
Venezuelan equine encephalomyelitis : the goat as a ... - CORE
107
Retrospective eses and Dissertations Iowa State University Capstones, eses and Dissertations 1-1-1973 Venezuelan equine encephalomyelitis : the goat as a sentinel for virus activity and as a serum donor for fluorescent antibody conjugates Gene Allan Erickson Iowa State University Follow this and additional works at: hps://lib.dr.iastate.edu/rtd is esis is brought to you for free and open access by the Iowa State University Capstones, eses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective eses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Recommended Citation Erickson, Gene Allan, "Venezuelan equine encephalomyelitis : the goat as a sentinel for virus activity and as a serum donor for fluorescent antibody conjugates" (1973). Retrospective eses and Dissertations. 18367. hps://lib.dr.iastate.edu/rtd/18367
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Venezuelan equine encephalomyelitis : the goat as a ... - CORE

Retrospective Theses and Dissertations Iowa State University Capstones, Theses andDissertations
1-1-1973
Venezuelan equine encephalomyelitis : the goat as asentinel for virus activity and as a serum donor forfluorescent antibody conjugatesGene Allan EricksonIowa State University
Follow this and additional works at: https://lib.dr.iastate.edu/rtd
This Thesis is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University DigitalRepository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University DigitalRepository. For more information, please contact [email protected].
Recommended CitationErickson, Gene Allan, "Venezuelan equine encephalomyelitis : the goat as a sentinel for virus activity and as a serum donor forfluorescent antibody conjugates" (1973). Retrospective Theses and Dissertations. 18367.https://lib.dr.iastate.edu/rtd/18367

Venezuelan equine encephalomyelitis:
The goat as a sentinel for virus
activity and as a serum donor for
fluorescent antibody conjugates
by
Gene Allan Erickson
A Thesis Submitted to the
Graduate Faculty in Partial Fulfi 1 lment of
The Requirements for the Degree of
MASTER OF SCIENCE
Department: Veterinary Microbiology and Preventive Medicine
Major: Veterinary Microbiology
Signatures have been redacted for privacy
lewa State University Ames, Iowa
1973
I I

.ISl-l 1'173 £..,. 44 G,b INTRODUCTION
REVIEW OF LITERATURE
MATERIALS AND METHODS
RESULTS
DISCUSS I ON
SUMMARY
APPENDIX
B I B LI OGRAPHY
ACKNOWLEDGEMENTS
i i
TABLE OF CONTENTS
Page
3
28
43
68
74
76
89
101

i i i
TABLE OF CONTENTS
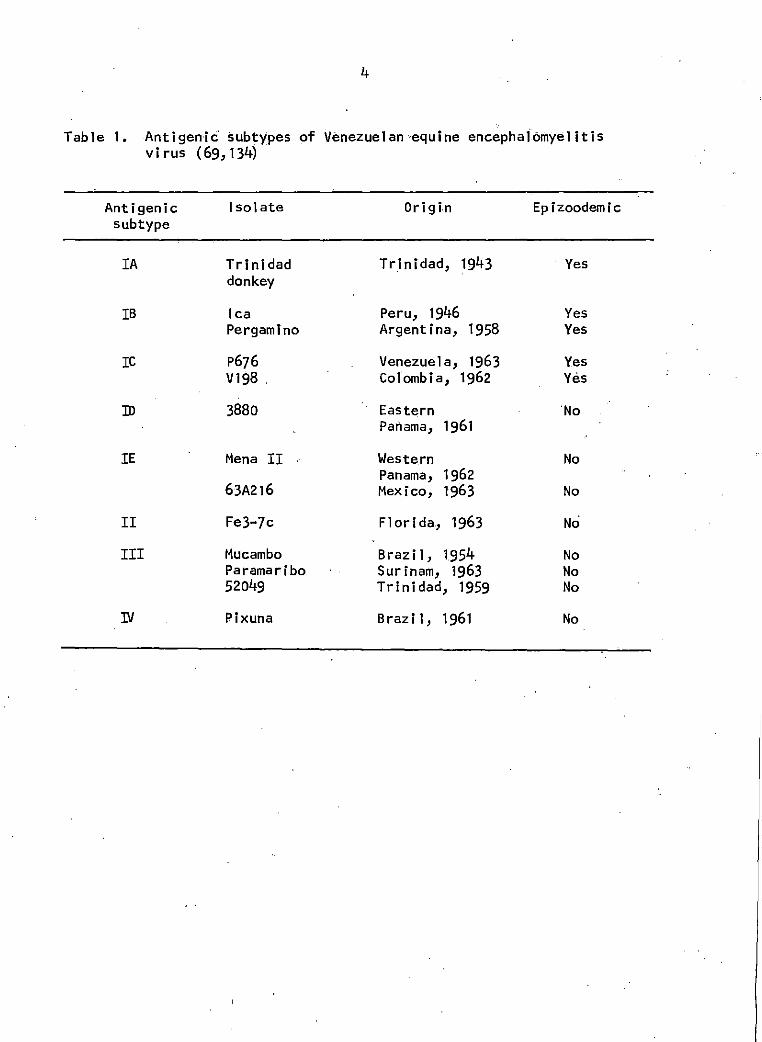
Table 1. Antigenic subtypes of Venezuelan equine encephalomyelitis virus
Table 2. Mosquito species from which epizoodemic VEE viruses have been isolated
Table 3. Mosquito infection and transmission rates for VEE virus IA
Page
4
8
10
Table 4. Mosquito infection thresholds for VEE virus IB 10
Table 5. Mammals considered to be involved in the 21 maintenance of endemic VEE virus activity
Table 6. Mosquito species from which endemic VEE virus have been isolated
23
Table 7. Susceptibility of different cell types to 44 GJ9-1BJ VEE virus
Table 8. Virus·titers in serum of goats inoculated with 53 GJ9~1BJ VEE virus
Table 9. Body temperature of goat number 159 inoculated 53 with IB VEE virus
Table 10. Virus detection by the fluorescent antibody test 67 (FAT) and by intracerebral inoculation of suck! ing mice

Figure 1.
Figure 2.
Figure 3.
Figure 4.
iv
LIST OF FIGURES
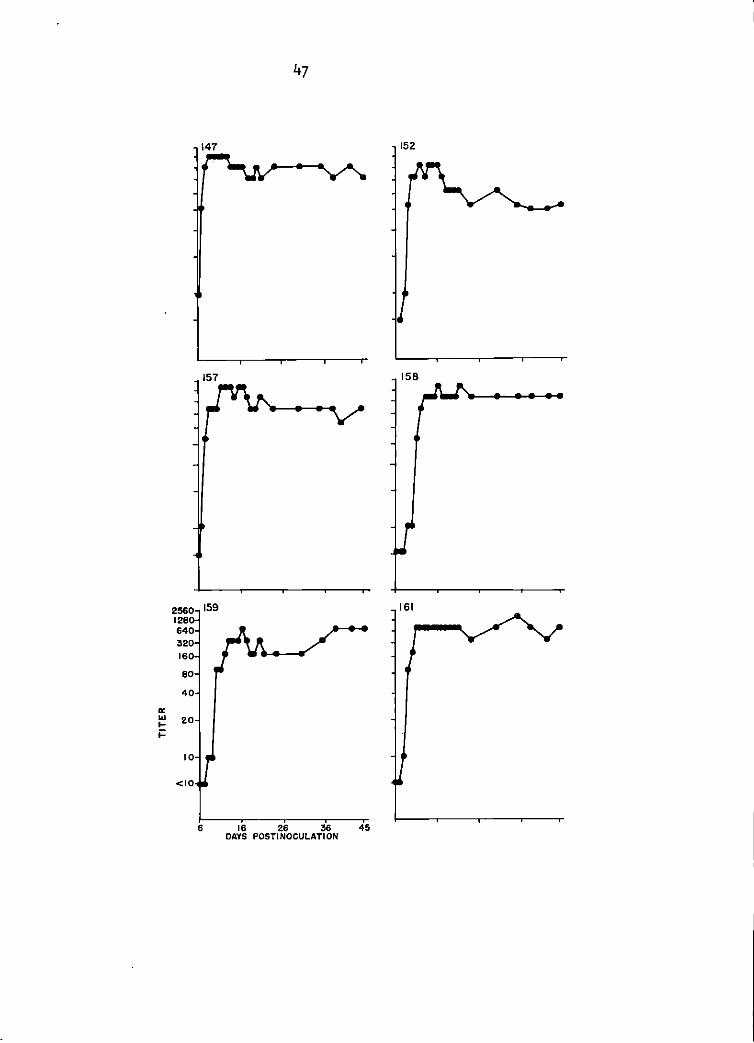
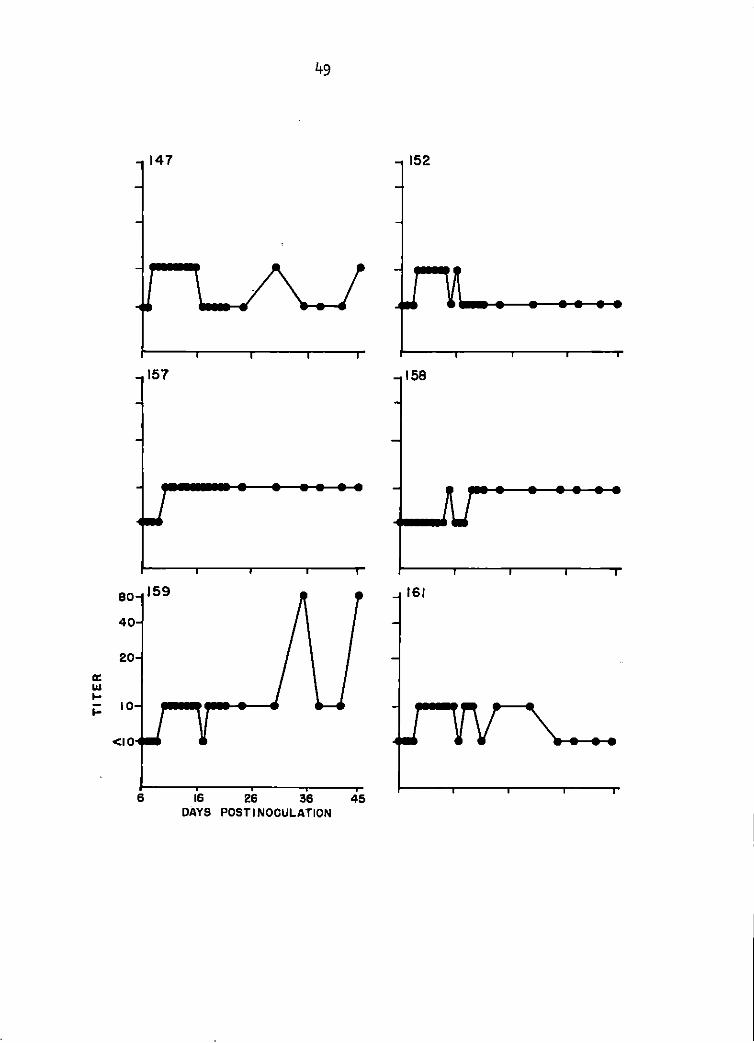
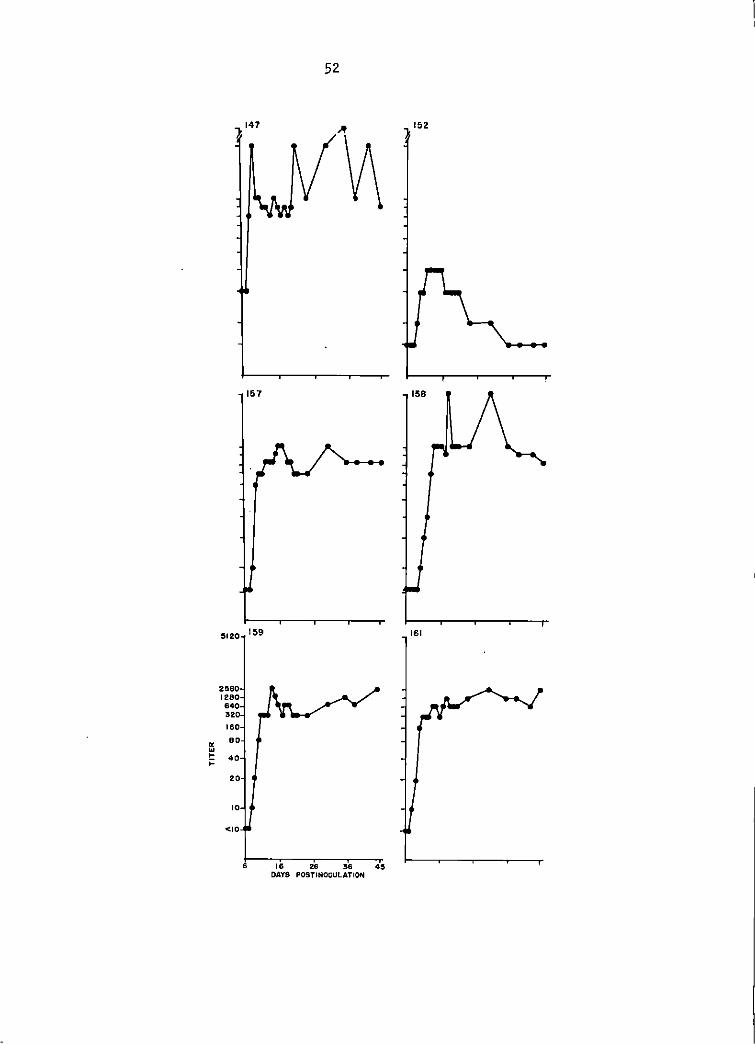
Sentinel goat study. TC-83 SN antibody titers of goats 147, 152, 157, 158, 159, and 161 between days 6 and 45 postinoculation. Titers 1 i sted were the highest serum <! i 1 ut ion that produced at least 90% plaque reduction. See Appendix for specific titers.
Sentinel goat study. GJ9-1BJ SN antibody titers of goats 147, 152, 157, 158, 159, and 161 between days 6 and 45 postinoculation. Titers listed were the highest serum dilution that produced at least 90"/o plaque reduction. See Appendix for speci~ic titers.
Sentinel goat stud)'.. Hi antibody titers of goats 147, 152, 157, 158, 159 and 161 between days 6 and 45 post inoculation. See Appendix for specific titers.
Absence of clinical signs in a domestic goat three days after infection with the GJ9-1BJ strain of Venezuelan equine encephalomyelitis (VEE) virus.
Figure 5. VEE conjugate applied to normal BHK-21 cell monolayer. Note absence of specific fluorescence. 125x
Page
47
49
52
56
56
Figure 6. VEE conjugate applied to GJ9-1BJ VEE-virus- 58 i.nfected BHK-21 ce 11 mono 1 ayer at 24 hours postinoculation. Note specific perlnuclear and cytoplasmic fluorescence. 125x
Figure 7. Normal conjugate applied to GJ9-1BJ VEE-virus- 58 infected BHK-21 cell monolayer at 24 hours postinoculation. Note absence of specific fluorescence. 125x
Figure 8. VEE conjugate applied to TC-83-VEE-virus- 60 infected BHK-21 cell monolayer at 24 hours postinoculation. Note specific, granular perinuclear and cytoplasmic fluorescence. 500x

v
Figure 9. VEE conjugate applied to GJ9-1BJ-,VEE-virus-infected BHK-21 cell monolayer at 24 hours postinoculation. Note specific, granular perinuclear fluorescence. 1188x
Figure 10. VEE conjugate applied to TC-83-VEE-virus-infected BHK-21 cell monolayer at 48 hours' postinoculation. Note specific cytoplasmic fluorescence of rounded up cells. 125x
Page
60
62
Figure 11. VEE conjugate applied to TC-83-VEE-virus- 62 infected BHK-21 cell monolayer at 48 hours postinoculation. Note specific, granular cytoplasmic fluorescence of rounded up cells. SOOx
Figure 12. VEE conjugate applied to western equine en-cephalomyelitis virus-infected BHK-21 cell, monolayer at 24 hours postlnoculatlon. Note absence of cross-staining. 125x
Figure 13. VEE conjugate applied to eastern equine en-cephalomyelitis virus-infected BHK-21 cell monolayer at 24 hours postinoculation. Note absence of cross-staining. 125x
64
64

INTRODUCTION
The insidious spread of Venezuelan equine encephalomyelitis virus
(VEE) from South America through Central America to the.United States has
aroused great concern within the horse industry and among agricultural
officials in the United States. Indeed the disease in the epizoodemic
form appears to have been eliminated from the continental borders of this
country but the mechanisms by which this virus can apparently reside
within a given region are unknown. The term epizoodemic as employed in
the text above refers to the concurrent presence of an epidemic and an
epizootic·due to a single disease agent (56,57). There is at present a
great confusion of terminology in the literature due to the remarkable
ability of the VEE virus complex to give rise to several forms of disease
among man, domestic animals, and fauna of the countryside. Therefore, in
an attempt to reduce this confusion, the term epizoodemic shall be em-
ployed in this study.
Due to the explosiveness of epizoodemic VEE it has become imperative
to have rapid diagnostic.techniques available to the virologist for con-
firmation of the disease agent in domestic livestock and in wildlife
populations. Such a technique is readily available in the form of the
fluorescent antibody test whf.ch can be used to obtain positive or negative.
results within a 48 hour period as compared to suckling mouse inoculation.
with complement fixation (CF) test confirmation which requires from 72
hours to one week for comp 1 et ion of diagnosis •.

-'
2
The invasion of the United States by VEE with subsequent eradication
in 1971 has emphasized the need for a natural sentinel to detect the
virus should it reenter the country in the future. Several animals have
been investl'gated as possible sentinels by other workers. These include
the canine (9), bovine, porcine (33), equine (25), laboratory (39,43,105,
107,108) and wild ungulate (67) species. Despite such intensive work, the
suitability of the goat, (Capra hircus), as a sentinel species has not
been investigated.
In hypothesizing the means by which epizoodemic strains of VEE run
rampant through the countryside, reference has occasionally been made to
the possibility of the goat acting as ,a silent amplifier of the virus
(57,104,110). ·Jt was the purpose of this. study, firstly, to investigate
the suitability of the goat as a sentinel for virus activity, and
secondly, to determine the sensitivity of the fluorescent antibody test
on cell cultures as compared· to suckling mouse inoculation with con-
firmatory CF test for identification of VEE virus from tissue specimens.

3
REVIEW OF LITERATURE
History
An encephalitic disease of equines which appeared in 1935 in the
river valleys of Huila, Tolima, Valle, and Bolivar in the Andes mountains
of western Colombia was tentative I y diagnosed as Barna' s disease or
European equine encephalitis. From there the disease spread to Magdalena,
Colombia in 1936 and later that year appeared in the Guajira peninsula of
Colombia and Venezuela ( 125). In 1938 Kubes. and Rios of the Venezuelan
Ministry of Agriculture and Animal Husbandry isolated a filterable v.irus
from the brain of a horse that had died with encephalitic signs (77).
Subsequent characterization of the agent was done by Kubes (76),
Kubes and R las ( 77) and by Beck and Wyckoff ( 6) in 1939 and 1944. The
virus was found to be of greater virulence for equidae than any of the
eastern or western equine encephalomyelitis viru.ses previously ·identified·
in South America. It also differed immunologically from these viruses.
Due to its origin the encephalitic agent was designated "Venezuelan equine
encephalomyel it is virus. 11 A uniform system of nomenclature was not
available for the complex of endemic (enzootic) and epizoodemic strains
of Venezuelan equine encephalomyelitis (VEE) virus that later arose
until Youn,g and Johnson (134) in 1969 published their system embracing
the antigenic variants of VEE (Table 1).
Since 1939, either epizoodemics or epidemics of VEE have been
described in Peru, Ecuador, Columbia, Venezuela, Trinidad, Costa Rica,
Nicaragua, Honduras, El Salvador, Guatemala, Mexico and the United States.
4
Table 1. Anti gen i c· subtypes of Venezuelan ·equine enceph a 1omye1 it is virus (69, 134)
Antigenic Isolate Origin Epizoodemic subtype
IA Trinidad Tr.inidad, 1943 Yes donkey
IB lea Peru, 1946 Yes Pergamlno Argentina, 1958 Yes
re p676 Venezuela, 1963 Yes V198 . Colombia, 1962 Yes
ID 3880 Eastern 'No Panama, 1961
IE Mena II Western No Panama, 1962
63A216 Mexico, 1963 No
II Fe3-7c Florida, 1963 No
III Mucambo Brazil, 1954 No Paramaribo Surinam, 1963 No 52049 Trinidad, 1959 No
IV Pixuna Brazil, 1961 No

5
Endemic foci have also been found to be widely distributed in the western
hemisphere including Brazil, Trinidad, Surinam, Colombia, Panama,
Nicaragua, Honduras, Guatemala, British Honduras, Mexico and Florida (57).
Current virus activity
In 1969 a severe epizoodemic of VEE arose in Guatemala and eventually
reached Texas in late June, 1971 (22,24,_37,39,42,56,75,114,124,13P). The
source of VEE virus for this and other epizoodemics remains an enigma.
Three possible alternatives for the introduction of the virulent IB
subtype of VEE virus into Guatemala have recently been proposed by
Franck and Johnson (42). They are (1) mutation of an endemic virus
subtype, (2) sudden emergence of a previously silent unrecognized virus,
and (3) introduction of the virus from another region. Introduction of
the IB subtype of VEE virus into Guatemala from Ecuador has been suggested
by several authors (37,42,86,114, 1~9).
Equine deaths were first reported along the Pacific coast of
Guatemala near El Salvador and shortly thereafter in northeastern
El Salvador. VEE then spread north and south from each focus. The
southward spread of the disease included Honduras (1969), Nicaragua
(1969), and Costa Rica (1970). Susceptible equines were vaccinated with
the attenuated TC-83 strain of VEE virus in Guatemala in an attempt to
halt the northward spread of epizoodemic VEE. However, in August, 1969,
epizoodemic VEE virus was isolated in the upper Grijalva River Valley
of Chiapas, Mexico near the Guatemalan border. Equine deaths continued
through the month of February and increased with the start of the rainy

