
Incidence of Equine Hoof Derangements in Malaysian Horse Population
Upload
khangminh22Category
view
0download
0


Equine viral arteritis: in vivo and ex vivo
pathogenesis of a European isolate
Sabrina Vairo
Thesis for obtaining the degree of Doctor in Veterinary Sciences (Ph.D.)
2013
Promoters:
Prof. H. Nauwynck
Prof. A. Scagliarini
Laboratory of Virology
Department of Virology, Parasitology and Immunology
Faculty of Veterinary Medicine, Ghent University
Salisburylaan 133, B-9820 Merelbeke

To the experimental animals, and in particular to my ten Shetland ponies, which sacrificed their l i fe for science.
I hope this PhD worthes your l ives.
A mi abuela: la roca sobre la que fundé mi moral y mis principios.
Te amo con todo mi corazón

TABLE OF CONTENTS
I. LIST OF ABBREVIATIONS
II. INTRODUCTION
1. EQUINE ARTERITIS VIRUS
1.1. History
1.2. Taxonomy
1.3. Morphology
1.3.1. General structure of the virion
1.3.2. Genomic organization
1.3.3. Nucleocapsid protein
1.3.4. Major envelope proteins
1.3.5. Minor envelope proteins
2. EQUINE ARTERITIS VIRUS - CELL INTERACTION: The replication cycle
2.1. Virus attachment and entry
2.2. Genome replication and gene expression
2.2.1. Genome translation and processing of polyproteins in non-structural proteins
2.2.2. Genome replication
2.2.3. Subgenomic-length RNA transcription and expression of structural proteins
2.3. Virus assembly, budding and release
3. EQUINE ARTERITIS VIRUS - ANIMAL INTERACTION
3.1. Prevalence
3.2. Pathogenesis of infection with North American EAV strains
3.2.1. Routes of infection
3.2.2. Pathogenesis of generalized infection following respiratory uptake of the virus
3.2.3. Pathogenesis of abortion following respiratory infection
3.2.4. Pathogenesis following venereal route
3.2.5. Mechanism of persistence in stallions
3.3. Clinical signs
3.4. Pathology
3.4.1. Gross lesions
3.4.2. Histopathology
1
3
3
3
3
5
5
6
7
7
8
9
10
11
11
12
12
13
14
14
17
17
18
19
20
20
21
24
24
24

3.5. Clinical signs and pathology of infection with European EAV strains
3.6. Immunity
3.7. Diagnosis
3.7.1. Clinical diagnosis
3.7.2. Detection of EAV virus, viral components or antibodies
4. EQUINE ARTERITIS VIRUS - UPPER RESPIRATORY MUCOSA INTERACTION:
EX VIVO MODEL
4.1. Mucosa explants in research
4.2. General characteristics of the respiratory mucosa
4.3. Histology of the upper respiratory tract mucosa
4.3.1. Respiratory epithelial components and their functions
4.3.2. Cell-cell and cell-matrix adhesions
4.3.3. Extracellular matrix
4.3.3.1. The basement membrane
4.3.3.2. The lamina propria
4.4. Mucosal immune cells
REFERENCES
III. AIMS OF THE THESIS
IV. Clinical and virological outcome of an infection with the Belgian equine arteritis virus
strain 08P178
V. Identification of target cells of a European equine arteritis virus strain in experimentally
infected ponies
VI. Development and use of a polarized equine upper respiratory tract mucosal explant
system to study the early phase of pathogenesis of a European strain of equine arteritis virus
VII. GENERAL DISCUSSION
VIII. SUMMARY-SAMENVATTING
IX. CURRICULUM VITAE
X. ACKNOWLEDGEMENTS
25
26
27
27
27
29
29
29
30
30
31
32
32
33
34
37
47
49
68
80
94
107
113
118

1
I. LIST OF ABBREVIATIONS
aa: Amino acid
BM: Basement membrane
CD: Cluster of differentiation
CF: Complement fixation test
CoV: Coronavirus
CPE: Cytopathic effect
DC: Dendritic cells
MALT: Mucosa-associated lymphoid tissue
dpi: days post inoculation
DRT: Deep respiratory tract
EAV: Equine arteritis virus
EDTA: Ethylenediamine tetraacetic acid
EHV: Equine herpesvirus
ELISA: Enzyme-linked immunosorbent assay
ER: Endoplasmic reticulum
EVA: Equine viral arteritis
FAE: Follicle-associated epithelium
GP: Glycoprotein
HIV: Human immunodeficiency virus
hpi: hours post inoculation
Ig: Immunoglobulin
IL: Interleukin
Kb: Kilobase
kDa: Kilodalton
LDV: Lactate dehydrogenase-elevating virus
mAbs: Monoclonal antibodies

2
mRNA: Messenger ribonucleic acid
N protein: Nucleocapsid protein
nm: Nanometer
nsp: Non-structural protein
ORF: Open reading frame
PBMC: Peripheral blood mononuclear cell
PCR: Polymerase Chain Reaction
pp: Polyprotein
PRRSV: Porcine reproductive and respiratory syndrome virus
RdRp: RNA-dependent RNA-polymerase
RK13: Rabbit kidney 13
RNA: Ribonucleic acid
SARS: Severe acute respiratory syndrome
sg RNA: Subgenomic ribonucleic acid
SHFV: Simian hemorrhagic fever virus
SN: Serum neutralizing
TCID50: Tissue culture infectious dose with 50% endpoint
URT: Upper respiratory tract

INTRODUCTION

Introduction _
3
1. EQUINE ARTERITIS VIRUS
1.1. History
Equine arteritis virus (EAV) is the etiological agent of the disease currently known as equine viral arteritis
(EVA). EAV was first isolated in 1953 from an outbreak of respiratory disease and abortion on a
Standardbred breeding farm near Bucyrus (Ohio, USA). However, descriptions of disease outbreaks that
most likely were EVA, have first been published in the late 18th and early 19
th centuries and were called
“pinkeye”, “infectious or epizootic cellulites”, “influenza erysipelatosa”, “pferdestaupe”, and “equine
influenza” (Bergman et al., 1913 and Pottie et al., 1888). In 1957, the agent isolated from the Ohio outbreak
was named as EAV because of the distinctive vascular lesions leading to arteritis (Doll et al., 1968). A first
and important outbreak in Europe was reported in Bern (Switzerland) in 1964 involving over 400 remount
horses (Bürki and Gerber, 1966). Recently, the number of EAV outbreaks, has drastically increased
worldwide. EVA is an economically important viral disease of equids and its prevalence is increasing,
possibly due to intensified transportation of horses and semen. Although deaths are very rare in infected
adults, acute illness may occur, pregnant mares may abort and very young foals may die of a fulminating
pneumonia and enteritis. Further, stallions may become carriers and transmit EAV during breeding (Timoney
and McCollum, 1993a). Although the global dissemination and incidence of EAV have increased during the
last decades, several aspects of its pathogenesis remain uninvestigated. A better understanding of EAV
pathogenesis can lead to an effective control of the disease and a reduction of economic losses.
1.2. Taxonomy
EAV is the prototype virus of the genus Arterivirus, family Arteriviridae, order of Nidovirales. Members of
the order Nidovirales contain a positive-sense ssRNA genome within an external lipid bilayer (envelope)
with associated proteins which encloses the internal nucleocapsid structure (Perlman et al., 2012). Virions of
Nidovirales vary in morphology from spherical to bacilliform depending on the family to which they belong
(Lai and Holmes, 2001). Based on phylogenetic analysis of the RNA-dependent RNA-polymerase (RdRp),
the order of the Nidovirales was divided in three families: Arteriviridae (1 genus), Roniviridae (1 genus) and
Coronaviridae (2 subfamilies: Coronavirinae and Torovirinae). Nidovirales cause important diseases in a
broad range of hosts including humans, other mammals, birds, shrimps, and fishes (de Groot et al., 2012;
Granzow et al., 2001; Siddell and Snijder, 2008 and Walker et al., 2005). Nidovirales are characterized by an
extraordinary genetic complexity which allows them to expand the host range and to adapt rapidly to
changing environmental conditions (Ziebuhr et al., 2000). The ORFs located in the 3’-part of the nidoviruses
genome are expressed from a nested set of subgenomic (sg) mRNAs, a property that was reflected in the
name of the virus order (nidus in Latin means nest) (Cavanagh, 1997). Since the multinuclear zinc-binding
domain (contained in nsp10) and uridylate-specific endoribonuclease domain (contained in nsp11) have not
been identified in other RNA virus families, they are used to discriminate between nidoviruses and other
RNA viruses (Posthuma et al., 2006).

Introduction
4
Besides EAV, the family of Arteriviridae contains 3 other members: lactate dehydrogenase-elevating virus
(LDV; mice), porcine reproductive and respiratory syndrome virus (PRRSV; pigs) and simian hemorrhagic
fever virus (SHFV; monkeys) (Snijder and Meulenberg, 1998). Arteriviruses have a highly restricted host
range and may cause prolonged or lifelong infections in their natural host (Timoney and McCollum, 1993a).
The ability to persist in their host suggests that they can escape to some extend from recognition and
elimination by the host immunity. However, the immune-evasion mechanisms are largely unknown.
Although only one neutralization serotype of EAV has been identified so far (Golnik et al., 1986), there is
considerable genetic variation among EAV field strains as demonstrated by comparative sequence analysis
of ORFs 2 to 7 (Hornyak et al., 2005). Since ORF 5 contains several variable regions, it became the main
target for tracing the origin of EAV strains (Balasuriya et al., 1999 and 2004a). Phylogenetic analyses based
on M (ORF 6) and N protein (ORF 7) genes confirmed EAV strain variation (Chirnside et al., 1994).
Sequence analysis of the ORF 6 was used to separate distinct groups of EAV isolates from Europe and the
USA. Comprehensive phylogenetic analyses have identified two phylogenetic groups: Group I consists of
viruses originally isolated in North America and Group II consists of viruses originally isolated in Europe
(Stadejek et al., 1999). In addition, several subgroups have been identified within each group. Particularly,
the European group can be further divided into two subgroups: EU-1 and EU-2 (Zhang et al., 2010). More
recently, North American lineage viruses have been isolated in Europe and vice versa, indicating interchange
of viruses between the two continents. In South America, isolates of both EAV groups have been identified
(González et al., 2003). EAV strains belonging to the North American group are present in Australia. This
geographic exchange is most probably the outcome of the movement of carrier stallions and/or shipment of
virus-contaminated semen.
The nucleotide identity between North American and European isolates of EAV is about 85% (Balasuriya et
al., 1995a and 2004a). A genetically very diverse strain of EAV has been isolated from the semen of a
donkey in South Africa having only 60-70% of nucleotide identity with EAV strains isolated from North
American and European horses and donkeys (Stadejek et al., 2006). Experimental infection of horses and
donkeys with the South African asinine strain has demonstrated that it is poorly transmissible to horses.
Although there is a widespread distribution of this South African strain among South African donkeys
(Paweska et al., 1997), the number of seropositive horses is very low.
Genetically, EAV remains rather stable during horizontal and vertical transmission in the course of a disease
outbreak but genetic variants can emerge during persistent infections of stallions (Balasuriya et al., 1999 and
2004a). EAV isolates may markedly vary in their severity to induce clinical signs and in their abortigenic
potential (Balasuriya and MacLachlan 2004). The genetic determinants of virulence have not yet been
defined.

Introduction _
5
Figure 1: Electron photomicrograph (a) and schematic representation (b) of EAV morphology. The
nucleocapsid core consists of a capsid that encloses the viral genome. The nucleocapsid, composed of
nucleocapsid protein (N), is surrounded by an envelope that contains 6 (glyco)proteins: envelope protein E,
GP2b, GP3, GP4, GP5 and matrix protein M. GP5 and M proteins are present in the envelope as
heterodimers and are the backbone of the envelope. GP2b, 3 and 4 are connected and form heterotrimers.
Protein E is suggested to interact with the GP2b/GP3/GP4 heterotrimer on the one hand and with the GP5/M
heterodimer and/or the nucleocapsid on the other hand.
1.3. Morphology
1.3.1. General structure of the virion
EAV is a small, spherical virus with a diameter of 40-60 nm. EAV virions contain an icosahedral
nucleocapsid core of 25 to 35 nm in diameter (Horzinek et al., 1971). This nucleocapsid consists of capsid
proteins (N) that enclose the viral genome. Virions are enclosed in an envelope with tiny surface projections
(Horzinek et al., 1971). Seven structural proteins have been identified in EAV virions: the 14-kDa
phosphorylated nucleocapsid (N) protein and six envelope proteins: glycoprotein (GP) 2b (previously named
Gs), envelope protein E, GP3, GP4, GP5 (previously named GL due to its larger dimensions compared to the
other structural proteins) and M protein (Figure 1).

Introduction
6
1.3.2. Genomic organization
The EAV genome consists of an approximately 12.7 kilobases (Kb) single-stranded positive-sense RNA
molecule and is divided in 9 open reading frames (ORFs): 2 large replicase ORFs and 7 smaller ORFs
coding for structural proteins. Besides the coding regions, the genome contains a 5’ non-coding region which
carries a cap at its 5’ end and a 3’ non-coding region to which a poly-A tail is attached (Figures 1 and 2). The
two largest 5’ ORFs, ORF 1a and ORF 1b, occupy three-quarters of the genome and overlap each other in a
small area containing a ribosomal frameshift signal (Figure 2b). ORF 1a and 1b are processed in 13 non-
structural proteins (nsps), including the RNA-dependent RNA-polymerase (RdRp; nsp9). The RNA sequence
downstream of ORF 1a and 1b contains 7 overlapping ORFs: 2a, 2b, 3, 4, 5, 6, and 7 encoding respectively
the structural proteins E, GP2b, GP3, GP4, GP5, M, and N (Figure 2) (de Vries et al., 1992).
A detailed computational analysis revealed an additional ORF which overlaps the 5’ end of ORF 5, named
ORF 5a, that is conserved in all arterivirus species. The ORF 5a is predicted to be a type III membrane
protein (59aa) and is thought to be expressed from the same subgenomic mRNA (sg mRNA5). The function
of this protein is yet to be characterized but studies, using reverse genetics, suggested that it is the eighth
structural protein of arteriviruses and may be important for arterivirus infection (Firth et al., 2011).
Figure 2: Schematic diagram of the genome organization and expression of EAV (a) and of the EAV
ORF1a/1b frameshift-directing signals: the ‘shifty’ codons (5’ GUUAAAC 3’) and RNA pseudoknot
structure (b). Adapted from Snijder and Meulenberg, 1998.

Introduction _
7
1.3.3. Nucleocapsid protein
The nucleocapsid (N) protein is a small protein, encoded by ORF 7, with a high content of basic amino acids
and a hydrophilic nature (de Vries et al., 1992). The N protein is phosphorylated and present in the virion as
a monomer (Snijder and Meulenberg, 1998). The N protein is expressed abundantly in infected cells and
constitutes about 35 to 40% of the protein moiety in the virion (de Vries et al., 1992). The nucleus normally
contains only trace amounts of EAV N protein, which targets to the nucleus immediately after translation.
The EAV N protein is localized in the nucleus as foci, most likely in nucleoli in view of the data obtained
with PRRSV (Rowland et al., 1999). Although only small quantities of the N protein are normally present in
the nucleus, all N protein molecules are initially transported to the nucleus. Genome replication and mRNA
synthesis can continue with the N protein trapped in the nucleus and, thus EAV N protein is dispensable for
viral RNA synthesis in the cytoplasm (Molenkamp et al., 2000a). However, since it plays an important role
in encapsidating the viral genomic RNA and in interacting with envelope proteins during virus assembly, N
protein is necessary for virus assembly and for production of infectious virus particles (Wieringa et al.,
2004). Since both nucleocapsid formation and budding of arteriviruses are assumed to be strictly cytoplasmic
events (Snijder and Meulenberg, 1998) the N protein has to be shuttled back to the cytoplasm to fulfill its
role in the virion biogenesis. The function of the nucleo-cytoplasmic shuttling of the arterivirus N protein is
still unclear. Firstly, nuclear shuttling frequently involves protein phosphorylation and this pathway could
thus be used to achieve an essential post-translational modification. Secondly, since nucleoli are implicated
in a variety of host cell processes (Olson et al., 2000), the nuclear/nucleolar localization of the N protein may
be part of a strategy to modulate host cell functions. Studies on Coronaviridae have also revealed nuclear and
nucleolar import of the N protein, a process that was postulated to disrupt host cell division (Wurm et al.,
2001). These observations suggest that the nuclear import of the N protein is important for a mechanism
common to other nidoviruses.
1.3.4. Major envelope proteins
The non-glycosylated membrane protein (M) and the large envelope glycoprotein GP5 are the two major
envelope proteins and are encoded by ORF 6 and 5, respectively (de Vries et al., 1992).
The M protein (16-kDa) is assumed to span the viral envelope three times with its internal trans-membrane
segments, leaving a short stretch of 10-18aa exposed at the outside of the virion (ectodomain) and an
approximately 72-residue buried at the inside (endodomain). In EAV infected cells, disulfide-linked M
protein homodimers are also observed but they are not incorporated into virions (Snijder and Meulenberg,
1998).
The GP5 protein (30 to 42-kDa; 255aa) is a heterogeneously glycosylated protein with an ectodomain (19-
116aa), three membrane-spanning domains and an endodomain of about 64 amino acids (Balasuriya and
MacLachlan, 2004). The GP5 protein expresses the neutralization determinants of the virus, all located on
the ectodomain of the GP5 protein (Balasuriya et al., 1997 and 2004b).

Introduction
8
M and GP5 proteins are present in virions in equimolar amounts. When expressed individually, GP5 and M
proteins are retained only in the ER. In contrast, when co-expressed, the M protein localizes both in ER and
the Golgi complex and the GP5 protein consistently co-localizes with the M protein in the Golgi complex
showing that their transport is dependent upon the formation of a GP5/M heterodimer. The ectodomain of
GP5 forms a disulfide bond with the M ectodomain, which mediates GP5/M heterodimerization (Snijder et
al., 2003). The GP5/M heterodimers constitute the basic protein matrix of the envelope (de Vries et al., 1992
and 1995). Further, GP5 and M proteins are indispensable for both virus assembly and the production of
infectious virus particles (Wieringa et al., 2004).
1.3.5. Minor envelope proteins
Besides the three major structural proteins, the EAV virion contains four minor envelope proteins: the GP4
(28-kDa), the GP3 (36 to 42-kDa), the GP2b (25-kDa), and the protein E (8-kDa).
GP4 and GP2b proteins are encoded by ORF 4 and 2b, respectively and are type I integral membrane
proteins with a 22-24aa residue cleaved off during transport through the ER (Wieringa et al., 2002).
The GP3 protein is encoded by ORF 3 and is a heavily glycosylated integral membrane protein with
hydrophobic sequences. Hydrophobic terminal domains anchor the GP3 protein to the membrane and no part
of its structure is detectably exposed cytoplasmically (Wieringa et al., 2002).
The protein E is a small (67aa) unglycosylated integral membrane protein encoded by ORF 2a with a central
hydrophobic domain (40aa). The E protein does not form covalently linked multimers and associates with
intracellular membranes (both the ER and Golgi complex) (Snijder et al., 1999).
GP2b, GP3, and GP4 are abundantly expressed in EAV-infected cells, but only a small fraction of them is
assembled into the virion (Wieringa et al., 2003a). GP2b, GP3 and GP4 form heterotrimers (Snijder et al.,
2003). It has been postulated that the GP2b and GP4 protein first form a heterodimer which then interacts
with GP3 protein. The GP2b/GP4/GP3 complex is finally assembled into the virion (Wieringa et al., 2003a
and 2003b). When one of the GP2b, GP3, or GP4 proteins is missing, incorporation of the remaining proteins
is blocked. Further, since absence of the E protein entirely prevents incorporation of the GP2b, GP3 and GP4
proteins into the virion, the existence of a GP2b/GP4/GP3/(E) complex was suggested (Wieringa et al.,
2004). The E protein is thought to be the component, which interacts with the GP2b/GP4/GP3 heterotrimer
on the one hand and interacts with the GP5/M heterodimer and/or the nucleocapsid, on the other hand. It has
been shown that the E, GP2b, GP3, and GP4 proteins are dispensable for the formation of virus-like particles
while they are essential for the production of infectious virus particles suggesting that the
GP2b/GP4/GP3/(E) complex may be involved in the virus attachment and cell entry process (Molenkamp et
al., 2000a).

Introduction _
9
2. EQUINE ARTERITIS VIRUS - CELL INTERACTION: The replication
cycle
EAV can be propagated in a variety of primary cell cultures such as equine macrophages (Moore et al.,
2003a), equine endothelial cells (Moore et al., 2003b), equine kidney cells (McCollum et al., 1961), and
hamster kidney cells (Wilson et al., 1962). The virus can also replicate efficiently in several continuous cell
lines such as rabbit kidney (RK13) (McCollum et al., 1962), baby hamster kidney (BHK-21) (Hyllseth,
1969), and African green monkey kidney (Vero) (Konishi et al., 1975). EAV infection of primary cells and
continuous cell lines is highly cytocidal. The cytopathic effect (CPE) is characterized by rounding of cells
and cell detachment from the culture plate surface (McCollum et al., 1962). In primary equine cells, one
replication cycle takes 4 to 6 h and maximum virus yield is obtained at 36 hours post inoculation (Moore et
al., 2002).
The replication of positive-stranded RNA (+RNA) viruses of eukaryotes is schematically presented in Figure
3 and depends on a unique process of cytoplasmic RNA-dependent RNA synthesis. A common feature is the
involvement of host cell membranes, which are often modified to accommodate the +RNA virus replication
complex (Pedersen et al., 1999). Like other viruses, EAV infection of cells involves virus attachment (Figure
3.1), entry (Figure 3.2), viral genome replication (Figure 3.3 and 3.4), mRNA transcription (Figure 3.5), viral
protein synthesis (Figure 3.6 a-b-c), virus assembly (Figure 3.7), budding (Figure 3.8 and 3.9), and release
(Figure 3.10).

Introduction
10
2.1. Virus attachment and entry
The first steps of the EAV replication cycle are viral attachment (Figure 3.1) to a specific receptor on a
susceptible cell and internalization (Figure 3.2). Virus attachment molecule(s), specific cell receptor(s) or
mechanism of fusion between EAV envelope and endosomal membrane have not yet been identified. It is
still unknown how many receptors or co-receptors are needed for EAV attachment, whether EAV utilizes the
same receptor(s) in different cells or whether different EAV strains use the same receptor(s). Therefore, the
attachment and entry processes of EAV still need further study. In analogy with many other animal RNA
viruses and in view of its recognition by neutralizing antibodies, the EAV GP5 protein has been postulated to
serve as the virus attachment protein and to mediate receptor recognition. However, exchange of the
ectodomain of the EAV GP5 protein with that of PRRSV or LDV did not alter the cell tropism of the mutant
virus (Dobbe et al., 2001). Similarly, exchange of the ectodomain of the PRRSV M protein with that of EAV
or LDV still retained their ability to infect porcine alveolar macrophages and did not acquire tropism for cells
susceptible to the respective viruses from which the foreign ectodomains were derived (Verheije et al.,
Figure 3: Schematic representation of EAV replication cycle. (3.1): attachment; (3.2): entry and
translation; (3.3 and 3.4): viral genome replication; (3.5): mRNA transcription; (3.6 a-b): viral protein
synthesis; (3.6 c): formation of the nucleocapsid; (3.7): virus assembly; (3.8 and 3.9): budding; and
(3.10): release.

Introduction _
11
2002). This suggests that, in case of arteriviruses, GP5 and M proteins are not responsible for receptor
binding. Since E, GP2b, GP3, and GP4 proteins are not required for the formation of EAV particles but are
essential for ensuring that the virus particles are infectious (Wieringa et al., 2004), it was proposed that the
GP2b/GP4/GP3/(E) complex might be involved in the EAV attachment/entry process. However, it is
possible that EAV, similarly to PRRSV, utilizes more than one attachment molecule for binding to cells and
that, as PRRSV (Delputte and Nauwynck, 2004), EAV uses different attachment molecules to attach to
different cells. Asagoe et al. (1997) showed that heparin can reduce EAV infection of RK13 cells and that
this inhibition was due to the direct interaction between heparin and EAV rather than to the interaction
between heparin and RK13 cells. Furthermore, treatment of RK13 cells with heparinase before virus
inoculation decreased EAV infection of the cells. These data suggested that, similarly to PRRSV (Delputte et
al., 2002), a heparin-like molecule on the surface of RK13 cells might serve as a cell receptor for EAV.
However, heparinase treatment of RK13 cells could not reduce EAV infection below the 13% even in the
presence of a very high concentration of heparin (Asagoe et al., 1997). This implies that, as demonstrated for
PRRSV (Delputte et al., 2002), other molecules on the cell surface might serve as EAV receptors.
Little is known on the mechanism(s) used by EAV to enter cells and to uncoat the envelope. In analogy with
PRRSV which uses a mechanism of clathrin-dependent-receptor-mediated endocytosis to enter cells
(Nauwynck et al., 1999), EAV is assumed to use a process of receptor-mediated endocytosis. Further, since
for PRRSV it was demonstrated that low pH is necessary for the fusion between the endosomal membrane
and the viral envelope and subsequent virus uncoating (Nauwynck et al., 1999), it is believed that, to uncoat
the envelope, EAV needs similar conditions.
2.2. Genome replication and gene expression
Once the viral RNA is released into the cytoplasm, arteriviruses start their replication cycle in the cell. The
EAV replication cycle commences with translation of the replicase polyproteins (RdRp) from the genome,
followed by genome replication and transcription and translation of structural proteins from subgenomic (sg)
mRNAs (Figure 3.3) (Snijder and Meulenberg, 1998).
2.2.1. Genome translation and processing of polyproteins in non-structural proteins
EAV genome translation is initiated via a cap-dependent mechanism. The EAV replicase (RdRp) is
expressed directly from the viral genome in the form of polyprotein (pp) 1a and 1ab. The pp1a is translated
directly from ORF 1a while ORF 1b translation requires a ribosomal frame-shift just before ORF 1a
translation is terminated (den Boon et al., 1991). Two RNA structures, a slippery sequence (7 nucleotides)
located upstream of the ORF 1a stop codon and a pseudo-knot structure downstream of the slippery
sequence, are considered to be essential for efficient ribosomal frame-shift (Snijder and Meulenberg, 1998)
(Figure 2). Once pp1a and pp1ab are synthesized, they are cleaved 7 and 11 times, respectively by three
different viral proteases localized in non-structural proteins (nsps) 1, 2 and 4. In total, 13 end-products (1, 2,

Introduction
12
3, 4, 5, 6, 7α, 7β, 8, 9, 10, 11, 12 nsps) and multiple processing intermediates are generated (van Aken et al.,
2006). In particular, the nsps 1 to 8 are encoded by ORF 1a and nsps 9 to 12 by ORF 1b (van Dinten et al.,
1999). Besides its proteolytic role, nsp 2, together with nsp 3 is necessary and sufficient to induce the
formation of virus-induced host cell-derived double-membrane vesicles (DMVs) from paired endoplasmic
reticulum membranes (Snijder et al., 2001). Once DMVs are created, the nsps 2, 4, 7, 7-8, 8, 9, and 10
assemble on them creating the viral replication complex (Figure 3.3c). Afterwards, the hydrophobic domains
located in nsp2, nsp3 and nsp5 mediate the association between the viral replication complex and the
intracellular membrane (Figure 3.3d) (van der Meer et al., 1998). As result of this association, the
membrane-bound scaffold, which will direct the replication and the transcription of both viral genome and sg
RNAs, is formed (Figure 3.4 and 3.5) (Snijder et al., 2001). The formation of paired membranes and,
consequently, DMVs in the perinuclear region is a typical feature of EAV and other arterivirus infections
(Pol et al., 1997).
2.2.2. Genome replication
EAV RdRp is the only viral protein required for genome replication (Molenkamp et al., 2000b). However it
is still uncertain whether other host cell proteins are involved in genomic replication. The RdRp copies (+)
genomic strands into full-length (-) genomic strands (Figure 3.4) and then utilizes (-) genomic strands as
templates to synthesize (+) genomic strands (Figure 3.5). EAV RNA synthesis is directed by the viral
replication complex assembled on the membrane of DMVs and associated with the intracellular membrane
as described in the previous section.
2.2.3. Subgenomic-lenght RNA transcription and expression of structural proteins
Structural proteins are derived through the expression of subgenomic-length RNAs (sg mRNAs). As for
genomic replication, synthesis of sg mRNAs is also directed by the RdRp complex (Molenkamp et al.,
2000b). The sg mRNAs transcription occurs at the same intracellular location and follows the same
principles of RNA synthesis (den Boon et al., 1996 and Godeny et al., 1998). The only difference is that
genomic replication is a continuous process while sg mRNA transcription involves a discontinuous
mechanism where the RdRp has to stop transcription at one site and reinitiate transcription at another site
(den Boon et al., 1995). As a result, 17 small transcription-regulating sequences (5’UCAAC3’) are created of
which only 6 will attach to sg mRNAs directing the expression of the structural proteins. Structural proteins
E, GP2b, GP3, GP4, GP5, M, and N, are derived through the expression of 2a, 2b, 3, 4, 5, 6 and 7 sg
mRNAs, respectively (Figure 3.6 a-b-c) (den Boon et al., 1991).

Introduction _
13
2.3. Virus assembly, budding and release
EAV acquires its envelope from internal membranes of the infected cell rather than from the plasma
membrane and its assembly takes place at the cytoplasmic faces of the ER and/or the Golgi complex
(Magnusson et al., 1970). All seven EAV structural proteins (E, GP2b, GP3, GP4, GP5, M, and N proteins)
are indispensable for the production of infectious progeny virus (Snijder et al., 1999 and 2003) but only the
structural proteins GP5, M, and N are essential for the formation of virus-like particles. Besides the GP5, M
and N proteins, additional unknown factors are involved in EAV particle formation as demonstrated by the
unsuccessful attempts to produce virus-like particles by co-transfection of cells with expression plasmids
encoding the EAV GP5, M and N proteins (Wieringa et al., 2004). It is generally thought that disulfide-
linked GP5/M heterodimers constitute the basic protein matrix of the envelope and, since transport of EAV
GP5 and M proteins from ER to the Golgi complex is dependent upon the formation of a GP5/M
heterodimer, the latter is a prerequisite for virus assembly (Snijder et al., 2003). The cytoplasmically exposed
domains of the GP5/M heterodimers may interact with the synthesized nucleocapsid. Thus, the EAV
nucleocapsid buds into the lumen of ER or Golgi network and acquire a lipid membrane carrying viral
envelope proteins. Since the E, GP2b, GP3, and GP4 proteins are also integral membrane proteins and are
anchored to the lipid membrane of ER or Golgi complex, the EAV nucleocapsid also acquires these envelope
proteins forming virus particles. Afterwards, virus particles are transported from the intracellular
compartments to the plasma membrane where they are released via exocytosis. After the release of virus
particles, the non-covalent linkage between the GP3 protein and the GP2b/GP4 heterodimer becomes
covalently linked forming the GP2b/GP4/GP3 heterotrimers (Figure 3.7 to 3.10) (Wieringa et al., 2003a).

Introduction
14
3. EQUINE ARTERITIS VIRUS - ANIMAL INTERACTION
3.1. Prevalence
EVA is an infectious disease of Equidae. Antibodies to EAV have been reported in horses, ponies, donkeys
and mules. Illness occurs mainly among horses and ponies but clinical signs have also been reported in
experimentally infected donkeys (McCollum et al., 1995 and Timoney and McCollum, 1993a). EAV-
specific neutralizing antibodies were detected in 51 zebras (24%) of the Burchell’s zebra population in the
Serengeti National Park (Borchers et al., 2005) but not in free-ranging or captive zebra species in southern
Africa (Paweska et al., 1997). There is only one reported EAV infection in alpaca through the detection of
EAV nucleic acid from aborted fetal tissues by reverse-transcription polymerase chain reaction (RT-PCR)
and the demonstration of neutralization antibodies in a high percentage of its cohorts (Weber et al., 2006).
Retrospective serological investigations revealed that the virus was present in nearly all countries before the
disease was recognized. Antibodies to EAV have been found in most countries where sero-surveillance has
been carried out. Seropositive horses have been reported in North and South America, Europe, Asia, Africa
and Australia. This virus has not been reported in Iceland and Japan. In the Netherlands, examination of sera
collected from horses between 1963 and 1966 and from 1972 onwards showed an incidence of EAV
infection of 14%. Equine sera collected in Europe between 1966 and 1976 demonstrated that, in Austria,
England and France 59%, 14% and 14% of the horses were seropositive, respectively. In Africa, the rate of
infection was high in Morocco (52.5%), average in Egypt (14%) and low in Ethiopia and Senegal (7.7% and
2.5%, respectively) (Moraillon and Moraillon, 1978). Further, examination of stored sera demonstrated that
EAV has been present in Australia since at least 1975 (Huntington et al.,1990). EAV (European strain) was
introduced in South Africa in 1981 by importing a Lipizzaner stallion from Yugoslavia (Guthrie et al., 2003).
A schematic representation of EVA distribution is given in Figure 4.
The seroprevalence of EAV infection varies not only among countries but also among equine breeds. In the
USA, the infection is particularly common among Standardbreds. In a survey done in the USA in 2001, 85%
of Standardbreds, 5% of Thoroughbreds, 0.6% of Quarter horses, and 3.6% of Warmblood horses had
antibodies to this virus (Hullinger et al., 2001). In Europe, 55 to 93% of Austrian Warmblood stallions are
positive for antibodies to EAV (Newton et al., 1999). Breed-related differences in seroprevalence might be
due to genetic differences, but they are more likely to be caused by different management practices. In
experimentally infected horses, the breed has no apparent effect on susceptibility to infection or on the
establishment of carrier’s state.
In 1964 a very large EVA outbreak occurred in Switzerland where over 400 horses showed clinical signs
(Bürki and Gerber, 1966). In 1984, a widespread EVA outbreak occurred in 41 Thoroughbred breeding
farms in Kentucky (Timoney, 1984). As a result, concerns about EVA and its economic impact in equine
industry increased. In 1986 and 1995, EAV was isolated from an outbreak of epidemic abortion of mares in
Germany (Eichhorn et al., 1995 and Golnik et al., 1986). During 1992, a widespread epidemic occurred in a
riding center in Spain with a total of 31 out of 186 horses showing severe clinical signs (Monreal, et al.,

Introduction _
15
1995). In 1993, six premises and around 100 horses were infected in UK (Wood et al., 1995). Between 1997
and 1999, three different fatal outbreaks resulted in three dead foals and an aborted fetus in Denmark (Larsen
et al., 2001). Between 1998 and 2000, eight cases of abortion in Hungary in six different herds were
attributed to EAV infection (Szeredi et al., 2005). In 2006, outbreaks occurred in a breeding farm of Quarter
Horses in New Mexico in the USA and subsequently extended to premises in five other states (Kansas,
Montana, Oklahoma, Utah, and Alabama) where a quarantine period of 7 months was subsequently imposed.
In France, an outbreak was registered in the summer of 2007. Both incidences were associated with artificial
insemination of cool-shipped semen (Holyoak et al., 2008). The global dissemination and rising incidence of
EAV probably reflects the intensified national and international movement of horses for competition and
breeding.
In Belgium, a first serological survey carried out between 1997 and 1998 showed that 16 out of 165 sera
(10%) tested positive for antibodies against EAV (Lauwers, 1999; University of Ghent, unpublished data).
Equine sera collected between 2009 and 2010 demonstrated that the seroprevalence of EAV had increased to
29% (Lauwers, 2011; University of Ghent, unpublished data). The first disease outbreak of EVA abortion
occurred in Belgium in an Arabian stud farm in 2001 (Van der Meulen et al., 2001). A second outbreak in
Belgium occurred in 2008 (Gryspeerdt et al., 2009) and started with one foal that suffered from acute
dyspnea and died four days after birth. Three weeks later, another foal, born on the same farm in a healthy
condition but with placental edema, showed acute respiratory distress, severe dyspnea at the age of 10 days
and died shortly thereafter. Post-mortem examination of both foals revealed consolidated diaphragmatic lung
lobes with compensatory emphysema. Histologic lesions consisted of a mild acute interstitial pneumonia.
Microscopic examination of the allantochorion of the second foal showed a focal necrotizing vasculitis, and
immunolabeling demonstrated the presence of EAV-antigens in chorionic vascular endothelial cells and
macrophages.

Introduction
16
Figure 4: Schematic representation of EAV cases reported in the World Organization for Animal Health
(OIE) manual of 2012 (a) worldwide and (b) in Europe.
a
b

Introduction _
17
3.2. Pathogenesis of infection with North American EAV strains
3.2.1. Routes of infection
A schematic representation of the routes of infection is given in Figure 5.
The two major routes are the respiratory and venereal ones.
Aerosol spreading is the most important and main route of EAV spreading during an outbreak of the disease.
Throughout the acute phase of infection, significant amounts of virus are shed in the respiratory tract fluids.
Direct and close contact appears to be necessary for aerosol transmission (Timoney, 1988).
Sexual transmission is another important route of virus infection. The virus can be transmitted venereally by
mares and stallions not only during the acute phase but also through persistently infected stallions (Timoney
et al., 1986 and 1987). From 30 to 70% of infected stallions become persistently infected and constantly shed
the virus in their semen. The virus is associated with the sperm-rich and not the pre-ejaculatory fluid fraction
of semen. Duration of virus persistence can vary, carrier stallions can stop shedding virus in their semen
weeks to years after infection with no evidence of reversion to a shedding status later. Frequency of the
carrier state varies between different groups of stallions but no breed predisposition was demonstrated
(Timoney et al., 1986 and 1987). EAV can efficiently spread through artificial insemination and the use of
fresh or frozen semen. Approximately 85 to 100% of seronegative mares become infected when they are
bred to persistently infected stallions or artificially inseminated with semen containing virus. The venereal
infection and transmission is most relevant from an epidemiological point of view. Reciprocal venereal
transmission from an acutely infected mare to a seronegative stallion, though plausible, has not been
documented (Timoney et al., 1987). The viral shedding in semen has also been demonstrated in donkeys
(Paweska et al., 1996). A carrier state has never been reported in mares, geldings or sexually immature colts.
Genetic variants of EAV can emerge during the persistence of the infection in stallions (Balasuriya et al.,
2004a) giving a potential hazard for new EVA outbreaks. Therefore, carrier stallions may occupy a special
niche in the epidemiology of EAV infection since they are not only a natural virus reservoir, but can also be
a natural source of genetic diversity of EAV. Outbreaks of EAV occur when one of these variants is
transmitted to a susceptible cohort. Thus, the percentage of actively shedding carrier stallions likely
determines the prevalence of EAV infection in horse breeds (Balasuriya et al., 2004a).
Acutely infected horses also shed virus in their urine, feces, vaginal and other body secretions, although in
smaller amounts (Timoney and McCollum, 1993a). Virus can be detected from 3 to 14 days post infection
(dpi) in respiratory secretions and from 5 to 19dpi in urine (McCollum et al., 1971).
EAV can also be found in aborted fetuses, fetal membranes, placenta, and fluids of a mare that has aborted
(Timoney and McCollum, 1993b). These sources of virus may contribute to aerosol transmission but also to
indirect transmission via fomites.
EAV can also, but less commonly, be transmitted by other means such as indirect contact with virus-
contaminated fomites or by an infected teaser stallion or by a nurse mare (Timoney and McCollum, 1996). A
recent study by Broaddus et al. (2011) demonstrated that there could also be a risk of EAV transmission

Introduction
18
Figure 5: Schematic representation of routes of infection and transmission of EAV. Carrier stallions (a)
may transmit EAV to mares through infected semen. Acutely infected mares (b) shed the virus in respiratory
secretions infecting animals of the same cohort. As a result, newly infected animals shed virus through the
respiratory route, disseminating the infection (c). Further, naïve stallions can become carrier (d) and infect
mares through the venereal route. If a pregnant mare is acutely infected, beside shedding the virus in
respiratory secretions, she can abort (e). Aborted fetuses and fetal membranes contain high quantities of
EAV and, therefore, they can contribute to the EAV epidemiology (f).
resulting from in vivo embryo transfer from a donor mare inseminated with EAV infective semen (Broaddus
et al. 2011 and Timoney et al., 1987). Finally, a vertical trans-placental transmission was also documented
(Vaala et al.,1992).
3.2.2. Pathogenesis of generalized infection following respiratory uptake of the virus
The pathogenesis of North American EAV strains was studied following the distribution of viral antigens
and lesions in horses experimentally infected with virulent EAV strains via the respiratory route (Crawford
and Henson, 1972 and Del Piero, 2000). The first cells to be infected at the site of entry are not yet
characterized. At 24 hours post infection (hpi), the virus infects the alveolar macrophages and pneumocytes
with viral-antigens localized within their cytoplasm (Wilkins et al., 1995). At 48hpi, the virus can be found
in the satellite lymph nodes, especially in the bronchial lymph nodes. EAV-antigens are contained within
stromal dendrite-like cells and within the macrophages of the lymph node sinuses (Jones et al., 1957).
Viraemia starts from 2 to 3dpi depending on the animal and the virus strain. Therefore, at 3dpi, the virus
replicates in broncho-pulmonary lymph nodes, endothelium and circulating monocytes. Different authors
described a cell-associated EAV viraemia with sporadic isolations of the virus from the non-cellular fraction.

