 107, 1881-1891. Wayner, E. A., Garcia-Pardo, A., Humphries, M. J., McDonald, J. A., and Carter, W. G. (1989). ldenthication and characterization of the lym- phocyte adhesion receptor for an alternative cell attachment domain in plasma fibronectin. J. Cell Biol. 709, 1321-1336.




