6
season in June. From there the disease spread westward through the
mountain canyons to the state,of Oaxaca on the Pacific coast and was
confined to that region by vaccination until September when the disease
appeared just outside th'e vaccination zone in the state of Veracruz.
Vaccination was resumed only to have epizoodemic VEE occur again just
outside the barrier to the north.
Extension of the disease followed a similar pattern on the Pacific
coast as it progressively appeared beyond each new vaccination barrier.
Epizoodemic VEE had spread northward along the Pacific coast through the
state of Guerrero to the state of Michoacah by November, 1970.
Virus activity continued unreported through the end of 1970 and
into Apri 1 of 1971. Equine deaths were then confirmed· as VEE in Veracruz
and a new vaccination barrier was established to the no'rth. Despite al 1
vaccination barriers epizoodemic VEE continued its northward spread across
salt water lagoons of Mexico's gulf coast some 400 kilometers to Texas
( 125) •
First case of epizoodemic VEE in the United States
The first confirmed case of epizoodemic VEE .in the United States was
reported on June 23, 1971, near Three Rivers, Five Oak County, Texas,
approximately 320 kilometers northwest of Brownsville. The Texas
epizoodemic peaked in the equine population by mid-July (30 virus
isolations per day) and by the end of July had decreased to fewer than
5 viral isolates of epizoodemic VEE virus per day. A combination of
ultra-low-volume aerial insecticide application and vaccination (TC-83

7
virus) of all susceptible equines in Texas effectively brought the in-.
vasion of Texas' equine and human populations to an end by November 7,
1971 (75). A total of 1,620 equine deaths and an additional 2,000
clinical cases of epizoodemic VEE were recorded in Texas in 1971 (86).
Epizoodemic virus activity in Mexico in 1972
Confirmed (virus isolation) epizoodemic VEE virus activity recurred
in Mexico in the states of Durango and Sonora in 1972. Other noncon-
finned reports of VEE outbreaks came from the states of Nayarit, Guerrero,
and Morelos. Intensification of vaccination of susceptible equines with
the TC-83 strain of VEE virus in the state of Sonora in addition to that
done in 1971 appears to have prevented epizoodemic VEE from advancing
beyond Hermosillo, which is approximately 265 ·kilometers south of the
Arizona-Mexico border (22,23,24,25).
Arthropod vectors and their possible role in the spread of epizoodemic VEE
Epizoodemic strains of VEE have been isolated from at least 25
species of mosquitoes consisting of 7 genera and one subgenus (22,1151
125} (Table 2). Isolation of epizoodemic VEE virus from field-trapped
mosquitoes in itself is insufficient evidence to incriminate a particular·
species as a vector of the disease. Three additional factors must be
evaluated prior to the assignment of vector status (115). They are as
fol lows:
(1) Determination of the infection threshold and infection rate of the mosquito. Infection threshold may be defined as that level of host viremia sufficient to infect 1 to 5 percent of the mosquitoes feeding upon the virus source. Infection ra.te is the percent of mosquitoes that are viremic 14 to 21 days after

8
Table 2. Mosquito species from which epizoodemic VEE viruses have been isolated ( 115,117,123,12~
Mosquito species
Aedes aegypti Aedes angustivittatus Aedes scapularis Aedes serratus Aedes sollicitans Aedes taeniorhynchus
Aedes thelcter
Anopheles aquasalis Anopheles crucians Anopheles neomaculipennis Anopheles pseudopunctipennls. Anopheles punctimacula
Cul ex aikeni i Culex cor.niger Culex niqripalpus Culex guinguefasciatus Culex (Melanoconion) sp.
Culiseta inornata
Deinocerites pseudes
Mansonia indubitans Mansonia tltillans
Psorophora ciliata . Psorophora c i Ii pes
Psorophora confinn is Psorophora discolor
Country
Venezue 1 a Colombia Venezuela Venezuela Texas, U.S.A. Colombia, Venezuela, Costa Rica; Texas, U.S.A. Texas, U.S.A.
Venezuela Texas, U.S.A. Trinidad Texas, U.S.A. Colombia
Colombia Colombia Guatemala Colombia Guatemala; Texas, U.S.A.
Mexico
Costa Rica
Colombia Colombia, Guatemala, Trinidad
Texas, U.S.A • Guatemala Venezuela, Guatemala; Texas, U.S.A. Texas, U.S.A.

9
having fed upon a viremic host. Infection is determined by trituration of each mosquito with subsequent inoculation of one 2- to 4-day-old mouse per mosquito.
(2) Determination of transmission rate of the mosquito. This is obtained by allowing individuals one opportunity to infect a susceptible host .following an incubation period (14-21 days) after the original blood meal. The transmission rate is then calculated by determining the percentage of infected individuals that transmitted the disease agent. Infection is assayed as described ·in (1) after al lowing the arthropod to engorge.
(3) Determination of the extent of mosquito-host interaction. Field studies are employed to define this parameter of the possible vector as determined by (1) and (2). Areas of mosquito-host interaction usually considered are proximity of the host to breeding sites of the mosquito, blood meal identification, abundance of the potential vector species, attraction rate to various sentinel hosts, longevity, and flight range of the mosquito.
Some information on potential epizoodemic VEE virus vectors has been
presented (Tables 3 and 4), but much remains to be done. Simulium spp.
have also been implicated as biologic vectors of VEE virus. However, no
I aboratory data are avai I ab le to substantiate th is hypothesis (16,7D, 121).
The average host viremia needed to infect a suitable ~osquito vector
species with VEE virus is 5.D log1D suckling mouse intracerebral lethal
dose for 5D% of those inoculated per ml (SMICLD50) of viremic ·blood (14,
16) and an average VEE virus inoculum per mosquito bite has been stated 1 to be 3.0 log 10 SMICLD50 For establishment of infection and trans-
mission capability of a mosquito two major barriers must be overcome (17).
When a mosquito feeds upon a host the blood meal moves directly to the
glandular midgut for digestion. To retain the blood meal within the
1chamberlain, R. W., Communicable Disease Center, Atlanta, Georgia. Personal communication. October, 1971.
10
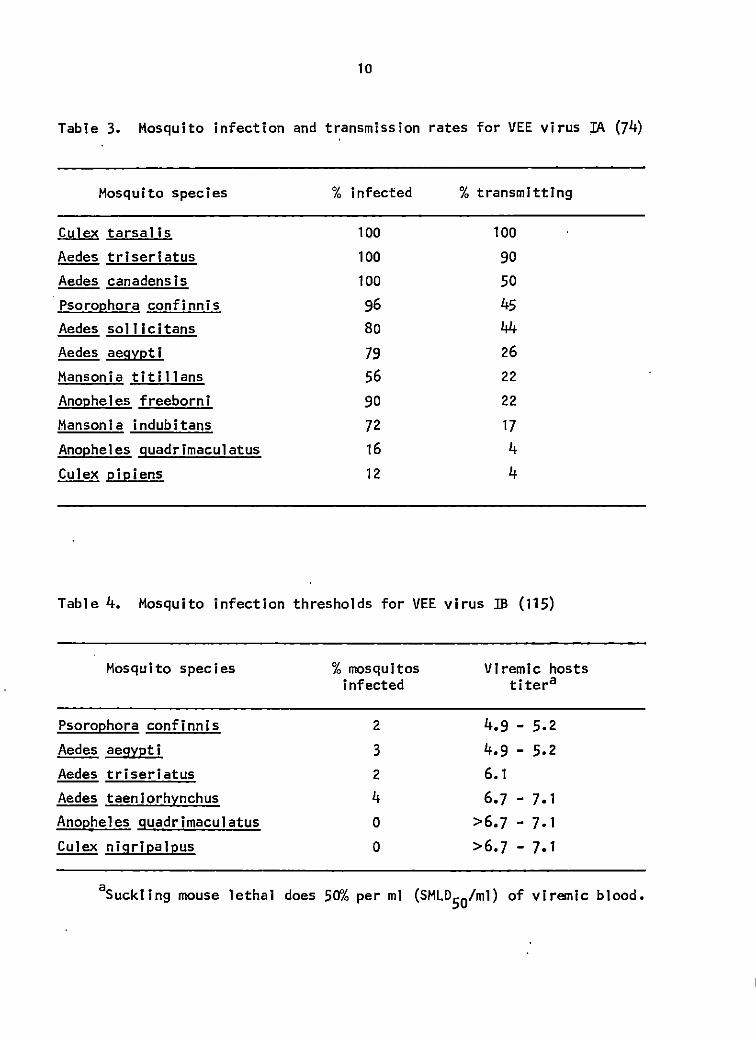
Table 3. Mosquito infection and transmission rates for VEE virus IA (74)
Mosquito species % infected % transmitting
Cy]ex tarsal is 100 100 Aedes triseriatus 100 90 Aedes canadensis 100 50 Psorophora confinnis 96 45 Aedes sol 1 lcitans Bo 44 Aedes aegypti 79 26 Manson i a tit i 11 ans 56 22 Anopheles freeborni 90 22 Mansonia indubltans 72 17 Anopheles guadrimaculatus 16 4 Culex pipiens 12 4
Table 4. Mosquito infection thresholds for VEE virus IB (115)
Mosquito species
Psorophora confinnis Aedes aegvpti Aedes triseriatus Aedes taenlorhynchus Anopheles guadrimaculatus Culex nigripalpus
% mosquitos infected
2 3 2 4 0 0
Vlremic hosts titera
4.9 - 5.2 4.9 - 5.2 6.1 6.7 - 7.1
>6.7 - 7.1 >6.7-7.1
aSuckling mouse lethal does 50"/o per ml (SML050tml) of viremic blood.

11
digestive tract a peritrophic membrane is usually secreted around it
which then serves as a minor barrier to passage of virus from a viremic
blood meal to the midgut epithelial cells. The virus may then attach to
suitable receptor sites if they are ava.ilable. These represent the first
major barrier to infection of the mosquito. Assuming the mosquito is a
suitable host species, the virus adsorbs to the midgut cells and is likely
taken into the cell by the process of pinocytosis or viropexis. Upon
entering the cell it is subjected to the action of a series of enzymes
which uncoat the virion. Replication of the viral nucleic acid and coat
materials then occurs with eventual assembly and maturation of progeny
virions. In the process of maturation the assembled viral nuclear mate-
rial is extruded through the cel 1 's cytoplasmic membrane containing
virus-specified envelope components. Hence, with viral maturation the
progeny virions pass out of the midgut epithelial cell to enter the
hemolymph of the mosquito. Having entered the primitive circulatory
system the virus is then able to infect the organs of the mosquito that
are bathed by the hemolymph. Viral replication ensues, but appears to
continue only in the salivary glands. Within the salivary glands the
virus replicates to high titers where it is considered to persist for
the lifespan of the mosquito. The second major barrier is now at hand.
In order for the mosquito to be infective the virus must be able to bud
out from the acinar cells of the salivary glands into the salivary ducts
or their precursors. If the mosquito in question is a vector species
the virus buds into the salivary gland ducts and the mosquito is then
infective indefinitely according to laboratory studies (16,17). The

12
extrinsic incubation period In the mosquito can be defined as the interval
required for virus to appear in the salivary juices after feeding upon a
viremic host. This period is inversely proportional to the ambient
temperature of the mosquito's habitat. For species of Aedes and
Mansonia the period is probably no more than 8 to 12 days or about-the
span of time between the first and the third blood meals. The period
would probably be even less for a species such as Psorophora confinnis
due to its relatively short lifespan as compared to longer-lived mosqui-
toes such as Aedes sp. (16).
According to the previously listed cri·teria for vector status only
Psorophora confinnis and Aedes taeniorhynchus have been proven as likely
vectors of the IB strains of VEE virus. Insufficient data are available
at this time to unequivocally incriminate these species as vectors of
epizoodemic VEE. Other probable vectors species according to habitat are
as follows (115):
(1) Permanent fresh water with vegetation. Mansonia indubitans and _tl. titillans.
(2) Temporary fresh water pools, sunny and grassy. Psorophora conflnnis, f· discolor, and Aedes thelcter.
(3) Coastal beaches, crab hole terrain. Deinocerites pseudes.
(4) Coastal areas, brackish water, inland 80 to 110 kilometers. Aedes sollicitans and [1. taeniorhynchus.
(5) Woodlands, fresh water, shady pools. Aedes scapularis.
Although the role of mosquitoes as vectors of VEE virus is gradually
being defined, the question of origin of epizoodemics of VEE remains
unknown. Migratory birds have been considered to be a potential means

13
of transport of eastern and western equine encephalomyelitis viruses from
one country to another, but few epizoodemic VEE virus isolations have
been obtained from migratory birds to substantiate such a concept for
VEE virus movement. According to Grayson (51) the only naturally-in-
fected, vlremic (epizoodemic VEE) bird reported has been a fledgling
green heron (Butorldes virescens), also referred to as the striated heron
(B. striatus). On the basis of that isolation Grayson and co-workers (51)
inoculated 9 wild-caught, serologically negative green herons with 100
suckling mouse lntraperitoneal lethal dose 50% (SMIPLD50) of the ~880,
ID (endemic) strain of VEE virus.· The 9 birds were all viremic by the
4th and 5th day postinoculatlon and were used for experimental trans-
mission studies with Deinocerites pseudes as the vector and golden
hamsters as the recipient hosts~ Positive transmission was obtained
indicating a possible role for the green heron in the movement of
epizoodemlc VEE virus strains (IA, IB, IC) from country to country along
the coastal areas as occurred in the epizoodemic that spread from
Guatemala to the United States. Additional supporting evidence is that
green herons commonly inhabit areas where the crab-hole-breeding
Deinocerltes pseudes lives.
To determine If rodents (57,63) could be involved in the main-
tenance of epizoodemlc VEE virus Zarate (135) and Walton (128) inoculated
cotton rats (Sigmodon hlspidus) with epizoodemic strains IC and IB
respectively. With 4 to 1000 plaque-forming units of virus administered
subcutaneously Zarate obtained death patterns of 1 out of 5 and 3 out
of 5 rats inoculated. Walton inoculated 3 weanling~· hispidus with

14
1000 SMICLD50 units of virus and had no survivors by 6 days post-
inoculation. The conflicting results of these two studies and the
statistically small number of experimental animals indicate further
studies in this area are needed to define the possible role rodents
may have· in the maintenance of epizoodemic VEE virus.
Fossaert (40), Franck (41) and Johnson (68) have likewise cited
the possibility of epizoodemic virus residing in natural foci much
as endemic strains of VEE are known to exist. Little evidence other
than that of Grayson (51) and Walton (128) exists to substantiate such
a hypothesis.
Bats have also been proposed as a transport and maintenance mechanism
for epizoodemic VEE by several workers (65). According to Baer, cited
by Calisher (12), the Mexican freetail bat (Tadarida brasiliensls
mexicana) cohabits with the vampire bat (Desmodus rotundus) and
migrates hundreds of kilometers to the United States and to other
regions. In August, 1970, Q. rotundus was found infected with the IB
strain of VEE in the state of Oxaca, Mexico during an epizoodemic (27).
Epizoodemic VEE virus has also been isolated in Ecuador from Q. rotundus
in 1969 (51).
Vampire bats subsist on a strict blood diet and frequently feed upon
equines, consuming as much as 20 to 25 ml of blood per day. While feeding
the bats may either ingest viremic blood or be bitten by infectious
mosquitoes (27). Sanmartin (99) has fed heparinized viremic horse blood
to Q. rotundus and has determined VEE virus to be present in the bat's
oral cavity for!48 to 168 hours postinoculation. Transmission studies

15
have not yet been reported. However, after feeding vampire bats will
often rest in hollow trees which may also serve as resting areas for
mosquitoes. If those mosquitoes were also vector species they could
then feed upon the resting, possibly viremic bats, and later become
Infectious to exacerbate an epizoodemic of VEE (1,29).
Another means of maintenance of epizoodemic strains of VEE virus
that has been postulated is that of silent amplification. Groot (55)
investigated various races of sheep from La Guijara, Colombia and found
that they either developed a transient, minimal viremia or were not
viremlc. However, all of the sheep exhibited serologic conversion from
negative to positive.
Goats have also been considered as possible silent amplifiers of VEE
(57) but confirmatory data are not available at present (104). Serological
surveys conducted during outbreaks of epizoodemic VEE have on the other
hand shown a fairly high ratio of serologic conversion of domestic goats
tested for VEE antibody by the hemagglutination-inhibition and serum-
neutral lzation tests. Bergold (7) found a high incidence of antibody in
goats in Venezuela from 1962 to 1969. By the hemagglutination-inhibition
tests 102 of 332 goats were positive at a serum dilution of 1:20.
Eplzoodemic VEE virus was also obtained from a goat in Zulia, Venezuela
over the eplzoodemlc period of 1968 to 1969.
In an earlier publication, Sellers, et al. (110) described ~ ~
hemagglutlnation-lnhibition and serum-neutralizing antibody titers in
approximately 59 percent of 39 goats sampled from November, 1962 to
January, 1964 in Venezuela. During epizoodemic virus activity in

16
Colombia from 1967 to 1968, Mackenzie (85) obtained an overall antibody
incidence of 20 percent of 31 goats tested. Serum neutralization
antibodies were also found in goats in Texas during the 1971 outbreak
but detailed results are not yet available (94).
The role of the dog as a silent amplifier has.been evaluated by
several groups. Taber, et~· (118) infected beagles with the IA
subtype of VEE virus and obtained adolescent (21-23 day old) mouse
viremia titers ranging from 3.1 to 4.3 log 10 mouse IPLD50 per ml :0f
viremic dog blood. Bivin, et al. (9) were able to transmit IA VEE virus --to beagles with A· triseriatus mosquitoes which had been infected by
the hanging-drop method. Davis, et al. (30) transmitted IA VEE virus --from beagles to guinea pigs with A· triseriatus at viremia titers
ranging from 3.7 to 6.0 log 10 adolescent (21-23 day old) mouse 1PLD50 per ml of blood. However, work done by Sudla (115) indicates
that the threshold for infection of A· triseriatus with the IB
subtype of VEE virus is considerably higher than that indicated by the
work of Davis, et~· (30) with the IA subtype of VEE virus. Based on
the information available it does not seem likely that the dog would
serve as a silent amplifier of IB VEE virus.
Chamberlain (13) has proposed several areas which should be studied
to elucidate the role of alteration of viral virulence in the sudden
eruption of epizoodemic VEE in regions that previously were·11 free" of
v I rus ac:tt vJ t·y. They a re as fo 1 I ows:
(1) Passage in vertebrates with either higher or lower bo~y tempera.tures than the usual hosts.

17
(2) Incubation l.n vectors for either exceedingly long or very short periods.
(3) Passage by the pharyngeal route (abnormal).
(4) Reproductive organ infection and transmission by sexual contact.
(5) Simultaneous infection of either mosquitoes or vertebrates with two strains of virus.
Due to the high magnitude of viremia produced in man by VEE virus
infection he is also subject to consideration as an amplification
mechanism. Viremias in excess of 5.0 .log 10 SMICLD50 per ml of blood
with a r,ange of 3,5 to 6.0 log10 per ml have been reported. Man i·s
therefore a possible vehicle of virus movement but is of secondary
importance when compared to the great mobility of equines (11,36,64,101,
111, 126) •
The movement of viremic equines has very likely played a major role
in the dissemination of epizoodemic VEE in this day of modern, rapid
transportation facilities (103,109,125). McConnell (86) has stated that
about 50 percent of all equines infected by the epizoodemic virus will
be clinically normal. Consequently, frantic owners have been known to
ship their valuable horses out of regions where virus activity is prev-
alent and is responsible for dally reports of equine mortality and
morbid l ty ( 120). Race horses have a 1 so been i ncr im i nated as 1ike1 y
prospects due to their widespread travels from ranches to racetracks (93).
Many factors point to equine movement as a major mode of spread of
epizoodemic VEE virus but little specific evidence has been accumulated
to date.

18
During the 1970 VEE outbreak in Central America an inactivated
vaccine of poor quality was prepared in Nicaragua from IB VEE virus
and distributed for use. Shortly thereafter breaks of the disease
appeared to follow the path of the vaccine. When use of the preparation
was suspended the equine cases of VEE immediately ceased (89).
When 11 new11 epizoodemics of VEE appear the poor monitoring of equine
deaths or complete lack of monitoring must be taken into consideration.
The probability would seem to be great that epi zoodemic virus activity
could have been occurring undetected in isolated regions of a country
with a small but significant number of equine deaths (122) . The con-
current presence of endemic and epi zoodemic VEE viruses in a given
region would present a partially immune equine population and would
likely keep equine disease to a minimum. In this type of situation
the epizoodemic virus activity would probably be detected only on the
periphery of the endemic virus area resulting in a very low level or
complete lack of virus activity being reported.
The mechanism by which the epizoodemic virus overwinters in a given
region is not known. On January 10, 1973, IB VEE virus was isolated
from a pool of 50 Culiseta melanura in Xochimilco, Distrito Federal,
Mexico, the first isolate of an overwintering study initiated in coop-
eration with the Communicable Disease Center (CDC) , Atlanta, Georg i a, by
the Animal Health Department, Ministry of Agriculture and Livestock,
Mexico (22).