Introduction _
19
It was noticed that the highest EAV titers were associated with platelets although significant virus titers were
also present in red blood cells and PBMC fraction. EAV can be harvested from the buffy coat from 1 to
19dpi while serum can yield the virus from 1 to 9dpi (MacLachlan et al., 1996). Experimental infections with
EAV can cause a cell-associated viraemia that lasts several weeks after serum SN antibodies become
detectable (Fukunaga et al., 1981 and Neu et al., 1987). However, the systemic distribution of the virus
through viraemia results in infection of macrophages and dendritic cells of associated lymphoid tissues of
several organ systems. Approximately from 6 to 8dpi, the virus localizes within endothelium and myocytes
of blood vessels and mesothelium (McCollum et al., 1971). At 10dpi, EAV-antigens localize in endothelium,
myocytes and pericytes of blood vessels. Marginating macrophages containing intracytoplasmic EAV-
antigens may be seen, occasionally associated with infected endothelial cells. Apparently, the last site to be
invaded is the renal tubular epithelium. Abundant intracytoplasmic viral-antigens can be found within
morphologically intact and necrotic tubular epithelial cells, intratubular cellular hyaline casts, glomerular
endothelium and stellate and fusiform cells located in the renal interstitium (Del Piero, 2006). After 10dpi,
EAV-antigens have decreased in all the locations except the tunica media of small muscular arteries.
Infectious EAV is no longer detectable in most tissues after 28dpi, with the exception of the reproductive
tract of some stallions (Del Piero, 2000).
3.2.3. Pathogenesis of abortion following respiratory infection
Timoney and McCollum (1993a) showed that transplacental infection, although rare, can occur when
seronegative pregnant mares are exposed to EAV through the respiratory route. If the transplacental
transmission occurs in the first months of gestation, it will result in abortion. In the case a pregnant mare is
exposed to the virus in the last third of gestation, the foal will be congenitally infected but no abortion will
occur (Vaala et al., 1992). EAV-antigens are inconsistently detectable within tissues of aborted fetuses and
when present, they are localized within the cytoplasm of the trophoblast, allantochorionic mesenchyma,
thymus epithelium, splenic reticular cells, endothelium of visceral blood vessels, and enterocytes (Johnson et
al., 1991). Coignoul and Cheville (1984) attributed EAV-induced abortion to decreased blood supply to the
fetus as a consequence of blood vessel compression by endometrial edema, alteration of vascular tone by
various inflammatory mediators and/or virus-induced injury to the myometrium rather than to any direct
effect of the virus on the fetus itself. Further, Del Piero (2000) reported that the progesterone levels in the
mare’s serum constantly diminish from 48 to 6 hours before abortion. The decreased production of
progesterone, due to hypoxia in the placenta, combined with a local release of prostaglandins, may trigger
chorionic detachment. In addition, ischemia induced by vasculitis and thrombosis may also play a role. As a
consequence of chorionic detachment and ischemia, expulsion of an infected or uninfected fetus may follow.
Furthermore, Coignoul and Cheville (1984) indicated that virus in the fetus may reflect only contamination
attributable to increased permeability of the placenta. However, considerably higher EAV titers were found
in fetal than in maternal blood and a relative abundance of viral antigens was detected in fetal tissues, as
compared to those of the dam (MacLachlan et al, 1996) indicating that the presence of EAV in fetal tissues

Introduction
20
does not simply reflect contamination but rather points towards EAV infection of the fetus itself. Fetal
infection and pathology can activate the normal process of parturition. In addition, fetal death itself can also
release the inhibitory effects of pregnancy on the myometrium, which may result in abortion (Norwitz et al.,
1999).
3.2.4. Pathogenesis following venereal route
Up to now, there is no recorded study on the pathogenesis of EAV following venereal exposure to the virus.
However, it is presumed that the virus is taken up from vaginal mucosa and is transported to the local lymph
glands where it likely replicates and is released into the bloodstream and the lymphatic circulation (Timoney,
personal communication).
3.2.5. Mechanism of persistence in stallions
EAV reaches the male reproductive tract through viraemia resulting in an acute infection and/or in a carrier
state. Two situations have been demonstrated in the stallion: a short-term virus excretion period lasting 4-5
weeks and a long-term carrier state after clinical recovery persisting for years to life long (Timoney et al.,
1986 and 1987). In carrier stallions, EAV persists exclusively in the reproductive tract and not in other sites
of the body. The ampulla of the vas deferens and other accessory sex glands have been identified as the main
sites of viral persistence (Neu et al., 1987). The virus is associated with the sperm-rich fraction of the
ejaculate and the virus is not present in the pre-sperm fraction of semen (Timoney and McCollum, 2000).
The detailed mechanisms of this virus persistence are not clearly understood. It has been proposed that viral
persistence may be the result of two essential ingredients: the first could be that the virus has a unique
strategy of viral replication by which, instead of killing its host cell, it causes little to no damage and thus can
reside in some infected cells; the second is that the immune response does not or insufficiently reach the
virus in the host or, for unknown reasons, is unable to eliminate it (Oldstone, 1989 and 1991). Since EAV in
the reproductive tract of carrier stallions can be venereally transmitted to susceptible mares, it is clear that
EAV has not acquired a restricted tropism for the reproductive tract of stallions (McCollum et al., 1988). It
appears that humoral immunity does not prevent the establishment and the maintenance of EAV infection in
the reproductive tract of stallions. In fact, it was observed that carrier stallions have moderate to high titers of
serum neutralizing antibodies (Timoney and McCollum, 1993a) and that the sera of persistently infected
stallions consistently recognize the GP5, N and M viral proteins while sera of mares, geldings and non-
persistently infected stallions only recognize the M viral proteins (MacLachlan et al., 1998).
Comparison between sequential isolates recovered at regular intervals from the same naturally infected
stallions, revealed ongoing oligonucleotide variation in the virus, which may be another mechanism for the
establishment of the carrier state (Murphy et al., 1992). Also, down-regulation of viral antigens presented on
the host-cell surface could mask EAV for antibodies allowing the virus to persist. However, since the virus
variants emerging in carrier stallions can still be neutralized by polyclonal neutralizing antibodies, it seems
unlikely that immune escape of viral mutants plays a significant role (Balasuriya and MacLachlan, 2004).

Introduction _
21
Another hypothesis put forward to explain virus persistence may be related to the immunologically
privileged site of the male reproductive tract. It may be that EAV in the male reproductive tract is
inaccessible to circulating neutralizing antibodies or that neutralizing antibodies can only partially reduce
virus replication.
There is convincing evidence that establishment and maintenance of the carrier state in the stallion is
testosterone-dependent. When persistently infected stallions are castrated and treated with testosterone, they
continue to shed virus into the semen while castrated stallions given a placebo cease to shed virus (Little et
al., 1991). Investigation of infected prepuberal and peripuberal colts showed that the virus, after clinical
recovery, continues to replicate in the reproductive tract in a significant proportion of these colts for a
variable period of time. This occurs as well in the absence of circulating concentrations of testosterone as in
the presence of testosterone levels equivalent to those found in sexually mature stallions. However, long-
term persistent EAV infection did not occur in these colts (Holyoak et al., 1993a). The mechanism by which
testosterone contributes to the establishment and maintenance of the persistent EAV infection in stallions
remains undetermined. It is speculated that testosterone may be involved by stimulating the development of a
mature reproductive tract and in the production of androgen-dependent cells in the reproductive tract of
stallions. It thus remains to be determined which host or viral factor(s) contribute to the establishment and
maintenance of persistent EAV infection in stallions. Analysis of two carrier stallions and a number of their
male offspring did not demonstrate a significant association between inherited MHC haplotype and the
carrier state (Albright-Fraser, 1998). Moreover, the mechanism accounting for the spontaneous clearance of
EAV in some of the carrier stallions is not clear.
3.3. Clinical signs
Although the confirmed cases of clinical EVA have increased in recent years, the majority of infections are
subclinical. The occurrence of clinical signs depends on the age of the animal, the route of exposure, the
virus strain and the virus dose. The clinical signs are generally most severe in old and very young animals
and in horses that are immune-compromised or in poor condition. Clinical signs typically develop between 1
and 10dpi (Cole et al., 1986) and may include any combination or all of the following clinical signs: pyrexia
(2-12 days), depression, anorexia, conjunctivitis, with lacrimal discharge, rhinitis with serous to mucoid
nasal discharge, rhinorrhea, epiphora, lower limb edema, and stiffness of gait. Edema of the periorbital and
supraorbital areas, mid-ventral regions, scrotum, prepuce, mammary gland, and urticarial rash may also
occur. Less frequently observed are: severe respiratory distress, ataxia, mucosal eruptions, photophobia,
diarrhea, icterus, submaxillary lymphadenopathy, and intermandibular and shoulder edema may be present.
In general, animals recover completely (Timoney and McCollum, 1993a and 1996). A schematic
representation of the main and most common clinical signs, laboratory findings and localization of EVA
following respiratory infection is given in Figure 6.
The abortion rate varies from 10% to 50-60% and an outbreak may be characterized as “epidemic abortion”.
Susceptible mares infected by EAV between 2 and 11 months of gestation can experience abortion and the

Introduction
22
ages of the aborted fetuses may range from 90 to 337 days. At the time of abortion, mares usually exhibit no
clear signs but it is possible that they have manifested anorexia, lameness, fever (41°C), conjunctivitis, and
nasal discharge prior to abortion (Timoney and McCollum, 1993a). Normally, abortion can occur during
either the acute stage of the infection or soon thereafter (indicatively between 1 to 3 weeks following
exposure to the virus). There is no evidence that mares can abort more than once due to EAV infection
(Timoney and McCollum, 1987). Experimentally, mares aborted fetuses enveloped within their fetal
membranes between 10 and 12dpi (Wada et al., 1996) while non-inoculated in contact mares aborted 23 to
57 days after the infection had been started in the inoculated mares (Cole et al., 1986).
While mortality is very rare in healthy adult horses, it is nearly 100% in newborn foals. Thus, EAV infected
neonates, not protected by maternal immunity, may die suddenly or shortly after showing severe respiratory
distress (Timoney and McCollum, 1993a and 1996). Foals infected within a few months of age may develop
a life-threatening pneumonia or pneumoenteritis.
After experimental EAV infection and particularly during the acute stage, stallions may undergo a period of
temporary subfertility, associated with reduced libido, increase of scrotal temperature and change in sperm
quality as manifested by decrease in motility, concentration, and percentage of morphologically normal
spermatozoa. These abnormalities may persist for up to 16 weeks before returning to pre-exposure levels
(Neu et al., 1992). Semen quality is normal in persistently infected stallions, despite presence of the virus.
Venereal infection of mares by persistently infected stallions may result in decreased fertility at the initial
cycle, but it does not appear to result in subsequent fertility problems (Timoney and McCollum, 1993a).

Introduction _
23
Fig
ure
6:
Sch
emat
ic r
epre
sen
tati
on
of
clin
ical
and l
abora
tory
fin
din
gs
and l
oca
liza
tion o
f E
VA
foll
ow
ing r
esp
irat
ory
infe
ctio
n.
Ver
tica
l bar
s re
pre
sent
the
chro
nolo
gic
al o
ccurr
ence
of
the
resp
ecti
ve
clin
ical
or
labora
tory
fi
ndin
gs
and th
e dis
trib
uti
on o
f vir
us
in b
od
y ti
ssues
an
d
secr
etio
ns
(ad
apte
d fr
om
Bal
asuri
ya
and M
acL
achla
n,
2004).

Introduction
24
3.4. Pathology
The North American EAV isolates appear to differ in virulence and, consequently, induce lesions that differ
in severity (McCollum, 1981). Data on gross and histological lesions result from studies of natural and
experimental infections with North American strains of EAV.
3.4.1. Gross lesions
The gross and microscopic lesions reflect the extensive and considerable vascular damage at the level of
lymphatic vessels, large and small arteries, veins and capillaries. Edema, congestion, and hemorrhages in the
subcutaneous tissues, lymph nodes, and viscera are the most frequently observed gross lesions. The body
cavities may contain moderate to abundant amounts of peritoneal, pleural, and pericardial clear to yellowish
exudate. Congestion and lymphadenomegaly, edema, and hemorrhages can be observed along the course of
the colonic and cecal vessels and are also evident in systemic organs. In lymph nodes, there may be a
prominent subcapsular sinus and dilated medullary sinuses. Lungs are wet and increased in weight, with
edema, emphysema, interstitial pneumonia and show a prominent lobular pattern. These findings are more
severe in infected neonates. When congestion and hemorrhages are present, lungs can be multifocally or
diffusely reddish. Enteritis and infarcts in the spleen have been described in fatal cases of the disease in
foals. The uterine endometrial surface of aborting mares can be swollen and diffusely congested, sometimes
with hemorrhages (Del Piero, 2006 and Prickett et al., 1972).
3.4.2. Histopathology
In general, the histopathologic lesions are observed in many organs but the blood vessels are the principal
target. Mild vascular lesions include lymphocytic infiltration and endothelial cell hypertrophy. Severe
vascular changes include (i) vasculitis with fibrinoid necrosis of the tunica media, (ii) abundant vascular and
perivascular lymphocytic and lesser granulocytic infiltration, (iii) loss of endothelium, and (iv) formation of
large fibrinocellular stratified thrombi with associated tissue infarction. Capillary vessels are often
obliterated by swollen endothelial cells, platelet thrombi, or neutrophils (Estes and Cheville, 1970).
Detailed descriptions of the histopathological changes in different organs were provided following
experimental infections with EAV North American isolates (Del Piero, 2006; Jones et al., 1957 and Prickett
et al., 1972). The type and severity of lesions described and the organs affected vary from one animal to
another and from one virus strain to another. In lungs, mild to severe interstitial pneumonia characterized by
alveolar infiltration with macrophages and neutrophils, hyaline membrane formation, and fluid-filled alveoli
are common lesions. In addition, pulmonary arteritis and phlebitis are frequently observed. Within lymphoid
organs, lymphoid follicle necrosis, edema and slight hemorrhage with histiocytic erythrophagocytosis are
common findings. In lymph node sinuses, prominent and sometimes highly pleomorphic histiocytic cells and
lymphocytes are often detected. Cardiac vasculitis with myocyte necrosis and associated EAV-antigens is
rarely observed. In the liver, a portal vasculitis consisting of a severe inflammatory cell infiltrate that erodes

Introduction _
25
the hepatocellular periportal limiting plate, is occasionally observed. Distention of the submucosal
lymphatics of the large intestine, with mild crypt and lamina propria necrosis, is sporadically observed. The
adrenal gland occasionally show multifocal vasculitis, hemorrhages and infarcts. Renal lesions occur when
the infection is at an advanced stage and consists of tubular necrosis, lymphocytic interstitial nephritis,
glomerular tuft disorganization, and hypercellularity. The dermis is occasionally involved with vasculitis
with or without association of thrombosis and ulcerative dermatitis. The nervous system is generally not
affected, although cerebral vascular necrosis has been reported in fetuses. The uterine propria and
submucosa are sometimes edematous with infiltration of neutrophils and macrophages and endothelial cell
swelling, while necrotic myocytes, and macrophages are commonly found (Jones et al., 1957 and Coignoul
and Cheville, 1984).
Experimentally infected prepuberal and peripuberal colts, euthanized between the 7th and the 14th day
following viral inoculation, show necrotizing vasculitis in the testes, epididymides, vasa deferentia,
ampullae, prostate glands, and vesicular and bulbourethral glands. This vasculitis is characterized by severe
fibrinoid necrosis of the small muscular arteries with edema and hemorrhage. In colts examined between the
28th and 180
th dpi, lymphocytic and plasmacytic infiltrates were found in the lamina propria and tunica
muscularis of the epididymides and accessory genital glands. One of the prepuberal foals, infected for 15
months, had marked lymphoplasmacytic infiltration of ampullae (Holyoak et al., 1993b).
Fetuses and fetal membranes are often expelled without premonitory signs of abortion, either autolyzed or
well preserved. Lesions in the fetus, when present, consist of mild perivascular lymphocytic infiltrate and
mild interstitial pneumonia. Mild vasculitis involving the allantochorion, brain, liver, spleen, and lung is
sporadically found. In foals, where the pulmonary lesions generally prevail, a pneumoenteric syndrome with
pathologic changes involving intestinal crypts, intestinal mucosal blood vessels and gastrointestinal tract-
associated lymphoid tissues is sometimes observed; infarcts of caecum and colon are rare (Golnik et al.,
1981 and Johnson et al., 1991).
3.5. Clinical signs and pathology of infection with European EAV strains
A natural outbreak of EAV caused by European strains may be characterized by fever, anorexia, depression,
edema of the limbs, conjunctivitis with lacrimation, rhinitis and nasal discharge, urticaria of the head,
abortion and death in foals (Hans et al., 2008). In a Bulgarian outbreak for example (Chenchev, 2008) the
first clinical signs were depression, anorexia and conjunctivitis with tearing from the medial canthus. The
body temperature increased to approximately 40-40.2°C. In some animals dermatitis was noted. Sick animals
were retarded in growth and frequently experienced secondary bacterial infections, such as salmonellosis and
pneumonia. The percentage of cases of abortion was 18%. Abortions were mainly observed during the first
part of gestation and the mares recovered without treatment. Pathological changes included: cyanosis of skin;
typical lesions of interstitial pneumonia with multiple nodular masses in the lungs which were pink to
bloody-red in color. The trachea and bronchi contained frothy fluid.

Introduction
26
3.6. Immunity
Since natural EAV infection occurs via respiratory or reproductive tract, the innate response of the mucosa
lining the respiratory and genital tracts provides the first line of defense. Although little is known on the
innate immune response evoked by EAV, it is assumed not to differ from that induced by other viruses
(Balasuriya and MacLachlan, 2004).
The adaptive immune response is divided in humoral and cell-mediated.
Antibodies have been detected after infection using the complement fixing (CF) and serum neutralization
(SN) tests. The CF antibodies peak 2–3 weeks post infection and persist for 8 months. EAV-neutralizing
antibodies, induced after natural or experimental infection or by vaccination, peak within 1-2 months
(Chirnside, 1992) and last for several years (possibly life-long). With the exception of persistently infected
carrier stallions, EAV is generally eliminated from the tissues of infected horses by 28 days after infection
and virus clearance correlates with the appearance of antibodies in serum (Timoney and McCollum, 1993a).
Carrier stallions generally have particularly high titers of neutralizing antibodies (Timoney and McCollum,
1993a and 2000 and Balasuriya and MacLachlan, 2004). SN antibodies protect horses against re-infection
upon subsequent challenge (McCollum, 1970; 1976 and 1986).
Maternally derived SN antibodies appear a few hours after colostrum uptake and are detectable until 2 to 6
months of age. The mean biological half-life of the maternally derived antibodies in foals is estimated to be
32 days (Hullinger et al., 1998).
The immunoblotting assay showed that sera of non-carrier animals most consistently recognize the
conserved M envelope protein, that the serological response of horses to the GP5 and N proteins is variable
and that the GP2b protein is rarely recognized. Persistently infected stallions and horses immunized
repeatedly with vaccines develop antibodies specific for GP3 (100 and 81%, respectively), while such
antibodies were found only in 16–22% of animals that were in contact with the virus only once. These
findings clearly suggest that GP3 is immunogenic in horses but only after repeated exposure (Balasuriya and
MacLachlan, 2004). Several laboratories have developed neutralizing monoclonal antibodies to EAV and all
of them recognize the GP5 protein using Western blot and/or immunoprecipitation assays (Glaser et al.,
1995). Several studies have demonstrated that the ectodomain (aa 19-116) of the GP5 protein is immune-
dominant and that, more specifically, the amino acid residues (i) 49 and 61; (ii) from 67 to 90 and (iii) from
99 to 106 are the sites for induction of virus neutralization (Balasuriya et al., 1993; 1995b and 1997). Taken
together, these studies demonstrated that the ectodomain of the GP5 protein contains EAV neutralization
determinants (Balasuriya et al., 1997 and 2004b). The mechanism by which antibodies act to neutralize viral
infectivity are not clear but it is shown that neutralization of EAV is complement-dependent and that the
addition of 10% guinea pig complement into the SN assay increases its sensitivity (Fukunaga et al., 1994).
Cell-mediated immunity to EAV is poorly studied. The detection of CD8+ CTL precursors from EAV-
convalescent animals indicates that cell-mediated immunity may play an important role in the ultimate

Introduction _
27
clearance of the viraemia. It was noticed that upon induction with EAV, an activation occurred of both CD8+
and CD4+ cells and an increase in the CD8
+/CD4
+ ratio which may indicate that CD8
+ cells can be
responsible for the lysis of EAV-infected target cells (Castillo-Olivares et al., 2003).
Taking into consideration that (i) EAV replication in chronically infected stallions persists for several
months to years despite high levels of circulating SN Abs; and (ii) experimental infections with EAV can
cause a cell-associated viraemia that lasts several weeks after serum SN antibodies become detectable, it can
be deduced that EAV, as well as the other members of its family, is able to escape to some extend from the
host immunity.
3.7. Diagnosis
3.7.1. Clinical diagnosis
EVA cannot be diagnosed based solely on clinical or clinical-pathological findings. Therefore, virological or
serological laboratory confirmation is required. EAV should be considered when the clinical signs include
fever, depression, edema, conjunctivitis, nasal discharge and abortion.
Differential diagnoses are numerous and include other viral infections such as EHV-1 and 4, equine
influenza viruses, equine rhinoviruses, equine infectious anemia, Hendra disease, African horse sickness, and
Getah virus. Also diseases of non-infectious nature such as urticaria, shock, purpura hemorrhagica, Hoary
alyssum toxicity, hemolytic uremic syndrome may resemble EAV infection (Del Piero, 2000).
3.7.2. Detection of EAV virus, viral components or antibodies
When EAV is suspected, virus isolation should be performed on nasopharyngeal and conjunctival swabs,
bronchoalveolar lavage fluid, unclotted citrated or EDTA blood samples from live animals (Timoney and
McCollum, 1993a). To optimize the chances of virus isolation, the relevant specimens should be obtained as
soon as possible after the onset of fever in affected horses. In cases of mortality in young foals or older
animals, virus isolation can be attempted from a variety of tissues, especially the alimentary tract with its
associated lymph nodes, the lungs, liver and spleen (McCollum et al., 1971). In outbreaks of EVA-related
abortion and/or cases of stillborn foals, placenta, fetal fluids and therefore, tissues (especially lungs) are
sources of virus (Timoney and McCollum, 1993a). Often, an aborted fetus does not contain detectable levels
of EAV-antigens and virus isolation and PCR on endometrial tissue should be performed.
The presence of EAV in body fluid, tissue samples, blood, semen and placenta can usually be detected by
virus isolation in cell culture. The most appropriate specimens for virus isolation from live animals are
nasopharyngeal swabs, conjunctival swabs and citrated or EDTA blood samples for separation of buffy coat
cells. Isolation of EAV is currently performed using RK-13 cell line. Virus isolation (VI) is the current gold
standard test, approved by the World Organization for Animal Health (OIE), for the detection of EAV in
semen from stallions and is the prescribed test for international trade. The cytopathic effect (CPE) and virus

Introduction
28
titer can vary significantly among cell lines. The viral cytopathic effects (CPE) in EAV infected cells appear
within 2–6 days and are usually characterized by rounding, vacuolation, increased optical density, refraction
and detachment from culture vessels. It should be considered that while the vast majority of isolations of
EAV are made on the first passage in cell culture, a small minority will only become evident during the
second or subsequent passages.
Immunocytochemical methods (Little et al., 1995), such as indirect immunofluorescence (Crawford and
Henson, 1972) or the avidin–biotin–peroxidase staining (Del Piero et al., 1997) have also been used to detect
viral antigens in fetal membranes, placenta, fetal tissues (especially lungs, lymph nodes, liver, and intestine)
and tissues (such as lungs, lymph nodes, heart, liver, spleen, intestine and testis or uterus) collected from
experimentally infected animals, as well as in skin biopsies from acutely infected horses.
RT-PCR can detect viral nucleic acids in clinical samples such as body fluids and semen from live horses or
tissues (fetal membranes, placenta, fetal lungs, lymph nodes, heart, liver, spleen, intestine, testis or uterus)
collected at necropsy. RT-PCR is a powerful and sensitive diagnostic technique even if genetic variability of
EAV quasi-species has to be taken into consideration. Multiple PCRs using different couples of primers are
advised to reduce false negatives (Balasuriya et al., 1998).
Finally, seropositive stallions without vaccination history or with a certificate that the animal was
seronegative before the first vaccination, should be screened for carrier status. The carrier status of a stallion
can be determined by RT- PCR on the sperm-rich fraction or by breeding two seronegative mares and to test
the mares for development of antibodies. If the mares seroconvert four weeks after breeding, then the virus
was present in the semen and the stallion is diagnosed as a carrier.
A variety of serological tests, including virus neutralization (VN), complement fixation (CF), agar gel
immuno-diffusion, enzyme-linked immunosorbent assay (ELISA), and fluorescent microsphere
immunoassay (MIA) have been used to detect antibodies to EAV. In acute cases, a four-fold increase in titers
in paired serum samples should be observed. The VN test is considered the “gold standard” for detection and
titration of antibodies to EAV and, currently, is the only validated test accepted for international trade.
Although, the VN test is highly sensitive and accurate, it has several disadvantages: it is expensive, labor-
intensive and time-consuming to perform (Westcott et al., 1995). In addition, results can vary among
laboratories when adequate attention is not paid to standardization of both test reagents and procedure.
Moreover, some sera when used undiluted or at low dilutions, may induce cytotoxicity in the cell cultures
possibly leading to misinterpretation. Further, the VN test cannot differentiate the antibody response of
vaccinated from naturally infected horses. To overcome these disadvantages, several ELISAs have been
developed. Even though they have not been as extensively validated as the VN test, some appear to offer
comparable specificity and close to equivalent sensitivity (Cho et al., 2000). The CF test is less sensitive than
the aforementioned assays, but can be used for diagnosing recent infections.

Introduction _
29
4. EQUINE ARTERITIS VIRUS - UPPER RESPIRATORY MUCOSA
INTERACTION: EX VIVO MODEL
4.1. Mucosal explants in research
Experiments in animals should be limited as much as possible for ethical reasons. In vivo work involving
large animals is difficult, as purchase and maintenance costs are high and suitable experimental animals are
often difficult to obtain. Further, for the horse, specific-pathogen-free animals are not available. Thus, there is
a constant search for in vitro models that minimize the number of in vivo experiments and the use of
experimental animals, as required by the principles of the 3 R’s (Russel and Burch, 1959). These include
reduction of the number of experimental animals, refinement of the experiments to minimize pain and distress
and replacement of experimental animals by using in vitro cell or tissue models. In some models such as in
one- or two-dimensional monolayer cultures, the possibility exists that the normal behavior of cells is
compromised if they are removed from their surrounding micro-environment and lose their normal three-
dimensional association. Explant models may fulfill many of the above mentioned requirements. In fact, the
three-dimensional structure and normal cell-cell contacts are maintained in these models, hereby providing
accessible means to mimic the in vivo situation. Explant models are powerful ex vivo tools permitting
controlled experimental manipulation while maintaining micro-environmental architecture (Anderson and
Jenkinson, 1998). Also, the use of explant models minimizes inter-animal and inter-experiment variations as
tissues can be obtained from the same animal and samples can be collected at different time points. In
general, the organ culture method can provide a valuable research tool for physiological and pathological
studies of the respiratory mucosa, for studying the effect of numerous non-infectious and infectious
respiratory agents upon interaction with the respiratory mucosa. Explant models of nasal mucosa have
already been described for a wide variety of species including pigs, horses, cows, chickens, rats, and humans
(Ali et al., 1996; Butler and Ellaway, 1972; Fanucchi et al., 1999; Glorieux et al., 2007; Steukers et al., 2011
and Vandekerckhove et al., 2009). All these models have two important features in common: (i) the
cultivation of the mucosa at an air-liquid interface which implies that the tissue is in contact with air on one
side, while it is in contact with culture medium on the other side, hence creating a physiologically relevant
environment (Middleton et al., 2003) and (ii) serum-free conditions used for all cultivation medium since
serum-supplemented medium causes an enlargement of the epithelial cells with loss of cells-cells contacts
and, consequently, reduced integrity of the epithelium and a decreased cultivation period of epithelial cells
(Glorieux et al., 2007).
4.2. General characteristics of the respiratory mucosa
The respiratory mucosa lines the respiratory tract, including nasal cavity, nasopharynx, larynx, trachea, and
bronchial tree and represents the first line of defense against pathogens such as EAV. The respiratory mucosa
consists of the luminal surface epithelium and the underlying connective tissue or lamina propria separated

Introduction
30
A B
from each other by a firm barrier, the basement membrane (BM). Epithelium, BM and lamina propria are
firm barriers that a virus needs to cross prior to cause a generalized infection.
4.3. Histology of the upper respiratory tract mucosa
4.3.1. Respiratory epithelial components and their functions
The upper respiratory tract (URT) and deep respiratory tract (DRT) are covered by an epithelium designated
as respiratory epithelium, which varies in composition depending on the site in the respiratory tract. Since
the ex vivo research part of this thesis was mainly conducted on respiratory epithelium of the nasal cavities
and nasopharynx, the respiratory epithelium of these two regions will be described here in detail.
The left and right nasal cavities are completely separated by a cartilaginous nasal septum and extend from
the external nares to the nasopharynx with which they communicate caudally (Kumar et al., 2000). The main
portion of the nasal cavities is formed by the pars respiratoria, which is characterized by ciliated
pseudostratified columnar respiratory epithelium. The nasopharynx has also a ciliated pseudostratified
columnar respiratory epithelium. An epithelium is defined as pseudostratified when, although all cells make
contact with the BM and represent a single layer of cells, the nuclei are not aligned in the same plane (Eurell
and Frappier, 2006). The nasal and nasopharyngeal ciliated pseudostratified epithelium contains several
morphologically different cell types, including basal cells, ciliated cells, brush cells and goblet cells, which
are unevenly distributed along the mucosal surface (Figure 7) (Plopper and Adams, 1993).
Figure 7. Schematic representation (A) and haematoxylin-eosin photogram (B) of the different cell types
present in the equine respiratory epithelium. (a): ciliated cell, (b): goblet cell, (c): brush cell, (d): basal cell,
(e): basement membrane, (f): mucus blanket. (A: adapted from Vandekerckhove PhD dissertation, 2011).

Introduction _
31
Basal cells may function as progenitor cells and, through division and differentiation, replace epithelial cells.
By flattening out and covering the BM, basal cells can act as a defense mechanism when neighboring
columnar cells are lost. Furthermore, they are firmly attached at the base of the epithelium to the basal
lamina by hemidesmosomes and to adjacent columnar cells by desmosomes (Evans et al., 2001).
Ciliated cells are the main cell type in the ciliated pseudostratified columnar epithelium. They are supporting,
columnar epithelial cells with 200 to 300 motile cilia and numerous microvilli projecting into the nasal
lumen (Plopper and Adams, 1993). Their rod-shaped nuclei are distributed irregularly, occupying the entire
thickness of the epithelium (Kumar et al., 2000). They have a wide apical side and a narrow basal side
directed towards the BM.
Brush cells, also known as tuft cells, represent a population of epithelial cells scattered throughout the
epithelial lining of the respiratory apparatus. Their most characteristic morphologic features are the brush of
relatively long, rigid and thick microvilli on their apical cell surface and a cytoplasm containing many
filaments. Despite numerous morphological studies, the function of brush cells remains obscure. The two
currently proposed functions of tuft cells are secretion and absorption (Sato, 2007).
Goblet cells, so called because they are shaped like a wine goblet, are widely distributed throughout the
mammalian airway tract. They are located towards the supranuclear zone of the epithelium and present a
strong periodic acid Schiff’s (PAS) reaction indicating the presence of neutral mucopolysaccharides (Kumar
et al., 2000). They are columnar epithelial cells containing membrane-bound mucous granules and secreting
mucin, which dissolves in water to form mucus. Mucus in the upper airway is important for defense,
maintenance of epithelial moisture and filtering the inhaled air by trapping inhaled particles and pathogens
(Davis and Dickey, 2008). Synchronized beating of surface cilia propels the mucus with the entrapped
materials to the naso- and oropharynx. It is then swallowed into the esophagus and hence, cleared from the
respiratory tract through the digestive apparatus (Harkema et al., 2006).
4.3.2. Cell-cell and cell-matrix adhesions
Specialized cell junctions are abundantly present at the level of cell-cell and cell-matrix contact. These
contacts can be partially disassembled and reassembled to facilitate physiological processes such as tissue
turnover, leukocyte extravasation, wound healing and tissue repair (Ebnet, 2008). Three functional groups of
cell junctions have been described: tight junctions, anchoring junctions and communicating junctions.
Tight junctions (or zonula occludens) are the sites where the membranes of two cells come very close
together. They function as a gate, limiting movement between adjacent epithelial cells and as a barrier
preventing diffusion of proteins from the apical to the basolateral surface of the cell (Van Itallie and
Anderson, 2006). Epithelial cells can alter their tight junctions to permit an increased flow of solutes and
water through breaches in the junctional barriers.

Introduction
32
Anchoring junctions are responsible for the mechanical attachment of the cytoskeleton of a cell either to the
cytoskeleton of a neighboring cell or to the extracellular matrix. Functionally, anchoring junctions can be
classified in adherens junctions (or zonula adherens) and desmosomes (or macula adherens). Adherens
junctions are responsible for cell-cell adhesion forming a continuous adhesion belt just below the tight
junctions (Gumbiner, 1996). Desmosomes are button-like points of intercellular contact and connect
intermediate filaments of adjacent cells forming a structural framework of great strength and resistance
against mechanical stress (Alberts et al., 2002).
Communicating junctions (or gap junctions) put cells in communication with their neighboring cells by
creating a channel between the membranes of contacting cells, hence connecting the cytoplasm of two
neighboring cells. Gap junctions enable small molecules to pass directly from cell to cell and are
indispensable for cell synchronization, growth, differentiation and migration.
4.3.3. Extracellular matrix
The extracellular matrix (ECM) provides the scaffolding, support and strength to tissue and organs. The ECM
contains collagens, proteoglycans and glycoproteins and provides a framework for cell adhesion and tissue
development (Tanzer, 2006). The ECM can be subdivided in the BM and the ECM of the lamina propria.
4.3.3.1. The basement membrane
The basement membrane (BM) is a thin, sheet-like structure of fibers underlying the epithelium which
individual components are regulators of biological activities such as cell growth, differentiation and
migration, influencing tissue development and repair (Aumailley and Gayraud, 1998). With a pore size of
approximately 50 nm, only small molecules are able to passively diffuse across this thin, rugged barrier
(Kalluri, 2003). Nonetheless, there are specific mechanisms that permit normal cells to traffic freely and
rapidly across the BM during morphogenesis and immune surveillance (Rowe and Weiss, 2008). Thus,
leukocytes can breach the BM barrier when they are recruited from capillaries for body defense tasks. The
BM is the fusion of two laminae: the lamina densa (or lamina basalis) and the lamina reticularis (Ham and
Cormack, 1979). Between the epithelium and the lamina densa, a clear area is visible: the lamina lucida,
which functions as the region of attachment between the epithelium and the lamina densa. The lamina densa
is a sheet of connective tissue predominantly made up of type IV collagen, adhesive glycoproteins (laminin
and fibronectin), proteoglycan (perlecan) and entactin. Type IV collagen, a non-fibrous polymer, is the major
constituent of the lamina densa and thus represents the backbone of the BM, which provides mechanical
strength and stability. The lamina reticularis is considered to be a specialized extension of the extracellular
matrix (Alberts et al., 2002). With transmission electron microscopy, it was seen that the lamina reticularis is
made up of numerous collagen fibers. The predominant collagen is type III, followed by collagens I, V, VI
and VII. In particular, collagen VII is the main structural component of anchoring fibrils, linking the lamina
densa to the underlying lamina propria (Evans et al., 2000).

Introduction _
33
4.3.3.2. The lamina propria
The lamina propria of the URT consists of proteoglycans and fibrous proteins (collagen fibers, elastin and
fibronectin), which are forming a three-dimensional network wherein cells, mostly fibroblasts, and matrix are
situated.
Proteoglycans are heavily glycosylated glycoproteins. They can resist compressive forces by forming a
highly hydrated gel. This gel forms a physicochemical environment for free diffusion of nutrients and
chemical messengers (LeBleu et al., 2007). Furthermore, proteoglycans have a major role in chemical
signaling between cells since they can bind various secreted signal molecules, such as protein growth factors,
enhancing or inhibiting their signaling activity. Proteoglycans can also bind and, as such, regulate the
activities of other types of secreted proteins, including proteolytic enzymes and protease inhibitors (Alberts
et al., 2002).
Collagens are the structural macromolecules and the most abundant proteins of the ECM. Collagens, mainly
type I, II, III, V and VI, form fibers made by alloys of fibrillar collagens. Their most important task is to give
structural support to resident cells (van der Rest and Garrone, 1991).
Fibronectins are proteins connected with collagen fibers. They facilitate cell movement by binding collagen
and cell surface integrins, hence reorganizing the cytoskeleton of the cell allowing them to migrate through
the ECM. Furthermore, they have a function in wound healing by binding to platelets during blood clotting
and facilitating cell movement to the affected area (Plopper, 2007).
Fibroblasts secrete the precursors of all the components of the extracellular matrix and a variety of fibers.
The main function of fibroblasts is to maintain the structural integrity of connective tissues by continuously
secreting precursors of the extracellular matrix.

Introduction
34
4.4. Mucosal immune cells
The mucosal surface of the respiratory tract is a cellular barrier that faces environments rich in pathogens.
Several of these pathogens have developed effective mechanisms for colonization of epithelial surfaces and
invasion of mucosal tissues. In defense, the respiratory mucosa is heavily populated with cells of the immune
system (Figure 8) (Neutra et al., 1996). The mucosal immune system consists of specialized local inductive
sites, the organized mucosa-associated lymphoid tissue or O-MALT and a widespread network of effector
sites, the diffuse mucosa-associated lymphoid tissue or D-MALT (Kraenenbuhl and Neutra, 1992). In the
respiratory epithelium, the D-MALT consists of lymphocytes in diapedesis between epithelial cells. It lies in
an intercellular position and at all depths of the epithelium with greater concentration towards the base. A
diffuse distribution of lymphocytes can also be found throughout the lamina propria and submucosa with
greater concentration just beneath the epithelium (Figure 8) (Mair et al., 1987a). D-MALT includes CD3+ T
lymphocytes, CD172a+ macrophages, dendritic cells and IgM
+ B lymphocytes.
Specifically, the CD3+ lymphocytes located between epithelial cells are mainly CD8
+ T cells, whereas the
CD4+ subset of T cells mainly localize in the stroma (Brandtzaeg, 1989 and 1996). CD8
+ T cells (or
Figure 8: Schematic representation of the different immune cell types present in the equine respiratory
epithelium and in the respiratory mucosa (adapted from Vandekerckhove Ph.D. dissertation, 2011).

Introduction _
35
cytotoxic T cells, killer T cells) are a subgroup of CD3+ T lymphocytes, capable of recognizing antigens
presented in association with major histocompatibility (MHC) class I molecules and thus inducing
dysfunction or damage or death of infected cells. CD4+ T cells (or helper T cells) are a subgroup of CD3
+ T
lymphocytes that generally do not display cytotoxic effects against pathogens. They assist other leukocytes
in processes such as maturation of B cells and activation of CD8+ T lymphocytes and macrophages
(Harrington et al., 2005).
CD172a+ macrophages are produced by differentiation of monocytes and their main function is phagocytosis
of cellular debris and pathogens. In addition, they stimulate lymphocytes and other immune cells. They are
present between the epithelium and in the lamina propria of the URT mucosa. Macrophages in the airways
are capable of effectively eliminating invading antigens or allergens by phagocytosis (Holt, 1993).
Numerous MHC class II and CD172a-positive DCs are located within the airway epithelium. DCs form a
network of antigen-presenting cells (APC) in the respiratory mucosa (Holt et al., 1990). DCs of mucosal
surfaces can serve as APCs after migration out of mucosal tissues to draining lymph nodes (Hamilton-Easton
and Eichelberger, 1995). The DC most extensively studied in the nasal mucosa is the Langerhans cell (LC),
found in both the epithelial layer and in the lamina propria (Fokkens et al., 1989 and 1991). Immature LCs
are well equipped for antigen binding and processing, but weak in stimulating resting T cells (Romani and
Schuler, 1992). After maturation, LCs lose their typical characteristics changing their structure, phenotype
and functional capacities into those of DCs. Mature DCs are weak in binding and processing antigens, but
extremely powerful in stimulating resting T cells (de la Salle et al., 1997). The DCs act as ‘sensory cells’ of
the immune system recognizing danger signals. In addition to their central role in the sensitization against
environmental antigens, their ability to activate T cells in the airway mucosa enables them to maintain a
prolonged inflammation (Huh et al., 2003). Because of their ability to take up antigens, their migratory
activities and their ability to establish close associations with T cells, DCs run the risk of becoming infected
by and serving as transport vehicles for viruses that infect mucosal surfaces (e.g. human immunodeficiency
virus) (Pope et al., 1994).
IgM+ B lymphocytes that produce immunoglobulins (Ig) have been identified in the respiratory mucosa,
lamina propria and submucosa of many domestic animals, revealing a predominance of IgA-secreting
plasma cells, over IgG-secreting cells and low numbers of IgM-secreting cells. In the lamina propria
adjacent to mucosal surfaces, plasma cells act as a first line of defense by producing IgA in a dimeric form
(Mair et al., 1987b and 1988). IgA then binds to the immunoglobulin receptor on the basolateral surface of
epithelial cells and is taken up via endocytosis. The receptor-IgA complex then passes through the cells in a
process called transcytosis and is secreted at the luminal surface of epithelial cells. After proteolysis of the
receptor, the dimeric IgA can diffuse throughout the lumen along with the secretory component of the
receptor (Snoeck et al., 2006).
Natural killer (NK) cells are an important part of the innate immune system and they contribute to host
defenses against tumors and infections, particularly with intracellular pathogens (Lodoen and Lanier, 2006).

Introduction
36
NK cells are lymphocytes which do not express a T or B cell receptor. A wide array of cell surface receptors
allows them to recognize different self-proteins and non-self-proteins on other cells. Some of these receptors
induce NK cells activation, while other receptors inhibit NK cells function (Lanier, 2005). The balance
between these activating and inhibitory signals determines whether or not an NK cell becomes activated by a
particular cell. Although cell cytotoxicity is the hallmark of NK cells and the basis for their name as ‘natural
killer’ cells, NK cells are able to produce several different cytokines, many of which are proinflammatory
(interferon-γ, tumor necrosis factor-α, granulocyte macrophage-colony stimulating factor and interleukins)
(Cooper et al., 2001). There are significant clinical data showing that human individuals with defects in NK
cell activity are more susceptible to infections, especially with herpes-type viruses (Vossen et al., 2005).
Mast cells are resident cells, present at all levels of the respiratory tract, with the greatest density in the
nasopharynx. Up to 94% of this cell population is located within the connective tissue of the lamina propria,
while only small numbers are present in the surface epithelium. They contain granules rich in histamine and
heparin and mast cells armed with IgE are common in allergic patients, both in the connective tissue and in
the epithelium. They are also involved in defense against (mainly parasitic) pathogens and wound healing
(Mair et al., 1988).
Neutrophils form an essential part of the innate immune response and are the hallmark of acute
inflammation, being the first-responders to migrate towards the site of inflammation, attracted by
chemokines (Nathan, 2006).