19
Young (133) has proposed the phenomenon of recrudescence as a source
of epidemics. Supportive evidence presented was the recurrence of
clinical illness due to the IE subtype of VEE virus in a laboratory field
worker one year after initial illness in a .region of Panama where IE VEE
virus had never been isolated.
Endemic (enzootic) VEE
As listed in Table 1, antigenic groups ID, IE, II, III, and IV are
not epizoodemic strains. Due to their focal nature involving primarily
small rodents and possibly birds they have collectively been denoted as
either endemic or enzootic VEE (4,18,19,46,47,66,72,74,95,107,108,134).
Endemic virus strains have been isolated from 7 species of birds, while
natural antibodies against the virus have been found in at least 23 avian
species. In various laboratory trials all bird species studied developed
low to moderate viremias, usually of 2 or more days• duration. In
mosquito-transmission -experiments, some mosquitoes have become infected
by feeding on birds with viremias as low as 2.6 log10 SMICLD50 of endemic
VEE virus per ml. However, the presence of antibodies against endemic
VEE virus in field-trapped birds has been highly variable. For given
regions of endemic virus activity investigators have reported a complete
lack of serological evidence in field-trapped birds while other groups
have found a very high incidence of endemic VEE virus antibodies in
their field-trapped birds from other endemic regions. Due to the marked
discrepancies at present in this area of study, the role of birds in the
maintenance of endemic VEE virus activity cannot be discounted as

20
insignificant (32,53,54,82,83).
Endemic foci usually do not exceed one qua.rter of a square mile and
are usually found in high rainfall areas with either a tropical or sub-
tropical climate (71). The foci may be in wooded regions and have often
been found to be proximal to fresh-water swamps. The endemic virus cycle
is mosquito-rodent-mosquito with occasional bird hosts comingled as noted
earlier. Marsupials, bats, and raccoons have recently been incriminated
in the cycle with small rodents (8,52,71,80,82,100,102,106). In Florida
the mammals primarily involved in the cycle with Culex (Melanoconion) spp.
appear to be the cotton rat (Sigmodon hispidus), the cotton mouse
(Peromyscus gossypinus), the raccoon (Procyon lotor), and the opossum
(Didelphis marsupialis)(8,15,82). See Table 5 for mammals involved in
the maintenance of endemic VEE virus;
A total of 38 species of mosquitoes in 14 genera and subgenera have
been implicated in the transmission of endemic VEE virus (Table 6).
Among them only 3 species have been reported in the literature as well-
substantiated vectors. Galindo (45) indicated that 2 species of mosquitoes
proven to be efficient natural vectors of endemic VEE virus are Culex
(Melanoconion) aikenii and.£. (tl.) portesi. Mosquito groups other than
Culex (tl.) spp. are probably involved as secondary vectors in the natural
transmission of endemic VEE virus. The third species which has been
clearly implicated as a primary vector of endemic VEE is Culex (tl.)
cedeci (15). These vector species would seem to possess a threshold for
the endemic strains of VEE virus not unlike that of Culex tarsal is and
western equine encephalomyelitis. Naturally-infected rodent species have

21
Table 5. Mammals considered to be involved in the maintenance of endemic VEE virus activity (47' 52, 53,54, 71,80,82,85, 1oo,102, 112, 113, 119, 1 31)
Species Virus Antibody isolation Hla CFb
Rodent Cotton rat
(Si smodon hisEidus} x x Terrestrial rice rat
(Orzom)!S 1 at i ceEs) x x Rice rat
(Orzom)!S Ealustris) x Spiny rat
(Proechlm)!S semisEinosus} x x R Ice rat
(Orzom)!S cali9nosus) x Common rat
(Rattus rattus) x Thorny rat
(HoElomys !J)!mnurus) x Spiny rat
(Proechlm)!S !JU)!annensis or is) x x Rice rat
(Orzomys caEito soeldii) x x Chisel-toothed kangaroo rat
(DiEodom}:'.s microEs) x Cotton mouse
(Perom)!SCUS !JOSS)!Einus) x Forest pocket mouse
(Heterom)!S anomalus} x x Short-tailed cane mouse
(Z)!9odontom)!S brevicauda) x x Deer mouse
(Perom)!Scus maniculatus} x Western harvest mouse
(Reithrodontom)!S me9alotis) x Mouse
(Perom}:'.scus mexicanus) x
aPresence of hemagglutination-inhibition antibodies. bComplement-fixation titers <?: 1: 16. cPresence of serum-neut ra 1 i zing an·t i bodies.
SNc
x
x

Table 5. (Continued)
Species
Marsupial Common opossum
(Didelphis marsupial is) Philander opossum
(Philander sp.) Woolly opossum
(Cal uromys derb I.anus) Murlne opossum
(Marmosa mitis)
Bats --;ii;"t I be us 11 turatus
Artibeus turp is Carollia subrufa Caroll la persplcillata Glossophaga sorcina Artibeus sp. Caroll la sp.
Other.: Raccoon
(Procyon 1 otor) Forest rabbit
(Sylvilagus brasiliensis) Desert cottontail
(Sylvilagus audubonii) Nuttal cottontail
(Syl vi 1 agus nuttal 1 ii) Black-tailed jackrabbit
(Lepus callfornicus) Paca
(Agouti pa ca) Red squirrel
(Sci urus granatens is) Vari~gated squirrel
(Sciurus variegatoides) White-ta i 1 ed ante·l ope squ i rre 1
(Citellus leucurus) Weasel
(Mustella frenata) Kit fox
(Vulpes macrotus)
22
Virus isolation Hla
x x x x x x x x
x x x x
x x x x
x x
x x x
x
Antibody CFb
x x x
x
x
SNc
x
I
.I

23
Table 6. ·Mosquito species from which endemic VEE.viruses have been isolated (2,3,4,15,19,46,47,53172,74,105,113,131,135,136)
Mosquito species
Aedes angustivittatus Aedes atlantlcus Aedes scapularis Aedes serratus Aedes taenlorhynchus
Anopheles aguasalis Anophe 1 es n lmbus Anopheles punctimacula
Coguillettldla (Rhynchotaenla) albicosta
Cogu i 11 ett id i a (Rhynchotaen i a) venezuelensls
Culex corniger Culex coronator Culex nlgrlpalpus Culex guinguefasciatus Culex thriambus Culex (Eubonnea) accelerans Culex (Eubonnea) amazonensls Cu lex· (Me 1 anocon ion) a i ken ii Culex (Melanoconion) albinensis Culex (MelanoC:onion) cedeci Culex (Melanoconion) epanastasis Culex (Melanoconion) iolambdls Cu·lex (Melanoconion) opisthopus Culex (Melanocon.ion) portesi Cu·l ex (Me 1 anocon I on) sp i ss i pes Culex (Melanoconion) taeniopus Culex (Melanoconlon) thriambus Culex (Melanoconlon) vomerifer Culex (Melanoconion) ybarmls
Deinocerltes pseudes
Hemagogus mesodentatus Hemagogus (stegoconops) spegazzinli
Country
Brazil, Colombia, Ven~zuela Florida, U.S.A. Brazil, Mexico, Venezuela Brazil, Trinidad• Venezuela French Gu1ana; Florida, U.S.A.
Brazil, Venezuela Braz i 1 Colombia
French Guiana
Brazil, French Guiana, Trinidad
Colombia Mexico Trinidad; Florida, U.S.A. Brazil, Colombia, Mexico, Panama Mexico Trinidad Trinidad Brazil, Colombia, Panama French Guiana Florida, U.S.A. Trinidad Mexico Mexico Brazil, French Guiana, Trinidad Trinidad Braz 11, French Guiana, Panama, Tr'in i.dad Mexico Brazil, Panama, Trinidad Trinidad
Mexico, Panama
Mexico Braz i 1

Table 6. (Continued)
Mosquito species
Limatus durhami Llmatus fl avisetosus
Mansonla fasciolata Mansonla titillans
Psorophora ferox
Sabeth i n.i sp.
Wyeomyia medioalbipes Wyeomyla mltchelli Wyeoinyla (Dendromy.ia) occulta
24
Trinidad Trin.idad
Mexico Trinidad
Country
Brazil, Mexico, Trinidad
Braz i 1
Trinidad Mexico French Guiana

25
been found to deve.lop vlremlas of 3.9 to 5.0 log 10 SMLD50 per ml of
viremlc blood which persist from 4 to 5 days (45,71). Within endemic
foci vector species have been found to be infected throughout the year
(81). The presence of susceptible host spec.ies for maintenance of the
virus is greatly facilitated by the average lifespan of 6 months for
ground-dwelling rodents. Continuous virus activity is further assisted
by the rodents' selective habit of breeding during the rainy season when
large populations of the vector species are available (71).
The fact that certain species of Culex of the subgenus Melanoconion
such as f.. (tl.) portesi (71) will readily feed upon man presents tne
potential of outbreaks of disease in man due to endemic VEE. Mosquito
species in the genera Aedes, Anopheles, Deinocerites, Mansonia, and
Psorophora are also known to feed avidly on equines and man .. (115).
Since species of these genera are involved in the transmission of both
endemic and ep.izoodemic strains of VEE virus, man is indeed a very 1 ikely
candidate for disease in the presence of high virus activity in a given
region. The first human case of VEE in the United States was due to
endemic virus activity in Florida as reported by Ehrenkranz, et~· in
1968 (35). More recently, however, the epizoodemic of VEE in Texas
resulted in 88 laboratory-confirmed cases of human VEE with al 1 but 2 of
the cases occurring in July, 1971 (24).
The fluorescent antibody .test
The swift spread of epizoodemic VEE and its high morbidity rate
necessitates a rapid di agnost I c system. The fluorescent anti body test

26
(FAT) as developed by Coons, et~· (26) with subsequent modifications
meets nearly all the requirements for a rapid, sensitive diagnostic
test when performed in a tissue culture system.
Several types of globulin separation and purification have been
employed in preparation of fluorescent-antibody conjugates. The two
more commonly employed techniques at present are 50 percent saturated
ammonium sulfate precipitation and chromatographic separation of a
ganvna-globul in-rich (lgG) fra.ction from hyperimmune sera.
Coons,~~· orginally used fluorescein isocyanate for conjugation
(26) but this compound has the disadvantages of being unstable and
dangerous to prepare. The synthesis of fluorescein isothiocyanate (FITC)
by Riggs, et~· in a more stable powder form has essentia.1 ly replaced the
isocyanate compound for use in antibody conjugation (79,97 ,98). · The
labeling of gamma globulin fractions with FITC can be done by either
direct addition or by dialysis labeling. For direct conjugation FITC
Is ordinarily added dropwise to the globulin fraction at a ratio that
has varied from 1:20 (1 mg of FITC to 20 mg of protein) up to 1:200.
Current literature indicates the most suitable range for optimal tagging
to occur Is from 1:100 to 1:200 (44,132).
Dialysis labeling, as described by Clark and Shepard (20), is gener-
ally considered to result in more uniform tagging of the antibody mole-
cules of the globulin preparation. Uniform labeling is also enhanced by
the lack of albumin and any traces of macroglobulins (lgM). If these
proteins are present their affinities for FITC are somewhat greater than
that of the lgG In the globulin preparation resulting in less FITC-tagging

27
of lgG and a final conjugate with high background fluorescence and
decreased specificity of fluorescence (88).
Dilution, tissue powder adsorption, and anion-exchange column
chromatographic purification of conjugates are three methods of removal
or marked reduction of antibody molecules excessively tagged with FITC
after unreacted FITC has been removed by either Sephadex G-25 1 chromato-
graphy or dialysis against a buffered saline solution. Dilution is one
of the most widely used. techniques to reduce.nonspecific staining due
to excessively-tagged antibody molecules and may readily be employed.
with hlgh-titered conjugates. Tissue powder adsorption and anion-
exchange chromatography both reduce the antibody content of conjugates
but they also result in a conjugate that is more specific in its staining
characteristics. The· disadvantage of the tissue powder adsorption
technique is that it can lead to bacterial contamination of the finished
conjugate. The advantage of anion-exchange chromatography is that only
optimally-labeled antibodies are left in the conjugate (28,31,49,87).
1Pharmacia Ltd., Uppsala, Sweden.

28
MATERIALS AND METHODS
Cell cultures
To determine the optimal in vitro cell culture system for the iso-
lation and identification of the IB subtype of VEE virus a compar.ative
propagation study was undertaken. Eight types of ceil cultures were
analyzed for their ability to support growth of the virus with the
production of visible cytopathic effects under a standard nutrient agar
overlay. The cell cultures employed in this study were Vero African
green monkey kidney cell line1, human amnion (FL) cell line1, baby hamster
kidney (BHK-21) eel I 1 ine1, L eel I 1 ine1, goat kidney secondary cel 1
culture2, bovine turbinate cell line2, horse kidney secondary cell culture2,
and duck embryo fibroblast (DEF) primary cell culture. All cell cultures
with the exception of the BHK-21 were grown and maintained with Gibco F-1S3
medium supplemented with 10.0 ml L-glutamate and 10.0 ml sodium pyruvate
per liter of medium plus serum as indicated below. The BHK-21 cell line
was grown and maintained with Stoker's modification4 of Eagle's BME 3 which
is the addition of tryptose phosphate broths to a concentration of 10
percent of the final medium. Antibiotics employed in the two culture
media were 10,000 IU penicillin, 0.13 g streptomycin, and 2.S mg nystatin
per liter. For growth a 10 percent concentration of fetal calf serum
1As obtained from the American Type Culture Collection, Rockville, Md. 2Developed and maintained by Diagnostic Virology, APHIS, USDA, Ames, la. 3Grand Island Biological Co., Grand Island, N.Y. 4stoker, M. 1962. Virology 1S:147-1S1. Soi fco Laboratories, Detro it,. Mi ch.

29
was used, and cells were maintained with medium containing 5 percent fetal
calf serum with the exception of the BHK-21 cell culture which was main-
tained with the growth medium.
For the comparative propagation study each of the 8 cell lines above
was seeded into 25 cm2 Falcon flasks 1 and was inoculated when confluent
with o.1 ml of a tenfold dilution of either cell culture or suckling mouse
brain origin virus. The inoculated flasks wer.e placed in a 37C incubator
for one hour to allow virus adsorption. They were then overlaid with
5.0 ml of 1 percent Noble agar2 containing lx Earles 6553, 0.5 percent
lactalbumin hydrolysate, 0.22 g' sodium bicarbonate per liter, 2 percent
fetal calf serum, 3.3 ml of 1:100 neutral red per 200 ml, and 400 IU
·penicillin, 200 µg streptomycin, and 100 µg nystatin per ml. Plaques
were first counted after 48 hours incubation at 37C and again after an
additional 24 hours incubation at 25C. The endpoint was calculated and
expressed as the number of plaque-forming units per 0.1 ml (PFU/0.1 ml).
Titers were then compared to virus titers obtained by suckling mouse
inoculation to determine the relative sensitivity of each cell culture
to IB VEE virus.
Vi ruses
The GJ9-1BJ strain of VEE virus was isolated from a pool of Psorophora
confinnis mosquitoes collected near Parcelamiento Montufar, Guatemala, in
1Falcon Plastics, Division of BioQuest, Los Angeles, Ca. 2Difco Laboratories, Detroit, Mich. 3Grand Island Biological Co., Grand Island, N. Y.

30
1969, during the course of an epizoodemic (62,116). Working stocks of
this virus were prepared by inoculation of suckling mouse seed virus as
supplied to Diagnostic Virology, APHIS, USDA, Ames, la., by CDC, Atlanta
Ga., into suckling mice and two cell lines, Vero and goat esophagus 1• A
0.5 ml quantity of a 1:100 dilution of 10 percent mouse brain suspension
prepared in 0.01 M, pH 7.2, calcium-and magnesium-free phosphate-buffered
saline containing 0.75 percent bovine serum albumin (BAPBS) was inoculated
into each 75 cm2 Falcon flask of confluent cells. After a one hour
adsorption period at 37C, 25 ml of maintenance medium containing one
percent serum was pipetted into each flask and they were returned to a
37C incubator. Fetal calf serum and normal goat serum were used in the
respective cell cultures at a lower than normal concentration to allow
use of the same stock viruses for tissue culture and goat inoculation
studies. After a 48 hour incubation period the culture fluids were
harvested as stock viruses and were maintained at -70C.
The suckling mice inoculated for stock virus preparation were
observed daily for signs of encephalitis. When 5 to 15 percent of the
suckling mice were either prostrate, moribund, or dead the litters were
harvested, A 10 percent suspension of the pooled mouse brains was
prepared in BAPBS, divided.into 1.0 ml aliquots, and maintained at -70C as
virus stock.
1As obtained from the American Type Culture Collection, Rockville, Md.

31
Experimental animals
Twelve domestic goats of mixed sex from 4 to 18 months of age were
used in the experiments. Prior to purchase the goats were screened by
the hemagglutination-inhibition test and were found to be negative for
antibodies agai·nst eastern equine encephalomyl itis, western equine
encephalomyelitis, and Venezuelan equine encephalomylitis viruses. They
were housed in maximum biological security stalls with a filtered air
intake and exhaust supply system that.is more than 99,6 percent effective
in the removal of viral disease agents.
Albino suckling mice, CF-1 strain, 2 to 4 days of age, obta.ined from
Carworth Farms, Division of Becton, Dickinson and Co., New City, N. Y.,
were used for viremia assays and for titration of virus strains. The
viruses were titrated in tenfold serial dilutions using 8 mice per dilu-
tion. Each mouse was inoculated intracerebrally with 0.02 ml of a given
virus preparation and was then observed daily for 7 days. Moribund,
prostrate, and dead mice were removed and stored at -70C until their
brains could be processed for preparation of·complement fixation (CF) test
antigen to confirm the presence of VEE virus in the affected mice. The
log 10 SMICLD 50 per ml endpoint for each specimen was determined according
to the method of Reed and Muench, cited by Lenette and Schmidt (79).
Serology
Hemagglutination-inhibition (HI) tests were performed by a microtiter
modification of the method of Clarke and Casals (21) in twofold serum
dilutions from 1:10. The test antigen was prepared from GJ9-1BJ VEE

32
virus infected mouse brains by sucrose acetone extraction. Ninety-six
well, u-bottom plastic plates 1 with an arrangement of 8 by 12 wells were
used for the test. A 0.25 percent suspension of goose red blood cells
prepared in dextrose-gelatin-veronal buffer was employed for all hemagglu-
tination-inhibition tests. The mouse brain antigen mentioned above was
then titrated for 4 to 8 hemagglutination units per 0.025 ml using a 0.2%
bovalbumin, pH 6.o borate saline (BBS). Sera to be tested were likewise
diluted in BBS for the HI test. Initially 0.05 ml of a 1:10 serum dilu-
tion was placed in the first well of the plate and 0.025 ml of BBS was
dispensed in the succeeding wells. Twofold dilutions were then made with
0.025 ml microdiluters 1, 0.025 ml of titrated antigen was placed in each
well, and the test was incubated overnight at 4C. The following morning
0.05 ml of a freshly prepared 0.25 percent suspension of goose red blood
cells was added to each test well and the plate was briefly agitated for
mixing of the test reagents. After one hour incubation at 37C the plate
was read for the absence of hemagglutination, indicating a positive HI
test.
Serum virus neutralization (SN) testing was performed in 2 virus-
tissue culture systems. The GJ9-1BJ strain of VEE virus was used with the
BHK-21 cell line and the TC-83 strain of VEE virus was used with DEF cells
for titration of goat sera. All sera tested were heat-inactivated at 56C
for twentymfoutes. The 0.2 ml al lquots of tenfold dilutions of sera being
tested were aseptically dispensed into sterile 12 x 75 mm stoppered tubes
1cooke Engineering Co., Alexandria, Virginia.

33
to each of which were added 0.2 ml of virus containing 120 to 200 PFU
per 0.1 ml. The resultant mixtures were then agitated briefly and placed
in a 37C waterbath for a one hour period of virus neutralization. At
the end of the incubation period the tubes were removed and placed in a
wet ice waterbath. One tenth ml of each mixture was then pipetted into
2 25 cm2 Falcon flasks of either BHK-21 or DEF cells according to the
strain of virus being used in that test. Following a one-hour period of
virus adsorption at 37C, 5 ml of the Noble agar preparation previously
described was dispensed into each flask. The flasks were then placed on
a level surface at 25C for 15 to 25 minutes for agar solidification. When
the agar had hardened the flasks were inverted and were incubated at 37C.
After 48 hours the flasks were removed and the first plaque coun~ was made.
A final count of plaques was made after 24 hours• incubation at 25C. The
endpoint of a serum was that dilution producing at least a 90 percent
plaque reduction.
The complement fixation test was used to confirm that suckling mice
employed for virus titrations which had been either moribund, prostrate,
or dead at the time of harvest were infected with VEE virus. A 10 percent
suspension of mouse brain was prepared in veronal buffer diluent (VBD),
and was centrifuged at 10,000 rpm for 30 minutes. The supernatant fluid
was retained as the 4x CF test antigen. The microtiter CF test' was
performed with either VEE antiserum or mouse ascitic fluid employing
dilutions of 1:8 through 1:256, a 1:20 dilution of unkown antigen, 7
hemolytic units of complement, and sensitized sheep red blood cells. The

34
VEE ant I serum was di 1 uted in the p 1 ates with subsequent add! ti on of
antigen and complement prior.to incubation overnight at 4C for complem~nt
fixation. The following morning sensitized sheep red blood cells were
added and the plate was incubated at 37C for 30 minutes, followed by
centrifugation at approximately 250 g for 10 to 15 minutes to pack the
remaining ghost and red blood cells. The test was read by comparing
hemolysls against known color standards. From zero to 30 percent hemolysis
was considered to be a positive test. Wells displaying greater than 30
percent hemolysls were cons.idered to be negative.
Sentinel study
Six goats were inoculated intradermally with 1000 SMICLD 50 of goat
esophagus cell culture origin GJ9-1BJ VEE virus (GEV). The 6 goats were
divided into 2 groups with one noninfected goat per group to determine if
contact transmission of GEV from goat to goat was possible. Blood samples
were taken with and without heparin from the 8 goats from zero to 14 days
postinoculation (DPI) for determination of viremia and for serologic assay.
An additional aliquot of blood was obtained at zero DPI from each goat to
provide normal goat serum for conjugation. Nasal, oral, and genitourinary
swabs were also obtained 0-7 DPI to determine if VEE virus was shed by
the infected goats. Serum samples were obtained from the 8 goats on a
dally basis from 15-21 DPI and biweekly thereafter through 49 DPI. Rectal
temperatures were recorded dally from 0-21 DPI. Goat viremia assay was
done by BHK-21 plaque assay and intracerebral inoculation of suckling mice.
Serologic responses were determined by HI and SN tests using viruses TC-83

35
and GJ9-1BJ.
Antiserum production
Two groups of goats were employed for the 2 immunization protocols.
Two goats were inoculated subcutaneously (SC) with 1000 SMICLD50 of GEV
In Freund's complete adjuvant 1 at o, 10, and 20 DPI. The 2 goats of the
second group were inoculated SC with 1000 SMICLo50 of GEV, which was
followed by 4 SC injections of the virus-adjuvant mixture described above
at 8, 10, 12, and 18 DPl. Serum samples were obtained at o, 21, 30, 42,
and 49 DPI for antibody assay. By 49 DPI the response of the 4 goats was
not adequate {1:1000) by the SN test with virus GJ9-1BJ and the goats were
subjected to challenge with 25,000 SMICLD50 of GEV intravenously (IV) at
50 DPI. Serologic response was then monitored at 59, 63, 70, 72, and 80
DPI to evaluate the 4 goats' response following challenge. The 2 goats
of the first group displayed the greatest serological response by SN
testing with vi~us GJ9-1BJ at 63 DPI.· Serum neutralization tests were
done in the latter half of each week which meant that the 63 DPI results
were not available until approximately 69 DPI. Serum harvest of the 2
goats by exsanguination was consequently done at 72 DPI. The goats of the
second group were exsanguinated with serum harvest at the termination of
this study at 90 DPI.
Seven of the 8 goats in the sentinel study were also used for the
production of hyperimmune serum. One of the contact transmission goats
was given 40,000 SMICLD50 of GEV IV at 50 DPI and rectal temperature was
1Dlfco Laboratories, Detroit, Mich.

36
taken daily for 7 days. The 6 previously infected goats were challenged
at 50 DPI with 25,000 SMICLD50 GEV given IV. Serologic monitoring was
continued on a biweekly basis through 90 DPI. Exsanguination with serum
harvest of the 7 goats was done at 70 (2 goats), 72 (1 goat), and 90
(4 goats) DPI.
VEE conj ugat_e preparation
The serum of goat 152, which had the highest neutralizing-antibody
titer to VEE using GJ9-1BJ virus, was selected for f.luorescent antibody
(FA) conjugate preparation. The FA conjugate was prepared by 2 different
techniques to assure that one of the conjugates would be of very high
quality. The first technique used was the method described by Goldman (48).
Four m.1 of the goat 152 serum were placed in an Amicon ultrafiltration
ce11 1 equipped with a membrane designed to retain materials of greater
than 100,000 molecular weight. Thirty-six ml of 0.1 M, pH 8 TRIS buffered
saline were added to the cell and the cell was then placed on a magnetic
stirrer set at a low speed to avoid denaturation of serum proteins. A
pressure of 10 psi was applied to the cell for a period of approximately
45 minutes to reduce the mixture to a volume of 4 ml and to remove more
than 80 percent of the serum albumin originally present. The air pressure
was then removed and the fi 1 tered protein so 1 ut ion was app 1 i ed to a 400 cc
co 1 umn packed with Sephadex G-2002equi1 i brated with the TR IS buffer 1 i sted
above. The gamma globulin (lgG) fraction was eluted overni9ht by reverse
1Scientific Systems Division, Amicon Corporation, Lexington, Mass. 2Pharmacia Ltd., Uppsala, Sweden.

37
flow chromatography and was recorded with an ultraviolet scanning device1•
The Lowry technique (84) was used to determine the protein concentration
of the lgG fraction. The lgG fraction was then pressure dialyzed against
0.05 M carbonate.-blcarbonate buffer, pH 9.0, to a 1.5 percent (15.0 mg
protein per ml) concentration and was ·placed in 8.0 mm dialysis membrane
tublng 2• A solution of fluorescein isothiocyanate3 (FITC) equal to 10
times the volume of the lgG fraction was prepared in a 250 ml beaker
with 0.05 M, pH 9.0 carbonate-bicarbonate buffer at an FITC concentration
of 0.1 mg per ml of buffer. The dialysis tubing containing the lgG and a
magnetic stirring bar were placed in the FITC solution at 4C. The beaker
was covered with aluminum foll to prevent evaporation and was stirred
slowly overnight with a magnetic mixer.
The FITC-tagged lgG preparation was then removed from the dialysis
tubing and was placed on a 2,0 x 20.0 cm column of Sephadex G-254 which
was equilibrated with 0,01 M, pH 6.8 phosphate buffered saline (PBS). T~e
conjugate was then eluted with the same buffer and was collected as a
single colored band from the column.
A 2.0 x 20.0 cm column of QAE-Sephadex4 (diethylaminoethyl-Sephadex)
was prepared in 0.01 M, pH 6.8 PBS for purification of the conjugate. The
conjugate was adsorbed to the column and was subsequently eluted by step-
wise addition of 0.1, 0.2, and 0.3 M NaCl to the PBS. Two column bed
1 instrumentat.ion Specialties Co., Lincoln, Nebraska. 2 Union Carbide Corporation, Films-Packaging Division, Chicago, 111. 3 The Sylvana Company, Milburn, N. J, Lot No. 1043. 4 Pharmacia, Ltd,, Uppsala, Sweden.

38
volumes, approximately 130 ml, were collected for each change of buffer.
The fractions were then concentrated to the volume of the original lgG
fraction by pressure dialysis, evaluated for fluorescent antibody activity,
and stored at -20C.
The second technique involved globulin precipitation with anvnonium
sulfate, direct tagging with FITC,·and tissue powder adsorption of the
conjugate (90). A 15.0 ml aliquot of antiserum from goat 152 was frac-
tionated at 4C by dropwise addition of an equal volume of 100 percent
saturated ammonium sulfate solution to obtain a gamma-globulin precipitate.
The resultant solution was stirred overnight at 4C and was then centrifuged
at 250g for twenty minutes. The supernatant fluid was discarded and the
precipitate was dissolved in distilled water to a volume of 15.0 ml. The
protein solution was then reprecipltated twice at 4C over a 3-hour period
as described earlier and the final precipitate was redissolved.in distilled
water to a volume of 8.o ml. The resultant protein solution was diaiyzed
against repeated changes of 0.85% NaCl solution at 4C until free of sulfate
ions (so4) as determined by reacting a portion of the o.85% NaCl solution
with a saturated solution of barium chloride. If so4 ions were present
a white precipitate would have been observed.
The protein concentration of the globulin solution was determined by
the bi uret method (50) using a Co 1 eman spectrophotometer at 540 and 560 m µ..
The protein solution was then adjusted to a concentration of 1% by the
addition of 0.1 M, pH 7.2 PBS (PBS'). The globulin solution was tagged
!
!

39
1 directly at a ratio of 1:20 (0.50 mg FITC per mg globulin). A solution
of FITC equal to one-tenth the volume of the globulin solution was prepared
with 0.5 M, pH 9.0 carbonate-bicarbonate buffer and was added dropwise to
the globulin solutton with constant stirring. The FITC-globulin mixture
was then stirred overnight at 4C with a magnetic mixer.
A 2.0 x 20.0 cm column of Sephadex G-25 equilibrated with PBS' was
used to remove FITC as described for the first conjugation preparation
technique. The conjugate was next adsorbed with acetone-extracted rabbit
liver powder. One g of liver powder was dissolved in 2.5 ml of PBS' for
each 20 ml volume of conjugate. The rabbit liver powder slurry and con•
jugate were then mixed overnight at 4C in a 250 ml beaker with the aid of
a magnetic stirring bar. The following morning the mixture was centrifuged
at 30,000 rpm for hour and the supernatant fluid was retained for further
treatment. After dialysis against PBS• for 60 hours the conjugate was
dispensed Into 1.0 dram screw-cap vials and maintained at -20C •. Conjugate
staining specificity of two conjugate preparations was evaluated by staining
VEE-virus-Infected BHK-21 cells.
Normal conjugate preparation
Four ml of pooled normal goat serum were fractionated by reverse flow
Sephadex G-200 chromatography. The resulting lgG fraction was then con-
jugated according to the first technique described. To obtain a working
dilution the normal conjugate was diluted to equal the intensity of
1 The Sylvana Company, Milburn, N. J. Lot. No. 1043.

40
background fluorescence of the VEE conjugate when both conjugates were
used to stain normal BHK-21 cell sheets.
Specimens for examination by FAT
In addition to FA endpoint titrations of suckling mouse GJ9-1BJ and
DEF TC-83 VEE viruses, 10 percent tissue suspensions were examined by FAT.
The VEE (GJ9-1BJ)-infected equine tissue suspensions were prepared from
spleen, tonsil, liver, salivary gland, pancreas, adrenal gland, thxmus,
lung, cerebellum, renal lymph nodes, splenic lymph nodes, mesenteric
lymph nodes, and prescapular lymph nodes.
Preparation of slides
Baby hamster kidney (BHK-21) cells grown on coverslips in Leighton
tubes were used for al I fluorescent antibody tests (FAT). Four tubes
were employed for each test. When the cells were 85 to 100 percent
confluent the growth medium was decanted and each tube was inoculated with
0.1 ml of the material to be tested. The tubes were then returned to a
37C incubator for a one hour period of adsorption. At that time a 1.25 ml
aliquot of maintenance medium was dispensed Into each tube. Two tubes per
test were decanted and rinsed twice with PBS' at 24 and 48 hours postin-
oculation. The cell monolayers were then fixed at 4C for a minimum of
twenty-four hours by the addition of 10 ml of acetone to each tube.
For staining the fixed BHK-21 coverslips were removed from their
tubes and were allowed to air dry at 25C. Approximately 0.05 ml of VEE
conjugate was applied to each coverslip as a thin film. The cells were
then stained for a minimum of 20 minutes in a moist chamber at 37c. After

41
incubation the coversl ips were rinsed first in PBS and next in distil Jed
water. After drying at 37C, the stained coverslips were mounted"cell
surface down on 2.5 x 7.5 cm clear glass microscope slides with a mounting
medium of 50% glycerin in PBS'.
Two"conjugate specificity controls were employed for FAT, Normal
conjugate was appJ ied to VEE-virus-infected eel I monolayers, and VEE
conjugate was applied to normal cell monolayers. Eastern and western
equine encephalomyelitis virus-infected cell monolayers were also stained
with VEE conjugate to evaluate the conguate•s specificity of staining.
Microscopy
Incident-I ight fluorescence microscopy was used for examination of
fluorescent antibody stained preparations (96). A Leitz Orthoplan
microscope stand with a Leitz fluorescence vertical illuminator and
mercury arc lamp (OSRAM HBO 200) was
system: The excitation filters were
equipped with 1 1 2.0mm KG-1
the following filter
to block the infrared
spectrum, 2 4.0mm BG-38 1 filters to absorb the red spectrum, 2 KP-4902
short-wave pass-interference filters to obtain an excitation wavelength of
490nm, and a 45 degree angle beam splitter TK-510 3 which reflected the
490nm light beam through the objective to the specimen and allowed only
I ight of"= 510nm to be transmitted back to the eyepieces. The barrier
filter system included the beam splitter as described, ~ne TK-515 3 barrier
1schott and Gen., Mainz, Germany. 2Balzars, Vaduz, Lichtenstein. 3E. Leitz, Wetzler, Germany.

42
1 'filter to pass light of :.:515nm, and one K-530. barrier passing light of
::.: 530nm to the eyepieces. The 2 KP-490 filters combined with the TK510
beam spl ltter ,produced a narrow-band filter system with both high.
transmittance (::.: 80%) around 490nm and very low transmittance (:<: 0.0001%)
around 525nm (96).
Fluorescence photomicrography
The Leitz Orthomat fully automatic camera system and Kodak high speed
Ektachrome film2 (ASA rating of 160) were used to record FAT results.
Exposure times ranged from 15 to 45 minutes.
1 E. Leitz, Wetzlar, Germany. 2Kodak, Rochester, N. Y.

43
RESULTS
Comparative propagation of VEE virus
Despite repeated attempts neither cytopathic effect (CPE) or
plaqulng was obtained with the L cell line. Up to 5.0 x 105 SMICLD50 of
the GJ9-1BJ strain of VEE virus were used in combination with a moist
37C 5% co2 Incubation system as employed by Hardy and Brown (60). In a
similar fashion, repeated attempts failed to produce either CPE or
plaquing in goat kidney, human amnion (FL), bovine turbinate, and horse
kidney cell cultures.
The Vero African green monkey, duck embryo, and BHK-21 cell cultures
were determined to be of approximately equal sensitivity in the propagation
of virulent VEE virus. Cytopathic effects (CPE). were not readily apparent
at 24 hours postinoculatlon under agar overlay. By 48 hours postinoculation
CPE were evident and were somewhat different for each of the 3 cell
cultures (Table 7).
The CPE of the Vero .ce 11 1 i ne consisted of 70 to 90 percent cytolysis·
of the plaque regions with rounding up of the remaining cells. Plaques
became more difficult to detect between the 146th and 150th cell passages
and the cells essentially lost their sensitivity to VEE-virus-induced CPE
after 150 to 155 passages.
Duck embryo cells seemed to be more resistant than the other 2 cell
types to VEE-induced cytolysis. As few as 5 percent and a maximum of 60
percent of the eel ls· within a plaque were lysed due to virus replication
with 20 to 95 percent of the remaining cells rounding up. Rounded up and
morphologically normal cells also had very fine cytoplasmic vacuoles

44
Table 7. Susceptibility of different cell types to GJ9-lBJ VEE virus
Ce 11 Mean titer Suck I ing Cytopathic Mean plaque culturea effectsb culture
Vero
Duck embryo
L eel I
Goat kidney
Human amnion
Bovine turbinate
BHK-21
Horse k I dney
a PFU/ml.
cell
6.4 x 108
2.7 x 108
Negative
.Negative
Negative
Negative
4. 3 x 10 8
Negative
mouse ICLD50
5 x 1010 48 hr, 70-90%
5 x 1010 48 hr, 5-60%
5 x 1010 ------9.1 x 10 10 ------9. 1 x 1010 ------9. 1 x 1010 ------
5 x 1010 48 hr, 30-75%
9. 1 x 1010 ------
bHours postlnoculation, % cells affected in the plaques. cDetermined during final 72 hr postinoculation plaque count.
diameterc
3.7mm
2.7mm
-------------------------3.3mm
------

45
present.
The overall cytopathic effect on BHK-21 cells was not as marked as
that se.en in Vero cells. Cytolysis varied from 10 to 45 percent of the
plaque cell population with 5 to 75 percent rounding up. Cytoplasmic
vacuolization was more marked in BHK-21 cells than that observed in duck
embryo cells and was usually observed in greater than 30 percent of the
plaque cells.
The goat as a sentinel
Serum neutralization (SN, TC-83 VEE virus) antibody titers of 1:10
or greater were detected in of the 6 infected goats at 6 days postin-
oculation (DPI), 2 goats at 7 DPI, 5 goats at 8 DPI, and all goats by 9
DPI. The SN titers in 5 of the 6 goats rose very rapidly and by the
second day of response had increased from 1:10 to 1:100 (Figure 1).
Serum-neutralization testing of the sera with GJ9-1BJ VEE virus did
not detect a serological response until 8 DPI in 1 goat. By 10 DPI 5 of
the 6 goats had 1: 10 titers. The 6th goat seroconverted by 15 DP I, was
negative at 1: 10 for the next 3 days, and regained a 1: lO titer at 19 DPI.
The 5 goats with 1:10 titers at 10 DPI responded in a serologically uniform
manner for the remainder of the 45 day monitoring period (Figure 2).
Maximal SN response with GJ9-1BJ virus was 1:100 in 1 goat as compared
with TC-83 virus SN titers of 1:1600 in 3 goats, and 1:800 in the other 3
goats of the group (Figure 1).
Hemagglutination-inhibition (HI) titers did not appear until 7 DPI
(1 goat). The maximal HI response was 1: 10,240 in 1 goat but another I

Figure 1. Senti·nel goat study. TC-83·SN antibody titers of goats 147, 152, 157, 158, 159, and· 161 between days 6 and 45 postinoculation. Titers listed were the highest serum di 1 ut ion that produced at 1 east 90% p 1 aque reduction'. See Append i'x for specific titers.
47
147 152
157 158
2560 159 161 128 640 020 160
80
40
"' ... 20 I-;:
10
<10
6 16 26 36 45 DAYS POSTINOCULATION

\
Figure 2. Sentinel goat study. GJ9-1BJ SN antibody titers of goats 147, 152, 157, 158, 159, and 161 between days 6 and 45 postinoculati-on. Titers listed were the highest serum dilution that produced at least 90% plaque reduction. See Appendix for specific titers.

50
goat did not exceed a titer of 1:40 during the observation period (Figure
3). The 2 contact 'transmission goats did not display detectable VEE viral
antibodies by the 3 serological assay methods employed in this study.
(See Appendix for titers by HI and SN).
The goat as a silent amplifier of IB VEE virus
Viremia was not detected by the plaque assay technique using BHK-21
cells,, By suckling mouse inoculation 5 of the inoculated goats were
found to be viremic for 1 to 3 days, viremia commencing between the 1st
and 5th days postinoculation. One of these goats was viremic from day 1
to day 3 postinoculation, another from day 2 to day 4, and a 3rd from day
3 to day 4. One goat was viremic on days 3 and 5, and one goat was viremic
only on day 4. Peak viremia did not exceed 4.08 log 10 SMICLD50 per ml of serum. Respective SMICLo50tml of the sera are listed in Table 8.
Oral, nasal, and genitourinary swabs obtained from day zero through day 7
were negative for VEE virus by suckling mouse inoculation.
Clinical evidence of disease in the domestic goat was lacking (Figure
4) during the 21-day observation period with one exception. Goat number
159 displayed an increased body temperature from day 2 through day 10,
which peaked on day 5 at 41.1C (105.9F), The goat 1 s normal body temperature
was 39.lC (102.3F) as determined from baseline data (Table 9),
VEE antiserum production
After intravenous challenge of the 3 groups of goats at 50 days post·
inoculation, the maximum antibody titers as determined by the 3 tests were

Figure 3.
)
Sentinel goat study. HI antibody titers of goats 147, 152, 157, 158, 159 and 161 between days 6 and 45 postinoculation. See Appendix for specific titers.
'
~ w ~
~
52
157
!5120 159
2!560 1280 640 320 160
60
40
20
10
<10
6 16 26 :56 4!5 DAYS POSTINOCULATION
152
108
161
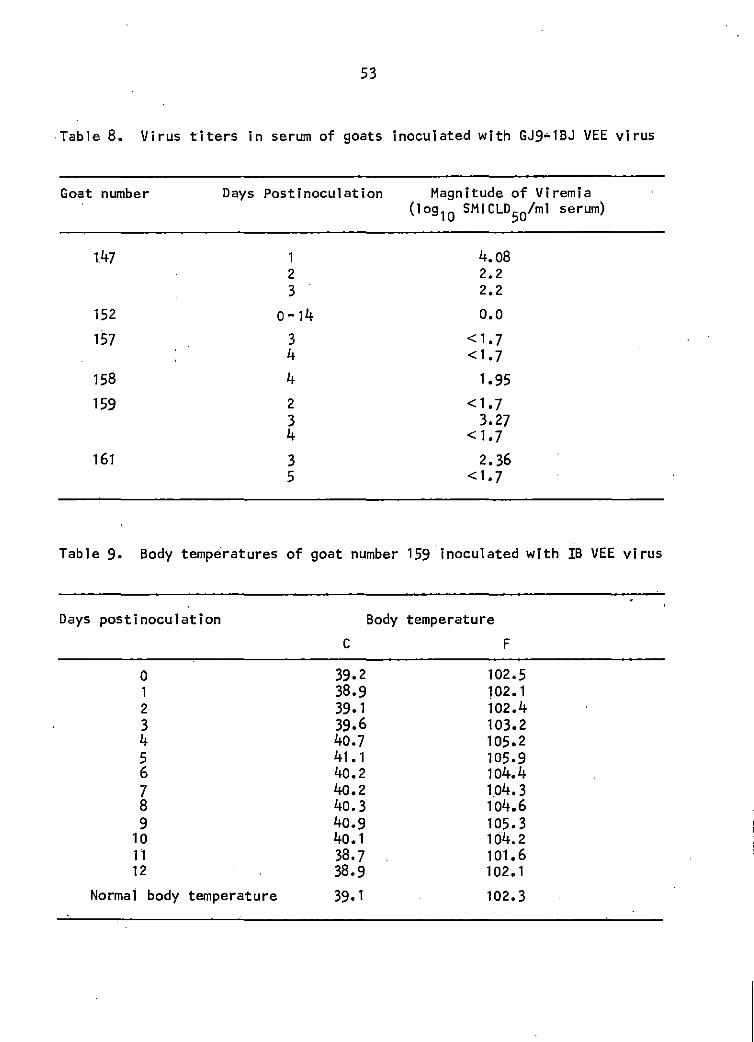
53
Table 8. Virus titers in serum of goats inoculated with GJ9•1BJ VEE virus
Goat number Days Postinoculation Magnitude of Viremia (log 10 SMICLD50tml serum)
147 1 4.08 2 2.2 3 2.2
152 0-14 o.o 157 3 < 1. 7
4 < 1. 7 158 4 1.95 159 2 < 1.7
3 3.27 4 < 1. 7
161 3 2. 36 5 < 1. 7
Table 9. Body temperatures of goat number 159 inoculated with IB VEE virus
Days postinoculation Body temperature
0 1 2 3 4 5 6 7 8 9
10 11 12
Normal body temperature
C F
39.2 38.9 39. 1 39.6 40.7 41.1 40.2 40.2 40.3 40.9 40.1 38.7 38.9 39. 1
102.5 102.1 io2.4 103.2 105.2 105.9 104.4 1.04. 3 104.6 105,3 104. 2 101.6 102. 1 102.3

54
1:51,200 (SN~ TC-83, 7 of 12 goats), 1:1280 (SN, GJ9-1BJ 2 of 12 goats),
and 1:64,000 (HI, 5 of 12 goats). See Appendix for listing of titers from
zero through 90 days postinoculation. Serum from goat number 152 exan-
guinated at 72 DPI was selected for conjugation on the basis of a
GJ9~1BJ SN titer of 1:1000.
Conjugate evaluation
After the QAE-Sephadex conjugate fractions had been concentrated by
pressure dialysis they were examined for specific fluorescence on TC-83
VEE virus-infected BHK-21 cells at 24 and 48 hours postinoculation. Only
that fraction obtained by addition of 0.2 M NaCl demonstrated specific
fluorescence. The fluorescence was finely granular in appearance and
occurred only in the cytoplasm of infected cells (Figures 8, 10, and 11).
Specificity of staining was evaluated by 2 methods. Baby hamster
kidney (BHK-21) cells were infected with approximat~ly 103 PFU per ml
of eastern equine encephalomyelitis or western equine encephalomyelitis
viruses. At 24 hours postinoculation the infected cell cultures were
fixed in acetone at 4C and were examined 1 day later with the conjugate.
Cross-staining resulting in unwanted specific fluorescence did not occur
(Figures 12 and 13).
The second conjugate specificity parameter was the staining of normal
BHK-21 cells 24 hours after they had been subjected to the same regimen as
that employed for infecting cells. Nonspecific staining of normal cells
was not observed (Figure 5).