Introduction _
37
REFERENCES
Albright-Fraser D.G. (1998). Studies on the organization and polymorphism of equine MHC Class II
genes. Ph.D. dissertation, University of Kentucky.
Ali M., Maniscalco J. and Baraniuk J.N. (1996). Spontaneous release of submucosal gland serous and
mucous cell macromolecules from human nasal explants in vitro. Am J Physiol, 270, 595-600.
Anderson G. and Jenkinson E.J. (1998). Use of explants technology in this study of in vitro immune
responses. J Immunol Methods, 216, 155-163.
Asagoe T., Inaba Y., Jusa E.R., Kouno M., Uwatoko K. and Fukunaga Y. (1997). Effect of heparin on
infection of cells by equine arteritis virus. J Vet Med Sci, 59, 727-728.
Aumailley M. and Gayraud B. (1998). Structure and biological activity of the extracellular matrix. J Mol
Med, 76, 253-265.
Balasuriya U.B. and MacLachlan N.J. (2004). The immune response to equine arteritis virus: potential
lessons for other arteriviruses. Vet Immunol Immunop, 102, 107-129.
Balasuriya U.B., Rossitto P.V., DeMaula C.D. and MacLachlan N.J. (1993). A 29K envelope
glycoprotein of equine arteritis virus expresses neutralization determinants recognized by murine
monoclonal antibodies. J Gen Virol, 74, 2525-2529.
Balasuriya U.B., Timoney P.J., McCollum W.H. and MacLachlan N.J. (1995a). Phylogenetic analysis of
open reading frame 5 of field isolates of equine arteritis virus and identification of conserved and
nonconserved regions in the GL envelope glycoprotein. Virology, 214, 690-697.
Balasuriya U.B., Maclachlan N.J., De Vries A.A., Rossitto P.V. and Rottier P.J. (1995b). Identification
of a neutralization site in the major envelope glycoprotein (GL) of equine arteritis virus. Virology, 207,
518-527.
Balasuriya U.B., Patton J.F., Rossitto P.V., Timoney P.J., McCollum W.H. and MacLachlan N.J.
(1997). Neutralization determinants of laboratory strains and field isolates of equine arteritis virus:
identification of four neutralization sites in the amino-terminal ectodomain of the G(L) envelope
glycoprotein. Virology, 232, 114-128.
Balasuriya U.B., Evermann J.F., Hedges J.F., McKeirnan A.J., Mitten J.Q., Beyer J.C., McCollum
W.H., Timoney P.J. and MacLachlan N.J. (1998). Serologic and molecular characterization of an
abortigenic strain of equine arteritis virus isolated from infective frozen semen and an aborted equine
fetus. J Am Vet Med Assoc, 213, 1586-1589.
Balasuriya U.B., Hedges J.F., Nadler S.A., McCollum W.H., Timoney P.J. and MacLachlan N.J.
(1999). Genetic stability of equine arteritis virus during horizontal and vertical transmission in an
outbreak of equine viral arteritis. J Gen Virol, 80, 1949-1958.
Balasuriya U.B., Hedges J.F., Smalley V.L., Navarrette A., McCollum W.H., Timoney P.J., Snijder
E.J. and MacLachlan N.J. (2004a). Genetic characterization of equine arteritis virus during persistent
infection of stallions. J Gen Virol, 85, 379-390.
Balasuriya U.B., Dobbe J.C., Heidner H.W., Smalley V.L., Navarrette A., Snijder E.J. and
MacLachlan N.J. (2004b). Characterization of the neutralization determinants of equine arteritis virus
using recombinant chimeric viruses and site-specific mutagenesis of an infectious cDNA clone.
Virology, 321, 235-246.
Bergman A.M. (1913). Beiträge zur Kenntnis der Virustrager bei Rotlaufseuche, Influenza erysipelatosa,
des Pferdes. Z Infektionskrankh, 13, 161-174.
Borchers K., Wiik H., Frolich K., Ludwig H. and East M.L. (2005). Antibodies against equine
herpesviruses and equine arteritis virus in Burchell's zebras (Equus burchelli ) from the Serengeti
ecosystem. J Wild Dis, 41, 80-86.
Brandtzaeg P. (1989). Overview of the mucosal immune system. Curr Top Microbiol Immunol, 146, 13-25.
Brandtzaeg P., Jahnsen F.L. and Farstad I.N. (1996). Immune functions and immunopathology of the
mucosa of the upper respiratory airways. Acta otolaryngol, 116, 149-159.
Broaddus C.C., Balasuriya U.B., Timoney P.J., White J.L., Makloski C., Torrisi K., Payton M. and
Holyoak G.R. (2011). Infection of embryos following insemination of donor mares with equine arteritis
virus infective semen. Theriogenology, 76, 47-60.

Introduction
38
Bürki F. and Gerber H. (1966). Ein virologisch gesicherter Grossansbruck von equiner Arteritis. Berl.
Münch Tierär Woch, 79, 391-395.
Butler M. and Ellaway W.J. (1972). Comparative studies on the infectivity of avian respiratory viruses for
eggs, cell cultures and tracheal explants. J Comp Pathol, 82, 327-332.
Castillo-Olivares J., Tearle J.P., Montesso F., Westcott D., Kydd J.H., Davis-Poynter N.J. and
Hannant D. (2003). Detection of equine arteritis virus (EAV)-specific cytotoxic CD8+ T lymphocyte
precursors from EAV-infected ponies. J Gen Virol, 84, 2745-2753.
Cavanagh D. (1997). Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch Virol,
142, 629-633.
Chenchev I. (2008). Studies on the Equine Viral Arteritis Situation in Bulgaria. Second international
workshop on equine arteritis virus, Lexington (KY), 30-31.
Chirnside E.D. (1992). Equine arteritis virus: An overview. Br Vet J, 148, 181-197.
Chirnside E.D., Wearing C.M., Binns M.M. and Mumford J.A. (1994). Comparison of M and N gene
sequences distinguishes variation amongst equine arteritis virus isolates. J Gen Virol, 75, 1491-1497.
Cho H.J., Entz S.C., Deregt D., Jordan L.T., Timoney P.J. and McCollum W.H. (2000). Detection of
antibodies to equine arteritis virus by a monoclonal antibody-based blocking ELISA. Can J Vet Res, 64,
38-43.
Coignoul F.L. and Cheville N.F. (1984). Pathology of maternal genital tract, placenta and fetus in equine
viral arteritis. Vet Pathol, 21, 333-340.
Cole J.R., Hall R.F., Gosser H.S., Hendricks J.B., Pursell A.R., Senne D.A., Pearson J.E. and Gipson
C.A. (1986). Transmissibility and abortogenic effect of equine viral arteritis in mares. J Am Vet Med
Assoc, 189, 769-771.
Cooper M.A., Fehniger T.A. and Caligiuri M.A. (2001). The biology of human natural killer-cell subsets.
Trends Immunol, 22, 633-640.
Crawford T.B. and Henson J.B. (1972). Immunofluorescent, light-microscopic and immunologic studies of
equine arteritis. In: Proc Int Conf Equine Infect Dis 3, 282-302.
Davis C.W. and Dickey B.F. (2008). Regulated airway goblet cell mucin secretion. Annu Rev Physiol, 70,
487-512.
de Groot R.J., Cowley J.A., Enjuanes L., Faaberg K.S., Perlman S., Rottier P.J., Snijder E.J., Ziebuhr
J., Gorbalenya A.E. (2012). Order Nidovirales. In: Virus taxonomy. Ninth report of the international
committee on taxonomy of viruses. King A.M.Q., Adams M.J., Carstens E.B. and Lefkowitz E.J.
editors. Elsevier Academic Press, Amsterdam, 785-795.
de la Salle H., Haegel-Kronenberger H., Bausinger H., Astier A., Cazenave J.P., Fridman W.H., Sautès
C., Teillaud J.L., Hanau D. and Bieber T. (1997). Functions of Fc receptors on human dendritic
Langerhans cells. International Rev Immunol, 16, 187-203.
de Vries A.A., Chirnside E.D., Horzinek M.C. and Rottier P.J. (1992). Structural proteins of equine
arteritis virus. J Virol, 66, 6294-6303.
de Vries A.A., Post S.M., Raamsman M.J., Horzinek M.C. and Rottier P.J. (1995). The two major
envelope proteins of equine arteritis virus associate into disulfide-linked heterodimers. J Virol, 69,
4668-4674.
Del Piero F. (2000). Equine viral arteritis. Vet Pathol, 37, 287-296.
Del Piero F. (2006). Equine Viral Arteritis: Signs, Lesions, Pathogenesis and Diagnoses. In: Proc
ACVP/ASCVA Annual, 2006.
Del Piero F., Wilkins P.A., Lopez J.W., Glaser A.L., Dubovi E.J., Schlafer D.H. and Lein D.H. (1997).
Equine viral arteritis in newborn foals: clinical, pathological, serological, microbiological and
immunohistochemical observations. Equine Vet J, 29, 178-185.
Delputte P.L., Vanderheijden N., Nauwynck H.J. and Pensaert M.B. (2002). Involvement of the matrix
protein in attachment of porcine reproductive and respiratory syndrome virus to a heparinlike receptor
on porcine alveolar macrophages. J Virol, 76, 4312-4320.
Delputte P.L. and Nauwynck H.J. (2004). Porcine arterivirus infection of alveolar macrophages is
mediated by sialic acid on the virus. J Virol, 78, 8094-8101.

Introduction _
39
den Boon J.A., Snijder E.J., Chirnside E.D., de Vries A.A., Horzinek M.C. and Spaan W.J. (1991).
Equine arteritis virus is not a togavirus but belongs to the coronaviruslike superfamily. J Virol, 65,
2910-2920.
den Boon J.A., Spaan W.J. and Snijder E.J. (1995). Equine arteritis virus subgenomic RNA transcription:
UV inactivation and translation inhibition studies. Virology, 213, 364-372.
den Boon J.A., Kleijnen M.F., Spaan W.J. and Snijder E.J. (1996). Equine arteritis virus subgenomic
mRNA synthesis: analysis of leader-body junctions and replicative-form RNAs. J Virol, 70, 4291-4298.
Dobbe J.C., van der Meer Y., Spaan W.J. and Snijder E.J. (2001). Construction of chimeric arteriviruses
reveals that the ectodomain of the major glycoprotein is not the main determinant of equine arteritis
virus tropism in cell culture. Virology, 288, 283-294.
Doll E.R., Bryans J.T. and Wilson J.C. (1968). Immunization against equine viral arteritis using modified
live virus propagated in cell cultures of rabbit kidney. Cornell Vet, 48, 497.
Ebnet K. (2008). Organization of multiprotein complexes at cell-cell junctions. Histochem Cell Biol, 130, 1-
20.
Eichhorn W., Heilmann M. and Kaaden O.R. (1995). Equine viral arteritis with abortions: Serological and
virological evidence in Germany. J Vet Med, 42, 574-576.
Estes P.C. and Cheville N.F. (1970). The ultrastructure of vascular lesions in equine viral arteritis. Am J
Pathol, 58, 235-253.
Eurell J.A. and Frappier B.L. (2006). In: Textbook of Veterinary Histology. Dellmann H.D. editor, 6th
edition, Philadelphia, Lea and Febiger, 416.
Evans M.J., Van Winkle L.S., Fanucchi M.V., Toskala E., Luck E.C., Sannes E.C. and Plopper C.G.
(2000). Three-dimensional organization of the lamina reticularis in the rat tracheal basement membrane
zone. Am J Respir Cell Mol Biol, 22, 393-397.
Evans M.J., Van Winkle L.S., Fanucchi M.V. and Plopper C.G. (2001). Cellular and molecular
characteristics of basal cells in airway epithelium. Exp Lung Res, 27, 401-415.
Fanucchi M.V., Harkema J.R., Plopper C.G. and Hotchkiss J.A. (1999). In vitro culture of
microdissected rat nasal airways tissues. Am J Resp Cell Mol Biol, 20, 1274-1285.
Firth A.E., Zevenhoven-Dobbe J.C., Wills N.M., Go Y.Y., Balasuriya U.B.R., Atkins J.F., Snijder E.J.
and Posthuma C.C. (2011). Discovery of a small arterivirus gene that overlaps the GP5 coding
sequence and is important for virus production. J Gen Virol, 92, 1097-1106.
Fokkens W.J., Vroom T.M., Rijntjes E. and Mulder P.G.H. (1989). CD1a (T6), HLA-DR-expressing
cells, presumably Langerhans cells, in nasal mucosa. Allergy, 44, 167-172.
Fokkens W.J., Broekhuis-Fluitsma D.M., Rijntjes E., Vroom T.M. and Hoefsmet E.C. (1991).
Langerhans cells in nasal mucosa of patients with grass pollen allergy. Immunobiology, 182, 135-142.
Fukunaga Y., Imagawa H., Tabuchi E. and Akiyama Y. (1981). Clinical and virological findings on
experimental equine viral arteritis in horses. Bull Equine Res Inst, 18, 110-118.
Fukunaga Y., Matsumura T., Sugiura T., Wada R., Imagawa H., Kanemaru T. and Kamada M.
(1994). Use of the serum neutralization test for equine viral arteritis with different virus strains. Vet
Rec, 134, 574-576.
Glaser A.L., de Vries A.A. and Dubovi E.J. (1995). Comparison of equine arteritis virus isolates using
neutralizing monoclonal antibodies and identification of sequence changes in GL associated with
neutralization resistance. J Gen Virol, 76, 2223-2233.
Glorieux S., Van den Broeck W., Van der Meulen K., Van Reeth K., Favoreel H. and Nauwynck H. (2007). In vitro culture of porcine respiratory nasal mucosa explants for studying the interaction of
porcine viruses with the respiratory tract. J Virol Methods, 142, 105-112.
Godeny E.K., de Vries A.A., Wang X.C., Smith S.L. and de Groot R.J. (1998). Identification of the
leader-body junctions for the viral subgenomic mRNAs and organization of the simian hemorrhagic
fever virus genome: evidence for gene duplication during arterivirus evolution. J Virol, 72, 862-867.
Golnik W., Michalska Z. and Michalak T. (1981). Natural equine viral arteritis in foals. Schweiz Arch
Tierheilkd, 123, 523-533.

Introduction
40
Golnik W., Moraillon A. and Golnik J. (1986). Identification and Antigenic Comparison of Equine
Arteritis Virus Isolated from an Outbreak of Epidemic Abortion of Mares. Zoonoses and public health,
33, 413-417.
Gonzalez LJ, De la sota M, Barrandeguy M, Trono K, Ayerbe M and Tarantelli M. (2003).
Relevamiento serológico de arteritis viral equina en padrillos registrados en argentina. In Proceedings of
the VIII Congreso of the World Equine Vet Assoc, 194-195.
Granzow H., Weiland F., Fichtner D., Schutze H., Karger A., Mundt E., Dresenkamp B., Martin P.
and Mettenleiter T.C. (2001). Identification and ultrastructural characterization of a novel virus from
fish. J Gen Virol, 82, 2849-2859.
Gryspeerdt A., Chiers K., Govaere J., Vercauteren G., Ducatelle R., Van de Walle G. and Nauwynck
H. (2009). Neonatal foal death due to infection with equine arteritis virus in Belgium. Vlaams
Diergeneeskundig Tijdschr, 78, 189-193.
Gumbiner B.M. (1996). Cell adhesion: the molecular basis of tissue architecture and morphogenesis. Cell,
84, 345-357.
Guthrie A.J., Howell P.G., Hedges J.F., Bosman A.M., Balasuriya U.B., McCollum W.H., Timoney
P.J. and MacLachlan N.J. (2003). Lateral transmission of equine arteritis virus among Lipizzaner
stallions in South Africa. Equine Vet J, 35, 596-600.
Ham A.W. and Cormack D.H. (1979). Loose connective tissue and basement membrane. In: Histology.
Lippincott, Williams and Wilkins, Philadelphia, PA, 222-224.
Hamilton-Easton A. and Eichelberger M. (1995). Virus-specific antigen presentation by different subsets
of cells from lung and mediastinal lymph node tissues of influenza virus-infected mice. J Virol, 69,
6359-6366.
Hans A., Gaudaire D., Tapprest J., Guix E., Marcillaud-Pitel C., Legrand L., Pronost S., Fortier C.
Laugier S. and Zientara G. (2008). French 2007 Equine Viral Arteritis Outbreak: Epidemiological and
Pathological Features. Second international workshop on equine arteritis virus, Lexington (KY), 40.
Harkema J.R., Carey S.A. and Wagner J.G. (2006). The nose revisited: a brief review of the comparative
structure, function, and toxicologic pathology of the nasal epithelium. Toxicol Pathol, 34, 252-269.
Harrington L.E., Halton R.D., Mangan P.R., Turner H., Murphy T.L., Murphy K.M. and Weaver
C.T. (2005). Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T
helper type 1 and 2 lineages. Nat Immunol, 6, 1123-1132.
Holt P.G. (1993). Regulation of antigen-presenting cell function(s) in lung and airway tissues. Eur Respir J,
6, 120-129.
Holt P.G., Schon-Hegrad M.A. and McMenamin P.G. (1990). Dendritic cells in the respiratory tract. Int
Rev Immunol, 6, 139-149.
Holyoak G.R., Giles R.C., McCollum W.H., Little T.V. and Timoney P.J. (1993a). Pathological changes
associated with equine arteritis virus infection of the reproductive tract in prepubertal and peripubertal
colts. J Comp Pathol, 109, 281-293.
Holyoak G.R., Little T.V., McCollum W.H. and Timoney P.J. (1993b). Relationship between onset of
puberty and establishment of persistent infection with equine arteritis virus in the experimentally
infected colt. J Comp Pathol, 109, 29-46.
Holyoak G., Balasuriya U.B., Broaddus C. and Timoney P.J. (2008). Equine viral arteritis: Current status
and prevention. Theriogenology, 70, 403-414.
Hornyak A., Bakonyi T., Tekes G., Szeredi L. and Rusvai M. (2005). A novel subgroup among genotypes
of equine arteritis virus: genetic comparison of 40 strains. J Vet Med B Infect Dis Vet Public Health, 52,
112-118.
Horzinek M., Maess J. and Laufs R. (1971). Studies on the substructure of togaviruses. II. Analysis of
equine arteritis, rubella, bovine viral diarrhea, and hog cholera viruses. Arch Gesamte Virusforsch, 33,
306-318.
Huh J.C., Strickland D.H., Jahnsen F.L., Turner D.J., Thomas J.A., Napoli S., Tobagus I., Stumbles
P.A., Sly P.D. and Holt P.G. (2003). Bidirectional interactions between antigen-bearing respiratory
tract dendritic cells (DCs) and T cells precede the late phase reaction in experimental asthma: DC
activation occurs in the airway mucosa but not in the lung parenchyma. J Exp Med, 198, 19-30.

Introduction _
41
Hullinger P.J., Wilson W.D., Rossitto P.V., Patton J.F., Thurmond M.C. and MacLachlan N.J. (1998).
Passive transfer, decay and protein specificity of equine arteritis virus antibodies in an endemically-
infected standardbred herd. J Am Vet Med Assoc, 213, 839-842.
Hullinger P.J., Gardner I.A., Hietala S.K., Ferraro G.L. and MacLachlan N.J. (2001). Seroprevalence
of antibodies against equine arteritis virus in horses residing in the United States and imported horses. J
Am Vet Med Assoc, 219, 946-949.
Huntington P.J., Forman A.J. and Ellis P.M. (1990). The occurrence of equine arteritis virus in Australia.
Aust Vet J, 67, 432-435.
Hyllseth B. (1969). A plaque assay of equine arteritis virus in BHK21 cells. Arch Gesamte Virusforsch, 28,
26-33.
Johnson B., Baldwin C., Timoney P. and Ely R. (1991). Arteritis in equine fetuses aborted due to equine
arteritis virus. Vet Pathol, 28, 248.
Jones T.C., Doll E.R. and Bryans J.T. (1957). The lesion of equine viral arteritis. Cornell Vet, 47, 52-68.
Kalluri R. (2003). Basement membranes: structure, assembly and role in tumour angiogenesis. Nat Rev
Cancer, 3, 422-433.
Konishi S., Akashi H., Sentsui H. and Ogata M. (1975). Studies on equine viral arteritis. I.
Characterization of the virus and trial survey on antibody with vero cell cultures. Nippon Juigaku
Zasshi, 37, 259-267.
Kraenenbuhl J.P. and Neutra M.N. (1992). Molecular and cellular basis of immune protection of mucosal
surfaces. Rev Physiol, 72, 853-879.
Kumar P., Timoney J.F., Southgate H.H.P. and Sheoran A.S. (2000). Light and scanning electron
microscopic studies of the nasal turbinates of the horse. Anat, Histol, Embryol, 29, 103-109.
Lai M.M.C. and Holmes K.V. (2001). Coronaviruses. In: Fields Virology. Knipe D.M., Howley P.M.
Editors. Lippincott, Williams and Wilkins, Philadelphia, PA, 1163-1185.
Lanier L.L. (2005). NK cell recognition. Annu Rev Immunol, 23, 225-274.
Larsen L.E., Storgaard T. and Holm E. (2001). Phylogenetic characterization of the G(L) sequences of
equine arteritis virus isolated from semen of asymptomatic stallions and fatal cases of equine viral
arteritis in Denmark. Vet Mic, 80, 339-346.
LeBleu V.S., MacDonald B. and Kalluri R. (2007). Structure and function of basement membranes. Exp
Biol Med, 232, 1121-1129.
Little T.V., Holyoak G.R., McCollum W.H. and Timoney P.J. (1991). Output of equine arteritis virus
from persistently infected stallion is testosterone dependent. In: Proc Int Conf Equine Infect Dis 6, 225-
229.
Little T.V., Deregt D., McCollum W.H. and Timoney P.J. (1995). Evaluation of an immunocytochemical
method for rapid detection and identification of equine arteritis virus in natural cases of infection. In:
Proc Int Conf Equine Infect Dis 7, 27-31.
Lodoen M.B. and Lanier L.L. (2006). Natural killer cells as an initial defense against pathogens. Curr Opin
Immunol, 18, 391-398.
MacLachlan N.J., Balasuriya U.B., Rossitto P.V., Hullinger P.A., Patton J.F. and Wilson W.D. (1996).
Fatal experimental equine arteritis virus infection of a pregnant mare: immunohistochemical staining of
viral antigens. J Vet Diagn Invest, 8, 367-374.
MacLachlan N.J., Balasuriya U.B., Hedges J.F., Schweidler T.M., McCollum W.H., Timoney P.J.,
Hullinger P.J. and Patton J.F. (1998). Serologic response of horses to the structural proteins of equine
arteritis virus. J Vet Diagn Invest, 10, 229-236.
Magnusson P., Hyllseth B. and Marusyk H. (1970). Morphological studies on equine arteritis virus. Arch
Gesamte Virusforsch, 30, 105-112.
Mair T.S., Batten E.H., Stokes C.R. and Bourne F.J. (1987a). The histological features of the immune
system of the equine respiratory tract. J Comp Pathol, 17, 575-586.
Mair T.S., Stokes C.R. and Bourne F.J. (1987b). Quantification of immunoglobulins study in respiratory
tract secretions of the horse. Vet Immunol and Immunop, 14, 197-203.

Introduction
42
Mair T.S., Stokes C.R. and Bourne F.J. (1988). Distribution and ultrastructure of mast cells in the equine
respiratory tract. Equine Vet J, 20, 54-58.
McCollum W.H. (1970). Vaccination for equine viral arteritis. In: Proc Int Conf Equine Infect Dis 2, 143-
151.
McCollum W.H. (1976). Studies of passive immunity in foals to equine viral arteritis. Vet Mic, 1, 45-54.
McCollum W.H. (1981). Pathologic features of horses given avirulent equine arteritis virus intramuscularly.
Am J Vet Res, 42, 1218-1220.
McCollum W.H. (1986). Responses of horses vaccinated with avirulent modified-live equine arteritis virus
propagated in the E. Derm (NBL-6) cell line to nasal inoculation with virulent virus. Am J Vet Res, 47,
1931-1934.
McCollum W.H., Doll E.R., Wilson J.C. and Johnson C.B. (1961). Propagation of equine arteritis virus in
monolayer cultures of equine kidney. Am J Vet Res, 22, 731-735.
McCollum W.H., Doll E.R., Wilson J.C. and Cheatham J. (1962). Isolation and propagation of equine
arteritis virus in monolayer cell cultures of rabbit kidney. Cornell Vet, 52, 452-458.
McCollum W.H., Prickett M.E. and Bryans J.T. (1971). Temporal distribution of equine arteritis virus in
respiratory mucosa, tissues and body fluids of horses infected by inhalation. Res Vet Sci, 12, 459-464.
McCollum W.H., Timoney P.J. and Roberts A.W. (1988). Response of vaccinated and non-vaccinated
mares to artificial insemination with semen from stallions persistently infected with equine arteritis
virus. In: Proc Int Conf Equine Infect Dis 5, 13-18.
McCollum W.H., Timoney P.J. and Tengelsen L.A. (1995). Clinical, virological and serological responses
of donkeys to intranasal inoculation with the KY-84 strain of equine arteritis virus. J Comp Pathol, 112,
207-211.
Middleton A.M., Chadwick M.V., Nicholson A.G., Dewar A., Feldman C. and Wilson R. (2003).
Investigation of mycobacterial colonization and invasion of the respiratory mucosa. Thorax, 58, 246-
251.
Molenkamp R., van Tol H., Rozier B.C., van der Meer Y., Spaan W.J. and Snijder E.J. (2000a). The
arterivirus replicase is the only viral protein required for genome replication and subgenomic mRNA
transcription. J Gen Virol, 81, 2491-2496.
Molenkamp R., Rozier B.C., Greve S., Spaan W.J. and Snijder E.J. (2000b). Isolation and
characterization of an arterivirus defective interfering RNA genome. J Virol, 74, 3156-3165.
Monreal L., Villatoro A.J., Hooghuis H., Ros I. and Timoney P.J. (1995). Clinical features of the 1992
outbreak of equine viral arteritis in Spain. Equine Vet J, 27, 301-304.
Moore B.D., Balasuriya U.B., Hedges J.F. and MacLachlan N.J. (2002). Growth characteristics of a
highly virulent, a moderately virulent, and an avirulent strain of equine arteritis virus in primary equine
endothelial cells are predictive of their virulence to horses. Virology, 298, 39-44.
Moore B.D., Balasuriya U.B., Watson J.L., Bosio C.M., MacKay R.J. and MacLachlan N.J. (2003a).
Virulent and avirulent strains of equine arteritis virus induce different quantities of TNF-alpha and other
proinflammatory cytokines in alveolar and blood-derived equine macrophages. Virology, 314, 662-670.
Moore B.D., Balasuriya U.B., Nurton J.P., McCollum W.H., Timoney P.J., Guthrie A.J. and
MacLachlan N.J. (2003b). Differentiation of strains of equine arteritis virus of differing virulence to
horses by growth in equine endothelial cells. Am J Vet Res, 64, 779-784.
Moraillon A. and Moraillon R. (1978). Results of an epidemiological investigation on viral arteritis in
France and some other European and African countries. Ann Rech Vet, 9, 43-54.
Murphy T.W., McCollum W.H., Timoney P.J., Klingeborn B.W., Hyllseth B., Golnik W. and Erasmus
B. (1992). Genomic variability among globally distributed isolates of equine arteritis virus. Vet Mic, 32,
101-115.
Nathan C. (2006). Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol, 6, 173-182.
Nauwynck H.J., Duan X., Favoreel H., Van Oostveldt P. and Pensaert M.B. (1999). Entry of porcine
reproductive and respiratory syndrome virus into porcine alveolar macrophages via receptor-mediated
endocytosis. J Gen Virol, 80, 297-305.

Introduction _
43
Neu S.M., Timoney P.J. and McCollum W.H. (1987). Persistent infection of the reproductive tract in
stallions experimentally infected with equine arteritis virus. In: Proc Int Conf Equine Infect Dis 5, 14.
Neu S.M., Timoney P.J. and Lowry S.R. (1992). Changes in semen quality following experimental equine
arteritis virus infection in the stallion. Theriogenology, 37, 407-431.
Neutra M.R., Pringault E. and Kraehenbuhl J.P. (1996). Antigen sampling across epithelial barriers and
induction of mucosal immune responses. Ann Rev Immunol, 14, 275-300.
Newton J.R., Wood J.L., Castillo-Olivares F.J. and Mumford J.A. (1999). Serological surveillance of
equine viral arteritis in the United Kingdom since the outbreak in 1993. Vet Rec, 14, 5511-5516.
Norwitz E.R., Robinson J.N. and Challis J.R.G. (1999). The control of labor. N Engl J Med, 341, 660-
666.
Oldstone M.B. (1989). Viral persistence. Cell, 56, 517-520.
Oldstone M.B. (1991). Molecular anatomy of viral persistence. J Virol, 65, 6381-6386.
Olson M.O., Dundr M. and Szebeni A. (2000). The nucleolus: an old factory with unexpected capabilities.
Trends Cell Biol, 10, 189-196.
Paweska J.T., Aitchison H., Chirnside E.D. and Barnard B.J. (1996). Transmission of the South African
asinine strain of equine arteritis virus (EAV) among horses and between donkeys and horses. J Vet Res,
63,189-196.
Paweska J.T., Binns M.M., Woods P.S.A. and Chirnside E.D. (1997). A survey for antibodies to equine
arteritis virus in donkeys, mules and zebra using virus neutralisation (VN) and enzyme linked
immunosorbent assay (ELISA). Equine Vet J, 29, 40-43.
Pedersen K.W., van der Meer Y., Roos N. and Snijder E.J. (1999). Open reading frame 1a-encoded
subunits of the arterivirus replicase induce endoplasmic reticulum-derived double-membrane vesicles
which carry the viral replication complex. J Virol, 73, 2016-2026.
Perlman S., Poon L., Rottier P.J., Talbot P.J., Woo P.C. and Ziebuhr J. (2012). Family Coronaviridae.
In: Virus Taxonomy, Classification and Nomenclature of Viruses. Ninth Report of the International
Committee on Taxonomy of Viruses. King A.M.Q., Adams M.J., Carstens E.B. and Lefkowitz E.J.
editors. Elsevier Academic Press, Amsterdam, 806-828.
Plopper G. (2007). The extracellular matrix and cell adhesion. In: Cells. Lewin B., Cassimeris L., Lingappa
V. and Plopper G. editors. Jones and Bartlett, Sudbury, 645-702.
Plopper C.G. and Adams D.R. (1993). Respiratory System. In: Textbook of Veterinary Histology.
Dellmann H.D. editor, 4th edition, Philadelphia, Lea and Febiger, 136-152.
Pol J.M., Wagenaar F. and Reus J.E. (1997). Comparative morphogenesis of three PRRS virus strains. Vet
Mic, 55, 203-208.
Pope M., Betjes M.G.H., Romani N., Hirmand H., Cameron P.U., Hoffman L., Gezelter S., Schuler G.
and Steinman R.M. (1994). Conjugates of dendritic cells and memory T lymphocytes from skin
facilitate productive infection with HIV-1. Cell, 78, 389-398.
Posthuma C., Nedialkova D., Zevenhoven-Dobbe J., Blokhuis J., Gorbalenya A. and Snijder E. (2006).
Site-directed mutagenesis of the nidovirus replicative endoribonuclease NendoU exerts pleiotropic
effects on the arterivirus life cycle. J Virol, 80, 1653-1661.
Pottie A. (1888). The propagation of influenza from stallions to mares. J Comp Pathol, 1, 37-38.
Prickett M.E., McCollum W.H. and Bryans J.T. (1972). The gross and microscopic pathology observed in
horses experimentally infected with the equine arteritis virus. In: Proc Int Conf Equine Infect Dis, 3,
265-272.
Romani N. and Schuler G. (1992). The immunological properties of epidermal Langerhans cells as a part
of the dendritic cell system. Springer Semin Immunopathol, 13, 265-279.
Rowe R.G. and Weis S.J. (2008). Breaching the basement membrane: who, when and how? Trends Cell
Biol, 18, 560-574.
Rowland R.R., Kervin R., Kuckleburg C., Sperlich A. and Benfield D.A. (1999). The localization of
porcine reproductive and respiratory syndrome virus nucleocapsid protein to the nucleolus of infected
cells and identification of a potential nucleolar localization signal sequence. Virus Res, 64, 1-12.

Introduction
44
Russel W.M.S. and Burch R.L. (1959). The principles of humane experimental technique. Methuen,
London, UK.
Sato A. (2007). Tuft cells. Anat Sci Int, 82, 187-199.
Siddell S. and Snijder E.J. (2008). An introduction to nidovirus. In: Nidoviruses. Perlman S., Gallagher T.
and Snijder E.J. editors. ASM Press, Washington DC, 1-13.
Snijder E.J. and Meulenberg J.J. (1998). The molecular biology of arteriviruses. J Gen Virol, 79, 961-979.
Snijder E.J., van Tol H., Pedersen K.W., Raamsman M.J. and de Vries A.A. (1999). Identification of a
novel structural protein of arteriviruses. J Virol, 73, 6335-6345.
Snijder E.J., van Tol H., Roos N. and Pedersen K.W. (2001). Non-structural proteins 2 and 3 interact to
modify host cell membranes during the formation of the arterivirus replication complex. J Gen Virol,
82, 985-994.
Snijder E.J., Dobbe J.C. and Spaan W.J. (2003). Heterodimerization of the two major envelope proteins is
essential for arterivirus infectivity. J Virol, 77, 97-104.
Snoeck V., Peters I.R. and Cox E. (2006). The IgA system: a comparison of structure and function in
different species. Vet Res, 37, 455-467.
Stadejek T., Bjorklund H., Bascunana C.R., Ciabatti I.M., Scicluna M.T., Amaddeo D., McCollum
W.H., Autorino G.L., Timoney P.J., Paton D.J., Klingeborn B. and Belák S. (1999). Genetic
diversity of equine arteritis virus. J Gen Virol, 80, 691-699.
Stadejek T., Mittelholzer C., Oleksiewicz M.B., Paweska J. and Belak S. (2006). Highly diverse type of
equine arteritis virus (EAV) from the semen of a South African donkey: short communication. Acta Vet
Hung, 54, 263-270.
Steukers L., Vandekerckhove A., Van den Broeck W., Glorieux S. and Nauwynck H.J. (2011).
Comparative analysis of replication characteristics of BoHV-1 subtypes in bovine respiratory and
genital mucosa explants: a phylogenetic enlightenment. Vet Res, 42, 1-11.
Szeredi L., Hornyák Á., Pálfi V., Molnár T., Glávits R. and Dénes B. (2005). Study on the epidemiology
of equine arteritis virus infection with different diagnostic techniques by investigating 96 cases of
equine abortion in Hungary. Vet Mic, 108, 235-242.
Tanzer M.L. (2006). Current concepts of extracellular matrix. J Orthop Sci, 11, 326-331.
Timoney P.J. (1984). Clinical, virological and epidemiological features of the 1984 outbreak of equine viral
arteritis in the Thoroughbred population in Kentucky, USA. In: Proceedings of the Grayson Found
International Conference Thoroughbred Breeders Org on Equine Viral Arteritis, 24-33.
Timoney P.J. (1988). Equine viral arteritis: epidemiology and control. J Equine Vet Sci, 8, 54-59.
Timoney P.J. and McCollum W.H. (1987). Equine viral arteritis. Can Vet J, 28, 693-695.
Timoney P.J. and McCollum W.H. (1993a). Equine viral arteritis. Vet Clin North Am Equine Pract, 9,
295-309.
Timoney P.J. and McCollum W.H. (1993b). Equine viral arteritis in perspective in relation to International
trade. J Equine Vet Sci, 13, 50-52.
Timoney P.J. and McCollum W.H. (1996). Equine viral arteritis. Equine Vet Educ, 8, 97-100.
Timoney P.J. and McCollum W.H. (2000). Equine viral arteritis: Further characterization of the carrier
state in stallions. J Reprod Fertil Suppl, 56, 3-11.
Timoney P.J., McCollum W.H., Roberts A.W. and Murphy T.W. (1986). Demonstration of the carrier
state in naturally acquired equine arteritis virus infection in the stallion. Res Vet Sci, 41, 279-280.
Timoney P.J., McCollum W.H., Murphy T.W., Roberts A.W., Willard J.G. and Carswell G.D. (1987).
The carrier state in equine arteritis virus infection in the stallion with specific emphasis on the venereal
mode of virus transmission. J Reprod Fertil Suppl, 35, 95-102.
Vaala W.E., Hamir A.N., Dubovi E.J., Timoney P.J. and Ruiz B. (1992). Fatal, congenitally acquired
infection with equine arteritis virus in a neonatal thoroughbred. Equine Vet J, 24, 155-158.
van Aken D., Snijder E.J. and Gorbalenya A.E. (2006). Mutagenesis analysis of the nsp4 main proteinase
reveals determinants of arterivirus replicase polyprotein autoprocessing. J Virol, 80, 3428-3437.

Introduction _
45
van der Meer Y., van Tol H., Locker J.K. and Snijder E.J. (1998). ORF1a encoded replicase subunits are
involved in the membrane association of the arterivirus replication complex. J Virol, 72, 6689-6698.
Van der Meulen K., Caij A., Nauwynck H. and Pensaert M. (2001). An outbreak of equine viral arteritis
abortion in Belgium. Vlaams Diergeneeskd Tijdschr, 70, 221-222.
Van der Rest M. and Garrone R. (1991). Collagen family of proteins. The FASEB Journal, 5, 2814-2823.
van Dinten L.C., Rensen S., Gorbalenya A.E. and Snijder E.J. (1999). Proteolytic processing of the open
reading frame 1b-encoded part of arterivirus replicase is mediated by nsp4 serine protease and Is
essential for virus replication. J Virol, 73, 2027-2037.
Van Itallie C.M. and Anderson J.M. (2006). Claudins and epithelial paracellular transport. Annu Rev
Physiol, 68, 403-429.
Vandekerckhove A. (2011). Equine nasal mucosal explants, a valuable tool to study early events of the
pathogenesis of equine herpesvirus infections. Ph.D. dissertation 2011.
Vandekerckhove A., Glorieux S., Van den Broeck W., Gryspeerdt A., Van der Meulen K. and
Nauwynck H.J. (2009). In vitro culture of equine respiratory mucosa explants. Vet J, 181, 280-287.
Verheije M.H., Welting T.J., Jansen H.T., Rottier P.J. and Meulenberg J.J. (2002). Chimeric
arteriviruses generated by swapping of the M protein ectodomain rule out a role of this domain in viral
targeting. Virology, 303, 364-373.
Vossen M.T., Biezeveld M.H., de Jong M.D., Gent M.R., Baars P.A., von Rosenstiel I.A., van Lier R.A.
and Kuijpers T.W. (2005). Absence of circulating natural killer and primed CD8+ cells in life-
threatening varicella. J Infect Dis, 191, 198-206.
Wada R., Fukunaga Y., Kanemaru T. and Kondo T. (1996). Histopathological and immunofluorescent
studies on transplacental infection in experimentally induced abortion by equine arteritis virus. Zentralbl
Veterinarmed B, 43, 65-74.
Walker P.J., Bonami J.R., Boonsaeng V., Chang P.S., Cowley J.A., Enjuanes L., Flegel T.W., Lightner
D.V., Loh P.C., Snijder E.J. and Tang K. (2005). Family Roniviridae. In: Virus Taxonomy. Eighth
Report of the International Committee on Taxonomy of Viruses. Fauquet C.M., Mayo M.A., Maniloff
J., Desselberger U. and Ball L.A. editors. Elsevier, Academic Press, Amsterdam, 975-979.
Weber H., Beckmann K. and Haas L. (2006). Case report: equine arteritis virus (EAV) as the cause of
abortion in alpacas?. Deutsche tierarztliche Wochenschrift, 113, 162-163.
Westcott D., Lucas M. and Paton D. (1995). Equine arteritis virus: antigenic analysis of strains variation.
In: Proc Congr Eur Soc Vet Virol, 3, 479-483.
Wieringa R., de Vries A.A., Raamsman M.J. and Rottier P.J. (2002). Characterization of two new
structural glycoproteins, GP(3) and GP(4), of equine arteritis virus. J Virol, 76, 10829-10840.
Wieringa R., de Vries A.A. and Rottier P.J. (2003a). Formation of disulfidelinked complexes between the
three minor envelope glycoproteins (GP2b, GP3, and GP4) of equine arteritis virus. J Virol, 77, 6216-
6226.
Wieringa R., De Vries A.A., Post S.M. and Rottier P.J. (2003b). Intra- and intermolecular disulfide bonds
of the GP2b glycoprotein of equine arteritis virus: relevance for virus assembly and infectivity. J Virol,
77, 12996-13004.
Wieringa R., de Vries A.A., van der Meulen J., Godeke G.J., Onderwater J.J., van Tol H., Koerten
H.K., Mommaas A.M., Snijder E.J. and Rottier P.J. (2004). Structural protein requirements in
equine arteritis virus assembly. J Virol, 78, 13019-13027.
Wilkins P.A., Del Piero F., Lopez J. and Cline M. (1995). Recognition of bronchopulmonary dysplasia in
a newborn foal. Equine Vet J, 27, 398.
Wilson J.C., Doll E.R., Mc C.W. and Cheatham J. (1962). Propagation of equine arteritis virus previously
adapted to cell cultures of equine kidney in monolayer cultures of hamster kidney. Cornell Vet, 52, 200-
205.
Wood J.L., Chirnside E.D., Mumford J.A. and Higgins A.J. (1995). First recorded outbreak of equine
viral arteritis in the United Kingdom. Vet Rec, 136, 381-385.

Introduction
46
Wurm T., Chen H., Hodgson T., Britton P., Brooks G. and Hiscox J.A. (2001). Localization to the
nucleolus is a common feature of coronavirus nucleoproteins, and the protein may disrupt host cell
division. J Virol, 75, 9345-9356.
Zhang J., Timoney P.J., Shuck K.M., Seoul G., Go Y.Y., Lu Z., Powell D.G., Meade B.J. and
Balasuriya U.B. (2010). Molecular epidemiology and genetic characterization of equine arteritis virus
isolates associated with the 2006–2007 multi-state disease occurrence in the USA. J Gen Virol, 91,
2286-2301.
Ziebuhr J., Snijder E.J. and Gorbalenya A.E. (2000). Virus-encoded proteinases and proteolytic
processing in the Nidovirales. J Gen Virol, 81, 853-879.