Ffgure 4. Absence of clinical signs in a domestic goat three days after infectfon with the GJ9-1BJ straf n of Venezuelan equine encephalomyelltis (VEE) virus.
Figure 5. VEE conjugate applied to normal BHK-21 cell monolayer. Note absence of specific fluorescence. 125x
r

56

Figure 6. VEE conjugate applied to GJ9-1BJ VEE-virus-infected BHK-21 cell monolayer at 24 hours postinoculation. Note specific perinuclear and cytoplasmic fluoresence. 125x
Figure 7. Normal conjugate applied to GJ9-1BJ VEE-virus-infected BHK-21 cell monolayer at 24 hours postinoculation. Note absence of specific fluorescence. 125x
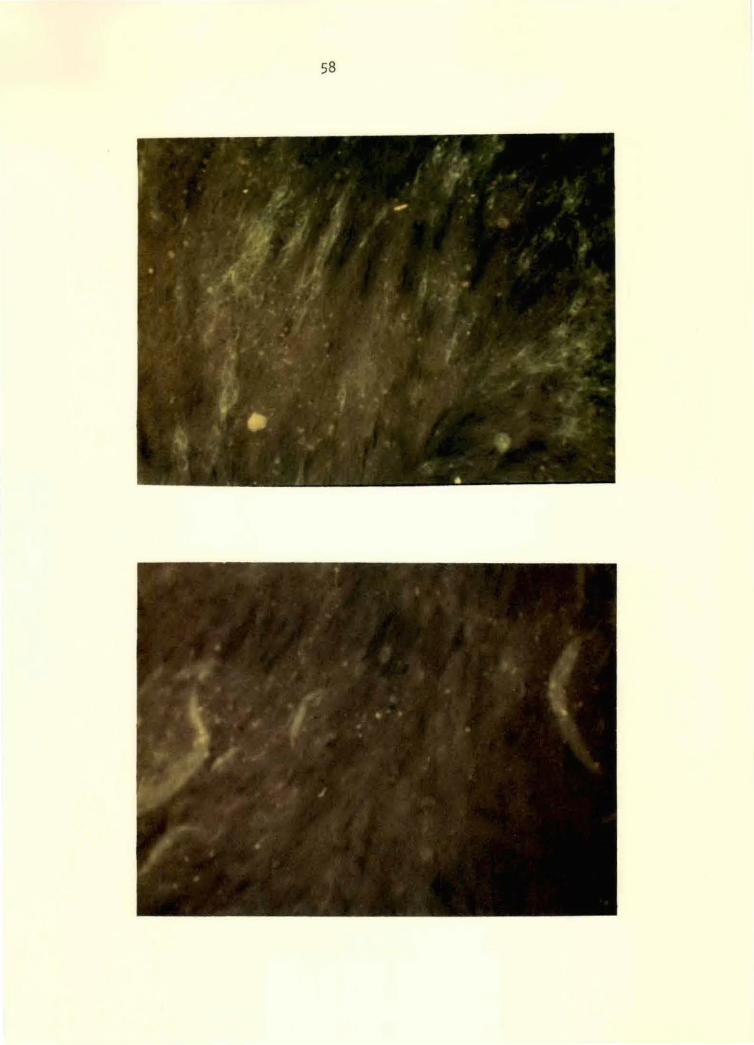
58

Ffgure 8. VEE conjugate applied to TC-83-VEE-virus-infected BHK-21 cell monolayer at 24 hours posti nocul ation. Note specific, granular perlnuclear and cytoplasmic fluorescence. 500x
Figure 9. VEE conjugate app lied to GJ9-1BJ-VEE-virus-infected BHK-21 cell mono l ayer at 24 hours postinoculation. Note specific, granular perinuclear f luorescence. 1188x
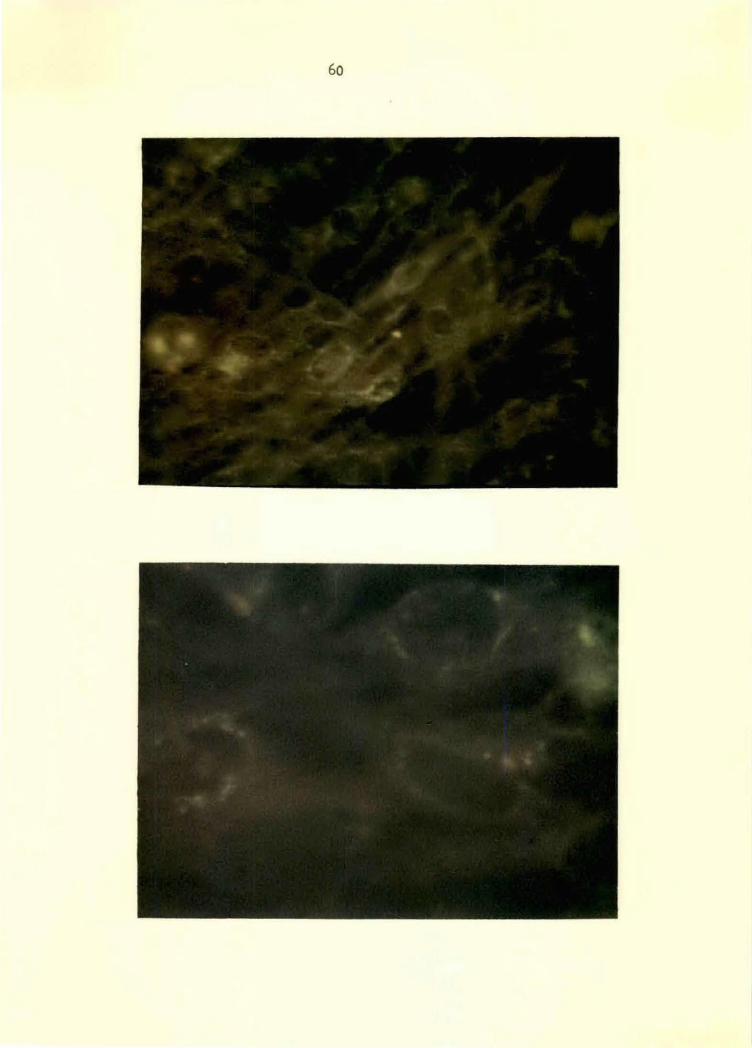
60

Figure 10. VEE conjugate applied to TC-83-VEE-virus-infected BHK-21 cell monolayer at 48 hours postinoculation. Note spec i fic cytoplasmic fluorescence of rounded up cells. 125x
Figure 11. VEE conjugate applied to TC-83-VEE-virus-infected BHK-21 cell monolayer at 48 hours postinoculation. Note specific, granular cytoplasmic fluorescence of rounded up cells. SOOx
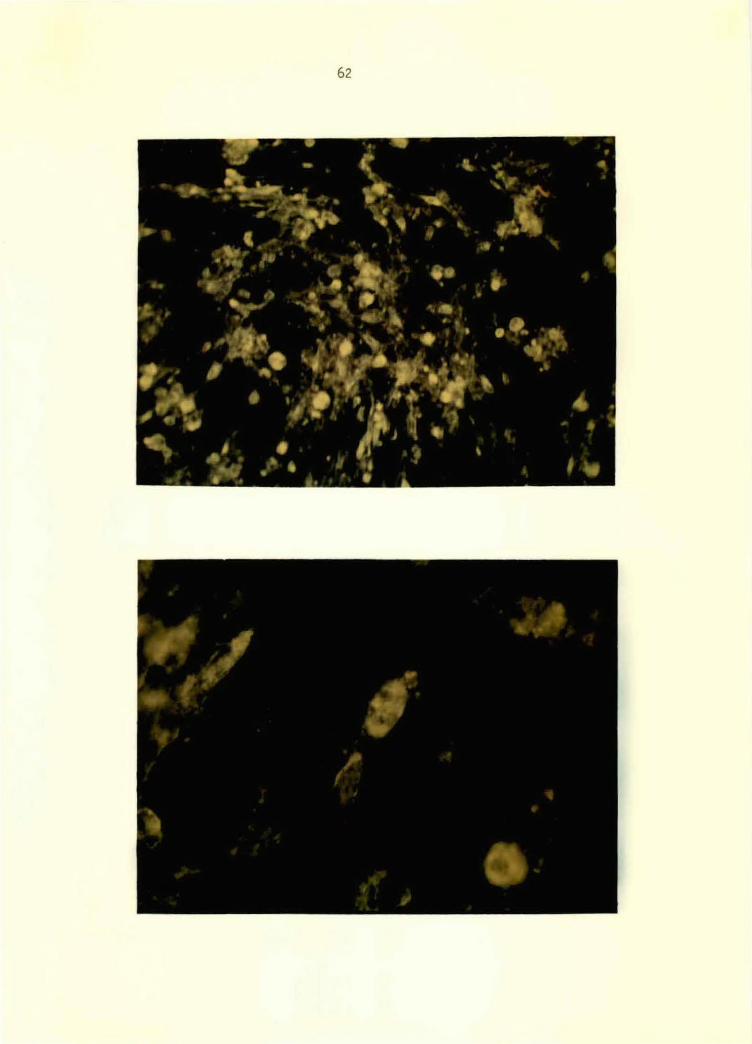
62
- ..... .. ... '· .
:-.. . • .. !...r • . •1 .. .
~
·~"' • ... . ~ .. t •" I 4 ,.
.:t. ..,, # " • •
• • .. "" ...
' I 9 ,. • • :-• ~ '
~· • t
' • • ' ' • ,,,, ·. . • ,, ., .\ I! ' ). ' • • • ; •
;~. "' ' "' ' • ,, •,
4t I
• • ,..

Figure 12.
Figure 13.
VEE conjugate applied to western equine encephalomyelitis virus-infected BHK-21 cell monolayer at 24 hours postin-oculation. Note absence of cross-sta ining. 125x
VEE conjugate applied to eastern equine encephalomyelitis virus-infected BHK-21 cell monolayer at 24 hours postin-oculatton. Note absence of cross-staining. 125x

64

65
When the direct 1 y-tagged conjugate prepa_rat ion was used to stain
TC-83 virus-infected BHK-21 cells marked nonspecific staining was
observed. Dilution of the conjugate beyond 1:8 was not feasible due to
markedly diminished specific staining and at the 1:8 dilution the level
of nonspecific fluorescence was not tolerable. On that basis further
fluorescent antibody studies on VEE virus were done with Q.AE-Sephadex-
treated conjugate.
The n<irma 1 -goat serum conjugate was prepared from Sephadex G-200
fractionated lgG which was tagged with FITC and purified by Q.AE-
Sephadex column chromatography. The working dilution of 1:4 was
derived by colll'arison of background fluorescence of both the normal
and VEE conjugates when VEE virus-infected and normal BHK-21 cell
monolayers were stained (Figures 5, 6, and 7). This conjugate
was then employed as a specificity of staining control when conducting
fluorescent antibody tests (FAT) on VEE virus-infected pony tissue
suspensions.
Comparison of sensitivity of FAT to suckling
mouse inoculation for virus identification
Initially the 2 stock virus preparations employed in this study were
assaye_d by the fluorescent antibody test and intracerebral inoculation of
suckling mice. Fluorescent antiboqy assay in BHK-21 cells was found to
have approximately 1.0 log 10 per ml greater sensitivity.
When 20 viremic pony tissues were tested by the 2 systems suckling
mice were the most sensitive virus detection system. The threshold of

66
sensitivity of the fluorescent antibody test appeared to be between 1.6
and 2.0 log 10 SMICLD50/ml of 10 percent pony t.issue suspension. By
FAT all tissue suspensions were positive by 24 hours postinoculation
with the exception of 4 of 6 tissues in the 2.0 to 2.8 log10 SMICLD501ml
virus titer range which were positive at 48 hours postinoculation. Two
tissues with a titer of 1.6 log 10 SMICLD50tml were negative at 24 and 48
hours postlnoculatlon.by the fluorescent antibody test (Table 10).
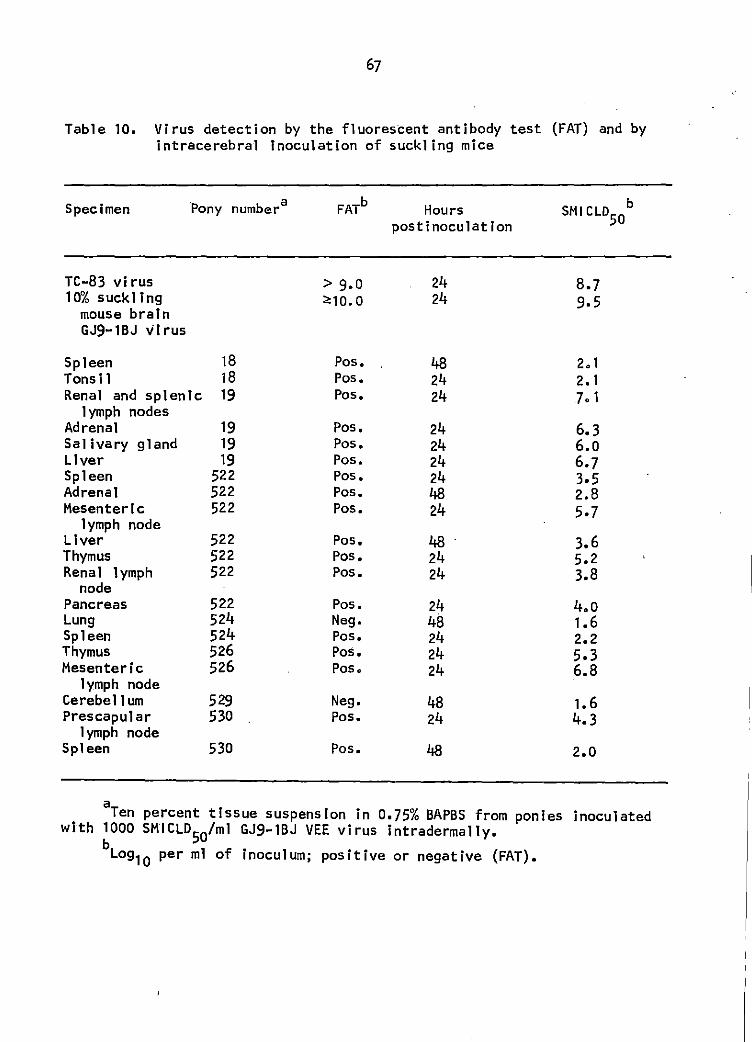
Table 10. Virus detection by the fluorescent antibody test (FAT) and by intracerebral inoculation of suckling mice
Specimen . a Pony number
TC-83 virus 10% suckling
mouse brain GJ9-1BJ virus
Spleen 18 Tons i 1 18 Renal and splenlc 19
lymph nodes Adrenal 19 Salivary gland 19 Liver 19 Spleen 522 Adrenal 522 Mesenteric 522
lymph node Liver 522 Thymus 522 Renal lymph 522
node Pancreas 522 Lung 524 Spleen 524 Thymus 526 Mesenteric 526
lymph node Cerebellum 529 Prescapular 530
lymph node Sp 1 een 530
FATb Hours post inoculation
> 9.0 24 :i:10.o 24
Pos. 48 Pas, 24 Pos, 24
Pos. 24 Pos, 24 Pas. 24 Pos. 24 Pas. 48 Pas. 24
Pos. 48 Pos. 24 Pas. 24
Pas. 24 Neg. 48 Pas. 24 Pos. 24 Pas. 24
Neg. 48 Pas. 24
Pos. 48
b SMI CLo50
8.7 9,5
2. 1 2. 1 7. 1
6.3 6.o 6.7 3,5 2.8 5,7
3,6 5,2 3.8
4.o 1. 6 2.2 5,3 6.8
1. 6 4.3
2.0
with aTen percent tissue suspension in 0.753 BAPBS from ponies inoculated 1000 SMI CLD50/ml GJ9-1BJ VEE virus intradermal ly. bLog10 per ml of inoculum; positive or negative (FAT).

68
DIS CUSS I ON
The baby hamster kidney (BHK-21) cell line has been proven superior
to either primary duck embryo or Vero African green monkey cell lines for
titration of Venezuelan equine encephalomyelitis (VEE) virus. The Vero
cell line is slightly more sensitive for the titration of the GJ9-1BJ
strain of VEE virus but It lacks the potential for continuous serial
passage that the BHK-21 line has as supplied from the American Type
Culture Collection. Baby hamster kidney cells have been passaged in
excess of 200 times at the National Animal Disease Laboratory, Ames, Iowa,
without loss of sensitivity for VEE virus. In contrast, the Vero cell
line is supplied at the 122nd passage level and has been found in this
laboratory to lose its susceptibility to VEE virus between the 150th
and 155th passage level. Earley, et~· (34) also did not find Vero cells
to be sufficiently susceptible after 30 serial passages. Thus, the time
saved by not having to restart the cell line from the 122nd passage level
every three months more than justifies the choice of the BHK-21 cell line
over the Vero cell 1 ine.
Primary duck embryo cell cultures are slightly less sensitive for the
detection of the epizoodemlc strain of VEE virus than BHK-21 cells. The
mean VEE plaque diameter in the duck cells is also 1.0 mm less than in
BHK-21 cells which makes enumeration of plaque-forming units more difficult.
The BHK-21 cell line is thus the cell culture of choice of the 8 evaluated
in this study.

Experimental lnoC:u.lation of the goat with the minimal exposure of
1000 SMICLD 50, an average mosquito inoculum, revealed the goat to be·an
excellent sentinel for VEE virus activity. The 6 inoculated goats
developed titers greater than 1:10 as determined by hemagglutionation-
inhlbltlon (HI) and serum neutralization (SN) tests. Both persisted in
excess of 40 days. In view of these findings the investigation of
Dickerman, ~ ~· (33) would seem to have provided questionable information
on the bovine as a possible sentinel species. In that· study 8 native HI
and SN seronegative Colombian bovines were inoculated with a great excess
of IB VEE virus, 7,1 log 10 primary chicken embryo cell. plaque-forming
units (CEC-PFU), to "insure establishment of infection." The 7.1 log1·0 CEC-PFU of virus would be roughly equivalent to 9 log 10 SMICLD50 •
Considering an average mosquito inoculum of 1000 SMICLD50, approximately
1,000,000 average IB VEE viremic mosquitoes would have had to feed upon ' a single animal over a given period. In view of the Information obtai.ned
herei.n with another domestic ruminant species, the goat, it would appear
necessary to repeat that study with a lower inoculum of IB VEE virus.
However, sufficient numbers of goats must be available to the
potential vector to be able to detect VEE virus activity in an area.
Hayes, et ~· (61) have demonstrated in a moquito host preference study
in Hale County, Texas, that the predominant feeding habit of mosquitoes
of the genera Aedes, Anopheles, Culiseta, and Psorophora is to feed
upon mammalian species. Sudla (115) has stated that only Psorophora
confinnis and Aedes taeniorhynchus have been proven as possible vectors

70
of the IB strain of VEE virus. He has also reported that 83 virus
isolates compatible with VEE were obtained from Psorophora confinnis
during the Texas epizoodemic (117). The bionomics off· confinnis have
been shown to be associated with temporary fresh water pools located in
sunny, grassy areas (115). Such areas can often be found in livestock
pastures. Therefore, in regions where large goat populations are present
the likelihood of the goat being a-sentinel of epizoodemic VEE virus
activity would appear to be very good.
Viremic goats on the other hand, do not offer a source of silent
amplification of epizoodemic VEE virus according to the results of this
study. Goat viremia did not exceed 4.08 log 10 SMICLD50!ml, with a mean
peak viremia of 2.67 log10 SMICLD50/ml. The average viremia required to
infect vector species of mosquitoes has been stated by Chamberlain (14,
16) to be 5.0 log 10 SMICi.o50/ml. Experimentally the Infection threshold
of Psorophora confinnis has been found to be 4.9 to 5.2 log10 SMICLD50/ml
(115). Consequently, the goat is not likely to be an amplification
mechanism of ep i zoodem i c VEE virus.
Due to the excellent serologic response of the goats inoculated as
sentinels they were hyperimmunized in an attempt to produce a more specific
VEE antiserum than that which Was likely produced by the goats given
multiple virus injections to stimulate antibody formation. The sero-
logical response of the 4 antiserum goats was no better than that of the
goats of the sentinel study given only one injection of virus. The serum
of goat number 152 was chosen for conjugate preparation because of its
high.antibody titer as determined by the GJ9-1BJ SN test. Serological

71
testing by the other two techniques, HI and TC-83 SN later revealed an
obvious lack of correlation of serum titers. The most plausible
explanation of such differences is the presence of residual infectivity
representing nonneutralized virus. Several workers have reported a
similar phenomenon of nonneutralized virus in other virus-serum
neutralization test systems. Ashe and Notkins (5), Bradish, et ~· (10),
and Wall is and Melnick (127) have theorized residual infectivity to be
due to the formation of infectious virus antibody complexes which resist
further neutralization by antibody. Lafferty (78) used kinetic serum
neutralization testing to show that residual infectivity was due to
lack of antibody avidity. Fazekas 'de St. Groth, et~· (38) set forth
the concept that residual infectivity was due to dissociation of the
virus-antibody complex. Through extensive kinetic serum neutralization
testing with VEE virus Hahon (58,59) has found that resid.ual infectivity
of VEE virus is not due to dissociation of noninfective antigen-antibody
complexes, to the presence of a genetically stable persistent viral
subpopulation, or to the presence of free infectious ribonucleic acid in
the virus preparation used for the serum neutralization test. His
experiments with TC-'83 and Trinidad IA strains of VEE virus (58) have
clearly demonstrated 2 factors contributing to VEE virus SN residual
infectivity. One factor is the formation of viral aggregates of the virus
preparation employed in the test system resulting in incomplete neutraliza-,
tion. Hahon•s main concept was, however, that residual infectivity was
due to the presence of sensitized VEE virions that were not neutralized '.