AIMS OF THE THESIS

Aims of the study _
47
Although equine arteritis virus (EAV) is not often included as etiologic agent in the differential diagnosis of
abortion and neonatal mortality, equine viral arteritis (EVA) is an important disease that can result in severe
economic losses for the equine industry. Breeders, racehorse and show horse owners have strong economic
reasons to prevent and control this disease. While it does not kill mature horses, EVA can markedly disturb the
breeding season by causing abortion in numerous mares. In addition, stallions may become persistent carriers of
the virus and transmit it during breeding, with a consequent decrease in their desirability as breeding animals.
EVA has recently increased in prevalence worldwide, possibly due to an increased transportation of infected
horses and infectious semen.
Outbreaks of EVA were first and most frequently described in the USA and, therefore, the main body of EAV
literature and studies on the virus-animal interactions originates from that continent. Virus strains show a wide
variety of virulence characteristics as manifested by the type and severity of clinical signs and lesions upon
experimental inoculation of horses. The appearance of EVA in Europe is more recent. The phylogenetic analysis
of ORF5 of different EAV isolates has revealed that they can be clustered in North American and European
clades. However, it is not clear if such genetic differences have an impact on the virulence and the pathogenetic
features of the European strains compared to the North American ones.
At the occasion of the isolation of an EAV strain (designated 08P178) from an infected foal that died shortly
after birth in Belgium in 2008, the question rose how this European strain would behave with regard to organ
tropism, clinical signs, virological aspects of pathogenesis and development of lesions when inoculated in
experimental horses. These points were particularly interesting since no clinical signs were observed in young or
adult horses on the stud where the Belgian isolate originated from. By performing such experimental infections,
an answer on the different aspects of the course of infection with this European strain was sought.
All the experimental studies in horses performed using North American EAV isolates showed that the virus first
caused an infection of the respiratory tract followed by spread to internal organs via a cell-associated viraemia.
Pathogenetic events of viral infections may be a result of virulence genes expression. It is unknown whether or
not a European strain, such as the Belgian isolate, show a profile of infection with regard to tropism for cell
types and organs that could be similar or different from that previously published for North American strains.
In general, the type of cells that become infected with EAV and that may be responsible for further virus
invasion at the respiratory tract as portal of entry is largely unknown. More detailed information in these initial
phases of EAV pathogenesis are essential and could give some insights not only on which anatomical
structure(s) of the mucosal layer is (are) involved but also on the local immunological processes allowing to
adapt approaches for prevention at the primary site of replication.

Aims of the study
48
Also, as the name “Equine Viral Arteritis “ implies, it has almost automatically been accepted that blood vessels
and possibly endothelial cells can be heavily involved in the infection and the genesis of lesions. It is intriguing
to find out if such generally accepted concept is justified. Also, it could be that the degree of the endothelial cell
involvement and the lesions in internal organs are directly related to the virulence characteristics of the virus
strain. It was thus interesting to know if, and possibly to which extent, endothelial cells in blood vessels of
infected horses serve as target cells for this European EAV strain.
Thus, the general aim of the research carried out for this doctoral thesis was to obtain a better understanding of
the EAV pathogenesis with a European strain starting from virus replication at the portal of entry, over the
viraemia and including replication in the internal organs.
The more specific aims were
(i) to inoculate horses with the Belgian field EAV strain (08P178) and follow the animals, at different time
intervals after inoculation, for virus quantities in different organ tissues and secretions, for humoral immune
response and for clinical signs and gross pathology.
(ii) to define the cell type(s) in the EAV inoculated horses (in vivo) in which the virus replicates at the portal of
entry and in internal organs, in the latter with special reference to endothelial cells.
(iii) to evaluate a polarized explant system of equine nasal and nasopharyngeal mucosae (ex vivo) for
determining and characterizing the cellular population in which EAV replicates at the level of the upper
respiratory mucosal tissues and, thus, to learn about the usefulness of this system for studying viral pathogenesis
at the respiratory portal of entry.


Sabrina Vairo, Annelies Vandekerckhove, Lennert Steukers, Sarah Glorieux, Wim Van den Broeck,
Hans Nauwynck
Clinical and virological outcome of an infection with the Belgian equine arteritis virus strain
08P178
Published in VET MIC 2012; 333-344.

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
49
Abstract
Equine viral arteritis (EVA) is an infectious disease with variable clinical outcome. Outbreaks, causing
important economic losses, are becoming more frequent. Currently, there is a shortage of pathogenesis
studies performed with European strains. In the present study, eight seronegative ponies were experimentally
inoculated with the Belgian strain of equine arteritis virus (EAV) 08P178 (EU-1 clade) and monitored daily
for clinical signs of EVA. Nasopharyngeal swabs, ocular swabs, bronchoalveolar cells and blood were
collected for virological and serological testing. At 3, 7, 14, and 28 days post inoculation (dpi) two ponies
were euthanized After necropsy, specimens were collected for virus titration and immunofluorescence. EVA
clinical signs such as fever and lymphadenomegaly were evident from 3 to 10dpi. Virus was isolated in nasal
secretions from 2 to 9dpi and in bronchoalveolar cells from 3 to 7dpi. A cell-associated viraemia was
detected from 3 to 10dpi. After replication in the respiratory tract and draining lymph nodes, EAV reached
secondary target organs (high virus titers in internal organs sampled at 7dpi). At 14dpi, virus titers dropped
drastically and, at 28dpi, only tonsils were positive. Immunofluorescence revealed both individual and
clustered EAV-infected cells. Antibodies were detected starting from 7dpi. It can be concluded that the
Belgian strain 08P178 is a European mildly virulent subtype. At present, most European EAV strain
infections were thought to run a subclinical course. This study is a proof that mildly virulent European EAV
strains do exist in the field.
1. Introduction
Equine viral arteritis (EVA) is a horse infectious disease caused by equine arteritis virus (EAV). EAV is a
single-stranded, positive-sense RNA virus with a genome of approximately 12.7kb (Snijder and Meulenberg,
1998 and Cavanagh, 1997). Within the order of the Nidovirales, EAV belongs to the family Arteriviridae.
EAV was first isolated from lungs of an aborted fetus following an extensive abortion outbreak in Bucyrus,
Ohio (USA, 1953) (Doll et al., 1957). Since then, several different strains have been identified.
Serological and virological studies have indicated that EAV is widely distributed in equine populations
around the world (Huntington et al., 1990 and Moraillon and Moraillon, 1978). Based on phylogenetic
analysis of EAV-ORF5 sequences, isolates of EAV are clustered into two distinct clades: a North American
and a European group. The latter can be further divided into two subgroups: EU-1 and EU-2 (Zhang et al.,
2007). It has been previously reported that North American and European EAV isolates have 85% nucleotide
identity. This genetic diversity among different field isolates may lead to a clinical outcome with varying
severity (McCollum and Timoney, 2003; Timoney, 2000a and Patton et al., 1999). The vast majority of EAV
strains causes subclinical infections. Nevertheless, in the past, extensive clinical outbreaks caused by certain
EAV strains have been described in various countries (Timoney et al., 2006; Van der Meulen et al., 2001;
Eichhorn et al., 1995; Wood et al., 1995 and Boer et al., 1979). A clinical infection with EAV is
characterized by an influenza-like illness in adult horses, abortion in 10 to 50% of susceptible mares and
interstitial pneumonia in newborn animals (Timoney, 2000a). Mortality is rare, except in foals and in old and
debilitated horses. The direct consequences of EVA outbreaks are financial losses mainly caused by the

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
50
previously mentioned clinical signs in mares and newborn animals. Up to 10-70% of the stallions infected
with EAV become carrier (Holyoak et al., 1993a and Holyoak et al., 1993b), constantly shedding virus into
the semen (Timoney, 2000a). The commercial value of persistently infected stallions decreases as the export
of these animals and their infected semen is prohibited in many countries (Timoney, 2000b). EAV can be
transmitted via the respiratory and venereal route. Aerosol transmission predominates, especially when
horses are gathered at racetracks, sales, shows and other events. Venereal transmission from carrier stallions
is particularly significant on breeding farms (Timoney, 2000b). The virus can be found in nasal secretions of
acutely infected animals for up to 16 days (Del Piero, 2000) and in infected semen for several years.
Although most EAV infections are subclinical, one outbreak in 2000 (Van der Meulen et al., 2001) and one
in 2008 (Gryspeerdt et al., 2009) entailed important economic losses in Belgium. In addition, a serological
survey, performed on samples submitted over the last ten years to the Faculty of Veterinary Medicine of
Ghent University (Belgium), reported an increase from 10 to 30% in seropositivity for EAV in horses. In this
decade, the authors obtained 3 EAV isolates from horses with different clinical signs. The first was isolated
from newborn animals which have died during the EAV outbreak in 2008 (Gryspeerdt et al., 2009). A
second strain was isolated from an aborted fetus during an abortion outbreak on a stud farm in 2000 (Van der
Meulen et al., 2001). The third strain was isolated from a healthy but persistently EAV shedding stallion.
The worldwide spread of EAV and its genetic variability increase the risk of new clinical outbreaks of EVA.
Currently, there is a lack of data on the pathogenic mechanisms of European EAV strains. The aim of this
research was to elucidate the pathogenesis of an infection with the Belgian EAV strain 08P178. Full
knowledge of this pathogenesis (i) will improve epidemiological insights in EVA, (ii) allow the
identification of clinical signs that can be attributed to EAV, (iii) ameliorate approaches to diagnose EVA
and (iv) enable the setting up of improved control strategies (blocking of invasion, modeling for the testing
of vaccines and antivirals).
2. Materials and methods
2.1. Animals
Ten Shetland ponies, 4 to 8 months old, were used in this study. Prior to the start of the experiment, an
acclimatization period of two weeks was respected. During this period, the animals, 6 males (M) and 4
females (F), were tested weekly for their seronegativity to EAV by means of a complement-dependent
seroneutralisation (SN)-test and an immunoperoxidase monolayer assay (IPMA). The ponies were housed in
isolation units and fed daily with a commercial, complete food. Drinking water and hay were supplied ad
libitum. EAV-infected animals were euthanized at different time points post inoculation: M3 and F3 at 3
days post inoculation (dpi); M7 and F7 at 7dpi; M14 and F14 at 14dpi and M28a and M28b at 28dpi. Two
mock inoculated, control animals M28ac and M28bc were euthanized at the end of the experiment.
The experiment was approved by the ethical committee of the veterinary faculty, Ghent university (EC
2009/008).

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
51
2.2. Virus and virus inoculation
2.2.1. Virus
The strain used in this study was isolated from neonatal foals that died after EAV infection in Belgium in
2008. The outbreak started with one foal suffering from acute dyspnea four days after birth. Despite
intensive treatment, this foal died within a few hours. Three weeks later, on the same farm, another foal was
born in healthy condition, although placental edema was found. Ten days later, the foal showed acute
respiratory distress with severe dyspnea and died shortly after (Gryspeerdt et al., 2009). Upon inoculation of
rabbit kidney (RK13) cells with a 20% suspension of lung tissue of these foals, a cytopathic effect (CPE)
was observed. The agent was identified as EAV by means of immunofluorescence staining using a mixture
of three mouse monoclonal antibodies (mAbs): two mAbs against the GP5 protein and one mAb against the
M protein of EAV (Van der Meulen et al., 2001). In addition, the virus was confirmed to belong to the
subgroup EU-1 by partially sequencing (916 nucleotides) the ORF5 (GenBank accession number: JN25761)
(Department of Health Care and Biotechnology, KATHO, Catholic University College of South-West
Flanders, Belgium).
2.2.2. Virus inoculation
Eight animals were inoculated both intranasally and orally with 20 ml of phosphate buffered saline (PBS)
containing 107.6
tissue culture infectious dose 50% endpoint (TCID50) of EAV [08P178 respiratory strain, 4°
passage on RK13 cells] using a fenestrated polypropylene 6.8 French, 400 mm length catheter. A dose of 5
ml of the viral suspension was administered into each nostril and 10 ml into the mouth. The exact virus titer
was controlled by titration of the inoculum. Two control animals were mock inoculated with 20 ml of PBS.
2.3. Scoring of clinical signs
On a daily basis, body temperature, heart and respiratory rates were determined. The animals were also
scored for appetite, behavior, swelling of the lymph nodes, congestion and/or presence of petechiae at the
level of mucosal membranes, respiratory distress, ocular and nasal discharge, conjunctivitis, edema, skin rash
and ataxia. The scoring system used ranged from 0 (no clinical signs or normal) to 3 (severe clinical signs)
(Table 1).

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
52
2.4. Collection of nasopharyngeal and ocular swabs, blood and tissues
Nasopharyngeal swabs, ocular swabs and 30 ml of blood on EDTA (VWR, Bridgeport, CT, USA) were
collected daily during the first 14dpi and then at 16, 18, 20, 22, 24, 26, and 28dpi. Bronchoalveolar cells
were harvested by bronchoalveolar lavage (BAL) as previously described (Clark et al., 1995) at -1, 3, 7, 10,
14, 21, and 28dpi. At euthanasia, forty-four different samples (Table 2) were collected for histopathology,
virus titration and immunofluorescence staining. Abdominal and pleural fluids, when present, were
submitted for virus titration and antibody detection (IPMA). Lungs, liver, spleen and kidneys were collected
in duplicate with one superficial and one deep portion.
2.5. Virus titration
Nasal and ocular swabs, plasma and tissues were processed for virus titration. Briefly, RK13 cells were
inoculated for 1h with serial 10-fold dilutions (100-10
-7 in quadruplicate) of nasal and ocular secretions,
plasma and 20% tissue suspensions, overlaid with medium and observed daily for CPE for 5-7 days.
Presence of EAV-infected cells was confirmed by indirect immunohistochemistry (IHC) staining using
polyclonal antibodies against EAV 08P178 strain (1:160) as primary antibodies and peroxidase-labeled goat
anti-horse immunoglobulins (Molecular Probes, Oregon, USA) (1:500) as secondary antibodies. Viral
antigen-positive cells were detected with a light microscope (Olympus Optical Co., Hamburg, Germany) and
TCID50 was determined with the method of Reed and Muench (Reed and Muench, 1938).
Table I: Scoring system used to evaluate the clinical manifestations of the Belgian EAV strain 08P178.
Symptom Description of clinical signs that get a score …
0 1 2 3 Appetite Normal Diminished Anorexia Cachexia
Behaviour Alert Mild
depression
Moderate
depression
Severe
depression
Swelling lymph nodes None Mild Moderate Severe
Congestion of mucosal
membranes None Mild Moderate Severe
Petechiation of mucosal
membranes None Local Moderate Diffuse
Respiratory distress None Moderate
tachypnea
Severe
tachypnea Dispnea
Nasal discharge None Serous Mucous Purulent
Ocular discharge None Serous Mucous Purulent
Conjunctivitis / lacrimation None Mild Moderate Severe
Periorbital / supraorbital edema None Mild Moderate Severe
Edema of ventral abdomen / legs None Mild Moderate Severe
Edema of mammary glands or
preputium/ scrotum None Mild Moderate Severe
Diarrhoea None Mild Moderate Severe
Urticarial skin rash None Mild Moderate Severe
Ataxia None Slight Mild Moderate
Death None Yes

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
53
2.6. Co-culture of equine bronchoalveolar and peripheral blood mononuclear cells
Cells in the aspirated BAL fluid were pelleted by centrifugation at 400g for 20 min, washed twice and serial
five-fold dilutions of each sample were co-cultured with a monolayer of RK13 cells at 37°C and 5% CO2 for
5 days. Peripheral blood mononuclear cells (PBMC) were isolated from 30 ml of EDTA anticoagulated
blood by density gradient centrifugation on 95% Ficoll-Paque (Sigma-Aldrich, Bornem, Belgium).
Five-fold dilutions of the PBMC were co-cultured with a monolayer of RK13 cells at 37°C and 5% CO2 for
5 days. Afterward, the plates were air-dried and submitted for IHC staining. Briefly, the plates were
incubated with 4% paraformaldehyde for 10min at room temperature, washed twice with PBS and further
incubated with methanol supplemented with 0.1% H2O2. After 1h of incubation with polyclonal antibodies
against EAV strain 08P178 (1:160), the plates were further incubated for 1h with peroxidase-labeled goat
anti-horse immunoglobulin (Molecular Probes) (1:500). Primary and secondary antibodies were diluted in
PBS supplemented with 0.8% Tween and 10% complement-inactivated horse serum. Before each incubation
step, the wells were extensively washed with PBS supplemented with 0.8% Tween. Infected cells were
visualized with a substrate solution of 3-amino-9-ethylcarbazole in 0.05 M acetate buffer (pH 5) with 0.05%
H2O2. Viral antigen-positive cells were detected with a light microscope (Olympus Optical Co., Hamburg,
Germany) and TCID50 was determined with the method of Reed and Muench (Reed and Muench, 1938).
2.7. Immunofluorescence staining
Only tissues with titers equal or higher than 105.0
TCID50/g were investigated by means of
immunofluorescence staining. After fixation with 100% methanol for 20min at -20 °C, 8µm tissue
cryosections were incubated for 1h at 37°C with monoclonal antibodies (17D3), specific for the nucleocapsid
(N) protein of EAV (VMRD, Pullman, USA), diluted 1:100 in PBS containing 10% negative goat serum.
Afterwards, the samples were incubated for 1h at 37°C with goat anti mouse-fluorescein isothiocyanate
(FITC) antibodies (Molecular Probes) (dilution 1:100 in PBS containing 10% negative goat serum). After
fixation with 100% methanol for 20min at -20°C, PBMC cytospins were incubated with biotinylated
polyclonal antibodies against EAV (dilution 1:5 in PBS containing 10% negative horse serum) for 1h at
37°C and, afterwards, with streptavidin-FITC (dilution 1:100 in PBS containing 10% negative horse serum)
for another hour at 37°C. In order to detect the presence and the localization of viral antigen-positive cells,
the samples were analyzed with a Leica DMI 4000 B inverted fluorescence microscope (Leica microsystems,
Bannockburn, USA).
2.8. Quantification of the humoral immune response
2.8.1. Serology
Antibody titers were determined at -14, -7, 0, 7, 14, 21, and 28dpi by means of a complement-dependent SN-
test and IPMA.
The standard OIE SN test was used for the detection of EAV specific neutralizing antibodies. The SN titer
was calculated following the OIE instructions (cut off 1:2).
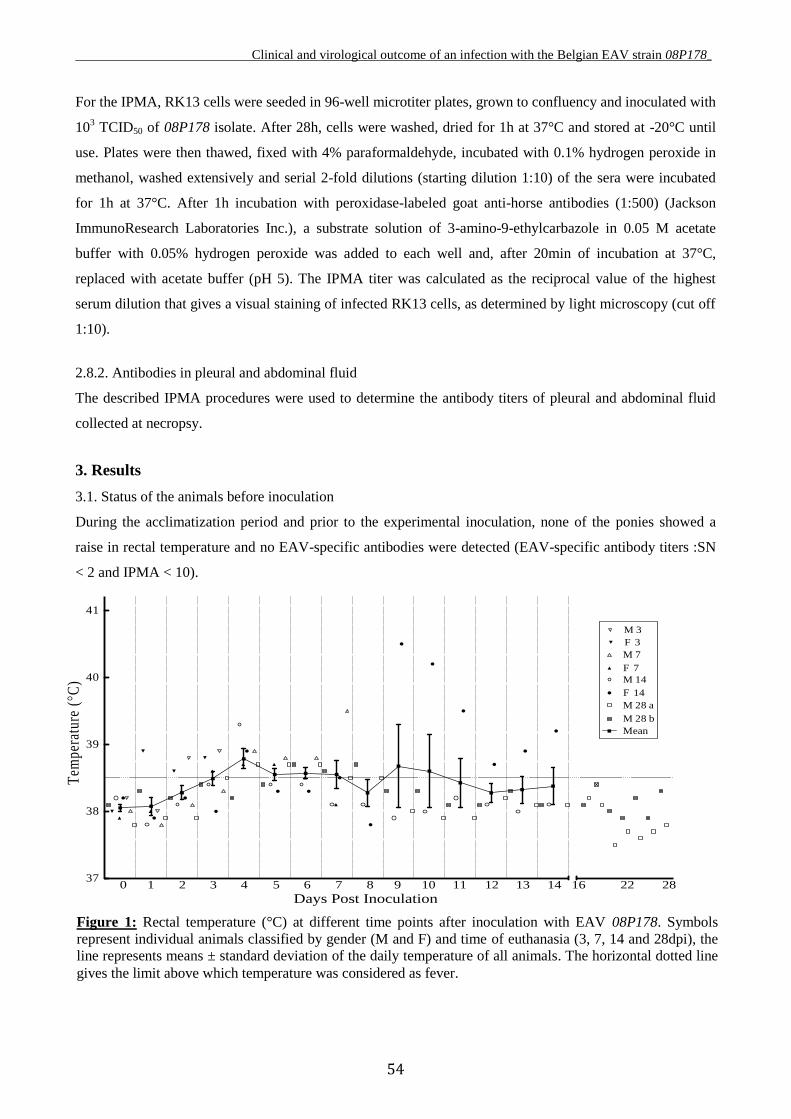
Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
54
For the IPMA, RK13 cells were seeded in 96-well microtiter plates, grown to confluency and inoculated with
103 TCID50 of 08P178 isolate. After 28h, cells were washed, dried for 1h at 37°C and stored at -20°C until
use. Plates were then thawed, fixed with 4% paraformaldehyde, incubated with 0.1% hydrogen peroxide in
methanol, washed extensively and serial 2-fold dilutions (starting dilution 1:10) of the sera were incubated
for 1h at 37°C. After 1h incubation with peroxidase-labeled goat anti-horse antibodies (1:500) (Jackson
ImmunoResearch Laboratories Inc.), a substrate solution of 3-amino-9-ethylcarbazole in 0.05 M acetate
buffer with 0.05% hydrogen peroxide was added to each well and, after 20min of incubation at 37°C,
replaced with acetate buffer (pH 5). The IPMA titer was calculated as the reciprocal value of the highest
serum dilution that gives a visual staining of infected RK13 cells, as determined by light microscopy (cut off
1:10).
2.8.2. Antibodies in pleural and abdominal fluid
The described IPMA procedures were used to determine the antibody titers of pleural and abdominal fluid
collected at necropsy.
3. Results
3.1. Status of the animals before inoculation
During the acclimatization period and prior to the experimental inoculation, none of the ponies showed a
raise in rectal temperature and no EAV-specific antibodies were detected (EAV-specific antibody titers :SN
< 2 and IPMA < 10).
Figure 1: Rectal temperature (°C) at different time points after inoculation with EAV 08P178. Symbols
represent individual animals classified by gender (M and F) and time of euthanasia (3, 7, 14 and 28dpi), the
line represents means ± standard deviation of the daily temperature of all animals. The horizontal dotted line
gives the limit above which temperature was considered as fever.
37
38
39
40
41
Mean
M 3
F 3
M 7
F 7
M 14
F 14
M 28 a
M 28 b
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 16 22 28
Days Post Inoculation
Tem
pera
ture
(°C
)
Figure 1: Rectal temperature (°C) at different time poi nts after inoculation with EAV 08P178. Rectal temperature (°C) was measured daily up to 28 days after inoculation with EAV 08P178.
Symbols represent individual animals classified by gender (M and F) and time of euthanasia (3, 7, 14 and 28 DPI), the line represents mean ± standard deviation of the daily temperature of all animals. The
horizontal dotted line gives the limit above which we considered the temperature as fever.

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
55
3.2. Clinical signs
After inoculation with EAV 08P178, all ponies showed signs of illness characterized by pyrexia from 3 to
14dpi with a maximum temperature of 40.5°C (Figure 1) and submandibular lymphoadenomegaly,
noticeable during the first 14dpi (Figure 2). In all EAV-inoculated animals, heart and respiratory rates
remained normal during the experiment. Ocular and nasal discharges were observed. Five out of eight ponies
(M3, F3, M14, M28a and M28b) showed depression but only from 1 to 3dpi. One pony (M28b) developed a
mild to moderate scrotal edema from 7 to 16dpi. None of the control animals did show any clinical signs
during the whole experiment.
3.3. Post mortem findings
At necropsy, straw-colored fluid was present in the thoracic cavity of 7 out of 8 infected animals, with the
exception of F3. Abdominal yellowish clear effusion was also present in all infected animals, except in F7.
Lymphoadenomegaly, involving principally submandibular, cervical, bronchial and mediastinal lymph
nodes, was evident in ponies euthanized at 3, 7 and 14dpi. Inguinal lymph nodes were normal at any time
point. Tonsils were enlarged and congested at 7 and 14dpi. Lungs showed a prominent lobular pattern,
especially at 7 and 14dpi. Congestion and lymph node enlargement were observed along the course of the
big intestinal vessels at 7dpi. The right adrenal cortex was edematous in F7 but no alterations were noticed in
any other animal at any other time point. Spleens collected from animals killed at 7 and 14dpi were
hemorrhagic and enlarged in size. Liver, kidneys and testicles did not manifest gross lesions at any time
point.
Figure 2: Submandibular lymph node enlargement at different time points after inoculation with EAV
08P178. The lymphoadenomegaly was scored as 0 when it was absent, 1 when it was mild, 2 when it was
moderate and 3 when it was severe (Table 1). Symbols represent individual animals classified by gender (M
and F) and time of euthanasia (3, 7, 14 and 28dpi).
0
1
2
3
M 3
F 3
M 7
F 7
M 14
F 14
M 28 a
M 28 b
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 16 22 28
Days Post Inoculation
Sw
elli
ng
sco
re
Figure 2: Submandibular lymph node enlargement at different time points after inoculation with EAV
08P178. Submandibular lymph node enlargement was examined daily up to 28 days after inoculation with EAV 08P178. The swelling of the lymph nodes was scored as 0 when it was absent, 1 when it was mild, 2
when it was moderate and 3 when it was severe (Table I). Symbols represent individual animals
classified by gender (M and F) and time of euthanasia (3, 7, 14 and 28 DPI).

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
56
In animal F14, euthanized at 14dpi, abscesses were detected in the ethmoid and in the right lung. After
bacteriological examination, Pasteurella pneumotropica was isolated from these abscesses.
Control animals did not show any gross lesions.
3.4. Virus replication
3.4.1. Virus shedding in nasal and ocular secretion
Virus was shed in both ocular and nasal secretions. In ocular secretions, virus was detected during 1 to 3
days from 3 to 7dpi with a maximum titer of 104 TCID50/g (Figure 3). In nasal secretions, virus was isolated
during 1 to 7 days from 2 to 9dpi with a maximum titer of 104.9
TCID50/g (Figure 4).
3.4.2. Quantification of bronchoalveolar infected cells in lungs
Co-cultivation of bronchoalveolar cells with RK13 cells showed the presence of infected cells from 3 to 7dpi
(Figure 5).
Figure 3: Ocular virus shedding after inoculation with EAV 08P178. Ocular swabs were taken daily from
each pony during the first 14dpi and then at 16, 18, 20, 22, 24, 26, and 28dpi. Virus titer was measured by
titration. Symbols represent individual ponies identified by gender (M and F) and time of euthanasia (3, 7,
14 and 28dpi), the horizontal dotted line gives the detection limit for the titration assay.
2
3
4
M 3
F 3
M 7
F 7
M 14
F 14
M 28 a
M 28 b
Figure 3: Ocular virus shedding after inoculation with EAV 08P178.
Ocular swabs were taken daily from each pony during the first 14 DPI and then every other day until euthanasia. Virus titer was measured by titration. Symbols represent individual ponies identified by
gender (M and F) and time of euthanasia (3, 7, 14 and 28 DPI), the horizontal dotted line gives the
detection limit for the titration assay.
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 16 22 28
Days Post Inoculation
Vir
us ti
ter
(log
10 T
CID
50/g
)

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
57
2
3
4
5
6 M 3
F 3
M 7
F 7
M 14
F 14
M 28 a
M 28 b
Figure 4: Nasal virus shedding after inoculation with EAV 08P178.
Nasal swabs were taken daily from each pony during the first 14 DPI and then every other day
until euthanasia. Virus titer was measured by titration. Symbols represent individual ponies identified by gender (M and F) and time of euthanasia (3, 7, 14 and 28 DPI), the horizontal dotted
line gives the detection limit for the titration assay.
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 16 22 28
Days Post Inoculation
Vir
us ti
ter
(log
10 T
CID
50/g
)
Figure 4: Nasal virus shedding after inoculation with EAV 08P178. Nasal swabs were taken daily
from each pony during the first 14dpi and then every other day until euthanasia. Virus titer was
measured by titration. Symbols represent individual ponies identified by gender (M and F) and time
of euthanasia (3, 7, 14 and 28dpi), the horizontal dotted line gives the detection limit for the titration.
Figure 5: Number of infected bronchoalveolar cells at different time points after inoculation with
EAV 08P178. Bronchoalveolar cells collected by bronchoalveolar lavage at -1, 3, 7, 10, 14, 21 and
28dpi were co-cultured on RK13 cells. The titer is expressed as the reciprocal of the highest
bronchoalveolar cell dilution that was able to give cytopathic effects in 50% of the wells. Symbols
represent individual ponies classified by gender (M and F) and time of euthanasia (3, 7, 14 and
28dpi), the horizontal dotted line gives the detection limit of the titration assay.
0
1
2
3
4M 3
F 3
M 7
F 7
M 14
F 14
M 28 a
M 28 b
Figure 5: Number of infected bronchoalveolar cells at different time points after inoculation with EAV
08P178. Bronchoalveolar cells collected by bronchoalveolar lavage at -1, 3, 7, 10, 14, 21 and 28 DPI were co-
cultured on RK13 cells. The titer is expressed as the reciprocal of the highest bronchoalveolar cell
dilution that was able to give cytopathic effects in 50% of the wells. Symbols represent individual ponies classified by gender (M and F) and time of euthanasia (3, 7, 14 and 28 DPI), the horizontal
dotted line gives the detection limit of the titration assay.
Nu
mb
er o
f in
fect
ed b
ron
cho
alv
eola
r
ce
lls
(lo
g1
0/2
*1
05)
-1 3 7 10 14 21 28
Days Post Inoculation

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
58
3.4.3. Viraemia
A strong cell-associated viraemia was detected from 3 to 10dpi (Figure 6). Only animals M14, F14 and M
28b were virus-positive in plasma on 6dpi, from 2 to 4dpi and on 5dpi, respectively (data not shown).
3.4.4. Virus titers in tissues
Contradictory results were detected in the two animals euthanized at 3dpi (Table 2). Nearly all organs of M3
were positive for EAV, whereas F3 showed a certain amount of virus in a limited number of samples, the
main part of which were taken from the upper respiratory tract (distal septum, nasopharynx and tubal-
nasopharyngeal tonsils). In M3, the highest virus titer (106.5
TCID50/g) was found at the level of the tubal-
nasopharyngeal tonsils and in the right intermediate superficial lobe of the lungs, followed by high virus
titers in the nasopharynx, submandibular and bronchial lymph nodes, distal trachea, colon and abdominal
fluid. At 7dpi, the virus was detected in all samples collected from both sacrificed horses. Particularly, high
titers were found in the respiratory tract, intestines and kidneys of both animals. A titer of 106.0
TCID50/g was
also found at the level of the testicles. Furthermore, the superficial regions of lungs, spleen and liver
generally presented higher titers than the corresponding deep sections. At 14dpi, only the tubal-
nasopharyngeal tonsils and the cranial superficial lobe of the right lung of F14 were positive with a very low
titer (101.7
TCID50/g), while in the male M14, the proximal septum, the nasopharynx, the liver and the colon
were positive (highest titer of 103.5
TCID50/g). Animals euthanized at 28dpi showed the presence of virus
only in the tonsils (peak of 105.5
TCID50/g) and the cranial part of the trachea. Virus could not be isolated
from control animals.
0
1
2
3M 3
F 3
M 28 a
M 28 b
M 7
F 7
M 14
F 14
Figure 6: Cell-associated viraemia determined by co-culture of Peripheral Blood Mononuclear Cells
(PBMC) at different time points after inoculation with EAV 08P178. Serial 5-fold dilutions of PBMC were co-cultured on RK13 cells. The titer is expressed as the
reciprocal of the highest PBMC dilution that was able to give cytopathic effects in 50% of the wells.
Symbols represent individual ponies classified by gender (M and F) and time of euthanasia (3, 7, 14
and 28 DPI), the horizontal dotted line gives the detection limit of the titration assay.
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 16 22 28
Days Post Inoculation
Num
ber
of in
fect
ed P
BM
C (
log 1
0/10
6 )
Figure 6: Cell-associated viraemia after inoculation with EAV 08P178. Serial 5-fold dilutions of Peripheral
Blood Mononuclear Cells (PBMC) were co-cultured on RK13 cells. The titer is expressed as the reciprocal
of the highest PBMC dilution that was able to give cytopathic effects in 50% of the wells. Symbols represent
individual ponies classified by gender (M and F) and time of euthanasia (3, 7, 14 and 28dpi), the horizontal
dotted line gives the detection limit of the titration assay.

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
59
Table II: EAV strain 08P178 replication in different organs and tissues collected at 3, 7, 14, 28 days post
inoculation (dpi).
Virus titers (log10 TCID50/gram tissue)
System Organs / tissues 3dpi 7dpi 14dpi 28dpi
M3 F3 M7 F7 M14 F14 M28a M28b
Upper respiratory
tract and
Proximal septum 3.0 2.7 3.8 5.6 2.5 <1.7 <1.7 <1.7 tract and Intermedial septum 4.7 4.7 4.0 3.3 <1.7 <1.7 <1.7 <1.7
associated Distal septum <1.7 6.0 3.8 3.8 <1.7 <1.7 <1.7 <1.7
lymphoid tissues Ethmoid 4.3 4.0 3.8 3.8 <1.7 <1.7 <1.7 <1.7
Nasopharynx 6.0 6.5 3.7 6.0 2.7 <1.7 <1.7 <1.7
Trachea (cranial part) 4.0 <1.7 5.2 4.2 <1.7 <1.7 <1.7 2.5
Tubal-nasopharyngeal tonsils 6.3 5.3 4.0 4.8 <1.7 1.7 <1.7 <1.7
Soft palate tonsils 2.7 <1.7 4.2 4.7 <1.7 <1.7 2.0 2.0
Lingual tonsils 4.5 <1.7 4.5 4.5 <1.7 <1.7 <1.7 5.5
Palatine tonsils 4.7 1.7 4.7 3.7 <1.7 <1.7 2.0 5.5
Retropharyngeal Ln. 4.5 3.0 3.0 2.5 <1.7 <1.7 <1.7 <1.7
Submandibular Ln. 5.7 4.3 2.7 4.8 <1.7 <1.7 <1.7 <1.7
Trachea (distal part) 5.3 <1.7 4.5 5.3 <1.7 <1.7 <1.7 <1.7
Deep respiratory
tract and Left cranial lobe (superf.) 2.3 <1.7 4.5 5.0 <1.7 <1.7 <1.7 <1.7
tract and Right cranial lobe (superf.) 3.6 <1.7 4.6 6.7 <1.7 1.7 <1.7 <1.7
associated Left cranial lobe (deep) 3.3 <1.7 4.5 4.5 <1.7 <1.7 <1.7 <1.7
lymphoid tissues Right cranial lobe (deep) 4.0 <1.7 3.7 6.5 <1.7 <1.7 <1.7 <1.7
Left interm.* lobe (superf.) 4.5 <1.7 4.8 / <1.7 <1.7 <1.7 <1.7
Right interm.* lobe (superf.) 6.2 <1.7 4.7 6.0 <1.7 <1.7 <1.7 <1.7
Left interm.* lobe (deep) 4.7 2.7 3.0 3.0 <1.7 <1.7 <1.7 <1.7
Right interm.* lobe (deep) 3.0 <1.7 3.5 5.2 <1.7 <1.7 <1.7 <1.7
Left distal lobe (superf.) 4.7 <1.7 5.0 4.0 <1.7 <1.7 <1.7 <1.7
Right distal lobe (superf.) 3.0 <1.7 5.5 5.8 <1.7 <1.7 <1.7 <1.7
Left distal lobe (deep) 3.0 <1.7 3.8 3.4 <1.7 <1.7 <1.7 <1.7
Right distal lobe (deep) 4.0 <1.7 3.7 5.4 <1.7 <1.7 <1.7 <1.7
Cervical Ln. <1.7 1.7 4.3 2.8 <1.7 <1.7 <1.7 <1.7
Mediastinal Ln. 4.7 <1.7 3.5 4.0 <1.7 <1.7 <1.7 <1.7
Bronchial Ln. 5.7 2.0 3.8 5.0 <1.7 <1.7 <1.7 <1.7
Abdominal organs Liver (superf.) 3.5 <1.7 3.3 4.4 2.5 <1.7 <1.7 <1.7
Liver (deep) 3.0 <1.7 3.0 3.0 <1.7 <1.7 <1.7 <1.7
Spleen (capsule) 4.3 4.5 3.4 4.0 <1.7 <1.7 <1.7 <1.7
Spleen (pulp) 4.7 4.5 2.8 4.2 <1.7 <1.7 <1.7 <1.7
Adrenal cortex 4.7 3.5 3.8 4.3 <1.7 <1.7 <1.7 <1.7
Kidney (cortex) 4.7 <1.7 3.8 2.8 <1.7 <1.7 <1.7 <1.7
Kidney (medulla) <1.7 <1.7 5.7 5.0 <1.7 <1.7 <1.7 <1.7
Small intestine-->Ileum 1.7 <1.7 5.0 5.0 <1.7 <1.7 <1.7 <1.7
Large intestine-->Caecum 4.7 <1.7 6.0 5.3 <1.7 <1.7 <1.7 <1.7
Large intestine-->Colon 5.0 1.7 5.2 3.5 3.5 <1.7 <1.7 <1.7
Inguinal Ln. 3.0 <1.7 4.0 4.2 <1.7 <1.7 <1.7 <1.7
Body fluids Pleural fluid 3.5 / 3.8 <1 <1 <1 <1 <1
Abdominal fluid 5.6 <1 4.2 / <1 <1 <1 <1
Blood Plasma <1 <1 <1 <1 <1 <1 <1 <1
Genital organs Testicles 4.5 / 6.0 / <1.7 / <1.7 <1.7
*Intermediate lobe included the middle and the accessory lobe.
Ln.= lymph nodes; superf.= superficial.

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
60
3.4.5. Viral antigen-positive cells in tissues
Viral antigens were not detected in PBMC. In general, immunofluorescence staining on tissue cryosections
revealed the presence of individual EAV-infected cells and clusters of EAV-infected cells (Figure 8). The
gross morphology of EAV-positive cells consisted of a big round or oval unstained nucleus surrounded by a
small rim of positive cytoplasm usually with blunt pseudopods (Figure 7). At the level of the nose, the
majority of single EAV-infected cells was found below the basement membrane (BM). In addition, clusters
of EAV-infected cells were noticed in the underlying connective tissue. The tonsils presented a high density
of individual EAV-infected cells, mostly localized in the parafollicular area. Small clusters of EAV-infected
cells were frequently detected. Only one big cluster of EAV-infected cells with an area of 800 µm2 was
observed. In the lymph nodes, single EAV-infected cells were detected, mostly localized in the paracortical
area. Lungs and intestines showed a high number of both individual and clusters of EAV-infected cells. The
pulmonary pattern had a homogeneous distribution of EAV infection around the alveoli, while the intestines
presented a higher concentration of EAV-positive cells and clusters close to lymphoid follicles. At 3 and
7dpi, the testicles presented a high number of EAV-positive clusters and some individual EAV-infected cells
(Figure 8).
Figure 7: Single and clustered EAV 08P178 viral antigen-positive cells in the colon.
Scale bar represents 50 µm.

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
61
3 DPI 7 DPI Nasal mucosa
Tubal naso- pharyngeal tonsils
Lungs
Bronchial lymph nodes
Colon
Testicles
Figure 8: EAV 08P178 viral antigen-positive cells in different organs. Scale
bar represents 100 µm.
3dpi 7dpi

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
62
3.5. Humoral immune response
3.5.1. Serology
At 7dpi, antibodies against EAV appeared with values of 10-20 in the IPMA and 2-8 in the SN-test.
Afterwards, the antibody titers increased progressively to reach a peak at 28dpi (IPMA-titer 640; SN-titer
512) (Figure 9).
3.5.2. Antibodies in pleural and abdominal fluid
In order to differentiate exudate from transudate effusion, the presence of antibodies in pleural and
abdominal fluid was investigated. At 7dpi, IPMA antibodies appeared in both pleural and abdominal liquid.
The IPMA antibody titers increased in time from 2 at 7dpi to 128 at 28dpi.
4. Discussion
In the present study, all animals inoculated with EAV 08P178 became infected and developed clinical signs
such as fever, ocular and nasal discharge, lymphoadenomegaly, with one animal showing scrotal edema.
This clinical picture was much less severe than the one seen in the field. Despite intensive treatments, two
neonatal foals died shortly after developing acute and severe respiratory distress (Gryspeerdt et al., 2009).
This age-dependent difference in clinical outcome of EAV infections has been previously described for other
strains (McCollum and Timoney, 1999 and Jones et al., 1957). The authors strongly believe that EVA is
often not diagnosed in the field. In fact, at present, animals presented with flu-like clinical signs and/or
Figure 9: Curves of whole (IPMA) and neutralizing (SN) antibody titers after inoculation with EAV
08P178. IPMA and SN antibodies against EAV were determined at -14, -7, -1, 7, 14, 21, and 28dpi.
Black symbols indicate IPMA-titers, while white symbols represent SN-titers. The full and the dotted
lines show, respectively, the means of IPMA and SN titers at each time point. Individual ponies were
classified by gender (M and F) and time of euthanasia (3, 7, 14, and 28dpi).