72
by the antiserum employed in the SN test. Supporting evidence for such a
theory was obtained when neutralized suspensions were incubat~d at 35C
for 1 hour with a 1: 10 dilution of anti-lgG serum. Addition of the anti-
lgG serum to a neutralized suspension yet containing 5.4 log10 units of
residual virulent IA VEE virus infectivity further reduced viral in-
fectivity of the suspension by an additional 3.7 log10 units. Similar
work has al!\o been done with lactic dehydrogenase virus and pol iovirus
(73,91,92).
A third component of Hahon's study was the determination of residual
infectivity of Trinidad IA VEE virus as compared to the TC-83 attenuated
strain. The percentage of original lnfectivity remaining after serum
neutralization with a 1:50 dilution' of monkey VEE antiserum to Trinidad
IA VEE virus was approximately 20% as compared to 2.2"/o residual activity
for TC-83 VEE virus after 15 minutes incubation at 35C. Clearly, the low
titers obtained with. the GJ9'-1BJ SN test as compared to those of the
TC-83 SN tests would·appear to have been the result of residual infectivity
of the virulent IS.strain of virus. The minor amount of residual infec-
tivity of the TC-83 SN test system made it possible to obtain good
correlation with the VEE HI test. The information subsequently derived
from the TC-83 SN and VEE HI tests indicated that several goats could
have served as .a source for high-ti tered anti sera for conjugation.
The fluorescent antibody conjugate prepared from the serum harvested
from goat number 152 22 days after intravenous challenge With 25,000

73
SMICL050 of GJ9-1BJ VEE v.irus was determined to b.e a highly specific
preparation. Although antigenic interrelationships with eastern equine
encephalomyel it is (EEE) and western equine encephalomyel it is (WEE)
viruses were detected by EEE and WEE HI tests, they could not be demon-
strated with the fluorescent antibody test.
Pooled normal goat serum was used to prepare a normal serum conjugate
for use as a VEE conjugate specificity of staining controi. When the VEE
conjugate was used on the BHK-21 cell line in a fluorescent antibody test,
as little virus as 2.1 log10 SMICLD50to.1 ml inoculum from GJ9-1BJ VEE-
virus-infected .pony tissue suspensions could be detected. If the Leighton
tube inoculum would have been increased to 1.0 ml of tissue suspension·
the sensitivity of the VEE fluorescent antibody test (FAT) would have
likely been within 1 log10 dilution of that of intracerebral inoculation
of 2- to 4-day-old suckling mice. The smaller inoculum was chosen for
use in FAT to make it a more rapid diagnostic test. If the 1 arger
inoculum·had been employed at least two washings with tissue culture
medium would have been required to remove toxic tissue components from
the BHK-21 monolayer after the 1 hour virus adsorption period, An
additional factor to be considered was that of personnel safety. The
use of a smaller inoculum with no washing of monolayers decreased the
possibility of aerosol exposure to laboratory personnel.

74
SUMMARY
The baby hamster kidney (BHK-21) cell line was shown to be superior
to either primary duck embryo cells or Vero African green monkey cells
as a host system for propagation of the epizoodemi·c strain, GJ9-1BJ, of
Venezuelan equine. encephalomyelitis (VEE) virus. For that reason the
BHK-21 cell line was chosen for evaluation of VEE fluorescent-antibody
conjugates.
Surveillance of possible VEE virus activity is of critical importance
in avoidance of major epizoodemics. The goat has been evaluated as a
sentinel for IB VEE virus activity in this study and was found to be a
very sensitive sentinel animal. Six goats of mixed sex from 4 to ·18
months of age when exposed to a single average mosquito inoculum of VEE .. . virus began to seroconvert within 6 days. Serological evaluation using
the TC-83 and GJ9-1BJ serum neutralization tests and the VEE hemagglutina:..
tion-inhibition test showed that the 6 goats were seropositive for VEE
antibody for 49 days. A minimum of VEE virus thus stimulated a very
adequate serological response in the goat.
Following challenge of the sentinel goats with 25,000 SMICLo50 of
the GJ9-1BJ strain of VEE virus a suitable serum for conjugation was
obtained. The VEE antiserum and normal goat serums were conjugated and
evaluated for staining specificity in the BHK-21 cell line. No cross-
staining of the VEE conjugate with eastern equine encephalomyel it is virus
or western equine encephalomyelltis virus was observed despite a 1:10
cross reaction for both vi r.uses as detected by EEE and WEE hemagglutina-
tion-inhibition testing of the same serum.

75
The conjugate was assayed for sensitivity for the detection of VEE
virus in cell cultures inoculated with equine tissue suspensions. The
(BHK-21) cell line when combined· with a conjugate highly specific for VEE
virus has been proven to be nearly as sensitive as intracerebral inoculation
of the suckling mouse for the detection or identification of VEE virus.
In a diagnostic sense the test can now be applied to routine screening of
VEE diagnostic specimens. All negative specimens should then be tested in
suckling mice by the intracerebral route for isolation of VEE virus not
detected by the fluorescent antibody test. The fluorescent ant.ibody test·
wou.ld be of greatest value in the event of an another outbreak of
eplzoodemic VEE among the equidae of the United States. Large numbers
of specimens likely to be positive for VEE virus would be encountered and
a rapid, efficient technique such as the fluorescent antibody test would
then be of critical importance.

76
APPENDIX
TRIS Buffered Saline pH 8.0
1. O. SM TR IS:
Dissolve 60.57 g TRIS and 4.o g sodium azide (NaN3) in 200 ml triple-distilled, deionized water and q,s. to 1000 ml.
2. o.sM HCl:
q.s. 42 ml of 11.9 M HCl to 1000 ml.
3. Buffer preparation:
a. two parts TRIS (133.3 ml) and one part HCI (66.6 ml) are mixed.
b. add. 1.19 g of NaCl to 200 ml of TRIS buffered saline for a final NaCl concentration of O.lM.
c. add 4.0 g NaN 3 per liter of final buffer.

77
0.5 M Carbonate-Bicarbonate Buffer
1 • Reagents required:
a, 0.5 M solution of sodium bicarbonate (NaHC0 3)
b, 0.5 M solution of sodium carbonate (Na2co3)
2. Mix 50 volumes reagent a with 4.5 volumes reagent b (pH should be 9.0 when di luted 10 times in triple-disti 1 led water},

78
0,1 M Phosphate-Buffered .Saline
pH 7. 2:
1. Prepare stock solutions a and b:
a, 0.2 M monobasic sodium phosphate (NaH2Po4) Dissolve 27.8 g NaH 2Po4 in 1000 ml distilled water.
b. 0.2 M dibaslc sodium phosphate (Na2HP04) Dissolve 53.65 g Na2HP04 · 7H2o in 1000 ml distilled water.
2. Mix 28.0 ml of Solution a with 72.0 ml of solution b. Di 1 ute to a total of 200 ml with distilled water.
pH 6.8:
1. Prepare stock solutions a and b as· shown.
2. Mix 51.0 ml of solution a with 49.0 ml of solution b. Di 1 ute to a total of 200 ml with distil led water.

79
Dextrose-Gelatin-Verona! Buffer
1. Dissolve 0.58 g 5,5-diethyl barbiturate acid and 0.6 g gelatin in 250 ml distilled water by heating.
2. Combine the following reagents with solution 1:
Sodium 5,5-diethyl-barbiturate CaC1 2 (anhydrous) MgS04 • 7H 20 NaCl Dextrose
3. q.s. to 1000 ml with distilled water.
0.38 g 0.02 g 0.12 g 8.50 g
10.00 g
\

80
0.2% Bovalbumin, pH 6.o Borate Saline (BBS)
1. Prepare pH 9, 0 borate saline as fol lows:
a. mix 80 ml of 1.5 M NaCl, 100 ml of 0.5 M H3Bo3, and 24.0 ml of 1.0 M NaOH.
b. q,s. to 1000 ml with distilled water.
2, Add Cohn fraction V bovine serum albumin to borate saline (1) to a concentration of o.4%.
3. Prepare 0.15 M NaCl-0.2 M Na 2HP04 as follows:
a. mix 100 ml of 1.5 M NaCl and 100 ml of 2.0 M Na 2HP04 b. q,s, to 1000 ml with distilled water.
4. Prepare 0.15 M NaCJ-0.2 M NaH/04 as fol lows:
a, mix 100 ml of 1.5 M NaCl and 100 ml of 2.0 M NaH2Po4 b. q.s. to 1000 ml with disti 1 led water.
5, Mix 11 ml of solution 3 with 89 ml of solution 4.
6. · Mix equal volumes of solutions 2 and 5, check pH, and BBS is then ready for use.

81
Veronal Buffer (VBD)
1. Dissolve 42.5 g NaCl ahd 1.87 g sodium 5,5-diethyl-barbiturate · in 700 ml distilled water.
2. Dissolve 2.875 g 5,5-diethyl barbiturate acid in 250 ml hot distilled water.
3. Dissolve 20,333 g MgC1 2 (1.0 M) and 4.411 g CaC1 2 (0.3 M) in 1000 ml distil led water.
4. 5 x VBD:
a. Mix solutions 1 and 2 and cool to room temperature.
b. Add 2.5 ml of solution 3 to a and q.s. to 1000 ml with distilled water.
c. To use, di 1 ute 1: 5 and add 1.0 g gelatin per 1000 ml of final diluent.
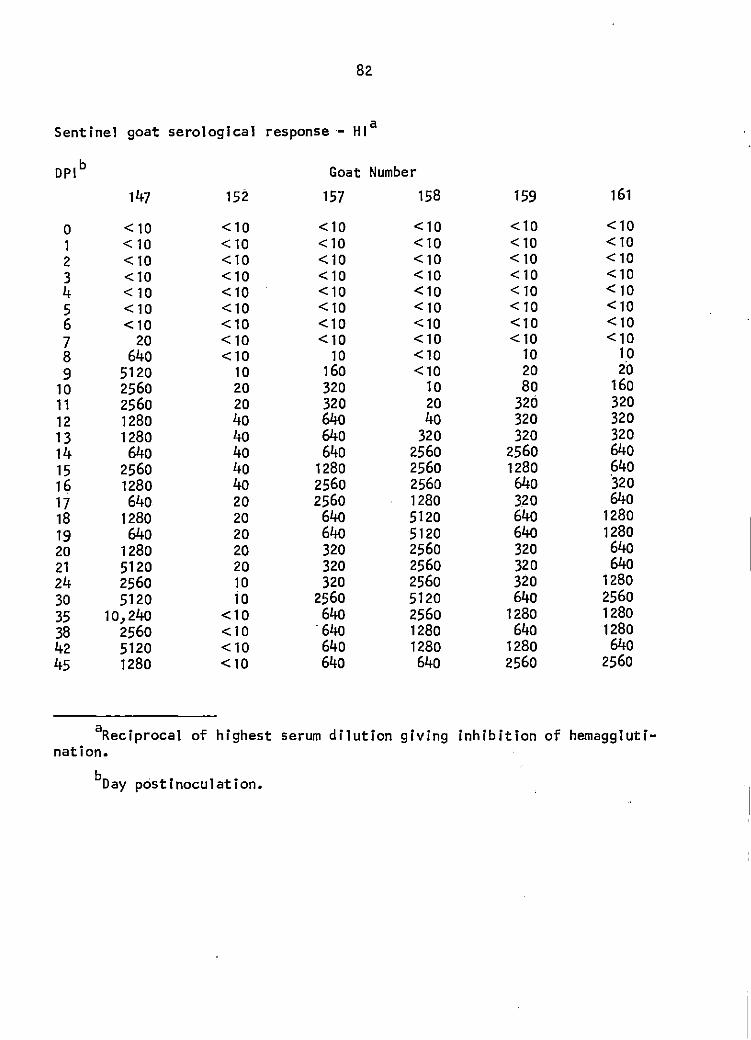
82
Sentinel goat serological response·- Hla
DPI b Goat Number 147 152 157 158 159 161
0 < 10 <10 <10 <10 <10 <10 1 < 10 <10 <10 <10 <10 <10 2 <10 <10 <10 <10 <10 <10 3 <10 <10 <10 <10 <10 < 10 4 <10 <10 <10 <10 <10 <10 5 <10 <10 <10 <10 <10 <10 6 <10 < 10 <10 <10 <10 <10 7 20 <10 <10 <10 < 10 <10 8 640 <10 10 <10 10 10 9 5120 10 160 <10 20 20
10 2560 20 320 10 80 160 11 2560 20 320 20 320 320 12 1280 40 640 40 320 320 13 1280 40 640 320 320 320 14 640 40 640 2560 2560 640 15 2560 40 1280 2560 1280 640 16 1280 40 2560 2560 640 320 17 640 20 2560 1280 320 640 18 1280 20 640 5120 640 1280 19 640 20 640 5120 640 1280 20 1280 20 320 2560 320 640 21 5120 20 320 2560 320 64o 24 2560 10 320 2560 320 1280 30 5120 10 2560 5120 64o 2560 35 10,240 <10 640 2560 1280 1280 38 2560 < 10 . 640 1280 640 1280 42 5120 <10 640 1280 1280 640 45 1280 < 10 640 640 2560 2560
aReciprocal of highest serum dilution giving inhibition of hemaggluti-nation.
bDay postinoculation.
I I
·.I !
I
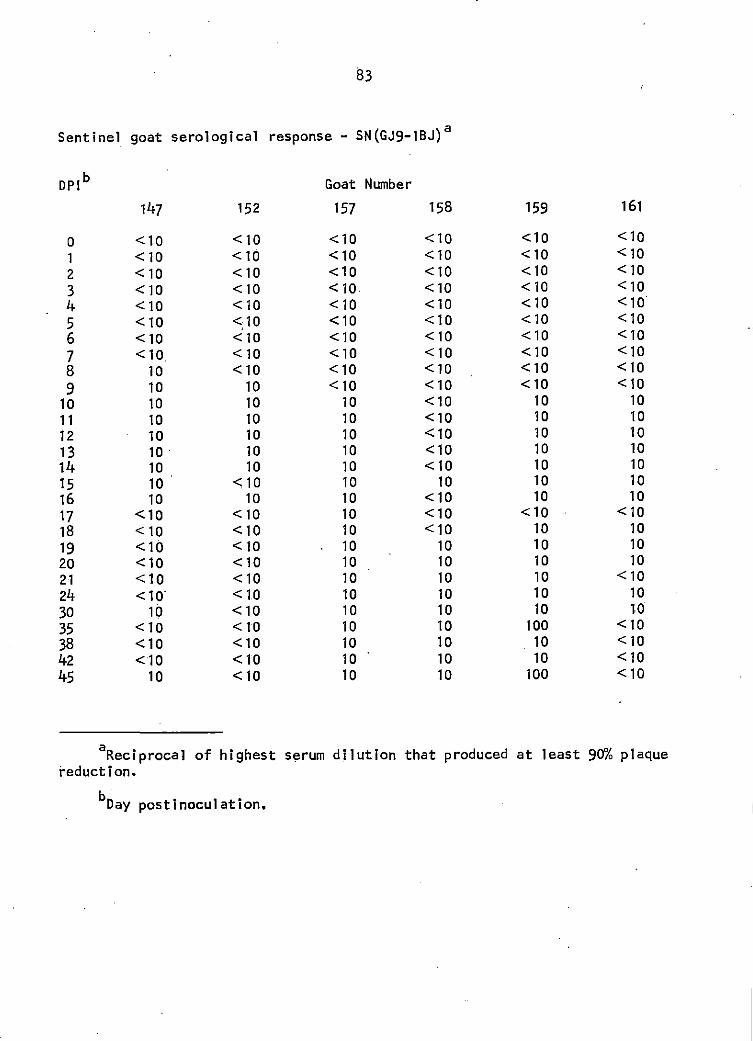
83
Sentinel goat serological response - SN(GJ9-1BJ)a
DP lb Goat Number 147 152 157 158 159 161
0 <10 <10 < 10 <10 <10 <10 1 <10 < 10 <10 < 10 <10 <10 2 < 10 < 10 <10 <10 < 10 < 10 3 <10 <10 <10 <10 < 10 <10 4 <10 <10 < 10 < 10 <10 <10 5 <10 <,10 < 10 <10 <10 <10 6 <10 <10 <10 <10 < 10 <10 7 <10 <10 <10 < 10 < 10 <10 8 10 <10 <10 <10 <10 <10 9 10 10 <10 <10 <10 < 10
10 10 10 10 < 10 10 10 11 10 10 10 <10 10 10 12 10 10 10 <10 10 10 13 10 10 10 <10 10 10 14 10 10 10 <10 10 10 15 10 < 10 10 10 10 10 16 10 10 10 <10 10 10 17 <10 <10 10 <10 <10 < 10 18 <10 <10 10 <10 10 10 19 <10 <10 10 10 10 10 20 <10 <10 10 10 10 10 21 <10 <10 10 10 10 <10 24 <10' < 10 10 10 10 10 30 10 <10 10 10 10 10 35 <10 < 10 10 10 100 <10 38 <10 <10 10 10 10 <10 42 <10 <10 10 10 10 <10 45 10 <10 10 10 100 <10
aReciprocal of highest serum dilution that produced at least 90% plaque reduction.
bDay postinoculation.
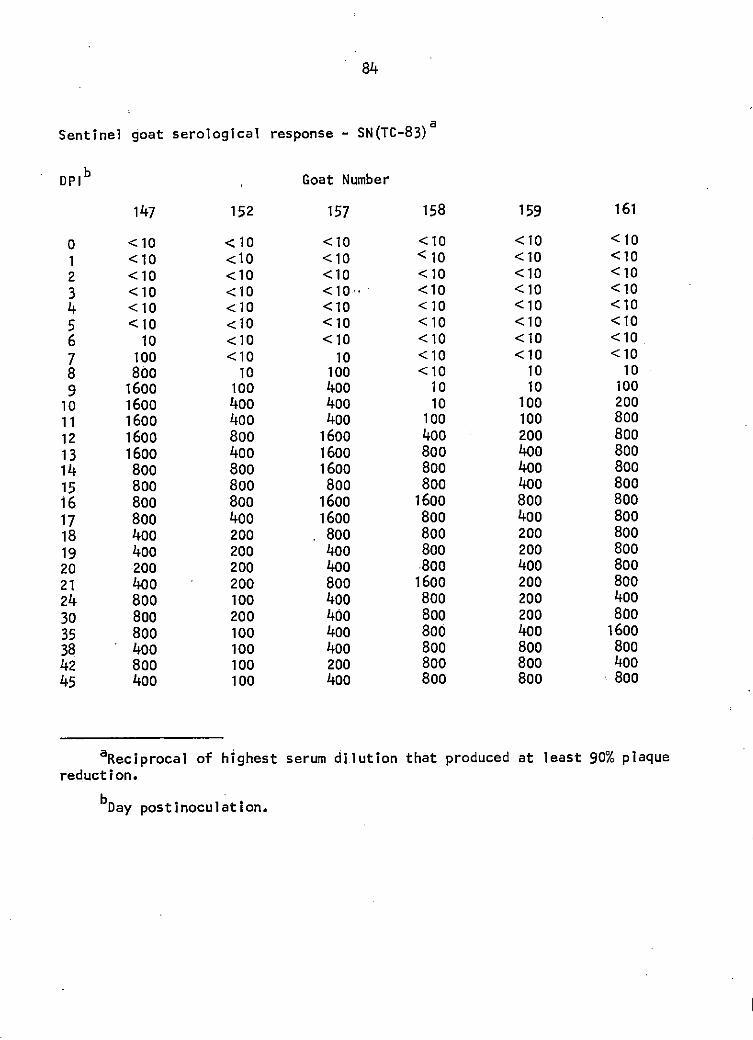
84
Sentinel goat serological response - SN(TC-83)a
DP lb Goat Number
147 152 157 158 159 161
D <10 < 10 < 10 < 10 <10 < 10 1 <10 <10 <10 <10 <10 < 10 2 < 10 < 10 < 10 <10 <10 <JO 3 <10 < 10 < 10 .. <10 < 10 <10 4 <10 <10 <10 <10 <10 < 10 5 <10 <10 < 10 <10 <10 <10 6 10 <10 < 10 <10 <10 <10 7 100 <10 10 <10 < 10 <10 8 800 10 100 <10 10 10 9 1600 100 400 10 10 100
10 1600 400 400 10 100 200 11 1600 400 400 100 100 800 12 1600 800 1600 400 200 800 13 1600 400 1600 800 400 800 14 800 800 1600 800 400 800 15 800 800 800 800 400 800 16 800 800 1600 1600 800 800 17 800 400 1600 800 400 800 18 400 200 800 800 200 800 19 400 200 400 800 200 800 20 200 200 400 800 400 800 21 400 200 800 1600 200 800 24 800 100 400 800 200 400 30 800 200 400 800 200 800 35 800 100 400 800 4oo 1600 38 400 100 400 800 800 800 42 800 100 200 Boo 800 400 45 400 100 400 800 800 . 800
aReciprocal of highest serum di.lution that produced at least 90% plaque reduction.
bDay postinoculation.
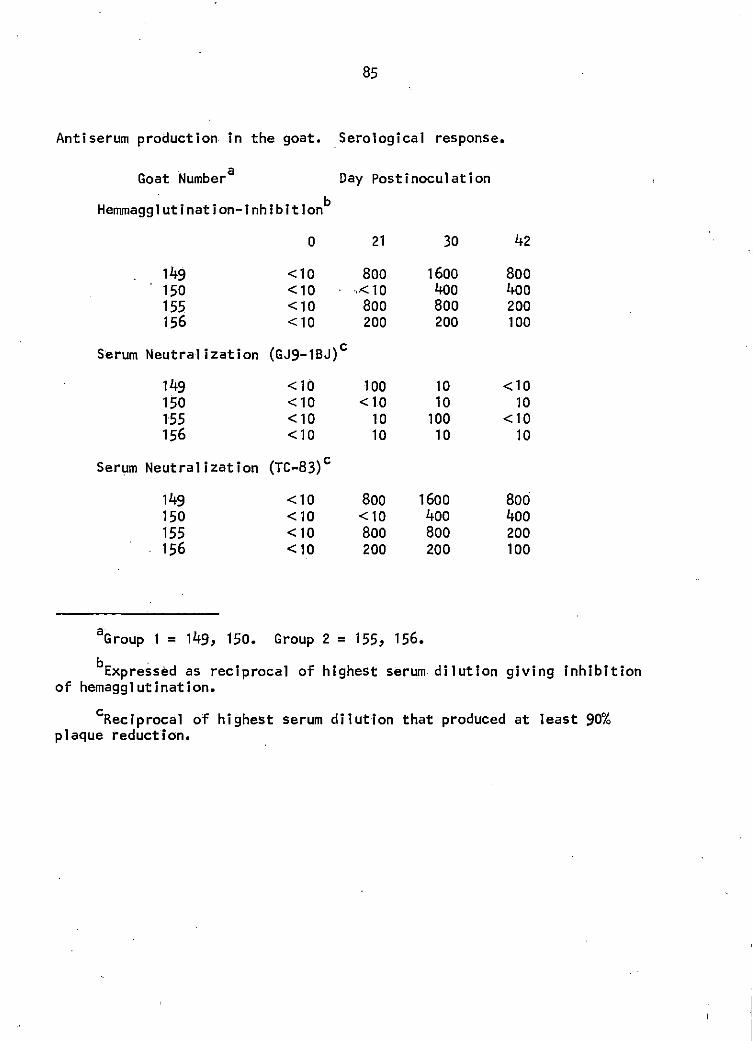
B5
Antiserum production in the goat. Serological response.
Goat Number a Day Postinoculation
Hernrnagglutination-inhibition b
0 21 30 42
149 <10 Boo 1600 Boo 150 <10 -.. <10 4oo 400 155 <10 Boo Boo 200 156 < 10 200 200 100
Serum Neutralization (GJ9-1BJ)c
149 < 10 100 10 <10 150 <10 <10 10 10 155 <10 10 100 <10 156 < 10 10 10 10
Serum Neutralization (TC-B3) c
149 <10 Boo 1600 8oci 150 <10 <10 400 400 155 <10 800 800 200 156 <10 200 200 100
aGroup 1 = 149, 150. Group 2 = 155, 156.
bExpressed as reciprocal of highest serum dilution giving inhibition of hemagglutination.
cReciprocal of highest serum dilution that produced at least 90% plaque reduction.
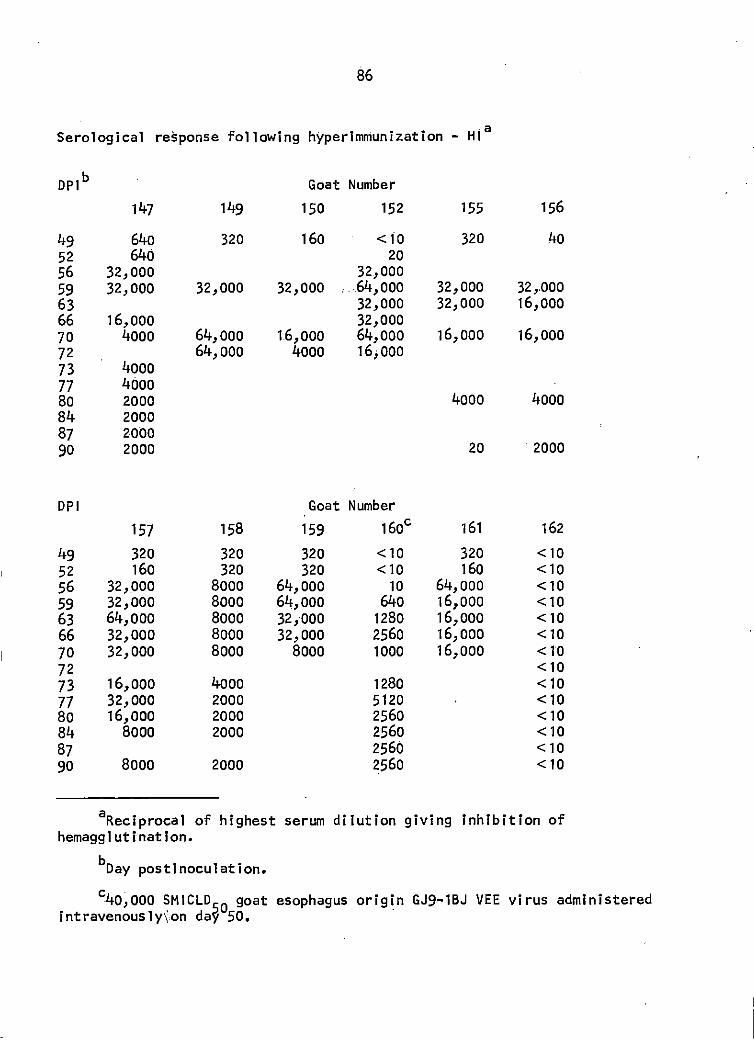
86
Serological response following hyperimmunization - Hla
DP lb Goat Number 147 149 150 152 155 156
49 640 320 160 < 10 320 40 52 64Ci 20 56 32,000 32,000 59 32,000 32,000 32,000 64,ooo 32,000 32,000 63 32,000 32,000 16,000 66 16,000 32,000 70 4000 64,ooo 16,000 64,ooo 16, 000 16,000 72 64,000 4000 16,000 73 4000 77 4Cioo 80 2000 4000 4000 84 2000 87 2000 90 2000 20 . 2000
DPI Goat Number 157 158 159 160c 161 162
49 320 320 320 < 10 320 <10 52 160 320 320 <10 160 <10 56 32,000 8000 64,ooo 10 64,ooo < 10 59 32,000 8000 64,ooo 640 16,000 <10 63 64,ooo 8000 32,000 1280 16,000 < 10 66 32,000 8000 32,000 2560 16,000 <10 70 32,000 8000 8000 1000 16,000 < 10 72 <10 73 16,.000 4000 1280 <10 77 32,000 2000 5120 <10 80 16,000 2000 2560 <10 84 8000 2000 2560 <10 87 2560 <10 90 8000 2000 2560 <10
aReciprocal of highest serum dilution giving inhibition of hemagglutination.
b Day postinoculation. c . GJ9-1BJ VEE administered 40,000 SMICLD O goat esophagus orig in virus
intravenously\on da9 50.
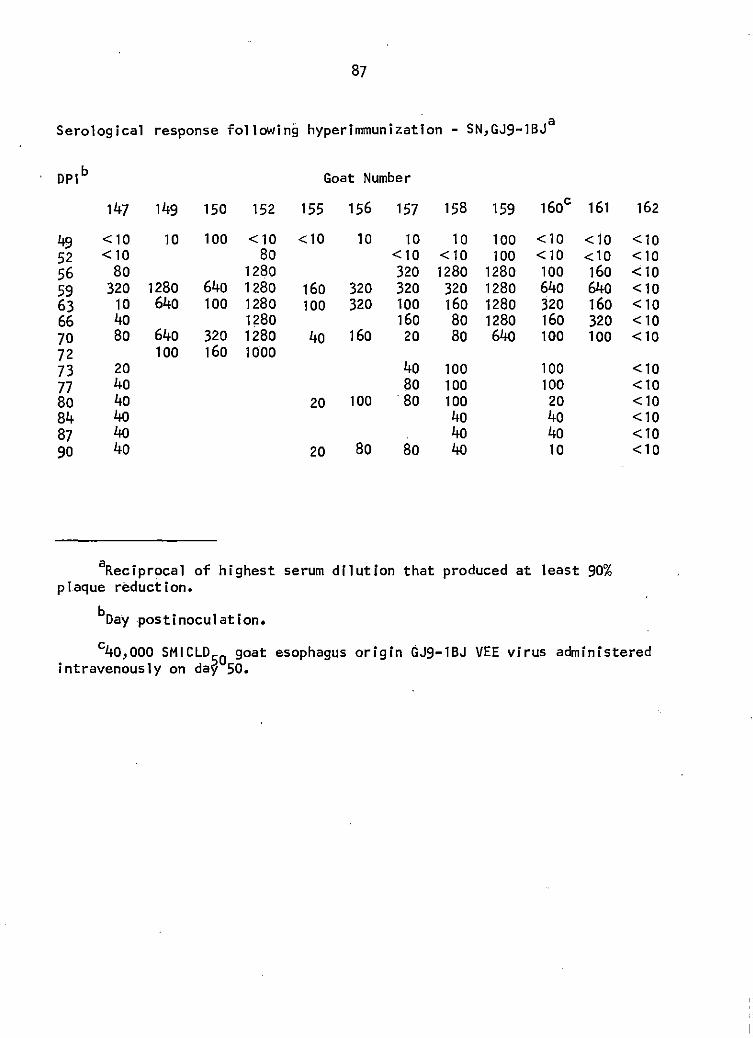
87
Serological response following hyperimmunization - SN,GJ9-1BJ a
DP lb Goat Number
147 149 150 152 155 156 157 158 159 16oc 161
4g <10 10 100 <10 <10 10 10 10 100 <10 <10 52 <10 80 < 10 <10 100 <10 <10 56 80 1280 320 1280 1280 100 160 59 320 1280 640 1280 160 320 320 320 1280 640 640 63 10 640 100 1280 100 320 100 160 1280 320 160 66 4o 1280 160 80 1280 160 320 70 80 640 320 1280 40 160 20 80 640 100 100 72 100 160 1000 73 20 40 100 100 77 40 80 100 100 80 40 20 100 80 100 20 84 4o 40 40 87 4o 4o 40 90 40 20 80 80 4o 10
aReciprocal of highest serum dilution that produced at least 90% plaque reduction.
bDay postinoculation.
162
< 10 < 10 <10 <10 <10 <10 <10
<10 <10 <10 <10 <10 <10
c4o,ooo SMICLD 0 goat esophagus origin GJ9-1BJ VEE virus administered intravenously on da~ 50.
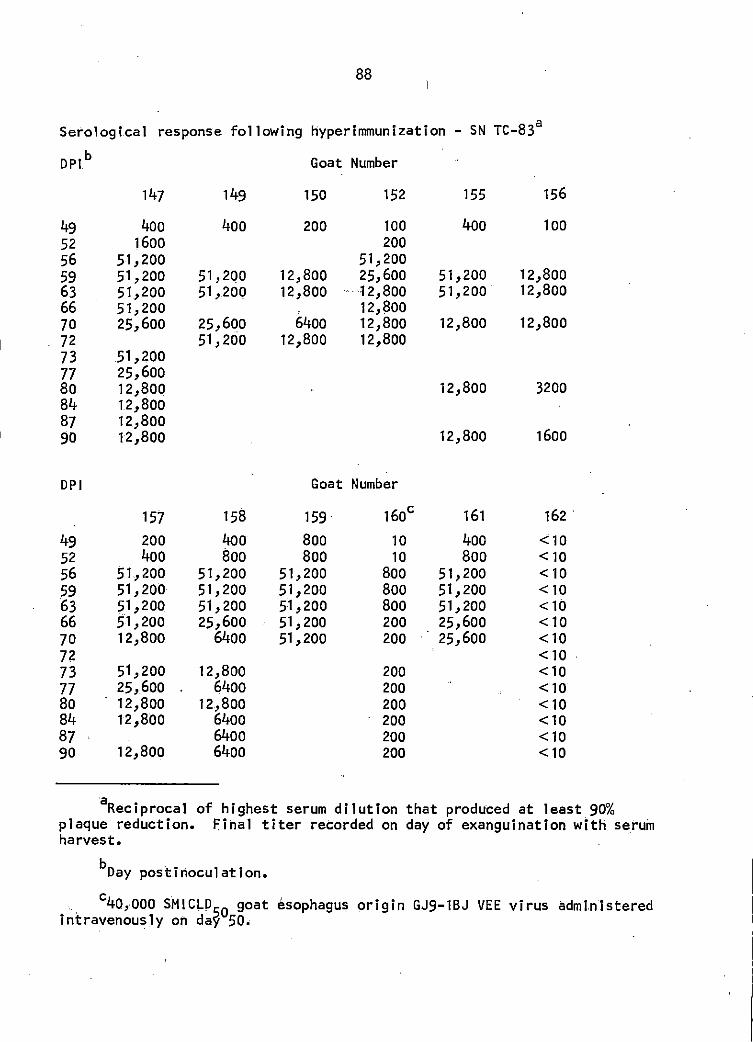
BB
Serological response following hyperimmunization - SN TC-B3a
DP lb Goat Number
147 149 150 152 155 156
49 400 400 200 100 400 100 52 1600 200 56 51,200 51,200 59 51,200 51,200 12,Boo 25,600 51,200 12,Boo 63 51,200 51,200 12,BOO 12,Boo 51,200· 12,Boo 66 51,200 12,Boo 70 25,600 25,600 6400 12,Boo 12,BOO 12,Boo 72 51,200 12,Boo 12,BOO 73 51,200 77 25,600 Bo 12,BOO 12,Boo 3200 B4 12,BOO B7 12,Boo 90 12,BOO 12,BOO 1600
DPI Goat Number
157 15B 159 16oc 161 162 49 200 4oo Boo 10 400 <10 52 400 Boo Boo 10 Boo <10 56 51,200 51,200 51, 200 Boo 51,200 <10 59 51,200 51,200 51,200 Boo 51,200 <10 63 51,200 51,200 51,200 Boo 51,200 <10 66 51,200 25,600 51,200 200 25,600 <10 70 12,Boo 6400 51,200 200 25,600 <10 72 <10 73 51,200 12,BOO 200 <10 77 25,600 6400 200 <10 Bo 12,BOO 12,Boo 200 <10 B4 12,Boo 6400 200 <10 B7 6400 200 <10 90 12,Boo 6400 200 <10
aReciprocal of highest serum dilution that produced at least 90% plaque reduction. Final titer recorded on day of exanguination with serum harvest.
bDay postinoculatlon.
c4o,ooo SMICLD 0 goat intravenously on da? 50.
esophagus origin GJ9-1BJ VEE virus administered

l.
89
BIBLIOGRAPHY
Aitken, T. H. G. 1971. Discussion. Symp. Venezuelan Encephalitis Virus, Health Organ. Sci. Publ. No. 243.
p. 363. In Proc. Workshop Washington, D.C. Pan Amer.
2. Aitken, T. H. G. 1971. Habits of some mosquito hosts of VEE (Mucambo) virus from northeastern South America, including Trinidad. p. 254-255. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
3. Aitken, T. H. G. 1971. VEE (Mucambo) virus vectors in northeastern South America. p. 148-149. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub 1. No. 243.
4. Aitken, T. H. G., L. Spence, A. H. Jonkers, and w. G. Downs. 1969. A 10-year survey of Trinidadian arthropods for natu'ral vi'rus in-fections. J. Med. Entomol. 6:207-215.
5. Ashe, W; K., and A. L. Notkins. 1966. Neutralization of an in-fectious.herpes simplex virus-antibody complex by anti-7 globulin. Proc. Nat. Acad. Sci. U.S.A. 56:447-451.
6. Beck, C. E., and W. G. Wyckoff. 1938. Venezuelan equine encephalo-myel itis. Science 88:530.
7. Bergold, G. H. 1971. Discussion. p. 164-167. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D. C. Pan Amer. Health Organ. Sci. Publ. No. 243.
8. Big 1 er, W. J. 1971. Sero 1 og i c evidence of Venezue 1 an equine encephalitis virus infections in raccoons of south cent,ral Florida. J. WI ldl. Dis. 7: 166-170.
9. Bivin, W. s., C. Barry, A. L. Hogge, Jr., and E. C. Corristan. 1967. Mosquito-induced infection with equine encephalomyel it is virus in dogs. Amer. J. Trap. Med. Hyg. 16:544-547.
10. Bradish, C. J., J. O. Farley, and H. E. N. Ferrier. 1962. Studies
11.
on the nature of the neut ra 1 i zat ion reaction and the .components of the virus system of foot-and-mouth disease. Virology 18:378-400.
Cal isher, C. H. 1971. Discussion. Symp. Venezue 1 an Encepha 1 it is Virus, Health Organ. Sci. Publ. No. 243.
p. 360. In Proc. Workshop Washington, D.C. Pan Amer.

\ 90
12. Calisher, C. H. 1971. Discussion. p. 363. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Anier. Health Organ. Sci. Publ. No. 2.43.
13. Chamberlain, R. W. 1971. Discussion. p. 347. In Proc. Workshop D.C. Pan Amer. , ·Symp. Venezue 1 an Encepha 1 it is Virus, Washington,
Health Organ. Sci. Publ. No. 243.
14. Chamberlain, R. W. 1971. Discussion. p. 357-358. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
15. Chamberlain, R. W. 1971. Sylvatic vectors of Venezuelan virus in south Florida. p. 257-258. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub 1. No. 243.
16. · Chamberlain, R. W. 1971. Venezuelan equine encephalitis: Infection of mosquitoes and transmission. p. 144-147. In Proc •. Workshop Symp. Venezuelan Encephaliti's Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
17. Chamberlain, R. W., and W. D. Sudia. 1961. Mechanism of transmission of viruses by mosquitoes. Annu. Rev. Entomol. 6:371-390.
18. Chamberlain, R. W.~ W. D. Sudia,. P. H. Coleman, and T. H. Work. 1964. Venezuelan equine encephalitis virus from south Florida. Science 145: 272-274,
19. Chamberlain, R. W., W. D. Sudia, T. H. Work, P. H. Coleman, V. F.
20.
Newhouse, and J. G. Johnston, Jr. 1969. Arbovirus studies in.south Florida,. with emphasis on Venezuelan equine encephalomel it.is virus. Amer. J. Epidemiol. 89: 197-210.
Clark, H. F., and C. C. Shepard. preparing fluorescent antibody.
1963. A dialysis technique for Virology 20:642-644.
21. Clarke, D. H., and J. Casals. 1958. Techniques for hemagglutination and hemagglutinatlon-inhibition with arthropod-borne viruses .. Amer. J. Trop. Med. Hyg. 7:561-573.
22. Commun. Dis. Center. litis Surveillance.
1973. p. 1-5. In Venezue I an Equine Encepha-VEE Annu. Summary.
23. Commun. Dis. Center. 1972. Venezuelan Equine Encephalitis . Survei I lance. VEE Summary 8: I.
24. Commun. Dis. Center. 1972. Zoonoses Surveillance. Venezuelan Equine Encephalitis. VEE Monthly Summary l:J-6.