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
63
edema, are mostly treated with routine standard therapies (broad-spectrum antibiotics and antipyretic drugs)
without an appropriate diagnosis. Because of the clinical similarity of EVA to other equine respiratory and
certain non-respiratory diseases, EAV should be included as a possible causative agent of flu-like clinical
signs and/or edema in a differential diagnosis list. As such, in cases of breathing problems and/or edema,
efforts should be made to demonstrate or exclude EAV infection (virus isolation (infectious virus),
immunostaining (viral antigens), RT-PCR (viral genetic material) or serology (seroconversion)). Whenever
an EVA outbreak is suspected, both blood and nasal swabs should be collected. Indeed, in some animals,
EAV can be recovered first in nasal secretions and afterwards in blood, but in other cases, blood becomes
positive for EAV before nasal secretions. In addition, co-culture of bronchoalveolar cells on RK13 cells
showed to be an appropriate method for early detection of EAV infections. Finally, infection with EAV may
also be demonstrated by sero-conversion using an IPMA or SN test.
Viral shedding in nasal and ocular secretions and presence of EAV in BAL fluids at early time points (1, 3
and 3dpi, respectively) indicates that the primary local replication occurs at the level of the upper respiratory
tract (URT) and the lungs. The most important primary replication site of EAV is the nasopharynx region as
the highest virus titers of tissues, collected at 3dpi, were detected in the nasopharynx (106.5
TCID50/g) and
tubal-nasopharyngeal tonsil (106.3
TCID50/g). Noteworthy, the URT continued to present high virus titers at
7dpi. The high virus titers in URT secretions provide an efficient way for horizontal transmission of EAV.
Immunofluorescence staining on nasal tissues showed both the presence of individual EAV-infected cells in
between epithelial cells and below the BM and clusters of EAV-infected cells in the underlying connective
tissue. Further studies will be performed to identify the target cells and to elucidate how EAV 08P178
invades the host. A possible explanation, although speculative at this moment, could be that, similar to
equine herpesvirus 1 (EHV-1) (Vandekerckhove et al., 2010), EAV uses individual cells as Trojan horses to
cross the BM. Identification of the target cells involved in viral passage through the BM of the URT is a
crucial step in understanding EAV pathogenesis. Afterwards, the virus invades the underlying lamina
propria where it can reach blood vessels to rapidly disseminate throughout the body. The positive results
obtained from the PBMC co-culture and the negative results of the plasma titration suggest that viral
spreading in the blood occurs mainly via a cell-associated viraemia since extracellular virus was a rare
finding. Previous studies demonstrated the presence of EAV in both buffy coat and plasma during viraemia,
with higher concentrations of virus in the buffy coat (MacLachlan et al., 1996; Cole et al., 1986 and Bryans
et al., 1957). The basis for the difference between our and other studies is unclear but may be strain-
dependent. Viraemia enables EAV to reach secondary target organs where replication can cause clinical
signs such as edema and abortion. In our study, all tissues collected at 7dpi from both animals were highly
positive. At 14dpi, the titers dropped substantially (≤103.5
TCID50/g) in nearly all tissues collected. The
mechanisms responsible for the potential clearance of viral-infected cells are unknown, but it was earlier
hypothesized that both humoral and cell-mediated immunity might play a role in the ultimate clearance of
the infection (Castillo-Olivares et al., 2003 and McCollum, 1976). The drop in virus titers after 7dpi could be
due to the appearance of neutralizing antibodies starting at the same time point. Virus-neutralizing antibodies

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
64
in equids are believed to play an important role in immunity against EAV. It has been reported that SN-
antibodies persist in the serum of recovered animals long after infection conferring a long-term, possibly
lifelong immunity against EAV (Balasuriya et al., 1995; McCollum, 1976 and McCollum, 1969). In previous
studies using North American strains, it was reported that the virus could be isolated from renal tissues long
after it had disappeared from other tissues of the body (McCollum et al., 1971). In our study, EAV was not
isolated from the kidneys after 7dpi. This finding is also suggestive for differences between European and
North American strains. The presence of EAV in renal tissues may lead to viral shedding in the urine causing
lateral spread of the virus to other susceptible horses. Starting from 7dpi, antibodies were also detected in
pleural and abdominal fluids, indicating that the fluids are exudates. This exudate most probably originates
from extravasation of plasma due to an increased permeability of the capillaries. EAV strains with different
virulence differ in both their cytopathogenicity and their ability to induce pro-inflammatory cytokines in
endothelial cells in vitro (McCollum and Timoney, 2003). However, in the present study, endothelial cells
did not appear to be the main target of EAV.
Finally, although virus titers dropped substantially in nearly all tissues starting from 14dpi, high virus titers
(105.0
TCID50/g) of EAV were found in the tonsillar tissues at 28dpi. Longer time intervals should be
enclosed to examine if EAV really persists at this site. The latter hypothesis, if confirmed, could have
important epizootiological significance, as not only stallions may be carriers of EAV but also other horses
including mares. Therefore, if this supposition will be proven, also mares could represent a long-term
reservoir of EAV and, in case of recrudescence, shed the virus through nasal secretions. However, since the
study only lasted 28dpi, no firm conclusions can be drawn without further investigations.
The virological and clinical outcome of the EAV strain 08P178 resembles the outcome of an experimental
infection with the American strain KY-84 (McCollum and Timoney, 1984). Therefore, it can be concluded
that the Belgian strain 08P178 is a European mildly virulent subtype. At present, most EAV infections were
thought to run a subclinical course. This study is a proof that mildly virulent European EAV strains do really
exist in the field. In addition, the present study gives new information on the primary replication site of a
European subgroup one EAV strain.
5. Acknowledgements
We thank M. Bauwens, C. Boone, N. Dennequin, G. Opsomer, C. Vanmaercke, L. De Bels and J. De Craene
for the technical support. We acknowledge S. Gleich, A. Gryspeerdt, G. Van de Walle, D. De Clercq, S.
Costers, H. Favoreel, K. van der Meulen and L. Lefère for their scientific contribution, helpful suggestions
and fruitful discussions. Permission to use the facilities of the Laboratory of Phytopathology, Faculty of
Bioscience Engineering, Ghent University and, in particular, Dr. D. De Vleesschauwer is gratefully
acknowledged. S. Vairo is supported by a doctoral grant from the special research fund of Ghent University
(V72-0209).

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
65
6. References
Balasuriya U.B. (2010). Molecular epidemiology and genetic characterization of equine arteritis virus
isolates associated with the 2006-2007 multi-state disease occurrence in the USA. J Gen Virol, 91, 2286-
2301.
Balasuriya U.B., MacLachlan N.J. and de Vries A.A. (1995). Identification of a neutralization site in the
major envelope glycoprotein (GL) of equine arteritis virus. Virology, 51, 8-527.
Boer G.F., Osterhaus A.D.M.E., Oirschot J.T. and Wemmenhove R. (1979). Prevalence of antibodies to
equine viruses in the Netherlands. Vet Quart, 1, 65-74.
Bryans J.T., Doll E.R., Crowe M.E. and McCollum W.H. (1957). The blood picture and thermal reaction
in experimental viral arteritis of horse. Cornell Vet, 47, 42-52.
Castillo-Olivares J., Tearle J.P., Montesso F., Westcott D., Kydd J.H., Davis-Poynter N.J. and
Hannant D. (2003). Detection of equine arteritis virus EAV-specific cytotoxic CD8+ T lymphocyte
precursors from EAV-infected ponies. J Gen Virol, 84, 2745-2753.
Cavanagh D. (1997). Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch Virol,
142, 629-633.
Clark C.K., Lester G.D., Vetro T. and Rice B. (1995). Bronchoalveolar lavage in horses: effect of exercise
and repeated sampling on cytology. Aust Vet J, 72, 249-252.
Cole J.R., Hall R.F. and Gosser H.S. (1986). Transmissibility and abortogenic effect of equine viral
arteritis in mares. J Am Vet Med Assoc, 89, 769-771.
Del Piero F. (2000). Equine Viral Arteritis. Vet Pathol, 37, 287-296.
Doll E.R., Knappenberger R.E. and Bryans J.T. (1957). An outbreak of abortion caused by equine
arteritis virus. Cornell Vet, 47, 69-75.
Echeverría M.G., Díaz S., Metz G.E., Serena M.S., Panei C.J. and Nosetto E. (2010). Evaluation of
neutralization patterns of the five unique Argentine equine arteritis virus field strains reported. Rev argent
Microbial, 42, 11-17.
Eichhorn W., Heilmann M. and Kaaden O.R. (1995). Equine viral arteritis with abortion: serological and
virological evidence in Germany. J Vet Med, 42, 574-576.
Gryspeerdt A., Chiers K., Govaere J., Vercauteren G., Ducatelle R., Van de Walle G.R. and
Nauwynck H.J. (2009). Neonatal foal death due to infection with equine arteritis virus in Belgium. Vlaams
Diergeneeskd Tijdschr, 78, 189-193.
Holyoak G.R., Giles R.C., Little T.V., McCollum W.H. and Timoney P.J. (1993a). Pathological changes
associated with equine arteritis virus infection of the reproductive tract in prepubertal and peripubertal colts.
J Comp Pathol, 3, 281-293.
Holyoak G.R., Little T.V., McCollum W.H. and Timoney P.J. (1993b). Relationship between onset of
puberty and establishment of persistent infection with equine arteritis virus in the experimentally infected
colt. J Comp Pathol, 1, 29-46.

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178_
66
Huntington P.J., Forman A.J. and Ellis P.M. (1990). The occurrence of equine arteritis virus in Australia.
Aust Vet J, 67, 432-435.
Jones T.C., Doll E.R. and Bryans J.T. (1957). The lesion of equine viral arteritis. Cornell Veterinarian, 47,
52-68.
MacLachlan N.J., Balasuriya U.B., Rossitto P.V., Hullinger P.A., Patton J.F. and Wilson W.D. (1996).
Fatal experimental equine arteritis virus infection of a pregnant mare: immunohistochemical staining of viral
antigens. J Ve. Diag. Inves, 8, 367-374.
McCollum W.H. (1969). Development of modified virus strain and vaccine for equine viral arteritis. J Am
Vet Med Assoc, 155, 322-348.
McCollum W.H. (1976). Studies of passive immunity in foals to equine viral arteritis. Vet Microbiol, 1, 45-
54.
McCollum W.H. and Timoney P.J. (1984). The pathogenic qualities of the 1984 strain of equine arteritis.
In: Proceeding of the Grayson foundation int conf thoroughbred breeders organization on equine viral
arteritis, 34-37.
McCollum W.H. and Timoney P.J. (1999). Experimentally observation on the virulence of isolates of
equine arteritis virus. Proceedings of the eight int conf equine infect dis, 8, 558-559.
McCollum W.H. and Timoney P.J. (2003). Differentiation of strains of equine arteritis virus of differing
virulence to horses by growth in equine endothelial cells. Am J Vet Res, 64, 779-784.
McCollum W.H., Prickett M.E. and Bryans J.T. (1971). Temporal distribution of equine arteritis virus in
respiratory mucosa, tissues and body fluid of horses infected by inhalation. Res Vet Sci, 2, 459-464.
Moraillon A. and Moraillon R. (1978). Results of an epidemiological investigation on viral arteritis in
France and some other European and African countries. Ann Rech Vet, 9, 43-54.
Patton J.F., Balasuriya U.B., Hedges J.F., Schweidler T.M., Hullinger P.J. and MacLachlan N.J.
(1999). Phylogenetic characterization of a highly attenuated strain of equine arteritis virus from the semen of
a persistently infected Standardbred stallion. Arch Virol, 144, 817-827.
Reed L.J. and Muench H. (1938). A simple method of estimating fifty percent endpoints. Am J Hygiene,
27, 493-497.
Snijder E.J. and Meulenberg J.J. (1998). The molecular biology of arteriviruses. J Gen Virol, 79, 961-979.
Timoney P.J. (2000a). Factors influencing the international spread of equine diseases. Vet Clin North Am
Equine Pract, 16, 537-551.
Timoney P.J. (2000b). The increasing significance of international trade in equids and its influence on the
spread of infectious diseases. Ann N Y Acad Sci, 916, 55-60.
Timoney P.J., Creekmore L., Meade B., Fly D., Rogers E. and King B. (2006). Multi-state occurrence of
EVA. Report of the Committee on Infectious Diseases of Horses: addressing equine viral arteritis in the
United States. In: Proceedings of the one-hundred tenth annual meeting of the United States animal health
association. Minneapolis, Minnesota: Pat Campbell & Associates and Spectrum Press. 354-362.

Clinical and virological outcome of an infection with the Belgian EAV strain 08P178 _
67
Van der Meulen K., Caij A., Nauwynck H. and Pensaert M. (2001). An outbreak of equine viral arteritis
abortion in Belgium. Vlaams Diergeneeskd Tijdschr, 70, 221-222.
Vandekerckhove A.P., Glorieux S., Gryspeerdt A., Steukers L., Duchateau L., Osterrieder N., Van de
Walle G.R. and Nauwynck H. (2010). Replication kinetics of neurovirulent versus non-neurovirulent
equine herpesvirus type 1 strains in equine nasal mucosal explants. J Gen Vir, 91, 2019-2028.
Wood J.L., Chirnside E.D., Mumford J.A. and Higgins A.J. (1995). First recorded outbreak of equine
viral arteritis in the United Kingdom. Vet Rec, 136, 381-385.
Zhang J., Miszezak F., Pronost S., Fortier C., Balasuriya U.B., Zientara S., Fortier G. and Timoney
P.J. (2007). Genetic variation and phylogenetic analysis of 22 French isolates of equine arteritis virus. Arch
Virol, 152, 1977-1994.

Sabrina Vairo, Herman Favoreel, Alessandra Scagliarini, Hans Nauwynck
Identification of target cells of a European equine arteritis virus strain in experimentally
infected ponies
Submitted to Veterinary Microbiology

esn experimentally infected ponitrain iIdentification of target cells of a European equine arteritis virus s
68
Abstract
Currently, little is known on the cellular pathogenesis of equine arteritis virus (EAV). The purpose of the
present study was to identify the target cells in ponies experimentally inoculated with EAV 08P178 (EU,
clade-1). Organs collected at 3 and 7 days post inoculation (dpi) with virus titers >105.0
TCID50/g were
processed with double immunofluorescence stainings for the detection of the EAV N protein and at the same
time for the identification of the cell (specific markers for myeloid cells (CD172a+), T-lymphocytes (CD3
+),
B-lymphocytes (IgM+) or endothelial cells (von Willebrand factor
+)). At 3 and 7dpi, 31-58% and 47%-63%
of the EAV-positive cells, respectively, were mononuclear leukocytes. In all these samples, with exception
of the lungs, the majority of EAV-positive cells were CD172a+ followed by CD3
+; the IgM
+ cells represented
a small percentage. EAV-positive endothelial cells were not found in 3.200 large blood vessels (≥ 3
endothelial cells). In terminal capillaries (1-2 endothelial cells), however, 15% to 51% of the endothelial
cells were EAV-positive. In conclusion, the present study demonstrates that (i) EAV 08P178 has a main
tropism for CD172a+ and CD3
+ mononuclear leukocytes; (ii) EAV 08P178 does not infect endothelial cells
of large blood vessels but does infect a considerable number of endothelial cells in terminal capillaries (iii)
EAV 08P178 infection in these capillaries is most probably the cause of an increased vascular permeability
leading to leakage of fluid (edema-serous exudate) but not to severe vasculitis and hemorrhages.
1. Introduction
Equine viral arteritis (EVA) is an infectious disease first etiologically defined in 1953, when equine arteritis
virus (EAV) was isolated during an abortion outbreak in Ohio (Doll et al., 1957). Since then, serological
surveys indicate that the virus is widely distributed in equine populations around the world (Holyoak et al.,
2008). Different EVA outbreaks have been reported in several European countries such as Switzerland
(Bürki and Gerber, 1966), Germany (Golnik et al., 1986 and Eichhorn et al., 1995), Spain (Monreal et al.,
1995), UK (Wood et al., 1995), Denmark (Larsen et al., 2001), Hungary (Szeredi et al., 2005) and France
(Holyoak et al., 2008). In Belgium, a first outbreak of EVA was described in 2000 (Van der Meulen et al.,
2001) and another followed in 2008, when two neonatal foals died shortly after birth (Gryspeerdt et al.,
2009).
Although only one neutralization serotype of EAV has been identified so far (McCollum, 1970 and Golnik et
al., 1986), a considerable genetic variation among EAV field strains was demonstrated by comparative
sequence analysis of ORFs 2 to 7 (Stadejek et al., 1999 and Hornyak et al., 2005). Based on ORF 5
phylogenetic analysis, EAV isolates are clustered into two distinct clades: a North American and a European
cluster (Echeverría et al., 2010). The latter can be further divided into two subgroups: EU-1 and EU-2
(Zhang et al., 2010). Distinct EAV isolates vary markedly in their capacity to induce clinical signs. It is
generally believed that the North American strains are more virulent than the European isolates (Balasuriya
and MacLachlan, 2004).
Since outbreaks of moderate to clinically severe EVA have been recorded primarily in the USA, although
often traced to the use of imported frozen semen from stallions standing in European stud, the scientific and

Identification of target cells of a European equine arteritis virus strain in experimentally infected ponies
69
economic interest on EAV has mainly been focused on North American strains with the exception of a prior
challenge study with an Italian isolate (Autorino et al., 1994). After natural and experimental infection with
such North American strains, viral antigens have been demonstrated, by means of immunohistochemistry
staining, in the cytoplasm of cells resembling macrophages, endothelial cells, myometrial and cardiac
myocytes, chorionic mesenchymal stromal cells and epithelial cells (Este and Cheville, 1970, Crawford and
Henson, 1972, and Del Piero, 2006). However, up till now no studies have been performed to identify EAV
infected target cells in equine tissues by the use of cell markers.
Recently, we studied the outcome of an experimental infection with EAV European strain 08P178 (EU-1
clade) isolated from a Belgian field outbreak of 2008, in eight seronegative Shetland ponies (Vairo et al.,
2012), which were followed until 28 days post inoculation (dpi). After replication in the respiratory tract and
associated lymphoid tissues, EAV caused a strong cell-associated viraemia from 3 to 10dpi and reached
secondary target organs, such as large intestines, at 3dpi. The infection resolved spontaneously at later time
points (14 and 28dpi). Mild EVA symptoms, such as fever, lymphadenomegaly, and nasal and ocular
discharge were reproduced. In addition, one animal manifested a transitory mild scrotal edema. Signs of
severe vascular damage such as hemorrhages, petechiae and infarcts were not seen (Vairo et al., 2012).
Histopathologically, the most prominent histological alteration was the intraepithelial, subepithelial,
perivascular lymphoplasmocytic infiltration, ranging from mild-multifocal to extensive-diffuse. In addition,
congestion and edema were consistently present in the majority of the organs (article submitted). Therefore,
the aim of the present study was to identify the target cells of this European strain of EAV in an attempt to
obtain better insights in the pathogenesis and in the mechanisms of occurrence of the clinical signs and
lesions. For this purpose, target organs collected from these ponies were examined using cell markers.
Special attention was given to mononuclear cells and endothelial cells in the primary and secondary target
organs.
2. Materials and methods
2.1. Animals
This study was carried out on samples collected during our previous experimental in vivo study (Vairo et al.,
2012). Briefly, 8 animals were oronasally inoculated with 20 ml of phosphate buffered saline (PBS)
containing 107.6
tissue culture infectious dose 50% endpoint (TCID50) of EAV 08P178 (isolated in Belgium
(Gryspeerdt et al., 2009), 4th passage on RK13 cells) [GenBank: JN25761]. Two other animals were mock
inoculated with 20 ml of PBS. EAV-infected animals were euthanized at different time points post
inoculation: M3 and F3 at 3dpi; M7 and F7 at 7dpi; M14 and F14 at 14dpi and M28a and M28b at 28dpi.
The two control animals were euthanized at the end of the experiment (28dpi of the others). This experiment
was approved by the ethical committee of the faculty of veterinary medicine, Ghent (EC 2009/008).
2.2. Sampling
Immediately after euthanasia, numerous tissue samples were collected from each animal for virus titration,

esn experimentally infected ponitrain iIdentification of target cells of a European equine arteritis virus s
70
histopathology and IF investigations (Vairo et al., 2012). In the present study, only the tissues collected at 3
and 7dpi, which contained virus titers ≥105.0
TCID50/g, were examined for identification of EAV-positive
cells. Specifically, the cellular tropism was studied in the following tissues (i) the nasal septum representing
the upper respiratory tract (URT); (ii) nasopharynx; (iii) tubal-nasopharyngeal tonsils representing lymphoid
tissues associated with the URT; (iv) lungs; (v) the bronchial lymph nodes as representative of the lymphoid
tissues associated with the deep respiratory tract (DRT); (vi) large intestines (caecum and colon).
2.3. Immunofluorescence staining
To quantify and characterize individual EAV-positive cells, a double immunofluorescence staining was
performed. Thirty 8µm-thick cryosections of each tissue, mentioned in section 2.2., were cut and fixed in
methanol at -20°C for 20min. For each tissue, ten cryosections were stained for each cell surface marker
separately. In the first step, optimal dilutions of mAb DH59B (VMRD), UC F6G-3 (California University,
Davis, USA) or 1.9/3.2 (VMRD) were used as markers for CD172a+ cells (myeloid lineage), CD3
+ cells (pan
T lymphocytes) and IgM+ cells (B lymphocytes), respectively. Afterwards, slides were incubated with Texas
Red®-labeled goat anti-mouse IgG antibodies (Molecular Probes). In the second step, EAV nucleocapsid (N)
protein was visualized with mAb 17D3 (VMRD), directly labeled with Zenon® Alexa Fluor
® 488 Mouse
IgG1 Labeling Kit (Molecular Probes), according to the manufacturer’s instructions. As negative control,
8µm-thick cryosections of nasopharynx, lungs and colon of mock-inoculated animals were stained following
the aforementioned protocol. In addition, ten extra sections of EAV inoculated animals were incubated with
CD172a+ as primary antibody and Texas Red
®-labeled goat anti-mouse IgG antibodies as secondary
antibody. In a second step, the slides were further incubated with an irrelevant IgG1 directly labeled with
Zenon® Alexa Fluor
® 488 Mouse IgG1 Labeling Kit. All antibodies were diluted in PBS and incubated for
1h at 37°C. The nuclei were counterstained with Hoechst 33342 (Molecular Probes) for 10min. The number
of viral-antigen positive cells was determined in an area of 8mm2 for each tissue investigated. Further, at
each time point, the percentage of double positive cells (EAV and surface marker+) was calculated on the
total number of EAV-positive cells counted in the 10 sections stained for that specific cell marker.
To identify EAV-positive endothelial cells, ten 8µm-thick cryosections of each of the tissues mentioned in
section 2.2. plus of animals sacrificed at 3 and 7dpi, were fixed in methanol at -20°C for 20min, incubated
for 1h at 37°C with the endothelial cell marker sheep anti human von Willebrand factor (vWF) directly
labeled with FITC (AbDserotec, 1:100) and, afterwards, further incubated with mAb 17D3 (VMRD) directly
labeled with Zenon® Alexa Fluor
® 594 Mouse IgG1 Labeling Kit (Molecular Probes), according to the
manufacturer’s instructions. In each of the tissues mentioned above, 100 vessel profiles in which ≥ 3
endothelial cells were considered large vessels (arteries, veins, arterioles, venules and large capillaries) and
analyzed at 20x with a confocal microscope (Leica TCS SP2 Laser scanning spectral confocal system, Leica
microsystems GmbH, Wetzlar, Germany). In addition, vessels with 1 or maximum 2 endothelial cells were
considered as terminal capillaries and investigated at 63x using the confocal microscope.

Identification of target cells of a European equine arteritis virus strain in experimentally infected ponies
71
To confirm the capacity of vWF marker to bind endothelial cells of equine arteries, veins and capillaries,
samples of aorta, vena cava and lungs were collected from a horse in the slaughterhouse and stained with
vWF marker directly labeled with FITC
3. Results
3.1. Quantification and identification of EAV-positive cells in tissues
Samples from mock-inoculated ponies were negative for EAV at any time point. Samples of aorta, vena cava
and lungs, which were collected from the slaughterhouse horse to test the activity of the endothelial marker
(vWF) were consistently positive. Table I summarizes the number and the identification of EAV-infected
cells.
Three days post inoculation - In nasal septum, nasopharynx, URT associated lymphoid tissues, lungs, DRT
associated lymphoid tissues, and large intestines 60, 170, 102, 182, 130 and 231 EAV-positive cells,
respectively, were found in 8mm2 of tissue; 19%, 24%, 27%, 20%, 25% and 36%, respectively, were
CD172a+, and 10%, 20%, 10%, 25%, 24% and 22%, respectively, were CD3
+.
Seven days post inoculation - In nasal septum, nasopharynx, URT associated lymphoid tissues, lungs, DRT
associated lymphoid tissues, and large intestines 122, 146, 171, 149, 272 and 470 EAV-positive cells,
respectively, were found in 8mm2 of tissue; 35%, 28%, 29%, 20%, 26% and 32%, respectively, were
CD172a+, and 22%, 26%, 19%, 25%, 17% and 25%, respectively, were CD3
+.
In all specimens investigated, only a small percentage of EAV-positive cells were IgM+ B-lymphocytes.
Endothelial cells were not infected in the 3.200 large vessels (arteries, veins, arterioles, venules and large
capillaries) examined in all organs at 3 and 7dpi (Figure 1). In contrast, in the nasal septum, nasopharynx,
URT associated lymphoid tissues, lungs, DRT associated lymphoid tissues, and large intestines 15%, 33%,
35%, 34%, 40% and 30% of EAV-infected cells, respectively, were identified as endothelial cells of terminal
capillaries at 3dpi and 18%, 28%, 38%, 51%, 45% and 35% at 7dpi (Figure 2).

esn experimentally infected ponitrain iIdentification of target cells of a European equine arteritis virus s
72
Figure 1: Representative photographs of nasopharyngeal mucosa collected at 3dpi, showing
EAV-positive cells (red) in the perivascular area (vessel = green).

Identification of target cells of a European equine arteritis virus strain in experimentally infected ponies
73

esn experimentally infected ponitrain iIdentification of target cells of a European equine arteritis virus s
74
Figure 2: Representative photographs of lungs (a and a') collected at 3dpi, large intestines collected at 3dpi
(b) and at 7dpi (c), showing EAV-infected cells (red), endothelial cells (green) and nuclei (blue). EAV-
infected capillaries (co- localization = yellow) are indicated with arrow heads in the overlaid pictures.

Identification of target cells of a European equine arteritis virus strain in experimentally infected ponies
75
4. Discussion
Viruses which use the respiratory mucosal surfaces as primary replication site and then proceed to cause a
generalized infection via blood, have to overcome several barriers. An important prerequisite for successful
host invasion is to cross the epithelial cell layer and the underlying basement membrane (BM). Immune cells
frequently traverse the BM barrier and make projections in between the epithelial cells reaching the lumen as
part of normal physiological processes. Although the primary function of such immune cells is to sample
pathogens to initiate an immune response, several pathogens can infect these cells and abuse them as Trojan
horses to cross the epithelial cell layer and BM barrier and to disseminate towards internal organs in the host
(Steukers et al., 2012). We have previously hypothesized (Vairo et al., 2012) that also EAV is hijacking
immune cells to transverse the BM prior to reach the blood stream. Consistent with this hypothesis, 52% of
EAV-positive cells were mononuclear leukocytes in the nasopharyngeal tissues collected at 3dpi, of which
24% were CD172a+ myeloid cells, 20% were CD3
+ T lymphocytes and 8% were IgM
+ B lymphocytes.
CD172a is expressed on monocytes, macrophages, dendritic cells (DCs) and granulocytes. In particular,
DCs, unlike differentiated macrophages, are migratory cells (Lipscomb and Masten, 2002) and are exploited
by several pathogens, such as human (Ayehunie et al., 1994) and simian (Hu et al., 1998) immunodeficiency
viruses, to facilitate their spread into the host. Hence, it is possible that EAV is using DCs as Trojan horses
to cross the epithelial cell layer and BM and invade into its host.
It is generally believed that the tropism of EAV is mainly directed to cells of the monocytic lineage (Holyoak
et al., 2008). However, the present study demonstrates that EAV also has a strong tropism for T
lymphocytes. These findings agree with earlier results obtained with URT mucosal explants infected with
EAV 08P178 (Vairo et al., 2013). Go et al. (2010) also described a tropism for T lymphocytes isolated from
PBMC of infected animals. In general, T lymphocytes play an important role in the clearance of viruses
during infections (Keynan et al., 2008). Therefore, infection of T lymphocytes might functionally hinder
those cells, which could result in a longer lasting persistence of EAV in its host during infection.
An EAV 08P178 infection is characterized by a strong cell-associated viraemia, with peripheral blood
mononuclear cells (PBMCs) carrying the virus (Vairo et al., 2012), and an elevated percentage of infected
mononuclear leukocytes in different primary and secondary replication sites. An important step in the
pathogenesis of generalized infections is the extravasation of infected PBMCs from blood towards the
secondary organs. In the present pathogenesis study, only endothelial cells of terminal capillaries were
consistently involved in the infection process with EAV, while larger vessels were not. It is known that small
vessels represent the site for increased paracellular permeability (Michel and Curry, 1999) and for leukocyte
extravasation during inflammation (Luscinskas et al., 2002). Currently, nothing is known on the
mechanism(s) by which EAV crosses the endothelial barrier to reach secondary organs. Considering the low
number of free infectious virus (Vairo et al., 2012) and the considerable number of infected endothelial cells,
it is hypothesized that the numerous infected blood leukocytes adhere to the endothelial cells of the small
capillaries and cause cell to cell transfer of virus to infect endothelial cells. In such case, endothelial cells are
not to be considered as true target cells but rather as cells, which are collaterally infected. In fact, the low

esn experimentally infected ponitrain iIdentification of target cells of a European equine arteritis virus s
76
viral titers detected in the plasma of the infected ponies may suggest that EAV replication in endothelial cells
is not lytic with virus release into the blood but, most probably, a cell to cell infection. However, various
EAV strains have been previously shown to produce a fully, productive, lytic cycle of replication in the
diploid equine endothelial cell line (Zhang, 2005).
The EAV infection of endothelial cells of capillaries together with the migration of EAV infected leukocytes
in between endothelial cells and through the underlying basement barrier is most probably the origin of the
increased permeability of vessels during an EAV infection (edema, serous exudate).
The interaction of EAV with endothelial cells appears to result in an alteration of the vessel homeostasis as
shown by the congestion and edema which were consistent findings in the infected ponies. However,
hemorrhages, thrombi or infarcts were not found in the organs collected at necropsy (article submitted).
Therefore, EAV 08P178 seems to provoke only minor alterations in the endothelial cell layer and in the
capillaries. In general, small changes of the endothelium layer will cause an increased vascular permeability
with passage of water, salts and some small proteins, such as albumin (Dejana et al., 2009). Subsequently,
pro-inflammatory cytokines can cause swelling and a partially retraction of endothelial cells which no longer
form a completely intact internal lining. Thereafter, intercellular gaps are formed and larger soluble
molecules, such as fibrinogen, leak into the extravascular compartment (Dejana et al., 2009). As a
consequence, much more fluid leaves the vessels than is returned, resulting in the net escape of protein-rich
(<50 g/l) fluid (exudate). When the integrity of the endothelial monolayer is severely affected, intracellular
junctions are disrupted, and endothelial cells detach from the vessel wall, creating areas of vascular damage
and hemorrhages (Dejana et al., 2009).
In conclusion, the present study demonstrates that (i) EAV 08P178 shows a main tropism for CD172a+ and
CD3+ mononuclear leukocytes; (ii) EAV 08P178 infection does not involve endothelial cells in large vessels
but occurs in a considerable number of endothelial cells in terminal capillaries probably via adherence of
infected blood monocytes and cell to cell virus transfer; (iii) EAV infection in these capillaries alters the
vascular permeability leading to leakage (edema/serous exudate) but not to vasculitis and hemorrhages.
5. Acknowledgements
The authors thank M. Bauwens, C. Boone and A. Mészáros for technical support. The authors acknowledge
K. De Hert, S. Sivilia, A. Giuliani and L. Szeredi for their practical help. Prof. M. Pensaert is acknowledged
for his scientific contribution, helpful suggestions and fruitful discussions. Permission to use the facilities of
the Department of Scienze Mediche Veterinarie, Faculty of Veterinary Medicine, Alma Mater Studiorum,
Bologna University and the laboratories of the National Food Chain Safety, Budapest is gratefully
acknowledged. S. Vairo is supported by a doctoral grant from the special research fund of Ghent University
(V72-0209).

Identification of target cells of a European equine arteritis virus strain in experimentally infected ponies
77
6. References
Alon R. and Ley K. (2008). Cells on the run: shear-regulated integrin activation in leukocyte rolling and
arrest on endothelial cells. Curr Opin Cell Biol, 20, 525.
Ayehunie S., Garcia-Zepeda E.A., Hoxie J.A., Horuk R., Kupper T.S., Luster A.D. and Ruprecht R.M.
(1997). Human immunodeficiency virus-1 entry into purified dendritic cells through CC and CXC
chemokine coreceptors. Blood, 90, 1379-1386.
Balasuriya U., MacLachlan N., 2004. The immune response to equine arteritis virus: potential lessons for
other arteriviruses. Vet. Immunol. Immunop., 102: 107-129;
Dejana E., Tournier-Lasserve E. and Weinstein B.M. (2009). The control of vascular integrity by
endothelial cell junctions: molecular basis and pathological implications. Dev Cell. 16, 209-221
Del Piero F. (2006). Equine viral arteritis: signs, lesions, pathogenesis and diagnoses. In: Proc
ACVP/ASCVA Annual, 2006.
Doll E.R., Bryans J.T. and Wilson J.C. (1968). Immunization against equine viral arteritis using modified
live virus propagated in cell cultures of rabbit kidney. Cornell Vet, 48, 497.
Echeverría M.G., Díaz S., Metz G.E., Serena M.S., Panei C.J. and Nosetto E. (2010). Evaluation of
neutralization patterns of the five unique Argentine equine arteritis virus field strains reported. Rev Argent
Microbiol, 42,11-17.
Eichhorn W., Heilmann M. and Kaaden O.R. (1995). Equine viral arteritis with abortions: Serological and
virological evidence in Germany. J Vet Med, 42, 574-576.
Feldmann H., Bugany H., Mahner F., Klenk H.D., Drenckhahn D. and Schnittler H.J. (1996).
Filovirus-induced endothelial leakage triggered by infected monocytes/macrophages. J Virol, 70, 2208-2214.
Go Y.Y., Zhang J., Timoney P.J., Cook R.F., Horohov D.W. and Balasuriya U.B. (2010). Complex
interactions between the major and minor envelope proteins of equine arteritis virus determine its tropism for
equine CD3+ T Lymphocytes and CD14+ Monocytes. J Virol, 84, 4898-4911.
Golnik, W., Moraillon A., . and Golnik J., 1986. Identification and antigenic comparison of equine arteritis
virus isolated from an outbreak of epidemic abortion of mares. Central bl Vet med B., 33: 413-417.
Gryspeerdt A., Chiers K., Govaere J., Vercauteren G., Ducatelle R., Van de Walle G. and Nauwynck
H. (2009). Neonatal foal death due to infection with equine arteritis virus in Belgium. Vlaams
Diergeneeskundig Tijdschr, 78, 189-193
Hahn G., Jores R. and Mocarski E.S. (1998). Cytomegalovirus remains latent in a common precursor of
dendritic and myeloid cells. PNAS, 95, 3937-3942.
Holyoak G.R., Balasuriya U.B., Broaddus C.C. and, Timoney P.J. (2008) Equine viral arteritis: current
status and prevention. Theriogenology, 70,: 403-414.
Hu J., Pope M., Brown C., O’Doherty U. and Miller C.J. (1998). Immunophenotypic characterization of
SIV-infected dendritic cells in cervix vagina and draining lymph nodes of rhesus monkeys. Lab
Investigation, 78, 435-451.

esn experimentally infected ponitrain iIdentification of target cells of a European equine arteritis virus s
78
Keynan Y., Card C.M., McLaren P.J., Dawood M.R., Kasper K. and Fowke K.R. (2008). The role of
regulatory T cells in chronic and acute viral infections. Clin Infect Dis, 46, 1046-1052.
Larsen L.E., Storgaard T. and Holm E. (2001). Phylogenetic characterization of the G(L) sequences of
equine arteritis virus isolated from semen of asymptomatic stallions and fatal cases of equine viral arteritis in
Denmark. Vet Mic, 80, 339-346.
Lipscomb M.F. and Masten B.J. (2002). Dendritic Cells: Immune Regulators in Health and Disease.
Physiol Rev, 82, 97-130.
Luscinskas F.W., Ma S., Nusrat A., Parkos C.A. and Shaw S.K. (2002). The role of endothelial cell
lateral junctions during leukocyte trafficking. Immunol Rev, 186, 57-67.
Marsac D., García S., Fournet A., Aguirre A., Pino K., Ferres M., Kalergis A.M., Lopez-Lastra M. and
Veas F. (2011). Infection of human monocyte-derived dendritic cells by ANDES Hantavirus enhances pro-
inflammatory state, the secretion of active MMP-9 and indirectly enhances endothelial permeability. J Virol,
13, 223.
Michel C.C. and Curry F.E. (1999). Microvascular permeability. Physiol Rev, 79, 703-761.
Monreal L., Villatoro A.J., Hooghuis H., Ros I. and Timoney P.J. (1995). Clinical features of the 1992
outbreak of equine viral arteritis in Spain. Equine Vet J, 27, 301-304.
O'Sullivan M.A. and Killen H.M. (1994). The differentiation state of monocytic cells affects their
susceptibility to infection and the effects of infection by dengue virus. J Gen Virol, 75, 2387 2392.
Ruggiero E., Toschi E. and Federico M. (2011). The activity of matrix metalloproteinase-9 is part of the
mechanism of cell-to-cell HIV-1 endocytosis in dendritic cells. Curr Drug Discov Technol, 8, 112-118.
Schnittler H., Stroeher U., Afanasieva T. and Feldmann H. (2004). The role of endothelial cells in
filovirus hemorrhagic fever. In: Ebola and Marburg viruses: molecular and cellular biology. Feldmann H.
editors, Horizon Biosciences, Norfolk, United Kingdom, 279-304.
Smith J. (2003). Mycobacterium tuberculosis pathogenesis and molecular determinants of virulence. Clin
Microbiol Rev, 16, 463-496.
Span A.H., Mullers W., Miltenburg A.M. and Bruggeman C.A. (1991). Cytomegalovirus induced PMN
adherence in relation to an ELAM-1 antigen present on infected endothelial cell monolayers. Immunology,
72, 355-360.
Steukers L., Glorieux S., Vandekerckhove A., Favoreel H. and Nauwynck H. (2012). Diverse microbial
interactions with the basement membrane barrier. Trends Microb, 20, 147-155.
St-Louis M. and, Archambault D., 2007. The equine arteritis virus induces apoptosis via caspase-8 and
mitochondria-dependent caspase-9 activation. Virology, 367:147-155.
Szeredi L., Hornyák Á., Pálfi V., Molnár T., Glávits R. and Dénes B. (2005). Study on the epidemiology
of equine arteritis virus infection with different diagnostic techniques by investigating 96 cases of equine
abortion in Hungary. Vet Mic, 108, 235-242.

Identification of target cells of a European equine arteritis virus strain in experimentally infected ponies
79
Vairo, S., Vandekerckhove, A., Steukers, L., Glorieux, S., Van den Broeck, W. and, Nauwynck, H.J.,
(2012). Clinical and virological outcome of an infection with the Belgian equine arteritis virus strain
08P178.8, Vet. Mic., 157, 333-344;
Vairo S., Van den Broeck W., Favoreel H., Scagliarini A., Nauwynck H. (2013). Development and use of
a polarized equine upper respiratory tract mucosal explant system to study the early phase of pathogenesis of
a European strain of equine arteritis virus. Vet Res
Van der Meulen K., Caij A., Nauwynck H. and Pensaert M. (2001). An outbreak of equine viral arteritis
abortion in Belgium. Vlaams Diergeneeskd Tijdschr, 70, 221-222.
Wagner D.D. and Frenette P.S. (2008). The vessel wall and its interactions. Blood, 111, 5271.
Wood J.L., Chirnside E.D., Mumford J.A. and Higgins A.J. (1995). First recorded outbreak of equine
viral arteritis in the United Kingdom. Vet Rec, 136, 381-385.
Zhang J. (2005) Permissiveness of selected cell lines to equine arteritis virus: establishment, characterization,
and significance of persistent infection in HELA cells. Ph.D. dissertation, University of Kentucky Doctoral
Dissertations. Paper 478.
Zhang J., Timoney P.J., Shuck K.M., Seoul G., Go Y.Y., Lu Z., Powell D.G., Meade B.J. and
Balasuriya U.B..R. (2010). Molecular epidemiology and genetic characterization of equine arteritis virus
isolates associated with the 2006–2007 multi-state disease occurrence in the USA. J Gen Virol, 91, 2286-
2301.


Sabrina Vairo, Wim Van den Broeck, Herman Favoreel, Alessandra Scagliarini, Hans Nauwynck
Development and use of a polarized equine upper respiratory tract mucosal explant system to
study the early phase of pathogenesis of a European strain of equine arteritis virus
Accepted by Veterinary Research

Equine polarized mucosal explants to study the early phase of EAV pathogenesis
80
Abstract
The upper respiratory tract mucosa represents the first line of defense, which has to be overcome by
pathogens before invading the host. Considering the economical and ethical aspects involved in using
experimental animals for pathogenesis studies, respiratory mucosal explants, in which the three-dimensional
architecture of the tissue is preserved, may be ideal alternatives. Different respiratory mucosal explant
cultures have been developed. However, none of them could be inoculated with pathogens solely at the
epithelium side. In the present study, equine nasal and nasopharyngeal explants were embedded in agarose
(3%), leaving the epithelium side exposed to allow apical inoculation. Morphometric analysis did not show
degenerative changes during 72h of cultivation. The number of apoptotic cells in the mucosa slightly
increased over time. After validation, the system was used for apical infection with a European strain
(08P178) of equine arteritis virus (EAV) (107.6
TCID50/ml per explant). Impermeability of agarose to virus
particles was demonstrated by absence of labeled microspheres (40nm) and lack of EAV-antigens in RK13
cells seeded underneath the agarose layer in which inoculated explants were embedded. In nasal explants,
27% of the EAV-positive cells were CD172a+ and 19% were CD3
+. In nasopharyngeal explants, 45% of the
EAV-positive cells were CD172a+ and 15% were CD3
+. Only a small percentage of EAV-positive cells were
IgM+
B lymphocytes This study validates the usefulness of a polarized mucosal explant system and shows
that mucosal CD172a+ myeloid cells and CD3
+ T lymphocytes represent important target cells for EAV in
the respiratory mucosa.
1. Introduction
The respiratory mucosa consists of epithelium and the underlying connective tissue or lamina propria which
are separated by a firm barrier, the basement membrane (BM). It lines the respiratory tract, including the
nasal cavity, the nasopharynx, the larynx, the trachea and the bronchial tree and represents the first line of
defense against respiratory infections. However, several pathogens, including equine arteritis (EAV) virus,
use the respiratory tract as portal of entry to invade the host (Vairo et al., 2012). At the level of the
respiratory mucosa, the mechanisms of replication and invasion of many of these pathogens are largely
unknown. In vivo experiments are difficult to perform due to their costs and ethical constraints. Recently,
Glorieux et al. (2007), Vandekerckhove et al. (2009), and Steukers et al. (2011) have developed ex vivo
explant models using gauzes to cultivate respiratory mucosae of pigs, horses and cattle, respectively. These
models were successfully used to get better insights in the invasion mechanisms of alphaherpesviruses
(Steukers et al., 2012). However, the impossibility to inoculate the mucosa solely at the epithelial side was a
limitation. In fact, inoculation of equine explants cultivated on gauzes with pseudorabies virus (PRV)
resulted in infection of endothelial cells while the epithelium above them was unaffected and, therefore, the
author concluded that infection occurred via the side regions of the explants (Vandekerckhove et al., 2011).
In line with this, when equine explants cultivated on gauzes are experimentally infected with a European
EAV strain (08P178), the majority of infected cells were localized in the connective tissue mainly at the
bottom and lateral sides of the explants. In an in vivo situation, viruses have to overcome the epithelial layer

Equine polarized mucosal explants to study the early phase of EAV pathogenesis _
81
before invading the host. In the explant system using only gauzes, all sides of the mucosal explants become
exposed to the virus and this barrier phenomenon is, therefore, lost. As a consequence, early pathogenic
events of certain respiratory agents, such as EAV, may be quite different in these ex vivo models compared
to those in the in vivo situation. Thus, it was necessary to create a model where the virus could only be
delivered at the apical side. The present study describes a polarized agarose embedded explant model which
allows cultivation of equine nasal and nasopharyngeal mucosae in a semi-closed system where only the
epithelium side of the explant is exposed to virus upon inoculation. Agarose is a linear galactose polymer
obtained by purification of agar (Araki, 1958) and it is frequently used as an alternative to agar in viral
plaque assays (de Maeyer and Schonne, 1964) and microbiological investigations (Hamilton et al., 2006). In
the present study, agarose embedded respiratory mucosal explants were tested for morphometric integrity
and cell viability. Additionally, this polarized ex vivo model was used to investigate EAV replication sites
and to identify the mucosal target cells.
2. Material and methods
2.1. Collection of nasal and nasopharyngeal mucosae and preparation of agarose embedded explant cultures
Nasal septum, nasopharynx and blood from seven horses of both genders and different ages (3-6 years old)
were collected at the slaughterhouse as previously described (Vandekerckhove et al., 2009). The collected
tissues were immediately transported to the laboratory in transport medium: phosphate buffered saline (PBS)
supplemented with 10µg/mL gentamicin (Invitrogen, Ghent, Belgium), 1mg/mL streptomycin (Certa, Braine
l'Alleud, Belgium), 1mg/mL kanamycin (Sigma, Bornem, Belgium), 1000U/mL penicillin (Continental
Pharma, Puurs, Belgium) and 5μg/mL fungizone (Bristol-Myers Squibb, New York, USA). The mucosae
were stripped from the underlying layers and divided in equal fractions of 2cm2. The nasal explants were
cultured on gauzes for 24h in 50% D-MEM (Invitrogen)/50% F12 (Invitrogen) serum-free medium
supplemented with 1μg/mL gentamicin, 0.1mg/mL streptomycin and 100U/mL penicillin. The
nasopharyngeal explants were cultured on gauzes for 24h in 50% D-MEM (Invitrogen)/50% RPMI
(Invitrogen) serum-free medium supplemented with 1nM Ca++
, 1μg/mL gentamicin, 0.1mg/mL streptomycin
and 100U/mL penicillin.
Six-well culture dishes were pre-seeded with rabbit kidney cells (RK13) and incubated at 37°C in the
presence of 5% CO2 until confluency was reached. Afterwards, RK13 medium was replaced with 3ml of a
solution containing 50% of sterile 3% agarose (low temperature gelling; Sigma) and 50% of 2X medium
(50% 2X D-MEM/50% 2X F12 supplemented with 2μg/mL gentamicin, 0.2mg/mL streptomycin and
200U/mL penicillin for nasal explants; 50% 2X D-MEM/50% 2X RPMI supplemented with 2nM Ca++
,
2μg/mL gentamicin, 0.2mg/mL streptomycin and 200U/mL penicillin for nasopharyngeal explants). Once
the agarose layer solidified, 1.5cm2 of stripped mucosa was placed on top of it with the epithelium upwards.
Additional agarose was added until the lateral surfaces of the mucosa were fully occluded. Finally, to avoid
drying out during incubation (37°C and 5% CO2), the nasal and nasopharyngeal mucosae were covered with
a tiny film of nasal or nasopharyngeal serum-free medium, respectively. Nasal and nasopharyngeal mucosae

Equine polarized mucosal explants to study the early phase of EAV pathogenesis
82
collected from three horses were used to examine the structural integrity and viability of the agarose
embedded explants. Upper respiratory mucosae of the remaining four animals were included to investigate
EAV 08P178 replication and target cells in the agarose embedded explants. To exclude previous EAV
infection of the donor animals, blood samples collected at the slaughterhouse were tested for EAV specific
antibodies by means of a complement-dependent virus neutralization (VN) test and an immunoperoxidase
monolayer assay (IPMA) as previously described (Vairo et al., 2012). Further, to exclude recent EAV
infection, 20% suspensions of nasal and nasopharyngeal mucosae were processed for virus titration on RK13
cells immediately after collection at the slaughterhouse.
2.2. Morphometric analysis
At 0, 24, 48 and 72h of cultivation, nasal and nasopharyngeal explants were fixed in phosphate buffered
3.5% formaldehyde solution for 24h. After fixation, the tissues were further processed with an automated
system (Thermo scientific - Microm STP 420D) and embedded in paraffin. Afterwards, haematoxylin-eosin
(HE), reticulin and Van Gieson stainings were performed. At each time point, the effect of ex vivo cultivation
on the mucosa architecture was evaluated in five randomly selected fields. The structure and thickness of the
epithelium were analyzed in the HE stained sections. The thickness and the continuity of the basement
membrane were evaluated in the reticulin stained sections. The relative amounts of collagen and nuclei were
calculated in a defined region of interest from five randomly chosen fields in the Van Gieson stained
sections. All measurements and calculations were performed using the Cell F Software linked to a BX61
light microscope (Olympus Optical Co., Hamburg, Germany).
2.3. Evaluation of the cell viability
The effect of ex vivo cultivation on cell viability was evaluated using an in-situ cell death detection kit
(Roche Diagnostics Corporation, Basel, Switzerland), based on terminal deoxynucleotidyl transferase
mediated dUTP nick end labeling (TUNEL). The TUNEL test was performed according to the
manufacturer's guidelines on cryosections preserved in methocel® (Sigma). In both epithelium and
connective tissues, the percentage of TUNEL-positive cells was assessed in five randomly chosen fields of
100 cells each.
2.4. Validation of the impermeability of agarose to virus particles
To confirm the agarose impermeability to virus particles, a preliminary experiment with fluorescent
microspheres was performed. Briefly, a dual-chamber system was created by placing a transwell insert (pore
size 0.4µm, 12mm of diameter; Millipore corporation, Bedford, MA) filled with 3ml of 50% agarose (3%)
and 50% 2X medium into a culture dish of a six-well plate. Three ml of PBS were added to the bottom
compartment. One ml of PBS containing 107.6
streptavidin-labeled microspheres (40nm) (Invitrogen) with
slightly smaller dimensions compared to EAV particles (50-100nm) (Bürki et al., 1992) was brought in the
upper compartment, on top of the agarose layer. Zero, 1, 12, 48, 72 and 96h later, 50µl of fluid was collected

Equine polarized mucosal explants to study the early phase of EAV pathogenesis _
83
from both the upper and the bottom compartment and analyzed with a fluoroskan ascent FL (Thermo lab
system, Altrincham, GB).
To verify that the virus did not cross the agarose layer during the experimental inoculation, wells used to
culture agarose embedded EAV-inoculated explants were pre-seeded with RK13 cells. After collection of the
explants, the agarose layer was gently removed from the wells and the underlying RK13 monolayer was
washed, air dried and submitted for indirect immunohistochemistry (IHC). Briefly, the monolayers were
fixed with 4% paraformaldehyde for 10min at room temperature, washed twice with PBS and further
incubated with methanol supplemented with 0.1% H2O2. The monolayers were subsequently incubated for
1h with mouse monoclonal antibody (mAb) 17D3 (1:100), specific for the nucleocapsid (N) protein of EAV
(VMRD, Pullman, USA) and with a peroxidase-labeled goat anti-mouse IgG immunoglobulin (Molecular
Probes, Oregon, USA) (1:500). Viral antigen-positive RK13 cells were visualized with 0.05M of 3-amino-9-
ethylcarbazole in acetate buffer (pH5) supplemented with 0.05% H2O2 and detected by light microscopy
(Olympus Optical Co.).
2.5. Virus inoculation
After validation of the agarose embedded explant system, the model was used to investigate EAV replication
and to identify the target cells. Nasal and nasopharyngeal agarose embedded mucosal explants of three
horses were inoculated with 1ml of medium containing 107.6
50% endpoint tissue culture infectious dose
(TCID50) of a European respiratory strain of EAV (isolated in Belgium, designated 08P178, 4th passage on
RK13 cells) [GenBank: JN25761].(Vairo et al., 2012). The agarose embedded explants of a fourth animal
were mock-inoculated with 1ml of medium. After 1h of incubation at 37°C and 5% CO2, the inoculum was
removed, the explants were gently washed twice and further incubated with fresh medium. At 0, 24, 48 and
72hpi, specimens were collected for quantification and identification of EAV infected cells by double
immunofluorescence staining (IF).
2.6. Identification of EAV infected cells
To quantify and identify individual EAV-positive cells, a double immunofluorescence staining was
performed. Of each donor animal and at each collection time, sixty 8µm-thick cryosections of nasal mucosa
and sixty 8µm-thick cryosections of nasopharyngeal mucosa were cut and fixed in methanol at -20°C for
20min. For each tissue, twenty cryosections were stained for each cell surface marker separately. In the first
step, optimal dilutions of mAb DH59B (VMRD), UC F6G-3 (California University, Davis, USA) or 1.9/3.2
(VMRD) were used as markers for CD172a+ cells (monocyte lineage), CD3
+ cells (pan T lymphocytes) and
IgM+
cells (B lymphocytes), respectively. Afterwards, slides were incubated with Texas Red®-labeled goat
anti-mouse IgG antibodies (Molecular Probes). In the second step, EAV nucleocapsid (N) protein was
visualized with mAb 17D3 (VMRD), directly labeled with Zenon® Alexa Fluor
® 488 Mouse IgG1 Labeling
Kit, according to the manufacturer’s instructions. As negative control, sections of the mock-inoculated nasal
mucosa were stained following the aforementioned protocol. In addition, ten extra sections of inoculated

Equine polarized mucosal explants to study the early phase of EAV pathogenesis
84
nasal mucosa were incubated with CD172a+ as primary antibody and Texas Red
®-labeled goat anti-mouse
IgG antibodies as secondary antibody. In a second step, the glass was further incubated with an irrelevant
IgG1 (13D12) directly labeled with Zenon® Alexa Fluor
® 488 Mouse IgG1 Labeling Kit. All antibodies were
diluted in PBS and incubated for 1h at 37°C. The nuclei were counterstained with Hoechst 33342 (Molecular
Probes) for 10min followed by a further fixation step of 5min with 4% paraformaldehyde. The number of
viral-antigen positive cells was determined in an area of 8mm2 for each nasal and nasopharyngeal mucosae.
Further, at each time point, the percentage of double positive cells (EAV-surface marker+) was calculated on
the total amount of EAV-positive cells counted in the 20 sections stained for that specific cell marker. The
samples were analyzed with a confocal microscope (Leica TCS SP2 Laser scanning spectral confocal
system, Leica microsystems GmbH, Wetzlar, Germany).
2.7. Statistical analysis
Morphometric parameters and viability were analyzed using prisma software to evaluate the variance
(ANOVA). The data are presented as means ± standard deviation of triple independent experiments. The
results were considered to be significantly different when p < 0.05.
3. Results
3.1. Morphometry
In both nasal and nasopharyngeal mucosal explants, the architecture of the epithelium (Figure 1a), and the
continuity of the basement membrane (Figure 2a) were preserved throughout the entire experiment. The
thickness of the epithelium and the basement membrane (Figures 1b and 2b, respectively) did not
significantly change over time.
In both nasal and nasopharyngeal mucosae, the relative amounts of nuclei (Figures 3a and 3b) and collagen
(Figures 3a and 3c) were conserved over time.
Table I. Percentage of TUNEL-positive cells in epithelium and underlying connective tissue
Percentage of TUNEL-positive cells at…h of cultivation
0
24
48
72
Nasal mucosa epithelium 0.2 ± 0.4 0.6 ± 0.6 0.4 ± 0.2 0.4 ± 0.8
lamina propria 0.8 ± 0.7 1.3 ± 1 3 ± 0.7 4.9 ± 2.7
Nasopharyngeal
mucosa
epithelium 0.4 ± 0.3 0.8 ± 1 0.7 ± 1.3 0.7 ± 0.5
lamina propria 0.5 ± 0.8 1.7 ± 1.8 3 ± 0.4 5.5 ± 1.9

Equine polarized mucosal explants to study the early phase of EAV pathogenesis _
85
Figure 1. Evaluation of the continuity (a) and the thickness (b) of nasal and nasopharyngeal epithelium (HE-
staining). The epithelial layer remained intact during the entire experiment (72h). The thickness (indicated by
arrow) of the epithelial layer did not show significant differences over time. Data are represented as means ±
S.D. of triplicate independent experiments.
Figure 2. Evaluation of the continuity (a) and the thickness (b) of nasal and nasopharyngeal basement
membrane (reticulin-staining). The collagen type III reticular fiber layer of the basement membrane (lamina
reticularis) (indicated by arrows) remained continuous during the entire experiment (72h). No significant
differences in the thickness of the basement membrane were found at any time point. Data represent means ±
S.D. of triplicate independent experiments.

Equine polarized mucosal explants to study the early phase of EAV pathogenesis
86
3.2. Viability
The effect of ex vivo culture on explant viability is given in Table I. The nasal and nasopharyngeal mucosal
explants showed a slight, not statistically significant, increase in the number of apoptotic epithelial and
stromal cells at 72h of cultivation.
3.3. Ability of agarose to restrain virus particles
The bead assay as specified in the material and methods indicated the ability of agarose to restrain virus
particles. None of the samples collected from the bottom compartment of the dual-chamber system contained
streptavidin-labeled microspheres at any time point. Microspheres remained present in the fluid collected
from the upper compartment up to 96hpi. In the absence of agarose, microspheres were readily observed in
the lower compartment.
In addition, IHC staining performed on the RK13 monolayer seeded underneath the EAV-inoculated agarose
embedded explants did not reveal EAV-positive cells at any time point.
3.4. Quantification and identification of EAV-antigen positive cells in agarose embedded explants
Samples included as negative control did not show EAV-positive cells at any time point. The number of
EAV-positive cells increased in time with a mean of 68 in 8mm2 of nasal mucosa and 225 in 8mm
2 of
nasopharyngeal mucosa at 72hpi. In particular, 74, 76 and 54 antigen-positive cells/8mm2 were counted in
nasal explants (Figure 4a) and 201, 254 and 220 EAV-positive cells/8mm2 were quantified in
nasopharyngeal explants (Figure 4b) of horse I, II and III, respectively. Specifically, in nasal mucosa, a mean
Figure 3. Percentage of cells (a-b) and collagen (a-c) in nasal and nasopharyngeal mucosae (Van Gieson-
staining). By giving different colors to collagen and nuclei (setting a threshold), relative amounts of collagen
and nuclei were measured during cultivation (72h) by means of light microscopy. No significant differences
in percentage of cells and collagen were found at any time point. Data represent means ± S.D. of triplicate
independent experiments.

Equine polarized mucosal explants to study the early phase of EAV pathogenesis _
87
of 22, 34 and 27% of EAV-positive cells were identified as CD172a+ at 24, 48 and 72hpi, respectively. The
percentage of EAV infected cells positive for CD3 increased linearly from 10 (24hpi) to 19% (48 and 72hpi)
during the experiment (Figure 5a). In the nasopharyngeal mucosal explants, the percentage of EAV infected
cells positive for CD172a increased from 22 (24hpi) to 26 (48hpi) to 45% (72hpi) over time, while the
percentage of EAV infected CD3 cells reached 22% at 24hpi and decreased to 15% afterwards (48 and
72hpi) (Figure 5b). At 24hpi, EAV-positive cells were mainly localized in the upper layers of both nasal and
nasopharyngeal mucosae and, at later time points (48 and 72hpi), were scattered all over the connective
tissue. In general, individual EAV-infected cells were mostly found under the basement membrane with
sporadic presence in between epithelial cells. In addition, in nasopharyngeal explants individual EAV-
positive cells were frequently localized in the parafollicular area. Viral antigen-positive cells were not found
at the cutting edges of the agarose embedded explants. Intracellularly, viral antigens were seen in the
cytoplasm (Figure 6), in the form of small dots (Figure 6a) but mostly in the form of large masses occupying
most of the cytoplasm (Figures 6b and c).
Figure 4. Number of antigen-positive cells/8mm2 in agarose embedded nasal (a) and nasopharyngeal (b)
explants.
Hours post inoculation

Equine polarized mucosal explants to study the early phase of EAV pathogenesis
88
Figure 6. Representative confocal photomicrographs illustrating viral tropism for leukocyte subpopulations.
Two different types of EAV-positive cells can be distinguished: big cells with a big round or oval nucleus
surrounded by large cytoplasmic extensions, mostly CD172a+ (a) and rather small cells with a big round or
oval nucleus surrounded by a small rim of cytoplasm, mostly CD3+
or IgM+ (b-c). The viral antigens could
be found in the form of small dots (a) or in the form of large masses occupying most of the cytoplasm (b-c).
Scale bar represents 10 µm.
Figure 5. Identification of EAV-positive leukocytes in agarose embedded nasal (a) and nasopharyngeal (b)
explants. Lines show the means ± S.D. of triplicate independent experiments.
Hours post inoculation Hours post inoculation

Equine polarized mucosal explants to study the early phase of EAV pathogenesis _
89
4. Discussion
The mucosa of the upper respiratory tract (URT) represents the first line of defense against respiratory
pathogens but often also serves as portal of entry for several microorganisms, whether or not causing a
generalized infection. The principal viral agents responsible for respiratory disorders in horses are equine
herpesvirus-1 (EHV-1) and equine herpesvirus-4 (EHV-4) (Van Maanen, 2002), equine arteritis virus (EAV)
(Del Piero, 2000), and equine influenza virus (Mumford et al., 1998). A previous system used by
Vandekerckhove et al. (2009) allowed to cultivate four representative tissues of the equine URT up to 96h
without significant changes in morphology and viability (Vandekerckhove et al., 2009). In that model, the
explants were cultivated on gauzes allowing nutrients to penetrate through all sides of the mucosa. Because
of the small size of the explants (0.5cm2, few mm thick) and the open edges, nutrients have free access to the
inner parts of the tissues. In this system, pathogens, just like nutrients, do not only enter the mucosa at the
upper epithelium side, but also at the edges through the open spaces of the connective tissue. In fact,
inoculation of such explant systems with PRV (Vandekerckhove et al., 2011) or EAV 08P178 (own studies,
not published) resulted in an extensive spread of the virus mainly at the bottom and lateral sides of the
mucosal explants. Therefore, in order to better mimic the in vivo situation, we established a new model in
which, through agarose embedded explants, a polarized system was obtained so that pathogens could only
enter through the upper epithelium. The results of morphometric analysis of epithelium, basement membrane
and connective tissue and of TUNEL assays demonstrated that the agarose system allows to cultivate
explants up to 72h without substantial alterations in the morphology and the viability of the mucosa of the
explants. Further, an assay based on streptavidin-labeled microspheres together with the absence of EAV
antigens in RK13 cells seeded underneath the agarose layer in which inoculated explants were embedded and
the lack of viral antigen-positive cells at the cutting edges of the agarose embedded explants, demonstrated
that the access of the pathogens in the system is restricted to the apical side of the epithelium.
We have recently shown that, upon in vivo oro-nasal inoculation, EAV 08P178 starts its replication in nasal
and nasopharyngeal regions and lungs (Vairo et al., 2012). Therefore, to gain more insights into the early
phase of EAV respiratory infection, kinetic studies were performed by inoculating the agarose embedded
nasal and nasopharyngeal mucosal explants with EAV 08P178. Currently, little is known about the early
phases of EAV pathogenesis such as (i) which strategy EAV uses to invade the host, (ii) what are the first
cells to be infected at the entry site or (iii) how the virus reaches blood vessels to disseminate throughout the
body. The current study showed that in the explant model, the majority of EAV-positive cells were CD172a+
cells followed by CD3+ T lymphocytes, whereas only a small percentage were IgM
+ B lymphocytes.
Previously, we have speculated that, at the level of the URT, EAV hijacks mononuclear leukocytes to
penetrate the BM and evade the immune response (Vairo et al., 2012). In possible support of this, the present
study showed a high percentage of EAV-infected CD172a+ cells. CD172a is expressed on equine monocytes,
macrophages, dendritic cells (DCs) and granulocytes. DCs have specific characteristics: (i) they have a major
responsibility in activating naïve and resting antigen-specific T cells (Ingutti et al., 1997) (ii) they are
migratory cells, unlike differentiated macrophages (Lipscomb and Masten, 2002) and (iii) they are

Equine polarized mucosal explants to study the early phase of EAV pathogenesis
90
abundantly present within the airway epithelium (Holt and Stumbles, 2000) forming a network of antigen
presenting cells in the respiratory mucosa (Banchereau and Steinman, 1998 and Holt et al., 1989). They play
a central role in initiating an immune response against infecting agents (Banchereau and Steinman, 1998) but
may also contribute to the pathogenesis and the spreading of several respiratory pathogens as has been
shown for human (Ayehunie, 1997) and simian (Hu et al., 1998) immunodeficiency viruses,
cytomegalovirus (Hahn et al., 1998), and Mycobacterium tuberculosis (Smith, 2003). Hence, it is tempting to
speculate that DCs could play an important role in the early phases of EAV pathogenesis. This theory could
corroborate the findings of Del Piero (2000) that described EAV-antigens within stromal dendrite-like cells
of lymph node sinuses and spleen (Del Piero, 2000). On the other side, since the main function of myeloid
cells is to capture antigens, CD172a+
cells may be positive for the EAV nucleocapsid protein due to
phagocytosis rather than to infection.
It was generally believed that EAV has a tropism mainly for monocyte/macrophage population. However,
Castillo-Olivares and collaborators (2003) showed that EAV-infected PBMC were negative for the human
monocyte/granulocyte marker L1 (calprotectin) (Castillo-Olivares et al., 2003). In the present study, both T-
lymphocytes and cells of the monocyte/macrophage lineage were found to be infected with EAV. Other
studies also demonstrated that EAV can infect CD3+ T lymphocytes in PBMC in vitro (Go et al., 2010 and
2011)
In the present study, 37 to 64% of EAV-positive cells were identified. Since EAV has a wide tropism (Del
Piero, 2006), the percentage of non-identified EAV-positive cells may be epithelial cells, endothelial cells,
mesenchymal stromal cells, and natural killer cells. In fact, in vivo, EAV-antigens were demonstrated within
the cytoplasm of endothelial cells, myometrial and cardiac myocytes, chorionic mesenchymal stromal cells
and epithelial cells such as alveolar pneumocytes, enterocytes, adrenal cortical cells, trophoblast, thymus
stroma, and renal tubular cells (Del Piero, 2006). Therefore, it will be interesting to further identify the
remaining EAV-positive cells in future experiments using the agarose explant system.
Although polarized mucosal explants are much more physiological than non-polarized ones and in vitro cell
cultures, they are only suitable to study the early stages of the pathogenesis of viruses that use the respiratory
tract as portal of entry. It must be kept in mind that the mucosal explants not fully represent the whole scope
of virus-animal interactions as the inflammatory processes including invasion of blood elements are absent.
However, preliminary results obtained with nasal mucosa from experimentally EAV-inoculated ponies were
very similar to those of the present in vitro study, confirming the relevance of the polarized mucosal explant
system. In conclusion, the present study demonstrates that the agarose embedded explant model represents a
valid tool to study some aspects of the early steps in the pathogenesis of equine respiratory viruses such as
EAV and that mucosal CD172a+ myeloid cells and CD3
+ T lymphocytes represent important target cells for
EAV.

Equine polarized mucosal explants to study the early phase of EAV pathogenesis _
91
5. Acknowledgements
The authors thank M. Bauwens, L. De Bels, L. Standaert and P. De Matteo for the technical support. The
authors acknowledge M. Pensaert, K. De hert, D. Saha, A. Vandekerckhove, L. Steukers, S. Glorieux, L.
Gallina, S. Sivilia, C. Mangano and A. Giuliani for their scientific contribution, helpful suggestions and
fruitful discussions. Permission to use the facilities of the department of scienze mediche veterinarie, Faculty
of Veterinary Medicine, Alma Mater Studiorum, Bologna University and, in particular, prof. L. Calzà is
gratefully acknowledged. S. Vairo is supported by a doctoral grant from the special research fund of Ghent
University (V72-0209).
6. References
Araki C. (1958). Carbohydrate chemistry of substances of biological interest. In: Fourth Int Congress of
Biochemistry 4, 1-15.
Ayehunie S., Garcia-Zepeda E.A., Hoxie J.A., Horuk R., Kupper T.S., Luster A.D. and Ruprecht R.M.
(1997). Human immunodeficiency virus-1 entry into purified dendritic cells through CC and CXC
chemokine co-receptors. Blood, 90, 1379-1386.
Banchereau J. and Steinman R.M. (1998). Dendritic cells and the control of immunity. Nature, 392, 245-
252.
Bürki F., Hofer A. and Nowotny N. (1992). Objective data plead to suspend import-bans for seroreactors
against equine arteritis virus except for breeder stallions. J Appl Anim Res, 1, 31-42.
Castillo-Olivares J., Tearle J., Montesso F., Westcott D., Kydd J., Davis-Poynter N. and Hannant D.
(2003). Detection of equine arteritis virus (EAV)-specific cytotoxic CD8+ T lymphocyte precursors from
EAV-infected ponies. J Gen Virol, 84, 2745-2753.
de Maeyer E. and Schonne E. (1964). Starch gel as an overlay for the plaque assay of animal viruses.
Virology, 24, 13-18.
Del Piero F. (2000). Equine Viral Arteritis. Vet Pathol, 37, 287-296.
Del Piero F. (2006). Equine Viral Arteritis: Signs Lesions Pathogenesis and Diagnoses. In: Proc Annual
ACVP and ASVCP 2006.
Glorieux S., Van den Broeck W., van der Meulen K., van Reeth K., Favoreel H. and Nauwynck H.J.
(2007). In vitro culture of porcine respiratory nasal mucosa explants for studying the interaction of porcine
viruses with the respiratory tract. J Virol Methods, 142, 105-112.
Go Y., Bailey E., Cook D., Coleman S., Macleod J., Chen K., Timoney P.J. and Balasuriya U.B. (2011).
Genome-Wide Association Study among Four Horse Breeds Identifies a Common Haplotype Associated
with In Vitro CD3+ T Cell Susceptibility/Resistance to Equine Arteritis Virus Infection. J Virol, 85, 13174-
13184.
Go Y., Zhang J., Timoney P.J., Cook R., Horohov D. and Balasuriya U.B. (2010). Complex Interactions
between the Major and Minor Envelope Proteins of Equine Arteritis Virus Determine Its Tropism for Equine
CD3+ T Lymphocytes and CD14
+ Monocytes. J Virol, 84, 4898-4911.

Equine polarized mucosal explants to study the early phase of EAV pathogenesis
92
Hahn G., Jores R. and Mocarski E.S. (1998). Cytomegalovirus remains latent in a common precursor of
dendritic and myeloid cells. PNAS, 95, 3937-3942.
Hamilton A., Robinson C., Sutcliffe C., Slater J., Maskell J., Davis-Poynter N., Smith K., Waller A.
and Harrington J. (2006). Mutation of the maturase lipoprotein attenuates the virulence of streptococcus
equi to a greater extent than does loss of general lipoprotein lipidation. Infection and Immunity, 74, 6907-
6919.
Holt P.G. and Stumbles P.A. (2000). Characterization of dendritic cell populations in the respiratory tract. J
Aerosol Med, 13, 361-367.
Holt P.G., Schon-Hegard M.A., Phillips M.J. and McMenamin P.G. (1989). Ia-positive dendritic cells
form a tightly meshed network within the human airway epithelium. Clinical Exp Allergy, 19, 597-601.
Hu J., Pope M., Brown C., O’Doherty U. and Miller C.J. (1998). Immunophenotypic characterization of
SIV-infected dendritic cells in cervix vagina and draining lymph nodes of rhesus monkeys. Lab Invest, 78,
435-451.
Ingutti E., Mondino A., Khotuts A. and Jenkins M.K. (1997). In vivo detection of dendritic cell antigen
presentation to CD4T cells. J Exp Med, 185, 2133-2141.
Lipscomb M.F. and Masten B.J. (2002). Dendritic Cells: Immune Regulators in Health and Disease.
Physiol Rev, 82, 97-130.
Mumford E., Traub-Dargatz J., Salman M., Collins J., Getzy D. and Carman J. (1998). Monitoring and
detection of acute viral respiratory tract disease in horses. J Am Vet Med Assoc, 213, 385-390.
Schon-Hegrad M.A., McMenamin O.J. and Holt P.G. (1991). Studies on the density distribution and
surface phenotype of intraepithelial class II major histocompatibility complex antigen (la)-bearing dendritic
cells (DC) in the conducting airways. J ExpMed, 173, 1345-1356.
Smith J. (2003). Mycobacterium tuberculosis Pathogenesis and Molecular Determinants of Virulence. Clin
Microbiol Rev, 16, 463-496.
Steukers L., Glorieux S., Vandekerckhove A., Favoreel H. and Nauwynck H.J. (2012). Diverse
microbial interactions with the basement membrane barrier. Trends Microbiol, 20, 147-155.
Steukers L., Vandekerckhove A., Van den Broeck W., Glorieux S. and Nauwynck H.J. (2011).
Comparative analysis of replication characteristics of BoHV-1 subtypes in bovine respiratory and genital
mucosa explants: a phylogenetic enlightenment. Vet Res, 42, 1-11.
Vairo S., Vandekerckhove A., Steukers L., Glorieux S., Van den Broeck W. and Nauwynck H.J.
(2012). Clinical and virological outcome of an infection with the Belgian equine arteritis virus strain
08P178. Vet Mic, 157, 333-344.
Van Maanen C. (2002). Equine herpesvirus 1 and 4 infections: an update. Vet Q, 24, 58-78.
Vandekerckhove A. (2011). Equine nasal mucosal explants, a valuable tool to study early events of the
pathogenesis of equine herpesvirus infections. A PhD thesis Gent University Virology Immunology and
Parassitology Department; http://hdl.handle.net/1854/LU-1187433.

Equine polarized mucosal explants to study the early phase of EAV pathogenesis _
93
Vandekerckhove A., Glorieux S., Van den Broeck W., Gryspeerdt A., Van der Meulen K. and
Nauwynck H.J. (2009). In vitro culture of equine respiratory mucosa explants. Vet J, 181, 280-287


GENERAL DISCUSSION

General discussion
94
EVA - clinical signs and lesions with European versus North American strains
In this thesis, different aspects of EAV infection were investigated by inoculating eight EAV-free Shetland
ponies with a European field strain (08P178) of EAV, which was isolated from a neonatal foal that died shortly
after birth. Studies on the clinical signs, pathology or pathogenesis of a European EAV strain obtained from a
field outbreak and inoculated in experimental ponies had not been performed before. The most recent in vivo
studies with EAV North American strains date from at least 10 years ago (McCollum and Timoney, 1984). In
1998, McCollum and Timoney were able to classify 14 EAV strains in 4 different virulence classes based on the
severity and duration of clinical signs upon experimental inoculation of a total of 100 different horses. Virulence
was defined on the basis of the clinical signs which were absent, or mild with short lasting fever and any
combination of slight nasal or ocular discharge, mild depression and moderate appetite, or moderate with horses
being febrile for 3 or more days and developing any combination of abundant nasal or ocular discharge,
depression, lack of appetite, and pronounced edema, or severe with high prolonged fever, severe depression,
anorexia, copious nasal or ocular discharge, respiratory distress, diarrhea, stiffness, severe widespread edema,
and sometimes death. Thus strains were classified as follows: 1) avirulent (KY63, PA76, KY77, and ARVAC
vaccine); 2) mildly virulent (Swiss64, Austr68, SS85, PR85, and Ill94); 3) moderately virulent (KY84, ARIZ87,
Nebr89, and Ill93); and 4) severely virulent (VB53).
Upon oro-nasal inoculation with our EAV 08P178 strain, all the experimental ponies became infected and
developed clinical signs such as fever, ocular and nasal discharge, lymphadenomegaly, with only one of the
eight animals showing scrotal edema. The clinical picture of EAV 08P178 was consistently less severe
compared to the one shown by animals experimentally infected with the highly virulent velogenic EAV Bucyrus,
which caused 33% and 100% of fatal hypovolemia and hypotension in the studies reported by Bryans et al. in
1957 and Estes and Cheville in 1970, respectively. The duration of virus excretion in nasal secretions and
viremia were 2-3 days shorter with the EAV 08P178 compared to the moderately virulent North American strain
KY84. Further, horses experimentally inoculated with KY84 showed leukopenia, which is often described as a
very consistent clinical feature of EVA (Timoney and McCollum, 1993). No alterations in the white blood cell
counts were evidenced during an EAV 08P178 infection. It was proposed that, in animals acutely infected with
EAV, the reduction in the number of circulating leukocytes is due to apoptotic processes and, therefore,
indicative of the strain’s virulence (Balasurya et al., 2007) but currently, no consistent studies can support this
theory. The most common gross lesions observed in the EAV 08P178 infected animals were straw-colored fluid
present in the thoracic and abdominal cavities, congestion, prominent lobular pattern of the lungs, and lymph
node enlargement. The same findings, together with edema and petechial hemorrhages, were also described after
natural and experimental infection with North American strains, such as Bucyrus. It is believed that the
distribution and the extension of gross lesions are associated with the strain virulence (Timoney and McCollum,
1993). In our study, the lack of edema, except for a moderate scrotal edema for a few days in one animal, and the

General discussion _
95
absence of petechial hemorrhages indicate that large vessels were not or only minimally damaged during a
European EAV infection.
In conclusion, most of these features indicate that typical signs of the disease and the pathological changes
associated with EAV 08P178 can be experimentally reproduced through oro-nasal inoculation of ponies. In
addition, our results show that, although rather typical, signs and lesions were mild. It was, therefore, interesting
to study the European strain's organ and cellular tropisms both in vivo and ex vivo with the attempt to correlate
them with clinical and pathological observations made during an EAV 08P178 infection and to compare our
findings with those obtained with EAV North American strains.
EVA - pathogenesis
Studies in experimental horses with European EAV strains are scarce in Europe. One study has been performed
with an Italian strain (Autorino et al., 1994) which caused disease of significant clinical severity. Thus, it has
been assumed that results previously obtained with North American isolates would also be applicable to
European strains. It was, therefore, interesting to find similarities/dissimilarities in the pathogenesis and in the
course of infection of the European strains when compared to the published information on the North American
EAV isolates. The present studies address some of the main points of EAV pathogenesis such as the replication
characteristics over time at the portal of entry in ponies and “ex vivo” in nasal and nasopharyngeal explants.
Furthermore, the invasion and the dissemination of the virus in the body were followed and attempts were made
to identify the internal target organs and cells. These studies were intended to give better insights in the EAV
infection with regard to the virus-host interactions in a European context.
Primary replication site and invasion
Virus was present at 1, 3 and 3dpi in nasal and ocular secretions and BAL fluids of infected ponies, respectively.
This indicates that the primary local replication occurs at the level of the upper respiratory tract (URT),
conjunctiva and lungs. More specifically, since the highest virus titers in the tissues collected from animals
sacrificed at 3dpi were detected in the nasopharynx (106.5
TCID50/g) and tubal-nasopharyngeal tonsils (106.3
TCID50/g), it can be concluded that the most important site of viral replication at the portal of entry is the
nasopharyngeal region. To further substantiate the role of the URT in the early phases of EAV pathogenesis,
polarized nasal and nasopharyngeal mucosal explants were inoculated with EAV 08P178 and followed over
time. At 6, 12, 18, and 24 hpi, the virus titers in nasopharyngeal explants were significantly higher (nearly 1 log)
than in nasal explants, again confirming the main role of the nasopharynx as the preferable replication site of
EAV. Currently all the available literature on EAV pathogenesis studies states that “initial multiplication of the
virus takes place in the lungs and particularly into the alveolar macrophages” (Del Piero, 2000; Timoney, 2003
and Timoney and McCollum, 1993). Thus, our results show that, most likely, EAV 08P178 uses the
nasopharynx as primary portal of entry in the body.

General discussion
96
The URT can rather easily serve as portal of entry. The nasopharyngeal epithelium consists of a thin
pseudostratified columnar ciliated epithelium forming crypts and containing lymphoid follicles (Kumar and
Timoney, 2001). These areas, called follicle-associated epithelium (FAE), have a thin layer of epithelial cells
with reduced numbers of cilia, presence of leukocytes and absence of goblet cells. Towards the base of the
crypts, the epithelial cell height is drastically reduced. A study on human nasopharyngeal tonsils has
demonstrated a direct antigen transmigration from the cryptal epithelium to the underlying lymphoid tissue
(Hafeez et al., 2009). Further, FAE is able to absorb and transfer soluble antigens by pinocytosis (Tenner-Racz
et al., 1979 and Bockman and Stevens, 1977) and can also allow transport of particulate material (Richardson et
al., 1976) towards the subepithelial lymphoid tissue (Chen and Morrow, 1989). It is now suggested that EAV
may also use the FAE to invade the host. From a pathogenetic point of view, EAV replication at the level of the
URT could explain why virus spread through aerosolized virus occurs rather easily between horses in close
contact (Timoney, 1988 and Collins et al., 1987).
In conclusion, the studies carried out with the European EAV indicate that the virus uses the URT tract as portal
of entry into the host and that, in particular, the FAE of the nasopharynx likely serves as site of invasion.
Both the in vivo and the ex vivo studies showed that, at the level of the URT mucosae, EAV infected cells appear
as clusters and as individual cells. Clusters of EAV-positive cells were mainly detected in nasal mucosa while
individual infected cells were mostly present at the level of nasopharyngeal mucosa. Ex vivo samples collected at
24, 48 and 72hpi demonstrated a high percentage of CD172a and CD3 EAV-positive cells which, when studied
in detail time wise, appeared to migrate from the superficial to the deeper parts of the mucosa. The URT tissues
collected from inoculated animals sacrificed at 3dpi, showed large numbers of EAV-positive cells which were
mainly CD172a and CD3 positive and a significant perivascular infiltration of mononuclear cells which were
highly EAV-positive. From that time point, infected mononuclear cells were present in blood, indicating that
EAV may use individual mononuclear cells as Trojan horse to reach the blood stream and spread into the host.
In general, microorganisms, which cause generalized infections, have to overcome a number of barriers that may
block their invasion process during primary contact with the respiratory mucosal surfaces. An important
prerequisite for successful host invasion is to cross the basement membrane (BM), a 50-100nm-thick crucial
barrier underlying epithelia and endothelia. In both in vivo and ex vivo studies, the majority of EAV-positive
cells were CD172a+
cells followed by CD3+ T lymphocytes and only a small percentage were IgM
+ B
lymphocytes. CD172a is expressed on equine monocytes, macrophages, dendritic cells (DCs) and granulocytes.
Within the CD172a+ population only macrophages and DCs are known to play a role in carrying the virus and
are candidates to play an important role to transport the virus through the BM and to contribute to EAV invasion
in the body. To breach the BM, some microorganisms (e.g. pseudorabies virus) have developed ingenious
mechanisms such as enzymatic lysis (Glorieux et al., 2011). Others hijack host cells that are able to transverse
the BM (e.g. dendritic cells and other leukocytes) (Vandekerckhove et al., 2010). During physiological

General discussion _
97
processes, immune cells, and in particular DCs, frequently cross the BM barrier. Although the primary function
of immune cells and in particular DCs is to sample pathogens to initiate an immune response several pathogens
have developed mechanisms to use these cells as Trojan horses to cross the BM barrier and disseminate
throughout the host. This strategy is quite common within viruses such as herpesviruses, immunodeficiency
viruses, human papilloma virus, measles virus, dengue virus, and many others (reviewed by Steukers et al.,
2012). In equine respiratory mucosa, it was recently demonstrated both in in vivo (Gryspeerdt et al., 2010) and
ex vivo (Vandekerckhove et al., 2010) that, at the level of the URT, EHV-1 is able to spread via individual T
lymphocytes and myeloid cells.
It is generally believed that EAV has a particular tropism for cells of the monocytic lineage. However, already in
2003 it was shown that EAV-infected peripheral blood mononuclear cells (PBMC) were negative for the human
monocyte/granulocyte marker L1 (calprotectin). This indicates that EAV-infected target cells may not
preferentially belong to the monocyte/macrophage lineage or that only a subpopulation of differentiated
monocytes/macrophages is susceptible to EAV infection (Castillo-Olivares et al., 2003). DCs have specific
characteristics: (i) they play a major role in activating naïve and resting antigen-specific T cells (Ingutti et al.,
1997); (ii) they are migratory cells, unlike differentiated macrophages (Lipscomb and Masten, 2002) and (iii)
they are abundantly present within the airway epithelium (Holt et al., 1990), forming a network of antigen
presenting cells in the respiratory mucosa (Schon-Hegrad et al., 1991 and Holt et al., 1989). DCs play a central
role in initiating an immune response against infecting agents (Banchereau and Steinman, 1998) but may also
contribute to the pathogenesis and the spreading of several respiratory pathogens as it has been shown for human
(Ayehunie et al., 1997) and simian (Hu et al., 1998) immunodeficiency viruses, cytomegalovirus (Hahn et
al.,1998), and Mycobacterium tuberculosis (Smith, 2003). Hence, it is tempting to believe that DCs are playing
an important role in the early phases of EAV pathogenesis. This theory could corroborate the findings of Del
Piero (2000) who described EAV-antigens within stromal dendrite-like cells of lymph node sinuses and spleen.
The finding that EAV can infect CD3+ T lymphocytes both in vivo and ex vivo, provides the first report of a
member of this virus family being associated with T lymphocytes in vivo. In vitro studies (Go et al., 2010 and
2011) performed in PBMC demonstrated that virulent strains of EAV have the ability to infect CD3+ T
lymphocytes as well as monocytes, while avirulent strains retained the capacity to infect monocytes but not
CD3+ T lymphocytes, suggesting that either directly or indirectly tropism for CD3
+ T lymphocytes may have a
significant role in the pathogenesis of EAV infections in vivo. In general, T lymphocytes play an important role
in the clearance of viral infections (Keynan et al., 2008). In particular, it was shown that CD8+ T lymphocytes
are determinant in the clearance of EAV from the horse (Castillo-Olivares et al., 2003). Therefore, EAV
infection of T lymphocytes could result in a longer presence of the virus in its host.
In the present study, nearly 60% of EAV-positive cells at the level of the URT were identified as mononuclear
leukocytes and around 30% as endothelial cells. To fully understand the EAV mechanism of entry, the remaining