25.
26.
27.
28.
Conimun. Dis. Center. Equine Encephalitis.
91
1972. Zoonoses Surveillance. VEE Summary 1:1.
Venezuelan
Coons, A. H., H. J. Creech, and R. N. Jones. l941. Immunological properties of an antibody containing a fluorescent group. Proc. Soc. Exp. Biol. 47:200-202.
Correa-Giron, P., C. H. Cali sher,. and G. M. Baer. 1972. Epidemic strain of Venezuelan equine encephalornyelitis virus from a vampire· bat captured in Oxaca, Mexico, 1970. Science 175:546-547.
Curtain, C. c, 1961. fluorescein-antibody.
The chromatographic purification of J. Histochern. Cytochern. 9:484-486.
29. Davies, J. B. 1971. Discussion. p. 363. Jn Proc. Workshop Syrnp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Arner. Health Organ. Sci. Publ. No. 243.
30. Davis, M. H., A. L. Hogge, Jr., E. C. Corristan, and J. F. Fernell. 1966. Mosquito transmission of Venezuelan equine encephalornyel it is virus from experirnental ly infected dogs. Arner. J, Trop. Med. Hyg. 15:227-230.
31. Dedmon, R. E., A. W. Holmes, and F. Deinhardt.· 1965. Preparation of fluorescein isothiocyanate-labeled 7-globulin by dialysi.s, gel filtration, and ion-exchange chromatography in combination.· J. Bacterial. 89:734-739.
32. Dickerman, R. W. 1971. Silent hosts: Birds. Workshop Syrnp. Venezuelan Encephalitis Virus, Arner. Health Organ. Sci. Publ. No. 243.
p. 281-283. In Proc. Washington, D.C. Pan
33. Dickerman, R. W., G. J. Baker, J. V •. Ordonez, and W. F. Scherer. 1973. Venezuelan equine encephalornyelitis virernia and antibody responses of pigs and cattle. Arner. J, Vet. Res. 34:357-361.
34. Earley, E., P. H. Peralta, and K. M. Johnson. 1967. A plaque neutralization method for arboviruses. Proc. Soc. Exp. Biol. Med. 125:741-747.
35. Ehrenkranz, N. J., M. C. Sinclair, E. Buff, and D. O. Lyman. 1970. The natural occurrence of Venezuelan equine encephalitis in the United States. New Engl. J. Med. 282:298-302,
36. Eldridge, B. F., B. A. Schiefer, and W. G. Pearson. 1972. U.S. Army activities in the 1971 Venezuelan equine encephalitis outbreak in Mexico and Texas. p·. 5-7. In Proc. 40th Annu. Conf. Calif. Mosquito Control Ass., Inc.

37. Encephalitis surveillance for the Americas. 1972. Centro Panamericano de Zoonosis 1: 1-2.
38.
39.
40.
Fazekas de St. Groth, S., G. S. Watson, and A. F •. Reid. 1958. The neutralization of animal viruses. I. A model of virus-antibody interaction. J. I mmuno 1. 80: 215-224.
Foreign Animal Diseases Report. 1972. myelitis. p. 5-7. In September issue.
Venezuelan equine encephalo-Emergency Programs,
Veterinary Services, APHIS, USDA.
Fossaert, H. 1971. Endeinic behavior in man. Workshop Symp. Venezuelan Encephalitis Virus, Amer. Health Organ. Sci. Publ. No. 243.
p. 291-296. In Proc. Washington) D.C. Pan
41. Franck, P. T. 1971. Significance of geographic distribution of VEE vi.rus variants. p. 322-328 •. In Proc. Workshop· Symp. Venezuelan Encephal ltis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub 1. No. 243.
42. Franck, P. T., and K. M. Johnson. 1971. An outbreak of Venezuelan equine encephalomyelitis in Central America. Amer. J. Epidemiol. 94:487-495.
43. Franck, P. T., and K. M. Johnson. 1970. An outbreak of Venezuelan. encephalitis in man in the Panama Canal Zone. Amer. J: Trap. Med. Hyg. 19: 860-865.
44. Frommhagen, L. H. 1965. The solubility and other physicochemical properties of human r-globulin labeled with fluorescein iso-
45.
th i ocyanate. J. Immune l. 95: 442-445.
Galindo, P. p. 249-253. Washington,
1971. Endemic vectors of Venezuelan encephalitis. In Proc. Workshop Symp. Venezuelan Encephalitis Virus,
D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
46. Galindo, P., and M.A. Grayson. 1971. Culex (Melanoconion) aikenii: Natural vector in Panama of endemic Venezuelan equine encephalitis. Science 172:594-595.
47. Galindo, P., S. Syrihongse, E. De Rodaniche, and M. A. Grayson. 1966. An ecological survey for arboviruses in Almirante, Panama, 1959-1962. Amer. J. Trop. Med. Hyg. 15:385-400.
48. Goldman, M. 1968. Fluorescent Antibody Methods. Academic Press, New York. 503 p.

49.
so.
93
Goldstein, G., I. S. Sllzys, and M. W. Chase. 1961.· Studies on fluorescent antibody staining. I. Non-specific fl·Liorescence with fluorescein-coupled ·sheep anti-rabbit globulins. J. Exp. Med. 114:89-111.
G 11 A G D J B·ardaw"1ll, and M. W. David. 1949·~ Detennina-orna , • ., • • tion of serum proteins by means of the biuret reaction. J. Biol. Chem. 177:751-756.
51. Grayson, M. A. 1971. Discussion. p. 284-285, 289. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pubi. No. 243.
52. Grayson, M. A. 1971. Open discussion. p. 276-277. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
53.
54.
55.
Grayson, M. A., and P. Galindo. 1969. Ecology of Venezue]a.n equine encephal it ls virus in Panama. J. Amer. Vet. Med. Ass. 155:2141-2145.
Grayson, M. A., and P. Galindo. 1968. Epidemiologic studies of Venezuelan equine encephalitis virus in Almirante, Panama. Amer. J. Ep i demi o 1. 88: 80-96.
Groot, H. 1971. Discussion. Venezuelan Encephalitis Virus, Organ. Sci. Publ. No. 243.
p. 244. ln·Proc. Workshop Symp. Washington, D.C. Pan Amer. Health
56. Groot, H. 1972. Epidemiology of the epizootic form of Venezuelan equine encephalitis and its impact on the economies and health of the nations of the Americas. p. 117-124. In IV. Inter-American Meeting on· Foot-and-Mouth Disease and Zoonoses Control. Pan Amer. Health Organ., World Health Organ., Washington, D.C.
57. Groot, H. 1971. The health and economic impact of Venezuelan equine encephalitis (VEE). p. 7-16. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub 1. No. 243. ·
58. Hahon, N. 1970. Neut ra 1 i zat ion of residua 1 infect iv i ty of Venezue 1 an
59.
60.
equine encephalomyelitis virus by anti-gamma globulin. J. Gen. Viral. 6:361-372.
Hahon, N. antiserum 4: 77-88.
1969. The kinetics of neutralization of VEE virus by and the reversibility of the reaction. J. Gen. Viral.
Hardy, F. M., and A. Brown. 196 I. encepha 1omye1 i ti s virus i n L ce I I s. J. Bacterial. 81:20-27.
Growth of Venezuelan equine I. Growth in monolayer cultures.

94
61. Hayes, R. O., C. H. Ternpelis, A. D. Hess, and W. C. Reeves. 1973.
62.
Mosquito host preference studies in Hale County, Texas. Amer. J. Trop. Med. Hyg. 22:270-277.
Henderson, B. Sudia. 1970. of Venezuelan 93: 194-205.
E.", W. A. Chappel I, J. G. Johnston, Jr., and W. D. Experimental infection of horses with three strains
equine encephalomyelitis virus. Amer. J. Epidemiol.
63. Hess, A. D. 1971. Discussion. p. 354. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub!. No. 243.
64. Hess, A. D. 1971. Discussion. p. 360. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub I. No. 243.
65. Hess, A.·o. 1971. Discussion. p. 365. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health. Organ. Sci. Pub!. No. 243.
66. Hinman, A. R~, J. E. McGowan, Jr., and B. E. Henderson. 1971. Venezuelan equine encephalomyelitis; Surveys of human ·illness during an epizootic in Guatemala and El Salvador. Amer. J. Epidemiol. 93: 130-136.
67. Hoff, G. L., and D. o. Trainer. 1972. Experimental infection of Venezuelan equine encephalomyelitis virus in white-tailed deer. Amer. J. Epidemiol. 96:379-382.
68. Johnson, K. M. 1971. Discussion. p. 330. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub]. No. 243.
69. Johnson, K. M. 1972. Properties and pathogenicity of Venezuelan equine encephalitis viruses. p. 113-116. In IV. Inter-American Meeting on Foot-and-Mouth .Disease and Zoonoses Control. Pan Amer. Health Organ;, World Health Organ., Washington, D.C.
70. Jones, R. H., .H. W. Potter, Jr., and H. A. Rhodes. 1972. Ceratopogonidae attacking horses in south Texas during the 1971 VEE epidemic. Mosquito News 32:507-509.
71. Jonkers, A. H. 1971. Si lent hosts of Venezuelan equine encephalitis (VEE) virus. p. 263~267. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.

72.
73.
74.
75.
95
Jonkers, A. H., L. Spence, W. G. Downs, T. H. G. Aitken, and C. E. ·Worth. 1968. Arbovi rus studies in Bush-Bush Forest, Trinidad, W. I., September 1959-December 1964. VI. Rodent-associated viruses (VEE and agents of groups C and Guama): Isolations and further studies. Amer. J. Trap. Med. Hyg. 17:285-298.
Ke 11 er, R. 1968. Stu di es on the mechanism of the enzymatic reactivation of antibody-neutralized pol iovi rus. J. lmmunol. 100: 1071-1079.
Kissling, R. E., and R. ~. Chamberlain. encephalomyelitis. In C. A. Brandly and in. Veterinary Science 11 :65-84.
1967. Venezuelan equine C. Cornelius (ed.) Advances
Knowles, R. C. United States. 209.
1971. VEE in South and Central America,. Mexico, Amer. Ass. Equine Practitioners Annu. Proc. 17:200-
76. Kubes, V. 1944. Venezue 1 an type of equine encepha 1omye1 it is virus in Trinidad. Amer. J. Vet. Res. 5:30-31.
77. Kubes, V., and F. A. Rios. In Venezuela. Advance data Comp. Med. 3: 43-44.
1939. Infectious equine encephalomyel it is concerning the causative agent. Can. J •.
7B. Lafferty, K. J. 1963. The interaction between virus .and antibody. I. Ki.netic studies. Virology 21:61-75.
79. Lenette, E. H., and N. J. Schmidt. Viral and Rickettsial Infections. Ass., Inc., New York. 369 p.
1969. Diagnostic.Procedures for Fourth edition. Amer. Pub. Health
80. Lord, R. D. 1971. Open discussion. p. 277. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
81. Lord, R. D. 1970. Some vertebrate host aspects of arbovirus ecology. J, Wi ldl. Dis. 6:236-238. '
82. Lord, R. D., C. H. Cal isher, W. D. Sudia, and T. H. Work. 1973. Ecological lnvestlgatlons of vertebrate hosts of Venezuelan. equine encephalomyelitis virus in Florida. Amer. J. Trap. Med. Hyg.
83.
22: 116-123.
Lovejoy, T. 1971. Discussion. p. 286-288. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.

84. Lowry, O. H., N. J. Rosebrough, A. L. Farr, and R. J. Randal 1. 1951. Protein measurement with the Falin phenol reagent. J. Biol. Chem. 193:265-275.
85. Mackenzie, R. B. 1971. The role of silent vertebrate hosts in epidemics of Venezuelan encephalitis. p. 239-243. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
86.
87.
McConnell, S. 1972. Present, and Future.
McDavitt, H. o., J. A. H. Coons. 1963. labeled antisera by
Venezuelan equine encephalomyelitis: Past, J. Amer. Vet. Med. Ass. 161: 1579-1583.
H. Peters, L. W. Pol lard, J. G. Harter, and Purification and analysis of fluorescein-
column chromatography. J. lmmunol. 90:634-642.
88. McKinney, R. M., J. T. Spillane, and G. W. Pearce. 1964. Factors affecting the rate of reaction of fluorescein isothiocyanate with serum proteins. J. lmmunol. 93:232-243.
89. McKinney, R. W. 1971. Discussion. Symp. Venezuelan Encephalitis Virus, Health Organ. Sci. Publ. No. 243.
p. 361-362. In Proc. Workshop Washington, D.C. Pan Amer:.
90. Meng~ling, W. L., D. E. Gutekunst, A. L. Fernelius, and E. D. Pirtle. 1963. Demonstration of hog cholera and bovine viral diarrhea . vi ruses by immunofluorescence. Can. J. Comp. Med. Vet. Sci. 27: 162-164.
91. Notkins, A. L., M. Mage, W. K. Ashe, and S. Mahar. 1968. Neutraliza-tion of sensitized lactic dehydrogenase by anti-r globulin. J. lmmunol. 100:314-320.
92. Notkins, A. L., S. Mahar, C. Scheele, and J. Goffman. 1966. In-fectious virus-antibody complex in the blood of chronically infected mice. J. Exp. Med. 124:81-97.
·93. Ordonez, J. V. 1971. Open discussion. p. 289. In Proc. Workshop Symp.· Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
94. Parker, R. L., P. B. Dean, and R. B. Zehmer. 1973. Public health aspects of Venezuelan equine encephalitis. J. Amer. Vet. Med, Ass. 162: 777-779.
95. Peralta, P. H., and A. Shelekov. 1966. Isolation and characteriza-tion of arboviruses from Almirante, Republic of Panama. Amer. J, Trap. Med. Hyg. 15:369-378.

96.
97.
97
Ploem, J, S. 1971. A study of filters and I ight sources. in immunofluorescence microscopy. Ann. N. Y. Acad. Sci. 177:414-429.
Riggs, J. L., P. C. Loh, and W. C. Eveland. 1960. A simple fractionation method for preparation of fluorescein-labeled gamma globulin. Proc. Soc. Exp. Biol. Med. 105:655-658.
98, Riggs, J. L., R. J, Seiwald, J. H. Burckhalter, C. M. Downs, and T. G. Metcalf. 1958. lsothiocyanate compounds as fluorescent labeling agents for immune serum. Amer. J. Pathol. 34:1081-1091.
99, Sanmartin, c. 1971. Open discussion. p. 247-248. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
JOO. Sanmarti·n, C. 1971. Open discussion. p. 276. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub!. No. 243.
10 I.
. 102.
Scherer, W. F. 1971. Discussion. p. 361. In Proc. Symp. Venezuelan Encephalitis Virus, Washington, D.C. Health Organ. Sci. Pub!. No. 243.
Workshop Pan Amer.
Scherer, W. F. 1971 .. Open discussion. p. 276; Symp. Venezue 1 an Encepha 1 it.is Virus, Washington, Health Organ. Sci. Pub!. No. 243.
In Proc. Workshop D.C. Pan Amer.
103. Scherer, W. F. 1971. Open discussion. p. 353. In Proc. Workshop Symp. Vene.zuelan Encephalitis Virus, Washington, D.C. Pan Amer. Hea I th 0 rgan. Sci. Pub 1. No. 243.
104. Scherer, W. F. 1972. Problems and prospects for control of animal and human disease caused by Venezuelan equine encephalitis viruses. p. 130-132. In IV. Inter-American Meeting on Foot-and-Mouth Disease and Zoonoses Control, Pan Amer. Health Organ., World Health Organ., Wash I ngton, D. C.
105. Scherer, W. F., R. W. Dickerman, A. Diaz-Najera, B. A. Ward, M. H. Mi lier, and P. A. Schaffer. 1971. Ecologic stud.ies of Venezuelan encepha 1 it is in southeastern Mexico. I 11 . Infection of mosquitoes. Amer. J. Trap. Med. Hyg. 20:969-979.
106 •. S.cherer, W. F., R. W. Dickerman, R. P. LaFiandra, C. Wong Chia, and J. Terrian. 1971. Ecologic studies of Venezuel.an encephalitis in. southeastern Mexico. IV. Infections of wild· mammals. Amer. J. Trap. Med. Hyg. 19:703-711. .

98
107. Scherer, W. F., R. W. Dickerman, and J. V. Ordonez. 1970. Discovery and geographic distribution of Venezuelan encephalitis virus in Guatemala, Honduras, and British Honduras during 1965-1968 and its possible movement to Central America and Mexico. Amer. J. Trop. Med. Hyg. 19:703-711.
108. Scherer, W. F., R. W. Dickerman, C. Wong Chia, A. Ventura, A. Moorhouse, R. Geiger, and A. Diaz-Najera. 1964. Venezue.l an equ i rie encephalitis virus in Veracruz, Mexico and the use of sentinels. Science 145:274-275.
109. Scherer, W. F., J. V. Ordonez, P. B. Pancake, and R. W. Dickerman. 1972. Observations of equines, humans and domestic and wild vertebrates during the 1969 equine epizootic and epidemic of Venezuelan encephalitis in Guatemala. Amer. J. Epidemiol. 95: 255-266.
110. Se! lers, R. F., G. H. Bergold, O. M. Suarez, and A. Morales. 1965.
Ill.
Investigations during Venezue I an equine encephalitis outbreaks in Venezuela-1962-1964. Amer. J. Trop. Med. Hyg. 14:460-469.
Shelekov, A. 1971. 'Discussion. p. 362. Venezuelan Encephalitis Virus, Washington, Organ. Sci. Pub!. No. 243.
In Proc. Workshop Symp. D.C. Pan Amer. Health
112. Shope, R. E. 1971. Discussion. p. 271-273. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. · Pan Amer. Health Organ. Sci. Pub!. No. 243.
113. Si.dwell, R. W., L. P. Gebhardt, and B. D. Thorpe. 1967. Epidemi-ological aspects of Venezuelan equine encephalitis virus infections. Bacteriol. Rev. 31:65-81.
114. Spertzel, R. O. 1971. Venezuelan equine encephalomyelitis. p. 162-165. In. Proc. 75th Annual Meeting U.S. Animal Health Ass., Oklahoma City, Oklahoma.
115. Sudia, W. D. 1971. Arthropod vectors of Venezuelan equine encephalitis. p. 157-161. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub I. No. 243.
I 16. S~dia~ W. D., R. D. Lord, V. F. Newhouse, O. L. Mi 1 ler, and R. E. Kissi 1ng. 1971. Vector-host studies of an eplzootic of Venezuelan equine encephalomyelitis in Guatemala, 1969. Amer. J. Epidemiol. 93: 137-243.
117. Sudla, W. D., V. F. Newhouse, and L. D. Beadle. 1971. equine encephalitis. p. 1-2. In July 23 issue Vector Briefs, Cornnun. Dis. Center, Atlanta, Georgia.
Venezuelan Control

99
118. Taber, L. E., A. L. Hogge, Jr., and R. W. McKinney. 1965. Experi-menta I infection of dogs with two strains of Venezue·I an equ roe encephalomyelitis virus. Amer. J. Trop. Med. Hyg. 15:647-651.
119. Thorpe, B. D., K. L. Smart, and R. W. Sidwel I. 1965. Arbovi rus complement fixi.ng antibodies in sera of wi Id! ife in west central Utah. Proc. Soc. Exp. Biol. Med. 118:179-181.
120. Trapido, H. 1971. Discussion. p. 358. In Proc. Workshop Symp. Venezuelan Encephalitis Virus; Washington, D.C. Pan Amer. Health Organ. Sci. Pub!. No. 243.
121. Trapido, H. 1971. Discussion. p. 163-164, In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
122. Trapido, H. 1971. Discussion. p. 348-349. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub!. No. 243.
123. Vene~uelan equine encephalomyelitis. 1972. Report of the USAHA cormnlttee on infectious diseases of horses. U.S. Animal Health Ass. ?5th Annual Meeting. J. Amer. Vet. Med. Ass. 160:87.
124, Ventura, A. K. 1971. Open discussion. p. 153. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Pub I. No. 243.
125. Veterinary Services. 1973. The origin and spread of Venezuelan equine encephalomyelitis. APHIS 91-10, U.S.D.A., Hyattsville, Maryland. 51 p.
126. Vilchis Villasenor, A. 1971. Discussion. p. 360. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
127. Wallis, C., and J. L. Melnick. 1967. Virus aggregation as the cause of the non-neut ra Ii zab I e persistent fraction. J. Vi ro I. I: 478-488.
128. Walton, T. E. 1971. Open discussion. p. 357. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Hea I th Organ. Sci. Pub I. No. 243.
129. Walton, T. E., and K. M. Johnson. 1972. Epizootiology of Venezuelan equine encephalomyel it is in the Americas. J. Amer. Vet. Med. Ass. 161:1509-1515.

100
130. Walton, T. E., F. E. Brautigam, J. A. Ferrer, and K •. M. _Johnson. 1972. Epizootic·venezuelan equine encephalomyelitis in Central America. Disease·patterh and vaccine evaluation i.'n Nicaragua, 1969-1970. Amer. J. Epidemiol. 95:247-254.
131. Work, T. H. 1971. On the natural history of Venezuelan equine encephalitis: Conclusions and correl~tions. p. 333-343. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
132. Yamamoto, A., and A. Kawamura, Jr. 1970. Ass.essment of conjugates. p. 181-183. In E. J. Holborow (ed.) Standardization in immuno-fluorescence. Blackwei l Scientific Publ I cations, Oxford and Edinburgh.
133. Young, N. A. 1972. Origin of epidemics of Venezuelan encephalitis. J. Inf. Dis. 125:565-567.
134. Young, N. A., and K. M. Johnson. 1969. Antigenic variants of Venezuelan equine encephalomyelitis virus: Their geographic distribu-tion and epidemiologic significance. Amer. J. Epidemiol. 89:286-307.
135. Zarate, M. L. 1971. Virulence aspects of VEE viruses. p. 124-139. In Proc. Workshop Symp. Venezuelan Encephalitis Virus, Washington, D.C. Pan Amer. Health Organ. Sci. Publ. No. 243.
136. Zarate, M. L., and W. F. Scherer. 1969. A comparative study of virulence, plaque morpholog.ies and ant"igenic characteristics of Venezuelan encephalitis virus strains. Amer. J. Epidemiol. 89:489-502.

101
ACKNOWLEDGEMENTS
The author wishes to thank Doctor c. J. Mare for his counsel and
guidance in the development and completion of this thesis.
This study was conducted with the guidance and general supervision
of Doctor E. A. Carbrey.
The author is indebted to the"'late Doctor E. P.· Pope who through.
his vision established this two-year advanced veterinary· training
program.
The author extends his appreciation to Doctor E. C. Sharman and
the Emergency Programs staff who made this developmental program
possible by their support.
Appreciation is expressed to Doctor J. E. Pearson and Messrs. G.
Gustafson, D. Senne, and M. Snyder for their assistance in the effectuation
of laboratory principles and techniques, and to Mister R. M. Glaz·ier and
his staff for preparation .of the excellent graphs and photographs.




