General discussion
98
virus-positive cells need to be identified. The unidentified EAV-positive cells could be epithelial cells,
mesenchymal stromal cells and/or natural killer cells. Even though a specific marker for epithelial cells was not
used in the present studies, the double IF staining performed against EAV N protein on the one hand and against
collagen VII on the other, revealed that clustered and individual EAV-positive cells were located in the
connective tissue below the BM, which allowed us to exclude that epithelial cells are being involved in the
infection.
In conclusion, this is the first time that EAV entry and tropism were investigated at the portal of entry in the
horse. Although several aspects need further investigation, our results show that EAV 08P178 targets myeloid
cells and T lymphocytes in nasal and nasopharyngeal mucosae, while epithelial cells are not involved.
Furthermore, indications were found that EAV may use individually infected cells, most probably DCs, as a kind
of Trojan horse to invade the host.
Viraemia and target organs
Viraemia is an important feature of EAV pathogenesis. In the ponies inoculated with the EAV 08P178, the virus
was isolated from PBMC between 3 and 10dpi and only sporadically from plasma. At maximum, 103 out of 10
6
PBMC co-cultured on RK13 were infected. Although positive results were obtained by means of co-culture,
EAV-positive PBMC were not detected after performing IF against EAV N protein on PBMC cytospin. Since
EAV was harvested from PBMC while the plasma was mostly negative, it can be concluded that the EAV
08P178 spreads in the blood mainly in a cell-associated form. Previous studies using the velogenic EAV
Bucyrus also demonstrated, although for longer periods and at higher titers, EAV in buffy coat and, to a lesser
extent, in plasma (Bryans et al., 1957 and MacLachlan et al., 1996). The finding that PBMC, while infectious for
RK cells, were negative for viral N protein by IF could be due to a temporary block of EAV replication. Go et al.
(2010) have found in PBMC, isolated from animals infected with the velogenic EAV Bucyrus, an indication of a
block of the later stages of the EAV replication cycle in the T lymphocyte subpopulation but a full replication
cycle in the CD14+
monocyte/macrophage subpopulation. A temporary block of the viral replication cycle in
blood mononuclear cells could support the hypothesis of EAV hiding in mononuclear leukocytes during the
dissemination phase. Other viruses, such as human cytomegalovirus (Singer and Jahn, 1996) and EHV-1
(Gryspeerdt et al., 2012) are also hampering expression of late proteins in migrating cells to reach secondary
replication sites through viraemia. Although the temporary block of EAV replication in PBMC is only
hypothetical at present, it may deserve further investigation.
From our findings we can conclude that, similarly to North American strains, EAV 08P178 disseminates to
internal target organs mainly through a cell-associated viraemia.
In our study, an increased vascular permeability was present in all tissues already at 3dpi and the percentage of
EAV- positive endothelial cells ranged between a minimum of 15% in the nasal septum to a maximum of 40% in
the lymphoid tissues associated with the lower respiratory tract. No endothelial cells were infected in the 3.200

General discussion _
99
vessels examined in which 3 or more endothelial cells were recognized (arteries, veins, arterioles, venules and
large capillaries). However, a minimum of 23% (in the intestines) and a maximum of 53% (in the lungs) of
terminal capillaries (less than 3 endothelial cells) were infected. Signs of an increased vascular permeability,
such as edema, congestion and inflammatory cell infiltration, were common findings in nearly all tissues and at
all time points. In addition, the presence of pleural and abdominal fluids at necropsy confirmed an increased
permeability of the capillaries. Another important step in the pathogenesis of generalized infections is the
migration of infected cells, such as PBMCs, from the blood stream towards the secondary organs. In these
studies, only endothelial cells of terminal capillaries were consistently involved in EAV infection processes,
while larger vessels were not. A similar finding was evidenced for other viruses such as Ebola virus (Schnittler
et al., 2004). It is known that small vessels represent the site for an increased paracellular permeability (Michel
and Curry, 1999) and for leukocyte extravasation during inflammation (Luscinskas et al., 2002). Currently,
nothing is known on the mechanism(s) by which EAV crosses the endothelial barrier to reach secondary organs.
Considering that the infection of endothelial cells is restricted to the capillaries, it is hypothesized that infected
blood leukocytes may adhere to the endothelium in the small capillaries and cause cell to cell transfer of virus to
infect endothelial cells during an extravasation process. In such a case, endothelial cells could be considered as
collaterally infected cells. The low viral titers detected in the plasma of the infected ponies at one side and the
relative high percentage of EAV-positive endothelial cells at the other side, can imply that the EAV replication
cycle is possibly not fully productive in the endothelial cells. A direct infection of endothelial cells via receptors
on their cell surface can be excluded. This would have led to a more general infection of endothelial cells in
vessels of different size. This hypothesis could also explain why only the small capillaries serve as site for
leukocyte adhesion and are involved in the EAV pathogenesis and why the number of endothelial cells and
capillaries remains restricted in the pathogenetic process. In general, viruses that are known to directly or
indirectly interact with endothelial cells may alter the permeability of vessels. Marburg and dengue viruses may
induce a reorganization of endothelial junctions with formation of intercellular gaps by misusing infected
monocytes/macrophages (Feldmann et al., 1996). In monocytes, infected with dengue virus, adherence to
endothelial cells is sufficient to trigger the release of pro-inflammatory cytokines (O'Sullivan and Killen, 1994).
Andes virus and human immunodeficiency virus can induce activation of DCs leading to secretion of
gelatinolytic matrix metalloproteases 2 and 9 that modulate the vascular permeability for their trafficking
(Marsac et al., 2011 and Ruggiero et al., 2011). Cytomegalovirus activates endothelial cells to express cellular
glycoproteins, such as selectins, on their membranes resulting in the increase of adherence of polymorphonuclear
leucocytes to the endothelial layer (Span et al., 1989), a slow rolling of leukocytes along vessel walls (Wagner
and Frenette, 2008) and an activation of integrins on the leukocyte surface which leads to increased binding on
endothelial cells (Alon and Ley, 2008). The interaction of EAV with endothelial cells appears to result in an
alteration of the vessel homeostasis as shown by the congestion and edema which were consistent findings in all

General discussion
100
the infected ponies. However, hemorrhages, thrombi or infarcts were never found. Therefore, EAV 08P178
seems to provoke only minor alterations in the endothelial cell layer and in the capillaries. However, various
strains of EAV have been shown to produce a fully productive, lytic cycle of replication in the diploid equine
endothelial cell line (Zhang, 2005).
Possibly as a consequence of the extravasation of infected cells, EAV reaches the internal organs where
secondary infections will start. The in vivo study showed that, already at 3dpi, the EAV infection started to be
generalized. At 7dpi, all internal organs in both euthanatized animals were positive, while at 14dpi, the infection
was controlled by the increased immunity. Only the nasopharyngeal tonsils and part of the lungs still contained
virus in one of the two animals, and the proximal septum, the nasopharynx, the liver, and the colon in the second
pony. At 28dpi, only tonsils and trachea were still positive. In the selected tissues with high virus titers, a double
IF revealed that approximately 50% of the infected cells belonged to the mononuclear leukocyte population and
around 45% to the endothelial cells of small capillaries. Histologically, the most prominent features observed in
all collected organs were the intraepithelial, subepithelial, perivascular and peribronchiolar infiltration of
lymphocytes and plasma cells, sporadic edema and congestion (data not shown). However, in contrast to the
North American strain findings, infarcts, hemorrhages or necrosis of vessel walls were not observed. Several
differences in lesions were apparently found in our study when compared to what has been described in the EAV
literature on North American strains. Experimental infection with the velogenic EAV Bucyrus not only resulted
in an extensive lymphocyte and plasma cell infiltration but also in panvasculitis with edema, thrombosis,
infarcts, hemorrhages, and necrosis. Thrombosis, infarcts, hemorrhages, and necrosis are a reflection of vascular
damage (Del Piero, 2000). In particular, Crawford and Henson (1973) described degeneration of the blood
vessels due to endothelial cell damage followed by leukocyte infiltration of the tunica intima. This apparently led
to fragmentation of the internal elastic-membrane (a condensation of elastic fibers separating the tunica intima
from the tunica media) allowing infectious virus to reach the tunica media of the leaking vessels. Infection of the
medial cells and subsequent active influx of neutrophils lead to a necrotizing process. The same authors reported
that the concomitant vascular hyperpermeability was responsible for the inflammation of the tunica media and
adventitia of the smaller blood vessels (Crawford and Henson,1973 and Estes and Cheville, 1970). Sustained
presence of edema, congestion and infiltration of lymphocytes and plasma cells induced by EAV 08P178 were
similar to those described for North American strains (Neu et al., 1992; Doll et al., 1968 and Jones et al., 1957 ).
The difference between the microscopic lesions associated to EAV 08P178 and the ones resulting from North
American strain infections may be due to a different cellular tropism of different EAV strains. In conclusion,
EAV 08P178 is able to alter the permeability of the blood vessels giving edema, but it does not create
intercellular gaps with consequent hemorrhages as highly virulent EAV strains do. Typical signs of
inflammation and increased vascular permeability have been reported as a consequence of lymphocyte,
neutrophil and macrophage infiltration in perivascular and interstitial spaces, leading to edema (Stokes et al.,
2011). Typical lesions induced by viruses with a high tropism for endothelial cells, consist of vasculitis,

General discussion _
101
hemorrhages, thrombosis, hypoxia and secondary ischemic degeneration (Borcher et al., 2010). It is thus
plausible that, with the European EAV strain, disease and lesions are induced more by inflammatory processes
rather than by direct virus induced degeneration of vascular walls. Different EAV strains vary in both their
cytopathogenicity and their ability to induce pro-inflammatory cytokines in endothelial cells in vitro (Timoney
and McCollum, 1993). Moore et al. (2003) have found that EAV infection of macrophages resulted in cell
activation with increased transcription of genes encoding pro-inflammatory mediators, including interleukin
(IL)-1b, IL-6, IL-8 and tumor necrosis factor (TNF)-α. Furthermore, Balasuriya and MacLachlan (2004) reported
that virulent and avirulent strains of EAV induce different quantities of TNF-α and other pro-inflammatory
cytokines (IL-1b, IL-6, IL-8) which are critical in determining the outcome of the infection.
The respiratory tract and its associated lymphoid tissues, but also the intestines (at 3, 7 and 14dpi) and the
kidneys (at 7dpi) were among the organs containing the highest viral titers. Ponies infected with EAV 08P178
did not present colic and feces did not change in consistency. However, with the velogenic EAV Bucyrus, it is
frequently reported that animals develop signs of colic, followed by diarrhea and, at necropsy, infarcts of the
intestinal walls are found which can explain the colic episodes. Although EAV 08P178 did not provoke infarcts
nor colic, the high virus titers and prolonged virus presence in the intestines indicate that this organ is also one of
the main internal target organs of EAV. At later time points in the course of infection (14 and 28dpi), virus titers
dropped drastically in all tissues (except the tonsils), while lesions continued to persist (data non shown). It is
possible that those late histologic changes, characterized by massive infiltration of lymphocytes and plasma
cells, may be still due to the immunological response rather than to a direct cytocidal viral effect.
Specimens of internal organs investigated with double IF, revealed that 58 and 63% of EAV-positive cells were
identified as mononuclear leukocytes and 30 and 35% as endothelial cells at 3 and 7dpi, respectively. Taken
together, we have identified around 90% of EAV-positive cells. Parenchymatous organs such as lungs, liver,
spleen, and kidneys are surrounded by a capsula, connective tissue rich of vessels and stromal cells such as
immune cells. On the other side the parenchyma comprises the functional parts of an organ (e.g. alveolar tissue,
hepatocyte, white and red pulp, and nephron). Thus EAV tropism for leukocytes and endothelial cells may
explain why the superficial regions which include the capsula of parenchymatous organs, had generally higher
virus titers than the deep regions (parenchyma). In particular, on a total of 29 EAV-positive specimens belonging
to parenchymatous organs, 20 had a higher virus titer in the capsula compared to the parenchyma. These findings
were in line with the publication of Crawford and Henson (1973) which reported that “organs with large
numbers of tissue macrophages and blood vessels showed the most abundant fluorescence. These included the
omentum, mesentery, lymph nodes and the capsule and the connective tissue of most parenchymatous organs”.
In conclusion, our results suggest that (i) EAV 08P178 infection does not involve large vessels but rather occurs
in a limited number of endothelial cells in terminal capillaries probably via adherence of infected blood
mononuclear cells and cell to cell virus transfer; (ii) EAV infection in these capillaries alters the vascular

General discussion
102
permeability, without creating intercellular gaps, leading to leakage of solutes but not to vasculitis and
hemorrhages; (iii) lesions caused by the EAV 08P178 may be a consequence of cell lysis in the early phase of
infection and immunological cytokines in the later stages. In addition, EAV 08P178 appears to have a main
tropism for large intestines and for stromal regions within the parenchymatous organs.
Usefulness of the equine polarized mucosal explant system
By the use of polarized mucosal explants, the access of EAV was restricted to the apical side of the epithelium.
This model allowed us to compare the in vivo results obtained upon inoculation of EAV in ponies with the ex
vivo ones and demonstrated that the results obtained with the ex vivo system are valuable. The number and the
identification of EAV-positive cells obtained in the agarose embedded explants were in agreement with those
obtained in vivo. Consistently, in both ex vivo and in vivo systems, it was shown that EAV can spread through
individual cells and forms clusters of infected cells in the nasopharyngeal and nasal mucosae. The excellent
correlation between the ex vivo model and the in vivo experiment indicates that our respiratory mucosal explant
system is a valuable tool to provide novel information on the pathogenesis of EAV and possibly other respiratory
viruses, such as equine influenza, EHV-1 and 4.
Although other equine explant systems have been previously developed (Vandekerckhove et al., 2009), this is
the first time that a polarized equine URT mucosal system was established. Several advantages have already
been described for the non-polarized explant systems (Vandekerckhove, 2011). Experiments performed on ex
vivo materials allow on the one hand, to reduce the ethical and economic issues linked to in vivo studies and, on
the other hand, to preserve the three-dimensional architecture and the physiology of the in vivo situation. In
addition, the possibility to use the mucosa of the same animal for kinetic studies and for comparison of viral
strains, minimizes the inter-animal variations. Apart from the aforementioned advantages, the polarized explant
systems has the ability to restrict the access of the pathogens to the apical side of the epithelium. Following
natural infection, pathogens have to overcome the epithelial layer before invading the host. In a non-polarized
explant system, all sides of the mucosal explants become exposed to the virus and this barrier phenomenon is,
therefore, lost. Indeed, inoculation of non-polarized explants with EHV-1 (Vandekerckhove, 2011) and EAV
08P178 (own studies, not published) resulted in an extensive spread of the virus mainly at the bottom and lateral
sides of the mucosal explants. The use of polarized explants prevents this artifact and resembles most closely the
in vivo situation with regard to viral events upon mucosal contact. Since the pathogen is obliged to overcome the
BM in a polarized system and does not have the possibility to bypass this barrier through the open edges of the
mucosa, it represents a reliable tool to investigate the interactions between the pathogen and the BM and the
mechanisms used by the virus to cross this barrier. Although polarized mucosal explants are much more
physiological than non-polarized explants systems and in vitro cell cultures, they are still far away from the
animal. Therefore, it is important to keep in mind that current detailed ex vivo findings do not always translate to

General discussion _
103
the in vivo situation as axiomatic truth. The absence of immunological factors coming from the blood should be
considered in explants.
In conclusion, polarized mucosal explants constitute a very valuable tool to get better insights of the early phases
of the pathogenesis of respiratory viruses, such as EAV, allowing a reduction of in vivo experiments, minimizing
the inter-animal variations and guaranteeing an apical inoculation of pathogens.
General conclusions
EVA can be experimentally reproduced through oro-nasal inoculation of ponies with the European EAV 08P178
field strain. The signs of disease such as fever, ocular and nasal discharge, lymphoadenomegaly, and one case of
scrotal edema and pathological changes such as intraepithelial, subepithelial, perivascular and peribronchiolar
infiltration of lymphocytes and plasma cells with sporadic edema and congestion, associated with EAV 08P178
infection are rather typical but moderate. Using the classification system defined by McCollum and Timoney
(1998), EAV 08P178 can be classified as moderately virulent as based on the presence of pyrexia for 3 or more
days, presence of nasal or ocular discharge and depression.
EAV uses the URT tract as portal of entry. In particular, migrating CD172a+
and CD3+ cells of the
nasopharyngeal and nasal mucosae may be used by EAV as Trojan horse to breach through the basement
membrane and migrate to the blood vessels. Through a cell-associated viraemia, EAV disseminates into
secondary organs. At early time points of infection, histological changes are likely due to virus-mediated injuries
in infected cells. Afterwards, with the appearance of EAV neutralizing antibodies and consequent drop of virus
titers, microscopic lesions could be caused by cytokines which increase vascular permeability.
EAV 08P178 does not seem to have a strong tropism for epithelial and endothelial cells of big vessels, while a
high tropism for CD172a+
and CD3+ mononuclear leukocytes was present in the portal of entry and for CD172a
+
and CD3+ mononuclear leukocytes and small capillaries in the internal organs.
In addition, it was demonstrated that polarized mucosal explants are an ideal tool to get better insights in the
early phases of the pathogenesis of respiratory viruses or viruses that have the respiratory tract as primary site of
replication prior to enter the body, such as EAV. This explant system allows a reduction of in vivo experiments
and guarantees an exclusive apical entry of pathogens, minimizing the inter-animal variations.

General discussion
104
REFERENCES
Alon R. and Ley K. (2008). Cells on the run: shear-regulated integrin activation in leukocyte rolling and
arrest on endothelial cells. Curr Opin Cell Biol, 20, 525.
Ayehunie S., Garcia-Zepeda E.A., Hoxie J.A., Horuk R., Kupper T.S., Luster A.D. and Ruprecht R.M. (1997). Human immunodeficiency virus-1 entry into purified dendritic cells through CC and CXC
chemokine coreceptors. Blood, 90, 1379-1386.
Autorino G., Cardeti G., Rosati R., Ferrari G., Vucano G., Ammeddeo D., McCollum W. and Timoney
P. (1994). Pathogenetic and virological properties of an Italian strain of equine arteritis virus in stallions
following intranasal challenge with infective raw semen. Proceedings of the seventh international
conference on equine infectious disease, edited by Nakajima H. and Plowright W. Publications R&W
publications, 335-336.
Balasuriya U.B. and MacLachlan N.J. (2004). The immune response to equine arteritis virus: A potential
lessons for other arteriviruses. Vet Immunol and Immunopat, 102, 107-129.
Balasuriya U.B., Snijder E.J., Heidner H.W., Zhang J., Zevenhoven-Dobbe J.C., Boone J.D.,
McCollum W.H., Timoney P.J. and MacLachlan N.J. (2007). Development and characterization of an
infectious cDNA clone of the virulent Bucyrus strain of equine arteritis virus. J Gen Virol, 88, 918-924.
Banchereau J. and Steinman R.M. (1998). Dendritic cells and the control of immunity. Nature, 392, 245-
252.
Bockman D.E. and Stevens W. (1977). Gut-associated lymphoepithelial tissue: bidirectional transport of
tracer by specialized epithelial cells associated with lymphoid follicles. J Reticuloendothel Soc, 21, 245-
254.
Borcher M., Roddy P., Pablo P., Jeffs B., Alberto O., Thomas S., Feldmann H., Strong J.E. and Villa
L. (2010). Factors associated with Marburg hemorrhagic fever: analysis of patient data from Uige,
Angola. J Infect Dis, 12, 1909-1918.
Bryans J.T., Doll E.R., Crowe M.E.W. and McCollum W.H. (1957). The blood picture and thermal
reaction in experimental viral arteritis of horses. Cornell Vet, 47, 42-52.
Castillo-Olivares J., Tearle J.P., Montesso F., Westcott D., Kydd J.H., Davis-Poynter N.J. and
Hannant D. (2003). Detection of equine arteritis virus (EAV)-specific cytotoxic CD8+ T lymphocyte
precursors from EAV-infected ponies. J Gen Virol, 84, 2745-2753.
Chen Y.C.P. and Morrow P.E. (1989). An analysis of alveolar macrophage mobility kinetics at dust
overloading of the lungs. Fundam Appl Toxicol, 13, 452-459.
Cole J.R., Hall R.F., Gosser H.S., Hendricks J.B., Pursell A.R., Senne D.A., Pearson J.E. and Gipson
C.A. (1986). Transmissibility and abortogenic effect of equine viral arteritis in mares. J Am Vet Med
Assoc, 189, 769-771.
Collins J.K., Kari S., Ralston S.L., Bennet D.G., Traub Dargatz J.L. and McKinnon A.O. (1987).
Equine viral arteritis in a veterinary teaching hospital. Prev Vet Med, 4, 389-397.
Crawford T.B. and Henson J.B. (1973). Immunofluorescent, light-microscopic and immunologic studies of
equine viral arteritis. In: Proc Int Conf Equine Infect Dis 3, 282-302.
Del Piero F. (2000). Equine viral arteritis. Vet Pathol, 37, 287-296.
Doll E.R., Bryans J.T. and Wilson J.C. (1968). Immunization against equine viral arteritis using modified
live virus propagated in cell cultures of rabbit kidney. Cornell Vet, 48, 497.
Estes P.C. and Cheville N.F. (1970). The ultrastructure of vascular lesions in equine viral arteritis. Am J
Pathol, 58, 235-253.
Feldmann H., Bugany H., Mahner F., Klenk H.D., Drenckhahn D. and Schnittler H.J. (1996).
Filovirus-induced endothelial leakage triggered by infected monocytes/macrophages. J Virol, 70, 2208-
2214.
Glorieux S., Favoreel H.W., Steukers L., Vandekerckhove A.P. and Nauwynck H.J. (2011). A trypsin-
like serine protease is involved in pseudorabies virus invasion through the basement membrane barrier of
porcine nasal respiratory mucosa. Vet Res, 42, 58.

General discussion _
105
Go Y.Y., Zhang J., Timoney P.J., Cook R.F., Horohov D.W. and Balasuriya U.B. (2010). Complex
interactions between the major and minor envelope proteins of equine arteritis virus determine its tropism
for equine CD3+ T Lymphocytes and CD14
+ Monocytes. J Virol, 84, 4898-4911.
Go Y.Y., Bailey E., Cook D., Coleman S., Timoney P.J., Chen K. and Balasuriya U.B. (2011). Genome-
wide association study identifies a common haplotype in horses associated with CD3+ T cell susceptibility
to equine arteritis virus infection. J Virol, 85, 13174-13184.
Gryspeerdt A., Vandekerckhove A., Garré B., Barbé F., Van de Walle G.R. and Nauwynck H.J.
(2010). Differences in replication kinetics and cell tropism between neurovirulent and non-neurovirulent
EHV-1 strains during the acute phase of infection in horses. Vet Mic, 142, 242-253.
Gryspeerdt A., Vandekerckhove A., Baghi H.B., Van de Walle G.R. and Nauwynck H.J. (2012).
Expression of late viral proteins is restricted in nasal mucosal leucocytes but not in epithelial cells during
early-stage equine herpes virus-1 infection. Vet J, 19, 3576-3578.
Hafeez A., Yunus Khan M. and Minhas L.A. (2009). Reticular epithelium and underlying high endothelial
venules of human nasopharyngeal and palatine tonsils: a comparative histological study. JSZMC, 3, 305-
309.
Hahn G., Jores R. and Mocarski E.S. (1998). Cytomegalovirus remains latent in a common precursor of
dendritic and myeloid cells. PNAS, 95, 3937-3942.
Holt P.G., Schon-Hegard M.A., Phillips M.J. and McMenamin P.G. (1989). Ia-positive dendritic cells
form a tightly meshed network within the human airway epithelium. Clin Exp Allergy, 19, 597-601.
Holt P.G., Schon-Hegrad M.A. and McMenamin P.G. (1990). Dendritic cells in the respiratory tract. Int
Rev Immunol, 6, 139-149.
Hu J., Pope M., Brown C., O’Doherty U. and Miller C.J. (1998). Immunophenotypic characterization of
SIV-infected dendritic cells in cervix vagina and draining lymph nodes of rhesus monkeys. Lab
Investigation, 78, 435-451.
Ingutti E., Mondino A., Khotuts A. and Jenkins M.K. (1997). In vivo detection of dendritic cell antigen
presentation to CD4+T cells. J Exp Med, 185, 2133-2141.
Jones T.C., Doll E.R. and Bryans J.T. (1957). The lesion of equine viral arteritis. Cornell Vet, 47, 52-68.
Keynan Y., Card C.M., McLaren P.J., Dawood M.R., Kasper K. and Fowke K.R. (2008). The role of
regulatory T cells in chronic and acute viral infections. Clin Infect Dis, 46, 1046-1052.
Kumar P. and Timoney P.J. (2001). Light and electron microscope studies on the nasopharynx and
nasopharyngeal tonsil of the horse. Anat Histol Embryol, 30, 77-84.
Lipscomb M.F. and Masten B.J. (2002). Dendritic Cells: Immune Regulators in Health and Disease.
Physiol Rev, 82, 97-130.
Luscinskas F.W., Ma S., Nusrat A., Parkos C.A. and Shaw S.K. (2002). The role of endothelial cell
lateral junctions during leukocyte trafficking. Immunol Rev, 186, 57-67.
MacLachlan N.J., Balasuriya U.B., Rossitto P.V., Hullinger P.A., Patton J.F. and Wilson W.D. (1996).
Fatal experimental equine arteritis virus infection of a pregnant mare: immunohistochemical staining of
viral antigens. J Vet Diagn Invest, 8, 367-374.
Marsac D., García S., Fournet A., Aguirre A., Pino K., Ferres M., Kalergis A.M., Lopez-Lastra M. and
Veas F. (2011). Infection of human monocyte-derived dendritic cells by ANDES Hantavirus enhances
pro-inflammatory state, the secretion of active MMP-9 and indirectly enhances endothelial permeability. J
Virol, 13, 223.
McCollum W.H. and Timoney P.J. (1984). The pathogenic qualities of the 1984 strain of equine arteritis
virus. In: Proc Grayson Found Int Conf Thoroughbred Breeders Organization, 34-47.
McCollum W.H. and Timoney P.J. (1998). Experimental observations on the virulence of isolates of
equine arteritis virus. In: Proc Int Conf Equine Infect Dis 8, 558-559.
Michel C.C. and Curry F.E. (1999). Microvascular permeability. Physiol Rev, 79, 703-761.
Minshall R.D., Tiruppathi C., Vogel S.M. and Malik A.B. (2002). Vesicle formation and trafficking in
endothelial cells and regulation of endothelial barrier function. Histochem Cell Biol, 117, 105-112.

General discussion
106
Moore B.D., Balasuriya U.B., Nurton J.P., McCollum W.H., Timoney P.J., Guthrie A.J. and
MacLachlan N.J. (2003). Differentiation of strains of equine arteritis virus of differing virulence to
horses by growth in equine endothelial cells. Am J Vet Res, 64, 779-784.
Neu S.M., Timoney P.J. and Lowry S.R. (1992). Changes in semen quality following experimental equine
arteritis virus infection in the stallion. Theriogenology, 37, 407-431.
O'Sullivan M.A. and Killen H.M. (1994). The differentiation state of monocytic cells affects their
susceptibility to infection and the effects of infection by dengue virus. J Gen Virol, 75, 2387-2392.
Richardson J., Bouchard T. and Ferguson C. (1976). Uptake and transport of exogenous proteins by
respiratory epithelium, Lab Invest, 35, 307-314.
Ruggiero E., Toschi E. and Federico M. (2011). The activity of matrix metalloproteinase-9 is part of the
mechanism of cell-to-cell HIV-1 endocytosis in dendritic cells. Curr Drug Discov Technol, 8, 112-118.
Schnittler H., Stroeher U., Afanasieva T. and Feldmann H. (2004). The role of endothelial cells in
filovirus hemorrhagic fever. In: Ebola and Marburg viruses: molecular and cellular biology. Feldmann H.
editors, Horizon Biosciences, Norfolk, United Kingdom, 279-304.
Schon-Hegrad M.A., McMenamin O.J. and Holt P.G. (1991). Studies on the density distribution and
surface phenotype of intraepithelial class II major histocompatibility complex antigen (la)-bearing
dendritic cells (DC) in the conducting airways. J Expe Med, 173, 1345-1356.
Singer C. and Jahn G. (1996). Human cytomegalovirus cell tropism and pathogenesis. Intervirology, 39,
302-319.
Smith J. (2003). Mycobacterium tuberculosis pathogenesis and molecular determinants of virulence. Clin
Microbiol Rev, 16, 463-496.
Span A.H., Mullers W., Miltenburg A.M. and Bruggeman C.A. (1991). Cytomegalovirus induced PMN
adherence in relation to an ELAM-1 antigen present on infected endothelial cell monolayers.
Immunology, 72, 355-360.
Steukers L., Glorieux S., Vandekerckhove A., Favoreel H. and Nauwynck H. (2012). Diverse microbial
interactions with the basement membrane barrier. Trends Microb, 20, 147-155.
Stokes K.L., Chi M.H., Sakamoto K., Newcomb D.C., Currier M.G., Huckabee M.M., Lee S.,
Goleniewska K., Pretto C., Williams J.V., Hotard A., Sherrill T.P., Peebles R.S. and Moore M.L. (2011). Differential pathogenesis of respiratory syncytial virus clinical isolates in BALB/c mice. J Virol,
85, 5782-5793.
Tenner-Racz K., Racz Myrvik P., Ockers Q. and Geister R. (1979). Uptake and transport of horseradish
peroxidase by lymphoepithelium of the bronchus associated lymphoid tissue in normal and Bacillus
Calmette-Guerin immunized and challenged rabbits. Lab Invest, 41, 106-115.
Timoney P. J. (1988). Equine viral arteritis: epidemiology and control. J Equine Vet Sci, 8, 54-59.
Timoney P.J. (2003). Equine viral Arteritis. In: Equine Respiratory Diseases, Lekeux P. (Ed.) Publisher:
International Veterinary Information Service, Ithaca, NewYork, USA.
Timoney P.J. and McCollum W.H. (1993). Equine viral arteritis in perspective in relation to international
trade. J Equine Vet Sci, 13, 50-52.
Vandekerckhove A. (2011). Equine nasal mucosal explants, a valuable tool to study early events of the
pathogenesis of equine herpesvirus infections. Ph.D. dissertation 2011.
Vandekerckhove A., Glorieux S., Van den Broeck W., Gryspeerdt A., Van der Meulen K. and
Nauwynck H.J. (2009). In vitro culture of equine respiratory mucosa explants. Vet J, 181, 280-287.
Vandekerckhove A., Glorieux S., Gryspeerdt A., Steukers L., Duchateau L., Osterrieder N., Van de
Walle G.R. and Nauwynck H.J. (2010). Replication kinetics of neurovirulent versus non-neurovirulent
equine herpesvirus type 1 strains in equine nasal mucosal explants. J Gen Virol, 91, 2019-2028.
Wagner D.D. and Frenette P.S. (2008). The vessel wall and its interactions. Blood, 111, 5271.
Zhang J. (2005) Permissiveness of selected cell lines to equine arteritis virus: establishment,
characterization, and significance of persistent infection in HELA cells. Ph.D. dissertation, University of
Kentucky Doctoral Dissertations. Paper 478.

SUMMARY-SAMENVATTING

Summary-Samenvatting _
107
SUMMARY-SAMENVATTING
Equine viral arteritis (EVA) is an important disease that can result in severe economic losses for the equine
industry. EVA is associated with abortion outbreaks, high mortality in neonatal animals and persistence in
stallions. In addition, equine arteritis virus (EAV) has recently increased in prevalence, possibly due to increased
transportation of infected horses and infectious semen. Despite the economic importance and the widespread
presence of EAV, little is known on its pathogenesis and the knowledge is based on very old studies performed
with a velogenic variant of EAV obtained by serially passaging of the 1953 Bucyrus field isolate in horses.
In Chapter I, an introduction was given on the history, the taxonomy, the virion morphology, and the replication
cycle of EAV. In addition, an overview was provided on the prevalence, the pathogenesis, the clinical signs, the
pathology, the histopathology, the immunity, and the diagnosis of EAV. Finally, the histology and the immunity
of the respiratory mucosae and the application of mucosal explants in research were briefly introduced.
In Chapter II, the specific aims of this thesis were given. The general aim of the thesis was to achieve a better
understanding of the pathogenesis of a wild-type European EAV strain (08P178) isolated from a Belgian natural
outbreak in 2008. The aims were divided into two major parts: (1) to investigate the virological, clinical and
pathological outcome of an experimental infection of 8 Shetland ponies. Samples collected at necropsy were
used to investigate the replication characteristics over time and to define the cellular tropism of EAV in primary
and secondary replication sites; (2) to investigate the early stages of EAV infection using the isolate 08P178
with emphasis on the cellular tropism at the level of the upper respiratory tract by the use of a polarized explant
system.
In Chapter III, the outcome of an infection with the EAV strain 08P178 was assessed in 8 seronegative Shetland
ponies through oro-nasal inoculation. Animals were monitored daily for clinical signs and, at different time
points (3, 7, 14 and 28 days post inoculation (dpi)), two ponies were euthanized. Two mock inoculated, control
ponies were sacrificed at the end of the experiment. Nasopharyngeal and ocular swabs and blood were collected
daily during the first 14 days and afterwards every other day until euthanasia. Bronchoalveolar cells were
harvested at -1, 3, 7, 10, 14, 21 and 28dpi. At euthanasia, 44 tissues were collected for virus titration and
immunofluorescence staining. All experimentally inoculated animals manifested, from 3 to 10dpi, symptoms
such as fever, swelling of lymph nodes, nasal and ocular discharge. Nasal swabs were positive from 2 to 9dpi
and ocular swabs were occasionally positive from 3 to 7dpi. Infected bronchoalveolar cells were evident from 3
to 7dpi and a strong cell-associated viraemia was detected from 3 to 10dpi. Hence, typical clinical signs and
pathological changes associated to EAV 08P178 were experimentally reproduced through oro-nasal inoculation
of ponies. In addition, our results showed that clinical signs and lesions, although rather typical, were relatively
mild. Further, the data showed that after a local replication at the level of the upper respiratory tract and the
lungs, EAV disseminated in the body via a cell-associated viraemia reaching secondary target organs.

Summary-Samenvatting
108
Accordingly, at 3dpi, high virus titers were found in the nasopharynx, tonsils, submandibular and bronchial
lymph nodes. At 7dpi, all collected samples, particularly lungs, intestines, kidneys and testicles manifested high
virus titers. At 14dpi, virus titers dropped drastically and, at 28dpi, only tonsils were highly positive.
Immunofluorescence staining revealed both individual EAV-infected cells and clusters of antigen-positive cells.
At the level of the nose, the majority of single EAV-infected cells were found below the basement membrane
and clusters of cells were noticed in the underlying connective tissue. As a result, the following hypothetical
model was forwarded: EAV may use individual cells, probably leukocytes, as Trojan horses to cross the
basement membrane at the level of the upper respiratory tract and then, enters the blood stream reaching the
internal organs.
In Chapter IV, organs collected at 3 and 7dpi with virus titers ≥105.0
TCID50/g were processed with double
immunofluorescence staining for the detection of the EAV N protein and, at the same time, for the identification
of the cell (specific markers for myeloid cells (CD172a+), T lymphocytes (CD3
+), B lymphocytes (IgM
+) or
endothelial cells (von Willebrand factor+)). At 3 and 7 dpi, 31 to 58% and 47 to 63% of the EAV-positive cells,
respectively, were mononuclear leukocytes. In all samples, with the exception of the lungs, the majority of EAV-
positive cells were CD172a+ followed by CD3
+; the IgM
+ cells represented a small percentage. No EAV-positive
endothelial cells were found out of 3.200 large blood vessels (≥ 3 endothelial cells) investigated. In terminal
capillaries (1-2 endothelial cells), however, 18 to 53% of the endothelial cells were EAV-positive. In conclusion,
it was demonstrated that (i) EAV 08P178 has a main tropism for CD172a+
and CD3+ mononuclear leukocytes;
(ii) EAV 08P178 does not infect endothelial cells of large blood vessels but does infect a considerable number of
endothelial cells lining terminal capillaries (iii) EAV 08P178 infection in these capillaries is most probably the
cause of an increased vascular permeability leading to leakage of fluid (edema-serous exudate) but not to severe
vasculitis and hemorrhages.
In Chapter V, a polarized mucosal explant system was designed and an exclusive apical entry of inoculated virus
was demonstrated by an assay based on streptavidin-labeled microspheres together with the absence of EAV-
antigens in RK13 cells seeded underneath the agarose layer in which inoculated explants were embedded and the
lack of viral antigen-positive cells at the cutting edges of the agarose embedded explants. Upon infection with
EAV 08P178, the number of EAV-positive cells increased over time with a mean of 68 in 8mm2 of nasal mucosa
and 225 in 8mm2 of nasopharyngeal mucosa at 72hpi, confirming the main role of the nasopharynx as first
replication site of EAV. Individual and clustered EAV-positive cells were located in the connective tissue below
the basement membrane, also at early time points, indicating that epithelial cells are not likely to be directly
involved in the early phases of EAV pathogenesis. Further, the majority of EAV-positive cells were CD172a+
cells followed by CD3+ T lymphocytes, whereas only a small percentage were IgM
+ B lymphocytes, suggesting
that EAV 08P178 has a main tropism in nasal and nasopharyngeal mucosae for myeloid cells and T lymphocytes
while no epithelial cells are involved. This study validates the usefulness of a polarized mucosal explant system

Summary-Samenvatting _
109
and shows that CD172a+ myeloid cells and CD3
+ T lymphocytes represent important EAV target cells in the
respiratory mucosa.
From this dissertation, it can be concluded that (1) EAV uses the upper respiratory tract as portal of entry. In
particular, the CD172a+
and CD3+ cells of the nasopharyngeal follicle associated epithelium are most likely used
by EAV as Trojan horses to breach through the basement membrane and migrate to the blood vessels. Through a
cell-associated viraemia, EAV disseminates into secondary organs. At early times of infection, histologic
changes are likely due to virus-mediated injuries in infected cells. Afterwards, with the appearance of EAV
neutralizing antibodies and consequent diminishment of virus replication, microscopic lesions may be mediated
by immunological cytokines which increase vascular permeability rather than by direct cytocidal effect of the
virus; (2) EAV 08P178 does not seem to have a strong tropism for epithelial cells and endothelial cells of big
vessels while a high tropism for CD172a+
and CD3+ mononuclear leukocytes and endothelial cells of small
capillaries was present both in the portal of entry and in the internal organs; (3) polarized mucosal explants are a
very valuable tool to get better insights in the early phases of the pathogenesis of respiratory viruses or viruses
that have the respiratory tract as primary site of replication prior to entering the body, such as EAV. This explant
system allows a reduction of in vivo experiments, minimizes the inter-animal variations and guarantees an apical
inoculation of pathogens.

Summary-Samenvatting
110
SAMENVATTING
Equine virale arteritis (EVA) is een belangrijke ziekte die kan leiden tot ernstige economische verliezen voor de
paardenindustrie. EVA wordt geassocieerd met abortusuitbraken, hoge mortaliteit bij pasgeboren dieren en
persistentie bij hengsten. Daarnaast is de prevalentie van equine arteritis virus (EAV) recent toegenomen,
mogelijk te wijten aan een toegenomen transport van geïnfecteerde paarden en infectieus sperma. Ondanks het
economisch belang en de wijdverspreide aanwezigheid van EAV, is er weinig bekend over de pathogenese en de
weinige beschikbare kennis is gebaseerd op zeer oude studies uitgevoerd met een velogene variant van EAV
verkregen door seriepassages van het 1953 Bucyrus veldisolaat bij paarden.
In Hoofdstuk I werd een inleiding gegeven over de geschiedenis, de taxonomie, de virion morfologie en de
replicatie cyclus van EAV. Daarnaast werd een overzicht gegeven van de prevalentie, de pathogenese, de
klinische symptomen, de pathologie, de histopathologie, de immuniteit en de diagnose van EVA. Tot slot werd
een inleiding gegeven over de histologie en immuniteit van de luchtweg mucosa en de toepassing van mucosa
explanten in het onderzoek.
In Hoofdstuk II werden de specifieke doelstellingen van dit proefschrift toegelicht. De algemene doelstelling van
het proefschrift was om een beter begrip te verwerven over de pathogenese van een wild-type Europese EAV
stam (08P178) geïsoleerd van een natuurlijke uitbraak in 2008. De specifieke doelstellingen werden opgesplitst
in twee grote onderdelen: (1) onderzoek naar de virologische, klinische en pathologische uitkomst van een
experimentele infectie van 8 Shetland pony’s met EAV 08P178. Stalen verzameld bij autopsie werden gebruikt
om de vermeerderingskenmerken in de loop van tijd te onderzoeken en om het cellulair tropisme van EAV in
primaire en secundaire vermeerderingsplaatsen te definiëren; (2) om de vroege stadia van infectie met het
Europese EAV isolaat 08P178 te onderzoeken met nadruk op het cellulair tropisme ter hoogte van de bovenste
luchtwegen door middel van een gepolariseerd explant systeem.
In Hoofdstuk III werd de uitkomst van een infectie met de EAV stam 08P178 geëvalueerd bij 8 seronegatieve
Shetland pony's. De dieren werden dagelijks gecontroleerd voor klinische symptomen en op verschillende
tijdstippen (3, 7, 14, en 28 dagen post inoculatie (dpi)) werden telkens twee pony's geëuthanaseerd. Twee mock-
geïnoculeerde controle pony's werden geëuthanaseerd op het einde van het experiment. Nasofaryngeale en
oculaire swabs en bloed werden dagelijks verzameld tijdens de eerste 14 dagen en daarna om de andere dag tot
euthanasie. Bronchoalveolaire cellen werden verzameld op -1, 3, 7, 10, 14, 21, en 28dpi. Na euthanasie werden
44 weefsels verzameld voor virustitratie en immunofluorescentie kleuring. Alle experimenteel geïnoculeerde
dieren vertoonden van 3 tot 10dpi symptomen zoals koorts, zwelling van de lymfeklieren, nasale en oculaire
uitvloei. Nasale swabs waren positief van 2 tot 9dpi en oculaire swabs waren soms positief van 3 tot 7dpi.
Geïnfecteerde bronchoalveolaire cellen waren aanwezig van 3 tot 7dpi en een sterke cel-geassocieerde viremie
werd gedetecteerd van 3 tot 10dpi. Typische symptomen en pathologische veranderingen geassocieerd met EAV

Summary-Samenvatting _
111
08P178 werden dus experimenteel gereproduceerd door middel van oro-nasale inoculatie van pony's. Daarnaast
toonden de resultaten aan dat symptomen en letsels, hoewel eerder typisch, relatief mild waren. Verder bleek uit
de gegevens dat na een lokale replicatie ter hoogte van de bovenste luchtwegen en de longen, EAV in het
lichaam verspreidde door middel van een cel-geassocieerde viremie en zo de secundaire doelwitorganen
bereikte. Overeenkomstig werden op 3dpi hoge virustiters aangetroffen in de nasofarynx, tonsillen,
submandibulaire en bronchiale lymfeklieren. Op 7dpi vertoonden alle verzamelde stalen, in het bijzonder de
longen, darmen, nieren en testikels hoge virustiters. Op 14dpi daalden de virustiters drastisch en op 28dpi waren
alleen de tonsillen nog sterk positief. Met de immunofluorescentie kleuring konden zowel individuele EAV-
geïnfecteerde cellen als clusters van antigeen-positieve cellen gedetecteerd worden. Ter hoogte van de neus,
werd de meerderheid van afzonderlijke, EAV-geïnfecteerde cellen gevonden onder de basaalmembraan en
clusters van cellen werden waargenomen in het onderliggende bindweefsel. Als gevolg hiervan werd het
volgende hypothetische model naar voren gebracht: EAV gebruikt mogelijk individuele cellen, waarschijnlijk
leukocyten, als Trojaans paard om de basaalmembraan te passeren ter hoogte van de bovenste luchtwegen, en
vervolgens de bloedsomloop binnen te dringen om zo de inwendige organen te bereiken.
In Hoofdstuk IV werden de organen, verzameld op 3 en 7dpi, met virustiters > 105 TCID50/g verwerkt voor
dubbele immunofluorescentie kleuring voor de detectie van het EAV N eiwit en, tegelijkertijd, voor de
identificatie van de cel (specifieke merkers voor myeloide cellen (CD172a+), T lymfocyten (CD3
+), B
lymfocyten (IgM+) of endotheliale cellen (von Willebrand factor
+)). Van de EAV-positieve cellen waren op 3 en
7dpi, 31 tot 58% en 47 tot 63%, respectievelijk, mononucleaire leukocyten. In alle stalen, met uitzondering van
de longen, waren de meerderheid van EAV-positieve cellen CD172a+ gevolgd door CD3
+; de IgM
+ cellen
vertegenwoordigden een klein percentage. Uit 3.200 onderzochte grote bloedvaten (> 3 endotheliale cellen)
werden geen EAV-positieve endotheliale cellen gevonden. Bij de terminale capillairen (1-2 endotheliale cellen)
echter waren 18 tot 53% van de endotheliale cellen positief voor EAV. Uit dit onderzoek kan worden
geconcludeerd dat (i) EAV 08P178 hoofdzakelijk een tropisme heeft voor CD172a+ en CD3
+ mononucleaire
leukocyten; (ii) EAV 08P178 geen endotheliale cellen van grote bloedvaten infecteert maar wel een aanzienlijk
aantal endotheliale cellen infecteert die de terminale capillairen aflijnen (iii) EAV 08P178 infectie in deze
capillairen hoogstwaarschijnlijk de oorzaak is van een verhoogde vasculaire permeabiliteit die leidt tot het
lekken van vocht (oedeem-sereus exsudaat) maar niet tot ernstige vasculitis en bloedingen.
In Hoofdstuk V werd een gepolariseerd mucosaal explant systeem op punt gesteld. Een uitsluitend apicale
intrede van geïnoculeerd virus werd aangetoond door een assay gebaseerd op streptavidine-gemerkte
microsferen, samen met het ontbreken van EAV-antigenen in RK13 cellen geplant onder de agarose laag waarin
geïnoculeerde explanten werden ingebed, en het ontbreken van viraal antigeen-positieve cellen aan de snijranden
van de agarose ingebedde explanten. Na infectie met EAV 08P178 steeg het aantal EAV-positieve cellen met de

Summary-Samenvatting
112
tijd met een gemiddelde van 68 in 8mm2 nasale mucosa en 225 in 8mm
2 nasofaryngeale mucosa op 72hpi, wat
de belangrijke rol van de nasofarynx als primaire replicatie plaats van EAV bevestigt. Geclusterde en individuele
EAV-positieve cellen bevonden zich in het bindweefsel onder de basaalmembraan, ook op vroege tijdstippen,
wat aangeeft dat epitheelcellen vermoedelijk niet direkt betrokken zijn bij de vroege fasen van EAV
pathogenese. Verder waren de meeste EAV-positieve cellen CD172a+ cellen gevolgd door CD3
+ T lymfocyten,
terwijl slechts een klein percentage IgM+ B lymfocyten waren. Dit suggereert dat de belangrijkste doelwitcellen
van EAV 08P178, in nasale en nasofaryngeale slijmvliezen, myeloide cellen en T lymfocyten zijn terwijl geen
epitheliale cellen betrokken zijn. Deze studie bevestigt de bruikbaarheid van een gepolariseerd mucosaal explant
systeem en toont aan dat CD172a+ myeloide cellen en CD3
+ T lymfocyten belangrijke EAV-doelwitcellen in de
luchtweg mucosa vertegenwoordigen.
Uit dit proefschrift kan geconcludeerd worden dat (1) EAV de bovenste luchtwegen gebruikt als toegangspoort.
Waarschijnlijk worden voornamelijk de CD172a+
en CD3+ cellen van de nasofaryngeale en nasale mucosae door
EAV als Trojaans paard gebruikt om de basaalmembraan te passeren en naar de bloedvaten te migreren. Door
middel van een cel-geassocieerde viremie spreidt EAV naar de secundaire organen. Op een vroeg tijdstip na
infectie zijn de histologische veranderingen waarschijnlijk te wijten aan virus-gemedieerde letsels in
geïnfecteerde cellen. Daarna, samen met het verschijnen van EAV neutralizerende antilichamen en
daaropvolgende vermindering van de virustiter, kunnen microscopische laesies mogelijk worden gemedieerd
door immunologische cytokines die de vasculaire permeabiliteit verhogen, eerder dan door een direct cytocidaal
effect van het virus; (2) EAV 08P178 lijkt geen sterk tropisme te hebben voor epitheliale cellen en endotheliale
cellen van grote bloedvaten, terwijl er een sterk tropisme was voor CD172a+
en CD3+ mononucleaire leukocyten
en endotheelcellen van kleine capillairen, zowel in de toegangspoort als in de inwendige organen; (3)
gepolariseerde mucosale explanten zijn een zeer geschikt hulpmiddel om een beter inzicht te krijgen in de vroege
fasen van de pathogenese van respiratoire virussen of virussen die de luchtwegen als primaire replicatie plaats
hebben voordat ze het lichaam binnendringen, zoals EAV. Dit explant systeem maakt een vermindering van in
vivo experimenten mogelijk, en vermindert tegelijk de variaties tussen dieren en tussen experimenten, en
garandeert ook een uitsluitend apicale intrede van ziekteverwekkers.


CURRICULUM VITAE

Curriculum Vitae _
113
Personal Information
Name and surname: Sabrina Vairo
Date and place of birth: January 14th, 1980 Moncalieri, (TO)
Citizenship: Italian
Languages: Italian (native language)
Spanish (native language)
English (2007) (First certificate passed with B)
French (good knowledge)
Dutch (basic knowledge)
Education:
Dates Degree Institution
1993-1998 High school graduate Scientific Lycée, Turin, Italy.
(57/60)
1998–2003 Dottore in Medicina Veterinaria University of Turin, Italy.
(DMV) (grade: 110/110 Cum Laude)
2008-today PhD in equine virology Ghent University, Belgium.
Bologna University, Italy.
Pregraduate Experience
Sept 2002- July 2003 Undergraduate veterinary student- Part time large animal scholarship: CISRA,
University of Turin, School of Veterinary Medicine, Turin, Italy.
Supervisor: DMV Mattone.
Sept 2002- July 2003 Undergraduate veterinary student- Equine night service, “Le Fontanette”
Equine farm , Vigone (TO), Italy.
Supervisor: DMV Rossi R.
Jan 2002-October 2003 Undergraduate veterinary student- Part time equine practice. S. Siro, Milan and
Vinovo (TO) racetracks, Italy.
Supervisor: DMV Vigliani A.
Sept 2001- July 2002 Undergraduate veterinary student- Part time genetic laboratory scholarship:
Department of genetics and animal production, University of Turin, School of
Veterinary Medicine, Turin, Italy.
Supervisor: DMV, PhD Sartore S.
Sept 2001- July 2002 Undergraduate veterinary student- Equine night service, “Il Grifone” Equine
farm Vigone (TO), Italy.
Supervisor: DMV Romano G.

Curriculum Vitae
114
2000-2001 Undergraduate veterinary student part-time small animal assistance Monticone
Small Animal Clinic, (Turin, Italy).
Supervisor: DMV Monticone R.
Postgraduate Experience
Aug 2011- today Doctoral fellowship in the Department of Veterinary Medical Science, Alma
Mater Studiorum, Bologna University, Italy.
Supervisor: Prof. Scagliarini A.
Jun 2009- Aug 2011 Doctoral fellowship in the Department of Virology, Immunology and
Parasitology, Ghent University, Belgium.
Supervisor: Prof. Nauwynck H.
Feb 2008- Jun 2009 Pre-doctoral fellowship in vivo drugs test for the pharmaceutical company
Barrier Therapeutics, Inc.
Supervisors: Prof. Nauwynck H. and Van Rossem K.
Breading season 2007 Equine reproduction Veterinary and embryo-transfer center Keros
(www.keros.be), Passendale, Belgium.
Supervisors: DMV, PhD ACVR, ECVT Daels P., DMV Vandale H. and DMV
Vandale G.
Jan 2006- Jan 2007 Large animal veterinarian.
Fellowship in Equine Clinical Studies, (equivalent to Internship) in all duties
of the Large Animal Clinic at the Veterinary Teaching Hospital of the
University of Turin, Grugliasco, (TO), Italy.
Supervisors: DMV, PhD Zarucco L. and DMV, PhD Bertuglia A.
Feb 2006- Oct 2006 Research fellow (Sinapsi Grant 9.000 € “Miglioramento delle risorse umane
nel settore della Ricerca e Sviluppo tecnologico” del Programma Operativo -
F.S.E. 2000-2006 - Obiettivo 3 – Periodo 2006).
Research project: Valutazione delle potenzialità di una miscela di aminoacidi
ed acido ialuronico nell’inibire il grado di artrite indotta da carragenina in un
modello sperimentale di artrite (Evaluation of the potentiality of aminoacid and
hyaluronic acid mix in inhibitioning the carragenine-arthritic stage in an
experimental arthritis mice model).
Supervisor: DMV, PhD Bertuglia A.
July 2004- August 2005 Equine veterinarian.
Equine Surgery Preceptorship, Hagyard Davidson McGee Equine Medical
Institute, Lexington, Kentucky, USA.
Supervisor: DVM Spirito M.

Curriculum Vitae _
115
Employment
2008- today Ghent University, 9000 Belgium.
Breading season 2007 Equine reproduction Veterinary and embryo-transfer center Keros
(www.keros.be), Passendale, Belgium.
Supervisors: DMV, PhD ACVR, ECVT Daels P., DMV Vandale H. and DMV
Vandale G.
Jan 2007- May 2007 Veterinarian: Equine private practitioner.
Reproduction assistance in “Violette” Equine Farm, Lagnasco (CN), Italy.
January 2006 Relief Veterinarian.
Small Animal Practice Dr.ssa Barbero R., Turin, Italy.
July 2004- August 2005 Equine veterinarian.
Equine Surgery Preceptorship, Hagyard Davidson McGee Equine Medical
Institute, Lexington, Kentucky, USA.
Supervisor: Spirito M., DVM.
Oct 2003- June 2004 Veterinarian: Small animal practice (after-hours emergency service) “Val
Susa” Small Animal Clinic, Avigliana (TO), Italy.
Community Service
Oct 2006- 2007 Veterinarian: Civil service for LIPU (Lega Italiana Protezione Uccelli) (Italian
Association for Bird Protection, Italian Ministry for the Environment and
Territory.
Dec 2005- Feb 2006 Tourist hostess- XX Winter Olympic Games, Turin, Italy.
June 2006 Veterinarian, AUFP (Allievo ufficiale in ferma prefissata) Centro
Addestramento Militare Foligno, (Roma).
Completed military veterinary entrance test (rank 4/40).
Licensure: Italy and all countries of the European Union.
Membership in Professional and Scientific Societies:
2004 - 2008 Ordine dei Medici Veterinari della Provincia di Turin, Italy (Italian Veterinay Medical
Association).
2006- 2008 SIVE:Società Italiana Veterinari per Equini (Italian Association of Equine
Practitioners).
2006- 2008 Società Italiana Fecondazione Artificiale.
2006- 2008 LIPU:Lega Italiana Protezione Uccelli (Italian Association for the Protection of Birds).

Curriculum Vitae
116
Bibliography
Papers published
Vairo S (2003) Indagine clinica ed ecomorfometrica nella tendinite del flessore superficiale delle falangi in 82
cavalli da corsa (Clinical and ecomorphometric evaluation of superficial digital flexor tendon injuries in 82 race
horses). [Italian] [DMV Thesis] Universita' degli Studi di Turin, Facoltà di Medicina Veterinaria; Anno
Accademico 2002-03, Turin, Italy: pp.1-192.
Vigliani A, Boniperti E, Scudo E, Vairo S (2005) Statistical and ultrasonographical evaluation of superficial
digital flexor tendon injuries in 190 race horses. (Italian) Ippologia, 16 (1): 9-14.
S. Vairo, A.P. Vandekerckhove, L. Steukers, S. Glorieux, W. Van den Broeck, H.J. Nauwynck (2011). Clinical
and virological outcome of an infection with the Belgian equine arteritis virus strain 08P178. (english).
PUBLISHED IN VET MIC 2012; 333-344.
S. Vairo, V. Saey, C. Bombardi, R. Ducatelle (2013). The European strain 08P178 of equine arteritis virus
causes inflammation but not vasculitis in experimentally infected ponies. (english). SUBMITTED to COMP.
PATHOL.
S. Vairo, H. Favoreel, A. Scagliarini, H. Nauwynck. (2013). Identification of target cells of a European strain of
equine arteritis virus in experimentally infected ponies. (english). SUBMITTED to VET MIC.
S Vairo, W. Van den Broeck, H. Favoreel, A. Scagliarini, H. Nauwynck. (2013). Development and use of a
polarized equine upper respiratory tract mucosal explant system to study the early phase of pathogenesis of a
European strain of equine arteritis virus. (english). ACCEPTED IN VET RES.
Abstract
S. Vairo, A. Vandekerckhove, L. Steukers, S. Glorieux, W. Van den Broeck, H. Nauwynck (2011). Clinical,
pathological and virological outcome of an infection with the belgian equine arteritis virus strain 08P178. The
XIIth
International Nidovirus Symposium. Michigan.
S. Vairo, W. Van den Broeck, H. Favoreel, A. Scagliarini, H. Nauwynck (2012). Replication characteristics of
equine arteritis virus 08P178 in equine polarized upper respiratory tract mucosal explants. IX International
Conference on Equine Infectious Diseases. Lexington, Kentucky.
S. Vairo, W. Van den Broeck, H. Favoreel, A. Scagliarini, H. Nauwynck (2012). Replication characteristics of
equine arteritis virus 08P178 in equine polarized upper respiratory tract mucosal explants. Symposium of the
Belgian Society for Microbiology, 30 November, Brussels, Belgium.
Oral presentation
S. Vairo (2009). Equine viral arteritis: implication in horse industry. Nationaal Agrarisch Centrum.

Curriculum Vitae _
117
S. Vairo (2010). Equine viral arteritis: implication in horse industry. Nationaal Agrarisch Centrum.
Externships/ Courses
2008- 2012 Doctoraal school Ghent University, Belgium.
Sept 2008- Jan 2009 Laboratory animals course.
July 2006 BiostAT 2006 – XIII Biostatistic and medical statistic summer class International school
onstatistics biology and human sciences, “Asti university” (Turin), Italy.
May 2005 Large animal anesthesiology master, “The Ohio State University, College of Veterinary
Medicine, Ohio, USA.
Continuing Education
2012 Symposium of the Belgian Society for Microbiology.
2012 Clinical studies: study design, implementation and reporting.
2011 XIII international nidoviridae symposium (participated with a poster which gained a prize of
500 dollars).
2010 Professional English writing skills.
2009 Summer course mucosal immunity.
2008 Professional English communication skills.
Jan 2007 XII SIVE Congress, Jan 26th-28
th, 2007, Meeting Centergross, Bologna, Italy.
Jan 2006 XI SIVE Congress, Jan 26th-28
th, 2006, Meeting Centergross, Bologna, Italy.
July 2003 XII Annual Scientific Meeting European College of Veterinary Surgeons, July 11 th-
13th, 2003,
Moat House Hotel, Glasgow, Scotland, GB.

ACKNOWLEDGEMENTS

Acknowledgements
118
Vorrei essere un poeta per poter descrivere in poche linee il mio affetto e gratitudine a tutti coloro che han costellato
la via del mio Ph.D.
The completion of my doctorate would not have been possible without the support, patience and guidance of many
people that I have met and worked with during my graduate studies. It is a pleasure to convey my gratitude to
everyone in my humble acknowledgment.
Prof. H. Nauwynck, I would like to thank you for opening the doors of your lab for me and for the brain-storms we
had in these 4 years. Prof, A. Scagliarini, grazie per l’aiuto scientifico e pratico con cui mi hai sempre sostenuto e per
aver creduto in me quando nemmeno più io ne ero capace. In Belgio son convinti che un dottorando debba ringraziare
solo per l’aspetto scientifico, tu sei la dimostrazione del contrario: pur non conoscendo molto il “mio virus” né le
tecniche da me utilizzate, la tua umanità ed il tuo supporto morale si son dimostrati essenziali per il completamento
del mio dottorato…Uno dei ricordi più vividi che ho di te è quando mi portasti a visitare l’università…già allora il tuo
entusiasmo e la tua vitalità mi contagiarono. I owe my deepest gratitude to Prof M. Pensaert for your advice and
supervision. I am indebted for all your contributions of time, ideas and help. I sincerely appreciate your friendship. I
am heartily thankful to Prof. P. Timoney for your advice and support throughout my studies on equine arteritis virus,
for having believed in me and in my capacities, for having always push me to go forward and never lose my
enthusiasm. Your encouragement inspired and enriched my growth as a Ph.D. student and I reached the goal also
thanks to you. I truly appreciate your guidance as a scientist and our philosophical talk on the meaning of life. I hold
you in the highest regard. Prof. Ducatelle, I would like to thank you for the teaching on pathology and for the
guidance you gave me… for having always answered to my several questions with a smile and for never having
considered a question a stupid question…even when, maybe, it was…thank you for your continuous support,
guidance and inspiring suggestions during my research and for your time, effort, and patience during my academic
growth. Prof. C. Bombardi, che dire…grazie per tutto il supporto tecnico per definire protocolli e colorazioni, per
l’aiuto nelle revisioni del secondo articolo e per la stesura del quarto articolo….ma soprattutto grazie e ancora grazie
per la disponibilità con cui mi hai sempre accolto e per l’atmosfera goliardica che riesci sempre a ricreare intorno a te,
sei davvero una persona fantastica e ti auguro tutto il meglio sul fronte lavoro e su quello familiare. Prof. H. Favoreel,
I have benefited from your guidance, your immuno-fluorescence advises and fruitful ideas which helped me in
shaping up the research and writing. I am grateful for your helpful discussions and valuable input. Prof. W. Van den
Broeck, thank you for all the help in setting the agarose system and in the morphology matters. Prof. Dubovi thank
you for everything you have done concerning the IHC protocol and staining….it impressed me a lot all the help and
the time you have spent for a stranger you will probably never meet. Dr. Levente Szeredi, it was a pleasure to work
with you and your team in Budapest, I learned a lot from you and I really appreciate the care you took of me in my
free time. I will remember you with sympathy.
I would like to express my sincere gratitude to the other members of my Ph.D. dissertation committee: Prof. Deprez
and Prof. Claerebout for their time, interest, helpful comments and insightful questions during the doctoral
examination. I gratefully acknowledge my BOF funds that made my Ph.D. work possible and provided my personal
financial stipend.
Chantal, Lieve, Nele, Dries, Tim, Ytse, Bart, Zeger, Mieke, Ann, Marijke, Magda, Fernand, Chris, Lobke, Liliana,
and Christian, I am deeply in debt with all of you for the technical support and the practical help you gave me.
Without your help the accomplishment of my Ph.D. would not have been possible. In particular, my sincere gratitude
to Geertje, for all the times you came to work at 4 am, just to hold my horses, for all the weekend and the holidays
you spent at the university to help me with my in vivo experiment. Carine, you are the person I own the most in the
lab. You were always patient with me and always ready to explain, teach and, in general, help me in any matter I had
during my research. For me you are more than a technician, but a sincere friend and a careful mother (I still remember
when you brought me food at 10 pm because you knew I was still working ). Melanie, you were a great help in my
practical work and a wonderful person to talk with, it was interesting to learn your point of view on many aspects of
life. Gert, thank you very much for all your bureaucratic support and help, you teach me different way to exploit
BUBBA, I am really graceful for it . Dirk, me and my pc own you a lot…thank you very much for all your technical
support both for research and personal matters but thank you overall for your friendship.
Karl, I have plenty of memory with you…all the mess we created and then we fix when it was almost impossible to
do. Thank you for all the Chinese evenings, for the ride, for the laughs, the serious and the silly talks, for all the care
you gave to me and overall to Alicia who is still remember your hot chocolate. These few lines are far away to
express my gratitude, I will remember you as my Belgium brother and you know you can always count on me. I love
you a lot. Mireia, tu eres una mujer estupenda. Yo te respeto mucho y aprecio tu extraordinaria fuerza, determinación

Acknowledgements _
119
y ambición. Gracias por honrarme con tu amistad y todo lo que has hecho y continúa haciendo por mí. Dipi, my other
big, big friend and companion of this Belgian experience. I thank you for all the scientific advices, for the reading and
the correction of my articles but overall for your guidance on personal matters. I am sure one day you will become a
good Guru and you will help a lot of people. Sarah Costers, I still remember the first day I arrived in the lab…you
bought me the lunch…a big black sandwich…I really felt welcome and you warmed up my heart. Sure I have then
thousands of memories linked to you and to the amazing person you are. I really love you a lot. Saruska, my cherry on
the cake…what I can say to you….sure I will never do enough or say enough to show how much I own you…I will
never forget all your support during my personal problems and your sweet character. I wish you all the best in life.
Annebel, if I have to describe you…you are the archer who always scores….you were mostly silent but when you
were telling something then it was the right thing to say and in the right moment. I am happy you have found your
way and I hope you will have the strength to follow it. I have a lot of good memories linked to you and I hope you
will soon visit us in Italy. Kalina..you cannot even imagine how much I miss you in the lab….over all after 10pm….I
never found anybody else doing such a convincing striptease on the lab desks….I am still laughing for it and for all
our dirty talk . David and Debby, I had such a good time with both of you and I really felt something missing in the
lab when you left. I thank you for all the scientific support and also for all the dirty talks that you were always able to
bring in any kind of discussion. Annelies, you teached me a lot, you represent the opposite of me and seeing you I
always remember how I should have acted. You really surprised me with some of your decisions and I have the prove
that, sometimes, following your heart, even if it is difficult and against society, brings you happiness. I wish you all
the best with Lieven and Bas. Amy, I am proud to see how strong you have grown in these years…I was always sure
you were a winner and now I am happy you realize yourself… Lennert I thank you for your contribution in scientific
and practical matters, to be a good friend who was always willing to help and give the best suggestions in several
aspect of life. I thank you for your hospitality and for the concerning you had for me and Alicia. You will always be
welcome. Hossein, you were the perfect colleague for the alternative Ph.D., with you it was really nice to have brain
storms concerning “THE TOPIC” and the 6 articles we have wrote concerning it. Thank you for all the laughs and all
the adventures we had. I wish your driving skills will improve soon . Sjouke or better Choco, Choco-toff or Choco-
milk you are the perfect researcher, a real model for me…it is impressive you always know what you are doing and
why you are doing…I learned a lot from ! Sabine, I wish you all the best with Alejandro and you’re little Paulina. I
really thank you for the patience you had to listen to my problems and all the care you provided to Zarrillo…they
meant a lot for me. Marc, I thank you for being a good companion during the long nights of experiments and most
importantly for the dirty talks and the many laughs we had during these years. Mieke, I mean Attila, every day I am
realizing how much I was lucky to survive to your destructive power . I thank you for have being my desk mate and
for the carnival masks you helped me to create. I wish you a lot of success with your Ph.D. Kathlyn, je vous remercie
beaucoup pour votre aide. Grâce à vous, j'ai pu avoir un repos comfortable. Je vous remercie aussi pour les longues
conversations que nous avions et le peu repas ensemble. Je vous vraiment souhaite beaucoup de chance avec votre
doctorat et votre vie personnelle. Karen O., it was always nice to hear you singing, while working, walking,
thinking….even when having difficult time in research, you were always smiling and ready to encourage people.
Annick, I am really happy to see that you finally found your way and that you are happy. Our relationship was not
easy, still I appreciate you and your professionalism. Nicolaas, thank you very much for the company in the icy
meeting room during the long “feestdagen” spent working in the lab…with you there I was feeling less lonely. Thank
you also for the crazy ride to the airport…it was fun and it ended well. Filip, it was nice to have you as colleague in
the first phase of my Ph.D., I thank you for the funny meals we had together and for always answering to my
questions with “in fact….” . Ivan, thank you very much for all your help and your fruitful suggestion, you are an
extreme smart person. I wish you all the best. Ben and Lowiese, thank you very much for all the fun we had in
Chicago, I really have a lot of memories linked to both of you and to our trip. Dominique, it was nice to work with
you during my Guinea pigs experiments and you really have grown a lot as a researcher but also as person. If you
keep going in this way, I am sure you will become a fantastic scientist. Eveline, thank you for helping me in finding a
house in the very early phase of my Ph.D. It is amazing to see the strength you had in facing difficulties and I wish
you a lot of health for you and overall for your little angel. Pepe, I wish you all the best with your PhD. Gerlinde and
Hannah, you were the perfect example of how a woman can be a super researcher and in the meantime a super
mummy. You were an example and source of inspiration for me. I really admire you a lot. Hanne and David, it was an
honor to have known a couple as you, great researchers but even greater people. You were always ready to help me
with the practical (titration) matters and with the writing skills. Thank you. Leslie, I admire you and your strength. I
wish you to preserve your perseverance and determination and to apply it in every aspect of life. Mathias, Dries and
Joao, I am grateful for all the evenings spent together, for all the activities you were always busy to organize and for
the good time we had together. Miet, I am really happy you find a nice job! It was nice to have you as a colleague and

Acknowledgements
120
with you, every time we went out, it was an evening full of fun. Nick, the gym man of the lab, you are the perfect
example that men with muscles can still preserve some brain . I thank you for all the scientific help you were always
willing to give me at the beginning of my Ph.D. and for your contagious passion and enthusiasm for research.
Zhongfang, you arrived in the lab and you left very soon, still it was amazing knowing somebody like you. You have
the capacity to do not let life break you down, even in terrifying situation. I am really happy you were able to create a
new family and I wish you all the best with your new job and baby. Eva. Maria y Irene gracias por toda la diversión
que teníamos y todo lo que me has enseñado. Muy agradable conocerte y tener la posibilidad de pasar algún tiempo
con ti. Te deseo todo lo mejor. Elisa barcellonese, anche detta “c’ho la vena occlusa”, quante risate ci siamo fatte
anche dopo 14-18 ore non-stop di lavoro in laboratorio, trovavamo ancora la forza di ridere e scherzare. Di te ricordo
soprattutto il viaggio a Parigi e tutto il divertimento legato a quel weekend. Ti ammiro tanto e sono davvero onorata di
averti conosciuta. Ti auguro di trovare la tua strada ed il lavoro dei tuoi sogni, perché nessuno come te lo merita. Elisa
padovana, son contenta che le tue ricerche ti abbiano portato a Gent dandomi così la possibilità di conoscerti. La vita
non è davvero stata generosa con te ma, nonostante tutto, sei riuscita a laurearti e a finire un dottorato. Ti stimo
davvero e ti auguro con tutto il cuore di trovare un lavoro appagante, un buon compagno ma soprattutto pace interiore
e tanta serenità. Ti voglio bene. Shrimp group, thank you very much for your hospitality in the warm and smelly
basement office…without it I would have been sick every time I was coming to Belgium. Veronique, thank you very
much for your help in the histopathology field. It was nice to collaborate with you. I wish you all the best. Veerle,
Inge, Sebastiaan, Matthias D., Isaura, Iris, An S., Ann D., Els, Tù, Lang, Liping, Wenfeng, Yu, Charlie, Kevin,
Constantinos, Nina, Maria, Thary, Jochen, Korneel, Merijn, Wander, Angela, Annelike, Uladzimir, and Ilias, thank
you for all the nice moments and the more or less scientific discussions we had during the past years. It was an honor
being your colleague. A special thanks to all the cleaning lady group to have always clean every mess in the lab and
have make it a nice environment to work in.
Marc and all the Antares’ team, thank you very much for everything you have done for me and the very good care you
have always provided me. I will really miss you a lot. Thank you very much to my lawyer Karin, you helped me in
one of the most difficult period of my life and you always showed me humanity and sympathy. I hold you in the
highest consideration and I own you more than I can describe. You will be always welcome and I hope we will stay in
contact. Barsha, you were a fantastic roommate and the younger sister I always desired. I still remember our long talks
and the fun we had. You were my Belgian family and a second mother for Alicia. I really love you and I wish you a
wonderful life. Cristian, Maria and Valentine it was great to live with you and it was nice to have a family bondage
with you in this Belgium experience. Wasana, you were a really good friend and a valuable person. I own you a lot
and I still remember you with love and affection. I am sorry life has brought us so far away and, although we do not
hear each other since long, I hope you are fine. Thank you for everything you have done for me. Lina, I really estimate
you and I own you a lot. You were an example for me and often I regret to do not have listened to you and to your
advices. Bertin, pour moi tu es plus qu' un ami. Je t' aime comme un frère et je possède plus que je peux expliquer.
Vous m’ avez montré la mentalité africaine de vivre, la joie pour les petites choses et la valeur incommensurable d'un
sourire. Tu m' as toujours aidé pour les questions pratiques et tu as toujours écouté à mes problèmes et tu étais là pour
moi. Encore une fois, je ne suis pas un poète, et même pas mille mots peuvent vous expliquer ma profonde gratitude.
Frank, you are a fantastic person and a precious friend. I own you a lot and I sincerely love you. I hope you will be
able to create a family with Nang and finally find happiness and the purpose of your life. Philipe, tu mi salvavida
cuando me estaba cayendo. Me ayudaste a ponerme de pie y me has apoyado siempre. Gracias por todo lo que has
hecho por mí, te quiero. Pedro y Gabiella, muchas gracias para ser mis amigos a pesar de la distancia y el tiempo. Te
quiero mucho. A big thank to all the member of the S Marino restaurant team, working with you was funny and I
always had good food at the end of the evening . In particolare grazie a Tony per aver cercato di aiutarmi e per
essersi preso a cuore la mia situazione. Grazie Maria per avermi sempre fatto ridere e avermi svelato i segreti della
tecnica sandwich…anche se in Italia non sembra funzionare….vi ricordo sempre con tanta ilarità. OBSG, with you
and in particular thanks to Silvia, Elisa, Piotre and Eric my weekends were always full of joy and fun. I am honored to
have been your friend and to have had the possibility to grown with you. You teach me a lot, overall the importance
and the joy to help people unconditionally. Learning from you, now I am a better person and I like myself much more.
Ale (Vigliani) e Prof. Zarucco, tutto è cominciato da e grazie a voi. Non riuscirò mai ad esprimere tutta la gratitudine
che provo per voi. Siete un esempio per me e vi stimo infinitamente. Grazie, grazie e ancora grazie.
Prof. Gatta, grazie per avermi permesso di lavorare nel suo dipartimento e soprattutto grazie per l’umanità
dimostratami, il pensiero delle sue parole mi scalda ancora il cuore. Prof. Vincenti, un grazie per aver sempre
dimostrato interesse per il mio lavoro e la mia vita personale, le auguro serenità e un ambiente lavorativo rilassato e
costruttivo. Prof. Mara, prima di conoscere te avevo la profonda convinzione che il trucco e gli accessori andassero

Acknowledgements _
121
strettamente abbinati all’ outfit….grazie per i tuoi insegnamenti su moda e shopping . Prof. A. Grandis, a casa mia
la si chiamerebbe “l’Helton John di photoshop”…grazie ancora per avermi lasciato lavorare sul suo computer e per
avermi svelato i segreti per migliorare le immagini….tanto che addirittura Prof. Chiocchetti può sembrare bello (ipse dixit). Prof. L. Calzà e L. Giardino, grazie per avermi permesso di usare i vostri microscopi e le vostre strutture.
Laura, Federica e Andrea son contenta di avervi avuto come colleghi e vi ringrazio per le battute e gli scherzi che han
sempre tenuto alto il morale dell’ufficio. Vi auguro tutto il meglio per il vostro futuro. Valeria, Caterina Michela e
Giovanni spero di riuscire a interagire un po’ di più con voi quando l’ansia di questo dottorato sarà un ricordo. Un
grazie anche al team di anatomia, per avermi accolto nei vostri laboratori e avermi sempre aiutato con gentilezza e
disponibilità. In particolare un grazie a Davide e al suo intercalare preferito (meeerdà), sei uno dei pochi maschi che
conosco che accetti di buon grado la propria inutilità . Alessandro e Sandra mi avete accolto a braccia aperte come
collega, genitore ma soprattutto come amica. Vi devo davvero molto e son contenta di avervi incontrato non solo
professionalmente ma anche dal punto di vista umano. Con voi in dipartimento è sempre stato divertente
lavorare…persino al microscopio!!!!!!!!!! Sandra, grazie anche per le feste, le cene deliziose che mi hai preparato, le
chiacchierate frivole e i lavaggi di capo che, da buona amica, ti sei presa la briga di fare. Ti voglio bene e ti auguro di
realizzare tutte le tua aspettative e di riuscire a vivere serenamente. Nicola, grazie per esserti preso a cuore il “mio
caso” personale e per il tempo e le energie che vi hai dedicato. Merci, sei stata il mio sfogo spagnolo e parlare con te
mi faceva sentire “ a casa”… muchas gracias y muy buena suerte para ti y tu familia. Grazie anche al resto del
Calzà/Giardino team: Chiara, Luisa Stella, Roberta, Luca e Antonio, la vostra presenza e allegria ha reso addirittura
piacevole un “double step-staining”, grazie per tutto l’aiuto tecnico, per le cioccolate e per il cameratismo. Auguro ad
ognuno di voi di trovare la propria strada e di essere felice. Gigi, il mio barista preferito, grazie ai tuoi caffè riuscivo
ad affrontare una nuova giornata lavorativa con brio e spirito…grazie per essere sempre stato di buon umore e aver
sempre scambiato battute bonarie con me. Meris, Lucia e Roberto grazie per tutto il supporto tecnico che mi avete
fornito durante i miei esperimenti ma soprattutto grazie per essere sempre stati empatici con me. Siete delle belle
persone e un team formidabile. Meris, grazie anche per tutte le attività extra-universitarie passate insieme: canto, feste,
la frutta e la verdura biologica, la gita sul trattore… Patrizia grazie per l’aiuto tecnico, per le colorazioni istologiche,
per i buon piatti che hai cucinato e per gli orecchini regalatemi alla festa della donna. Nadia, ti ringrazio per i mille
consigli sulle colorazioni istologiche, sul taglio e le inclusioni in paraffina ma anche per le risate e per le battute.
Franca, spero di non averti sconvolto troppo con le mie idee…anche se devo ammettere che molte volte le portavo
all’estremo solo per provocarti…Nico, cosa posso dire per dimostrarti il mio affetto e la mia gratitudine? Sei sempre
stato pronto ad aiutarmi, a prenderti cura di Alicia, a cucinare per noi e a portarci all’aeroporto o in stazione. Con te
abbiamo riso e giocato un sacco e ti abbiamo preso tanto in giro… Sei stato come un fratello per noi e se sono riuscita
a finire questo dottorato lo devo anche a te. Ti ricorderò sempre come il mio fratellone-tortellone. Lorenzo, la
sottoscritta e soprattutto il mio pc ti devono molto…grazie per i numerosi salvataggi in extremis di files
misteriosamente scomparsi e per avermi insegnato che, quando un computer si surriscalda, una buona alternativa è
lavorare fuori (quando naturalmente sta nevicando)…secondo me potresti far domanda alla NASA …ma ti ringrazio
anche per i caffè e le chiacchierate sul senso della vita e dei sogni, ti auguro di trovare la forza di vedere la vita con
ottimismo. Un grazie grande grande anche al resto del team informatico, per tutti i problemi tecnici risolti e i mitici
programmi installati.
Ringrazio tutte le maestre dell’asilo, il dott. Pietramala e tutti coloro che si son presi cura di Alicia quando i miei mille
impegni di lavoro non mi permettevano di stare con lei. Grazie a voi e alla fiducia con cui vi lasciavo Alicia, ho potuto
concentrarmi sul mio lavoro e finire questo dottorato. Mamma di Ludovica, di Gabriele e di Matteo, vi ricordo con
affetto e vi ringrazio di aver riempito i momenti bolognesi con feste, merende, ritrovi e gite …è bello vedervi così
vitali ed energetiche. Mamma Giorgia, tu mi hai insegnato tanto e sei sempre stata pronta ad ascoltarmi quando ne
avevo bisogno, mi hai dato fiducia e mi hai fatto di nuovo credere nel valore della famiglia. Son stata davvero
fortunata ad incontrare un’amica come te. Ringrazio te, Donato e Gaia per tutti i bei momenti passati insieme.
Beppe, Ghiacciolina, Joy, le numerose Ilarie di villa passeri, Sara, Ludovica, Francesca e Mariano è stato bello
condividere il giardino e i momenti estivi con voi, le cene, le pizze e le torte per celebrare i vostri esami passati son
stati momenti dolcissimi…con voi a corte non mi son mai sentita sola. Giovanna e babau Luna, grazie per le
passeggiate, il babysitteraggio e le cene, ti auguro di realizzare tutti i tuoi sogni e di avere un futuro brillante. Nena e
Alvise siete stati molto di più che semplici coinquilini per me e per Alicia…siete stati un grande supporto e ringrazio
ogni giorno per avervi incontrati. Vi auguro di tutto cuore un futuro insieme e tanta, tanta felicità. Teresa, sei una
coinquilina fantastica e la “zia” preferita di Alicia. Con te in casa, l’atmosfera è sempre serena. Grazie per tutto ciò
che fai per noi e grazie ad Andrea per tutti i nuovi cartoni che porta sempre ad Alicia.
La palestra ed in particolare Gianluca, Daniele, Carmine e Astrid, siete un team formidabile. Lavorare con voi e per
voi mi ha fatto crescere come insegnante e anche come persona. Vi ringrazio per tutti i sorrisi donatimi e per avermi

Acknowledgements
122
subito accolto come “una di casa”. Un grazie “inturgidente” soprattutto a Gianluca per avermi sempre ascoltato e
dedicato tempo. Un pensiero di riconoscenza a tutti i miei allievi, italiani e fiamminghi, per avermi sempre trasmesso
grinta, tenacia e determinazione. E’ stato un onore allenarvi e vedervi crescere nella conoscenza del vostro corpo.
Grazie! In particolare, Alessia grazie per tutte le cure materne e fraterne che ti sei sempre assicurata di darmi, per i
compleanni celebrati con te, per le serate passate insieme e le uscite…ma soprattutto grazie per avermi fatto provare
l’ebrezza del lucidalabbra inturgidente….penso non lo dimenticherò facilmente .
Giovanna, anche se le nostre vite son sempre state piene e non abbiamo mai avuto molto tempo per frequentarci, so
che posso sempre contare su di te e mi son sempre sentita ascoltata e capita da te. Sei sempre stata un’ottima amica e
ti ricordo con tanto affetto e nostalgia. Esmeralda, amica d’infanzia e amica tuttora, sei una delle persone che mi
conosce meglio e sei sempre stata saggia nel consigliarmi e guidarmi. Ti voglio bene e mi sei mancata tanto in questo
ultimo periodo. Spero che le difficoltà e le amarezza che stai provando non riescano ad abbatterti perché una delle tue
grandi qualità è l’essere positiva. Carolina, grazie per essere stata il sostegno e l’ancora di salvezza nel mio lungo
isolamento a Castagneto, mi hai aiutato non solo nelle cose pratiche ma sei stata una saggia consigliera e una paziente
ascoltatrice. Spero la nostra amicizia possa continuare nonostante non ci si riesca a frequentare molto. Un grazie
anche a “Piccoli” per le deliziose cenette, il capodanno e le discussioni su politica e companatico. Mauro, sei una delle
persone più importanti della mia vita. Sei un amico fedele e, sebbene tu mi conosca dalla prima elementare, non mi
hai mai deluso né mai mi hai giudicato. Ti voglio davvero bene e mi manchi da morire. Ti auguro tutta la felicità del
mondo e spero che il Brasile ti abbia accolto come un figlio. Spero che presto tu ed Anna riusciate ad ampliare la
vostra famiglia. Un pensiero di gratitudine anche alla mamma di Mauro, per avermi sempre accolto come una figlia.
Marco, altro grande amico d’infanzia…se dovessi scrivere tutte le cose per cui ti sono grata occuperei pagine e
pagine. Ti ricordo sempre con affetto e un profumo di vacanze e di mare nelle narici. Sono fiera di te e di quello che
sei diventato.
Cugina Laura, fra tutti i miei parenti sei quella a cui mi sento più vicina. Ti stimo molto per tutto quello che riesci a
fare, per riuscire sempre a reinventarti, per riuscire a mandare avanti una famiglia e trovare sempre tempo da dedicare
alla zia. Sei una donna bionica e spesso mi sei stata d’esempio. Un pensiero a mio fratello Edoardo, perché è sempre
stato un esempio per me. Spero un giorno di poter comprendere ciò che si cela dietro al tuo ermetismo. Mi auguro che
la vita non ci faccia dimenticare di essere fratelli e non soffochi l’amore che ogni fratello deve sentire per l’altro
fratello. Ti voglio bene. Michela, ormai sei parte della nostra famiglia da parecchi anni e ti sei sempre dimostrata
pronta ad aiutarmi, ti ringrazio per questo e spero tu possa essere felice. Alicia, sei un regalo ancora più meraviglioso
di quanto potessi mai immaginare e sognare, ogni tua carezza o bacio sono linfa vitale per me. Purtroppo la vita ci
ruberà la preziosa genuinità con cui ci scambiamo tanto amore ma vorrei che ricordassi sempre che sei quanto di più
bello potessi mai chiedere in 10.000 vite e che sono estremamente fiera di te. Grazie di esistere e di essere come sei.
Infine un grazie ai miei genitori che sebbene delusi e feriti da alcune mie scelte non hanno mai mancato di
supportarmi e volermi bene. Spero che un giorno Alicia nutrirà per me i sentimenti e la stima che io nutro per voi
perché allora vorrà dire che sarò stata un ottimo genitore. Vi voglio bene. Gattut, Zar e Zarrillo, grazie per il vostro
amore incondizionato, la vostra fedeltà e il vostro affetto, osservandovi e conoscendovi ho imparato a non aver paura
di amare.
.


Copyright © 2022 FDOKUMEN




















